
Occurrence of Enterococcus species with virulence markers in an urban flow-influenced tropical...
13
Occurrence of Enterococcus species with virulence markers in an urban flow-influenced tropical recreational beach Asmat Ahmad a,⇑ , Ayokunle Christopher Dada a,b,⇑ , Gires Usup c , Lee Yook Heng d a School of Biosciences and Biotechnology, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysia b Institute of Ecology and Environmental Studies, Obafemi Awlowo University, Ile-Ife, Nigeria c School of Environmental & Natural Resource Sciences, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysia d School of Chemical Sciences and Food Technology, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysia article info Keywords: Antibiotic resistance Beach water Enterococcus Sequence type, Virulence factors abstract Median enterococci counts of beach water samples gradually increased at statistically significant levels (v2: 26.53, df: 4; p < 0.0001) with increasing proximity to river influx. The difference in proportion of antibiotic resistant enterococci in beach water and river water samples was statistically significant (p < 0.05) for the tested antibiotics with river isolates generally presenting higher resistance frequencies. Virulence genes cyl, esp, gelE and asa were detected at varying frequencies (7.32%, 21.95%, 100% and 63.41% respectively) among river isolates. On the other hand, the prevalence of these genes was lower (0%, 20%, 67.27% and 41.82% respectively) among beach water isolates. Multi-Locus-Sequence-Typing analysis of Enterococcus faecalis presented four sequence types (ST) one of which shared six out of seven tested loci with ST6, a member of the clonal complex of multi-drug resistant strains associated with hos- pital outbreaks. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction The impact of external influence on surface water quality is well documented (Bhaduri et al., 2001; Schippmann et al., 2013; Mallin et al., 2000). Anthropogenic influences (urbanization, industrial and agricultural activities) as well as complex natural processes (changes in precipitation inputs, erosion, and weathering of crustal materials) are all implicated in the degradation of surface water quality (Zhang et al., 2009). These characteristically impair their use for recreation or other purposes (Peng et al., 2005). Surface water contamination in tropical and subtropical environments has been associated with waterborne diseases (Peng et al., 2005). Specifically, rivers in Malaysia have been reportedly treated as open sewers with attendant waste discharges which ultimately reach the sea (DailyExpress: Rubbish Ending Up in the sea, 2013). Pollutants from municipally-influenced sources may carry diverse bacteria which could directly pose threats to the health of recreational beach users (Barrell et al., 2000; Hamilton et al., 2010). Enterococci are commonly used as indicators of faecal contamination in recreational waters (Barrell et al., 2000). Studies that highlight enterococci diversity may play critical epidemiolog- ical roles in source pollution monitoring of surface waters. To date, there is no published study on the genetic variability of enterococci recovered from recreational beach water in Malaysia. Enterococcus spp. are documented to be intrinsically resistant to a number of antibiotics including cephalosporins, penicillinase- resistant penicillins, and clinically available concentrations of lincosamides and aminoglycosides (Fisher and Phillips, 2009). Apart from the notoriety of members of this genus for antibiotic resistance, enterococci may possess a number of virulence factors which are associated with the severity and duration of infections caused by them. These include gelatinase, enterococcal surface protein (Esp), aggregation substance (asa), cytolysin (cyl) and hyaluronidase (hyl) Jett et al., 1994; Vergis et al., 2002; Semedo et al., 2003. While the occurrence of virulence strains has been extensively studied among clinical enterococci (Padilla and Lobos, 2013), studies are only recently beginning to emerge on the elucidation of virulence genes among enterococci recoverable from recreational beach waters is (Brownell et al., 2007; Santiago-Rodriguez et al., 2013; Layton et al., 2009; Rathnayake et al., 2012). The scanty published information partly explains why the ecology of antibiotic resistance among environ- mental strains of enterococci is still not well understood (Santiago-Rodriguez et al., 2013; Rathnayake et al., 2012). Furthermore, there is a dearth of information on the molecular http://dx.doi.org/10.1016/j.marpolbul.2014.03.028 0025-326X/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding authors. Address: School of Biosciences and Biotechnology, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysia. Tel.: +60 149390730. E-mail addresses: [email protected] (A. Ahmad), [email protected](A.C. Dada). Marine Pollution Bulletin xxx (2014) xxx–xxx Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococcus species with virulence markers in an urban flow-influenced tropical recreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marpolbul.2014.03.028
Transcript of Occurrence of Enterococcus species with virulence markers in an urban flow-influenced tropical...

Marine Pollution Bulletin xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Occurrence of Enterococcus species with virulence markers in an urbanflow-influenced tropical recreational beach
http://dx.doi.org/10.1016/j.marpolbul.2014.03.0280025-326X/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding authors. Address: School of Biosciences and Biotechnology,Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKMBangi, Malaysia. Tel.: +60 149390730.
E-mail addresses: [email protected] (A. Ahmad), [email protected] (A.C. Dada).
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococcus species with virulence markers in an urban flow-influenced trecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marpolbul.2014.03.028
Asmat Ahmad a,⇑, Ayokunle Christopher Dada a,b,⇑, Gires Usup c, Lee Yook Heng d
a School of Biosciences and Biotechnology, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysiab Institute of Ecology and Environmental Studies, Obafemi Awlowo University, Ile-Ife, Nigeriac School of Environmental & Natural Resource Sciences, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysiad School of Chemical Sciences and Food Technology, Faculty of Science & Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Malaysia
a r t i c l e i n f o
Keywords:Antibiotic resistanceBeach waterEnterococcusSequence type, Virulence factors
a b s t r a c t
Median enterococci counts of beach water samples gradually increased at statistically significant levels(v2: 26.53, df: 4; p < 0.0001) with increasing proximity to river influx. The difference in proportion ofantibiotic resistant enterococci in beach water and river water samples was statistically significant(p < 0.05) for the tested antibiotics with river isolates generally presenting higher resistance frequencies.Virulence genes cyl, esp, gelE and asa were detected at varying frequencies (7.32%, 21.95%, 100% and63.41% respectively) among river isolates. On the other hand, the prevalence of these genes was lower(0%, 20%, 67.27% and 41.82% respectively) among beach water isolates. Multi-Locus-Sequence-Typinganalysis of Enterococcus faecalis presented four sequence types (ST) one of which shared six out of seventested loci with ST6, a member of the clonal complex of multi-drug resistant strains associated with hos-pital outbreaks.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The impact of external influence on surface water quality is welldocumented (Bhaduri et al., 2001; Schippmann et al., 2013; Mallinet al., 2000). Anthropogenic influences (urbanization, industrialand agricultural activities) as well as complex natural processes(changes in precipitation inputs, erosion, and weathering of crustalmaterials) are all implicated in the degradation of surface waterquality (Zhang et al., 2009). These characteristically impair theiruse for recreation or other purposes (Peng et al., 2005).
Surface water contamination in tropical and subtropicalenvironments has been associated with waterborne diseases (Penget al., 2005). Specifically, rivers in Malaysia have been reportedlytreated as open sewers with attendant waste discharges whichultimately reach the sea (DailyExpress: Rubbish Ending Up in thesea, 2013). Pollutants from municipally-influenced sources maycarry diverse bacteria which could directly pose threats to thehealth of recreational beach users (Barrell et al., 2000; Hamiltonet al., 2010). Enterococci are commonly used as indicators of faecalcontamination in recreational waters (Barrell et al., 2000). Studies
that highlight enterococci diversity may play critical epidemiolog-ical roles in source pollution monitoring of surface waters. To date,there is no published study on the genetic variability of enterococcirecovered from recreational beach water in Malaysia.
Enterococcus spp. are documented to be intrinsically resistant toa number of antibiotics including cephalosporins, penicillinase-resistant penicillins, and clinically available concentrations oflincosamides and aminoglycosides (Fisher and Phillips, 2009).Apart from the notoriety of members of this genus for antibioticresistance, enterococci may possess a number of virulence factorswhich are associated with the severity and duration of infectionscaused by them. These include gelatinase, enterococcal surfaceprotein (Esp), aggregation substance (asa), cytolysin (cyl) andhyaluronidase (hyl) Jett et al., 1994; Vergis et al., 2002; Semedoet al., 2003. While the occurrence of virulence strains has beenextensively studied among clinical enterococci (Padilla and Lobos,2013), studies are only recently beginning to emerge on theelucidation of virulence genes among enterococci recoverablefrom recreational beach waters is (Brownell et al., 2007;Santiago-Rodriguez et al., 2013; Layton et al., 2009; Rathnayakeet al., 2012). The scanty published information partly explainswhy the ecology of antibiotic resistance among environ-mental strains of enterococci is still not well understood(Santiago-Rodriguez et al., 2013; Rathnayake et al., 2012).Furthermore, there is a dearth of information on the molecular
ropical

2 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
determination of virulence markers among enterococci recover-able from tropical recreational beach waters (Santiago-Rodriguezet al., 2013).
The aim of the current study is, thus, to determine via molecularcharacterization, the species prevalence, diversity, antimicrobialresistance and virulence markers among enterococci recoveredfrom a tropical recreational beach in the East Coast of Malaysia.
2. Materials and methods
2.1. Study area description
The coast of northern Pahang, Malaysia is composed of the threeprominent bays (Cendur, Cerating and Beserah) and the minor baysof Pelindung and Chempedak (Tiong, 2001). Teluk Chempedak isKuantan’s most popular beach located 5 km east from Kuantantown centre, Pahang, Malaysia. Teluk Chempedak is usuallycrowded during the weekends with people engaging in a numberof sporting activities. Along the Chempedak bay, River Chempedakdrains into the sea just overhead the main bathing area at TelukChempedak. There is also a stormwater drainage system that emp-ties into this river at the brink of influx into the sea (Fig. 1).
2.2. Sample collection
Bathing water samples for both beaches were taken from areasclose to the swash zone of the bathing beach at locations that coverthe length of the beach (Fig. 1). Water samples were collected alsofrom the river draining into the sea at various locations and at thearea of influx into the seawater. Sterile glass bottles (1000 ml)were used to collect water samples in triplicates. Sand sampleswere collected in sterile plastic containers. Faecal samples werecollected at toilets proximate to the beach.
2.3. Isolation and enumeration of enterococci
Bacterial densities of enterococci from seawater samples weredetermined by membrane filtration method (A.P.H.A, 1999), usingSlanetz and Bartley (S + B) culture media. Plates were incubated at37 �C for 24–48 h. Preliminary tests on presumptive enterococciwere performed as previously described (Dada et al., 2013). All iso-lates were designated as members of the genus Enterococcus by thePCR detection of the tuf gene as described by Creti et al. (2004). Iso-lates were also confirmed by the amplification of sodA and ddlgenes (Enterococcus faecalis and Enterococcus faecium) (Jacksonet al., 2004; Dutka-Malen et al., 1995) and 16SrRNA sequencing(for identification of other Enterococcus spp.). Primers are listedin Table 1.
2.4. Multi Locus Sequence Typing
Multi-drug resistant E. faecalis isolates with virulence traits wereselected for Multi Locus Sequence Typing (MLST). MLST analysisinvolved a total of seven housekeeping genes: gdh (glucose-6-phos-phate dehydrogenase), gyd (glyceraldehyde-3-phosphate dehydro-genase), pstS (phosphate ATP binding cassette transporter), gki(putative glucokinase), aroE (shikimate 5-dehydrogenase),xpt (shikimate 5-dehydrogenase), and yiqL (acetyl-coenzyme A ace-tyltransferase) (Table 1) (Ruiz-Garbajosa et al., 2006). PCR condi-tions for all amplification reactions were as follows: initialdenaturation at 94 �C for 5 min; 30 cycles at 94 �C for 30 s, 52 �Cfor 30 s, and 72 �C for 1 min; and final extension at 72 �C for7 min. Reactions were performed in 25-ll volumes using Taq poly-merase (FirstBase Sdn Bhd). The resulting PCR amplicons were puri-fied with a kit (Qiagen) and sequenced using an ABI 3130XL 20
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
genetic analyzer (Applied Biosystems). For each locus, a distinct al-lele number was assigned to the obtained nucleotide sequences inaccordance with the E. faecalis MLST database (http://efaecal-is.mlst.net/).
2.5. Nucleotide sequence accession numbers
Nucleotide sequences determined in this study were submittedto GenBank (http://www.ncbi.nlm.nih.gov/genbank/). The depos-ited information was simultaneously made available to EMBL inEurope and the DNA Data Bank of Japan. sodA gene sequenceswere submitted under GenBank accession numbers KC603859-KC603862; ddl gene sequences were submitted under GenBankaccession numbers KC594678-KC594682; 16S rRNA genesequences were submitted under GenBank accession numbersKC890838-KC890842, KC707577-707586.
2.6. Detection of antibiotics resistance and resistance determinantsamong Enterococci
Antibiotic resistance among enterococci isolated from thebeaches was determined by the single disc Kirby-Bauer diffusionmethod using Mueller–Hinton Agar (Oxoid, UK) NCCLS: Perfor-mance Standards for Antimicrobial Disk Susceptibility Testing,2003. A total of 96 enterococci isolates recovered from TelukChempedak beach water and beach sand, River Chempedak waterand sand were tested for resistance to seven (7) antibiotic groups.Antibiotics tested include vancomycin (V) (glycopeptides) (30 lg),kanamycin (K) (30 lg), streptomycin (S) (25 lg) (aminoglycoside),ampicillin (A) (10 lg) (B-lactam), tetracycline (T) (30 lg) (tetracy-cline), chloramphenicol (C) (30 lg) (chloramphenicol) and nitrofu-ranoin (N) (50 lg) (nitrofuran) (Oxoid, UK). Diameters of zones ofinhibition were recorded in mm and interpreted as sensitive or resis-tant using breakpoints for enterococci as proposed by the NationalCommittee for Clinical Laboratory Standards (NCCLS: PerformanceStandards for Antimicrobial Disk Susceptibility Testing, 2003).
2.7. Determination of virulence-markers distribution in enterococci
Polymerase chain reaction (PCR) assays were applied to amplifyvirulence determinants {aggregation substance (asa), cytolysin(cylA), enterococcal surface protein (esp), gelatinase (gelE)}(Vankerckhoven et al., 2004). Details of primers are listed inTable 2. For each primer, initial optimization experiments wereconducted to ascertain optimal pcr conditions for MgCl2 andannealing temperatures. Multiplex PCR conditions used in thisstudy included an initial activation step at 95 �C for 4 min followedby 30 cycles of denaturation at 94 �C for 1 min, annealing at 55 �Cfor 1 min, extension at 72 �C for 1 min and a single cycle of 7 min at72 �C. Reference strains were gratefully provided by Prof. Shankarof the Department of Medicinal Chemistry and Pharmaceutics,University of Oklahoma and Dr. Fatimah Lopez, Institute ofBioscience, Brazil. PCR amplicons of asa, cylA, esp, gelE genes wereconfirmed by DNA sequencing with an ABI 3130XL 20 geneticanalyzer (Applied Biosystems). The DNA sequences were blastedfor sequence similarity with annotated sequences at http://www.ncbi.nlm.nih.gov.
2.8. Phenotypic assays
All isolates were subjected to gelatinase assay as described byCariolato et al. (2008) using Brain–Heart Infusion agar platessupplemented with 10 g/L peptone and 30 g/L gelatine. Assay forproteolytic activity was conducted by detecting casein hydrolysisin MHA containing 3% (w/v) skimmed milk. Biofilm formationwas determined by crystal violet assay in polystyrene microtiter
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

Fig. 1. (a) Map of sampled area (b) Teluk Chempedak showing the bathing beach and the combined loadings of river water and stormwater discharge.
A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 3
plates and subsequent examination at an optical density of 570 nm(OD570) (Mohamed et al., 2004).
2.9. Statistical analysis
Strata�, gratefully provided by the Harvard School of PublicHealth in collaboration with Strata Inc. (USA), was used for theanalyses. Each of the responses from the phenotypic and geno-typic tests were fed into the software program as conscriptedvariables in a binary coding system. These were thereafter used
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
to generate two-way tables with measures of association for allvariables considered. In each case, an enumeration of the with-in-row, within-column frequencies and Fisher’s exact analysiswere performed. In some instances, Spearman’s rank of correla-tion was used for hypothesis testing using unadjusted significantlevels. Using this test, all possible pair-wise correlation coeffi-cients were calculated and the results presented in a matrix tableoutput. Wilcoxon’s tests were conducted using GraphPad Prism.For all analyses, statistical significance was inferred when P val-ues were less than 0.05.
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028
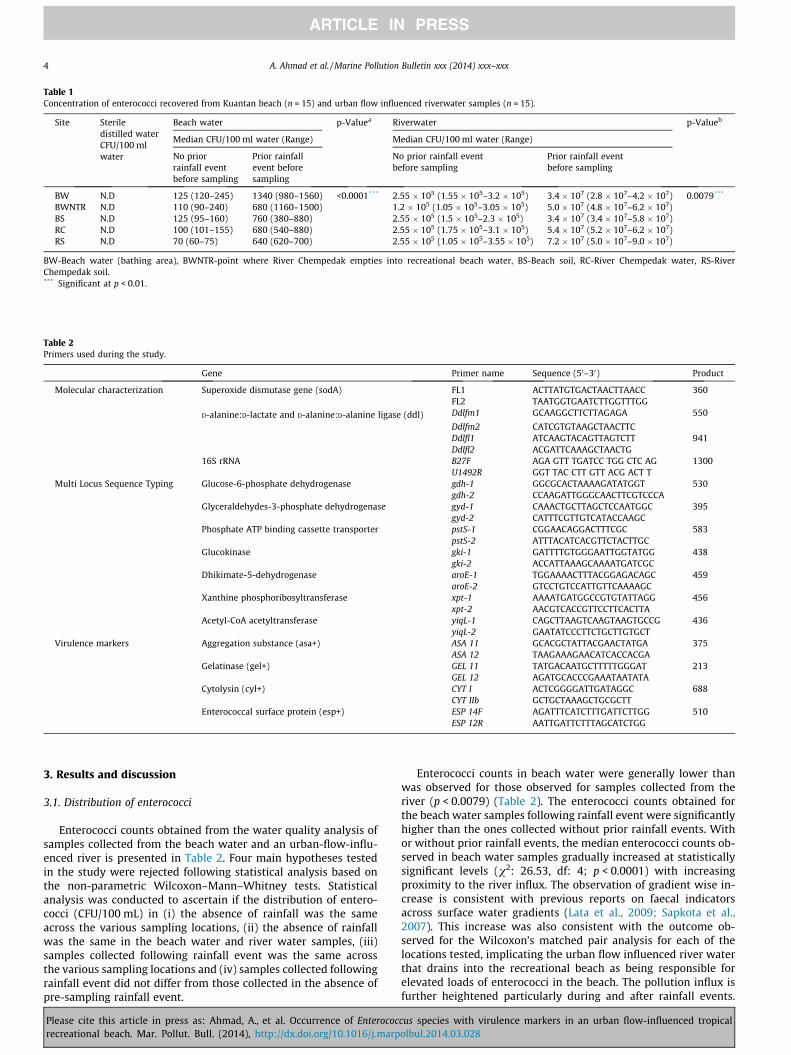
Table 1Concentration of enterococci recovered from Kuantan beach (n = 15) and urban flow influenced riverwater samples (n = 15).
Site Steriledistilled waterCFU/100 mlwater
Beach water p-Valuea Riverwater p-Valueb
Median CFU/100 ml water (Range) Median CFU/100 ml water (Range)
No priorrainfall eventbefore sampling
Prior rainfallevent beforesampling
No prior rainfall eventbefore sampling
Prior rainfall eventbefore sampling
BW N.D 125 (120–245) 1340 (980–1560) <0.0001*** 2.55 � 105 (1.55 � 105–3.2 � 105) 3.4 � 107 (2.8 � 107–4.2 � 107) 0.0079***
BWNTR N.D 110 (90–240) 680 (1160–1500) 1.2 � 105 (1.05 � 105–3.05 � 105) 5.0 � 107 (4.8 � 107–6.2 � 107)BS N.D 125 (95–160) 760 (380–880) 2.55 � 105 (1.5 � 105–2.3 � 105) 3.4 � 107 (3.4 � 107–5.8 � 107)RC N.D 100 (101–155) 680 (540–880) 2.55 � 105 (1.75 � 105–3.1 � 105) 5.4 � 107 (5.2 � 107–6.2 � 107)RS N.D 70 (60–75) 640 (620–700) 2.55 � 105 (1.05 � 105–3.55 � 105) 7.2 � 107 (5.0 � 107–9.0 � 107)
BW-Beach water (bathing area), BWNTR-point where River Chempedak empties into recreational beach water, BS-Beach soil, RC-River Chempedak water, RS-RiverChempedak soil.*** Significant at p < 0.01.
Table 2Primers used during the study.
Gene Primer name Sequence (50–30) Product
Molecular characterization Superoxide dismutase gene (sodA) FL1 ACTTATGTGACTAACTTAACC 360FL2 TAATGGTGAATCTTGGTTTGG
D-alanine:D-lactate and D-alanine:D-alanine ligase (ddl) Ddlfm1 GCAAGGCTTCTTAGAGA 550
Ddlfm2 CATCGTGTAAGCTAACTTCDdlfl1 ATCAAGTACAGTTAGTCTT 941Ddlfl2 ACGATTCAAAGCTAACTG
16S rRNA B27F AGA GTT TGATCC TGG CTC AG 1300U1492R GGT TAC CTT GTT ACG ACT T
Multi Locus Sequence Typing Glucose-6-phosphate dehydrogenase gdh-1 GGCGCACTAAAAGATATGGT 530gdh-2 CCAAGATTGGGCAACTTCGTCCCA
Glyceraldehydes-3-phosphate dehydrogenase gyd-1 CAAACTGCTTAGCTCCAATGGC 395gyd-2 CATTTCGTTGTCATACCAAGC
Phosphate ATP binding cassette transporter pstS-1 CGGAACAGGACTTTCGC 583pstS-2 ATTTACATCACGTTCTACTTGC
Glucokinase gki-1 GATTTTGTGGGAATTGGTATGG 438gki-2 ACCATTAAAGCAAAATGATCGC
Dhikimate-5-dehydrogenase aroE-1 TGGAAAACTTTACGGAGACAGC 459aroE-2 GTCCTGTCCATTGTTCAAAAGC
Xanthine phosphoribosyltransferase xpt-1 AAAATGATGGCCGTGTATTAGG 456xpt-2 AACGTCACCGTTCCTTCACTTA
Acetyl-CoA acetyltransferase yiqL-1 CAGCTTAAGTCAAGTAAGTGCCG 436yiqL-2 GAATATCCCTTCTGCTTGTGCT
Virulence markers Aggregation substance (asa+) ASA 11 GCACGCTATTACGAACTATGA 375ASA 12 TAAGAAAGAACATCACCACGA
Gelatinase (gel+) GEL 11 TATGACAATGCTTTTTGGGAT 213GEL 12 AGATGCACCCGAAATAATATA
Cytolysin (cyl+) CYT I ACTCGGGGATTGATAGGC 688CYT IIb GCTGCTAAAGCTGCGCTT
Enterococcal surface protein (esp+) ESP 14F AGATTTCATCTTTGATTCTTGG 510ESP 12R AATTGATTCTTTAGCATCTGG
4 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
3. Results and discussion
3.1. Distribution of enterococci
Enterococci counts obtained from the water quality analysis ofsamples collected from the beach water and an urban-flow-influ-enced river is presented in Table 2. Four main hypotheses testedin the study were rejected following statistical analysis based onthe non-parametric Wilcoxon–Mann–Whitney tests. Statisticalanalysis was conducted to ascertain if the distribution of entero-cocci (CFU/100 mL) in (i) the absence of rainfall was the sameacross the various sampling locations, (ii) the absence of rainfallwas the same in the beach water and river water samples, (iii)samples collected following rainfall event was the same acrossthe various sampling locations and (iv) samples collected followingrainfall event did not differ from those collected in the absence ofpre-sampling rainfall event.
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
Enterococci counts in beach water were generally lower thanwas observed for those observed for samples collected from theriver (p < 0.0079) (Table 2). The enterococci counts obtained forthe beach water samples following rainfall event were significantlyhigher than the ones collected without prior rainfall events. Withor without prior rainfall events, the median enterococci counts ob-served in beach water samples gradually increased at statisticallysignificant levels (v2: 26.53, df: 4; p < 0.0001) with increasingproximity to the river influx. The observation of gradient wise in-crease is consistent with previous reports on faecal indicatorsacross surface water gradients (Lata et al., 2009; Sapkota et al.,2007). This increase was also consistent with the outcome ob-served for the Wilcoxon’s matched pair analysis for each of thelocations tested, implicating the urban flow influenced river waterthat drains into the recreational beach as being responsible forelevated loads of enterococci in the beach. The pollution influx isfurther heightened particularly during and after rainfall events.
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 5
This possibly explains the 2log and 1log increase in enterococcidensities observed in river and beach water respectively whensamples were collected following a prior rainfall event. Given thehigher counts associated with the river water following dischargeof stormwater though the stormwater conveyance system(Fig. 1), there is considerable evidence that the stormwater convey-ance system in addition to the river flow may also be acting as areservoir, or breeding basin for indicator bacteria.
During rainfall events, high bacterial loads in urban influencedstorm water are thus flushed through the system. This resuspendsbacteria living in the sediments of the stormwater system furtherelevating the load discharged to the beach. Without the influenceof rainfall, enterococci counts of the considered bathing beachwater were still found to be above the decisive action level of104 enterococci per 100 ml in salt water as recommended byUSEPA (Bacterial Water Quality Standards for Recreational Waters(Freshwater and Waters), 2003). The values were also greater thanthe 640 per 100 mL water (95th percentile) benchmark stipulatedby the WHO (2003) for intestinal enterococci in category B water.The counts obtained for the river water samples too were abovethe international single-sample advisory limit of 61 CFU/100 mLfor enterococci in fresh water (Bacterial Water Quality Standardsfor Recreational Waters (Freshwater and Waters), 2003). There isno indicator bacteria standard for bathing water in Malaysia(Hamzah et al., 2011).
3.2. Species diversity of enterococci isolates
There was also a marked difference in the diversity of entero-cocci species recovered from beach sand (BS) and beach water(BW). Among the beach water isolates, Enterococcus casseflavusand E. faecalis were detected at the highest proportions (47.37%and 42.11% respectively) while the least occurring species wereEnterococcus hirae and E. faecium (both 5.26%). On the other hand,beach sand had a different species diversity profile, E. hirae beingthe most predominant (65.39%) followed by E. faecalis (26.92%).Notably, a high proportion of Enterococcus gallinarum was detectedin the river water. While it was the most predominant species inthe population tested (45.45%), E. gallinarum was not detectablein beach water and beach sand samples (Table 3). E. hirae and E.faecalis isolates were also present at varying proportions (27.27%and 22.72%, respectively) among the tested population of entero-cocci recovered from river chempedak water samples. Analysis of2 � 2 contingency tables of E. faecalis and E. faecium frequenciesin beach water and beach sand showed no association in the occur-rence of the two species (p = 0.212). Chi square analysis of thediversity data however revealed significant differences(X2 = 70.86, df = 16, p < 0.0001) in the frequencies of occurrenceof enterococci diversity from the various sites sampled duringthe study. The diversity of enterococci species recovered was thussignificantly associated with the sampling sites.
Table 3Frequency of the distribution of Enterococcus species diversity among the five sampled sit
Species Number of isolates (%) from each isolation site (n = 5
BW BWNTR BS
E. casseflavus 9 (9.38) 2 (2.08) 2 (2.08)E. hirae 1 (1.04) 1 (1.04) 18 (18.75)E. gallinarum 0 0 0E. faecium 1 (1.04) 0 0E. faecalis 8 (8.33) 7 (7.29) 6 (6.25)Total Enterococci per site 19 (19.79) 10 (10.42) 26 (27.08)
Pearson chi2 (v2) = 70.8603, df = 16, ���Statistically significant at alpha <0.05.BW-Beach water (bathing area), BWNTR- point where River Chempedak empties intChempedak soil.
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
In the current study, E. faecalis and E. casseliflavus dominatedbeach water isolates. In a similar study conducted at a tropicallocation, the dominant enterococci were identified as E. casselifla-vus in both river and beach samples regardless of the season orlocation suggesting their adaptation to this environment whencompared to the other enterococci. Also, among the non-pig-mented, E. faecalis was the dominant enterococci (Hernández,2011). This observation is in concert with our findings. Preliminaryinformation on the beach where samples were collected for entero-cocci isolation revealed the presence of a zoo near to the beach.Previous studies have highlighted E. hirae as commonly associatedwith the intestinal flora of several animals (Kolbjørnsen et al.,2011; Devriese et al., 1994). This probably explains the predomi-nance of E. hirae in beach sand isolates. A number of studies haveattempted tracing movements of sediments in the coastal zone.In a pioneering study by Trask (1952) on the investigation of themovement of beach sand along the southern California coast, itwas demonstrated that sand at the considered beach may havecome from a distance of more than 160 km up the coast. More re-cently, variations in the degree of grain rounding have been used totrace sand movements, or to obtain additional information con-cerning the history of the sediment particles (Allan and Hart,2007). The markedly differing characteristics observed amongbeach water and beach sand isolates in this study may be usefulfor application in microbial source tracking studies that aim todecipher if faecal enterococci loadings in adjacent waters areattributable to beach soil.
3.3. Antibiotic resistance among encountered enterococci
Two-tailed t-test analysis revealed differences in the levels ofantibiotic resistance demonstrated by isolates recovered from thebeach area and the river area. In this case, beach area was takenas the combination of isolates from beach water (BW), beach soil(BS) and the river influx region on the beach (BWNTR). On theother hand, river area was defined as the combination of riverwater (RC) and river soil (RS) isolates. The differences in proportionof antibiotic resistance were statistically significant (p < 0.05) formost of the antibiotics tested with the river area isolates generallypresenting higher resistance proportions (Table 4).
The level of antibiotic resistance was highest for kanamycin(90.2% and 85.45% for river area and beach areas, respectively).However, none of these isolates were found to be resistant to highlevels of aminoglycoside (500 mcg/ml and 2000 mcg/ml). Tetracy-cline and chloramphenicol resistance levels were at 44.79% and37.59% respectively. Fisher’s exact test revealed significant associ-ations at p-values less than 0.05 for ampicillin and vancomycinresistance. Nitrofurantoin resistance among the pool of testedenterococci (n = 96) was found to be associated significantly withresistance to chloramphenicol. Similar associations were also ob-servable for chloramphenicol and streptomycin, kanamycin and
es.
) Enterococci spp distribution p-Value
RC RS
1 (1.0 4) 0 14 (14.58) 0.0001���
6 (6.25) 5 (5.21) 31 (32.29)10 (10.42) 8(8.33) 18 (18.75)0 0 1 (1.04)5 (5.21) 6(6.25) 32 (33.33)22(22.92) 19 (19.79) 96 (100)
o recreational beach water, BS-Beach soil, RC-River Chempedak water, RS-River
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

Table 4Differences in antibiotic resistance and virulence factor carriage among beach water and river water emptying into the recreational beach water.
River area (n = 41) Beach area (n = 55) Two-tailed t-value p-Value
Resistance to antibiotics V 8 (19.51)a,b,c,d 15(27.27)a 0.8902 0.3757T 28 (68.29)a,e,f 15(27.27)b 4.3032 <0.0001 ***
C 27 (65.85) b,g,h 9(16.36)c 5.4797 <0.0001 ***
A 10 (24.39)e,g,I,j 14(25.45)d 0.1176 0.9067K 37 (90.24) c,I,k,l 47(85.45)a,b,c,d,e,f 0.7138 0.4771N 11 (26.83) f,h,k 4(7.27)e 2.4923 0.0155 *
S 21 (51.22) d,j,l 4(7.27)f 5.0758 <0.0001 ***
Possession of virulence factors cyl+ 3 (7.32) 0 (0.00) N.D N.Desp+ 9 (21.95) 11(20.00) 0.2291 0.8194gel+ 41 (100.0) 37(67.27) ND NDasa+ 26 (63.41) 23(41.82) 2.1266 0.0363 *
ND-not determined because one of the variables compared had standard deviations equals to zero. V, Vancomycin (glycopeptides), K, kanamycin, S-Streptomycin (amino-glycoside), A-ampicillin (B-lactam), T, tetracycline (tetracycline), C-chloramphenicol (chloramphenicol), N, nitrofurantoin (nitrofuran). Same letters within column arestatistically significant based on Turkey Crammers multiple mean comparison test.* Significant at p < 0.05*** Significant at p < 0.01
6 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
nitrofurantoin, and between kanamycin and streptomycin resis-tance levels. Nitrofurantoin is a common drug of choice for thetreatment of urinary tract infections. However, the rate at whichpathogens are developing resistance to the drug is exerting its tollon the reduction of the usefulness of this drug in the treatment ofuncomplicated UTIs. While resistance to nitrofurantoin was low forbeach isolates (7.27%), a significantly higher level of resistance(26.83%) to this drug was observed among river area isolates.Chloramphenicol resistance was not detected among beach waterisolates although a high proportion of RC isolates (72.73%) wereresistant to the antibiotics. Previous studies on the prevalence ofantibiotic resistance among environmental strains of enterococciin Malaysia are few. Notwithstanding, Toosa et al. (2001) docu-mented high chloramphenicol resistance levels (61%) among poul-try enterococci isolates in Malaysia.
In addition to testing for high level gentamicin resistance at500 lg/ml and 2000 lg/ml concentrations, streptomycin resis-tance was also evaluated since enterococcal resistance to gentami-cin and streptomycin occur by different mechanisms. Althoughgentamicin resistance may be a good predictor of resistance to anumber of aminoglycosides, this may not be the case with strepto-mycin (Cetinkaya et al., 2000). Streptomycin-resistant enterococci,particularly due to the production of streptomycin adenyltransfer-ase, are susceptible to gentamicin. Two types of streptomycinresistance are observable among enterococci. The first type is gen-erally limited to moderate-level resistance to 62–500 lg/ml con-centration of the drug simply because of low permeability anddrug uptake. On the other hand, the second type of resistance isusually either ribosomally mediated or as a result of the produc-tion of aminoglycoside-inactivating enzymes that allow suchstrains to demonstrate high-level resistance at concentrations ator higher than 2000 lg/ml (Cetinkaya et al., 2000). On the otherhand, gentamicin resistance is predominantly the result of thepresence of the inactivating enzyme 20 0-phosphotransferase-60-acetyltransferase. Although all strains in our study were found tobe susceptible to gentamicin, 7.27% and 51.22% of enterococcistrains recovered from beach area and river area respectively wereresistant to streptomycin.
Of all tested beach area isolates (n = 45), 27.27% were found tobe vancomycin resistant. This proportion was however not signif-icantly different (p = 0.3757) from the proportion of river area iso-lates (19.51%) that were resistant to the drug. Apart from a fewstudies that have reported on the occurrence of VRE in samplesfrom clinical patients, chickens, pigs and poultries in Malaysia(Son et al., 1999; Raja et al., 2005; Getachew et al., 2013), thereis a dearth of published information on the prevalence of VRE in
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
coastal beach water used for recreational purposes. The occurrenceof VRE in the environment presents threats to public health. Be-sides the commonly reported nosocomial acquisition and subse-quent colonization of vancomycin-resistant enterococci (VRE) inhospital settings, colonization among non-hospitalized personsand individuals outside the health-care setting have been docu-mented to occur frequently (McDonald et al., 1997). There is thusthe need for further studies on the prevalence and communitytransmission of VRE in Malaysia. If transmission with VRE fromhitherto unrecognized community sources could be identifiedand controlled, increased incidence of colonization and infectionboth in the community and among hospitalized patients may beprevented.
Notably, there was an extremely significant difference(p < 0.0001) in the observable levels of tetracycline resistance withthe river isolates presenting higher levels (68.29%) as compared tothe beach area isolates. Cheong et al. (1994), in a study on antimi-crobial resistance among some hospital enterococci isolates re-ported a prevalence level of 60.6% for tetracycline resistance.There is no published information on the prevalence of tetracyclineresistance among enterococci recovered from recreational beacheson Malaysia apart from a recent report (Dada et al., 2013) that re-ported a prevalence level of 3.3% at a separate beach. Consistentwith our study, the occurrence of tetracycline resistant beachenterococci was also reported by Rathnayake et al. (2012).
The levels of ampicillin resistance recorded in this study(24.39% and 25.45% respectively for the river and beach area) weremuch lower than was previously reported (Dada et al., 2013). Inanother study on public beaches (Turgeon et al., 2012), tetracyclineand ampicillin were among the three antimicrobials with the high-est frequency of resistance. Pronounced resistance to ampicillinhas also been reported by previous studies and they appear com-mon in samples from untreated sewage or in isolates from the hu-man and animal intestines (Silva et al., 2006; Adria et al., 2011).
Several enterococci isolates were observed with the ability togrow on oxacillin screen agar. However, based on PCR detection,all of these isolates were found to be devoid of mecA gene carriageassociated with MRSA that are resistant to oxacillin antibiotics inthe agar. Lata et al. (2009) also highlighted enterococci with abilityto grow on oxacillin agar recovered from a polluted river. mecAgene codes for a protein with a low affinity to penicillin (PBP2a)and confers methicillin resistance is located on a mobile genomicelement, the staphylococcal cassette chromosome (SCCmec).Although it was previously reported only among Staphylococ-cus genus from clinics, some studies (Turgeon et al., 2012;Tsubakishita et al., 2010) have also reported the carriage of mecA
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 7
gene in non-staphylococcal isolates recovered from surface watersand other environmental sources. Acquisition by transduction ofheterologous genes, particularly of antibiotic resistance genes,might represent an important mechanism of horizontal gene trans-fer in water bodies (de Vera and Simmons, 1996).
3.4. Occurrence of virulence determinants among tested bacteria
Biofilm formation was highest among beach soil isolates(61.54%) followed by beach water (31.58%) and river water(22.73%). Biofilm production was significantly associated withthe carriage of either asa or gelE genes. Conversely, fisher’s exacttest revealed significant associations between caseinase and car-riage of either esp or gelE gene. Virulence genes cyl, esp, gelE andasa were detected in 7.32%, 21.95%, 100% and 63.41% respectivelyamong river isolates but at lower proportions of 0%, 20%, 67.27%and 41.82% respectively among beach water isolates. Notably, cylpositive isolates were detected only among river area isolatesand not among beach water (BW) isolates thus implicating urban
Table 5Correlation observed between the enterococcus species isolation source and prevalence of
S/no
Combination of single/multiple antimicrobial resistance (groups/class)
1 Va Glycopeptide2 T Tetracycline3 C Chloramphenicol4 A B-lactam5 K Aminoglycoside6 N Nitrofuran7 S Aminoglycoside8 Va–K Glycopeptide–aminoglycoside9 K–S Aminoglycoside10 A–K B-lactam–aminoglycoside11 T–K Aetracycline–aminoglycoside12 C–K Chloramphenicol–aminoglycoside13 T–C Tetracycline–chloramphenicol14 Va–T Glycopeptide–tetracycline15 T–A Tetracycline–B-lactam16 C–N Chloramphenicol–nitrofuran17 C–K–S Chloramphenicol–aminoglycoside18 T–K–S Tetracycline–aminoglycoside19 Va–C–A–K Glycopeptide–chloramphenicol–B-lactam–aminoglycoside20 Va–C–A–K–S Glycopeptide–chloramphenicol–B-lactam–aminoglycoside21 T–K–N Tetracycline–aminoglycoside–nitrofuran22 T–A–K Tetracycline–B-lactam–aminoglycoside23 T–C–K Tetracycline–chloramphenicol–aminoglycoside24 Va–T–C–A–K Glycopeptide–tetracycline–chloramphenicol–B-lactam–amino25 T–C–A–K Tetracycline-chloramphenicol–B-lactam–aminoglycoside26 C–A–K–N Chloramphenicol–B-lactam–aminoglycoside–nitrofuran27 C–A–K–N–S Chloramphenicol–B-lactam–aminoglycoside–nitrofuran28 Va–T–A–N Glycopeptide–tetracycline–B-lactam–nitrofuran29 A–K–N B-lactam–aminoglycoside–nitrofuran30 Va–C–A Glycopeptide–chloramphenicol–B-lactam31 T–C–A Tetracycline–chloramphenicol–B-lactam32 T–A–K–N–S Tetracycline–B-lactam–aminoglycoside–nitrofuran33 T–C–A–K–S Tetracycline–chloramphenicol–B-lactam–aminoglycoside34 T–A–N–S–E Tetracycline–B-lactam–nitrofuran–aminoglycoside35 C–A–N Chloramphenicol–B-lactam–nitrofuran36 A–K–N–S B-lactam–aminoglycoside–nitrofuran–aminoglycoside37 Va–T–C–A–K–
SGlycopeptide–tetracycline–chloramphenicol–B-lactam–amino
38 Va–T–C–K–S Glycopeptide–tetracycline–chloramphenicol–aminoglycoside39 Va–T–C–A–K–
N–SGlycopeptide–tetracycline–chloramphenicol–B-lactam–aminoglycoside–nitrofuran
40 T–C–K–S Tetracycline–chloramphenicol–aminoglycoside41 Va–T–C–K–N–
SGlycopeptide–tetracycline–chloramphenicol–aminoglycoside–nitrofuran–aminoglycoside
42 T–C–K–N–S Tetracycline–chloramphenicol–aminoglycoside–nitrofuran–aminoglycoside
* Correlation significant at p � 0.05** Correlation significant at p � 0.01*** Correlation significant at p � 0.001**** Correlation significant at p � 0.0001
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
flow as potential source of cyl carrying enterococci influx into rec-reational bathing water. This is the first study that reports thisobservation. Some beach water isolates (57.89%) also possessedthe esp gene which was notably absent among all beach soil iso-lates. Unlike the beach water isolates, only the river isolates werefound to carry the esp gene (22.73%). This observation also presentsevidence to implicate the river emptying into the beach wateralong with other faecal sources as responsible for the preponder-ance of esp gene carrying enterococci in beach water. In developingnations, urban rivers often serve as sewers for communities whosimply discharge waste into the flowing water body. Some of thesemay be rich in faecal contamination and may explain for the espstrains that eventually reach the sea.
In terms of comparative levels of prevalence, asa gene wasdetectable in all faecal isolates, followed by the beach water(68.42%), river water (63.64%) and least among beach soil isolates(11.54%). GelE gene carriage was always present in all isolatesrecovered from BW (beach water), BWNTR (beach water at riverinflux region) and RC (river water). However, only 30.77% of beach
resistance to single/multiple classes of antibiotics.
No. ofenterococci
(%) Spearmancorrelation (rs)
p-Value
23 (23.95833) �0.08994 0.191843 (44.79167) 0.408 <0.0001 ****
36 (37.5) 0.5057 <0.0001 ****
24 (25) �0.01216 0.453284 (87.5) 0.07163 0.24415 (15.625) 0.2664 0.0043 **
25 (26.04167) 0.4953 <0.0001 ****
20 (20.83333) �0.02809 0.392924 (25) 0.4742 <0.0001 ****
18 (18.75) 0.01686 0.435238 (39.58333) 0.4207 <0.0001 ****
32 (33.33333) 0.4616 <0.0001 ****
23 (23.95833) 0.5021 <0.0001 ****
11 (11.45833) 0.2183 0.0163 *
13 (13.54167) 0.08911 0.1949 (9.375) 0.3003 0.0015 **
19 (19.79167) 0.4168 <0.0001 ****
17 (17.70833) 0.4821 <0.0001 ****
6 (6.25) 0.03806 0.35644 (4.166667) 0.1361 0.0938 (8.333333) 0.273 0.0036 **
10 (10.41667) 0.1192 0.123721 (21.875) 0.511 <0.0001 ****
glycoside 4 4.166667) 0.1361 0.0937 (7.291667) 0.1628 0.05655 (5.208333) 0.1767 0.0425 *
3 (3.125) 0.08699 0.19972 (2.083333) 0.0215 0.41767 (7.291667) 0.1628 0.05657 (7.291667) 0.0008437 0.49679 (9.375) 0.3003 0.0015 **
2 (2.083333) 0.1689 0.0499 *
4 (4.166667) 0.1361 0.0933 (3.125) 0.208 0.021 *
5 (5.208333) 0.1767 0.0425 *
4 (4.166667) 0.1361 0.093glycoside 3 (3.125) 0.208 0.021 *
8 (8.333333) 0.3492 0.0002 ***
1 (1.041667) 0.1188 0.1244
14 (14.58333) 0.4189 <0.0001 ****
2 (2.083333) 0.1689 0.0499 *
2 (2.083333) 0.1689 0.0499 *
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

8 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
sand isolates possessed the gene. Fisher’s exact test revealed signif-icant associations between carriage of esp and asa genes, esp andgelE genes and between asa and gelE genes. Examining species var-iation in the carriage of virulence genes, no notable differencehowever was observable between virulence factors detectableamong E. faecalis as compared with other enterococci species apartfrom the case of asa gene carriage. Also, Wilcoxon paired test didnot reveal any significant correlation of virulence marker preva-lence with species diversity (Table 5). However, statistically signif-icant correlations were observable between the carriage ofvirulence markers and the site of isolation. This was extremely sig-nificant for gelE gene as carriage of this factor was mostly associ-ated with strains from the river area.
A multivariate typing approach which involved clustering of 18different parameters covering resistance to antibiotics, demonstra-tion of virulence phenotypes and carriage of virulence genes wereanalysed using conglomerate hierarchical clustering (Fig. 2). Thedendogram obtained successfully clustered more than 70% of eachof the isolates recovered from various sources together on a sourcebasis. Interestingly when datasets of enterococci isolates recoveredfrom the portion of the beach proximate to the influx of riverchempedak were included in the analysis, they were found to clus-ter with the river chempedak isolates, predominantly; riverchempedak water isolates (Fig. 2). Using a 75% cut-off point, anal-ysis of the obtained dendogram revealed two main clusters ofbeach area and river area strains.
3.5. Evidence of co-selection of virulence markers and antibioticresistance
The highest levels of resistance to combination of antibioticswas observed for T–K (tetracycline–aminoglycoside) (39.58%) fol-lowed closely by C–K (chloramphenicol–aminoglycoside) whilethe least levels was observed for combined resistance to chloram-phenicol and nitrofurantoin (C–N) (9.38%) (Table 4). For resistanceto multiple antibiotic groups (P3), the highest proportion was ob-served for T–C–K (tetracycline–chloramphenicol–aminoglycoside)(21.88%). The lowest proportion of combined resistance observedwas that of V–T–C–A–K–N–S, i.e. resistance to a combination ofall 7 classes of antibiotics tested (1.04%). Considering co-selectionof two groups of antibiotics at the species level (Fig. 3a), the high-est level of combined resistance was observed for T–K–S (tetracy-cline–aminoglycoside) among 64.71% of E. gallinarum isolates. Onthe other hand, the least prevalence of combined resistance totwo groups of antibiotics was observed among E. casseflavus iso-lates (2.63%). E. faecalis isolates recovered in the study were foundto demonstrate resistance to a combination of two groups of anti-biotics at prevalence levels that ranged from 15% (as in V–K {glyco-peptide–aminoglycoside combination}) to a maximum of 53.85%(as in T–A {tetracycline–aminoglycoside}). For a combination ofthree groups of antibiotics (Fig. 3b), E. hirae strains presented thehighest prevalence levels as 75% of the strains were observed tobe resistant to a combination of A–K–N–S (B-lactam–aminoglyco-side–nitrofuran). For combinations of more than three groups ofantibiotics (Fig. 3c), E. gallinarum topped the list presenting a vari-ety of 15 combination possibilities. It is critical to affirm at thisjuncture that E. gallinarum strains dominated the species diversityof strains recovered from the river. Notably, combined resistanceto V–T–C–A–K–N–S, i.e. resistance to a combination of all 7 classesof antibiotics tested (1.04%) was only identified among the riverisolates. The current study provides empirical evidence that multi-ple-resistant strains are being disseminated along with river flowinto the considered recreational beach water. Evidently, an analy-sis of resistance to combination of groups of antibiotics vis-à-visthe considered isolation sites points also in this direction. For acombination of two groups (Fig. 3d), three groups (Fig. 3e) and four
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
or more groups of antibiotics (Fig. 3f), river water isolates werenotably the highest in terms of prevalence.
Considering the whole library of environmental enterococci(n = 96) tested in this study, Fishers exact analysis revealed evi-dence of co-selection of vancomycin resistance and a number ofresistance to combinations of groups of antibiotics. These includeresistance against V–K (P = 0.000), V–T (=0.000), V–C–A–K–S(P = 0.003), C–K–S (P = 0.011), V–T–C–A–K (P = 0.003), V–C–A(P = 0.000), T–C–A–K–S (P = 0.042), V–T–C–A–K–S (P = 0.012) andT–C–K–S (P = 0.007). Similarly evidence of co-selection of antibi-otic resistance and virulence was found between the carriage ofgelE and resistance to a number of combinations of antibioticgroups.
The correlation observed between prevalence of resistance tosingle/multiple classes of antibiotics was also explored using a bin-ary coding system that designated 0 and 1 for antimicrobial sus-ceptibility and resistance respectively. Answers to two mainquestions were sought in the analysis. Could it be inferred by cor-relation analysis that at every instance, co-selection of antibioticresistance (coder = 1) was always associated with E. faecalis or E.faecium species (coded = 1) and not the rest of the enterococci spe-cies (coded = 0)? At every instance resistance was observed, was italways associated with enterococci recovered from a particularsource (codes = 1) and not from the other sources (codes = 0)? Nostatistically significant correlation was observable for the firsthypotheses. However, for the second hypothesis that aimed to cor-relate resistance observed with source, several extremely signifi-cant correlations (p < 0.0001) were observed (Table 5). Resistanceto tetracycline, chloramphenicol, B-lactam, aminoglycoside werelinked with strains from the river draining into the recreationalbeach water (p-value < 0.0001). Furthermore, co-selection ofresistance to T–K (tetracycline–aminoglycoside), C–K (chloram-phenicol–aminoglycoside), T–C (tetracycline–chloramphenicol),C–K–S (chloramphenicol–aminoglycoside), T–K–S (tetracycline–aminoglycoside), T–C–K (tetracycline–chloramphenicol–aminogly-coside), T–C–K (tetracycline–chloramphenicol–aminoglycoside)and T–C–K–N–S (tetracycline–chloramphenicol–aminoglycoside–nitrofuran) were observed to be linked to strains from the riverarea at extremely significant probability levels (p < 0.0001). Theseobservations are in concert with Thong et al. (2011) that reportedchloramphenicol resistance as linked always with resistanceto streptomycin (aminoglycoside), tetracycline and ampicillin(B-lactam).
In attempting to provide similar answers for virulence, it wasobserved that the carriage of virulence markers was not statisti-cally correlated with species diversity. However, an extremely sig-nificant correlation (p < 0.0001) was observed for gelE carriage andmembership of the RA class (pooled population of river water andriver sand) (Table 6). Relatively lower but significant correlationswere also observed for cyl (p = 0.021) and asa (p = 0.018) gene car-riage. With carriage of three of four tested virulence markers in thisstudy found to correlate with membership of the RA class (pooledpopulation of river water and river sand), our study presents evi-dence again to implicate the river in the preponderance of entero-cocci with virulence traits in the tested recreational beach water.Added to our observations of potential recombination events fromsequence-based recombination detection analysis (data not in-cluded), suggestively these strains are able to exchange geneticmaterials with indigenous beach water strains.
3.6. Multi-Locus Sequence Typing
The aim of this part of the study was to determine if any of se-lected strains with multidrug resistance and multiple virulencegene carriage could be linked to a globally recognised clonalcomplex that has been previously implicated in enterococcal infec-
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

Fig. 2. Strain typing based on a combintion of phenotypic characteristics and the carriage of virulence genes.
A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 9
tion outbreaks. Based on a combination of their harbourage ofvirulence genes and antibiotic resistance profiles, four E. faecalisisolates were selected for multilocus sequence typing analysis(MLST) (Table 7). The first strain tested (RC19) was observed tobe ST181. ST181 shared six out of the seven tested loci with thepopular ST6, a member of the clonal complex 2 (CC2). CC2 corre-sponds to a clone of widely distributed multi-drug resistant strainthat has been previously associated with hospital outbreaks. It wasinteresting to note that the isolate was recovered from the low tideriver which drains into the seawater available for recreational pur-poses. Our observation of recovery of CC2 strains in non-hospitalsettings corroborate a recent report that also recovered sequencetype strains from this clonal complex from liquid manure and sew-age (Freitas et al., 2009). A single representative of CC2 was re-
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
ported also from a farm animal (pig) (Freitas et al., 2011).Enrichment of STs belonging to CC2 among hospital-associated iso-lates has been previously documented in a number of countries(Freitas et al., 2009; Ruiz-Garbajosa et al., 2006; Kawalec et al.,2007). Notably, the ST181 strain was observed to be resistant totetracycline, chloramphenicol, kanamycin and streptomycin. Theobservation of multi drug resistance is in concert with a previousstudy (Freitas et al., 2009). In addition to asa and gelE genes,ST181 possessed two other putative virulence genes, cyl and esp.These observations support previous findings from other authorsthat highlight the role of cyl and esp genes in the pathogenicityof enterococci (Freitas et al., 2009; Shankar et al., 2006; McBrideet al., 2007). Furthermore, full genome sequencing of a representativestrain of this CC (ST6) revealed the presence of several mobile ele-
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

V-K
A-K T-K C-K T-C V-T T-A C-NC-K
-ST-K
-S
V-K
A-K T-K
C-K T-C V-T T-A C-NC-K
-ST-K
-S
0
20
40
60
80
100
% R
esis
tanc
e
BW
BS
BWNTR
RC
RS
T-K-N
T-A-K
T-C-K
V-C-A
T-C-A
T-A-N
-SC-A
-N
A-K-N
-S
T-C-K
-S
T-K-N
T-A-K
T-C-K
V-C-A
T-C-A
T-A-N
-SC-A
-N
A-K-N
-S
T-C-K
-S
0
20
40
60
80
100
% R
esis
tanc
e
BS
BWNTR
RC
RS
BW
V-C-A
-K
V-C-A
-K-S
T-C-A
-K
C-A-K
-N
C-A-K
-N-S
V-T-A-N
A-K-N
-T
T-A-K
-N-S
T-C-A
-K-S
V-T-C-K
-N-S
T-C-K
-N-S
V-T-C-A
-K
V-T-C
-A-K
-S
V-T-C
-K-S
V-T-C
-A-K
-N-S
0
V-C-A
-K
V-C-A
-K-S
T-C-A
-K
C-A-K
-N
C-A-K
-N-S
V-T-A-N
A-K-N
-T
T-A-K
-N-S
T-C-A
-K-S
V-T-C-K
-N-S
T-C-K
-N-S
V-T-C-A
-K
V-T-C
-A-K
-S
V-T-C
-K-S
V-T-C
-A-K
-N-S
20
40
60
80
100
% R
esis
tanc
e
BW
BS
BWNTR
RC
RS
0
20
40
60
80
100
% R
esis
tanc
e
E._casseflavus
E.hiraeE.gallinarum
E.faecalisE.faecium
0
20
40
60
80
100%
Res
ista
nce
E._casseflavus
E.hirae
E.gallinarum
E.faecium
E.faecalis
0
20
40
60
80
100
Combination of four to six groups of
% R
esis
tanc
e
E._casseflavus
E.hirae
E.gallinarum
E.faecium
E.faecalis
Combination of two groups of antimicrobialsalong with isolation site Combination of three groups of antimicrobials
along with isolation site Combination of six groups of antimicrobialsalong with isolation site
antimicrobials along with diversity
Combination of three groups ofantimicrobials along with diversity
Combination of two groups ofantimicrobials along with diversity
(a) (b) (c)
(d) (e) (f)
Fig. 3. (a–c) Distribution of single or multiple antimicrobial resistance in different Enterococci species isolates based on isolation source and (d–f) species diversity. V,Vancomycin (glycopeptides), K, kanamycin, S-Streptomycin (aminoglycoside), A-ampicillin (B-lactam), T, tetracycline (tetracycline), C-chloramphenicol (chloramphenicol), N,nitrofuranoin (nitrofuran).
Table 6Correlation of the prevalence of single/multiple virulence markers with species diversity and isolation site.
Beach area (n = 55) proportions(%)
River area (n = 41) proportions (%) TotalEnterococci (%)
Correlation withspecies diversity
Correlation withisolation site
S/no Virulencemarkers
Ec Eh Eg Efm Efs Ec Eh Eg Efm Efs (rs) p-Value (rs) p-Value
1 cyl+ 0 0 – 0 0 0 9.09 11.11 – 0 3 (3.13) �0.01464 0.4437 0.208 0.021*
2 esp+ 46.15 5 – 0 19.04 0 27.27 27.78 – 9.09 20 (20.83) �0.1255 0.1116 0.02377 0.40913 asa+ 69.23 20 – 0 47.61 0 72.72 61.11 – 63.64 49 (51.04) 0.0149 0.4427 0.2137 0.0183*
4 gelE+ 92.31 40 – 100 76.19 100 100 100 – 100 78 (81.25) 0.1034 0.158 0.4148 <0.0001***
5 asa+gelE+ 69.23 20 – 0 47.62 0 72.73 61.11 – 63.64 49 (51.04) 0.0149 0.4427 0.2137 0.0183*
6 esp+gelE+ 46.15 5 – 0 19.05 0 27.27 27.78 – 9.09 20 (20.83) �0.1255 0.1116 0.02377 0.40917 esp+asa+gelE+ 46.15 5 – 0 19.05 0 27.27 27.78 – 9.09 20 (20.83) �0.1255 0.1116 0.02377 0.40918 cyl+esp+asa+gelE+ 0 0 – 0 0 0 9.09 11.11 – 0 3 (3.13) �0.01464 0.4437 0.208 0.021*
Notes: Ec-Enterococcus casseflavus, Eh-Enterococcus hirae, Eg- Enterococcus gallinarum, Efs-Enterococcus faecalis, Efm-Enterococcus faecium, cyl+: strains carrying gene coding forcytolysin; esp+: strains carrying gene coding for enterococcal surface protein; asa+: strains carrying gene coding for aggregation substance; gelE+: strains carrying gene codingfor gelatinase.
Table 7Characteristics of the four E. faecalis isolates selected for multilocus sequence typing analysis (MLST).
Strain gdh gyd pstS gki aroE xpt yqil Sequence Type (ST) Colonal complex (CC) Antibiotic resistance phenotype Virulence marker
E. faecalis RC9 14 2 18 10 16 2 12 59 N/A Tet, Chl, Kan, Nit asa, gelEE. faecalis BW15 14 1 18 57 16 35 12 474 N/A Kan asa, gelEE. faecalis BW17 1 1 9 6 1 1 1 117 CC21 Kan esp, asa, gelEE. faecalis RC19 12 5 3 7 6 1 5 181 CC2 Tet, Chl, Kan, Str esp, asa, cyl, gelE
10 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
ments, a pathogenicity island and an operon for capsular polysac-charide biosynthesis which have been demonstrated to aid resis-tance to phagocytic killing (Hancock and Gilmore, 2002).
Analysis of the MLST data revealed another isolate (RC59) asbeing a ST59 strain. As with RC19, the isolate was recovered fromthe low tide river which drains into the seawater available for rec-reational purposes. The occurrence of ST59 has been previously re-ported among chickens, faeces sample from a healthy pig and fromhospitalised patients (Ruiz-Garbajosa et al., 2006; Kawalec et al.,2007). The strain was found to harbour asa and gel genes. ST59
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
was found to be resistant to a number of antibiotics; tetracycline,chloramphenicol, kanamycin and nitrofurantoin. This is in agree-ment with a recent study that reported the isolation of ciprofloxa-cin resistant ST59 strains from Australian waterways (Rathnayakeet al., 2012). The past years of abuse of chloramphenicol and otherantibiotics in agricultural settings might have played a role in thespread of antibiotic resistance among these STs in Malaysia. ASouth East Asian study (Huys et al., 2007) reported that due tothe decrease in therapeutic efficiency of inexpensive antibioticssuch as oxytetracycline, many Asian aquaculture activities have
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 11
gradually switched during the past decade to the use of otherbroad-spectrum antibiotics such as chloramphenicol and the syn-thetically derived florfenicol. Although the lack of exact data onusage pattern of antimicrobials in human and veterinary medicinelimits the study in potentially exploring the quantitative nature ofcause-effect relationships, it remains generally accepted that theselection of resistant bacteria in agricultural settings might be con-tributing to the spread of resistance (Cabello, 2006).
One of the four isolates (BW17) recovered from recreationalbathing water in the current study was found to be of the sequencetype 117. A previous study (Freitas et al., 2009) isolated membersof this clonal complex from both hospital patients and from pig-gery. CC21 isolates have also been frequently observed in the glo-bal E. faecalis collection among isolates of diverse origin, includinghospitalized patients, non-hospitalized individuals, meat and farmanimals (Ruiz-Garbajosa et al., 2006). This seems to highlight thedispersion of this CC in different ecological settings. The ST117strain recovered from recreational bathing water in this studyexhibited resistance to low level kanamycin resistance. Quinoneset al. (2009) also observed the absence of multi drug resistanceamong ST117 strains tested. It was also reported that ST117 is adouble-locus variant of ST21 in CC21 (one of the biggest CCs de-tected in Spain), which include strains from animals, hospitalizedpatients, and apparently healthy volunteers in the community(Quinones et al., 2009). The ST117 strain in our study harbouredthe esp, asa and gel genes. Although previously characterizedCC21 strains usually harboured fewer antibiotic resistance and vir-ulence traits than isolates of other CCs (Ruiz-Garbajosa et al., 2006;Kawalec et al., 2007; McBride et al., 2007), the possibility of recom-bination in beach water settings could herald successful acquisi-tion of diverse genetic elements that might facilitate theirpersistence and spread in environments under selective antibioticpressure. Among STs reported in this study, ST117 and ST181 wereonly previously reported as novel STs in a study conducted in Cuba(Quinones et al., 2009) and another study on waterways in Austra-lia (Rathnayake et al., 2011). The regional geographical proximityof the latter to Malaysia may be a useful premise to present aST181 clonal regional spread hypothesis. Notwithstanding, the lo-cal availability of virulence genes is a crucial determinant on theextent to which these factors are acquired by epidemic clonesthrough horizontal gene transfer (Novais et al., 2004; Nilssonet al., 2009). It is difficult however to suggest a clonal regionalspread hypothesis for the occurrence of the novel ST117 in beachenvironment settings in Malaysia.
Only three studies have previously reported on the genetic var-iability of enterococci in Malaysia. All of these studies focused onclinical and animal strains. Weng et al. (2012) reported the occur-rence of ST18 and ST596 strains among isolates recovered fromhospital patients. Also, another study (Weng et al., 2013) reportedST17, ST78, ST203 and ST601 strains isolated from clinical speci-mens. In a most recent study, Getachew et al. (2013) reportedST4, ST6, ST87, ST108, ST274 and ST244 from a total of 11 isolatesrecovered from humans, chickens and pigs. To the best of ourknowledge, this is the first report on the use of MLST to delineatestrains recovered from beach water environment in Malaysia intoSTs and clonal complexes. Considering the growing evidences thatclinical resistance is intimately associated with environmental bac-teria (de Vera and Simmons, 1996; Abriouel et al., 2008), researchactivities need to be expended to include environmental microor-ganisms. In the current study, the isolation of a strain discoveredto be a member of the hospital adapted clonal complex (CC2) isan issue of concern. A recent study also reported the occurrenceof hospital adapted clonal complex 17 E. faecium from a beachenvironment. It could be that both the river from which the strainwas isolated along with the storm water drainage which emptiesinto it are acting as reservoirs for this ST and other potentially
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
virulent strains. Subsequent rainfall events dislodge them andultimately herald their dispersal into recreational beach water.Suggestively, the findings of our study emphasize the need formore studies on the characterization of enterococci strains frombeaches, particularly those recovered from beaches receiving sig-nificant influx from polluting rivers and storm drainage systems.Arguably, there is also the need for inclusion of these environmentsin the global discourse of epidemiological research.
Some limitations were observed during the study. First, there isa dearth of precise data on usage pattern of antimicrobials in hu-man and veterinary medicine in Malaysia. This arguably makes ita challenge to explore the quantitative nature of cause-effect rela-tionships. This observation however continues to remain a globalchallenge for environmental health researchers particularly in lessdeveloped nations where relevant regulatory institutions are frail(Dada et al., 2012). Another limitation in this study is the financialrestrictions that limited the number of isolates sequenced. In anideal situation, a very large library of sequences obtained from var-ious potential contributory sources of pollution ought to have beenanalysed in this study. This could have increased the confidencelevels of our analysis. Despite this restriction however, our studyprovides useful insight into the prevalence and dispersion of anti-biotic resistance and virulence makers in the studied beach. Finan-cial restrictions also limited the number of strains subjected toMLST, it is thus difficult to generalise on an overall representationof the E. faecalis population based on the prevalence of certain clo-nal complexes neither is it feasible to identify a possible clonalexpansion of a particular strain in the study location.
4. Conclusion
The presence of antibiotic resistant enterococci with virulencetraits in surface recreational water could be a public health risk.
Conflict of Interest
All authors declare that they have no competing interests.
Acknowledgements
The corresponding author (ACD) is in receipt of an EducationTax Fund PhD research grant. Thanks to Encik Alias for assistanceduring sample collection. Financial support for sampling and labo-ratory analysis from the Universiti Kebangsaan Malaysia MarinePathogen Program Grant (Science Fund 04-01-02-SF0754) underthe auspices of the School of Bioscience and Biotechnology, Facultyof Science and Technology is appreciated. Institute of Ecology andEnvironmental Studies, Obafemi Awolowo University, Ile-Ife, Nige-ria gratefully granted study leave for the research.
References
A.P.H.A., 1999. Standard Methods for the Examination of Water and Wastewater,20th ed. American Public Health Association (Apha), Washington, DC.
Abriouel, H., Omar, N.B., Molinos, A.C., López, R.L., Grande, M.J., Martínez-Viedma, P.,Ortega, E., Cañamero, M.M., Galvez, A., 2008. Comparative analysis of geneticdiversity and incidence of virulence factors and antibiotic resistance amongenterococcal populations from raw fruit and vegetable foods, water and soil,and clinical samples. Int. J. Food Microbiol. 123, 38–49.
Adria, R.I., Harry, H., Steve, N., 2011. Occurrence and antimicrobial drug resistanceof potential bacterial pathogens from shellfish, including Queen Conchs(Strombus Gigas) and Whelks (Cittarium pica) in Grenada. WebMedCentralArticle ID: WMC001943. <http://wwwwebmedcentralcom/wmcpdf/Article_WMC001943pdf>.
Allan, J.C., Hart, R., 2007. Profile dynamics and particle tracer mobility of a cobbleberm constructed on the oregon coast. In: Proceedings of the Sixth InternationalSymposium on Coastal Engineering and Science of Coastal Sediment Processes,New Orleans, USA, pp. 1–14.
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

12 A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx
USEPA: Bacterial Water Quality Standards for Recreational Waters (Freshwater andMarine Waters), 2003. <http://www.epa.gov/waterscience/beaches/local/statrept.pdf>. webcite EPA-823-R-03-008. U.S. Environmental ProtectionAgency, Washington, DC.
Barrell, R., Hunter, P., Nichols, G., 2000. Microbiological standards for water andtheir relationship to health risk. Commun. Dis. Public Health 3, 8–13.
Bhaduri, B., Minner, M., Tatalovich, S., Harbor, J., 2001. Long-term hydrologic impactof urbanization: a tale of two models. J. Water Resour. Plan. Manage. 127, 13–19.
Brownell, M., Harwood, V., Kurz, R., McQuaig, S., Lukasik, J., Scott, T., 2007.Confirmation of putative stormwater impact on water quality at a Florida beachby microbial source tracking methods and structure of indicator organismpopulations. Water Res. 41, 3747–3757.
Cabello, F.C., 2006. Heavy use of prophylactic antibiotics in aquaculture: a growingproblem for human and animal health and for the environment. Environ.Microbiol. 8, 1137–1144.
Cariolato, D., Andrighetto, C., Lombardi, A., 2008. Occurrence of virulence factorsand antibiotic resistances in Enterococcus faecalis and Enterococcus faeciumcollected from dairy and human samples in North Italy. Food Control 19, 886–892.
Cetinkaya, Y., Falk, P., Mayhall, C.G., 2000. Vancomycin-resistant enterococci. Clin.Microbiol. Rev. 13, 686–707.
Cheong, Y., Lim, V., Jegathesan, M., Suleiman, A., 1994. Antimicrobial resistance in 6Malaysian general hospitals. Med. J. Malaysia 49 (4), 317.
Creti, R., Imperi, M., Bertuccini, L., Fabretti, F., Orefici, G., Di Rosa, R., et al., 2004.Survey for virulence determinants among Enterococcus faecalis isolated fromdifferent sources. J. Med. Microbiol. 53, 13–20.
Dada, A.C., Asmat, A., Gires, U., Heng, L.Y., Deborah, B.O., 2012. Bacteriologicalmonitoring and sustainable management of beach water quality in malaysia:problems and prospects. Global J. Health Sci. 4, 126–138.
Dada, A.C., Ahmad, A., Usup, G., Heng, L.Y., 2013. Speciation and antimicrobialresistance of Enterococci isolated from recreational beaches in Malaysia.Environ. Monit. Assess. 185, 1583–1599.
DailyExpress: Rubbish ending up in the sea, 2013. News published on Thursday,August 15. <http://www.dailyexpress.com.my/news.cfm?NewsID=86412>.(accessed 02.09.2013).
de Vera, M.E., Simmons, R.L., 1996. Antibiotic-resistant enterococci and thechanging face of surgical infections. Arch. Surg. 131, 338.
Devriese, L., Hommez, J., Pot, B., Haesebrouck, F., 1994. Identification andcomposition of the streptococcal and enterococcal flora of tonsils, intestinesand faeces of pigs. J. Appl. Microbiol. 77, 31–36.
Dutka-Malen, S., Evers, S., Courvalin, P., 1995. Detection of glycopeptide resistancegenotypes and identification to the species level of clinically relevantenterococci by PCR. J. Clin. Microbiol. 33, 24–27.
Fisher, K., Phillips, C., 2009. The ecology, epidemiology and virulence ofEnterococcus. Microbiology 155, 1749–1757.
Freitas, A.R., Novais, C., Ruiz-Garbajosa, P., Coque, T.M., Peixe, L., 2009. Clonalexpansion within clonal complex 2 and spread of vancomycin-resistantplasmids among different genetic lineages of Enterococcus faecalis fromPortugal. J. Antimicrob. Chemother. 63, 1104–1111.
Freitas, A.R., Coque, T.M., Novais, C., Hammerum, A.M., Lester, C.H., Zervos, M.J.,Donabedian, S., Jensen, L.B., Francia, M.V., Baquero, F., 2011. Human and swinehosts share vancomycin-resistant Enterococcus faecium CC17 and CC5 andEnterococcus faecalis CC2 clonal clusters harboring Tn1546 on indistinguishableplasmids. J. Clin. Microbiol. 49, 925–931.
Getachew, Y., Hassan, L., Zakaria, Z., Aziz, S.A., 2013. Genetic variabilityof vancomycin-resistant Enterococcus faecium and Enterococcus faecalisisolated from humans, chickens and pigs in Malaysia. Appl. Environ.Microbiology.
Hamilton, M.J., Hadi, A.Z., Griffith, J.F., Ishii, S., Sadowsky, M.J., 2010. Large scaleanalysis of virulence genes in Escherichia coli strains isolated from Avalon Bay,CA. Water Res. 44, 5463–5473.
Hamzah, A., Kipli, S.H., Rahil Ismail, S.I.I., 2011. Microbiological study in coastalwater of Port Dickson, Malaysia. Sains Malaysiana 40, 93–99.
Hancock, L.E., Gilmore, M.S., 2002. The capsular polysaccharide of Enterococcusfaecalis and its relationship to other polysaccharides in the cell wall. Proc. Nat.Acad. Sci. 99, 1574–1579.
Ríos Hernández LA, 2011. Report as of FY2011 for 2011PR129B. The populationdynamics of the dominant Enterococci in the water systems of Puerto Rico.<http://water.usgs.gov/wrri/2011grants/progress/2011PR129B.pdf.
Huys, G., Bartie, K., Cnockaert, M., Hoang Oanh, D.T., Phuong, N.T., Somsiri, T.,Chinabut, S., Yusoff, F.M., Shariff, M., Giacomini, M., 2007. Biodiversity ofchloramphenicol-resistant mesophilic heterotrophs from Southeast Asianaquaculture environments. Res. Microbiol. 158, 228–235.
Jackson, C.R., Fedorka-Cray, P.J., Barrett, J.B., 2004. Use of a genus-and species-specific multiplex PCR for identification of enterococci. J. Clin. Microbiol. 42,3558–3565.
Jett, B.D., Huycke, M.M., Gilmore, M.S., 1994. Virulence of enterococci. Clin.Microbiol. Rev. 7, 462–478.
Kawalec, M., Kedzierska, J., Gajda, A., Sadowy, E., Wegrzyn, J., Naser, S., Skotnicki, A.,Gniadkowski, M., Hryniewicz, W., 2007. Hospital outbreak of vancomycin-resistant enterococci caused by a single clone of Enterococcus raffinosus andseveral clones of Enterococcus faecium. Clin. Microbiol. Infect 13, 893–901.
Kolbjørnsen, Ø., David, B., Gilhuus, M., 2011. Bacterial osteomyelitis in a 3-week-oldbroiler chicken associated with Enterococcus hirae. Veter. Pathol. Online 48,1134–1137.
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
Lata, P., Ram, S., Agrawal, M., Shanker, R., 2009. Enterococci in river Ganga surfacewaters: propensity of species distribution, dissemination of antimicrobial-resistance and virulence-markers among species along landscape. BMCMicrobiol. 9, 140.
Layton, B., Walters, S., Boehm, A., 2009. Distribution and diversity of theenterococcal surface protein (esp) gene in animal hosts and the Pacific coastenvironment. J. Appl. Microbiol. 106, 1521–1531.
Mallin, M.A., Williams, K.E., Esham, E.C., Lowe, R.P., 2000. Effect of humandevelopment on bacteriological water quality in coastal watersheds. Ecol.Appl. 10, 1047–1056.
McBride, S.M., Fischetti, V.A., LeBlanc, D.J., Moellering Jr, R.C., Gilmore, M.S., 2007.Genetic diversity among Enterococcus faecalis. PLoS ONE 2, e582.
McDonald, L.C., Kuehnert, M.J., Tenover, F.C., Jarvis, W.R., 1997. Vancomycin-resistant enterococci outside the health-care setting: prevalence, sources, andpublic health implications. Emerg. Infect. Dis. 3, 311.
Mohamed, J.A., Huang, W., Nallapareddy, S.R., Teng, F., Murray, B.E., 2004.Influence of origin of isolates, especially endocarditis isolates, and variousgenes on biofilm formation by Enterococcus faecalis. Infect. Immun. 72, 3658–3663.
NCCLS: Performance Standards for antimicrobial disk susceptibility testing. NCCLSdocument M2–A8. Wayne, PA: National Committee for Clinical LaboratoryStandards.
Nilsson, O., Greko, C., Top, J., Franklin, A., Bengtsson, B., 2009. Spread without knownselective pressure of a vancomycin-resistant clone of Enterococcus faeciumamong broilers. J. Antimicrob. Chemother. 63, 868–872.
Novais, C., Coque, T.M., Sousa, J.C., Baquero, F., Peixe, L., 2004. Local genetic patternswithin a vancomycin-resistant Enterococcus faecalis clone isolated in threehospitals in Portugal. Antimicrobial Agents Chemother. 48, 3613–3617.
Padilla, C., Lobos, O., 2013. Virulence, bacterocin genes and antibacterialsusceptibility in Enterococcus faecalis strains isolated from water wells forhuman consumption. SpringerPlus 2, 1–6.
Peng, X., Zhang, G., Mai, B., Hu, J., Li, K., Wang, Z., 2005. Tracing anthropogeniccontamination in the Pearl River estuarine and marine environment of SouthChina Sea using sterols and other organic molecular markers. Mar. Pollut. Bull.50, 856–865.
Quinones, D., Kobayashi, N., Nagashima, S., 2009. Molecular epidemiologic analysisof Enterococcus faecalis isolates in Cuba by multilocus sequence typing.Microbiol. Drug Resist. 15, 287–293.
Raja, N., Karunakaran, R., Ngeow, Y., Awang, R., 2005. Community-acquiredvancomycin-resistant Enterococcus faecium: a case report from Malaysia. J.Med. Microbiol. 54, 901–903.
Rathnayake, I., Hargreaves, M., Huygens, F., 2011. SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterway,Australia, and associated antibiotic resistance gene profiles. BMC Microbiol.11, 201.
Rathnayake, I., Hargreaves, M., Huygens, F., 2012. Antibiotic resistance andvirulence traits in clinical and environmental Enterococcus faecalis andEnterococcus faecium isolates. Syst. Appl. Microbiol. 35, 326–333.
Ruiz-Garbajosa, P., Bonten, M.J., Robinson, D.A., Top, J., Nallapareddy, S.R., Torres, C.,Coque, T.M., Cantón, R., Baquero, F., Murray, B.E., 2006. Multilocus sequencetyping scheme for Enterococcus faecalis reveals hospital-adapted geneticcomplexes in a background of high rates of recombination. J. Clin. Microbiol.44, 2220–2228.
Santiago-Rodriguez, T.M.R.J., Coradin, M., Toranzos, A., 2013. Antibiotic-resistanceand virulence genes in Enterococcus isolated from tropical recreational waters. J.Water Health 11, 387–396.
Sapkota, A.R., Curriero, F.C., Gibson, K.E., Schwab, K.J., 2007. Antibiotic-resistantenterococci and fecal indicators in surface water and groundwater impacted bya concentrated swine feeding operation. Environ. Health Perspect. 115, 1040.
Schippmann, B., Schernewski, G., Gräwe, U., 2013. Escherichia coli pollution in aBaltic Sea lagoon: A model-based source and spatial risk assessment. Int. J. Hyg.Environ. Health.
Semedo, T., Santos, M.A., Lopes, M.F., Marques, J.J.F., Crespo, M.T., Tenreiro, R., 2003.Virulence factors in food, clinical and reference enterococci: a common trait inthe genus? Syst. Appl. Microbiol. 26, 13–22.
Shankar, N., Baghdayan, A.S., Willems, R., Hammerum, A.M., Jensen, L.B., 2006.Presence of pathogenicity island genes in Enterococcus faecalis isolates frompigs in Denmark. J. Clin. Microbiol. 44, 4200–4203.
Silva, J., Castillo, G., Callejas, L., López, H., Olmos, J., 2006. Frequency of transferablemultiple antibiotic resistance amongst coliform bacteria isolated from a treatedsewage effluent in Antofagasta, Chile. Electron. J. Biotechnol. 9.
Son, R., Nimita, F., Rusul, G., Nasreldin, E., Samuel, L., Nishibuchi, M., 1999. Isolationand molecular characterization of vancomycin-resistant Enterococcus faeciumin Malaysia. Lett. Appl. Microbiol. 29, 118–122.
Thong, K.-L., Hoe, C.H., Koh, Y.T., Yasin, R.M., 2011. Prevalence of multidrug-resistant Shigella isolated in Malaysia. J. Health Popul. Nutr. (JHPN) 20, 356–358.
Tiong, S.T., 2001. Integrated shoreline management plan of North Pahang. Coast.Geomorphol. 1, 1–112, <http://Geogallers.Com/Geo/Download.Php?Path=Ismps&Download_File=North%20pahang%20ismp.Pdf>.
Toosa, H., Radu, S., Rusul, G., Latif, A.R.A., Rahim, R.A., Ahmad, N., Ling, O.W., 2001.Detection of Vancomycin-Resistant Enterococcus Spp.(VRE) from Poultry.Malaysian J. Med. Sci. MJMS 8, 53.
Trask, P.D., 1952: Source of beach sand at Santa Barbara, California, as indicated bymineral grain studies. In: Book Source of Beach Sand at Santa Barbara,California, City: Dtic Document.
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028

A. Ahmad et al. / Marine Pollution Bulletin xxx (2014) xxx–xxx 13
Tsubakishita, S., Kuwahara-Arai, K., Sasaki, T., Hiramatsu, K., 2010. Origin andmolecular evolution of the determinant of methicillin resistance instaphylococci. Antimicrob. Agents Chemother. 54, 4352–4359.
Turgeon, P., Michel, P., Levallois, P., Chevalier, P., Daignault, D., Crago, B., Irwin, R.,McEwen, S.A., Neumann, N.F., Louie, M., 2012. Antimicrobial-resistantEscherichia coli in public beach waters in Quebec. Can. J. Infect. Dis. Med.Microbiol. 23, e20.
Vankerckhoven, V., Van Autgaerden, T., Vael, C., Lammens, C., Chapelle, S., Rossi, R.,Jabes, D., Goossens, H., 2004. Development of a multiplex PCR for the detectionof asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulencedeterminants among European hospital isolates of Enterococcus faecium. J. Clin.Microbiol. 42, 4473–4479.
Vergis, E.N., Shankar, N., Chow, J.W., Hayden, M.K., Snydman, D.R., Zervos, M.J.,Linden, P.K., Wagener, M.M., Muder, R.R., 2002. Association between the
Please cite this article in press as: Ahmad, A., et al. Occurrence of Enterococrecreational beach. Mar. Pollut. Bull. (2014), http://dx.doi.org/10.1016/j.marp
presence of enterococcal virulence factors gelatinase, hemolysin, andenterococcal surface protein and mortality among patients with bacteremiadue to Enterococcus faecalis. Clin. Infect. Dis. 35, 570–575.
Weng, P.L., Hamat, R.A., MPath, Y.K.C., Zainol, N., Aziz, M.N., Shamsudin, M.N., 2012.Vancomycin-resistant Enterococcus faecium of multi locus sequence type 18 inMalaysia. Med. J. Malaysia 67, 639.
Weng, P.L., Ramli, R., Shamsudin, M.N., Cheah, Y.-K., Hamat, R.A., 2013. High geneticdiversity of enterococcus faecium and Enterococcus faecalis clinical isolates bypulsed-field gel electrophoresis and multilocus sequence typing from a hospitalin Malaysia. BioMed Res. Ints.
Zhang, Q., Li, Z., Zeng, G., Li, J., Fang, Y., Yuan, Q., Wang, Y., Ye, F., 2009. Assessment ofsurface water quality using multivariate statistical techniques in red soil hillyregion: a case study of Xiangjiang watershed, China. Environ. Monit. Assess. 152,123–131.
cus species with virulence markers in an urban flow-influenced tropicalolbul.2014.03.028




















