
Nuclear Factor of Activated T Cells Regulates Transcription of the Surfactant Protein D Gene (Sftpd)...
11
Nuclear Factor of Activated T Cells Regulates Transcription of the Surfactant Protein D Gene (Sftpd) via Direct Interaction with Thyroid Transcription Factor-1 in Lung Epithelial Cells* Received for publication, April 19, 2004, and in revised form, May 21, 2004 Published, JBC Papers in Press, June 1, 2004, DOI 10.1074/jbc.M404296200 Vrushank Dave ´ ‡§, Tawanna Childs‡, and Jeffrey A. Whitsett‡ From the ‡Division of Pulmonary Biology, Cincinnati Children’s Hospital Medical Center, Cincinnati, Ohio 45229-3039 Surfactant protein D (SP-D) plays critical roles in host defense, surfactant homeostasis, and pulmonary immu- nomodulation. Here, we identify a role of nuclear factor of activated T cells (NFATs) in regulation of murine SP-D gene (Sftpd) transcription. An NFAT-dependent enhancer modulated by NFATs or calcineurin and sen- sitive to cyclosporin was identified in the Sftpd pro- moter. Ionomycin and phorbol 12-myristate 13-acetate further increased the activity of this enhancer, whereas VIVIT, a potent NFAT inhibitor peptide, selectively in- terfered with the calcineurin-NFAT interaction and abolished enhancer function. Gel supershift and DNase I protection assays identified DNA elements that bind NFAT in the Sftpd promoter. Calcineurin and NFATc3 proteins were detected in the embryonic and adult mouse lung epithelium, and the mRNA expression pro- files of the NFATs were similar in immortalized mouse lung epithelial cells and alveolar epithelial type II cells. NFATc3 and TTF-1 activated the Sftpd promoter, syner- gized transcription, co-immunoprecipitated from mouse lung epithelial cells, and physically interacted in vitro. Components of the calcineurin/NFAT pathway were identified in respiratory epithelial cells of the lung that potentially augment rapid assembly of a multiprotein transcription complex on Sftpd promoter inducing SP-D expression. Surfactant protein D (SP-D), 1 is a 43-kDa member of the calcium-dependent host defense “collectin” (collagen-lectin) family of proteins that includes surfactant protein A, serum mannose-binding protein, conglutinin, and bovine serum lec- tins CL-34 (1). In the lung, SP-D is synthesized and secreted primarily by type II and other nonciliated bronchiolar respira- tory epithelial cells (2– 4). As a component of the innate im- mune system, SP-D inhibits the proliferation of Gram-negative bacteria, binds and aggregates a variety of microorganisms including respiratory viruses and fungi, and enhances their presentation to resident phagocytic cells for opsonization and intracellular killing (1, 2, 5–7, 9 –12). In addition, SP-D influ- ences surfactant homeostasis, reduces alveolar macrophage apo- ptosis, stimulates a variety of immune cells, and enhances oxygen radical production (13–16). Recent findings, however, implicate SP-D in immunomodu- lation during inflammation and allergic response. SP-D binds alveolar macrophages and circulating leukocytes (17) and stim- ulates the migration of human neutrophils, monocytes, and macrophages (18, 19), influencing recruitment following lung injury and inflammation. SP-D stimulates immune cell activity following exposure to pathogen-associated molecular pattern recognition. Increased expression in the lungs of transgenic mice enhanced host defense function and decreased inflamma- tory responses following exposure to pulmonary pathogens (7, 9, 20, 21). Conversely, Sftpd / mice develop emphysema as- sociated with chronic inflammation (22) and exhibit enhanced inflammatory responses to a variety of stimuli (23). Thus, SP-D plays a role as a surveillance molecule that facilitates killing and clearance of pathogens, serving to control inflammatory responses. Despite the important protective roles of SP-D in pulmonary homeostasis, transcriptional regulation of SP-D is poorly un- derstood. Recently, a role for SP-1-like transcription factors has been implicated in regulation of the human SP-D gene (SFTPD) promoter, but the identity of these factors is unknown (24). AP-1 family members regulate SFTPD promoter activity. JunB and JunD enhanced, whereas c-Jun and c-Fos inhibited, transcription (24). Mutation of DNA binding sites for forkhead transcription factors FoxA1 and FoxA2 decreased transcription of SFTPD. In contrast, TTF-1 did not activate its expression in H441 cells in vitro (24). SP-D expression is developmentally regulated and increases dramatically prior to birth. In mouse and rat lung, SP-D mRNA is first detected at midgestation and increases abruptly before birth and during the immediate postnatal period (25–27). Glu- cocorticoid treatment in utero precociously increased SP-D mRNA and accelerated lung maturation. Whereas the mecha- nism of SP-D induction is not known, it has been suggested that dexamethasone treatment increases promoter occupancy of CCAAT enhancer-binding protein- (C/EBP-) (24, 28). Ret- inoblastoma protein stimulated SFTPD activation by directly forming a complex with C/EBPs bound to the NF-IL-6 (C/ EBP-) consensus site in the SFTPD promoter. Cotransfection of C/EBP-,-, and - cDNAs increased transcription of the human SFTPD gene in H441 cells (29). Following a variety of lung infection and injury, an acute phase response similar to that of liver is observed in the lung * This work was supported by American Lung Association Grant RG-155-N (to V. D.) and NHLBI, National Institutes of Health, Grant HL 63329 (to J. A. W.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. § To whom correspondence should be addressed: Division of Pulmo- nary Biology, 4403, Cincinnati Children’s Hospital Research Founda- tion, 3333 Burnet Ave., Cincinnati, OH 45229-3039. Tel.: 513-636-8410 (office) and 513-636-3323 (laboratory); Fax: 513-636-7868; E-mail: [email protected]. 1 The abbreviations used are: SP-D, surfactant protein D; C/EBP-, CCAAT enhancer-binding protein-; IL, interleukin; ORF, open reading frame; CA, constitutively active; CMV, cytomegalovirus; RT, reverse transcription; CnB1, calcineurin B1; EMSA, electrophoretic mobility shift assay; NFAT, nuclear factor of activated T cells; Tricine, N-[2- hydroxy-1,1-bis(hydroxymethyl)ethyl]glycine; PMA, phorbol 12-myris- tate 13-acetate; HD, homeodomain; GST, glutathione S-transferase; CREB, cAMP-response element-binding protein. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 33, Issue of August 13, pp. 34578 –34588, 2004 © 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 34578
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Nuclear Factor of Activated T Cells Regulates Transcription of the Surfactant Protein D Gene (Sftpd)...

Nuclear Factor of Activated T Cells Regulates Transcription of theSurfactant Protein D Gene (Sftpd) via Direct Interaction withThyroid Transcription Factor-1 in Lung Epithelial Cells*
Received for publication, April 19, 2004, and in revised form, May 21, 2004Published, JBC Papers in Press, June 1, 2004, DOI 10.1074/jbc.M404296200
Vrushank Dave‡§, Tawanna Childs‡, and Jeffrey A. Whitsett‡
From the ‡Division of Pulmonary Biology, Cincinnati Children’s Hospital Medical Center, Cincinnati, Ohio 45229-3039
Surfactant protein D (SP-D) plays critical roles in hostdefense, surfactant homeostasis, and pulmonary immu-nomodulation. Here, we identify a role of nuclear factorof activated T cells (NFATs) in regulation of murineSP-D gene (Sftpd) transcription. An NFAT-dependentenhancer modulated by NFATs or calcineurin and sen-sitive to cyclosporin was identified in the Sftpd pro-moter. Ionomycin and phorbol 12-myristate 13-acetatefurther increased the activity of this enhancer, whereasVIVIT, a potent NFAT inhibitor peptide, selectively in-terfered with the calcineurin-NFAT interaction andabolished enhancer function. Gel supershift and DNaseI protection assays identified DNA elements that bindNFAT in the Sftpd promoter. Calcineurin and NFATc3proteins were detected in the embryonic and adultmouse lung epithelium, and the mRNA expression pro-files of the NFATs were similar in immortalized mouselung epithelial cells and alveolar epithelial type II cells.NFATc3 and TTF-1 activated the Sftpd promoter, syner-gized transcription, co-immunoprecipitated from mouselung epithelial cells, and physically interacted in vitro.Components of the calcineurin/NFAT pathway wereidentified in respiratory epithelial cells of the lung thatpotentially augment rapid assembly of a multiproteintranscription complex on Sftpd promoter inducing SP-Dexpression.
Surfactant protein D (SP-D),1 is a 43-kDa member of thecalcium-dependent host defense “collectin” (collagen-lectin)family of proteins that includes surfactant protein A, serummannose-binding protein, conglutinin, and bovine serum lec-tins CL-34 (1). In the lung, SP-D is synthesized and secretedprimarily by type II and other nonciliated bronchiolar respira-tory epithelial cells (2–4). As a component of the innate im-
mune system, SP-D inhibits the proliferation of Gram-negativebacteria, binds and aggregates a variety of microorganismsincluding respiratory viruses and fungi, and enhances theirpresentation to resident phagocytic cells for opsonization andintracellular killing (1, 2, 5–7, 9–12). In addition, SP-D influ-ences surfactant homeostasis, reduces alveolar macrophage apo-ptosis, stimulates a variety of immune cells, and enhancesoxygen radical production (13–16).
Recent findings, however, implicate SP-D in immunomodu-lation during inflammation and allergic response. SP-D bindsalveolar macrophages and circulating leukocytes (17) and stim-ulates the migration of human neutrophils, monocytes, andmacrophages (18, 19), influencing recruitment following lunginjury and inflammation. SP-D stimulates immune cell activityfollowing exposure to pathogen-associated molecular patternrecognition. Increased expression in the lungs of transgenicmice enhanced host defense function and decreased inflamma-tory responses following exposure to pulmonary pathogens (7,9, 20, 21). Conversely, Sftpd�/� mice develop emphysema as-sociated with chronic inflammation (22) and exhibit enhancedinflammatory responses to a variety of stimuli (23). Thus, SP-Dplays a role as a surveillance molecule that facilitates killingand clearance of pathogens, serving to control inflammatoryresponses.
Despite the important protective roles of SP-D in pulmonaryhomeostasis, transcriptional regulation of SP-D is poorly un-derstood. Recently, a role for SP-1-like transcription factors hasbeen implicated in regulation of the human SP-D gene(SFTPD) promoter, but the identity of these factors is unknown(24). AP-1 family members regulate SFTPD promoter activity.JunB and JunD enhanced, whereas c-Jun and c-Fos inhibited,transcription (24). Mutation of DNA binding sites for forkheadtranscription factors FoxA1 and FoxA2 decreased transcriptionof SFTPD. In contrast, TTF-1 did not activate its expression inH441 cells in vitro (24).
SP-D expression is developmentally regulated and increasesdramatically prior to birth. In mouse and rat lung, SP-D mRNAis first detected at midgestation and increases abruptly beforebirth and during the immediate postnatal period (25–27). Glu-cocorticoid treatment in utero precociously increased SP-DmRNA and accelerated lung maturation. Whereas the mecha-nism of SP-D induction is not known, it has been suggestedthat dexamethasone treatment increases promoter occupancyof CCAAT enhancer-binding protein-� (C/EBP-�) (24, 28). Ret-inoblastoma protein stimulated SFTPD activation by directlyforming a complex with C/EBPs bound to the NF-IL-6 (C/EBP-�) consensus site in the SFTPD promoter. Cotransfectionof C/EBP-�, -�, and -� cDNAs increased transcription of thehuman SFTPD gene in H441 cells (29).
Following a variety of lung infection and injury, an acutephase response similar to that of liver is observed in the lung
* This work was supported by American Lung Association GrantRG-155-N (to V. D.) and NHLBI, National Institutes of Health, GrantHL 63329 (to J. A. W.). The costs of publication of this article weredefrayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18U.S.C. Section 1734 solely to indicate this fact.
§ To whom correspondence should be addressed: Division of Pulmo-nary Biology, 4403, Cincinnati Children’s Hospital Research Founda-tion, 3333 Burnet Ave., Cincinnati, OH 45229-3039. Tel.: 513-636-8410(office) and 513-636-3323 (laboratory); Fax: 513-636-7868; E-mail:[email protected].
1 The abbreviations used are: SP-D, surfactant protein D; C/EBP-�,CCAAT enhancer-binding protein-�; IL, interleukin; ORF, open readingframe; CA, constitutively active; CMV, cytomegalovirus; RT, reversetranscription; CnB1, calcineurin B1; EMSA, electrophoretic mobilityshift assay; NFAT, nuclear factor of activated T cells; Tricine, N-[2-hydroxy-1,1-bis(hydroxymethyl)ethyl]glycine; PMA, phorbol 12-myris-tate 13-acetate; HD, homeodomain; GST, glutathione S-transferase;CREB, cAMP-response element-binding protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 33, Issue of August 13, pp. 34578–34588, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org34578

(4). For example, haptoglobin, C-reactive protein, serum lipopo-lysaccharide-binding protein, and respiratory epithelial de-rived fibrinogen and �1-acid glycoprotein increased followinglung injury (29–33). SP-D mRNA and protein in lung lavageincreased within 6 h after endotracheal endotoxin treatment inrats (32), following Pseudomonas infection in mice (34), a briefO2 exposure to rats (35), and treatment of type II cells withFGF-7 (36). SP-D mRNA was markedly increased followingexposure to IL-4 and IL-13 in vivo (34, 37, 38). These observa-tions and other data support the concept that SP-D is rapidlyinduced following lung injury, perhaps serving a protective roleby modulating inflammation and enhancing host defense.Strikingly, Sftpd proximal promoter revealed cis-elements forC/EBPs, nuclear factor of activated T cells (NFAT), AP-1, andthe signal transducers and activators of transcription family oftranscription factors that are known to rapidly activate tran-scription of a large number of genes during an effective im-mune response in several organs (39–41). In addition, severalTTF-1 sites were identified that may play critical roles in SP-Dtranscription following lung injury (42).
Since, SP-D expression is induced following lung injury andexposure to IL-4 and IL-13, we speculated that the calcineurinpathway, known to regulate T cell activation (43) mediated bydephosphorylation of NFATs, may be operative in the regula-tion of SP-D expression. Calcineurin is a heterodimeric Ca2�/calmodulin-dependent protein phosphatase activated by cal-cium ionophores such as ionomycin and inhibited bycyclosporin and tacrolimus (FK-506). It consists of one catalytic(CnA) and one regulatory (CnB) subunit (44). Changes in ex-pression of cytokines and other immunomodulatory moleculesactivate T-cell receptors, increasing intracellular calcium con-centrations, activating calcineurin. Once activated, calcineurindephosphorylates NFAT and translocates them into the nu-cleus. NFATs bind cooperatively with other nucleoproteins topromoters of target genes to induce transcription (41, 45).
In the present work, we identified an endogenous, cal-cineurin/NFAT pathway in respiratory epithelial cells of thelung that assembles NFATs, AP-1, and TTF-1 on a proximalenhancer located in the 5�-region of the Sftpd gene, forming amultiprotein transcription complex inducing SP-D expression.
EXPERIMENTAL PROCEDURES
Plasmid Construction and Mutagenesis—Sftpd promoter-reporterconstructs were made in pGL3-basic vector, a luciferase reporter plas-mid (Promega). Using TRANSFEC (Biobase; Biologische DatenbankenGmbH, Wolfenbuttel, Germany), a promoter sequence analysis anddata base program, the Sftpd promoter was identified from a 5.7-kbSftpd genomic clone (a kind gift from Dr. T. Korfhagen, CincinnatiChildren’s Hospital, Cincinnati, OH) that contained a 3-kb sequenceupstream of the first exon that included a consensus TATA box. A679-bp BtgI fragment (�79 to �600 bp) was Klenow end-filled andcloned into the SmaI site of pGL3-basic, the correct orientation wasdetermined, and it was used to generate a series of promoter deletionmutants by PCR cloning bp �79 to �557, �357, �246, �167, and �82into NheI and XhoI sites of pGL3-basic (see Table I for primers used).Rat TTF-1 expression plasmid pRC-CMV-TTF-1 was provided by Dr. R.DiLauro (Naples, Italy). Full-length human NFATc3 (the original clonefrom Nakoi Aroi), NFATc4 with a 3.1-kb cDNA containing the ORFcloned in the KpnI and NotI site in expression vector pREp4, constitu-tively active (CA) CA-NFATc3 and CA-NFATc4 in pAC-CMV pLpA-5�expression vector, and constitutively active calcineurin (pECE-FLAG-�CnA) were provided by Dr. J. Molkentin (Cincinnati Children’s Hos-pital, Cincinnati, OH). Green fluorescent protein (GFP)-VIVIT expres-sion plasmid containing an in-frame ORF for peptide sequenceMAGPHPVIVITGPHEE fused to enhanced green fluorescent protein inpEGFP.N1 (Clontech), was a kind gift from Dr. A. Rao (Harvard Uni-versity, Boston, MA). pcDNA3.1 with CMV promoter (Invitrogen) andpCMV �-galactosidase (Clontech) vectors were used to normalize DNAand transfection efficiency, respectively. All subcloned DNAs were con-firmed by sequencing.
Cell Culture, Transfection, and Reporter Gene Assays—HeLa cells
were cultured in Dulbecco’s modified Eagle’s medium (Invitrogen) with10% fetal bovine serum. Mouse lung epithelial cells, MLE-15, an im-mortalized mouse lung epithelial cell line that maintains some morpho-logical and functional characteristics of type II epithelial cells, werecultured in HITES medium (46) for functional characterization of Sftpdpromoter. A series of Sftpd promoter-luciferase constructs were used intransient transfection assays using Fugene 6 at a DNA/Fugene ratio of1:2 according to the manufacturer’s instructions (Roche Applied Sci-ence). Briefly, 6-well plates at 30–50% confluence were transfected witha fixed amount of Sftpd promoter-luciferase plasmid and variousamounts of CMV-based cDNA expressing transactivator plasmids. Forco-transfection experiments, the promoter-reporter plasmid was kept at15 ng/well unless stated otherwise. Total DNA was normalized withcorresponding CMV-empty vectors, and transfection efficiency was nor-malized to �-galactosidase activity using 100 ng/well of pCMV �-galac-tosidase. Two days after transfection, luciferase and �-galactosidaseassays were performed using 20 �l of the supernatant according to aprevious protocol (47). The light units were assayed by luminometry(Monolight 2010, Analytical Luminescence Laboratory, San Diego, CA).Data obtained represent the average of three transfection experiments,each carried out in duplicate (n � 6) and depicted as means � S.D.unless stated otherwise.
Isolation of RNA and Reverse Transcription (RT)-PCR—MLE-15cells grown in a T75 flask to 80% confluence were washed and sus-pended in 10 ml of phosphate-buffered saline. 2.5 ml of cells werepelleted at 3,000 rpm and lysed in 0.5 ml of 1� RNA lysis buffer (50 mM
Tris�Cl, pH 7.5, 50 mM NaCl, 5 mM EDTA, 0.5% SDS) by 2 min ofvigorous vortexing. Total RNA was extracted in 100 �l of 5 M ammo-nium acetate (Ambion), 2 �l of glycogen and 600 �l of acid phenol/chloroform (5:1) solution, pH 4.5 (Ambion), precipitated in 2 volumes ofethanol, washed in 70% ethanol, dried, and suspended in 60 �l ofdiethyl pyrocarbonate-treated H2O. 10 �l of RNA suspension was re-verse transcribed for first strand cDNA synthesis using SuperScriptII-RT, with procedures provided by the manufacturer (Invitrogen).RNA from purified mouse alveolar epithelial type II cells (AE II) waskindly provided by Dr. John Shannon (Cincinnati Children’s Hospital,Cincinnati, OH). AE II cells were prepared from 6-week-old femaleC57B/6 mice as described previously (48). RT products were producedby PCR using primers described in Table I. Primers and reactionconditions were optimized using cDNA made from reverse transcribedtotal RNA of adult mouse lung. In studies of mouse alveolar type II cells(AE II cells), PCR was performed on RT product using Amplitaq GoldDNA polymerase (Roche Applied Science) under the following condi-tions: 95 °C for 10 min for the denaturation cycle and 35 cycles at 95 °C(30 s), 58 °C (30 s) annealing, and 72 °C for extension, with a finalextension of 10 min at 72 °C. �-Actin was used as a positive control.Reactions containing RNA that was not reverse transcribed were usedas a control to detect genomic DNA contamination.
Immunohistochemistry of Embryonic and Adult Lungs—Embryoswere obtained at different gestational stages, processed according toprevious methods (49), and embedded in paraffin. For adult lung tis-sues, lungs were inflation fixed with 4% paraformaldehyde at 25-cmwater pressure and processed for paraffin embedding. Serial 5-�msections were prepared for adult lung. For fetal lung, whole embryoswere fixed with 4% paraformaldehyde and sectioned at 5-�m intervals.To detect the regulatory subunit calcineurin B1 (CnB1), anti-mouseCnB1 rabbit polyclonal antitibody (PA3-025; Affinity Bioreagents,Goldon, CO) at a 1:500 dilution as primary antibody and anti-rabbit IgGmade in goat as secondary antibody (Vector Laboratories, Inc.) wereused, respectively, for immunohistochemistry as described previously(50). Biotinylated secondary antibodies and a streptavidin-biotin-per-oxidase detection system (Vector Laboratories) were used to localize theantibody-antigen complexes in the tissues, as previously described (51).A mouse-on-mouse blocking kit (Vector Laboratories) was used withprimary mouse monoclonal antibodies for NFATc3, (F-1, SC-8405X) at1:100 dilution. Antigen detection was enhanced with nickel-diamino-benzidine and Tris-cobalt, followed by counterstaining with NuclearFast Red.
Electrophoretic Mobility Shift Assays (EMSAs)—Nuclear extractfrom MLE-15 cells were prepared as previously described (52) exceptthat nuclear extracts were stored before dialyses. 32P-Labeled double-stranded DNA probes were made by a Klenow end-filling reaction using[�-32P]dCTP. EMSAs were performed by incubating 10 nM double-stranded DNA oligonucleotide probes corresponding to the �130(NFAT-I) and �170 (NFAT-II) sites (see Table I for oligonucleotidesequences) with 3.5 �l (�15 �g) of nuclear extract for 40 min at 4 °C in30 �l of binding buffer (15 mM HEPES, 0.1 mM EDTA, 0.5 mM dithio-threitol, 40 mM KCl, 5% glycerol, pH 7.9) with 1 �g of poly(dI)�poly(dC)
NFAT, TTF-1, and SP-D Gene Transcription 34579

(Amersham Biosciences), and reactions were electrophoresed on a 4%,0.5� TBE native polyacrylamide gel at room temperature for 2–3 h at150 V. Unlabeled competitor double-stranded DNA oligonucleotides,NFAT-I and NFAT-II from the Sftpd promoter, TTF-1 (Oligo-C), with astrong TTF-1 binding site from the rat thyroglobulin gene (rTg) pro-moter spanning �82 to �56 bp (53), and a strong NFAT site-containingoligonucleotide from the IL-2 promoter (54) were added at a 100-foldmolar excess. Antibody supershift assays were carried out in the samebuffer by the further addition of 2 and 4 �l of either anti-NFATc3antibodies (SC-1152X; Santa Cruz Biotechnology, Inc., Santa Cruz, CA)or albumin as mock, and the reaction continued at 4 °C for 1 h. Gelswere autoradiographed to detect specific DNA-protein complexformation.
DNase I Footprinting—DNA fragment spanning the �246 to �79 bpregion of the Sftpd promoter was subjected to DNase I reaction (see Fig.4B) using MLE-15 nuclear extracts. The (TG)n region in the �246/�79Sftpd promoter had four nucleotides (ACAC) less than the originalpromoter (GenBankTM accession number AF047742) as confirmed bysequencing. The Sftpd promoter (�246/�79) cloned in pGL3 was di-gested with NheI and labeled at the 3�-end using [�-32P]dCTP (6,000Ci/mmol) in a Klenow reaction, DNA-purified, and released with XhoI.The DNase I footprinting assay contained 10 fmol (�5,000 cpm) of theend-labeled fragment in 200 �l of 1� binding buffer (52) containing 150-and 300-�g nuclear extracts, and a protein binding reaction was carriedout for 30 min at 4 °C. DNase I reaction conditions and purification ofdigested DNA were as described previously (52). Samples were heat-denatured and loaded on to an 8% sequencing gel. DNA ladders for Gand A � G were made by Maxam-Gilbert chemical sequencing.
Glutathione S-Transferase GST Pull-down Assays—Full-lengthGST-TTF-1 fusion protein was made in two steps. TTF-1 cDNA wasfirst PCR-amplified with a stop codon from pRC-CMV-TTF-1, digested,and subcloned into EcoRI and HindIII sites (see Table I for primerdetails) within the multiple cloning sites of pCMV-Tag 2A vector (Clon-tech, Palo Alto, CA). TTF-1 cDNA was then excised from pCMV-Tag 2Awith EcoRI and SalI and ligated in frame between the same enzymesites in pGEX-4T-3, a bacterial expression vector (Amersham Bio-sciences), and sequenced. The GST-TTF-1 deletion constructs weremade from a series of pVP16-TTF-1 plasmids (47). DNA fragmentscontaining TTF-1 deletion derivatives were digested with XbaI andKlenow end-filled before cutting with BamHI. The DNA fragments werethen subcloned into the BamHI and SmaI sites in pGEX-5X-2, a bacte-rial expression vector (Amersham Biosciences). The plasmids weretransformed into BL21-DE3 bacterial strains for protein expression.GST fusion protein expression, cell lysis, and immobilization of proteinson glutathione-Sepharose beads were performed as described previ-ously (55). Immobilized proteins were detected on a Tris-glycine 10–20% SDS-polyacrylamide gel by Coomassie staining and total protein-normalized for protein-protein interaction using empty glutathione-Sepharose-beads. Beads (20 �l) with immobilized proteins were washedseveral times with 1� phosphate-buffered saline, pH 7.4, incubated for2 h at 4 °C with 15 �l of in vitro transcribed and translated radiolabeled[35S]Met-CA-NFATc3 in 1 volume binding buffer (10 mM Tris, pH 7.5, 1mM EDTA, 100 mM NaCl and 0.1% Nonidet P-40) on a rotator. Beadswere washed three times with 1.5 ml of wash buffer (same as bindingbuffer) and boiled in 1 volume of SDS Laemmli buffer containing 10%�-mercaptoethanol. Proteins released were separated by electrophore-sis under reducing conditions on SDS 10–20% Tris-glycine polyacryl-amide gradient gel (Novex, San Diego, CA). Gels were dried, and radio-labeled [35S]Met-CA-NFATc3 was detected on a PhosphorImager(Storm 860; Amersham Biosciences).
Co-Immunoprecipitation Assays and Western Blotting—The 3�FLAG-tagged CA-NFATc3 expression vector used in the co-immunopre-cipitation experiment was constructed as follows. CA-NFATc3 in pAC-CMV pLpA-5� expression vector was digested with HindIII and Kle-now end-filled and then excised with NotI containing the SV40 3�-untranslated region and inserted into EcoRV and NotI sites of 3�FLAG-CMV expression vector. The 3� FLAG-tagged CA-NFATc3 vec-tor was transfected (50 �g) into 40% confluent MLE-15 cells in T75flask, and nuclear extracts were prepared after 48 h as described (56),except that nuclear proteins were extracted by suspending nuclei for 1 hin 1 volume of high salt extraction buffer (20 mM HEPES, pH 7.9, 550mM NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM dithiothreitol, and 10mM phenylmethylsulfonyl fluoride) with one protease inhibitor mini-tablet (Roche Applied Science). Nuclear extracts from MLE-15 cells(�15 �g/�l) were diluted in 10 volumes of 1� phosphate-buffered salineand incubated for 2 h on a rotator at 4 °C with 100 �l of mousemonoclonal anti-FLAG M2-IgG-tagged Sepharose beads (Sigma). Thebeads were washed three times with 1.5 ml of wash buffer (10 mM Tris,
pH 7.5, 1 mM EDTA, 100 mM NaCl, and 0.1% Nonidet P-40) and boiledin 1 volume of SDS buffer (Laemmli sample buffer) containing 10%�-mercaptoethanol. The co-immunoprecipitated proteins released wereseparated by electrophoresis under reducing conditions onSDS-10�20% Tricine polyacrylamide gradient gels (Novex) and trans-ferred to nylon membranes (Bio-Rad). Protein blots were blocked with5% nonfat dry milk in TBST (10 mM Tris, pH 8, 150 mM NaCl, 0.1%Tween 20) and incubated with anti-TTF-1 monoclonal antibody pro-duced in our laboratory, followed by horseradish peroxidase-conjugatedrabbit anti-mouse IgG (Calbiochem). The presence of FLAG-taggedCA-NFATc3 on beads was assessed using primary anti-FLAG mousemonoclonal antibodies directly conjugated to horseradish peroxidase(Sigma). Western blots were developed with the enhanced chemilumi-nescence system (Amersham Biosciences) and exposed to autoradio-graphic film X-Omat (Eastman Kodak Co.).
RESULTS
TTF-1 and NFATs Activate Transcription from the SftpdPromoter—A promoter-luciferase reporter plasmid (Fig. 1A)comprising �600 to �79 bp (D-600-Luc) of the mouse surfac-tant protein D gene (Sftpd), was transiently transfected intoHeLa and MLE-15 cells. The �600/�79 bp Sftpd promoter washighly active in MLE-15 but not in HeLa cells (Fig. 1B). HeLacells co-transfected with D-600-Luc and increasing amounts ofan expression plasmid encoding TTF-1 activated the Sftpdpromoter by �15-fold (Fig. 1C). The proximal region of theSftpd promoter contains consensus elements for both TTF-1and NFATs. We therefore hypothesized that TTF-1- and NFAT-dependent pathways may directly influence Sftpd promoteractivity in respiratory epithelial cells. Full-length NFATc3 (Fu-NFATc3), full-length NFATc4 (Fu-NFATc4), constitutively ac-tive NFATc3 (CA-NFATc3), or constitutively active NFATc4(CA-NFATc4) expression plasmids (Fig. 1D) activated Sftpdpromoter activity in MLE-15 cells. CA-NFATc3 was more ac-tive than CA-NFATc4 (Fig. 1D, lanes 4 and 5). Full-lengthNFATs Fu-NFATc3 and Fu-NFATc4 also activated the �600/�79 Sftpd promoter, albeit to a much lesser extent (Fig. 1D,lanes 2 and 3), suggesting endogenous but weak calcineurinactivity in respiratory epithelial cells. Interestingly, this NFATactivation was not observed in HeLa cells (data not shown),suggesting the requirement of cofactors present in MLE-15cells but not in HeLa cells.
NFATs Are Present in the Respiratory Epithelial Cells—Since NFATs activated the �600/�79 Sftpd promoter inMLE-15 cells, we tested whether NFATs were expressed inMLE-15 and alveolar epithelial type II cells (AE-II) that areknown to express SP-D (27, 57). RT-PCR was carried out usingtotal RNA from MLE-15 and purified mouse AE-II cells.Unique sets of primers designed for murine NFATc1, NFATc2,NFATc3, NFATc4, and NFATc5 cDNA (Table I) gave PCRproducts of expected sizes (Fig. 2A). The expression profiles ofNFATs were similar in MLE-15 and AE-II cells. NFATc1 andNFATc3 were the predominant forms (Fig. 2, lanes 2, 4, 8, and10), NFATc4 was weakly expressed in AE-II cells (Fig. 2, lanes5 and 11), whereas NFATc2 was not expressed in either celltype (Fig. 2, lanes 3 and 9). NFATc5, a ubiquitously expressedgene not part of the classical NFAT family (58), was detected atlow levels in MLE-15 and AE-II cells (Fig. 2, lanes 6 and 12).
NFAT Binds Sftpd Promoter Elements—Since NFATc3 wasmost active on �600/�79 Sftpd promoter and was expressed athigh levels in MLE-15 and AE-II cells, we tested whetherNFATc3 directly bound to the �600/�79 Sftpd promoter. Twoperfect NFAT consensus sites (NFAT-I and NFAT-II) (Fig. 3B)present in the Sftpd promoter region at �170 to �164 and�130 to �124 bp from the transcription start site with a coresimilarity of 1.0 and a matrix similarity of 0.99 and 0.97,respectively, were identified using MatInspector 7.0 (Geno-matix Software GmbH, Munchen, Germany). Two DNA probescomprising NFAT-I or NFAT-II (see Table I) were used in
NFAT, TTF-1, and SP-D Gene Transcription34580
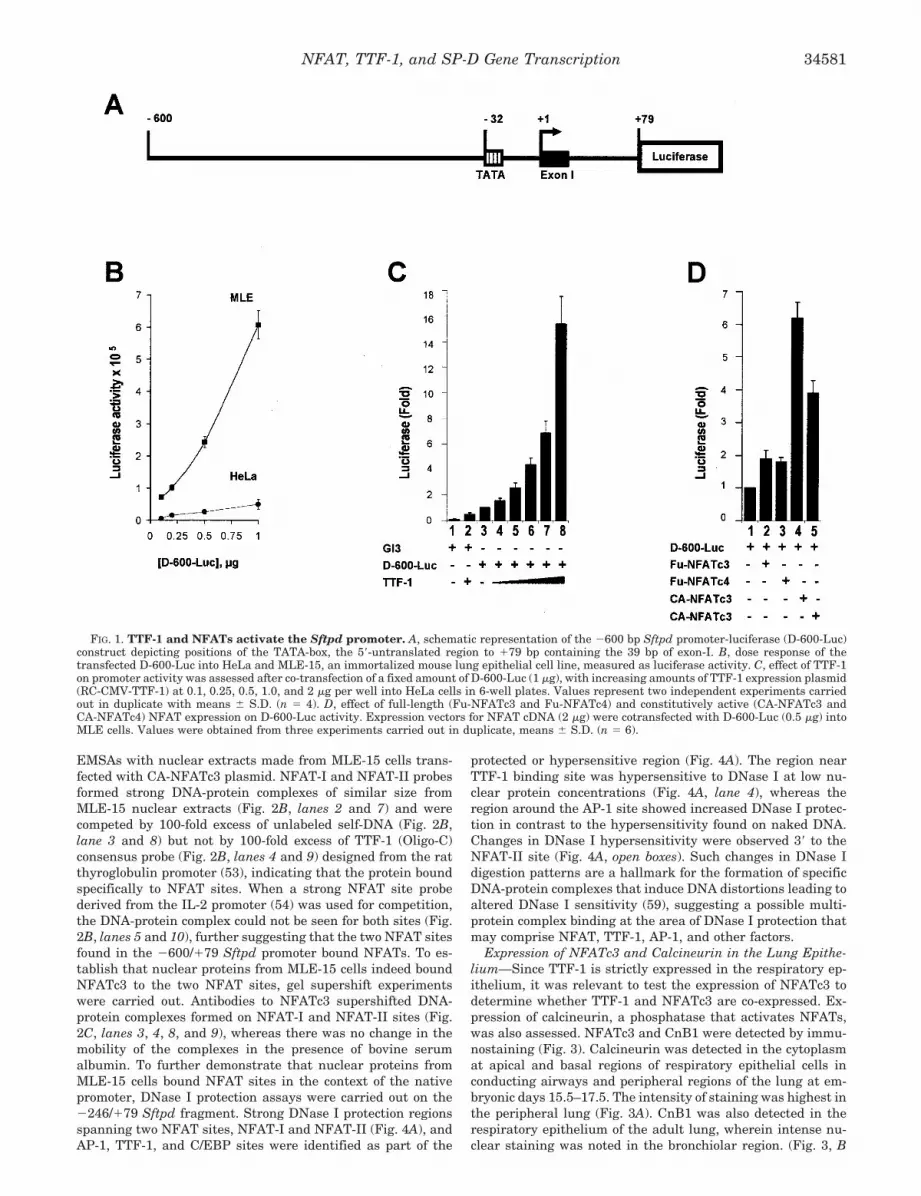
EMSAs with nuclear extracts made from MLE-15 cells trans-fected with CA-NFATc3 plasmid. NFAT-I and NFAT-II probesformed strong DNA-protein complexes of similar size fromMLE-15 nuclear extracts (Fig. 2B, lanes 2 and 7) and werecompeted by 100-fold excess of unlabeled self-DNA (Fig. 2B,lane 3 and 8) but not by 100-fold excess of TTF-1 (Oligo-C)consensus probe (Fig. 2B, lanes 4 and 9) designed from the ratthyroglobulin promoter (53), indicating that the protein boundspecifically to NFAT sites. When a strong NFAT site probederived from the IL-2 promoter (54) was used for competition,the DNA-protein complex could not be seen for both sites (Fig.2B, lanes 5 and 10), further suggesting that the two NFAT sitesfound in the �600/�79 Sftpd promoter bound NFATs. To es-tablish that nuclear proteins from MLE-15 cells indeed boundNFATc3 to the two NFAT sites, gel supershift experimentswere carried out. Antibodies to NFATc3 supershifted DNA-protein complexes formed on NFAT-I and NFAT-II sites (Fig.2C, lanes 3, 4, 8, and 9), whereas there was no change in themobility of the complexes in the presence of bovine serumalbumin. To further demonstrate that nuclear proteins fromMLE-15 cells bound NFAT sites in the context of the nativepromoter, DNase I protection assays were carried out on the�246/�79 Sftpd fragment. Strong DNase I protection regionsspanning two NFAT sites, NFAT-I and NFAT-II (Fig. 4A), andAP-1, TTF-1, and C/EBP sites were identified as part of the
protected or hypersensitive region (Fig. 4A). The region nearTTF-1 binding site was hypersensitive to DNase I at low nu-clear protein concentrations (Fig. 4A, lane 4), whereas theregion around the AP-1 site showed increased DNase I protec-tion in contrast to the hypersensitivity found on naked DNA.Changes in DNase I hypersensitivity were observed 3� to theNFAT-II site (Fig. 4A, open boxes). Such changes in DNase Idigestion patterns are a hallmark for the formation of specificDNA-protein complexes that induce DNA distortions leading toaltered DNase I sensitivity (59), suggesting a possible multi-protein complex binding at the area of DNase I protection thatmay comprise NFAT, TTF-1, AP-1, and other factors.
Expression of NFATc3 and Calcineurin in the Lung Epithe-lium—Since TTF-1 is strictly expressed in the respiratory ep-ithelium, it was relevant to test the expression of NFATc3 todetermine whether TTF-1 and NFATc3 are co-expressed. Ex-pression of calcineurin, a phosphatase that activates NFATs,was also assessed. NFATc3 and CnB1 were detected by immu-nostaining (Fig. 3). Calcineurin was detected in the cytoplasmat apical and basal regions of respiratory epithelial cells inconducting airways and peripheral regions of the lung at em-bryonic days 15.5–17.5. The intensity of staining was highest inthe peripheral lung (Fig. 3A). CnB1 was also detected in therespiratory epithelium of the adult lung, wherein intense nu-clear staining was noted in the bronchiolar region. (Fig. 3, B
FIG. 1. TTF-1 and NFATs activate the Sftpd promoter. A, schematic representation of the �600 bp Sftpd promoter-luciferase (D-600-Luc)construct depicting positions of the TATA-box, the 5�-untranslated region to �79 bp containing the 39 bp of exon-I. B, dose response of thetransfected D-600-Luc into HeLa and MLE-15, an immortalized mouse lung epithelial cell line, measured as luciferase activity. C, effect of TTF-1on promoter activity was assessed after co-transfection of a fixed amount of D-600-Luc (1 �g), with increasing amounts of TTF-1 expression plasmid(RC-CMV-TTF-1) at 0.1, 0.25, 0.5, 1.0, and 2 �g per well into HeLa cells in 6-well plates. Values represent two independent experiments carriedout in duplicate with means � S.D. (n � 4). D, effect of full-length (Fu-NFATc3 and Fu-NFATc4) and constitutively active (CA-NFATc3 andCA-NFATc4) NFAT expression on D-600-Luc activity. Expression vectors for NFAT cDNA (2 �g) were cotransfected with D-600-Luc (0.5 �g) intoMLE cells. Values were obtained from three experiments carried out in duplicate, means � S.D. (n � 6).
NFAT, TTF-1, and SP-D Gene Transcription 34581
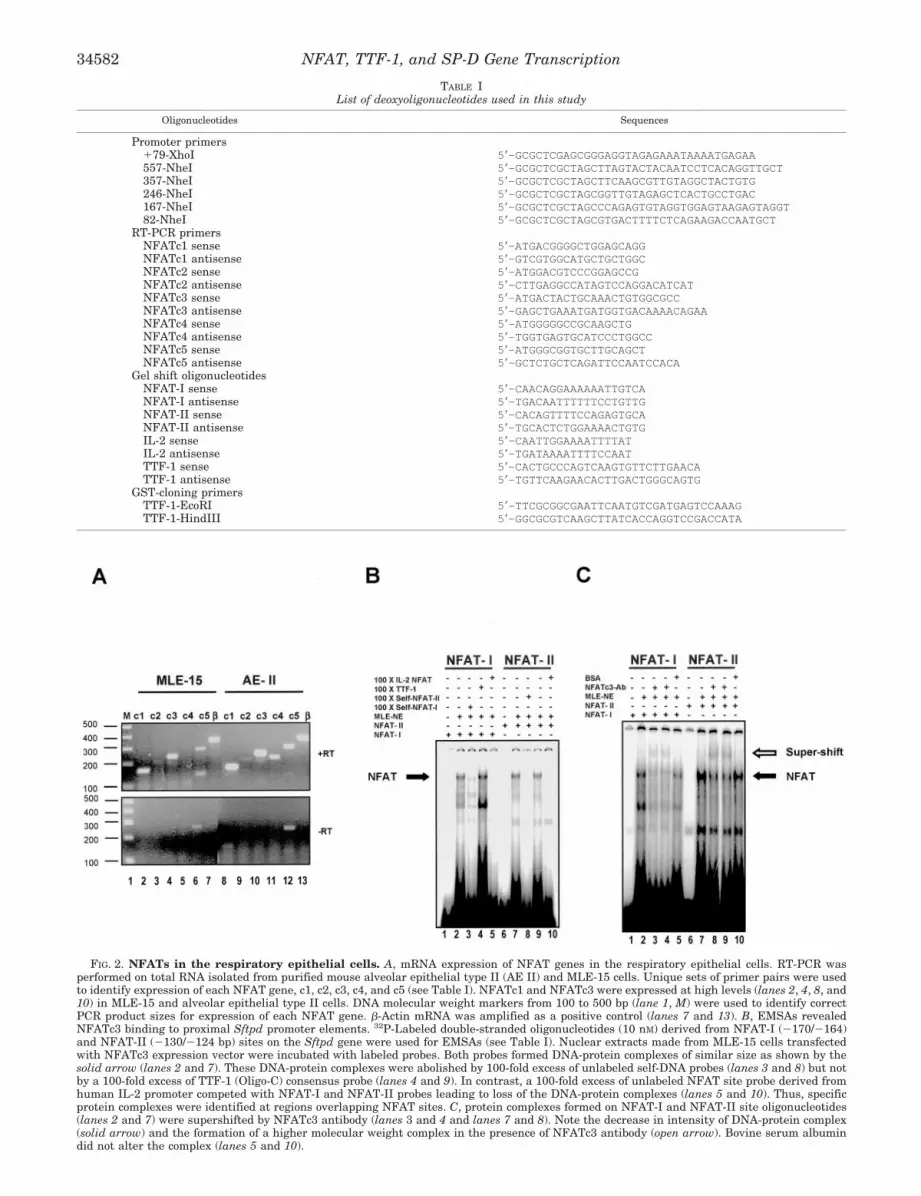
FIG. 2. NFATs in the respiratory epithelial cells. A, mRNA expression of NFAT genes in the respiratory epithelial cells. RT-PCR wasperformed on total RNA isolated from purified mouse alveolar epithelial type II (AE II) and MLE-15 cells. Unique sets of primer pairs were usedto identify expression of each NFAT gene, c1, c2, c3, c4, and c5 (see Table I). NFATc1 and NFATc3 were expressed at high levels (lanes 2, 4, 8, and10) in MLE-15 and alveolar epithelial type II cells. DNA molecular weight markers from 100 to 500 bp (lane 1, M) were used to identify correctPCR product sizes for expression of each NFAT gene. �-Actin mRNA was amplified as a positive control (lanes 7 and 13). B, EMSAs revealedNFATc3 binding to proximal Sftpd promoter elements. 32P-Labeled double-stranded oligonucleotides (10 nM) derived from NFAT-I (�170/�164)and NFAT-II (�130/�124 bp) sites on the Sftpd gene were used for EMSAs (see Table I). Nuclear extracts made from MLE-15 cells transfectedwith NFATc3 expression vector were incubated with labeled probes. Both probes formed DNA-protein complexes of similar size as shown by thesolid arrow (lanes 2 and 7). These DNA-protein complexes were abolished by 100-fold excess of unlabeled self-DNA probes (lanes 3 and 8) but notby a 100-fold excess of TTF-1 (Oligo-C) consensus probe (lanes 4 and 9). In contrast, a 100-fold excess of unlabeled NFAT site probe derived fromhuman IL-2 promoter competed with NFAT-I and NFAT-II probes leading to loss of the DNA-protein complexes (lanes 5 and 10). Thus, specificprotein complexes were identified at regions overlapping NFAT sites. C, protein complexes formed on NFAT-I and NFAT-II site oligonucleotides(lanes 2 and 7) were supershifted by NFATc3 antibody (lanes 3 and 4 and lanes 7 and 8). Note the decrease in intensity of DNA-protein complex(solid arrow) and the formation of a higher molecular weight complex in the presence of NFATc3 antibody (open arrow). Bovine serum albumindid not alter the complex (lanes 5 and 10).
TABLE IList of deoxyoligonucleotides used in this study
Oligonucleotides Sequences
Promoter primers�79-XhoI 5�-GCGCTCGAGCGGGAGGTAGAGAAATAAAATGAGAA557-NheI 5�-GCGCTCGCTAGCTTAGTACTACAATCCTCACAGGTTGCT357-NheI 5�-GCGCTCGCTAGCTTCAAGCGTTGTAGGCTACTGTG246-NheI 5�-GCGCTCGCTAGCGGTTGTAGAGCTCACTGCCTGAC167-NheI 5�-GCGCTCGCTAGCCCAGAGTGTAGGTGGAGTAAGAGTAGGT82-NheI 5�-GCGCTCGCTAGCGTGACTTTTCTCAGAAGACCAATGCT
RT-PCR primersNFATc1 sense 5�-ATGACGGGGCTGGAGCAGGNFATc1 antisense 5�-GTCGTGGCATGCTGCTGGCNFATc2 sense 5�-ATGGACGTCCCGGAGCCGNFATc2 antisense 5�-CTTGAGGCCATAGTCCAGGACATCATNFATc3 sense 5�-ATGACTACTGCAAACTGTGGCGCCNFATc3 antisense 5�-GAGCTGAAATGATGGTGACAAAACAGAANFATc4 sense 5�-ATGGGGGCCGCAAGCTGNFATc4 antisense 5�-TGGTGAGTGCATCCCTGGCCNFATc5 sense 5�-ATGGGCGGTGCTTGCAGCTNFATc5 antisense 5�-GCTCTGCTCAGATTCCAATCCACA
Gel shift oligonucleotidesNFAT-I sense 5�-CAACAGGAAAAAATTGTCANFAT-I antisense 5�-TGACAATTTTTTCCTGTTGNFAT-II sense 5�-CACAGTTTTCCAGAGTGCANFAT-II antisense 5�-TGCACTCTGGAAAACTGTGIL-2 sense 5�-CAATTGGAAAATTTTATIL-2 antisense 5�-TGATAAAATTTTCCAATTTF-1 sense 5�-CACTGCCCAGTCAAGTGTTCTTGAACATTF-1 antisense 5�-TGTTCAAGAACACTTGACTGGGCAGTG
GST-cloning primersTTF-1-EcoRI 5�-TTCGCGGCGAATTCAATGTCGATGAGTCCAAAGTTF-1-HindIII 5�-GGCGCGTCAAGCTTATCACCAGGTCCGACCATA
NFAT, TTF-1, and SP-D Gene Transcription34582
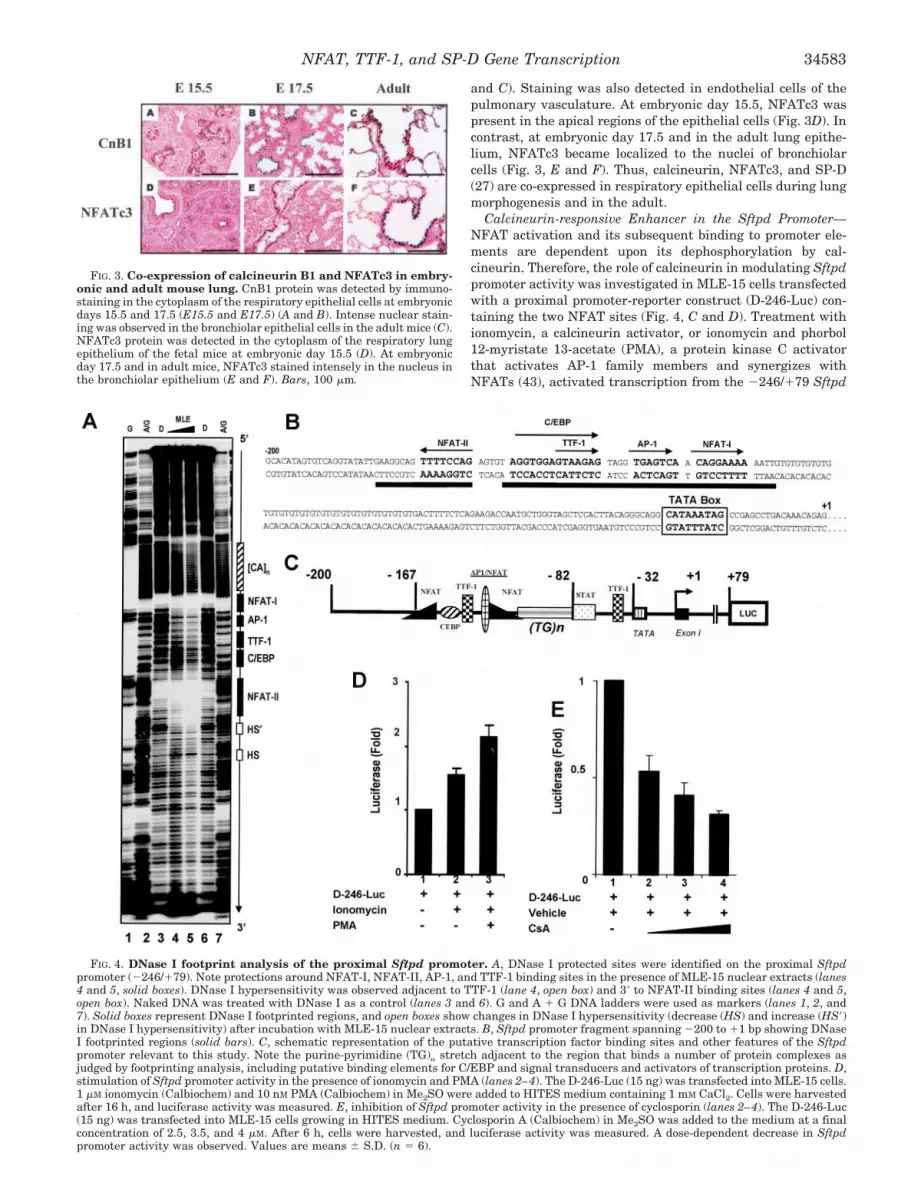
and C). Staining was also detected in endothelial cells of thepulmonary vasculature. At embryonic day 15.5, NFATc3 waspresent in the apical regions of the epithelial cells (Fig. 3D). Incontrast, at embryonic day 17.5 and in the adult lung epithe-lium, NFATc3 became localized to the nuclei of bronchiolarcells (Fig. 3, E and F). Thus, calcineurin, NFATc3, and SP-D(27) are co-expressed in respiratory epithelial cells during lungmorphogenesis and in the adult.
Calcineurin-responsive Enhancer in the Sftpd Promoter—NFAT activation and its subsequent binding to promoter ele-ments are dependent upon its dephosphorylation by cal-cineurin. Therefore, the role of calcineurin in modulating Sftpdpromoter activity was investigated in MLE-15 cells transfectedwith a proximal promoter-reporter construct (D-246-Luc) con-taining the two NFAT sites (Fig. 4, C and D). Treatment withionomycin, a calcineurin activator, or ionomycin and phorbol12-myristate 13-acetate (PMA), a protein kinase C activatorthat activates AP-1 family members and synergizes withNFATs (43), activated transcription from the �246/�79 Sftpd
FIG. 4. DNase I footprint analysis of the proximal Sftpd promoter. A, DNase I protected sites were identified on the proximal Sftpdpromoter (�246/�79). Note protections around NFAT-I, NFAT-II, AP-1, and TTF-1 binding sites in the presence of MLE-15 nuclear extracts (lanes4 and 5, solid boxes). DNase I hypersensitivity was observed adjacent to TTF-1 (lane 4, open box) and 3� to NFAT-II binding sites (lanes 4 and 5,open box). Naked DNA was treated with DNase I as a control (lanes 3 and 6). G and A � G DNA ladders were used as markers (lanes 1, 2, and7). Solid boxes represent DNase I footprinted regions, and open boxes show changes in DNase I hypersensitivity (decrease (HS) and increase (HS�)in DNase I hypersensitivity) after incubation with MLE-15 nuclear extracts. B, Sftpd promoter fragment spanning �200 to �1 bp showing DNaseI footprinted regions (solid bars). C, schematic representation of the putative transcription factor binding sites and other features of the Sftpdpromoter relevant to this study. Note the purine-pyrimidine (TG)n stretch adjacent to the region that binds a number of protein complexes asjudged by footprinting analysis, including putative binding elements for C/EBP and signal transducers and activators of transcription proteins. D,stimulation of Sftpd promoter activity in the presence of ionomycin and PMA (lanes 2–4). The D-246-Luc (15 ng) was transfected into MLE-15 cells.1 �M ionomycin (Calbiochem) and 10 nM PMA (Calbiochem) in Me2SO were added to HITES medium containing 1 mM CaCl2. Cells were harvestedafter 16 h, and luciferase activity was measured. E, inhibition of Sftpd promoter activity in the presence of cyclosporin (lanes 2–4). The D-246-Luc(15 ng) was transfected into MLE-15 cells growing in HITES medium. Cyclosporin A (Calbiochem) in Me2SO was added to the medium at a finalconcentration of 2.5, 3.5, and 4 �M. After 6 h, cells were harvested, and luciferase activity was measured. A dose-dependent decrease in Sftpdpromoter activity was observed. Values are means � S.D. (n � 6).
FIG. 3. Co-expression of calcineurin B1 and NFATc3 in embry-onic and adult mouse lung. CnB1 protein was detected by immuno-staining in the cytoplasm of the respiratory epithelial cells at embryonicdays 15.5 and 17.5 (E15.5 and E17.5) (A and B). Intense nuclear stain-ing was observed in the bronchiolar epithelial cells in the adult mice (C).NFATc3 protein was detected in the cytoplasm of the respiratory lungepithelium of the fetal mice at embryonic day 15.5 (D). At embryonicday 17.5 and in adult mice, NFATc3 stained intensely in the nucleus inthe bronchiolar epithelium (E and F). Bars, 100 �m.
NFAT, TTF-1, and SP-D Gene Transcription 34583
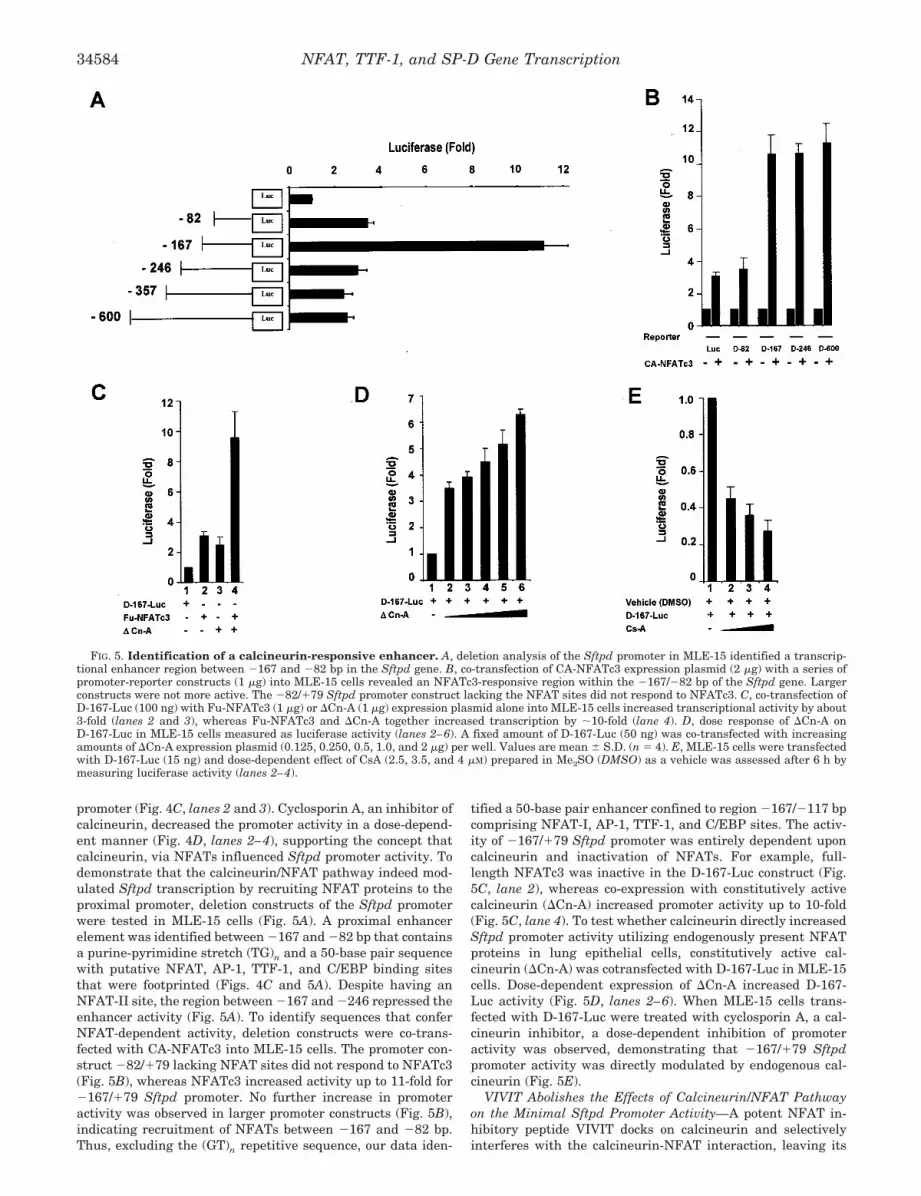
promoter (Fig. 4C, lanes 2 and 3). Cyclosporin A, an inhibitor ofcalcineurin, decreased the promoter activity in a dose-depend-ent manner (Fig. 4D, lanes 2–4), supporting the concept thatcalcineurin, via NFATs influenced Sftpd promoter activity. Todemonstrate that the calcineurin/NFAT pathway indeed mod-ulated Sftpd transcription by recruiting NFAT proteins to theproximal promoter, deletion constructs of the Sftpd promoterwere tested in MLE-15 cells (Fig. 5A). A proximal enhancerelement was identified between �167 and �82 bp that containsa purine-pyrimidine stretch (TG)n and a 50-base pair sequencewith putative NFAT, AP-1, TTF-1, and C/EBP binding sitesthat were footprinted (Figs. 4C and 5A). Despite having anNFAT-II site, the region between �167 and �246 repressed theenhancer activity (Fig. 5A). To identify sequences that conferNFAT-dependent activity, deletion constructs were co-trans-fected with CA-NFATc3 into MLE-15 cells. The promoter con-struct �82/�79 lacking NFAT sites did not respond to NFATc3(Fig. 5B), whereas NFATc3 increased activity up to 11-fold for�167/�79 Sftpd promoter. No further increase in promoteractivity was observed in larger promoter constructs (Fig. 5B),indicating recruitment of NFATs between �167 and �82 bp.Thus, excluding the (GT)n repetitive sequence, our data iden-
tified a 50-base pair enhancer confined to region �167/�117 bpcomprising NFAT-I, AP-1, TTF-1, and C/EBP sites. The activ-ity of �167/�79 Sftpd promoter was entirely dependent uponcalcineurin and inactivation of NFATs. For example, full-length NFATc3 was inactive in the D-167-Luc construct (Fig.5C, lane 2), whereas co-expression with constitutively activecalcineurin (�Cn-A) increased promoter activity up to 10-fold(Fig. 5C, lane 4). To test whether calcineurin directly increasedSftpd promoter activity utilizing endogenously present NFATproteins in lung epithelial cells, constitutively active cal-cineurin (�Cn-A) was cotransfected with D-167-Luc in MLE-15cells. Dose-dependent expression of �Cn-A increased D-167-Luc activity (Fig. 5D, lanes 2–6). When MLE-15 cells trans-fected with D-167-Luc were treated with cyclosporin A, a cal-cineurin inhibitor, a dose-dependent inhibition of promoteractivity was observed, demonstrating that �167/�79 Sftpdpromoter activity was directly modulated by endogenous cal-cineurin (Fig. 5E).
VIVIT Abolishes the Effects of Calcineurin/NFAT Pathwayon the Minimal Sftpd Promoter Activity—A potent NFAT in-hibitory peptide VIVIT docks on calcineurin and selectivelyinterferes with the calcineurin-NFAT interaction, leaving its
FIG. 5. Identification of a calcineurin-responsive enhancer. A, deletion analysis of the Sftpd promoter in MLE-15 identified a transcrip-tional enhancer region between �167 and �82 bp in the Sftpd gene. B, co-transfection of CA-NFATc3 expression plasmid (2 �g) with a series ofpromoter-reporter constructs (1 �g) into MLE-15 cells revealed an NFATc3-responsive region within the �167/�82 bp of the Sftpd gene. Largerconstructs were not more active. The �82/�79 Sftpd promoter construct lacking the NFAT sites did not respond to NFATc3. C, co-transfection ofD-167-Luc (100 ng) with Fu-NFATc3 (1 �g) or �Cn-A (1 �g) expression plasmid alone into MLE-15 cells increased transcriptional activity by about3-fold (lanes 2 and 3), whereas Fu-NFATc3 and �Cn-A together increased transcription by �10-fold (lane 4). D, dose response of �Cn-A onD-167-Luc in MLE-15 cells measured as luciferase activity (lanes 2–6). A fixed amount of D-167-Luc (50 ng) was co-transfected with increasingamounts of �Cn-A expression plasmid (0.125, 0.250, 0.5, 1.0, and 2 �g) per well. Values are mean � S.D. (n � 4). E, MLE-15 cells were transfectedwith D-167-Luc (15 ng) and dose-dependent effect of CsA (2.5, 3.5, and 4 �M) prepared in Me2SO (DMSO) as a vehicle was assessed after 6 h bymeasuring luciferase activity (lanes 2–4).
NFAT, TTF-1, and SP-D Gene Transcription34584
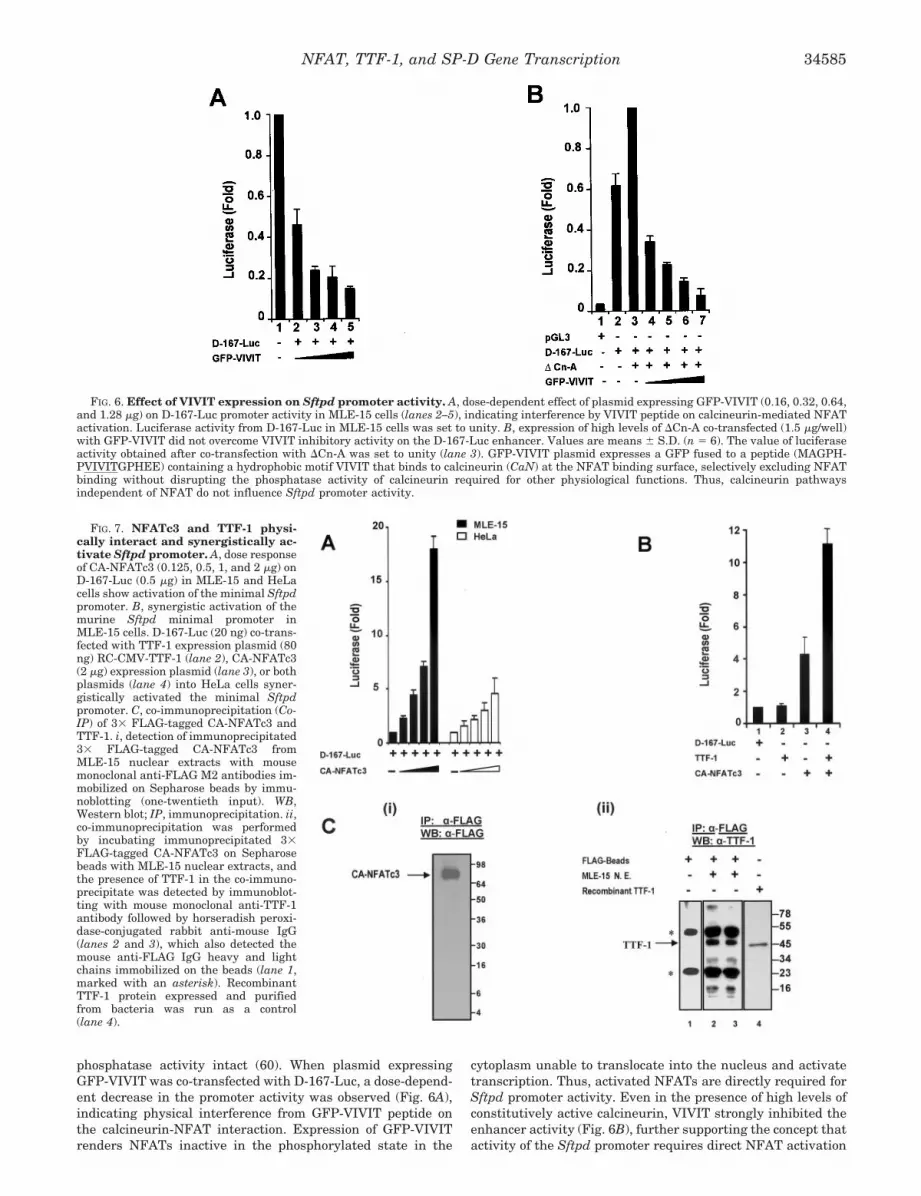
phosphatase activity intact (60). When plasmid expressingGFP-VIVIT was co-transfected with D-167-Luc, a dose-depend-ent decrease in the promoter activity was observed (Fig. 6A),indicating physical interference from GFP-VIVIT peptide onthe calcineurin-NFAT interaction. Expression of GFP-VIVITrenders NFATs inactive in the phosphorylated state in the
cytoplasm unable to translocate into the nucleus and activatetranscription. Thus, activated NFATs are directly required forSftpd promoter activity. Even in the presence of high levels ofconstitutively active calcineurin, VIVIT strongly inhibited theenhancer activity (Fig. 6B), further supporting the concept thatactivity of the Sftpd promoter requires direct NFAT activation
FIG. 7. NFATc3 and TTF-1 physi-cally interact and synergistically ac-tivate Sftpd promoter. A, dose responseof CA-NFATc3 (0.125, 0.5, 1, and 2 �g) onD-167-Luc (0.5 �g) in MLE-15 and HeLacells show activation of the minimal Sftpdpromoter. B, synergistic activation of themurine Sftpd minimal promoter inMLE-15 cells. D-167-Luc (20 ng) co-trans-fected with TTF-1 expression plasmid (80ng) RC-CMV-TTF-1 (lane 2), CA-NFATc3(2 �g) expression plasmid (lane 3), or bothplasmids (lane 4) into HeLa cells syner-gistically activated the minimal Sftpdpromoter. C, co-immunoprecipitation (Co-IP) of 3� FLAG-tagged CA-NFATc3 andTTF-1. i, detection of immunoprecipitated3� FLAG-tagged CA-NFATc3 fromMLE-15 nuclear extracts with mousemonoclonal anti-FLAG M2 antibodies im-mobilized on Sepharose beads by immu-noblotting (one-twentieth input). WB,Western blot; IP, immunoprecipitation. ii,co-immunoprecipitation was performedby incubating immunoprecipitated 3�FLAG-tagged CA-NFATc3 on Sepharosebeads with MLE-15 nuclear extracts, andthe presence of TTF-1 in the co-immuno-precipitate was detected by immunoblot-ting with mouse monoclonal anti-TTF-1antibody followed by horseradish peroxi-dase-conjugated rabbit anti-mouse IgG(lanes 2 and 3), which also detected themouse anti-FLAG IgG heavy and lightchains immobilized on the beads (lane 1,marked with an asterisk). RecombinantTTF-1 protein expressed and purifiedfrom bacteria was run as a control(lane 4).
FIG. 6. Effect of VIVIT expression on Sftpd promoter activity. A, dose-dependent effect of plasmid expressing GFP-VIVIT (0.16, 0.32, 0.64,and 1.28 �g) on D-167-Luc promoter activity in MLE-15 cells (lanes 2–5), indicating interference by VIVIT peptide on calcineurin-mediated NFATactivation. Luciferase activity from D-167-Luc in MLE-15 cells was set to unity. B, expression of high levels of �Cn-A co-transfected (1.5 �g/well)with GFP-VIVIT did not overcome VIVIT inhibitory activity on the D-167-Luc enhancer. Values are means � S.D. (n � 6). The value of luciferaseactivity obtained after co-transfection with �Cn-A was set to unity (lane 3). GFP-VIVIT plasmid expresses a GFP fused to a peptide (MAGPH-PVIVITGPHEE) containing a hydrophobic motif VIVIT that binds to calcineurin (CaN) at the NFAT binding surface, selectively excluding NFATbinding without disrupting the phosphatase activity of calcineurin required for other physiological functions. Thus, calcineurin pathwaysindependent of NFAT do not influence Sftpd promoter activity.
NFAT, TTF-1, and SP-D Gene Transcription 34585

and is not activated by calcineurin pathways independently ofNFATs. Taken together, the activity of the proximal Sftpdpromoter is modulated by the calcineurin/NFAT pathway inrespiratory epithelial cells.
NFATc3 and TTF-1 Physically Interact and SynergisticallyActivate the Sftpd Promoter—Since the functional NFAT sitesare adjacent to the TTF-1 site (Fig. 4B), functional cooperativ-ity and physical interaction of NFATc3 and TTF-1 proteins onthe D-167-Luc proximal enhancer were tested. Unlike D-600m-Luc, NFATc3 activated the �167/�79 Sftpd promoter in HeLacells, thus making it amenable for functional studies (Fig. 7A).CA-NFATc3 increased the activity of the �167/�79 Sftpd pro-moter and synergistically enhanced transactivation by TTF-1in HeLa cells (Fig. 7B). To demonstrate physical associationbetween NFATc3 and TTF-1 in respiratory epithelial cells, 3�FLAG-tagged CA-NFATc3 protein expressed in MLE-15 cellswas immunoprecipitated with anti-FLAG mouse monoclonalantibodies attached to Sepharose beads (Fig. 7C, i). Co-immu-noprecipitation of TTF-1 with FLAG-tagged NFATc3 was con-firmed by blotting with mouse monoclonal antibody to TTF-1(Fig 7C, ii).
The TTF-1 Homeodomain (HD) Is Required for Direct Phys-ical Interaction with NFATc3—To determine whether directprotein-protein interactions mediated the observed cooperationbetween TTF-1 and NFATc3, GST pull-down assays were per-formed with GST fused to full-length TTF-1 (GST-TTF-1) andin vitro transcribed and translated radiolabeled [35S]Met-CA-NFATc3. GST-TTF-1 full-length protein strongly interactedwith CA-NFATc3 (Fig. 8A, lanes 2 and 3), indicating directphysical interaction between TTF-1 and NFATc3 in vitro. To
identify the TTF-1 domains involved in physical interactionwith NFATc3, a series of GST-TTF-1 protein deletion con-structs were made (Fig. 8C) and incubated with in vitro tran-scribed and translated radiolabeled [35S]Met-CA-NFATc3.TTF-1 HD (residues 161–223) alone (Fig. 8B, lanes 5 and 6),TTF-1 HD with the amino-terminal (residues 1–223) region(Fig. 8B, lanes 7 and 8), and TTF-1 HD with the carboxyl-terminal (residues 161–372) region (Fig. 8B, lanes 9 and 10)strongly interacted with CA-NFATc3. In contrast, GST-TTF-1amino-terminal (residues 1–160) region (lanes 3 and 4) andGST-TTF-1 carboxyl-terminal (residues 224–372) region (lanes11 and 12), which do not contain the HD, did not bind [35S]Met-CA-NFATc3. TTF-1 HD was necessary and sufficient for directphysical interaction with NFATc3, whereas the flanking ami-no-terminal and carboxyl-terminal activation domains did notphysically interact with NFATc3.
DISCUSSION
Expression of SP-D is highly regulated in the respiratoryepithelium, increasing prior to birth, following a variety of lunginjury, exposure to pathogens, and in response to Th2-cyto-kines. Transcriptional pathways regulating SP-D expressionare largely unknown. In this study, we identified an endoge-nous calcineurin/NFAT pathway in the respiratory epithelialcells that regulated Sftpd transcription. Calcineurin andNFATc3 co-expressed in the respiratory epithelial cells of thedeveloping and postnatal lung at sites of SP-D expression (27).A calcineurin-dependent transcriptional enhancer was identi-fied in the proximal Sftpd promoter. This region contained anNFAT�AP-1 composite element adjacent to a TTF-1 binding
FIG. 8. TTF-1 HD is required for direct physical interaction with NFATc3. GST pull-down assays were performed with bacteriallyexpressed GST or GST fused to full-length TTF-1 (GST-TTF-1) or various TTF-1 domains. Proteins were immobilized on glutathione-Sepharosebeads and incubated with in vitro transcribed and translated radiolabeled [35S]Met-CA-NFATc3. A, beads with GST protein alone (lane 1) did notbind, whereas beads with GST-TTF-1 full-length protein (lanes 2 and 3) bound [35S]Met-CA-NFATc3. Lane 4 shows one-tenth of the totalradiolabeled [35S]Met-CA-NFATc3 protein input. B, TTF-1 HD alone (lanes 5 and 6), TTF-1 HD with the amino-terminal region (lanes 7 and 8),and TTF-1 HD with the carboxyl-terminal region (lanes 9 and 10) strongly interacted with CA-NFATc3. In contrast, Sepharose beads without GSTprotein (lanes 1 and 2), with GST protein alone (lanes 13 and 14), with the GST-TTF-1 amino-terminal region (lanes 3 and 4), and with theGST-TTF-1 carboxyl-terminal region (lanes 11 and 12) did not bind [35S]Met-CA-NFATc3. C, schematic diagram of various GST-TTF-1 proteindomains and their interaction with CA-NFATc3 protein.
NFAT, TTF-1, and SP-D Gene Transcription34586

site. NFAT inhibitory peptide VIVIT (60) abolished the en-hancer activity, demonstrating the requirement of activatedNFATs to stimulate the Sftpd promoter. TTF-1 and NFATc3physically interacted and synergistically stimulated the Sftpdpromoter activity, supporting the concept that they are a partof a transcription complex regulating Sftpd transcription. Atranscriptional pathway mediated by interactions between ac-tivated NFATc3 and TTF-1 regulate SP-D expression in therespiratory epithelial cells.
Whereas NFATs were first identified in T-cell developmentin the immune system, they are expressed in many cell typesand contribute to diverse cellular functions (41). Calcineurinand NFATs have recently been implicated in development anddifferentiation of the nervous system, bone, cartilage, muscle,heart, skin, and fat tissues (41). Our studies demonstrate thatcalcineurin, NFATc3, and SP-D are coincident with TTF-1 pro-tein expression in the respiratory epithelium during fetal lungdevelopment. Whereas neither NFATc3 nor TTF-1 expressionis confined to the lung during development, these transcriptionfactors are co-expressed in the respiratory cells of the develop-ing lung, where they physically interacted and synergisticallyactivated Sftpd transcription. Consistent with this finding, al-veolar epithelial type II cells that co-express TTF-1 andNFATc3 mRNA secrete SP-D protein in culture (48). The role ofTTF-1 and its phosphorylation mediated by several kinasesplay a critical role in lung development (61, 62). Thus, dispar-ate signaling events that post-translationally influence TTF-1and NFATc3 function may converge on target genes to initiatetheir spatial, temporal, and cell-specific expression. Taken to-gether, our data support the hypothesis that the calcineurin/NFAT pathway interacts with TTF-1 to regulate SP-Dexpression.
Although SP-D has been regarded as constitutively ex-pressed (1), recent studies demonstrate activation of SP-D ex-pression following inflammation by exposure to pathogens, al-lergens, and pollutants (4, 32). The present study demonstratesthat Sftpd promoter is regulated by the calcineurin/NFATpathway. NFAT activates transcription of a number of genesfollowing an effective immune response (41). NFAT binds co-operatively with transcription factors of the AP-1 family tocomposite NFAT�AP-1 sites found in the regulatory regions ofimmunomodulatory genes (43). Indeed, an NFAT�AP-1 compos-ite site in the Sftpd proximal promoter (�139/�125) producedDNase I footprints with nuclear proteins from lung epithelialcells and was responsive to ionomycin and PMA, a hallmark ofNFAT�AP-1 sites that bind activated NFAT and AP-1 family oftranscription factors in target genes (41). Human SFTPD isregulated by AP-1 family members (24). Because AP-1 activityisalsomodulatedbydiversesignalingpathways(63),NFAT�AP-1-dependent transcription may integrate responses to lunginjury.
TTF-1 regulates transcription of genes involved in lung mor-phogenesis and surfactant protein production (64). TTF-1 syn-ergistically activates transcription of lung surfactant genes andseveral genes involved in lung development by direct protein-protein interaction or functional cooperation with a number oftranscription factors (47, 65, 66). For example, TTF-1 interactswith GATA6 to activate SP-C and Wnt7b transcription (47, 67).Transcriptional co-activator TAZ and NFI interact with TTF-1and synergistically activate the SP-C gene (65, 68), whereassurfactant protein A and SP-B transcription is regulated byinteraction of TTF-1 with CREB-binding protein and SRC-1(66, 69). TTF-1 also interacts with RAR/RXR and BR22 (66, 70)to activate SP-B transcription. TTF-1 is a homeodomain-con-taining transcription factor. In general, HD proteins have lowDNA binding specificity and require partners and co-activators
to bind specific regulatory regions of target genes (71). Thisinherent flexibility to interact with relevant cofactors allowsthem to execute precise spatial, temporal, and tissue-specificroles (71). Our findings demonstrate that TTF-1 acts as acofactor for NFATc3 and synergistically activates Sftpd tran-scription. NFAT proteins have relatively low affinity for DNAbinding sites (41). NFAT proteins interact with several familiesof transcription factors and cooperatively bind DNA as het-erodimers on native promoters of target genes and synergizetranscription (41). NFAT�AP-1 complexes have discrete andflexible polar patches providing interaction surfaces for bindingof different transcription factor (72). TTF-1-NFAT interactionsmay be facilitated by the close apposition of the TTF-1 andNFAT binding elements in the Sftpd proximal promoter. TTF-1HD alone interacted with NFATc3 and was required for pro-tein-protein interaction with NFATc3. Recently, TTF-1 home-odomain interaction with the carboxyl-terminal zinc finger do-main of GATA6 synergizing transcription from the Sftpcpromoter was demonstrated (47). Whether GATA6 andNFATc3 interact with the same region of the TTF-1 HD is aninteresting question. Nonetheless, transcriptional synergismdue to physical interaction of homeodomains with other tran-scription factors is well documented. For example, Pitx HDinteracts with basic helix-loop-helix proteins on the pro-opiomelanocortin promoter, whereas Pdx1 HD interacts withPan1 on the rat insulin promoter (8, 73).
The present study identifies active components of the cal-cineurin/NFAT pathway in the lung epithelial cells. Cal-cineurin and NFATc3 are expressed coincidentally with TTF-1and SP-D in respiratory epithelial cells of the fetal and adultlung. NFATc3 and TTF-1 physically interacted and synergisti-cally activated Sftpd transcription.
REFERENCES
1. Crouch, E., and Wright, J. R. (2001) Annu. Rev. Physiol. 63, 521–5542. Madsen, J., Kliem, A., Tornoe, I., Skjodt, K., Koch, C., and Holmskov, U. (2000)
J. Immunol. 164, 5866–58703. Hawgood, S., and Poulain, F. R. (2001) Annu. Rev. Physiol. 63, 495–5194. Crouch, E. C. (1998) Biochim. Biophys. Acta 1408, 278–2895. Reid, K. B. (1998) Immunobiology 199, 200–2076. Mason, R. J., Greene, K., and Voelker, D. R. (1998) Am. J. Physiol. 275,
L1–L137. Gardai, S. J., Xiao, Y. Q., Dickinson, M., Nick, J. A., Voelker, D. R., Greene,
K. E., and Henson, P. M. (2003) Cell 115, 13–238. Ohneda, K., Mirmira, R. G., Wang, J., Johnson, J. D., and German, M. S.
(2000) Mol. Cell. Biol. 20, 900–9119. McCormack, F. X., and Whitsett, J. A. (2002) J. Clin. Invest. 109, 707–712
10. Wu, H., Kuzmenko, A., Wan, S., Schaffer, L., Weiss, A., Fisher, J. H., Kim,K. S., and McCormack, F. X. (2003) J. Clin. Invest. 111, 1589–1602
11. Wright, J. R., Borron, P., Brinker, K. G., and Folz, R. J. (2001) Am. J. Respir.Cell Mol. Biol. 24, 513–517
12. LeVine, A. M., and Whitsett, J. A. (2001) Microbes Infect. 3, 161–16613. Yoshida, M., Korfhagen, T. R., and Whitsett, J. A. (2001) J. Immunol. 166,
7514–751914. Ikegami, M., Hull, W. M., Yoshida, M., Wert, S. E., and Whitsett, J. A. (2001)
Am. J. Physiol. 281, L697–L70315. Van Iwaarden, J. F., Shimizu, H., Van Golde, P. H., Voelker, D. R., and Van
Golde, L. M. (1992) Biochem. J. 286, 5–816. Clark, H., Palaniyar, N., Strong, P., Edmondson, J., Hawgood, S., and Reid,
K. B. (2002) J. Immunol. 169, 2892–289917. Crouch, E. C., Persson, A., Griffin, G. L., Chang, D., and Senior, R. M. (1995)
Am. J. Respir. Cell Mol. Biol. 12, 410–41518. Tino, M. J., and Wright, J. R. (1999) Am. J. Physiol. 276, L164–L17419. Madan, T., Eggleton, P., Kishore, U., Strong, P., Aggrawal, S. S., Sarma, P. U.,
and Reid, K. B. (1997) Infect. Immun. 65, 3171–317920. Palaniyar, N., Zhang, L., Kuzmenko, A., Ikegami, M., Wan, S., Wu, H., Korf-
hagen, T. R., Whitsett, J. A., and McCormack, F. X. (2002) J. Biol. Chem.277, 26971–26979
21. Wright, J. R. (2003) J. Clin. Invest. 111, 1453–145522. Wert, S. E., Yoshida, M., LeVine, A. M., Ikegami, M., Jones, T., Ross, G. F.,
Fisher, J. H., Korfhagen, T. R., and Whitsett, J. A. (2000) Proc. Natl. Acad.Sci. U. S. A. 97, 5972–5977
23. LeVine, A. M., Whitsett, J. A., Gwozdz, J. A., Richardson, T. R., Fisher, J. H.,Burhans, M. S., and Korfhagen, T. R. (2000) J. Immunol. 165, 3934–3940
24. He, Y., Crouch, E. C., Rust, K., Spaite, E., and Brody, S. L. (2000) J. Biol.Chem. 275, 31051–31060
25. Ogasawara, Y., Kuroki, Y., Tsuzuki, A., Ueda, S., Misaki, H., and Akino, T.(1992) Life Sci. 50, 1761–1767
26. Crouch, E., Rust, K., Marienchek, W., Parghi, D., Chang, D., and Persson, A.(1991) Am. J. Respir. Cell Mol. Biol. 5, 13–18
NFAT, TTF-1, and SP-D Gene Transcription 34587

27. Wong, C. J., Akiyama, J., Allen, L., and Hawgood, S. (1996) Pediatr. Res. 39,930–937
28. Gotoh, T., Chowdhury, S., Takiguchi, M., and Mori, M. (1997) J. Biol. Chem.272, 3694–3698
29. He, Y., and Crouch, E. (2002) J. Biol. Chem. 277, 19530–1953730. Dentener, M. A., Vreugdenhil, A. C., Hoet, P. H., Vernooy, J. H., Nieman, F. H.,
Heumann, D., Janssen, Y. M., Buurman, W. A., and Wouters, E. F. (2000)Am. J. Respir. Cell Mol. Biol. 23, 146–153
31. Crestani, B., Rolland, C., Lardeux, B., Fournier, T., Bernuau, D., Pous, C.,Vissuzaine, C., Li, L., and Aubier, M. (1998) J. Immunol. 160, 4596–4605
32. McIntosh, J. C., Swyers, A. H., Fisher, J. H., and Wright, J. R. (1996) Am. J.Respir. Cell Mol. Biol. 15, 509–519
33. Haidaris, P. J. (1997) Blood 89, 873–88234. Jain-Vora, S., LeVine, A. M., Chroneos, Z., Ross, G. F., Hull, W. M., and
Whitsett, J. A. (1998) Infect. Immun. 66, 4229–423635. Aderibigbe, A. O., Thomas, R. F., Mercer, R. R., and Auten, R. L., Jr. (1999)
Am. J. Respir. Cell Mol. Biol. 20, 219–22736. Xu, X., McCormick-Shannon, K., Voelker, D. R., and Mason, R. J. (1998) Am. J.
Respir. Cell Mol. Biol. 18, 168–17837. Ikegami, M., Whitsett, J. A., Chroneos, Z. C., Ross, G. F., Reed, J. A., Bachur-
ski, C. J., and Jobe, A. H. (2000) Am. J. Physiol. 278, L75–L8038. Homer, R. J., Zheng, T., Chupp, G., He, S., Zhu, Z., Chen, Q., Ma, B., Hite,
R. D., Gobran, L. I., Rooney, S. A., and Elias, J. A. (2002) Am. J. Physiol.283, L52–L59
39. Poli, V. (1998) J. Biol. Chem. 273, 29279–2928240. Alonzi, T., Maritano, D., Gorgoni, B., Rizzuto, G., Libert, C., and Poli, V. (2001)
Mol. Cell. Biol. 21, 1621–163241. Hogan, P. G., Chen, L., Nardone, J., and Rao, A. (2003) Genes Dev. 17,
2205–223242. Stahlman, M. T., Gray, M. E., and Whitsett, J. A. (1996) J. Histochem. Cyto-
chem. 44, 673–67843. Rao, A., Luo, C., and Hogan, P. G. (1997) Annu. Rev. Immunol. 15, 707–74744. Rusnak, F., and Mertz, P. (2000) Physiol. Rev. 80, 1483–152145. Crabtree, G. R., and Olson, E. N. (2002) Cell 109, (suppl.) 67–7946. Wikenheiser, K. A., Vorbroker, D. K., Rice, W. R., Clark, J. C., Bachurski, C. J.,
Oie, H. K., and Whitsett, J. A. (1993) Proc. Natl. Acad. Sci. U. S. A. 90,11029–11033
47. Liu, C., Glasser, S. W., Wan, H., and Whitsett, J. A. (2002) J. Biol. Chem. 277,4519–4525
48. Rice, W. R., Conkright, J. J., Na, C. L., Ikegami, M., Shannon, J. M., andWeaver, T. E. (2002) Am. J. Physiol. 283, L256–L264
49. Wert, S. E., Glasser, S. W., Korfhagen, T. R., and Whitsett, J. A. (1993) Dev.Biol. 156, 426–443
50. Mucenski, M. L., Wert, S. E., Nation, J. M., Loudy, D. E., Huelsken, J.,Birchmeier, W., Morrisey, E. E., and Whitsett, J. A. (2003) J. Biol. Chem.
278, 40231–4023851. Zhou, L., Lim, L., Costa, R. H., and Whitsett, J. A. (1996) J. Histochem.
Cytochem. 44, 1183–119352. Baker, D. L., Dave, V., Reed, T., and Periasamy, M. (1996) J. Biol. Chem. 271,
5921–592853. Civitareale, D., Lonigro, R., Sinclair, A. J., and Di Lauro, R. (1989) EMBO J.
8, 2537–254254. Molkentin, J. D., Lu, J. R., Antos, C. L., Markham, B., Richardson, J., Robbins,
J., Grant, S. R., and Olson, E. N. (1998) Cell 93, 215–22855. Dave, V., Zhao, C., Yang, F., Tung, C. S., and Ma, J. (2000) Mol. Cell. Biol. 20,
7673–768456. Schreiber, E., Matthias, P., Muller, M. M., and Schaffner, W. (1989) Nucleic
Acids Res. 17, 641957. Mason, R. J., Lewis, M. C., Edeen, K. E., McCormick-Shannon, K., Nielsen,
L. D., and Shannon, J. M. (2002) Am. J. Physiol. 282, L249–L25858. Graef, I. A., Gastier, J. M., Francke, U., and Crabtree, G. R. (2001) Proc. Natl.
Acad. Sci. U. S. A. 98, 5740–574559. Suck, D. (1994) J. Mol. Recognit. 7, 65–7060. Aramburu, J., Yaffe, M. B., Lopez-Rodriguez, C., Cantley, L. C., Hogan, P. G.,
and Rao, A. (1999) Science 285, 2129–213361. Kimura, S., Hara, Y., Pineau, T., Fernandez-Salguero, P., Fox, C. H., Ward,
J. M., and Gonzalez, F. J. (1996) Genes Dev. 10, 60–6962. DeFelice, M., Silberschmidt, D., DiLauro, R., Xu, Y., Wert, S. E., Weaver, T. E.,
Bachurski, C. J., Clark, J. C., and Whitsett, J. A. (2003) J. Biol. Chem. 278,35574–35583
63. Davis, R. J. (2000) Cell 103, 239–25264. Whitsett, J. (1998) Nat. Genet. 20, 7–865. Bachurski, C. J., Yang, G. H., Currier, T. A., Gronostajski, R. M., and Hong, D.
(2003) Mol. Cell. Biol. 23, 9014–902466. Yan, C., Naltner, A., Conkright, J., and Ghaffari, M. (2001) J. Biol. Chem. 276,
21686–2169167. Weidenfeld, J., Shu, W., Zhang, L., Millar, S. E., and Morrisey, E. E. (2002)
J. Biol. Chem. 277, 21061–2107068. Park, K. S., Whitsett, J. A., Di Palma, T., Hong, J. H., Yaffe, M. B., and
Zannini, M. (2004) J. Biol. Chem.69. Yi, M., Tong, G. X., Murry, B., and Mendelson, C. R. (2002) J. Biol. Chem. 277,
2997–300570. Yang, M. C., Wang, B., Weissler, J. C., Margraf, L. R., and Yang, Y. S. (2003)
Eur. Respir. J. 22, 28–3471. Mannervik, M. (1999) BioEssays 21, 267–27072. Chen, L., Glover, J. N., Hogan, P. G., Rao, A., and Harrison, S. C. (1998) Nature
392, 42–4873. Poulin, G., Lebel, M., Chamberland, M., Paradis, F. W., and Drouin, J. (2000)
Mol. Cell. Biol. 20, 4826–4837
NFAT, TTF-1, and SP-D Gene Transcription34588




















