
Nuclear expression of a cytoplasmic male sterility gene modifies mitochondrial morphology in yeast...
13
Nuclear expression of a cytoplasmic male sterility gene modifies mitochondrial morphology in yeast and plant cells Yann Duroc 1 , Catherine Gaillard 1 , Sophie Hiard, Corinne Tinchant, Richard Berthome ´, Georges Pelletier, Franc ¸oise Budar * Station de Ge ´ne ´tique et d’Ame ´lioration des Plantes, Institut Jean-Pierre Bourgin, INRA, Route de Saint-Cyr, 78026 Versailles cedex, France Received 22 August 2005; received in revised form 14 November 2005; accepted 14 November 2005 Available online 5 December 2005 Abstract ORF138 is the mitochondrion-encoded protein responsible for Ogura cytoplasmic male sterility (cms) in radish (Raphanus sativus) and Brassica sp. As a means of developing technical tools for analyzing the mechanism of cms in relation to the structure of the sterility protein, we generated gene constructs for ORF138, the green fluorescent protein GFP and an ORF138-GFP fusion, to give nuclear expression with mitochondrial targeting of the encoded proteins. These genes were introduced into the yeast Saccharomyces cerevisiae and were transiently and stably expressed in plant cells. The targeting of nuclear-encoded ORF138 protein to the mitochondria did not prevent the growth of yeast cells on fermentable or non-fermentable media, but modified the cytological appearance of mitochondria in both yeast and plant cells. However, the production from a nuclear gene and mitochondrial targeting of ORF138 did not induce male sterility in transgenic A. thaliana plants. This may be due to the submitochondrial location of nuclearly expressed ORF138. # 2005 Elsevier Ireland Ltd. All rights reserved. Keywords: Cytoplasmic male sterility; GFP fusion; Mitochondrial targeting; Mitochondrial morphology; Saccharomyces cerevisiae; Submitochondrial localization 1. Introduction Cytoplasmic male sterility (cms) is a phenomenon specific to plant reproduction that has been extensively exploited for hybrid seed production. It can occur in natural populations and reflects a conflict between the maternally inherited mitochon- drial genome, which induces male sterility without affecting the overall phenotype of the plant, and the nuclear genome, which restores male fertility [1–3]. In most studied cms systems, a specific mitochondrial gene, constitutively expressed, causes pollen abortion [4]. There are probably few mechanisms by which a constitutively expressed mitochondrial gene could impair male reproductive function without affecting vegetative development or female gameto- genesis. Thus, unraveling the molecular mechanism of a natural cms system should provide insight into the role played by the mitochondria in male gametogenesis in plants. However, before this can be achieved, we need to improve our knowledge of the structure of the cms-associated protein, its interaction with mitochondrial membranes and the structure-function relationship of this protein. A number of mitochondrial genes have been shown to be associated with cms in the last 15 years, but very few of the encoded proteins have been studied experimentally [4–6]. There is no evidence to suggest that the physiological mechanism of cms is similar in all systems. Mitochondrial genes associated with cms generally display no similarity, but similarities have been found between the pol and nap cms- associated open reading frames (orf) in Brassica [7] and between the cms-associated orf79 in rice and orf107 in Sorghum [8]. However, cms-associated proteins have been shown to have similar features. They are generally small (less than 30 kDa) and contain at least one hydrophobic domain. This may be responsible for the association of these proteins with mitochondrial membranes, as has been observed in the rare studies carried out on these proteins. Structure-function relationships for a given protein can be studied by investigating the differences in function of known variants. For cms, we could produce sterile plants by genetic www.elsevier.com/locate/plantsci Plant Science 170 (2006) 755–767 * Corresponding author. Tel.: +33 1 30 83 31 80; fax: +33 1 30 83 33 19. E-mail address: [email protected] (F. Budar). 1 These authors contributed equally to this work. 0168-9452/$ – see front matter # 2005 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.plantsci.2005.11.008
Transcript of Nuclear expression of a cytoplasmic male sterility gene modifies mitochondrial morphology in yeast...

Nuclear expression of a cytoplasmic male sterility gene modifies
mitochondrial morphology in yeast and plant cells
Yann Duroc 1, Catherine Gaillard 1, Sophie Hiard, Corinne Tinchant,Richard Berthome, Georges Pelletier, Francoise Budar *
Station de Genetique et d’Amelioration des Plantes, Institut Jean-Pierre Bourgin, INRA, Route de Saint-Cyr, 78026 Versailles cedex, France
Received 22 August 2005; received in revised form 14 November 2005; accepted 14 November 2005
Available online 5 December 2005
Abstract
ORF138 is the mitochondrion-encoded protein responsible for Ogura cytoplasmic male sterility (cms) in radish (Raphanus sativus) and
Brassica sp. As a means of developing technical tools for analyzing the mechanism of cms in relation to the structure of the sterility protein, we
generated gene constructs for ORF138, the green fluorescent protein GFP and an ORF138-GFP fusion, to give nuclear expression with
mitochondrial targeting of the encoded proteins. These genes were introduced into the yeast Saccharomyces cerevisiae and were transiently and
stably expressed in plant cells. The targeting of nuclear-encoded ORF138 protein to the mitochondria did not prevent the growth of yeast cells on
fermentable or non-fermentable media, but modified the cytological appearance of mitochondria in both yeast and plant cells. However, the
production from a nuclear gene and mitochondrial targeting of ORF138 did not induce male sterility in transgenic A. thaliana plants. This may be
due to the submitochondrial location of nuclearly expressed ORF138.
# 2005 Elsevier Ireland Ltd. All rights reserved.
Keywords: Cytoplasmic male sterility; GFP fusion; Mitochondrial targeting; Mitochondrial morphology; Saccharomyces cerevisiae; Submitochondrial localization
www.elsevier.com/locate/plantsci
Plant Science 170 (2006) 755–767
1. Introduction
Cytoplasmic male sterility (cms) is a phenomenon specific
to plant reproduction that has been extensively exploited for
hybrid seed production. It can occur in natural populations and
reflects a conflict between the maternally inherited mitochon-
drial genome, which induces male sterility without affecting
the overall phenotype of the plant, and the nuclear genome,
which restores male fertility [1–3]. In most studied cms
systems, a specific mitochondrial gene, constitutively
expressed, causes pollen abortion [4]. There are probably
few mechanisms by which a constitutively expressed
mitochondrial gene could impair male reproductive function
without affecting vegetative development or female gameto-
genesis. Thus, unraveling the molecular mechanism of a
natural cms system should provide insight into the role played
by the mitochondria in male gametogenesis in plants.
* Corresponding author. Tel.: +33 1 30 83 31 80; fax: +33 1 30 83 33 19.
E-mail address: [email protected] (F. Budar).1 These authors contributed equally to this work.
0168-9452/$ – see front matter # 2005 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.plantsci.2005.11.008
However, before this can be achieved, we need to improve
our knowledge of the structure of the cms-associated protein,
its interaction with mitochondrial membranes and the
structure-function relationship of this protein.
A number of mitochondrial genes have been shown to be
associated with cms in the last 15 years, but very few of the
encoded proteins have been studied experimentally [4–6].
There is no evidence to suggest that the physiological
mechanism of cms is similar in all systems. Mitochondrial
genes associated with cms generally display no similarity, but
similarities have been found between the pol and nap cms-
associated open reading frames (orf) in Brassica [7] and
between the cms-associated orf79 in rice and orf107 in
Sorghum [8]. However, cms-associated proteins have been
shown to have similar features. They are generally small (less
than 30 kDa) and contain at least one hydrophobic domain.
This may be responsible for the association of these proteins
with mitochondrial membranes, as has been observed in the
rare studies carried out on these proteins.
Structure-function relationships for a given protein can be
studied by investigating the differences in function of known
variants. For cms, we could produce sterile plants by genetic

Y. Duroc et al. / Plant Science 170 (2006) 755–767756
transformation and study the activity of cms proteins in these
plants. This strategy provides an opportunity to test directed
changes in protein structure directly in a plant model. As
mitochondrial transformation is not technically feasible in
plants, such a strategy would require the production of nuclear
transformants with genes encoding cms proteins fused to
mitochondrial targeting signals. However, the only reported
example of the successful production of male sterile plants by
nuclear expression of a cms gene concerns the orf239 gene
from bean (Phaseolus vulgaris) [9]. Similar attempts with
Texas-maize and Petunia cms-associated proteins did not give
rise to sterile plants [10–12].
Alternatively, the biochemical properties of the cms protein
could also be studied in a more easy to handle, albeit
heterologous, system. An attractive model species for this
strategy is the yeast Saccharomyces cerevisiae, which has been
an exceptional model system for mitochondrial function for
many years. Our extensive knowledge of the biogenesis and
function of mitochondria in yeast, and the possibility of studying
cells with impaired mitochondrial function make this model
very attractive for explorations of the possible effects of a cms
protein on normal mitochondrial functioning. The Texas maize
T-URF13 protein has been successfully produced and targeted to
mitochondria in yeast, and the uncoupling induced by its
interaction with methomyl and fungal toxins was then analyzed
[13–15].
A mitochondrial gene, orf138, encoding a protein associated
with mitochondrial inner membranes [16,17] has been shown to
be responsible for Ogura cms in Brassica and Raphanus species
[18,19]. It is present in natural populations of wild radish
(Raphanus raphanistrum) in Asia and Europe [20]. As in most
cms systems studied, the protein thought to be responsible for
the sterile phenotype is present in all organs of sterile plants,
including hypocotyls, leaves, roots and buds [16], but no
morphological or respiratory defect is detected in the vegetative
tissues of these plants [21]. This feature is common to several
cms systems and accounts for the difficulties encountered in
studies of the mechanism of cms [6].
We constructed genes for the production of ORF138, GFP
and an ORF138-GFP fusion from the nucleus and their
targeting to mitochondria. We present here the results obtained
following the introduction of these genes into yeast and plant
cells. These results concern the phenotypes induced by the
ORF138 and ORF138-GFP proteins, when targeted to
mitochondria, particularly concerning mitochondrial morphol-
ogy. The submitochondrial distribution of the produced
proteins may account for these phenotypes.
2. Material and methods
2.1. S. cerevisiae strains
We used S. cerevisae strain BY384 (Mata; his3D25;
leu2D1; lys2D2102; trp1D63; ura3-52), provided by J. Boeke.
It was grown in synthetic complete (SC) medium without uracil
and methionine. We added 100 mM methionine to repress the
expression of genes located downstream from the Met25
promoter. The carbon source in the medium was 2% glucose,
2% galactose, 2% raffinose, or 3% glycerol.
2.2. Synthesis of a nuclear-like form of the orf138 gene
(orf138n) and construction of fusion genes
The orf138n coding sequence was synthesized by PCR,
using overlapping long primers in four successive amplification
reactions. The final PCR product was cloned into pBluescript
and sequenced. Undesirable point mutations introduced during
synthesis were corrected by site-directed mutagenesis. The
resulting gene encodes the ORF138 protein (Fig. 1A).
The orf138n coding sequencewas fused to the mitochondrial
targeting sequence of the b-subunit of the ATP synthase of
Nicotiana plumbaginifolia (preb), kindly provided by Marc
Boutry [22].
A chimeric gene consisting of the preb and orf138n coding
sequences followed by the S65T mutated gfp [23,24] coding
sequencewas also constructed. A control fusion was constructed
from the preb targeting sequence and GFP (S65T). Schematic
representations of the fusion proteins are shown in Fig. 1B.
2.3. Introduction of the gene constructs into yeast
expression vectors
The preb-gfp and preb-orf138n-gfp genes were inserted
between the EcoRI and EcoRV sites and the preb-orf138n gene
was inserted between the HindIII and SalI sites of pRS426Met.
The resulting plasmids were named Met-preb-gfp, Met-preb-
orf138n-gfp and Met-preb-orf138n, respectively. The
pRS426Met vector [25] is essentially pRS426 [26] carrying
an expression cassette with the Met25 promoter and the PGK
terminator.
2.4. Yeast transformation and determination of plating
efficiency
We used 1.5 mg of DNA corresponding to plasmids Met-
preb-gfp, Met-preb-orf138n, Met-preb-orf138n-gfp and
pRS426Met to transform strain BY384, as previously described
[27]. For each transformation, we plated 250 mL of the mixture
on SC (-URA) (2%, w/v, glucose) medium supplemented with
methionine. After 2 days of incubation at 30 8C, the plates werereplicated on SC (-URA) medium containing 3% (v/v) glycerol
or 2% (w/v) glucose, with or without methionine.
2.5. Extraction of total yeast proteins
Yeast cells carrying Met-preb-gfp, Met-preb-orf138n and
Met-preb-orf138n-gfp were cultured at 30 8C in 25 mL of SC
medium (2%, w/v, glucose) with or without 100 mM of
methionine until an OD600 of about 1.5 was reached. To ensure
that equivalent numbers of cells were used, we used a volume
corresponding to an OD600 of 15 (about 10 mL of culture),
which was centrifuged for 5 min at 3000 � g. The pellet was
resuspended in 200 mL of Laemmli loading buffer, vortexed for
5 min in the presence of 0.45 mm glass beads and boiled for

Y. Duroc et al. / Plant Science 170 (2006) 755–767 757
Fig. 1. Construction of the genes used for the transformation of yeast and plant cells. (A) Sequence alignment of mitochondrial (wild type) and ‘‘nuclear’’ (synthetic)
orf138 genes. orf138 is the original mitochondrial gene, and orf138n has been modified to resemble a nuclear gene, as described in Section 2, to optimize nuclear
expression. Substituted nucleotides are indicated in bold and the putative polyadenylation signal (AATAAA) is underlined. The peptide sequence of ORF138 is given
below in the one-letter code. (B) Schematic representation of the constructs designed for nuclear expression in S. cerevisiae and plant cells. The orf138n coding
sequence is represented by hatched bars, the gene encoding GFP(S65T) is represented by white rectangles and the preb mitochondrial targeting sequence is
represented by black rectangles. The names of the genes are indicated on the left: preb-gfp encodes preb-GFP(S65T), preb-orf138n-gfp encodes the preb-ORF138-
GFP fusion protein and preb-orf138n encodes preb-ORF138.
5 min. Themixturewas centrifuged for 5 mn at 10,000 � g, and
an aliquot of the supernatant was used for Western-blot
analysis.
2.6. Yeast mitochondrion isolation and fractionation
Yeast mitochondria were isolated from 4 L of yeast culture
in SC medium containing 2% (w/v) raffinose, as described by
Guerin et al. [28].
Mitochondrial membranes were fractionated as follows.
Mitochondria were resuspended in 250 mM sucrose, 1 mM
EDTA, 1 mM EGTA, 10 mM HEPES pH 7.2. Low-osmotic
strength buffer (10 mM HEPES, 1 mM EDTA, 1 mM EGTA,
pH 7.2) was added until a sucrose concentration of 10 mM was
obtained. The suspension was then shaken gently for 20 min at
4 8C, after which the concentration of sucrose was adjusted to
250 mM by adding high-strength osmotic buffer (2 M sucrose,
10 mM HEPES, 1 mM EDTA, 1 mM EGTA, pH 7.2).
Mitochondria were then shaken gently for a further 20 min
at 4 8C. The sample was loaded onto a stepped sucrose gradient
(3 mL 60%, 7 mL 35%, 4 mL 25% sucrose in 10 mM HEPES
1 mM EDTA, 1 mM EGTA, pH 7.2) and centrifuged at

Y. Duroc et al. / Plant Science 170 (2006) 755–767758
150,000 � g for 1 h at 4 8C in a swingout rotor. Outer
membrane-enriched vesicles were collected from the 25/35
interface; the mitoplast-enriched fraction was collected from
the 35/60 interface. The two fractions were diluted in low-
osmotic strength buffer and the membranes were concentrated
by centrifugation at 100,000 � g for 1 h at 4 8C.Isolated mitochondria were treated with carbonate to assess
the strength of the association between ORF138 and the
membrane [29]. Samples were incubated for 30 min on ice in
100 mM Na2CO3 pH 11.5 and centrifuged at 4 8C for 30 min at
100,000 � g. The supernatant (soluble fraction) was collected
and the pellet (membrane fraction) was resuspended in 100 mM
Tris–HCl pH 7.5.
2.7. Introduction of the gene constructs into plant
expression vectors
The preb-orf138n, preb-gfp, and preb-orf138n-gfp genes
were inserted into the single HindIII site of an expression
cassette containing the CaMV 35S promoter (with a duplicated
enhancer sequence) and terminator (T. Michael, unpublished
personal communication) in a derivative of pBluescript
(Stratagene). We checked that the orientation of the inserts
was correct by restriction analysis. We used these plasmids for
transient expression in onion and N. benthamiana cells.
The promoter of the A9 gene of Arabidopsis thaliana [30]
was amplified by PCR from the Landsberg erecta ecotype,
using the following primers:
Forward primer: ATCTAGACATAACGGTGAGAGT-
TAATA
Reverse primer: TGGTACCTCTAATTAGATACTATATTG
The XbaI and KpnI sites used for cloning are underlined. The
XbaI site is present in the genomic sequence, the KpnI site was
introduced to facilitate subsequent manipulations. After
sequence verification, an expression cassette was constructed
with the A9 promoter and the CaMV 35S terminator, flanked by
two NotI restriction sites. The preb-orf138n, preb-gfp, and
preb-orf138n-gfp genes were inserted into the HindIII cloning
site between the promoter and terminator. We checked that the
orientation of the inserts was correct by restriction analysis.
For the stable transformation of A. thaliana, NotI restriction
fragments containing the genes of interest were inserted into the
unique NotI site of the pEC2 binary vector [31]. The resulting
plasmids were verified by restriction analysis and used to
transform an Agrobacterium tumefaciens strain (C58C1)
carrying the pMP90 helper plasmid [32].
2.8. Transient and stable transformation of plant cells
Tungsten particles (diameter 0.5 mm) were sterilized by
incubation in 70% ethanol for 10 min, rinsed with 1.5 mL
sterile water and kept on ice in 50% glycerol until use. We
mixed 100 mL of these particles (15–20 mg) with 5–10 mg of
plasmid DNA (for 3–6 shots). Spermidine (40 mL of 0.1 M
solution) and CaCl2 (100 mL of 2.5 M solution) were then
added and the mixture kept on ice for 10 min, with vortexing
every 3 min. The particles were centrifuged for 30 s in a
benchtop centrifuge. The supernatant was discarded and the
particles were rinsed twice in cold ethanol and resuspended in
60 mL of 100% ethanol. The DNA-coated particles were then
transferred to macrocarrier disks.
Onion slices or Nicotiana benthamiana leaves were placed
on solid medium [33] in Petri dishes and bombarded with the
DNA-coated particles using a Biorad PDS-1000/He Biolistic
Particle Delivery system (900 psi rupture disks) according to
the manufacturer’s instructions.
Explants were then incubated for 48–72 h in a culture
chamber and analyzed by confocal microscopy.
Stable A. thaliana transformants (ecotype Wassilewskija)
carrying the verified constructs were generated using Agro-
bacterium strain C58C1 (pMP90), as previously described [34].
2.9. Extraction of A. thaliana mitochondrial proteins
We grew the F2 progeny of several transformants carrying
the preb-orf138n gene under the control of the 35S promoter on
soil in the greenhouse for 5 weeks. We ground about 20–50
plants (leaves and stems) together in extraction buffer (0.3 M
sucrose, 25 mM tetrasodium pyrophosphate, 2 mM EDTA,
10 mM KH2PO4, 1% (w/v) PVP-40, 1% (w/v) BSA, 20 mM
ascorbate, pH 7.5). The homogenate was then filtered through
four layers of Miracloth (Calbiochem) and subjected to three
cycles of differential centrifugation as previously described
[35]. The organelle-enriched pellet was resuspended in the
minimum volume of washing buffer (0.3 M sucrose, 10 mM
HEPES, 0.2% (w/v) BSA, pH 7.5). Vesicles were ruptured by
five cycles of freezing and thawing involving successive
immersions in liquid nitrogen and incubations at room
temperature. The insoluble fraction was collected by centri-
fugation at 20,000 � g for 30 min at 4 8C and resuspended in a
minimum volume of washing buffer.
2.10. Affinity purification of antibodies
Mitochondria were prepared from male-sterile Brassica
napus plants as previously described [36]. Purified mitochon-
dria obtained from 10 g of flower buds from greenhouse-grown
plants were lysed in 62.5 mM Tris–HCl (pH 6.8) supplemented
with 2% (w/v) SDS, 5% (v/v) b-mercaptoethanol, and 10% (v/
v) glycerol. Proteins were precipitated in acetone, collected by
centrifugation and resuspended in 50 mM Tris–HCl (pH 7.5).
Rabbit anti-ORF138 IgG was affinity-purified using a
modified version of a previously described procedure [37].
Total mitochondrial protein extracts from B. napus sterile
plants were spotted onto a nitrocellulose membrane (HybondC,
Amersham). The membrane was incubated for 1 h at 37 8Cwith
50 mM Tris–HCl (pH 7.5) supplemented with 150 mM NaCl,
0.05% (v/v) Tween 20 and 3% (w/v) nonfat milk powder (buffer
A). The membrane was then incubated with crude rabbit anti-
ORF138 immune serum [16] (1 mL/cycle) for 1 h at 37 8C. Themembrane was washed four times at 37 8C for 1 min each with
buffer B (buffer B = buffer A minus milk powder). The bound

Y. Duroc et al. / Plant Science 170 (2006) 755–767 759
antibodies were eluted with threewashes (1 min each) in 0.5 mL
5 mM glycine–HCl (pH 3) containing 500 mMNaCl, 0.25% (v/
v) Tween 20 and 1% (w/v) BSA. Eluates were pooled and
neutralized with solid Tris. The membrane was washed three
times, in 3 mL of buffer B.We then carried out six further cycles
of incubation with the ORF138 antiserum, followed by washing
and elution. Affinity-purified IgG was divided into 0.25 mL
aliquots, frozen in liquid nitrogen and stored at �80 8C.
2.11. SDS–PAGE and immunoblot analysis of protein
extracts
An aliquot of each culture or fraction was processed by
SDS–PAGE. The concentration of acrylamide used for PAGE is
given in the legend to each figure. The proteins were
electroblotted onto nitrocellulose membranes (HybondC,
Amersham) or PVDF (Immobilon, Millipore). Immunoblotting
was carried out with the following antibodies: unpurified
(diluted 1:5000; plant extracts) or purified (diluted 1:200; yeast
extracts) rabbit ORF138 antisera; a mixture of two monoclonal
antibodies against GFP (Boehringer; diluted 1:1000); a
polyclonal rabbit antiserum against ferrochelatase (Hem15;
diluted 1:1000); a monoclonal mouse antibody against porin
(Molecular Probes; diluted 1:500). The ORF138 and ferroche-
latase antibodies were then detected with peroxidase-con-
jugated anti-rabbit IgG (Sigma). The GFP and porin antibodies
were detected with peroxidase-conjugated secondary anti-
mouse IgG (Caltag). Secondary antibodies were detected with
the ECL kit (Amersham). The apparent molecular weights of
the proteins were estimated with prestained low-molecular
weight or broad-range molecular weight markers (Biorad) or
with a prestained MW marker (Fermentas).
2.12. Cytological observations
Yeasts carrying Met-preb-gfp, and Met-preb-orf138n-gfp
were cultured for 2 days at 30 8C in 2 mL of SC medium
(containing 2% galactose) with or without 100 mMmethionine.
Cells were centrifuged for 5 min at 4000 � g, and resuspended
in 20 mL of culture medium. An aliquot (3 mL) of this cell
suspension was then immobilized in 3 mL of 0.8% low-melting
point agarose at 36 8C on glass slides.
Onion slices were peeled and epiderm fragments mounted
on microscope slides. N. benthamiana leaf fragments were
mounted directly on microscope slides.
Cells producing GFP were observed under a Leica TCS-NT
confocal laser scanning microscope (Leica Microsystems,
Heidelberg, Germany) with an argon/krypton laser (Omni-
chrome, Chino, CA). GFP fluorescence was observed using an
FITC filter set (BP530/30) with an argon (488 nm) laser.
Medium-scan (450 lines per second) images (1024 � 1024
pixels) were taken using a 63 � 1.32 NA PL APO objective for
yeasts, and a 40 � 0.8 w HCX APO L objective for plant cells.
In some experiments, a sequential filter set with a second
photomultiplier was used to collect the transmitted light. In
these cases, the light emitted by GFP was collected using the
configuration described above.
3. Results
3.1. Construction of genes for production and targeting to
mitochondria of ORF138, GFP and ORF138-GFP proteins
The orf138 coding sequence has a high A + T content (65%)
and carries putative internal polyadenylation site (AATAAA).
We used PCR to synthesize a version of this gene (orf138n)
encoding exactly the same peptide, but with an A + T content of
only 50% and no internal polyadenylation signal, mimicking a
nuclear gene (Fig. 1A). We constructed a chimeric gene in
which the orf138n and gfp coding sequences were fused. The
proteins were targeted to the mitochondria by fusing their
coding sequences to the mitochondrial presequence of the N.
plumbaginifolia b-ATPase gene (preb). Duby et al. [38]
precisely dissected the sequence necessary for efficient import
of GFP into plant and yeast cells and showed that the first 47
aminoacids of the signal peptide were sufficient for efficient
mitochondrial import. The same group previously showed that
a presequence extending to the 59th residue was also efficient
for mitochondrial targeting of fusion proteins [11]. The
presequence of b-ATPase from N. plumbaginifolia was also
used for the import of GFP into mitochondria of stable
Arabidopsis transformants [39]. The presequence used in this
study encodes the first 59 amino acids of the b-subunit
precursor and includes the cleavage site. We constructed three
genes encoding fusion proteins (Fig. 1B). These constructs
were designed to ensure targeting of the ORF138, GFP and
ORF138-GFP proteins to the mitochondria.
3.2. Production of mitochondrially targeted GFP, ORF138,
and ORF138-GFP in yeast
We used the yeast S. cerevisiae as a heterologous
eukaryotic model, to investigate whether ORF138 had a
general effect on mitochondria. As we did not know whether
ORF138 would impair yeast growth, we introduced the preb-
orf138n, preb-gfp and preb-orf138n-gfp genes downstream
from the Met25 promoter, which is repressed in the presence
of methionine. The plasmids carrying the preb-gfp, preb-
orf138n and preb-orf138n-gfp genes were introduced into
yeast strain BY384, as was the insert-less vector, pRS426Met.
Transformants were selected on glucose-containing selective
(synthetic complete, URA minus) medium and replicated on
glycerol-containing selective medium. Plating efficiency and
colony size were similar in strains transformed with the three
constructs and in strains transformed with the insert-less
vector. We repeated the experiment, adding 100 mM of
methionine to the glucose-containing medium used for the
selection of transformants, to prevent selection against cells
expressing the genes. Plates were then replicated in parallel,
on glucose- or glycerol-containing selective media, with or
without methionine. In these conditions, no significant
difference in growth or colony size was detected for any of
the constructs.
We checked that the various proteins were produced in vivo,
by extracting total proteins from several transformants,

Y. Duroc et al. / Plant Science 170 (2006) 755–767760
Fig. 2. Immunodetection of the preb-ORF138, preb-GFP, and preb-ORF138-GFP fusion proteins in S. cerevisiae. Extracts equivalent to 1.1 OD units of culture were
run on a 10% acrylamide gel. Lanes 1, 3, and 5 contain protein extracts from repressed cells whereas lanes 2, 4 and 6 contain protein extracts from non- repressed cells.
Lanes 1 and 2: extracts from preb-gfp harboring cells. Lanes 3 and 4: extracts from preb-orf138n harboring cells. Lanes 5 and 6: extracts from preb-orf138n-gfp
harboring cells. Numbers on the left indicate sizes of molecular weight markers (kDa). Two identical gels were processed in parallel and the corresponding
membranes hybridized with antibodies directed against GFP (A) or ORF138 (B).
carrying out SDS–PAGE and immunoblotting with antibodies
against ORF138 and/or GFP. Yeast strains carrying the various
constructs were found to produce the fusion proteins at
detectable levels (Fig. 2A and B). These antibodies detected
smaller amounts of ORF138-GFP fusion protein than of GFP
and ORF138.
3.2.1. Production and mitochondrial targeting of ORF138-
GFP modifies mitochondrial morphology
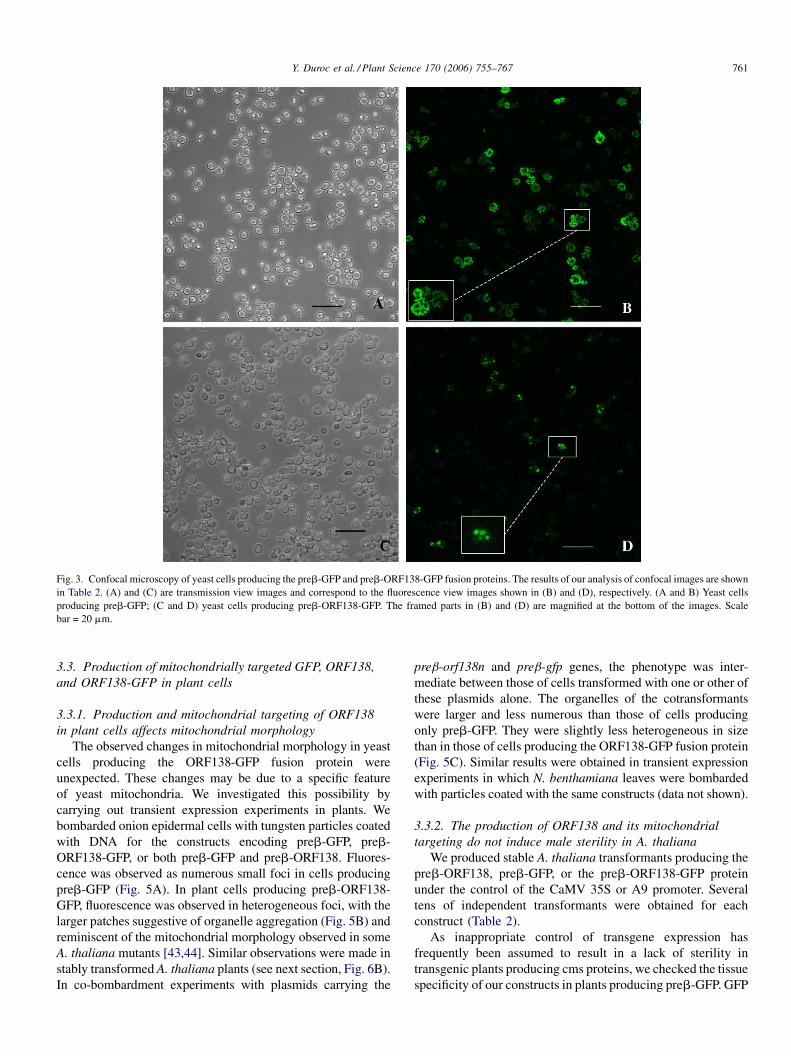
We observed yeast cells producing mitochondrion-targeted
GFP and ORF138–GFP fusion proteins by fluorescence
microscopy, after the cells had been cultured in galactose
synthetic complete (SC) medium without methionine (Fig. 3).
Yeast cells producing preb-GFP displayed numerous fluor-
escent organelles, associated in tubular branched networks, as
previously described [40,41]. In contrast, cells producing preb-
ORF138-GFP showed fluorescence concentrated in globular
organelles, reminiscent of some yeast mdm mutants [41] or of
rapamycin-treated cells displaying autophagy of mitochondria
[42]. Light and fluorescence micrographs were used to count
the number of GFP-producing cells. We calculated the
percentage of GFP-containing cells that had globular mito-
chondria. We counted GFP-containing cells on six fields for
preb-GFP and ten fields for preb-ORF138-GFP (Table 1).
Significantly, fewer fluorescent cells were detected for preb-
ORF138-GFP than for preb-GFP. The normal mitochondrial
tubular network was observed in a very small percentage of
Table 1
Percentage of yeast cells presenting a globular mitochondrial phenotype
following the production of preb-ORF138-GFP or preb-GFP fusion proteins
Construct name % GFP-expressing cells % globular
Preb-GFP 15 � 4 10 � 3
Preb-ORF138-GFP 6 � 2 85 � 15
We counted fluorescent cells on 6 or 10 independent images obtained by light
and fluorescence microscopy of the same field, and estimated the % of cells
expressing GFP. The number of cells with globular organelles was estimated as
a percentage of GFP-expressing cells. Examples of the images analyzed are
given in Fig. 3.
fluorescent cells producing preb-ORF138-GFP, mostly being
replaced by globular organelles.
3.2.2. Submitochondrial distribution of the ORF138 and
ORF138-GFP proteins in yeast
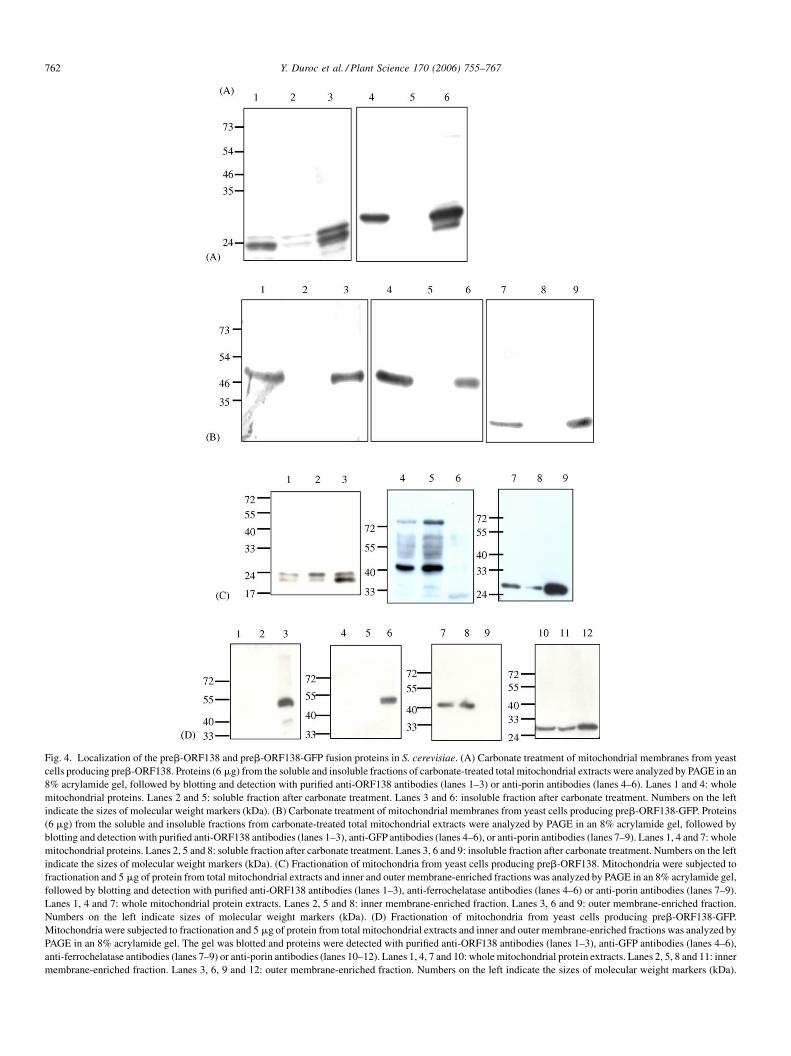
Mitochondria were purified from yeast strains producing
either preb-ORF138 or preb-ORF138-GFP. Following alka-
line carbonate treatment and centrifugation, both proteins
were found to be intrinsically associated with mitochondrial
membranes (Fig. 4A and B), like ORF138 in sterile plants
[16]. The ORF138 signal detected in immunoblots performed
with preb-ORF138-producing cells appears has a doublet
(Fig. 4A and C). We sometimes observe doublet bands for
ORF138 in immunoblots from other cell types, including
plant cells (data not shown). We interpret this observation has
a partial proteolysis of the protein in some extracts. In
Fig. 4A, the porin also appears as a doublet in the insoluble
fraction.
We fractionated mitochondrial extracts into outer mem-
brane- and inner membrane-enriched fractions, to determine
the precise location of the fusion proteins. We assessed the
efficiency of fraction enrichment by immunolocalization of
ferrochelatase (an inner membrane protein) or porin (an outer
membrane protein). In preb-ORF138-producing cells, the
ORF138 protein was shown to be present in both the outer and
inner membranes (Fig. 4C). Similar levels of ORF138 were
detected in both fractions, strongly suggesting that the protein
was associated with both mitochondrial membranes in these
cells. This pattern differs from that observed in the
mitochondria of sterile plants, in which ORF138 is associated
solely with the inner membrane [17]. The difference in relative
abundance of the upper and lower signals of the doublet in
inner and outer membrane fraction (Fig. 4C) might be due to
different abundance or efficiency of proteases in the respective
fractions. In preb-ORF138-GFP-producing yeast cells, the
ORF138 protein was found exclusively in the outer membrane
fraction (Fig. 4D). These results were confirmed by
immunodetection of the GFP moiety of the fusion protein
(Fig. 4D).
Y. Duroc et al. / Plant Science 170 (2006) 755–767 761
Fig. 3. Confocal microscopy of yeast cells producing the preb-GFP and preb-ORF138-GFP fusion proteins. The results of our analysis of confocal images are shown
in Table 2. (A) and (C) are transmission view images and correspond to the fluorescence view images shown in (B) and (D), respectively. (A and B) Yeast cells
producing preb-GFP; (C and D) yeast cells producing preb-ORF138-GFP. The framed parts in (B) and (D) are magnified at the bottom of the images. Scale
bar = 20 mm.
3.3. Production of mitochondrially targeted GFP, ORF138,
and ORF138-GFP in plant cells
3.3.1. Production and mitochondrial targeting of ORF138
in plant cells affects mitochondrial morphology
The observed changes in mitochondrial morphology in yeast
cells producing the ORF138-GFP fusion protein were
unexpected. These changes may be due to a specific feature
of yeast mitochondria. We investigated this possibility by
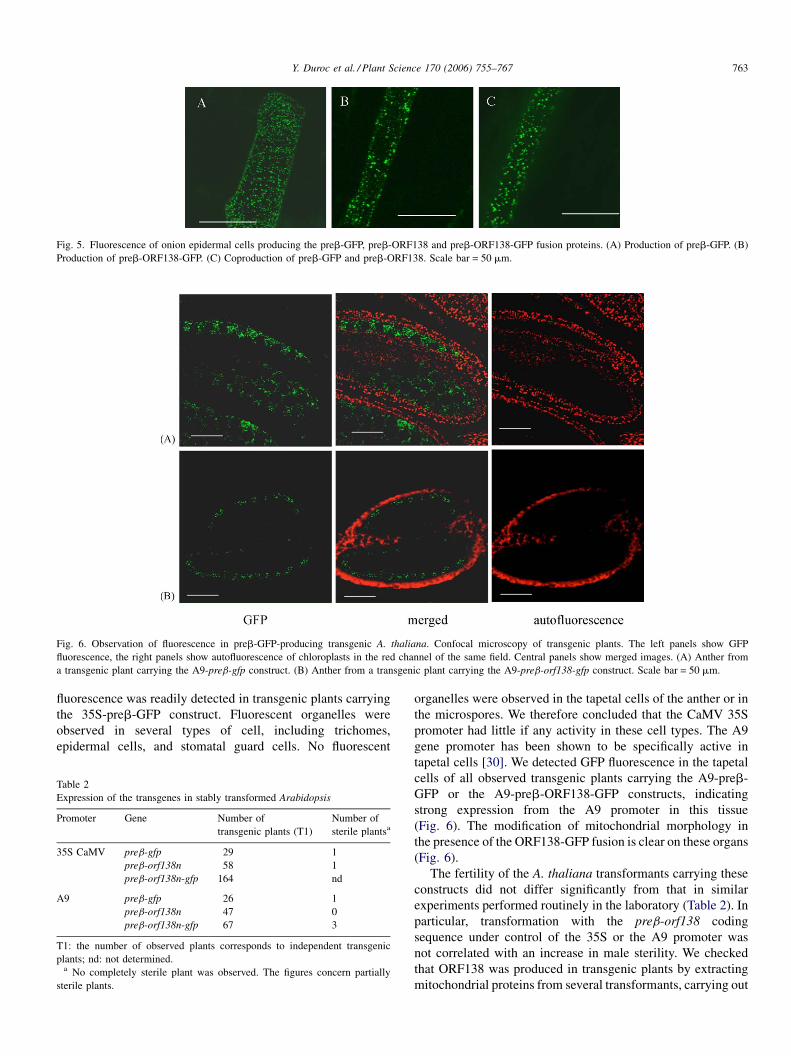
carrying out transient expression experiments in plants. We
bombarded onion epidermal cells with tungsten particles coated
with DNA for the constructs encoding preb-GFP, preb-
ORF138-GFP, or both preb-GFP and preb-ORF138. Fluores-
cence was observed as numerous small foci in cells producing
preb-GFP (Fig. 5A). In plant cells producing preb-ORF138-
GFP, fluorescence was observed in heterogeneous foci, with the
larger patches suggestive of organelle aggregation (Fig. 5B) and
reminiscent of the mitochondrial morphology observed in some
A. thaliana mutants [43,44]. Similar observations were made in
stably transformed A. thaliana plants (see next section, Fig. 6B).
In co-bombardment experiments with plasmids carrying the
preb-orf138n and preb-gfp genes, the phenotype was inter-
mediate between those of cells transformed with one or other of
these plasmids alone. The organelles of the cotransformants
were larger and less numerous than those of cells producing
only preb-GFP. They were slightly less heterogeneous in size
than in those of cells producing the ORF138-GFP fusion protein
(Fig. 5C). Similar results were obtained in transient expression
experiments in which N. benthamiana leaves were bombarded
with particles coated with the same constructs (data not shown).
3.3.2. The production of ORF138 and its mitochondrial
targeting do not induce male sterility in A. thaliana
We produced stable A. thaliana transformants producing the
preb-ORF138, preb-GFP, or the preb-ORF138-GFP protein
under the control of the CaMV 35S or A9 promoter. Several
tens of independent transformants were obtained for each
construct (Table 2).
As inappropriate control of transgene expression has
frequently been assumed to result in a lack of sterility in
transgenic plants producing cms proteins, we checked the tissue
specificity of our constructs in plants producing preb-GFP. GFP
Y. Duroc et al. / Plant Science 170 (2006) 755–767762
Fig. 4. Localization of the preb-ORF138 and preb-ORF138-GFP fusion proteins in S. cerevisiae. (A) Carbonate treatment of mitochondrial membranes from yeast
cells producing preb-ORF138. Proteins (6 mg) from the soluble and insoluble fractions of carbonate-treated total mitochondrial extracts were analyzed by PAGE in an
8% acrylamide gel, followed by blotting and detection with purified anti-ORF138 antibodies (lanes 1–3) or anti-porin antibodies (lanes 4–6). Lanes 1 and 4: whole
mitochondrial proteins. Lanes 2 and 5: soluble fraction after carbonate treatment. Lanes 3 and 6: insoluble fraction after carbonate treatment. Numbers on the left
indicate the sizes of molecular weight markers (kDa). (B) Carbonate treatment of mitochondrial membranes from yeast cells producing preb-ORF138-GFP. Proteins
(6 mg) from the soluble and insoluble fractions from carbonate-treated total mitochondrial extracts were analyzed by PAGE in an 8% acrylamide gel, followed by
blotting and detection with purified anti-ORF138 antibodies (lanes 1–3), anti-GFP antibodies (lanes 4–6), or anti-porin antibodies (lanes 7–9). Lanes 1, 4 and 7: whole
mitochondrial proteins. Lanes 2, 5 and 8: soluble fraction after carbonate treatment. Lanes 3, 6 and 9: insoluble fraction after carbonate treatment. Numbers on the left
indicate the sizes of molecular weight markers (kDa). (C) Fractionation of mitochondria from yeast cells producing preb-ORF138. Mitochondria were subjected to
fractionation and 5 mg of protein from total mitochondrial extracts and inner and outer membrane-enriched fractions was analyzed by PAGE in an 8% acrylamide gel,
followed by blotting and detection with purified anti-ORF138 antibodies (lanes 1–3), anti-ferrochelatase antibodies (lanes 4–6) or anti-porin antibodies (lanes 7–9).
Lanes 1, 4 and 7: whole mitochondrial protein extracts. Lanes 2, 5 and 8: inner membrane-enriched fraction. Lanes 3, 6 and 9: outer membrane-enriched fraction.
Numbers on the left indicate sizes of molecular weight markers (kDa). (D) Fractionation of mitochondria from yeast cells producing preb-ORF138-GFP.
Mitochondria were subjected to fractionation and 5 mg of protein from total mitochondrial extracts and inner and outer membrane-enriched fractions was analyzed by
PAGE in an 8% acrylamide gel. The gel was blotted and proteins were detected with purified anti-ORF138 antibodies (lanes 1–3), anti-GFP antibodies (lanes 4–6),
anti-ferrochelatase antibodies (lanes 7–9) or anti-porin antibodies (lanes 10–12). Lanes 1, 4, 7 and 10: wholemitochondrial protein extracts. Lanes 2, 5, 8 and 11: inner
membrane-enriched fraction. Lanes 3, 6, 9 and 12: outer membrane-enriched fraction. Numbers on the left indicate the sizes of molecular weight markers (kDa).
Y. Duroc et al. / Plant Science 170 (2006) 755–767 763
Fig. 5. Fluorescence of onion epidermal cells producing the preb-GFP, preb-ORF138 and preb-ORF138-GFP fusion proteins. (A) Production of preb-GFP. (B)
Production of preb-ORF138-GFP. (C) Coproduction of preb-GFP and preb-ORF138. Scale bar = 50 mm.
Fig. 6. Observation of fluorescence in preb-GFP-producing transgenic A. thaliana. Confocal microscopy of transgenic plants. The left panels show GFP
fluorescence, the right panels show autofluorescence of chloroplasts in the red channel of the same field. Central panels show merged images. (A) Anther from
a transgenic plant carrying the A9-preb-gfp construct. (B) Anther from a transgenic plant carrying the A9-preb-orf138-gfp construct. Scale bar = 50 mm.
fluorescence was readily detected in transgenic plants carrying
the 35S-preb-GFP construct. Fluorescent organelles were
observed in several types of cell, including trichomes,
epidermal cells, and stomatal guard cells. No fluorescent
Table 2
Expression of the transgenes in stably transformed Arabidopsis
Promoter Gene Number of
transgenic plants (T1)
Number of
sterile plantsa
35S CaMV preb-gfp 29 1
preb-orf138n 58 1
preb-orf138n-gfp 164 nd
A9 preb-gfp 26 1
preb-orf138n 47 0
preb-orf138n-gfp 67 3
T1: the number of observed plants corresponds to independent transgenic
plants; nd: not determined.a No completely sterile plant was observed. The figures concern partially
sterile plants.
organelles were observed in the tapetal cells of the anther or in
the microspores. We therefore concluded that the CaMV 35S
promoter had little if any activity in these cell types. The A9
gene promoter has been shown to be specifically active in
tapetal cells [30]. We detected GFP fluorescence in the tapetal
cells of all observed transgenic plants carrying the A9-preb-
GFP or the A9-preb-ORF138-GFP constructs, indicating
strong expression from the A9 promoter in this tissue
(Fig. 6). The modification of mitochondrial morphology in
the presence of the ORF138-GFP fusion is clear on these organs
(Fig. 6).
The fertility of the A. thaliana transformants carrying these
constructs did not differ significantly from that in similar
experiments performed routinely in the laboratory (Table 2). In
particular, transformation with the preb-orf138 coding
sequence under control of the 35S or the A9 promoter was
not correlated with an increase in male sterility. We checked
that ORF138 was produced in transgenic plants by extracting
mitochondrial proteins from several transformants, carrying out

Y. Duroc et al. / Plant Science 170 (2006) 755–767764
Fig. 7. Immunodetection of the ORF138 protein in mitochondrial extracts of
transgenic A. thaliana. Insoluble fractions of mitochondrion-enriched extracts
(50 mg) of A. thaliana were analyzed by PAGE in a 15% acrylamide gel,
followed by blotting and detection with anti-ORF138 antibodies. (1) Positive
control: mitochondrial proteins from sterile rapeseed (5 mg). (2) Negative
control: non transgenic A. thaliana WS. (3–6) Four independent transgenic
plants carrying the 35S-preb-orf138n. Horizontal arrows on the right indicate
the position of specific signals from monomers and dimers of ORF138.
Numbers on the left indicate the sizes of molecular weight markers (kDa).
SDS–PAGE on the insoluble fraction and immunoblotting with
antibodies against ORF138 (Fig. 7). We found that the ORF138
protein was produced and targeted to mitochondria in
transgenic plants, but did not induce male sterility.
4. Discussion
Transgenic A. thaliana plants producing ORF138 targeted to
the mitochondria are not male sterile. This lack of male sterility
following the nuclear expression of a cms gene is shared by
several cms systems [10–12]. Inappropriate timing and/or
spatial control of transgene expression is often put forward as
the main explanation for this lack of sterility. Use of the GFP
reporter gene showed that the 35S promoter was only weakly
active, if at all, in tapetal cells. Similar observations have been
reported in previous studies [45]. The tapetum has been shown
to be the target tissue of the sterility mechanism, as
development is initially impaired in this highly active cell
layer [46]. It is possible that the induction of male sterility by a
nuclear orf138 genewould require specific expression in tapetal
cells. We therefore generated constructs including the tapetum-
specific promoter A9, which has the ideal spatial and temporal
expression profile for this purpose [30]. The GFP reporter gene
was used to check the efficiency of the A9 promoter for driving
tapetum-specific expression of GFP in our experiments.
However, the A9-preb-orf138 construct did not induce male
sterility in the transgenic plants. Thus, a lack of expression of
the cms gene in the correct tissue and at the correct time is not
the primary reason for the lack of male sterility in these
transgenic plants.
Other non-mutually exclusive hypotheses can be proposed
to account for the lack of male sterility in transgenic plants
expressing nuclear copies of cms genes. Too little of the protein
may accumulate in cases in which a quantitative threshold must
be exceeded for activity. Alternatively, inappropriate folding or
submitochondrial location of the cms protein may be
responsible. If the cms gene is expressed in a plant species
different from that in which it originated, the absence of specific
partners of the cms protein necessary for sterility may also be a
problem. This seems unlikely in this case, but the possibility
cannot be formally excluded. The Ogura cms originates from
Raphanus sativus and has been successfully transferred to
Brassica species [47]. As Arabidopsis belongs to the same
family as Raphanus and Brassica, possible partners conserved
in these two genera are probably also present in A. thaliana.
We previously showed that the male sterility induced by
orf138 depends on a threshold gene copy number (and probably
protein production) being exceeded in rapeseed cybrids [48].
The production of too small a quantity of the sterility protein
would therefore probably not result in a sterility phenotype.
However, only quantitative analysis of ORF138 production in
the tapetal cells of transgenic plants would be able to confirm or
reject this hypothesis and such analysis is not technically
feasible.
It is also possible that the import of the cms protein from the
cytosol results in its incorrect submitochondrial targeting. Our
results, obtained in yeast cells producing mitochondrially
targeted GFP, ORF138 and ORF138-GFP proteins, are
consistent with this hypothesis. In previous studies, we have
shown that ORF138 is an intrinsic mitochondrial membrane
protein, associated with the inner mitochondrial membrane of
sterile plants [16,17]. We report here that both ORF138 and
ORF138-GFP are intrinsic mitochondrial membrane proteins
in yeast cells. ORF138 was also associated with the insoluble
fraction of mitochondrial proteins from transgenic A. thaliana
plants. We performed fractionation experiments on yeast
mitochondrial extracts and found that the preb-ORF138
protein was present in both mitochondrial membranes,
whereas the preb-ORF138-GFP fusion protein was detected
only in the outer membrane-enriched fraction. This was
confirmed by immunodetection of theGFPmoiety of the fusion
protein. These results indicate that the proteins produced are
efficiently targeted to mitochondria, with a significant
proportion remaining in the outer mitochondrial membrane.
We therefore conclude that only some of the ORF138 protein
produced in the cytosol and targeted to the mitochondria of
yeast cells ends up in the correct compartment of mitochondria
(i.e. that in which it is found in the mitochondria of sterile
plants). We suggest that the inappropriate distribution in the
mitochondria of the produced ORF138 may contribute to the
absence of a male sterility phenotype in transgenic plants. A
similar explanation may apply to the similar results obtained
with other cms genes. The bean cms-associated protein is
unique in not being located in the mitochondria. It is instead
found in cell walls and in the callose layer of microspore
tetrads; it is also the only cms production for which the
successful induction of male sterility by the production of the
cms protein from a nuclear transgene has been reported [9]. As
some of the ORF138 protein targeted to the mitochondria was
found in the innermembrane, we cannot exclude the possibility
that the absence of phenotype results from too small an amount
of protein being correctly distributed. Nevertheless, we feel
that correct submitochondrial targeting of cms proteins

Y. Duroc et al. / Plant Science 170 (2006) 755–767 765
produced from nuclear transgenes may have been under-
estimated in previous studies.
The use of GFP fusion proteins made it possible to observe
mitochondria by fluorescence microscopy. GFP has proved to
be a powerful tool for monitoring mitochondrial shape in vivo
and changes in mitochondria morphology in various condi-
tions [39–41,43,44,49]. We observed mitochondrial morphol-
ogy in yeast and plant cells producing preb-GFP and preb-
ORF138-GFP. We found that preb-ORF138-GFP had a
dramatic effect on mitochondrial morphology in both cell
types. Yeast mitochondria rapidly change shape during cell
growth and can be modified by mutations affecting the
division or fusion of mitochondria [50] and by mutations
affecting other mitochondrial functions, including respiration
[41]. However, strains producing preb-ORF138 (or preb-
ORF138-GFP) showed no inhibition of respiration, at least in
the conditions tested in this study. The morphological
phenotype observed when ORF138-GFP was targeted to
yeast mitochondria was similar to that of the mutants mdm32
and mdm34 [41]. Interestingly, this phenotype is reminiscent
of autophagy in rapamycin-treated yeast cells [42]. In yeast,
autophagy is induced by starvation conditions and involves
the sequestration of organelles, including mitochondria, in a
lytic vacuole [51–55].
It is difficult to assess the similarity in mitochondrial
morphology between yeast and plant cells producing preb-
ORF138-GFP. In plant cells, cotransformants producing the
preb-ORF138 and preb-GFP proteins also displayed clear
changes in mitochondrial morphology, although these changes
were less marked than with the fusion protein. Thus, the
observed morphology is not exclusively linked to the ORF138-
GFP fusion. It is also seen when the ORF138 protein is present
in the outer membrane of the mitochondria. The presence of
ORF138 in the outer mitochondrial membrane therefore leads
to an abnormal mitochondrial morphology, via a direct
aggregative effect or indirectly via a more complex pathway.
The more marked effect of preb-ORF138-GFP may be due to
aggravation due to the fusion with GFP, or due to all the protein
being present in the outer membrane, whereas the preb-
ORF138 was present in both the inner and outer membranes.
The submitochondrial location of the ORF138 produced
from a nuclear gene in our experiments is not analogous to that
in sterile plants. Any interpretation of the mechanism of male
sterility based on our results would therefore not be valid.
Future studies on cms should take this into account and
experimental systems should be selected with care to ensure
that the topology of the proteins concerned in the mitochondrial
membranes is analogous to that in sterile plants. This could be
achieved, for instance, by generating transgenic yeast cells in
which the orf138 gene has been inserted into the mitochondrial
genome. As the genetic code used by yeast mitochondria is
different from that used by nuclear genes and by plant
mitochondrial genes, this experiment would require the entire
recoding of the orf138 gene.
This study clearly shows that inappropriate control of
expression is not the only reason for the lack of induction of
male sterility by production of mitochondrially targeted cms
proteins in transgenic plants. Very few studies have dealt with the
complementation of mitochondrial mutations by a mitochond-
rially targeted protein encoded by a nuclear gene. This is the case
for var1p and Aap1p (ATP synthase subunit 8) in S. cerevisiae
[56,57], and the product of urfa in Schizosaccharomyces pombe
[58]. Another example concerns a mitochondrial mutation in the
subunit 6 of the human ATPase. This mutation is involved in
Leigh syndrome, and was successfully complemented by a
protein imported into the mitochondria in transfected cells [59].
Recently, the complementation of a complex I N. sylvestris
mitochondrial mutant was realized by targeting to mitochondria
the NAD7 subunit, whose gene was deleted in the mitochondrial
genome of the mutant [60]. To our knowledge, these are the only
published reports of successful experiments of this type. Their
scarcity is remarkable, especially in yeast, for which mitochon-
drial mutants have been known and studied for more than 50
years. The lack of studies reporting the relocation of
mitochondrial genes to the nuclear genome may be due to the
necessity of recoding mitochondrial yeast and human genes into
the universal genetic code to ensure correct translation. This may
also be due to the folding requirements and membrane
interactions of the proteins, which may be incompatible with
their import from the cytosol. Law et al. [57] also reported that
ATP9 produced from a nuclear gene and targeted to
mitochondria was unable to complement a mitochondrial atp9
mutation, even though in vitro import of the precursor was
observed. This work and our present study suggest that
addressing of a protein to mitochondria might not be sufficient
for its correct intramitochondrial targeting and folding, hence to
its proper function.
Acknowledgments
This work was supported by a grant awarded to C. Gaillard
by the French ‘‘Association pour la Promotion des Recherches
sur les Oleagineux’’ (PROMOSOL). Y. Duroc was supported
by the ‘‘Centre Technique Interprofessionnel des Oleagineux
Metropolitains’’ (CETIOM). We would like to thank E.
Lesuisse for sharing antibodies and M. Boutry for providing
the preb targeting signal-carrying plasmid. We would also like
to thank O. Grandjean for help with confocal microscopy and
R. Berthome, I. Small, M. Grelon, S. Bonhomme, C. Mezard,
H. Mireau and F. Nogue for stimulating discussions and critical
reading of the manuscript.
References
[1] J.H. Werren, L.W. Beukeboom, Sex determination, sex ratios, and genetic
conflict, Ann. Rev. Ecol. Syst. 29 (1998) 233–261.
[2] S.A. Frank, Polymorphism of attack and defense, Trends Ecol. Evol. 15
(2000) 167–171.
[3] F. Budar, P. Touzet, R. de Paepe, The nucleo-mitochondrial conflict in
cytoplasmic male sterilities revisited, Genetica 117 (2003) 3–16.
[4] M.R. Hanson, S. Bentolila, Interactions of mitochondrial and nuclear
genes that affect male gametophyte development, Plant Cell 16 (Suppl.)
(2004) S154–S169.
[5] P.S. Schnable, R.P. Wise, The molecular basis of cytoplasmic
male sterility and fertility restoration, Trends Plant Sci. 3 (1998) 175–
180.

Y. Duroc et al. / Plant Science 170 (2006) 755–767766
[6] F. Kempken, D.R. Pring, Plant breeding: male sterility in higher plants-
fundamentals and applications, in: K. Esser (Ed.), Progress in Botany,
Springer-Verlag, Berlin Heidelberg, 1999, pp. 139–166.
[7] Y. L’Homme, R.J. Stahl, X.-Q. Li, A. Hameed, G.G. Brown, Brassica nap
cytoplasmic male sterility is associated with expresion of a mtDNA region
containing a chimeric gene similar to the polCMS-associated orf224 gene,
Curr. Genet. 31 (1997) 325–335.
[8] H.V. Tang, D.R. Pring, L.C. Shaw, R.A. Salazar, F.R. Muza, B. Yan, K.F.
Schertz, Transcript processing internal to a mitochondrial open reading
frame is correlated with fertility restoration in male-sterile sorghum, Plant
J. 10 (1996) 123–133.
[9] S. He, A.R. Abad, S.B. Gelvin, S.A. Mackenzie, A cytoplasmic male
sterility-associated mitochondrial protein causes pollen disruption in
transgenic tobacco, Proc. Natl. Acad. Sci. U.S.A. 93 (1996) 11763–
11768.
[10] J.M. von Allmen, W.H. Rottmann, B.G. Gengenbach, A.J. Harvey, D.M.
Lonsdale, Transfer of methomyl and hmt-toxin sensitivity from T-cyto-
plasm maize to tobacco, Mol. Gen. Genet. 229 (1991) 405–412.
[11] F. Chaumont, B. Bernier, R. Buxant, M.E. Williams, C.S. Levings III, M.
Boutry, Targeting the maize T-urf13 product into tobacco mitochondria
confers methomyl sensitivity to mitochondrial respiration, Proc. Natl.
Acad. Sci. U.S.A. 92 (1995) 1167–1171.
[12] H. Wintz, H.C. Chen, C.A. Sutton, C.A. Conley, A. Cobb, D. Ruth, M.R.
Hanson, Expression of the CMS-associated urfS sequence in transgenic
petunia and tobacco, Plant Mol. Biol. 28 (1995) 83–92.
[13] J. Huang, S.-H. Lee, C. Lin, R. Medici, E. Hack, A.M. Myers, Expression
in yeast of the T-URF13 protein from Texas male-sterile maize mitochon-
dria confers sensitivity to methomyl and to Texas-cytoplasm-specific
fungal toxins, EMBO J. 9 (1990) 339–347.
[14] N. Glab, R.P. Wise, D.R. Pring, C. Jacq, P.P. Slonimski, Expression in
Saccharomyce cerevisiae of a gene associated with cytoplasmic male
sterility from maize: respiratory dysfunction and uncoupling of yeast
mitochondria, Mol. Gen. Genet. 223 (1990) 24–32.
[15] N. Glab, P.X. Petit, P.P. Slonimski, Mitochondrial dysfunction in yeast
expressing the cytoplasmic male sterility T-urf13 gene from maize:
analysis at the population and individual cell level, Mol. Gen. Genet.
236 (1993) 299–308.
[16] M. Grelon, F. Budar, S. Bonhomme, G. Pelletier, Ogura cytoplasmic male-
sterility (CMS)-associated orf138 is translated into a mitochondrial
membrane polypeptide in male-sterile Brassica cybrids, Mol. Gen. Genet.
243 (1994) 540–547.
[17] Y. Duroc, C. Gaillard, S. Hiard, M.C. Defrance, G. Pelletier, F. Budar,
Biochemical and functional characterization of ORF138, a mitochondrial
protein responsible for Ogura cytoplasmic male sterility in Brassiceae,
Biochimie 87 (2005) 1089–1100.
[18] S. Bonhomme, F. Budar, D. Lancelin, I. Small, M.-C. Defrance, G.
Pelletier, Sequence and transcript analysis of the Nco2.5 Ogura-specific
fragment correlated with cytoplasmic male sterility in Brassica cybrids,
Mol. Gen. Genet. 235 (1992) 340–348.
[19] S. Krishnasamy, C.A. Makaroff, Characterization of the radish mitochon-
drial orfB locus: possible relationship with male sterility in Ogura radish,
Curr. Genet. 24 (1993) 156–163.
[20] H. Yamagishi, T. Terachi, Molecular and biological studies on male-sterile
cytoplasm in the Cruciferae. IV. Ogura-type cytoplasm found in the wild
radish, Raphanus raphanistrum, Plant Breed. 116 (1997) 323–329.
[21] J. Farineau, L. Pascal, G. Pelletier, Study of respiratory and photosynthetic
activities in several cytoplasmic hybrids of rapeseed with cytoplasmic
male sterility, Plant Physiol. Biochem. 28 (1990) 333–342.
[22] M. Boutry, N.-H. Chua, A nuclear gene encoding the beta subunit of the
mitochondrial ATP synthase gene in Nicotiana plumbaginifolia, EMBO J.
4 (1985) 340–342.
[23] J. Haseloff, B. Amos, GFP in plants, Trends Genet. 11 (1995) 328–329.
[24] R. Heim, R.Y. Tsein, Engineering green fluorescent protein for improved
brightness, longer wavelengths and fluorescence resonance energy trans-
fer, Curr. Biol. 6 (1996) 178–182.
[25] M.A. Freire, C. Tourneur, F. Granier, J. Camonis, A. El Amrani, K.S.
Browning, C. Robaglia, Plant lipoxygenase 2 is a translation initiation
factor-4E binding protein, Plant Mol. Biol. 44 (2000) 129–140.
[26] T.R. Christianson, R. Sikorski, M. Dante, J. Schero, P. Hieter, Multi-
fonctional yeast high copy-number shuttle vectors, Gene 110 (1992) 119–
122.
[27] R.D. Gietz, R.H. Schiestl, A.R. Willems, R.A. Woods, Studies on the
transformation of intact cells by the LiAc/ss-DNA/PEG procedure, Yeast
11 (1995) 355–360.
[28] B. Guerin, P. Labbe, M. Somlo, Preparation of yeast mitochondria
(Saccharomyces cerevisiae) with good P/O and respiratory control ratios,
Method Enzymol. 55 (1979) 149–159.
[29] Y. Fujiki, A.L. Hubbard, S. Fowler, P.B. Lazarow, Isolation of intracellular
membranes by means of sodium carbonate treatment: application to
endoplasmic reticulum, J. Cell Biol. 93 (1982) 97–102.
[30] W. Paul, R. Hodge, S. Smartt, J. Draper, R. Scott, The isolation and
characterisation of the tapetum-specific Arabidopsis thaliana A9 gene,
Plant Mol. Biol. 19 (1992) 611–622.
[31] M.E. Cartea, M.Migdal, A.M. Galle, G. Pelletier, P. Guerche, Comparison
of sense and antisense methodologies for modifying the fatty acid
composition of Arabidopsis thaliana oilseed, Plant Sci. 136 (1998)
181–194.
[32] C. Koncz, J. Schell, The promoter of TL-DNA gene 50 controls the
tissue specific expression of chimeric genes carried by a novel type
of Agrobacterium binary vector, Mol. Gen. Genet. 204 (1986) 383–
396.
[33] T.Murashige, F. Skoog, A revisedmedium for rapid growth and bio-assays
with tobacco tissue cultures, Physiol. Plant 15 (1962) 473–497.
[34] N. Bechtold, J. Ellis, G. Pelletier, In Planta, Agrobacteriummediated gene
transfer by integration of adult Arabidopsis thaliana plants, C.R. Acad.
Sci. Paris 316 (1993) 1194–1199.
[35] A. Millar, A. Liddell, C. Leaver, Isolation and subfractionation of mito-
chondria from plants, Method Cell Biol. 65 (2001) 53–74.
[36] M. Bellaoui, G. Pelletier, F. Budar, The steady-state level of mRNA from
the Ogura cytoplasmic male sterility locus in Brassica cybrids is deter-
mined post-transcriptionally by its 30 region, EMBO J. 16 (1997) 5057–
5068.
[37] M. Lin, D.H. Turpin, W.C. Plaxton, Pyruvate kinase isozymes from the
green alga, Selenastrum minutum, Arch. Biochem. Biophys. 269 (1989)
219–227.
[38] G. Duby, M. Oufattole, M. Boutry, Hydrophobic residues within the
predicted N-terminal amphiphilic alpha-helix of a plant mitochondrial
targeting presequence play a major role in in vivo import, Plant J. 27
(2001) 539–549.
[39] D.C. Logan, C.J. Leaver, Mitochondria-targeted GFP highlights the
heterogeneity of mitochondrial shape, size and movement within living
plant cells, J. Exp. Bot. 51 (2000) 865–871.
[40] B. Westermann, W. Neupert, Mitochondria-targeted green fluorescent
proteins: convenient tools for the study of organelle biogenesis in Sac-
charomyces cerevisiae, Yeast 16 (2000) 1421–1427.
[41] K. Dimmer, S. Fritz, F. Fuchs, M. Messerschmitt, N. Weinbach, W.
Neupert, B. Westermann, Genetic basis of mitochondrial function and
morphology in Saccharomyces cerevisiae, Mol. Biol. Cell 13 (2002) 847–
853.
[42] I. Kissova, M. Deffieu, S. Manon, N. Camougrand, Uth1p is involved in
the autophagic degradation of mitochondria, J. Biol. Chem. 279 (2004)
39068–39074.
[43] D.C. Logan, I. Scott, A.K. Tobin, The genetic control of plant mitochon-
drial morphology and dynamics, Plant J. 36 (2003) 500–509.
[44] X. Feng, S. Arimura, H.Y. Hirano,W. Sakamoto, N. Tsutsumi, Isolation of
mutants with aberrant mitochondrial morphology from Arabidopsis thali-
ana, Genes Genet. Syst. 79 (2004) 301–305.
[45] S.L. Medberry, B.E. Lockhart, N.E. Olszewski, The Commelina yellow
mottle virus promoter is a strong promoter in vascular and reproductive
tissues, Plant Cell 4 (1992) 185–192.
[46] J. Gourret, R. Delourme, M. Renard, Expression of ogu cytoplasmic male
sterility in cybrids of Brassica napus, Theor. Appl. Genet. 83 (1992) 549–
556.
[47] G. Pelletier, C. Primard, F. Vedel, P. Chetrit, R. Remy, P. Rousselle, M.
Renard, Intergeneric cytoplasmic hybridization in Cruciferae by proto-
plast fusion, Mol. Gen. Genet. 191 (1983) 244–250.

Y. Duroc et al. / Plant Science 170 (2006) 755–767 767
[48] M. Bellaoui, A. Martin-Canadell, G. Pelletier, F. Budar, Low-copy-
number molecules are produced by recombination, actively maintained
and can be amplified in the mitochondrial genome of Brassicaceae:
relationship to reversion of the male sterile phenotype in some cybrids,
Mol. Gen. Genet. 257 (1998) 177–185.
[49] S.W. Gorsich, J.M. Shaw, The importance of mitochondrial dynamics
during meiosis and sporulation, Mol. Biol. Cell (2004).
[50] H. Sesaki, R.E. Jensen, Division versus fusion: Dnm1p and Fzo1p antag-
onistically regulate mitochondrial shape, J. Cell Biol. 147 (1999) 699–706.
[51] Y. Ohsumi, Molecular mechanism of autophagy in yeast, Saccharomyces
cerevisiae, Philos. Trans. R. Soc. Lond. B Biol. Sci. 354 (1999) 1577–
1580 (discussion 1580–1).
[52] D.J. Klionsky, S.D. Emr, Autophagy as a regulated pathway of cellular
degradation, Science 290 (2000) 1717–1721.
[53] A. Abudugupur, K. Mitsui, S. Yokota, K. Tsurugi, An ARL1 mutation
affected autophagic cell death in yeast, causing a defect in central vacuole
formation, Cell Death Differ. 9 (2002) 158–168.
[54] N. Camougrand, A. Grelaud-Coq, E. Marza, M. Priault, J.J. Bessoule, S.
Manon, The product of the UTH1 gene, required for Bax-induced cell
death in yeast, is involved in the response to rapamycin, Mol. Microbiol.
47 (2003) 495–506.
[55] M. Priault, N. Camougrand, K.W. Kinnally, F.M. Vallette, S. Manon, Yeast
as a tool to study Bax/mitochondrial interactions in cell death, FEMS
Yeast Res. 4 (2003) 15–27.
[56] M. Sanchirico, A. Tzellas, T.D. Fox, H. Conrad-Webb, P.S. Periman, T.L.
Mason, Relocation of the unusual VAR1 gene from the mitochondrion to
the nucleus, Biochem. Cell Biol. 73 (1995) 987–995.
[57] R.H. Law, L.B. Farrell, D. Nero, R.J. Devenish, P. Nagley, Studies on the
import into mitochondria of yeast ATP synthase subunits 8 and 9 encoded
by artificial nuclear genes, FEBS Lett. 236 (1988) 501–505.
[58] R. Neu, S. Goffart, K. Wolf, B. Schafer, Relocation of urf a from the
mitochondrion to the nucleus cures the mitochondrial mutator phenotype
in the fission yeast Schizosaccharomyces pombe, Mol. Gen. Genet. 258
(1998) 389–396.
[59] G. Manfredi, J. Fu, J. Ojaimi, J.E. Sadlock, J.Q. Kwong, J. Guy, E.A.
Schon, Rescue of a deficiency in ATP synthesis by transfer of MTATP6, a
mitochondrial DNA-encoded gene, to the nucleus, Nat. Genet. 30 (2002)
394–399.
[60] B. Pineau, C. Mathieu, C. Gerard-Hirne, R. De Paepe, P. Chetrit, Targeting
the NAD7 subunit to mitochondria restores a functional complex I and a
wild type phenotype in the Nicotiana sylvestris CMS II mutant lacking
nad7, J. Biol. Chem. 280 (2005) 25994–26001.



















