
Sequence and Structural Convergence of Broad and Potent HIV Antibodies That Mimic CD4 Binding
Upload
independentCategory
view
2download
0
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Oct. 2009, p. 4283–4291 Vol. 53, No. 100066-4804/09/$08.00�0 doi:10.1128/AAC.01709-08Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Novel Broad-Spectrum Bis-(Imidazolinylindole) Derivatives with PotentAntibacterial Activities against Antibiotic-Resistant Strains�†
Rekha G. Panchal,1* Ricky L. Ulrich,1 Douglas Lane,2 Michelle M. Butler,3 Chad Houseweart,3Timothy Opperman,3 John D. Williams,3 Norton P. Peet,3 Donald T. Moir,3
Tam Nguyen,2 Rick Gussio,4 Terry Bowlin,3 and Sina Bavari1*United States Army Medical Research Institute of Infectious Diseases, 1425 Porter Street, Fort Detrick, Frederick, Maryland 21702-50111;
Target Structure Based Drug Discovery Group, SAIC-Frederick, Inc., NCI-Frederick, Frederick, Maryland 21702-12012;Microbiotix Inc., One Innovation Dr., Worcester, Massachusetts 016053; and Target Structure Based Drug Discovery Group,
Information Technology Branch, Developmental Therapeutic Program, National Cancer Institute,Frederick, Maryland 21702-12014
Received 23 December 2008/Returned for modification 10 February 2009/Accepted 1 July 2009
Given the limited number of structural classes of clinically available antimicrobial drugs, the discovery ofantibacterials with novel chemical scaffolds is an important strategy in the development of effective therapeu-tics for both naturally occurring and engineered resistant strains of pathogenic bacteria. In this study, severaldiarylamidine derivatives were evaluated for their ability to protect macrophages from cell death followinginfection with Bacillus anthracis, a gram-positive spore-forming bacterium. Four bis-(imidazolinylindole) com-pounds were identified with potent antibacterial activity as measured by the protection of macrophages and bythe inhibition of bacterial growth in vitro. These compounds were effective against a broad range of gram-positive and gram-negative bacterial species, including several antibiotic-resistant strains. Minor structuralvariations among the four compounds correlated with differences in their effects on bacterial macromolecularsynthesis and mechanisms of resistance. In vivo studies revealed protection by two of the compounds of micelethally infected with B. anthracis, Staphylococcus aureus, or Yersinia pestis. Taken together, these resultsindicate that the bis-(imidazolinylindole) compounds represent a new chemotype for the development oftherapeutics for both gram-positive and gram-negative bacterial species as well as against antibiotic-resistantinfections.
Bacillus anthracis is a serious bioterrorism threat because itsspores are stable under extreme conditions in the environment,easily cultured and produced, easily distributed by aerosol (ina powder form), and highly fatal via inhalation, as dramaticallydemonstrated in 2001 (4, 16, 17). While B. anthracis cells aresensitive to several antibiotics (3, 13), naturally occurring orintentionally engineered drug resistance is a concern. Antibi-otic resistance is also a growing problem in the clinic (10), andthe recent increased prevalence of community-acquired methi-cillin (meticillin)-resistant Staphylococcus aureus (MRSA) hasadded to the concern (11, 12). While effective new agents arein the pipeline, they are all new analogs of existing classes ofantibiotics (22). The development of new antibiotics againstunexploited targets with novel mechanisms of action is a vitalpart of the solution to these problems because such antibioticsare unlikely to be affected by preexisting target-based resis-tance alleles.
In order to explore potential new chemotypes of antibacte-rial agents with a variety of possible mechanisms of action, we
developed a cell-based screen for rescue of macrophages fromB. anthracis-mediated death and applied it to a focused librarycontaining diarylamidine compounds. This class of compoundshas been evaluated previously for antimicrobial properties (2),as well as for antiproteolytic, anticoagulant (23), and antipro-liferative activity (5). In our study, four bis-(imidazolinylin-dole) compounds from the diarylamidine library exhibited verypotent activity in the macrophage rescue screen. Mechanism ofaction studies indicated that these compounds are neither non-specific inhibitors of macromolecular synthesis nor membraneactive but are rapidly bactericidal inhibitors of a broad spec-trum of bacterial species. Finally, we demonstrate activities ofthese inhibitors in animal models of infection.
MATERIALS AND METHODS
Small molecule library. A focused library containing �70 compounds sharinga diarylamidine chemical scaffold was obtained from the National Cancer Insti-tute and used for screening (see Table S1 in the supplemental material).
Bacterial strains. The different bacterial species and strains used in this studyinclude Bacillus anthracis (Sterne), Bacillus anthracis (Ames), Bacillus brevis,Bacillus licheniformis, Bacillus megaterium, Bacillus anthracis (Vollum), Bacillussubtilis, Bacillus pumilus, Staphylococcus aureus NCTC8325, S. aureus (Smithstrain), MRSA 1094, Enterococcus faecalis ATCC 29212, Mycobacteria smegmatisATCC 19420, M. smegmatis ATCC 35798, M. smegmatis ATCC 700009, Esche-richia coli J53, Klebsiella pneumoniae 5657, Pseudomonas aeruginosa PA01, Yer-sinia pestis CO92, Y. pestis KIM (�pgm pCD1�), Burkholderia mallei ATCC 3344,Burkholderia pseudomallei DD503, Burkholderia thailandensis, and Burkholderiacepacia.
Cell-based infection assays. J774A.1 macrophages (6 � 105) were infectedwith B. anthracis Sterne spores at a multiplicity of infection (MOI) of 5, in the
* Corresponding author. Mailing address: United States Army Med-ical Research Institute of Infectious Diseases, 1425 Porter Street, FortDetrick, Frederick, MD 21702-5011. Phone for Rekha G. Panchal:(301) 619-4985. Fax: (301) 619-2348. E-mail: [email protected]. Phone for Sina Bavari: (301) 619-4246. Fax: (301) 619-2348.E-mail: [email protected].
† Supplemental material for this article may be found at http://aac.asm.org/.
� Published ahead of print on 27 July 2009.
4283

presence of dimethyl sulfoxide (DMSO; 1%, as control) or test compounds (10�M). After 4 h incubation at 37°C, bacterial growth was inhibited by the additionof the antibiotics penicillin (100 IU) and streptomycin (100 �g/ml). To determinecell viability, Sytox green dye that is impermeant to live cells was added andincubated for 15 min at 37°C. The cells were centrifuged at 2,000 rpm for 2 minand then washed two times with complete medium containing antibiotics. Thecells were then analyzed by flow cytometry.
In vitro inhibition of bacterial growth. MICs were determined by the brothmicrodilution method (20). B. anthracis Sterne spores or bacterial cultures(5 �105 CFU/ml) in log-phase growth were seeded in 96-well plates and treatedwith DMSO (1%) or compound at concentrations ranging from 0 to 20 �M.Plates were incubated at 37°C for 16 to 20 h, and cell growth determined bymeasuring the absorbance at 600 nm.
The minimal bactericidal concentration (MBC) was determined by a modifi-cation of the MIC method. Bacteria from MIC wells and 4 dilutions above theMIC were diluted, and bacteria were plated onto sheep blood agar plates. Thenext day, colonies were counted, and MBCs reflecting a 99.9% reduction inviable counts were determined.
Spore germination assay. Sterne spores (5 �105 CFU/ml) were germinated inMueller-Hinton broth in the presence of DMSO (control) or compounds (1�MIC). At time intervals of 0, 15, and 30 min, samples were heated at 70°C for 30min to kill any germinated spores, and appropriate dilutions were plated ontosheep blood agar plates to quantify remaining viable spores.
Kinetics of bactericidality. To determine the kinetics of cidality, compoundswere diluted in Mueller Hinton broth and tested at concentrations equivalent to
4� their respective MICs against B. anthracis Sterne spores (5 �105 CFU/ml) orB. anthracis Sterne vegetative bacilli or an attenuated Yersinia pestis strain (KIM�pgm pCD1�) (1 �106 CFU/ml). The cultures were incubated and sampled atvarious time points (0, 1, 2, 4, 6, and 24 h), diluted appropriately into freshmedium, and then plated onto drug-free agar plates to determine the number ofCFU/ml present in the sample (CFU/ml is number of colonies on the platemultiplied by the dilution factor and adjusted for a volume of 1 ml). An addi-tional experiment was conducted, as described above, using B. subtilis BD54,MBX 1066, and the antibiotic ciprofloxacin, an inhibitor of bacterial DNAreplication, at concentrations of 5� their MICs. The minimum level of detectionin these experiments was 50 CFU/ml. The log10 value of CFU/ml was plottedversus time. Bactericidal activity is defined as a �3 log reduction in initial CFUcount within 24 h.
Determination of mammalian cytotoxicity. Cytotoxicity of the compounds wasmeasured as described previously, except that HeLa cells were used (9). Cyto-toxicity was quantified as the CC50, the concentration of compound that inhibited50% of conversion of MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt] to formazan (18). The “se-lectivity index” is defined as the ratio of the mammalian cell cytotoxicity to theMIC against B. anthracis Ames (i.e., CC50/MIC), both measured in the presenceof 10% fetal calf serum. The addition of 10% fetal calf serum had no effect onthe MICs.
Macromolecular synthesis assays. Compounds were examined at 5� MIC forinhibitory effects on bacterial DNA, RNA, protein, and cell wall biosynthesis inB. subtilis BD54, in 96-well polystyrene plates. Incorporation of the following
FIG. 1. Small molecules protect macrophages from B. anthracis-induced cell death. J774A.1 macrophages were treated with either DMSO (1%)control or compounds (10 �M) and infected with B. anthracis spores (MOI, 5) (A) or not infected (B). Cell viability was measured by uptake ofSytox green dye and analyzed by flow cytometry. The percentages of live and dead cells are indicated. (C) Chemical structures of the four mostpotent antimicrobial compounds.
4284 PANCHAL ET AL. ANTIMICROB. AGENTS CHEMOTHER.
radiolabeled precursors into macromolecules was measured: [methyl-3H]thymi-dine for DNA, [5-3H]uridine for RNA, L-[4,5-3H]leucine for protein, andN-acetyl-D-[1-3H]glucosamine for cell wall synthesis. At time zero, compound- orpathway-specific control antibiotic (in DMSO) at a concentration of 5� its MICand radioactive precursors were added to log phase cells. At various times,samples were collected from each 37°C culture, precipitated with an equal vol-ume of ice-cold 20% trichloroacetic acid, collected onto 96-well glass fiber filterplates, dried, and counted for radioactivity. Plots of counts (cpm) incorporatedversus incubation times were generated for each compound.
Membrane activity assays. The effect of compounds on bacterial membranepotential of B. subtilis BD54 was determined using a fluorescent assay essentiallyas described by Wu and Hancock (25). Briefly, B. subtilis BD54 cultures weregrown to exponential phase (optical density at 600 nm, 0.4 to 0.5) in LB media.Cells were harvested, washed, and resuspended in wash buffer (5 mM HEPES[pH 7.4], 20 mM glucose) to an optical density at 600 nm of 0.1. The fluorescentdye DiSC3(5) was added to the cell suspension (final concentration, 1 �M) andincubated at room temperature for 10 min to allow dye uptake. KCl was addedto a final concentration of 100 mM. The cell suspension was transferred to a96-well assay plate (200 �l/well) containing control and test compounds dissolvedin DMSO (final concentration, 2%). A total of eight wells were tested under eachcondition. Fluorescence intensity (RFU) was measured after 5 min, using aMolecular Devices SpectraMax fluorescence plate reader with an excitationwavelength of 622 nm and an emission wavelength of 670 nm. 2-4-Dinitrophenol(DNP), a protonophore, was used at a concentration of 200 �g/ml as a positivecontrol. The average RFU and standard deviation for eight assay wells werecalculated and are presented in Fig. 4F.
The effect of the compounds on mammalian cell membrane integrity wasdetermined by measuring the release of lactate dehydrogenase (LDH) fromHeLa cells treated with a concentration range of compounds. Briefly, HeLa cellswere grown to confluence in Dulbecco’s modified Eagle’s medium and weretreated for 1 h with 1% DMSO alone (untreated control), various concentrationsof test compounds, and a control antibiotic (32� MIC for vancomycin). The finalconcentration of DMSO in all samples was 1%. LDH activity in the supernatantwas measured using the CytoTox ONE homogenous membrane integrity assaykit (Promega, Madison, WI) according to the manufacturer’s instructions.
Selection for resistant mutants. Eight independent cultures of S. aureusNCTC8325 were grown in 96-well assay plates in the presence of several con-centrations of each of the compounds (0.125� to 128� MIC). Cultures wererecovered from the well with highest compound concentration that exhibitedrobust growth (�50% of untreated control). This process was repeated for 20days, and results were displayed as the highest sublethal compound concentra-tion for each culture for each day. Colonies were isolated from apparentlyresistant cultures and confirmed to be resistant by MIC assays.
Animal studies. Eight- to 10-week-old C57BL/6 mice were used in this study.The dosing regimen was based on the MICs and on initial pilot studies varyingdose and administration frequency. For in vivo B. anthracis studies, mice (n �10/group) were challenged via intraperitoneal (i.p.) injection with �300 CFU ofB. anthracis Ames. After 6 h postchallenge, mice were treated via i.p. injectionwith vehicle control, MBX 1066 (5 or 10 mg/kg/injection), or MBX 1090 (0.2, 0.5,or 1.0 mg/kg/injection). Mice were treated every 6 hours for 5 days, and survivalwas monitored for up to 20 days. Compounds in this study were dissolved at astock concentration of 50 mg/ml in DMSO and then diluted to an appropriateworking concentration in 5% dextrose in water.
To determine the protective effect of the compounds during the late stages ofinfection, treatment with compound MBX 1066 (10 mg/kg/injection) via i.p.injection was initiated 6, 12, 18, or 24 h postchallenge. Thereafter, mice weretreated every 6 hours for 5 days, and survival was monitored for up to 20 days.Compound MBX 1066 was dissolved at a stock concentration of 50 mg/ml inDMSO and then diluted to an appropriate working concentration in 5% dextrosein water.
To test the efficacy of the compounds in a Y. pestis infection model, C57BL/6mice (n � 10/group) were challenged via i.p. injection with �100 CFU of Y. pestis(strain CO92). After 6 h, mice were treated via i.p. injection with MBX 1066 (5or 10 mg/kg/injection) and treatment continued every 6 hours for 5 days. Survivalof the mice was monitored for 15 days.
To investigate the efficacy of the compounds administered via a different routefrom that of the pathogen challenge, Swiss Webster mice (n � 10/group) werechallenged via i.p. injection with 8.3 � 108 CFU of S. aureus (Smith strain). After15 min, mice were treated via intravenous (i.v.) injection with a single dose ofMBX 1066 (10 mg/kg), MBX 1090 (10 mg/kg) prepared in 10% dimethyl acet-amide-5% dextrose in water (pH 4.0), daptomycin control (10 mg/kg), or vehiclecontrol (10% dimethyl acetamide-5% dextrose in water [pH 4.0]). Survival wasmonitored for 48 h.
All research was conducted under an approved protocol and in compliancewith the Animal Welfare Act and other federal statutes and regulations relatedto animals and experiments involving animals and adhered to principles stated inthe Guide for the Care and Use of Laboratory Animals, National ResearchCouncil, 1996. The facilities in which this research was conducted are fullyaccredited by the Association for Assessment and Accreditation of LaboratoryAnimal Care International.
RESULTS
Small molecules protect macrophages from B. anthracis-induced cell death. B. anthracis, a gram-positive spore-formingbacterium, produces a virulent lethal toxin that causes death ofsusceptible macrophages (14). A cell-based B. anthracis infec-tion assay was used to test a focused library that containeddiarylamidine derivatives (see Table S1 in the supplementalmaterial for chemical structures). Macrophages were infectedwith B. anthracis spores (MOI of 5) in the presence of a DMSOcontrol (1%) or the compounds (10 �M), and cell death wasmonitored by the uptake of membrane-impermeant Sytoxgreen using flow cytometry. A number of compounds (28% hitrate) protected macrophages to various extents (�50%) fromB. anthracis-induced cell death. Representative results for thefour most potent compounds (MBX 1066, MBX 1090, MBX1113, and MBX 1128, also known as NSC 317881, NSC 317880,NSC 330687, NSC 369718, respectively, in Table S1 in thesupplemental material) are shown in Fig. 1A and B. Thesepotent inhibitors all share a bis-(imidazolinylindole) chemo-type; their chemical structures are shown in Fig. 1C. The ob-served cellular protection also suggests that the compounds arerelatively nontoxic to macrophages at the tested dose. In anindependent experiment, in the absence of bacteria, only MBX1128 caused more macrophage death than the DMSO control,and the effect was modest (Fig. 1B).
In vitro inhibition of bacterial growth. The protection ofmacrophages from cell death by the identified compounds sug-gests that they may be targeting bacterial growth or viability,bacterial virulence factors, or host factors vital to the bacteria.
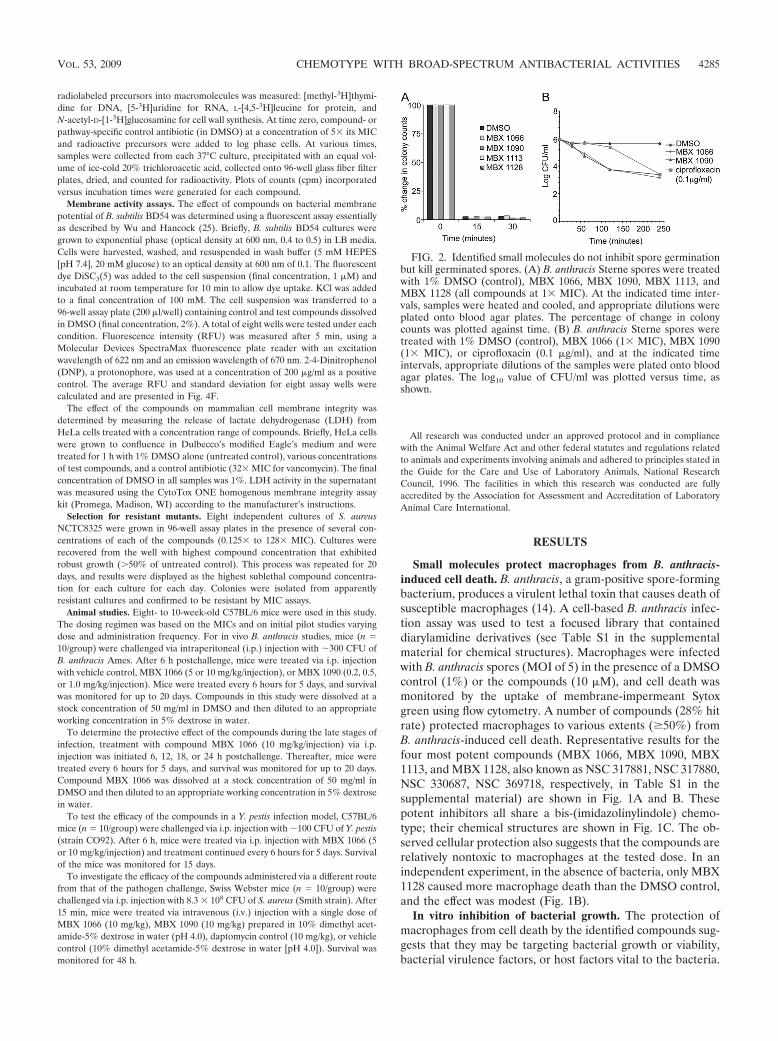
FIG. 2. Identified small molecules do not inhibit spore germinationbut kill germinated spores. (A) B. anthracis Sterne spores were treatedwith 1% DMSO (control), MBX 1066, MBX 1090, MBX 1113, andMBX 1128 (all compounds at 1� MIC). At the indicated time inter-vals, samples were heated and cooled, and appropriate dilutions wereplated onto blood agar plates. The percentage of change in colonycounts was plotted against time. (B) B. anthracis Sterne spores weretreated with 1% DMSO (control), MBX 1066 (1� MIC), MBX 1090(1� MIC), or ciprofloxacin (0.1 �g/ml), and at the indicated timeintervals, appropriate dilutions of the samples were plated onto bloodagar plates. The log10 value of CFU/ml was plotted versus time, asshown.
VOL. 53, 2009 CHEMOTYPE WITH BROAD-SPECTRUM ANTIBACTERIAL ACTIVITIES 4285

The cells infected with B. anthracis in the presence of theidentified hit compounds did not show any changes in the pHof the medium, and microscopic examination of these samplesshowed little to no outgrowth of the bacteria. These resultssuggest that the compounds are acting as antibacterials. Toinvestigate the antibacterial properties, all compounds fromthe focused library were tested in vitro for their ability toinhibit growth of B. anthracis Sterne spores and vegetativebacilli (see Table S1 in the supplemental material). The fourmost potent antibacterial compounds identified in the macro-phage cell-based rescue assay were also potent bacterialgrowth inhibitors. Compounds MBX 1066, MBX 1090, MBX1113, and MBX 1128 displayed MICs ranging from 0.2 to 1�g/ml on both spores and vegetative bacilli.
Small molecules do not inhibit B. anthracis spore germina-tion but rapidly kill bacteria. B. anthracis spores germinatewithin minutes following contact with a suitable medium (24),and their conversion into the vegetative form is essential foranthrax pathogenicity. Spore germination may be detected invitro by alterations in the spore refractility, heat resistance, andstaining. To evaluate the effects of the compound on sporegermination, the four most potent compounds were selectedfor further study. As shown in Fig. 2A, a dramatic reduction inthe CFU was seen as early as 15 min following treatment with
the compounds, suggesting that the compounds did not affectspore germination.
Since these compounds had no effect on spore germination,but inhibited spore outgrowth, we examined the time-depen-dent killing of B. anthracis Sterne spores during germinationand outgrowth. As shown in Fig. 2B, there was a greater than2 log reduction in the initial CFU count after 4 hours oftreatment with compound MBX 1066 or MBX 1090. A similarreduction was observed following treatment with ciprofloxacin(0.1 �g/ml).
Lead compounds exhibit broad-spectrum antibacterial ac-tivity. To determine if the compounds were capable of actingagainst a broad spectrum of gram-positive and gram-negativebacteria, the four most potent inhibitors, MBX 1066, MBX1090, MBX 1113, and MBX 1128, were tested for their anti-bacterial activity against a diverse panel of bacterial speciesand strains. Table 1 demonstrates that these four compoundsare active against a range of gram-positive (MICs, 0.1 to 4.9�g/ml) and gram-negative (MICs, 0.4 to �80 �g/ml) bacteria.The compounds were also tested on selected antibiotic resis-tant bacteria. As shown in Table 1, all four compounds wereactive against ciprofloxacin-resistant B. anthracis Ames (MICs,0.2 to 4.9 �g/ml), MRSA (MICs, 0.16 to 2.8 �g/ml), and van-comycin-resistant Enterococcus (MICs, 0.1 to 0.6 �g/ml).
TABLE 1. MICs and MBCs of the four most potent compounds against select gram-positive and gram-negative bacteria and antibiotic-resistant strains of selected species
Strain/speciesMIC (MBC) (�g/ml)b
MBX 1066 MBX 1090 MBX 1113 MBX 1128 Ciprofloxacin
Gram-positive bacteriaB. anthracis Ames 0.3 (0.7) 0.6 (2.3) 1.1 (2.2) 1.4 (2.8) NDB. brevisa 0.7 0.6 0.3 0.2 NDB. licheniformisa 0.7 2.3 2.2 1.4 0.1B. megateriuma 0.3 0.6 0.6 0.4 0.2B. pumilusa 0.1 0.2 0.1 0.4 NDB. anthracis Volluma 0.3 (ND) 0.6 (1.2) 0.6 (1.1) 0.4 (0.4) NDB. subtilisa 0.2 0.3 0.6 0.4 0.1B. anthracis Ames spores 0.2 (0.4) 0.2 (1.6) 0.3 (0.5) 0.2 (1.2) NDS. aureus ATCC 25923 0.1 0.9 0.3 0.3 0.3S. aureus 0.3 1.2 1.1 0.7 NDE. faecalisa 0.2 0.6 2.2 0.7 NDE. faecalis 29212 0.2 0.3 0.3 0.1 2.5M. smegmatis 19420 0.1 0.2 0.2 0.1 0.1M. smegmatis 35798 0.1 0.1 0.1 0.1 0.2M. smegmatis 700009 0.1 0.2 0.3 0.1 0.4Ciprofloxacin-resistant B. anthracis Ames (strain 105-6) 0.2 (0.8) 0.4 (1.6) 0.2 (1.8) 4.9 (�10) �100MRSAa 0.3 2.3 2.2 2.8 NDMRSA 1094 0.2 0.6 0.3 0.5 5.0Vancomycin-resistant E. faecium B42762 0.1 0.3 0.2 0.1 20Vancomycin-resistant E. faecalis ATCC 51575 0.1 0.6 0.5 0.1 0.6
Gram-negative bacteriaE. coli J53a 0.7 0.6 1.1 1.4 0.1K. pneumoniae 5657 3.8 1.3 0.4 16 0.2P. aeruginosa PAO1 7.5 25 25 �80 0.1Y. pestis 3.4 13 7.4 9.7 NDB. mallei ATCC 23344 1.7 1.6 1.8 �9.7 NDB. pseudomallei DD503 1.7 3.1 1.8 �9.7 NDB. thailandensisc 21 19 18 2.8 NDB. cepaciaa �20 19 4.4 22 ND
a Clinical isolate.b MBCs are shown in parentheses. ND, not determined.c Environment.
4286 PANCHAL ET AL. ANTIMICROB. AGENTS CHEMOTHER.

Thus, the identified compounds have broad-spectrum potentantibacterial activity and are more effective than many antibi-otics in current use.
Minimum bactericidal activities of the most potent antibac-terial compounds. To determine if the identified bioactivecompounds were bactericidal, the MBCs of MBX 1066, MBX1090, MBX 1113, and MBX 1128 against several strains ofgram-positive bacteria were determined. As shown in Table 1,three of the compounds (MBX 1066, 1090, 1113) demon-strated potent bactericidal (MBCs, 0.8 to 1.8 �g/ml) activityagainst ciprofloxacin-resistant Ames B. anthracis.
To further evaluate the time course of bactericidality of thecompounds, B. anthracis vegetative bacilli (Sterne strain) andattenuated Y. pestis (strain KIM �pgm pCD1�) (106 CFU/ml)were incubated in Mueller-Hinton broth in the presence ofMBX 1066 or MBX 1090 at 4� MIC concentrations over atime course of 24 h. As shown in Fig. 3A and B, both MBX1066 and MBX 1090 were rapidly bactericidal (1 to 4 h) to bothY. pestis and B. anthracis at concentrations of 4� MIC, gener-ating a reduction in CFU of �3 log within 6 h.
Lead compounds exhibit favorable selectivity indices. Thecytotoxicity of the compounds was determined using HeLacells in a 3-day incubation assay. The results, shown in Table 2,confirm that little or no cytotoxicity would be expected at thescreening concentration (10 �M or �6 �g/ml). However, somecytotoxicity is observed in this more stringent assay, with CC50
values of 50 �g/ml for all four compounds. When evaluatedin relation to the very potent antibacterial activity versus B.
anthracis Ames, the selectivity indices (CC50/MIC) for three ofthe compounds are �10, suggesting some preference for bac-terial versus mammalian cells in the mode of action of thesecompounds.
Lead compounds inhibit DNA synthesis. To identify themolecular mechanism of bacterial growth inhibition, MBX1066, MBX 1090, MBX 1113, and MBX 1128 were examinedat 5� MIC for their inhibitory effects on bacterial DNA, RNA,protein, and cell wall biosynthesis in B. subtilis BD54. The timecourse of bacterial killing, growth, and incorporation of radio-labeled precursors into macromolecules of cells incubated inthe presence or absence of the compounds MBX 1066 andMBX 1090 is shown in Fig. 3C and Fig. 4A to E. Controlantibiotics ciprofloxacin, rifampin (rifampicin), chloramphen-icol, and vancomycin displayed potent and specific inhibition ofincorporation of radiolabeled precursors into their target mac-romolecules DNA, RNA, protein, and cell wall, respectively.Cells appeared to double only once in the presence of MBX1066 and MBX 1090, and both compounds inhibited DNAsynthesis (Fig. 4A to E). Compounds MBX 1113 and MBX1128 also inhibited DNA synthesis but in addition inhibitedRNA and cell wall synthesis to about 40% of control values,although protein synthesis was less potently affected (notshown). The somewhat broader inhibitory activity of MBX1113 and MBX 1128 suggests that they are less specific thanare MBX 1066 and MBX 1090.
Lead compounds are not membrane active. The effect ofMBX 1066 and MBX 1090 on the membrane potential of B.subtilis BD54 was investigated using DiSC3(5), a membranepotential-sensitive fluorescent dye. As DiSC3(5) is taken up bycells with intact membrane potential, the fluorescence de-creases due to quenching. However, disruption of the mem-brane potential (or integrity) results in release of DiSC3(5)into the buffer, which can be detected due to an increase influorescence intensity. As shown in Fig. 4F, the untreated cellsexhibited low fluorescence signal, whereas cells treated withDNP, a compound that disrupts the membrane potential, pro-duced a significantly higher signal. Significantly, treatment withMBX 1066 or MBX 1090 at concentrations up to 10� MIC didnot result in increased fluorescence, indicating that these com-pounds do not perturb bacterial membranes.
The effect of three of the compounds on mammalian mem-brane integrity was measured by examining the degree of celllysis after incubation with the compounds (see Materials andMethods). Exposure of HeLa cells to MBX 1066, MBX 1090,and MBX 1113 at concentrations equivalent to 32� MIC (S.aureus, 9.6 �g/ml) did not result in the release of LDH at levels
FIG. 3. Rapid cidality of lead compounds. Time kill assay for twoMBX compounds tested at 4� their respective MICs against the B.anthracis Sterne strain (A) or the Y. pestis strain KIM �pgm, pCD1�
(B). Bacteria were incubated in the presence of compounds at 4� theknown MIC and sampled at the indicated time points. The log10 valueof CFU/ml was plotted versus time, as shown. (C) B. subtilis BD54 wasincubated in the presence of ciprofloxacin (Cipro) or MBX 1066 atconcentrations of 5� their MICs and sampled at the indicated timepoints.
TABLE 2. Cytotoxicity assessment of compounds against HeLacells in a 3-day exposure
CompoundCC50 (�g/ml)
with HeLacells
B. anthracisAmes with MIC
(�g/ml) of:
Selectivityindex
(CC50/MIC)a
MBX 1066 33 0.3 110MBX 1090 10 0.6 17MBX 1113 3 1.1 2.7MBX 1128 17 1.4 12
a CC50 values are divided by MICs for B. anthracis Ames in order to determinethe selectivity index.
VOL. 53, 2009 CHEMOTYPE WITH BROAD-SPECTRUM ANTIBACTERIAL ACTIVITIES 4287

significantly different from those of the control samples treatedwith no antibiotic or with vancomycin concentrations up to32� MIC (32 �g/ml) (data not shown). The results of thisexperiment indicate that neither MBX 1066 nor MBX 1090disrupts membranes of HeLa cells at high concentrations.
Selection of mutants resistant to bis-(imidazolinylindole)compounds. To assess the capability of cells to become resis-tant to the antibacterial effects of these compounds, we at-tempted to identify resistant mutants. Initial attempts to selectdirectly for colonies with spontaneous mutations to resistanceon agar plates containing MBX 1066 were unsuccessful. There-fore, mutations with decreased susceptibility to MBX 1066,MBX 1090, and MBX 1113 were selected in liquid media inserial passage experiments. After 20 days of growth in suble-thal concentrations of MBX 1066 and MBX 1113, mutantswith significant increase in resistance to these compounds(�4� MIC) were not isolated (Fig. 5). In contrast, mutants
able to grow in concentrations up to 16� MIC of MBX 1090appeared within 10 to 12 days of culture.
Individual clones from each of the resistant populationswere isolated, and MICs for MBX 1090 and MBX 1066 weredetermined in order to confirm decreased susceptibility toMBX 1090 and to test for cross-resistance to MBX 1066. MICsfor six independent MBX 1090-resistant mutants against MBX1090 were 16- to 32-fold higher (16 to 32 �g/ml) than for theparent strain (WT), confirming reduced susceptibility of themutants. Interestingly, these mutants were not cross resistantto MBX 1066, indicating that the resistance mutations arespecific for MBX 1090 (data not shown). This result indicatesthat MBX 1090 and MBX 1066 have distinct mechanisms ofresistance.
Lead compounds protect mice in B. anthracis and Y. pestisinfection models. To evaluate their in vivo efficacy, the com-pounds MBX 1066 and MBX 1090 were tested in B. anthracis
FIG. 4. Mechanism studies of inhibitors MBX 1066 and MBX 1090. (A) Effect of compounds on growth of B. subtilis BD54. Assays wereperformed in the absence or presence of compounds at 5� their MICs. OD600, optical density at 600 nm. (B to E) Effect of compounds onmacromolecular biosynthesis in B. subtilis BD54. Assays were performed using radiolabeled precursors and B. subtilis BD54 as described inMaterials and Methods with compounds MBX 1066 and MBX 1090 and appropriate control antibiotics present at 5� their MICs as indicated.Macromolecular pathways and control antibiotics are as follows: DNA synthesis, ciprofloxacin control (B); RNA synthesis, rifampin control (C);protein synthesis, chloramphenicol control (D); cell wall synthesis, vancomycin control (E). (F) Effect of compounds on bacterial membranepermeability. Compounds were incubated with B. subtilis BD54 at various concentrations (0.25�, 1�, 5�, and 10� their MICs) in the presenceof DiSC3(5) dye. Control is DNP.
4288 PANCHAL ET AL. ANTIMICROB. AGENTS CHEMOTHER.

and Y. pestis lethal infection mouse models. In B. anthracisstudies, the C57BL/6 control mice were highly susceptible to B.anthracis infection, with death being observed as early as 48 hfollowing challenge and 100% mortality within 4 to 5 days (Fig.6A to C). In mice treated with the compound MBX 1066 (Fig.6A) or MBX 1090 (Fig. 6B), 100% survival was observed at thehighest concentration of the compounds tested (10 mg/kg/in-jection and 1 mg/kg/injection, respectively). To determine theprotective effect of compounds at late stages of infection, micewere infected with B. anthracis Ames spores, and after 6, 12,18, and 24 h postinfection, treatment with MBX 1066 wasinitiated. Protective effects of the compound MBX 1066 wereobserved as late as 18 to 24 h postinfection (Fig. 6C). In Y.pestis studies, a 90% protective effect was observed in the micetreated with the higher dose (10 mg/kg/injection) of the com-pound MBX 1066 (Fig. 6D). Thus, these studies indicate thatMBX 1066 and MBX 1090 exhibit potent in vivo antibacterialactivity against representative gram-positive and gram-nega-tive bacteria.
Intravenous administration of the compounds protects micefrom Staphylococcus aureus infection. To begin to investigatethe efficacy of the compounds via an i.v. administration route,Swiss Webster mice were infected via i.p. injection with S.aureus (Smith strain), and after 15 min, treatment was initi-ated. A single i.v. injection of the compounds protected at least80% of the mice from death due to S. aureus infection (Fig.6E). These results indicate that i.v. administration of the com-pounds could be considered an alternative administration ap-proach for treatment with these antibacterial compounds.
DISCUSSION
The development of resistance to clinically important anti-biotics in bacterial pathogens and potential biowarfare agentsposes a major threat to public health (1, 6, 19, 21). Further-more, it is clear that resistance is more likely when newlyintroduced antibiotics are chemically similar to those that arealready ineffective. Therefore, new antimicrobial compounds,possessing novel scaffolds and unique mechanisms of action,are urgently needed to combat this growing incidence of anti-bacterial-resistant strains in the clinic (21).
In this study we describe the identification and initial char-
acterization of novel bis-(imidazolinylindole) compounds withpotent antibacterial activities. Four potent inhibitors, MBX1066, MBX 1090, MBX 1113, and MBX 1128, were identifiedin a cell-based B. anthracis infection assay of a focused libraryof diarylamidines. One member of this set, MBX 1090, hasbeen reported previously to display antibacterial activity (2),but no further investigations of mechanism or activity in animalinfection models have been described. In this report, we de-scribe in vivo studies demonstrating that two of the most po-tent compounds could protect mice following challenge with B.anthracis, a gram-positive spore-forming bacterium; Staphylo-coccus aureus, a gram-positive coccus; and Y. pestis, a gram-negative ovoid bacillus that is a facultative intracellularorganism.
The results described in this study offer some clues regardingthe mechanism of action of these compounds. They are potentinhibitors of DNA synthesis (Fig. 4B). While the moleculartarget of these compounds is not known, the fact that theyshare some structural features with compounds that bind in theminor grove of duplex DNA (7) suggests that these compoundsmay inhibit DNA synthesis by binding to DNA. However, theefficacy of MBX 1066 and MBX 1090 in live animal models ofinfection, together with favorable selectivity indices in vitro,suggests that these compounds prefer bacterial DNA overmammalian DNA. Similar moderate levels of species selectiv-ity have been observed previously for other compounds with alikely DNA binding mode of action, possibly resulting from apreference for AT-rich DNA (8, 15). While preliminary exper-iments indicate that MBX 1066 and MBX 1090 also preferAT-rich DNA (unpublished observations), it is unclear whetherfurther optimization of the structures can provide sufficient selec-tivity for clinical studies.
In contrast to MBX 1066 and MBX 1090, the other twocompounds, MBX 1113 and MBX 1128, exhibit some inhibi-tion of RNA and cell wall biosynthesis as well as DNA synthe-sis. While all four compounds exhibit detectable cytotoxicity ina stringent 3-day incubation with HeLa cells, compared to theirantibacterial activity, MBX 1066 and MBX 1090 exhibit higherselectivity indices (CC50/MIC, �15) than do the other twocompounds. These observed differences in macromolecularsynthesis specificity and selectivity indices could be related tostructural differences between these two pairs of compounds.
FIG. 5. Serial passage of S. aureus NCTC 8325 in MBX 1066, MBX 1090, and MBX 1113 selecting for increased resistance. The highestsublethal concentration of compound (fold MIC) is plotted versus the number of days of serial passage for eight independent cultures (A to H)for each compound.
VOL. 53, 2009 CHEMOTYPE WITH BROAD-SPECTRUM ANTIBACTERIAL ACTIVITIES 4289

The indole groups of MBX 1066 and MBX 1090 face eachother in a symmetrical fashion, while they are positioned in atandem arrangement in the other two compounds. Furtherstudies will be required to determine if this is an importantfeature for selective antibacterial activity. Attempts to selectmutants resistant to three of the compounds (MBX 1066,MBX 1090, and MBX 1113) were successful only for MBX1090. Furthermore, the MBX 1090-resistant mutants were notcross-resistant to MBX 1066, indicating that these two relatedcompounds do not share this mechanism of resistance. In sum-mary, the indications that MBX 1066 protects mice from lethalinfections with B. anthracis, Y. pestis, and S. aureus and doesnot readily select resistant mutations suggest that it representsa new antibacterial chemotype worthy of further explorationfor use against drug-resistant bacterial pathogens.
ACKNOWLEDGMENTS
We thank Jon Goguen, University of Massachusetts Medical School,for providing us with the attenuated Y. pestis strain (KIM �pgmpCD1�); Melani Ulrich, Clemson University, for providing the clinicalisolates of the listed bacterial strains; and Henry Heine, USAMRIID,for the ciprofloxacin-resistant strains of B. anthracis and helpful dis-cussions.
This project has been funded in part by HDTRA1-06-C-0042 and bythe Defense Threat Reduction Agency (R.G.P.) and with federal fundsfrom the National Cancer Institute, National Institutes of Health,under contract N01-CO-12400. This research was supported in part bythe Developmental Therapeutics Program in the Division of CancerTreatment and Diagnosis of the National Cancer Institute.
The content of this publication does not necessarily reflect the viewsor policies of the Department of Health and Human Services, nor doesmention of trade names, commercial products, or organizations implyendorsement by the U.S. government. Opinions, interpretations, con-
FIG. 6. Identified lead compounds protect mice against B. anthracis, Y. pestis, and S. aureus infection. C57BL/6 mice (n � 10) were infectedvia i.p. route with B. anthracis Ames spores (A to C) or Y. pestis CO92 strain (D), or Swiss Webster mice (n � 10) were infected i.p. with S. aureus(Smith) (E). (A) At 6 h postchallenge, the mice were treated with DMSO control (1%) or the indicated concentrations of MBX 1066. (B) At 6 hpostchallenge, the mice were treated with DMSO control (1%) or the indicated concentrations of MBX 1090. (C) Treatment with the compoundMBX 1066 (10 mg/kg/injection) was initiated at 6, 12, 18, and 24 h postchallenge. (D) At 6 h postchallenge with Y. pestis, mice were treated withDMSO control (1%) or the indicated concentrations of MBX 1066. In all studies, treatment with compound was every 6 h for 5 days. (E) At 15min postchallenge, mice were treated i.v. with the indicated doses of MBX 1066, MBX 1090, daptomycin, or vehicle control. Survival of mice wasmonitored for 48 h.
4290 PANCHAL ET AL. ANTIMICROB. AGENTS CHEMOTHER.

clusions, and recommendations are those of the authors and are notnecessarily endorsed by the U.S. Army.
REFERENCES
1. Alanis, A. J. 2005. Resistance to antibiotics: are we in the post-antibiotic era?Arch. Med. Res. 36:697–705.
2. Anne, J., E. De Clercq, H. Eyssen, and O. Dann. 1980. Antifungal andantibacterial activities of diarylamidine derivatives. Antimicrob. Agents Che-mother. 18:231–239.
3. Athamna, A., M. Massalha, M. Athamna, A. Nura, B. Medlej, I. Ofek, D.Bast, and E. Rubinstein. 2004. In vitro susceptibility of Bacillus anthracis tovarious antibacterial agents and their time-kill activity. J. Antimicrob. Che-mother. 53:247–251.
4. Baillie, L. W. 2006. Past, imminent and future human medical countermea-sures for anthrax. J. Appl. Microbiol. 101:594–606.
5. Balzarini, J., E. de Clercq, and O. Dann. 1983. Inhibitory activity of diaryla-midine derivatives on murine leukemia L1210 cell growth. Investig. NewDrugs 1:103–115.
6. Barrett, C. T., and J. F. Barrett. 2003. Antibacterials: are the new entriesenough to deal with the emerging resistance problems? Curr. Opin. Biotech-nol. 14:621–626.
7. Brosh, R. M., Jr., J. K. Karow, E. J. White, N. D. Shaw, I. D. Hickson, andV. A. Bohr. 2000. Potent inhibition of werner and bloom helicases by DNAminor groove binding drugs. Nucleic Acids Res. 28:2420–2430.
8. Burli, R. W., J. A. Kaizerman, J. X. Duan, P. Jones, K. W. Johnson, M.Iwamoto, K. Truong, W. Hu, T. Stanton, A. Chen, S. Touami, M. Gross, V.Jiang, Y. Ge, and H. E. Moser. 2004. DNA binding ligands with in vivoefficacy in murine models of bacterial infection: optimization of internalaromatic amino acids. Bioorg. Med. Chem. Lett. 14:2067–2072.
9. Butler, M. M., W. A. Lamarr, K. A. Foster, M. H. Barnes, D. J. Skow, P. T.Lyden, L. M. Kustigian, C. Zhi, N. C. Brown, G. E. Wright, and T. L. Bowlin.2007. Antibacterial activity and mechanism of action of a novel anilinouracil-fluoroquinolone hybrid compound. Antimicrob. Agents Chemother. 51:119–127.
10. Chastre, J. 2008. Evolving problems with resistant pathogens. Clin. Micro-biol. Infect. 14(Suppl. 3):3–14.
11. David, M. Z., D. Glikman, S. E. Crawford, J. Peng, K. J. King, M. A.Hostetler, S. Boyle-Vavra, and R. S. Daum. 2008. What is community-asso-ciated methicillin-resistant Staphylococcus aureus? J. Infect. Dis. 197:1235–1243.
12. Diep, B. A., G. G. Stone, L. Basuino, C. J. Graber, A. Miller, S. A. Etages, A.Jones, A. M. Palazzolo-Ballance, F. Perdreau-Remington, G. F. Sensabaugh,F. R. Deleo, and H. F. Chambers. 2008. The arginine catabolic mobile
element and staphylococcal chromosomal cassette mec linkage: convergenceof virulence and resistance in the USA300 clone of methicillin-resistantStaphylococcus aureus. J. Infect. Dis. 197:1523–1530.
13. Doganay, M., and N. Aydin. 1991. Antimicrobial susceptibility of Bacillusanthracis. Scand. J. Infect. Dis. 23:333–335.
14. Friedlander, A. M. 1986. Macrophages are sensitive to anthrax lethal toxinthrough an acid-dependent process. J. Biol. Chem. 261:7123–7126.
15. Hu, W., R. W. Burli, J. A. Kaizerman, K. W. Johnson, M. I. Gross, M.Iwamoto, P. Jones, D. Lofland, S. Difuntorum, H. Chen, B. Bozdogan, P. C.Appelbaum, and H. E. Moser. 2004. DNA binding ligands with improved invitro and in vivo potency against drug-resistant Staphylococcus aureus.J. Med. Chem. 47:4352–4355.
16. Hughes, J. M., and J. L. Gerberding. 2002. Anthrax bioterrorism: lessonslearned and future directions. Emerg. Infect. Dis. 8:1013–1014.
17. Inglesby, T. V., T. O’Toole, D. A. Henderson, J. G. Bartlett, M. S. Ascher, E.Eitzen, A. M. Friedlander, J. Gerberding, J. Hauer, J. Hughes, J. McDade,M. T. Osterholm, G. Parker, T. M. Perl, P. K. Russell, and K. Tonat. 2002.Anthrax as a biological weapon, 2002: updated recommendations for man-agement. JAMA 287:2236–2252.
18. Marshall, N. J., C. J. Goodwin, and S. J. Holt. 1995. A critical assessment ofthe use of microculture tetrazolium assays to measure cell growth and func-tion. Growth Regul. 5:69–84.
19. Monroe, S., and R. Polk. 2000. Antimicrobial use and bacterial resistance.Curr. Opin. Microbiol. 3:496–501.
20. National Committee for Clinical and Laboratory Standards (NCCLS). 2003.Methods for dilution antimicrobial susceptibility tests for bacteria that growaerobically. Approved standard, M7-A6. National Committee for ClinicalLaboratory Standards, Wayne, PA.
21. Pathania, R., and E. D. Brown. 2008. Small and lethal: searching for newantibacterial compounds with novel modes of action. Biochem. Cell Biol.86:111–115.
22. Rodríguez de Castro, F., O. R. Naranjo, J. A. Marco, and J. S. Violan. 2009.New antimicrobial molecules and new antibiotic strategies. Semin. Respir.Crit. Care Med. 30:161–171.
23. Tidwell, R. R., J. D. Geratz, O. Dann, G. Volz, D. Zeh, and H. Loewe. 1978.Diarylamidine derivatives with one or both of the aryl moieties consisting ofan indole or indole-like ring. Inhibitors of arginine-specific esteroproteases.J. Med. Chem. 21:613–623.
24. Woese, C. R., J. C. Vary, and H. O. Halvorson. 1968. A kinetic model forbacterial spore germination. Proc. Natl. Acad. Sci. USA 59:869–875.
25. Wu, M., and R. E. Hancock. 1999. Interaction of the cyclic antimicrobialcationic peptide bactenecin with the outer and cytoplasmic membrane.J. Biol. Chem. 274:29–35.
VOL. 53, 2009 CHEMOTYPE WITH BROAD-SPECTRUM ANTIBACTERIAL ACTIVITIES 4291
Copyright © 2022 FDOKUMEN




















