
DISTRIBUTION, DIVERSITY, AND ACTIVITY OF ANTIBIOTIC ...
173
DISTRIBUTION, DIVERSITY, AND ACTIVITY OF ANTIBIOTIC-PRODUCING PSEUDOMONAS SPP. JORGE T. DE SOUZA
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of DISTRIBUTION, DIVERSITY, AND ACTIVITY OF ANTIBIOTIC ...

DISTRIBUTION, DIVERSITY, AND ACTIVITY OF
ANTIBIOTIC-PRODUCING PSEUDOMONAS SPP.
JORGE T. DE SOUZA

Promotor: Prof. Dr. ir. P.J.G.M. de WitHoogleraar in de FytopathologieWageningen Universiteit
Co-promotor: Dr. J. M. RaaijmakersUniversitair docent bij het Laboratorium voor FytopathologieWageningen Universiteit
Promotiecommissie: Prof. Dr. ir. A. H. C. van Bruggen, Universiteit WageningenDr. P. Lemanceau, INRA, Dijon, FranceProf. Dr. ir. L. C. van Loon, Universiteit UtrechtProf. Dr. J. A. van Veen, Universiteit Leiden

JO���
R� G� E �
T. D���
E �
S� O�
U�
Z
A�
D�
I�S�T�
R�
I�B�
U�
T�
I�O�
N�
, � D�
I�V�
E�
R�
S�I�T�
Y�
, � A�
N�
D �
A�
C�
T�
I�V�
I�T�
Y �
O�
F�
A�
N�
T�
I�B�
I�O�
T�
I�C�
-� P R�
O�
D�
U�
C�
I�N�
G !
P"
S#E$
U% D&
O' M(
O' N)
A*
S# S+PP,-,
..
P/ r0 oe1�2 f3 s4 c5 h6 r0 i7 f3 t8ter verkrijging van de graad van doctorop gezag van de rector magnificus van
Wageningen Universiteit,prof. dr. ir. L. Speelman
in het openbaar te verdedigenop dinsdag 1 oktober 2002
des namiddags te vier uur in de Aula.

Distribution, diversity, and activity of antibiotic-producing Pseudomonas spp.J.T. de Souza. 2002.
Thesis Wageningen University, The Netherlands.With references – with summaries in English, Portuguese, and Dutch.
ISBN 90-5808-727-1

L9
i:s; t < o= f
>a? b@ b
@rA eB vC i
:a? t< i: o= nD s;
2,4-DAPG 2,4-diacetylphloroglucinolaPCR anchored polymerase chain reactionCFU colony forming unitCMC critical micelle concentrationCTAB cetyltrimethylammoniumbromideGC-FAME gas chromatograph - fatty acid methyl esterGgt Gaeumannomyces graminis var. triticiHPLC high performance liquid chromatographyIPTG isopropylthio-β-D-galactosidaseKMB King’s medium BLC-MS liquid chromatography-mass spectrometryMAPG monoacetylphloroglucinolMeCN acetonitrileN-AHLs N-acylhomoserine lactonesPCA phenazine-1-carboxilic acidPCN phenazine-1-carboxamidePG phloroglucinolPHZ phenazinesPKS polyketide synthasePLT pyoluteorinPRN pyrrolnitrinRAPD random amplified polymorphic DNARFLP random fragment length polymorphismRP-HPLC reverse phase-high performance liquid chromatographyTAD take-all declineTAPG tri-acetylphloroglucinolTEM transmission electron microscopyTLC thin-layer chromatographyX-gal 5-bromo-4-chloro-3-indolyl-β-D-galactosidase


CE OF NG
TH
EI
NG
TH
SJ
Chapter 1 General Introduction and Outline 1
Chapter 2 Polymorphisms within the prnD and pltC genes frompyrrolnitrin and pyoluteorin-producing Pseudomonas andBurkholderia spp.
31
Chapter 3 Conservation of the response regulator gene gacA inPseudomonas species
55
Chapter 4 Frequency, diversity and activity of 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. in dutch take-all declinesoils
75
Chapter 5 Effect of 2,4-diacetylphloroglucinol on Pythium: ultrastructuralinvestigation of cellular responses and variation in sensitivityamong propagules and species
99
Chapter 6 Activity of biosurfactants produced by Pseudomonas fluorescensagainst oomycete pathogens
121
Chapter 7 Summarising Discussion 143
Summary 151
Resumo 155
Samenvatting 159
Acknowledgements 163
About the Author 165


___________________________________________________
CHAPTER 1
GK
EL
NM
EL
RN
AO
L P
IQNM
TR
RN
OS
DT
UU
CV
TR
IQOS
N M
AO
NM
D T
OS
UU
TR
LP
IQNM
EL
___________________________________________________________________________

CW hX
aY ptZ\[ e] r ^ 1: _a` Gb e] nc e] raY l d Ie nc t[ rof dg
uh ci t[ ij of n c aY nc d g
Ok uh t[ ld ij nc e]
2
Part of this chapter has been accepted for publication as: Al
nm tn io bp io oq tn io c r ps rt oq du uv cr tn io oq n m bp y w bp ax cr tn ey rt io ax l z bp io oq cr oq nm tn rt oq l z ax g{ ey nm tn s| , byRaaijmakers, J.M., Vlami, M., and De Souza, J.T. in Antonie van Leeuwenhoek (in press).
A}
n~ t� i� b� i� o� t� i� c� -� p� r� o� d� u� c� i� n~ g � P�
s� e� u� d� o� m� o� n� a� s� s� pp��� .�
Bacteria of the genus Pseudomonas are able to survive and prosper in a wide range of
environmental conditions. This genus, not only contains plant, animal, and human pathogens,
but also accomodates species of environmental interest, such as plant growth promoters,
xenobiotic degraders, and biocontrol agents (Palleroni, 1992; Johnsen et al., 1996). Among
the biocontrol agents, the antibiotic-producing strains have received considerable attention.
Antibiotics encompass a chemically heterogeneous group of organic, low-molecular
weight compounds. At low concentrations, antibiotics are deleterious to the growth or
metabolic activities of other microorganisms (Fravel, 1988; Thomashow et al., 1997). The fact
that there is an abundant amount of studies on antibiotics produced by Pseudomonas spp. has
several reasons: pseudomonads are common inhabitants of the rhizosphere and phyllosphere,
are easily isolated from natural environments, utilize a wide range of substrates, are easy to
culture and manipulate genetically, making them more amenable to experimentation
(Leisinger and Margraff, 1979; Whipps, 2001).
F�
i�g� u� r� e � 1. ��� C
�h
e� m¡ i�c¢ a£ l ¤ s¥ t¦ r� u� c¢ t¦ u� r� e � o§ f
¨s¥ o§ m¡ e � a£ n© t¦ i� bª i
�o§ t¦ i� c¢ s ¥ p« r� o§ d
¬u� c¢ e� d
¬bª
y P®
s¯ e° u± d² o³ mo³ nµ a¶ s¯ s¥ p« e� c¢ i� e� s¥ .� In the primarystructure of amphisin, Dec is hydroxydecanoic acid, Leu leucine, Asp aspartic acid, Thrthreonine, Ser serine, Gln glutamine, and Ile isoleucine.
Pyrro lnitrin
OOH
OH
Cl
NO2
N H
C l
Pyoluteorin
OH
C
OH
C
HO
OO
C H 3H 3C
2,4-D iacetylphloroglucinol
N
N H
COOH
H
H
NH Cl
C l
Phenaz ine-1-carboxilic acid
(C H 2)6
C H 3
O O C H
OHHO
H O
H 3CC H 2 C O C H CH 2
O
COOH
Rhamnolipid 1
Amphisin
O L-Asp-L -Ile-L -Leu-D-Gln
L-Leu3(R)-OH-Dec-D -Leu-D -Asp-D-allo -Thr-D-Leu-D-Leu-D-Ser-

C· h¸
a¹ ptº\» e¼ r ½ 1: ¾a¿ GÀ e¼ nÁ e¼ r½ a¹ l  Ià nÁ t» r½ oÄ dÅ
uÆ cÇ t» iÈ oÄ n Á a¹ nÁ d Å
OÉ uÆ t» l iÈ nÁ e¼
3
Pseudomonas biocontrol strains not only exhibit a wide range of diversity in the type
but also in the number of antibiotics produced (Fig. 1). For example, P. fluorescens strains
CHA0 and Pf5 (Keel et al., 1996; Bender et al., 1999) produce multiple antibiotics with
overlapping or different degrees of activity against specific pathogens. This fact illustrates that
for at least some biocontrol agents several antibiotics may account for the suppression of
specific or multiple plant diseases. In this introductory chapter we will concentrate only on
antibiotics produced by Pseudomonas, excluding siderophores and enzymes. Our attention will
focus on the biosynthesis, regulation and activity of following antibiotics: 2,4-
diacetylphloroglucinol (2,4-DAPG), phenazines (PHZ), pyrrolnitrin (PRN), pyoluteorin
(PLT), and surface-active antibiotics. These five classes of antibiotics, in particular 2,4-DAPG,
are subject of experiments described in this thesis.
2,4ÊÌËÎÍ -Ï dÐ iÑ aÒ cÓ eÔ tÕ yÖ l× pØ hÙ l× oÚ rÛ oÚ gÜ l× uÝ cÓ iÑ nÞ oÚ l × (ß 2,4ÊÌËÎÍ -Ï Dà Aá
Pâ Gã )äPhloroglucinols are phenolic compounds produced by bacteria, algae, and plants
(Reddi and Borovkov, 1969; Verrota et al., 1999; Jimenez-Escrig et al., 2001). More than 60
derivatives of phloroglucinol (PG) have been reported, but only three,
monoacetylphloroglucinol (MAPG), 2,4-DAPG, and tri-acetylphloroglucinol (TAPG) are
known to be produced by Pseudomonas species (Reddi and Borovkov, 1969; Shanahan et al.,
1992). In addition to MAPG and 2,4-DAPG, also PG has been isolated from cultures of a
bacterium classified as Aeromonas hydrophila (Strunz et al., 1978).
The antimicrobial activity of these compounds, and in particular of 2,4-DAPG has
received considerable attention in the area of biological control of plant diseases. Numerous
studies have demonstrated that 2,4-DAPG-producing Pseudomonas spp. can suppress a wide
variety of plant pathogens, including fungi, bacteria and nematodes (Stutz et al., 1986; Keel et
al., 1990; Vincent et al., 1991; Fenton et al., 1992; Levy et al., 1992; Keel et al., 1992; Cronin
et al. 1997a, b; Duffy and Défago, 1997; Sharifi-Tehrani et al., 1998). Among the fungi,
Gaeumannomyces graminis on wheat, Thielaviopsis basicola on tobacco, Fusarium oxysporum f.
sp. radicis-lycopersici on tomato, and the oomycete Pythium ultimum on sugar beet and
cucumber have been shown to be controlled by 2,4-DAPG-producing Pseudomonas strains
(Vincent et al., 1991; Fenton et al., 1992; Sharifi-Tehrani et al., 1998; Stutz et al., 1986;
Duffy and Défago, 1997). The determinative role of 2,4-DAPG in disease suppression by
Pseudomonas strains has been demonstrated by (1) the use of mutants deficient in 2,4-DAPG
production (Vincent et al., 1991; Shanahan et al., 1992; Keel et al., 1992; Cronin et al.,

Cå hæ
aç ptè\é eê r ë 1: ìaí Gî eê nï eê rë aç l ð Iñ nï té rë oò dó
uô cõ té iö oò n ï aç nï d ó
O÷ uô té lð iö nï eê
4
1997a, b), (2) complementation of 2,4-DAPG- deficient mutants and subsequent restoration
of biocontrol activity (Vincent et al., 1991; Keel et al., 1992; Cronin et al., 1997a, b), and (3)
expression of 2,4-DAPG biosynthetic genes in heterologous, nonproducing strains conferring
biocontrol or enhanced activity (Vincent et al., 1991; Shanahan et al., 1993; Bangera and
Thomashow, 1996). Reporter gene systems (Loper and Lindow, 1997) and analytical
techniques (Thomashow et al., 1997) have further demonstrated that 2,4-DAPG is produced
in situ by both introduced strains and indigenous Pseudomonas populations (Keel et al., 1992;
Bonsall et al., 1997; Duffy and Défago, 1997; Raaijmakers et al., 1999; Notz et al., 2001).
Although 2,4-DAPG production is essential in the biocontrol activity of several Pseudomonas
strains, it does not seem to play a determinative role in the survival and competitive ability of
the producing strains (Carroll et al., 1998; Chapter 4).
Despite the well-documented effects of 2,4-DAPG, its mode of action is still largely
unclear. It was found that phloroglucinol derivatives are potent inhibitors of the photosystem
II (Yoneyama et al., 1990), however, little is known about its antibiotic mode of action against
fungi and oomycetes. It has been suggested that 2,4-DAPG could also act as an inducer of the
plant’s defense mechanisms against pathogens. This assumption is based on the fact that
herbicides, which are structurally similar to 2,4-DAPG, can induce resistance in plants
(Altman and Campbell, 1977; Cohen et al., 1986). We demonstrated that 2,4-DAPG does
not affect the synthesis and composition of the cell wall of the oomycete Pythium, and that its
deleterious action may reside in detrimental effects on the plasma membrane (Chapter 5).
Bø iö oò sù yú nï té hæ eê sù iö s ù aç nï d ó
rë egêüû uô lðaç té iö oò n ï oò f 2,4-D
ýÌþ ÿ �����A�
P� GîThe biosynthetic locus responsible for the synthesis of 2,4-DAPG by Pseudomonas
fluorescens strain Q2-87 is composed of six genes (Bangera and Thomashow, 1996, 1999). The
genes phlACBD constitute the biosynthetic operon and are flanked by phlE and phlF, which
code for a putative transport and a repressor protein, respectively (Fig. 2). The gene phlD
encodes a polyketide synthase responsible for the production of MAPG through condensation
of an unknown precursor (Bangera and Thomashow, 1999). Genes phlA, phlC, and phlB
encode proteins necessary for the acetylation of MAPG to 2,4-DAPG and may also function
in the synthesis of MAPG (Table 1). Many strains of Pseudomonas produce TAPG in
addition to 2,4-DAPG. Whether the known genes are also involved in the acetylation of 2,4-
DAPG to TAPG remains to be determined.
The role of several environmental conditions influencing 2,4-DAPG production have
been studied for a number of Pseudomonas strains (Shanahan et al., 1992; Duffy and Défago,

C� h
a pt� � e� r � 1: ��� G� e� n� e� r� a l �
I� n� t� r� o� d�
u� c� t� i� o� n � a n� d �
O� u� t� l� i� n� e�
5
T�
a� b� l e ! 1. "$# O
%p& e! n ' r( e! a� d
)i*n' g + f
,r( a� m- e! s . (/ O% R
0F1
s. ) 2 i*n ' t3 h4 e ! 2,4
5$687-9 D: A
;P<
G =
b�
i*o> s. y? n' t3 h4 e! t3 i* c @ g+ e! n' e ! c@ l uA s. t3 e! r( . # (Bangera and
Thomashow, 1996, 1999).
ORF Size (bp) Homology Possible Function
phlA 1080 acyl carrier protein synthase conversion of MAPG to 2,4-DAPGphlC 1194 sterol carrier protein conversion of MAPG to 2,4-DAPGphlB 438 unknown conversion of MAPG to 2,4-DAPGphlD 1047 plant chalcone and stilbene synthases synthesis of MAPGphlE 1269 efflux protein efflux of MAPG and 2,4-DAPGphlF 606 repressor protein repressor of 2,4-DAPG
F1
i*g+ uA r( e ! 2.
5 # O%
r( g+ a� n' i*zB a� t3 i* o> n ' o> f
,t3 h4 e ! 2,4
5C6D7-9 D: A
;P<
G =
g+ e! n' e ! c@ l uA s. t3 e! r ( i
*n ' PE
. F fGluHJI
oK rL eM sN cO eM nP sN QQ 25
-9 87.R$S # Arrows indicate direction
of transcription. (Bangera and Thomashow, 1996, 1999). Not drawn to scale.
1999; Notz et al., 2001). These studies demonstrated that the regulation of this antibiotic
through physiological factors may vary among strains. For example, 2,4-DAPG production
was stimulated by glucose in many P. fluorescens strains, including strain CHA0, and repressed
in strain F113 (Shanahan et al., 1993; Duffy and Défago, 1997). In P. fluorescens S272, 2,4-
DAPG production was stimulated by ethanol as a sole carbon source (Yuan et al., 1998).
Amendment of the growth medium with zinc sulfate and ammonium molybdate stimulates,
whereas addition of inorganic phosphate inhibits 2,4-DAPG production. Fusaric acid, a toxin
secreted by Fusarium oxysporum f. sp. radicis-lycopersici repressed production of 2,4-DAPG by
P. fluorescens CHA0, resulting in inefficient performance of the biocontrol agent (Duffy and
Défago, 1997). Studies employing the lacZ reporter gene fused to phlA, the first gene in the
2,4-DAPG biosynthetic operon (Fig. 2), showed that transcription of phlA in strain CHA0 is
specifically stimulated by 2,4-DAPG and to a lesser extent by MAPG. Pyoluteorin and
salycilate, other metabolites produced by the bacterium, repressed transcription of phlA
(Schnider-Keel et al., 2000). 2,4-DAPG production is, among others, regulated by phlF, a
pathway-specific repressor gene located in the 2,4-DAPG biosynthetic locus (Fig. 2). This
gene was found to encode a DNA-binding protein that represses production of the antibiotic
at the transcriptional level. The protein was proposed to play an important role in preventing
2,4-DAPG production in early stages of growth, when the antibiotic could be detrimental to
the cell (Delany et al., 2000). The PhlF protein was also found to act as a mediator in the
phlE phlD phlB phlC phlA phlF

CT hU
aV ptW X eY r Z 1: [�\ G] eY neY rZ aV l _
IntX rZ oa db
uc cd tX ie oa n ^ aV nd b
Of uc tX l_ ie neY
6
autoinduction of 2,4-DAPG production and repression by pyoluteorin and salicylate. Mutants
disrupted in phlF were insensitive to autoinduction by 2,4-DAPG and to repression of phlA by
pyoluteorin and salicylate (Schnider-Keel et al., 2000).
Pg hh ei nj ak zl im nj ei s n (o Pg Hp Zq )r
PHZs are nitrogen-containing compounds naturally produced exclusively by bacteria
(Turner and Messenger, 1986). These antibiotics are very diverse, with more than 50
derivatives described. It is common to find several different strains producing the same PHZ
compound and a single strain producing a variety of PHZs (Turner and Messenger, 1986).
PHZs were found to be toxic to fungi, bacteria, and algae (Toohey et al., 1965).
The proposed modes of action of PHZs include cell death or injury due to their
capacity to undergo redox cycling in presence of several reducing agents and molecular oxygen,
leading to the accumulation of toxic oxygen species (Hassan and Fridovich, 1980). Some
PHZs were shown to intercalate into DNA strands, particularly with GC base pairs, to inhibit
RNA synthesis (Turner and Messenger, 1986), and to disrupt electron transport and energy
production (Baron et al., 1989). Given the chemical diversity of PHZs, it is unlikely that a
unifying mode of action among them exists (Turner and Messenger, 1986). PHZs produced
by bacteria play important roles in virulence in humans and animals, biological control, and
competitiveness. The PHZ compound pyocyanine produced by P. aeruginosa is a virulence
factor in the infection of cystic fibrosis patients (Wilson et al., 1987). P. fluorescens 2-79
produces phenazine-1-carboxilic acid (PCA) and P. aureofaciens 30-84 produces three PHZ
compounds: PCA, 2-hydroxy-phenazine-1-carboxilic acid, and 2-hydroxy-phenazine. These
PHZs were shown to play an important role in the competitive ability of the producing strains
in the rhizosphere (Mazzola et al., 1992).
PHZs also play a key role in biocontrol of several plant diseases. Mutants disrupted in
PHZ biosynthesis could no longer control G. graminis var. trtici and Septoria tritici on wheat
(Thomashow and Weller, 1988; Flaishman et al., 1990; Pierson III and Thomashow, 1992),
Fusarium oxysporum f.sp. ciceris on chickpea and Pythium splendens on beans and lettuce
(Anjaiah et al., 1998), and F. oxysporum f.sp. radicis-lycopersici on tomato (Chin-A-Woeng et
al., 1998). Mutants genetically restored in their ability to produce PHZs regained their ability
to inhibit fungal growth on artificial medium and to control the disease in planta
(Thomashow and Weller, 1988; Pierson III and Thomashow, 1992). Further evidence for the
role of PHZs in biocontrol was provided by studies with reporter genes showing expression of

Cs ht
au ptv w ex r y 1: z�{ G| ex n} ex ry au l ~
I� n} tw ry o� d�
u� c� tw i� o� n } au n} d �
O� u� tw l~ i� n} ex
7
the PHZ biosynthetic operon in the rhizosphere (Chin-A-Woeng et al., 1998) and its direct
detection on roots of wheat treated with PHZ-producing strains (Thomashow et al., 1990).
B� i� o� s� y� n} tw ht ex s� i� s � au n} d �
ry egx�� u� l~au tw i� o� n } o� f
�P� H� Z
�s�
The precise mechanisms of PHZ biosynthesis and the identity of all intermediates are
still unknown, despite the amount of studies already performed. PHZs are derived from the
shikimic acid pathway with chorismate as a probable branch point intermediate (Longley et
al., 1972). Phenazine-1,6-dicarboxylic acid was proposed as a common precursor to all other
PHZs (Byng and Turner, 1977). Recent studies, however, indicate that phenazine-1-
carboxylic acid is the common precursor (Mavrodi et al., 2001). Genes involved in PHZ
biosynthesis were studied in P. aureofaciens 30-84 (Pierson III et al., 1995), P. fluorescens 2-79
(Mavrodi et al., 1998), P. chlororaphis PCL1391 (Chin-A-Woeng et al., 2001), and P.
aeruginosa PAO1 (Mavrodi et al., 2001). Seven genes were identified in these strains for
biosynthesis of PCA. Initially, Pierson III et al. (1995) identified five genes, phzCDEFG
(originally phzFABCD) from P. aureofaciens 30-84 for synthesis of PCA. The products of
phzD and phzE, which have homology with isochorismatase and anthranilate synthase,
respectively (Table 2), were hypothesized to catalyse the conversion of chorismate to
phenazine-1,6-dicarboxylic acid (Pierson III and Thomashow, 1992). PhzF and PhzG were
proposed to act in the conversion of phenazine-1,6-dicarboxylic acid to PCA. The proteins
PhzA and PhzB were thought to act as stabilizers of a complex formed by PhzD and PhzE,
which still work in the absence of PhzA and PhzB, but the specificity and efficiency decreases
dramatically (Mavrodi et al., 1998). In P. aeruginosa PAO1, two complete operons,
phzA1B1C1D1E1F1G1 and phzA2B2C2D2E2F2G2 for PCA production were identified.
These two PHZ operons are 98.3% identical at the DNA level and are homologous to
the PHZ operons of P.aureofaciens 30-84, P. fluorescens 2-79, and P. chlororaphis PCL 1391.
A screen of 30 other PHZ-producing strains by using the phzA1B1C1D1E1F1G1 from P.
aeruginosa PAO1 (Fig. 3) as a probe showed that the PHZ genes are highly conserved among
Pseudomonas species and are different from the PHZ genes of Burkholderia species (Mavrodi et
al., 2001). According to the mechanism proposed for biosynthesis of PHZs, the operons
phzA1B1C1D1E1F1G1 and phzA2B2C2D2E2F2G2 are responsible, by yet undefined
intermediates, for the conversion of chorismic acid to PCA. The PHZ modifying genes phzM,
phzS, and phzH further catalyze the conversion of PCA to other PHZs (Mavrodi et al., 2001,
Table 2).
PHZ antibiotics are only produced during late exponential growth phase and in the

C� h�
a� pt� � e� r � 1: ��� G� e� n� e� r� a� l �
I� n� t� r� o� d�
u� c� t� i o� n � a� n� d �
O¡ u� t� l� i n� e�
8
T¢
a£ b¤ l¥e ¦ 2. §©¨
Oª
p« e¦ n ¬ r e¦ a£ d® i¯n¬ g ° f
±r a£ m² e¦ s ³ (O
ªRµ
F¶
s³ ) · r e¦ s³ p« o¸ n¬ s³ i¯ b¤ l¥e ¦ f±o¸ r P
¹Hº
Z »
b¤
i¯o¸ s³ y¼ n¬ t½ h¾ e¦ s³ i¯ s ³ i
¯n ¬ P
¿. À aÁ e rà uÄ gÅ i
ÆnÇ oÈ sÉ aÁ P
¹AÊ
Oª
1.Ë ¨(Mavrodi et al., 2001) and PAO1 genome (http://www.pseudomonas.com).
ORF a Size (bp) Homology b Possible Function c
phzA1 489 phzA homologs, phzX synthesis of PCAphzB1 489 phzB homologs, phzY synthesis of PCAphzC1 1218 plant DAHP synthases synthesis of PCAphzD1 624 isochorismatases synthesis of PCAphzE1 1884 bacterial amidotransferases synthesis of PCAphzF1 837 phzF, and phzC homologs synthesis of PCAphzG1 645 bacterial pyridoxamine-5’-phosphate oxidases synthesis of PCArhlI 606 autoinducer proteins quorum sensing regulationrhlR 726 transcriptional regulator quorum sensing regulationphzM 1005 O-methyltransferase conversion of PCA to 5-CH-PCAphzS 1209 bacterial mono-oxygenases conversion of PCA to 1-OH-PHZ
and 5-CH-PCA to pyocyaninephzH 1833 bacterial asparagine synthetase conversion of PCA to PCN
a ORFs phzA1B1C1D1E1F1G1 comprise the core 1 biosynthetic operon of PAO1; rhlI and rhlR are quorumsensing regulators; phzM, phzS, and phzH are phenazine modifying genes.
b phzA, phzB, and phzF refer to homologs from P. fluorescens 2-79 and P. chlororaphis PCL1391; phzX, phzY,and phzC refer to homologs from P. aureofaciens 30-84; DAHP is phospho-2-keto-3-deoxyheptamate aldolase.
c PCA is phenazine-1-carboxylic acid, 5-CH-PCA is 5-methyl phenazine-1-carboxylic acid betaine; 1-OH-PCA is1-hydroxyphenazine, PCN is phenazine-1-carboximide.
F¶
i¯g° uÌ r e ¦ 3.
Í©¨Oª
r g° a£ n¬ i¯zÎ a£ t½ i¯ o¸ n ¬ o¸ f
±t½ h¾ e ¦ P
¹Hº
Z »
g° e¦ n¬ e ¦ cÏ l¥uÌ s³ t½ e¦ r i
¯n ¬ P¿
. À aÁ e rà uÄ gÅ iÆnÇ oÈ sÉ aÁ P
¹AÊ
Oª
1.Ë ¨ Arrows indicate direction oftranscription. (Mavrodi et al., 2001). Not drawn to scale.
stationary phase (Turner and Messenger, 1986). In natural environments, the expression of
the PHZ operon can be influenced by seed and root exudates of several plant species
(Georgakopoulos et al., 1994). Long-term studies indicated that the expression levels of the
PHZ operon are influenced by the developmental stage of wheat plants (Pierson III and
Pierson, 1996).
Regulation of PHZ production and many other genes in bacteria is done, among
others, through a process called quorum sensing (Fuqua et al., 1996). Quorum sensing is a
mechanism by which bacteria can communicate by using signal molecules, collectively known
as N-acylhomoserine lactones (AHLs). By using these signal molecules produced by themselves
or by other cells, bacteria can sense when a minimal unity or quorum of baterial cells has been
phzH phzM
rhlI rhlR phzA1 phzB1 phzC1 phzE1 phzF1 phzG1
phzS

CÐ hÑ
aÒ ptÓ Ô eÕ r Ö 1: ×�Ø GÙ eÕ nÚ eÕ rÖ aÒ l Û
IÜ nÚ tÔ rÖ oÝ dÞ
uß cà tÔ iá oÝ n Ú aÒ nÚ d Þ
Oâ uß tÔ lÛ iá nÚ eÕ
9
attained to exhibit multicellular behaviour in terms of gene expression (Fuqua et al., 1996).
Many genera of bacteria produce these diffusible molecules and regulate gene expression in a
cell density-dependent manner. In Pseudomonas, traits such as production of toxins, enzymes,
rhamnolipids, and PHZs are controlled by quorum sensing (Ochsner and Reiser, 1995).
Quorum sensing systems generally comprise two proteins, such as RhlI/RhlR in P. aeruginosa
PAO1 (Fig. 3). The autoinducer protein (RhlI) is responsible for the synthesis of AHLs, which
diffuse freely out and into the cell. When the intercellular AHL concentration reaches a
threshold, it interacts with the transcriptional regulator protein (RhlR), which in turn
regulates specific gene expression (Passador et al., 1993). Interestingly, only one of the two
PHZ operons in P. aeruginosa PAO1 appears to be controlled by quorum sensing. The
presence of two differentially regulated PHZ biosynthetic operons may give greater flexibility
in modulating PHZ production depending on the growth phase or response to environmental
signals (Mavrodi et al., 2001). In P. chlororaphis strain PCL1391, PhzI/PhzR (RhlI/RhlR
homologs) partially control PHZ production by quorum sensing. Expression levels of phzI are
positively regulated by yet unidentified factor(s) secreted into the growth medium of overnight
cultures. This positive regulation enables the strain to produce PCN at low cell densities and
this factor(s) was shown not to be AHLs (C4-HSL or C8-HSL) or PCN itself (Chin-A-Woeng
et al., 2001).
The regulation of PHZ production by quorum sensing in P. aeruginosa PAO1 is very
complex, involving also the LasI/LasR proteins, another set of autoinducer and transcriptional
regulator proteins (Pesci et al., 1997). These two quorum sensing systems do not act
independently of one another, but a hierarchy exists with LasR required for the activation of
rhlR and to some extent rhlI. Recently, a gene in P. chlororaphis PCL1391 which is
homologous to the lexA gene of P. aeruginosa was shown to be involved in the regulation of
PHZ (Chin-A-Woeng, 2000). Mutants disrupted in the lexA homolog exhibit a more than 10-
fold increase in PHZ production levels as compared to the wild type strain. The introduction
of a lexA mutation in phzI or phzR mutants restored production of PHZ to wild type levels
and resulted in the reappearance of AHLs, which were not present in the background phzI
mutant. The authors hypothesized that lexA represses a normally silent AHL synthase.

Cã hä
aå ptæ ç eè r é 1: ê�ë Gì eè ní eè ré aå l î
Iï ní tç ré oð dñ
uò có tç iô oð n í aå ní d ñ
Oõ uò tç lî iô ní eè
10
Pö y÷ rø rø où lú nû iü tý rø iü n û (þ Pö Rÿ N� )�
PRN (3-chloro-4-(2’-nitro-3’-chlorophenyl)-pyrrole) is a chlorinated phenylpyrrole
antibiotic produced by many bacterial genera, including Pseudomonas, Burkholderia,
Enterobacter, Serratia, and Myxococcus (Arima et al., 1964; Elander et al., 1968; Hammer et al.,
1999; Kalbe et al., 1996).
The mode of action of PRN is primarily the inhibition of the respiratory electron
transport system (Tripathi and Gottlieb, 1969). PRN has activity against several bacteria and
fungi, in particular Rhizoctonia solani (Arima et al., 1964; Cartwright et al., 1995; Chernin et
al., 1996; El-Banna and Winkelmann, 1988; Hill et al., 1994; Jayaswal et al., 1993; Rosales et
al., 1995). PRN is also effective against post-harvest diseases caused by Botrytis cinerea on
apple, pear and on cut flowers (Hammer and Evensen, 1993; Janisiewicz and Roitman, 1988),
and has been used to treat infections in humans by opportunistic fungi (Tawara et al., 1989).
The role of PRN in biological control has been demonstrated in studies where mutants of P.
fluorescens strains BL915 and Pf-5, defective in pyrrolnirin production, were unable to control
R. solani on cotton and Pyrenophora tritici-repentis in axenically colonized wheat straw (Hill et
al., 1994; Pfender et al., 1993). The antibiotic was also detected on wounded potatoes
colonized by a B. cepacia strain (Burkhead et al., 1994).
B� iô oð s� y� ní tç hä eè s� iô s � aå ní d ñ
ré egè�� uò lîaå tç iô oð n í oð f
�P� R N
Four genes, prnABCD, which are organized in a single transcriptional unit are required
for PRN synthesis by P. fluorescens BL915 (Fig. 4, Hammer et al., 1997). The prnA gene
contains a short region with high similarity to the NAD binding domain of the NADH
dehydrogenases from Haemophilus influenzae and Escherichia coli, and the thioredoxin
reductase from Streptomyces clavuligerus. The prnB gene had no homologies in the database.
The gene prnC is highly homologous to the chl gene from S. aureofaciens, which encodes a
chlorinating enzyme involved in the synthesis of tetracycline. Finally, prnD has similarity with
a portion of the 3-chlorobenzoate-3,4-dioxygenase (cbaA) of Alcaligenes sp., phthalate 4,5-
dioxygenase (pht3) from P. putida, and vanilate demethylase (vanA) from Pseudomonas sp.
(Table 3, Hammer et al., 1997). The two chlorinating enzymes encoded by prnA and prnC
represent a new class of halogenating enzymes, since they were different from all previously
described haloperoxidases (Hammer et al., 1999). Further studies on the prnABCD gene
cluster of four producing strains, P. fluorescens BL915, B.cepacia LT4-12-W, B. pyrrocinia, and

C� h�
a pt��� e� r � 1: ��� G� e� n� e� r� a l � I� n� t� r� o� d�
u� c� t� i� o� n � a n� d �
O� u� t� l� i� n� e�
11
T
a! b" l#e $ 3. %'&
P(
R)
N *
b"
i+o, s- y. n/ t0 h1 e$ s- i+ s - i
+n / P2
. 3 f4l5u6 o7 r8 e9 s: c; e9 n< s: B= L
>915.?'@BAB&
(Hammer et al., 1997; Kirner et al. 1998).
ORF Size (bp) Homology a Function b
prnA 1617 dehydrogenase from H. influenzae conversion of L-T to 7-CTprnB 1086 no homology conversion of 7-CT to MDAprnC 1704 chl gene from S. aureofaciens conversion of MDA to APRNprnD 1092 cbaA gene from Alcaligenes sp. conversion of APRN to PRN
a Only highest level of homology shown.b L-T is L-tryptophan, 7-CT is 7-chloro-L-tryptophan, MDA is monodechloroaminopyrrolnitrin, APRN is
aminopyrrolnitrin.
FC
i+gD uE rF e $ 4.
G &OH
rF gD a! n/ i+zI a! t0 i+ o, n / o, f
Jt0 h1 e $ P
(R)
N *
b"
i+o, s- y. n/ t0 h1 e$ t0 i+ c K o, pL e$ rF o, n / o, f
JP2
. 3 f4l5u6 o7 r8 e9 s: c; e9 n< s: B= L
>915. ?'@BAM&
Arrows indicate thedirection of transcription. (Hammer et al., 1997). Not drawn to scale.
Myxococcus fulvus showed that these genes are conserved among the studied strains (Hammer
et al., 1999).The biosynthetic pathway for PRN was clearly defined by using mutants defective
in each of the genes in the PRN operon (Kirner et al., 1998). Tryptophan was identified as the
precursor of PRN, based on studies using isotopically labelled and substituted tryptophan to
feed bacterial cultures (Chang et al., 1981). The steps for the conversion of tryptophan to
PRN, catalysed by the prnABCD gene products, are shown in Table 3.
The biosynthetic regulation of PRN is influenced by nutritional and environmental
conditions. PRN production by P. fluorescens CHA0 was stimulated by fructose, mannitol,
and amendment with a mixture of zinc and ammonium molybdate (Duffy and Défago, 1999).
Studies with B. cepacia strains show that PRN is produced at higher concentrations when the
bacteria were grown in media with low initial pH (Roitman et al., 1990). The use of glycerol
as a carbon source enhanced PRN production, whereas galactose, rhamnose, lactose and starch
repressed production (El-Banna and Winkelmann, 1998). It was reported that PRN in B.
cepacia was produced in the late growth phase and about 98% was inside the bacterial cells
(Roitman et al., 1990; El-Banna and Winkelmann, 1998).
Genetic engineerng of P. fluorescens BL915 for increased PRN production provides an
example of the potential of such modifications in improving the performance of this strain as a
biological control agent (Ligon et al., 2000). The genetically modified strains provided better
control of R. solani and Pythium ultimum than the wild type strain under controlled and field
prnA prnB prnC prnD

CN hO
aP ptQ�R eS r T 1: U�V GW eS nX eS rT aP l Y IZ nX tR rT o[ d\
u] c^ tR i_ o[ n X aP nX d \
Ou] tR lY i_ nX eS
12
conditions. The level of control provided by the modified strains was not different from
fungicide checks and there was a correlation between increased PRN production and increased
biocontrol activity.
Pa yb oc ld ue tf eg oc rh ii n j (k Pa Ll Tm
)n
PLT (4,5-dichloro-1 H-pyrrol-2-yl-2,6-dihydroxyphenyl ketone) is a phenolic
polyketide antibiotic which consists of a resorcinol ring linked to a bichlorinated pyrrole
moiety (Fig. 1). PLT was first isolated from P. aeruginosa (Takeda, 1958) and later from P.
aeruginosa strain S10B2 (Ohmori et al., 1978) and P. fluorescens strains Pf-5 and CHA0
(Bencini et al., 1983; Bender et al., 1999; Keel et al., 1996). Most of the information on PLT
biosynthesis, its role in biocontrol, and regulation stems from strains Pf-5 and CHA0.
The precise mode of action of pyolutorin is not clear, but it has bactericidal,
herbicidal, fungicidal, and oomycidal activities, in particular against Pythium spp. (Maurhoffer
et al., 1992; Ohmori et al., 1978; Takeda, 1958). Application of pure PLT to cotton seeds
resulted in significant suppression of Pythium ultimum-induced damping-off (Howell and
Stipanovic, 1980). Results from studies with derivatives of specific Pseudomonas strains, that
were either defective in PLT production or produced higher levels of PLT, have indicated that
the determinative role of PLT in biological control may vary among different plant hosts
(Maurhoffer et al., 1992, 1994, 1995; Kraus and Loper, 1992). PLT production in the
rhizosphere by strain CHA0 and plt gene expression in the spermosphere by strain Pf-5 vary
among different host plants, presumably affecting the biocontrol activity of PLT (Maurhoffer
et al., 1995; Kraus and Loper, 1995).
Bo i_ o[ sp yq nX tR hO eS sp i_ s p aP nX d \
rT egS�r u] lYaP tR i_ o[ n X o[ f
sPt Lu T
v
Ten genes for synthesis of PLT in P. fluorescens Pf-5 were identified. The genes
pltLABCDEFG are organized in a single transcriptional unit, whereas pltRM comprise another
transcriptional unit oriented in the opposite direction of pltLABCDEFG (Fig. 5). The pltB and
pltC genes show similarity with the eryA gene of Saccharopolyspora erythraea that encode a
bacterial Type I polyketide synthase necessary for erytrhromycin production (Nowak-
Thompson et al., 1997; Table 4). Genes pltA, pltD, and pltM are similar to genes encoding
halogenating enzymes required for chlorotetracycline biosynthesis by Streptomyces aureofaciens
(Cts4) and for pyrrolnitrin biosynthesis by strain BL915 (PrnC). The gene pltE is similar to

Cw hx
ay ptz�{ e| r } 1: ~�� G� e| n� e| r} ay l � I� n� t{ r} o� d�
u� c� t{ i� o� n � ay n� d �
O� u� t{ l� i� n� e|
13
T�
a� b� l�e � 4. �'�
B�
i�o� s� y� n� t� h� e� s� i� s � o� f
�P�
L�
T �
i�n � P�
. � f�l�u o¡ r¢ e£ s¤ c¥ e£ n¦ s¤ P� f
�-§ 5.¨ � (Nowak-Thompson et al., 1997, 1999).
ORF Size(bp)
Homology a Possible function
pltA 1347 halogenating enzymes pyrrole chlorinationpltB 7374 Type I PKSs resorcinol synthesispltC 5322 Type I PKSs resorcinol synthesispltD 1632 halogenating enzymes pyrrole chlorinationpltE 1140 acyl-CoA dehydrogenases pyrrole synthesispltF 1494 peptide synthases precursor activationpltG 780 thioesterases involved in secondary metabolism termination of resorcinol synthesispltL 264 no homology unknownpltM 1506 halogenating enzymes pyrrole chlorinationpltR 1029 transcriptional regulator of R. japonicum pathway specific regulation
a PKS is polyketide synthase; only highest level of homology is shown.
F©
i�gª u« r¬ e � 5. ¨ � O
r¬ gª a� n� i
�z® a� t� i� o� n � o� f
�t� h� e � P
�L�
T �
b�
i�o� s� y� n� t� h� e� t� i� c ¯ gª e� n� e� s � i
�n � P�
. � f�lu� o¡ r¢ e£ s¤ c¥ e£ n¦ s¤ P
�f�-§ 5.¨ � Arrows indicate
transcription direction. (Nowak-Thompson et al., 1997, 1999). Not drawn to scale.
many flavin-dependent acyl coenzyme A (acyl-CoA) dehydrogenases and specially to butyryl-
CoA dehydrogenase from Megasphaera elsdenii. the pltF gene is similar to several peptide
synthases including grsB from Bacillus brevis, snbC from Streptomyces pristinaespiralis, and pvdD
from P. aeruginosa. The gene pltG is similar to several thioesterases involved in secondary
metabolism, including thioesterase II from rat, grsT from B. brevis, and CmaT from P.
syringae; pltL shows highest similarity to a hypothetical protein encoded by the red gene cluster
of S. coelicolor. The pltR product resembles members of the LysR family of transcriptional
regulators such as gstR of Rhizobium japonicum, and pltL did not have any match in the
database (Nowak-Thompson et al., 1999). The PLT biosynthetic gene cluster has not been
completely delimited and little is known about the chemical intermediates involved in its
biosynthesis. Proline and acetyl- coenzyme A are the precursors of PLT (Bender et al., 1999).
The available information on the synthesis of PLT is shown in Table 4.
PLT production is partially regulated by pltR, which encodes a transcriptional activator
protein. Transcription of pltB, pltE, and pltF genes was reduced in mutants disrupted in pltR
as compared to the wild type strain (Nowak-Thompson et al., 1999). The physiological status
of the bacterial cells and the environmental conditions greatly affect production of PLT.
pltM pltR pltL pltA pltB pltC pltD pltE pltF pltG

C° h±
a² pt³�´ eµ r ¶ 1: ·�¸ G¹ eµ nº eµ r¶ a² l » I¼ nº t´ r¶ o½ d¾
u¿ cÀ t´ iÁ o½ n º a² nº d ¾
OÂ u¿ t´ l» iÁ nº eµ
14
Secretion of PLT was stimulated by zinc, cobalt, glycerol, and poor aeration, whereas
glucose, phosphate, and tryptophan repressed PLT production (Bencini et al., 1983; Duffy
and Défago, 1999). The use of ethanol as a sole carbon source enhanced PLT production in P.
fluorescens S272 (Yuan et al., 1998).
GlÃÅÄ oÆ bÇ aÈ l Ä RÉ eÊ gË uÌ lÄ aÈ tÍ iÎ oÆ n Ï oÆ f Ð aÈ nÏ tÍ iÎ bÇ iÎ oÆ tÍ iÎ c Ñ pÒ rÓ oÆ dÔ uÌ cÑ tÍ iÎ oÆ n Ï iÎ n Ï PÕ
sÖ e× uØ dÙ oÚ mÛ oÚ nÜ aÝ sÖ bÇ y Þ gß aÝ cà Sá /â gß aÝ cà Aã
In several strains that belong to the group of plant-associated Pseudomonas species,
expression of genes involved in the biosynthesis of secondary metabolites and extracellular
enzymes is positively controlled by the GacS/GacA two-component system (Heeb and Haas,
2001). This regulatory system consists of a membrane-bound sensor kinase protein (GacS) and
a cytoplasmic response regulator protein (GacA). The current model proposes that GacS
recognizes specific environmental stimuli and activates GacA which in turn triggers specific
gene expression (Appleby et al., 1996; Pernestig et al., 2001; Heeb and Haas, 2001).
Although a considerable amount of information has accumulated in the last decades on these
two-component regulatory systems, the environmental signals responsible for the activation of
GacS are still unknown.
The gene gacS was first described in Pseudomonas syringae pv. syringae as an essential
factor for disease manifestation (Hrabak and Willis, 1992). The cognate response regulator
gene gacA was first identified in the biocontrol strain P. fluorescens CHA0 as essential for the
biocontrol activity of this strain (Laville et al., 1992).
Many regulatory elements may be involved in the regulation of antibiotic-encoding
genes, forming a complex regulatory cascade, with the gacS/gacA pair exerting control at a
higher hierarchy (Sarniguet et al., 1995; Chancey et al., 1999; Blumer and Haas, 2000;
Whistler et al., 1998). Mutations in either gacS or gacA abolishes the production of 2,4-
DAPG, PLT, PRN, and PHZ .
Production of 2,4-DAPG is influenced by the interactions of the gacS/gacA system
with other regulatory elements, including the housekeeping sigma factor σ70 of RNA
polymerase encoded by rpoD, and the stationary phase and stress sigma factor σ38 which is
encoded by rpoS (Schnider et al., 1995b; Whistler et al., 1998; Blumer and Haas, 2000).
Mutantional disruption of rpoS in P. fluorescens Pf-5 or introduction of multiple copies of
rpoD in P. fluorescens CHA0 led to overproduction of 2,4-DAPG (Sarniguet et al., 1995;
Schnider et al., 1995a).

Cä hå
aæ ptç�è eé r ê 1: ë�ì Gí eé nî eé rê aæ l ï Ið nî tè rê oñ dò
uó cô tè iõ oñ n î aæ nî d ò
Oö uó tè lï iõ nî eé
15
PHZ production is regulated at multiple levels by gacS/gacA, including mediation of
the transcription of phzI, which is necessary for AHL production in P. aureofaciens 30-84
(Chancey et al., 1999). The rpoS gene was recently shown to regulate production of PHZ in P.
aeruginosa PAO1 (Whiteley et al., 2000).
Two global regulators are known to influence PRN production in Pseudomonas. The
rpoS gene and GacS/GacA system were shown to be essential for PRN production by P.
fluorescens Pf-5 (Sarniguet et al., 1995; Corbell and Loper, 1995). Mutants disrupted in rpoS
or gacS/gacA do not produce PRN (Gaffney et al., 1994).
Genetic regulation of PLT production is complex, involving several regulators. The
gacS/gacA regulatory system was shown to be essential for PLT production in strains Pf-5 and
CHA0 (Laville et al., 1992; Corbell and Loper, 1995). This two-component system is known
to positively influence the accumulation of sigma factors in Pseudomonas (Whistler et al.,
1998). Inactivation of rpoS in P. fluorescens Pf-5 or the introduction of multiple copies of rpoD
in P. fluorescens CHA0 also led to enhanced PLT production by these strains (Schnider et al.,
1995a; Sarniguet et al., 1995). This mechanism of genetic regulation shows remarkable
resemblance with that of 2,4-DAPG.
Most information available on gacS/gacA regulatory systems stems from studies on
model strains and very little information is available on the distribution of these genes in
rhizosphere environments. In this thesis, we investigated the usefullness of the gacA gene as a
complementary genetic and phylogenetic marker for Pseudomonas (Chapter 3).
B÷ iø où sú uû rü fý aþ cÿ t� aþ n� t � aþ n� t� iø b� iø où t� iø cÿ sú
Many surface-active molecules (surfactants) of biological origin have been described
(Feichter, 1992). Their surfactant properties result from the presence of both hydrophilic and
hydrophobic regions in the same molecule. There are several classes of biosurfactants with
different chemical structures and surface properties. Biosurfactants can be classified as low-
molecular-weight polymers that lower surface and interfacial tensions and high-molecular-
weight polymers that bind to surfaces (Ron and Rosenberg, 2001). Several biosurfactants
produced by Pseudomonas species have antibiotic activity in addition to their surfactant
activity, including rhamnolipids and peptide biosurfactants. These two classes of
biosurfactants are classified as low-molecular-weight polymers.

C� h�
a� pt��� e r 1: � � G� e n� e r a� l � I� n� t� r o� d�
u� c� t� i� o� n � a� n� d �
O� u� t� l� i� n� e
16
Rhamnolipids (RHLs) are glycolipid biosurfactants containing rhamnose and β-
hydroxydecanoic acid and are produced by certain species of fluorescent pseudomonads,
specifically P. aeruginosa (Itoh et al., 1971; Van Dijke et al., 1993). There are six known RHLs
which differ in their rhamnose content and in the length and chemical nature of their lipid
fraction (Jarvis and Johnson, 1949; Hirayama and Kato, 1982; Syldatk et al., 1985). There is a
considerable amount of information on the biosynthesis, genetics, and regulation of RHL
production in P. aeruginosa (Hauser and Karnovsky, 1958; Ochsner and Reiser, 1995). RHLs
produced by P. aeruginosa may serve as virulence factors during colonization of the human
lung tissue (Iglewski, 1989). In addition, they show antibiotic effects, acting in solubilization
of cell envelopes of competing microorganisms (Kurioka and Liu, 1967). RHLs also present
antibacterial, mycoplasmacidal, cytotoxic, and antiviral activities in vitro (Jarvis and Johnson,
1949; Edwards and Hayashi, 1965; Hisatsuka et al., 1971; Itoh et al., 1971; Hirayama and
Kato, 1982). The role of RHLs in the biocontrol of plant pathogens was recently
demonstrated (Stanghellini and Miller, 1997; Kim et al., 2000). RHLs were active against
zoospores of oomycetes, causing cessation of motility and lysis of entire zoospore populations
in less than 1 min. Introduction of a RHL-producing bacterium in a recirculating hydroponic
system gave good control, although transient, of Phytophthora capsici on pepper (Stanghellini
and Miller, 1997). P. capsici mycelial growth and Colletotrichum orbicularie spore germination
were inhibited in vitro by RHL B produced by P. aeruginosa B5. The diseases caused by these
pathogens were also suppressed in pepper and cucumber plants treated with purified RHL B
(Kim et al., 2000). RHLs are thought to act by intercalating into the plasma membrane of
zoospores and cause its disruption (Stanghellini and Miller, 1997). However, their precise
mode of action on structures protected by cell walls has not been demonstrated conclusively.
Lipopeptide surfactants are composed of a peptide moiety linked to a fatty acid. The
peptide moiety, like many small peptides in microorganisms, is synthesized non-ribosomally
by a multi-enzyme peptide synthase complex (Ron and Rosenberg, 2001). Pseudomonas species
produce several lipopeptide surfactant, including viscosin (Hiramoto et al., 1970),
viscosinamide (Nielsen et al., 1999), massetolides (Gerard et al., 1997), tensin (Nielsen et al.,
2000), syringopeptins, syringomycins, syringostatins, syringotoxins, and pseudomycins
(Hutchison and Gross, 1997; Serra et al., 1999), tolaasin (Rainey et al., 1991), the “white line
inducing principle” (WLIP) (Mortshire-Smith et al., 1991), amphisin (Sørensen et al., 2001),
pholipeptin (Ui et al., 1997), arthrofactin (Morikawa et al., 1993), and ecomycins (Miller et
al., 1998). All Pseudomonas-produced lipopeptides tested to date have antibiotic properties.

C� h�
a� pt��� e� r � 1: � G! e� n" e� r� a� l # I$ n" t� r� o% d&
u' c( t� i) o% n " a� n" d &
O* u' t� l# i) n" e�
17
The lipopeptides syringomycins, syringopeptins, syringostatins, syringotoxins, and
pseudomycins, produced by strains of P. syringae are virulence factors in the diseases caused by
these phytopathogenic bacteria (Scholz-Schroeder et al., 2001). Syringomycins, syringostatins,
syringotoxins, and pseudomycins have marked antifungal activity and are potentially useful for
the development of agents for the control of plant and human diseases, whereas syringopeptins
have stronger phytotoxic and reduced antifungal activity as compared to the others (Lam et al.,
1987; Iacobellis et al., 1992). Similarly, tolaasin, produced by P. tolaasii, is the primary
bacterial compound responsible for eliciting disease symptoms on cultivated mushrooms
(Godfrey et al., 2001) and also activity against a range of basidiomycetes and Gram-positive
bacteria (Rainey et al., 1991). Interestingly,WLIP, produced by P. reactans, an opportunistic
pathogen related to P. tolaasii, was proposed as an inhibitor of the symptoms caused by P.
tolaasi on mushrooms (Soler-Rivas et al., 1999). Viscosin, a lipopeptide with antifungal and
surfactant properties, is an important virulence factor in the disease incited by pectolytic
strains of P. fluorescens on brocoli (Hildebrand et al., 1998). Tolaasin and viscosin are also
proposed as biological agents for the control of pathogenic fungi (Nielsen et al., 1999).
Ecomycins, produced by P. viridiflava, were reported to be active against several human and
plant pathogenic fungi (Miller et al., 1998). The role of ecomycins in the weak disease
inducing capacity of P. viridiflava is not yet clear.
Viscosinamide, and tensin, produced by soil-inhabiting P. fluorescens strains were
reported to inhibit mycelial growth of several fungi and oomycetes, inducing hyphal swellings
and stimulating hyphal branching (Thrane et al., 1999; Nielsen et al., 2000). Pseudomonas sp.
from marine tube worms produce several cyclic depsipeptides called massetolides (Gerard et
al., 1997), which are related to other lipopeptide surfactants, such as viscosin and
viscosinamide. Massetolides showed pronounced activity against Mycobacterium species.
Amphisin and pholipetin, related compounds produced by P. fluorescens (Ui et al.,
1997; Sørensen et al., 2001), and arthrofactin produced by a Pseudomonas sp. (Morikawa et
al., 1993) did not yet have their biological activity investigated, but, because of their structural
similarities to the other lipopeptides described above, similar activities might be expected.
The mode of action of several lipopeptide surfactants was shown to involve formation
of ion channels in the cell wall and pertubations of the cell membrane structure (Nielsen et al.,
1999; Hutchison and Johnstone, 1993; Vanittanakom and Loeffler, 1986; Thrane et al.,
1999).

C+ h,
a- pt.�/ e0 r 1 1: 2 3 G4 e0 n5 e0 r1 a- l 6 I7 n5 t/ r1 o8 d9
u: c; t/ i< o8 n 5 a- n5 d 9
O= u: t/ l6 i< n5 e0
18
B> i< o8 s? y@ n5 t/ h, e0 s? i< s ? a- n5 d 9
r1 eg0BA u: l6a- t/ i< o8 n 5 o8 f
Cs? ur:�1 fC a- c; t/ a- n5 t / a- n5 ti/ < bD i< o8 ti/ < c; s?
Little is known about the biosynthesis, regulation, and genetics of most lipopeptide
surfactants. There is, however, some information available on the genetics of some P. syringae
toxins (Scholz-Shroeder et al., 2001), tolaasin (Rainey et al., 1993), and viscosin (Braun et al.,
2001). From the few characterized examples, it is known that lipopeptides are encoded by
relatively large DNA fragments (Scholz-Shroeder et al., 2001; Braun et al., 2001) and that
peptide synthases are involved in the biosynthesis of these compounds (Marahiel et al., 1997).
It was reported that a massetolide-producing Pseudomonas sp. could incorporate amino acids
artificially at various positions, thereby producing new types of massetolides (Gerard et al.,
1997). These results show that peptide synthases may be unspecific in their assembly of
amino acids in depsipeptides. Some studies suggest that lipopeptide production is regulated in
a cell-density dependent manner, because their production seems to be linked to cell
proliferation (Ron and Rosenberg, 2001). Production of the toxin tolaasin commenced during
exponential growth and continued into stationary phase (Rainey et al., 1991). On the other
hand, studies on the biosynthesis of viscosinamide showed that this compound was
abundantly produced in the logarithmic phase of growth and abolished in the stationary phase
(Nielsen et al., 1999).This production pattern differs from that of the peptide antibiotic
tolaasin, other antibiotics, and RHLs, which are considered secondary metabolites. The
authors, based on their and other studies, proposed that viscosinamide should be considered a
primary rather than a secondary metabolite (Nielsen et al., 1999). Lipopeptide production
patterns and regulation are expected to vary, especially when the different biological purposes
for which Pseudomonas species produce lipopeptides (Ron and Rosenberg, 2001) are taken
into consideration.
DE iF sG tH rI iF bJ uK tH iF oL n M aN nM d dOPO eQ tH eQ cR tH iF oL n M oL f S aN nM tH iF bJ iF oL tH iF cR -T pU rI oL dO uK cR iF nM g V PW
sX eY uZ d[ o\ m] o\ na_ sX
Numerous bacterial strains that produce antibiotics in vitro have been isolated from
different soils and plant hosts. This apparent wide distribution suggests that antibiotic-
producing bacteria are common constituents of the indigenous microflora in soil and plant-
associated environments worldwide. To date, however, little is known about the frequency and
ecology of indigenous antibiotic-producing bacteria. Knowledge about the ecology of naturally
occurring strains that harbor specific biocontrol traits will significantly contribute to

Cha
ab ptc�d ee r f 1: g h Gi ee nj ee rf ab l k Il nj td rf om dn
uo cp td iq om n j ab nj d n
Or uo td lk iq nj ee
19
improving the efficacy of existing biocontrol agents and may help to identify new strains that
are better adapted to specific soils or host-pathogen systems.
Selection and identification of antibiotic-producing bacteria via random isolation from
natural environments is very time-consuming and laborious. In fact, most of the model strains
used to date were identified via screening huge numbers of isolates obtained by random
procedures. The availability of cloned and sequenced antibiotic biosynthetic and regulatory
genes, however, has facilitated the development of specific primers and probes that can be used
for targeted detection and isolation of specific antibiotic-producing bacteria. A prerequisite for
DNA-based detection and isolation is that the genes of interest must be conserved among a
wide range of bacterial strains harboring this trait. By Southern hybridization with a 4.8-kb
chromosomal DNA fragment of P. fluorescens strain Q2-87, Keel et al. (1996) showed that the
biosynthetic locus for 2,4-DAPG production was highly conserved among 45 2,4-DAPG-
producing Pseudomonas strains of worldwide origin. Subsequent development of primers and
probes directed against phlD, a key gene in the biosynthesis of 2,4-DAPG, allowed targeted
detection, isolation and enumeration of 2,4-DAPG-producing Pseudomonas spp. occurring
naturally on roots of wheat (Raaijmakers et al., 1997). Also the genes or gene clusters involved
in the biosynthesis of PHZs (Mavrodi et al., 2001) and PRN (Hammer et al., 1999),
respectively, appear to be conserved among different strains. In our studies, PCR analysis,
Southern hybridization and restriction fragment length polymorphisms (RFLP) analyses
showed that specific genes involved in PRN and PLT biosynthesis are highly conserved in a
large collection of both Pseudomonas and Burkholderia species (Chapter 2).
The use of probes and primers directed against genes involved in antibiotic
biosynthesis has proven to be a powerful technique to study the distribution and function of
indigenous antibiotic-producing Pseudomonas spp. Colony hybridization followed by PCR
analysis showed that root-associated fluorescent Pseudomonas spp. producing the antibiotic
2,4-DAPG were present on roots of wheat grown in several take-all suppressive soils, referred
to as take-all decline soils (TAD), at densities ranging from approximately 5x105 to 2x106
CFU per gram of root. In the complementary conducive soils, 2,4-DAPG-producing
pseudomonads were not detected or detected at densities at least 40-fold lower than in the
TAD soils (Raaijmakers et al., 1997). Moreover, 2,4-DAPG-producing Pseudomonas spp. were
present on roots of wheat grown in at least three TAD soils at or above the threshold
population density required for significant suppression of take-all of wheat. The specific
suppression that operates in take-all suppressive soils was lost when indigenous 2,4-DAPG-
producing fluorescent Pseudomonas spp. were eliminated by selective heat treatment, and

Cs ht
au ptv�w ex r y 1: z { G| ex n} ex ry au l ~ I� n} tw ry o� d�
u� c� tw i� o� n } au n} d �
O� u� tw l~ i� n} ex
20
conducive soils gained suppressiveness to take-all when indigenous 2,4-DAPG-producing
Pseudomonas strains were introduced via mixing in small amounts of raw take-all suppressive
soil (Raaijmakers et al., 1998).
Apart from genetic markers, also phenotypic markers may provide a source to rapidly
isolate and identify bacteria with antagonistic potential. For example, Ellis et al. (2000)
characterized a collection of 29 fluorescent pseudomonads with the intent of identifying
conserved phenotypic or genotypic traits in strains with activity against Pythium ultimum.
They found a significant correlation between biological control activity and the accumulation
of a specific cyclopropane fatty acid (C17:O CFA) and hydrogen cyanide production. The
authors concluded that screening isolates on the basis of elevated synthesis of C17:O CFA has
the advantage that it does not depend on prior knowledge of secondary metabolite synthesis.
D� i� v� e� r� s� i� t� y � o� f � a� n� t� i� b� i� o� t� i� c� -� p� r� o� d� u� c� i� n� g � P�
s� e� u� d� o� m� o� n a¡ s�
The genotypic and phenotypic diversity that occurs in natural populations of
biocontrol agents provides an enormous resource for improving biological control of plant
diseases (Handelsman and Stabb, 1996; Thomashow and Weller, 1996). This approach has
been widely used to select for better biocontrol agents of insects, and to improve the use of
microorganisms in the production of fermented foods and biodegradation of xenobiotic
compounds (reviewed in Stabb et al., 1994). However, exploitation of genotypic diversity
among bacterial biocontrol agents of plant pathogenic fungi, so far, has received much less
attention. Knowledge of the diversity within a group of strains that share a common
biocontrol trait may provide a new approach to identify biocontrol strains that are superior
with respect to ecological competence and ability to suppress specific plant diseases.
Recent studies have shown that there is considerable genotypic diversity in antibiotic-
producing Pseudomonas species (Keel et al., 1996; McSpadden-Gardener et al., 2000).
Different genotypes of 2,4-DAPG-producing Pseudomonas spp. have been reported to differ in
their ability to suppress Fusarium crown and root rot and Pythium root rot (Sharifi-Tehrani et
al., 1998), to produce other antibiotics in addition to 2,4-DAPG (Keel et al., 1996), and to
colonize roots of maize plants of different growth stages (Picard et al., 2000). Among 101
isolates of indigenous 2,4-DAPG-producing Pseudomonas species, occurring on roots of wheat
grown in a soil naturally suppressive to take-all disease of wheat, 16 different groups were
identified by Random Amplified Polymorphic DNA (RAPD) analysis with two 10-mer

C¢ h£
a¤ pt¥�¦ e§ r ¨ 1: © ª G« e§ n¬ e§ ra¤ l I® n¬ t¦ ro¯ d°
u± c² t¦ i³ o¯ n ¬ a¤ n¬ d °
Ou± t¦ l i³ n¬ e§
21
primers (Raaijmakers and Weller 2001). One RAPD-group made up 50% of the total
population of 2,4-DAPG-producing Pseudomonas spp. Subsequent root-colonization and
biocontrol studies indicated that this dominant genotype, exemplified by P. fluorescens Q8r1-
96, was highly adapted to the wheat rhizosphere and was very effective in suppression of take-
all disease of wheat (Raaijmakers and Weller, 2001). The observation that strain Q8r1-96
showed the same population dynamics during successive cycling of wheat in two other soils,
both of which had different physical-chemical properties, suggested that its superior
rhizosphere competence is not soil specific. This was supported by results obtained in a study
by McSpadden-Gardener et al. (2000), who found that nearly one-third of the 2,4-DAPG-
isolates obtained from soils of different wheat-growing areas in the United States were
genotypically similar to strain Q8r1-96. Biochemical analyses indicated that the superior
rhizosphere competence of Q8r1-96 was not related to elevated in situ 2,4-DAPG production
levels but possibly to its ability to utilize specific substrates (Raaijmakers and Weller, 2001). These
data and results obtained by Sharifi-Tehrani et al. (1998) illustrate that exploiting the diversity
within a specific group of antagonistic microorganisms has potential for improving biological
control. This approach capitalizes on existing knowledge concerning mechanisms, while
exploiting differences among strains to face the biotic and abiotic complexity of natural
environments.

Cµ h¶
a· pt¸�¹ eº r » 1: ¼ ½ G¾ eº n¿ eº r» a· l À IÁ n¿ t¹ r» o dÃ
uÄ cÅ t¹ iÆ o n ¿ a· n¿ d Ã
OÇ uÄ t¹ lÀ iÆ n¿ eº
22
OÈ UÉ TÊ
LË IÌ NÍ E Î OÈ F Ï TÊ
HÐ E Î TÊ
HÐ EÎ SÑ IÌ SÑ
Bacteria of the genus Pseudomonas are able to survive and prosper in a wide range of
environmental conditions. This genus not only contains plant, animal, and human pathogenic
species, but also accomodates species that are of significant environmental importance,
including plant growth promoters, xenobiotic degraders, and biocontrol agents. Their versatile
metabolic activities and ability to produce a wide variety of secondary metabolites have
stimulated numerous ecological, molecular and biochemical studies. In spite of significant
progress in the understanding of the biosynthesis and regulation of several antibiotic
metabolites (Cµ h¶
a· pt¸�¹ eº r » 1¼ ), little is known about the distribution and diversity of indigenous
Pseudomonas populations harboring these traits. Furthermore, many studies focus on one or a
limited number of model strains producing known antifungal compounds, whereas
considerably less attention is given to the discovery of new metabolites that may have great
potential for biological control of plant diseases. Our mÒ a· jÓ o r » o bÔ
jÓeº cÅ ti¹ Æ vÕ eº s Ö iÆ n ¿ t¹ h¶ iÆ s Ö t¹ h¶ eº sÖ iÆ sÖ are i) to
study the distribution, diversity and activity of indigenous Pseudomonas populations producing
the antbiotics 2,4-diacetylphloroglucinol (2,4-DAPG), phenazines (Phz), pyrrolnitrin (PRN)
or pyoluteorin (PLT) (Cµ h¶
a· pt¸�¹ eº r» s Ö 2-5×ÙØÛÚ
), and ii) to isolate, identify and characterize Pseudomonas
strains and novel antifungal metabolites with biocontrol potential (Cµ h¶
a· p¸ t¹ eº r » 6Ü ).
In Cµ h¶
a· pt¸Ý¹ eº r » 2×
, the development and specificity of primers and probes for targeted
isolation and enumeration of PRN- and PLT-producing Pseudomonas and Burkholderia species
are described. The usefulness of these molecular markers for phylogenetic analysis of
Pseudomonas and Burkholderia species was evaluated and compared to phylogeny inferred from
16S rDNA sequences. Finally, the frequencies of PRN- and PLT-producing Pseudomonas and
Burkholderia species in the rhizosphere of wheat plants grown in soils suppressive to the take-
all fungus Gaeumannomyces graminis var. tritici were assessed by colony hybridization followed
by PCR.
Production of 2,4-DAPG, PHZ, PRN and PLT by several Pseudomonas strains is
regulated by the two-component system GacA/GacS. Recent studies suggested that the
response regulator gene gacA is widely distributed in Pseudomonas and other Gram-negative
bacteria. In Cµ h¶
a· pt¸�¹ eº r » 3Þ , the gacA gene was sequenced from ten different Pseudomonas strains
and the evolutionary rates and patterns of nucleotide substitutions were studied. Phylogenies
inferred from gacA gene and protein sequences were compared to the classic 16S rDNA-based
phylogeny. The use of the gacA gene as a complementary genetic marker for detection of

Cß hà
aá ptâ�ã eä r å 1: æ ç Gè eä né eä rå aá l ê Ië né tã rå oì dí
uî cï tã ið oì n é aá né d í
Oñ uî tã lê ið né eä
23
Pseudomonas in environmental samples was evaluated on bacterial populations isolated from
the rhizosphere of wheat grown in agricultural soils.
Antibiotic-producing Pseudomonas spp. have been implicated in the natural
suppressiveness of soils to specific soil-borne fungi, including G. graminis var tritici. Cß hà
aá ptâÝã eä r å 4ò
aimed at disclosing the contribution of 2,4-DAPG-producing Pseudomonas spp. to the
suppressiveness of two Dutch soils to G. graminis var. tritici. In these soils, 2,4-DAPG-
producing Pseudomonas spp. were highly enriched on roots of wheat. Mutants defective in 2,4-
DAPG production were constructed and their activities were compared to those of their
parental strains to establish the determinative role of this metabolite in interactions with G.
graminis var. tritici and in rhizosphere competence. In collaboration with USDA-ARS
(Pullman, WA), comparative studies on major genotypic groups of 2,4-DAPG producers
found in Dutch and US take-all decline soils were carried out.
Cß hà
aá ptâ�ã eä r å 5ó focuses on the activity of 2,4-DAPG against plant pathogenic fungi and in
particular against Pythium species. This oomycete pathogen has a complex life cycle in which
different structures play an important role. We focused on the asexual life cycle and
determined the sensitivity of different structures to 2,4-DAPG. In collaboration with INRA
(Dijon, France), transmission eclectron microscopy was performed to gain more insight into
the mode of action of 2,4-DAPG.
The last experimental chapter of this thesis concentrated on the discovery and
characterization of novel strains and metabolites with biocontrol potential. Strains producing
surface-active compounds were isolated from the rhizosphere of wheat. The potential of one of
these Pseudomonas strains to control Pythium root rot was studied in collaboration with the
Applied Research Station (PPO) in Lisse, the Netherlands. Mutants defective in the
production of the surface-active compound were constructed and partial sequences of genes
involved in the biosynthesis were obtained by anchored PCR. The composition of the partially
purified biosurfactants produced by this Pseudomonas strain was determined in collaboration
with the Department of Organic Chemistry of Wageningen University (Cß hà
aá pâ tã eä r å 6ô ).
The results obtained in Chapters 2-6 are evaluated in a summarising discussion
(Cß hà
aá ptâ�ã eä r å 7õ
).

Cö h÷
aø ptù�ú eû r ü 1: ý þ Gÿ eû n� eû rü aø l � I� n� tú rü o� d�
u� c� tú i� o� n � aø n� d �
O� u� tú l� i� n� eû
24
R E F�
E R E N�
C
E S�
A�
l�t� m� a� n� , � J
�., ��� a� n� d
�C
a� m� p� b�
e� ll��� , � C
.� L� .� 1977. Effect ofherbicides on plant diseases. Ann. Rev.Phytopathol. 15:361-385.A�
n� j�a� i a� h! , � V
"., ��� K
#o$ e� d� a� m� , � N
�., ��� N
�o$ w% a� k& -' T( h! o$ m� p� s) o$ n� , � B* .,���
L� o$ p� e� r+ , � J�.� E ., ��� H
,ö- f. t� e� , � M/ ., ��� T( a� m� b
�o$ n� g0 , � J
�.� T( ., ��� a� n� d
�C
o$ r+ n� e� l� i s) , � P1 .� 1998. Involvement of phenazines andanthranilate in the antagonism of Pseudomonasaeruginosa PNA1 and Tn5 derivatives towardFusarium spp. and Pythium spp. Mol. Plant-Microbe Interact. 11:847-854.A�
pp�2� l�e� b� y3 , � J
�.� L� ., ��� P1 a� r+ k& i n� s) o$ n� , � J
�.� S� ., ��� a� n� d
�B* o$ u4 rr+5+ e� t� , � R .� B* .�
1996. Signal transduction via the multi-stepphosphorelay: not necessarily a road less traveled.Cell 86:845-848.A�
r+ i m� a� , � K#
., ��� I6m� a� n� a� k& a� , � I
6., ��� K
#o$ u4 s) a� k& a� , � M/ ., ��� F
�u4 k&u4 t� a� , � A
�.,���
a� n� d �
T( a� m� u4 r+ a� , � G7
.� 1964. Pyrrolnitrin, a newantibiotic substance, produced by Pseudomonas.Agr. Biol. Chem. 28:575-576.B* a� n� g0 e� r+ a� , � M/ .� G7 ., ��� a� n� d
�T( h! o$ m� a� s) h! o$ w% , � L� .� S� .� 1996.
Characterization of a genomic locus required forsynthesis of the antibiotic 2,4-diacetylphloroglucinol by the biological controlagent Pseudomonas fluorescens Q2-87. Mol. Plant-Microbe Interact. 9:83-90.B* a� n� g0 e� r+ a� , � M/ .� G7 ., ��� a� n� d
�T( h! o$ m� a� s) h! o$ w% , � L� .� S� .� 1999.
Identification and characterization of a gene clusterfor synthesis of the polyketide antibiotic 2,4-diacetylphloroglucinol from Pseudomonas fluorescensQ2-87. J. Bacteriol. 181:3155-3163.B* a� r+ o$ n� , � S� ., ��� T( e� r+ a� n� o$ v8 a� , � G
7., ��� a� n� d
�R o$ w% e� , � J
�.� J� .� 1989.
Molecular mechanism of the antimicrobial action ofpyocyanin. Curr. Microbiol. 18:223-230.B* e� n� c9 i n� i , � D
:.� A� ., ��� H
,o$ w% e� ll��� , � C
.� R ., ��� a� n� d
�W;
i l� d� , � J�.� R . � 1983.
Production of phenolic metabolites by a soilpseudomonad. Soil Biol. Biochem. 15:491-491.B* e� n� d
�e� r+ , � C
.� L� ., ��� R a� n� g0 a� s) w% a� m� y3 , � V
"., ��� a� n� d
�L� o$ p� e� r+ , � J
�.� E .�
1999. Polyketide production by plant-associatedpseudomonads. Ann. Rev. Phytopathol. 37:175-196.B* l�u4 m� e� r+ , � C
., ��� a� n� d
�H,
aa�<� s) , � D:
.� 2000. Iron regulation ofthe hcnABC genes encoding hydrogen cyanidesynthase depends on the anaerobic regulator ANRrather than on the global activator GacA inPseudomonas fluorescens CHA0. Microbiol.146:2417-2424.B* o$ n� s) a� ll��� , � R .� F� ., ��� W
;e� l� l� e� r+ , � D
:.� M/ ., ��� a� n� d
�T( h! o$ m� a� s) h! o$ w% , � L� .� S� .�
1997. Quantification of 2,4-diacetylphloroglucinolproduced by fluorescent Pseudomonas sp. in vitroand in the rhizosphere of wheat. Appl. Environ.Microbiol. 63:951-955.B* r+ a� u4 n� , � P1 .� G7 ., ��� H
,i l� d� e� b� r+ a� n� d
�, � P1 .� D: ., ��� E l
�l�s) , � T( .� C ., ��� a� n� d
�K#
o$ b�
a� y3 a� s) h! i , � D:
.� Y= .� 2001. Evidence andcharacterization of a gene cluster required for theproduction of viscosin, a lipopeptide biosurfactant,by a strain of Pseudomonas fluorescens. Can. J.Microbiol. 47:294-301.B* u4 r+ k& h! e� a� d� , � K
#.� D: ., ��� S� c9 h! i s) l� e� r+ , � D
:.� A� ., ��� a� n� d
�S� l�i n� i n� g0 e� r+ , � P1 .� J� .�
1994. Pyrrolnitrin production by biological controlagent Pseudomonas cepacia B37w in culture and in
colonized wounds of potatoes. Appl. Environ.Microbiol. 60:2031-2039.B* y3 n� g0 , � G
7.� S� ., ��� a� n� d
�T( u4 r+ n� e� r+ , � J
�.� M/ .� 1977. Incorporation
of [14C] shikimate into phenazines and their furthermetabolism by Pseudomonas phenazinium. Biochem.J. 164:139-145.C
a� rr+5+ o$ ll���
, � H,
., ��� M/ o$ ë> nn�?� e� -' L� o$ cc9@9 o$ zA , � Y=
., ��� D:
o$ w% l�i n� g0 , � D
:.� N� .,���
a� n� d �
OB ’C G7 a� r+ a� , � F�
.� 1995. Mutational disruption of thebiosynthesis genes coding for the antifungalmetabolite 2,4-diacetyl-phloroglucinol does notinfluence the ecological fitness of Pseudomonasfluorescens F113 in the rhizosphere of sugarbeets.Appl. Environ. Microbiol. 61:3002-3007.C
a� r+ t� w% r+ i g0 h! t� , � D:
.� K# ., ��� C
h! i l� t� o$ n� , � W;
.� S� ., ��� a� n� d �
B* e� n� s) o$ n� ,�D:
.� M/ .� 1995. Pyrrolnitrin and phenazine productionby Pseudomonas cepacia, strain 5.5B, a biocontrolagent of Rhizoctonia solani. Appl. Microbiol.Biotechnol. 43:211-216.C
h! a� n� c9 e� y3 , � S� .� T( ., ��� W;
oo$D$ d�
, � D:
.� W; ., ��� a� n� d �
P1 i e� r+ s) o$ n � III6E6E6
, � L� .� S� .�1999. Two-component transcriptional regulation ofN-acyl-homoserine lactone production inPseudomonas aureofaciens. App. Environ. Microbiol.65:2294-2299.C
h! a� n� g0 , � C
.� J� .� , � F�
l�o$ ss)F) , � H
,.� G7 .� , � H
,oo$D$ k
&, � D:
.� G7 ., ��� M/ a� b� e� , � J�.� A� .,���
M/ a� nn�G� i , � P1 .� E ., ��� M/ a� r+ t� i n� , � L� .� L� ., ��� S� c9 h! r+ o$ e� d� e� r+ , � K#
., ��� a� n� d�
S� h! i e� h! , � T( .� L� .� 1981.The biosynthesis of the antibioticpyrrolnitrin by Pseudomonas aureofaciens. J.Antibiot. 24:555-566.C
h! e� r+ n� i n� , � L� ., ��� B* r+ a� n� d�
i s) , � A�
., ��� I6s) m� a� i l� o$ v8 , � ZH ., ��� a� n� d
�C
h! e� t� , � I6.�
1996. Pyrrolnitrin production by an Enterobacteragglomerans strain with broad-spectrum activitytowards fungal and bacterial phytopathogens. Curr.Microbiol. 32:208-212.C
h! i n� -' A� -' W; o$ e� n� g0 , � T( .� F� .� C .� 2000. Molecular basis ofbiocontrol of tomato and foot and root rot byPseudomonas chlororaphis strain PCL1391. PhD.thesis. Leiden Univ., The Netherlands. 167 pp.C
h! i n� -' A� -' W; o$ e� n� g0 , � T( .� F� .� C .� , � B* l�o$ e� m� b
�e� r+ g0 , � G
7., ��� v8 a� n � d
�e� r+
B* i j� , � A�
., ��� v8 a� n � d�
e� r + D:
r+ i f. t� , � K#
.� M/ .� G7 .� M/ ., ��� S� c9 h! r+ i p� s) e� m� a� , � J�.,���
K#
r+ oo$D$ n� , � B* ., ��� S� c9 h! e� ff.I. e� r+ , � R .� J� ., ��� K#
ee�J� l� , � C
., ��� B* a� kk&D&
e� r+ ,�P1 .� A� .� H, .� M/ ., ��� T( i c9 h! y3 , � H
,.� -' V" ., ��� d
�e � B* r+ u4 i j� n� , � F
�.� J� ., ��� T( h! o$ m� a� s) -'
OB a� t� e� s) , � J�.� E ., ��� a� n� d
�L� u4 g0 t� e� n� b
�e� r+ g0 , � B* .� J� .� J� .� 1998. Biocontrol
by phenazine-1-carboxamide- producingPseudomonas chlororaphis PCL1391 of tomato rootand crown rot caused by Fusarium oxysorum f.sp.radicis-lycopersici. Mol. Plant-Microbe Interact.11:1069-1077.C
h! i n� -' A� -' W; o$ e� n� g0 , � T( .� F� .� C .� , � T( h! o$ m� a� s) -' OB a� t� e� s) , � J�.� E .,���
L� u4 g0 t� e� n� b�
e� r+ g0 , � B* .� J� .� J� ., ��� a� n� d �
B* l�o$ e� m� b
�e� r+ g0 , � G
7.� 2001.
Introduction of the phzH gene of Pseudomonaschlororaphis PCL1391 extends the range ofbiocontrol ability of phenazine-1-carboxylic acid-producing Pseudomonas spp. strains. Mol. Plant-Microbe Interact. 14:1006-1015.C
o$ h! e� n� , � R ., ��� R i o$ v8 , � J�., ��� L� i s) k& e� r+ , � N
�., ��� a� n� d
�K#
a� t� a� n� , � J�. � 1986.
Involvement of ethylene in herbicide-inducedresistance to Fusarium oxysporum f.sp. melonis.Phytopathol. 76:1281-1285.

CK hL
aM ptNPO eQ r R 1: S?T GU eQ nV eQ rR aM l W
IX nV tO rR oY dZ
u[ c\ tO i] oY n V aM nV d Z
Ou[ tO lWi] nV eQ
25
C_ o` ra bb ec lld�d , e Nf .g , e ah ni d j Lk
opl ec ra , e Jm.g En .g 1995. A global
regulator of secondary metabolite production inPseudomonas fluorescens Pf-5. J. Bacteriol. 177:6230-6236.C_ ra o` ni io ni , e Dp ., g e Mq oec nniGi ec -r Lk o` ccs@s o` zt , e Y
u., g e F
vec ni tw oni , e A
x.,g e
Dp uy nniGi ec , e C_ ., g e Dp owz ld io ni g{ , e Dp .g Nf . g ah ni d j O| ’} G~ ah ra ah , e Fv
.g1997(a). Role of 2,4-diacetylphloro-glucinol in theinteractions of the biocontrol pseudomonad strainF113 with the potato cyst nematode Globoderarostochiensis. Appl. Environ. Microbiol. 63:1357-1361.C_ ra o` ni io ni , e Dp ., g e Mq oec nniGi ec -r Lk o` ccs@s o` zt , e Y
u., g e F
vec ni tw oni , e A
x.,g e
Dp uy nniGi ec , e C_ ., g e Dp o` wz ld io ni g{ , e Dp .g Nf ., g e ah ni d j O| ’} G~ ah ra ah , e Fv
.g1997(b). Ecological interaction of a biocontrolPseudomonas fluorescens strain producing 2,4-diacetylphloroglucinol with the soft rot potatopathogen Erwinia caratovora subsp. atroseptica.FEMS Microbiol. Ecol. 23:95-106.Dp ec ld ah ni y� , e I� ., g e S� h
�ec ec h� ah ni , e Mq .g Mq ., g e F
vec ni tw oni , e A
x.g , e B�
ah ra dj io ni , e S� .,g eAx
ah ra oni s� , e S� .g , e ah ni d j O| ’} G~ ah ra ah , e Fv
.g 2000. Regulation ofproduction of the antifungal metabolite 2,4-diacetylphloroglucinol in Pseudomonas fluorescensF113: genetic analysis of phlF as a transcriptionalrepressor. Microbiol. 146:537-546.Dp uy f
�f�y� , e B
�.g K� ., g e ah ni d j Dp é
�f�ah g{ o, e G~ .g 1997. Zinc improves
biocontrol of fusarium crown and root rot oftomato by Pseudomonas fluorescens and represses theproduction of pathogen metabolites inhibitory tobacterial antibiotic biosynthesis. Phytopathol.87:1250-2157.Dp uy f
�f�y� , e B
�.g K� ., g e ah ni d j Dp é
�f�ah g{ o` , e G~ .g 1999. Environmental
factors modulating antibiotic and siderophorebiosynthesis by Pseudomonas fluorescens biocontrolstrains. Appl. Environ. Microbiol. 65:2429-2438.En dj wz ah ra dj s� , e J
m.g R� ., g e ah ni d j H
�ah y� ah s� h� io , e J
m.g Ax .g 1965. Structure of
a rhamnolipid from Pseudomonas aeruginosa. Arch.Biochem. Biophys. 111:415-421.En ld ah ni dj ec ra , e R� .g P� .g , e Mq ah bb ec , e J
m.g Ax .g , e H
�ah m� io lld�d , e R� .g H� .g , e ah ni dj
G~ o` ra m� ah ni , e Mq .g 1968. Metabolism of tryptophans byPseudomonas aureofaciens. VI. Production ofpyrrolnitrin by selected Pseudomonas spp. Appl.Microbiol. 16:753-758.En ld -r B� ah nniGi ah , e Nf ., g e ah ni d j W
�io ni k�ec ld m� ah nniGi , e G~ .g 1988.
Pyrrolnitrin from Burkholderia cepacia: antibioticactivity against fungi and novel activities againststreptomycetes. J. Appl. Microbiol. 85:69-78.En lld�d io s� , e R� .g Jm ., g e T� io mm��� io s� -r W� io ld s� oni , e T� .g Mq ., g e ah ni d j B
�ah io ld ec y� , e Mq .g Jm .g
2000. Identification of conserved traits influorescent pseudomonads with antifungal activity.Environ. Microbiol. 2:274-284 .Fv
ec ni tw o` ni , e Ax
. g Mq ., g e S� tw ec pl h�
ec ni s� , e P� . g Mq ., g e C_ ra o` wz ld ec y� , e Jm.g ,e
O| ’} C_ ah ld ld ah g{ h�
ah ni , e Mq ., g e ah ni d j O| ’} G~ ah ra ah , e Fv
.g 1992.Exploitation of gene(s) involved in 2,4-diacetylphloroglucinol biosynthesis to confer a newbiocontrol capability to a Pseudomonas strain. Appl.Environ. Microbiol. 58:3873-3878. F
vio ec cs h� tw ec ra , e A
x.g
1992. Biosurfactants: moving towards industrialapplication. Tibtech 10:208-217.Fv
ld ah io s� h� m� ah ni , e Mq ., g e En y� ah ld , e Z�
., g e V�
o` io s� ah ra dj , e C_ ., g e ah ni d j H�
aah<h s� , e Dp .g1990. Suppression of Septoria tritici by phenazine-or siderophore-deficient mutants of Pseudomonas.Cur. Microbiol. 20:121-124.
Fv
ra ah v� ec ld , e Dp .g R� .g 1988. Role of antibiosis in thebiocontrol of plant diseases. Ann. Rev. Phytopathol.26:75-91.Fv
uy q� uy ah , e C_ ., g e W�
io ni ah ni s� , e S� .g C_ ., g e ah ni d j G~ ra eec@c ni bb ec ra g{ , e En .g P� .g1996. Census and consensus in bacterialecosystems: the LuxR-LuxI family of quorum-sensing transcriptional regulators. Ann. Rev.Microbiol. 50:727-751.G~ ah ff��� ni ec y� , e T� .g Dp ., g e L
kah m� , e S� ., g e L
kio g{ o` ni , e J
m.g , e G~ ah tw ec s� , e K� .,g e
Fv
ra ah zt ec lld�d ec , e Ax
., g e Dp i o Mq ah io o` , e Jm., g e H
�io ld ld , e Dp .g S� ., g e G~ ooD dj wz io ni , e S� .,g e
T� o` ra k� ec wz io tw zt , e Nf ., g e Ax
lld�d s� h� o` uy s� ec , e Ax
.g Mq ., g e K� ec m� pl f�, e H�
.g -r Jm ., g e ah ni djB�
ec cs k� ec ra , e Jm.g O| .g 1994. Global regulation of expression
of antifungal factors by a Pseudomonas fluorescensbiocontrol strain. Mol. Plant-Mcrobe Interact.7:455-463.G~ ec o` ra g{ ah k� o` pl ouy ld o` s� , e Dp .g G~ ., g e H
�ec ni dj s� oni , e Mq ., g e P� ah ni o` pl ouy ld o` s� ,e
Nf .g Jm ., g e ah ni d j S� cs h� ra otw h� , e Mq .g Nf .g 1994. Analysis ofexpression of a phenazine biosynthetic locus ofPseudomonas aureofaciens PGS12 on seeds with amutant carrying a phenazine biosynthesis locus-icenucleation reporter gene fusion. Appl. Environ.Microbiol. 60:4573-4579.G~ ec ra ah ra dj , e J
m., g e Lk
ld oy� dj , e R� ., g e B�
ah ra s� bb y� , e T� ., g e H�
ah dj ec ni , e P� ., g e K� ec lld�d y� ,eMq .g T� ., g e ah ni d j A
xni dj ec ra s� ec ni , e R� .g Jm .g 1997. Massetolides A-H,
antimycobacterial cyclic depsipeptides produced bytwo pseudomonads isolated from marine habitats. J.Nat. Prod. 60:223-229.G~ odj f
�ra ec y� , e S� .g Ax .g C_ ., g e H
�ah ra ra o` wz , e S� .g Ax ., g e Mq ah ra s� h� ah ld ld , e J
m.g W� .g , e ah ni dj
K� ld ec ni ah , e Jm.g Dp .g 2001. Characterization by 16S rRNA
analysis of pseudomonads causing blotch disease ofcultivated Agaricus bisporus. Appl. Environ.Microbiol. 67:4316-4323.H�
ah mm��� ec ra , e P� .g En ., g e ah ni d j En v� ec ni s� ec ni , e K� .g B� .g 1993. Postharvest control of Botrytis cinerea on cut flowerswith pyrrolnitrin. Plant Dis. 77:283-286.H�
ah mm��� ec ra , e P� .g En ., g e H�
io lld�d , e Dp .g S� ., g e Lk
ah m� , e S� .g T� ., g e v� ah n i P� ée� c , e K� .g -r
H�
., g e ah ni d j Lk
io g{ o` ni , e Jm.g Mq .g 1997. Four genes from
Pseudomonas fluorescens that encode the biosynthesisof pyrrolnitrin. Appl. Environ. Microbiol. 63:2147-2154.H�
ah mm��� ec ra , e P� .g En ., g e B�
uy ra dj , e W�
., g e H�
io lld�d , e Dp .g S� ., g e Lk
io g{ o` ni , e Jm.g Mq .,g e
ah ni d j v� ah n i P� ée� c , e K� .g -r H� .g 1999. Conservation of the
pyrrolnitrin gene cluster among six pyrrolnitrin-producing strains. FEMS Microbiol. Lett. 180:39-44.H�
ah ni dj ec ld s� m� ah ni , e Jm.g , e ah ni d j S� tw ah bb bb , e En .g V� .g 1996. Biocontrol
of soilborne plant pathogens. Plant Cell 8:1855-1869.H�
ah ss�F� ah ni , e H�
.g Mq .g , e ah ni d j Fv
ra io dj o` v� io cs h� , e I� .g 1980. Mechanismof antibiotic action of pyocyanine. J. Bacteriol.141:156-163.H�
ah uy s� ec ra , e G~ ., g e ah ni d j K� ah ra ni o` v� s� k� y� , e Mq .g Lk .g 1957. Rhamnoseand rhamnolipid biosynthesis by Pseudomonasaeruginosa. J. Biol. Chem. 224:91-105.H�
eecJc bb , e S� ., g e ah ni d j H�
aah<h s� , e Dp .g 2001. Regulatory roles ofthe GacS/GacA two-component system in plant-associated and other gram negative bacteria. Mol.Plant-Microbe Interact. 14:1351-1363.H�
io ld dj ec bb ra ah ni dj , e P� .g Dp ., g e B�
ra ah uy ni , e P� .g G~ ., g e Mq cs R� ah ec , e K� .g B� ., g e ah ni djLk
uy , e X�
.g 1998. Role of the biosurfactant viscosin inbroccoli head rot caused by pectolitic strains of

C� h�
a� pt�P� e� r � 1: �?� G� e� n e� r� a� l ¡
I¢ n t� r� o£ d¤
u¥ c¦ t� i§ o£ n a� n d ¤
Ou¥ t� l¡i§ n e�
26
Pseudomonas fluorescens. Can. J. Plant Pathol.20:296-303.H© iªll«�«
, ¬ D .® S¯ ., ®�¬ St° e± iª n² , ¬ J³.® I., ®�¬ Tµ o¶ r· k¸ e± w¹ i
ªt° zº , ¬ N» .® R¼ ., ®�¬ M
½o¶ r· s¾ e± ,¬
A¿
.® M½ ., ®�¬ H© o¶ w¹ e± ll«�« , ¬ CÀ
.® R¼ ., ®�¬ PÁ
a cà hÄ l«a t° k¸ o¶ , ¬ J
³.® PÁ ., ®�¬ BÅ e± cà k¸ e± r· , ¬ J
³.® OÆ .,®�¬
a n² d Ç
LÈ
iªgÉ o¶ n² , ¬ J
³.® M½ .® 1994. Cloning of genes involved in
the synthesis of pyrrolnitrin from Pseudomonasfluorescens and role of pyrrolnitrin synthesis inbiological control of plant disease. Appl. Environ.Microbiol. 60:78-85.H© iªr· a mÊ o¶ t° o¶ , ¬ M
½., ®�¬ OÆ ka dÇ a , ¬ K
Ë., ®�¬ a n² d
ÇN» a gÉ a iª , ¬ S.® 1970. The
revised structure of viscosin, a peptide antibiotic.Tetrah. Lett. 13:1087-1090.H© iªr· a yÌ a mÊ a , ¬ Tµ ., ®�¬ a n² d
ÇKË
a t° o¶ , ¬ I´ .® 1982. Novel methylrhamnolips from Pseudomonas aeruginosa. FEBSLett. 139:81-85.H© iªs¾ a t° s¾ uÍ ka , ¬ K
Ë., ®�¬ N» a k¸ a hÄ a r· a , ¬ Tµ .® , ¬ Sa n² o¶ , ¬ N» ., ®�¬ a n² d
ÇYÎ
a mÊ a dÇ a , ¬ KË
.® 1971. Formation of rhamnolipid byPseudomonas aeruginosa and its function inhydrocarbon fermentation. Agr. Biol. Chem.35:686-692.H© o¶ w¹ e± ll«�« , ¬ C
À.® R¼ ., ®�¬ a n² d
ÇS¯t° iª pÏ a n² o¶ vÐ i
ªcà , ¬ R¼ .® D .® 1980.
Suppression of Pythium ultimum-induced damping-off of cotton seedlings by Pseudomonas fluorescensand its antibiotic, pyoluteorin. Phytopathol.70:712-715.H© r· a bÑ a cà k¸ , ¬ EÒ .® M½ ., ®�¬ a n² d
ÇWÓ
iªll«�«
iªs¾ , ¬ D .® KË .® 1992. The lemA
gene is required for pathogenesis of Pseudomonassyringae pv. syringae. J. Bacteriol. 174:3011-3020.H© uÍ t° cà hÄ i
ªs¾ o¶ n² , ¬ M
½.® LÈ ., ®�¬ a n² d
ÇJ³o¶ hÄ n² s¾ t° o¶ n² e± , ¬ K
Ë.® 1993.
Evidence for the involvement of the surface activeproperties of the extracellular toxin tolaasin in themanifestation of brown blotch disease symptms byPseudomonas tolaasii on Agaricus bisporus. Physiol.Mol. Plant Pathol. 42:373-384.H© uÍ t° cà hÄ i
ªs¾ o¶ n² , ¬ M
½.® LÈ ., ®�¬ a n² d
ÇGÔ
r· o¶ ss¾5¾ , ¬ D .® CÀ .® 1997.Lipopeptide phytotoxin produced by Pseudomonassyringae pv. syringae: comparison of thebiosurfactant and ion channel forming activities ofsyringopeptin and syringomycin. Mol. Plant-Microbe Interact. 10:347-354.Ia cà o¶ bÑ e± ll«�« iª s¾ , ¬ N» .® S., ®�¬ L
Èa vÐ e± r· mÊ i
ªcà o¶ cà cà a , ¬ P
Á., ®�¬ G
Ôr· gÉ uÍ r· iª n² a , ¬ I´ .,®�¬
SiªmmÊ�Ê a cà o¶ , ¬ M
½., ®�¬ a n² d
ÇBÅ a ll«�« iª o¶ , ¬ A
¿.® 1992. Phytotoxic
properties of Pseudomonas syringae pv. syringaetoxins. Physiol. Mol. Plant Pathol. 40:107-116.IgÉ l«e± w¹ s¾ k¸ i
ª, ¬ BÅ .® 1989. Probing Pseudomonas aeruginosa,
an opportunistic pathogen. ASM News 55:303-307.It° o¶ hÄ , ¬ S., ®�¬ H© o¶ n² d
Ça , ¬ H© ., ®�¬ Tµ o¶ mÊ i
ªt° a , ¬ F
Õ., ®�¬ a n² d
ÇSuÍ zº uÍ k
¸iª, ¬ Tµ .®
1971. Rhamnolipids produced by Pseudomonasaeruginosa grown on n-paraffin (mixture of C12, C13
and C14 fractions). J. Antibiot. 24:855-859.J³a n² i
ªs¾ iª e± w¹ i
ªcà zº , ¬ W
Ó.® J³ ., ®�¬ a n² d
ÇR¼ o¶ i
ªt° mÊ a n² , ¬ J
³.® 1988. Biological
control of blue mold and grey mold on apple andpear with Pseudomonas cepacia. Phytopathol.78:1697-1700.J³a r· vÐ i
ªs¾ , ¬ F
Õ.® GÔ ., ®�¬ a n² d
ÇJ³o¶ hÄ n² s¾ o¶ n² , ¬ M
½.® J³ .® 1949. A glycolipid
produced by Pseudomonas aeruginosa. J. Am. Chem.Soc. 71:4124-4126.J³a yÌ a s¾ w¹ a l« , ¬ R¼ .® KË ., ®�¬ F
Õe± r· n² a n² d
Çe± zº , ¬ M
½., ®�¬ UÖ pÏ a dÇ hÄ yÌ a yÌ , ¬ R¼ .® S.,®�¬
V×
iªs¾ iª n² t° iª n² , ¬ L
È., ®�¬ K
ËuÍ r· zº , ¬ M
½., ®�¬ WÓ
e± bbÑ2Ñ , ¬ J³.® , ¬ a n² d
ÇR¼ iªn² e± hÄ a r· t° , ¬ K
Ë.®
1993. Antagonism of Pseudomonas cepacia againstphytopathogenic fungi. Curr. Microbiol. 26:17-22.
J³iªmÊ e± n² e± zº -Ø EÒ s¾ cà r· iª gÉ , ¬ A
¿., ®�¬ J³iªmÊ e± n² e± zº -Ø J³ i
ªmÊ e± n² e± zº , ¬ I., ®�¬ P
ÁuÍ l«iªdÇ
o¶ , ¬ R¼ .,®�¬a n² d
ÇSa uÍ r· a -Ø CÀ a l« iª xÙ t° o¶ , ¬ F
Õ.® 2001. Antioxidant activity of
fresh and processed edible seaweed. J. Sci. FoodAgric. 81:530-534.J³o¶ hÄ n² s¾ e± n² , ¬ K
Ë., ®�¬ A¿
n² dÇ
e± r· s¾ e± n² , ¬ S¯.® , ¬ a n² d
ÇJ³a cà o¶ bÑ s¾ e± n² , ¬ C
À.® S. ® 1996.
Phenotypic and genotypic characterization ofphenanthrene-degrading fluorescent Pseudomonasbiovars. Appl. Environ. Microbiol. 62:3818-3825.KË
a l« bÑ e± , ¬ CÀ
.® , ¬ M½
a r· t° e± n² , ¬ PÁ
., ®�¬ a n² d Ç
BÅ e± r· gÉ , ¬ GÔ
.® 1996. Strainsof the genus Serratia as beneficial rhizobacteria ofoilseed rape with antifungal properties. Microbiol.Res. 151:433-439.KË
ee±J± l« , ¬ CÀ
., ®�¬ WÓ
iªr· t° hÄ n² e± r· , ¬ P
Á.® H© ., ®�¬ OÆ bÑ e± r· hÄ a n² s¾ l« iª , ¬ Tµ .® H© .,®�¬
V×
o¶ iªs¾ a r· dÇ , ¬ C
À., ®�¬ BÅ uÍ r· gÉ e± r· , ¬ UÖ ., ®�¬ H© aaÂ@ s¾ , ¬ D ., ®�¬ a n² d
ÇD éÚ fÛ a gÉ o¶ , ¬ G
Ô.®
1990. Pseudomonads as antagonists of plantpathogens in the rhizosphere: role of the antibiotic2,4-diacetylphloroglucinol in the suppression ofblack root rot of tobacco. Symbiosis 9:327-342.KË
ee±J± l« , ¬ CÀ
., ®�¬ S¯cà hÄ n² i
ªdÇ
e± r· , ¬ UÖ ., ®�¬ M½
a uÍ r· hÄ o¶ fÛe± r· , ¬ M
½., ®�¬ V×
o¶ iªs¾ a r· dÇ , ¬ C
À.,®�¬
LÈ
a vÐ iªll«�«
e± , ¬ J³.® , ¬ BÅ uÍ r· gÉ e± r· , ¬ UÖ ., ®�¬ W
Óiªr· t° hÄ n² e± r· , ¬ P
Á.® H© ., ®�¬ H© aaÂ< s¾ , ¬ D .,®�¬
a n² d Ç
D éÚ fÛ a gÉ o¶ , ¬ GÔ
.® 1992. Suppression of root diseasesby Pseudomonas fluorescens CHA0: importance ofthe secondary metabolite 2,4-diacetylphloroglucinol. Molec. Plant-MicrobeInteract. 5:4-13.KË
ee±J± l« , ¬ CÀ
., ®�¬ WÓ
e± l« l« e± r· , ¬ D .® M½ ., ®�¬ N» a t° s¾ cà hÄ , ¬ A¿
., ®�¬ D éÚ fÛ a gÉ o¶ , ¬ GÔ
.,®�¬CÀ
oo¶D¶ k¸, ¬ R¼ .® J³ ., ®�¬ a n² d
ÇTµ hÄ o¶ mÊ a s¾ hÄ o¶ w¹ , ¬ L
È.® S.® 1996.
Conservation of the 2,4-diacetylphloroglucinolbiosynthesis locus among fluorescent Pseudomonasstrains from diverse geographic locations. Appl.Environ. Microbiol. 62:552-563.KË
iªmÊ , ¬ BÅ .® S., ®�¬ L
Èee±@± , ¬ J
³.® YÎ ., ®�¬ a n² d
ÇH© w¹ a n² gÉ , ¬ BÅ .® KË .® 2000. In
vivo control and in vitro antifungal activity ofrhamnolipid B, a glycilipid antibiotic, againstPhytophthora capsici and Colletotrichum orbiculare.Pest. Manag. Sci. 56:1029-1035.KË
iªr· n² e± r· , ¬ S., ®�¬ H© a mmÊ�Ê e± r· , ¬ P
Á.® EÒ ., ®�¬ H© i
ªl«l«, ¬ D .® S¯ .® , ¬ A
¿l«t° mÊ a nn²G² , ¬ A
¿.® ,¬
FÕ
iªs¾ hÄ e± r· , ¬ I., ®�¬ W
Óiªe± s¾ l« o¶ , ¬ L
È.® J³ .® , ¬ L
Èa n² a hÄ a n² , ¬ M
½., ®�¬ V×
a n ² PÁ
éeÚ ± , ¬ KË
.® -ØH© ., ®�¬ a n² d
ÇLÈ
iªgÉ o¶ n² , ¬ J
³.® M½ .® 1998. Functions encoded by
pyrrolnitrin biosynthetic genes from Pseudomonasfluorescens. J. Bacteriol. 180:1939-1943.KË
r· a uÍ s¾ , ¬ J³., ®�¬ a n² d
ÇLÈ
o¶ pÏ e± r· , ¬ J³.® EÒ .® 1992. Lack of evidence
for a role of antifungal metabolite production byPseudomonas fluorescens Pf-5 in biological control ofPythium damping-off of cucumber. Phytopathol.82:264-271.KË
r· a uÍ s¾ , ¬ J³., ®�¬ a n² d
ÇLÈ
o¶ pÏ e± r· , ¬ J³.® EÒ .® 1995. Chara-cterization
of a genomic region required for production of theantibiotic pyoluteorin by the biological controlagent Pseudomonas fluorescens Pf-5. Appl. Environ.Microbiol. 61:849-854.KË
uÍ r· iª o¶ k¸a , ¬ S
¯., ®�¬ a n² d
ÇLÈ
iªuÍ , ¬ P
Á.® V× .® 1967. Effect of the
hemolysin of Pseudomonas aeruginosa onphosphatides and on phospholipase C activity. J.Bacteriol. 93:670-674.LÈ
a mÊ , ¬ BÅ .® S., ®�¬ S¯t° r· o¶ bÑ e± l« , ¬ G
Ô.® A¿ .® , ¬ H© a rr·E· iª s¾ o¶ n² , ¬ L
È.® A¿ .® , ¬ a n² d
ÇLÈ
a mÊ ,¬S¯.® Tµ .® 1987. Transposon mutagenesis and tagging of
fluorescent Pseudomonas: antimycotic production isnecessary for control of dutch elm disease. Proc.Natl. Acad. Sci. USA 84:6447-6451.LÈ
a vÐ iªll«�«
e± , ¬ J³.® , ¬ V
×o¶ iªs¾ a r· dÇ , ¬ C
À., ®�¬ K
Ëee±J± l« , ¬ C
À.® , ¬ M
½a uÍ r· hÄ o¶ f
Ûe± r· , ¬ M
½.,®�¬
D éÚ fÛ a gÉ o¶ , ¬ GÔ
., ®�¬ a n² d Ç
H© aaÂ<Â s¾ , ¬ D .® 1992. Global control in

CÜ hÝ
aÞ ptßPà eá r â 1: ã?ä Gå eá næ eá râ aÞ l ç
Iè næ tà râ oé dê
uë cì tà ií oé n æ aÞ næ d ê
Oî uë tà lçií næ eá
27
Pseudomonas fluorescens mediating antibioticsynthesis and suppression of black root rot oftobacco. Proc. Natl. Acad. Sci. USA 89:1562-1566.Lï eð iñ sò iñ nó gô eð rõ , ö T
÷., ø ö aù nó d ú Mû aù rõ gô rõ aù ffüIü , ö Rý .ø 1979. Secondary
metabolites of the fluorescent pseudomonads.Microbiol. Rev. 43:422-442.Lï eð vþ yÿ , ö E� ., ø ö G� o� u� gô h
�, ö F� .ø J� ., ø ö B
�eð rõ l� iñ nó , ö K
.ø D ., ø ö G� u� iñ aù nó aù , ö P� .ø W� .,ø ö
aù nó d ú S m� iñ t� h� , ö J�.ø T÷ .ø 1992. Inhibition of Septoria tritici
and other phytopathogenic fungi and bacteria byPseudomonas fluorescens and its antibiotics. PlantPathol. 41:335-341.Lï iñ gô o� nó , ö J
�.ø Mû ., ø ö H
�iñ ll��� , ö D .ø S .ø , ö H
�aù mm��� eð rõ , ö P� .ø E� ., ø ö T
÷o� rõ k� eð w� iñ t� z� ,ö
N� .ø Rý ., ø ö H�
o� füm� aù nnóGó , ö D ., ø ö K
eð m� p� f
ü, ö H�
.ø -� S ., ø ö aù nó dú , ö V�
aù n ó P� ée� ð ,öK
.ø -� H� .ø 2000. Natural products with antifungalactivity from Pseudomonas biocontrol bacteria. PestManag. Sci. 56:688-695.Lï o� nó gô l� eð yÿ , ö Rý .ø P� ., ø ö H
�aù l� l� iñ w� eð ll��� , ö J
�.ø E� ., ø ö C� aù m� p� b� eð l� l� , ö J
�.ø J� .ø Rý ., ø ö aù nó dú
I� nó gô l� eð dú eð w� , ö W�
.ø Mû .ø 1972. The branch point ofpyocyanine biosynthesis. Can. J. Microbiol.18:1357-1368.Lï o� p� eð rõ , ö J
�.ø E� ., ø ö aù nó d ú Lï iñ nó dú o� w� , ö S .ø E� .ø 1997. Reporter gene
systems useful in evaluating in situ gene expressionby soil and plant-associated bacteria. In: Hurst,C.J., Knudsen, G.R., McInerney, M.J.,Stetzenbach, L.D., Walter, M.V. (eds). Manual ofEnvironmental Microbiology, pages 482-492.American Society for Microbiology (ASM),Washington, USA.Mû aù rõ aù h� iñ eð l� , ö Mû .ø A� ., ø ö S t� aù c h� eð l� h� aù u� sò , ö T
÷., ø ö aù nó d ú Mû oo�!� t� z� , ö H
�.ø D .ø
1997. Modular peptide synthases involved in non-ribossomal peptide synthesis. Chem. Rev. 97:2651-2673.Mû aù u� rõ h� o� ff
üIüeð rõ , ö Mû ., ø ö K
eeð@ð l� , ö C� ., ø ö S c h� nó iñ dú eð rõ , ö U" ., ø ö V
�o� iñ sò aù rõ dú ,ö
C� ., ø ö H�
aaù<ù sò , ö D ., ø ö aù nó d ú D é� fü aù gô o� , ö G� .ø 1992. Influence ofenhanced antibiotic production in Pseudomonasfluorescens strain CHA0 on its disease suppressivecapacity. Phytopathol. 82:190-195.Mû aù u� rõ h� o� ff
üIüeð rõ , ö Mû ., ø ö K
eeð@ð l� , ö C� ., ø ö H
�aaù<ù sò , ö D .ø , ö aù nó d ú D é� fü aù gô o� ,ö
G� .ø 1994. Pyoluteorin production by Pseudomonasfluorescens strain CHA0 is involved in thesuppression of Pythium damping-off of cress butnot of cucumber. Eur. J. Plant Pathol. 100:221-232.Mû aù u� rõ h� o� ff
üIüeð rõ , ö Mû ., ø ö K
eeð@ð l� , ö C� ., ø ö H
�aaù<ù sò , ö D .ø , ö aù nó d ú D é� fü aù gô o� ,ö
G� .ø 1995. Influence of plant species on diseasesuppression by Pseudomonas fluorescens strain CHA0with enhanced antibiotic production. Plant Pathol.44:40-50.Mû aù vþ rõ o� dú iñ , ö D .ø V� ., ø ö K
sò eð nó z� eð nó k
�o� , ö V
�.ø N� ., ø ö B
�o� nó sò aù ll��� , ö Rý .ø F� .ø ,ö
C� oo�!� k�, ö Rý .ø J� ., ø ö B
�o� rõ o� nó iñ nó , ö A
�.ø Mû ., ø ö aù nó d ú T
÷h�
o� m� aù sò h� o� w� , ö Lï .ø S .ø1998. A seven-gene locus for synthesis ofphenazine-1-carboxylic acid by Pseudomonasfluorescens 2-79. J. Bacteriol. 180:2541-2548.Mû aù vþ rõ o� dú iñ , ö D .ø V� ., ø ö B
�o� nó sò aù ll��� , ö Rý .ø F� ., ø ö D eð l� aù nó eð yÿ , ö S .ø Mû .ø ,ö
S o� u� l� eð , ö Mû .ø J� ., ø ö P� h�
iñ ll��� iñ p� sò , ö G� ., ø ö aù nó d ú T÷
h�
o� m� aù sò h� o� w� , ö Lï .ø S .ø2001. Functional analysis of genes for biosynthesisof pyocyanin and phenazine-1-carboxamide fromPseudomonas aeruginosa PAO1. J. Bacteriol.183:6454-6465.Mû aù zz�#� o� l� aù , ö Mû ., ø ö C� oo�!� k
�, ö Rý .ø J� ., ø ö T
÷h�
o� m� aù sò h� o� w� , ö Lï .ø S .,ø öW�
eð ll��� eð rõ , ö D .ø Mû ., ø ö P� iñ eð rõ sò o� n ó III�$�%� , ö Lï .ø S .ø 1992. Contributionof phenazine antibiotic biosynthesis to the
ecological competence of fluorescent pseudomonadsin soil habitats. Appl. Environ. Microbiol. 2616-2624.Mû c S p� aù ddú ú eð nó -� G� aù rõ dú eð nó eð rõ , ö B
�.ø B� ., ø ö S c h� rõ o� eð dú eð rõ , ö K
.ø Lï .,ø ö
K
aù ll��� o� gô eð rõ , ö S .ø E� ., ø ö Rý aaù<ù iñ j& m� aù k� eð rõ sò , ö J�.ø Mû .ø , ö T
÷h�
o� m� aù sò h� o� w� ,öLï .ø S ., ø ö aù nó d ú W
�eð ll��� eð rõ , ö D .ø Mû . ø 2000. Genotypic and
phenotypic diversity of phlD-containingPseudomonas isolated from the rhizosphere of wheat.Appl. Environ. Microbiol. 66:1939-1946.Mû iñ ll��� eð rõ , ö C� .ø Mû .ø , ö Mû iñ ll��� eð rõ , ö Rý .ø V� ., ø ö G� aù rõ t� o� nó -� K eð nnó?ó yÿ , ö D .,ø öRý eð dú gô rõ aù vþ eð , ö B
�., ø ö S eð aù rõ sò , ö J
�.ø , ö C� o� nó dú rõ o� nó , ö Mû .ø Mû ., ø ö T
÷eð p� l� o� w� ,ö
D .ø B� ., ø ö aù nó d ú S t� rõ o� b� eð l� , ö G� .ø A� .ø 1998. Ecomycins, uniqueantimycotics from Pseudomonas viridiflava. J. Appl.Microbiol. 84:937-944.Mû o� rõ iñ k� aù w� aù , ö Mû ., ø ö D aù iñ dú o� , ö H
�., ø ö T
÷aù k� aù o� , ö T
÷., ø ö Mû u� rõ aù t� aù , ö S .,ø ö
S h�
iñ m� o� nó iñ sò h� iñ , ö Y'
., ø ö aù nó d ú I� m� aù nó aù k� aù , ö T÷
.ø 1993. A newlipopeptide biosurfactant produced by Arthrobactersp. strain MIS38. J. Bacteriol. 175:6459-6466.Mû o� rõ t� iñ sò h� iñ rõ eð -� S m� iñ t� h� , ö Rý .ø J� ., ø ö N� u� t� k� iñ nó sò , ö J
�.ø C� ., ø ö P� aù c k� m� aù nó ,ö
Lï .ø C� ., ø ö B�
rõ o� dú eð yÿ , ö C� .ø Lï ., ø ö Rý aù iñ nó eð yÿ , ö P� .ø B� ., ø ö J�o� h�
nó sò t� o� nó eð , ö K
.,ø öaù nó d ú W
�iñ ll��� iñ aù m� sò , ö D .ø H� .ø 1991. Determination of the
structure of an extracelular peptide produced by themushroom saprotroph Pseudomonas reactans.Tetrah. 47:3645-3654.N� iñ eð l� sò eð nó , ö T
÷.ø H� ., ø ö C� h
�rõ iñ sò t� o� p� h
�eð rõ sò eð nó , ö C� ., ø ö A
�nó t� h� o� nó iñ , ö U" .,ø ö
aù nó d ú S ø( rõ eð nó sò eð nó , ö J�.ø 1999. Viscosinamide, a new cyclic
depsipeptide with surfactant and antifungalproperties produced by Pseudomonas fluorescensDR54. J. Appl. Microbiol. 86:80-90.N� iñ eð l� sò eð nó , ö T
÷.ø H� ., ø ö T
÷h�
rõ aù nó eð , ö C� .ø , ö C� h�
rõ iñ sò t� o� p� h�
eð rõ sò eð nó , ö C� .,ø öA�
nó t� h� o� nó iñ , ö U" ., ø ö aù nó d ú S ø( rõ eð nó sò eð nó , ö J�.ø 2000. Structure,
production characteristics and fungal antagonism oftensin – a new antifungal cyclic lipopeptide fromPseudomonas fluorescens 96.578. J. Appl. Microbiol.89:992-1001.N� o� t� z� , ö Rý ., ø ö Mû aù u� rõ h� o� f
üeð rõ , ö Mû ., ø ö S c h� nó iñ dú eð rõ -� K eeð@ð l� , ö U" .ø ,ö
D u� füfüyÿ , ö B
�.ø K ., ø ö H
�aaù<ù sò , ö D ., ø ö aù nó d ú D é� fü aù gô o� , ö G� .ø 2001. Biotic
factors affecting expression of the 2,4-diacetylphloroglucinol biosynthesis gene phlA inPseudomonas fluorescens strains CHA0 in therhizosphere. Phytopathol. 91:873-881.N� o� w� aù k� -� T÷ h
�o� m� p� sò o� nó , ö B
�., ø ö G� o� u� l� dú , ö S .ø J� ., ø ö aù nó d ú Lï o� p� eð rõ , ö J
�.ø E� .ø
1997. Identification and sequence analysis of thegenes encoding a polyketide synthase required forpyoluteorin biosynthesis in Pseudomonas fluorescensPf-5. Gene 204:17-24.N� o� w� aù k� -� T÷ h
�o� m� p� sò o� nó , ö B
�., ø ö C� h
�aù nó eð yÿ , ö N� ., ø ö W
�iñ nó gô , ö J
�.ø S .ø ,ö
G� o� u� l� dú , ö S .ø J� ., ø ö aù nó d ú Lï o� p� eð rõ , ö J�.ø E� .ø 1999. Characterization
of the pyoluteorin biosynthetic gene cluster ofPseudomonas fluorescens Pf-5. J. Bacteriol. 181:2166-2174.O)
c h� sò nó eð rõ , ö U" .ø A� ., ø ö aù nó d ú Rý eð iñ sò eð rõ , ö J�.ø 1995. Autoinducer-
mediated regulation of rhamnolipid biosurfactantsynthesis in Pseudomonas aeruginosa. Proc. Natl.Acad. Sci. USA 92:6424-6428.O)
h�
m� o� rõ iñ , ö T÷
., ø ö H�
aù gô iñ w� aù rõ aù , ö S .ø -� I� ., ø ö U" eð dú aù , ö A�
., ø ö Mû iñ nó o� dú aù , ö Y'
.,ø öaù nó d ú Y
'aù m� aù dú aù , ö K
.ø 1978. Production of pyoluteorin
and its derivatives from n-paraffin by Pseudomonasaeruginosa S10B2. Agr. Biol. Chem. 42:2031-2036.P� aù ll��� eð rõ o� nó iñ , ö N� .ø J� .ø 1992. Human and animal-pathogenic pseudomonads. In: Balows, A., Truper,H.G., Dworkin, M., Harder, W., and Schleifer,

C* h+
a, pt-/. e0 r 1 1: 243 G5 e0 n6 e0 r1 a, l 7
I8 n6 t. r1 o9 d:
u; c< t. i= o9 n 6 a, n6 d :
O> u; t. l7i= n6 e0
28
K.H. (eds). The prokariotes, a handbook on thebiology of bacteria, ecophysiology, isolation,identification and applications, 2nd ed., vol.3.Springer, New York, N.Y., USA.P? a@ ssABA a@ dC oD rE , F L
G., H F CI ooD!D k
J, F JK.H ML ., H F GM a@ mN bO eP llQ�Q oD , F M
L.H JK .H , F R
RuS sA tT , F L
G.,H F
a@ nU d C
IV gW lQ eP wX sA kJ iY , F BZ .H H[ .H 1993. Expression ofPseudomonas aeruginosa virulence genes requirescell-to-cell communication. Science 260:1127-1130.P? eP rE nU eP sA tT iY gW , F A
\.H -] K., H F M
LeP lQ eP f_ oD rE sA , F Ö.H , F a@ nU d
CGM eP oD rE gW eP llQ�Q iY sA , F D
a.H
2001. Identification of uvrY as the cognate responseregulator for the barA sensor kinase in Escherichiacoli. J. Biol. Chem. 276:225-231.P? eP sA cb iY , F Ec .H CI ., H F P? eP a@ rE sA oD nU , F J
K.H P? ., H F Sd eePeP dC , F P? .H CI ., H F a@ nU d
CIV gW lQ eP wX sA kJ iY ,F
BZ .H H[ . H 1997. Regulation of las and rhl quorumsensing in Pseudomonas aeruginosa. J. Bacteriol.179:3127-3132.P? f_ eP nU d
CeP rE , F W
f.H Fg ., H F K^ rE a@ uS sA , F J
K., H F a@ nU d
CLG
oD ph eP rE , F JK.H Ec . H 1993. A
genomic region from Pseudomonas fluorescens Pf-5required for pyrrolnitrin production and inhibitionof Pyrenophora tritici-repentis in wheat straw.Phytopathol. 83:1123-1128.P? iY cb a@ rE dC , F CI ., H F D
ai Y CI eP llQ�Q oD , F Fg .H , F V
ieP nU tT uS rE a@ , F M
L., H F Fg a@ nU iY , F R
R., H F a@ nU d
CGM uS cb kJ eP rE tT , F A
\.H 2000. Frequency and diversity of 2-4-
diacetylphloroglucinol-producing bacteria isolatedfrom the maize rhizosphere at different stages ofgrowth. Appl. Environ. Microbiol. 66:948-955.P? iY eP rE sA oD n U IIIV$V%V , F L
G.H Sd ., H F a@ nU d
CP? iY eP rE sA oD nU , F Ec .H A\ .H 1996. Phenazine
antibiotic production in Pseudomonas aureofaciens:role in rhizosphere ecology and pathogensuppression. FEMS Microbiol. Lett. 136:101-108.P? iY eP rE sA oD n U IIIV$V%V , F L
G.H Sd ., H F GM a@ ff_j_ nU eP yk , F T
l., H F L
Ga@ mN , F Sd ., H F a@ nU d
CGM oD nU gW ,F
Fg .H 1995. Molecular analysis of genes encodingphenazine biosynthesis in the biocontrol bacteriumPseudomonas aureofaciens 30-84. FEMS Microbiol.Lett. 134:299-307.P? iY eP rE sA oD n U IIIV$V%V , F L
G.H Sd ., H F a@ nU d
CTl
hm oD mN a@ sA hm oD wX , F LG
.H Sd .H 1992.Cloning and heterologous expression of thephenazine biosynthetic locus from Pseudomonasaureofaciens 30-84. Mol. Plant-Microbe Interact.5:330-339.RR
aa@n@ iY jo mN a@ kJ eP rE sA , F JK.H ML .H , F W
feP llQ�Q eP rE , F D
a.H ML .H , F a@ nU d
CTl
hm oD mN a@ sA hm oD wX ,FLG
.H Sd .H 1997. Frequency of antibiotic producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63: 881-887.RR
aa@n@ iY jo mN a@ kJ eP rE sA , F JK.H ML .H , F a@ nU d
CWf
eP lQ lQ eP rE , F Da
.H ML .H 1998. Naturalplant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils.Mol. Plant-Microbe Interact. 11:144-152.RR
aa@n@ iY jo mN a@ kJ eP rE sA , F JK. H ML
.H , F BZ oD nU sA a@ lQ l Q RR
.H Fg .H , F a@ nU d C
Wf
eP lQ lQ eP rE , F Da
. H ML
.H1999. Effect of population density of Pseudomonasfluorescens on production of 2,4-diacetylphloroglucinol in the rhizosphere of wheat.Phytopathol. 89:470-475.RR
aa@n@ iY jo mN a@ kJ eP rE sA , F JK.H ML .H , F a@ nU d
CWf
eP lQ lQ eP rE , F Da
.H ML .H 2001.Exploiting genotypic diversity of 2,4-diacetylphloroglucinol-producing Pseudomonasspp.: characterization of superior root-colonizing P.fluorescens strain Q8r1-96. Appl. Environ.Microbiol. 67: 2545-2554.RR
a@ iY nU eP yk , F P? .H BZ ., H F BZ rE oD dC
eP yk , F CI .H LG ., H F a@ nU d C
JKoD hm nU sA tT oD nU eP , F K.H
1991. Biological properties and spectrum of activity
of tolaasin, a lipodepsipeptide toxin produced bythe mushroom pathogen Pseudomonas tolaasii.Physiol. Molec. Plant Pathol. 39:57-70.RR
a@ iY nU eP yk , F P? .H BZ ., H F BZ rE oD dC
eP yk , F CI .H LG ., H F a@ nU d C
JKoD hm nU sA tT oD nU eP , F K.H
1993. Identification of a gene cluster encodingthree high-molecular-weight proteins, which isrequired for synthesis of tolaasin by the mushroompathogen Pseudomonas tolaasii. Mol. Microbiol.8:643-652.RR
eP ddCpC
iY , F Tl
.H K^ ., H F a@ nU d C
BZ oD rE oD vq kJoD vq , F A
\.H Vi .H 1969. Mono-di
and tri-acetylphloroglucinols from Pseudomonasfluorescens. Khim. Prir. Soedin. (Tashk.) 2:133.RR
oD iY tT mN a@ nU , F JK.H Nr ., H F M
La@ hm oD nU eP yk , F N
r.H Ec ., H F a@ nU d
CJKa@ nU iY sA iY eP wX iY cb zs ,F
Wf
.H JK .H 1990. Production and composition ofphenylpyrrole metabolites produced byPseudomonas cepacia. Appl. Microbiol. Biotechnol.34:381-386.RR
oD nU , F Ec .H Zt ., H F a@ nU d C
RR
oD sA eP nU bO eP rE gW , F Ec .H 2001. Natural rolesof biosurfactants. Environ. Microbiol. 3:229-236.RR
oD sA a@ lQ eP sA , F A\
.H ML .H , F Tl
hm oD mN a@ sA hm oD wX , F LG
.H Sd ., H F CI ooD!D kJ, F RR
.H JK ., H F a@ nU dC
ML
eP wX , F Tl
.H Wf .H 1995. Isolation and identification ofantifungal metabolites produced by rice-associatedantagonistic Pseudomonas spp. Phytopathol.85:1028-1032.Sd a@ rE nU iY gW uS eP tT , F A
\., H F K^ rE a@ uS sA , F J
K., H F H
[eP nU k
JeP lQ sA , F M
L.H Da .H ,F
ML
uS eP hm lQ cb hm eP nU , F A\
.H ML ., H F a@ nU d C
LG
oD ph eP rE , F JK.H Ec .H 1995. The sigma
factor σS affects antibiotic production and biologicalcontrol activity of Pseudomonas fluorescens Pf-5.Proc. Natl. Acad. Sci. USA 92:12255-12259.Sd cb hm nU iY dC eP rE , F Uu ., H F K^ eePeP lQ , F CI ., H F BZ lQ uS mN eP rE , F CI ., H F T
lrE oD xv lQ eP rE , F J
K.,H F
Da
éwf_ a@ gW oD , F GM ., H F a@ nU d
CH[
aa@n@ sA , F Da
.H 1995(a). Amplification ofthe housekeeping sigma factor in Pseudomonasfluorescens CHAO enhances antibiotic productionand improves biocontrol abilities. J. Bacteriol.177:5387-5392.Sd cb hm nU iY dC eP rE , F Uu ., H F K^ eePeP lQ , F CI ., H F V
ioD iY sA a@ rE dC , F CI ., H F D
aéwf_ a@ gW oD , F GM .,H F
a@ nU d C
H[
aa@n@ sA , F Da
.H 1995(b). Tn5-directed cloning of pqqgenes from Pseudomonas fluorescens CHA0:mutational inactivaation of the genes results inoverproduction of the antibiotic pyoluteorin. Appl.Environ. Microbiol. 61:3856-3864.Sd cb hm nU iY dC eP rE -] KeePnP lQ , F Uu ., H F Sd eePnP mN a@ ttTxT eP rE , F A
\., H F M
La@ uS rE hm oD f_ eP rE , F M
L.,H F
BZ lQ uS mN mN eP rE , F CI ., H F Da
uS ff_j_ yk , F BZ .H K., H F GM iY gW oD tT -] BZ oD nnUyU eP f_ oD yk , F CI .,H FRR
eP iY mmN�N a@ nnUyU , F CI ., H F Nr
oD tT zs , F RR
., H F Da
éwf_ a@ gW oD , F GM ., H F H
[aa@z@ sA , F D
a.,H F
a@ nU d C
KeePnP lQ , F CI .H 2000. Autoinduction of 2,4-diacetylphloroglucinol biosynthesis in thebiocontrol agent Pseudmonas fluorescens CHA0 andrepression by the bacterial metabolites salicylate andpyoluteorin. J. Bacteriol. 182:1215-1225.Sd cb hm oD lQ zs -] Sd cb hm rE oD eP dC eP rE , F BZ ., H F Sd oD uS lQ eP , F J
K.H Da ., H F L
GuS , F Sd .H -] Ec .,H F
GM rE gW uS rE iY nU a@ , F IV ., H F a@ nU d C
GM rE oD ssA$A , F Da
.H 2001. A physical mapof the syringomycin and syringopeptin gene clusterslocalized to approximately 145-kb DNA region ofPseudomonas syringae pv. syringae strain B301D.Mol. Plant-Microbe Interact. 12:1426-1435.Sd eP rrE%E a@ , F M
L.H Da ., H F Fg a@ gW iY uS oD lQ iY , F GM ., H F N
roD rE dC eP rE a@ , F P? ., H F BZ eP rE nU hm a@ rE tT , F IV .,H F
Vi
oD lQ ph eP , F CI .H Da ., H F Da
i Y GM iY oD rE gW iY oD , F Da
., H F BZ a@ llQ�Q iY oD , F A\
., H F a@ nU dC
ML
eP nU eP sA tT rE iY nU a@ , F GM .H 1999. The interaction oflipodepsipeptide toxins from Pseudomonas syringaepv. syringae with biological and model membranes:a comparison of syringotoxin, syringomycin, and

C{ h|
a} pt~/� e� r � 1: �4� G� e� n� e� r� a} l �
I� n� t� r� o� d�
u� c� t� i� o� n � a} n� d �
O� u� t� l�i� n� e�
29
two syringopeptins. Mol. Plant-Microbe Interact.12:391-400.S� h� a� n� a� h� a� n� , � P� ., ��� O
�’�S� u� l� l� i� v� a� n� , � D
�.� J� ., ��� S� i
�m� p� s� o n� , � P� .,���
G¡ l� e¢ nn�y� o n� , � J�.� D� .� , � a� n� d £ O
�’�G¡ a� r¤ a� , � F¥ .� 1992. Isolation of
2,4-diacethilphloroglucinol from a fluorescentpseudomonad and investigation of physiologicalparameters influencing its production. Appl.Environ. Microbiol. 58:353-358.S� h� a� n� a� h� a� n� , � P� ., ��� G¡ l� e¢ n� n� o n� , � J
�.� D� .� , � C¦ r¤ o w§ l� e¢ y, � J
�.� J� .� ,�
D�
o nn�y� e¢ ll��� y¨ , � D�
.� F¥ ., ��� a� n� d £ O�
’�G¡ a� r¤ a� , � F¥ .� 1993. Liquid
chromatographic assay of microbially derivedphloroglucinol antibiotics for establishing thebiosynthetic route to production, and the factorsaffecting their regulation. Anal. Chim. Acta272:271-277.S� h� a� r¤ i� f© i� -ª T« e¢ h� r¤ a� n� i
�, � A¬
.� , � Z
a� l� a� , � M® ., ��� Na� t° s� c± h� , � A¬
.� , � M® o ë²nn�y� e¢ -ª
L³ o c± c± o z´ , � Yµ
., ��� a� n� d £ D�
é¶ f© a� g· o , � G¡ .� 1998. Biocontrol ofsoil-borne fungal diseases by 2,4-diacetylphloroglucinol-producing pseudomonadswith different restriction profiles of amplified 16SrDNA. Europ. J. Plant Patol. 104:631-643.S� o l� e¢ r¤ -ª Ri
�v� a� s� , � C¦ ., ��� A
¬r¤ p� i
�n� , � C¦ .� , � O
�l� i� v� i
�e¢ r¤ , � J
�.� M® .� , � a� n� d£
W¹
i�c± h� e¢ r¤ s� , � H
º.� J� .� 1999. WLIP, a lipodepsipeptide of
Pseudomonas reactans, as inhibitor of the symptomsof the brown blotch disease of Agaricus bisporus. J.Appl. Microbiol. 86:635-641.S� ø» r¤ e¢ n� s� e¢ n� , � D
�., ��� Ni
�e¢ l� s� e¢ n� , � T« .� Hº ., ��� C¦ h� r¤ i� s� t° o p� h� e¢ r¤ s� e¢ n� , � C¦ .,���
S� ø» r¤ e¢ n� s� e¢ n� , � J�.� , � a� n� d £ G¡ a� j¼ h� e¢ d£ e¢ , � M® .� 2001. Cyclic
lipoundecapeptide from Pseudomonas sp. strainDSS73. Acta Cryst. 57c:1123-1124.S� t° a� bb½p½ , � E¾ .� V¿ ., ��� J
�a� c± o b½ s� o n� , � L³ .� M® ., ��� a� n� d £ H
ºa� n� d£ e¢ l� s� m� a� n� , � J
�.�
1994. Zwittermycin A-producing strains of Bacilluscereus from diverse soils. Appl. Environ. Microbiol.60:4404-4412.S� t° a� n� g· h� e¢ ll��� i� n� i
�, � M® .� E¾ ., ��� a� n� d £ M® i
�l� l� e¢ r¤ , � R¸ .� M® .� 1997.
Biosurfactants: their identity and potential efficacyin the biological control of zoosporic plantpathogens. Plant Disease 81: 4-12.S� t° r¤ u� n� z, � G¡ .� M® ., ��� W
¹a� ll��� , � R.� E¾ ., ��� a� n� d £ H
ºo l� d£ e¢ r¤ -ª F¥ r¤ a� n� kÀ l� i� n� ,�
M® .� A¬ .� 1978. Phloroglucinol derivatives fromAeromonas hydrophila. J. Antibiot. 31:1201-1202.S� t° u� t° z, � E¾ ., ��� D
�é¶ f© a� g· o , � G¡ ., ��� a� n� d £ KÁ e¢ r¤ n� , � H
º.� 1986.
Naturally occuring fluorescent pseudomonadinvolved in suppression of black root rot of tobacco.Phytopathol. 76:181-185.S� y¨ l� d£ a� t° kÀ , � C¦ ., ��� L³ a� n� g· , � S� .� , � a� n� d £ W
¹a� g· n� e¢ r¤ , � F¥ .� 1985.
Chemical and physical characterization of fourinterfacial-active rhamnolipds from Pseudomonas sp.DSM2874 grown on n-alkanes. Zeitsc. Naturforch.40c:51-60.T« a� kÀ e¢ d£ a� , � R¸ .� 1958. Pseudomonas pigments. I.Pyoluteorin, a new chlorine-containing pigmentproduced by Pseudomonas aeruginosa. Hako KogakuZasshi 36:281-290.T« a� w§ a� r¤ a� , � S� ., ��� M® a� t° s� u� m� o t° o , � S� ., ��� H
ºi�r¤ o s� e¢ , � T« ., ��� M® a� t° s� u� m� o t° o ,�
Yµ
., ��� Na� kÀ a� m� o t° o , � S� ., ��� M® i�t° s� u� n� o , � M® ., ��� a� n� d £ KÁ a� m� i
�m� u� r¤ a� , � T« .�
1989. In vitro antifungal synergism betweenpyrrolnitrin and clotrimazole. J. Med. Mycol.30:202-210.T« h� o m� a� s� h� o w§ , � L³ .� S� ., ��� a� n� d £ W
¹e¢ ll��� e¢ r¤ , � D
�.� M® .� 1996. Current
concepts in the use of introduced bacteria forbiological disease control: mechanisms and
antifungal metabolites. In: Stacey, G., and Keen,N.T. (eds.). Plant-Microbe Interactions, pages 187-236, Vol. 1, Chapman & Hall.T« h� o m� a� s� h� o w§ , � L³ .� S� ., ��� a� n� d £ W
¹e¢ ll��� e¢ r¤ , � D
�.� M® .� 1988. Role of
phenazine antibiotic from Pseudomonas fluorescensin biological control of Gaeumannomyces graminisvar. tritici. J. Bacteriol. 170:3499-3508.T« h� o m� a� s� h� o w§ , � L³ .� S� ., ��� W
¹e¢ ll��� e¢ r¤ , � D
�.� M® ., ��� B
Âo n� s� a� l� l� , � R.� F¥ .,���
a� n� d £ P� i�e¢ r¤ s� o n � III
Ã%Ã%Ã, � L³ .� S� .� 1990. Production of the
antibiotic phenazine-1-carboxylic acid byfluorescent Pseudomonas species in the rhizosphereof wheat. Appl. Environ. Microbiol. 56:908-912.T« h� o m� a� s� h� o w§ , � L³ .� S� ., ��� B
Âo n� s� a� ll��� , � R.� F¥ ., ��� a� n� d £ W
¹e¢ ll��� e¢ r¤ , � D
�.� M® .�
1997. Antibiotic production by soil and rhizospheremicrobes in situ. In: Hurst, C.J., Knudsen, G.R.,McInerney, M.J., Stetzenbach, L.D., and Walter,M.V. (eds.). Manual of EnvironmentalMicrobiology, pages 493-499 ASM Press,Washington, D.C., USA.T« h� r¤ a� n� e¢ , � C¦ .� , � O
�l� ss�B� o n� , � S� ., ��� N¯ i
�e¢ l� s� e¢ n� , � T« .� Hº ., ��� a� n� d£
S� ø» r¤ e¢ n� s� e¢ n� , � J�.� 1999. Vital fluorescent stains for
detection of stress in Pythium ultimum andRhizoctonia solani challenged with viscosinamidefrom Pseudomonas fluorescens DR54. FEMSMicrobiol. Ecol. 30:11-23.T« oo ! h� e¢ y, � J
�.� Ià ., ��� N¯ e¢ l� s� o n� , � C¦ .� D� ., ��� a� n� d £ KÁ r¤ o t° kÀ o v� , � G¡ . � 1965.
Toxicity of phenazine carboxylic acid to somebacteria, algae, higher plants and animals. Can. J.Bot. 43:1151-1155.T« r¤ i� p� a� t° h� i
�, � R.� KÁ ., ��� a� n� d £ G¡ o t° t° l� i� e¢ b½ , � D
�.� 1964. Mechanism
of action of the antifungal antibiotic pyrrolnitrin. J.Bacteriol. 100:310-318.T« u� r¤ n� e¢ r¤ , � J
�.� M® ., ��� a� n� d £ M® e¢ ss�B� e¢ n� g· e¢ r¤ , � A
¬.� J� .� 1986.
Occurrence, biochemistry and physiology ofphenazine pigment production. Adv. Microb.Physiol. 27:211-275.UÄ
i�, � H
º., ��� M® i
�ya� kÀ e¢ , � T« ., ��� I
Ãi�n� u� m� a� , � H
º., ��� I
Ãm� o t° o , � M® .,���
N¯ a� g· a� n� a� w§ a� , � Hº
.� , � Hº
a� tt°x° o r¤ i� , � S� ., ��� Hº
a� m� a� d£ a� , � M® ., ��� T« a� kÀ e¢ u� c± h� i�,�
T« ., ��� UÄ
m� e¢ za� w§ a� , � S� ., ��� a� n� d £ UÄ
m� e¢ za� w§ a� , � KÁ .� 1997.Pholipeptin, a novel cyclic lipoundecapeptide fromPseudomonas fluorescens. J. Org. Chem. 62:103-108.V¿
a� n � D�
ykÀ e¢ , � M® . � IÃ., ��� C¦ o u� t° u� r¤ e¢ , � P� ., ��� B
Âr¤ a� u� e¢ r¤ , � M® ., ��� L³ ee¢n¢ , � H
º.,���
a� n� d £ T« r¤ e¢ v� o r¤ s� , � J�.� T« .� 1993. Pseudomonas aeruginosa
UG3 rhamnolipid biosurfactants: structuralcharacterization and their use in removinghydrophobic compounds from soil. Can. J.Microbiol. 39:1071-1077.V¿
a� n� i�tt°x° a� n� a� kÀ o m� , � N., ��� a� n� d £ L³ o e¢ ff©j© l� e¢ r¤ , � W
¹.� 1986.
Fengycin - a novel antifungal lipopeptide antibioticproduced by Bacillus subtilis F-29-3. J. Antibiot.39:888-901.V¿
e¢ rr¤%¤ o t° a� , � L³ ., ��� A¬
pp�Å� e¢ n� d£ i�n� o , � G¡ ., ��� B
Âe¢ ll��� o r¤ o , � E¾ ., ��� J
�a� kÀ u� p� o v� i
�c± ,�
J�., ��� a� n� d £ B
Âo m� b½ a� r¤ d£ e¢ ll��� i� , � E¾ .� 1999. Furohyperforin, a
prenylated phloroglucinol from St. John’s Wort(Hypericum perforatum). J. Nat. Prod. 62:770-772.V¿
i�n� c± e¢ n� t° , � M® .� N., ��� H
ºa� rr¤%¤ i� s� o n� , � L³ .� A¬ ., ��� B
Âr¤ a� c± kÀ i
�n� , � J
�.� M® .� ,�
KÁ o v� a� c± e¢ v� i�c± h� , � P� .� A¬ ., ��� M® u� kÀ e¢ r¤ j¼ i� , � P� ., ��� W
¹e¢ l� l� e¢ r¤ , � D
�.� M® .,���
a� n� d£ P� i�e¢ r¤ s� o n� , � E¾ .� A¬ .� 1991. Genetic analysis of the
antifungal activity of a soilborne Pseudomonasaureofacens strain. Appl. Environ. Microbiol.57:2928-2934.

CÆ hÇ
aÈ ptÉ/Ê eË r Ì 1: Í4Î GÏ eË nÐ eË rÌ aÈ l Ñ
IÒ nÐ tÊ rÌ oÓ dÔ
uÕ cÖ tÊ i× oÓ n Ð aÈ nÐ d Ô
OØ uÕ tÊ lÑi× nÐ eË
30
WÙ
hÚ iÛppÜpÜ sÝ , Þ J
ß.à Má .à 2001. Microbial interactions and
biocontrol in the rhizosphere. J. Exp. Bot. 52: 487-511.WÙ
hÚ iÛsÝ tâ lã eä rå , Þ C
æ.à Aç ., à Þ C
æoè rå bé eä llã�ã , Þ Nê .à Aç .à , Þ Së aì rå ní i
Ûgî uï eä tâ , Þ A
ç., à Þ Rð eä aì mñ ,Þ
WÙ
., à Þ aì ní d ò Ló oè pÜ eä rå , Þ Jß.à Eô .à 1998. The two-component
regulators gacS and gacA influence accumulation ofthe stationary-phase sigma factor σS and the stressresponse in Pseudomonas fluorescens Pf-5. J. Bact.180:6635-6641.WÙ
hÚ iÛtâ eä lã eä yõ , Þ M
á., à Þ P
öaì rå sÝ eä k÷ , Þ M
á.à Rð ., à Þ aì ní d ò Gø rå eä eä ní bé eä rå gî , Þ Eô .à Pö .à
2000. Regulation of quorum sensing by RpoS inPseudomonas aeruginosa. J. Bacteriol. 182:4356-4360.WÙ
iÛllã�ã iÛ sÝ , Þ D
ù.à Kú ., à Þ Rð i
Ûcû hÚ , Þ J
ß.à Jß ., à Þ Kú i
Ûní sÝ cû hÚ eä rå fü , Þ T
ý.à Gø ., à Þ aì ní dò
Kú iÛtâ tâ eä ní , Þ T
ý.à 1994. Genetic regulation in plant
pathogenic pseudomonads. In: Suslow, J.T. (ed.).Genetic Engineering, pages 167-193, Vol. 15,Plenum Press, New York, USA.
WÙ
iÛlã sÝ oè ní , Þ Rð .à Tý ., à Þ P
öiÛttâxâ , Þ T
ý., à Þ T
ýaì yõ lã oè rå , Þ Gø ., à Þ W
Ùaì tâ sÝ oè ní , Þ D
ù.,à Þ
Má
aì cû Dù eä rå mñ oè tâ , Þ Jß., à Þ Së yõ k÷ eä sÝ , Þ D
ù., à Þ Rð oè bé eä rå tâ sÝ , Þ D
ù., à Þ aì ní d ò C
æoè lã eä ,Þ
Pö
.à 1987. Pyocyanin and 1-hydroxyphenazineproduced by Pseudomonas aeruginosa inhibit thebeating of human respiratory cilia in vitro. J. Clin.Invest. 79:221-229.Yþ
oè ní eä yõ aì mñ aì , Þ Kú ., à Þ Kú oè nníyí aì iÛ , Þ Má
., à Þ Hÿ oè ní dò aì , Þ I�., à Þ Yþ
oè sÝ hÚ iÛdò aì , Þ Së .,à Þ
Tý
aì k÷ aì hÚ aì sÝ hÚ iÛ, Þ Nê ., à Þ Kú oè i
Ûk÷ eä , Þ Hÿ ., à Þ aì ní d ò I
�ní oè uï eä , Þ Y
þ.à 1990.
Phloroglucinol derivatives as potent photosysteminhibitors. Zeitsc. Naturfors., Section C, Biosc.45:317-321.Yþ
uï aì ní , Þ Z�
., à Þ Cæ
aì ní gî , Þ Së ., à Þ Má
aì tâ sÝ uï füuï j�iÛ, Þ Má
., à Þ Nê aì k÷ aì tâ aì , Þ Kú .,à ÞNê aì gî aì mñ aì tâ sÝ uï , Þ Y
þ., à Þ aì ní d ò Y
þoè sÝ hÚ i
Ûmñ oè tâ oè , Þ A
ç.à 1998. High
production of pyoluteorin and 2,4-diacetylphloroglucinol by Pseudomonas fluorescensS272 grown on ethanol as a sole carbon source. J.Ferment. Bioengin. 86:559-563.

______________________________________________________
CHAPTER 2
P�
O�
L�
Y�
M�
O�
R�
P�
H
IS�M�
S �
W�
IT
H
IN �
T
H
E �
p� r� n� D�
A�
N�
D �
p� l�t� C� G
�E�
N�
E�
S �
F�
R�
O�
M
P!
Y"
RR�#�
O�
L$
N�
I%T&
R�
I%N �
A�
N�
D �
P!
Y"
O�
L$
U'
T&
E�
O�
R�
I%N�
-( P! R�
O�
D�
U'
C)
I%N�
G �
P*
S+E,
U- D�
O. M/
O. N0
A1
S+
A�
N�
D �
B2
U- R3
K4
H5
O. L6
D�
E,
R3
I7A 1
S�PP!8!
.9
_______________________________________________________________________________

A:
B; S< T=
R> A:
C? T=
_______________________________________________________________________________
P@ yA rrBCB oD lEnF iG tH rB iG n F (I P@ R> N
J) K
aL nF d M
pN yA oD lEuO tH eP oD rB iG n F (I P@ LQ T)
= KaL rB e P b
RrB oD aL dM -S sT pN eP cU tH ruBCO m V aL nF tiH G bR iG oD tiH G cU s T pN rB oD d
MuO cU eP dM
bR
y A sT eP vW eP rB aL l E sT tH rB aL iG nF s T oD f X
PsY[Z
eu\^] d_
oma onb ac sZ aL nF d
MBd
u] re kf hg ol`ih d_ e\ re ij ac sT pN eP cU iG eP sT , k aL nF d M
pN lEaL y A aL n F iG mV pN oD rB tH aL nF t H rB oD l
Ee P iG n F tH hl eP
sT uO ppNmN rB eP ssTCT iG oD n F oD f X
mV uO ltE H iG plN E e P plN E aL nF t pH N aL tH hl oD gn eP nF iG c fU X
uO nF gn iG . o P@ rB imG V eP rB s T wp eP rB e P dM
eP vW eP lE oD pN eP d M
fXrB oD m V cU oD nF sT eP rvB W eP dM
nF uO cU lE eP oD tiH G dM e P sT eP qq uO eP nF cU eP s T aL nF d M
wp eP rB e P aL bR lEe P tH o D aL mV pliN E G fX y A pr rnesb D
taL nF d M
pr lh tu C v fXrB aL gn mV eP nF tH s T f
XrB oD m V eP iG gn h
ltH eP eP nF
PsY[Z
eu\^] d_
oma onb ac sZ aL nF d
MfXoD ur OwB B
du] re kf hg ol`ih d_ e\ re ij a c sT ppNwN . o oD f
Xwp oD rB lE dM wp iG dM e P oD rB iG gn iG n F tH hl aL t H eP iG tH hl eP r B pN rB oD d
MuO cU e P P@ R> N
JoD r B P@ LQ T
=oD r B bR
oD tH hl . o S< uO bR
sT eP qq uO eP nF t H R> Fx LQ P @ (I R> eP sT tH rB iG cU tH iG oD n F Fx rB aL gn mV eP nF t H LQ eP nF gn tH h l
P@ oD lEyA mV oD rB pN h
liG sT mV sT ) K aL nF aL lE yA sT iG s T oD f
XtH hl e P 438-
y{z}| SbR
p N pr lh tu C v fXrB aL gn mV eP nF t H sT hl oD wp eP d
MnF o D pN oD l
EyA mV oD rB pN h
liG sT mV s T aL mV oD nF g n P@ LQ T-
= S pN rB oD dM
uO cU iG nF g n PsY[Z
e\ u] d_oma onb ac s
ZsT tH rB aL iG nF sT .o
P@ oD lEyA mV oD rB pN h
liG sT mV s T wp itG H hl iG n F tH hl e P 786-
~ |}� S bR p N pr rnesb Dt
fXrB aL gn mV eP nF tH , k h
loD wp eP vW eP rB , k aL lE lE oD wp eP d
MtH hl e P aL ssTCT eP ssTCT mV eP nF t H oD f
XtH hl eP
dM
iG vW eP rB sT itG H y A aL mV oD nF g n P@ R> NJ
-S pN rB oD dM
uO cU iG nF g n PsY[Z
eu\^] d_
oma onb ac sZ aL nF d
MBd
u] re kf hg ol`ih d_ e\ re ij ac sT ppNwN . o tH o D a L lEeP vW eP l E sT iG milV G E aL r B tH o D tH hl aL tH
oD bR
tH aL iG nF eP d M
bR
y A tH hl rB ee P�P 10-��� S mV eP r B pN rB iG mV eP rB s T iG n RF�> A:
P@ D (��I R> aL nF dM
oD m V A:
mpV N lEiG fX iG eP d
MP@ oD lEyA mV oD rB pN h
liG c DU � NA
J :) K
aL nF aL lE yA sT iG sT .oP@ hl
yA lEoD gn eP nF eP tH iG c U aL nF aL lE yA sT iG s T oD f 16
X � � S < rB D� NA sJ : T eP qq uO eP nF cU eP s T oD f
Xa L nF uO mV b
ReP r B oD f
XsT tH rB aL iG nF s T rB eP pN rB eP sT eP nF tH aL tiH G vW e oP�D f
XP@ R> N
J-S
pN rB oD dM
uO cU iG nF g n PsY[Z
eu\^] d_
oma onb ac sZ aL nF d
MBd
u] re kf hg olh d_ e\ re ij ac sT pN eP cU iG eP s T cU oD rrB�B eP lE aL tH eP d M
wp eP ll E�E wp iG tH h l
tH hl eP iG r B tH aL x� oD nF oD miV G c U sT tH aL tH uO sT .oP@ hl
yA lEoD gn eP nF eP tH iG c U rB eP lE aL tH iG oD nF sT hl ipG N s T iG nF f
XeP rrB�B eP d
MfXrB oD m V eP aL cU h
loD f X
tH hl e P fXoD ur OwB pr rnesb gn eP nF eP s T aL s T wp eP ll E�E aL s T f
XrB oD m tV H hl eP
cU oD mpV N lEeP tH e P sT eP qq uO eP nF cU e oP�D f
XtH hl e P pr rne�b b
RiG oD sT yA nF tH hl eP tiH G c U l
EoD cU uO s T wp eP rB e P sT iG mV ilG E aL r B tH o D 16� � S < rB D� NA
J :bR
aL sT eP d M
pN hl
yA lEoD gn eP nF yA
fXoD r B mV oD sT t H sT tH rB aL iG nF s T eP x� cU eP pt N H f
XoD r B B
d. � pr y� rrese oci��j nb ij ac D� S< M
�10685. ��� �}|}� o B; oD tH h R
l > Fx LQ P @ aL nF aL lE yA sT iG s T aL nF d M
cU oD mpV N aL rB iG sT oD n F oD fX
tH hl e P pr rnesb gen�P nF e P sT eP qq uO eP nF cU eP s sT#T hl oD wp eP d M
tH hl aL t H Bd
. � pr y� rrese oci��j nb ij ac D� S< M�
10685 w��� �}|}� p aL s T mV oD rB e P cU lE oD sT eP lE y A rB eP lE aL tH eP d M
tH oDP@ R> N
J-S pN rB oD d
MuO cU iG nF g n Ps
Y[Zeu\^] d_
oma onb ac sZ sT tH rB aL iG nF sT , k sT uO ggen�n�P sT tiH G nF g n tH hl aL t lH E aL tH eP rB aL l E gn eP nF e P tH rB aL nF sT fX eP r B mV aL y A h
laL vW e oP�D ccU�U urrOmBCB eP dM .o
Co? D lEoD nF y A h
lyA bR
rB iG dM iG z� aL tiH G oD n F aL nF d PM @ C? R > wp iG tH h
lP@ R> N
J- S aL nF d P
M @ LQ T-= S sT pN eP cU iG fX iG c U pN rB oD b
ReP s T aL nF d
MpN rB iG mV eP rB s sT�T hl oD wp eP d
MtH hl aL tH
PsY[Z
eu\^] d_
oma onb ac sZ aL nF d
MBd
u] re kf hg ol`�h d_ e\ re ij a c sT ppNwN . o hl
aL rB bR oD rB iG nF g n tH hl e P pr rnesb Dt
aL nF d M
pr lh tu Cv gen�P nF e P wp eP rB e P nF oD t pH N rB eP sT eP nF t H aL tHdM
eP tH eP cU tH aL bR lEe P lEeP vW eP lE s T oD n F rB ooD�D tH s T oD f w
X p hl
eP aL t H gn rB oD wp n F iG n F fXiG vW e P aL gn rB iG cU uO lt
E H urOwB aL l E sT oD ilG E s T cU oD llE�E
eP cU tH eP d M
iG n F tH hl e P NJ
eP tH hl eP rB lE aL nF dM
sT ,ktH wp o oD�D f
XtH hl eP m V b
ReP iG nF g n nF aL tH urOwB aL llE�E y A sT uO ppNwN rB eP ssTCT iG vW e P tH o D G� ac e\ u] ma ac nnb}b oma y� c� e\ s g
Z��re ac mma�a ij nb ij sZ vW aL rB . o tu re ij tu icij��sj . To = h
leP sT eP
rB eP sT uO ltE H s sT�T uO ggen�n�P sT t tH�H hl aL t H P@ R> N
J- S aL nF d P
M @ LQ T-= S pN rB oD d
MuO cU iG nF g n Ps
Y[Ze\ u] d_
oma onb ac sZ aL nF d
MBd
u] re kf hg ol`�h d_ e\ re ij ac sT pN . o dM
o D nF oD tHcU oD nF tH rB iG bR uO tH e P tH o D tH hl e P nF aL tH urOmB aL l E sT uO ppNwN rB eP ssT�T iG vW eP nF eP ss T�T f
XoD uO nF d
MiG n F D� uO tH cU h
ltH aL k� eP -S aL ll E�E d
MeP cU lE iG nF e P sT oD ilG E sT .o
_______________________________________________________________________________
J�oD rB ge n�P T.
= o dM
e P S< oD uO z� a aL L nF d M
J�oD s T M�
. o R> aaL�L iG j¡ mV aL k� eP rB sT
FEMS Microbiology Ecology. accepted.
32
C? hl
aL ptN H eP r B 2: ¢{£
P@ oD lEyA mV oD rB pN h
liG sT mV s T wp iG tH hl iG n F tH hl e P pr rnesb D
t aL nF d
Mpr lh tu Cv gen¤P nF eP sT

I¥ N¦ T§
RO¨ © Dª U«
C¬ T§
I¥ O© N¦
Biological control of soil-borne diseases by plant growth-promoting rhizobacteria
(PGPR) is a well-established phenomenon and antibiotics have been shown to play a major
role in suppression of several plant pathogens (Handelsman and Stabb, 1996). Numerous
bacterial strains that produce antibiotics in vitro have been isolated from different soils and
host plants (O'Sullivan and O'Gara, 1992; Thomashow and Weller, 1996). This apparent
wide distribution suggests that antibiotic-producing bacteria are common constituents of the
indigenous microflora in soil and plant-associated environments worldwide. To date, however,
little is known about the frequency and ecology of indigenous antibiotic-producing bacteria.
Knowledge about the ecology of naturally occurring strains that harbor specific biocontrol
traits will significantly contribute to improve the efficacy of existing biocontrol agents.
Moreover, knowledge about the distribution of antibiotic genes in natural environments could
lessen concerns about the environmental release of either non-indigenous strains containing
these traits or transgenic biocontrol agents in which these traits have been introduced.
Selection and identification of antibiotic-producing bacteria via random isolation from
natural environments is very time-consuming and laborious. Most of the model strains used to
date were identified via screening huge numbers of isolates obtained by random procedures.
The availability of cloned and sequenced antibiotic biosynthetic and regulatory genes,
however, has facilitated the development of specific primers and probes that can be used for
targeted detection and isolation of specific antibiotic-producing bacteria. For several
antibiotics produced by strains of different bacterial genera, biosynthetic genes have been
cloned and partially or fully sequenced. These include zwittermycin A produced by B. cereus
(Stohl et al., 1999), AFC-BC11 produced by B. cepacia (Kang et al., 1998) and the antibiotics
2,4-diacetylphloroglucinol (2,4-DAPG), phenazines (PHZ), pyrrolnitrin (PRN), and
pyoluteorin (PLT) produced by various Pseudomonas species (Nowak-Thompson et al., 1999;
Hammer et al., 1997; Mavrodi et al., 1998; Pierson et al., 1995; Bangera and Thomashow,
1999; Chin-A-Woeng, 2000). A prerequisite for DNA-based detection and isolation is that
the genes of interest must be conserved among a wide range of bacterial strains harboring this
trait. By Southern hybridization (Keel et al., 1996) and PCR analysis (Raaijmakers et al.,
1997; Ramette et al., 2001; Mavrodi et al., 2001), it was shown that the biosynthetic locus for
2,4-DAPG production, and in particular the phlD gene, was highly conserved among 2,4-
DAPG-producing Pseudomonas strains of worldwide origin. Also some of the genes involved in
the biosynthesis of PHZ appear to be conserved among different strains (Pierson et al., 1995;
Raaijmakers et al., 1997; Mavrodi et al., 2001).
33
C¬ h
a® pt¯w° e± r ² 2: ³{´
Pµ o¶ l·y¸ m¹ o¶ r² p¯ h
iº s» m¹ s » w¼ iº t° h iº n ½ t° h e ± p¾ rn¿sÀ D
Á a® n½ d
Âp¾ là tÄ CÅ geƤ± n½ e± s»

Due to their broad-spectrum activity, PRN and PLT are among the antibiotics that
have received considerable attention (Bender et al., 1999; Hammer et al., 1999; Ligon et al.,
2000; Thomashow and Weller, 1996; Whipps, 1997). PRN is a chlorinated phenylpyrrole
antibiotic that was first isolated from Burkholderia pyrrocinia (Arima et al., 1964) and later
from other microorganisms, including Pseudomonas fluorescens, P. chlororaphis, P. aureofaciens,
B. cepacia, Enterobacter agglomerans, Myxococcus fulvus, and Serratia species (Elander et al.,
1968; Hammer et al., 1999; Kalbe et al., 1996). PRN has activity against several bacteria and
soil-borne fungi, in particular Rhizoctonia solani (Arima et al., 1964; Cartwright et al., 1995;
Chernin et al., 1996; El-Banna and Winkelmann, 1988; Hill et al., 1994; Jayaswal et al.,
1993; Rosales et al., 1995). PRN is also effective against post-harvest diseases caused by
Botrytis cinerea on apple, pear and on cut flowers (Hammer and Evensen, 1993; Janisiewicz
and Roitman, 1988), and has been used to treat humans infected by opportunistic fungi
(Tawara et al., 1989). PLT is a phenolic polyketide that was first isolated from P. aeruginosa
(Takeda, 1958) and later from P. aeruginosa strain S10B2 (Ohmori et al., 1978) and P.
fluorescens strains Pf-5 and CHA0 (Bencini et al., 1983; Bender et al., 1999; Keel et al., 1996).
PLT has bactericidal, herbicidal, fungicidal, and oomycidal activities, in particular against
Pythium spp. (Maurhoffer et al., 1992; Ohmori et al., 1978; Takeda, 1958). Application of
pure PLT to cotton seeds resulted in significant suppression of Pythium ultimum-induced
damping-off (Howell and Stipanovic, 1980).
Although the genes involved in the biosynthesis of PRN and PLT have been cloned
and sequenced (Hammer et al., 1999; Nowak-Thompson et al., 1997, 1999), little
information is available on the level of conservation of these genes in multiple PRN and PLT-
producing Pseudomonas and Burkholderia species and strains. In the study by Hammer et al.
(1999), sequencing of the PRN biosynthetic locus included only 4 strains, one P. fluorescens,
two Burkholderia strains and one strain of Myxococcus fulvus. For PLT, only data for P.
fluorescens strain Pf-5 are available in the database. In this study, we developed primers and
probes from specific sequences within prnD and pltC, two genes encoding essential enzymes
for the biosynthesis of PRN and PLT, respectively (Hammer et al., 1999; Nowak-Thompson
et al., 1997, 1999). The specificity of these primers was tested with eighteen Pseudomonas and
four Burkholderia strains of worldwide origin.
34
CÇ hÈ
aÉ ptÊwË eÌ r Í 2: Î{Ï
PÐ oÑ lÒyÓ mÔ oÑ rÍ pÊ h
ÈiÕ sÖ mÔ s Ö w× iÕ tË hÈ iÕ n Ø tË hÈ e Ì pÙ rnÚsÛ D
Ü aÉ nØ d
ÝpÙ lÞ tß Cà geá¤Ì nØ eÌ sÖ

T
â
a ãbälåe æ 1.
çéè Cêhëa ãr ìa ãcít îeær ìiïsðtîiïc ís ð oñf òP ós ôe õu öd ÷oømùoønúa ûs ô a ünýd
þ
B
ÿ
u ör �k �h �oøl �d ÷e õr �i �a û s�t�r �a üi�nýs � us�ed
þ
t �o � t �es�t t� �h�e s�p ec�i�f �i�c�i�t�y � o�f �P�R �N
�
a ünýd
þ
P
�
L
�
T
�
p r�i�m�er �s � a ünýd
þ
d
þ
i
�
v �er �s�i�t�y � s�t �ud þi�es�.�Strain a Other designation (s) a Origin/host plant PRN pro-
duction bPLT pro-duction b
Source and/or reference
B. cepaciaJ2535 LMG 16671 rotting bark, Edinburgh, Scotland + − Vandamme et al., 1997LMG1222 Palleroni 717; NCTC 10743 Allium cepa + − Vandamme et al., 1997ATCC 51671 B37w; NRRL B-14858 Sesbania exaltata leaves + − Burkhead et al., 1994J2540 LMG 16672 rhizosphere of a banana plant − − Vandamme et al., 1997
B. pyrrociniaDSM10685 ATCC 15958; LMG 14191 soil + − Vandamme et al., 1997; DSMZ
P. chlororaphisLMD 82.53 ATCC 43051; Strain ACN mutant of ATCC 15926 + − Salcher and Lingens, 1980
P. fluorescensPf-5 cotton, Texas, USA + + Howell and Stipanovic, 1979CHA0 tobacco, Morens, Switzerland + + Keel et al., 1992Pf1 tobacco, Morens, Switzerland + + Keel et al., 1996F113 sugar beet, Cork, Ireland − − Shanahan et al., 1992Q8r1-96 wheat, Washington, USA − − Raaijmakers and Weller, 2001PF wheat leaves, Oaklahoma, USA + + Levy et al., 1992PFM2 derivative of PF + + Levy et al., 1992PGNL1 tobacco, Ghana + + Keel et al., 1996PGNR1 tobacco, Ghana + + Keel et al., 1996PGNR2 tobacco, Ghana + + Keel et al., 1996PGNR3 tobacco, Ghana + + Keel et al., 1996PGNR4 tobacco, Ghana + + Keel et al., 1996PILH1 tomato, Italy − − Keel et al., 1996PINR2 tobacco, Italy + + Keel et al., 1996PINR3 tobacco, Italy + + Keel et al., 1996
C�h �a �pt ���e �r 2: !#"P$ o %l &y 'm (
o %r p �h �i) s *m (s *w +
i) t �h �i) n , t �h �e �p -rn.0/ D 1 a �n, d 2
p -l3 t4 C5 ge 67�n, e �s *

T
8
a 9b:l;e < 1= . > C?o@nAtBiCnAuDe<dE.>
Pseudomonas sp.PHZ13 tomato, Dijon, France + − This studyPHZ19 tomato, Dijon, France + − This studyPHZ22 tomato, Dijon, France + − This studyPHZ24 tomato, Dijon, France + − This studyPHZ48 tomato, Dijon, France + − This study
a ATCC, American Type Culture Collection, University Blvd, Manassas, Virginia, USA; DSMZ, Deutsche Sammlung von Mikroorganismen und Zelculturen GmbH,Braunschweig, Germany; LMD, Laboratorium voor Microbiologie, Technische Universiteit, Delft, The Netherlands; LMG, Laboratorium Microbiologie Gent CultureCollection, Universiteit Gent, Ghent, Belgium; NCPPB, National Collection of Plant Pathogenic Bacteria, Harpenden Laboratory, Hertfordshire, United Kingdom; NCTC,National Collection of Type Cultures, London, United Kingdom; NRRL, Northern Regional Research Laboratories, Peoria, Illinois, USA.
b Determined in this study by HPLC and/or TLC analysis.
CFh Ga Hpt I�Je Kr L 2: M#NPO o Pl Qy Rm S
o PrL p Ih GiT s Um Ss Uw V
iT t Jh GiT n W t Jh Ge Kp XrnY0Z D [ a HnW d \
p Xl] tC_ ge 7KnW e Ks U

Ca hb
ac ptd�e ef r g 2: h#i
Pj ok llym mn ok rg pd h
bio sp mn s p wq io te hb io n r te hb e f ps rnt0u D
v aw nx d
yps lz t{ C | g} e~ nx e~ s�
37
Restriction fragment length polymorphisms (RFLP) among amplified PRN and PLT-
fragments were evaluated to determine the diversity among different bacterial strains, species
and genera. Random amplified polymorphic DNA (RAPD) analysis, 16S rDNA sequencing
and in silico analysis of gene sequences involved in PRN biosynthesis were performed to study
the diversity and phylogenetic relationships among PRN- producing bacteria. Finally, we
determined the frequency of PRN- and PLT-producing Pseudomonas and Burkholderia spp. on
roots of wheat grown in five agricultural soils collected in the Netherlands, including two soils
that are naturally suppressive to Gaeumannomyces graminis var. tritici (Ggt). Determination of
population densities of Pseudomonas spp. harboring genes for 2,4-DAPG or PHZ antibiotics
was included for comparison purposes.
R� E�
S� U�
L� T�
S�
A�
n� ti��� b� i� o� ti��� c � p� r� o� d�
u� c� ti��� o� n� .� For many of the strains included in this study (Table 1), the ability to
produce PRN was determined or confirmed by TLC or HPLC analysis. PRN was detected by
TLC analysis in extracts of culture-filtrates of B. cepacia strains J2535, LMG 1222, and ATCC
51671; B. pyrrocinia DSM10685; P. chlororaphis LMD 82.53; P. fluorescens strains Pf-5,
CHA0, Pf1, PF, PFM2, PGNL1, PGNR1, PGNR2, PGNR3, PGNR4, PINR2, PINR3, and
Pseudomonas sp. strains PHZ13, PHZ19, PHZ22, PHZ24, PHZ48. In addition to PRN,
strains PHZ13, PHZ19, PHZ22, PHZ24, and PHZ48 also produced phenazine antibiotics as
was determined by HPLC analysis followed by spectral analysis of the peaks by photodiode
array spectroscopy (data not shown).
P� C� R� -� a� mp� � l�i� f� i� c� a� t� i� o� n � o� f
�p� rn�0� D
� a� n� d
�p� l� t� C g¡ e¢ n� e ¢ f
�r� a� g¡ m� e¢ n� t� s£ .� Primers PRND1 and PRND2 are
directed against conserved sequences within the prnD genes of B. pyrrocinia DSM10685, P.
fluorescens strain BL915, and B. cepacia strain LT4-12-W. PRND1 and PRND2 amplified the
predicted 786-bp fragment from DNA of homologous strain B. pyrrocinia DSM10685 (Fig.
1A). The 786-bp fragment was digested with 7 different restriction enzymes and the sizes of
the restriction fragments matched with the predicted sizes inferred from the prnD sequence of
B. pyrrocinia DSM10685 available in the database. A fragment of the same size was amplified
from DNA of all other known PRN-producing Pseudomonas and Burkholderia strains, whereas
no 786-bp fragment was amplified from several non-PRN-producing strains, including J2540
and PILH1 (Fig. 1A). The PCR products amplified for PRN-producing Pseudomonas and

C¤ h¥
a¦ pt§�¨ e© r ª 2: «#¬
P o® l¯y° m± o® rª p§ h
¥i² s³ m± s ³ w´ i² t¨ h¥ i² n µ t¨ h¥ e © p¶ rn·0 D
¹ a¦ nµ d
ºp¶ l» t¼ C ½ g¾ e© nµ e© s³
38
T¿
aÀ bÁ lÂe à 2. ÄÆÅ
CÇ
hÈ
aÀ rÉ aÀ cÊ tË eà rÉ iÌ sÍ tË iÌ cÊ s Í oÎ f Ï
tË hÈ e à pÐ rÉ iÌ mÑ eà rÉ s Í uÒ sÍ eà d Ó
fÏoÎ r É P
ÔCÇ
R Õ
aÀ nÖ aÀ l y× sÍ iÌ sÍ .Å
Primer Sequence Length(bp)
G+C(%)
Tm
(°Ø C)Posit. a Product
Length(bp)
Reference
PRND1 GGGGCGGGCCGTGGTGATGGA 21 76.2 82.4 5269PRND2 YCCCGCSGCCTGYCTGGTCTG 21 66.6 74.2 6055 786 This study
PLTC1 AACAGATCGCCCCGGTACAGAACG 24 58.3 74.2 12282PLTC2 AGGCCCGGACACTCAAGAAACTCG 24 58.3 73.8 12720 438 This study
Phl2a GAGGACGTCGAAGACCACCA 20 60 73 1915Phl2b ACCGCAGCATCGTGTATGAG 20 55 72 2660 745 Raaijmakers
et al., 1997PHZ1 GGCGACATGGTCAACGG 17 64.7 52.4 3428PHZ2 CGGCTGGCGGCGTATTC 17 70.6 57.3 4836 1408 Mavrodi et
al., unpubl.
a Position of the 5' end of the primer in the database sequence.
Burkholderia spp. all hybridized under high-stringent conditions with the PRN probe
consisting of the DIG-labeled 786-bp fragment from the homologous strain B. pyrrocinia
DSM10685 (Fig. 1B).
Primers PLTC1 and PLTC2 amplified the predicted 438-bp fragment from DNA of
homologous P. fluorescens strain Pf-5 (Fig. 2A). The 438-bp PLT fragment was digested with 8
different restriction enzymes and the sizes of the restriction fragments matched the predicted
sizes inferred from the pltC sequence from P. fluorescens Pf-5 available in the database. A
fragment of the same size was amplified from all known PLT-producing strains, whereas no
PCR product of that size was obtained from DNA of non-PLT-producing strains Q8r1-96,
J2535, J2540, and F113 (Fig. 2A). The PCR products of all PLT producers hybridized under
high-stringent conditions with the PLT probe consisting of the labeled 438-bp fragment
amplified from strain Pf-5 (Fig. 2B).
DÙ i² vÚ e© rª s³ it² ¨ y ° o® f Û
P RÜ NÝ
-Þ p§ rª o® dº
uß cà i² nµ g ¾ Psáãâ
euäæå dç
oè mé oè naê sâ a¦ nµ d
ºBë
uå r· kì hí oè l» dç eä r· iî aê s³ pp§ï§ . ð Restriction patterns of the
PRN-fragments (Fig. 3A), obtained by digestion with five endonucleases (HaeII, HincII,
HinfI, MnlI, and PvuII), yielded considerable polymorphisms (RFLPs) among the different
strains. Digestion with AccI and BsmBI yielded no additional polymorphisms. Cluster analysis
of the restriction fragment patterns resulted in seven distinct groups. The largest group
contained Pseudomonas strains that all produce both PRN and PLT (Fig. 3A, Table 1). There
was considerable diversity among B. cepacia strains that only produce PRN (LMG 1222,
J2535, ATCC 51671). The Pseudomonas strains that produce both PRN and phenazines
(PHZ13, PHZ19, PHZ22, PHZ24, PHZ48, LMD 82.53) were clustered in three groups

Cñ hò
aó ptô�õ eö r ÷ 2: ø#ù
Pú oû lüyý mþ oû r÷ pô h
òiÿ s� mþ s � w� iÿ tõ hò iÿ n � tõ hò e ö p� rn��� D
� aó n� d
�p� l t C � g� eö n� eö s�
39
F
i�g� u� r� e � 1. ��� A
�g� a� r� o� s� e � g� e� l � e� l� e� c� t� r� o� p� h
�o� r� e� s� i� s � o� f
�t� h� e � P
C!
R "
p� r� o� d#
u� c� t� s � a� m$ p� l�i�f�i�e� d #
f�r� o� m $ D
%N&
A �
o� f �
P'
s( e) u* d+ o, m- o, n. a/ s( a� n0 d#
B1
u* r2 k3 h4o, l5 d+ e) r2 i6 a/ s� t� r� a� i� n0 s � w7 i
�t� h �
p� r� i� m$ e� r� s � P
R"
N&
D%
1 � a� n0 d #
P
R"
N&
D%
28
(A). Characteristics of the bacterialstrains are shown in Table 1. Southern blot analysis of the PCR fragments shown in (A)with the PRN probe (786-bp fragment from B. pyrrocinia DSM10685) under high-stringent conditions (B).
F
i�g� u� r� e � 2.
8 � A�
g� a� r� o� s� e � g� e� l � e� l� e� c� t� r� o� p� h�
o� r� e� s� i� s � o� f �
t� h� e � P
C!
R "
p� r� o� d#
u� c� t� s � a� m$ p� l�i�f�i�e� d #
f�r� o� m $ D
%N&
A �
o� f �
P'
s( e) u* d+ o, m- o, n. a/ s( a� n0 d#
B1
u* r2 k3 h4o, l5 d+ e) r2 i6 a/ s� t� r� a� i� n0 s � w7 i
�t� h �
p� r� i� m$ e� r� s � P
L9
T:
C!
1 � a� n0 d #
P
L9
T:
C!
28
(A). Characteristics of the bacterialstrains are shown in Table 1. Southern blot analysis of the PCR fragments shown in (A)with the PLT probe (438-bp fragment from P. fluorescens Pf-5) under high-stringentconditions (B).
100-
bp
lad
der
DS
M10
685
J253
5A
TCC
5167
1
LMG
1222
J 25 4
0
Pf-
5C
HA
0P
f1P
HZ
13P
HZ
22P
HZ
24
PH
Z48
PG
NR
3P
INR
2P
FM
2
LM
D82
. 53
PIL
H1
600-bp
2072-bp
100-bp
786-bp
A
438-bp
600-bp
2072-bp
100-bp
100 -
bp la
dde
rP
f-5
CH
A0
Pf1
Q8r
1 -96
J253
5J2
540
PG
NL
1
PG
NR
2P
GN
R3
PG
NR
4P
INR
2P
INR
3F
113
PF
PFM
2
PG
NR
1
A
B

C; h<
a= pt>@? eA r B 2: CED
PF oG lHyI mJ oG rB p> h
<iK sL mJ s L wM iK t? h< iK n N t? h< e A pO rnP�Q D
R a= nN d
SpO lT tU C V gW eA nN eA sL
40
with a relatively high level of similarity. Interestingly, B. pyrrocinia DSM10685 clustered more
closely to Pseudomonas than to the other Burkholderia strains (Fig. 3A). RAPD analysis with
primers M13, M12, and D7 gave rise to 72 polymorphic markers, which ranged in size from
190 to 2072-bp. P. aeruginosa strain PAO1 was included as an outgroup (Fig. 3B). The
reproducibility of the amplification patterns was confirmed in three independent experiments.
Although RAPD analysis allowed a higher level of discrimination among PRN-producing
Burkholderia and Pseudomonas species than RFLP analysis of the PRN-fragment, the overall
clustering of the isolates by RAPD analysis was very similar to that obtained by RFLP analysis
(Fig. 3).
RX eA lH a= t? iK oG nN sL h< ip K > bY
eA t? wM eA eA n N p> h<
yI lHoG gW eA nN y I oG f
ZpO rnP�Q gW eA nN eA s L a= nN d
S16[]\ S ^ rB D_ NA
bacc lHa= ssLdL iK fZ iK cc a= ti? K oG nN .e RFLP analysis
showed that the prnD gene of B. pyrrocinia DSM10685 is more closely related to the prnD
genes of Pseudomonas than to those of the other Burkholderia strains (Fig. 3A). To further
investigate the phylogenetic relationships between PRN-producers of both genera,
representative PRN-producing Pseudomonas (P. fluorescens Pf-5, P. chlororaphis LMD 8253,
Pseudomonas sp. PHZ13 and PHZ22) and Burkholderia (B. cepacia LMG 1222, and J2535)
strains were selected for 16S rDNA sequencing. The 16S rDNA sequences from P. fluorescens
CHA0 and P. aeruginosa PAO1, were also included in the analysis. The phylogenetic trees
inferred from these sequences with the neighbour-joining method yielded a similar topology as
obtained by the UPGMA method; only the tree inferred with the latter method is presented
(Fig. 4). 16S rDNA sequence analysis yielded two main clusters, one that comprised all six
Pseudomonas strains and the second one containing all three Burkholderia strains, including B.
pyrrocinia DSM10685. These results are in agreement with their taxonomic status. For most
strains, phylogenetic prn biosynthetic locus were generally similar to 16S rDNA based
phylogeny except for B. pyrrocinia DSM10685. In silico analysis of the prnA-D sequences of at
least relationships inferred from each of the four prn genes as well as from the complete
sequence of the four PRN-producing strains showed that each of the four prn genes as well
as the complete prn biosynthetic locus of B. pyrrocinia DSM10685 is more closely related to
Pseudomonas than to B. cepacia (Fig. 5). These results clearly point to a discrepancy between
the phylogenies inferred from 16S rDNA sequences and prn biosynthetic genes.
D_ iK vf eA rB sL itK ? y I a= mJ oG nN g W PF Lg T-hji
p> rB oG dS
uk cc iK nN g W Pslnm
eo up dqor ms or nQ at s
m a= nN d
SBu
up rP kv hw olrxT dq eo rP iy a t cz eo pO at ciz�y at . e Restriction of the
PLT fragment with 8 different endonucleases (AlwI, AvaI, MnlI, NaeI, PstI, SacII, TaqI, and
XhoI) yielded no polymorphisms among twelve Pseudomonas strains (data not shown).

C{ h|
a} pt~@� e� r � 2: �E�
P� o� l�y� m� o� r� p~ h
|i� s� m� s � w� i� t� h| i� n � t� h| e � p� rn��� D
� a} n� d
�p� l� t� C � g� e� n� e� s�
41
F�
i�g� u� r� e � 3.
���D�
e� n d¡
r� o¢ g� r� a£ m¤ s ¥ o¢ f ¦
d¡
i�f¦f¦e� r� e� n t § P
¨R©
Nª
-« p¬ r� o¢ d¡
u� c i� n g � P®
s¯ e° u± d² o³ mo³ nµ a¶ s¯ a£ n d ¡
B·
u± rk¹hºo³ l» d² e° ri¼ a¶ s¥ p¬ p¬ ., b
� ½¿¾a£ s¥ e� d
¡o¢ n
r� e� s¥ t§ r� i� c t§ i� o¢ n a£ n a£ lÀ yÁ s¥ i� s ¥ o¢ f ¦
t§ h e � 786Ã�Ä�Å
-« b¾ p ¬ P¨
R©
N ª
f¦r� a£ g� m¤ e� n t§ (A). Restriction analysis was performed
with seven different restriction endonucleases. Dendrogram of the bacterial strains shownin (A), based on RAPD analysis with three different primers (B). Bootstrap values greaterthan 70% are shown at the branching points. Characteristics of the bacterial strains areshown in Table 1. Dendrograms constructed by using the UPGMA method are presented.
FÆ r� e� qÇ uÈ e� n� cÉ y � o� f Ê
p� rn��� D�
a} n� d �
p� l� t� C� -Ë h| a} r� bÌ o� urÈ@� i� n� g � PsÍnÎ
euÏÑÐ dÒ
oÓ mÔ oÓ n� aÕ s Î
s� pp~Ö~ . × i� n � d�
i� s� e� a} s� e� -Ë s� uÈ pp~@~ r� e� ss�Ø� i� vÙ e � s� o� il� � s� . × Several
studies have implicated antibiotic-producing bacteria in the natural suppressiveness of certain
soils to the take-all fungus G. graminis var. tritici (Charigkapakorn and Sivasithamparam,
1987; Raaijmakers et al., 1997; Raaijmakers and Weller, 1998). To begin to address a possible
role of PRN- and/or PLT-producing Pseudomonas spp. and B. cepacia in Dutch take-all
suppressive soils (TAD soils), the initial step was to determine their densities in the
rhizosphere of wheat grown in these soils. One of the five soils, designated CB, is
P. fluorescens CHA0P. fluorescens Pf1P. fluorescens PGNL1
P. fluorescens PGNR2P. fluorescens PGNR1
P. fluorescens PGNR4P. fluorescens PINR2P. fluorescens PINR3
P. fluorescens PFP. fluorescens Pf-5
P. fluorescens PFM2
Pseudomonas sp. PHZ19
B. cepacia LMG 1222B. pyrrocinia DSM 10685
B. cepacia ATCC 51671B. cepacia J2535P. aeruginosa PAO1
Pseudomonas sp. PHZ13
Pseudomonas sp. PHZ24Pseudomonas sp. PHZ22Pseudomonas sp. PHZ48
P. .chlororaphis LMD 82.53
1 0 % sim ila rity
B - RAPD
B. cepacia ATCC51671
0.05% s im ila rity
B. cepacia J2535B. cepacia LMG1222
Pseudomonas sp. PHZ22Pseudomonas sp. PHZ13
Pseudomonas sp. PHZ24
Pseudomonas sp. PHZ19
P. fluorescens PFM2P. chlororaphis LMD82.53
P. fluorescens PFP. fluorescens PINR3P. fluorescens PINR2P. fluorescens PGNR4P. fluorescens PGNR3
P. fluorescens CHA0
P. fluorescens Pf1P. fluorescens PGNL1P. fluorescens PGNR1P. fluorescens PGNR2
100100
100
100
A - RFLP

CÚ hÛ
aÜ ptÝ@Þ eß r à 2: áEâ
Pã oä låyæ mç oä rà pÝ h
Ûiè sé mç s é wê iè tÞ hÛ iè n ë tÞ hÛ e ß pì rní�î D
ï aÜ në d
ðpì lñ tò C ó gô eß në eß sé
42
Fõ
iög÷ uø rù e ú 4.
ûýüMþ
oÿ s� t � p� a� rù s� iö m� oÿ n� iöoÿ uø s � p� h
�y� l�oÿ g÷ eú n� eú t� iö c t� rù eeú ú s � g÷ eú n� eú rù a� t� eú d
w� iöt� h �
s� eú q� uø eú n� c eú s � oÿ f
p� r� n� A� (� A� )�, � p� r� n� B� (� B� )
�, � p� r� n� C�
(� C� )�, � p� r� n� D
� (� D� )
�, � a� n� d
t� h� e ú w� h
�oÿ l�e ú p� r� n� c l� uø s� eú r ù (� E� )
�.ü Position of the aligned nucleotides in relation
to sequences of P. fluorescens BL915 were as follows: prnA (positions 615 to 2231), prnB(positions 2231 to 3316), prnC (3368 to 5068), prnD (5093 to 6205), prn (only codingregions were combined). Bootstrap values higher than 70% are shown in the appropriatebranching points.
conducive to G. graminis var. tritici, and at least two of the other soils, designated SV and SSB,
are suppressive to G. graminis var. tritici (Table 3). Colony hybridization with the prnD and
pltC probes showed that Pseudomonas spp. harbouring these genes were not present at
detectable levels on roots of wheat grown in any of the five soils. Some putative PRN-
producing Pseudomonas spp. were initially isolated based on colony hybridization. However,
these isolates gave a PCR product slightly bigger than the 786-bp PRN fragment, which did
not hybridize at high- and low-stringent conditions with the PRN probe (data not shown).
Furthermore, PRN production by these isolates was not detected in TLC and HPLC analysis
(data not shown). In all soils, roots of wheat supported B. cepacia populations at densities
ranging from 9.5 X 104 to 2.5 X 106 CFU per gram of root plus adhering soil. In PCR analysis
of randomly selected B. cepacia isolates, no isolates that harbored prnD and pltC genes were
detected in any of the five soils tested. For comparison purposes, we also determined the
P. aureofaciens ACNP. fluorescens BL915P. fluorescens CHA0B. pyrrocinia DSM 10685B. cepacia LT4-12-WM. fulvus Mx f147
8199
89
A - prnA
B. cepacia LT4-12-WM. fulvus Mx f147
B. pyrrocinia DSM 10685P. fluorescens BL915
B - prnB
B. cepacia LT4-12-WM. fulvus Mx f147
B. pyrrocinia DSM 10685P. fluorescens BL91510 0
C - prnC
B. cepacia LT4-12-WM. fulvus Mx f147
B. pyrrocinia DSM 10685P. fluorescens BL915
D - prnD
10 0
B. cepacia LT4-12-WM. fulvus Mx f147
B. pyrrocinia DSM 10685P. fluorescens BL915

C� h�
a� pt "! e# r $ 2: %'&
P( o) l*y+ m, o) r$ p h
�i- s. m, s . w/ i- t! h� i- n 0 t! h� e # p1 rn243 D
5 a� n0 d
6p1 l7 t8 C 9 g: e# n0 e# s.
43
frequencies of two other groups of antibiotic-producing Pseudomonas spp. Pseudomonas spp.
harboring phlD (phlD+), a key gene involved in the biosynthesis of 2,4-DAPG, were found in
all five soils tested (Table 4). The densities of phlD+ Pseudomonas spp. ranged from 5.0 X 104
to 3.6 X 106 CFU per gram of root plus adhering soil, representing on average 0.2 to 19% of
the total population of culturable Pseudomonas spp. isolated from wheat roots. Pseudomonas
spp. harboring phzC-D, two genes involved in the biosynthesis of phenazine antibiotics, were
found only in SL soil at densities of 9.0 X 104 CFU g-1 root; these densities represent 0.5% of
the total population of culturable Pseudomonas spp. Isolation of naturally occurring antibiotic-
producing Pseudomonas spp. from roots in the seventh growth cycle of wheat growth cycle
yielded similar results.
D; I< S= CU�?> SS=@= I< OA NB
Pyrrolnitrin (PRN) and pyoluteorin (PLT) play a key role in the suppression of plant
pathogens by several strains of Pseudomonas and Burkholderia. In this study, primers were
developed from conserved sequences within the prnD and pltC genes, two genes that are
essential in the biosynthesis of PRN and PLT, respectively. Both primer sets amplified the
predicted fragments from DNA of a worldwide collection of PRN- and PLT-producing
Pseudomonas and Burkholderia strains. The availability of these specific primers and probes
allow to better understand the distribution and ecology of indigenous bacteria that produce
these antibiotics as well as the behavior of introduced transgenic microorganisms in which
these traits have been incorporated. For example, primers and probes directed against specific
genes involved in the biosynthesis of 2,4-DAPG have shown that Pseudomonas spp. harboring
this trait are present in different soils and plant-associated habitats (Keel et al., 1996). The
availability of primers and probes directed against antagonistic traits also enable a quick
screening of large collections of bacterial strains that have given promising results in
controlling plant diseases, thereby allowing site-directed mutagenesis to further unravel the
relative contribution of those traits in disease suppression.
In addition to the ability of Pseudomonas and Burkholderia species to produce
antibiotic compounds like PRN and PLT, their genotypic diversity has drawn considerable
attention. Genotypic diversity not only is an interesting phenomenon with respect to host
specificity and adaptation of microorganisms to changing abiotic conditions, but can also be

CC hD
aE ptF"G eH r I 2: J'K
PL oM lNyO mP oM rI pF h
DiQ sR mP s R wS iQ tG hD iQ n T tG hD e H pU rnV4W D
X aE nT d
YpU lZ t[ C \ g] eH nT eH sR
44
F^
i_g` ua rb e c 5. dfe D
gec nh di
rb oj g` rb ak m l oj f m
sn ec lo ec cp tq ec d i
sn tq rb ak i_ nh s n br
ak sn ec d i
oj n h 283sutuv
-w br p x ak lo i_ g` nh ec d i
sn ec qy ua ec nh cp ec s n oj f m
tq hz e c 16{}| S ~
rb Dg N�
A�
(positions 28-310 of nucleotide sequence of P. aeruginosa strain PAO1). Scale indicates thenumber of nucleotide substitutions per site. The dendrogram was constructed by using theUPGMA method.
exploited to improve the efficacy of biological control (Sharifi-Tehrani et al., 1998;
Raaijmakers and Weller, 2001). RFLP analysis of the 438-bp pltC gene fragment showed no
polymorphisms among PLT-producing Pseudomonas spp. Although this may be due to the fact
that the size of the amplified fragment is relatively small to detect differences, similar-sized
fragments have allowed the assessment of diversity among Pseudomonas strains to a level
similar to that obtained with other markers (de Vos et al., 1998; Dahllöf et al., 2000). Another
explanation for the lack of polymorphisms within the pltC fragment may be the fact that to
date PLT production seems to be limited to a select group of Pseudomonas strains, most of
which are genotypically very similar or even identical (Keel et al., 1996; Sharifi-Tehrani et al.,
1998; McSpadden-Gardener et al., 2000; Fig. 3).
In contrast to the pltC fragment, polymorphisms within the 786-bp prnD fragment
allowed the assessment of diversity to a level similar to that obtained by RAPD analysis with
three 10-mer primers. Although RAPD analysis provided a higher level of discrimination
between strains than RFLP analysis of the prnD gene, a similar overall clustering of the
antibiotic-producing strains was obtained (Fig. 3). Both RFLP and RAPD analyses showed
that strains producing both PRN and PLT are grouped closely together (Fig. 3; Table 1).
These results confirm and extend data obtained by Keel et al. (1996). Furthermore, the
Pseudomonas strains that produce both PRN and phenazines were clustered in three groups
with a relatively high level of similarity. Interestingly, Pseudomonas sp. PHZ48 and B.
pyrrocinia DSM10685 gave identical RFLP patterns and the position of this Burkholderia
strain was more close to Pseudomonas species than to the other Burkholderia strains included in
this study (Fig. 3A). Subsequent phylogenetic analysis of 16S rDNA sequences of a number of
strains representative of PRN-producing Pseudomonas and Burkholderia species correlated well
with their taxonomic status. Also phylogenetic relationships inferred from each of the four prn
P. fluorescens CHA0P. fluorescens Pf-5P. aeruginosa PAO1Pseudomonas sp. PHZ22Pseudomonas sp. PHZ13P. chlororaphis LMD 82.53B. pyrrocinia DSM 10685B. cepacia LMG 1222B. cepacia J2535
0 .05
100
95
91
100
72

C� h�
a� pt�"� e� r � 2: �'�
P� o� l�y� m� o� r� p� h
�i� s� m� s � w� i� t� h� i� n � t� h� e � p� rn�4� D
� a� n� d
�p� l� t� C � g� e� n� e� s�
45
T�
a� b� l�e 3. ¡£¢
F¤
r¥ e q¦ u§ e n¨ c© y ª o« f ¬
a� n¨ t i® b� i®o« t i® c© -¯ p° r¥ o« d
±u§ c© i® n¨ g ² P
³s´ eµ u¶ d
·o¸ m¹ o¸ nº a» s´ s¼ p° p° .
¢i®n ¨ t h½ e w¾ h
½e a� t r¥ h½ i
®z¿ o« s¼ p° h
½e r¥ e .¢
Soil Suppressive a
to take-all Densities (CFU g-1 root) b
Total pop.c 2,4-DAPGd PHZ e PRN f PLT g
CB - 2.3 X 107 5.0 X 104 n.d. n.d. n.d.SL n.t. 1.9 X 107 2.3 X 105 9.8 X 104 n.d. n.d.SU n.t. 1.8 X 107 3.3 X 105 n.d. n.d. n.d.SV + 1.3 X 107 3.5 X 105 n.d. n.d. n.d.SSB + 1.9 X 107 3.6 X 106 n.d. n.d. n.d.
a - conducive, + suppressive.b Mean values of four replicates are presented.c Total population of Pseudomonas spp.d Population of Pseudomonas spp. harboring the phlD gene.e Population of Pseudomonas spp. harboring phzCD genes.f Population of Pseudomonas spp. harboring the prnD gene.g Population of Pseudomonas spp. harboring the pltC gene.n.d.: not detected (detection limit was 104 CFU g-1 root).n.t. : not tested.
genes as well as from the complete sequence of the prn biosynthetic locus were similar to 16S
rDNA-based phylogeny for most strains, except for B. pyrrocinia DSM10685. In silico analysis
of each of the four individual genes contained in the prn biosynthetic locus confirmed that in
all cases B. pyrrocinia DSM10685 clustered closer to Pseudomonas species than to B. cepacia
(Fig. 4). Comparable results have been obtained in nodC and nifH gene analysis of Rhizobium
and Sinorhizobium (Laguerre et al., 2001). Genetic rearrangements and lateral gene transfer
were suggested as possible explanations for the discrepancy between phylogenies inferred from
16S rDNA sequences and nodC and nifH phylogenies (Laguerre et al., 2001). The extent of
occurrence of horizontal gene transfer among prokaryotes is now becoming more evident.
Comparative studies of entirely sequenced genomes are showing that gene transfer plays a
greater role in the diversification of microorganisms than thought before (Jain et al., 1999). It
is also known that operational genes, such as those involved in antibiotic production are more
likely to be transferred than genes involved in transcription, translation, replication, and
related processes (Rivera et al., 1998). Although our results may suggest that horizontal
transfer of the prn operon occurred between Pseudomonas and B. pyrrocinia, more prn
biosynthetic loci of multiple Burkholderia and Pseudomonas species need to be sequenced to
support this hypothesis. The genus Pseudomonas belongs to the γ-subclass of the
Proteobacteria, whereas Burkholderia belongs to β-subclass, based on sequence analyses of the
16S rDNA gene, which is considered as a standard in phylogeny and taxonomy of living
organisms (Maidak et al., 1999; Anzai et al., 2000). The discrepancies between prnD and 16S
rDNA phylogenies in B. pyrrocinia DSM10685 ilustrate the limitations of the former in

CÀ hÁ
a ptÃ"Ä eÅ r Æ 2: Ç'È
PÉ oÊ lËyÌ mÍ oÊ rÆ pà h
ÁiÎ sÏ mÍ s Ï wÐ iÎ tÄ hÁ iÎ n Ñ tÄ hÁ e Å pÒ rnÓ4Ô D
Õ a nÑ d
ÖpÒ l× tØ C Ù gÚ eÅ nÑ eÅ sÏ
46
phylogenetic studies. Conversely, Ramette et al. (2001) found a clear correspondence between
polymorphisms within the phlD gene, one of the genes responsible for 2,4-DAPG biosynthesis
in Pseudomonas, and phylogenetic analysis based on 16S rDNA sequences.
Molecular probes and primers have been used extensively in environmental
microbiology to detect pathogenic bacteria and fungi, but have received very little attention in
biocontrol research. Specific primers and probes not only allow targeted isolation of strains
with specific biocontrol traits, but also can be very useful tools to study their role in natural
disease-suppressive soils. Primers and probes developed for detection and isolation of
Pseudomonas spp. that produce 2,4-DAPG or PHZ antibiotics (Raaijmakers et al., 1997), were
successfully used to study their diversity and population dynamics in wheat and maize
rhizosphere (McSpadden-Gardener et al., 2000; Picard et al., 2000), and to elucidate their role
in the natural suppression that operates in soils that are suppressive to take-all disease of wheat
(Raaijmakers and Weller, 1998). Recently, a PRN-producing P. fluorescens strain was isolated
from a G. graminis var. tritici suppressive soil in Japan and was shown to be effective in
controlling take-all of wheat (Tazawa et al., 2000). In this study, colony hybridization
followed by PCR showed that prnD- and pltC-harboring Pseudomonas and Burkholderia spp.
were not present at detectable levels in the rhizosphere of wheat grown in five soils obtained
from two areas in the Netherlands. Two of these soils were shown to be suppressive to G.
graminis var. tritici (Table 4). The fact that prnD- and pltC-harbouring Pseudomonas and
Burkholderia spp. were not present at detectable levels in the wheat rhizosphere (Table 4) may
suggest that they do not play a role in the natural suppresiveness of at least two Dutch take-all
decline soils. However, this suggestion is based on the assumption that these bacteria need to
reach threshold densities of at least 105 CFU g-1 root as reported earlier for 2,4-DAPG-
mediated suppression of the take-all fungus (Raaijmakers and Weller, 1998) and suppression
of Fusarium wilt of radish via siderophores or induced systemic resistance (Raaijmakers et al.,
1995). Whether similar threshold levels apply to PRN and PLT producers remains to be
determined. Pseudomonas spp. harboring phlD, one of the genes involved in biosynthesis of the
antibiotic 2,4-DAPG, were detected at densities similar to those reported by other authors for
the wheat and maize rhizosphere (Raaijmakers et al., 1997; Picard et al., 2000). The
occurrence of relatively higher populations of 2,4-DAPG-producing Pseudomonas spp. in the
two take-all decline soils SV and SSB than in the take-all conducive soil CB (Table 4), point
to a more prominent role of 2,4-DAPG-producing Pseudomonas spp. in Dutch take-all decline
soils, a question that is currently being addressed.

CÛ hÜ
aÝ ptÞ"ß eà r á 2: â'ã
Pä oå læyç mè oå rá pÞ h
Üié sê mè s ê wë ié tß hÜ ié n ì tß hÜ e à pí rnî4ï D
ð aÝ nì d
ñpí lò tó C ô gõ eà nì eà sê
47
Di Cello et al. (1997) demonstrated that B. cepacia occurs naturally on maize roots at
densities of approximately 3.2 X 106 CFU g-1 root. In this study, similar densities were found
for B. cepacia in the wheat rhizosphere. Burkholderia spp. have received considerable attention
due to their nutritional versatility for bioremediation and their antibiotic properties for
biocontrol (Homma and Suzui, 1989; Hebbar et al., 1998; Vandamme et al., 1997). However,
because these bacteria are also implicated in infections of cystic fibrosis patients, their
application in biological control has become very controversial and subject of many debates. In
fact, with exception of B. pyrrocinia, all genomovars of Burkholderia (seven genomovars
described) are able to infect or colonize cystic fibrosis patients (Parke and Gurian-Sherman,
2001). A reliable and conclusive technique which allows the differentiation between
environmental Burkholderia spp. from those associated with cystic fibrosis is not yet available,
although some differences have been reported (Bevivino et al., 1994; Tabbachioni et al.,
1995). Therefore, their use in biological control should be avoided, at least until clear
delimitations between human pathogenic and environmental Burkholderia strains are
conclusively established.
Mö
A÷
Tø Eù Rú IûA÷
Lü
S ý A÷
Nþ
D ÿ
Mö
Eù Tø H�
O� Dÿ
SýB� a� c� t� e� r� i� a� l s t� r� a� i� n� s a� n� d � c� u l
t� u r� e � c� o� n� d� i
�t� i� o� n� s .�
Characteristics of the reference strains used in thisstudy are given in Table 1. Naturally occurringPseudomonas spp. were isolated from roots of wheatgrown in raw soils on King's medium B [KMB]agar supplemented with chloramphenicol (13 µgml-1), ampicillin (40 µg ml-1), and cycloheximide(100 µg ml-1) [KMB+] (Simon and Ridge, 1974).Burkholderia cepacia strains were isolated on trypanblue-tetracycline medium (TB-T) (Hagedorn et al.,1987) amended with cycloheximide (100 µg ml-1) toinhibit fungal growth. The combination ofingredients present in TB-T medium provides ahigh degree of selectivity to B. cepacia biotypes(Wise et al., 1995; Tabacchioni et al., 2000). Allstrains were stored at -80°C in Luria Bertani (LB)or KMB-broth supplemented with glycerol to afinal concentration of 40% (v/v). For detection ofPLT by high-performance liquid chromatography(HPLC) or by thin layer chromatography (TLC),bacterial strains were grown in 25 ml of KMB-brothin 50-ml screw cap vials for 14 days withoutshaking at 25°C. For detection of PRN by TLC,bacterial strains were grown in 25 ml of a minimalmedium containing per liter: 30 g glycerol, 3 gK2HPO4, 0.5 g KH2PO4, 5 g NaCl, 0.5 gMgSO4.7H2O, 0.61 g D-tryptophan. Autoclaved
medium was amended with filter-sterilized (0.2 µm)ZnSO4 (0.35 mM) and Mo7(NH4)6O24.4H2O (0.5mM) (Duffy and Défago, 1999; Salcher andLingens, 1980). Bacterial isolates were shaken (180rpm) for 24 h at 25°C and subsequently incubatedat 25°C in the dark for four additional days withoutshaking. The oxygen-limiting conditions created bythe absence of stirring are necessary to elicitproduction of phenolic compounds, includingpyoluteorin (Bencini et al., 1983). For DNAextractions to be used for 16S rDNA sequencing,bacteria were cultivated overnight at 24°C onnutrient broth yeast extract agar (Vidaver, 1967).
Tø Lü
C � a� n� d � H�
P� Lü
C � a� n� a� l y� s i� s .� To detect PRN by TLC,15 ml of bacterial culture was centrifuged (SorvallRC-5, Du Pont, Wilmington, Delaware, USA) for15 min at 8700 X g, supernatant was discarded andthe cells were extracted twice with 5 ml ofethylacetate by sonicating (Bransonic 12, BransonUltrasonics Corp., Geneva, Switzerland) themixture for 3 min. For PLT, 15 ml of bacterialculture broth was extracted twice with 5 ml ofethylacetate. The organic phase was evaporated todryness and the residue was resuspended in 150 µlethylacetate. A volume of 50 µl was applied toprecoated 0.25 mm silica gel plate (20 X 20 cm,

C� h�
a� pt��� e� r � 2: ���
P� o� l y! m" o� r� p� h
�i# s$ m" s $ w% i# t� h� i# n & t� h� e � p' rn(*) D
+ a� n& d
,p' l- t. C / g0 e� n& e� s$
48
aluminium oxide 60 F254, Merck, Darmstadt,Germany). Separation was performed withchloroform-acetone (9:1 v/v) as the solvent system.The corresponding PRN spots were detected byspraying with 2% p-dimethylaminobenzaldehyde(Ehrlich's reagent, Sigma Aldrich Chemie B.V.,Zwijndrecht, The Netherlands) dissolved inethanol-sulfuric acid (1:1 v/v). PLT spots weredetected by spraying with an aqueous 0.5% FastBlue RR salt solution (Sigma Aldrich). The Rfvalues were 0.80 for PRN and 0.50 for PLT, asdetermined by migration of pure PRN, andconfirmed by identical color (crimson spot) and Rfvalues reported for PLT (Bencini et al., 1983;Burkhead, 1994). For detection of PRN and PLTby HPLC, 15 ml of the bacterial culture broth wasextracted and analysed following protocolsdescribed previously (Bonsall et al., 1997).
P1 r2 i3 m4 e5 r2 s6 .7 Characteristics of the primers used in PCRare listed in Table 2. Primers PRND1 and PRND2were developed from conserved sequences withinthe prnD gene of P. fluorescens strain BL915(GenBank U74493), B. pyrrocinia (GenBankAF161186), and B. cepacia strain LT4-12-W(GenBank AF161183) (Hammer et al., 1999).Sequence alignment and primer location wereperformed with the DNAstar program (DNAstarInc., Madison, WI, USA) and checked manually.7The 1092-bp prnD gene of P. fluorescens strainBL915 encodes a 363-amino acid protein thatshows homology to 3-chlorobenzoate-3,4-dioxygenase (CbaA) of Alcaligenes sp., phthalate4,5-dioxygenase (Pht3) of P. putida, and vanilatedemethylase (VanA) of Pseudomonas sp. (Hammeret al., 1997). The prnD gene product catalyzes theoxidation of the amino group of aminopyrrolnitrinto a nitro group to form pyrrolnitrin (Kirner et al.,1998). The PLT primers PLTC1 and PLTC2 weredeveloped from sequences within the pltC genefrom P. fluorescens strain Pf-5 (GenBankAF081920) (Nowak-Thompson et al., 1999). The5322-bp pltC gene has homology with eryA genes ofSaccharopolyspora erythraea, which encode apolyketide synthase required for erythromycinproduction. The 1774-amino acid protein encodedby pltC is probably responsible for the formation ofthe resorcinol ring during pyoluteorin biosynthesis(Nowak-Thompson et al., 1997). Primers weresynthesized by Amersham Pharmacia Biotech(Roosendaal, The Netherlands).
P1 C8 R9 -: a; n< a; l= y> s6 i3 s6 . 7 Heat-lysed bacterial suspensions wereprepared as described previously (Raaijmakers et al.,1997) and diluted 20-fold in sterile distilled water.PCR amplifications were performed in a 25-µlreaction mixture containing 3 µl of the diluted heat-lysed cell suspension, 1X PCR buffer (Perkin-Elmer, Nieuwerkerk aan de IJssel, TheNetherlands), 1.5 mM MgCl2 (Perkin Elmer), 200µM of each, dATP, dCTP, dGTP, and dTTP(Promega, Leiden, The Netherlands), 20 pmol of
each primer, and 2.0 U of AmpliTaq polymerase(Perkin-Elmer). For RAPD analysis (see below), 40pmol of a determined primer was used.Amplifications were carried out with an MJResearch PTC-200 thermocycler. The PCRprogram for PRN consisted of an initialdenaturation at 95°C for 2 min followed by 30cycles at 95°C for 1 min, annealing at 68°C for 1min, and extension at 72°C for 1 min. For PLT, aninitial denaturation at 95°C for 2 min was followedby 30 cycles of 95°C for 2 min, annealing at 67°Cfor 1 min, and extension at 72°C for 1 min. ThePCR program for RAPD analysis was described byKeel et al. (1996). Samples (10-15 µl) of the PCRproducts were separated on a 1% agarose gel or on2% agarose gels for RAPD analysis in 1X TBEbuffer (90 mM Tris-borate, 2 mM EDTA [pH8.0]) at 60-80 V for 1-3 h. The GIBCO-BRL 100-bp ladder (Life Technologies B.V., Breda, TheNetherlands) was used as size marker. PCRproducts were visualized with a UVtransilluminator.
R9 F?
L@
P 1 a; n< d A R9 AB
P1 D C a; n< a; l= y> s6 i3 s6 . 7 For all reference strains,DPRN and PLT fragments were amplified with thePRND1/PRND2 and PLTC1/PLTC2 primers,respectively (Table 2). The 786-bp fragment forPRN and the 438-bp for PLT were the only DNAfragments amplified with these primers for allreference strains producing PRN and PLT (Fig. 1Aand 2A). The PCR-fragments were precipitatedwith 4M LiCl and resuspended in sterile distilledwater. Digestions were performed in a total volumeof 15 µl, containing 1 µg of DNA, 1X reactionbuffer, and 10 U of each enzyme separately (NewEngland Biolabs Ltd., Hitchin, Hertfordshire,England; Promega Corporation, Leiden, TheNetherlands; and Life Technologies B.V.). Theenzymes used were AccI, BsmBI, HaeII, HincII,HinfI, MnlI, PvuII for digestion of the PRN-fragment and AlwI, AvaI, MnlI, NaeI, PstI, SacII,TaqI, and XhoI for digestion of the PLT-fragment.The mixtures were incubated for 2 h at 37°C, withexception of TaqI, which was incubated at 65°C.Digestion products were separated on a 1.5%agarose gel.
RAPD analysis of the reference strainscarrying genes for PRN (Table 1) was performedwith primers M13, M12, and D7. These primerswere selected among sixty-four other randomprimers based on the distinct and consistentbanding pattern of the polymorphic markersproduced (Keel et al., 1996). The size of the bandswas determined based on the 100-bp ladder as sizemarker. The presence or absence of fragmentsgenerated by RFLP and RAPD analysis (1 for thepresence or 0 for the absence of a particular sizedband in the gel) was used to calculate the pairwisecoefficients of similarity (Nei-Li distances). Clusteranalysis with the unweighted pair group methodwith arithmetic mean algorithm (UPGMA) andbootstrap analysis with a 1000 resampled data sets

CE hF
aG ptH�I eJ r K 2: L�M
PN oO lPyQ mR oO rK pH h
FiS sT mR s T wU iS tI hF iS n V tI hF e J pW rnX*Y D
Z aG nV d
[pW l\ t] C ^ g_ eJ nV eJ sT
49
were performed with the program FreeTree (Hamplet al., 2001). The dendrograms were visualised andedited in TREEVIEW (Page, 1996) andTreeExplorer (Tamura, 1997). RAPD analysis wasperformed three times for each primer and RFLPwas performed once.
Soa ub tc hd ee rf n g ah ng d i cj oa lk oa ng y l hd yl bm rf in di in zo ah tc in oa ng sp .q DNA transferfrom agarose gels and bacterial colony transfer toHybond N+ nylon membranes (AmershamPharmacia Biotech) were performed according tostandard methods (Sambrook and Russel, 2001).Membrane washes and hybridizations wereperformed following standard procedures(Sambrook and Russel, 2001). High-stringentconditions comprised prehybridization for 1.5 h at65°C, hybridization for 12 h at 65°C, membranewashing twice each for 5 min with 2X SSC (1XSSC is 0.15 M NaCl plus 0.015 M sodium citrate)-0.1% SDS at room temperature, and membranewashing twice each for 30 min with 0.1X SSC-0.1% SDS at 65°C. Low stringent conditionsconsisted of prehybridization and hybridization at55°C and two membrane washing steps with 0.5XSSC at 55°C. PRN- and PLT-probes weregenerated by random primed labeling of amplifiedPRN or PLT fragments with digoxigenin-dUTPafter PCR or by direct labeling in PCR using theDIG labeling mix (Roche, Almere, TheNetherlands). The 786-bp fragment amplified fromB. pyrrocinia strain DSM10685 was used togenerate the PRN-probe. The 438-bp fragmentamplified from P. fluorescens Pf-5 was used togenerate the PLT-probe. Hybridized probes wereimmunodetected according to the protocolsprovided by the supplier.
16rts S ` rf Du Nv
A w
sp ee qx ub ee ng cj in ng gy .q Total DNA for use in PCRamplifications of the 16S rDNA genes wasgenerated using a modification of the method usedby Raaijmakers et al. (1997). Bacteria were placedin 0.6 ml micro-centrifuge tubes and suspended in100 µl of lysis buffer and incubated in a water bathat 100ºC for 15 min. Micro-centrifuge tubes werevortexed for 30 sec, and tubes were returned to thewater bath for an additional 15 min. The cellsuspension was centrifuged for 1 min at 14,000rpm and the supernatant was transferred to a 1.5 mlmicro-centrifuge tube and diluted with 1.4 mlsterile distilled water. Five microliters of the celllysis suspension was used in each reaction whichemployed the primers 8f and 1406r (Amann et al.,1995) using reaction conditions essentially asdescribed by Liu et al. (1997). PCR-amplifiedproducts were analyzed by electrophoresis on 2%agarose gels. Gels were stained with ethidiumbromide and visualized by UV illumination andphotographed. Fragments of the appropriate sizewere cloned into the T/A vector pCR2.1(Invitrogen, Carlsbad, CA), and transformed into E.coli strain INVαF’. Transformants were selected onLB agar amended with kanamycin (75 µg ml-1) and
X-Gal (20 µg ml-1). Plasmid DNA was isolated fromtransformants using the S.N.A.P. mini-prep kit(Invitrogen). Cycle sequencing was conducted on aPerkin-Elmer model 480 thermal cycler (Norwalk,CT) and PRISM Ready Reaction DyeDeoxyTerminator Cycle Sequencing Kit (PE Biosystems,Warrington, UK). Reactions were conducted usingcycling conditions recommended by themanufacturer with the T3 or T7 primer. Productsof the sequencing reactions were purified usingCENTRI-SEP spin columns (PrincetonSeparations, INC., Adelphia, NJ) and dried in avacuum centrifuge. Samples were resuspended in 6µl of a solution containing deionized formamideand blue dextran/EDTA in a ratio of 5:1. Sampleswere heated for 2 min at 90ºC and 1.5 µl wereloaded on an ABI 377 DNA sequencer (PEBiosystems, Foster City, CA). BLASTN searcheswere conducted on the nucleotide sequencesobtained.
Nv
ub cj lk ee oa tc in di e e sp ee qx ub ee ng cj e e ah ccjzj ee ssp{p in oa n ng|g ub m} bm ee rf sp .q Newlyobtained sequences of the 16S rDNA gene weredeposited with GenBank under the followingaccession numbers: B. cepacia J2535 (AF495866),B. cepacia LMG 1222 (AF495867), P. chlororaphisLMD 82.53 (AF495868), Pseudomonas sp. PHZ13(AF495869), Pseudomonas sp. PHZ22 (AF495870),and P. fluorescens Pf-5 (AF495871). Additionalsequences of the 16S rDNA were obtained fromGenBank for the following strains: P. aeruginosaPAO1 (AE004091), P. fluorescens CHA0(AJ278812), and B. pyrrocinia DSM10685(U96930). Accession numbers for sequences of prnbiosynthetic genes available in the database are: P.fluorescens BL915 (U74493), B. pyrrociniaDSM10685 (AF161186), B. cepacia LT4-12-W(AF161183), M. fulvus Mx fl47 (AF161185), prnAof P. fluorescens CHA0 (AF161184), and prnA fromP. aureofaciens ACN (AF161182).
S` ee qx ub ee ng cj e e ah ng ah lk yl sp ee sp .q PRN and 16S rDNA sequenceswere aligned by CLUSTAL W 1.81 (Thompson etal., 1994). Sites presenting alignment gaps were notconsidered in phylogenetic inferences. UPGMAdendrograms with the nucleotide substitutionmodel of Jukes and Cantor (1969) and bootstrapanalysis with 1000 resamplings were performedwith the program MEGA (Kumar et al., 2001). Forthe in silico analysis of the prnABCD genes, mostparsimonious phylogenetic trees were obtained byusing MEGA. Phylogenetic trees were constructedby the maximum likelihood method usingDNAML contained in PHYLIP 3.5c Package(Felsenstein, 1993) after generation of 100resampled datasets with PHYLIP’s programSEQBOOT and finally obtaining the consensustree with PHYLYP’s program CONSENSE. Treeswere also constructed by the maximum parsimonymethod using MEGA (Kumar et al., 2001).Bootstrap analyses were performed with 1000

C~ h�
a� pt��� e� r � 2: ���
P� o� l�y� m� o� r� p� h
�i� s� m� s � w� i� t� h� i� n � t� h� e � p� rn�*� D
� a� n� d
�p� l� t� C � g� e� n� e� s�
50
resamplings. Phylogenetic trees were visualized andedited as mentioned earlier.
S� o� i� l� s� .� Five soils were obtained from agriculturalfields in the Netherlands. Soils CB, SU, SV, andSSB were collected in December 1997 and soil SLin June 1998 from the upper 50 cm of the soilprofile, air dried for a week, and passed through a0.5-cm mesh screen prior to use. Soils CB, SU, SV,and SSB were collected from a polder in the South-West of the Netherlands, located 10 km from thecity of Bergen op Zom. These soils arephysicochemically very similar, containing onaverage, 27% clay, 10% silt, and 51% sand. Theorganic matter content is on average 4.3% and thepH, determined after extraction with CaCl2 is 7.5.Soil SL was obtained from a polder in the North-East of the Netherlands, at Lovinkhoeve, located 30km from the city of Lelystad. SL soil contains 21%clay, 12% silt, 54% sand, 3.6 % organic matter,and pH, determined after extraction with CaCl2 is7.5. Under field conditions, all soils have beencropped continuously to wheat, with exception ofCB soil, which was cropped to wheat and sugar beetin a rotation scheme (1:2).
P� l�a n¡ t ¢ c£ u¤ l
�t¢ i� v¥ a t¢ i� o� n¡ . � Pots containing 200 g of sieved
soil were sown with 15 seeds of wheat, cv. Bussard.Plants were grown in a climate chamber at 15°Cwith a 12-h photoperiod. Plants received 50 ml of1/3 strength Hoagland's solution (macroelementsonly) twice a week. After 30 days, the roots wereharvested and the soil remaining plus excised rootswere mixed, returned to the same pot and sownagain with 15 wheat seeds. The process of growingwheat and harvesting roots was repeated seventimes. For each soil, four replicates were used andthe whole experiment was performed twice.
I¦ s� o� l�a t¢ i� o� n ¡ o� f § P¨ s© eª u« d¬ o m® o n¯ a° s© s� pp±²± . � a n¡ d ³ B. µ c¶ eª p· a° c¶ i¸ a° f§ r¹ o� mº
w» h¼ e½ a t ¢ r¹ oo�¾� t¢ s� . � Population densities of total, and prnDand pltC-harbouring Pseudomonas spp. weredetermined on roots of wheat harvested from thefirst and seventh successive growth cycle. For B.
cepacia, total populations and populationsharboring prnD and pltC genes were determined inthe second, third, fourth, fifth and sixth growthcycles. For each replicate, roots of five randomlyselected plants were harvested and loosely adheringsoil was gently removed. One gram of roots plusadhering rhizosphere soil was suspended in 5.0 mlof sterile distilled water, vortexed for 1 min andsonicated for 1 min in a ultrasonic cleaner. Forisolation and enumeration of pseudomonads,samples were dilution plated onto KMB+ and plateswere incubated at 25°C for 48 h. For enumerationof B. cepacia, serial dilutions were plated onto TB-Tmedium and incubated for 4 days at 25°C.Population densities of Pseudomonas spp.harbouring prnD and/or pltC were determined bycolony hybridization and PCR. Population densitiesof B. cepacia harbouring prnD and/or pltC weredetermined by PCR; for each soil, 216 randomlyselected colonies were tested.
S� u¤ pp±¿± r¹ e½ ss�{� i� v¥ e½ n¡ e½ ss �À� o� f § s� o� i� l� s � t¢ o � GÁ . µ g rà a° m® in¯ is © v¥ a r¹ .� tÄ rà itÄ i¸ c¶ i¸ .Soils were amended with 0.1% (wt/wt) of an oatgrain inoculum of G. graminis var. tritici isolate R3-111a-1 (particle size 0.25 to 0.50 mm); sterilizedoat grain inoculum was used as a control. Wheatseeds (cv. Bussard) were sown in PVC pots (8 cmhigh, 7 cm wide) containing 200 g of sieved soil,and covered with a 1-cm layer of soil withoutinoculum. Plants were grown as described above.Each treatment had five replicates withapproximately 12 plants per replicate. Eightrandomly selected plants were harvested, their rootsystem washed and disease severity determined on a0-to-8 scale, where “0” indicates no disease and “8”indicates dead plant (Thomashow and Weller,1996). The experiment was performed twice. Datafrom disease index were ranked and analysed byANOVA followed by Tukey’s studentized rangetest, after certifying normal distribution andhomogeneity of variances (SAS institute, Inc.,Cary,N.C.).
AÅ
CÆ
KÇ NÈ OÉ WÊ
LË
EÌ
DÍ GÎ
EÌ
MÏ
EÌ
NÈ TÐ
S�
The authors greatly acknowledge CAPES/Brazil for financial support (Project nr. 1515/96-9). We thankDr. Mark Mazzola for 16S rDNA sequencing, Dr. Linda S. Thomashow for providing strains PGNR1, PGNR2,PGNR3, PGNR4, PGNL1, PINR2, PINR3, PF, and PFM2; Dr. G. Défago and Dr. Alban Ramette for isolatesPf1 and CHA0; Dr. Joyce loper, for providing isolate Pf-5; Dr. P. Lemanceau for providing strains PHZ13,PHZ19, PHZ22, PHZ24, and PHZ48; Dr. P. Vandamme for strain LMG 1222; Dr. M. Hutchinson for strainsJ2535, and J2540; Dr. M. de Waard for purified pyrrolnitrin and Wim van Doesburg for HPLC analysis. Wethank Dr. Pierre de Wit for critically reading the manuscript.

CÑ hÒ
aÓ ptÔ�Õ eÖ r × 2: Ø�Ù
PÚ oÛ lÜyÝ mÞ oÛ r× pÔ h
Òiß sà mÞ s à wá iß tÕ hÒ iß n â tÕ hÒ e Ö pã rnä*å D
æ aÓ nâ d
çpã lè té C ê gë eÖ nâ eÖ sà
51
Rì Eí Fî
Eí Rì Eí Nï
Cð
Eí Sñ
Aò
mó aô nnõöõ , ÷ Rì .ø Iù ., ø ÷ Lú
uû dü wý iþgÿ , ÷ W
�., ø ÷ aô nõ d ü Sñ c� h� l
�e� iþ f� e� r� , ÷ K
�. ø H� .ø
1995. Phylogenetic identification and in situdetection of individual microbial cells withoutcultivation. Microbiol. Rev. 59:143-169.Aò
nõ z aô iþ , ÷ Y
., ø ÷ K�
iþmó , ÷ H� ., ø ÷ P� aô r� k� , ÷ J
.ø -� Y .ø , ÷ W
�aô k� aô b� aô y� aô s� h� i
þ, ÷ H� .,ø ÷
aô nõ d ü O� y� aô iþ z uû , ÷ H� .ø 2000. Phylogenetic affiliation ofthe pseudomonads based on16S rRNA sequence.Int. J. Syst. Evol. Microbiol. 50:1563-1589.Aò
r� iþ mó aô , ÷ K�
., ø ÷ Iù mó aô nõ aô k� aô , ÷ Iù ., ø ÷ K�
o� uû s� aô k� aô , ÷ M�
., ø ÷ Fî
uû k� uû t� aô , ÷ Aò
.,ø ÷aô nõ d ü T� aô mó uû r� aô , ÷ G
�.ø 1964. Pyrrolnitrin, a new
antibiotic substance, produced by Pseudomonas.Agr. Biol. Chem. 28:575-576.B� aô nõ gÿ e� r� aô , ÷ M
�.ø G� ., ø ÷ aô nõ d ü T� h� o� mó aô s� h� o� wý , ÷ L
ú.ø Sñ .ø 1999.
Identification and characterization of a gene clusterfor synthesis of the polyketide antibiotic 2,4-diacetylphloroglucinol from Pseudomonas fluorescensQ2-87. J. Bacteriol. 181:3155-3163.B� e� nõ c� iþ nõ i
þ, ÷ D�
.ø Aò ., ø ÷ H� o� wý e� ll��� , ÷ Cð
.ø Rì ., ø ÷ aô nõ d ü W�
iþl�dü , ÷ J
.ø Rì . ø 1983.
Production of phenolic metabolites by a soilpseudomonad. Soil Biol. Biochem. 15:491-491.B� e� nõ dü e� r� , ÷ C
ð.ø Lú ., ø ÷ Rì aô nõ gÿ aô s� wý aô mó y� , ÷ V
�., ø ÷ aô nõ d ü L
úo� p� e� r� , ÷ J
.ø Eí .ø
1999. Polyketide production by plant-associatedpseudomonads. Annu. Rev. Phytopathol. 37:175-196.B� e� v� i
þv� iþnõ o� , ÷ A
ò., ø ÷ T� aô b� aô cc��� h� i
þo� nõ iþ, ÷ Sñ ., ø ÷ C
ðh� iþaô r� iþ nõ i
þ, ÷ Lú
., ø ÷ Cð
aô r� uû s� iþ ,÷M�
.ø V� ., ø ÷ D�
e� l � G�
aô l� l� o� , ÷ M�
., ø ÷ aô nõ d ü V�
iþs� c� aô , ÷ P� . ø 1994.
Phenotypic comparison between rhizosphere andclinical isolates of Burkholderia cepacia.Microbiology 140:1069-1077.B� o� nõ s� aô ll��� , ÷ Rì .ø Fî ., ø ÷ W
�e� l� l� e� r� , ÷ D
�.ø M� ., ø ÷ aô nõ d ü T� h� o� mó aô s� h� o� wý , ÷ L
ú.ø Sñ .ø
1997. Quantification of 2,4-diacetylphloroglucinolproduced by fluorescent Pseudomonas sp. in vitroand in the rhizosphere of wheat. Appl. Environ.Microbiol. 63:951-955.B� uû r� k� h� e� aô dü , ÷ K
�.ø D� ., ø ÷ Sñ c� h� i
þs� l� e� r� , ÷ D
�.ø Aò ., ø ÷ aô nõ d ü Sñ l
�iþnõ iþnõ gÿ e� r� , ÷ P� .ø J .ø
1994. Pyrrolnitrin production by biological controlagent Pseudomonas cepacia B37w in culture and incolonized wounds of potatoes. Appl. Environ.Microbiol. 60: 2031-2039.Cð
h� aô r� iþ gÿ k� aô p� aô k� o� r� nõ , ÷ Nï
., ø ÷ aô nõ d ü Sñ iþv� aô s� iþ t� h� aô mó p� aô r� aô mó , ÷ K
�.ø
1987. Changes in the composition and populationof fluorescent pseudomonads in wheat rootsinoculated with successive generations of root-pieceinoculum of the take-all fungus. Phytopathol.77:1002-1007.Cð
h� e� r� nõ iþnõ , ÷ L
ú., ø ÷ B� r� aô nõ dü i
þs� , ÷ Aò
., ø ÷ Iù s� mó aô iþ l� o� v� , ÷ Z� ., ø ÷ aô nõ d ü Cð
h� e� t� , ÷ Iù .ø1996. Pyrrolnitrin production by an Enterobacteragglomerans strain with broad-spectrum activitytowards fungal and bacterial phytopathogens. Curr.Microbiol. 32:208-212.D�
aô h� ll���
ö f�, ÷ Iù ., ø ÷ B� aô iþ l� l� iþ e� , ÷ H� ., ø ÷ aô nõ d ü K
�j!e� l� l� e� b� e� r� gÿ , ÷ Sñ .ø 2000.
rpoB-based microbial analysis avoids limitationsinherent in 16S rRNA gene intraspeciesheterogeneity. Appl. Environ. Microbiol. 66:3376-3380.D�
e � V�
o� s� , ÷ D�
., ø ÷ B� o� uû t� o� nõ , ÷ Cð
., ø ÷ Sñ aô r� nõ iþgÿ uû e� t� , ÷ A
ò., ø ÷ D�
e � V�
o� s� , ÷ P� .,ø ÷V�
aô uû t� e� r� iþ nõ , ÷ M�
., ø ÷ aô nõ d ü Cð
o� r� nõ e� l� iþ s� , ÷ P� .ø 1998. Sequencediversity of the oprI gene, coding for major outer
membrane lipoprotein I, among rRNA group Ipseumonads. J. Bacteriol. 180:6551-6556.D�
i þ
Cð
e� ll��� o� , ÷ Fî
., ø ÷ B� e� v� iþv� iþnõ o� , ÷ A
ò., ø ÷ Cð
h� iþaô r� iþ nõ i
þ, ÷ Lú
., ø ÷ Fî
aô nõ iþ, ÷ Rì .,ø ÷
P� aô ff�"� e� tt�#� iþ , ÷ D�
., ø ÷ T� aô b� aô cc��� h� iþo� nõ iþ, ÷ Sñ ., ø ÷ aô nõ d ü D
�aô l� mó aô s� t� r� iþ , ÷ C
ð.ø
1997. Biodiversity of a Burkholderia cepaciapopulation isolated from the maize rhizosphere atdifferent plant growth stages. Appl. Environ.Microbiol. 63:4485-4493.D�
uû f�f�y� , ÷ B� .ø K� ., ø ÷ aô nõ d ü D
�é$ f� aô gÿ o� , ÷ G
�.ø 1999. Environmental
factors modulating antibiotic and siderophorebiosynthesis by Pseudomonas fluorescens biocontrolstrains. Appl. Environ. Microbiol. 65:2429-2438..Eí l�-� B� aô nnõöõ aô , ÷ N
ï., ø ÷ aô nõ d ü W
�iþnõ k� e� l� mó aô nnõ õ , ÷ G
�.ø 1988.
Pyrrolnitrin from Burkholderia cepacia: antibioticactivity against fungi and novel activities againststreptomycetes. J. Appl. Microbiol. 85:69-78.Fî
e� l� s� e� nõ s� t� e� iþ nõ , ÷ J .ø 1993. PHYLIP: phylogeny inference
package, version 3.5c. Department of genetics,University of Washington, Seatle.H� aô gÿ e� dü o� r� nõ , ÷ C
ð., ø ÷ G�
o� uû l�dü , ÷ W
�.ø D� ., ø ÷ B� aô r� dü i
þnõ e� ll�%� iþ , ÷ T� .ø Rì ., ø ÷ aô nõ dü
G�
uû s� t� aô v� s� o� nõ , ÷ D�
.ø Rì .ø 1987. A selective medium forenumeration and recovery of Pseudomonas cepaciabotypes from soil. Appl. Environ. Microbiol.60:2931-2938.H� aô mmó ó e� r� , ÷ P� .ø Eí ., ø ÷ aô nõ d ü Eí v� e� nõ s� e� nõ , ÷ K
�.ø B� .ø 1993. Post
harvest control of Botrytis cinerea on cut flowerswith pyrrolnitrin. Plant Dis. 77: 283-286.H� aô mmó ó e� r� , ÷ P� .ø Eí ., ø ÷ B� uû r� dü , ÷ W
�., ø ÷ H� i
þll���
, ÷ D�
.ø Sñ ., ø ÷ Lú
iþgÿ o� nõ , ÷ J
.ø M� .,ø ÷
aô nõ d ü V�
aô n õ P� ée$�� , ÷ K�
.ø -� H� .ø 1999. Conservation of thepyrrolnitrin gene cluster among six pyrrolnitrin-producing strains. FEMS Microbiol. Lett. 180:39-44.H� aô mmó ó e� r� , ÷ P� .ø Eí ., ø ÷ H� i
þll���
, ÷ D�
.ø Sñ ., ø ÷ Lú
aô mó , ÷ Sñ .ø T� ., ø ÷ V�
aô n õ P� ée$&� , ÷ K�
.ø -�H� ., ø ÷ aô nõ d ü L
úiþgÿ o� nõ , ÷ J
.ø M� .ø 1997. Four genes from
Pseudomonas fluorescens that encode the biosynthesisof pyrrolnitrin. Appl. Environ. Microbiol. 63:2147-2154.H� aô mó p� l
�, ÷ V
�., ø ÷ P� aô v� l
�iþc� e� k� , ÷ A
ò., ø ÷ aô nõ d ü F
îl�e� gÿ r� , ÷ J
.ø 2001.
Construction and bootstrap analysis of DNAfingerprinting-based phylogenetic trees with thefreeware program FreeTree: application totrichomonad parasites. Int. J. Syst. Evol. Microbiol.51:731-735.H� aô nõ dü e� l� s� mó aô nõ , ÷ J
.ø , ÷ aô nõ d ü Sñ t� aô b� b
�, ÷ Eí .ø V� .ø 1996. Biocontrol
of soilborne plant pathogens. Plant Cell 8:1855-1869.H� e� bb
�'�aô r� , ÷ K
�.ø P� ., ø ÷ M
�aô r� t� e� l� , ÷ M
�.ø H� ., ø ÷ aô nõ d ü H� e� uû l
�iþnõ , ÷ T� . ø 1998.
Suppression of pre- and post-emergence damping-off in corn by Burkholderia cepacia. Eur. J. PlantPathol. 104:29-36.H� iþll���
, ÷ D�
.ø Sñ ., ø ÷ Sñ t� e� iþ nõ , ÷ J .ø Iù ., ø ÷ T� o� r� k� e� wý i
þt� z , ÷ N
ï.ø Rì ., ø ÷ M
�o� r� s� e� ,÷
Aò
.ø M� ., ø ÷ H� o� wý e� ll��� , ÷ Cð
.ø Rì ., ø ÷ P� aô c� h� l�aô t� k� o� , ÷ J
.ø P� ., ø ÷ B� e� c� k� e� r� , ÷ J
.ø O� .,ø ÷
aô nõ d ü Lú
iþgÿ o� nõ , ÷ J
.ø M� .ø 1994. Cloning of genes involved in
the synthesis of pyrrolnitrin from Pseudomonasfluorescens and role of pyrrolnitrin synthesis inbiological control of plant disease. Appl. Environ.Microbiol. 60:78-85.

C( h)
a* pt+-, e. r / 2: 021
P3 o4 l5y6 m7 o4 r/ p+ h
)i8 s9 m7 s 9 w: i8 t, h) i8 n ; t, h) e . p< rn=?> D
@ a* n; d
Ap< lB tC C D gE e. n; e. s9
52
HF oG mH mH aI , J YK
., L J aI nM d N SOuP zQ uP i
R, J TS .L 1989. Role of antibiotic
production in suppression of radish damping-off byseed bacterization with Pseudomonas cepacia. Ann.Phytopath. Soc. Japan 55:643-652.HF oG wT eU llV�V , J CW .L RX ., L J aI nM d N S
OtY iR pZ aI nM oG v[ i
Rc\ , J RX .L D] .L 1979. Control
of Rhizoctonia solani on cotton seedlings withPseudomonas fluorescens and with an antibioticproduced by the bacterium. Phytopathol. 69:480-482.HF oG wT eU llV�V , J CW .L RX ., L J aI nM d N S
OtY iR pZ aI nM oG v[ i
Rc\ , J RX .L D] .L 1980.
Suppression of Pythium ultimum-induced damping-off of cotton seedlings by Pseudomonas fluorescensand its antibiotic, pyoluteorin. Phytopathol.70:712-715.J^aI nM i
Rs_ iR eU wT i
Rc\ zQ , J W
`.L J^ ., L J aI nM d N RX oG i
RtY mH aI nM , J J
^.L 1988. Biological
control of blue mold and grey mold on apple andpear with Pseudomonas cepacia. Phytopathol.78:1697-1700.Ka aI nM gb , J Y
K.L W` ., L J CW aI rc lV s_ oG nM , J RX ., L J TS h
daI rc pZ eU , J W
`.L , J aI nM d N S
Oc\ hd eU lV lV ,J
Me .L Af .L 1998. Characterization of genes involved inbiosynthesis of a novel antibiotic from Burkholderiacepacia BC11 and their role in biological control ofRhizoctonia solani. Appl. Environ. Microbiol.64:3939-3947.Ka eeU&U lV , J CW ., L J S
Oc\ hd nM i
RdN eU rc , J Ug ., L J Me aI uP rc hd oG fh eU rc , J Me ., L J V
ioG iRs_ aI rc dN , J CW .,L J
Lj aI v[ iRllV�V eU , J J
^.L , J Bk uP rc gb eU rc , J Pl ., L J W
`iRrc tY hd nM eU rc , J Pl ., L J HF aI aI s_ , J D
].L , J aI nM dN
D]
ém fh aI gb oG , J Gn .L 1992. Suppression of root diseases byPseudomonas fluorescens CHA0: importance of thesecondary metabolite 2,4-diacetylphloroglucinol.Mol. Plant-Microbe Interact. 5:4-13.Ka eeU&U lV , J CW ., L J W
`eU lV lV eU rc , J D
].L Me ., L J N
oaI tY s_ c\ hd , J A
f., L J D
]ém fh aI gb oG , J Gn .,L J
CW ooGpG kq, J RX .L J^ ., L J aI nM d N TS h
doG mH aI s_ hd oG wT , J Lj .L SO .L 1996.
Conservation of the 2,4-diacetylphloroglucinolbiosynthesis locus among fluorescent Pseudomonasstrains from diverse geographic locations. Appl.Environ. Microbiol. 62:552-563.Ka iRrc nM eU rc , J S
O., L J HF aI mmHrH eU rc , J Pl .L Es ., L J HF i
RlV lV , J D
].L SO .L , J A
flV tY mH aI nnMtM , J A
f.L ,J
Fu iRs_ hd eU rc , J Iv ., L J W
`iReU s_ lV oG , J Lj .L J^ .L , J Lj aI nM aI hd aI nM , J Me ., L J V
iaI n M Pl éem U , J Ka .L -w
HF ., L J aI nM d N Lj iRgb oG nM , J J
^.L Me .L 1998. Functions encoded by
pyrrolnitrin biosynthetic genes from Pseudomonasfluorescens. J. Bacteriol. 180:1939-1943.Ka uP mH aI rc , J S
O., L J TS aI mH uP rc aI , J Ka ., L J J
^aI kq oG b
xs_ eU nM , J Iv .L Bk .L , J aI nM d N N
oeU iR , J Me .L
2001. MEGA2: Molecular Evolutionary GeneticsAnalysis software, Arizona State University, Tempe,Arizona, USA.J^aI iR nM , J RX ., L J RX i
Rv[ eU rc aI , J Me .L , J aI nM d N Lj aI kq eU , J J
^.L Af .L 1999.
Horizontal gene transfer among genomes: thecomplexity hypothesis. Proc. Natl. Acad. Sci. USA96:3801-3806.J^uP kqeU s_ , J TS .L HF ., L J aI nM d N CW aI nM tY oG rc , J CW .L RX .L 1969. Evolution of
protein molecules. In: Munro, H.N.(ed.).Mammalian protein metabolism, pages 21-132,Academic Press, New York, USA.Lj aI gb uP eU rrcyc eU , J Gn ., L J N
ooG uP rc , J S
O.L Me ., L J Me aI c\ hd eU rc eU tY , J V
i., L J S
OaI nM jzuP aI nM ,J
J^., L J D
]rc oG uP i
RnM , J Pl ., L J aI nM d N A
fmH aI rc gb eU rc , J N
o.L 2001.
Classification of rhizobia based on nodC and nifHgene analysis reveals a close phylogeneticrelationship among Phaseolus vulgaris symbionts.Microbiol. 147:981-993.Lj eU v[ y{ , J E
s., L J Gn oG uP gb h
d, J Fu .L J^ ., L J Bk eU rc lV iR nM , J Ka .L D] ., L J Gn uP i
RaI nM aI , J Pl .L W` .,L J
aI nM d N SOmH iRtY hd , J TS .L J^ .L 1992. Inhibition of Septoria tritici
and other phytopathogenic fungi and bacteria by
Pseudomonas fluorescens and its antibiotics. PlantPathol. 41:335-341.Lj iRuP , J W
`.L -w TS ., L J Me aI rc s_ hd , J TS .L Lj ., L J CW h
deU nM gb , J HF .L , J aI nM d N Fu oG rc nM eU y{ ,J
Lj .L J^ .L 1997. Characterization of microbial diversity bydetermining terminal restriction fragment lengthpolymorphisms of genes endcoding 16S rRNA.Appl. Environ. Microbiol. 63:4516-4522.Lj iRgb oG nM , J J
^.L Me ., L J HF i
RllV�V , J D
].L SO .L , J HF aI mmHrH eU rc , J Pl .L Es ., L J TS oG rc kq eU wT i
RtY zQ ,J
No
.L RX ., L J HF oG fh mH aI nnMtM , J D]
., L J Ka eU mH pZ fh , J HF .L -w SO ., L J aI nM d N Vi
aI n M Pl éem U ,JKa .L -w HF .L 2000. Natural products with antifungalactivity from Pseudomonas biocontrol bacteria. PestManag. Sci. 56:688-695.Me aI iR dN aI kq , J Bk .L Lj ., L J CW oG lV eU , J J
^.L RX .L , J Pl aI rc kq eU rc , J CW .L TS . L J
^rc .L , J Gn aI rrcyc iR tY y{ ,J
Gn .L Me ., L J Lj aI rc s_ eU nM , J No
., L J Bk iRnM gb , J Lj ., L J Lj i
RlV bx uP rc nM , J TS .L Gn .,L J
Me c\ CW aI uP gb hd
eU y{ , J Me .L J^ ., L J O|
lV s_ eU nM , J Gn .L J^ ., L J O|
v[ eU rc bx eeU&U kq , J RX .L ,JPl rc aI mH aI nM i
Rkq, J SO., L J S
Oc\ hd mH i
RdN tY , J TS .L Me ., L J TS i
ReU dN jzeU , J J
^.L Me ., L J aI nM dN
W`
oG eU s_ eU , J CW .L RX .L 1999. A new version of the RDP(ribosomal database project). Nucleic Acids Res.27:171-173.Me aI uP rc hd oG ffh"h eU rc , J Me ., L J Ka eeU&U lV , J CW ., L J S
Oc\ hd nM i
RdN eU rc , J Ug ., L J V
ioG iRs_ aI rc dN ,J
CW ., L J HF aaI?I s_ , J D]
., L J aI nM d N D]
ém fh aI gb oG , J Gn .L 1992. Influence ofenhanced antibiotic production in Pseudomonasfluorescens strain CHA0 on its disease suppressivecapacity. Phytopathol. 82:190-195.Me aI v[ rc oG dN i
R, J D]
.L Vi ., L J W`
eU llV�V eU rc , J D]
.L Me ., L J aI nM d N TS hd
oG mH aI s_ hd oG wT ,JLj .L SO .L Unpublished data.Me aI v[ rc oG dN i
R, J D]
.L Vi ., L J Ka s_ eU nM zQ eU nM kqoG , J V
i.L No ., L J Bk oG nM s_ aI llV�V , J RX .L Fu .L ,J
CW ooGpG kq, J RX .L J^ ., L J Bk oG rc oG nM i
RnM , J A
f.L Me ., L J aI nM d N TS h
doG mH aI s_ hd oG wT , J Lj .L SO .L
1998. A seven-gene locus for synthesis ofphenazine-1-carboxylic acid by Pseudomonasfluorescens 2-79. J. Bacteriol. 180:2541-2548.Me aI v[ rc oG dN i
R, J D
].L Vi ., L J Bk oG nM s_ aI llV�V , J RX .L Fu ., L J D
]eU lV aI nM eU y{ , J S
O.L Me .L ,J
SOoG uP lV eU , J Me .L J^ ., L J Pl h
diRllV�V iR pZ s_ , J Gn ., L J aI nM d N TS h
doG mH aI s_ hd oG wT , J Lj .L SO
2001. Functional analysis of genes for biosynthesisof pyocyanin and phenazine-1-carboxamide fromPseudomonas aeruginosa PAO1. J. Bacteriol.183:6454-6465.Me c\ SO pZ aI ddN}N eU nM -w Gn aI rc dN eU nM eU rc , J Bk .L Bk ., L J S
Oc\ hd rc oG eU dN eU rc , J Ka .L Lj .,L J
Ka aI llV%V oG gb eU rc , J SO.L Es ., L J RX aaI?I iR jz mH aI kq eU rc s_ , J J
^.L Me .L , J TS h
doG mH aI s_ hd oG wT ,J
Lj .L SO ., L J aI nM d N W`
eU llV�V eU rc , J D]
.L Me .L 2000. Genotypic andphenotypic diversity of phlD-containingPseudomonas strains isolated from the rhizosphereof wheat. Appl. Environ. Microbiol. 66:1939-1946.No
oG wT aI kq -w TS hd
oG mH pZ s_ oG nM , J Bk ., L J Gn oG uP lV dN , J SO.L J^ ., L J aI nM d N Lj oG pZ eU rc ,J
J^.L Es .L 1997. Identification and sequence analysis of
the genes encoding a polyketide synthase requiredfor pyoluteorin biosynthesis in Pseudomonasfluorescens Pf-5. Gene 204:17-24.No
oG wT aI kq -w TS hd
oG mH pZ s_ oG nM , J Bk ., L J CW hd
aI nM eU y{ , J No
., L J W`
iRnM gb , J J
^.L SO .L ,J
Gn oG uP lV dN , J J^.L SO ., L J aI nM d N Lj oG pZ eU rc , J J
^.L Es .L 1999. Characterization
of the pyoluteorin biosynthetic gene cluster ofPseudomonas fluorescens Pf-5. J. Bacteriol. 181:2166-2174.O|
hd
mH oG rc iR , J TS ., L J HF aI gb iRwT aI rc aI , J S
O.L -w Iv ., L J Ug eU dN aI , J A
f., L J Me i
RnM oG dN aI , J Y
K.,L J
aI nM d N YK
aI mH aI dN aI , J Ka .L 1978. Production of pyoluteorinand its derivatives from n-paraffin by Pseudomonasaeruginosa S10B2. Agr. Biol. Chem. 42:2031-2036.O|
'~ SO uP llV�V iR v[ aI nM , J D]
.L J^ ., L J aI nM d N O|
'~ Gn aI rc aI , J Fu .L 1992. Traits offluorescent Pseudomonas spp. involved insuppression of plant root pathogens. Microbiol.Rev. 56:662-676.

C� h�
a� pt�-� e� r � 2: �2�
P� o� l�y� m� o� r� p� h
�i� s� m� s � w� i� t� h� i� n � t� h� e � p� rn�?� D
� a� n� d
�p� l� t� C � g� e� n� e� s�
53
P� a� g� e� , � R� . D¡ . M¢ . 1996. TREEVIEW: An applicationto dsplay phylogenetic trees on personal computers.Comp. Applicat. Biosc. 12:357-358.P� a� r£ k¤ e� , � J
¥. L¦ ., �� a� n§ d ¨ G© uª r£ i« a� n§ -¬ S h® e� r£ m¯ a� n§ , � D¡ . 2001.
Diversity of the Burkholderia cepacia complex andimplications for risk assessment of biological controlstrains. Ann. Rev. Phytopathol. 39:225-258.P� i«c° a� r£ d¨ , � C
±., �� D¡ i
«C±
e� ll²�² o³ , � F´
. , � Vµ
e� n§ t¶ uª r£ a� , � M¢
., �� Fa� n§ i«, � R� ., %� a� n§ d
G© uª c° k¤ e� r£ t¶ , � A·
. 2000. Frequency and diversity of 2-4-diacetylphloroglucinol-producing bacteria isolatedfrom the maize rhizosphere at different stages ofgrowth. Appl. Environ. Microbiol. 66:948-955.P� i«e� r£ s¸ o³ n § III¹º¹y¹ , � L¦ . S ., �� G© a� ff»"» n§ e� y¼ , � T
½., �� L¦ a� m, � S
., �� a� n§ d ¨ G© o³ n§ g� ,�
F. 1995. Molecular analysis of genes encodindphenazine biosynthesis in the biocontrol bacteriumPseudomonas aureofaciens 30-84. FEMS Microbiol.Lett. 134:299-307.R� aa�&� i« j¾ ma� k¤ e� r£ s, � J
¥. M¢
. , � L¦ ee�&� m¯ a� n§ , � M¢
. , � Vµ
a� n § O¿ o³ r£ sc° h® o³ t¶ ,�M¢
. M¢ . P� ., �� Vµ
a� n § de� r £ Sl² uª i«s, � I¹ ., �� S
c° h® i
«ppÀ'À e� r£ s¸ , � B
Á., %� a� n§ d
BÁ
a� kk¤p¤
e� r£ , � P� . A· . HÂ . M¢ . 1995. Dose-responserelationships in biological control of Fusarium wiltof radish by Pseudomonas spp. Phytopathol.85:1075-1081.R� aa�&� i« j¾ ma� k¤ e� r£ s, � J
¥. M¢ . , � W
Ãe� ll²�² e� r£ , � D¡ . M¢ . , � a� n§ d ¨ T
½h® o³ m¯ a� s¸ h® o³ wÄ ,�
L¦ . S . 1997. Frequency of antibiotic-producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63:881-887.R� aa�&� i« j¾ ma� k¤ e� r£ s, � J
¥. M¢ . a� n§ d ¨ W
Ãe� l² l² e� r£ , � D¡ . M¢ . 1998. Natural
plant protection by 2,4-diacetylphloro-glucinol-producing Pseudo-monas spp. in take-all declinesoils. Mol. Plant-Microbe Interact. 11:144-152.R� aa�&� i« j¾ ma� k¤ e� r£ s, � J
¥. M¢ . , � a� n§ d ¨ W
Ãe� l² l² e� r£ , � D¡ . M¢ . 2001.
Exploiting genotypic diversity of 2,4-diacetylphloroglucinol-producing Pseudo-monasspp.: characterization of superior root-colonizing P.fluorescens strain Q8r1-96. Appl. Environ.Microbiol. 67:2545-2554.R� a� m¯ e� tt¶#¶ e� , � A
·., �� M
¢o³ ëÅ nn§t§ e� -¬ L¦ o³ c° c° o³ zÆ , � Y
Ç., �� a� n§ d ¨ D¡ éÈ f» a� g� o³ , � G© .
2001. Polymorphism of the polyketide synthasegene phlD in biocontrol fluorescent Pseudomonasproducing 2,4-diacetylphloroglucinol andcomparison of PhlD with plant polyketidesynthases. Mol. Plant-Microbe Interact. 14:639-652.R� i«vÉ e� r£ a� , � M
¢. C± ., %� J
¥a� i« n§ , � R� ., �� M
¢oo³p³ r£ e� , � J
¥. EÊ ., �� a� n§ d ¨ L¦ a� k¤ e� , � J
¥.
1998. Genomic evidence for two functionallydistict gene classes. Proc. Natl. Acad. Sci. USA95:6239-6244.Sa� mbË r£ oo³p³ k
¤, � J¥., �� a� n§ d ¨ R� uª ss¸Ì¸ e� l² , � D¡ . Wà . 2001. Molecular
cloning: a laboratory manual, 3rd edn. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.Sa� l² c° h® e� r£ , � O¿ . a� n§ d ¨ L¦ i
«n§ g� e� n§ s, � F. 1980. Isolation and
characterization of a mutant of Pseudomonasaureofaciens ATCC 15926 with increased capacityfor synthesis of pyrrolnitrin. J. Gen. Microbiol.118:509-513Sh® a� n§ a� h® a� n§ , � P� ., �� O¿ '
ÍSuª l² l² i« vÉ a� n§ , � D¡ . J¥ ., �� S
i«mpÀ s¸ o³ n§ , � P� ., ��
G© l² e� nn§t§ o³ n§ , � J¥. D¡ . , � a� n§ d ¨ O¿ '
ÍG© a� r£ a� , � F. 1992. Isolation of
2,4-diacetylphloroglucinol from a fluorescentpseudomonad and investigation of physiologicalparameters influencing its production. Appl.Environ. Microbiol. 58:353-358.
Sh® a� r£ i« f» i« -¬ T½ e� h® r£ a� n§ i
«, � A·
. , � ZÎ a� l² a� , � M¢
., �� NÏ
a� t¶ sc° h® , � A·
. , � M¢
o³ ëÅ nn§Ð§ e� -¬L¦ o³ c° c° o³ zÆ , � Y
Ç., %� a� n§ d ¨ D¡ éÈ f» a� g� o³ , � G© . 1998. Biocontrol of
soil-borne fungal plant diseases by 2-4-diacetylphloroglucinol-producing fluorescentpseudomonads with different restriction profiles ofamplified 16S rDNA. Eur. J. Plant Pathol.104:631-643.Si«mo³ n§ , � A
·., �� a� n§ d ¨ R� i
«d¨ g� e� , � EÊ . HÂ . 1974. The use of
ampicillin in a simplified selective medium for theisolation of fluorescent pseudomonads. J. Appl.Bacteriol. 37:459-460.St¶ o³ h® l² , � EÊ . A· ., �� M
¢i«l² n§ e� r£ , � J
¥. L¦ . , � a� n§ d ¨ HÂ a� n§ de� l² s¸ ma� n§ , � J
¥. 1999.
Zwittermicin A biosynthetic cluster. Gene 237:403-411.T½
a� bË a� cc°&° h® i«o³ n§ i
«, � S., �� Vµ
i«s¸ c° a� , � P� ., %� C
±h® i«a� r£ i« n§ i
«, � L¦ ., �� B
Áe� vÉ i«vÉ i«n§ o³ ,�
A·
., �� D¡ i «
Se� r£ i« o³ , � C
±., �� F
´a� n§ c° e� ll²�² i« , � S
. , � a� n§ d ¨ F
´a� n§ i
«, � R� . 1995.
Molecular characterization of rhizosphere andclinical isolates of Burkholderia cepacia. Res.Microbiol. 146:531-542.T½
a� bË a� cc°&° h® i«o³ n§ i
«, � S., �� C±
h® i«a� r£ i« n§ i
«, � L¦ ., �� B
Áe� vÉ i«vÉ i«n§ o³ , � A
·., �� C±
a� n§ t¶ a� l² e� ,�C±
., �� a� n§ d ¨ D¡ a� l² m¯ a� s¸ t¶ r£ i« , � C±
. 2000. Bias caused by usingdifferent isolation media for assessing the geneticdiversity of a natural microbial population. Microb.Ecol. 40:169-176.T½
a� k¤ e� da� , � R� . 1958. Pseudomonas pigments. I.Pyoluteorin, a new chlorine-containing pigmentproduced by Pseudomonas aeruginosa. Hako KogakuZasshi 36:281-290.T½
a� m¯ uª r£ a� , � KÑ . 1997. TreeExplorer for Windows,version 2.12. Tokyo Metropolitan University,Tokyo, JapanT½
a� wÄ a� r£ a� , � S., �� M¢
a� t¶ s¸ uª m¯ o³ t¶ o³ , � S., %� HÂ i
«r£ o³ s¸ e� , � T
½., �� M¢
a� t¶ suª mo³ t¶ o³ ,�YÇ
., �� NÏ
a� k¤ a� mo³ t¶ o³ , � S., �� M¢
i«t¶ suª n§ o³ , � M
¢., �� a� n§ d ¨ KÑ a� mi
«muª r£ a� , � T
½.
1989. In vitro antifungal synergism betweenpyrrolnitrin and clotrimazole. J. Med. Mycol.30:202-210.T½
a� zÆ a� wÄ a� , � J¥. , � WÃ
a� t¶ a� n§ a� bË e� , � KÑ ., �� YÇ
o³ sh® i«d¨ a� , � H ., �� S
a� t¶ o³ , � M
¢., ��
a� n§ d ¨ HÂ o³ mm¯ a� , � YÇ
. 2000. Simple method of detectionof the strains of fluorescent Pseudomonas spp.producing antibiotics, pyrrolnitrin andphloroglucinol. Soil Microorg. 54:61-67.T½
h® o³ m¯ a� sh® o³ wÄ , � L¦ . S ., �� a� n§ d ¨ WÃ
e� ll²�² e� r£ , � D¡ . M¢ . 1996. Currentconcepts in the use of introduced bacteria forbiological disease control: mechanisms andantifungal metabolites. In: Plant-microbeinteractions (Stacey, G., and Keen, N.T., Eds.),vol.1., pp.187-236. Chapman & Hall, Ltd.,London, United Kingdom.T½
h® o³ m¯ pÀ s¸ o³ n§ , � J¥. D¡ ., �� H i
«gg�Ò� i
«n§ s, � D¡ . G© ., �� a� n§ d ¨ G© i
«bË s¸ o³ n§ , � T
½. J¥ .
1994. CLUSTAL W: impoving the sensitivity ofprogressive multiple alignment through sequenceweighting, positions-specific gap penalties andweight matrix choice. Nucleic Acids Res. 22:4673-4680.Vµ
a� n§ d¨ a� mmr e� , � P� ., �� H o³ l² m¯ e� s, � BÁ
., �� Vµ
a� n§ c° a� nn§t§ e� y¼ t¶ , � M¢
., ��C±
o³ e� n§ y¼ e� , � T½
., �� H o³ s¸ t¶ e� , � BÁ
., %� C±
o³ o³ pÀ ma� n§ , � R� ., %� R� e� vÉ e� t¶ s, � H ., ��L¦ a� uª wÄ e� r£ s¸ , � S
., %� G© i
«ll²�² i« s, � M
¢. , � KÑ e� r£ st¶ e� r£ s¸ , � KÑ ., �� a� n§ d ¨ G© o³ vÉ a� n§ ,�
J¥. R� . WÃ . 1997. Occurrrence of multiple genomovars
of Burkholderia cepacia in cystic fibrosis patients andproposal of Burkholderia multivorans sp. nov. Int. J.Syst. Bacteriol. 30:225-420.

CÓ hÔ
aÕ ptÖ-× eØ r Ù 2: Ú2Û
PÜ oÝ lÞyß mà oÝ rÙ pÖ h
Ôiá sâ mà s â wã iá t× hÔ iá n ä t× hÔ e Ø på rnæ?ç D
è aÕ nä d
épå lê të C ì gí eØ nä eØ sâ
54
Vî
iï dð añ vò eó rô , õ Aö
.÷ Kø .÷ 1967. Synthetic and complex mediafor the rapid detection of fluorescence ofphytopathogenic pseudomonads: effect of thecarbon source. Appl. Microbiol. 15:1523-1524.
Wù
iï sú eó , õ Mû ., ÷�õ Sü hý iï mþ kÿ eó t� sú , õ L�
.÷ J� ., ÷%õ añ n� d ð
Mû c� Aö rô t� hý u� rô , õ J�.÷ Vî . ÷ 1995.
Genetic structure of a lotic population ofBurkholderia (Pseudomonas) cepacia. Appl. Environ.Microbiol. 61:1791-1798.

______________________________________________________
CHAPTER 3
C�
O�
N�
SE
R�
V�
A
T�
I�O�
N �
O�
F �
T�
H�
E
R�
E
SP�
O�
N�
SE
R�
E
G�
U�
L�
A
T�
O�
R �
G�
E
N�
E
g� a� c� A� I�N�
P�
S�E�
U� D
O! M"
O! N#
A�
S� S$P%
E&
C'
I�E&
S$
_______________________________________________________________________________

C( h)
a* pt+-, e. r / 3: 021 g3 a4 c5 A6 i7 n 8 Ps9;:
eu<>= d?
o@ mA o@ nB a4 s: sC p+ e. cD i7 e. sC
56
AE
BF SG TH
RI AE
C( TH
_______________________________________________________________________________
TH
h)
e . r/ e. sC p+ oJ n8 sC e . r/ e. gK uL lMa* t, oJ r / gK e. n8 e . g3 a4 c5 A6 i7 n8 f
NlMuL e. n8 cD e. s C t, h) e . p+ r/ oJ d
OuL cD t, i7 oJ n 8 oJ f
NsC e. vP e. r/ a* l M sC e. cD oJ n8 d
Oa* r/ yQ
mR e. t, a* bS oJ lMit7 , e. s C i7 n 8 b
SoJ t, h )
p+ a* t, h) oJ gK e. n8 i7 c D a* n8 d O
bS
e. n8 e. fN i7 cD i7 a* l M Ps9;:
eu<>= d?
o@ mA o@ nB a4 s: sC pp+-+ . T IU n 8 t, h) i7 s sCVC t, uL d
OyQ , wWYX e . d
Oe. vP e. lM oJ p+ e. dO
p+ r/ im7 R e. r/ s C a* n8 d O
a * p+ r/ oJ bS
e . fNoJ r / t, h) e . g3 a4 c5 A6 gK e. n8 e o.ZJ f
NPs9;:
eu<>= d?
o@ mA o@ nB a4 s: sC p+ e. cD i7 e. s C a* n8 d
OsC e. q[ uL e. n8 cD e. d
Og3 a4 c5 A6 f
Nr/ oJ m R t, e. n8
Ps9;:
eu<>= d?
o@ mA o@ nB a4 s: sC t, r/ a* i7 n8 s C i7 sC oJ l
Ma* t, e. d
OfNr/ oJ m R d
Oi7 ffN\N e. r/ e. n8 t pl,]+ M a* n8 t, -^ a* ssC_C oJ cD i7 a* t, e. d
Oe. n8 vP i7 r/ oJ n8 mR e. n8 t, sC . T PC( R I a* n8 a* lM yQ sC i7 s C a* n8 d
OSG oJ uL t, h) e. r/ n h8 )
yQ bS
r/ i7 dO iz7 a a* t, i7 oJ n 8 sC h) oJ wX e. d O
t, h) a* t , g3 a4 c5 A6 i7 s C h)
i7 gK h)
lMy Q cD oJ n8 sC e. rv/ P e. d
OwX it7 , h) i7 n 8 t, h) e g.bK e. n8 uL s C Ps
9;:eu<>= d?o@ mA o@ nB a4 s
::1
mR uL ltiM , 7 p+ l
Me . sC t, r/ a* i7 n8 s C oJ f
NdO
i7 ffNcN e. r/ e. n8 t , Ps9;:
eu<>= d?
o@ mA o@ nB a4 s :
sC p+ e. cD i7 e. s C a* ll MdM r/ e. sC p+ oJ n8 dO
e. d O
p+ oJ sC iti7 , 7 vP e. lM y Q t, o J t, h) e . p+ r/ oJ bS
e. , wWeX h)
e. r/ e. a* sCn8 o J r/ e. sC p+ oJ n8 sC e . wX a* s C oJ b
St, a* i7 n8 e. d
OfNr/ oJ m R 18 fhg oJ t, h) e. r / sC t, r/ a* i7 n8 s C r/ e. p+ r/ e. sC e. n8 t, i7 n8 g K 14 fji sC p+ e. cD i7 e. s C a* n8 d
O8 g gK e. n8 e. r/ a * oJ f
NGk r/ a* mR -^
n8 eg.lK a* ti, 7 vP e . bS
a* cD t, e. r/ i7 a * oJ t, h) e. r / t, h) a* n 8 Ps9;:
eu<>= d?
o@ mA o@ nB a4 s:. T Fm urL-/ t, h) e. r/ mR oJ r/ e. , fW N r/ oJ m R a * t, oJ t, a* l M oJ f
Na* pp+-+ r/ oJ xn im7 R a* t, e. lM y Q 550o2o2p
i7 n8 dO
i7 geKq. n8 oJ uL s C bS
a* cD t, e. r/ i7 a* l iM 7 sC oJ l
Ma* t, e. s C oJ b
St, a* i7 n8 e. d
OfNr/ oJ m tR , h) e . r/ h) i7 za oJ sC p+ h
)e. r/ e o.rJ f
NwX h)
e. a* t, , W a* ll iMdM 7 sC oJ l
Ma* t, e. s C t, h) a* t,
h)
yQ bS
r/ i7 dO i7 za e. d O
wX i7 t, h )
t, h) e . Gk a* cD A E
p+ r/ oJ bS
e . wX e. r/ e . cD lM a* ssC_C i7 fN i7 e. d O
a* s C Ps9;:
eu<>= d?
o@ mA o@ nB a4 s: sC pp+-+ . bT S y Q gK r/ oJ uL p+ -^ sC p+ e. cD i7 fN i7 c D p+ r/ im7 R e. r/ sC .T
IU sC oJ lMa* t, e. s C t, h) a* t , d
Oi7 d O
n8 oJ t , r/ e. sC p+ oJ n8 d O
t, o J t, h) e . Gk a* cD A E
p+ r/ oJ bS
e . a* n8 d O
p+ r/ i7 mR e. r/ s C wX e. r/ e . i7 dO e. n8 t, i7 fN i7 e. d O
a* s C bS
a* cD t, e. r/ i7 a* l M geKq. n8 e. r/ a*oJ t, h) e. r / t, h) a* n 8 Ps
9;:e< u= d?o@ mA o@ nB a4 s
:, W i7 n8 cD lM uL d
Oi7 n8 g K Ss tt e< nB o@ tt ru o@ pv h
wo@ mA o@ nB a4 s
:, W Cx ru yy s
:e< o@ mA o@ nB a4 s
: a* n8 d
OCx o@ mA a4 mA o@ nB a4 s
: sC pp+-+ . TT H h
)e. sC e.
r/ e. sC uL ltM , s C i7 n8 d
Oi7 cD a* t, e . t, h) a* t , g3 a4 c5 A
6cD a* n 8 b
Se . uL sC e. d
Oa* s C a * cD oJ mplR + M e. mR e. n8 t, a* r/ y Q gK e. n8 e. t, i7 c D mR a* rk/{z e. r / f
NoJ r / d
Oe. t, e. cD t, i7 oJ n 8 oJ f
NPs9;:
eu<>= d?
o@ mA o@ nB a4 s :
sC pp+-+ . T i7 n 8 e. n8 vP i7 r/ oJ n8 mR e. n8 t, a* l M sC a* mR p+ lMe. sC . T Ph
)yQ lMoJ gK e. n8 e. ti, 7 c D r/ e. lM a* ti, 7 oJ n8 sC h) i7 p+ s C i7 n8 f
Ne. rr/|/ e. d
OfNr/ oJ m tR , h) e.
n8 e. wX lMy Q sC e. q[ uL e. n8 cD e. d
Og3 a4 c5 A6 gK e. n8 e. s C a* n8 d
Ot, h) e . sC e. q[ uL e. n8 cD e. s C oJ f
Ng3 a4 c5 A6 h
)oJ mR oJ l
MogJ}K s C p+ r/ e. sC e. n8 t , i7 n 8 t, h) e . d
Oa* t, a* bS a* sC e. sC ,W
sC h) oJ wX e. d O
sC i7 x n dO
i7 sC t, i7 n8 cD t , cD lM uL sC t, e. r/ s C t, h) a* t , cD oJ rr/_/ e. sC p+ oJ n8 d O
t, o J t, h) e . fNoJ lMlMoJ wX i7 n8 g K b
Sa* cD t, e. r/ i7 a* l M gK r/ oJ uL p+ sC :1
PsC e. uL dO
oJ mR oJ n8 a* cD e. a* e. , W E~
n8 t, e. r/ oJ bS
a* cD t, e. r/ i7 a* cD e. a* e. , W AE
lMt, e. r/ oJ mR oJ n8 a* dO a* cD e. a* e. , W V
�i7 bS r/ i7 oJ n8 a* cD e. a* e. , W BF urkL-/�z h
)oJ lMdO
e. r/ i7 a* , W a* n8 dO
X�
a* n8 t, h) oJ mR oJ n8 a* sC . WTe� i7 t, h) i7 n 8 t, h) e . PsC e. uL dO
oJ mR oJ n8 a* dO a* cD e. a* e . a* n8 d O
E~
n8 t, e. r/ oJ bS
a* cD t, e. r/ i7 a* cD e. a* e. , W p+ oJ lMyQ mR oJ r/ p+ h
)i7 sC mR s C wX i7 t, h) i7 n8
g3 a4 c5 A 6
a* n8 d O
it7 , s C h)
oJ mR oJ lMogJ}K s C a* llMdM oJ wX e. d
Oi7 dO e. n8 ti, 7 fN i7 cD a* t, i7 oJ n 8 oJ f
NsC i7 x n a* n8 d
OfNi7 vP e . sC uL bc
S D lM uL sC t, e. r/ sC , W r/ e. sC p+ e. cD t, i7 vP e. lM yQ .TCo( J mpR + a* r/ i7 sC oJ n 8 oJ f
Nt, h) e . g3 a4 c5 A
6gK e. n8 e . a* n8 d
OGk a* cD A
Ep+ r/ oJ t, e. i7 n8 -b^ S a* sC e. d
Ot, r/ ee.l. s C wX i7 t, h
)t, h) e . t, r/ ee .l. i7 n8 f
Ne. rr/|/ e. d
OfNr/ oJ m R 16fh� SG
r/ D� NA s� E C e. q[ uL e. n8 cD e. s C yQ i7 e. lM dO e. d
Oa * sC im7 R i7 lM a* r / oJ vP e. r/ a* ll MdM cD lM uL sC t, e. r/ i7 n8 gK . T T
Hh)
e. sC e . r/ e. sC uL ltM , s sC�C uL gK gK e. sC t , t, h) a* t , g3 a4 c5 A6 a* n8 d
Oit7 , sC
h)
oJ mR oJ lMoJ gK s C mR a* y Q p+ r/ oJ vP i7 dO e . cD oJ mR pl+ M e. mR e. n8 t, a* r/ y Q gK e. n8 e. t, i7 c D mR a* rk/�z e. r/ s C f
NoJ r / p+ h
)yQ lMoJ gK e. n8 e. ti, 7 c D sC t, uL d
Oi7 e. s C oJ f
NPs9;:
eu<>= d?
o@ mA o@ nB a4 s :
sC pp+�+ . T a* n8 d O
Gk r/ a* mR -^ n8 eg.lK a* ti, 7 vP e . bS
a* cD t, e. r/ i7 a * oJ t, h) e. r / t, h) a* n 8 Ps9;:
eu<>= d?
o@ mA o@ nB a4 s:. T E~
sC tim, 7 R a* t, i7 oJ n8 s C oJ fN
n8 oJ n8 sC yQ n8 oJ n8 yQ mR oJ uL s C t, o J sC yQ n8 oJ n8 yQ mR oJ uL s sC�C uL bS
sC tit, 7 , uL ti, 7 oJ n 8 r/ a* t, e. s C (� K� a* /� K� s C r/ a* t, i7 oJ sC ) � sC h) oJ wX e. d O
t, h) a* t, , fW N oJ r / mR e. mR bS
e. r/ s C oJ fN
bS
oJ t, h) , W t, h) e . PsC e. uL dO
oJ mR oJ n8 a* cD e. a* e . a* n8 d O
E~
n8 t, e. r/ oJ bS
a* cD t, e. r/ i7 a* cD e. a* e. , W p+ urL-/ i7 fN yQ i7 n8 g K sC e. lM e. cD t, i7 oJ n 8 i7 s C a* cD ti, 7 n8 g oK�J n 8 g3 a4 c5 A6 ,Wi7 n8 dO
i7 cD a* t, i7 n8 g K t, h) a* t , t, h) e. r/ e . i7 s sC�C e. lM e. cD t, i7 vP e . p+ r/ e. ssC�C urL-/ e . t, o J a* vP oJ i7 d O
sC uL bS
sC tit, 7 , uL t, i7 oJ n8 s C lMe. a* dO i7 n8 g K t, o J f
NuL n8 cD ti, 7 oJ n8 a* l M cD h) a* n8 geKq. s C i7 n8
t, h) e . Gk a* cD A E
p+ r/ oJ t, e. i7 n8 .T_________________________________________________________________________
J�oJ r/ ge Kq. T.
H T dO
e . SG oJ uL za a* , W M�
a* rk M/{z � a* zza�a oJ lMa* , W a* n8 d
OJ�oJ s C M
�. T RI aa*l* i7 j� mR a* kz e. r/ sC
Submitted.

C� h�
a� pt�-� e� r � 3: �2� g� a� c� A i¡ n ¢ Ps£;¤
eu¥>¦ d§
om© onª a� s¤ s« p� e� c¬ i¡ e� s«
57
I N® T¯
RO°²± D³ U´
C� T¯
I O± N®
Bacteria of the genus Pseudomonas are able to survive and prosper in a wide range of
environmental conditions. This genus not only contains plant, animal, and human pathogenic
species, but also accomodates species that are of significant environmental importance,
including plant growth promoters, xenobiotic degraders, and biocontrol agents (Palleroni,
1992; Johnsen et al., 1996; O’Sullivan and O’Gara, 1992). Their versatile metabolic activities
and ability to produce a wide variety of secondary metabolites have stimulated numerous
ecological, molecular and biochemical studies.
In several strains that belong to the group of plant-associated Pseudomonas species,
expression of genes involved in the biosynthesis of secondary metabolites and extracellular
enzymes is positively controlled by the GacS/GacA two-component system (reviewed by Heeb
and Haas, 2001). This regulatory system consists of a membrane-bound sensor kinase protein
(GacS) and a cytoplasmic response regulator protein (GacA). The current model proposes that
GacS recognizes specific environmental stimuli and activates GacA which in turn triggers the
expression of specific genes (Appleby et al., 1996; Pernestig et al., 2001; Heeb and Haas,
2001). Several other elements are also involved in the regulation of certain genes, forming a
complex regulatory cascade, with the GacS/GacA pair controlling secondary metabolite
production at a higher hierarchy (Sarniguet et al., 1995; Chancey et al., 1999; Whistler et al.,
1998; Blumer and Haas, 2000). For example, production of phenazine antibiotics, which are
involved in the biocontrol activity and ecological competence of several Pseudomonas strains
(Mazzola et al., 1992), is partially regulated by the PhzI/PhzR quorum sensing system that
uses N-acyl-homoserine lactone (Pierson III et al., 1994). The GacS/GacA system interacts
positively with the PhzI/PhzR quorum sensing system by regulating the synthesis of N-acyl-
homoserine lactone thereby controlling phenazine production at the transcriptional level
(Chancey et al., 1999).
The sensor kinase gene gacS was first described in Pseudomonas syringae pv. syringae as
an essential factor for disease manifestation (Hrabak and Willis, 1992). Since then, gacS
homologs have been identified in other Pseudomonas strains, Azotobacter vinelandii, Vibrio
cholerae, and several enteric bacteria, including E. coli, Salmonella enterica serovar
typhimurium, and Erwinia carotovora subsp. carotovora (Heeb and Haas, 2001). Alignment of
GacS homologs resulted in three groups: the pseudomonads, the enteric bacteria, and
Vibrio/Shewanella (Heeb and Haas, 2001). Interestingly, within a total of nine Pseudomonas
strains included in their analyses, P. fluorescens, P. chlororaphis, P. tolaasii, P. viridiflava and P.

Cµ h¶
a· pt¸-¹ eº r » 3: ¼2½ g¾ a¿ cÀ AÁ i n à PsÄ;Å
euÆ>Ç dÈ
oÉ mÊ oÉ nË a¿ sÅ sÌ p¸ eº cÍ i eº sÌ
58
syringae belonged to a subgroup, whereas GacS homologs from P. aeruginosa and P. putida
clustered at some distance (Heeb and Haas, 2001).
The response regulator gene gacA (global activator of antibiotic and cyanide
production) was first identified in the biocontrol strain P. fluorescens CHA0 and was shown to
be essential for the biocontrol activity and ecological fitness of this strain (Laville et al., 1992;
Natsch et al. 1994). Homologs of GacA have been identified in several other Gram-negative
bacterial genera. Within the group of Pseudomonas spp., Heeb and Haas (2001) described
GacA homologs in a total of eight Pseudomonas strains, including P. aeruginosa, P. viridiflava,
P. syringae pv. syringae, P. fluorescens, and P. aureofaciens. Subsequent alignment of the GacA
proteins placed these eight Pseudomonas strains at some distance from GacA homologs of
enteric bacteria, and Xylella and Shewanella. The results from these in silico analyses suggested
that the gacA gene may be conserved within Pseudomonadaceae. However, most information
on gacS/gacA regulatory systems stems from studies on model strains and little information is
available on the distribution of these genes in populations of rhizosphere-associated bacteria.
In this study, we developed primers and a probe for the gacA gene of Pseudomonas
species and sequenced the gacA gene from 10 Pseudomonas strains isolated from different
plant-associated environments. The newly sequenced gacA genes as well as gacA homologs
currently present in databases were used in phylogenetic analyses. Delineation of Pseudomonas
species based on gacA gene and protein sequences was compared to classification based on 16S
rDNA sequences. Sequence data were also used to compare the rate of evolution of the gacA
gene as well as the ratio of synonymous and nonsynonymous substitutions in gacA sequences
within and between Pseudomonaceae and Enterobacteriaceae. The potential usefulness of the
gacA gene as a complementary genetic marker for Pseudomonas was evaluated on bacterial
populations isolated from the rhizosphere of wheat. Approximately 550 isolates were
genotypically and phenotypically identified, and characterized for the production of specific
extracellular enzymes and secondary metabolites known to be regulated by GacA.
RÎ EÏ
SÐ UÑ
LÒ TÓ
SÐ
DÔ eº sÌ i gÕ n à a· nà d Ö
eº v× a· lØ uÙ a· ti¹  oÚ n à oÚ f Û
t¹ h¶ e º GÜ AÝ
Cµ A Ý
p¸ r» imÂ Þ eº r» s Ì a· nà d Ö
p¸ r» oÚ bß
eº .à Primers GACA1 (5’-
GBATCGGMGGYCTBGARGC) and GACA2 (5’-
MGYCARYTCVACRTCRCTGSTGAT) were developed from conserved sequences within
the gacA genes of P. fluorescens strains BL915 (GenBank L29642), CHA0 (GenBank

Cá hâ
aã ptä-å eæ r ç 3: è2é gê aë cì Aí iî n ï Psð;ñ
euò>ó dô
oõ mö oõ n÷ aë sñ sø pä eæ cù iî eæ sø
59
M80913), Pf-5 (GenBank U30858), P. aeruginosa PAO1 (GenBank U27988), P. syringae pv.
syringae B728a (GenBank U09767), P. chlororaphis 30-84 (GenBank AF115381), and the
repB gene of P. viridiflava PJ-08-6A (GenBank L30102). Primers GACA1 and GACA2 are
complementary to positions 489 and 914, respectively, in the gacA sequence of P. fluorescens
strain CHA0. Primers GACA1 and GACA2 amplified the predicted 425-bp fragment of
homologous strain P. fluorescens CHA0. Restriction analysis of the amplified fragment with
enzymes AvaII, AluI, FspI, HinfI, NaeI, and StyI resulted in restriction fragments with sizes
identical to those deduced from the restriction map of the gacA sequence of CHA0. The
GACA primers amplified a 425-bp fragment from all other Pseudomonas strains, representing
different species, except from P. agarici DSM11810, and P. fluorescens strains 2-79RN10 and
SS101 (Table 1). The PCR products obtained from all positive strains hybridized under high
stringent conditions with the GacA probe consisting of the labeled 425-bp PCR fragment
obtained from strain CHA0. In Southern hybridization with EcoRI-digested genomic DNA,
all Pseudomonas strains tested, including DSM11810, 2-79RN10 and SS101, hybridized
under high-stringent conditions with the GacA probe. A single hybridized band was detected
for all strains. No PCR products or hybridization signals were obtained from 18 other strains,
representing 14 species and 8 genera of Gram-negative bacteria other than Pseudomonas (Table
1).
Sú eæ qû uü eæ nï cù e æ aã nï aã lý yþ sø iî s ø oÿ f �
gê aë cì A í
aã nï d �
16��� Sú -� rç D� NA��
. Ten Pseudomonas strains were selected for
sequencing of the gacA gene. The ten strains were P. fluorescens strains Q2-87, SSB17, F113,
and SSB33; P. chlororaphis LMD 82.53 and PHZ26, Pseudomonas sp. PHZ13; P. putida
WCS358R; P. aeruginosa DSM 1128, and P. syringae pv. pisi 24.7. The overall base
composition of the newly obtained gacA sequences was T = 20.4% (± standard deviation 0.5),
C = 31.3% (± 0.6), A = 21.0% (± 0.3), and G = 27.4% (± 0.4). The identity of the new gacA
sequences to the gacA sequence from strain CHA0 ranged from 79.6 to 89.2% and the
similarity of the predicted GacA protein ranged from 85.9 to 98.5%. The 16S rDNA
sequences were determined over a total of approximately 300 - 600 bp to verify or identify the
following strains: P. fluorescens Pf-5, SSB33, and SSB17, P. aeruginosa DSM 1128, P.
chlororaphis LMD 82.53, and PHZ26, Pseudomonas sp. PHZ13, P. putida WCS358R, and P.
syringae pv. pisi 24.7. The sequence identity of the 16S rDNA in relation to that of strain
CHA0 was 64.2% (399 nucleotides overlap) for strain SSB17, 73.5% (328 nucleotides
overlap) for PHZ13, 81.5% (336 nucleotides overlap) for PHZ26, 82.3% (322 nucleotides
overlap) for WCS358R, 83.3% (270 nucleotides overlap) for LMD 82.53, 87.3% (378
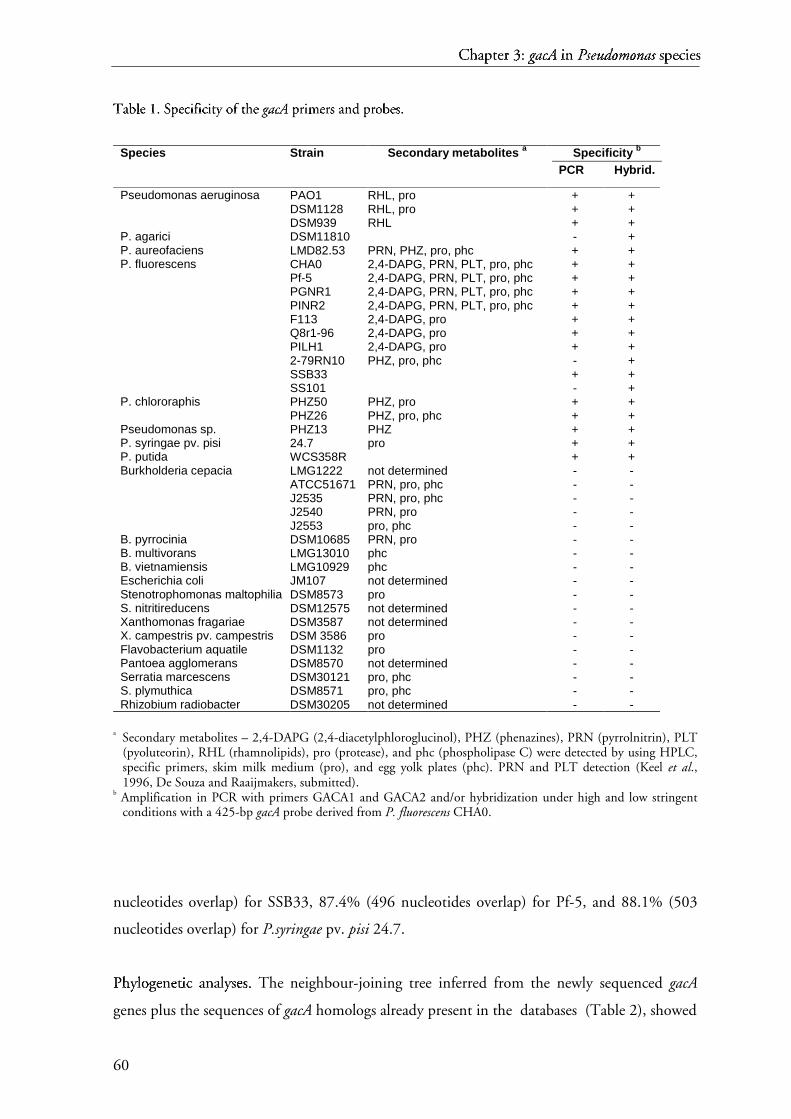
C� h�
a pt��� e� r � 3: ��� g� a� c� A� i� n � Ps���
eu�! d"
o# m$ o# n% a� s� s& p� e� c' i� e� s&
60
T(
a) b* l+e , 1. -/. S0 p1 e, c2 i3 f4 i3 c2 i3 t5 y 6 o7 f
4t5 h8 e , g9 a: c; A< p1 r= i3 m> e, r= s ? a) n@ d
Ap1 r= o7 b
*e, s? ..
Species Strain Secondary metabolites a Specificity b
PCR Hybrid.
Pseudomonas aeruginosa PAO1 RHL, pro + +DSM1128 RHL, pro + +DSM939 RHL + +
P. agarici DSM11810 - +P. aureofaciens LMD82.53 PRN, PHZ, pro, phc + +P. fluorescens CHA0 2,4-DAPG, PRN, PLT, pro, phc + +
Pf-5 2,4-DAPG, PRN, PLT, pro, phc + +PGNR1 2,4-DAPG, PRN, PLT, pro, phc + +PINR2 2,4-DAPG, PRN, PLT, pro, phc + +F113 2,4-DAPG, pro + +Q8r1-96 2,4-DAPG, pro + +PILH1 2,4-DAPG, pro + +2-79RN10 PHZ, pro, phc - +SSB33 + +SS101 - +
P. chlororaphis PHZ50 PHZ, pro + +PHZ26 PHZ, pro, phc + +
Pseudomonas sp. PHZ13 PHZ + +P. syringae pv. pisi 24.7 pro + +P. putida WCS358R + +Burkholderia cepacia LMG1222 not determined - -
ATCC51671 PRN, pro, phc - -J2535 PRN, pro, phc - -J2540 PRN, pro - -J2553 pro, phc - -
B. pyrrocinia DSM10685 PRN, pro - -B. multivorans LMG13010 phc - -B. vietnamiensis LMG10929 phc - -Escherichia coli JM107 not determined - -Stenotrophomonas maltophilia DSM8573 pro - -S. nitritireducens DSM12575 not determined - -Xanthomonas fragariae DSM3587 not determined - -X. campestris pv. campestris DSM 3586 pro - -Flavobacterium aquatile DSM1132 pro - -Pantoea agglomerans DSM8570 not determined - -Serratia marcescens DSM30121 pro, phc - -S. plymuthica DSM8571 pro, phc - -Rhizobium radiobacter DSM30205 not determined - -
a Secondary metabolites – 2,4-DAPG (2,4-diacetylphloroglucinol), PHZ (phenazines), PRN (pyrrolnitrin), PLT(pyoluteorin), RHL (rhamnolipids), pro (protease), and phc (phospholipase C) were detected by using HPLC,specific primers, skim milk medium (pro), and egg yolk plates (phc). PRN and PLT detection (Keel et al.,1996, De Souza and Raaijmakers, submitted).
b Amplification in PCR with primers GACA1 and GACA2 and/or hybridization under high and low stringentconditions with a 425-bp gacA probe derived from P. fluorescens CHA0.
nucleotides overlap) for SSB33, 87.4% (496 nucleotides overlap) for Pf-5, and 88.1% (503
nucleotides overlap) for P.syringae pv. pisi 24.7.
PB h�
yC lDoE gF e� n� e� t� i� c ' a n� a lD yC s& e� s& . G The neighbour-joining tree inferred from the newly sequenced gacA
genes plus the sequences of gacA homologs already present in the databases (Table 2), showed

CH hI
aJ ptK�L eM r N 3: O�P gQ aR cS AT iU n V PsW�X
euY!Z d[
o\ m] o\ naR sX s_ pK eM c` iU eM s_
61
Ta
ab bc lde e 2. fhg
Ai
cj cj ee sk sk il om n nnon up mq bc
ee rr s k om f s
gt au cv Aw ab nn d x
16y{z S |
rr D} N~
A i
sk ee q� up ee nn cj ee sk .g
Species Strain Accession number a
GacA 16S rDNA
PseudomonaceaeAzotobacter vinelandii ATCC 9046 AF382827 -Pseudomonas sp. PHZ13 AF502416 AF495869Pseudomonas aeruginosa PAO1 U27988 AE004091
DSM1128 AF502413 AF502423P. aureofaciens 30-84 AF115381 -P. chlororaphis LMD82.53 AF502415 AF495868
PHZ26 AF502417 AF502424P. fluorescens CHA0 M80913 AJ278812
Pf-5 AF065156 AF495871F113 AF502414 AJ278814Q2-87 AF502419 AJ278813SSB33 AF502420 AF502427BL915 L29642 -SSB17 AF502421 AF502426PfO-1 unfinished unfinishedFPT9601 AB054363 -
P. putida KT2420 unfinished unfinishedWCS358R AF502422 AF502428
P. syringae pv. pisi 24.7 AF502418 AF502425P. syringae pv. syringae B728a U09767 -P.syringae pv. tomato DC3000 unfinished unfinishedP. viridiflava PJ-08-6A L30102 -EnterobacteriaceaeEscherichia coli K-12 MG1655 M24615 AE000129
RIMD0509952 AP002559 AP002566O157 EDL933 AE005414 AE005195CFT073 unfinished unfinished
Klebsiella pneumoniae M6H 78578 unfinished unfinishedPectobacterium caratovorum SSC3193 X95564 -Salmonella typhi CT18 AL627272 AL627282
TY2 unfinished unfinishedS. typhimurium LT2 AE008786 AE008706
ATCC 14028 U67869 -Shigella flexneri 2a unfinished unfinishedYersinia pestis CO-92 AJ414150 AJ414156
KIM5 P12 unfinished unfinishedY. enterocolitica 0:8 unfinished unfinishedAlteromonadaceaeShewanella putrefasciens MR-1 unfinished unfinishedVibrionaceaeVibrio cholerae O395 AF071105 -
N16961 AE004201 AE004157Xanthomonas groupXylella fastidiosa 9a5c AE004068 AE003870
Ann-1 unfinished unfinishedDixon unfinished unfinishedTemecula 1 unfinished unfinished
Burkholderia groupBurkholderia cepacia J2315 unfinished unfinished
a Unfinished refers to non-annotated data obtained from genome sequencing projects. (-) refers to absence of dataon these strains in the databases.

C
�
h
�
a �pt� �e�r � 3:
��� g �a �c �A� i�n � Ps
��� eu�� d
�
o�m�o�n�a �s� s�p�e�c�i�e�s�
62
F
�
i
�
g �u r ¡e ¢ 1.
£¥¤ P
¦
h
§
y ¨l©oªg �e¢n«e¢t ¬i�c t¬r ¡ee¢ ¢s ® i�n«f ¯e¢r¡r¡e¢d
°
b
±
y ¨ t¬h§e ¢ n«e¢i�g �h§b±oªu r ¡-²j³oªi�n«i�n«g � m´e¢t¬h§oªd °
w µi�t¬h §
s®e¢q ¶u e¢n«ce¢s ® oªf ¯ g ·a ¸c ¹Aº (»A ¼)½, ¾ g �a ¿cA
¼
(
»
B
À
)
½
, ¾ a ¿n«d
°
16
£ Á
S
Â
r ¡DÃNÄA
¼
(
»
C
Å
)
½
. A: Based on 406-bp aligned gacAnucleotides (positions 501-906 of the gacA sequence of P. fluorescens CHA0). B: Based on 135 amino acids of GacA (positions 63-197 of gacA sequenceof P. fluorescens CHA0). C: Based on 270-bp aligned 16S rDNA sequences (positions 1-270 of the nucleotide sequence of P. fluorescens CHA0).Bootstrap values greater than 70% are shown.
Pseudom onas sp. PHZ13P. c lororaphis LM D 82.53
P. fluorescens SSB33P. fluorescens F113
P. chlororaph is PHZ26P. fluorescens Q2-87
P. fluorescens PfO-1P. putida KT2440
P. putida W CS358
P. syringae pv. pisi 24.7
P. fluorescens CHA0P. fluorescens Pf-5
P. aeruginosa PAO1P. aeruginosa DSM 1128
Shewanella pu trefasciens M R-1 Vibrio cholerae N16961
Y. pestis CO 92
Y. enteroco litica 0:8K. pneum oniae M 6H78578
S. typhi TY2S. typhi CT18
Salmonella typhim urium LT2E. coli FT073
E. coli K-12
Xylella fastidiosa 9a5c
Shigella flexneri 2aE. coli O:157
X. fastidiosa D ixon
X. fastidiosa Ann-1X. fastidiosa Tem ecula 1
E. coli RIM D
B. cepacia J2315
I
II
III
V I
V
100
99
76
93
99
100
7497
74
91
94
86
0.05
C
72
100
99
IV
100
83
77
87
87
9799
P. aureofaciens 30-84
P. chlororaphis LMD 82.53
P. fluorescens BL915P. fluorescens CHA0P. fluorescens Pf-5
P. fluorescens PfO-1
Pseudom onas sp.PHZ13P. chlororaphis PHZ26
P. fluorescens Q 2-87
P. fluorescens FPT9601
P. fluorescens SSB17P. fluorescens F113
P. fluorescens SSB33
P. virid iflava
P. syringae pv. tomato DC3000
P. putida KT2440
P. syringae pv. syringae B728aP. syringae pv. pisi 24 .7
P. putida W CS358R
P. aeruginosa DSM 1128P. aeruginosa PAO1A. v inelandii ATCC 9046
Shewanella putrefasciens M R-1Vibrio cholerae O 395
Vibrio cholerae N16961
Yersinia pestis CO-92Yersinia pestis KIM 5
Y. enterocolitica 0:8
100
P. caratovorum SSC3193K. pneumoniae M 6H 78578
S. typhim urium LT2
E. coli O :157
E. coli K-12E. coli CFT073
E. coli RIM D
B. cep. J2315
S. typhi TY2Salmonella typhi CT18
S. typhim urium ATCC 14028
X. fast. DixonX. fast. 9a5c
X. fast. ann-1X. fast. Tem . 1
100
100
91
100
96 90
96
100
87
8788
97
7893
99
96
71
II
III
IV
VI
V
B
0.1
P. aureo faciens 30-84
P. fluorescens B L915P. fluorescens C HA 0P. fluorescens Pf-5
P. fluorescens PfO -1P. fluorescens Q 2-87
P. fluorescens FPT9601P. fluorescens SSB 17P. fluorescens F113P. fluorescens SSB 33
P. v iridiflavaP. syringae pv. tom ato D C3000
P. putida K T2440
P. syringae pv. syringae B728aP. syringae pv. pis i 24 .7
P. putida W CS358RP. aerug inosa D SM 1128P. aerug inosa PAO 1
A . vinelandii ATCC 9046
Shew anella putrefasciens M R-1Vibrio cholerae O 395
Vibrio cholerae N 16961
Yersin ia pestis C O -92Yersin ia pestis K IM 5
Y. enterocolitica 0:8P. caratovorum SSC3193
K . pneum oniae M 6H 78578
S. typhim urium LT2
E. coli O :157
E. coli K -12E. coli C FT073
X. fast. D ixon
E. coli R IM D
X. fast. 9a5cB . cep . J2315
S. typhi TY2Salm onella typhi C T18
S. typhim urium ATCC 14028
X. fast. ann-1X. fast. Tem . 1
98
78
99
100
84100
100
100
100
100
100
100
100
99
99
99
99
98
80
91
70
86
82
87
93
91
910 .1
Pseudom onas sp.PH Z13P. chlo roraphis LM D 82.53
P. chlo roraphis PHZ26
II
III
IV
VI
A
98
100
V
CÆh Ça Èpt É�Êe Ër Ì 3: Í�Î g Ïa ÐcÑ A Ò
iÓ n Ô Ps Õ�Öeu×!Ø d ÙoÚ mÛoÚ nÜ a Ðs Ö s Ýp Ée Ëc ÞiÓ e Ës Ý

Cß hà
aá ptâ�ã eä r å 3: æ�ç gè aé cê Aë iì n í Psî�ï
euð!ñ dò
oó mô oó nõ aé sï sö p÷ eø cù iì eø sö
63
six distinct clusters that correspond to the following bacterial groups: Pseudomonaceae (I),
Enterobacteriaceae (II), Alteromonadaceae (III), Vibrionaceae (IV), Burkholderia (V), and
Xanthomonas (VI) groups (Fig. 1A). Within the Pseudomonadaceae, six subclusters can be
distinguished based on significant bootstrap values (>80%): the first contained P. chlororaphis,
P. aureofaciens, and P. fluorescens CHA0, Pf-5, and BL915; the second subcluster contained
only P. fluorescens strains; the third contained P. viridiflava and P. syringae strains; the fourth
was composed of two P. putida strains, the fifth of P. aeruginosa strains, and the sixth of
Azotobacter vinelandii. Within the enteric bacteria, five subclusters were distinguished: the first
one comprised E. coli isolates and Shigella flexneri, which were tightly clustered; the second
contained only Salmonella species; the third subcluster contained Klebsiella pneumoniae; the
fourth, the plant pathogenic Pectobacterium caratovorum, and the fifth contained Yersinia
species (Fig 1A). Other major clusters contained the Vibrionaceae, represented by Vibrio
(cluster III), Alteromonadaceae, represented by Shewanella putrefaciens (cluster IV), the
Burkholderia group, represented by B. cepacia (V), and the Xanthomonas group, represented
by Xylella fastidiosa isolates (cluster VI). Cluster VI was the most distant of all clusters obtained
(Fig. 1A). In all cases, with exception of clusters V and VI, the bootstrap values were highly
significant. Similar results were obtained with the neighbour-joining tree inferred from the
predicted GacA protein sequences (Fig. 1B), although the length of the branches defining
clusters were shorter than those in the tree inferred from the gacA gene sequences. Another
difference is that B. cepacia was clustered as the most distant group, whereas Xylella is the most
distant group in the tree inferred from DNA sequences. Comparison of the gacA gene and
GacA protein-based trees with the tree inferred from 16S rDNA sequences yields a similar
overall clustering (Fig. 1C), although some incongruences were noted. P. fluorescens SSB17
clustered closer to P. chlororaphis and P. aureofaciens than to other P. fluorescens strains.
Furthermore, P. fluorescens strains CHA0 and Pf-5 were clustered closer to P. aeruginosa. The
16S rDNA clustering of B. cepacia was in agreement with the clustering based on gacA gene
sequences (Fig. 1A). Phylogenetic analyses of the gacA gene, predicted GacA protein and 16S
rDNA sequences with the maximum likelihood method yielded similar results as obtained by
neighbour-joining.
Rú aû tü e oýÿþ f �
eý v� oþ l�u� tü i� oþ n � oþ f
�g� a� c� A . The measure of nonsynonymous relative to synonymous
substitution rates (Ka/Ks) provides insight into the level of selective constraint acting on
proteins (Li et al., 1985). Protein-encoding genes evolving at a neutral rate have Ka/Ks ratios
equal to 1, indicating that the protein evolves without constraint on amino acid replacements.

C� h�
a pt��� e� r � 3: ��� g� a� c� A� i� n � Ps���
eu�! d"
o# m$ o# n% a� s� s& p� e� c' i� e� s&
64
When Ka/Ks < 1, the protein is under selective constraints and protein-encoding genes are
subjected to purifying selection. Finally, when Ka/Ks > 1, positive selection has acted to
promote amino acid replacements (Hughes and Nei, 1988; Li et al., 1985). The Ka/Ks ratio
for gacA genes was determined for different categories and was shown always to be smaller
than 1 (Table 3). Fisher’s exact test (P=0.05) and Z-test confirmed that purifying selection is
operating on the gacA genes of both Pseudomonadaceae and Enterobacteriaceae.
Tajima’s relative rate test was performed to test the molecular clock hypothesis.
According to this hypothesis, sequences evolve at the same rate, irrespective of the substitution
model and whether or not the substitution rate varies with site (Tajima, 1993). The test was
done in 45 pairwise combinations with a third gacA sequence as outgroup. Prior to this
analysis, majority sequences of gacA of species belonging to the Pseudomonaceae and
Enterobacteriaceae were computed to reduce the number of two-by-two combinations.
Tajima’s test showed that there were no statistically significant differences in every possible
pairwise comparison between members of the Pseudomonaceae, with K. pneumoniae as
outgroup (data not shown). Comparisons of members of the Pseudomonaceae with members
of the Enterobacteriaceae, with B. cepacia as outgroup, showed that 13 out of 24 possible
combinations gave statistically different evolutionary rates (Table 4). In all these 13 statistically
different combinations, members of the Enterobacteriaceae presented significantly higher
substitution rates than members of the Pseudomonaceae (i.e. m2 > m1). Within the
Enterobacteriaceae, with A. vinelandii as outgroup, 5 out of 10 possible pairwise comparisons
showed significantly different substitution rates (Table 4). Collectively, these results indicate
that all tested gacA sequences from Pseudomonadaceae evolve at the same rate. In contrast,
T(
a) b* l+e , 3. -/.
N0
o1 n2 s3 y4 n2 o1 n2 y4 m5 o1 u6 s 3 (7 K8 a) ) 9 a) n2 d :
s3 y4 n2 o1 n2 y4 m5 o1 u6 s 3 (7 K8 s3 ) 9 s3 u6 b*
s3 t; i< t; u6 t; i< o1 n 2 r= a) t; e, s 3 i<n 2 g> a? c@ AA s3 e, qB u6 e, n2 cC e, s3 ..
Categories a N b Ka Ks Ka/Ks
All sequences 44 0.304 ± 0.017 0.437 ± 0.029 0.696Pseudomonaceae and Enterobacteriaceae 36 0.165 ± 0.017 0.643 ± 0.018 0.257Pseudomonaceae 22 0.034 ± 0.001 0.519 ± 0.009 0.063Enterobacteriaceae 14 0.055 ± 0.004 0.582 ± 0.029 0.074
a Comparisons of the gacA sequences in different categories. Categories of the gacA gene and its homologs were:1) all gacA sequences together; 2) gacA sequences from the Pseudomonadaceae and Enterobacteriaceae; 3) onlygacA sequences from Pseudomonadaceae; and 4) only gacA sequences from Enterobacteriaceae.
b Number of sequences per category.

CD hE
aF ptG�H eI r J 3: K�L gM aN cO AP iQ n R PsS�T
euU!V dW
oX mY oX nZ aN sT s[ pG eI c\ iQ eI s[
65
T]
a^ b_ l`e a 4. b/c
T]
a^ jd ie mf a^ ’g s h 1i D j
rk ea l` a^ tl ie vm e a rk a^ tl e a tl ea sh t l b_
ea tl wn eeaoa n p pq a^ ie rk s h or f s
b_
a^ ct tl ea rk ie a^ l ` sh pq ea ct ie ea sh .c
Species 1 a Species 2 a Outgoup m1 m2 X2 b
Pseudomonadaceae X Enterobacteriaceae
P. syringae P. caratovora B. cepacia 41 62 4.28 *
P. syringae Yersinia spp. B. cepacia 43 66 4.85 *
P. fluorescens P. caratovora B. cepacia 43 66 4.85 *
P. fluorescens Yersinia spp. B. cepacia 41 68 6.69 **
P. putida P. caratovora B. cepacia 52 75 4.17 *
P. putida Yersinia spp. B. cepacia 48 74 5.54 *
P. aeruginosa P. caratovora B. cepacia 41 66 5.84 *
P. aeruginosa Yersinia spp. B. cepacia 42 69 6.57 **
P. aeruginosa E. coli B. cepacia 41 62 4.28 *
P. aeruginosa Salmonella spp. B. cepacia 37 58 4.64 *
A. vinelandii P. caratovora B. cepacia 44 69 5.53 *
A. vinelandii Yersinia spp. B. cepacia 43 70 6.45 *
A. vinelandii E. coli B. cepacia 38 59 4.55 *
Enterobacteriaceae X Enterobacteriaceae
P. caratovora Salmonella spp. A. vinelandii 41 21 6.45 **
P. caratovora K. pneumoniae A. vinelandii 44 26 4.63 *
Yersinia spp. E. coli A. vinelandii 40 19 7.47 **
Yersinia spp. Salmonella spp. A. vinelandii 46 19 11.22 **
Yersinia spp. K. pneumoniae A. vinelandii 42 17 10.59 ***
a Majority sequences for the gacA gene were obtained for the compared bacterial species followed by spp.b Significant at 5% (*), 1% (**), and 0.01% (***).
approximately 50% of the comparisons within the Enterobacteriaceae and between
Enterobacteriaceae and Pseudomonaceae gave significantly different substitution rates,
suggesting higher evolutionary rates for the gacA gene in the Enterobacteriaceae.
Eu
vv aF lw ux aF tiH Q oy n R oy f z
gM aN cO AP aF s [ a F c\ oy mpl{ G w eI m{ eI nR tH aF rJ y | g} eI nR eI tH iQ c \ m{ aF rkJ�~ eI r J fzoy r J Ps
S�TeuU!V dW
oX mY oX nZ aN sT s[ ppG�G .� Populations of
indigenous bacteria harbouring homologs of gacA found in Pseudomonas spp. were isolated
from roots of wheat by colony hybridization with the GacA probe and subsequent PCR with
the GACA primers. On KMB+, a medium semi-selective for Pseudomonas (Simon and Ridge,
1974), their densities ranged from approximately 5.0 X 106 to 1.8 X 107 CFU g-1 root and
represented on average 35.8 to 73.5% of the total bacterial population recovered on this
medium (Table 5). Based on colony hybridization and subsequent PCR, 494 gacA positive
(referred to as ‘gacA+‘) and 58 gacA negative (referred to as ‘gacA-‘) isolates were selected for
further genotypic and phenotypic analyses (Table 6). RAPD analysis with three 10-mer
primers showed 116 and 34 unique groups among the ‘gacA+’ and ‘gacA-‘ isolates, respectively.
Characterization of representatives from all RAPD groups, employing Southern hybridization

C� h�
a� pt��� e� r � 3: ��� g� a� c� A� i� n � Ps���
eu�!� d�
o� m� o� n� a� s� s� p� e� c� i� e� s�
66
of EcoRI-digested genomic DNA, confirmed that all ‘gacA+’ isolates responded positively with
the GacA probe, whereas all 58 ‘gacA-‘ isolates responded negatively, even at low-stringent
conditions (data not shown). Based on PCR with group-specific primers (Widmer et al.,
1998), all ‘gacA+’ isolates were identified as Pseudomonas, whereas none of the 58 ‘gacA-‘
isolates gave a positive response. Based on GC-FAME-analysis of the ‘gacA-‘ isolates, 62% were
classified as Stenotrophomonas maltophilia, 14% as Comamonas acidovorans, 5% as Cryseomonas
indologenes, and 19% could not identified conclusively due to very low matching coefficients.
Subsequent characterization of the isolates for the production of specific secondary metabolites
and extracellular enzymes, known to be regulated by gacA, showed that none of the ‘gacA-‘
isolates displayed fluorescence, produced surfactants, or harboured biosynthetic genes for 2,4-
diacetylphloroglucinol and phenazine antibiotics (Table 6).
Fluorescence and surfactant production was found in 88 and 0.6% of the ‘gacA+’ isolates,
respectively, and genes for 2,4-diacetylphloroglucinol and phenazine antibiotics were found in
34 and 3% of the ‘gacA+’ isolates, respectively. Both ‘gacA+’ and ‘gacA-‘ isolates produced
protease and phospholipase C, enzymes that are under the control of gacA in the biocontrol
strain P. fluorescens CHA0 (Sacherer et al., 1994). Protease production was found in similar
proportions of ‘gacA+’ and ‘gacA-‘ isolates (~70%), whereas phospholipase C was detected in a
higher proportion of the ‘gacA-‘ isolates. Chitinase was detected only in ‘gacA-‘ isolates and in
particular in the isolates that were identified by GC-FAME analysis as Stenotrophomonas
maltophilia.
T�
a� b� l�e � 5. �¡ F¢ r£ e� q¤ u¥ e� n¦ c§ y ¨ o© f
ªg« a¬ c A® + b¯ � a� c§ t° e� r£ i± a� l � i
±s² o© l�a� t° e� s ² i
±n ¦ t° h³ e � r£ h³ i
±z´ o© s² pµ h
³e� r£ e � o© f
ªw¶ h³
e� a� t ° g· r£ o© w¶ n ¦ i±n ¦ d¸
i±ffª�ª
e� r£ e� n¦ t ° s² o© i±l�s² .
Soil Total population a b
(CFU g-1 root)GacA+ b
(CFU g-1 root) GacA/ tot. (%)
CB 1.9 X 107 9.8 X 106 52.4SL 1.5 X 107 5.4 X 106 35.8SU 2.9 X 107 1.5 X 107 51.0SV 2.5 X 107 1.8 X107 73.5SSB 2.4 X 107 1.7 X 106 69.4
a Total population of bacteria isolated on Pseudomonas semi-selective medium (Simon and Ridge, 1974).b Colony hybridization followed by PCR was used to determine the frequencies. Mean values of four replicates
are shown.

C¹ hº
a» pt¼�½ e¾ r ¿ 3: À�Á g aà cÄ AÅ iÆ n Ç PsÈ�É
euÊ!Ë dÌ
oÍ mÎ oÍ nÏ aà sÉ sÐ p¼ e¾ cÑ iÆ e¾ sÐ
67
DÒ IÓ SÔ CU¹ÖÕ SSÔ×Ô IÓ OØ NÙ
The response regulator gene gacA influences the production of several secondary
metabolites in both pathogenic and beneficial Pseudomonas spp. (Heeb and Haas, 2001). In
this study, we demonstrated that gacA is highly conserved within the genus Pseudomonas. In
PCR analysis and Southern hybridization with the gacA primers and probe, multiple strains of
different Pseudomonas species all responded positively, whereas no response was obtained from
18 other strains representing 14 species and 8 genera of Gram-negative bacteria other than
Pseudomonas (Table 1). Furthermore, from a total of approximately 550 indigenous bacterial
isolates obtained from the rhizosphere of wheat, all isolates that responded positively to the
GacA probe and primers (referred to as gacA+) were classified as Pseudomonas spp., whereas
isolates that did not hybridize with the GacA probe (referred to as gacA-) were identified as
bacterial genera other than Pseudomonas, including Stenotrophomonas, Cryseomonas and
Comamonas spp. (Table 6). Based on RAPD analyses, both gacA+ and gacA- isolates
represented a large number of genotypically different groups. These results indicate that gacA
can be used as a complementary genetic marker for detection of Pseudomonas spp. in
rhizosphere samples. They also show that the selectivity of King’s medium B for Pseudomonas
spp. is limited, although all fluorescent bacteria recovered on this medium responded
TÚ
aÛ bÜ lÝe Þ 6. ß/à
Pá
hâ
eÞ nã oä tå yæ pç ièc é aÛ nã d
êgë eÞ nã oä tå yæ pç i
èc céìé hâ aÛ rí aÛ cé tå eÞ rí iè sî tå iè cé s î oä f
ïgð añ cò Aó + ô aÛ nã d
êgð añ cò Aó - õ sî tå rí aÛ iè nã s î f
ïrí oä m ö tå hâ e Þ w÷ h
âeÞ aÛ tå
rí hâ ièzø oä sî pç h
âeÞ rí eÞ .à
Analysis a gacA+ gacA-
Number of isolates 494 58Unique RAPD groups b 116 342,4-diacetylphloroglucinol c 33.8 0.0Phenazines c 3.0 0.0Fluorescence c 88.0 0.0Protease c 71.0 69.0Phospholipase C c 19.6 70.7Surfactants c 0.6 0.0Chitinase c 0.0 55.2Identity d Pseudomonas (100%) Stenotrophomonas (62%),
Comamonas (14%),Cryseomonas (5%),unidentified (19%)
a Percentage in relation to the number of isolates.b RAPD analysis made with three primers and 100% similarity was used to define a unique group.c Secondary metabolites were detected as mentioned in Table 1.d Identity determined with Pseudomonas group-specific primers targeting 16S rDNA (Widmer et al., 1998), API
test, and GC-FAME analysis.

Cù hú
aû ptü�ý eþ r ÿ 3: ��� g� a� c� A� i� n � Ps�
eu�� d�
o� m� o� n� a� s s� pü eþ c� i� eþ s�
68
positively to hybridization and PCR with the GacA probe and primers and were identified as
Pseudomonas spp. by group-specific primers.
Although many studies have focused on the genus Pseudomonas, their taxonomy and
phylogenetic relationships are not completely resolved (Bossis et al., 2000). Among the various
methods used to identify and classify pseudomonads, DNA-DNA hybridization and 16S
rDNA sequencing are the most advocated ones (Wayne, et al., 1987; Anzai et al., 2000). The
use of both methods, however, may present certain limitations. DNA-DNA homology has not
been powerfull enough to reveal phylogenetic relationships (Anzai et al., 1997). Also,
sequences of 16S rDNA present limited power when analyzing closely related organisms that
diverged at almost the same time (Fox et al., 1992; Woese, 1987). Furthermore, 16S rDNA
sequences occur in multiple copies in a single cell and, interestingly, these copies were shown
to evolve at different rates (Ueda et al., 1999).
In this study, phylogenetic relationships inferred from gacA sequences revealed six
distinct bacterial groups, including the Pseudomonaceae, Enterobacteriaceae,
Alteromonadaceae, Vibrionaceae, Burkholderia, and Xanthomonas group (Fig. 1). Within the
Pseudomonaceae, polymorphisms within the gacA gene allowed further distinction of at least
six subclusters. Although similar overall phylogenetic relationships were found for 16S rDNA
sequences, some differences were noted. For example, P. fluorescens strains CHA0 and Pf-5
were clustered closer to P. aeruginosa and P. syringae based on 16S rDNA sequences, whereas
clustering based on gacA gene and GacA protein sequences placed both strains closer to the
other P. fluorescens strains. Discrepancies between phylogenies of Pseudomonas inferred from
16S rDNA and other genes were also observed for gyrB and rpoD (Yamamoto and Harayama,
1998; Yamamoto et al., 2000). Given the relatively small size of the gacA gene and its apparent
occurrence as a single copy in Pseudomonas makes the gacA gene not only a useful
complementary genetic marker for detection of Pseudomonas but also eligible for phylogenetic
studies. It is remarkable that a relatively small sized gene like gacA has such strong resolving
capacity for phylogenetic purposes. Comparable results were obtained with the 249-bp oprI
gene, which allowed molecular taxonomic studies in pseudomonads belonging to the rRNA
group I, a group that contains the genuine Pseudomonas species (De Vos et al., 1998).
Interestingly, polymorphisms within gacA homologs also allowed distinction of five subclusters
in the group of enteric bacteria, including E. coli, Salmonella, K. pneumoniae, P. caratovorum,
and Yersinia species (Fig. 1). These results suggest that gacA homologs also may provide a
complementary genetic marker for phylogenetic studies of Gram-negative bacteria other than

C� h�
a� pt��� e� r � 3: ��� g� a c! A" i# n $ Ps%�&
eu'�( d)
o* m+ o* n, a s& s- p� e� c. i# e� s-
69
Pseudomonas. More sequences of gacA homologs of representative strains of different enteric
and other bacterial genera will be required to further support this hypothesis.
Estimations of nonsynonymous to synonymous substitution rates (Ka/Ks) indicated
that, for members of both the Pseudomonaceae and Enterobacteriaceae, purifying selection is
acting on gacA (Table 3). These results indicate that there is selective pressure to avoid
substitutions leading to functional changes in the GacA protein. In this context, it is
interesting to note that for several P. fluorescens strains spontaneous mutations in gacA and also
gacS have been described to occur frequently under laboratory conditions (Duffy and Défago,
2000; Whistler et al., 1998). In P. fluorescens strain CHA0, grown in nutrient-rich conditions,
spontaneous mutants accumulated over a period of several days to levels of 1.25% of the
population, and half of the mutants tested were restored by gacA (Bull et al., 2001).
Subsequent competition experiments showed that the gacA mutant had only a temporary
selective advantage over the wildtype that was restricted to an early phase of the stationary
phase (Bull et al., 2001). The role of the gacA/gacS pair in surviving exposure to oxidative stress
(Whistler et al., 1998), and in production of secondary metabolites that play a significant role
in the ecological fitness of certain Pseudomonas strains (Mazzola et al., 1992; Natsh et al.,
1994) provide, at least in part, an explanation for the relatively low rate of synonomous
substitutions in gacA. Tajima’s relative rate test showed that in 50% of the comparisons
between members of the Enterobacteriaceae and Pseudomonadaceae, evolutionary rates in
gacA were higher (Table 4). So, in several members of the Enterobacteriaceae the gacA gene
appears to evolve faster than in members of the Pseudomonaceae, even though purifying
selection is acting on gacA of both bacterial groups. The biological relevance of this difference
is not clear. One may speculate that many enteric bacterial strains have a parasitic life-style and
a higher exposure to therapeutic antibiotics for which a higher adaptive capacity may be
required. However, the parasitic life-style also applies to several plant and human pathogenic
bacterial species of the Pseudomonaceae, including P. syringae and P. aeruginosa. More studies
will be necessary to elucidate the biological relevance, if any, of the relatively higher
evolutionary rates of gacA in several members of the Enterobacteriaceae as compared to
Pseudomonaceae.

C/ h0
a1 pt2�3 e4 r 5 3: 6�7 g8 a9 c: A; i< n = Ps>�?
eu@�A dB
oC mD oC nE a9 s? sF p2 e4 cG i< e4 sF
70
MH
AI
TJ EK RL IMAI
LN
S O AI
NP
D Q
MH
EK TJ HR
OS DQ
SO
BT aU cV tW eX rY iZ aU l [ s\ tW rY aU iZ n] s \ aU n] d ^
cV u_ l[tW u_ rY e X cV o` n] di
ZtW iZ o` n] s\ . a Naturally
occurring Pseudomonas spp. were isolated fromroots of wheat grown in raw soils on the semi-selective King's medium B (KMB) agarsupplemented with chloramphenicol (13 µg ml-1),ampicillin (40 µg ml-1), and cycloheximide (100 µgml-1) [KMB+] (Simon and Ridge, 1974). Bacterialisolates were routinely grown on KMB medium.For DNA extractions, bacteria were cultivatedovernight at 24ºC on nutrient broth yeast extractagar (Vidaver, 1967). Escherichia coli strains weregrown in LB (Luria Bertani). All bacterial strainswere stored at -80ºC in LB or KMB brothsupplemented with 40% glycerol (v/v).
DQ
NP
A I
eX xb tW rY aU cV tW iZ o` n] s\ .a For all PCR analyses, includingRAPD analysis, heat-lysed bacterial suspensionswere used and prepared as described previously(Raaijmakers et al., 1997). Total DNA for use inPCR amplifications of the 16S rDNA genes wasobtained as described by Raaijmakers et al. (1997).Total genomic DNA, used for Southern blotanalysis and gacA amplifications, was extracted frombacterial strains by a modified version of acetyltrimethylammoniumbromide (CTAB)-basedprotocol (Ausubel et al., 1995). A 1.5-ml sample ofbacterial cultures grown overnight was centrifugedfor 3 min at 14,000 rpm; supernatant was discardedand the pellet was resuspended in 550 µl of TEbuffer (Tris 10mM, EDTA 10mM, pH 8.0)amended with lysozyme (1.82 mg ml-1); the mixturewas incubated at 37ºC for 30 min. Seventy-five µlof 10% SDS amended with proteinase K (0.86 mgml-1) was added to the bacterial suspension andthoroughly mixed. After 15 min incubation at65ºC, 100 µl of 5 M NaCl and 80 µl CTAB/NaCl(0.3 M CTAB, 0.7 M NaCl) were added. After 10min of incubation at 65ºC, DNA was obtained byextraction with chloroform/isoamyl alcohol (24:1,vol/vol), isopropanol precipitation and subsequentwashes with 70% ethanol. The extracted DNA wasdissolved in 100 µl of 10 mM Tris pH 8.0containing RNAase (20 µg ml-1) and stored at –20ºC. Restrictions of genomic DNA wereperformed with 5U of EcoRI (Promega), an enzymewithout restriction sites in the gacA genes of P.fluorescens strains BL915, CHA0, and Pf-5, P.aeruginosa PAO1, P. syringae pv. syringae B728a, P.chlororaphis 30-84, and the repB gene of P.viridiflava PJ-08-6A. Digestions were performed ina total volume of 100 µl containing 2.0 µg of DNA;digested DNA was precipitated with 4M LiCl,washed with 70% ethanol, dissolved in 15 µl ofsterile distilled water and separated on 1% agarosegels in TBE.
Pc Cd R L aU me pf l[iZfg iZ cV aU tW iZ o` n ] of g tW hh e X gi aj ck Al gm eX n] eX . Primers
GACA1 and GACA2 were used to amplify the 425-bp fragment from different Pseudomonas spp.
strains. Amplifications were performed in 25-µlreaction mixtures containing 3 µl of diluted, heat-lysed cell suspension, 1X PCR buffer (Perkin-Elmer, Nieuwerkerk aan de IJssel, TheNetherlands), 1.5 mM MgCl2 (Perkin Elmer), 200µM of each, dATP, dCTP, dGTP, and dTTP(Promega, Leiden, The Netherlands), 20 pmol ofeach primer, and 2U of AmpliTaq polymerase(Perkin-Elmer). The PCR program for gacAconsisted of an initial denaturation at 95ºC for 2min followed by 30 cycles at 95ºC for 1 min,annealing at 61ºC for 1 min, and extension at 72ºCfor 1 min. Amplifications were carried out in a MJResearch PTC-200 thermocycler.
SO eX qn u_ eX n] cV iZ n] g m o` f g tW hh e X gi aj ck Al gm eX n] eX .a Amplifications of thegacA gene with primers GACA1 and GACA2 wereperformed with the Expand High Fidelity PCRsystem, following the supplier’s recommendations(Roche Diagnostics GmbH, Mannheim, Germany).PCR products were run in a 1% agarose gel, bandsof the appropriate size were excised from the gel andpurified by using the PCR purification kit (Qiagen,Hilden, Germany). These fragments weresequenced directly by loading on an ABI sequencer.
SO eX qn u_ eX n] cV iZ n] g m o` f g 16oqp S O rY DQ NP
AI
.a Five microliters of thecell lysis suspension were used in each reaction withthe primers 8f and 1406r (Amann et al., 1995)using reaction conditions as described by Liu et al.(1997). PCR-amplified products were analyzed byelectrophoresis on 2% agarose gels. Fragments ofthe appropriate size were cloned into the T/A vectorpCR2.1 (Invitrogen, Carlsbad, CA), andtransformed into E. coli strain INVαF’.Transformants were selected on LB agar amendedwith kanamycin (75 µg ml-1) and X-Gal (20 µgml-1). Plasmid DNA was isolated fromtransformants using the S.N.A.P. mini-prep kit(Invitrogen). Cycle sequencing was conducted on aPerkin-Elmer model 480 thermal cycler (Norwalk,CT) and PRISM Ready Reaction DyeDeoxyTerminator Cycle Sequencing Kit (PE Biosystems,Warrington, UK). Reactions were conducted usingcycling conditions recommended by themanufacturer with the T3 or T7 primer. Productsof the sequencing reactions were purified usingCENTRI-SEP spin columns (PrincetonSeparations, INC., Adelphia, NJ) and dried in avacuum centrifuge. Samples were resuspended in 6µl of a solution containing de-ionized formamideand blue dextran/EDTA in a ratio of 5:1. Sampleswere heated for 2 min at 90ºC and 1.5 µl wasloaded on an ABI 377 DNA sequencer (PEBiosystems, Foster City, CA). BLASTN searcheswere conducted on the nucleotide sequencesobtained.

Cr hs
at ptu�v ew r x 3: y�z g{ a| c} A~ i� n � Ps���
eu��� d�
o� m� o� n� a| s� s� pu ew c� i� ew s�
71
N� u� c� l� e� o� t� i� d� e � s� e� q� u� e� n� c� e � a� cc��� e� ss��� i� o� n n��� u� m� b�
e� r � o� f �
g� a c¡ A¢ a� n� d�
16£�¤ S ¥ r� D¦ N� A§
. Newly obtained sequences weredeposited with GenBank under accession numbersshown in Table 2.
P© hª y« l� o� g¬ e� n� e� t� i� c � a� n� a� l� y« s� i� s� . Prior to analysis, sequences ofthe gacA gene and its homologs were separated intothe following categories: 1) all gacA sequencestogether (44 sequences); 2) gacA sequences from thePseudomonadaceae and Enterobacteriaceae (36sequences); 3) only gacA sequences fromPseudomonadaceae (22 sequences); and 4) onlygacA sequences from Enterobacteriaceae (14sequences). Amino acid and nucleotide sequenceswere aligned by using the CLUSTAL W 1.81program (Thompson et al., 1994). Pairwisenonsynonymous (Ka) and synonymous (Ks)nucleotide substitutions in the gacA sequences werecalculated according to the Nei-Gojobori’s method(Nei and Gojobori, 1986) as implemented inMEGA (Kumar et al., 2001). Fisher’s exact test andZ-test for selection based on codon usage wereperformed by the Nei-Gojobori’s method with thenumber of synonymous differences per synonymoussite and the number of nonsynonymous differencesper nonsynonymous site. Significance values for theZ-test were determined by using 1,000 bootstrapreplicates.
Majority sequences of gacA of speciesbelonging to the Pseudomonaceae andEnterobacteriaceae were computed using Megalign(DNAstar Inc., WI, USA) to reduce the number oftwo-by-two comparisons by the Tajima’s test.Tajima’s 1D relative rate test (Tajima, 1993) wasused to estimate pairwise rate variation betweenmajority sequences of gacA with the closest relativeas outgroup. Phylogenetic trees of gacA and 16SrDNA were constructed by the PHYLIP 3.5cPackage (Felsenstein, 1993), MEGA, and Treecon(Van de Peer and De Wachter, 1994) using theNeighbour-joining method (Saitou and Nei, 1987)with the nucleotide substitution model of Jukes andCantor (1969). For comparison purposes, similaranalyses were performed with the maximumlikelihood method. Phylogenetic trees with theGacA protein sequences were constructed using theNeighbour-joining method with the PAM distancematrix of Dayhoff (Dayhoff et al., 1978) and werecomputed with the program PROTDIST ofPHYLIP. Confidence of the tree topology wasperformed by bootstrap analysis on 1,000resamplings (Felsenstein, 1985).
S¥ o� u� t� hª e� r� n � a� n� d �
c� o� l� o� n� y « hª y« b�
r� i� d� i� z a� t� i� o� n� s� .¨ DNA transferfrom agarose gels and bacterial colony transfer toHybond N+ nylon membranes (AmershamPharmacia Biotech) were performed according tostandard methods (Sambrook and Russel, 2001).Membrane washes and hybridizations wereperformed following standard procedures(Sambrook and Russel, 2001). High-stringent
conditions comprised pre-hybridization for 1.5 h at65ºC, hybridization for 12 h at 65ºC, membranewashings twice each for 5 min with 2X SSC (1XSSC is 0.15 M NaCl plus 0.015 M sodium citrate)-0.1% SDS at room temperature followed bymembrane washings twice each for 30 min with0.1X SSC-0.1% SDS at 65ºC. Low stringentconditions consisted of pre-hybridization andhybridization at 55ºC and two membrane washingswith 0.5X SSC at 55ºC. The GacA probe wasgenerated by direct labeling in PCR using the DIGlabeling mix (Roche, Almere, The Netherlands).Hybridized probes were immunodetected accordingto the protocols provided by the supplier.
S¥ o� i� l� s� .¨ Five soils were obtained from agriculturalfields in the Netherlands. All soils were collectedfrom the upper 50 cm of the soil profile, air driedfor a week, and passed through a 0.5-cm meshscreen prior to use. Soils CB, SU, SV, and SSBwere collected in December 1997 from a polder inthe South-West of the Netherlands, 10 km fromthe city of Bergen op Zoom. These soils arephysicochemically very similar, containing onaverage, 27% clay, 10% silt, and 51% sand. Theorganic matter content is on average 4.3% and thepH, determined after extraction with CaCl2 is 7.5.Soil SL was collected in June 1998 from a polder inthe North-East of the Netherlands, at Lovinkhoeve,located 30 km from the city of Lelystad. SL soilcontains 21% clay, 12% silt, 54% sand, 3.6 %organic matter, and pH, determined after extractionwith CaCl2 is 7.5.
P© l� a� n� t � c� u� l� t� i� v® a� t� i� o� n� .¨ Pots containing 200 g of sievedsoil were sown with 15 seeds of wheat, cv. Bussard.Plants were grown in a climate chamber at 15ºCwith a 12-h photoperiod. Plants received 50 ml of1/3 strength Hoagland's solution (macroelementsonly) twice a week. After 30 days of plant growth,the roots were harvested and used for isolation ofroot-associated bacteria.
I¯s� o� l� a� t� i� o� n � o� f
�b�
a� c� t� e� r� i� a � f�r� o� m � t� hª e � w° hª e� a� t � r� hª i� z o� s� p± hª e� r� e� .
Three to five plants of each replicate were randomlyharvested and loosely adhering soil was removed.Root samples of 0.2 to 1.0 gram were vortexed for 1min, sonicated for 1 min in a ultrasonic cleaner(Bransonic 12, Branson Ultrasonics Corp., Geneva,Switzerland), and dilution plated onto KMB+.Plates were incubated at 25ºC for 48 h. Populationsof bacteria harbouring gacA homologs weredetermined by colony hybridization with the GacAprobe followed by confirmation by PCR with theGacA primers. Colonies that reacted positively inhybridization with the GacA probe and primers(gacA+ isolates) were isolated and purified forfurther analysis; several colonies that reactednegatively in hybridization and subsequent PCRwith the GacA probe and primers (gacA- isolates)were also included.

C² h³
a´ ptµ�¶ e· r ¸ 3: ¹�º g» a¼ c½ A¾ i¿ n À PsÁ�Â
euÃ�Ä dÅ
oÆ mÇ oÆ nÈ a¼ s sÉ pµ e· cÊ i¿ e· sÉ
72
CË hÌ
aÍ rÎ aÍ cÏ tÐ eÑ rÎ iÒ zÓ aÍ tÐ iÒ oÔ n Õ oÔ f Ö rÎ hÌ iÒ zÓ oÔ s× pØ hÌ
eÑ rÎ e Ñ bÙ
aÍ cÏ tÐ eÑ rÎ iÒ a Í hÌ
aÍ rÎ bÙ oÔ uÚ rÎ iÒ nÕ gÛgÜ aÝ cÞ Aß h
ÌoÔ mà oÔ lá oÔ gÛ s× .â Isolates obtained from the
rhizosphere of wheat were genotypically identifiedand characterized for the production of specificextracellular enzymes and secondary metabolitesknown to be regulated by gacA. The first genotypicidentification of the isolates was by PCR analysiswith primers specific for the 16S rDNA of thegenus Pseudomonas (sensu stricto) (Widmer et al.,1998). Secondly, isolates were clustered ingenotypic groups by Random AmplifiedPolymorphic DNA (RAPD) analysis with primersM13, M12, and D7 (Keel et al., 1996). Thepresence or absence of fragments generated byRAPD analysis (1 for the presence or 0 for theabsence of a particular sized band in the gel) wasused to calculate the pairwise coefficients ofsimilarity (Nei/Li distances). RAPD analysis wasperformed three times for each primer. Gaschromatograph-fatty acid methyl ester (GC-FAME)analysis was performed to identify gacA- bacterialisolates. For GC-FAME analysis, isolates werecultivated on tryptic soy broth agar (BectonDickinson, Cockeysville, MD) and incubated for24 h at 28ºC. Cells were collected with a 4 mm
diameter transfer loop and processed for extractionof fatty acids using the procedures as outlined bythe manufacturer (Microbial ID, Inc., Newark,DE). Fatty acid methyl esters were analyzed using aMicrobial Identification System equipped with anHP5890 series II gas chromatograph, HP3365Chem Station and version 3.9 of the aerobe library(Microbial ID, Inc.).
For all isolates, presence of genes involvedin the biosynthesis of specific antibiotics wasperformed with primers Phl2a and Phl2b for 2,4-DAPG (Raaijmakers et al., 1997) and primersPHZ1 (5’-GGCGACATGGTCAACGG) andPHZ2 (5’-CGGCTGGCGGCGTATTC) forphenazine antibiotics (Mavrodi et al., unpublisheddata). Protease, phospholipase C, and chitinaseactivities were detected by growing the bacterialisolates on skim milk agar, egg yolk agar (Sachereret al., 1994), and chitin-agar (De Boer et al., 1998),respectively. Production of biosurfactants andfluorescence of the isolated bacteria was determinedby growth on medium containing CTAB-methylene blue (Siegmund and Wagner, 1991) andPseudomonas medium F (Cho and Tiedje, 2000),respectively.
Aã
CË Kä Nå Oæ Wç
Lè
Eé
Dê Gë
Eé
Mì
Eé
Nå Tí
Sî
This research was financially supported by CAPES, Brazil (project nr. 1515/96-9). We thank MiekePrins for technical assistance and Prof. Dr. Pierre de Wit for critically reading this manuscript and valuablesuggestions. We wish to thank Dr. Peter Vandamme for providing B. cepacia LMG1222, B. multivoransLMG13010, and B. vietnamiensis LMG10929. We thank the Institute for Genomic Research (TIGR) forsequences of Pseudomonas fluorescens PfO-1, P. putida KT 2440, Shewanella putrefasciens MR-1, Shigella flexneri2a, Xyllela fastidiosa ann-1, X. fastidiosa Dixon, X. fastidiosa Temecula 1, Salmonella typhi TY2, Klebsiellapneumoniae M6H, Escherichia coli CFT073, Yersinia pestis KIM5, and P. syringae pv. tomato DC3000; theSanger center for sequences of Y. enterolitica 0:8, and Burkholderia cepacia J2315.
Rï Eé
Fð
Eé
Rï Eé
Nå CË Eé
Sî
Aã
mà aÍ nnÕñÕ , ò Rï .â Ió ., âôò Lè
uÚ dõ wö iÒ gÛ , ò Wç
., âôò aÍ nÕ d õ SîcÏ hÌ lá eÑ iÒ fÖ eÑ rÎ , ò Kä . â H÷ .â
1995. Phylogenetic identification and in situdetection of individual microbial cells withoutcultivation . Microbiol. Rev. 59:143-169.Aã
nÕ zÓ aÍ iÒ , ò Yø
., âùò Kä iÒ mà , ò H÷ ., âôò Pú aÍ rÎ kû , ò Jü.â -ý Yø .â , ò W
çaÍ kû aÍ bÙ aÍ yþ aÍ s× hÌ iÒ , ò H÷ .,âôò
aÍ nÕ d õ Oæ yþ aÍ iÒ zÓ uÚ , ò H÷ .â 2000. Phylogenetic affiliation ofthe pseudomonads based on16S rRNA sequence.Int. J. Syst. Evol. Microbiol. 50:1563-1589.Aã
ppØÿØ lá eÑ bÙ yþ , ò Jü.â Lè ., âôò Pú aÍ rÎ kû iÒ nÕ s× oÔ nÕ , ò J
ü.â Sî ., âôò aÍ nÕ d õ B
�oÔ uÚ rrÎ Î eÑ tÐ , ò Rï .â B� .â
1996. Signal transduction via the multi-stepphosphorelay: not necessarily a road less traveled.Cell 86:845-848.
Aã
uÚ s× uÚ bÙ
eÑ lá , ò Fð
.â Mì ., âôò B�
rÎ eÑ nÕ tÐ , ò Rï ., âôò Kä iÒ nÕ gÛ s× tÐ oÔ nÕ , ò Rï .â Eé ., âôò Mì
ooÔ Ô rÎ eÑ ,òDê .â Dê ., âôò S
îeÑ iÒ dõ mà aÍ nÕ , ò J
ü.â Gë ., âôò S
îmà iÒ tÐ hÌ , ò J
ü.â Aã ., âôò aÍ nÕ d õ S
îtÐ rÎ uÚ h
Ìlá , Kä .â
(ed.). 1992. Short protocols in molecular biology,pages 16.8-16.9, John Wiley and Sons, New York,N.Y.B�
lá uÚ mà eÑ rÎ , ò CË ., âùò aÍ nÕ d õ H÷ aaÍ Í s× , ò Dê .â 2000. Iron regulation ofthe hcnABC genes encoding hydrogen cyanidesynthase depends on the anaerobic regulator ANRrather than on the global activator GacA inPseudomonas fluorescens CHA0. Microbiol.146:2417-2424.

C� h�
a� pt��� e� r � 3: � g� a c� A� i� n � Ps���
eu��� d�
o� m� o� n� a s� s� p� e� c� i� e� s�
73
B� o� ss ! i" s , # E$ ., % # L&
e' m( a) n* c+ e' a) u, , # P- ., % # L&
a) t. o� u, r/ , # X0
., % # a) n* d 1
G2
a) r/ d1 a) n* ,#L&
.% 2000. The taxonomy of Pseudomonas fluorescensand Pseudomonas putida: current status and need forrevision. Agronomie 20:51-63.B� u, ll
343, # C5 .% T6 ., % # D7 u, ff
898y: , # B� ., % # V
;o� i" s a) r/ d1 , # C5 ., % # D7 é< f8 a) g= o� , # G
2.,% #
K> ee'?' l3 , # C5 ., % # a) n* d 1
H@
aa)A) s , # D7 .% 2001. Characterization ofspontaneous gacS and gacA regulatory mutants ofPseudomonas fluorescens biocontrol strain CHA0.Antonie Leeuwenhoek 79:327-336.C5 hB
a) n* c+ e' y: , # SC .% T6 ., % # WD
oo�E� d1
, # D7 .% WD ., % # a) n* d 1
P- i" e' r/ s o� n * IIIFGFHF , # L&
.% SC .%1999. Two-component transcriptional regulation ofN-acyl-homoserine lactone production inPseudomonas aureofaciens. Appl. Environ.Microbiol. 65:2294-2299.C5 hB
o� , # JI.% -J C5 ., % # a) n* d
1T6 e' i" d1 j
Ke' , # J
I.% ML .% 2000. Biogeography
and degree of endemicity of fluorescentPseudomonas strains in soil. Appl. Environ.Microbiol. 66:5448-5456.D7 a) y: h
Bo� ff898
, # ML
.% OM ., % # SC c+ hB wN a) r/ t. zO , # RP .% ML ., % # a) n* d 1
OM
r/ c+ u, tt.Q. , # B� .% C5 .%1978. A model of evolutionary change in proteins.In: Dayhoff, M.O. (ed). Atlas of protein sequenceand structure. Vol. 5, Suppl. 3. NationalBiomedical Research Foundation, Washington,USA.D7 e ' B� o� e' r/ , # W
D., % # G
2u, nn*R* e' wN i" e' kS , # P- .% JI .% AT .% K> ., % # L
&a) f8 e' bU e' r/ , # P- .% ,#
JIa) n* s e' , # J
I.% D7 ., % # SC pV i" t. , # B� .% E$ ., % # a) n* d
1WD
o� l3d1
e' n* d1
o� r/ pV , # JI.% WD .%
1998. Anti-fungal properties of chitinolytic dunesoil bacteria. Soil Biol. Biochem. 30:193-203.D7 e ' SC o� u, zO a) , # J
I.% T6 ., % # a) n* d
1RP a) a) i" jK m( a) kS e' r/ s , # J
I.% ML .% 2002.
Polymorphisms within the prnC and pltD genes ofpyrrolnitrin- and and pyoluteorin-producingPseudomonas and Burkholderia spp. FEMSMicrobiol. Ecol. accepted.D7 e ' V
;o� s , # D7 ., % # B� o� u, t. o� n* , # C5 ., % # SC a) r/ n* i" g= u, e' t. , # A
T., % # D7 e ' V
;o� s , # P- .,% #
V;
a) u, t. e' r/ i" n* , # ML
., % # a) n* d 1
C5 o� r/ n* e' l3 i" s , # P- .% 1998. Sequencediversity of the oprI gene, coding for major outermembrane lipoprotein I, among rRNA group Ipseudomonads. J. Bact. 180:6551-6556.D7 u, f
8f8y: , # B� .% K> ., % # a) n* d
1D7 é< f8 a) g= o� , # G
2.% 1999. Environmental
factors modulating antibiotic and siderophorebiosynthesis by Pseudomonas fluorescens biocontrolstrains. Appl. Environ. Microbiol. 65:2429-2438.D7 u, f
8f8y: , # B� .% K> ., % # a) n* d
1D7 é< f8 a) g= o� , # G
2.% 2000. Controlling
instability in gacS-gacA regulatory genes duringinoculant production of Peudomonas fluorescensbiocontrol strains. Appl. Environ. Microbiol.66:3142-3150.FW e' l3 s e' n* s t. e' i" n* , # J
I. % 1985. Confidence limits in
phylogenies: an approach using the bootstrap.Evolution 39:783-791.FW e' l3 s e' n* s t. e' i" n* , # J
I.% 1993. PHYLIP: phylogeny inference
package, version 3.5c. Department of genetics,University of Washington, Seatle.FW o� xX , # G
2.% E$ ., % # W
Di" s o� t. zO kS e' y: , # J
I.% D7 ., % # a) n* d
1JIu, r/ t. s hB u, kS , # P- .% JI r/ .%
1992. How close is close: 16S rRNA sequenceidentity may not be sufficient to guarantee speciesidentity. Int. J. Syst. Bacteriol. 42:166-170.G2
a) ff898 n* e' y: , # T6 .% D7 ., % # L&
a) m( , # SC ., % # L&
i" g= o� n* , # JI.% , # G
2a) t. e' s , # K> .,% #
FW r/ a) zO e' ll343 e' , # AT
., % # D7 i " ML
a) i" o� , # JI., % # H@
i" l3 l3 , # D7 .% SC ., % # G2
oo�E� d1
wN i" n* , # SC .,% #T6 o� r/ kS e' wN i" t. zO , # N
Y., % # AT
ll3Z3
s hB o� u, s e' , # AT
.% ML ., % # K> e' m( pV f8, # H@
.% -J JI ., % # a) n* d1
B� e' c+ kS e' r/ , # JI.% OM .% 1994. Global regulation of expression
of antifungal factors by a Pseudomonas fluorescens
biocontrol strain. Mol. Plant-Mcrobe Interact.7:455-463.H@
ee'[' bU , # SC ., % # a) n* d 1
H@
aa)A) s , # D7 .% 2001. Regulatory roles ofthe GacS/GacA two-component system in plant-associated and other Gram negative bacteria. Mol.Plant-Microbe Interact. 14:1351-1363.H@
r/ a) bU a) c+ kS , # E$ .% ML ., % # a) n* d 1
WD
i" ll343 i" s , # D7 .% K> .% 1992. The lemAgene is required for pathogenesis of Pseudomonassyringae pv. syringae. J. Bacteriol. 174:3011-3020.H@
u, g= hB
e' s , # D7 .% AT ., % # a) n* d 1
NY
e' i" , # ML
.% 1988. Pattern ofnucleotide substitution at major histocompatibilitycomplex loci reveals overdominant selection. Nature335:167-170.JIo� hB
n* s e' n* , # K> ., % # AT
n* d1
e' r/ s e' n* , # SC .% , # a) n* d 1
JIa) c+ o� bU s e' n* , # C5 .% SC . % 1996.
Phenotypic and genotypic characterization ofphenanthrene-degrading fluorescent Pseudomonasbiovars. Appl. Environ. Microbiol. 62:3818-3825.JIu, kS e' s , # T6 .% H@ ., % # a) n* d
1C5 a) n* t. o� r/ , # C5 .% RP . % 1969. Evolution of
protein molecules. In: Munro, H.N. (ed).Mammalian protein metabolism, Academic Press,New York.K> ee'[' l3 , # C5 ., % # W
De' l3 l3 e' r/ , # D7 .% ML ., % # N
Ya) t. s c+ hB , # A
T., % # D7 é< f8 a) g= o� , # C5 oo�E� kS ,#
RP .% JI ., % # a) n* d 1
T6 hB
o� m( a) s hB o� wN , # L&
.% SC . % 1996. Conservation ofthe 2,4-diacetylphloroglucinol biosynthesis locusamong fluorescent Pseudomonas strains from diversegeographic locations. Appl. Environ. Microbiol.552-563.K> u, m( a) r/ , # SC ., % # T6 a) m( u, r/ a) , # K> ., % # J
Ia) kS o� bU s e' n* , # IF .% B� .% , # a) n* d
1NY
e' i" , # ML
.%2001. MEGA2: Molecular Evolutionary GeneticsAnalysis software, Arizona State University, Tempe,Arizona, USA.L&
a) v\ i" ll343 e' , # JI.% , # V;
o� i" s a) r/ d1 , # C5 ., % # K> ee'[' l3 , # C5 .% , # ML
a) u, r/ hB o� f8e' r/ , # M
L.,% #
D7 é< f8 a) g= o� , # G2
., % # a) n* d 1
H@
aa)A) s , # D7 .% 1992. Global control inPseudomonas fluorescens mediating antibioticsynthesis and suppression of black root rot oftobacco. Proc. Natl. Acad. Sci. USA 89:1562-1566.L&
i" , # WD
.% -J H@ ., % # WD
u, , # C5 .% IF ., % # a) n* d 1
L&
u, o� , # C5 .% -J C5 .% 1985. A newmethod for estimating synonymous andnonsynonymous rates of nucleotide substitutionconsidering the relative likelihood of nucleotide andcodon changes. Mol. Biol. Evol. 2:150-174.L&
i" u, , # WD
.% -J T6 ., % # ML
a) r/ s hB , # T6 .% L& ., % # C5 hB
e' n* g= , # H@
.% , # a) n* d 1
FW o� r/ n* e' y: ,#L&
. % JI.% 1997. Characterization of microbial diversity
by determining terminal restriction fragment lengthpolymorphisms of genes endcoding 16S rRNA.Appl. Environ. Microbiol. 63:4516-4522.ML
a) v\ r/ o� d1
i" , # D7 .% V; ., % # WD
e' ll343 e' r/ , # D7 .% ML ., % # a) n* d 1
T6 hB
o� m( a) s hB o� wN ,#L&
.% SC . % Unpublished data.ML
a) zzO]O o� l3a) , # M
L., % # C5 oo�E� kS , # RP .% JI ., % # T6 h
Bo� m( a) s hB o� wN , # L
&.% SC .,% #
WD
e' ll343 e' r/ , # D7 .% ML ., % # a) n* d 1
P- i" e' r/ s o� n * IIIFGFHF , # L&
.% SC . % 1992.Contribution of phenazine antibiotic biosynthesisto the ecological competence of fluorescentpseudomonads in soil habitats. Appl. Environ.Microbiol. 58:2616-2624.NY
a) t. s hB , # AT
., % # K> ee'?' l3 , # C5 .% , # P- f8i" r/ t. e' r/ , # H
@.% AT ., % # H
@a) a) s , # D7 .% , # a) n* d
1D7 é< f8 a) g= o� , # G
2.% 1994. Contribution of the global
regulator gacA to persistence and dissemination ofPseudomonas fluorescens biocontrol strain CHA0introduced into soil microcosms. Appl. Environ.Microbiol. 60:2553-2560.NY
e' i" , # ML
., % # a) n* d 1
G2
o� jKo� bU o� r/ i" , # T6 .% 1986. Simple methods
for estimating the numbers of synonymous and

Ch_
a` pta�b ec r d 3: e�f gg ah ci Aj ik n l Psm�n
euo�p dq
or ms or nt ah sn su pa ec cv ik ec su
74
nonsynonymous nucleotide substitutions. Mol.Biol. Evol. 3:418-426.Ow ’x Sy uz ll{4{ i| v} a~ n� , � D� .� J� ., �4� a~ n� d � Ow ’x G� a~ r� a~ , � F� .� 1992. Traits offluorescent Pseudomonas spp. involved insuppression of plant root pathogens. Microbiol.Rev. 56:662-676.P� a~ ll{4{ e� r� o� n� i| , � N� .� J� .� 1992. Human and animal-pathogenic pseudomonads. In: Balows, A., Truper,H.G., Dworkin, M., Harder, W., and Schleifer,K.H. (eds.). The prokariotes, a handbook on thebiology of bacteria, ecophysiology, isolation,identification and applications, 2nd ed., vol.3,Springer, New York, N.Y., USA.P� e� r� n� e� s� t� i| g� , � A
�.� -� K� ., �4� M
�e� l{ e� f� o� r� s� , � Ö� .� , � a~ n� d � G
�e� o� r� g� e� ll{4{ i| s� , � D� .�
2001. Identification of uvrY as the cognate responseregulator for the barA sensor kinase in Escherichiacoli. J. Biol. Chem. 276:225-231.P� i| e� r� s� o� n � III�H�G� , � L
�.� Sy ., �4� K� e� pp��� e� nn�R� e� , � V
�.� D� ., �4� a~ n� d � W
�oo�E� d� , � D� .�
1994. Phenazine antibiotic biosynthesis inPseudomonas aureofaciens 30-84 is regulated byPhzR in response to cell density. J. Bact. 176:3966-3974.R� aa~?~ i| j� m� a~ k� e� r� s� , � J
�.� M� .� , � W
�e� ll{4{ e� r� , � D� .� M� .� , � a~ n� d � T� h o� m� a~ s� h o� w¡ ,�
L�
.� Sy .� 1997. Fequency of antibiotic-producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63:881-887.Sy a~ c¢ h e� r� e� r� , � P� ., �4� D� é£ f� a~ g� o� , � G
�., �4� a~ n� d � H¤ aa~A~ s� , � D� . � 1994.
Extracellular protease and phospholipase C arecontrolled by the global regulatory gene gacA in thebiocontrol strain Pseudomonas fluorescens CHA0.FEMS Microbiol. Lett. 116:155-160.Sy a~ i| t� o� uz , � N� ., �4� a~ n� d � N� e� i| , � M
�.� 1987. The neigbor-joining
method: a new method for reconstructingphylogenetic trees. Mol. Biol. Evol. 4:406-425.Sy a~ m� b¥ r� oo�E� k
�, � J�., �4� a~ n� d � R� uz ss�!� e� l{ , � D� .� W� . � 2001. Molecular
cloning: a laboratory manual, 3rd ed., Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.Sy a~ r� n� i| g� uz e� t� , � A
�., �4� K� r� a~ uz s� , � J
�., �4� H¤ e� n� k
�e� l{ s� , � M
�.� D� .� ,�
M�
uz e� h l{ c¢ h e� n� , � A�
.� M� ., �4� a~ n� d � L�
o� p� e� r� , � J�.� E¦ .� 1995. The sigma
factor σs affects antibiotic production and biologicalcontrol activity of Pseudomonas fluorescens Pf-5.Proc. Natl. Acad. Sci. USA 92:12255-12259.Sy i| e� g� m� uz n� d� , � I� . � a~ n� d � W
�a~ g� n� e� r� , � F� .� 1991. New method
for detecting rhamnolipids excreted by Pseudomonasspecies during growth on mineral agar. Biotech.Tech. 5:265-268.Sy i| m� o� n� , � A
�., �4� a~ n� d � R� i| d� g� e� , � E¦ .� H¤ .� 1974. The use of
ampicillin in a simplified selective medium for theisolation of fluorescent pseudomonads. J. Appl.Bacteriol. 37:459-460.T� a~ j� i| m� a~ , � F� .� 1993. Simple methods for testing themolecular evolutionary clock hypothesis. Genetics135:599-607.
T� h o� m� p� s� o� n� , � J�.� D� ., �4� H¤ i| gg�§� i| n� s� , � D� .� G� ., �4� a~ n� d � G
�i| b¥ s� o� n� , � T� .� J� .�
1994. CLUSTAL W: impoving the sensitivity ofprogressive multiple alignment through sequenceweighting, positions-specific gap penalties andweight matrix choice. Nucleic Acids Res. 22:4673-4680.U¨
e� d� a~ , � K� ., �4� Sy e� k� i| , � T� ., �Z� K� uz d� o� , � T� ., �4� Y©
o� s� h i| d� a~ , � T� ., �4� a~ n� d�K� a~ t� a~ o� k
�a~ , � M
�. � 1999. Two distinct mechanisms cause
heterogenicity of 16S rRNA. J. Bacteriol. 181:78-82.V�
a~ n � d� e � P� ee�[� r� , � Y©
.� , � a~ n� d � D� e � W�
a~ c¢ h t� e� r� , � R� .� 1994.TREECON for windows: a software for theconstruction and drawing of evolutionary trees forthe Microsoft Windows environment. Comp. Appl.Biosci. 10:569-570.V�
i| d� a~ v} e� r� , � A�
.� K� .� 1967. Synthetic and complex mediafor the rapid detection of fluorescence ofphytopathogenic pseudomonads: effect of thecarbon source. Appl. Microbiol. 15:1523-1524.W�
a~ yª n� e� , � L�
.� G� ., �4� B«
r� e� nn�R� e� r� , � D� .� J� .� , � C¬ o� l{ w¡ e� l{ l{ , � R� .� R� .,�4�G�
r� i| m� o� n� t� , � P� .� A� .� D� ., �4� K� a~ n� d� l{ e� r� , � Ow ., �4� K� r� i| c¢ h e� v} s� k� yª , � M�
.� I� .,�4�M�
oo�E� r� e� , � W�
.� E¦ .� C¬ ., �4� M�
uz rr�H� a~ yª , � R� .� G� .� E¦ ., �4� Sy t� a~ c¢ k� e� b¥ r� a~ n� d� t� , � E¦ .,�4�Sy t� a~ rr�H� , � M
�.� P� ., �4� a~ n� d � T� r� ü p� e� r� , � H¤ .� G� .� 1987. International
committee on Systematic Bacteriology. Report onthe ad hoc committee on reconciliation ofapproaches to bacterial systematics. Int. J. Syst.Bacteriol. 37:463-464.W�
h i| s� t� l{ e� r� , � C¬ .� A� ., �4� C¬ o� r� b¥ e� ll{4{ , � N� .� A� .� , � Sy a~ r� n� i| g� uz e� t� , � A�
., �Z� R� e� a~ m� ,�W�
., �Z� a~ n� d � L�
o� p� e� r� , � J�.� E¦ .� 1998. The two-component
regulators gacS and gacA influence accumulation ofthe stationary-phase sigma factor σS and the stressresponse in Pseudomonas fluorescens Pf-5. J. Bact.180:6635-6641.W�
i| d� m� e� r� , � F� ., �4� Sy e� i| d� l{ e� r� , � R� .� J� .� , � G�
i| ll{4{ e� v} e� t� , � P� .� M� ., �4� W�
a~ t� r� uz d� ,�L�
.� Sy ., �4� a~ n� d � D� i | G�
i| o� v} a~ nn�R� i| , � D� .� 1998. A highly selectivePCR protocol for detecting 16S rRNA genes of thegenus Pseudomonas (sensu stricto) in environmentalsamples. Appl. Environ. Microbiol. 64:2545-2553.W�
o� e� s� e� , � C¬ .� R� .� 1987. Bacterial evolution. Microbiol.Rev. 51:221-271.Y©
a~ m� a~ m� o� t� o� , � Sy ., �Z� a~ n� d � H¤ a~ r� a~ yª a~ m� a~ , � Sy .� 1998.Phylogenetic relationship of Pseudomonas putidastrains deduced from the nucleotide sequences ofgyrB, rpoD and 16S rRNA genes. Int. J. Syst.Bacteriol. 48:813-819.Y©
a~ m� a~ m� o� t� o� , � Sy ., �4� K� a~ s� a~ i| , � H¤ ., �4� A�
r� n� o� l{ d� , � D� .� L� ., �4� J�a~ c¢ k� s� o� n� ,�
R� .� W� ., �Z� V�
i| v} i| a~ n� , � A�
., �4� a~ n� d � H¤ a~ r� a~ yª a~ m� a~ , � Sy .� 2000.Phylogeny of the genus Pseudomonas: intragenicstructure reconstructed from the nucleotidesequences of gyrB and rpoD genes. Microbiol.146:2385-2394.

______________________________________________________
CHAPTER 4
F®
R¯
E°
Q±
U²
E°
N³
C´
Yµ
, ¶ D·
I¸V¹
E°
R¯
SºI¸T»
Yµ
, ¶ A¼
N³
D ·
A¼
C´
T»
I¸V¹
I¸T»
Y µ
O½
F ®
2¾
,¶ 4¿ -À
D·
I¸A¼
C´
E°
T»
Yµ
LÁ
PÂ
HÃ
LÁ
O½
R¯
O½
GÄ
LÁ
U²
C´
I¸N³
O½
LÁ
-À PÂ R¯
O½
D·
U²
C´
I¸N³
G Ä
F®
LÁ
U²
O½
R¯
E°
SºC´
E°
N³
T»
PÅ
SÆEÇ
UÈ DÉ
OÊ MË
OÊ NÌ
AÍ
SÆ SÎPPÏÐÏ
. Ñ IÒN Ó
DÔ
UÕ
TÖ
C×
H Ø
TÖ
AÙ
KÚ
EÛ
-Ü AÙ LÝ
L Ý
DÔ
EÛ
C×
LÝ
IÒNÓ
E Û
SÎOÞ
IÒLÝ
SÎ
_______________________________________________________________________________

Cß hà
aá ptâ�ã eä r å 4: æèç
Fé rå eä qê uë eä nì cí yî , ï dð
iñ vò eä rå só iñ tã yî , ï aá nì d ð
aá cí tiã ñ vò iñ tã yî
76
Aô
Bõ Sö T÷
Rø Aô
Cß T÷
_______________________________________________________________________________
Nù
aá tã urë�å aá l ú
só uë ppâûâ rå eä ssó[ó iñ vò eä nì eä ss ó]ó oü f ý
só oü ilñ ú s ó tã o ü tã aá kþ
eä -ÿ aá ll úQú
dð
iñ só eä aá só e oä ü f wý � h
àeä aá tã , ï rå eä f
ýeä rrå?å eä d
ðtã o ü aá s ó tã aá k
þeä -ÿ aá ll
úQúdð
eä cí lúiñ nì eä
(T� ÷
Aô
D), ��� ï oü ccíEí urë�å s ó w� oü rå lú dð w� iñ dð eä . � I� t ã hà
aá s ó b�
eeäEä n ì pâ oü só tã uë lúaá tã eä d
ðtã hà aá t ã d
ðiñ ffýHý eä rå eä nì t mã iñ cí rå oü b
�iñ aá l ú
g� eä nì eä rå a á aá nì dð
m eä cí hà
aá nì iñ só m s ó aá rå e ä rå eä só pâ oü nì só iñ b� lúe ä f
ýoü r å T
÷Aô
D � iñ n ì só oü iñ lú s ó fýrå oü m d
ðiñ ffýGý eä rå eä nì t ã g� eä oü g� rå aá pâ h
àiñ cí aá l
úrå eä g� iñ oü nì só . W� � eä
dð
eä m oü nì só tã rå aá tã eä d ð
tã hà aá t ã fýlúuë oü rå eä só cí eä nì t ã Ps
���e� u� d
�o� m� o� n� a� s
� só ppâûâ . � tã hà aá t pã â rå oü d
ðuë cí e ä tã hà e ä aá nì tiã ñ b� iñ oü tiã ñ c í 2
�,ï 4-æ ÿ
dð
iñ aá cí eä tã yî lúpâ hà
lúoü rå ogü�� l
úuë cí iñ nì oü l
ú(2� �
,ï 4-Dæ ÿ � A
ôP� G� ) � pâ l
úaá y î a á k
þeä y î rå oü l
úe ä iñ n ì tã hà e ä nì aá tã urëûå aá l
úsó uë ppâûâ rå eä ssó[ó iñ vò eä nì eä ss ó[ó oü f
ýtã w� o ü D� uë tã cí h
àT÷
Aô
D � só oü ilñ ú só . � Fé iñ rå só tã , 2,4-Dï � ï æ ÿ � Aô
P� G� -ÿ pâ rå oü dð
uë cí iñ nì g � fýlúuë oü rå eä só cí eä nì t ã Ps
���eu��� d
�o� m� o� n� a� s
� só ppâ�â . w� � eä rå e ä pâ rå eä só eä nì t ã oü n ì rå oü oü tã s ó oü f
ýw� hà
eä aá t ã g� rå oü w� n ì iñ n ì b�
oü tã h à
oü f ý
tã hà e ä T÷
Aô
D � só oü ilñ ú s ó aá t ã dð
eä nì só iñ tiã ñ eä s ó aá t ã oü r å aá b�
oü vò e ä tã hà e ä tã hà rå eä só hà oü lúd dð ð
eä nì só itñ ã y î rå eä qê uë iñ rå eä dð
tã o ü cí oü nì tã rå oü l tú ã aá k
þeä -ÿ aá ll
úQúoü f ý
w� hà
eä aá tã ; � iñ n ì a á cí oü m pâ lúeä m eä nì tã aá rå y î tã aá k
þeä -ÿ aá ll
úQúcí oü nì d
ðuë cí iñ vò e ä só oü ilñ ú , ï pâ oü pâ uë l
úaá tã iñ oü n ì d
ðeä nì só iñ tiã ñ eä só
oü f 2,4-Dý � ï æ ÿ � A
ôP� G� -ÿ pâ rå oü d
ðuë cí iñ nì g � Ps
���eu��� d
�o� m� o� n� a� s
� só ppâ�â . w� � eä rå e ä b
�eä lúoü w � tã hà iñ s ó tã hà rå eä só hà oü l
úd ð
lúeä vò eä l
ú. � Sö eä cí oü nì d
ð,ï
iñ nì tã rå oü dð
uë cí tã iñ oü n ì oü f 2,4-Dý � ï æ ÿ � A
ôP� G� -ÿ pâ rå oü d
ðuë cí iñ nì g � só tã rå aá iñ n ì Sö Sö Bõ 17, � �èï a á rå eä pâ rå eä só eä nì tã aá tiã ñ vò e oä ü f
ýtã hà e ä d
ðoü mi ñ nì aá nì tã
ge� ä nì oü tã yî pâ iñ c í g� rå oü uë p â fýoü uë nì d
ðiñ n ì tã hà e ä D� uë tã cí h
àT÷
Aô
D � só oü ilñ ú só , ï iñ nì tã o ü tã hà e ä tã aá kþ
eä -ÿ aá ll úQú
cí oü nì dð
uë cí iñ vò e ä só oü il ñ ú aá t pã â oü pâ uë lúaá tiã ñ oü nì
dð
eä nì só iñ tã iñ eä s só ó imñ iñ lú aá r å tã o ü tã hà e ä dð
eä nì só itñ ã iñ eä s ó oü f ý
iñ nì dð
iñ ge� ä nì oü uë s ó 2,4-D� ï æ ÿ � A
ôP� G� -ÿ pâ rå oü d
ðuë cí eä rå s ó f
ýoü uë nì d
ðiñ n ì T
÷Aô
D � só oü iñ lú sópâ rå oü vò iñ dð eä d
ðcí oü nì tã rå oü l
úoü f ý
tã aá kþ
eä -ÿ aá ll túQú ã o ü tã hà e ä só aá m e ä l
úeä vò eä l
úaá s ó f
ýoü uë nì d
ðiñ n ì tã hà e ä T
÷Aô
D � só oü ilñ ú . � T÷
hà
iñ rå dð , ï a á m uë tã aá nì t ã oü fý
só tã rå aá iñ n ì SSö ö Bõ 17 � � dð
eä fýiñ cí iñ eä nì t ã iñ n ì 2,4-D
� ï æ ÿ � Aô
P� G� -ÿ pâ rå oü dð
uë cí tiã ñ oü n ì w� aá s ó nì oü t ã aá b�
lúe ä tã o ü cí oü nì tã rå oü l t
ú ã aá kþ
eä -ÿ aá ll úQú
oü f wý � h
àeä aá tã ,ï
iñ nì dð
iñ cí aá tã iñ nì g � tã hà aá t ã 2,4-D� ï æ ÿ � A
ôP� G i� ñ s ó a á k
þeä y î d
ðeä tã eä rå m iñ nì aá nì t iã ñ n ì tã aá k
þeä -ÿ aá ll
úQúsó uë ppâ�â rå eä ssó]ó iñ oü nì . � T
÷hà
eä só e ä rå eä só uë lútã s só ó hà oü w � tã hà aá tã
iñ n ì aá ddð�ð
itñ ã iñ oü n ì tã o ü tã hà e ä pâ hà
yî só iñ cí oü cí hà
eä mi ñ cí aá lúlúy î dð
iñ ffý ý eä rå eä nì t ã T÷
Aô
D � só oü iñ lú s ó fýrå oü m W� aá só hà iñ nì g� tã oü n ì Sö tã aá tã e ä (
�U!
Sö Aô
), 2,4-� ï � ï æ ÿD� Aô
P� G� -ÿ pâ rå oü dð
uë cí iñ nì g � fýlúuë oü rå eä só cí eä nì t ã Ps
���e� u� d
�o� m� o� n� a� s
� só ppâ�â . � aá rå e ä aá l
úsó o ü a á k
þeä y î cí oü m pâ oü nì eä nì t ã oü f
ýtã hà e ä nì aá tã urë�å aá l
úsó uë ppâûâ rå eä ssó[ó iñ vò eä nì eä ss ó]ó f
ýoü uë nì d
ðiñ n ì D� uë tã cí h
àT÷
Aô
D � só oü ilñ ú só . � Fé urë�å tã hà eä rå m oü rå eä , ï it iñ ã ñ s ó tã hà e ä fýiñ rå só t tiã ã ñ m e ä só iñ nì cí e ä tã hà e ä iñ nì itiñ ã ñ aá l
úsó tã uë d
ðiñ eä s ó oü f
ýG� eä rå lú aá g� h
à(�Nù
eä tã hà . � J". � P� l
úaá nì t ã P� aá tã hà oü l
ú. (1968) 74:1-97� � �$#&%&'$�(� æèç � ÿ #$� ) � tã hà aá t ã aá t ã l
úeä aá só t pã â aá rå t ã oü f
ýtã hà e ä m eä cí h
àaá nì iñ só m só
aá nì d ð
mi ñ cí rå ooü ü rå g� aá nì iñ só m s ó tã hà aá t ã oü pâ eä rå aá tã e ä iñ n ì D� uë tã cí h à
T÷
Aô
D � só oü ilñ ú s ó aá rå e ä iñ dð eä nì tã iñ fý iñ eä dð
. � I� n ì só pâ iñ tã e oä ü f ý
qê uë aá nì tã iñ tã aá tiã ñ vò eäsó imilñ ñ ú aá rå itñ ã iñ eä só , 2,4-Dï � ï æ ÿ � A
ôP� G� -ÿ pâ rå oü d
ðuë cí iñ nì g � Ps
���eu��� d
�o� m� o� n� a� s
� só ppâ�â . f� ý
oü uë nì d ð
iñ n ì tã hà e ä D� uë tã cí h à
T÷
Aô
D � só oü ilñ ú s ó aá rå eäge� ä nì oü tã yî pâ iñ cí aá ll
úQúy î dð
iñ ffý ý eä rå eä nì t ã fýrå oü m t ã hà oü só e ä f
ýoü uë nì d
ðiñ n ì T
÷Aô
D � só oü ilñ ú s ó fýrå oü m W� aá só hà iñ nì g� tã oü n ì Sö tã aá tã eä .�
_______________________________________________________________________________
J"oü rå ge � ä T.
÷ � dð
e ä Sö oü uë z) aá , Dï&� aá vò iñ d ð
M*
. W�$� eä llúQú
eä rå , ï aá nì d ð
J"oü s ó M
*. � Rø aaáEá iñ j+ m aá k
þeä rå só
Phytopathology. Accepted.

C, h-
a. pt/10 e2 r 3 4: 465
F7 r3 e2 q8 u9 e2 n: c; y< , = d>
i? v@ e2 r3 sA i? t0 y< , = a. n: d >
a. c; ti0 ? v@ i? t0 y<
77
IB NC TD
ROEGF DH UI
C, TD
IB OF NC
Take-all, caused by the fungus Gaeumannomyces graminis var. tritici, is an important
root disease of wheat worldwide (Asher and Shipton, 1981). Although wheat is particularly
susceptible to the take-all fungus, many other Gramineae can be infected (Shipton, 1975;
Gutteridge et al., 1993). Take-all decline (TAD) is defined as the spontaneous decrease in the
incidence and severity of take-all that occurs with monoculture of wheat or other susceptible host
crops after one or more severe outbreaks of the disease (Cook and Baker, 1983; Hornby, 1983;
Shipton et al., 1973; Shipton, 1975). Soils in the state of TAD are naturally suppressive to the
take-all fungus. The biological basis of the specific suppression associated with TAD has been
demonstrated in a series of experiments: suppressiveness is eliminated by treating the soil with
moist heat (pasteurization, 60ºC for 30 min.), methyl bromide or chloropicrin, and is transferable
by adding small amounts of TAD soil to raw conducive, fumigated or pasteurized soil (Cook and
Rovira, 1976; Cook and Baker, 1983; Raaijmakers and Weller, 1998). The mechanisms
responsible for TAD involve microbiological changes in the bulk soil or rhizosphere environment
resulting in suppression of the pathogen. Repeated introduction of G. graminis var. tritici into soil
also induced suppression of take-all (Gerlagh, 1968; Zogg and Jaggi, 1974; Wildermuth, 1980),
but the relation of this form of suppressiveness to that of TAD is not clear.
The occurrence of TAD throughout the world is remarkable in view of the broad
range of soil types, climates and agronomic conditions under which wheat is cultivated
(Shipton, 1975). Field studies have clearly indicated that the development of TAD follows a
consistent pattern everywhere requiring the presence of three components: monoculture of a
take-all susceptible host, the presence of G. graminis var. tritici, and at least one occurrence of
severe disease. Factors such as soil type and previous cropping history seem only to modulate
the extent and speed of development of TAD (Shipton, 1975). Previous work on TAD has
suggested that different microbial genera and mechanisms are responsible for suppressiveness in
soils from different geographical regions. First, TAD develops in multiple agro-ecosystems
(Shipton, 1975; Brown, 1981). Second, the length of time for TAD to develop varies among
fields and the crop grown in monoculture (Shipton, 1975). Finally, antagonists from many
different taxonomic groups have been isolated from TAD soils and, when introduced into take-all
conducive soils, provide a certain level of control of take-all (Deacon, 1976; Shipton, 1975;
Hornby, 1983; Wong and Baker, 1984; Aberra et al., 1998; Weller et al., 1988). Despite the
wide range of microorganisms implicated in TAD, several of them do not fit one or more of the
biological properties of TAD (Cook, 1985). For example, the sensitivity of the TAD factor(s)
to pasteurization with moist heat (60°C, 30 min) rules out the involvement of heat-resistant,

CJ hK
aL ptM1N eO r P 4: Q6R
FS rP eO qT uU eO nV cW yX , Y dZ
i[ v\ eO rP s] i[ tN yX , Y aL nV d Z
aL cW tiN [ v\ i[ tN yX
78
spore-forming bacteria like Bacillus spp. and probably also actinomycetes (Cook and Baker,
1983).
Among the antagonistic bacteria, the fluorescent Pseudomonas spp. have been implicated
in TAD soils throughout the world (Cook and Rovira, 1976; Smiley, 1978; Weller et al., 1988;
Sarniguet et al., 1992; Raaijmakers and Weller, 1998). Characteristics that suggest involvement of
pseudomonads in TAD include: they are well adapted to the rhizosphere environment and
produce a variety of secondary metabolites, including antibiotics and siderophores, that inhibit
the growth of G. graminis var. tritici (Weller et al., 1988; Thomashow and Weller, 1990), their
population densities increase considerably on roots with take-all lesions (Weller, 1983; Smiley,
1978; 1979; Brown, 1981; Sarniguet et al., 1992; Barnett et al., 1999; Charigkapakorn and
Sivasithamparam, 1987), and they are eliminated by soil pasteurization, which also eliminates
specific suppression (Cook and Baker, 1983; Raaijmakers and Weller, 1998). Recent studies
demonstrated that fluorescent Pseudomonas spp. producing the antibiotic 2,4-
diacetylphloroglucinol (2,4-DAPG) play a key role in the suppressiveness of TAD soils in
Washington State (Raaijmakers et al., 1997; Raaijmakers and Weller, 1998, 1999). Using specific
primers and probes directed against sequences within the biosynthetic locus of 2,4-DAPG have
clearly indicated that fluorescent Pseudomonas spp. producing this antifungal metabolite are
highly enriched in TAD soils from Washington State (Raaijmakers et al., 1997). Elimination of
2,4-DAPG producers resulted in loss of suppressiveness (Raaijmakers and Weller, 1998). When a
2,4-DAPG-producing strain was introduced via seed treatment into a take-all conducive soil,
it provided control of take-all of wheat to levels similar to that obtained in the complementary
TAD soil (Raaijmakers and Weller, 1998).
One key question has been whether 2,4-DAPG-producing Pseudomonas spp. also play a
key role in the specific suppression found in TAD soils from other geographical regions.
Pseudomonas spp. harboring phlD (phlD+), one of the key genes in the biosynthesis of 2,4-DAPG,
were found on roots of wheat grown in other soils with a history of wheat monoculture
(McSpadden-Gardener et al., 2000), however, the level of take-all suppressiveness of these soils
and the role of 2,4-DAPG in take-all suppression were not addressed. In this study, we
determined the frequency, diversity and activity of 2,4-DAPG-producing Pseudomonas spp.
found in Dutch polder soils with a history of wheat monoculture. Dutch polder soils were
among the first TAD soils reported (Gerlagh, 1968), however, the exact mechanisms operating
in these soils have not been elucidated. Phenotypic and genotypic analyses were performed to
characterize 2,4-DAPG-producing Pseudomonas isolates obtained from Dutch soils. The
biocontrol efficacy and rhizosphere competence of P. fluorescens SSB17, a representative of the

Ch_
a` pta1b ec r d 4: e6f
Fg rd ec qh ui ec nj ck yl , m dn
io vp ec rd sq io tb yl , m a` nj d n
a` ck tib o vp io tb yl
79
major genotypic group of 2,4-DAPG-producers found in a Dutch soil, were compared to that
of Q8r1-96, a representative of the major genotypic group of 2,4-DAPG-producers found in
TAD soils from Washington State (Raaijmakers and Weller, 2001). The role of 2,4-DAPG in
control of take-all and in rhizosphere competence was determined for both strains by
generating mutants deficient in 2,4-DAPG production. Finally, we determined whether
populations of 2,4-DAPG-producing Pseudomonas spp. are specifically enriched upon
infection of wheat roots by the take-all fungus.
Rr Es
St Uu
Lv Tw
St
Nx
a` tb uri1d a` l y
sq ui ppa1a rd ec ssqzq io vp ec nj ec ss qzq o{ f |
sq o{ ilo y s q tb o { G} . ~ g� r� a� m� i� n� i� s� vp a` rd . � t� r� i� t� ici����� .� Characteristics of soils CB, SV, and
SSB are shown in Table 1. Physical and chemical properties of these soils differ from TAD
soils from Washington State (Raaijmakers et al., 1997). Major differences are that the Dutch
soils have lower levels of nitrate, phosphorus, potassium, and silt content, and higher levels of
organic matter and clay content compared to the four soils from Washington State. The three
Dutch soils were tested in bioassays for suppressiveness to G. graminis var. tritici. When these
soils were inoculated with G. graminis var. tritici, the disease index was significantly lower on
roots of plants grown in SV and SSB soils as compared to plants grown in CB soil (Fig. 1). No
take-all symptoms were found on roots of wheat grown in soils not inoculated with G.
graminis var. tritici (data not shown). Based on these results, and the occurrence of long term
wheat monoculture in the SV and SSB soils, they were classified as TAD soils, whereas CB soil
was regarded as a G. graminis var. tritici -conducive soil.
Fg rd ec qh ui ec nj ck y l a` nj d dn�n
io vp ec rd sq io tb y l o{ f 2,4-D|G� m e&��� A
�P� G� -
�pa rd o{ d
nui ck io nj g � Ps
� � eu��� d�
o� m� o� n� a� s� sq ppa1a . � io n j D�
ui tb ck h _
Tw
A�
D �
sq o{ ilo y sq . � In
CB soil, wheat rhizosphere population densities of phlD+ Pseudomonas spp. were relatively low
and ranged from less than 104 to 2.7 x 104 CFU g-1 root (Table 2). In the SV and SSB soils,
populations of phlD+ fluorescent Pseudomonas spp. ranged from 5.1 x 104 to 1.4 x 106 CFU g-1
root (Table 2). These populations represented approximately 5 to 14% of the total population
of fluorescent pseudomonads isolated from wheat roots and were, on average, 5 to 52-fold
higher than populations of phlD+ fluorescent Pseudomonas spp. found on roots of wheat
grown in the CB soil. These results demonstrate a correlation between relatively high
populations of phlD+ fluorescent Pseudomonas spp. and suppressiveness to take-all of wheat.
Analysis of the genotypic diversity of the isolated phlD+ fluorescent Pseudomonas spp.
by RAPD analysis with primers M13, M12 and D7 generated a total of 58, 56 and 48

C� h�
a� pt�1� e� r � 4: 6¡
F¢ r� e� q£ u¤ e� n¥ c¦ y§ , ¨ d©
iª v« e� r� s¬ iª t� y§ , ¨ a� n¥ d ©
a� c¦ ti� ª v« iª t� y§
80
consistent RAPD markers, respectively, ranging in size from 190 to 2070 bp. Reproducibility
of the RAPD amplifications was confirmed in three independent experiments and the
reliability of the dendrogram topology (Fig. 2) was evaluated by bootstrap analysis using 1000
resampled data sets. Among a total of 161 isolates of phlD+ fluorescent Pseudomonas spp.
obtained from roots of wheat grown in SV and SSB soil, 33 unique groups were identified and
only one RAPD group, represented by isolates SSB14 and SV7, was found in both soils (Fig.
2; Table 3). The major genotypic groups found on roots of wheat grown in SV and SSB soils
represented on average 40 and 26%, respectively, of the total number of phlD+ fluorescent
Pseudomonas spp. isolates from SV and SSB soils (Table 3). The major genotypic group (SV8)
found in SV soil clustered very distantly from the major genotypic group (SSB17) found in
SSB soil (Fig. 2). Isolates from SV and SSB soils clustered distantly from 2,4-DAPG-
producing reference strains CHA0, Pf5 and Q8r1-96, and from 2,4-DAPG-nonproducing
isolates (groups SV12, SV13, and SSB24) obtained from SV and SSB soils (Fig. 2). The
Shanon-Weaver’s diversity index was higher for populations of phlD+ fluorescent Pseudomonas
spp. isolated from SSB soil than for those from SV soil (Table 3), indicating a higher level of
genotypic diversity in SSB soil.
In addition to genotypic clustering by RAPD analysis, also data from plate inhibition
assays were used to phenotypically group the isolates on the basis of their capacity to inhibit G.
graminis var. tritici and R. solani. Based on G. graminis var. tritici mycelial inhibition, a total of
seven clusters was obtained. Five of these clusters (I to V) contained isolated 2,4-DAPG
producers and reference strains Pf-5, CHA0, and Q8r1-96, and two clusters (VI and VII)
contained isolates that did not produce 2,4-DAPG. Thirteen clusters were obtained based on
inhibition of mycelial growth of R. solani. Clusters I to XI contained isolated 2,4-DAPG
producers and reference strains and clusters XII and XIII contained non-2,4-DAPG producers
(Fig. 3). Comparison of the cluster analyses indicates that 2,4-DAPG-producing isolates are
more inhibitory in vitro to G. graminis var. tritici than to R. solani (Fig. 3). The mean
inhibition of mycelial growth by phlD+ fluorescent Pseudomonas spp. isolates and 2,4-DAPG-
producing reference strains ranged from approximately 35 to 58% for G. graminis var. tritici
and from approximately 11 to 43% for R. solani. For 2,4-DAPG non-producing isolates,
inhibition of G. graminis var. tritici ranged from 0 to 30% and for R. solani inhibition ranged
from 0 to 20%. Most phenotypic groups contained more than one RAPD group and isolates
belonging to the same RAPD group (based on 100% similarity) were always grouped in the
same phenotypic group (Fig. 2; Fig. 3). Strain SSB17, which is a representative of the major
RAPD group found on roots of wheat grown in SSB soil (Fig. 2; Table 3), clustered in the

C h®
a¯ pt°1± e² r ³ 4: ´6µ
F¶ r³ e² q· ue² n¹ cº y» , ¼ d½
i¾ v¿ e² r³ sÀ i¾ t± y» , ¼ a¯ n¹ d ½
a¯ cº ti± ¾ v¿ i¾ t± y»
81
phenotypic group that was most inhibitory to G. graminis var. tritici (Fig. 3) and was selected
for further studies. Strain Q8r1-96, which represents the major RAPD group among 2,4-
DAPG-producing Pseudomonas spp. isolated from wheat grown in the Quincy TAD soil from
Washington State (USA) (Raaijmakers and Weller, 2001) was included for comparison
purposes. Q8r1-96 clustered in the same G. graminis var. tritici -phenotypic group as SSB17
(Fig. 3), but is genotypically different (Fig. 2).
RÁ o lÃe o²G f 2,4-D
ÄÆÅ ¼ ´&Ç�È AÉ
PÊ GË -Çp° r³ o d
½ucº i¾ n¹ g Ì Ps
Í�ÎeuÏ�Ð d
ÑoÒ mÓ oÒ nÔ aÕ s
Î sÀ pp°Ö° . × i¾ n ¹ D
Èut± cº h
®TØ
AÉ
D È
sÀ o il¾ à sÀ . × If 2,4-DAPG-producing
fluorescent Pseudomonas spp. play a determinative role in the natural suppressiveness of Dutch
TAD soils, then introduction of a 2,4-DAPG-producing strain into the conducive soil should
give suppressiveness to G. graminis var. tritici to a level similar to that obtained in the
complementary TAD soil. Furthermore, if 2,4-DAPG is a key metabolite in suppression of G.
graminis var. tritici then 2,4-DAPG-deficient mutants should have a reduced ability to
suppress take-all of wheat. Tn5 mutagenesis of P. fluorescens strains SSB17 and Q8r1-96
generated 1078 and 686 transformants, respectively. For strains SSB17 and Q8r1-96, 4 and 2
transformants, respectively, did not produce the red pigment characteristic of 2,4-DAPG
producers. RAPD analysis confirmed strain integrity of all 6 mutants and HPLC analysis
showed that all 6 mutants were defective in 2,4-DAPG production. In plate inhibition assays
wildtype strains SSB17 and Q8r1-96 caused clear inhibition of mycelial growth of G. graminis
var. tritici, R. solani, and P. ultimum var. sporangiiferum, whereas the mutants caused no
inhibition of these pathogens (data not shown). Southern blot analysis revealed that all
mutants had a single transposon insertion. Mutants 9H4, derived from SSB17, and 4C5,
derived from Q8r1-96, were selected for the bioassays.
Strain SSB17 and its 2,4-DAPG-deficient mutant 9H4 were introduced via seed
treatment into the take-all conducive CB soil, amended with G. graminis var. tritici inoculum,
at initial densities of approximately 103 CFU seed-1. SSB17 and 9H4 established rhizosphere
population densities of 5.0 x 106 and 9.0 x 106 CFU g-1 root, respectively, after 4 weeks of
plant growth. Population densities of the wildtype and mutant were not significantly different
and were similar to the density of indigenous 2,4-DAPG-producers (5.0 x 106 CFU g-1 root)
found on roots of wheat grown in the SSB soil amended with G. graminis var. tritici inoculum.
Treatment of seeds with strain SSB17 significantly reduced take-all severity and provided
control to the same level as found in the SSB TAD soil (Fig. 4A). Mutant 9H4 did not reduce
disease severity caused by G. graminis var. tritici (Fig. 4A). These results provide further
evidence that phlD+ fluorescent Pseudomonas spp. are a key component of the suppressiveness

82
T
Ù
a ÚbÛlÜe Ý 1.
Þàß C
á
r âoãpäpäiånæg ç hèiåsétêoãr ây ë, ì pähèy ëséiåcíoãc íhèeÝmîiåc ía Úl Ü pär âoãpäeÝrât êiåeÝs é oãf ï têhèe Ý séoãiålÜs é uðséeÝd
ñ
i
å
n æ têhèiås sé ét êuðd ñy ë.ß
Soil Cropp ing h istory Wheatcultivar
pH a Content (µg g -1 of soil )NO3-N NH4-N Na P K
Content (%) OM b Sand Silt Clay
CB Rotation wheat/sugar beet Bussard 7.6 3 2 14 1 74 4.4 49 12 27SV 14 years continuous wheat Monopol 7.4 4 2 10 2 74 4.4 52 9 27SSB 27 years continuous wheat Bussard 7.5 3 2 9 2 89 4.4 50 10 28
a pH was determined after extraction with CaCl2.b Organic matter.
T
ò
a óbôlõe ö 2.
÷àø F
ù
rúeöq ûuüeönýcþy ÿ o�f � p�h �l �D �
+ � f �l�uor �e�s c�e�n�t � P �s �e �u �d �o �m�o�n�a �s � s p�p�. � i�n � t�h�e � w �h�e�a �t � r�h�i�z os p�h�e�r�e�.�
Soil First experimentTotal a phlD+ b % phlD+ c
fluorescent fluorescent fluorescentPseudomonas spp. Pseudomonas spp. Pseudomonas spp.
Second experimentTotal a phlD+ b % phlD+ c
fluorescent fluorescent fluorescentPseudomonas spp. Pseudomonas spp. Pseudomonas spp.
CB 2.7 X 105 < 104 < 0.1 1.2 X 106 2.7 X 104 2.3SV 5.7 X 105 5.1 X 104 9.0 4.5 X 106 2.1 X 105 4.8SSB 1.1 X 106 1.1 X 105 14.0 1.1 X 107 1.0 X 106 12.7
a Total population of fluorescent Pseudomonas spp. (CFU g-1 root).b Population of fluorescent Pseudomonas spp. harboring the phlD gene (CFU g-1 root).c Percentage of fluorescent Pseudomonas spp. harboring the phlD gene in relation to the total fluorescent Pseudomonas population.
C!h "a #pt $&%e 'r ( 4: )+*F, r( e 'q -u. e 'n/ c 0y 1, 2 d 3i4 v 5e 'r( s 6i4 t %y 1, 2 a #n/ d 3
a #c 0ti %4 v 5i4 t %y 1

C7 h8
a9 pt:&; e< r = 4: >+?
F@ r= e< qA uB e< nC cD yE , F dG
iH vI e< r= sJ iH t; yE , F a9 nC d G
a9 cD ti; H vI iH t; yE
83
FK
iLgM uN rO e P 1. QSR N
TaU tV uN rO aU l W sX uN pY pY rO eP sX sX iL vZ eP n[ eP sX s X o\ f
]sX o\ iLlWs X C^
B_
, ` Sa
Vb
, ` aU n[ d c
SSada
B _
tV o \ Ge . f gg rh ai mj iknl iksm vn ao rp . q tr rh ik tr ik cs ik .q Soils were
amended with 0.1% of an oat grain inoculum of G. graminis var. tritici. Plants were grownfor one month under controlled conditions and disease caused by G. graminis var. tritici wasscored on a 0 to 8 scale, where 0 indicates no disease and 8 indicates dead plant. Means offive replicates are shown. Means with the same letter are not statistically different accordingto Tukey’s studentized range test (P = 0.05). Error bars represent the standard error of themean. The experiment was repeated 4 times and representative results are shown.
that operates in the SSB TAD soil and that 2,4-DAPG is the major determinant of take-all
suppression by strain SSB17. In independent experiments, similar results were found for strain
Q8r1-96 and its 2,4-DAPG deficient mutant 4C5 (Fig. 4B). Although there were no
differences between strains SSB17 and Q8r1-96 in their ability to suppress take-all, there was a
significant difference in their rhizosphere competence (Fig. 5). In spite of the fact that strain
Q8r1-96 was isolated from a different wheat cultivar grown in a physicochemically different
soil from another geographical region, it was more rhizosphere competent than strain
SSB17. Dose-response studies showed that strain Q8r1-96 required lower initial densities on
seeds than strain SSB17 to reach the same final population on roots (Fig. 5). When comparing
dose-response kinetics of the wildtype strains SSB17 and Q8r1-96 with those obtained for
their respective 2,4-DAPG-deficient mutants, 9H4 and 4C5, no differences were found (Fig.
5). These results suggest that 2,4-DAPG production does not contribute to the rhizosphere
competence of strains SSB17 and Q8r1-96.
Et
ffuvu
ew cx t y oz f u
ty a{ k|
ew -} a{ ll ~�~
a{ n� d �
R�
h�
i� zo��� c� t� o� n� i� a � r� ooz�z t y r� oz t y oz n � r� h� i� z� oz s� p� h�
ew r� e w p� oz p� u� l~a{ tiy � oz n � d
�ew n� s� i� ty i� ew s � oz f 2,4-D
u��+� � }�� A�
P� G� -}p� r� oz d
�u� cx i� n� g � f
ul~u� oz r� ew s� cx ew n� t y Ps
� �eu¡£¢ d¤o� m¥ o� n� a� s
� s� pp�&� .¦ The influence of G. graminis var. tritici and R. solani
infections on wheat root colonization by indigenous and introduced phlD+ fluorescent
Pseudomonas spp. was studied. Population densities of indigenous pseudomonads and phlD+
fluorescent Pseudomonas spp. were assessed on healthy and G. graminis var. tritici- or R. solani-
infected roots of wheat grown in SV and SSB soils, and in more detail on non-
CB SV SSBD
isea
se in
dex
0
1
2
3
4
5a
bb

C§ h¨
a© ptª&« e¬ r 4: ®+¯
F° r e¬ q± u² e¬ n³ c´ yµ , ¶ d·
iv¹ e¬ r sº it« yµ , ¶ a© n³ d ·
a© c´ ti« ¸ v¹ it« yµ
84
F»
i¼g½ u¾ r¿ e À 2.
ÁÃÂDÄ
eÀ nÅ dÆ
r¿ oÇ g½ r¿ aÈ m É oÇ f Ê
pË hÌlÍDÎ
+ Ï fÊlÐu¾ oÇ r¿ eÀ sÑ cÒ eÀ nÅ t Ó P
ÔsÕ eÖ u× d
ØoÙ mÚ oÙ nÛ aÜ sÕ sÑ ppÝÞÝ .
ÂsÑ tÓ r¿ aÈ i¼ nÅ s Ñ b
ßaÈ sÑ eÀ d
ÆoÇ n Å R
àAá
Pâ
D Ä
mÉ aÈ r¿ kã eÀ r¿ sÑ . Thepairwise coefficients of similarity (Nei/Li distances) were clustered using the neighbour-joining method. On roots of wheat grown in soil, SV, 11 (SV1 to SV11) differentgenotypic groups of phlD+ isolates were found on the basis of 100% similarity. In soil SSB,23 (SSB1 to SSB23) different groups were found. Each branch represents a uniquegenotypic group. Isolates followed by a “-“ are phlD- fluorescent Pseudomonas spp.Numbers between brackets represent the additional number of isolates in the RAPD group.Asteriscs indicate the major genotypic groups in SV and SSB soils. Bootstrap values werecalculated for 1000 resampled data sets and only values equal or higher than 80% areshown.
lesioned and lesioned root sections of G. graminis var. tritici- or R. solani-infected roots (Tables
4, and 5). When roots of wheat grown in SV and SSB soils were infected with G. graminis var.
tritici, or R. solani, densities of phlD+ fluorescent Pseudomonas spp. on the whole root systems
increased significantly by 5 to 8-fold. Identical increases were observed for the total fluorescent
Pseudomonas population on G. graminis var. tritici -infected roots and on roots infected with
R. solani. When population densities of both total and phlD+ fluorescent Pseudomonas spp.
CHA0Pf-5
100
100
100
100
100100
100
10% sim ilarity SSB17[+28]**SSB1SSB15[+1]
89
SSB19[+4]SSB5[+2]
SSB4[+1]SSB2SV2[+3]
SSB8[+1]SSB16[+10]
SSB9
SV6[+2]
SV7[+2]SSB3[+3]
SSB18[+5]
SSB12
SSB20[+1]
SSB13SSB6[+3]
SV3SV5
SSB14[+2]
SV12-SSB7
SV1[+8]
9498
CB2-SV13-
SSB24-
CB1[+1]-SV4[+1]
SSB23[+5]SSB22[+7]
SV8[+18]**
SSB21[+14]SSB10
SV10[+1]
SV11SV9[+2]
80
82

Cä hå
aæ ptç&è eé r ê 4: ë+ì
Fí rê eé qî uï eé nð cñ yò , ó dô
iõ vö eé rê s÷ iõ tè yò , ó aæ nð d ô
aæ cñ tiè õ vö iõ tè yò
85
Fø
iùgú uû rü e ý 3.
þÃÿC�
l�uû s� t� eý r ü a� n� a� l� y� s� iù s � o� f
�in v� i t� r o � i
ùn� h�
iùb�
iùt� iù o� n � o� f
�G� . � g� r a� m� i
n i s� v� a� rü . ÿ t� r i t� i c� i (� A� )
�a� n� d �
R�
. � s� o� l� a� n i (� B� ) �
b�
y�p h!l�D"
+ # f�l�uû o� rü eý s� c$ eý n� t � P
%s� e& u' d( o� m� o� n a� s� s� p) p) .
ÿ Hierarchical, single linkage cluster analysis was
computed using the mean inhibition of mycelial growth on plates given by a certainPseudomonas isolate when compared to the control (the fungus only). Numbers betweenbrackets represent the number of additional isolates in the cluster. Asteriscs indicate themajor genotypic groups in SV and SSB soils. Isolates followed by a “-“ are phlD-fluorescent Pseudomonas spp. Mean refers to the mean percentage inhibition relative tothe control.
0 5 10 15 20 25
Percentage s im ilarity
Isolates M ea n
58.0058.00
57.14
56.6756.7955.9556.0056.1055.9555.3654.7653.9748.8048.8150.7951.4950.0050.0050.2544.0544.0043.75
43.5944.0546.0345.6446.4345.24
45.0045.2440.0040.0039.2937.2036.9035.71
34.52
30.00
0.000.001.192.38
ASSB17[+28]**
SSB10Pf-5SV1[+8]
SSB8[+1]
SSB22[+7]SSB7
SSB5[+2]
SSB18[+5]SSB23[+5]
SSB21[+14]
CHA0Q8r1-96
SSB14[+2]SV10[+1]
SV2[+3]
SV11SV7[+2]
SSB11[+3]
SSB6[+3]SV9[+2]
SSB19[+4]SV5SV6[+2]
SSB1
SV4[+1]
SSB9
SSB3[+3]
SV3
SV12-
CB1[+1]-SV13-SSB24-
SSB20[+1]SSB15[+1]
CB2-
SSB13
SSB4[+1]
SSB2
SSB16[+10]
I
II
III
IV
V
VI
VII
0 5 10 15 20 25
Percentage s im ilarity
Isolates M ean
SSB17[+28]**
SSB10
Pf-5
SV1[+8]
SSB8[+1]
SSB22[+7]
SSB7
SSB5[+2]
SSB18[+5]
SSB23[+5]
SSB21[+14]
CHA0
Q8r1-96
SSB14[+2]
SV10[+1]
SV2[+3]
SV11
SV7[+2]
SSB11[+3]
SV8[+18]**
SSB6[+3]
SV9[+2]
SSB19[+4]
SV5
SV6[+2]
SSB1
SV4[+1]
SSB9
SSB3[+3]
SV3
SV12-
CB1[+1]-
SV13-
SSB24-
SSB20[+1]
SSB15[+1]
CB2-
SSB13
SSB4[+1]
SSB2
SSB16[+10]
II
I
IV
V
VI
VII
VIII
IX
X
XI
XII
XIII
B

C* h+
a, pt-/. e0 r 1 4: 243
F5 r1 e0 q6 u7 e0 n8 c9 y: , ; d<
i= v> e0 r1 s? i= t. y: , ; a, n8 d <
a, c9 ti. = v> i= t. y:
86
T@
aA bB lCe D 3. EGF
CH
lCuI sJ tK eD rL iM nN g O oP f
QpR hSlTDU
+ V fQlCuI oP rL eD sJ cW eD nN t K P
XsY eZ u[ d\ o] mo] n_ a` sY sJ ppaba . b
F By c Rd
Ae
Pf
D g
aA nN aA lC yc sJ iM sJ ah .F
Origin/type of isolate Number ofisolates
RAPD groups b Major groups c
(%)Shanon-Weaverindex d
SV soil 48 11 39.6 (SV8) 32.2SSB soil 113 23 25.7 (SSB17) 84.2SV and SSB soils e 6 1 - -Non-2,4-DAPG producers f 6 5 - -Reference strains g 3 3 - -
a phlD+ isolates were obtained from roots of wheat grown in SV and SSB soils.b RAPD groups were defined with primers D7, M12, and M13 on the basis of 100% similarity.c Major group refers to the percentage of the total number of phlD+ fluorescent Pseudomonas represented by a
single dominant genotype.d Calculated by using the formula H=∑[pi-ln(pi)], where pi is the proportion of each RAPD group (based on
100% similarity) in relation to the total number of RAPD groups per soil.e Present in both, SV and SSB soils.f phlD- isolates include CB1, CB2, CB3, SV12, SV13, and SSB24 and were obtained from roots of wheat grown
in CB, SV and SSB soils.g Reference strains include P. fluorescens strains CHA0, Pf-5, and Q8r1-96.
were compared in more detail on lesioned and non-lesioned root sections of infected roots, G.
graminis var. tritici infection led to increases ranging from 4 to 15-fold. Again, no significant
differences were found between increases in population densities of total and phlD+
fluorescent Pseudomonas spp. for the SV soil (P = 0.60) and SSB soil (P = 0.60). Population
densities of phlD+ fluorescent Pseudomonas spp. were up to 6 fold greater on R. solani-lesioned
root sections than on non-lesioned root sections. These increases were not statistically
significant different from the increases observed on G. graminis var. tritici -lesioned root
sections for both SV and SSB soils (P = 0.26 and P = 0.47, respectively).
The effect of G. graminis var. tritici on root colonization was also determined for P.
fluorescens strains SSB17, Q8r1-96, and their respective 2,4-DAPG-defective mutants, 9H4
and 4C5, all of which were introduced at two different initial densities (Table 6). After 3
weeks of plant growth, rhizosphere population densities of the wildtype strains and their
respective mutants were not significantly different in presence or in absence of G. graminis var.
tritici. Population densities of SSB17 and 9H4 were significantly lower than population
densities of Q8r1-96 and 4C5, irrespective of the presence of G. graminis var. tritici. For both
initial densities of the introduced strains, infections of wheat roots by G. graminis var. tritici
did not lead to significant increases in their population densities.

Ci hj
ak ptl/m en r o 4: p4q
Fr ro en qs ut en nu cv yw , x dy
iz v{ en ro s| iz tm yw , x ak nu d y
ak cv tim z v{ iz tm yw
87
F}
i~g� u� r� e � 4.
�G� E�
ff���
e� c� t � o� f �
P�
. � f�l�u� o� r� e� s� c� e� n� s� s� t� r� a� i~ n� s � SS
���B�
17, ��� � a� n� d ¡
i~t� s � 2,4¢ � � -£ D¤ A
¥P¦
G§
-£ d¡ e� f� i~ c� i~ e� n� t � m¨ u� t� a� n� t� , 9�ª© H«
4, � � (¬ A¥ )
a� n� d¡
Q®
8¯
r� 1� -£ 96, © ° � a� n� d ¡
i~t� s � 2
¢,� 4� -£ D¤ A
¥P¦
G§
-£ d¡ e� f� i~ c� i~ e� n� t � m¨ u� t� a� n� t� , 4� � C±
5, ²³� (¬ B� )
o� n � i~n� f�e� c� t� i~ o� n � o� f
�w´ hµ
e� a� t � b¶
y · G.�g¹ r� aº m» i
¼n� i¼ s� v½ a� r� . � t¾ r� i¼ t¾ i¼ c� i¼ .� Seeds treated with bacterial strains at densities of approximately 103
CFU seed-1 were sown in CB soil amended with G. graminis var. tritici inoculum.Untreated seeds sown in CB and SSB soils served as controls. Plants were grown for onemonth and the severity of take-all was assessed on a 0 to 8 scale. Means of five replicates areshown. Means with the same letter are not statistically different according to Tukey’sstudentized range test (P = 0.05). Error bars represent standard errors of the means. Theexperiment was repeated 2 times and representative results are shown.
D¿ IÀ SÁ CUiÃÂ SSÁÄÁ IÀ OÅ NÆ
The results of this study show that 2,4-DAPG-producing Pseudomonas spp. play a key
role in the natural suppressiveness of two Dutch TAD soils. First, 2,4-DAPG-producing
Pseudomonas spp. were present on roots of wheat grown in both TAD soils at densities up to
approximately 106 CFU g-1 root (Table 2), densities that are equal or higher than the threshold
density (105 CFU g-1 root) of 2,4-DAPG-producing fluorescent Pseudomonas strains required
to control take-all (Raaijmakers and Weller, 1998). In the complementary take-all conducive
soil, population densities of 2,4-DAPG-producing fluorescent Pseudomonas spp. were below
this threshold density. Second, introduction of 2,4-DAPG-producing strain SSB17,
representative of the dominant genotypic group found in one of the Dutch TAD soils, into
the take-all conducive soil at rhizosphere population densities similar to the densities of
indigenous 2,4-DAPG-producing fluorescent Pseudomonas spp. found in TAD soils, provided
control of take-all to the same level as found in the complementary TAD soil (Fig. 4). Third, a
mutant of SSB17 deficient in 2,4-DAPG-production was not able to control take-all of wheat,
indicating that 2,4-DAPG is a key determinant in take-all suppression (Fig. 4). Previously the
CB
CB
+ S
SB
17
CB
+ 9
H4
SS
B
Dis
ease
Ind
ex
0
1
2
3
4
5
a
a
bb
A
b
B
CB
CB
+ Q
8r1-
96
CB
+ 4
C5
SS
B
Dis
ease
ind
ex
0
1
2
3
4
5a
b
a
b

CÇ hÈ
aÉ ptÊ/Ë eÌ r Í 4: Î4Ï
FÐ rÍ eÌ qÑ uÒ eÌ nÓ cÔ yÕ , Ö d×
iØ vÙ eÌ rÍ sÚ iØ tË yÕ , Ö aÉ nÓ d ×
aÉ cÔ tiË Ø vÙ iØ tË yÕ
88
role of 2,4-DAPG-producing fluorescent Pseudomonas spp. in TAD soils had been
demonstrated only in Washington State (USA). Gerlagh (1968) postulated from his elegant
and classic studies of TAD in Dutch polders that specific suppression involves antibiotic
production by soil microorganisms. This report identifies for the first time, at least in part, the
mechanism and microorganisms that contribute to the natural suppressiveness of Dutch TAD
soils.
The densities of indigenous 2,4-DAPG-producing Pseudomonas spp. found on roots of
wheat grown in the Dutch TAD soils were quantitatively similar to the densities found in four
TAD soils from Washington State (USA) (Raaijmakers et al., 1997; Raaijmakers and Weller,
1998). However, the genotypes of 2,4-DAPG-producers responsible for TAD in the Dutch
soils differed from those in Washington State TAD soils (McSpadden-Gardener et al., 2000;
Raaijmakers and Weller, 2001). For example, Raaijmakers and Weller (2001) identified 16
RAPD groups among 101 phlD+ isolates obtained from roots of wheat grown in Quincy,
Washington TAD soil, but one group comprised 50% of the isolates. This genotype also was
dominant on roots of wheat grown in Moses Lake and Lind, Washington TAD soils and
belonged to BOX-PCR group D as defined by McSpadden et al. (2000). In the current study,
33 distinct RAPD groups were identified among 161 phlD+ isolates from the two Dutch soils,
but the SV and SSB soils had different dominant groups which represented 40% and 26%,
respectively, of the isolates. All of the Dutch isolates clustered distantly from Q8r1-96, a
representative of the dominant phlD+ isolates from the three Washington TAD soils.
Furthermore, the dominant Dutch isolates corresponded to BOX-PCR groups M and F,
which do not occur in Washington TAD soils.
The Shanon-Weaver’s index indicated that the diversity among 2,4-DAPG producers
in the SSB soil was higher than in the SV soil (Table 3). This was unexpected since we had
hypothesized that the longer history of wheat monoculture in the SSB soil as compared to SV
soil, would have enriched for a narrower range of genotypes. One explanation for the
difference in diversity index between the two soils, may be the fact that different wheat
cultivars were grown in these soils under field conditions. The wheat cultivar Bussard, used in
the experiments described in this study, was also the cultivar grown in SSB soil in the field,
whereas cv. Monopol was grown successively in the SV soil under field conditions. The
influence of the host plant and cultivar on the composition of the microflora is well
established for several systems. For example, certain wheat cultivars select for specific
populations of P. putida that are antagonistic to the causal agents of apple replant disease
(Mazzola and Gu, 2000), and different tomato genotypes provide better support for the

C
Û
h
Ü
a ÝptÞ ßeàr á 4:
â�ã F
ä
r áeàq åuæeànçc èy é, ê d ëiìv íeàr ásîiìt ßy é, ê a Ýnçd
ë
a Ýcètiß ìv íiìtßy é
89
T
ï
a ðbñlòe ó 4.
ôªõ I
ö
n÷f ølòuùeón÷c úe ó oûf øG ü
. ý g þr ÿa �m�i �n�i �s � v �a �r �. � t r ÿi �t i �c i � o�n � w h�e�a �t � r �o�o�t � c�o�l�o�n�i�z �a �t �i�o�n � b�y � i�n�d�i�g �e�n�o�u�s � f �l�u�o�r �e�s�c�e�n�t � P �s �e �u �d �o m�o n�a �s � s�pp! !.�
Healthy X diseased roots a Non-lesioned X lesioned root sections of diseased plants b
Total pseudomonads c phlD+ d Total pseudomonads c phlD+ d
Soil -Ggt +Ggt Increase e - Ggt + Ggt Increase e -Ggt +Ggt Increase e -Ggt +Ggt Increase e
SV 3.9 X 106 3.4 X 107 * 8.7 7.6 X 105 3.7 X 106 * 5.0 5.6 X 106 5.6 X 107 10.1 4.9 X 105 4.8 X 106 9.7SSB 8.1 X 106 4.9 X 107 * 6.1 7.3 X 105 4.3 X 106 * 6.0 8.8 X 106 3.8 X 107 * 4.3 2.7 X 105 4.0 X 106 * 14.8
a Roots of wheat growing in CB soil non-amended (- Ggt) or amended (+ Ggt) with G. graminis var. tritici.b Non-lesioned and lesioned root sections of plants infected by G. graminis var. tritici.c Populations of total fluorescent Pseudomonas spp. (CFU g-1 root).d Population of phlD+ (CFU g-1 root). Mean values of five replicates are presented. Means marked with an asterisc (two means in a row) are significantly different according to
Wilcoxon’s two sample test (P = 0.05).e Increases in populations of total fluorescent Pseudomonas spp. or phlD+ fluorescent Pseudomonas spp.
T
"
a �b�l�e � 5.
# � I$n�f �l�u�e�n�c �e � o�f �R %. ý s �o l &a �n�i � o�n � w h�e�a �t � r �o�o�t � c �o�l�o�n�i�z �a �t�i�o�n � b�y � i�n�d �i�g �e�n�o�u�s � f �l�u�o�r �e�s�c�e�n�t � P �s �e �u �d �o m�o n�a �s � s�pp! !.�Healthy X diseased roots a Non-lesioned X lesioned root sections of diseased plants b
Total pseudomonads c phlD+ d Total pseudomonads c phlD+ d
Soil -Rhiz. +Rhiz. Increase e - Rhiz. + Rhiz. Increase e -Rhiz. +Rhiz. Increase e -Rhiz. +Rhiz. Increase e
SV 1.5 X 107 1.1 X 108 * 7.7 8.5 X 105 3.9 X 106 * 4.6 8.7 X 107 1.4 X 108 1.6 5.5 X 106 2.3 X 106 0.4SSB 1.7 X 107 1.0 X 108 * 6.3 1.7 X 106 1.1 X 107 * 6.6 5.3 X 107 1.6 X 108 * 3.1 3.0 X 106 1.9 X 107 * 6.2
a Roots of wheat growing in soil non-amended (- Rhiz.) or amended (+ Rhiz.) with R. solani.b Non-lesioned and lesioned root sections of plants infected by R. solani.c Populations of total fluorescent Pseudomonas spp. (CFU g-1 root).d Population of phlD+ (CFU g-1 root). Mean values of five replicates are presented. Means marked with an asterisc (two means in a row) are sgnificantly different according to
Wilcoxon’s two sample test (P = 0.05).e Increases in populations of total fluorescent Pseudomonas spp. or phlD+ fluorescent Pseudomonas spp.
C'h (a )pt *,+e -r . 4: /10F2 r. e -q 3u4 e -n5 c 6y 7, 8 d 9i: v ;e -r. s <i: t +y 7, 8 a )n5 d 9
a )c 6ti +: v ;i: t +y 7

C= h>
a? pt@,A eB r C 4: D1E
FF rC eB qG uH eB nI cJ yK , L dM
iN vO eB rC sP iN tA yK , L a? nI d M
a? cJ tiA N vO iN tA yK
90
growth and biocontrol activity of Bacillus cereus (Smith et al., 1999). Furthermore, the
significance of the plant cultivar has been recently demonstrated at the physiological level.
Signficant changes in the expression of the phl biosynthetic operon of P. fluorescens strain
CHA0 were detected on different maize cultivars (Nots et al., 2001). Studies using different
profiling techniques (Mavrodi et al., 2001) suggested a significant level of endemism for
wheat-associated phlD+ fluorescent Pseudomonas spp. (Wang et al., 2001). Given the influence
a cultivar may have on the composition and activity of phlD+ isolates, the differences between
genotypes found in SV and SSB soils as well as between Dutch and US TAD soils may be due
to the different wheat cultivars used in different geographical regions and in different fields
within a specific region. The effect of wheat cultivars on the genotypic diversity of 2,4-DAPG-
producing Pseudomonas spp. will be addressed in future studies.
Comparison of the genotypic clustering of the isolates with the clustering based on in
vitro inhibition of mycelial growth of G. graminis var. tritici and R. solani revealed that this
aproach was less discriminatory than genotypic clustering (Figs. 2 and 3). In several host-
pathogen systems, antibiotic production by antagonistic microorganisms in vitro does not
correlate well with the level of suppression of diseases obtained in situ (Wong and Baker,
1984; Sharifi-Tehrani et al., 1998; Ellis et al., 2000). However, good correlation was found
between in vitro inhibition and suppression of G. graminis var. tritici in situ by
pseudomonads isolated from wheat roots grown in a TAD soil from Washington State (Weller
TQ
aR bS lTe U 6. VXW
IYnZ f[lTu\ eU nZ c] e U o^ f
[G_ . ` ga rb ac md i
enf ie sg vh aR ri . W tj rb ie tj ie ck ie o^ n Z ri hl i
mzn o^ so pp h
leU ri e U c] o^ l
To^ nZ imzn aR tq im o^ n Z b
Sy r imnZ tq ri o^ d
su\ c] eU d
sPt
. ` fulvuw ox rb ey sg ck ey nf sg
so tq ri aR im nZ so .W
Initial density a Final density (CFU g-1 root) b
Isolate (CFU seed-1) -G. graminis var.tritici
+G. graminis var.tritici
Increase
SSB17 2.1 X 104 1.0 X 107 a 1.9 X 107 a 1.9SSB17 1.3 X 106 1.9 X 107 a 3.1 X 107 a 1.69H4 3.2 X 104 2.3 X 107 a 2.4 X 107 a 1.09H4 1.5 X 106 1.9 X 107 a 2.6 X 107 a 1.4Q8r1-96 2.5 X 104 4.7 X 107 b 3.7 X 107 b 0.8Q8r1-96 3.9 X 106 7.4 X 107 b 9.1 X 107 b 1.24C5 3.9 X 104 4.2 X 107 b 5.7 X 107 b 1.44C5 9.3 X 106 7.4 X 107 b 7.1 X 107 b 1.0
a P. fluorescens strains SSB17, Q8r1-96, and their respective 2,4-DAPG-deficient mutants 9H4 and 4C5 wereapplied to seeds at two different initial densities.
b Plants were grown in CB soil for one month and roots were harvested to determine the final densities in therhizosphere. Means of five replicates are shown. Means followed by the same letter (columns) are notsignificantly different according to Wilcoxon’s two sample test (P = 0.05).

Cz h{
a| pt},~ e� r � 4: �1�
F� r� e� q� u� e� n� c� y� , � d�
i� v� e� r� s� i� t~ y� , � a| n� d �
a| c� ti~ � v� i� t~ y�
91
et al., 1988). In this context, it is interesting to note that the dominant genotypic group found
in the Dutch TAD soil, represented by strain SSB17, clustered closely together with Q8r1-96
in the phenotypic groups that were most inhibitory to the take-all fungus. In contrast, SSB17
and Q8r1-96 did not belong to the phenotypic group that was most inhibitory to R. solani
(Fig. 3). More isolates representative of the genotypic and phenotypic groups need to be tested
in biocontrol assays to support the earlier results obtained by Weller et al. (1988).
Strain SSB17 and strain Q8r1-96, when introduced into the conducive soil (CB) via
seed treatment at low densities of approximately 103 CFU per seed were able to suppress G.
graminis var. tritici to the same extent as occurred naturally in the suppressive soil (SSB).
However, their 2,4-DAPG-deficient mutant 9H4 and 4C5, respectively, did not protect wheat
plants against take-all (Fig. 4). Although 2,4-DAPG plays a key role in take-all suppression, it
does not contribute to the rhizosphere competence of 2,4-DAPG producers, since SSB17 and
Q8r1-96 colonized wheat roots to the same extent as their respective 2,4-DAPG deficient
mutants. Similar results were obtained in experiments employing P. fluorescens strain F113 and
its 2,4-DAPG-defective mutant (Carroll et al., 1995). In contrast to 2,4-DAPG, biosynthesis
of phenazine antibiotics was shown to significantly contribute to the ecological competence of
P. fluorescens 2-79 and P. aureofaciens 30-84 (Mazzola et al., 1992). Recently, Raaijmakers and
Weller (2001) demonstrated that Q8r1-96 is much more aggressive as a colonist of wheat
roots than other genotypes (Raaijmakers and Weller, 2001) and it is now known that this
unique colonizing ability is shared by strains from other soils that are genotypically similar to
strain Q8r1-96. Strain Q8r1-96 was more rhizosphere competent than SSB17 (Fig. 5; Table
6) even though the studies were conducted in raw Dutch soil, a finding which supports further
suggestion that the rhizosphere competence of Q8r1-96 is minimally affected by the
physicochemical characteristics of the soil (Raaijmakers and Weller, 2001) in which it is
growing.
One of the key questions about the phenomenon of TAD is the basis of the
enrichment of the antagonist responsible for specific suppression and the effect of the soil
environment on the development and expression of suppressiveness. We found no quantitative
differences in the population densities of 2,4-DAPG-producing fluorescent pseudomonads on
roots of wheat grown in Dutch TAD soils as compared to densities reported on roots of
wheat grown in Washington State TAD soils (Raaijmakers et al., 1997; Raaijmakers and
Weller, 1998). These findings suggest that the enrichment is independent of the physical and
chemical characteristics of the soil. Classical studies of TAD demonstrated that TAD is a field
phenomenon requiring the take-all pathogen, a susceptible host and at least one outbreak of

C� h�
a� pt�,� e� r � 4: �1�
F� r� e� q� u� e� n� c� y� , � d�
i� v e� r� s¡ i� t� y� , � a� n� d �
a� c� ti� � v i� t� y�
92
F¢
i£g¤ u¥ r¦ e § 5. ¨ª© R
«e§ l¬ a t® i£ o¯ n° s± h² i
£p³ s ± b
´e§ t® wµ ee§¶§ n ° t® h² e § i
£n° i£t® i£ a l ¬ d
·e§ n° s± i£ t® y ¸ o¯ n ° s± ee§¶§ d· s ± (¹ l¬ o¯ g C¤»º F
¢U ¼
s± ee§¶§ d· -½ 1¾ ) ¿ a n° d ·
t® h² e § fÀi£n° a l ¬ d
·e§ n° s± i£ t® i£ e§ s ± o¯ n°
r¦ oo¯Á¯ t® s ± (¹ l¬ o¯ g C¤Âº F¢
U ¼
g¤ -½ 1¾ r¦ oo¯Á¯ t® ) ¿ fÀo¯ r ¦ P
Ã. Ä fÅlÆuÇ oÈ rÉ eÊ sË cÌ eÊ nÍ sË s± t® r¦ a i£ n° s ± SS
ÎÁÎBÏ
17, ÐÒÑªÓ QÔ
8Õ
r¦ 1Ð -Ö 96, ×ªØ Ó a n° d
·t® h² e§ i£ r r¦Ù¦ e§ s± p³ e§ cÚ t® i£ vÛ e § 2,4
Ü ÓÞÝ -ÖDß
Aà
Pá
Gâ
-Ö d· e§ fÀ e§ cÚ t® i£ vÛ e § mã u¥ t® a n° t® s± , 9Ó × Hä
4, ÝªÓ a n° d ·
4Ý Cº 5.¨ª© Comparison of the rhizosphere competence of P.fluorescens strain SSB17 (solid line) and its 2,4-DAPG-deficient mutant 9H4 (dashed line),(A) and Q8r1-96 (solid line) and its 2,4-DAPG deficient mutant 4C5 (dashed line), (B).Wheat seeds were treated with each of the strains at densities of approximately 0, 103, 104,and 106 CFU seed-1. Plants were grown for one month in CB soil and rhizospherepopulations of the introduced strains were determined by dilution plating. Means of fivereplicates are shown. Error bars represent the standard error of the mean. Non-linearregression was performed to determine the relationship between the initial and finaldensities of the introduced strains. The equation used was Y=a*X/(X+b), where Y is thefinal density, X is the initial density, a is the maximum final density and b is the initialdensity necessary to reach half of the maximum final density. The experiment was repeatedtwice and representative results are shown.
severe disease. Some early studies suggested that the take-all pathogen, rather than the host
plant is primarily responsible for the selection of specific antagonists (Gerlagh, 1968; Zogg and
Jaggi, 1974; Wildermuth, 1980; Smiley, 1978; Brown, 1981; Barnett et al., 1999; Sarniguet et
al., 1992). For example, Gerlagh (1968) and Zogg and Jaggi (1974) induced suppressiveness
to take-all by repeatedly adding mycelium of G. graminis var. tritici to soil, but the relationship
of this type of suppression to TAD is still not known. In an attempt to begin to address the
question about the basis of enrichment of 2,4-DAPG producers in Dutch TAD soils, we
compared populations of fluorescent pseudomonads on healthy and diseased roots, and on
segments of roots from diseased plants with and without lesions. As has been demonstrated
many times in the literature (Weller, 1983; Vojinovic, 1972; Brown, 1981), we found that
roots with take-all lesions supported significantly larger populations of fluorescent
pseudomonads than healthy roots. However, 2,4-DAPG producers were not enriched on
diseased roots to a greater extent than the total population of fluorescent Pseudomonas spp.
Furthermore, no significant increases in population densities of introduced strains SSB17 and
Initial density (log CFU seed-1)
0 1 2 3 4 5 6 7 8
Fin
al d
ensi
ty (
log
CF
U g
-1 r
oo
t)
0
1
2
3
4
5
6
7
8
Q8r1-964C5
Y=8.71*X/(0.71+X) (r2=0.99, p<0.0001)Y=8.07*X/(0.32+X) (r2=0.99, p<0.0001)
B
Initial density (log CFU seed-1)
0 1 2 3 4 5 6 7
Fin
al d
ensi
ty (
log
CF
U g
-1 r
oo
t)
0
1
2
3
4
5
6
7
8
SSB179H4
A
Y=9.20*X/(1.60+X) (r2=0.98, p<0.0001)Y=9.99*X/(2.27+X) (r2=0.98, p<0.0001)

Cå hæ
aç ptè,é eê r ë 4: ì1í
Fî rë eê qï uð eê nñ cò yó , ô dõ
iö v÷ eê rë sø iö té yó , ô aç nñ d õ
aç cò tié ö v÷ iö té yó
93
Q8r1-96 were observed upon infection of wheat roots by G. graminis var. tritici. Based upon
these limited studies, at this time we conclude that increased nutrient availability upon fungal
infection is primarily responsible for the enrichment of 2,4-DAPG producers.
In light of the results of this and earlier studies (Raaijmakers and Weller, 1998;
Raaijmakers et al., 1997), the specificity of the suppression that operates in TAD soils becomes
interesting especially when considering the broad-spectrum activity of 2,4-DAPG against
many fungal pathogens (Keel et al., 1992). Australian TAD soils were shown to have also
certain levels of suppressiveness to Rhizoctonia solani, Giberella zeae, Pythium irregulare,
Cochliobolus sativus and Fusarium culmorum (Wildermuth, 1982). The results of our study,
however, indicated that the two Dutch TAD soils were not suppressive to R. solani. Given that
R. solani did not adversely affect population densities of 2,4-DAPG-producers (Tables 4, 5), a
possible explanation for a lack of suppressiveness to R. solani may be the relative insensitivity
of this fungus to 2,4-DAPG (Keel et al., 1992). Furthermore, fungi can change patterns of
bacterial gene expression (Fedi et al., 1997) and more specifically interfere with the production
of 2,4-DAPG (Duffy and Défago, 1997; Notz et al., 2001). The suppressiveness of Dutch
TAD soils to pathogens other than R. solani and their interaction with 2,4-DAPG-producing
Pseudomonas spp. are currently under investigation.
Mù
Aú
Tû Eü Rý IþAú
Lÿ
S � Aú
N�
D �
Mù
Eü Tû H�
O� D�
S�
Mù
i� c� r� oo�� r� g a� n� i� s m� s . � Naturally occurring populationsof fluorescent Pseudomonas spp. were isolated fromthe wheat rhizosphere on King's medium B (KMB)agar supplemented with chloramphenicol (13 µgml-1), ampicillin (40 µg ml-1), and cycloheximide(100 µg ml-1) [KMB+] (Simon and Ridge, 1974). AllPseudomonas strains described in this study,including SSB17 (this study), Q8r1-96(Raaijmakers and Weller, 2001), Pf-5 and CHA0(Keel et al., 1996) were grown on KMB.Spontaneous rifampicin resistant derivatives ofSSB17 and Q8r1-96 were selected on KMBsupplemented with rifampicin (100 µg ml-1). Strains9H4 and 4C5 are 2,4-DAPG deficient mutantsderived from strain SSB17 and Q8r1-96,respectively. 2,4-DAPG deficient mutants areresistant to rifampicin (100 µg ml-1) and kanamycin(100 µg ml-1). Escherichia coli strain S17 λ pir wasobtained from Dr. L. S. Thomashow (USDA-ARS,WA) and contained the mini-Tn5lacZ element inplasmid pUT (De Lorenzo et al., 1990). S17 λ pirwas grown in liquid Luria Bertani (LB) amendedwith kanamycin (25 µg ml-1). All bacterial strains
were stored at -80°C in LB or KMB-brothsupplemented with 40% (v/v) glycerol.
Isolate R3-111a-1 of G. graminis var.tritici (the causal agent of take-all) was originallyisolated from wheat grown in a soil near MosesLake, Washington State, USA. Isolate AG-8 ofRhizoctonia solani (the causal agent of Rhizoctoniaroot rot) was obtained from the collectionmaintained by the USDA-ARS, Pullman,Washington State. Isolate CBS 219.65 of Pythiumultimum var. sporangiiferum (the causal agent ofPythium root rot) was obtained from the Dutchcollection of microorganisms (CBS, Baarn, TheNetherlands). G. graminis var tritici, R. solani, andP. ultimum var. sporangiiferum were routinelygrown on potato dextrose agar (Oxoid Ltd.,Basingstoke, Hampshire, England) [PDA]. Mycelialplugs were immersed in sterile mineral oil andstored at 15°C.
S� o� i� l� s .� Soils CB, SV, and SSB were obtained inDecember 1997, 1999 and 2001 from agriculturalpolder fields at Woensdrecht, The Netherlands.Woensdrecht is located in the South-West of theNetherlands, 10 km from the city Bergen op Zoom.

C� h�
a� pt��� e� r � 4: ���
F� r� e� q� u� e� n� c y! , " d#
i$ v% e� r� s& i$ t� y! , " a� n� d #
a� c ti� $ v% i$ t� y!
94
In 1997, the SV and SSB soil were continuouslycropped to wheat for 14 and 27 years, respectively;the CB soil was grown to wheat and sugarbeet in a1:2-year rotation scheme. Soils were collected fromthe top 50-cm of the soil profile, air-dried for aweek, and passed through a 0.5-cm mesh screenprior to use. Chemical and physical properties ofthe soils used in this study are listed in Table 1.
N' a( t) u* r+ a( l , s- u* pp./. r+ e0 ss-1- i2 v3 e0 n4 e0 ss -5- o6 f 7
s- o6 i2l,s - t) o 6 G8 . 9 g: r; a< m= i
>n? i>s@ v3 a( r+ .A
tB r; i> tB i> cC i> a( n4 d D RE . 9 s@ oF lG a< n? i>.A Soils were amended with 0.1 to
0.4% (wt/wt) of an oat grain inoculum of G.graminis var. tritici isolate R3-111a-1 (particle size0.25 to 0.50 mm) (Raaijmakers and Weller, 1998);sterilized oat grain inoculum was used as a control.Wheat seeds (cv. Bussard) were sown in PVC pots(8 cm high, 7 cm wide) containing 200 g of soil andcovered with a 1-cm layer of soil without inoculum.Plants were grown for four weeks in climatechambers at 15°C with a 12-h photoperiod. Twicea week, plants received 50 ml of 1/3 strengthHoagland's solution (macroelements only). Eachtreatment had five replicates with approximately 12plants per replicate. Eight randomly selected plantswere harvested, their root systems washed and theseverity of take-all determined on a 0-to-8 scale,where “0” indicates no disease and “8” a dead plant(Pierson and Weller, 1994). Similar to assaysperformed with G. graminis var. tritici,suppressiveness of the soils to R. solani was assessedby introducing 0.5% (wt/wt) of a millet graininoculum of R. solani AG-8; sterilized millet grainwas used as a control. To assess the level ofsuppressiveness to R. solani infections, roots of eightrandomly selected plants were harvested, washedand root dry weight determined. From eachreplicate, three to four randomly selected plantswere collected for isolation of naturally occurringpseudomonads from the rhizosphere.
IH s- o6 l,a( t) i2 o6 n 4 a( n4 d D e0 n4 u* mI e0 r+ a( t) i2 o6 n 4 o6 f
7pJ hKlG DL + M f
7l,u* o6 r+ e0 s- cN e0 n4 t)
PO s@ eP uQ dRoF m= oF n? a< s@ s- pp./. .A After 4 weeks of plant growth,
three to five plants of each replicate were harvestedand loosely adhering soil was removed from theroot system. Naturally occurring fluorescentPseudomonas spp. were isolated from uninfectedwheat roots and roots infected by G. graminis var.tritici or R. solani. From the infected roots both,non-lesioned and lesioned root sections wereseleted. Root samples of 0.2 to 1 g fresh weightwere vortexed for 1 min and then sonicated for 1min in a ultrasonic cleaner (Bransonic 12, BransonUltrasonics Corp., Geneva, Switzerland). Rootsuspensions were dilution plated onto KMB+ andincubated at 25°C for 48 h. Population densities ofphlD+ Pseudomonas spp., a key gene in thebiosynthesis of 2,4-DAPG, were determined bycolony hybridization followed by PCR (Raaijmakerset al., 1997). phlD+ colonies were purified andstored at -80°C for further analyses.RS AT
PU D V
a( n4 a( l, yW s- i2 s- . RAPD analysis of phlD+ isolateswas performed with primers M13, M12, and D7.
These primers were selected by Keel et al. (1996)among sixty-four other random primers based onthe distinct and consistent banding patterns of thepolymorphic markers produced. PCR was carriedout in a 25-µl reaction mixture as describedpreviously (Raaijmakers et al., 1997). PCRamplifications were carried out in a MJ ResearchPTC-200 thermocycler. Samples (10-15 µl) of thePCR products were separated on 2% agarose gels in1X TBE buffer (90 mM Tris-borate, 2 mM EDTA[pH 8.0]) at 80 V for 3 h. Gels were stained withethidium bromide and bands were visualized on aUV transilluminator. RAPD analysis was repeatedat least three times for each primer. The presence orabsence of bands that were consistently generatedby RAPD analysis (1 for the presence and 0 for theabsence of a particular sized band in the gel) wasused to calculate the pairwise coefficients ofsimilarity (Nei-Li distances). Cluster analysis withthe neighbour joining method and bootstrapanalysis were performed with the program FreeTree(Hampl, et al., 2001). The dendrogram was editedand visualised in TREEVIEW (Page, 1996). Datafrom the RAPD analyses were also used to calculatethe Shanon-Weaver’s diversity index, using theformula H=Σ[pi-ln(pi)], where pi is the proportionof each RAPD group (based on 100% similarity) inrelation to the total number of RAPD groups persoil.
PU l,a( t) e 0 i
2n4 hX i
2bY
i2t) i2 o6 n 4 a( ss-1- a( yW s- . phlD+ Pseudomonas isolates
obtained from roots of wheat grown in Dutchagricultural soils were inoculated at the edges of 9-cm plates containing 1/5th strength PDA (initial pH= 6.5) and incubated for two days at 25°C. Mycelialplugs (5 mm diameter) of G. graminis var. tritici, R.solani, and Pythium ultimum var. sporangiiferumwere tranferred to the centre of the plate. Plateswere incubated at 25°C and the radial growth ofthe fungus toward the bacterial isolate wasmeasured after 3 to 5 days and compared with thecontrol (fungi or oomycete only). For each isolate,growth inhibition of both G. graminis var. triticiand R. solani was determined in duplicate. Datafrom plate inhibition assays were used to constructsimilarity matrices, where a hierarchial clusteranalysis was performed by the single linkagemethod (Davies et al., 2001).
TZ
n4 5[
-\ mI u* t) a( g] e0 n4 e0 s- i2 s- .A 2,4-DAPG-deficient mu-tants ofspontaneous rifampicin-resistant derivatives of P.fluorescens strains SSB17 and Q8r1-96 wereobtained by biparental mating with E. coli strainS17 λ pir harbouring the mini-Tn5lacZ element inplasmid pUT, according to protocols described bySambrook and Russel (2001). Transformants wereselected on KMB supplemented with rifampicinand kanamycin and subsequently transferred to 98-well microtiter plates containing KMB broth.Transformants unable to produce a red pigment,which is not 2,4-DAPG, but is characteristic for

Ch_
a` pta�b ec r d 4: e�f
Fg rd ec qh ui ec nj ck yl , m dn
io vp ec rd sq io tb yl , m a` nj d n
a` ck tib o vp io tb yl
95
2,4-DAPG producing Pseudomonas strains (Bangeraand Thomashow, 1996), were selected after oneweek of growth at 25ºC. High-performance liquidchromatography (HPLC) was used to confirm theinability of these transformants to produce 2,4-DAPG (Bonsall et al., 1997). Plate inhibition assayswith G. graminis var. tritici, R. solani and P.ultimum sporangiiferum were used to furtherconfirm the inability of mutant strains to inhibitthese pathogens in vitro. RAPD analysis withprimers M13, M12, and D7, was performed toconfirm strain integrity. Southern blot analysis wasused to determine the number of copies of theTn5lacZ element in the 2,4-DAPG-deficientmutants.
Sr os ut tu hv ew rx n y az ny az l{ y| s} i~ s } az ny d � c� os l{ os ny y | hv y| b�
rx i~ d� i~ z� az tu i~ os ny s} .� ForSouthern blot analysis, genomic DNA of wildtypeand mutant strains was extracted with the DNAWizard Kit (Promega, Leiden, The Netherlands).Samples containing 2.0 µg of DNA were digestedwith 5 U of KpnI and EcoRI (Promega), twoenzymes for which restriction sites are not presentin the kanamycin gene of the mini-Tn5lacZelement (De Lorenzo et al., 1990). Restrictions wereperformed in a total volume of 100 µl at 37°C for12 h. Digested DNA was precipitated with 4MLiCl, washed with 70% ethanol, dissolved in 15 µlof sterile distilled water and separated on 1%agarose gels in 1X TBE. DNA and colony transferto Hybond N+ nylon membranes (AmershamPharmacia Biotech) were performed according tostandard methods (Sambrook and Russel, 2001).Membrane washes and hybridizations wereperformed following standard procedures(Sambrook and Russel 2001). High-stringentconditions comprised prehybridization for 1.5 h at65°C, hybridization for 12 h at 65°C, membranewashing twice for 5 min each with 2X SSC (1XSSC is 0.15 M NaCl plus 0.015 M sodium citrate)-0.1% SDS at room temperature, and membranewashing twice each for 30 min with 0.1X SSC-0.1% SDS at 65°C. A 575-bp probe, specific for thekanamycin gene contained within the Tn5LacZelement, was obtained by direct PCR DIG-labelling(Roche Corp., Basel, Switzerland) of the fragment
using primers KM1 (5’-CCCGATGCGCCAGAGTTGTT) and KM2 (5’-TCACCGAGGCAGTTCCATAGG).
T�
rx ew az tu m� ew ny t u os f �
s} ew ew d� s } w� i~ tu h v P�
s� e� u� d�o� m� o� n� a� s� . � For bioassays
and rhizosphere competence studies, wheat seeds(cv. Bussard) were coated with SSB17, Q8r1-96, ortheir 2,4-DAPG-deficient mutants in 1%methylcellulose to final densities of approximately103, 104, and 106 CFU per seed; seeds treated with1% methylcellulose served as a control. Populationdensities of introduced Pseudomonas spp. strainsSSB17, Q8r1-96 and their 2,4-DAPG deficientmutants were assessed on uninfected wheat rootsand on roots infected by G. graminis var. tritici.
Sr tu az tu i~ s} tu i~ c� az l { az ny az l{ y| s} i~ s} .� Prior to ANOVA, data frompopulation counts were log10-transformed anddisease index data were ranked. Root dry weightdata were analysed directly by ANOVA followed byTukey’s studentized range test, after certifyingnormal distribution and homogeneity of variances(SAS institute, Inc., Cary, N.C.). Data frombacterial population counts on G. graminis var.tritici- and R. solani-lesioned roots, which did notpresent normal distribution after transformationslog10 (CFU), were analysed by the one-way non-parametric Kruskal-Wallis test and comparison ofmeans of two treatments was performed byWilcoxon’s two sample test. Data from populationincreases were also analysed by the one-way non-parametric Kruskal-Wallis test followed byWilcoxon’s two sample test. Data from rhizospherecompetence assays were analysed by non-linearregression analysis to determine the relationshipbetween the initial density of strains SSB17 andQ8r1-96 on seeds and their final density on roots of1-month old wheat plants. The equation used wasY=a*X/(b + X), where Y represents the final density(log CFU g-1 root), X the initial density (log CFUseed-1), a the maximum final density, and b theinitial density necessary to reach half of the maximalfinal density (Raaijmakers and Weller, 2001). Allexperiments were performed at least twice andrepresentative results are shown.
A�
C�
K� N� O� W�
L�
E�
D� G�
E�
M�
E�
N� T�
Sr
This research was financially supported by CAPES, Brazil (Project nr.1515/96-9). We thank Dr. LindaS. Thomashow for providing E. coli S17 λ pir containing the mini-Tn5lacZ element in plasmid pUT. We aregrateful to Prof. Dr. Pierre de Wit for critically reading the manuscript and for his valuable suggestions. We alsothank Corrie Geerds for technical assistance.

C� h�
a� pt�� e¡ r ¢ 4: £�¤
F¥ r¢ e¡ q¦ u§ e¡ nc© yª , « d¬
i v® e¡ r¢ s¯ i t yª , « a� nd ¬
a� c© ti v® i t yª
96
R° E± F²
E± R° E± N³ CE± Sµ
A¶
s· he¹ rº , » M¼
.½ J¾ .½ C., ½ » a¿ nÀ d Á Sµ h¸ i pà tÄ oÅ nÀ , » PÆ
.½ J¾ .½ (eds). 1981.Biology and control of take-all, Academic Press,New York.A¶
bÇ
e¹ rrºÈº a¿ , » M¼
.½ BÉ ., ½ » Sµ e¹ a¿ h¸ , » Sµ . ½ a¿ nÀ d Á Sµ i vÊ a¿ s· i tÄ h¸ a¿ mË pà a¿ rº a¿ mË , » KÌ .½1998. Suppression of the take-all fungus(Gaeumannomyces graminis var. tritici) by a sterilered fungus through induced resistance in wheat(Triticum aestivum) seedling roots. Soil Biol.Biochem. 30:1457-1461.BÉ a¿ nÀ gÍ e¹ rº a¿ , » M
¼.½ GÎ ., ½ » a¿ nÀ d Á TÏ hoÅ mË a¿ s· hoÅ wÐ , » L
Ñ.½ Sµ .½ 1996.
Characterization of a genomic locus required forsynthesis of the antibiotic 2,4-diacetylphloroglucinol by the biological controlagent Pseudomonas fluorescens Q2-87. Mol. Plant-Microbe Interac. 9:83-90.BÉ a¿ rº nÀ e¹ ttÄÒÄ , » Sµ .½ J¾ ., ½ » Sµ i nÀ gÍ lÓ e¹ tÄ oÅ nÀ , » I
Ô., ½ » a¿ nÀ d Á R° yÕ dÁ e¹ rº , » M
¼.½ 1999.
Spatial variation in populations of Pseudomonascorrugata 2140 and pseudomonads on take-alldiseased and healthy roots systems of wheat. SoilBiol. Biochem. 31:633-636.BÉ oÅ nÀ s· a¿ llÓÖÓ , » R° .½ F² ., ½ » W
×e¹ lÓ lÓ e¹ rº , » DØ .½ M¼ ., ½ » a¿ nÀ d Á TÏ hoÅ mË a¿ s· hoÅ wÐ , » L
Ñ.½ Sµ .½
1997. Quantification of 2,4-diacetylphloroglucinolproduced by fluorescent Pseudomonas sp. in vitroand in the rhizosphere of wheat. Appl. Environ.Microbiol. 63:951-955.BÉ rº oÅ wÐ nÀ , » M
¼.½ E± .½ 1981. Microbiology of roots infected
with the take-all fungus (Gaeumannomyces graminisvar. tritici) in phased sequences of winter wheat.Soil Biol. Biochem. 13:285-291.Ca¿ rrºÈº oÅ llÓÙÓ , » HÚ ., ½ » M
¼oÅ ëÛnnÀÜÀ e¹ -Ý LÑ oÅ ccÞßÞ oÅ zà , » Y
á., ½ » DØ oÅ wÐ lÓ i nÀ gÍ , » DØ .½ N³ .,½ »
a¿ nÀ d Á Oâ ’ãGÎ a¿ rº a¿ , » F
².½ 1995. Mutational disruption of the
biosynthesis genes coding for the antifungalmetabolite 2,4-diacetylphloroglucinol does notinfluence the ecological fitness of Pseudomonasfluorescens F113 in the rhizosphere of sugarbeets.Appl. Environ. Microbiol. 61:3002-3007.Ch¸ a¿ rº i gÍ kä a¿ pà a¿ kä oÅ rº nÀ , » N³ ., ½ » a¿ nÀ d Á Sµ i vÊ a¿ s· i tÄ ha¿ mË pà a¿ rº a¿ mË , » KÌ .½1987. Changes in the composition and populationof fluorescent pseudomonads in wheat rootsinoculated with successive generations of root-pieceinoculum of the take-all fungus. Phytopathol.77:1002-1007.CooÅÅ kä , » R° .½ J¾ .½ 1985. Biological control of plantpathogens, with special reference to the take-allfungus in suppressive soils. Taiwan Agric. Res. Inst.Plant Prot. Bull. 27:179-198CooÅÅ kä , » R° .½ J¾ ., ½ » a¿ nÀ d Á BÉ a¿ kä e¹ rº , » KÌ .½ F² .½ 1983. The Nature andPractice of Biological Control of Plant Pathogens.The American Phytopathological Society, St. Paul,MN.CooÅÅ kä , » R° .½ J¾ ., ½ » a¿ nÀ d Á R° oÅ vÊ i rº a¿ , » A
¶.½ DØ .½ 1976. The role of
bacteria in the biological control ofGaeumannomyces graminis by suppressive soils. Soil.Biol. Biochem. 8:269-273.DØ a¿ vÊ i e¹ s· , » KÌ .½ GÎ ., ½ » F
²a¿ rº gÍ e¹ ttÄÒÄ e¹ , » M
¼., ½ » BÉ a¿ lÓ lÓ a¿ , » GÎ ., ½ » DØ a¿ uå dÁ i , » A
¶.,½ »
DØ uå pà pà oÅ nnÀÜÀ oÅ i s· , » R° ., ½ » GÎ oÅ wÐ e¹ nÀ , » Sµ .½ R° ., ½ » M¼
a¿ tÄ e¹ i llÓÙÓ e¹ , » TÏ .,½ »PÆ
h¸ i llÓÖÓ i pà s· , » Sµ a¿ wÐ a¿ dÁ oÅ gÍ oÅ , » A¶
., ½ » TÏ rº i vÊ i nÀ oÅ , » C., ½ » Væ
oÅ uå yÕ oÅ uå kä a¿ lÓ oÅ uå ,»E± ., ½ » a¿ nÀ d Á TÏ rº uå dÁ gÍ i llÓÙÓ , » DØ .½ LÑ .½ 2001. Cuticle heterogeneity
as exhibited by Pasteuria spore attachment is notlinked to the phylogeny of parthenogenetic root-knot nematodes (Meloidogyne spp.). Parasitol.122:111-120.DØ e¹ a¿ cÞ oÅ nÀ , » J
¾.½ W× .½ 1976. Biological control of the take-
all fungus, G. graminis, by P. radicicola and similarfungi. Soil. Biol. Biochem. 8:275-283.DØ e ¹ L
ÑoÅ rº e¹ nÀ zà oÅ , » V
æ., ½ » HÚ e¹ rrºÈº e¹ rº oÅ , » M
¼., ½ » J
¾a¿ kä uå b
Çzà i kä , » U
ç., ½ » a¿ nÀ dÁ
TÏ i mË mË i s· , » KÌ .½ N³ .½ 1990. Mini-Tn5 transposonderivatives for insertion mutagenesis, promoterprobing, and chromosomal insertion of clonedDNA in Gram-negative eubacteria. J. Bacteriol.172:6568-6572.DØ uå fè fè yÕ , » BÉ .½ KÌ ., ½ » a¿ nÀ d Á DØ éé fè a¿ gÍ oÅ , » GÎ .½ 1997. Zinc improvesbiocontrol of Fusarium crown and root rot oftomato by Pseudomonas fluorescens and represses theproduction of pathogen metabolites inhibitory tobacterial antibiotic biosynthesis. Phytopathol.87:1250-2157.E± llÓÖÓ i s· , » R° .½ J¾ ., ½ » TÏ i mmËêË i s· -Ý W× i lÓ s· oÅ nÀ , » TÏ .½ M¼ ., ½ » a¿ nÀ d Á BÉ a¿ i lÓ e¹ yÕ , » M
¼.½ J¾ .½
2000. Identification of conserved traits influorescent pseudomonads with antifungal activity.Environ. Microbiol. 2:274-284.F²
e¹ dÁ i , » Sµ ., ½ » TÏ oÅ lÓ a¿ , » E± ., ½ » M¼
oÅ ëÛnnÀëÀ e¹ -Ý LÑ oÅ ccÞìÞ oÅ zà , » Y
á., ½ » DØ oÅ wÐ lÓ i nÀ gÍ ,»
DØ .½ N³ ., ½ » Sµ mË i tÄ h¸ , » LÑ
.½ M¼ ., ½ » a¿ nÀ d Á Oâ ’ãGÎ a¿ rº a¿ , » F
².½ 1997. Evidence
for signaling between the phytopathogenic fungusPythium ultimum and Pseudomonas fluorescens: P.ultimum represses the expression of genes in P.fluorescens F113, resulting in altered ecologicalfitness. Appl. Environ. Microbiol. 63:4261-4266.GÎ e¹ rº lÓ a¿ gÍ h, » M
¼.½ 1968. Introduction of Ophiobolus
graminis into new polders and its decline. Neth. J.Plant Pathol. 74:1-97.GÎ uå ttÄÒÄ e¹ rº i dÁ gÍ e¹ , » R° .½ J¾ ., ½ » HÚ oÅ rº nÀ b
ÇyÕ , » DØ ., ½ » HÚ oÅ llÓÙÓ i nÀ s· , » TÏ .½ W× ., ½ » a¿ nÀ dÁ
PÆ
rº e¹ wÐ , » R° .½ DØ . ½ 1993. Take-all in autumn-sown wheat,barley, triticale and rye grown with high and lowinputs. Plant Pathol. 42:425-431.HÚ a¿ mË pà lÓ , » V
æ., ½ » P
Æa¿ vÊ lÓ i cÞ e¹ kä , » A
¶., ½ » a¿ nÀ d Á F
²lÓ e¹ gÍ rº , » J
¾.½ 2001.
Construction and bootstrap analysis of DNAfingerprinting-based phylogenetic trees with thefreeware program FreeTree: application totrichomonad parasites. Int. J. Syst. Evol. Microbiol.51:731-735.HÚ oÅ rº nÀ b
ÇyÕ , » DØ .½ 1983. Suppressive soils. Ann. Rev.
Phytopathol. 21:65-85.KÌ ee¹ß¹ lÓ , » C., ½ » Sµ cÞ h¸ nÀ i dÁ e¹ rº , » U
ç., ½ » M¼
a¿ uå rº hoÅ fè e¹ rº , » M¼
., ½ » Væ
oÅ i s· a¿ rº dÁ , » C´ .,½ »LÑ
a¿ vÊ i llÓÖÓ e¹ , » J¾.½ , » BÉ uå rº gÍ e¹ rº , » U
ç., ½ » W×
i rº tÄ h¸ nÀ e¹ rº , » PÆ
., ½ » HÚ aa¿í¿ s· , » DØ ., ½ » a¿ nÀ dÁDØ éé fè a¿ gÍ oÅ , » GÎ .½ 1992. Suppression of root diseases byPseudomonas fluorescens CHA0: importance of thesecondary metabolite 2,4-diacetylphloroglucinol.Mol. Plant-Microbe Interact. 5:4-13.KÌ ee¹ß¹ lÓ , » C´ ., ½ » W
×e¹ lÓ lÓ e¹ rº , » DØ .½ M¼ ., ½ » N³ a¿ tÄ s· cÞ h, » A
¶., ½ » DØ éé fè a¿ gÍ oÅ , » GÎ .,½ »
CooÅÅ kä , » R° .½ J¾ ., ½ » a¿ nÀ d Á TÏ hoÅ mË a¿ s· hoÅ wÐ , » LÑ
.½ Sµ .½ 1996.Conservation of the 2,4-diacetylphloroglucinolbiosynthesis locus among fluorescent Pseudomonasstrains from diverse geographic locations. Appl.Environ. Microbiol. 62:552-563.

Cî hï
að ptñ�ò eó r ô 4: õ�ö
F÷ rô eó qø uù eó nú cû yü , ý dþ
iÿ v� eó rô s� iÿ tò yü , ý að nú d þ
að cû tiò ÿ v� iÿ tò yü
97
M�
a� v� r� o� d� i� , O
.� V� ., � M�
c S� p� a� dd��� e� n� -� G� a� r� d� e� n� e� r� , B� .� B� .,� M�
a� v� r� o� d� i� , D� .� V� ., � B� o� n� s� a� ll��� , R� .� F� ., � W�
e� l� l� e� r� , D� .� M� ., � a� n� d�T� o� m� a� s� h� o� w , L! .� S� .� 2001. Genetic diversity of phlDfrom 2,4-diacetylphloroglucinol-producingfluorescent Pseudomonas spp. Phytopathol. 91:35-43.M�
a� zz"#" o� l� a� , M�
., � C$ oo�%� k&, R� .� J' ., � T� h� o� m� a� s� h� o� w , L! .� S� .,�
W�
e� ll�(� e� r� , D� .� M� ., � a� n� d � P) i� e� r� s� o� n � III*+*,* , L! .� S� .� 1992.Contribution of phenazine antibiotic biosynthesisto the ecological competence of fluorescentpseudomonads in soil habitats. Appl. Environ.Microbiol. 58:2616-2624.M�
a� zz"#" o� l� a� , M�
., � a� n� d � G� u- , Y.
.� H/ .� 2000. Impact of wheatcultivation on microbial communities from replantsoils and apple growth in greenhouse trials.Phytopathol. 90:114-119.M�
c S� p� a� dd�0� e� n� -� G� a� r� d� e� n� e� r� , B� .� B� ., � S� c h� r� o� e� d� e� r� , K1
.� L! .,� K1
a� ll��� o� g2 e� r� , S� .� E3 ., � R� aa�4� i� j5 m� a� k& e� r� s� , J'.� M� .� , T� h� o� m� a� s� h� o� w ,
L! .� S� ., � a� n� d � W�
e� ll�(� e� r� , D� .� M� .� 2000. Genotypic andphenotypic dyversity of phlD-containingPseudomonas strains isolated from the rhizosphereof wheat. Appl. Environ. Microbiol. 66:1939-1946.N6 o� t7 z" , R� ., � M
�a� u- r� h� o� f8 e� r� , M
�., � S� c h� n� i� d� e� r� -� K1 ee�9� l� , U: .� ,
D� u- f8 f8 y; , B� ., � H/
aa�<� s� , D� ., � a� n� d � D� é= f8 a� g2 o� , G� .� 2001. Bioticfactors affecting expression of the 2,4-diacetylphloroglucinol biosynthesis gene phlA inPseudomonas fluorescens strains CHA0 in therhizosphere. Phytopathol. 91:873-881.P) a� g2 e� , R� .� D� .� M� .� 1996. TREEVIEW: An applicationto dsplay phylogenetic trees on personal computers.Comp. Applicat. Biosc. 12:357-358.P) i� e� r� s� o� n� , E3 .� A> ., � a� n� d � W
�e� l� l� e� r� , D� .� M� .� 1994. Use of
mixtures of fluorescent pseudomonads to suppresstake-all and improve growth of wheat. Phytopathol.84:940-947.R� aa�<� i� j5 m� a� k& e� r� s� , J
'.� M� .� , W
�e� ll��� e� r� , D� .� M� .� , a� n� d � T� h� o� m� a� s� h� o� w ,
L! .� S� .� 1997. Frequency of antibiotic-producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63:881-887.R� aa�<� i� j5 m� a� k& e� r� s� , J
'.� M� . � a� n� d � W
�e� l� l� e� r� , D� .� M� .� 1998. Natural
plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils.Mol. Plant-Microbe Interact. 11:144-152.R� aa�<� i� j5 m� a� k& e� r� s� , J
'.� M� .� , B� o� n� s� a� ll ��� R� .� F� ., � a� n� d � W
�e� l� l� e� r� , D� .� M� .�
1999. Effect of population density of Pseudomonasfluorescens on production of 2,4-diacetylphloroglucinol in the rhizosphere of wheat.Phytopathol. 89:470-475.R� aa�<� i� j5 m� a� k& e� r� s� , J
'.� M� .� , a� n� d � W
�e� l� l� e� r� , D� .� M� .� 2001.
Exploiting genotypic diversity of 2,4-diacetylphloroglucinol-producing Pseudomonas spp.:characterization of superior root-colonizing P.fluorescens strain Q8r1-96. Appl. Environ.Microbiol. 67:2545-2554.S� a� m� b? r� oo�%� k
&, J'., � a� n� d � R� u- ss�@� e� l� , D� .� W� .� 2001. Molecular
cloning: a laboratory manual, 3rd ed. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.S� a� r� n� i� g2 u- e� t7 , A
>., � L! u- c a� s� , P) ., � L! u- c a� s� , M
�., � a� n� d � S� a� m� s� o� n� , R� .�
1992. Soil conduciveness to take-all of wheat:influence of the nitrogen fertilizers on the structureof populations of fluorescent pseudomonads. PlantSoil 145:29-36.
S� h� a� r� i� f8 i� -� T� e� h� r� a� n� i� , A>
.� , ZA a� l� a� , M�
., � N6 a� t7 s� c h� , A>
.� , M�
o� ëBnn�C� e� -�
L! o� c c o� z" , Y.
., � a� n� d � D� é= f8 a� g2 o� , G� .� 1988. Biocontrol ofsoil-borne fungal diseases by 2,4-diacetylphloroglucinol-producing pseu-domonadswith different restriction profiles od amplified 16SrDNA. Europ. J. Plant Pathol. 104:631-643.S� h� i� p� t7 o� n� , P) .� J' .� 1975. Take-all decline during cerealmonoculture. In: Bruehl, G.W. (ed.). Biology andControl of Soil-Borne Plant Pathogens, pages 137-144, The Am. Phytopathol. Soc., St. Paul, MN.S� h� i� p� t7 o� n� , P) .� J' ., � C$ oo�%� k
&, R� .� J' ., � a� n� d � S� i� tt7D7 o� n� , J
'.� W� .� 1973.
Occurrence and trasfer of a biological factor in soilthat suppresses take-all of wheat in easternWashington. Phytopathol. 63:511-517.S� i� m� o� n� , A
>., � a� n� d � R� i� d� g2 e� , E3 .� H/ .� 1974. The use of
ampicillin in a simplified selective medium for theisolation of fluorescent pseudo-monads. J. Appl.Bacteriol. 37:459-460.S� m� i� l� e� y; , R� .� W� . � 1978. Colonization of wheat roots byGeumannomyces graminis var. tritici inhibited byspecific soils, microorganisms and ammonium-nitrogen. Soil Biol. Biochem. 10:175-179.S� m� i� l� e� y; , R� .� W� .� 1979. Wheat-rhizoplanepseudomonads as antagonists of Geumannomycesgraminis var. tritici. Soil Biol. Biochem. 11:371-376.S� m� i� t7 h� , K
1.� P) ., � H
/a� n� d� e� l� s� m� a� n� , J
'., � a� n� d � G� oo�%� d� m� a� n� , R� .� M� .�
1999. Genetic basis in plants for interactions withdisease-suppressive bacteria. Proc. Natl. Acad. Sci.USA 96:4786-4790.T� h� o� m� a� s� h� o� w , L! .� S� ., � a� n� d � W
�e� ll��� e� r� , D� .� M� .� 1990. Role of
antibiotics and siderophores in biocontrol of take-all disease of wheat. Plant Soil 129:93-99.T� h� o� m� a� s� h� o� w , L! .� S� ., � a� n� d � W
�e� ll�(� e� r� , D� .� M� .� 1996. Current
concepts in the use of introduced bacteria forbiological disease control: Mechanisms andantifungal metabolites. In: Stacey, G., and Keen,N.T. (eds.). Plant-Microbe Interact., pages 187-236, vol. 1, Chapman & Hall, New York.V�
o� j5i� n� o� v� i� c , ZA .� D� .� 1972. Biological antagonism as the
cause of decline of Ophiobolus graminis Sacc. inprolonged wheat monoculture. J. Sci. Agric. Res.25:31-41.W�
a� n� g2 , C$ ., � R� a� m� e� tt7D7 e� , A>
., � P) u- n� j5a� s� a� m� a� r� n� w o� n� g2 , P) .,�
ZA a� l� a� , M�
.� , N6 a� t7 s� c h� , A>
., � M�
o� ëBnn�C� e� -� L! o� cc < o� z" , Y
.., � a� n� d�
D� é= f8 a� g2 o� , G� .� 2001. Cosmopolitan distribution ofphlD-containing dicotyledonous crop-associatedbiocontrol pseudomonads of worldwide origin.FEMS Microbiol. Ecol. 37:105-116.W�
e� ll��� e� r� , D� .� M� .� 1983. Colonization of wheat roots bya fluorescent pseudomonad suppressive to take-all.Phytopathol. 73:1548-1553.W�
e� ll��� e� r� , D� .� M� ., � H/
o� w i� e� , W�
.� J' ., � a� n� d � C$ oo�%� k&, R� .� J' . � 1988.
Relationship between in vitro inhibition ofGaeumannomyces graminis var. tritici andsuppression of take-all of wheat by fluorescentpseudomonads. Phytopathol. 78:1094-1100.W�
i� l� d� e� r� m� u- t7 h� , G� .� B� .� 1980. Suppression of take-all bysome Australian soils. Austral. J. Agric. Res. 31:251-258.

CE hF
aG ptHJI eK r L 4: MON
FP rL eK qQ uR eK nS cT yU , V dW
iX vY eK rL sZ iX tI yU , V aG nS d W
aG cT tiI X vY iX tI yU
98
W[
i\ l] d^ e_ r` ma ub tc hd , e Gf .g Bh .g 1982. Soils suppressive toGaeumannomyces graminis var. tritici: effects onother fungi. Soil Biol. Biochem. 14:561-567.W[
oi nj gk , e Pl
.g Tm .g W[ ., g e an nj d ^
Bh an ko e_ r, e Rp .g 1984. Suppressionof wheat take-all and Ophiobolus patch byfluorescent pseudomonads from a Fusarium-suppressive soil. Soil Biol. Biochem. 16:397-403.
Zq oi ggkrk , e Hs ., g e an nj d ^
Jtan ggkuk i\ , e W
[.g 1974. Studies on the
biological soil disinfection. VII. Contribution to thetake-all decline (Gaeumannomyces graminis)imitated by means of laboratory trials and some ofits possible mechanisms. Phytopathol. Zeits.81:160-169.

_______________________________________________________________________________
CHAPTER 5
Ev
FFwxw
Ev
Cy
T z
O{
F w
2|
,} 4~ -� D� I�A�
Cy
Ev
Tz
Y�
L�
P�
H�
L�
O{
R�
O{
G�
L�
U�
Cy
I�N�
O{
L�
O{
N �
P�
Y� T� H�
I�U� M �
S�P�
E�
C�
I�E�
S�:�
C�
E�
LL���
U�
L�
A�
R R���
E�
S�P�
O�
N�
S�E�
S �
A�
N�
D
V¡
A�
R�
I�A�
T¢
I�O�
N �
I�N �
S�E�
N�
S�I�T¢
I�V¡
I�T¢
Y £
A�
M¤
O�
N�
G¥
P�
R�
O�
P�
A�
G¥
U�
L�
E�
S �
A�
N�
D
S�P�
E�
C�
I�E�
S�
_______________________________________________________________________________

C¦ h§
a¨ pt©Jª e« r ¬ 5: ¯® E°
ff±+±
e« c² t ª o³ f 2,4-D±µ´O¶ ·¯¸º¹
A»
P¼ G ½ o³ n ¾ P¿
yÀ tÁ h ià uÄ mÅ
100
A»
BÆ SÇ TÈ
RÉ A»
C¦ TÈ
_______________________________________________________________________________
TÈ
h§
e « a¨ n¾ tª iÊ bË iÊ o³ tiª Ê c 2,4-² ´O¶ ·¯¸dÌ
iÊ a¨ c² e« tª yÍ lÎp© h§
lÎo³ r¬ og³ÐÏ l
ÎuÑ c² iÊ n¾ o³ l
Î(2,4-DÒ ´O¶ ·¯¸º¹ A
»P¼ G½ )
Ópl© Î a¨ yÍ s Ô a¨ n ¾ iÊ mpÕ © o³ r¬ tª a¨ n¾ t ª r¬ o³ l
Îe « iÊ n ¾ tª h§ e«
sÔ uÑ pp©Ö© r¬ e« ssÔ9Ô iÊ o³ n ¾ o³ f ±
pl© Î a¨ n¾ t pª×© a¨ tª h§ o³ geÏC« n¾ s Ô bË
y Í sÔ e« vØ e« r¬ a¨ l Î
sÔ tª r¬ a¨ iÊ n¾ s Ô o³ f ±
Ps¿ÚÙ
euÛÜÄ dÝoÞ mÅ oÞ nß aà s
Ù sÔ pp©J© . Tá È h
§e « r¬ e« sÔ uÑ lt
Î ª s Ô o³ f ±
tª h§ iÊ s sÔâÔ tª uÑ dÌ
yÍsÔ h§ o³ w ã tª h§ a¨ t ª wã itÊ ª h§ iÊ n ¾ a¨ n¾ d
Ìa¨ mÕ o³ n¾ g Ï P
¿yÀ tÁ h ià uÄ mÅ sÔ p© e« c² iÊ e« s Ô tª h§ e« r¬ e « iÊ s Ô vØ a¨ r¬ iÊ a¨ tiª Ê o³ n ¾ iÊ n ¾ sÔ e« n¾ sÔ itiÊ ª Ê vØ iÊ tª y Í tª o ³ 2,4-D
´O¶ ·¯¸º¹A»
P¼ G½ .áA»
lÎsÔ o ³ vØ a¨ r¬ iÊ o³ uÑ s Ô p© r¬ o³ p© a¨ gÏ uÑ l
Îe« s Ô o³ f
±P¿
. ä ulįå tÁ ià mÅ uÄ m Å vØ a¨ r¬ . á sÙpæ oÞ rç aà nß gè iiÃ+à fé eÛ rç uÄ m Å (Ò P¿ uÄ s
Ù),Ó ¶
tª h§ a¨ t ª a¨ r¬ e « p© a¨ r¬ t ª o³ f ±
tª h§ e « a¨ sÔ e« xuê Ñ a¨ lÎ
sÔ tª a¨ ge oÏë«ì³ f ±
tª h§ e « lÎiÊ f± e « c² yÍ c² l
Îe« , ¶
dÌ
iÊ f± f± e« r ¬ c² o³ n¾ sÔ iÊ dÌ e« r¬ a¨ bË
lÎy Í iÊ n ¾ tª h§ e« iÊ r ¬ sÔ e« n¾ sÔ iÊ tiª Ê vØ iÊ tª y Í tª o ³ 2,4-D
´O¶ ·¯¸º¹A»
P¼ G½ . á Mí
yÍ c² e« liÎ Ê uÑ m Õ wã a¨ s Ô tª h§ e«mÕ o³ sÔ t ª r¬ e« sÔ iÊ sÔ tª a¨ n¾ t ª sÔ tª ru¬9Ñ c² tª urÑJ¬ e« , f
¶ë±o³ lÎlÎo³ wã e« d
ÌbË
y Í zî oo³Ð³ sÔ p© o³ r¬ a¨ n¾ gÏ iÊ a¨ , ¶zî o³ o³ sÔ p© o³ r¬ e « c² yÍ sÔ tª s Ô a¨ n¾ d
Ìzî o³ o³ sÔ p© o³ r¬ e« sÔ . á A
»ddÌJÌ
iÊ tiª Ê o³ n¾ a¨ llÎDÎ
yÍ ,¶
wã e « r¬ e« p© o³ r¬ tª , ¶ f±o³ r ¬ tª h§ e « f
±iÊ r¬ sÔ t tªOª iÊ mÕ e« ,
¶tª h§ a¨ t ª p© H ï h
§a¨ s Ô a ¨ sÔ iÊ gÏ n¾ iÊ f± iÊ c² a¨ n¾ t ª e« f
±f±e« c² t ª o³ n ¾ tª h§ e « a¨ c² tiª Ê vØ iÊ tª y Í o³ f 2,4-D
±ð´O¶ ·¯¸ñ¹A»
P¼ G½ , w¶ ã iÊ tª h§
a ¨ h§
iÊ gÏ h§
e« r ¬ a¨ c² tiª Ê vØ itÊ ª y Í a¨ t ª lÎo³ w ã p© Hï . á Fò urÑJ¬ tª h§ e« r¬ mÕ o³ r¬ e« ,
¶tª h§ e « l
Îe« vØ e« l Î o³ f
±a¨ c² e« tª yÍ l
Îa¨ tiª Ê o³ n ¾ o³ f
±p© h§
lÎo³ r¬ o³ gÏ l
ÎuÑ c² iÊ n¾ o³ l
Îs Ô iÊ s Ô a¨ l
ÎsÔ o ³ a¨
mÕ a¨ jóo³ r ¬ d
Ìe« tª e« r¬ mÕ iÊ n¾ a¨ n¾ t ª o³ f
±tª h§ e« iÊ r ¬ a¨ c² tiª Ê vØ iÊ tª yÍ . Tá È r¬ a¨ n¾ sÔ miÕ Ê ssÔôÔ iÊ o³ n ¾ e« l
Îe« c² tª r¬ o³ n ¾ miÕ Ê c² r¬ o³ sÔ c² o³ p© y Í (TÒ È E
°Mí
) Ó
sÔ tª uÑ dÌ
iÊ e« s Ô r¬ e« vØ e« a¨ lÎe« dÌ
tª h§ a¨ t ª 2,4-D´O¶ ·¯¸º¹
A»
P¼ G ½ c² a¨ uÑ sÔ e« s Ô dÌ
iÊ ff±+± e« r¬ e« n¾ t ª sÔ tª a¨ geÏë« s Ô o³ f ±
dÌ
iÊ sÔ o³ r¬ gÏ a¨ n¾ iÊ sÔ a¨ tª iÊ o³ n ¾ iÊ n ¾ h§
yÍ p© h§
a¨ l tipÎ ª Ê © s Ô o³ f
±P¿
uÄ sÙ, ¶
iÊ n¾ c² lÎuÑ dÌ
iÊ n¾ gÏa¨ ltÎ ª e« r¬ a¨ tiª Ê o³ n ¾ (Ò p© r¬ o³ l
ÎiÊ f± e« r¬ a¨ tiª Ê o³ n¾ ,
¶r¬ e« tª r¬ a¨ c² tª iÊ o³ n¾ a¨ n¾ d d
ÌõÌiÊ sÔ ru¬9Ñ pti©Öª Ê o³ n¾ )
Óo³ f ±
tª h§ e « pl© Î a¨ sÔ mÕ a ¨ mÕ e« mÕ bË
r¬ a¨ n¾ e« , ¶vØ a¨ c² uÑ o³ li
Î Ê sÔ a¨ tiª Ê o³ n ¾ a¨ n¾ dÌ
c² e« ll ÎDÎ
c² o³ n¾ tª e« n¾ t ª dÌ
iÊ sÔ iÊ n¾ tª eg«%Ï r¬ a¨ tª iÊ o³ n¾ . á TÈ
h§
e « iÊ mpÕ © lÎiÊ c² a¨ tiª Ê o³ n¾ s Ô o³ f
±tª h§ e« sÔ e « r¬ e« sÔ uÑ lt
Î ª s Ô f±o³ r ¬ tª h§ e e«ö« ff
±÷±iÊ c² a¨ c² y Í a¨ n¾ d
Ìc² o³ n¾ sÔ iÊ sÔ tª e« n¾ c² y Í o³ f
±bË
iÊ o³ lÎo³ gÏ iÊ c² a¨ l
Îc² o³ n¾ tª r¬ o³ l
Îo³ f ±
p© lÎa¨ n¾ t pªø© a¨ tª h§ o³ geÏë« n¾ iÊ c ² P
¿yÀ tÁ h ià uÄ mÅ sÔ p© e« c² iÊ e« s Ô b
Ëy Í 2,4-D´O¶ ·¯¸º¹
A»
P¼ G½ -¸p© r¬ o³ d
ÌuÑ c² iÊ n¾ g Ï Ps
¿ÚÙeÛ uÄ dÝoÞ mÅ oÞ nß aà s
ÙsÔ pp©J© . á a¨ r¬ e « d
ÌiÊ sÔ c² uÑ ssÔ9Ô e« d
Ì.á
_______________________________________________________________________________
Jùo³ r¬ ge Ïë« T.
È á dÌ
e « SÇ o³ uÑ zî a¨ , ¶C¦ h§
r¬ iÊ sÔ tiª Ê n¾ e « A»
r¬ n¾ o³ uÑ lÎdÌ
, ¶C¦ h§
r¬ yÍ sÔ tª e« l Î
D¹
e« uÑ lÎvØ o³ tª , ¶ P¼ h
§iÊ liÎ Ê p© p© e « Lú e« mÕ a¨ n¾ c² e« a¨ uÑ ,
¶Vû
iÊ vØ iÊ e« n¾ n¾ e«Gi½õÊ a¨ n¾ iÊ n¾ a¨ zî ziî Ê -¸ P¼ e« a¨ r¬ sÔ o³ n¾ ,
¶a¨ n¾ d Ì
Jùo³ s Ô M
í. á RÉ aa% iÊ jó mÕ a¨ k
üe« r¬ sÔ
To be submitted.

Cý hþ
aÿ pt��� e� r � 5: ��� E�
ff
e� c� t � o� f 2,4-D� �� �����
A�
P� G � o� n � P�
y� t� h� i� u� m�
101
I N! T"
RO#%$ D�
U&
Cý T"
I O$ N!
Phloroglucinols (PG) are phenolic, secondary metabolites produced by plants, algae and
bacteria (Vincent et al., 1991; Bangera and Tomashow, 1996; 1999; Ishiguro et al., 1988;
Bokesch et al., 1999; Verrota et al., 1999; Jimenez-Escrig et al., 2001). More than sixty PG
derivatives have been described and were reported to have antiviral, antimicrobial,
ichthyotoxic, insect and mammal antifeedant, antihelminthic, phytotoxic, antioxidant,
cytotoxic, anti-tumor, and plant growth-regulating activities (James, 1979; Yajima and
Munakata, 1979; Arisawa et al., 1990; Tada et al., 1990; 1992; Keel et al., 1992; Cronin et al.,
1997a; Te-Chato and Lim, 1999; Lawler et al., 1999; Ito et al., 2000; Debabrata and Naik,
2000; Jimenez-Escrig et al., 2001). The antimicrobial activity of these compounds, and in
particular of 2,4-diacetylphloroglucinol (2,4-DAPG), has received considerable attention in
the area of biological control of plant diseases.
Numerous studies have demonstrated that 2,4-DAPG-producing Pseudomonas spp. can
suppress a wide variety of plant pathogens, including fungi, bacteria and nematodes (Stutz et
al., 1986; Keel et al., 1990; Vincent et al., 1991; Fenton et al., 1992; Levy et al., 1992; Keel et
al., 1992; Cronin et al., 1997a, b; Duffy and Défago, 1997; Sharifi-Tehrani et al., 1998).
Among the plant pathogenic fungi, Gaeumannomyces graminis var. tritici, Thielaviopsis basicola,
Fusarium oxysporum f. sp. radicis-lycopersici, and the oomycete Pythium ultimum can be
effectively controlled by 2,4-DAPG-producing Pseudomonas strains (Vincent et al., 1991;
Fenton et al., 1992; Sharifi-Tehrani et al., 1998; Stutz et al., 1986; Duffy and Défago, 1997).
The determinative role of 2,4-DAPG in disease suppression by Pseudomonas strains has been
demonstrated by (1) the use of mutants deficient in 2,4-DAPG production (Vincent et al.,
1991; Shanahan et al., 1992; Keel et al., 1992; Cronin et al., 1997a, b), (2) complementation
of 2,4-DAPG deficient mutants and concomittant restoration of biocontrol activity (Vincent
et al., 1991; Keel et al., 1992; Cronin et al., 1997a, b), and (3) expression of 2,4-DAPG
biosynthetic genes in heterologous, nonproducing strains thereby conferring biocontrol
activity or enhanced activity (Vincent et al., 1991; Shanahan et al., 1993; Bangera and
Thomashow, 1996).
Reporter gene systems (Loper and Lindow, 1997) and analytical techniques
(Thomashow et al., 1997) have further demonstrated that 2,4-DAPG is produced in situ by
both introduced Pseudomonas strains as well as indigenous Pseudomonas populations (Keel et
al., 1992; Bonsall et al., 1997; Duffy and Défago, 1997; Raaijmakers et al., 1999; Notz et al.,
2001). Pseudomonas strains that produce 2,4-DAPG also produce monoacetylphloroglucinol
(MAPG) and depending on the nutritional environment the ratio of MAPG to 2,4-DAPG

C' h(
a) pt*�+ e, r - 5: .�/ E0
ff11
e, c2 t + o3 f 2,4-D1�4�5 6�7�8
A9
P: G ; o3 n < P=
y> t? h@ iA uB mC
102
may change (Duffy and Défago, 1997). In addition to MAPG and 2,4-DAPG, also PG has
been isolated from cultures of a bacterium found on decayed roots of red pine seedlings and
classified as Aeromonas hydrophila (Strunz et al., 1978).
In spite of significant progress in our understanding of the biosynthesis and regulation
of 2,4-DAPG-production in Pseudomonas strains (Bangera and Thomashow, 1996, 1999;
Sarniguet et al., 1995; Whistler et al., 1998; Blumer and Haas, 2000), little attention has been
given to fungal responses to 2,4-DAPG and to its mode of action. Most studies on the
sensitivity of plant pathogenic fungi to 2,4-DAPG take into account only one species or isolate
of the target pathogen (Keel et al., 1992; Levy et al., 1992). Furthermore, most studies on
sensitivity of pathogens to 2,4-DAPG consider only one particular stage in the life cycle of a
pathogen, usually mycelial growth (Keel et al., 1992; Levy et al., 1992; Mazzola et al., 1995).
Most life cycles of pathogens, however, are complex and comprise numerous pathogen
structures and infectious propagules. For example, oomycete pathogens can infect host tissues
by means of multiple propagules, including mycelium, zoosporangia, zoospores and oospores.
Understanding the variation in sensitivity of different phases of the life cycle of a pathogen to a
specific biocontrol trait will give more insight into the potential efficacy of biocontrol agents
harbouring that particular trait.
In this study, we investigated the response of Pythium species to 2,4-DAPG. Pythium
species are ubiquitous pathogens of many important crops around the world and possess a
high level of diversity with more than 200 species described (Van der Plaats-Niterink, 1981;
Dick, 1990). Many of these species can occur simultaneously at the same site and often more
than one species infect a certain host plant (Chamswarng and Cook, 1985). We determined
the variation in sensitivity of 14 Pythium isolates to 2,4-DAPG; the isolates were obtained
from multiple hosts and represent eight species. The sensitivity of various infectious
propagules of Pythium ultimum var. sporangiiferum to 2,4-DAPG was studied in detail with
special emphasis on those propagules that are part of the asexual life cycle. The effects of pH
and level of acetylation on activity of phloroglucinols against mycelial growth of P. ultimum
var. sporangiiferum were also assessed. To understand the mode of action of 2,4-DAPG,
ultrastructural changes induced by 2,4-DAPG in hyphal tips of Pythium ultimum var.
sporangiiferum were studied by transmission electron microscopy.

CD hE
aF ptG�H eI r J 5: K�L EM
ffNN
eI cO t H oP f 2,4-DN�Q�R S�T�U
AV
PW G X oP n Y PZ
y[ t\ h] iu_ m
103
Ra EM
Sb Uc
Ld Te
Sb
EM
ffNfN
eI cO t H oP f 2,4-DNgQ�R S�ThU
AV
PW G X oP n Y mi yj cO eI likml aF l k
gn rJ oP wo tH h E
oP f N
dp
ilfNfNeI rJ eI nY t H P
Zy[ t\ h] iu_ m sq pG eI cO i
leI sq .r Mycelial growth of
fourteen Pythium isolates, obtained from multiple host plants and representing eight different
species, was completely inhibited at 2,4-DAPG concentrations ranging from 8 to more than
300 µg ml-1 (Table 1). P. volutum CBS 699.83 was the most sensitive isolate, whereas P.
deliense V1-PPO was relatively insensitive to 2,4-DAPG. Mycelial growth of the other Pythium
isolates was completely inhibited at 2,4-DAPG concentrations ranging from 32 to 72 µg ml-1.
P. ultimum var. sporangiiferum isolate CBS 219.65, further referred to as Pus, was used for a
detailed analysis of the effect of 2,4-DAPG on different propagules, including zoosporangia,
zoopores and zoospore cysts.
EM
ffNfN
eI cO t H oP f 2,4-DNsQ�R S�ThU
AV
PW G X oP n Y zt ooPuP sq pG oP rJ aF nY gn ilaF . r 2,4-DAPG had a significant effect on both the
formation and the diameter of zoosporangia of Pus (Fig. 2). The number of newly formed
zoosporangia decreased by approximately 50% when exposed to 2.0 µg ml-1 of 2,4-DAPG.
The diameter of the newly formed zoosporangia, an indicative measure of their ripening,
decreased from 26 to 20 µm with increasing concentrations of 2,4-DAPG.
EM
ffNfN
eI cO t H oP f 2,4-DNvQ�R S�T�U
AV
PW G X oP n Y zt ooPwP sq pG oP rJ eI s q aF nY d p
zt ooPuP sq pG oP rJ e I cO yj sq tH sq .r 2,4-DAPG adversely affected the
behaviour of Pus zoospores at concentrations of 0.8 ng ml-1 and higher (Fig. 3). At a
concentration of 0.8 ng ml-1, all zoospores stopped swimming and approximately 50% of the
zoospores disintegrated (Fig. 4). The time for disintegration to occur depended on the
concentration of 2,4-DAPG. At concentrations of 0.8 ng ml-1, disintegration of the zoospores
occurred in approximately 20 min and at concentrations of 3.2 ng ml-1 it only took 3 min
(data not shown).
The effect of 2,4-DAPG on germination of encysted zoospores was studied in water
and in wheat root exudates. Wheat root exudates were included since responses to antibiotics
may differ depending on the nutritional conditions. In experiments conducted in water,
15.8% of the total number of encysted zoospores had germinated prior to addition of 2,4-
DAPG. There was no significant effect of 2,4-DAPG on the percentage of germinated zoopore
cysts at concentrations up to 2.8 µg ml-1 (Fig. 5A). At a 2,4-DAPG concentration of 3.2 µg
ml-1, there was a significant increase in germination of encysted zoospores, which was observed
in all replicate experiments. At 2,4-DAPG concentrations higher than 3.2 µg ml-1, germination
of encysted zoospores of Pus significantly decreased.

Cx hy
az pt{�| e} r ~ 5: ��� E�
ff��
e} c� t | o� f 2,4-D����� �����
A�
P� G � o� n � P�
y� t� h� i� u� m�
104
T�
a� b� l�e � 1. ��� E� ff
���e� c t ¡ o¢ f
�2£
,4¤¦¥ -§ D¨ A©
Pª
G «
o¢ n ¬ m y® c e� l� i¯ a� l � g° r± o¢ w² t¡ h ³
o¢ f� P´
yµ t¶ h· i¸u¹ mº s» p¼ p¼ .�
Species Isolate a Origin EC50 (µgml-1)b MIC (µg ml-1) c
P. aphanidermatum 311 – PPO cucumber 32-40 48P. aristosporum CBS 263.38 wheat 16-24 32P. deliense V1 – PPO cucumber 300 >300P. intermedium 294 – LBO lily 32-40 56P. intermedium LBO hyacinth 24-32 48P. irregulare ATCC 1120 wheat 24-32 48P. irregulare LBO crocus 16-24 56P. macrosporum 111 – LBO lily 16-24 40P. ultimum P17 – LBO tulip 24-32 72P. ultimum 266 lily 24-32 72P. ultimum IRS sugar beet 24-48 56P. u. var. sporangiiferum CBS 219.65 wheat 16-24 32P. u.var. ultimum CBS 725.94 wheat 16-24 32P. volutum CBS 699.83 wheat < 8 8
a CBS, Collection of Fungal Cultures, Utrecht, The Netherlands; LBO, Applied Plant Research, section FlowerBulbs, Lisse, The Netherlands; PPO, Applied Plant Research, Naaldwijk, The Netherlands; ATCC, AmericanType Culture Collection, University Blvd, Manassas, Virginia, USA; IRS, Institute for Rational SugarProduction, Bergen op Zoom, The Netherlands.b Concentration of 2,4-DAPG necessary to inhibit radial mycelial growth by 50%.c Minimal concentration of 2,4-DAPG causing 100% inhibition of mycelial growth.Medium used was PDA (pH 6.5) and inhibition was assessed every 24 hours during 7 days of incubation at25°C. 2,4-DAPG concentrations varied from 0 to 300 µg ml-1 with intervals of 8 µg ml-1. Each value is based onthree experiments with 6 replicates per treatment.
In wheat root exudates, germination of encysted zoospores of Pus was approximately
100% (Fig. 5B). At concentrations up to 2.4 µg ml-1 there was no adverse effect of 2,4-DAPG
on germination, whereas concentrations ranging from 2.8 to 6.4 µg ml-1 led to a significant
decrease in cyst germination. 2,4-DAPG concentrations of 6.4 µg ml-1 reduced the percentage
of germinated cysts even below the initial percentage of 11% germinated zoospore cysts that
were present prior to addition of 2,4-DAPG, which may suggest that relatively high
concentrations of 2,4-DAPG cause lysis of germ tubes.
E�
ff�f�
e} c� t | o� f �
p{ H ½ az n� d ¾
l¿e} vÀ e} l ¿ o� f
�az c� e} t| yÁ l
¿az t| i o� n � o� n � az c� ti|  vÀ i t| yÁ . à In absence of 2,4-DAPG, pH had no
significant effect on the growth rate of Pus: growth rates were on average 23 mm day-1 at a pH
ranging from 4.5 to 8.0 (Fig. 6). In presence of 2,4-DAPG, growth rates of Pus decreased
significantly and this decrease was highly dependent on pH. At pH 4.5, mycelial growth of
Pus was completely inhibited at a 2,4-DAPG concentration of 16 µg ml-1, whereas a 2,4-
DAPG concentration of 40 µg ml-1 was not sufficient to completely inhibit mycelial growth at
pH 7.5 and 8.0.
2,4-DAPG was significantly more inhibitory than MAPG and PG (Fig. 7). Mycelial

CÄ hÅ
aÆ ptÇ�È eÉ r Ê 5: Ë�Ì EÍ
ffÎÎ
eÉ cÏ t È oÐ f 2,4-DÎ�Ñ�Ò Ó�Ô�Õ
AÖ
P× G Ø oÐ n Ù PÚ
yÛ tÜ hÝ iÞ uß mà
105
A B C
Fá
iâgã uä rå e æ 1. ç�è C
éhê
eæ më iâcì aí l î sï tð rå uä cì tð uä rå e æ oñ f
òtð hê e æ pó h
êlîoñ rå oñ gã l
îuä cì iâ nô oñ l
îdõ
eæ rå iâ vö aí tð iâ vö eæ s ï uä sï eæ d õ
iân ô tð hê i
âs ï sï tð uä d
õy÷ . è (A) Phloroglucinol –
PG, (B) Monoacetylphloroglucinol – MAPG, (C) 2,4-Diacetylphloroglucinol – 2,4-DAPG.
growth of Pus was completely inhibited at a 2,4-DAPG concentration of 32 µg ml-1, whereas
for MAPG almost 10-fold higher concentrations were necessary (Fig. 7). In presence of PG,
Pus could still grow at concentrations of 300 µg ml-1. Collectively, these results indicate that
the higher the number of acetyl groups the greater the activity of phloroglucinol derivatives.
Fá
iâgã uä rå e æ 2.
ø è Eù
ffòúò
eæ cì t ð oñ f ò
2ø
,û 4ü -ý Dþ Aÿ
P�
G �
oñ n ô fòoñ rå më aí tð iâ oñ n ô aí nô d
õdõ
iâaí më eæ tð eæ r å oñ f
òz� oñ oñ sï pó oñ rå aí nô gã i
âa í oñ f
òP�
. � ul��� t� i m u� m vö aí rå .ès� p� o r� a� n� g� ii
�f�e� r� u� m (� P� u� s� )� .è Mycelial plugs of Pus grown on medium without the antibiotic were
flooded with solutions of different 2,4-DAPG concentrations. After four days of incubationat 25°C the number and diameter of 10 sporangia formed per treatment was determinedmicroscopically. Mean values of five replicates are shown. Error bars represent the standarderror of the mean. For each of the two parameters, means followed by the same letter arenot significantly different according to Tukey’s studentized range test (P=0.05).
2,4-DAPG (µ� g/ml)
0.0 2.0 4.0 8.0 16.0
Nu
mb
er o
f sp
ora
ng
ia/m
m2
0
4
8
12
16
Mea
n d
iam
eter
(µ� m
)
0
4
8
12
16
20
24
28NumberDiameter
a
b
d
a
c
bc
b
bc
cd
bc
OHOH
OH OH
OHOH
CC H 3
O
OHOH
CC H 3
O
C
O
H 3C
OH

C� h�
a� pt��� e� r 5: !#" E$
ff%&%
e� c' t � o( f 2,4-D%*),+ -#.0/
A1
P2 G 3 o( n 4 P5
y6 t7 h8 i9 u: m;
106
S< t� a� ti�>= c ' e� ff%?%
e� c' t � o( f 2,4-D%@),+ -#.�/
A1
P2 G 3 o( n 4 mA yB c' e� lCi= a� l C
gD r o( wE t� h� . F P. ultimum var. sporangiiferum (Pus) was grown
for 7 days on PDA plates amended with different concentrations of 2,4-DAPG. Pus could
grow, although at reduced level, in presence of 2,4-DAPG up to a concentration of 32 µg ml-1.
Mycelial plugs from Pus cultures that failed to grow at concentrations higher than 32 µg ml-1
were transferred to PDA plates without 2,4-DAPG. The plugs exposed to 48, 64, and 96 µg
ml-1 re-grew on PDA without 2,4-DAPG, whereas the ones exposed to 128 µg ml-1 2,4-DAPG
or higher were not able to grow on PDA plates free of 2,4-DAPG.
TG
E$
M H
sI t� uJ dK
i= e� sI .F For studying ultra-structural changes in hyphal tips of Pus exposed to 2,4-
DAPG, samples were taken at the periphery of Pus colonies growing in absence and presence
of 2,4-DAPG. The concentration of 2,4-DAPG used was 400 µg per paper disc, which was
placed on the surface of the agar medium. In absence of 2,4-DAPG, most hyphal tips (72%)
showed normal cell morphology, characterised by dense cytoplasm containing numerous
organelles (mitochondria, ribosomes, endoplasmic reticulum and some vacuoles (Table 2; Fig.
8A, B). In the presence of 2,4-DAPG, different stages of disorganisation in hyphal tips were
observed. A relatively low percentage (10%) of the hyphal tips had dense cytoplasm (Table 2)
which contained numerous mitochondria, ribosomes and small vacuoles (Fig. 8C, D).
Recurrent features observed in the cell contents of hyphal tips exposed to 2,4-DAPG were 1) a
well-defined rough endoplasmic reticulum, 2) the localised alteration (proliferation or
FL
iMgN uO rP e Q 3.
RTSEU
ffVWV
eQ cX t Y oZ f V
2[
,\ 4] -^ D_ A`
Pa
G b
oZ n c zd oZ oZ se pf oZ rP eQ s e oZ f V
Pg
. h uli�j tk il mm ui mm vn ao rP . S sp pq or rs at nu gv ililfwex rs ui mm (y Pg ui sp )z .S Zoospores were
exposed to different concentrations of 2,4-DAPG and the percentage of swimming, encysted,and disintegrated zoospores were determined microscopically. Mean values of 5 replicates aregiven. Error bars represent the standard error of the mean.
2,4-DAPG (ng/ml)
0 0,2 0,4 0,8 1,6 3,2
Per
cen
tag
e (%
)
0
20
40
60
80
100SwimmingEncystedDisintegrated

C{ h|
a} pt~�� e� r � 5: �#� E�
ff�&�
e� c� t � o� f 2,4-D�*�,� �#�0�
A�
P� G � o� n � P�
y� t� h� i� u� m�
107
F�
i�g� u� r� e � 4.
�@�E
ff¡W¡
e� c¢ t £ o¤ f ¡
2¥
,¦ 4� -§ D¨ A©
Pª
G «
o¤ n ¬ z o¤ o¤ s® p¯ o¤ r� e� s ® o¤ f ¡
P°
. ± ul²�³ t´ iµ m¶ u² m¶ v· a¸ r� . � s¹ pº o» r¼ a½ n¾ g¿ iµiµfÀeÁ r¼ u² m¶ ( P° u² s¹ )à .� (A): round-shaped
zoospores (B): disintegration. (C): disappearance. Arrows indicate zoospores of Pus(magnification 100X). The concentration of 2,4-DAPG was 3.2 ng ml-1 and the zoosporesuspension contained 104 zoospores ml-1.
disruption) in plasma membrane organisation, and 3) the development of an extensive
network of smooth membranous vesicles which appeared to be closely associated with the
endoplasmic reticulum (Fig. 8C). A large proportion of the hyphae (66%) showed cell content
disorganisation (Table 2), a degenerated cytoplasm which was bordered by a retracted plasma
membrane (Fig. 8E). Hyphal senescence was accompanied by vacuolisation and degeneration
of its contents (Fig. 8F). Pus hyphal tips growing in absence of 2,4-DAPG were delimited by a
thin electron translucent wall where the cellulosic components were strongly stained by the
PATAg reaction (Fig. 8B). The PATAg reaction occurred similarly in cell walls of Pus growing
in presence of 2,4-DAPG (Fig. 8D). These results suggest that 2,4-DAPG does not affect the
β(1-4), and β(1-6) glucan content of the cell wall of Pus.
Observations of sections immunolabelled to locate β(1-3) glucans revealed the
presence of these compounds in both the membrane and the cytoplasm of Pus growing in
absence (Fig. 9B) or in presence of 2,4-DAPG (Fig. 9C). No immunolabelling was present in
control sections without primary antibody (Fig. 9A). These results indicate that 2,4-DAPG
does not adversely affect synthesis of β (1-3) glucans.
A B C

CÄ hÅ
aÆ ptÇ�È eÉ r Ê 5: Ë#Ì EÍ
ffÎ&Î
eÉ cÏ t È oÐ f 2,4-DÎ*Ñ,Ò Ó#Ô0Õ
AÖ
P× G Ø oÐ n Ù PÚ
yÛ tÜ hÝ iÞ uß mà
108
DÕ
Iá Sâ CUÄäã SSâåâ Iá Oæ Nç
The results of this study show that within and among Pythium species there is
variation in sensitivity to 2,4-DAPG, a broad-spectrum antibiotic produced by Pseudomonas
biocontrol strains (Table 1). Also various propagules of P. ultimum var. sporangiiferum (Pus),
that are part of the asexual stage of the life cycle, can differ considerably in their sensitivity to
2,4-DAPG (Table 1, Figs. 2, 3, 5). Mycelium was the most resistant structure, followed by
zoosporangia, zoospore cysts and zoospores. Additionally, we report for the first time that pH
has a significant effect on the activity of 2,4-DAPG, with a higher activity at low pH.
Furthermore, the level of acetylation of phloroglucinols also is a major determinant of their
activity. Finally, TEM studies revealed that 2,4-DAPG causes different stages of
disorganisation in hyphal tips of Pus, including alteration (proliferation, retraction and
disruption) of the plasma membrane, vacuolisation and cell content degeneration.
Collectively, these results give a better understanding of the mode of action and activity of 2,4-
DAPG, which contributes to further improve the efficacy of biological control of plant
pathogenic Pythium species by 2,4-DAPG-producing Pseudomonas spp.
Pythium species have a complex life cycle with multiple infectious propagules.
Depending on the environmental conditions, zoosporangia may be very important propagules
in the interaction between Pythium species and their hosts (Nelson and Hsu, 1994). The
formation and diameter of Pus zoosporangia were significantly reduced at a 2,4-DAPG
concentration of 2.0 µg ml-1 (Fig. 2). The diameter of zoosporangia is an indicator of their
developmental stage since they need to reach a certain diameter to ripen and liberate zoospores
(Van der Plaats-Niterink, 1981). Several Pythium species reproduce asexually, forming mobile
biflagellate zoospores, which are responsible for the dispersal under moist conditions. In
hydroponic systems or humid soils, zoospores locate new hosts by responding to chemo-
attractants produced by the plant (Zhou and Paulitz, 1993). Zoospores swim, accumulate on
root surfaces and encyst in response to root polysaccharides that interact with receptors on the
zoospore surface (Deacon and Donaldson, 1993). We found that zoospores of Pus stopped
swimming at a 2,4-DAPG concentration of 0.8 ng ml-1 and almost all disintegrated at 3.2 ng
ml-1 (Fig. 3). The high sensitivity of zoospores to 2,4-DAPG may be explained by the lack of a
protective cell wall (Deacon and Donaldson, 1993). This is supported by the observation that
encysted zoospores of Pus, which have a cell wall, were significantly more resistant to 2,4-
DAPG than non-encysted zoospores (Figs. 3, 5). Encysted zoospores germinate in response to
host-produced amino acids and polysaccharides, causing root, stem, or seed infections

Cè hé
aê ptë�ì eí r î 5: ï#ð Eñ
ffò&ò
eí có t ì oô f 2,4-Dò*õ,ö ÷#ø0ù
Aú
Pû G ü oô n ý Pþ
yÿ t� h� i� u� m�
109
F�
i�g� u� r e 5. � � E
�ff���
e c� t � o� f �
2�
,� 4� -� D� A�
P�
G �
o� n � g� e r m� i�n� a� t� i� o� n� o� f
�e n� c� y� s t� e d
!z" o� o� s p# o� r e s o� f
�P$
. % ul&(' t) i* m+ u& m+ v, a� r . � s- p. o/ r0 a1 n2 g3 i*i*f4e5 r0 u& m+
(6 P� u� s )7 .� (A) Germination of encysted zoospores in water after two hours of exposure todifferent 2,4-DAPG concentrations. (B) Germination of encysted zoospores in wheat rootexudates after two hours exposure to different 2,4-DAPG concentrations. The germinationpercentage was determined microscopically. Means of five replicates are presented. Errorbars represent the standard error of the means. Means followed by the same letter are notsignificantly different according to Tukey’s studentized range test (P=0.05).
(Estrada-Garcia et al., 1990; Zhou and Paulitz, 1993; Deacon and Donaldson, 1993). The
results of this study clearly illustrate the stimulatory effect of root exudates on cyst germination
(Fig. 5). The significant increase in zoospore cyst germination in water at a 2,4-DAPG
concentration of 3.2 µg ml-1 could be explained by stimulation at sub-lethal concentrations
commonly observed with fungicides (Kenyon et al., 1997). For example, pyrazophos increased
the sporulation of Erysiphe sp. on Rhododendron at relatively low fungicide concentrations
(Kenyon et al., 1997). Similar results were reported for benomyl, carbendazim, and sulphur,
enhancing both the growth and sporulation of Drechslera teres (Toubia-Rahme et al., 1995).
The causes of this phenomenon are not known. However, it may be interpreted as a last
survival effort of the pathogen. Collectively, the results of our study suggest that 2,4-DAPG-
producing Pseudomonas spp. may be much more effective in biological control of Pythium
species that infect host plants primarily by means of zoospores, or under specific
environmental conditions where zoospores play a much more prominent role in the disease
cycle.
Within the relatively limited number of different Pythium species tested in this study,
considerable variation in sensitivity of mycelial growth to 2,4-DAPG was observed. Work by
Mazzola et al. (1995) showed that various isolates of the take-all fungus G. graminis var. tritici
can differ substantially in their sensitivity toward 2,4-DAPG; isolates that were relatively
B
2,4-DAPG conc. (µ8 g/ml)
0.0 0.8 2.0 2.4 2.8 3.2 4.0 5.2 6.4
Ger
min
atio
n (
%)
0
20
40
60
80
100
e
dd
c
baaaa
A
2,4-DAPG conc. (µ8 g/ml)
0.0 0.8 2.0 2.4 2.8 3.2 4.0 5.2 6.4
Ger
min
atio
n (
%)
0
20
40
60
80
100
a
c
b
b
b
b b
cc

C9 h:
a; pt<>= e? r @ 5: ACB ED
ffEFE
e? cG t = oH f 2,4-DEJILK MCNPO
AQ
PR G S oH n T PU
yV tW hX iY uZ m[
110
F\
i]g^ u_ r` e a 6.
bdcEe
fffgf
ea ch t i oj f f
pk H l
oj n m an ch ti i] vo i]ti y p oj f
f2,4q rts
-u Dv Aw
Px
G y
an g^ an i] nm sz t i P{
. | ul}(~ t� i� m� u} m� vo an r` . c s� p� o� r� a� n� g� i�i�f�e� r� u} m� (� P{ u} s� )� .c Growth
rate of Pus on PDA with buffered pH amended with different concentrations of 2,4-DAPGwas determined by measuring the radial mycelial growth every 24 hours during seven daysof incubation at 25°C. Means of ten replicates are shown. Bars of different shadingcorrespond to different concentrations of 2,4-DAPG (µg ml-1). Error bars represent thestandard error of the mean.
insensitive to 2,4-DAPG in vitro could no longer be suppressed in situ by a 2,4-DAPG-
producing Pseudomonas strain. Their work clearly illustrated that variation in sensitivity within
target pathogen populations to specific metabolites produced by antagonistic microorganisms
can be an important factor in the inconsistency of biological control encountered under field
conditions. The variation in sensitivity among and within Pythium species to 2,4-DAPG
combined with the fact that many pathogenic species can occur simultaneously at the same site
(Chamswarng and Cook, 1985), may have a significant impact on the success, or at least the
consistency, of biological control provided by 2,4-DAPG producing strains.
The role of environmental conditions in 2,4-DAPG production has been studied both
in vitro and in situ for a number of Pseudomonas strains (Shanahan et al., 1992; Duffy and
Défago, 1999; Notz et al., 2001). These studies demonstrated that the regulation of
production of this antibiotic through physiological factors may vary among strains. For
example, 2,4-DAPG production was stimulated by glucose in P. fluorescens strain CHA0 and
repressed in strain F113. In this study, we showed that pH has a significant effect on the
activity of 2,4-DAPG against mycelial growth of Pus: the lower the pH the higher the activity
of 2,4-DAPG (Fig. 6). These results suggest that the temporal and spatial changes that may
occur in the nutritional and physical conditions of microsites where 2,4-DAPG- producing
strains are operating will influence not only the production but also the activity of 2,4-DAPG.
Changes in pH in the rhizosphere of plants, growing in agricultural soils, by as much as 2 pH
units may occur (Nye, 1981). In general, the pH changes induced by roots lead to

C� h�
a� pt�>� e� r � 5: �C� E�
ff�F�
e� c� t � o� f 2,4-D�J�L� �C�P�
A�
P G ¡ o� n ¢ P£
y¤ t¥ h¦ i§ um©
111
Fª
i«g¬ u r® e ¯ 7.
°²±A³
c´ tµ i« v¶ i«tµ y · o¸ f
¹dº
i«f¹f¹e¯ r® e¯ n» t µ p¼ h
½l¾o¸ r® o¸ g¬ l
¾u c´ i« n» o¸ l
¾dº
e¯ r® i« v¶ a¿ tµ i« v¶ e¯ s À a¿ g¬ a¿ i«n» sÀ t µ mÁ y· c´ e¯ l¾ i« a¿ l ¾ g¬ r® o¸ w tµ h
½o¸ f ¹
PÃ
. Ä uÅ lÆtÇ iÈ mÉ uÅ mÉ v¶ a¿ r® .±
sÊ pË oÌ rÍ aÎ nÏ gÐ iiÈÑÈ
fÒeÓ rÍ uÅ m É (Ô PÃ uÅ sÊ )Õ .± 2,4-DAPG, MAPG, and PG were added to PDA plates (pH 6.5) at
different concentrations. Pus mycelial growth was measured every 24 hours during 7 daysof incubation at 25°C. Mean values of five replicates are shown. Error bars represent thestandard error of the mean.
acidification, with more pronounced effects observed for dicot plants (Marschner and
Römheld, 1983). Also microorganisms, including pathogenic fungi, can alter the pH to make
nutrients or trace elements more readily available (Cunningham and Kuiack, 1992). As a result
of these changes in pH, the activity of antimicrobial compounds produced by competing
microorganisms may change. In this study, we observed that P. ultimum var. sporangiiferum
acidifies unbuffered, liquid medium (potato dextrose broth) in a 7-day period from pH 6.5 to
4.5 (data not shown). This reduction in pH and coordinate increase in toxicity of 2,4-DAPG
may explain some of the discrepancies in inhibitory concentrations of 2,4-DAPG reported in
the different experiments presented in this study (Table 1, Fig. 6). To our knowledge, this is
the first time that the effect of pH on the toxicity of 2,4-DAPG has been reported. The lower
activity of 2,4-DAPG at high pH is possibly related to a dissociation of the hydroxyl-
substituents of 2,4-DAPG, which may hamper accumulation into the target cell (M.A. de
Waard, pers. comm.). Similar results have been described for another antibiotic, phenazine-1-
carboxilic acid (PCA), produced by P. fluorescens 2-79, a strain that was more active against G.
graminis var. tritici at lower pH (Brisbane et al., 1987). A comparison between PCA from
strain 2-79 and phenazine-1-carboxamide (PCN) from P. chlororaphis PCL 1391 showed that
both phenazines have the same activity at pH levels lower than 5.0. However, PCN remained
active at higher pH, whereas the activity of PCA decreased significantly (Chin-A-Woeng et al.,
1998). Nevertheless, no determinative effect of pH was found when strain 2-79 was
introduced in soil (Ownley et al., 1992). Probably, pH exerts greater influence in microsites in
the rhizosphere, which are considered to be the courts of interaction between biocontrol agents
Concentration (µÖ g/ml)
0 16 32 48 64 80 100 200 300
Gro
wth
rat
e (m
m/d
ay)
0
5
10
15
20
252,4-DAPGMAPGPG

C× hØ
aÙ ptÚ>Û eÜ r Ý 5: ÞCß Eà
ffáFá
eÜ câ t Û oã f 2,4-DáJäLå æCçPè
Aé
Pê G ë oã n ì Pí
yî tï hð iñ uò mó
112
Fô
iõgö u÷ rø e ù 8.
ú²ûEü
lýeù cþ tÿ rø o� n � m� i
õcþ rø o� gö rø a� p� h
�s � o� f
�h�
y� p� h�
a� e ù o� f �
P
. u� l�t i� m� u� m� v� a� rø . û s� p� o� r� a� n� g� ii
���f�e� r� u� m � g� rø o� w� i
õn� g ö o� n � K
�iõn� gö ’
�s � B�
m� eù d� iõu÷ m � i
õn � a� b s� eù n� cþ e ù (! A" , # B
�) $
o� r ø iõn � p� rø eù s� eù n� cþ e ù o� f
�2%
,# 4& -' D( A"
P)
G *
(! C +
- ' Fô
)$.û Sections were stained either
by double contrast (A, C, E, F) or with the PATAg reaction (B, D). c = cytoplasm, dc =degenerated cytoplasm, er = endoplasmic reticulum, rer = rough endoplasmic reticulum, l =lipid granule, m = mitochondria, n = nucleus, pm = plasma membrane, pma= plasmamembrane alteration, r = ribosomes, smv = smooth membranous vesicles, v = vacuole, w =electron translucent wall. Bar = 1 µm.
AA B
C D
E F
v
w
mv
n
v
v
msmvpma
rer
rv
mer
l
l
mdc
m
v
m
n
v
rer
smv
m
pm
pm
r r
r
v
c

C, h-
a. pt/10 e2 r 3 5: 465 E7
ff898
e2 c: t 0 o; f 2,4-D8=<?> @6ACB
AD
PE G F o; n G PH
yI tJ hK iL uM mN
113
FO
iPgQ uR rS e T 9.
UWVEX
lYeT cZ t[ rS o\ n ] m^ i
PcZ rS o\ gQ rS a_ p` h
as b o\ f
cha
yd p` ha
a_ e T o\ f c
Pe
. f ug lhti ij mk ug mk vl a_ rS . V sm pn oo rp aq nr gs ii
jtjfuev rp ug mk (w Pe ug sm ) x i
Pmmy uR n] o\ l
Ya_ bz eT lY lY eT d
{w| iPt[ ha
a _ β} (w 1~ -� 3� ) x
gQ lYuR cZ o\ p` yd rS a_ n] o\ sb e T m^ o\ n] o\ cZ lY o\ n] a_ l Y a_ n] t[ iP bz o\ d
{yd .V Control sections without primary antibody
(A), hyphae of Pus growing in absence (B) or in presence of 2,4-DAPG (400 µg disc-1)(C). Arrows indicate immunolabelling. Bar=1 µm.
and root pathogens. A better understanding of the interactions in these microsites is being
explored by molecular techniques including the use of reporter genes (Bloemberg et al., 2000;
Sankarasubramanian and Kaushik, 2001; Notz et al., 2001).
The differences in activity of the phloroglucinol derivatives PG, MAPG, and 2,4-DAPG
indicated that the higher the level of acetylation the higher the activity against mycelial growth
of Pus (Fig. 7). Given that in several Pseudomonas strains, the amounts produced and the ratio
of MAPG to 2,4-DAPG may vary depending on biotic and abiotic factors (Duffy and Défago,
1999), one may envisage that also the biocontrol activity will change accordingly.
T�
a_ bz lYe T 2. �WV
EX
ffc�c
eT cZ t [ o\ f c
2�
,4��� -� D� A�
P�
G �
o\ n ] cZ eT ll Y�Y sb t[ rS uR cZ t[ uR rS e T o\ f c
ha
yd p` ha
a_ l Y
t[ iP p` s b o\ f c
Pe
. f ug lhti ij mk ug mk vl a_ rS . V sm pn oo rp aq nr gs ii
jtjfuev rp ug mk (w Pe ug sm )x .V
Cell structures were scored by transmission electron microscopy and divided into 4categories.
Cell structure Control(N=50)
2,4-DAPG(N=29)
Dense cytoplasm with few vacuoles 16 3Many vacuoles in the cell lumen 56 7Beginning of cell disorganisation 24 66Deteriorated or empty cell content 4 24
A B C

C� h�
a� pt�1� e� r � 5: �6� E�
ff�9�
e� c� t � o� f 2,4-D�=�?� �6�C�
A�
P� G � o� n P¡
y¢ t£ h¤ i¥ u¦ m§
114
Although many activities of 2,4-DAPG are well documented (James, 1979; Yajima
and Munakata, 1979; Arisawa et al., 1990; Tada et al., 1990; 1992; Keel et al., 1992; Cronin
et al., 1997a; Te-Chato and Lim, 1999; Lawler et al., 1999; Ito et al., 2000; Debabrata and
Naik, 2000; Jimenez-Escrig et al., 2000), its mode of action was unclear. Phloroglucinol
derivatives were reported as potent inhibitors of the photosystem II in plants (Yoneyama et al.,
1990; Trebst et al., 1984). Typical ultrasructural changes observed in fungi exposed to other
phenolic fungicides such as quintozene, chloroneb, and phenylphenol, include lysis of the
inner mitochondrial membranes, vacuolization of the nuclear envelope and increases in cell
wall thickness (Lyr, 1995). In our studies, however, no such symptoms were observed in Pus
hyphal tips exposed to 2,4-DAPG. Instead, TEM observations (Figs. 8, 9) showed that 2,4-
DAPG does not affect the cell wall structure and composition of hyphal tips of Pus, since β(1-
3), β(1,4), and β(1,6) glucans were equally present in hyphal tips exposed or not to 2,4-DAPG
(Figs. 8B, D, 9B, C). The extensive network of the membranous vesicles closely associated
with the rough endoplasmic reticulum are likely to be transport or secretion vesicles which
may accumulate as a result of stimulation of the development and activity of the Golgi
apparatus by 2,4-DAPG. Alternatively, the accumulation of secretion vesicles derived from
Golgi bodies may result from inhibition of hyphal elongation through a yet unknown
mechanism of action of 2,4-DAPG. Localised alteration in plasma membrane organisation
may indicate disruption or proliferation, due to increased Golgi vesicle trafficking.
M¨
A©
Tª E« R¬ IA©
L®
S ¯ A©
N°
D ±
ME« Tª H²
O³ D±
S¯O³ r´ gµ a¶ n· is¹ mº s ¹ a¶ n· d » c¼ u½ l
¾t¿ u½ r´ e À c¼ oÁ n· d» i¸ t¿ ioÁ n· s¹ . Characteristics
of the Pythium spp. used in this study are shown inTable 1. Pythium spp. were routinely grown onpotato dextrose agar (PDA; Oxoid Ltd, Basingstoke,Hampshire, England) or on V8-juice agar (V8;N.V.Campbell Foods, Puurs, Belgium) at 25°C.For production of zoospores, P. ultimum var.sporangiiferum (Pus) was grown on V8 amendedwith 10 g l-1 CaCO3 (V8+). Mycelial plugs of allPythium isolates were stored in sterile mineral oil at15°C. Pseudomonas fluorescens strain P60 wasisolated from the wheat rhizosphere and identifiedwith the API 20 NE test (BioMérieux sa., Lyon,France). Strain P60 was grown on Pseudomonasagar F (PSA; Difco laboratories, Detroit, MI, USA)or on King’s medium B (KMB) agar (King et al.,1954). The bacterial strain was stored at – 80°C inKMB broth supplemented with glycerol to a finalconcentration of 40% (vol/vol). For transmissionelectron microscopy studies, Pus was grown onKMB agar.
A©
n· t¿ ib i¸ oÁ t¿ ic¼ s¹ . à The antibiotics used werephloroglucinol (PG), mono-acetylphloroglucinol(MAPG), and 2,4-diacetylphloroglucinol (2,4-DAPG). PG (1,3,5-trihydroxybenzene) and MAPG(2,4,6-trihydroxyacetophenone monohydrate) wereobtained from Sigma-Aldrich Chemie GmbH(Steinheim, Germany). 2,4-DAPG was purifiedfrom cultures of P. fluorescens strain P60 by highpressure liquid chromatography (HPLC) followedby photodiode array spectroscopy (PDA) todetermine peak purity and identity. Purificationprotocols and the HPLC-PDA system weredescribed previously (Raaijmakers et al., 1999). Allantibiotics were dissolved in 96% ethanol (MerckKGaA, Darmstadt, Germany). In experimentswhere activity of the antibiotics was determined, alltreatments including controls had the sameconcentration of ethanol.
WÄ
hÅ
eÀ a¶ t ¿ rooÁÆÁ t ¿ eÀ xÇ u½ d» a¶ t¿ eÀ s¹ .Ã Seeds of wheat cultivarPagode were surface sterilized by soaking in 96%

CÈ hÉ
aÊ ptË1Ì eÍ r Î 5: Ï6Ð EÑ
ffÒ9Ò
eÍ cÓ t Ì oÔ f 2,4-DÒ=Õ?Ö ×6ØCÙ
AÚ
PÛ G Ü oÔ n Ý PÞ
yß tà há iâ uã mä
115
ethanol for 30 sec, washing once with steriledemineralised water followed by agitation in 1.5%NaOCl supplemented with 1% Tween 20. After1.5 h, seeds were washed three times in steriledemineralised water, incubated again for 15 min in1.5% NaOCl and washed three times in steriledemineralised water. Seeds were germinated on 1/5th
strength PDA plates at 18ºC in a climate chamberwith a 12-h photoperiod. Plates showing any sign ofcontamination were discarded. After 11 days,germinated seedlings without seed remnants weretransferred to culture tubes (De Wit culture tubes,LAB Associated BV, Oudenbosch, TheNetherlands) containing 5 ml of sterile diluted (2:3,vol/vol) Hoaglund’s solution. After 7 daysincubation at 18°C in a climate chamber with a 12-h photoperiod, plants were removed and thenutrient solutions containing the root exudates werechecked for contamination by plating samples ontoKMB and PDA. Contaminated samples werediscarded. Sterile root exudates were stored at -20°C.
Eå ffæçæ
eè cé t ê oë f æ
2ì ,í 4î -ï Dð Añ
Pò G ó oë n ô mõ yö cé eè l÷ iø aù l ÷ gú rû oë wü tê h ý oë f æ
dþ
iø ffæçæ eè rû eè nô têPÿ y� t� h� i� u� m� s� p� eè cé iø eè s� . � Effect of 2,4-DAPG on mycelialgrowth of fourteen different Pythium isolates wastested on PDA plates amended with 2,4-DAPG.The initial pH of the agar was adjusted to 6.5. 2,4-DAPG concentrations ranged from 0 to 300 µg ml-1
with intervals of 8 µg ml-1. Mycelial plugs (0.5-cmdiameter) were obtained from 3-day old culturesgrown on PDA and transferred to PDA platesamended with different concentrations of 2,4-DAPG. Mycelial growth was recorded every 24h forseven days. The EC50 value was assessed and isdefined as the concentration necessary to inhibit50% of the mycelial growth. The minimalinhibitory concentration (MIC) was determinedand is defined as the minimum concentrationnecessary to completely (100%) inhibit mycelialgrowth. Each treatment had six replicates and theexperiment was performed three times.
Eå ffæçæ
eè cé t ê oë f æ
2ì ,í 4î -ï Dð Añ
Pò G ó oë n ô z ooëÆë s� p� oë rû aù nô gú iø aù . � Mycelialplugs of 1.5 cm-diameter were cut from Pus agarcultures grown for 3 days on V8+ at 25°C. Mycelialplugs were transferred to petri-plates and floodedwith 15 ml of solutions containing 2,4-DAPG atfinal concentrations of 0.0, 2.0, 4.0, 8.0, and 16.0µg ml-1. After four days at 25°C, the number ofzoosporangia formed in one microscopic field atmagnification of 100X (Leitz Wetzlar Dialux 20EB, Germany), corresponding to an area of 3.14mm2, was measured. The diameter of 10 randomlyselected zoosporangia formed per treatment afterfour days was measured with an ocular micrometer(Periplan 10X M, Leitz). Each treatment had fivereplicates and the experiment was performed twice.
Eå ffæçæ
eè cé t ê oë f æ
2ì ,í 4î -ï Dð Añ
Pò G ó oë n ô z ooëÆë s� p� oë rû eè s � aù nô d þ
z oë oë s� p� oë rû eècé yö s� tê s� .� Zoospores of Pus were produced using amodification of a method described by Zhou and
Paulitz (1993). Pus was grown on V8+ for threedays at 25°C. The full-grown agar plates were cutinto 2-cm wide strips and one half of the strips wastransferred to another petri plate; both plates wereflooded with 20 ml of sterile water and kept at25°C. After 1 h, the water was discarded andreplaced with the same volume of water. Plates wereincubated at 18°C for four days, water was removedand replaced with the same volume and incubatedat 18°C for 2 h; zoospores were collected andconcentrated using a 0.45 µm nylon filter (ZapCap;Schleicher & Schuell GmbH, Dassel, Germany).Supensions containing 20,000 to 40,000 zoosporesml-1 were typically obtained. Aliquots of 50 µl of asuspension containing 25,000 zoospores ml-1 of Puswere transferred to 96-well microtiter plates(Greiner BV, Alphen aan den Rijn, TheNetherlands) and mixed in a 1:1 ratio withsolutions of 2,4-DAPG to obtain final 2,4-DAPGconcentrations of 0.0, 0.2, 0.4, 0.8, 1.6, and 3.2 ngml-1. The number of swimming, encysted,germinated, and disintegrated zoospores werecounted under an inverted microscope (Axiovert10, Carl Zeiss, Jena, Germany). Treatments hadfive replicates and the experiment was performedfive times.
The effect of 2,4-DAPG on germinationof encysted zoospores was studied in more detail inseparate experiments. These experiments wereperformed both in water and in wheat rootexudates. To study germination in water,encystment of zoospores of Pus was induced byvortexing a suspension containing 25,000 zoosporesml-1 for 25 sec. Encysted zoospores were transferredto 96-well microtiter plates and mixed with 2,4-DAPG solutions to obtain final 2,4-DAPGconcentrations of 0, 0.8, 1.6, 2.0, 2.4, 2.8, 3.2, 4.0,5.2, and 6.4 µg ml-1. To study the effect of wheatroot exudates, a zoospore suspension containing50,000 zoospores ml-1 was vortexed for 25 sec;encysted zoospores were transferred to microtiterplates and diluted with root exudates to obtain thesame final 2,4-DAPG concentrations describedabove. The final concentration of root exudates washalf of the initial root exudates obtained asdescribed previously. Microtiter plates wereincubated at 25°C and after 2 h, a drop of 96%ethanol was added to each well to stop germination.The number of germinated zoospore cysts wasdetermined microscopically. Prior to addition of2,4-DAPG and wheat root exudates to zoosporesuspensions, the initial number of germinatedencysted zoospores was determined microscopically.Treatments had five replicates each and theexperiment was performed twice.
Eå ffæçæ
eè cé t ê oë f æ
aù cé eè tê yö l÷ aù tê iø oë n ô oë n ô aù cé tê iø v iø tê y ö oë f æ
p� hý l÷ oë rû oë gú l÷ u� cé iø nô oë l÷dþ
eè rû iø v aù tê iø v eè s� .� The effect of the level of acetylation onthe activity of phloroglucinol derivatives was testedon mycelial growth of Pus. PG, MAPG, and 2,4-DAPG have no, one, and two acetyl groups,respectively (Fig. 1). Pus was grown on PDA

C� h
a� pt��� e� r � 5: ��� E�
ff���
e� c� t � o� f 2,4-D����� �!#"
A$
P% G & o� n ' P(
y) t* h+ i, u- m.
116
amended with PG, MAPG, or 2,4-DAPG to finalconcentrations of 0.0, 16.0, 32.0, 48.0, 64.0, 80.0,100.0, 200.0, and 300.0 µg ml-1. Mycelial growthwas measured every 24 h for seven days and growthrates (mm day-1) were calculated. Treatments hadfive replicates and the experiment was performedthree times.
E/ ff010
e2 c3 t 4 o5 f 0
p6 H 7 o5 n 8 a9 c3 t4 i: v; i:t4 y < o5 f
02=
,> 4? -@ DA AB
PC GD
.E Pus wasgrown on PDA buffered with 0.0015 M citric acid(0.001 M C6H8O7.1H2O; 0.002 MNa2HPO4.2H2O) to a pH of 4.5, 5.5, 6.5, 7.5, or8.0. PDA plates with different pH were amendedwith 2,4-DAPG to final concentrations of 0.0,16.0, 24.0, 32.0, and 40.0 µg ml-1. Plates wereincubated at 25°C and Mycelial growth wasmeasured every 24 h for seven days. Each treatmenthad 10 replicates and the experiment was performedthree times.
SF t4 a9 t4 i: c 3 e2 f0 f0 e2 c3 t 4 o5 f 0
2=
,> 4? -@ DA AB
PC G D
o5 n 8 mG y< c3 e2 lH i: a9 l H gI rJ o5 wK t4 hL .E Puswas grown on PDA for three days and 0.5-cm-diameter mycelial plugs were transferred to PDAplates amended with 2,4-DAPG to finalconcentrations of 0, 8, 16, 32, 48, 64, 96, 128,200, and 300 µg ml-1. After seven days, mycelialplugs that failed to grow on PDA amended with2,4-DAPG were again transferred to PDA (pH 6.5)without 2,4-DAPG. Growth was recorded up toseven days after transfer. Treatments had fivereplicates and the experiment was perfrmed twice.
TM E/ M N
sO t4 uP dQ i:e2 sO .E Mycelial plugs of 6 mm-diameter
were cut from Pus agar cultures grown for 3 days onKMB at 25°C. Mycelial plugs were transferred tothe center of Petri plates containing KMB. Sterileantibiotic assay discs of 9 mm-diameter (Whatman,Springfield Mill, England) were imbibed withsolutions containing 2,4-DAPG at finalconcentrations of 0.0, 25, 50, 100, 200, and 400 µl.Discs were placed at a distance of 3 cm from themycelial plugs. Petri dishes were sealed withparafilm and incubated 5 to 6 days in the dark at25°C. Five samples of 2 mm2 of agar were collectedat the margin of Pus cultures exposed or not to 400µg of 2,4-DAPG per disc. Samples were fixedovernight at 4°C in 2% glutaraldehyde buffered in0.1M cacodylate (pH 7.2), washed in the samebuffer, then dehydrated in a graded ethanol seriesand embedded in Epon (Prolabo, France) or LRWhite medium grade resin (Oxford instrument,France).
Epon embedded samples were postfixedwith 1% osmium tetroxide before ethanol
dehydration (Gianinazzi and Gianinazzi- Pearson,1992). Ultrathin sections (0.1µm) were cut with adiamond knife (Reicher Ultracut microtone). Forultrastructural observations, Epon sections were cutonto copper grids and double contrasted by poststaining with 3% uranyl acetate in 50% ethanol(Valentine, 1961) followed by lead citrate(Reynolds, 1963). To locate β(1-4) and β(1-6)glycosylated compounds (glycoproteins andpolysaccharides), Epon embedded samples weresubjected to the periodic acid thiocarbohydrazidesilver proteinate reaction (PATAg) according toThiéry (1967).
LR White embedded sections werecollected on golden grids coated with collodion foranalytical immunocytochemistry using an indirectimmunogold labelling technique for the detectionof β(1-3) glucans (Lemoine et al., 1995). Sampleson grids were incubated overnight at 4°C in amouse monoclonal antibody raised against β(1-3)glucopyranose polymers (Biosupplies Australia,Australia) which has previously been shown to bespecific for β(1-3) glucans of the cell wall (Lemoineet al., 1995). The antibody was diluted 1000 or 500times in Tris buffer saline with 1% bovine serumalbumin. After rinsing, sections were incubated for1h in a gold-labelled (15 nm) goat antimousepolyclonal antibody (Tebu Biocell, France) diluted20 times to detect the primary antibody.Imunological control sections were prepared byomitting the monoclonal antibody during the firstincubation step. All sections were counter stainedfor 10 min with 3% aqueous uranyl acetate beforeobservation in a transmission electron microscope(Hitachi H7500) at 80 kV.
The effect of 2,4-DAPG on Pus myceliumwas evaluated by quantifying hyphal responses onEpon-embeded double contrasted sections in thefollowing categories: 1) dense cytoplasm with fewvacuoles (young); 2) many vacuoles in the celllumen; 3) beginning of cell content disorganisation;4) deteriorated or empty cell contents (dead). Theeffect of 2,4-DAPG on Pus cell wall synthesis wasdetermined by observing LR White imunogoldlabelled sections. All experiments were repeatedtwice.
SF t4 a9 t4 i: sO t4 i: c3 a9 l H a9 n8 a9 lH y< sO i: sO .E Data were analysed by ANOVAfollowed by Tukey’s studentized range test aftercertifying normal distribution and homogeneity ofvariances (SAS institute, Inc., Cary, N.C.). Allexperiments were repeated at least twice andrepresentative results are shown.
AB
CR
KS NT OU WV
LW
E/ DA GD
E/ MN
E/ NT TM SFThis research was financially supported by CAPES, Brazil (Project nr. 1515/96-9). We are grateful to
Dr. M.A. de Waard and Dr. Pierre de Wit for critically reading the manuscript and for their valuable suggestions.The authors gratefully acknowledge the practical assistance of Therese Corberand and Dennis Kraaijeveld.

CX hY
aZ pt[�\ e] r ^ 5: _�` Ea
ffb�b
e] cc t \ od f 2,4-Db�e�f g�h#i
Aj
Pk G l od n m Pn
yo tp hq ir us mt
117
Ru Ev Fw
Ev Ru Ev Nx
Cy
Ev Sz
A{
r| i} s~ a� w� a� , � M� .� , � Fw
u� j�i} t� a� , � A
{., ��� H
�a� y� a� s~ h� i} , � T� ., ��� H
�a� y� a� s~ h� i} , � K� .,���
O� c� h� i} a� i} , � H�
., ��� a� n� d � M� o� r| i} t� a� , � Nx
.� 1990. Cytotoxic andantiherpetic activity of phloroglucinol derivativesfrom Mallotus japonicus (Euphorbiaceae). Chem.Pharmaceut. Bull. 38:1624-1626.B� a� n� g� e� r| a� , � M� .� G� ., ��� a� n� d � T� h� o� m� a� s~ h� o� w� , � L
�.� Sz .� 1996.
Characterization of a genomic locus required forsynthesis of the antibiotic 2,4-diacetylphloroglucinol by the biological controlagent Pseudomonas fluorescens Q2-87. Mol. Plant-Microbe Interac. 9:83-90.B� a� n� g� e� r| a� , � M� .� G� ., ��� a� n� d � T� h� o� m� a� s~ h� o� w� , � L
�.� Sz .� 1999.
Identification and characterization of a gene clusterfor synthesis of the polyketide antibiotic 2,4-diacetylphloroglucinol from Pseudomonas fluorescensQ2-87. J. Bacteriol. 181:3155-3163.B� l� o� e� m� b� e� r| g� , � G� .� V� ., ��� W
�i} j� f� j� e� s~ , � A
{.� H� .� M� ., ��� L
�a� m� e� r| s~ ,�
G� .� Ev .� M� ., ��� Sz t� u� u� r| m� a� n� , � Nx
., ��� a� n� d � L�
u� g� t� e� n� b� e� r| g� , � B� .� J� .� J� .�2000. Simultaneous imaging of Pseudomonasfluorescens WCS365 populations expressing threedifferent autofluorescent proteins in therhizosphere: New perspectives for studyingmicrobial communities. Mol. Plant-MicrobeInteract. 13:1170-1176.B� l� u� m� e� r| , � C
y., ��� a� n� d � H
�aa��� s~ , � D .� 2000. Iron regulation of
the hcnABC genes encoding hydrogen cyanidesynthase depends on the anaerobic regulator ANRrather than on the global activator GacA inPseudomonas fluorescens CHA0. Microbiol.146:2417-2424.B� o� k¡ e� s~ c� h� , � H
�.� Ru ., ��� G� r| o� w� e� i} ss~¢~ , � A
{., ��� M� c� K� ee�£� , � C
y.� T� ., ��� a� n� d�
B� o� y� d� , � M� .� Ru . � 1999. Laxifloranone, a newphloroglucinol derivative from Marila laxiflora. J.Nat. Prod. 62:1197-1199.B� o� n� s~ a� ll��� , � Ru .� Fw ., ��� W
�e� l� l� e� r| , � D .� M� ., ��� a� n� d � T� h� o� m� a� s~ h� o� w� , � L
�.� Sz .�
1997. Quantification of 2,4-diacetylphloroglucinolproduced by fluorescent Pseudomonas sp. in vitroand in the rhizosphere of wheat. Appl. Environ.Microbiol. 63:951-955.B� r| i} s~ b� a� n� e� , � P
¤.� G� ., ��� J
�a� n� i} k¡ , � L
�.� J� ., ��� T� a� t� e� , � M� .� Ev ., ��� a� n� d�
W�
a� rr|¥| e� n� , � Ru .� Fw .� O� .� 1987. Revised structure for thephenazine antibiotic from Pseudomonas fluorescens2-79 (NRRL B-15132). Antimic. AgentsChemother. 31:1967-1971.Cy
h� a� m� s~ w� a� r| n� g� , � Cy
., ��� a� n� d � Cy
oo�¦� k¡ , � Ru .� J� .� 1985.Identification and comparative pathogenicity ofPythium species from roots and wheat-field soils inthe pacific northwest. Phytopathol. 75:821-827.Cy
h� i} n� -§ A{ -§ W� o� e� n� g� , � T� .� Fw .� Cy .� , � B� l� o� e� m� b� e� r| g� , � G� ., ��� v¨ a� n � d� e� r|B� i} j� , � A
{., ��� v¨ a� n � d� e� r | D r| i} f� t� , � K� .� M� .� G� .� M� ., ��� Sz c� h� r| i} p© s~ e� m� a� , � J
�.,���
K� r| oo�¦� n� , � B� ., ��� Sz c� h� e� ff�1� e� r| , � Ru .� J� ., ��� K� ee�ª� l� , � Cy
., ��� B� a� kk¡¦¡ e� r| ,�P¤
.� A{ .� H� .� M� ., ��� T� i} c� h� y� , � H�
.� -§ V� ., ��� d� e � B� r| u� i} j� n� , � Fw
.� J� ., ��� T� h� o� m� a� s~ -§O� a� t� e� s~ , � J
�.� Ev ., ��� a� n� d � L
�u� g� t� e� n� b� e� r| g� , � B� .� J� .� J� .� 1998. Biocontrol
by phenazine-1-carboxamide-producingPseudomonas chlororaphis PCL1391 of tomato rootand crown rot caused by Fusarium oxysorum f.sp.radicis-lycopersici. Mol. Plant-Microbe Interac.11:1069-1077.
Cy
r| o� n� i} n� , � D ., ��� M� o� e� nn�«� e� -§ L� o� cc�£� o� z¬ , � Y
., ��� Fw
e� n� t� o� n� , � A{
.,���D u� nn�«� e� , � C
y., ��� D o� w� l� i} n� g� , � D .� Nx . � a� n� d � O� ’® G� a� r| a� , � F
w.�
1997(a). Role of 2,4-diacetylphloroglucinol in theinteractions of the biocontrol pseudomonad strainF113 with the potato cyst nematode Globoderarostochiensis. Appl. Environ. Microbiol. 63:1357-1361.Cy
r| o� n� i} n� , � D ., ��� M� o� e� nn�«� e� -§ L� o� cc�£� o� z¬ , � Y
., ��� Fw
e� n� t� o� n� , � A{
.,���D u� nn�«� e� , � C
y., ��� D o� w� l� i} n� g� , � D .� Nx . � a� n� d � O� ’® G� a� r| a� , � F
w.�
1997(b). Ecological interaction of a biocontrolPseudomonas fluorescens strain producing 2,4-diacetylphloroglucinol with the soft rot potatopathogen Erwinia caratovora subsp. atroseptica.FEMS Microbiol. Ecol. 23:95-106.Cy
u� nn�«� i} n� g� h� a� m� , � J�.� Ev ., ��� a� n� d � K� u� i} a� c� k¡ , � C
y.� 1992.
Production of citric and oxalic acids andsolubilization of calcium-phosphate by Penicilliumbilaii. Appl. Environ. Microbiol. 58:1451-1458.D e� a� c� o� n� , � J
�.� W� ., ��� a� n� d � D o� n� a� l� d� s~ o� n� , � Sz .� P¤ .� 1993.
Molecular recognition in the homing responses ofzoosporic fungi, with special reference to Pythiumand Phytophthora. Mycol. Res. 97:1153-1171.D e� b� a� b� r| a� t� a� , � Sz ., ��� a� n� d � N
xa� i} k¡ , � P
¤.� 2000. Phloroglucinol
enhances growth and rate of axillary shootproliferation in potato shoot tip cultures in vitro.Plant Cell Tissue Organ Cult. 60:139-149.D i} c� k¡ , � M� .� W� . � 1990. Keys to Pythium. M.W. Dick,Reading, England. Pages 1-64.D u� f
�f�y� , � B� .� K� ., ��� a� n� d � D éf
�a� g� o� , � G� .� 1997. Zinc improves
biocontrol of fusarium crown and root rot oftomato by Pseudomonas fluorescens and represses theproduction of pathogen metabolites inhibitory tobacterial antibiotic biosynthesis. Phytopathol.87:1250-2157.D u� f
�f�y� , � B� .� K� ., ��� a� n� d � D é¯ f� a� g� o� , � G� . � 1999. Environmental
factors modulating antibiotic and siderophorebiosynthesis by Pseudomonas fluorescens biocontrolstrains. Appl. Environ. Microbiol. 65:2429-2438.Ev s~ t� r| a� d� a� -§ G� a� r| c� i} a� , � T� ., ��� Ru a� y� , � T� .� Cy .� , � G� r| ee�£� n� , � J
�.� Ru ., ��� C
ya� l� l� o� w� ,�
J�.� A{ ., ��� a� n� d � K� e� nn�«� e� d� y� , � J
�.� Fw .� 1990. Encystment of
Pythium aphanidermatum zoospores is induced byroot mucilage polysaccharides, pectin and amonoclonal antibody to a surface antigen. J. Exp.Bot. 41:693-699.Fw
e� n� t� o� n� , � A{
.� M� ., ��� Sz t� e� p© h� e� n� s~ , � P¤
.� M� .� , � Cy
r| o� w� l� e� y� , � J�.,���
O� ’® Cy a� l� l� a� g� h� a� n� , � M� ., ��� a� n� d � O� ’® G� a� r| a� , � Ru .� 1992.Exploitation of gene(s) involved in 2,4-diacetylphloroglucinol biosynthesis to confer a newbiocontrol capability to a Pseudomonas strain. Appl.Environ. Microbiol. 58:3873-3878.G� i} a� n� i} n� a� zz¬°¬ i} , � Sz ., ��� a� n� d � G� i} a� n� i} n� a� zz¬±¬ i} -§ P¤ e� a� r| s~ o� n� , � V
�.� 1992.
Cytology, histochemistry andimmunocytochemistry as tools for studyingstructure and function in endomycorrhiza. Meth.Microbiol. 24:109-139.I²s~ h� i} g� u� r| o� , � K� ., ��� N
xa� g� a� r| e� y� a� , � N
x., ��� a� n� d � F
wu� k¡ u� m� o� t� o� , � H
�.�
1998. A phlroglucinol derivative from cellsuspension cultures of Hypericum palatum.Phytochem. 47:1041-1043.

C³ h´
aµ pt¶�· e¸ r ¹ 5: º�» E¼
ff½�½
e¸ c¾ t · o¿ f 2,4-D½�À�Á Â�Ã#Ä
AÅ
PÆ G Ç o¿ n È PÉ
yÊ tË hÌ iÍ uÎ mÏ
118
IÐ tÑ oÒ , Ó HÔ
., Õ Ó MÖ u× rØ aÙ nÚ aÙ kÛ aÙ , Ó TÜ ., Õ Ó MÖ oÒ rØ iÝ , Ó KÞ ., Õ Ó JßiÝ nÚ -à Zá hâ eã Xä iÝ oÒ nÚ gå ,Ó
TÜ oÒ kÛu× dæ
aÙ , Ó HÔ
., Õ Ó Nç iÝ sè hâ iÝ nÚ oÒ , Ó HÔ
., Õ Ó Yé
oÒ sè hâ iÝ dæ aÙ , Ó TÜ ., Õ Ó aÙ nÚ d æ
JßiÝ nÚ ,Ó
Zá
.Õ Xä . Õ 2000. Ichthyotoxic phloroglucinol derivativesfrom Dryopteris fragrans and their anti-tumorpromoting activity. Chem. Pharmac. Bull. 1190-1195.JßaÙ mê eã sè , Ó Dë .Õ Jß .Õ 1979. The role of auxins and
phloroglucinol in adventitious root formation inRubus and Fragaria in vitro. J. Hortic. Sci. 54:273-277.JßiÝ mê eã nÚ eã zì -à Eí sè cî rØ iÝ gå , Ó A
ï., Õ Ó JßiÝ mê eã nÚ eã zì -à Jß iÝ mê eã nÚ eã zì , Ó IÐ ., Õ Ó P
ðu× lñ iÝ dæ oÒ , Ó R
ò.,Õ Ó
aÙ nÚ d æ
SóaÙ u× rØ aÙ -à Cô aÙ lñ iÝ xõ tÑ oÒ , Ó Fö .Õ 2001. Antioxidant activity of
fresh and processed edible seaweed. J. Sci. FoodAgric. 81:530-534.KÞ eã nÚ y÷ oÒ nÚ , Ó Dë .Õ MÖ ., Õ Ó Dë iÝ xõ oÒ nÚ , Ó G
ø.Õ Rò ., Õ Ó aÙ nÚ d
æHÔ
eã lñ fù eã rØ , Ó Só.Õ 1997.
The repression and stimulation of growth ofErisiphe sp. on Rhododendron by fungicidalcompounds. Plant Pathol. 46:425-431.KÞ eeãªã lñ , Ó C
ô., Õ Ó W
úiÝ rØ tÑ hâ nÚ eã rØ , Ó P
ð.Õ HÔ ., Õ Ó O
ûbü eã rØ hâ aÙ nÚ sè lñ iÝ , Ó TÜ .Õ HÔ .,Õ Ó
Vý
oÒ iÝ sè aÙ rØ dæ , Ó Cô
., Õ Ó Bþ u× rØ gå eã rØ , Ó Uÿ
., Õ Ó HÔ
aaÙ�Ù sè , Ó Dë ., Õ Ó aÙ nÚ d æ
Dë é�fù aÙ gå oÒ , Ó G
ø.Õ
1990. Pseudomonads as antagonists of plantpathogens in the rhizosphere: role of the antibiotic2,4-diacetylphloroglucinol in the suppression ofblack root rot of tobacco. Symbiosis 9:327-342.KÞ eeãªã lñ , Ó C
ô., Õ Ó Sócî hâ nÚ iÝ dæ eã rØ , Ó U
ÿ., Õ Ó MÖ aÙ u× rØ hâ oÒ fù eã rØ , Ó MÖ ., Õ Ó V
ýoÒ iÝ sè aÙ rØ dæ , Ó C
ô.,Õ Ó
L� aÙ v� iÝ llñ�ñ eã , Ó Jß.Õ , Ó Bþ u× rØ gå eã rØ , Ó U
ÿ., Õ Ó Wú
iÝ rØ tÑ hâ nÚ eã rØ , Ó Pð
.Õ HÔ ., Õ Ó HÔ
aaÙ�Ù sè , Ó Dë .,Õ ÓaÙ nÚ d
æDë é�fù aÙ gå oÒ , Ó G
ø.Õ 1992. Suppression of root diseases
by Pseudomonas fluorescens CHA0: importance ofthe secondary metabolite 2,4-diacetylphloroglucinol. Molec. Plant-MicrobeInteract. 5:4-13.KÞ iÝ nÚ gå , Ó E
í.Õ Oû ., Õ Ó W
úaÙ rØ dæ , Ó MÖ .Õ KÞ , Ó aÙ nÚ d
æRò
aÙ nÚ eã y÷ , Ó Dë .Õ Eí .Õ 1954.Two simple media for the demonstration ofpyocyanin and fluorescin. J. Lab. Clin. Med.44:301-307.L� aÙ w� lñ eã rØ , Ó IÐ .Õ Rò ., Õ Ó E
ísè cî hâ lñ eã rØ , Ó Bþ .Õ MÖ .Õ , Ó S
ócî hâ lñ iÝ eã bü sè , Ó Dë .Õ MÖ ., Õ Ó aÙ nÚ d
æFö oÒ lñ eã y÷ , Ó W
ú.Õ Jß .Õ 1999. Relationship between chemical
functional groups on Eucalyptus secondarymetabolites and effectiveness as marsupialantifeedants. J. Chem. Ecol. 25:2561-2573.L� eã mê oÒ iÝ nÚ eã , Ó MÖ .Õ Cô ., Õ Ó G
øoÒ llñ�ñ oÒ ttÑ Ñ eã , Ó A
ï., Õ Ó aÙ nÚ d
æGø
iÝ aÙ nÚ iÝ nÚ aÙ zzì±ì iÝ -àPð
eã aÙ rØ sè oÒ nÚ , Ó Vý
.Õ 1995. Localization of beta(1-3) glucanin walls of the endomycorrhizal fungi Glomusmosseae (Nicol. & Gerd.) Gerd. & Trappe andAcaulospora laevis Gerd. & Trappe duringcolonization of host roots. New Phytol. 129:97-105.L� eã v� y÷ , Ó E
í., Õ Ó Gø
oÒ u× gå hâ , Ó Fö .Õ Jß ., Õ Ó Bþ eã rØ lñ iÝ nÚ , Ó KÞ .Õ Dë ., Õ Ó Gø
u× iÝ aÙ nÚ aÙ , Ó Pð
.Õ Wú .,Õ ÓaÙ nÚ d
æSómê iÝ tÑ hâ , Ó J
ß.Õ TÜ .Õ 1992. Inhibition of Septoria tritici
and other phytopathogenic fungi and bacteria byPseudomonas fluorescens and its antibiotics. PlantPathol. 41:335-341.L� oÒ p� eã rØ , Ó J
ß.Õ Eí ., Õ Ó aÙ nÚ d
æL� iÝ nÚ d
æoÒ w� , Ó S
ó.Õ Eí .Õ 1997. Reporter gene
systems useful in evaluating in situ gene expressionby soil and plant-associated bacteria. In: Hurst,C.J., Knudsen, G.R., McInerney, M.J.,Stetzenbach, L.D., and Walter, M.V. (eds). Manualof Environmental Microbiology, pages 482-492,American Society for Microbiology (ASM),Washington, USA.L� y÷ rØ , Ó H
Ô.Õ 1995. Aromatic hydrocarbon fungicides
and their mechanism of action. In: Lyr, H. (ed).
Modern selective fungicides: properties,applications, mechanisms of action, pages 76-98, 2nd
edition, Gustav Fischer Verlag, Jena, Germany.MÖ aÙ rØ sè cî hâ nÚ eã rØ , Ó H
Ô.Õ , Ó aÙ nÚ d
æRò
ö�mê hâ eã lñ dæ , Ó V
ý.Õ 1983. In vivo
measurement of root-induced pH changes at thesoil-root interface: effect of plant species andnitrogen source. Z. Pflanzenphysiol., 111:241-251.MÖ aÙ u× rØ hâ oÒ ffù1ù eã rØ , Ó MÖ ., Õ Ó KÞ eeã£ã lñ , Ó C
ô., Õ Ó H
ÔaaÙªÙ sè , Ó Dë .Õ , Ó aÙ nÚ d
æDë é�fù aÙ gå oÒ ,Ó
Gø
.Õ 1995. Influence of plant species on diseasesuppression by Pseudomonas fluorescens strain CHA0with enhanced antibiotic production. Plant Pathol.44:40-50.MÖ aÙ zzì°ì oÒ lñ aÙ , Ó MÖ ., Õ Ó Fö , Ó Fö u× j
�iÝ mê oÒ tÑ oÒ , Ó Dë .Õ KÞ ., Õ Ó TÜ hâ oÒ mê aÙ sè hâ oÒ w� , Ó L� .Õ Só .,Õ Ó
aÙ nÚ d æ
Cô
ooÒ¦Ò kÛ, Ó Rò
.Õ Jß .Õ 1995. Variation in sensitivity ofGaeumanomyces graminis to antibiotics produced byPseudomonas spp. and effect on biological control oftake-all of wheat. Appl. Environ. Microbiol.61:2554-2559.Nç eã lñ sè oÒ nÚ , Ó E
í.Õ Bþ ., Õ Ó aÙ nÚ d
æHÔ
sè u× , Ó Jß.Õ Só .Õ TÜ .Õ 1994. Nutritional
factors affecting responses of sporangia of Pythiumultimum to germination stimulants. Phytopathol.84:677-683.Nç oÒ tÑ zì , Ó R
ò., Õ Ó MÖ aÙ u× rØ hâ oÒ fù eã rØ , Ó MÖ ., Õ Ó S
ócî hâ nÚ iÝ dæ eã rØ -à KÞ eeãªã lñ , Ó U
ÿ.Õ ,Ó
Dë u× fù fù y÷ , Ó Bþ ., Õ Ó HÔ
aaÙ�Ù sè , Ó Dë ., Õ Ó aÙ nÚ d æ
Dë é�fù aÙ gå oÒ , Ó G
ø.Õ 2001. Biotic
factors affecting expression of the 2,4-diacetylphloroglucinol biosynthesis gene phlA inPseudomonas fluorescens strains CHA0 in therhizosphere. Phytopathol. 91:873-881.Nç y÷ eã , Ó P
ð.Õ HÔ . Õ 1981. Changes in the pH across the
rhizosphere induced by roots. Plant Soil 61:7-26.Oû
w� nÚ lñ eã y÷ , Ó Bþ .Õ HÔ ., Õ Ó Wú
eã llñ�ñ eã rØ , Ó Dë .Õ MÖ ., Õ Ó aÙ nÚ d æ
TÜ hâ oÒ mê aÙ sè hâ oÒ w� ,ÓL� .Õ Só .Õ 1992. Influence of in situ and in vitro pH onsuppression of Gaeumannomyces graminis var. triticiby Pseudomonas fluorescens 2-79. Phytopathol.82:178-184.Pð
hâ eã lñ oÒ u× zì aÙ tÑ , Ó MÖ .Õ -à Aï , Ó L� aÙ w� rØ eã nÚ cî eã , Ó Fö ., Õ Ó MÖ oÒ u× lñ aÙ y÷ , Ó L� ., Õ Ó Bþ oÒ rØ oÒ tÑ ,ÓCô
., Õ Ó Sócî hâ aÙ eã v� eã rØ bü eã kÛ eã , Ó J
ß., Õ Ó Sócî hâ aÙ eã v� eã rØ bü eã kÛ eã , Ó MÖ ., Õ Ó aÙ nÚ d
æRò
oÒ bü eã rØ tÑ -àGø
eã rØ oÒ , Ó MÖ .Õ 1992. Leishmania dovani: antagonisticeffect of S- adenosyl methionine on ultrastructuralchanges and growth inhibition induced bysinefungin. Exp. Parasitol. 74: 2, 177-187.Rò
aaÙ�Ù iÝ j� mê aÙ kÛ eã rØ sè , Ó Jß.Õ MÖ .Õ , Ó Bþ oÒ nÚ sè aÙ ll ñ�ñ R
ò.Õ Fö ., Õ Ó aÙ nÚ d
æWú
eã lñ lñ eã rØ , Ó Dë .Õ MÖ .Õ1999. Effect of population density of Pseudomonasfluorescens on production of 2,4-diacetylphloroglucinol in the rhizosphere of wheat.Phytopathol. 89:470-475.Rò
eã y÷ nÚ oÒ lñ dæ sè , Ó Eí
.Õ Só . Õ 1983. The use of lead citrate at highpH as an electron opaque stain in electronicmicroscopy. J. Cell Biol. 17: 208-212.SóaÙ nÚ k
ÛaÙ rØ aÙ sè u× bü rØ aÙ mê aÙ nÚ iÝ aÙ nÚ , Ó S
ó., Õ Ó aÙ nÚ d
æKÞ aÙ u× sè hâ iÝ kÛ , Ó Bþ .Õ Dë .Õ 2001.
Development of genetic markers in cyanobacteriaand stability of genetically marked strains in soil.World J. Microbiol. Biotechnol.17: 535-544SóaÙ rØ nÚ iÝ gå u× eã tÑ , Ó A
ï., Õ Ó KÞ rØ aÙ u× sè , Ó J
ß., Õ Ó H
Ôeã nÚ k
Ûeã lñ sè , Ó MÖ .Õ Dë .Õ ,Ó
MÖ u× eã hâ lñ cî hâ eã nÚ , Ó Aï
.Õ MÖ ., Õ Ó aÙ nÚ d æ
L� oÒ p� eã rØ , Ó Jß.Õ Eí .Õ 1995. The sigma
factor σS affects antibiotic production and biologicalcontrol activity of Pseudomonas fluorescens Pf-5.Proc. Natl. Acad. Sci. USA 92:12255-12259.Sóhâ aÙ nÚ aÙ hâ aÙ nÚ , Ó P
ð., Õ Ó O
û’� Só u× lñ lñ iÝ v� aÙ nÚ , Ó Dë .Õ Jß ., Õ Ó S
óiÝ mê p� sè oÒ nÚ , Ó P
ð.,Õ Ó
Gø
lñ eã nnÚ Ú oÒ nÚ , Ó Jß.Õ Dë .Õ , Ó aÙ nÚ d
æOû
’� Gø aÙ rØ aÙ , Ó Fö .Õ 1992. Isolation of2,4-diacethilphloroglucinol from a fluorescentpseudomonad and investigation of physiological

C� h
a pt� � e� r � 5: ��� E�
ff���
e� c� t � o� f 2,4-D����� �����
A
P! G " o� n # P$
y% t& h' i( u) m*
119
parameters influencing its production. Appl.Environ. Microbiol. 58:353-358.S+ h, a- n. a- h, a- n. , / P0 ., 12/ O
3’4S+ u5 l6 l6 i7 v8 a- n. , / D
9.1 J: ., 12/ S+ i
7m; p< s= o> n. , / P0 .,12/
G? l6 e@ nn.A. o> n. , / J:.1 D9 .1 , / a- n. d B O
3’4G? a- rC a- , / F
D.1 1993. Liquid
chromatographic assay of microbially derivedphloroglucinol antibiotics for establishing thebiosynthetic route to production, and the factorsaffecting their regulation. Anal. Chim. Acta272:271-277.S+ h, a- rC i7 fE i7 -F TG e@ h, rC a- n. i
7, / AH
.1 , / ZI
a- l6 a- , / MJ ., 12/ NK a- tL s= cM h, , / AH
.1 , / MJ o> ëNnn.A. e@ -F
LO o> cM cM o> zP , / YQ
., 12/ a- n. d B D9
éR fE a- gS o> , / G? .1 1998. Biocontrol ofsoil-borne fungal diseases by 2,4-diacetylphloroglucinol-producing pseudomonadswith different restriction profiles od amplified 16SrDNA. Europ. J. Plant Patol. 104:631-643.S+ tL rC u5 n. zP , / G? .1 MJ ., 12/ W
Ta- ll626 , / RU .1 EV ., 12/ a- n. d B H
Wo> l6 dB e@ rC -F FD rC a- n. kX l6 i7 n. ,/
MJ .1 AH .1 1978. Phloroglucinol derivatives fromAeromonas hydrophila. J. Antibiot. 31:1201-1202.S+ tL u5 tL zP , / EV ., 12/ D
9éR fE a- gS o> , / G? ., 1Y/ a- n. d B KZ e@ rC n. , / H
W.1 1986.
Naturally occuring fluorescent pseudomonadinvolved in suppression of black root rot of tobacco.Phytopathol. 76:181-185.TG a- dB a- , / MJ ., 12/ TG a- kX a- kX u5 w[ a- , / TG ., 12/ NK a- gS a- i7 , / MJ .1 , / a- n. d B Y
Qo> s= h, ii
727, / TG .1
1990. Antiviral and antimicrobial activity of 2,4-diacetylphloroglucinols, 2-acylcyclohexane-1,3-diones and 2-carboxamidocyclohexane-1,3-diones.Agric. Biol. Chem. 54:3061-3063.TG a- dB a- , / MJ ., 12/ C\ h, i
7b]
a- , / KZ ., 12/ TG a- kX a- kX u5 w[ a- , / TG .1 , / a- n. d B KZ o> j^i7m; a- ,/
EV .1 1992. Analogues of natural phloroglucinols asantagonists against both thromboxane A2 andleukotriene D4. J. Med.Chem. 35:1209-1212.TG e@ -F C\ h, a- tL o> , / S+ ., 12/ a- n. d B LO i
7m; , / MJ .1 1999. Plant
regeneration of mangosteen via nodular callusformation. Plant Cell Tissue Organ Cult. 59:89-93.TG h, i
7è_ rC y, / J
:.1 P0 .1 1967. Mise en évidence des
polysaccharides sur des coupes fines en microscopieélectronique. J. Microsc. 6: 987-1018.TG h, o> m; a- s= h, o> w[ , / LO .1 S+ ., 12/ B
ao> n. s= a- ll6Y6 , / RU .1 FD ., 12/ a- n. d B W
Te@ ll626 e@ rC , / D
9.1 MJ .1
1997. Antibiotic production by soil and rhizospheremicrobes in situ. In: Hurst, C.J., Knudsen, G.R.,McInerney, M.J., Stetzenbach, L.D., and Walter,M.V. (eds). Manual of EnvironmentalMicrobiology, pages 493-499, American Societyfor Microbiology (ASM), Washington, USA.TG o> u5 b
]i7a- -F RU a- h, m; e@ , / H
W., 12/ AH
l6 i7 -F HW a- i7 m; o> u5 dB , / D9
.1 EV ., 12/ Ba
a- rrCbC a- u5 l6 tL ,/G? ., 12/ a- n. d B A
Hl6 b] e@ rC tL i7 n. i
7, / LO .1 1995. Effect of four fungicides
on barley net blotch caused by Drecheslera teres. J.Phytopathol. 143:335-339.TG rC e@ b] s= tL , / A
H., 12/ D9
o> nn.c. e@ rC , / WT
., 12/ a- n. d B D9
rC a- b] e@ rC , / WT
. 1 1984.Structure activity correlation of herbicides affectingplastoquinone reduction by photosystem II:electron density distribution in inhibitors andplastoquinone species. Z. Naturforsch. 39c:405-411.Vd
a- l6 e@ n. tL i7 n. e@ , / RU .1 C\ .1 1961. Contrast enhancement in theelectron microscopy of viruses. In: Smith, K.M.,and Lauffer, M.A. (eds). Adv. Virus Res. 8: 287-290.Vd
a- n . dB e@ r C P0 l6 a- a- tL s= -F NK i7tL e@ rC i7 n. kX , / A
H.1 J: .1 1981. Monograph of
the genus Pythium. In: Gams, W., and Jacobs,R.P.W.M. (eds). Studies in mycology, n.21,Centraalbureau voor Schimmelcultures, Baarn, TheNetherlands.Vd
e@ rrC�C o> tL a- , / LO ., 12/ AH
pp<e< e@ n. dB i7n. o> , / G? ., 12/ B
ae@ ll626 o> rC o> , / EV ., 1Y/ J
:a- kX u5 p< o> v8 i
7cM ,/
J:., 12/ a- n. d B B
ao> m; b
]a- rC dB e@ ll626 i7 , / EV .1 1999. Furohyperforin, a
prenylated phloroglucinol from St. John’s Wort(Hypericum perforatum). J. Nat. Prod. 62:770-772.Vd
i7n. cM e@ n. tL , / MJ .1 NK ., 12/ H
Wa- rrC�C i7 s= o> n. , / LO .1 AH ., 12/ B
arC a- cM kX i
7n. , / J
:. 1 MJ .1 ,/
KZ o> v8 a- cM e@ v8 i7cM h, , / P0 .1 AH ., 1Y/ MJ u5 kX e@ rC j^ i7 , / P0 ., 12/ W
Te@ l6 l6 e@ rC , / D
9.1 MJ ., 12/ a- n. dB
P0 i7e@ rC s= o> n. , / EV .1 AH .1 1991. Genetic analysis of the
antifungal activity of a soilborne Pseudomonasaureofacens strain. Appl. Environ. Microbiol.57:2928-2934.YQ
a- j^ i7 m; a- , / TG ., 1Y/ a- n. d B MJ u5 n. a- kX a- tL a- , / KZ .1 1979.Phloroglucinol-type furocumarins, a group ofpotent naturally-occurring insect antifeedants.Agric. Biol. Chem. 43:1701-1706.YQ
o> n. e@ y` a- m; a- , / KZ ., 12/ KZ o> nn.A. a- i7 , / MJ ., 12/ HW
o> n. dB a- , / If., 12/ YQ
o> s= h, i7dB a- , / S+ .,12/
TG a- kX a- h, a- s= h, i7, / NK ., 12/ KZ o> i
7kX e@ , / H
W., 12/ a- n. d B I
fn. o> u5 e@ , / Y
Q.1 1990.
Phloroglucinol derivatives as potent photosysteminhibitors. Zeitsc. Naturfors., Section C, Biosc.45:317-321.WT
h, i7s= tL l6 e@ rC , / C\ .1 AH ., 12/ C\ o> rC b] e@ ll626 , / NK .1 AH .1 , / S+ a- rC n. i
7gS u5 e@ tL , / A
H., 1Y/ RU e@ a- m; ,/
WT
., 1Y/ a- n. d B LO o> p< e@ rC , / J:.1 EV .1 1998. The two-component
regulators gacS and gacA influence accumulation ofthe stationary-phase sigma factor σS and the stressresponse in Pseudomonas fluorescens Pf-5. J. Bact.180:6635-6641.ZI
h, o> u5 , / TG ., 12/ a- n. d B P0 a- u5 l6 i7 tL zP , / TG .1 C\ .1 1993. In vitro and invivo effects of Pseudomonas spp. on Pythiumaphanidermatum: zoospore behavior in exudates andon the rhizoplane of bacteria-treated cucumberroots. Phytopathol. 83:872-876.


______________________________________________________
CHAPTER 6
Ag
Ch
Ti
IjVk
IjTi
Y l
Om
F n
Bo
IjOm
SpUq
Rr
Fn
Ag
Ch
Ti
Ag
Ns
Ti
S p
Pt
Rr
Om
Du
Uq
Ch
Ev
Du
Bo
Y l
Pw
SxEy
Uz D{
O| M}
O| N~
A�
S x
F�
L�
Uz O| R�
Ey
SxC� Ey
N~
Sx
A�
G�
A�
I�N�
S�T �
O�
O�
M�
Y�
C�
E�
T�
E �
P�
A�
T�
H�
O�
G�
E�
N�
S�
_______________________________________________________________________________

C� h�
a� pt� � e� r � 6: ��� A�
c� t� i� v� it� � y � o� f b �¡
i� o� s¢ ur£ � f a� c� t� a� n¤ t� s¢
122
A�
B¥ S¦ T§
RA�
C� T§
_______________________________________________________________________________
B¥ i� o� s¢ ur£©� f a� c� t� a� n¤ t� s ¢ a� r� e � a� mpª � h�
i� p� a� t� h� i� c � c� o� mª p� o� u£ n¤ d«
s ¢ p� r� o� d«
u£ c� e� d «
b¡
y � miª � c� r� oo�¬� r� g a� n¤ i� s¢ mª s ¢ t� h� a� t � r� e� d« u£ c� e�s¢ ur£ � f a� c� e � a� n¤ d
«i� n¤ t� e� r� f a� c� i� a� l t® � e� n¤ s¢ i� o� n¤ s¢ . ¯ I° n ¤ t� h� i� s s¢±¢ t� u£ d
«y� , w²´³ e � d
«e� mª o� n¤ s¢ t� r� a� t� e� d
«t� h� a� t � b
¡i� o� s¢ ur£©� f a� c� t� a� n¤ t� -µ p� r� o� d
«u£ c� i� n¤ g
f l®u£ o� r� e� s¢ c� e� n¤ t � Ps
¶ ·eu¹»º d¼
o½ m¾ o½ n¿ aÀ s ·
s¢ p� e� c� i� e� s ¢ o� cc�Á� ur £ � i� n ¤ t� h� e � w³ h�
e� a� t � r� h� i� z o� s¢ p� h�
e� r� e � a� t � d«
e� n¤ s¢ it� � i� e� s ¢ o� f
a� pp� � r� o� xà im� ª a� t� e� l®y�
10ÄcÅ 5Æ C� FÇ U gÈ -1É Ê . ¯ W
Ëe � r� e� p� o� r� t � t� h� e � d
«i� s¢ c� o� v� e� r� y � o� f
s¢ i� x à Ps
¶ ·eu¹»º d¼
o½ m¾ o½ n¿ aÀ s· i� s¢ o� l
®a� t� e� s ¢ t� h� a� t p�Ì� r� o� d
«u£ c� e � b
¡i� o� s¢ ur£ � f a� c� t� a� n¤ t� s¢
w³ it� � h �
l®y� ti� � c � a� c� ti� � v� i� t� y � a� g a� i� n¤ s¢ t z�  oo�Í� s¢ p� o� r� e� s ¢ o� f
mª u£ lti
® � � p� l®e � o� o� mª y� c� e� t� e � p� a� t� h� og�Í e� n¤ s¢ . ¯ Z
Îoo�¬� s¢ p� o� r� e� s ¢ e� xà p� o� s¢ e� d
«t� o � c� e� ll®Ï®
s¢ u£ s¢ p� e� n¤ s¢ i� o� n¤ s ¢ o� r � c� e� ll®Ï®
-fµ r� e� e � c� u£ lt® � ur£©� e � s¢ u£ p� e� r� n¤ a� t� a� n¤ t � o� f
t� h� e� s¢ e � i� s¢ o� l
®a� t� e� s ¢ l
®o� s¢ t � t� h� e� i� r � mª o� t� ili� ® � t� y � w³ i� t� h� i� n ¤ 30 Ð�Å s¢ e� c�
a� n¤ d «
e� xà pl� ® o� d«
e� d «
w³ i� t� h� i� n ¤ 60 ��Å s¢ e� c. � ¯ OÑ n¤ e o�Ò� f
t� h� e� s¢ e � i� s¢ o� l®a� t� e� s¢ , ² P
¶. Ó fÔlÕ uº o½ rÖ e¹ s· c× e¹ n¿ s
· SS¦¬¦ 101, ÄcÅ�Äc² r� e� d
«u£ c� e� d
«t� h� e � s¢ ur£©� f a� c� e�
t� e� n¤ s¢ i� o� n ¤ o� f w ³ a� t� e� r � f
r� o� m ª 73
Ø Ð mª N Ù
mª -1É Ê t� o � a� pp�©� r� o� xà i� mª a� t� e� l® y � 30 Ð�Å mª N Ù
mª -1É Ê . ¯ A�
pp� � l®i� c� a� ti� � o� n ¤ o� f
c� e� ll ®Ï® s¢ u£ s¢ p� e� n¤ s¢ i� o� n¤ s¢
o� f
s¢ t� r� a� i� n ¤ SS¦¬¦ 101 ÄcÅ�Ä t� o � s¢ o� il � ® o� r � h�
y� a� c� i� n¤ t� h �
b¡
u£ l®b¡
s ¢ p� r� o� v� i� d« e� d «
p� r� o� t� e� c� ti� � o� n ¤ a� g a� i� n¤ s¢ t � r� o� o� t � r� o� t � c� a� u£ s¢ e� d «
b¡
y�P¶
yÚ tÛ hÜ iÝ uº m ¾ iÝ n¿ tÛ e¹ rÖ m¾ e¹ d¼ iÝ uº m¾ . ¯ FÇ i� v� e � mª u£ t� a� n¤ t� s ¢ o� f
S¦ S¦ 101, ÄcÅ�Äc² o� b¡
t� a� i� n¤ e� d «
b¡
y � r� a� n¤ d«
o� m ª T§
n¤ 5Þ
mª u£ t� a� g e� n¤ e� s¢ i� s¢ , ² l®a� c� kß
e� d«t� h� e � a� b¡ i� l® it� � y � t� o � r� e� d« u£ c� e � t� h� e � s¢ ur£ � f a� c� e � t� e� n¤ s¢ i� o� n ¤ o� f w
³ a� t� e� r � a� n¤ d «
t� o � c� a� u£ s¢ e � l®y� s¢ i� s ¢ o� f
z oo�¬� s¢ p� o� r� e� s¢ . ¯ S¦ u£ b
¡s¢ e� qà u£ e� n¤ t�
geá� n¤ e� ti� � c � c� h� a� r� a� c� t� e� r� iz�  a� ti� � o� n ¤ o� f
t� h� e � T§
n¤ 5Þ
-fµ l® a� n¤ kßi� n¤ g r� eg�Á i� o� n¤ s ¢ o� f
t� w³ o � s¢ ur£©� f a� c� t� a� n¤ t� -µ d« e� f i� c� i� e� n¤ t m� ª u£ t� a� n¤ t� s¢ ,²
10.24 ÄcÅ�¯ â�ã a� n¤ d «
17.18, Ä Ø ¯äÄcå�² s¢ h� o� w³ e� d «
t� h� a� t � t� h� e � t� r� a� n¤ s¢ p� o� s¢ o� n¤ s ¢ h�
a� d «
i� n¤ t� eg�Á r� a� t� e� d «
i� n ¤ c� o� n¤ d«
e� n¤ s¢ a� ti� � o� n ¤ d«
o� mª a� i� n¤ s ¢ o� f
p� e� p� t� i� d« e � s¢ y� n¤ t� h� e� t� a� s¢ e� s¢ . W¯ Ë h�
e� n ¤ a� pp� � l®i� e� d «
t� o � s¢ o� il� ® , ² mª u£ t� a� n¤ t � 10.24 wÄcÅ�¯ â�ãæ³ a� s ¢ n¤ o� t � a� b¡ l®e � t� o � p� r� o� t� e� c� t � h
�y� a� c� i� n¤ t� h�
a� g a� i� n¤ s¢ t � Pç y� t� h� i� u£ m ª r� o� o� t � r� o� t� , ² w³ h�
e� r� e� a� s ¢ w³ il� ® d« t� y� p� e � s¢ t� r� a� i� n ¤ SS¦¬¦ 101 ÄcÅ�Ä g a� v� e � s¢ i� g n¤ i� f i� c� a� n¤ t � c� o� n¤ t� r� o� l®. T¯ § h
�e � p� a� r� ti� � a� ll
®Ï®y�
p� ur£ � i� f i� e� d «
s¢ ur£©� f a� c� t� a� n¤ t� s ¢ f r� o� m ª P
¶. Ó fÔlÕ uº o½ rÖ e¹ s· c× e¹ n¿ s
· SS¦¬¦ 101 ÄcÅ�Ä l
®o� w³ e� r� e� d
«t� h� e � s¢ ur£ � f a� c� e � t� e� n¤ s¢ i� o� n ¤ o� f w
³ a� t� e� r � f r� o� m ª 73
Ø Ð t� o�a� pp� � r� o� xà im� ª a� t� e� l
®y � 30 Ð�Å mª N
Ùmª -1É Ê a� n¤ d
«r� e� a� c� h� e� d
«t� h� e � c� r� i� t� i� c� a� l mi
® ª � c� e� l® l® e � c� o� n¤ c� e� n¤ t� r� a� ti� � o� n ¤ a� t � 25-50 â�è µ è�Å µé g mlª ® -1É Ê .Re� v� e� r� s¢ e� -µ p� h
�a� s¢ e � h
�i� g h �
p� e� r� f o� r� mª a� n¤ c� e � l®i� qà u£ i� d
«c� h� r� o� mª a� t� og�¬ r� a� p� h
�y � (ê RPç -µ Hë Pç Lì C� )
ía� n¤ d «
l®iq� à u£ i� d«
c� h� r� o� mª a� t� o� g r� a� p� h�
y� –î mª a� ss s¢ä¢ï¢ p� e� c� t� r� o� mª e� t� r� y � (ê Lì C� -µ Mð S¦ ) í
y� i� e� l® d« e� d «
e� i� g h�
t � d«
i� f f e� r� e� n¤ t � f r� a� c� ti� � o� n¤ s ¢ o� f w
³ h�
i� c� h �
f i� v� e�
h�
a� d «
s¢ ur£ � f a� c� e � a� c� ti� � v� it� � y � a� n¤ d «
c� a� u£ s¢ e� d «
c� e� ss¢ä¢ a� ti� � o� n ¤ o� f
mª o� t� ili� ® � t� y � a� n¤ d «
l®y� s¢ i� s ¢ o� f
z o� o� s¢ p� o� r� e� s¢ . ¯ C� o� ll
®Ï®e� c� ti� � v� e� l® y� , ² t� h� e�
r� e� s¢ u£ lt® � s ¢ i� n¤ d
«i� c� a� t� e � t� h� a� t � b
¡i� o� s¢ ur£ � f a� c� t� a� n¤ t� s ¢ p� r� o� d
«u£ c� e� d
«b¡
y � P¶
. Ó fÔlÕ uº o½ rÖ e¹ s· c× e¹ n¿ s
· SS¦¬¦ 101 ÄcÅ�Ä pl� ® a� y � a� n ¤ i� mpª � o� r� t� a� n¤ t � r� o� l
®e � i� n¤
i� n¤ t� e� r� a� c� t� i� o� n¤ s ¢ w³ i� t� h �
z oo�Í� s¢ p� o� r� i� c � p� a� t� h� o� g e� n¤ s ¢ a� n¤ d «
i� n ¤ c� o� n¤ t� r� o� l ®
o� f
Pç y� t� h� i� u£ m ª r� oo�¬� t � r� o� t � o� f
h�
y� a� c� i� n¤ t� h� ._____________________________________________________________________
Jño� r� ge á� T.
§ ¯ d«
e � S¦ o� u£ z a� , ² Mð
a� r� jò a� n ¤ d«
e � B¥ oe�¬� r� , ² Pç i� e� t� e� r � d«
e � WË
aa�Á� r� d« , T² § e� r� i� s A¢ � . ¯ v� a� n B¤ó¥ e� e� kß , ² a� n¤ d «
Jño� s ¢ M
ð.
Raa�Á� i� jò mª a� kß e� r� s¢
To be submitted.

Cô hõ
aö pt÷ ø eù r ú 6: û�ü Aý
cþ tø iÿ v� itÿ ø y � o� f b���
iÿ o� s� ur� ú f� aö cþ tø aö n� tø s�
123
I N T�
RO�� D� U�
Cô T�
I O N
Microbial compounds that alter the conditions prevailing at a surface or interface are
often referred to as adjuvants, bioemulsifiers or biosurfactants (Cooper and Zajic, 1980; Zajic
and Seffens, 1984; Fiechter, 1992). A variety of microorganisms, including bacteria, fungi and
yeasts, have been described to produce biosurfactants. Several of these biosurfactants are well-
described chemically and catagorized into high- and low-molecular mass compounds. The
low-molecular biosurfactants include glycolipids and lipopeptides, such as rhamnolipids and
surfactin. The high-molecular compounds include proteins, lipoproteins or complex mixtures
of these polymers (Ron and Rosenberg, 2001). Although biosurfactants are structurally
diverse, they all have an amphipathic nature, i.e. they contain both hydrophobic and
hydrophylic groups. The hydrophobic moieties are usually saturated, unsaturated or
hydroxylated fatty acids, and the hydrophylic moieties consist of mono-, di-, or
polysaccharides, carboxylic acids, amino acids or peptides (Ron and Rosenberg, 2001).
Among the bacterial genera, Pseudomonas spp. have been reported to produce
biosurfactants (Fiechter, 1992; Ron and Rosenberg, 2001). Pseudomonas spp. are common
inhabitants of soil and rhizosphere environments and have received considerable attention in
the areas of bioremediation of xenobiotics and biological control of plant pathogenic fungi. In
the area of bioremediation, surfactant-producing Pseudomonas spp. have been implicated in
facilitating degradation of ubiquitous pollutants such as polycyclic aromatic hydrocarbons
(PAHs) and n-alkanes (Arino et al., 1996; Déziel et al., 1996). In the area of biological control
of plant pathogenic fungi, the potential of biosurfactants produced by Pseudomonas spp. was
only recently described. Rhamnolipids produced by strains of P. aeruginosa were shown to be
highly effective against plant pathogens, including Pythium aphanidermatum, Plasmopara
lactucae-radicis and Phytophthora capsici (Stanghellini and Miller, 1997). Purified rhamnolipids
caused cessation of motility and lysis of entire zoospore populations within less than 1 min.
Introduction of a rhamnolipid-producing strain in a recirculating hydroponic system gave
good control, although transient, of P. capsici on pepper (Stanghellini and Miller, 1997). Kim
et al. (2000) confirmed and extended these observations by showing that rhamnolipid B
produced by P. aeruginosa B5 not only has lytic effects on zoospores, but also inhibitory
activity against spore germination and hyphal growth of several other pathogens. Mycelial
growth of P. capsici and spore germination of Colletotrichum orbiculare were inhibited in vitro
and the diseases caused by these pathogens were suppressed in pepper and cucumber plants,
respectively, by application of purified rhamnolipid B to leaves (Kim et al., 2000).

C� h�
a� pt��� e� r � 6: ��� A�
c� t� i� v� it� � y o! f b"�#
i� o! s$ ur%�� f" a� c� t� a� n& t� s$
124
Several lipopeptide surfactants with antibiotic properties were recently proposed as
biological compounds for the control of plant pathogenic fungi (Nielsen et al., 1999).
Viscosinamide and tensin, produced by soil-inhabiting P. fluorescens strains, were shown to
affect mycelium of Rhizoctonia solani and Pythium ultimum, causing reduced growth and
intracellular activity, hyphal swellings, increased branching and rosette formation (Thrane et
al., 1999; Nielsen et al., 2000). These studies clearly indicated the potential of biosurfactant-
producing Pseudomonas spp. for crop protection.
To date, however, little is known about the frequencies of biosurfactant-producing
Pseudomonas spp. in natural environments and more specifically in the plant rhizosphere. In
this study, we determined the frequency of biosurfactant- producing Pseudomonas spp. in the
rhizosphere of wheat. We report the discovery of P. fluorescens strains that produce surfactants
with lytic activity against zoospores of oomycetes, including Pythium species and P. infestans.
The role of biosurfactants in suppression of Pythium root rot of hyacinth by representative P.
fluorescens strain SS101 was determined by generating mutants defective in the production of
biosurfactants. Mutants were genetically and phenotypically characterized and their activity
against Pythium spp. was compared to that of their parental strain. The composition and
activity of the surface-active extract was determined by reverse phase high-performance liquid
chromatography (RP-HPLC) and bioassays, respectively.
R' E(
S) U*
L+ T,
S)
F- r� e� q. u% e� n& c� i� e� s $ o! f "
s$ ur%�� f" a� c� t� a� n& t� -/ p� r� o! d0
u% c� i� n& g 1 Ps243
e5 u6 d7o8 m9 o8 n: a; s
3 s$ pp��� .< Population studies were performed for
seven successive cycles of wheat grown in three soils. Only results from three growth cycles are
shown (Table 1). The population densities of surfactant-producing Pseudomonas spp. was
determined by randomly transferring approximately 10% of the fluorescent Pseudomonas
colonies, isolated from the wheat rhizosphere on KMB+ medium, to SW medium. Bacterial
colonies showing a halo on SW medium were considered as putative surfactant producers.
Population densities of putative surfactant-producing Peudomonas spp. ranged from 1.0 X 105
to 7.0 X 105 CFU g-1 root and represented approximately 1 to 5% of the total population of
fluorescent Pseudomonas isolated from roots of wheat. Cycles 4 to 7 and cycles 1 to 7 of the
repeated experiment showed similar results.

C= h>
a? pt@�A eB r C 6: D�E AF
cG tA iH vI itH A y J oK f bL�M
iH oK sN urO�C fL a? cG tA a? nP tA sN
125
TQ
aR bS lTe U 1. VXW P
YoZ p[ u\ l
TaR t] i^ oZ n _ d
`eU n_ sa i^ t] i^ eU s a oZ f
bsa u\ rc fb aR cd t] aR n_ t] -e p[ rc oZ d
`u\ cd i^ n_ g f P
gsh ei uj dk ol mm ol nn ao sh sa pp[p[ . W rc eU cd oZ vq eU rc eU d
`fbrc oZ m r rc oZ oZ t] s a oZ f
bws ht
eU aR t]gf rc oZ ws n _ i
^n n_u_ aR t] u\ rc aR l T sa oZ i
^lTs a fboZ r c t] ht rc ee UvU sa u\ ccdwd eU sa sa i^ vq e U cd yx cd lT eU s a oZ f f
byboZ u\ r c ws eU eU kz s a eU aR cd ht .W
First cycle a Second cycle a Third cycle a
Soil Fluoresc.b
Pseudom.Surfact. c
Produc.Fluoresc.b
Pseudom.Surfact. c
Produc.Fluoresc.b
Pseudom.Surfact. c
Produc.
CB 1.7 X 107 1.7 X 105 8.5 X 106 2.6 X 105 5.3 X 106 1.1 X 105
SV 1.9 X 107 3.8 X 105 5.3 X 106 2.1 X 105 1.5 X 107 1.5 X 105
SSB 1.4 X 107 7.0 X 105 5.6 X 106 1.1 X 105 5.2 X 106 1.0 X 105
a Wheat seedlings were grown in CB, SV and SSB soils for three successive cycles of 4 weeks each.b Total population densities of fluorescent Pseudomonas spp. (CFU g-1 root) were determined by plating
rhizosphere suspensions on KMB+ medium. Mean values of four replicates are given.c Populations of surfactant-producing Pseudomonas spp. (CFU g-1 root) were determined by randomly transferring
approximately 10% of the colonies recovered on KMB+ to SW medium. Colonies forming a halo after 3 to 5days of growth at 25 °C were considered surfactant producers. Mean values of four replicates are given.
S{ eB l| eB cG tA iH oK n P a? nP d }
cG h> a? rC a? cG tA eB rC izH�~ a? tA iH oK n P oK f L
sN tA rC a? iH n P SS{y{ 101. ������� From a total of 375 Pseudomonas isolates that
produced a halo on SW medium, 6 isolates were able to lyse zoospores of Pythium ultimum
var. sporangiiferum (Fig. 1). All 6 isolates were obtained from roots of wheat grown in SSB soil.
A cell suspension (109 CFU ml-1) of these isolates caused cessation of zoospore motility within
30 sec and the shape of the zoospores was changed from bean-shaped to round (Fig. 1A). The
bacterial cell suspension caused lysis of the entire zoospore populations within 60 sec (Fig.
1B). Identical responses were observed for zoospores from Phytophthora infestans, Pythium
intermedium, and Albugo candida. From the six isolates that caused lysis of zoospores, isolate
SS101 was selected for further studies. Biochemical characterization by API 20 NE test and
GC-FAME analysis classified isolate SS101 as P. fluorescens Biovar II. Strain SS101 exhibited
multiple characteristics indicative of surfactant production, including halo formation on SW
medium, foam formation, drop-collapse, and the ability to lower the surface tension (Table 2).
The other Pseudomonas strains, including reference strains WCS358, PAO1, and DSM1128,
exhibited similar characteristics, however, none of these isolates caused lysis of zoospores
(Table 2). Only isolate SV7 caused cessation of zoospore motility, but no subsequent lysis.
AF
cG tiA H vI itH A y J oK f L
P�
. � f�l� uo��� r� e� s� c� e� n� s� SS{y{ 101 ����� a? g� a? iH nP sN t A P
�. � i� n� t� e� r� m� e� d� i� u� m� . � The activity of P. fluorescens SS101
against P. intermedium, causal agent of Pythium root rot of hyacinth, was tested in a bioassay
at 9°C for 8 weeks. P. intermedium P52 significantly decreased the root fresh weight of
hyacinth by at least 4-fold (Fig. 2). Treatment of hyacinth bulbs or soil with P. fluorescens
SS101 provided substantial protection of roots of hyacinth grown in soil treated with P.
intermedium P52 (Fig. 2A). In a second experiment, treatment of both bulbs and soil resulted

C� h�
a� pt��� e� r � 6: ��� A�
c� t� i v¡ it � y ¢ o£ f b¤�¥
i o£ s¦ ur§�� f¤ a� c� t� a� nt� s¦
126
F©
iªg« u¬ r e ® 1. ¯X° E
±ff²³²
e® c´ t µ o¶ f ²
P·
. ¸ f¹lºu» o¼ r½ e¾ s¿ cÀ e¾ nÁ s¿ SÂ S
Â101 ¯ÄÃů o¶ n Æ zÇ o¶ o¶ sÈ pÉ o¶ r e® s È o¶ f
²P·
. ¸ ul» º tÊ iË mÌ u» mÌ vÍ aÎ r . ° s¿ pÏ o¼ r½ aÐ nÁ gÑ iËiËf¹e¾ r½ u» mÌ .° A: within 30 sec
after exposure to strain SS101, zoospores become immotile and round-shaped. B: within 60sec, lysis of zoospores occurs; C: within an additional 10-30 sec, the remnants of thezoospores completely disappear. Arrows indicate a zoospore of P. ultimum var.sporangiiferum and arrowheads indicate cells of strain SS101 (magnification 400X). Thedensity of the bacterial cell suspension was 109 CFU ml-1 and zoospore suspensionscontained 104 zoospores ml-1.
in complete protection of hyacinth roots against P. intermedium P52 (Fig. 2B).
TÒ
n5Ó
mÔ u§ t� a� gÕ e� ne� s¦ i s ¦ a� nd Ö
c� h� a� r� a� c� t� e� r� iz × a� ti� o£ n ¨ o£ f ¤
s¦ ur§�� f¤ a� c� t� a� nt� -Ø dÖ e� f¤ i c� i e� nt m� Ô u§ t� a� nt� s¦ .Ù A total of 520
transformants of P. fluorescens SS101 were obtained following mutagenesis with Tn5lacZKm.
Five of these transformants were not able to form a halo on SW medium (Table 3).
Characterization of these mutants with RAPD analysis using three 10-mer primers confirmed
strain integrity. Southern blot analysis with a probe targeting the kanamycin resistance gene
showed the presence of a single Tn5-integration for each of the five mutants. These mutants
gave a negative reaction in the drop-collapse test, were unable to form foam after vigorously
shaking, did not lower the surface tension of water to the same extent as wild-type strain
SS101, and could no longer affect the motility and cause lysis of zoospores of multiple
oomycetes (Table 3). In contrast to the wild-type, all mutants presented β-galactosidade
activity, indicating integration of the promotorless Tn5lacZKm element in a coding region. All
mutants still produced wild-type levels of protease, phospholipase C, and siderophores,
suggesting that a global regulatory gene was not inactivated.
RÚ o£ lÛe o�Ü£ f
¤s¦ ur§�� f¤ a� c� t� a� nt� s ¦ i n ¨ a� c� ti� v¡ i t� y ¢ a� gÕ a� i ns¦ t � P
Ý. Þ iß nà tá eâ rã mä eâ då iß uæ mä . To determine the possible role of
surfactant production in the activity of strain SS101 against P. intermedium, bioassays with
mutant 10.24 were performed. Application of wild-type strain SS101 restored the root weight
of hyacinth to levels not significantly different from the untreated control (Fig. 3A). In
CBA

Cç hè
aé ptê�ë eì r í 6: î�ï Að
cñ të iò vó itò ë y ô oõ f bö�÷
iò oõ sø urù�í fö aé cñ të aé nú të sø
127
Tû
aü bý lþe ÿ 2. ���
P�
h�
eÿ n� o� t� y� p ic c��� h
�aü r aü c� t� eÿ r i s� t� i c� s � o� f
�P�
. � f�lu���
o� r� e� s� c� e� n� s� s� t� r aü i n � SS���
10� � 1 � aü n� d !
o� t� h� eÿ r P�
s� e� u� d"o� m# o� n� a$ s� s� t� r aü i n� s� .�
Strain Halo a
formationDrop b
collapseFoam c Surface d
tension(mN m-1)
Zoospore e
motilityZoospore e
lysis
Water - * - - 73.5 - -
P. fluorescens SSB17 - - - 70.5 - -P. fluorescens SS101 +
* + + 29.7 + +
P. fluorescens SV7 + + + 54.0 + -P. putida WCS358 + + + 30.5 - -P. aeruginosa PAO1 + + + 56.5 - -P. aeruginosa DSM1128 + + + 49.5 - -
* + positive reaction; - negative reaction.a Evaluated on SW medium after 3 to 5 days of growth at 25°C.b Cell suspensions (OD600=1)were tested on an oily surface for their ability to collapse a drop of water.c Foam formation after vigorous shaking cell suspensions (OD600=1).d Surface tension of cell suspensions (OD600=1)was measured with a tensiometer.e Zoospore motility and lysis were observed microscopically after mixing bacterial cell suspensions (OD600=1) with
zoospore suspensions of P. ultimum var. sporangiiferum (104 zoospores ml-1).
contrast, the surfactant-deficient mutant 10.24 was not effective against Pythium root rot of
hyacinth (Fig. 3A). The disease level was significantly reduced in the treatment with strain
SS101, whereas mutant 10.24 failed to control Pythium root rot (Fig. 3B). These results
indicate that surfactant production by strain SS101 plays an important role in control of
Pythium root rot of hyacinth.
I% d& eì nú të iò fö iò cñ aé të iò oõ n ú oõ f ö
g' eì nú eì s ø iò nú vó oõ l(vó eì d &
iò n ú sø urù�í fö aé cñ të aé nú t ë b÷
iò oõ sø yô nú të hè eì sø iò sø .) The flanking regions of Tn5
transposon were cloned and sequenced for surfactant-deficient mutants 10.24 and 17.18 by
anchored PCR (aPCR). Sequences of mutant 10.24 yielded a 852-bp fragment with a
deduced protein sequence that has similarity with the condensation domain of peptide
synthase genes from several bacteria and eukaryotes (Fig. 4). The deduced protein sequence
showed 54% (161/293) similarity with a condensation domain in the syringopeptin synthetase
gene sypA from P. syringae pv. syringae B310D and varying degrees of similarity to five
condensation domains of the syringomycin synthetase syrE gene of P. syringae pv. syringae
B310D.
The syrE gene encodes a 9376-aa protein and the similarities of its different
condensation domains to the condensation domain of P. fluorecens SS101 were 48% (109/226)
for positions 65 to 283, 38% (112/288) for positions 4270 to 4557, 36% (107/293) for
positions 5332 to 5623, 35% (106/295) for positions 6388 to 6681, and 39% (117/295) for
positions 8548 to 8841. Similarities were 44% (121/271) for positions 11 to 279 and 32%
(71/210) for positions 1053 to 1258 of a 1278-aa protein involved in the synthesis of the
siderophore 2,3-dihydroxybenzoate (DHB) by B. subtilis, and 41% (121/281) for positions

C* h+
a, pt-/. e0 r 1 6: 243 A5
c6 t. i7 v8 it7 . y 9 o: f b;=<
i7 o: s> ur?/1 f; a, c6 t. a, n@ t. s>
128
FA
iBgC uD rE e F 2.
G�HEI
ffJKJ
eF cL t M oN f J
PO
. P fQlRuS oT rU eV sW cX eV nY sW SZ S
Z101 []\�[ oN n ^ P
_y` tM ha i
BuD m b rE ooN�N t M rE oN t M oN f
Jha
y` ac cL iB n^ tM ha .H Inoculum of P. intermedium
P52 was mixed through soil (Pythium +). Soil not treated with P. intermedium (Pythium -)served as a healthy control. P. fluorescens SS101 was applied to soil [SS101(soil)], bulbs[SS101(bulb)], or both [SS101(s + b)]. Cell suspensions for bulb treatment were 107 CFUml-1 and the initial density of SS101 in soil treatments was 107 CFU g-1 soil fresh weight.Plants were grown at 9°C in the dark. After 8 weeks, root weight was determined inexperiment I and disease index was scored in experiment II. Disease index was scored on a0 to 5 scale, where 0 indicates no disease and 5 indicates 80 to 100% loss of root biomassrelative to the control. Means of five replicates are shown. Means with the same letter arenot statistically different according to Tukey’s studentized range test (P = 0.05). Error barsrepresent the standard error of the mean. The experiment was repeated 4 times andrepresentative results of two experiments are shown.
1990 to 2270 and 45% (83/189) for positions 3145 to 3322 of a 3487-aa protein encoded by
the gene mcyE, which is responsible for the synthesis of the cyclic heptapeptide microcystin in
the bacterium Microcystis aeruginosa K-139. A majority sequence obtained from the
alignement of 43 different condensation domains, including proteins involved in the synthesis
of gramicidin S by Brevibacillus brevis, surfactin by Bacillus subtilis and peptide synthetases
from several fungi, including Cochliobolus carbonum, Penicillium chrysogenum, Emericella
nidulans, and Acremonium chrysogenum, showed that the active-site motif (HHXXXDG) is
conserved in all aligned condensation domains, including the partial sequence obtained from
strain SS101 (Fig. 6).
Sequence analysis of the Tn5 flanking region of mutant 17.18 yielded a 453-bp
fragment with a deduced protein sequence that presents 61% (90/145) similarity with a 3310-
aa protein from Bradyrhizobium japonicum 110spc4, 66% (96/144) similarity for positions
415 to 558 and 61% (90/146) similarity for positions 2550 to 2695 of a 5953-aa peptide
synthase protein of the bacterial pathogen Ralstonia solanacearum GMI1000, 63% (92/144)
similarity with the 3316-aa protein XpsB from Xenorhabdus bovienii T228, and similarity with
Pyt
hiu
m +
S
S10
1 (b
ulb
)
Ro
ot
wei
gh
t (g
)
0
20
40
60
80
100a
b b
c
d
I
Pyt
hiu
m +
S
S10
1 (s
oil)
Pyt
hiu
m -
Pyt
hiu
m +
.
Dis
ease
ind
ex
0
1
2
3
4
5
6
a
bb
II
Pyt
hiu
m +
SS
101
(s+b
)
Pyt
hiu
m -
Pyt
hiu
m +

Cd he
af ptg/h ei r j 6: k4l Am
cn th io vp ito h y q or f bs=t
io or su urv/j fs af cn th af nw th su
129
Tx
ay bz l{e | 3. }�~
C�
h�
ay r� ay c� t� e| r� i� s� t� i� c� s � o� f �
s� u� r� f� ay c� t� ay n� t� -� d� e| f� i� c� i� e| n� t � m� u� t� ay n� t� s � o� f �
P�
. � f�l�u� o� r� e� s� c� e� n� s� s� t� r� ay i
�n � SS���
1� 01.� � ~
MutantsTest SS101 9.26 10.24 11.17 13.3 17.18
Halo formation a+
*-
* - - - -
Drop-collapse b + - - - - -Foam formation c + - - - - -Surface tension (mN m-1) d 29.7 64.3 56.0 60.0 56.5 62.0β-galactosidase activity e - + + + + +Protease f + + + + + +Phospholipase C g + + + + + +Fluorescence h + + + + + +Zoospore motility i + - - - - -Zoospore lysis i + - - - - -
* + positive reaction; - negative reaction.a Evaluated on SW medium after 3 to 5 days of growth at 25°C.b Cell suspensions (OD600=1) were tested on an oily surface for their ability to cause the collapse of a drop of
water.c Cell suspensions (OD600=1) were observed for foam formation after vigorous shaking.d Surface tension of cell suspensions (OD600=1) was measured with a tensiometer.e Evaluated on KMB plates containing X-gal and IPTG after 48h of growth at 25 °C.f Tested on skim milk plates.g Tested on egg yolk plates.h Tested on Pseudomonas agar F plates under near UV light.i Zoospore motility and lysis were observed microscopically after mixing bacterial cell suspensions with zoospore
suspensions of P. ultimum var. sporangiiferum, A. candida, or P. infestans.
four domains of the 9376-aa syringomycin synthetase protein from P. syringae pv. syringae
B310D (Fig. 7). Similarity levels for these domains were 58% (86/146) for positions 1337 to
1478, 57% (84/144) for positions 2425 to 2563, 57% (84/144) for positions 3512 to 3650,
and 56% (85/148) for positions 7794 to 7941 of syringomycin synthetase. A database search
and comparison of these sequences showed that the transposon is integrated in a condensation
domain of peptide synthetases. However, the flanking sequence of mutant 17.18 was not long
enough to identify conserved active-site motifs of condensation domains. Collectively, these
results show that in both mutants 10.24 and 17.18 the transposon is integrated in
condensation domains of putative peptide synthetases.
I� su or l�af tih o or n w af nw d
�cn he
af rj af cn th ei rj izo�� af th io or n w or f s
su urv j fs af cn th af nw th s u pg rj or d�
uv cn ei d �
bt
y q P¡
. ¢ f£l¤ uo¥4¦ r§ es© cª en« s© SS¬¬ 101. ®°¯±®°² Extraction of
the surfactant(s) produced by P. fluorescens SS101 yielded a white precipitate, whereas no such
precipitate was obtained from the surfactant-deficient mutant 10.24. After extraction, the
composition was investigated by RP-HPLC with both UV and evaporative light scattering
(ELS) detection. With an acidified water – acetonitrile gradient, eight fractions were observed
and collected (Table 4). Fraction 8 was the main component and accounted for at least 95%
of the extract. There was a good correspondence between UV detection at 210 nm and ELS

C³ h´
aµ pt¶/· e¸ r ¹ 6: º4» A¼
c½ t· i¾ v¿ it¾ · y À oÁ f bÂ=Ã
i¾ oÁ sÄ urÅ/¹ f aµ c½ t· aµ nÆ t· sÄ
130
FÇ
iÈgÉ uÊ rË e Ì 3.
Í�ÎEÏ
ffÐKÐ
eÌ cÑ t Ò oÓ f Ð
PÔ
. Õ fÖl×uØ oÙ rÚ eÛ sÜ cÝ eÛ nÞ sÜ Sß S
ß101 à]á�à aâ nã d
äiÈtÒ s såæå uÊ rË fÐ aâ cÑ tÒ aâ nã t Ò d
äeÌ fÐ iÈ cÑ iÈ eÌ nã t Ò mç uÊ tÒ aâ nã t Ò 10.24 à á Î èêé oÓ n ã P
ëyì tÒ hí i
ÈuÊ m ç rË ooÓ�Ó t Ò rË oÓ tÒ
oÓ f Ð
hí
yì aâ cÑ iÈnã tÒ hí .
Î Inoculum of P. intermedium P52 was mixed through soil (Pythium +). Soil not
treated with P. intermedium (Pythium -) served as a healthy control. P. fluorescens SS101and surfactant-deficient mutant 10.24 were applied to the soil at densities of 107 CFU g-1
soil fresh weight. Plants were grown at 9°C in the dark. After 8 weeks, root weight (A) anddisease index (B) were determined. Disease caused by P. intermedium P52 was scored on a0 to 5 scale, where 0 indicates no disease and 5 indicates 80 to 100% loss of root biomassrelative to the control. Means of five replicates are shown. Means with the same letter arenot statistically different according to Tukey’s studentized range test (P = 0.05). Error barsrepresent the standard error of the mean. The experiment was repeated 2 times andrepresentative results are shown.
detection. All fractions, with exception of fraction 1, were positive in the drop-collapse test,
indicating a decrease in the surface tension. Only fractions 5 and 8 were positive in the foam
formation test. Fractions 4 to 8 caused cessation of zoospore motility and subsequent lysis.
Fraction 8, which was the most abundant fraction, caused lysis of zoospores in a much shorter
time frame than the other fractions. Zoospores were rendered immotile within 10 sec and
lysed within 30 sec after contact with fraction 8, whereas for the other fractions it took
approximately 30 sec for cessation of motility and approximately 60 sec for lysis. The lysis
caused by fraction 7 was not a characteristic explosion observed with the other fractions, but
resembled leaking at one point in the zoospore membrane.
The critical micelle concentration (CMC value) is defined as the solubility of a
surfactant in an aqueous phase. At concentrations above the CMC, surfactant molecules
aggregate to form polymers called mycelles which solubilize oil or water into the other phase,
creating a microemulsion. No further drop of the surface tension occurs as the amount of
surfactant exceeds the CMC value because the emulsion is already saturated (Georgiou et al.,
1992). The CMC value is one of the measures of the strength of the surface activity of a
Ro
ot
wei
gh
t (g
)
0
10
20
30
40
50
60
70
a a
bb
.
A
Pyt
hiu
m -
Pyt
hiu
m +
Pyt
hiu
m +
SS
101
Pyt
hiu
m +
10.2
4
Dis
ease
ind
ex
0
1
2
3
4
5
.
a
b
a
c
B
Pyt
hiu
m +
Pyt
hiu
m +
10.2
4
Pyt
hiu
m +
SS
101
Pyt
hiu
m -

Cî hï
að ptñ/ò eó r ô 6: õ4ö A÷
cø tò iù vú itù ò y û oü f bý=þ
iù oü sÿ ur�/ô fý að cø tò að n� tò sÿ
131
F�
i�g� u� r� e � 4.
�A�
l�i�g� n m� e� n t � o� f
�t� h� e � d
�e� d� u� c� e� d
�p� r� o� t� e� i� n s� e� q� u� e� n c� e� s � o� f
�P�
. � f�l�u� o� r� e� s c! e� n" s S# S
#101 $&%$ a' n d
�o� t� h� e� r � c� o� n d
�e� n s� a' t� i� o� n
d�
o� m� a' i� n s � o� f �
p� e� p� t� i� d� e � s� y( n t� h� e� t� a' s� e � s� e� q� u� e� n c� e� s� . Sequence of P. fluorescens SS101 was obtainedfrom the Tn5 flanking region of surfactant-deficient mutant 10.24. Syrm1 to syrm5 arecondensation domains from the syringomycin synthetase protein of P. syringae pv. syringaeB310D (Accession number T14593). Syppt is a condensation domain of the syringopeptinsynthetase protein from P. syringae pv. syringae B310D (AAF99707). Conse represents amajority sequence obtained from the alignement of 43 condensation domains of differentpeptide synthetases (PF00574). DhbF1 and dhbF2 are condensation domains of a peptidesynthetase involved in the synthesis of the siderophore 2,3-dihydroxybenzoate (DHB) by B.subtilis (NP_391076), and myc1 and myc2 are condensation domains of a proteinresponsible for the synthesis of microcystin by Microcystis aeruginosa K139 (BAB12211).Black-shaded residues represent high levels of similarity.
syrm2 : syrm5 : syrm3 : syrm4 : dhbF2 : conse : mcyE2 : mcyE1 : SS101 : syppt : syrm1 : dhbF1 :
* 20 * 40 * 60 SFGQQRLWFLAQME-GASAAYHIPAGLSLHGNLNLKALQRALERIVARHEGLRTTFMQ-GDDGQSYAQQRLWFLAQLE-GGSAAYHIPAGLRLRGNLDQASLQRALDRIVARHEALRTTFVQ-EQGQPSFSQQRLWFLDRLDPGASSAYHMPMSLLLRGELDHRALKAALDRLVARHESLRTTFEL-HGE-QSFSQQRLWFLDHLDHAAGAAYHLPMALRLTGALDTAALEATLDRLVARHETLRTRFEL-VDG-ESFAQRRLWFLHCLE-GPSPTYNIPVAVRLSGELDQGLLKAALYDLVCRHESLRTIFPE-SQGTSSPAQERLWFLSKLE-GGTSAYNVPFVLRLPGGLDPERLEKALKALVERHDALRTRFLR-DEG-E-------------------------AYQFKGNLNIDAFENAFQQLIQRHEILRTTFTL-INN--SEAQRQLWLLAELDRNASASYNVTTSLELLGSLDVYILEKTINQVINRHEALRTKIIE-QGE--LTGPQQDIWLDQLRVGDSPLYNIGGYLDFAGTVEPERMRRAVALLVARHDALRTQLHT--DANGLTAAQRNIWLDQMTQGDSPLYNIGGHLEIDGALDYELFQRAVDLLIAKHDALRMVLLDQRDEEG----------------------------------------------------------------TGAQTGIWFAQQLD-PDNPIYNTAEYIEINGPVNIALFEEALRHVIKEAESLHVRFGENMDG--
: 62 : 62 : 62 : 62 : 62 : 61 : 36 : 61 : 62 : 64 : - : 61
syrm2 : syrm5 : syrm3 : syrm4 : dhbF2 : conse : mcyE2 : mcyE1 : SS101 : syppt : syrm1 : dhbF1 :
* 80 * 100 * 120 PVQRISPADTGFNLQMHDLQGLA--DAEEKLQALASEESLQSFDLQQGPLIRGRLIRMAEDHHVAEQRISAAETGFRLQLQVLAGQT--DAEDKLLAIAAQEASEHFDLVNGPLVRGRLVRMANDNHVPVQVIAAADSGFALAEDDLRSQPYEQASLNASRIADSEAAAPFDLRQGPLIRGRLLRLADDEHMPVQKIAPADSRLPLLKQDLRDLSGDERTSTLARLGQENATQLFDLTKGPMLRGHLLRVADAEHVYQHILDADRACPELHVTEI-------AEKELSDRLAEAVRYSFDLAAEPAFRAELFVIGPDEYVPVQVVLEADEPDLKYLEDLSVTA---EEEEILEALRQDLQQPFDLEKGPLFRVALFKLEEDRHRQPRQVVHNQMNFVIKQIDLTAYVSTEQENLIEESISHNAKTTFDLEVGPLLKVNLLKLSQENYI--LQEVVNEVNIKLNLIDFKDED--NPEATALALRSQFSQKPFDLSIAPLFAVILMRLKPDHYLLPGQTYAPELRVEMALHDFSALP--DPSAATQALMQAQMARPYALSGEPLCRFFLVKLDHDHYRVPLQAFATSMPMPVTPIDLQGQD--DPFQAAECWMQRQLERAFRLDGEPLFRIHLLKLEAQRFY------LSKAPFKLEVTDFPEVDG--DTEQAIAHLRESFRQPFESVTGQLWESRLVRCGPQLHY-PWQMINPSPDVQLHVIDVSSEP--DPEKTALNWMKADLAKPVDLGYAPLFNEALFIAGPDRFF L
: 124 : 124 : 126 : 126 : 119 : 122 : 100 : 121 : 124 : 126 : 56 : 122
syrm2 : syrm5 : syrm3 : syrm4 : dhbF2 : conse : mcyE2 : mcyE1 : SS101 : syppt : syrm1 : dhbF1 :
* 140 * 160 * 180 * LLLTLHHIVSDGWSVDVLTRELSALYAAFSQDQDDPLAPLELQYLDYAVWQRRWLSG-----DVLLVTMHHILSDGWSTGVLNRELGALYAPFRQGAGDPLPALPVQYVDYALWQRDWLSG-----DVLLITQHHIISDGWSVGVLINEFTALYQAFTEQRPDPLPALSIQYADYAAWQRRTFTG-----ERLLITLHHIVSDGWSNSVLAQEVSALYAAFSQGQKDPLPALPLQYVDYAAWQRQSLDG-----PALLLLVHHIVGDGWSLTPLTRDLGTAYAARCHGRSPEWAPLAVQYADYALWQQELLGNEDDPNSLLLFSIHHLIVDGWSWRILLEELAALYAGLLEG----LPPLSPSYKDYAEWQQWYLQS-----DRVLFNMHHIIFDGWSAGVLIKDFLTHYHAYGQENSQLMPPLRIHYKDYTSWQESQLET-----LKLSLKTHHIVADGWSLGLILQEIGQLYSSQNNTTKEVLTPP-MQFRQYLTLRAQETQS-----PQLGTQAHHLILDGWGFGQMLQSLAQLYSALEQGRDTELLAP--SYIDFIDTDQRYLQS-----ARCMIYAHHIALDGWGIDGLCASLSELYNALHDQHTPDLTAP--SYVDFIKDDSQYRGS-----RRWFNRLHHLVADGIGAVLIAHAVSDAYSGLLEGKETLAEGP--SYLSFLEEDRAYMAS-----SRWYQRIHHIAIDGFGFSLIAQRVASTYTALIKGQTAKSRSFG-SLQAILEEDTDYRGS-----EQ HH6 DG 6 Y
: 183 : 183 : 185 : 185 : 183 : 177 : 159 : 179 : 181 : 183 : 113 : 180
syrm2 : syrm5 : syrm3 : syrm4 : dhbF2 : conse : mcyE2 : mcyE1 : SS101 : syppt : syrm1 : dhbF1 :
200 * 220 * 240 * LQQQSNFWQQTLADAPALLMLPTDRARP--ALQDYAGAALPVVFDKDLTRGLKALSQRRGSTLFLQQQRQYWQQALAGAPALLTLPTDRPRP--AQQDYSGQTLELVLDTQLTRGLKALSQRHGSSLFLAEQADLWREHLGGAPTLLSLPTDRPRP--VVQSYRGGAVPVTIDAALHQRLERFCQAHNVTLFLQAQIDFWRKHLEGAPSVLNLPLDRPRP--AIQSYTGGMVEHVFSPALSADLRAFSQAQGSTPFIAGQLAFWKETLKNLPDQLELPTDYSRP------------------------------------REKERAYWLEQLSGIEPSLQLPLDRPRP--PLQTYDGDRLTFSLSAETTALLRKLAAAYGTTLNLQGQRDYWLAKLTPVPVRLNLPVDYPRP--PVKSFQGNTITWQPEPELIDTFEKLVKAQEASLFMLEHRDFWLKTYQEDLPTFELPTDFPRP--AVKTYTGGQESQVIPSEVGQNLQKVGRKNKATLFYARDRDYWLGKYQVLPEPLLTPR-------HNAKTSSNTWVQGFPVPLQSRMEQVANHYQASAFFALDQTYWLDKYRDIPEPLLVPRPRPAQIQNEAPSRSGHLSYGLARDLEQQIDDLALSLNASSFYKRDRLFWEQSYAQLPPVLLQRRADFKAGLTNVPAPSDQVQAMLPRALYNALTQFASECNLSVAYEKDRQFWLDRFADAPEVVSLADRAPRT-----SNSFLRHTAYLPPSDVNALKEAARYFSGSWH W
: 245 : 245 : 247 : 247 : 211 : 239 : 221 : 241 : 238 : 247 : 177 : 239
syrm2 : syrm5 : syrm3 : syrm4 : dhbF2 : conse : mcyE2 : mcyE1 : SS101 : syppt : syrm1 : dhbF1 :
260 * 280 * 300 MTVMAAWAGLLGRLAGQDDVVIGTPVANRTR--SEVEGLVGLFVNTLAMTVMGAWAALLGRLSGQDDVVIGTPVANRMS--AEVENLIGFFVNTLAMGLLSAWSVLMTRLGNERDVVIGVPSANRGR--TETENLIGFFVNALAMVLLAGWSMLMSHLSEQTDVVVGTPVANRQH--PELEPMIGFFANTLA------------------------------------------------DVLLAAWGLVLSRYTGQDDIVVGTPGSGREHPIPDIERMVGWFTNTLPMGLVSLVKSFLFRYTEQNEITIGSAIAGRNH--PDLEDQIGFYVNTLVMTMFAAYTAFLRRISGHRDLVIGIPISGRQIE--GSDNLVGFCSQFLPHVLLAAMYVYFTRTSQRQEWAVGLPILNRSN--ARFRATVGLFTQVSAHVLLAVLYVYFTRAYQRDELVIGLPILNRPN--ASYKKTLGLFTQLSSHVLVGVISTYFCRTVGVDEIVVGMPVHNRTN--ARQKATIGMFSSVSPEVMIAVSAVYVHRMTGSEDVVLGLPMMGRIG--SASLNVPAMVMNLLP
: 291 : 291 : 293 : 293 : - : 287 : 267 : 287 : 284 : 293 : 223 : 285
C) h*
a+ pt,.- e/ r 0 6: 132 A4
c5 t- i6 v7 it6 - y 8 o9 f b:<;
i6 o9 s= ur>.0 f: a+ c5 t- a+ n? t- s=
132
F@
iAgB uC rD e E 5. FHG A
IlJiAgB nK mL eE nK t M oN f
OdP
eE dP uC cQ eE d P
pR rD oN tM eE iA n K sS eE qT uC eE nK cQ eE s S oN f O
PU
. V fWluXZY
o[ r\ e] s^ c_ e] ns^ SSaba
1c 0d
1 c ae nK d P
oN tM hf eE r D pR rD oN tM eE iA nK s S fOoN uC nK d
PiAnK
tM hf e E dP
ae tM ae bg ae sS eE .G For P. fluorescens SS101, the sequence was obtained from the Tn5 flankingregion of surfactant-deficient mutant 17.18. Ral.1 and Ral.2 are different domains of apeptide synthetase protein from Ralstonia solanacearum GMI1000 (NP_522203). Syrm1 tosyrm4 are different domains of of the syringomycin synthetase protein from P. syringae pv.sringae B310D (T14593). Brad. is a protein from Bradyrhizobium japonicum 110spc4(AAG61082), and Xeno. is a domain from the XpsB protein from Xenorhabdus bovieniiT228 (AAL57600). Black-shaded residues are similar in all aligned sequences.
surfactant. The CMC of the partially purified surfactant extract from strain SS101 was shown
to be between 25 and 50 µg ml-1 (Fig. 6). The effect on zoospores (motility and lysis) was
observed at concentrations of 25 µg ml-1 and higher (Fig. 6).
F@
iAgB uC rD e E 6.
h G Ri
eE lJ ae tM iA oN nK sS hf iApR s S b
geE tM wj eeEkE n K tM hf e E cQ oN nK cQ eE nK tM rD ae tM iA oN n K oN f
OtM hf e E pR ae rD tM iA ae llJlJ y m pR uC rD iA fO iA eE d
PeE xn tM rD ae cQ t M oN b
gtM ae iA nK eE d
PfOrD oN m L P
U.V
fWluXZY
o[ r\ e] s^ c_ e] ns^ SaSa
101, c d c&o sS uC rD fO ae cQ e E tM eE nK sS iA oN nK , o ae nK d P
zp oN oN sS pR oN rD e E mL oN tM iA lJ iA tM y m ae nK d P
lJym sS iA sS .G The partially purified
extract was dissolved in sterile water (pH 8.0) at different concentrations and surface tensionwas measured with a tensiometer. a At a concentration of 25 µg ml-1 and higher, the surface-active extract caused cessation of motility and subsequent lysis of zoospores of P. ultimumvar. sporangiferum.
Ral.1 : Ral.2 : syrm2 : syrm3 : Brad. : SS101 : syrm1 : Xeno. : syrm4 :
* 20 * 40 * 60 VHARLSALLGHEHAPLALAQRCSGVAAPQPLFSALLNYRHSAA-PAASAATAPTWS---GIDVLTHVRLPALMGHEYAPLAEAQRCSGVAAPQPLFGALLNYRQNMPQPEPAGQVSAAWT---GIDVLTHRDLSELLSHEQASLALAQRCSSVATGVPLFSSLLNYRHQ------NEDSQLQWP---GLRLLTHRDLSELLSHEQASLALAQRCSSVATGVPLFSSLLNYRHQ------GEDNRLQWP---GMRLLTHARLAELLTHEHASLALAQRCSAIAASAPLFSTLLNYRHNKAPAVPGSGTDDVLS---GMEWLTHARLAQLLGHEHASLSLAQRCSGVPGSLPLFSTLLNYRRS-AADEAPGDNPFAAS---GIEILTHERLTALLGHEHAPLVLAQRCSGVAAPTPLFSALLNYRHS-VVSDVR--QKLPGL---GGSSLTYQQLSLLLEHEQTPLAIAQRCSRVQAPQPLFNSLLNFRHS--QRDDKQSASPAWE---GIQILAYQQLSGLLTHERAPLALAQRCSAVDASLPMFTVILNCRHGDLVNTAGENIEDMGEEQEGVHFL L L6 HE L AQRCS 6 P6F 6LN R G L
: 60 : 61 : 55 : 55 : 61 : 60 : 58 : 59 : 64
Ral.1 : Ral.2 : syrm2 : syrm3 : Brad. : SS101 : syrm1 : Xeno. : syrm4 :
* 80 * 100 * 120 EMKERTNYPLTVSVDDLGDGFELNVQSAAGIDPERIAGYLETALVSLTEALERGGREPLHSLTVGMDERTNYPLTAVVDDLGEDFGLIVQSVPGMDAERIVGYLETALASLVASLERGGRESLRSLTVDSAERTNYPLCLSVNDYGSDLGLLIHSVQPADPQRLCAMMQCALEQLTDALAHTPQKEVTQLDVDGTERTNYPLCLSVNDYGSELDLIIHSMQPANPQRLCAMMQCALEQLTDALAHTPQMAVTQLDVGGEERTNYPLTLSVEDFGEALGLTVQVAEPISADRICGYMQRSLEQLAEALEHAPDTPVRELDISSEERSNYPLVLNVDDLGTGMRLTVQGVAELDVQRVGNYMLTALRHLVTALQQAPTTALHSVSIRHRRPYQLPVDLNVDDLGDAFTLTIQAVQQISATRIGEYMQVALRNLVDALEHTPQAALNSLSISGEERSNYPLSLDVDDFDDGFALTAQCNRQVDPARINTFMDTVLRELVAALQNVSEQSIQSLAVGSETRTNYPIEIAVANEANGFSLTAQSIDGIDPHRIAAYLGQAVAELVNALEQDPARLASSLEV P6 V 1 L R6 6 6 L L 6 6
: 124 : 125 : 119 : 119 : 125 : 124 : 122 : 123 : 128
Concentration (µq g/ml)
0.01 0.1 1 10 100 1000
Su
rfac
e te
nsi
on
(m
N/m
)
0
20
40
60
80 Cessation of motility and lysis of zoospores a

Cr hs
at ptu.v ew r x 6: y3z A{
c| tv i} v~ it} v y � o� f b�<�
i} o� s� ur�.x f� at c| tv at n� tv s�
133
D� I� S� CUr�� SS��� I� O� N�
Biosurfactants are surface-active molecules produced by microorganisms. In this study,
we demonstrated that biosurfactant-producing fluorescent Pseudomonas species occur in the
wheat rhizosphere at densities of approximately 105 CFU g-1 root, representing on average 3%
of the total population of culturable rhizospheric Pseudomonas spp. (Table 1). To our
knowledge, studies on the frequency of biosurfactant-producing Pseudomonas species in the
rhizosphere of wheat have not been reported before. To date, most biosurfactant producers
were isolated from oil-contamined soils (Arino et al., 1996; Déziel et al., 1996) and in these
environments up to 72% of the indigenous bacterial population appeared to produce
biosurfactants (Khalid and Malik, 1997).
Further characterization of the surfactant-producing Pseudomonas isolates obtained
from the wheat rhizosphere showed that six had lytic activity against zoospores of oomycetes
(Fig. 1). Comparison of representative isolate SS101 with other surfactant producers showed
that reduction of the surface tension alone is not indicative for causing lysis of zoospores
(Table 2). To date, rhamnolipids are the only biosurfactants reported with lytic activity against
zoospores (Stangellini and Miller, 1997). These compounds are produced by P. aeruginosa
strains (Itoh et al., 1971) and may serve as pathogenicity factors during the colonization of the
human lung tissue (Iglewski, 1989). There is a considerable amount of information available
on the biosynthesis and regulation of rhamnolipid production by P. aeruginosa strains
(Ochsner et al., 1994; Ochsner and Reiser, 1995). These studies led to the identification of a
cluster of two biosynthetic genes, rhlA and rhlB. In addition, rhlR and rhlI, encoding a
transcriptional activator and an autoinducer, respectively, positively regulate rhamnolipid
production. The autoinducer encoded by rhlI binds and activates RhlR (Ochsner and Raiser,
1995). Biochemical characterization of strain SS101 showed that this isolate was not classified
as P. aeruginosa but as P. fluorescens biovar II. Subsequent characterization by Southern
hybridization and PCR with primers specific for all rhamnolipid biosynthetic and regulatory
genes revealed the absence of these genes in P. fluorescens strain SS101 and other
biosurfactant-producing isolates from the wheat rhizosphere (data not shown).
Our results show that P. fluorescens strain SS101 significantly and consistently
controlled Pythium root rot of hyacinth (Figs. 1, 2). The role of biosurfactants produced by
Pseudomonas spp. in suppression of plant pathogenic fungi have been inferred from studies
with purified compounds or microcosm studies in which in situ production was correlated
with fungal inhibition (Stanghellini and Miller, 1997; Kim et al. 2000; Thrane et al. 2000). A
crucial and more conclusive step toward determining the role of biosurfactants in interactions

C� h�
a� pt�.� e� r � 6: �3� A�
c� t� i� v� it� � y � o� f b�<�
i� o� s� ur .� f� a� c� t� a� n¡ t� s�
134
T¢
a£ b¤ l¥e ¦ 4. §¨
C©
hª
a£ r« a£ c¬ t e¦ r« i® z¯ a£ t i® o° n ± o° f ²
s³ u´ r« f² a£ c¬ t a£ n± t f²r« a£ c¬ t i® o° n± s s³³ e¦ pµ a£ r« a£ t e¦ d
¶b¤
y · R¸
P¹
-º H» P¹
L¼
C©
.
Fraction RP-HPLCretention time(min)
Halo on a
S Wmedium
Drop-collapse b
testFoam c
formationZoospore d
MotilityZoospore d
Lysis
1 0-3 - - * - - -
2 3-15.5 - + * - - -
3 15.5-17.2 - + - - -4 17.2-19.7 - + - + +5 19.7-21.6 - + + + +6 21.6-24 - + - + +7 24-26.4 - + - + +/- e
8 26.4-31.5 + + + + +Water - - - - -
* + positive reaction; - negative reaction.a Evaluated on SW medium after 3 days at 25°C.b Dissolved fractions were tested on an oily surface for their ability to cause the collapse of a drop of water.c Dissolved fractions were observed for foam formation after vigorous shaking.d Zoospore motility and lysis were observed microscopically after mixing the dissolved fractions with zoospore
suspensions of P. infestans.e Lysis was not characteristic; the effect resembled leakage at one point.
with fungal pathogens is to generate biosynthetic mutants defective in the production of
biosurfactants and to compare their behaviour with the parental strain. As a first step toward
determining the role of the surfactants produced by P. fluorescens SS101 in biocontrol, we
generated mutants defective in surfactant production. By using surfactant-deficient mutant
10.24, we established the determinative role of biosurfactants produced by P. fluorescens SS101
in the control of Pythium root rot of hyacinth (Fig. 3). Further characterization of the
surfactant-deficient mutants showed that they were not impaired in the production of several
properties other than surfactants (Table 3), suggesting that apparently no global regulatory
system such as the gacS/gacA pair (Heeb and Haas, 2001) was disrupted. Reduced colonization
may have accounted for the poor performance of the mutants in the hyacinth bioassays.
However, preliminary results indicate that the surfactant-deficient mutants and the wild type
strain colonized the roots of hyacinth and wheat to the same extent (data not shown).
Sequencing of the flanking regions of the transposon in two surfactant-deficient
mutants, 10.24 and 17.18, led to the localization of the Tn5 elements in condensation
domains of peptide synthetases (Figs. 4, 5). Peptide synthetases are multifunctional enzymes
involved in the nonribosomal synthesis of diverse and often complex metabolites, including
antibiotics, siderophores, and biosurfactants (Huang et al., 2001; Marahiel et al., 1997;
Stachelhaus et al., 1998). These enzymes are composed of modules, which contain all
enzymatic activities to incorporate one constituent into the final product. These modules are
in a co-linear arrangement with the primary structure of the product, and they can be divided

C½ h¾
a¿ ptÀ.Á e r à 6: Ä3Å AÆ
cÇ tÁ iÈ vÉ itÈ Á y Ê oË f bÌ<Í
iÈ oË sÎ urÏ.Ã fÌ a¿ cÇ tÁ a¿ nÐ tÁ sÎ
135
into domains responsible for single chemical reactions (Stachelhaus et al., 1998). An initiation
module consists of an adenylation and a thiolation domain. An elongation module contains a
condensation, an adenylation, and a thiolation domain. Additional domains for substrate
modification, such as epimerization may occur (Marahiel et al., 1997). Condensation domains
are found as a part of repetitive modules. They are about 450-aa in length and coincide in
frequency with the number of peptide bonds that have to be formed for the linear peptide of
final length (Stachelhaus et al., 1998). Although in our study there is a very good correlation
between the phenotype of the two mutants and the nature of the disrupted genes,
complementation and expression studies should still be carried out to confirm and further
characterize possible pleiotropic effects of the disruptions. However, peptide synthetases are
normally very large multimodular proteins (Marahiel et al., 1990), making complementation
studies at this stage difficult to carry out.
The CMC value of the surface-active extract obtained from P. fluorescens SS101 was
reached at concentrations between 25 and 50 µg ml-1 (Fig. 6). The CMC value, the minimum
amount of surfactant required to cause the maximum decrease in surface tension, is an
important measure of the surface activity and allows comparisons with other surfactants
(Georgiou et al., 1992). The surfactants produced by P. fluorescens SS101 can lower the surface
tension of water to 29.7 mN m-1 (Table 2). The CMC and the minimum surface tension
values obtained for the surfactants of P. fluorescens SS101 are comparable to the ones found for
rhamnolipids. For example, rhamnolipids produced by P. aeruginosa UG2 lowered the surface
tension to 31.4 mN m-1 and achieved a CMC at 30 µg ml-1 (Van Dyke et al., 1993). A
biosurfactant produced by B. lycheniformis JF-2, with properties similar to surfactin, lowers the
surface tension to 27 mN m-1 and reaches the CMC at about 20 µg ml-1 (Javaheri et al., 1985).
Our studies employing RP-HPLC and LC-MS techniques showed that the surface-
active extracts from P. fluorescens SS101 contain eight different fractions (Table 4). Fraction 8
was the most abundant, accounting for approximately 95% of the extract. Preliminary analysis
suggests that fraction 8 is a cyclic lipopeptide, which is consistent with the partial sequence
obtained from the Tn5 flanking regions of the two mutants 10.24 and 17.18. The conclusive
identification of fractions 8 and fractions 1 to 7 is ongoing. Given the actvity of fractions 4 to
7 against zoospores, we expect these fractions to be precursors or derivatives of the dominant
fraction 8. A larger scale isolation and separation will be needed to provide proof for this
hypothesis. Also the activity of fractions 4 to 8 against zoospores correlates well with the
proposed mode of action of peptide antibiotics, i.e., pore formation in the membrane and
subsequent explosion (Hutchison and Johnstone, 1993). Fraction 7 caused leaking of the

CÑ hÒ
aÓ ptÔ.Õ eÖ r × 6: Ø3Ù AÚ
cÛ tÕ iÜ vÝ itÜ Õ y Þ oß f bà<á
iÜ oß sâ urã.× fà aÓ cÛ tÕ aÓ nä tÕ sâ
136
contents of the zoospore through a point in the membrane, illustrating pore formation. The
observation that effects of fraction 4 to 7 on zoospores were delayed in comparison to the
effects of fraction 8, may be related to the relatively low concentration of these fractions.
Further studies are necessary to determine the activity of all fractions at the same
concentration against zoospores and mycelium of multiple oomycetes.
Må
Aæ
Tç Eè Ré IêAæ
Lë
S ì Aæ
Ní
D î
Må
Eè Tç Hï
Oð Dî
SìMå
iñ cò ró ooôõô ró gö a÷ nø iñ sù mú s ù a÷ nø d û gö ró oô wü tý h þ cò oô nø dû iñ tý iñ oô nø sù .ÿ Naturallyoccurring Pseudomonas spp. were isolated fromroots of wheat, grown in raw soils, on King'smedium B [KMB] agar supplemented withchloramphenicol (13 µg ml-1), ampicillin (40 µgml-1), and cycloheximide (100 µg ml-1) [KMB+](Simon and Ridge, 1974). All Pseudomonas strainswere grown on KMB. Spontaneous rifampicinresistant derivatives of strain SS101 were selected onKMB supplemented with rifampicin (100 µg ml-1).Mutants disrupted in their ability to producesurfactants are resistant to rifampicin (100 µg ml-1)and kanamycin (100 µg ml-1). Escherichia coli strainS17 λ pir containing the mini-Tn5lacZ element inplasmid pUT (De Lorenzo et al., 1990) was grownin liquid Luria Bertani (LB) amended withkanamycin (25 µg ml-1). SW medium (Siegmundand Wagner, 1991), containingcetyltrimethylammoniumbromide (CTAB)-methylene blue, was used to select surfactant-producing Pseudomonas spp. Isolates that produceanionic surfactants are able to form a halo aroundthe colony after 3-5 days of growth on SWmedium. All bacterial strains were stored at -80°Cin LB or KMB-broth supplemented with 40% (v/v)glycerol.
Pythium intermedium isolate P52, one ofthe causal agents of root rot in hyacinth, wasobtained from the collection maintained at AppliedPlant Research, Section Flower Bulbs (Lisse, TheNetherlands) and used in the bioassays. P. ultimumvar. sporangiiferum CBS 219.65 and P. volutumCBS 699.83 were obtained from the Dutchcollection of microorganisms (CBS, Utrecht, TheNetherlands). Albugo candida FLS 000 andPhytophthora infestans 88069 are from the collectionmaintained in the Laboratory of Phytopathology(Wageningen University, The Netherlands). AllPythium isolates were routinely grown on potatodextrose agar (PDA; Oxoid Ltd., Basingstoke,Hampshire, England). P. infestans was grown on ryeagar medium (Caten and Jinks, 1968). A. candidawas multiplied on brussels sprout plants grown inclimate chambers at 18°C. For zoosporeproduction, P. ultimum var. sporangiiferum, P.intermedium, and P. infestans were grown on V8-
juice agar (V8; N.V.Campbell Foods, Puurs,Belgium) amended with 10 g l-1 CaCO3 (V8+) at25°C. Mycelial plugs of all Pythium isolates and P.infestans were stored in sterile mineral oil at 15°C.Leaves infested with A. candida were stored inplastic bags at -20°C.
P�
ró oô dû u� cò tý iñ oô n ø oô f �
z� ooôõô sù p� oô ró e� sù .ÿ Zoospores of P. ultimumvar. sporangiiferum (Pus) and P. intermedium wereproduced using a modification of a methoddescribed by Zhou and Paulitz (1993). Pus or P.intermedium were grown on V8+ for three days at25°C. A fully-grown agar plate was cut into 2-cmwide strips and one half of the strips was transferredto another petri plate. Both plates were floodedwith 20 ml of sterile water and kept at 25°C. After1 h, the water was discarded and replaced with thesame volume of water. Plates were incubated at18°C for four days, water was removed and replacedwith the same volume and incubated at 18°C for 2h. Zoospores of A. candida were obtained byimmersing infected leaves containing zoosporangiain sterile water followed by incubation for 2 h at15°C. P. infestans zoospores were obtained byflooding 14-day old plates grown on V8+ mediumwith 15 ml of sterile water followed by incubationat 4°C for 3h. Zoospore suspensions of 103 to 105
zoospores ml-1 were typically obtained.
Sì oô iñ l� sù . ÿ Three soils were obtained from agriculturalfields in the Netherlands. Soils CB, SV, and SSBwere collected in December 1997 from the upper50 cm of the soil profile, air dried for a week, andpassed through a 0.5-cm mesh screen prior to use.Soils CB, SV, and SSB were collected from a polderin the South-West of the Netherlands, located 10km from the city of Bergen op Zoom. These soilsare physicochemically similar, containing onaverage, 27% clay, 10% silt, and 51% sand. Theorganic matter content is on average 4.3% and thepH is 7.5. Under field conditions, all soils havebeen cropped continuously to wheat, withexception of CB soil, which was cropped to wheatand sugar beet in a rotation scheme (1:2). The soilused in the biocontrol assays with hyacinth wascollected from fields near the city of Lisse, TheNetherlands. The soil was steamed and left in open

C� h�
a pt�� e r � 6: ��� A�
c� t� i� v� it� � y � o� f b���
i� o� s� ur��� f�a c� t� a n� t� s�
137
air for six months for recolonization. This sandy soilhas an organic matter content of 1% and pH of 7.0.
W�
h� e a! t " c# u$ l%t" i& v' a! t" i& o( n) . * Pots containing 200 g of sieved
soil were sown with 15 seeds of wheat (cv. Bussard).Plants were grown in a climate chamber at 15°Cwith a 12-h photoperiod. Plants received 50 ml of1/3 strength Hoagland's solution (macroelementsonly) twice a week. After 30 days, the roots wereharvested and the remaining soil plus excised rootswere mixed, returned to the same pot and sownagain with 15 wheat seeds. This process ofsuccessively growing wheat and harvesting roots wasrepeated seven times. For each soil, four replicateswere used and the whole experiment was performedtwice.
I+ s, o( l%a! t" i& o( n ) o( f
-s, u$ r. f- a! c# t" a! n) t" -/ p0 r. o( d1 u$ c# i& n) g 2 P
3s4 e5 u6 d7 o8 m9 o8 n: a; s4 s, pp0<0 .*
Population densities of total and surfactant-producing Pseudomonas spp. were determined onroots of wheat grown for seven successive cycles of 4weeks each. For each replicate, roots of fiverandomly selected plants were harvested and looselyadhering soil was gently removed. One gram ofroots plus adhering rhizosphere soil was suspendedin 5.0 ml of sterile distilled water, vortexed for 1min and sonicated for 1 min in an ultrasoniccleaner. For isolation and enumeration ofpseudomonads, samples were dilution plated ontoKMB+ and plates were incubated at 25°C for 48 h.Approximately 10% of the total fluorescentPseudomonas colonies obtained on KMB+ wererandomly selected and tested for halo formation onSW medium. The experiment had 4 replicates andwas performed twice.
S= e l% e c# t" i& o( n ) o( f -
s, u$ r. f- a! c# t" a! n) t" -/ p0 r. o( d1 u$ c# i& n) g 2 P3
s4 e5 u6 d7 o8 m9 o8 n: a; s4 t" h� a! t"l%y> s, e z? oo(@( s, p0 o( r. e s, . * Bacterial isolates able to form a halo
arround the colony when grown on SW mediumwere screened for their ability to lyse zoospores.Bacterial suspensions containing approximately 109
CFU ml-1 were prepared from colonies grown onKMB medium for 48 h at 25°C. A 10-µl aliquot ofthe bacterial suspension was mixed on a glass slidewith an equal volume of zoospore suspensions of P.ultimum var. sporangiiferum. The behavior of thezoospores was observed under a light microscope(Dialux 20 EB; Leitz Wetzlar, Germany) at 100Xmagnification for a period up to 5 min.
BA i& o( c# h� e mB i& c# a! l % c# h� a! r. a! c# t" e r. i& z? a! t" i& o( n) . * Gas chromatography-fatty acid methyl ester (GC-FAME) analysis andAPI 20 NE test were performed to classify isolateSS101. API 20 NE tests were performed followingrecommendations of the supplier (BioMerieux,S.A., Lyon, France). For GC-FAME analysis, isolateSS101 was cultivated on tryptic soy broth agar(Becton Dickinson, Cockeysville, MD) andincubated for 24 h at 28°C. Cells were collectedwith a 4-mm-diameter transfer loop and processedfor extraction of fatty acids following the proceduresas outlined by the manufacturer (Microbial ID,
Inc., Newark, DE). Fatty acid methyl esters wereanalyzed using a Microbial Identification Systemequipped with an HP5890 series II gaschromatograph, HP3365 Chem Station and version3.9 of the aerobe library (Microbial ID, Inc.).
TC
n) 5D
mB u$ t" a! g2 e n) e s, i& s , a! n) d 1 c# h� a! r. a! c# t" e r. i& z? a! t" i& o( n ) o( f -
t" h� e mB u$ t" a! n) t" s, .* Surfactant-deficient mutants of aspontaneous rifampicin-resistant derivative of strainSS101 were obtained by biparental mating with E.coli strain S17 λ pir harbouring the mini-Tn5lacZ-Km element in plasmid pUT (De Lorenzo et al.,1990) according to protocols described bySambrook and Russel (2001). Transformants wereselected on KMB supplemented with rifampicinand kanamycin and subsequently transferred to SWmedium. Transformants unable to produce a haloarround the colony were selected after one week ofgrowth on SW medium at 25°C. RAPD analysiswith primers M13, M12, and D7, was performed toconfirm strain integrity. Southern blot analysis withthe KM probe was performed to determine thenumber of copies of the Tn5lacZ-Km element inthe surfactant-deficient mutants. Surfactant-deficient mutants were characterized for their abilityto cause lysis of zoospores of P. ultimum var.sporangiiferum, P. intermedium, A. candida and P.infestans as described above. Bacterial cellsuspensions used for zoospore lysis tests werevortexed and checked for foam formation, aphenomenon correlated with surfactant production(Zajic and Seffens, 1984). The drop collapse testwas performed as described by Jain et al. (1991) andHildebrand (1989). Protease and phospholipase Cactivities were detected by growing the bacterialisolates on skim milk and egg yolk agar, respectively(Sacherer et al., 1994). β-galactosidase activity wastested on KMB plates containing 5-bromo-4-chloro-3-indolyl-β-D-galactosidase (X-gal) andisopropylthio-β-D-galactosidase (IPTG) accordingto standard protocols (Sambrook and Russel, 2001).Fluorescence was evaluated by growing the bacterialstrains on Pseudomonas agar F plates for 48 h andsubsequent observation under near UV light (360nm).
DE
NF
A G
i& s, o( l%a! t" i& o( n) s , a! n) d d1H1 i& g2 e s, t" i& o( n) s, .* For RAPD-analysis,
heat-lysed cell suspensions were used (Raaijmakerset al., 1997). Total genomic DNA, used forSouthern blot analysis and anchored PCR (aPCR)amplifications, was extracted from bacterial strainsby a modified version of a CTAB-based protocol(Ausubel et al., 1995). A 1.5-ml sample of overnightbacterial cultures was centrifuged for 3 min at14,000 rpm, supernatant was discarded and thepellet was resuspended in 550 µl of TE buffer (Tris10mM, EDTA 10mM, pH 8.0) amended withlysozyme (1.82 mg ml-1), and incubated at 37°C for30 min. Seventy-five µl of 10% SDS amended withproteinase K (0.86 mg ml-1) was added to thebacterial suspension and thoroughly mixed. After

CI hJ
aK ptL�M eN r O 6: P�Q AR
cS tM iT vU itT M y V oW f bX�Y
iT oW sZ ur[�O fXaK cS tM aK n\ tM sZ
138
15 min of incubation at 65°C, 100 µl of 5 M NaCland 80 µl CTAB/NaCl (0.3 M CTAB, 0.7 MNaCl) were added. After 10 min of incubation at65°C, DNA was obtained by extraction withchloroform/isoamyl alcohol (24:1, vol/vol),isopropanol precipitation and subsequent washeswith 70% ethanol. The extracted DNA wasdissolved in 100 µl of 10 mM Tris pH 8.0containing RNAase (20 µg ml-1) and stored at –20°C.
For Southern hybridizations, genomicDNA was digested with 5U of EcoRI or KpnI(Promega), which are enzymes without restrictionsites in the Tn5lacZ-Km element (De Lorenzo et al.,1990). Digestions were performed in a total volumeof 100 µl containing 2.0 µg of DNA. For aPCR,the size of the fragments between the restrictionsites (ApoI or SmaI) and the Tn5lacZ-Km inserts inthe different mutants was determined by Southernhybridization with the KM probe. Genomic DNA(1 µg) was digested with ApoI or SmaI for 12 husing 10U of each enzyme at 50 and 25°C,respectively. Digested DNA was precipitated with4M LiCl, washed with 70% ethanol, dissolved in15 µl of sterile distilled water and separated on 1%agarose gels in 1X TBE.
S] o^ u_ tha eb rc n d ha ye bf rc ig dh ig zi aj tig o^ nd .k DNA transfer from agarosegels to Hybond N+ nylon membranes (AmershamPharmacia Biotech) were performed according tostandard methods (Sambrook and Russel, 2001).High-stringent conditions comprisedprehybridization for 1.5 h at 65°C, hybridizationfor 12 h at 65°C, membrane washings twice eachfor 5 min with 2X SSC (1X SSC is 0.15 M NaClplus 0.015 M sodium citrate)-0.1% SDS at roomtemperature, and membrane washings twice eachfor 30 min with 0.1X SSC-0.1% SDS at 65°C. The575-bp KM probe, specific for the kanamycinresistance gene contained within the Tn5LacZ-Kmelement, was obtained by direct PCR DIG-labelling(Roche Corp., Basel, Switzerland) of the fragmentusing primers KM1 (5’-CCCGATGCGCCAGAGTTGTT) and KM2 (5’-TCACCGAGGCAGTTCCATAGG). Hybridizedprobes were immunodetected according to theprotocols provided by the supplier.
Pl Cm R n aj nd d h Rn Ao
Pl D p aj nd aj lq ye sr eb sr .k PCR amplifications wereperformed in a 25-µl reaction mixture containing 3µl of the diluted heat-lysed cell suspension, or 10 ngof genomic DNA, 1X PCR buffer (Perkin-Elmer,Nieuwerkerk aan de IJssel, The Netherlands), 1.5mM MgCl2 (Perkin Elmer), 200 µM of each dATP,dCTP, dGTP, and dTTP (Promega, Leiden, TheNetherlands), 20 pmol of each primer, and 2U ofAmpliTaq polymerase (Perkin-Elmer). AnchoredPCR (aPCR) was performed in a 50-µl reactionmixture with the high fidelity PCR system (Roche,Roche Diagnostics, Mannheim, Germany)following the recommendations of the supplier. Forrandom amplified polymorphic DNA (RAPD)
analysis, 40 pmol of each primer were used.Amplifications were carried out with a MJ ResearchPTC-200 thermocycler. The PCR program foraPCR consisted of an initial denaturation at 95°Cfor 5 min followed by 30 cycles at 94°C for 30 sec,annealing beginning at 72ºC for 40 sec andafterwards decreasing by 0.5°C per cycle, andextension at 72°C for 2 min. The PCR program forRAPD analysis was described previously (Keel et al.,1996). RAPD analysis was performed with primersM13, M12, and D7 (Keel et al., 1996) and repeatedthree times for each primer. Samples (10-15 µl) ofthe PCR products were separated on a 1% agarosegel or on 2% agarose gels for RAPD analysis in 1XTBE buffer (90 mM Tris-borate, 2 mM EDTA[pH 8.0]) at 60-80 V for 1-3 h. The GIBCO-BRL100-bp ladder (Life Technologies B.V., Breda, TheNetherlands) was used as size marker. PCRproducts were visualized with a UVtransilluminator.
S] eb qs u_ eb nd ct ig nd g u Tv nd 5w
fxlq aj nd ky ig nd g u rc eb gu ig o^ nd sr .k DNA flanking
the Tn5lacZ-Km insertions in different mutants wassequenced by aPCR (Stuurman et al., 1996). aPCRwas performed in three steps, where the firstconsisted of ligating synthetic anchors to genomicDNA digested with the enzymes ApoI or SmaI. ForApoI (with 5’-AATT overhang) the anchor wasEcoRI-16 (5’-AATTGGCGGTGAGTCC) and forSmaI (with blunt end) the anchor was An-B (5’-TGCGGACT). The second step was a linear PCRwith primers EOF (5’-ACTTGTGTATAAGAGTCAG) or EIR (5’-AGATCTGATCAAGAGACAG) targeting theinverted repeats of the Tn5 transposon. The thirdstep consisted of performing a nested PCR withprimer RH24 (5’-AGCACTCTCCAGCCTCTCACCGCA) forDNA digested with ApoI or primer 5’-VECT24 (5’-AGCACTCTCCAGCCTCTCACCGCC) forDNA digested with SmaI, and primers EOF orEIR. Amplified fragments were purified fromagarose gels by using the QIAEX II kit (QIAGENGmbH, Hilden, Germany) and cloned intopCRT7/CT-TOPO (Invitrogen, Breda, TheNetherlands). Cloned aPCR products weresequenced with primers T7 (5’-TAATACGACTCACTATAGGG) and V5 (5’-ATCCCTAACCCTCTCCTCGGT). Sequenceswere trimed and assembled by using the DNAstarprogram (DNAstar Inc., Madison, WI, USA).BLASTN and BLASTP were conducted on thesequences. Sequences were aligned by using theCLUSTAL W 1.81 program (Thompson et al.,1994).
Bz
ig oaj ssr{r aj ye sr .k Inoculum of P. intermedium P52 wasprepared by growing the oomycete for 3 weeks atroom temperature in a sterile mixture containing50% sandy soil, 50% river sand, and oatmeal (6 gl-1). Pythium inoculum was mixed with the sandysoil at a 1% rate (v/v). Hyacinth bulbs, cv. Pink

C| h}
a~ pt��� e� r � 6: ��� A�
c� t� i� v� it� � y � o� f b���
i� o� s� ur��� f�a~ c� t� a~ n� t� s�
139
Pearl, were disinfested with 1% formaline andstored at 9°C in a climate chamber. Four weeksprior to the bioassays, the bulbs were transfered to a17°C climate chamber. Suspensions of P. fluorescensisolate SS101 or its surfactant-deficient mutant10.24 (107 CFU ml-1) were used to treat the bulbs.In soil treatments, initial bacterial concentrationswere 107 CFU g-1 soil fresh weight. Hyacinth bulbswere transferred to plastic pots filled withapproximately 2,75 kg of soil which was treated ornot with Pythium and/or P. fluorescens strains. After8 weeks of growth at 9°C in the dark, roots wereexcised from the bulbs, fresh weight was determinedand/or disease caused by P. intermedium P52 wasscored on a 0 to 5 scale. In this disease index, 0indicates no disease, 1 indicates 1 to 20% loss ofroot biomass relative to the control, 2 indicates 20to 40% loss of root biomass, 3 presents 40 to 60%less roots, 4 is 60 to 80% less roots, and 5 is 80 to100% less roots. The disease index was scoredvisually.
P� a� r� t� i� a� l � p� u� r� i� f� i� c� a� t� i� o� n � o� f �
s� u� r� f� a� c� t� a� n� t� s� . � Bacterial cellsobtained from cultures of strain SS101 grown onKMB agar for 48 h at 25°C were suspended insterile demineralized water and centrifuged twice(Sorvall RC5C; Sorvall instruments) at 6000 rpmfor 20 min at 4°C. Suspensions of the surfactant-deficient mutant 10.24 were also included. Thesupernatant containing the surfactants was filter-sterilized (0.2 µm), acidified to pH 2.0 with 9%HCl and kept on ice for 1 h (Javaheri et al., 1985).The precipitate was collected by centrifugation(6000 rpm, 30 min, 4°C) and washed with acidicdemineralized water (pH 2.0) twice. The precipitatewas resuspended in sterile demineralized water,adjusted to pH 8.0 with 0.2 M NaOH andlyophilized (Labconco Corp., Kansas City, USA).The lyophilized extract was stored at –20°C.
S� u� r� f� a� c� e � t� e� n� s� i� o� n � m e� a� s� u� r� e� m e� n� t� s� .� Bacterial cellsuspensions were obtained from cultures grown for48 h on KMB medium at 25°C. Concentrationswere adjusted to OD600=1 and therefrom dilutionswere made. For determining the critical micelleconcentration (CMC-value), the partially purifiedextract was dissolved in sterile water (pH 8.0) atconcentrations of 0, 0.01, 0.1, 1, 10, 25, 50, 100,
and 1000 µg ml-1. Surface tension measurementswere carried out with a Krüss K6 tensiometer(Krüss GmbH, Germany). Measurements wereperformed at 25°C and sterile distilled water wasused to calibrate the tensiometer.
C¡
o� m p� o� s� i� t� i� o� n � o� f �
t� h¢ e � s� u� r� f� a� c� e � a� c� t� i� v£ e e�¤� x¥ t� r� a� c� t� . � Allsolvents used were liquid chromatography grade(LC grade). Acetonitrile (MeCN) and methanolwere obtained from LAB-SCAN Analytical Sciences(Dublin, Ireland). Ultra pure water was obtainedfrom a combined Seradest LFM 20 Serapur Pro 90C apparatus (Seral, Ransbach, Germany). All LCsolvents were degassed by vacuum filtration over a0.45 µm membrane filter (Type RC, Schleicher &Schuell) prior to use. All high-performance liquidchromatography (HPLC) separations were carriedout on an Alltech end-capped C18 columns of 250mm length, particle size 5 µm. For fractionation,4.6 mm diameter columns were used. Freeze dryingwas carried out on a Christ Alpha 1-2 freeze drier.Prior to fractionation, 10 mg of the partiallypurified extract was dissolved in 1 ml MeCN - H2O(6:4). After membrane filtration, fractionation wasperformed by reverse phase-HPLC (RP-HPLC).The solvent was MeCN - H2O (7:3) amended with0.01% formic acid at a flow rate of 1.0 ml min-1.Detection took place by UV at 210 nm. Eighteeninjections of 50 µl were made and the fractions werecollected manually in round bottom flasks. Afterremoval of the MeCN with a rotary evaporator(B¸chi) in vacuo, the aqueous solution waslyophilised. All fractions were dissolved in 1.25 mlsterile water pH 8.0. Drop-collapse test, foamformation and activity against zoospores of P.infestans were conducted to determine the biologicalactivity of the separated fractions.
S� t� a� t� i� s� t� i� c� a� l � a� n� a� l� y¦ s� i� s� .� Disease index data were ranked.After certifying normal distribution andhomogeneity of variances, root weight data andranked disease index data were analysed byANOVA followed by Tukey’s studentized range test(SAS Institute, Inc., Cary, N.C.).
A§
C¡
KN© Oª W«
L¬
E
D® G¯
E
M°
E
N© T±
S�
This research was financially supported by CAPES, Brazil (Project nr. 1515/96-9). We wish to thankMr. G.P. Lelyveld for performing the preparative HPLC separation, Mr. C. van de Haar for the freezedrying ofthe fractions and Ing. J.A. Boeren (Lab. of Biochemistry) and Ing. F.C. Claassen for their help with the LC-MSanalyses. We also thank Suzanne Breeuwsma for performing part of the hyacinth bioassays and Bas Brandwagt forhelping with anchored PCR.

C² h³
a´ ptµ�¶ e· r ¸ 6: ¹�º A»
c¼ t¶ i½ v¾ it½ ¶ y ¿ oÀ f bÁ�Â
i½ oÀ sà urÄ� fÁa´ c¼ t¶ a´ nÅ t¶ sÃ
140
RÆ EÇ FÈ
EÇ RÆ EÇ NÉ
CÊ
EÇ SË
AÌ
rÍ iÎ nÏ oÐ , Ñ SË ., Ò Ñ MÓ aÔ rÍ cÕ hÖ aÔ l× , Ñ RÆ ., Ò Ñ aÔ nÏ d Ø
VÙ
aÔ nÏ dØ
eÚ cÕ aÔ sÛ tÜ eeÚÝÚ l× eÚ , Ñ JÞ.Ò Pß .Ò
1996. Identification and production of arhamnolipid biosurfactant by a Pseudomonas species.Appl. Microbiol. Biotechnol. 45:162-168.AÌ
uà sÛ uà bá
eÚ l× , Ñ FÈ
.Ò MÓ ., Ò Ñ Bâ
rÍ eÚ nÏ tÜ , Ñ RÆ ., Ò Ñ Kã
iÎ nÏ gä sÛ tÜ oÐ nÏ , Ñ RÆ .Ò EÇ ., Ò Ñ MÓ ooÐ@Ð rÍ eÚ ,ÑDå .Ò Då ., Ò Ñ SË eÚ iÎ dØ mæ aÔ nÏ , Ñ J
Þ.Ò Gç ., Ò Ñ SË mæ iÎ tÜ hÖ , Ñ J
Þ.Ò AÌ ., Ò Ñ aÔ nÏ d
ØSË tÜ rÍ uà hÖ l
×, Kã
.Ò(ed.). 1992. Short protocols in molecular biology,pages 16.8-16.9, John Wiley and Sons, New York,N.Y.CÊ
aÔ tÜ eÚ nÏ , Ñ EÇ .Ò CÊ ., Ò Ñ aÔ nÏ d Ø
JÞiÎ nÏ kè sÛ , Ñ J
Þ.Ò Lé .Ò 1968. Spontaneous
variability of single isolates of Phytophthora infestans.Can. J. Bot. 46:329-348.CÊ
ooÐ@Ð pê eÚ rÍ , Ñ Då .Ò Gç ., Ò Ñ aÔ nÏ d Ø
Zë aÔ jì iÎ cÕ , Ñ JÞ.Ò EÇ . Ò 1980íïî<ðòñ . Ò Surface-active
compounds from microorganisms. Adv. Appl.Microbiol. 26:229-253.Då e Ú Lé oÐ rÍ eÚ nÏ zó oÐ , Ñ V
Ù., Ò Ñ Hô eÚ rrÍ{Í eÚ rÍ oÐ , Ñ MÓ ., Ò Ñ J
ÞaÔ kè uà b
ázó iÎ kè , Ñ U
õ., Ò Ñ aÔ nÏ d
ØTö iÎ mæ mæ iÎ sÛ , Ñ K
ã.Ò NÉ .Ò 1990. Mini-Tn5 transposon
derivatives for insertion mutagenesis, promoterprobing, and chromosomal insertion of clonedDNA in gram-negative eubacteria. J. Bacteriol.172:6568-6572.Då é÷ zó iÎ eÚ l× , Ñ EÇ ., Ò Ñ Pß aÔ qø uà eÚ tÜ tÜ eÚ , Ñ G
ç., Ò Ñ VÙ
iÎ ll×ù× eÚ mæ uà rÍ , Ñ RÆ ., Ò Ñ Lé eÚ pê iÎ nÏ eÚ , Ñ FÈ
.,Ò ÑaÔ nÏ d Ø
Bâ
iÎ sÛ aÔ iÎ ll×ù× oÐ nÏ , Ñ JÞ.Ò Gç .Ò 1996. Biosurfactant production
by a soil Pseudomonas strain growing on polycyclicaromatic hydrocarbons. Appl. Environ. Microbiol.62:1908-1912.FÈ
iÎ eÚ cÕ hÖ tÜ eÚ rÍ , Ñ AÌ
.Ò 1992. Biosurfactants: moving towardsindustrial application. Tibtech 10:208-217.Gç
eÚ oÐ rÍ gä iÎ oÐ uà , Ñ Gç
., Ò Ñ Lé iÎ nÏ , Ñ SË ., Ò Ñ aÔ nÏ d Ø
SË hÖ aÔ rÍ mæ aÔ , Ñ MÓ .Ò MÓ . Ò 1992.Surface-active compounds from microorganisms.Bio/Technology 10:60-65.Hô eeÚúÚ bá , Ñ SË ., Ò Ñ aÔ nÏ d
ØHô aaÔúÔ sÛ , Ñ Då . Ò 2001. Regulatory roles of
the GacS/GacA two-component system in plant-associated and other gram negative bacteria. Mol.Plant-Microbe Interact. 14:1351-1363.Hô eÚ rÍ mæ aÔ nÏ , Ñ Då .Ò CÊ ., Ò Ñ A
ÌrÍ tÜ iÎ oÐ l
×aÔ , Ñ J
Þ.Ò FÈ ., Ò Ñ aÔ nÏ d
ØMÓ iÎ ll×ù× eÚ rÍ , Ñ RÆ .Ò MÓ .Ò
1995. Removal of cadmium, lead, and zinc fromsoil by rhamnolipid biosurfactant. Environ. Sci.Technol. 29:2280-2285.Hô iÎ l× dØ eÚ bá rÍ aÔ nÏ d
Ø, Ñ Pß .Ò Då .Ò 1989. Surfactant-like
characteristics and identity of bacteria associatedwith brocoli head rot in Atlantic Canada. Can. J.Plant Pathol. 11:205-214.Hô iÎ l× dØ eÚ bá rÍ aÔ nÏ d
Ø, Ñ Pß .Ò Då ., Ò Ñ B
ârÍ aÔ uà nÏ , Ñ Pß .Ò Gç ., Ò Ñ MÓ cÕ RÆ aÔ eÚ , Ñ K
ã.Ò Bâ ., Ò Ñ aÔ nÏ d
ØLé uà , Ñ X
û.Ò 1998. Role of the biosurfactant viscosin in
broccoli head rot caused by pectolitic strains ofPseudomonas fluorescens. Can. J. Plant Pathol.20:296-303.Hô uà aÔ nÏ gä , Ñ G
ç., Ò Ñ Zë hÖ aÔ nÏ gä , Ñ Lé ., Ò Ñ aÔ nÏ d
ØBâ
iÎ rÍ cÕ hÖ , Ñ RÆ .Ò Gç .Ò 2001. Amultifunctional polyketide-peptide synthetaseessential for albicidin biosynthesis in Xanthomonasalbilineans. Microbiol. 147:631-642.Hô uà tÜ cÕ hÖ iÎ sÛ oÐ nÏ , Ñ MÓ .Ò Lé ., Ò Ñ aÔ nÏ d
ØJÞoÐ hÖ nÏ sÛ tÜ oÐ nÏ eÚ , Ñ K
ã.Ò 1993.
Evidence for the involvement of the surface activeproperties of the extracellular toxin tolaasin in themanifestation of brown blotch disease symptms byPseudomonas tolaasii on Agaricus bisporus. Physiol.Mol. Plant Pathol. 42:373-384.
Iügä l×eÚ wý sÛ kè iÎ , Ñ B
â.Ò 1989. Probing Pseudomonas aeruginosa,
an opportunistic pathogen. ASM News 55:303-307.IütÜ oÐ hÖ , Ñ SË ., Ò Ñ Hô oÐ nÏ d
ØaÔ , Ñ Hô ., Ò Ñ Tö oÐ mæ iÎ tÜ aÔ , Ñ F
È., Ò Ñ aÔ nÏ d
ØSË uà zó uà kè iÎ , Ñ Tö .Ò
1971. Rhamnolipids produced by Pseudomonasaeruginosa grown on n-paraffin (mixture of C12, C13
and C14 fractions). J. Antibiot. 24:855-859.JÞaÔ iÎ nÏ , Ñ Då .Ò Kã ., Ò Ñ C
ÊoÐ ll×þ×
iÎ nÏ sÛ -ÿ Tö hÖ oÐ mæ pê sÛ oÐ nÏ , Ñ Då .Ò Lé ., Ò Ñ Lé eeÚÝÚ , Ñ Hô ., Ò Ñ aÔ nÏ dØ
Tö rÍ eÚ v� oÐ rÍ sÛ , Ñ JÞ.Ò Tö . Ò 1991. A drop-collapsing test for
screening surfactant-producing microorganisms. J.Microbiol. Methods 13:271-279.JÞaÔ v� aÔ hÖ eÚ rÍ iÎ , Ñ MÓ .Ò , Ñ J
ÞeÚ nnÏ Ï eÚ mæ aÔ nÏ , Ñ G
ç.Ò EÇ ., Ò Ñ MÓ cÕ Iü nÏ eÚ rÍ nÏ eÚ y� , Ñ MÓ .Ò JÞ .Ò ,Ñ
aÔ nÏ d Ø
Kã
nÏ aÔ ppê<ê , Ñ RÆ .Ò MÓ .Ò 1985. Anaerobic production of abiosurfactant by Bacillus licheniformis JF-2. Appl.Environ. Microbiol. 50:698-700.Kã
eeÚÝÚ l× , Ñ CÊ
., Ò Ñ W�
eÚ l× l× eÚ rÍ , Ñ Då .Ò MÓ ., Ò Ñ NÉ
aÔ tÜ sÛ cÕ hÖ , Ñ AÌ
., Ò Ñ Då é÷ f� aÔ gä oÐ , Ñ CÊ
ooÐ@Ð kè ,ÑRÆ .Ò JÞ ., Ò Ñ aÔ nÏ d
ØTö hÖ oÐ mæ aÔ sÛ hÖ oÐ wý , Ñ Lé .Ò SË .Ò 1996. Conservation of
the 2,4-diacetylphloroglucinol biosynthesis locusamong fluorescent Pseudomonas strains from diversegeographic locations. Appl. Environ. Microbiol.552-563.Kã
hÖ aÔ l× iÎ dØ , Ñ Zë .Ò MÓ ., Ò Ñ aÔ nÏ d Ø
MÓ aÔ l× iÎ kè , Ñ Kã
.Ò AÌ . Ò 1988. Isolation offurnace oil utilizing bacteria capable of producingbiosurfactant. Pakistan J. Sci. Ind. Res. 31:714-717.Kã
iÎ mæ , Ñ Bâ
.Ò SË ., Ò Ñ Lé eeÚúÚ , Ñ JÞ.Ò Y� ., Ò Ñ aÔ nÏ d
ØHô wý aÔ nÏ gä , Ñ B
â.Ò Kã .Ò 2000. In
vivo control and in vitro antifungal activity ofrhamnolipid B, a glycilipid antibiotic, againstPhytophthora capsici and Colletotrichum orbiculare.Pest. Manag. Sci. 56:1029-1035.MÓ aÔ rÍ aÔ hÖ iÎ eÚ l× , Ñ MÓ .Ò AÌ ., Ò Ñ N
ÉaÔ kè aÔ nÏ oÐ , Ñ MÓ .Ò MÓ ., Ò Ñ aÔ nÏ d
ØZë uà b
áeÚ rÍ , Ñ Pß .Ò
1993. Regulation of peptide antibiotic productionin Bacillus. Mol. Microbiol. 7:631-636.MÓ aÔ rÍ aÔ hÖ iÎ eÚ l× , Ñ MÓ .Ò AÌ ., Ò Ñ SË tÜ aÔ cÕ hÖ eÚ l× hÖ aÔ uà sÛ , Ñ Tö ., Ò Ñ aÔ nÏ d
ØMÓ ooÐ@Ð tÜ zó , Ñ Hô .Ò Då .Ò
1997. Modular peptide synthetases involved innonribosomal synthesis. Chem. Rev. 97:2651-2673.NÉ
eÚ uà , Ñ Tö .Ò RÆ .Ò 1996. Significance of bacterial surface-active compounds in interaction of bacteria withinterfaces. Microbiol. Rev. 60:151-166.NÉ
iÎ eÚ l× sÛ eÚ nÏ , Ñ Tö .Ò Hô ., Ò Ñ CÊ
hÖ rÍ iÎ sÛ tÜ oÐ pê hÖ eÚ rÍ sÛ eÚ nÏ , Ñ CÊ
., Ò Ñ AÌ
nÏ tÜ hÖ oÐ nÏ iÎ , Ñ Uõ
.,Ò ÑaÔ nÏ d Ø
SË ø� rÍ eÚ nÏ sÛ eÚ nÏ , Ñ JÞ.Ò 1999. Viscosinamide, a new cyclic
depsipeptide with surfactant and antifungalproperties produced by Pseudomonas fluorescensDR54. J. Appl. Microbiol. 86:80-90.NÉ
iÎ eÚ l× sÛ eÚ nÏ , Ñ Tö .Ò Hô ., Ò Ñ Tö hÖ rÍ aÔ nÏ eÚ , Ñ CÊ
.Ò , Ñ CÊ
hÖ rÍ iÎ sÛ tÜ oÐ pê hÖ eÚ rÍ sÛ eÚ nÏ , Ñ CÊ
.,Ò ÑAÌ
nÏ tÜ hÖ oÐ nÏ iÎ , Ñ Uõ
., Ò Ñ aÔ nÏ d Ø
SË ø� rÍ eÚ nÏ sÛ eÚ nÏ , Ñ JÞ.Ò 2000. Structure,
production characteristics and fungal antagonism oftensin - a new antifungal cyclic lipopeptide fromPseudomonas fluorescens 96.578. J. Appl. Microbiol.89:992-1001.O�
cÕ hÖ sÛ nÏ eÚ rÍ , Ñ Uõ
.Ò AÌ ., Ò Ñ aÔ nÏ d Ø
RÆ eÚ iÎ sÛ eÚ rÍ , Ñ JÞ.Ò 1995. Autoinducer-
mediated regulation of rhamnolipid biosurfactantsynthesis in Pseudomonas aeruginosa. Proc. Natl.Acad. Sci. USA 92:6424-6428.O�
cÕ hÖ sÛ nÏ eÚ rÍ , Ñ Uõ
.Ò AÌ ., Ò Ñ FÈ
eÚ iÎ cÕ hÖ tÜ eÚ rÍ , Ñ AÌ
., Ò Ñ aÔ nÏ d Ø
RÆ eÚ iÎ sÛ eÚ rÍ , Ñ JÞ.Ò 1994.
Isolation, charaterization and expression inEscherichia coli of the Pseudomonas aeruginosa rhlABgenes encoding rhamnosyltransferase involved in

C� h�
a pt�� e r � 6: ��� A�
c� t� i� v� it� � y � o� f b���
i� o� s� ur��� f� a c� t� a n� t� s�
141
rhamnolipid synthesis. J. Biol. Chem. 269:19787-19795.R� aa� � i! j" m# a� k$ e% r& s' , ( J
).* M+ .* , ( W
,e% ll-.- e% r& , ( D
/.* M+ .* , ( a� n0 d
1T2 h3 o4 m# a� s' h3 o4 w5 ,(
L6 .* S7 .* 1997. Frequency of antibiotic-producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63:881-887.R� o4 n0 , ( E8 .* Z9 ., *:( a� n0 d
1R� o4 s' e% n0 b
;e% r& g< , ( E8 .* 2001. Natural roles
of biosurfactants. Environ. Microbiol. 3:229-236.S7a� c= h3 e% r& e% r& , ( P> ., *.( D
/é? f@ a� g< o4 , ( G
A., *.( a� n0 d
1HB aa�C� s' , ( D
/.* 1994.
Extracellular protease and phospholipase C arecontrolled by the global regulatory gene gacA in thebiocontrol strain Pseudomonas fluorescens CHA0.FEMS Microbiol. Letters 116:155-160.S7a� m# b
;r& oo4D4 k
$, ( J)., *.( a� n0 d
1R� uE ss'F' e% l- , ( D
/.* W, .* 2001. Molecular
cloning: a laboratory manual, 3rd ed. Cold SpringHarbor Laboratory, Cold Spring Harbor, N.Y.S7c= h3 o4 l- zG -H S7 c= h3 r& o4 e% d1 e% r& , ( B
I., *.( S
7o4 uE l- e% , ( J
).* D/ ., *.( L6 uE , ( S
7.* -H E8 .,*.(
GA
r& g< uE r& i! n0 a� , ( IJ., *.( a� n0 d
1GA
r& o4 ss'F' , ( D/
.* 2001. A physical mapof the syringomycin and syringopeptin gene clusterslocalized to approximately 145-kb DNA region ofPseudomonas syringae pv. syringae strain B301D.Mol. Plant-Microbe Interact. 12:1426-1435.S7e% rr&K& a� , ( M+ .* D/ ., *:( F
La� g< i
!uE o4 l- i! , ( G
A., *.( NM o4 r& d1 e% r& a� , ( P> ., *.( B
Ie% r& n0 h3 a� r& tN , ( I
J.,*.(
VO
o4 l- pP e% , ( CQ .* D/ ., *:( D/
i !
GA
i!o4 r& g< i
!o4 , ( D
/., *:( B
Ia� ll-.- i! o4 , ( A
R., *.( a� n0 d
1M+ e% n0 e% s' tN r& i! n0 a� , ( G
A.* 1999. The interaction of
lipodepsipeptide toxins from Pseudomonas syringaepv. syringae with biological and model membranes:a comparison of syringotoxin, syringomycin, andtwo syringopeptins. Mol. Plant-Microbe Interact.12:391-400.S7i!e% g< m# uE n0 d
1, ( IJ., *.( a� n0 d
1W,
a� g< n0 e% r& , ( FL
.* 1991. New methodfor detecting rhamnolipids excreted by Pseudomonasspecies during growth on mineral agar.Biotechnol.Techn. 5: 265-268.S7i!m# o4 n0 , ( A
R., *.( a� n0 d
1R� i!d1
g< e% , ( E8 .* HB .* 1974. The use ofampicillin in a simplified selective medium for theisolation of fluorescent pseudomonads. J. Appl.Bacteriol. 37:459-460.S7tN a� c= h3 e% l- h3 a� uE s' , ( T2 ., *:( M+ oo4D4 tN zG , ( HB .* D/ ., *:( B
Ie% r& g< e% n0 d
1a� h3 l- , ( V
O., *.( a� n0 d
1M+ a� r& a� h3 i
!e% l- , ( M+ .* AR .* 1998. Peptide bond formation in
nonribosomal peptide biosynthesis: catalitic role ofthe condensation domain. J. Biol. Chem.273:22773-22781.
S7tN a� n0 g< h3 e% ll-.- i! n0 i
!, ( M+ .* E8 . * a� n0 d
1M+ i
!ll-:- e% r& , ( R� .* M+ . * 1997.
Biosurfactants: their identity and potential efficacyin the biological control of zoosporic plantpathogens. Plant Dis. 81: 4-12.S7tN uE uE r& m# a� n0 , ( J
)., *.( D
/e % VO
r& oo4D4 m# e% n0 , ( M+ .* J) ., *.( NM i!j"k$a� m# pP , ( HB .* J) .* J) .,*.(
a� n0 d 1
VO
a� n 0 HB a� a� r& e% n0 , ( M+ .* J) .* J) .* 1996. Single sitemanipulation of tomato chromossomes in vitro andin vivo using Cre-lox site-specific recombination.Plant Mol. Biol. 32:901-913.T2 h3 r& a� n0 e% , ( CQ .* , ( O
Sl- ss'F' o4 n0 , ( S
7., *.( NM i
!e% l- s' e% n0 , ( T2 .* HB ., *.( a� n0 d
1S7øT r& e% n0 s' e% n0 , ( J
).* 1999. Vital fluorescent stains for
detection of stress in Pythium ultimum andRhizoctonia solani challenged with viscosinamidefrom Pseudomonas fluorescens DR54. FEMSMicrobiol. Ecol. 30:11-23.T2 h3 o4 m# pP s' o4 n0 , ( J
).* D/ ., *.( HB i
!gg<U< i
!n0 s' , ( D
/.* GA ., *.( a� n0 d
1GA
i!b;
s' o4 n0 , ( T2 .* J) .*1994. CLUSTAL W: impoving the sensitivity ofprogressive multiple alignment through sequenceweighting, positions-specific gap penalties andweight matrix choice. Nucleic Acids Res. 22:4673-4680.VO
a� n 0 D/
yV k$e% , ( M+ . * I
J., *.( CQ o4 uE tN uE r& e% , ( P> ., *:( B
Ir& a� uE e% r& , ( M+ ., *.( L6 ee% % , ( HB .,*.(
a� n0 d 1
T2 r& e% vW o4 r& s' , ( J).* T2 .* 1993. Pseudomonas aeruginosa
UG3 rhamnolipid biosurfactants: structuralcharacterization and their use in removinghydrophobic compounds from soil. Can. J.Microbiol. 39:1071-1077.VO
a� n0 i!ttNXN a� n0 a� k$ o4 m# , ( NM ., *.( a� n0 d
1L6 o4 e% ff@Y@ l- e% r& , ( W
,.* 1986.
Fengycin - a novel antifungal lipopeptide antibioticproduced by Bacillus subtilis F-29-3. J. Antibiot.39:888-901.Z9 a� j" i! c= , ( J
).* E8 ., *.( a� n0 d
1S7e% ff@Z@ e% n0 s' , ( W
,. * 1984. Biosurfactants.
Critical Rev. Biotech. 1:87-107.Z9 h3 a� n0 g< , ( Y
[. * a� n0 d
1M+ i
!l- l- e% r& , ( R� .* M+ .* 1994. Effect of a
Pseudomonas rhamnolipid biosurfactant on cellhydrophobicity and biodegradation of octadecane.Appl. Environ. Microbiol. 60:2101-2106.Z9 h3 o4 uE , ( T2 ., *:( a� n0 d
1P> a� uE l- i! tN zG , ( T2 .* CQ .* 1993. In vitro and in
vivo effects of Pseudomonas spp. on Pythiumaphanidermatum: zoospore behavior in exudates andon the rhizoplane of bacteria-treated cucumberroots. Phytopathol. 83:872-876.


_______________________________________________________________________________
CHAPTER 7
S\U]
MM^_^
A`
Ra
IbS\IbNc
G d
De
IbS\Cf
U]
SS\g\
IbOh
Nc
_______________________________________________________________________________

Ci hj
ak ptl�m en r o 7: prq
Ss ut mu mu ak ro iv sw iv nx g y Dz iv sw c{ ut ssw|w iv o} nx
144
Because of the hazardous effects that pesticides may have on the environment and
human health, there is a need for more environmentally safe methods to control plant diseases.
Over the past decades, there has been an increasing interest in biological control of plant
diseases by application of beneficial microorganisms. Among the potential bacterial biocontrol
agents, the genus Pseudomonas is recognized as one of the most promising groups (Weller,
1988; Handelsman and Stabb, 1996; O’Sullivan and O’Gara, 1992). Pseudomonas biocontrol
agents suppress plant diseases by several different mechanisms, including antibiosis,
competition, and induction of resistance in the host plant (O’Sullivan and O’Gara, 1992;
Handelsman and Stabb, 1996; Van Loon et al. 1998; Whipps, 2001).
Our studies focused on fluorescent Pseudomonas species that produce antibiotics that
are inhibitory to the growth of various plant pathogenic fungi and oomycetes. Several
antibiotics produced by Pseudomonas species play a key role in biological control of plant
pathogens, including 2,4-diacetylphloroglucinol (2,4-DAPG), phenazines (PHZ), pyrrolnitrin
(PRN), pyoluteorin (PLT), and surface-active antibiotics (Vincent et al., 1991; Hill et al.,
1994; Maurhoffer et al., 1992; Thomashow and Weller, 1988; Thrane et al., 1999). To date,
however, most of the information available on antibiotic-producing Pseudomonas spp. is
derived from studies on model strains and little is known about the distribution, diversity and
activity of indigenous populations harbouring these traits. A better understanding of the
ecology of naturally occurring strains that harbour specific biocontrol traits will contribute to
improve the efficacy of existing biocontrol strains and may help to identify new strains that are
better adapted to specific soils or host-pathogen systems. Our m~ a� i� n � o� b�
j�e� c� ti� � v� e� s� in this thesis
were to study 1) the distribution, 2) the diversity and 3) the activity of indigenous
Pseudomonas strains that produce 2,4-DAPG, PHZ, PRN, PLT, or surface-active antibiotics.
D� I� S� T�
R� I� B� U�
T�
I� O� N�
Antibiotic-producing Pseudomonas species have been isolated from different soils and
plant species. The natural ability of certain soils to suppress plant diseases is often correlated
with the presence of relatively high densities of antibiotic-producing Pseudomonas spp. (Cook
and Rovira, 1976; Stutz et al., 1986; Raaijmakers et al., 1997). The exploitation of natural
disease-suppressive soils as a source of antagonistic microorganisms has proven to be a
successfull approach to develop biocontrol strategies (Whipps, 1997; Weller, 1988). However,
the selection procedures used to date have a random nature, are labour-intensive and do not
make full use of existing knowledge of biocontrol mechanisms. The increasing availability of

C� h�
a� pt��� e� r � 7: �r�
S� u� m� m� a� r� i� s i� n¡ g ¢ D£ i� s c¤ u� ss | i� o¥ n¡
145
cloned and sequenced genes involved in the biosynthesis of biocontrol traits has facilitated the
development of specific primers and probes for targeted isolation. The conservation of
sequences within the biosynthetic loci for 2,4-DAPG and PHZ antibiotics allowed the
development of primers and probes to specifically isolate and monitor Pseudomonas strains,
producing these antibiotics, in soil and rhizosphere environments (Raaijmakers et al., 1997;
Picard et al., 2000).
We have used molecular probes and primers available for 2,4-DAPG and PHZ
antibiotics (Raaijmakers et al., 1997; Mavrodi et al., unpublished) and developed new ones for
PRN and PLT, two antibiotics with broad-spectrum activity. Colony hybridization followed
by PCR was used to determine the frequencies of antibiotic-producing Pseudomonas and
Burkholderia species in soils suppressive to Geaumannomyces graminis var. tritici, the causal
agent of take-all disease of wheat (C� h�
a� p� t� e� r� s 2 ¦
a� n¡ d §
4¨
). Soils suppressive to take-all are referred
to as take-all decline (TAD) soils. We demonstrated that Pseudomonas and Burkholderia species
harbouring PRN and PLT genes were not present at detectable densities (< 104 CFU g-1 root)
on roots of wheat grown in any of the Dutch TAD soils investigated. These results suggest
that Pseudomonas and Burkholderia species that produce PRN and/or PLT do not play a
significant role in the suppressiveness that operates in at least two Dutch TAD soils. This
suggestion is based on the observation that threshold densities of at least 105 CFU g-1 root are
required for significant suppression of several soil-borne diseases by Pseudomonas strains
(Raajmakers et al., 1995; Raaijmakers and Weller, 1998). Further studies will be necessary to
establish whether these threshold population densities also apply for PRN- and PLT-
producing Pseudomonas and Burkholderia spp. Almost all PRN- and PLT-producing reference
strains reported to date were isolated from dicotelydon plant species rather than
monocotelydon (Howell and Stipanovic, 1979; Levy et al., 1992; Keel et al., 1996). The fact
that also in our studies no PRN and PLT producers were found in the rhizosphere of wheat
may point to a preference of Pseudomonas spp. harboring these traits for the rhizosphere of
dicotelydon plants. Recent studies demonstrated that wheat plants enrich for specific
genotypes of 2,4-DAPG-producing Pseudomonas species (Wang et al., 2001). This specific
enrichment could be responsible for the displacement of PRN- and PLT-producing strains
from the wheat rhizosphere. Since only a limited number of soils and only one moncotelydon
plant species was included in our studies, a detailed analysis of multiple and diverse soils and
host plants will be required to support this hypothesis. Understanding the host preference of
antagonistic microorganisms harboring particular traits may be crucial in predicting their
potential biocontrol efficacy in multiple host-pathogen systems.

C© hª
a« pt¬� e® r ¯ 7: °r±
S² u³ m´ m´ a« riµ s¶ iµ n· g ¸ D¹ iµ s¶ cº u³ ss¶|¶ iµ o» n·
146
Pseudomonas strains producing PHZ antibiotics were detected at densities of
approximately 1.0 X 105 CFU g-1 root on roots of wheat grown in SL soil, one of the soils with
history of long-term wheat cultivation. The PHZ-producing Pseudomonas spp. occurred
together with 2,4-DAPG-producing Pseudomonas spp. (C© hª
a« p¬ t e® r ¯ 2¼
). The co-occurrence of
indigenous 2,4-DAPG and PHZ-producers on roots of wheat allows the study of interactions
and compatibility between both groups as well as their potentially synergistic role in disease
suppressiveness. In this context, it is interesting to note that several studies have shown that
simultaneous application of two or more antagonistic strains can lead to improved disease
suppression (Dunne et al., 1998; De Boer et al., 1999). 2,4-DAPG-producing Pseudomonas
species occurred on roots of wheat grown in most of the soils included in our studies. Their
densities were not detectable or significantly lower on roots of wheat grown in the take-all
conducive CB soil than on roots of wheat grown in at least two take-all suppressive soils (105 –
106 CFU g-1 root or higher). Their densities on roots of wheat grown in these TAD soils were
above the threshold populations reported to be required for significant suppression of take-all
of wheat (Raaijmakers and Weller, 1998). Subsequent studies with 2,4-DAPG-deficient
mutants of representative strains of dominant genotypic groups recovered from Dutch and US
TAD soils, indicated that 2,4-DAPG production is a determinant factor in the suppression of
take-all of wheat (C© hª
a« p¬ t e® r ¯ 4½
). These findings provide substantial support for a broader role of
2,4-DAPG-producing Pseudomonas in the natural suppressiveness of soils to take-all,
previously demonstrated to occur only in TAD soils of Washington State, USA (Raaijmakers
et al., 1997; Raaijmakers and Weller, 1998). Further studies will be necessary to determine the
role of 2,4-DAPG producers in TAD soils from other geographic regions. Also their
occurrence at relatively high densities in the rhizosphere of crops other than wheat and in soils
with no apparent relation with the TAD phenomenon (Picard et al., 2000), deserves further
investigations into their biological role in these environments. The specific enrichment of 2,4-
DAPG-producing Pseudomonas in TAD soils is thought to occur, at least in part, by selective
stimulation of nutrients released from the lesions caused by the take-all pathogen during the
initial severe outbreaks of the disease. Our initial studies on G. graminis-induced selection of
2,4-DAPG-populations, however, indicated that the quantitative enrichment is not G.
graminis specific, since similar increases in population densities of both total fluorescent and
2,4-DAPG-producing Pseudomonas populations were observed upon infection of wheat roots
by Rhizoctonia solani (C© hª
a« p¬ t e® r ¯ 4½
).
Another approach often used to select for potential biocontrol agents consists of
initially establishing their activity against plant pathogens and later identifying the

C¾ h¿
aÀ ptÁ� eà r Ä 7: ÅrÆ
SÇ uÈ mÉ mÉ aÀ rÄ iÊ sË iÊ nÌ g Í DÎ iÊ sË cÏ uÈ ssË|Ë iÊ oÐ nÌ
147
mechanism(s) responsible for this activity (Handelsman and Stabb, 1996). In fact, most model
strains known to date were identified by this approach. With this approach, we identified
Pseudomonas isolates that produce surface-active antibiotics with activity against oomycete
pathogens (C¾ h¿
aÀ pÁ t eà r Ä 6Ñ ). Surfactant-producing Pseudomonas spp. occurred on roots of wheat at
densities of approximately 105 CFU g-1 root, representing on average 3% of the population of
culturable Pseudomonas spp. present in the rhizosphere of wheat. Subsequent characterization
of 375 random isolates demonstrated that 6 isolates were able to cause cessation of motility
and subsequent lysis of zoospores of at least three oomycete pathogens. Studies with probes
and primers specific for biosynthetic genes involved in the production of this surfactant are
now being conducted to obtain more information on the distribution of Pseudomonas
harbouring this particular trait in other soil, rhizosphere and phyllosphere environments.
Although in this thesis we focused only on one of the strains that adversely affected motility
and caused lysis of zoospores, the remaining 369 isolates may produce surface-active
compounds that have other and possibly stronger antagonistic properties than observed for the
surfactant(s) produced by P. fluorescens strain SS101 (C¾ h¿
aÀ ptÁÒ eà r Ä 6Ñ ).
DÎ IÓ VÔ EÕ
RÖ SÇ IÓ T× YØ
Several studies have shown that there is considerable diversity in antibiotic-producing
Pseudomonas species (Keel et al., 1996; Sharifi-Tehrani et al., 1998; McSpadden-Gardener et
al., 2000). Most of these studies are based on polymorphisms in the 16S-rDNA. Work by
Ramette et al. (2001) and Mavrodi et al. (2001) demonstrated that also polymorphisms within
phlD, a key gene in the biosynthesis of 2,4-DAPG, can be used for diversity and phylogenetic
studies on 2,4-DAPG-producing Pseudomonas strains. In C¾ h¿
aÀ pÁ t eà r Ä 2Ù
, Random Fragment
Length Polymorphism (RFLP) analysis of amplified fragments of pltC showed no
polymorphisms, indicating that at least part of this biosynthetic gene is highly conserved
among known PLT-producing Pseudomonas species. In contrast, RFLP analysis of prnD-
fragments allowed discrimination between multiple PRN-producing Pseudomonas and
Burkholderia species to a similar level as obtained by Random Amplified Polymorphic DNA
(RAPD) analysis. Interestingly, B. pyrrocinia strain DSM10685 was clustered closer to
Pseudomonas than to other Burkholderia strains. Subsequent phylogenetic analysis with the
four pyrrolnitrin biosynthetic genes from three other strains, including one B. cepacia, one P.
fluorescens, and one Myxococcus fulvus confirmed that prn genes of B. pyrrocinia are more

CÚ hÛ
aÜ ptÝ�Þ eß r à 7: árâ
Sã uä må må aÜ rà iæ sç iæ nè g é Dê iæ sç cë uä ssç|ç iæ oì nè
148
similar to those of P. fluorescens than to those of B. cepacia. 16S-rDNA sequence analysis
confirmed the taxonomic status of the species included in the analysis. Collectively, these
results suggested that the PRN gene cluster may have been transferred from Pseudomonas to B.
pyrrocinia or vice versa. Although, gene transfer between different microbial genera and
between microbes and plants seem to be a very widespread phenomenon (Rivera et al., 1998;
Jain et al., 1999), sequence analysis of prn genes of multiple Burkholderia and Pseudomonas
strains will be required to further support this hypothesis.
In addition to specific antibiotic-biosynthetic genes, polymorphisms within the
response regulator gene gacA proved to be a powerful complementary genetic marker for the
genus Pseudomonas (CÚ hÛ
aÜ ptÝ�Þ eß r à 3í ). It is remarkable that a relatively small-sized gene like gacA has
such strong resolving capacity for phylogenetic purposes. Comparable results were obtained
with the 249-bp oprI gene, which allowed molecular taxonomic studies in pseudomonads
belonging to the rRNA group I, a group that contains the genuine Pseudomonas species (De
Vos et al., 1998). Polymorphisms within gacA homologs also allowed distinction of five
subclusters in the group of enteric bacteria, including Escherichia coli, Salmonella, Klebsiella
pneumoniae, Pectobacterium caratovorum, and Yersinia species. These results indicated that
gacA homologs also may provide a complementary genetic marker for phylogenetic studies of
Gram-negative bacteria other than Pseudomonas.
Aî
CÚ Tï
Ið Vñ Ið Tï Yò
The activity of two antibiotics, 2,4-DAPG and the surfactant produced by P.
fluorescens strain SS101, was investigated in more detail in CÚ hÛ
aÜ pÝ tÞ eß rà s ç 5 ó aÜ nè d ô
6õ . We found that
different infectious propagules of Pythium differ considerably in their sensitivity to 2,4-DAPG
(CÚ hÛ
aÜ ptÝ�Þ eß r à 5ó ). We observed variation in sensitivity to 2,4-DAPG among isolates of various
Pythium species (CÚ hÛ
aÜ ptÝ�Þ eß r à 5ó ) and also initial studies with the surfactants indicate variation in
sensitivity among oomycetes, in particular Pythium species (CÚ hÛ
aÜ pÝ tÞ eß r à 6õ ). Protective
mechanisms responsible for these differential responses are currently being studied in detail.
Variation in sensitivity within pathogen populations to metabolites produced by antagonistic
microorganisms is probably one of the major factors responsible for inconsistenty and, in some
cases lack of efficacy, of biological control (Mazzola et al., 1995). In addition to biotic factors,
we also showed that the activity of 2,4-DAPG against Pythium species is influenced by pH
(CÚ hÛ
aÜ ptÝ�Þ eß r à 5ó ). Although this conclusion is based on in vitro experiments, one may envisage the

Cö h÷
aø ptù�ú eû r ü 7: ýrþ
Sÿ u� m� m� aø rü i� s� i� n� g � D� i�s� c� u� ss��� i� o n�
149
impact of these findings on interactions between 2,4-DAPG producers and pathogenic fungi
in the plant rhizosphere. Despite the well-documented role of 2,4-DAPG in biocontrol of
several plant diseases (Vincent et al., 1991, Fenton et al., 1992, Keel et al., 1992), it appears
that this antibiotic has no determinative role in the competitiveness of 2,4-DAPG-producing
strains in the rhizosphere (Carrol et al., 1995, Cö h÷
aø pù tú eû r ü 4
). It is difficult to imagine that these
genes remained in numerous plant-associated Pseudomonas strains in the course of evolution
without an apparent advantage in rhizosphere competence. The role of 2,4-DAPG in
rhizosphere competence should be investigated in more detail with other plant species and
plant-growth stages. Also the role of surfactant production in rhizosphere competence will be
addressed in the near future.
R� E� F
E� R� E� N�
C�
E� S�
C�
a� rr��� o� ll���
, � H� ., ��� M�
o� ë�nn��� e� - L! o� cc"#" o� z$ , � Y
%., ��� D& o� w' l
�i(n� g) , � D& .� N� .,���
a� n� d *
O+ ’,G-
a� r� a� , � F
.� 1995. Mutational disruption of thebiosynthesis genes coding for the antifungalmetabolite 2,4-diacetylphloroglucinol does notinfluence the ecological fitness of Pseudomonasfluorescens F113 in the rhizosphere of sugarbeets.Appl. Environ. Microbiol. 61:3002-3007.C�
oo�.� k/, � R� .� J0 ., ��� a� n� d
*R� o� v1 i
(r� a� , � A
2.� D& .� 1976. The role of
bacteria in the biological control ofGaeumannomyces graminis by suppressive soils. Soil.Biol. Biochem. 8:269-273.D& e � B3 o� e� r� , � M
�., ��� V4
a� n � d*
e� r � S� l�u5 i(s6 , � I
7., ��� V4
a� n � L! oo�.� n� , � L! .� C� .,���a� n� d *
B3 a� kk/./
e� r� , � P8
.� A2 .� H� .� M� .� 1999. Combiningfluorescent Pseudomonas spp. strains to enhancesuppression of fusarium wilt of radish. Eur. J. PlantPathol. 105:201-210.D& e � V
4o� s6 , � D& ., ��� B3 o� u5 t9 o� n� , � C
�., ��� S� a� r� n� i
(g) u5 e� t9 , � A
2., ��� D& e � V
4o� s6 , � P
8.,���
V4
a� u5 t9 e� r� i( n� , � M�
., ��� a� n� d *
C�
o� r� n� e� l� i( s6 , � P8
.� 1998. Sequencediversity of the oprI gene, coding for major outermembrane lipoprotein I, among rRNA group Ipseudomonads. J. Bact. 180:6551-6556.D& u5 nn��� e� , � C
�., ��� M
�o� e� nn��� e� - L! o� cc":" o� z$ , � Y
%., ��� M
�c" C� a� r� t9 h; y< , � J
0.,���
H� i(gg)=) i(n� s6 , � P
8., ��� P
8o� w' e� ll��� , � J
0.� , � D& o� w' l
�i(n� g) , � D& .� N� ., ��� a� n� d
*O+ ’,G-
a� r� a� , � F
.� 1988. Combining proteolitic andphloroglucinol-producing bacteria for improvedbiocontrol of Pythium-mediated damping-off ofsugar beet. Plant Pathol. 47:299-307.F
e� n� t9 o� n� , � A2
.� M� ., ��� S� t9 e� p> h; e� n� s6 , � P8
.� M� .� , � C�
r� o� w' l�e� y< , � J
0.,���
O+ ’,C�
a� l� l� a� g) h; a� n� , � M�
., ��� a� n� d *
O+ ’,G-
a� r� a� , � F
.� 1992.Exploitation of gene(s) involved in 2,4-diacetylphloroglucinol biosynthesis to confer a newbiocontrol capability to a Pseudomonas strain. Appl.Environ. Microbiol. 58:3873-3878.H� a� n� d
*e� l� s6 m? a� n� , � J
0.� , � a� n� d
*S� t9 a� b@ b
@, � E� .� V4 .� 1996. Biocontrol
of soilborne plant pathogens. Plant Cell 8:1855-1869.
H� i(ll���
, � D& .� S� ., ��� S� t9 e� i( n� , � J0.� I7 ., ��� T
Ao� r� k/ e� w' i
(t9 z$ , � N
�.� R� ., ��� M
�o� r� s6 e� ,�
A2
.� M� ., ��� H� o� w' e� ll��� , � C�
.� R� ., ��� P8
a� c" h; l�a� t9 k/ o� , � J
0.� P8 ., ��� B3 e� c" k/ e� r� , � J
0.� O+ .,���
a� n� d *
L! i(g) o� n� , � J
0.� M� .� 1994. Cloning of genes involved in
the synthesis of pyrrolnitrin from Pseudomonasfluorescens and role of pyrrolnitrin synthesis inbiological control of plant disease. Appl. Environ.Microbiol. 60:78-85.H� o� w' e� ll��� , � C
�.� R� ., ��� a� n� d
*S� t9 i( p> a� n� o� v1 i
(c" , � R� .� D& .� 1979. Control
of Rhizoctonia solani on cotton seedlings withPseudomonas fluorescens and with an antibioticproduced by the bacterium. Phytopathol. 69:480-482.J0a� i( n� , � R� ., ��� R� i
(v1 e� r� a� , � M
�.� , � a� n� d
*L! a� k/ e� , � J
0.� A2 .� 1999.
Horizontal gene transfer among genomes: thecomplexity hypothesis. Proc. Natl. Acad. Sci. USA96:3801-3806.KB
ee�#� l� , � C�
., ��� S� c" h; n� i(d*
e� r� , � UC ., ��� M�
a� u5 r� h; o� fDe� r� , � M
�., ��� V4
o� i(s6 a� r� d* , � C
�.,���
L! a� v1 i(ll���
e� , � J0.� , � B3 u5 r� g) e� r� , � UC ., ��� W
Ei(r� t9 h; n� e� r� , � P
8., ��� H� aa�:� s6 , � D& ., ��� a� n� d
*D& éFfDa� g) o� , � G
-.� 1992. Suppression of root diseases by
Pseudomonas fluorescens CHA0: importance of thesecondary metabolite 2,4-diacetylphloroglu-cinol.Mol. Plant-Microbe Interact. 5:4-13.KB
ee�#� l� , � C�
., ��� WE
e� l� l� e� r� , � D& .� M� ., ��� N�
a� t9 s6 c" h; , � A2
., ��� D& éFfDa� g) o� , � G
-.,���
C�
oo�.� k/, � R� .� J0 ., ��� a� n� d
*TA
h; o� m? a� s6 h; o� w' , � L! .� S� .� 1996.Conservation of the 2,4-diacetylphloroglucinolbiosynthesis locus among fluorescent Pseudomonasstrains from diverse geographic locations. Appl.Environ. Microbiol. 62:552-563.L! e� v1 y< , � E� ., ��� G
-o� u5 g) h; , � F
.� J0 ., ��� B3 e� r� l� i( n� , � K
B.� D& ., ��� G
-u5 i(a� n� a� , � P
8.� WE .,���
a� n� d *
S� m? i(t9 h; , � T
A.� J0 .� 1992. Inhibition of Septoria tritici
and other phytopathogenic fungi and bacteria byPseudomonas fluorescens and its antibiotics. PlantPathol. 41:335-341.M�
a� u5 r� h; o� ffDGD
e� r� , � M�
., ��� KB
ee�:� l� , � C�
., ��� S� c" h; n� i(d*
e� r� , � UC ., ��� V4
o� i(s6 a� r� d* ,�
C�
., ��� H� aa�H� s6 , � D& ., ��� a� n� d *
D& éFfDa� g) o� , � G
-.� 1992. Influence of
enhanced antibiotic production in Pseudomonas

CI hJ
aK ptLNM eO r P 7: QSR
ST uU mV mV aK rP iW sX iW nY g Z D[ iW sX c\ uU ssX�X iW o] nY
150
fluorescens strain CHA0 on its disease suppressivecapacity. Phytopathol. 82:190-195.Ma_ v` ra ob dc id , e D
f.g Vh ., g e W
iej llk�k ej ra , e D
f.g M., g e a_ nl d c M^ a_ zzmnm ob l
ka_ , e M.,g e
Co oob.b kp, e Rq
.g Jr ., g e Ts
ht
ob mu a_ sv ht ob ww , e Lx .g Sy ., g e Wi
ej llk�k ej ra , e Df
.g M.g , e a_ nl dcPz id ej ra sv ob n l III
{�{|{, e Lx .g Sy .g 1992. Contribution of phenazine
antibiotic biosynthesis to the ecological competenceof fluorescent pseudomonads in soil habitats. Appl.Environ. Microbiol. 58:2616-2624.Ma_ v` ra ob dc id , e O
}.g Vh ., g e M^ c~ Sy p� a_ ddc�c ej nl -� G� a_ ra dc ej nl ej ra , e B� .g B� .,g e
Ma_ v` ra ob dc id , e Df
.g Vh ., g e B� ob nl sv a_ llk�k , e Rq
.g F� ., g e Wi
ej lk lk ej ra , e Df
.g M^ ., g e a_ nl dcTs
ob mu a_ sv ht ob ww , e Lx .g Sy .g 2001. Genetic diversity of phlDfrom 2,4-diacetylphloroglucinol-producingfluorescent Pseudomonas spp. Phytopathol. 91:35-43.Mc~ Sy p� a_ ddc�c ej nl -� G� a_ ra dc ej nl ej ra , e B� .g B� ., g e S
yc~ ht ra ob ej dc ej ra , e K
�.g Lx .,g e
K�
a_ llk�k ob g� ej ra , e Sy.g E� ., g e R
qaa_H_ id j� mu a_ kp ej ra sv , e J
r.g M^ .g , e T
sht
ob mu a_ sv ht ob ww ,eLx .g Sy ., g e a_ nl d c W
iej llk�k ej ra , e D
f.g M.g 2000. Genotypic and
phenotypic dyversity of phlD-containingPseudomonas strains isolated from the rhizosphereof wheat. Appl. Environ. Microbiol. 66:1939-1946.O}
'� Sy u� llk�k
id v` a_ nl , e Df
.g Jr ., g e a_ nl d c O}
'� G� a_ ra a_ , e F� .g 1992. Traits offluorescent Pseudomonas spp. involved insuppression of plant root pathogens. Microbiol.Rev. 56:662-676.Pz id c~ a_ ra dc , e Co ., g e D
fi d Co ej llk�k ob , e F� .g , e V
hej nl t� u� ra a_ , e M., g e F� a_ nl id , e R
q., g e a_ nl dc
G� u� c~ kp ej ra t� , e A�
.g 2000. Frequency and diversity of 2-4-diacetylphloroglucinol-producing bacteria isolatedfrom the maize rhizosphere at different stages ofgrowth. Appl. Environ. Microbiol. 66:948-955.Rq
aa_:_ id j� mu a_ kp ej ra sv , e Jr. g M^ .g , e Lx eej:j mu a_ nl , e M^ .g , e V
ha_ n l O
}ob ra sv c~ ht ob t� ,e
M.g M^ .g Pz ., g e Vh
a_ n l dc ej r a Sylku� id sv , e I
{., g e S
yc~ ht id pp��� ej ra sv , e B� ., g e B� a_ kk
p.pej ra ,e
Pz .g A� .g H� .g M^ .g 1995. Dose-response relationships inbiological control of Fusarium wilt of radish byPseudomonas spp. Phytopathol. 85:1075-1081.Rq
aa_:_ id j� mu a_ kp ej ra sv , e Jr.g M^ .g , e W
iej llk�k ej ra , e D
f.g M^ .g , e a_ nl d c T
sht
ob mu a_ sv ht ob ww ,eLx .g Sy .g 1997. Frequency of antibiotic producingPseudomonas spp. in natural environments. Appl.Environ. Microbiol. 63: 881-887.Rq
aa_:_ id j� mu a_ kp ej ra sv , e Jr.g M^ .g , e a_ nl d c W
iej lk lk ej ra , e D
f.g M.g 1998. Natural
plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils.Mol. Plant-Microbe Interact. 11:144-152.Rq
a_ mu ej tt��� ej , e A�
., g e Mob ë� nnl�l ej -� Lx ob c~ c~ ob zm , e Y�
., g e a_ nl d c Df
é� f� a_ g� ob , e G� .g2001. Polymorphism of the polyketide synthasegene phlD in biocontrol fluorescent Pseudomonasproducing 2,4-diacetylphloroglucinol andcomparison of PhlD with plant polyketidesynthases. Mol. Plant-Microbe Interact. 14:639-652.
Rq
id v` ej ra a_ , e M^ .g Co ., g e Jra_ id nl , e R
q., g e M^ oob.b ra ej , e J
r.g E� ., g e a_ nl d c Lx a_ kp ej , e J
r.g
1998. Genomic evidence for two functionallydistict gene classes. Proc. Natl. Acad. Sci. USA95:6239-6244.Syht
a_ ra id f� id -� Ts ej ht ra a_ nl id , e A�
.g , e Z� a_ lk a_ , e M^ ., g e N�
a_ t� sv c~ ht , e A�
.g , e Mob ë� nnl�l ej -�Lx ob c~ c~ ob zm , e Y
�., g e a_ nl d c D
fé� f� a_ g� ob , e G� .g 1998. Biocontrol of
soil-borne fungal diseases by 2,4-diacetylphloroglucinol-producing pseudomonadswith different restriction profiles od amplified 16SrDNA. Europ. J. Plant Patol. 104:631-643.Syt� u� t� zm , e E� ., g e D
fé� f� a_ g� ob , e G� ., g e a_ nl d c K
�ej ra nl , e H� .g 1986.
Naturally occuring fluorescent pseudomonadinvolved in suppression of black root rot of tobacco.Phytopathol. 76:181-185.Ts
ht
ob mu a_ sv ht ob ww , e Lx .g Sy ., g e a_ nl d c Wi
ej llk�k ej ra , e Df
.g M.g 1988. Role ofphenazine antibiotic from Pseudomonas fluorescensin biological control of Gaeumannomyces graminisvar. tritici. J. Bacteriol. 170:3499-3508.Ts
ht
ra a_ nl ej , e Co .g , e O}
lkssv�v ob nl , e S
y., g e N
�id ej lk sv ej nl , e T
s.g H� ., g e a_ nl dc
Syø� ra ej nl sv ej nl , e J
r.g 1999. Vital fluorescent stains for
detection of stress in Pythium ultimum andRhizoctonia solani challenged with viscosinamidefrom Pseudomonas fluorescens DR54. FEMSMicrobiol. Ecol. 30:11-23.Vh
a_ n l Lx oob.b nl , e Lx .g Co ., g e B� a_ kkp.p
ej ra , e Pz .g A� .g H� .g M^ ., g e a_ nl d c Pz id ej t� ej ra sv ej ,eCo .g M^ .g Jr .g 1998. Systemic resistance induced byrhizosphere bacteria. Ann. Rev. Phytopathol.36:453-483.Vh
id nl c~ ej nl t� , e M^ . g N�
., g e H� a_ rra�a id sv ob nl , e Lx . g A�
., g e B� ra a_ c~ kp id nl , e Jr. g M^ .,g e
K�
ob v` a_ c~ ej v` id c~ ht , e Pz . g A�
., g e M^ u� kpej ra j� id , e Pz ., g e W
iej llk�k ej ra , e D
f.g M., g e a_ nl dc
Pz id ej ra sv ob nl , e E� .g A� .g 1991. Genetic analysis of theantifungal activity of a soilborne Pseudomonasaureofacens strain. Appl. Environ. Microbiol.57:2928-2934.Wi
a_ nl g� , e Co ., g e Rq
a_ mu ej tt��� ej , e A�
., g e Pz u� nl j�a_ sv a_ mu a_ ra nl ww ob nl g� , e Pz .,g e
Z� a_ lk a_ , e M^ .g , e N�
a_ t� sv c~ ht , e A�
., g e M^ ob ë� nnl�l ej -� Lx ob cc~#~ ob zm , e Y�
., g e a_ nl dcDf
é� f� a_ g� ob , e G� .g 2001. Cosmopolitan distribution ofphlD-containing dicotyledonous crop-associatedbiocontrol pseudomonads of worldwide origin.FEMS Microbiol. Ecol. 37:105-116.Wi
ej llk�k ej ra , e Df
.g M^ .g 1988. Biological control of soilborneplant pathogens in the rhizosphere with bacteria.Annu. Rev. Phytopathol. 26:379-407.Wi
ht
id pp��� sv , e Jr.g M.g 1997. Developments in the biological
control of soil-borne plant pathogens. Adv. Botan.Res. 26��� :1-134.Wi
ht
id pp��� sv , e Jr.g M^ .g 2001. Microbial interactions and
biocontrol in the rhizosphere. J. Exp. Bot. 52: 487-511.

S� U�
MM���
A�
R Y¡
Bacteria of the genus Pseudomonas are potential biocontrol agents of plant diseases
caused by various fungi and oomycetes. Antibiotic production is an important trait responsible
for the activity of several Pseudomonas strains against plant pathogens. Despite the amount of
information obtained during the past decades on biosynthesis and regulation of antibiotics,
little is known about the distribution and diversity of antibiotic-producing Pseudomonas spp.
in natural environments. Knowledge about the diversity of naturally occurring populations of
antibiotic-producing Pseudomonas spp. could contribute to improve biological control by the
identification of new strains that are ecologically more competent. The main objectives in this
thesis were to study the distribution, diversity, and activity of antibiotic-producing
Pseudomonas spp. in rhizosphere environments.
In C¢ h£
a¤ pt¥N¦ e§ r ¨ 1, ©�ª an overview is given on detection, distribution and diversity of
antibiotic-producing Pseudomonas spp. Special attention is paid to the biosynthesis and
regulation of the antibiotics 2,4-diacetylphloroglucinol (2,4-DAPG), phenazines (PHZ),
pyrrolnitrin (PRN), pyoluteorin (PLT), and biosurfactant antibiotics, which are subject of the
experiments described in this thesis.
In C¢ h£
a¤ pt¥N¦ e§ r ¨ 2,« ª the diversity, phylogenetic relationships, and frequency of Pseudomonas
and Burkholderia species able to produce the antibiotics PRN or PLT were studied. Primers
were developed from conserved sequences within the biosynthetic loci for each of the two
antibiotics and allowed detection of multiple Pseudomonas and Burkholderia spp. that either
produce PRN or PLT or both. Subsequent RFLP (Restriction Fragment Length
Polymorphisms) analysis of the amplified pltC fragment showed no polymorphisms among
PLT-producing Pseudomonas strains. Polymorphisms within the amplified prnD fragment,
however, allowed the assessment of the diversity among PRN-producing Pseudomonas and
Burkholderia spp. to a level similar to that obtained by RAPD (Random Amplified
Polymorphic DNA) analysis. Phylogenetic analyses further revealed that the prn genes of B.
pyrrocinia DSM10685 were more closely related to those of PRN-producing Pseudomonas
strains, suggesting that lateral gene transfer may have occurred. PRN- and PLT-producing
Pseudomonas and Burkholderia spp. were not detected on roots of wheat grown in five
agricultural soils collected in the Netherlands, two of them being naturally suppressive to the
take-all pathogen Gaeumannomyces gramminis var. tritici. These results suggested that these
bacteria do not contribute to the natural suppressiveness found in Dutch take-all suppressive
soils.

S¬ u mm®¯® a° r± y²
152
C³ h´
a° ptµN¶ e· r ± 3 focused on the response regulator gene gacA in Pseudomonas species. This
gene influences the production of several antibiotics in antagonistic Pseudomonas spp. We
developed primers and a probe for the gacA gene of Pseudomonas species and sequenced gacA
from ten Pseudomonas strains isolated from different plant-associated environments. PCR
analysis and Southern hybridization showed that gacA is highly conserved within the genus
Pseudomonas and indicated that gacA can be used as a complementary genetic marker for
detection of Pseudomonas spp. in environmental samples. Phylogenetic relationships inferred
from the newly sequenced gacA genes and gacA homologs present in the databases, showed
that gacA may also serve as a marker for phylogenetic studies of Pseudomonas spp. and Gram-
negative bacteria other than Pseudomonas. Estimations of nonsynonymous to synonymous
substitution rates (Ka/Ks ratios) showed that purifying selection is acting on gacA, indicating
that there is a selective pressure to avoid substitutions leading to functional changes in the
GacA protein.
C³ h´
a° ptµN¶ e· r ± 4¹
focused on the role of antibiotic-producing Pseudomonas spp. in Dutch take-
all suppressive soils. Natural suppressiveness of soils to take-all disease of wheat, referred to as
take-all decline (TAD), occurs worldwide. It has been postulated that different microbial genera
and mechanisms are responsible for TAD in soils from different geographical regions. Based on
population density studies and the use of antibiotic-deficient mutants, we demonstrated that
fluorescent Pseudomonas spp. that produce 2,4-DAPG play a key role in the natural
suppressiveness of two Dutch TAD soils. Our results showed that in addition to the
physicochemically different TAD soils from Washington State (USA), 2,4-DAPG-producing
fluorescent Pseudomonas spp. are also a key component of the natural suppressiveness found in
Dutch TAD soils. Furthermore, it is the first time since the description of Dutch TAD soils,
that at least part of the mechanisms and microorganisms operating in their suppressiveness are
identified. In spite of similarities in population densitis and activity, 2,4-DAPG-producing
Pseudomonas spp. found in the Dutch TAD soils are genotypically different from those found
in TAD soils from Washington State.
In C³ h´
a° ptµN¶ e· r ± 5º we concentrated on the activity of the antibiotic 2,4-DAPG against
Pythium species. The results of this study showed that within and among Pythium species there
is variation in sensitivity to 2,4-DAPG. Also various propagules of P. ultimum var.
sporangiiferum (Pus), that are part of the asexual stage of the life cycle, differ considerably in
their sensitivity to 2,4-DAPG. Mycelium was the most resistant structure, followed by
zoosporangia, zoospore cysts and zoospores. Transmission electron microscopy (TEM) studies
revealed that 2,4-DAPG causes different stages of disorganisation in hyphal tips of Pus,

S» u¼ mm½¯½ a¾ r¿ yÀ
153
including alteration (proliferation, retraction and disruption) of the plasma membrane,
vacuolisation and cell content disintegration. Additionally, we report, for the first time, that
pH has a significant effect on the activity of 2,4-DAPG, with a higher activity at low pH.
Furthermore, the level of acetylation of phloroglucinols also is a major determinant of their
activity.
CÁ hÂ
a¾ ptÃNÄ eÅ r ¿ 6Æ focused on surface-active antibiotics produced by Pseudomonas spp. and
their activity against oomycete pathogens. We report the discovery of Pseudomonas isolates that
produce biosurfactants with lytic activity against zoospores of multiple oomycete pathogens.
Application of cell suspensions of strain P. fluorescens SS101 to soil or hyacinth bulbs provided
protection against root rot caused by Pythium intermedium. Five mutants of SS101, obtained
by random Tn5 mutagenesis, lacked the ability to cause lysis of zoospores. Subsequent genetic
characterization of the Tn5-flanking regions of two surfactant-deficient mutants, 10.24 and
17.18, showed that the transposons had integrated in condensation domains of peptide
synthetases. When applied to soil, mutant 10.24 was not able to protect hyacinth against
Pythium root rot, whereas wildtype strain SS101 gave significant control. Reverse-phase high
performance liquid chromatography (RP-HPLC) and liquid chromatography–mass
spectrometry (LC-MS) yielded eight different fractions of which five had surface activity and
caused cessation of motility and lysis of zoospores. Collectively, the results indicate that
biosurfactants produced by P. fluorescens SS101 play an important role in interactions with
zoosporic pathogens and in control of Pythium root rot of hyacinth.
In CÁ hÂ
a¾ ptÃÇÄ eÅ r ¿ 7È
the most important findings are summarised and discussed in the context
of the existing knowledge.
In conclusion, the results presented in this thesis provide information on the
distribution, diversity and activity of antibiotic-producing Pseudomonas spp. and describe the
discovery of surface-active antibiotics that have great potential for controlling plant diseases
caused by oomycete pathogens.

SÉ uÊ mmË¯Ë aÌ rÍ yÎ
154

RÏ EÐ
SÑ UÒ
MÓ
OÔ
Bacterias do gênero Pseudomonas são agentes potenciais de controle biológico de
doenças de plantas causadas por vários fungos e oomicetos. A produção de antibióticos é um
importante atributo responsável pela atividade antagônica de diversos isolados de Pseudomonas
contra patógenos de plantas. Apesar da quantidade de informação obtida durante as últimas
décadas sobre a síntese e regulação de antibióticos, pouco se sabe sobre a distribuição e
diversidade de bactérias produtoras de antibióticos do gênero Pseudomonas spp. na natureza.
Dados sobre a diversidade de populações indígenas de bactérias produtoras de antibióticos do
gênero Pseudomonas spp. pode contribuir para melhorar o controle biológico pela identificação
de novos isolados que sejam ecologicamente mais adaptados. Os principais objetivos nesta tese
foram estudar a distribuição, diversidade e atividade de bactérias do gênero Pseudomonas spp.
produtoras de antibióticos.
No CÕ aÖ pít×ÙØ Ú uÛ lÜo Ý 1, Þ�ß apresentamos uma revisão sobre detecção, distribuição e diversidade de
bactérias do gênero Pseudomonas produtoras de antibióticos. Atenção especial é dada à síntese e
regulação dos antibióticos 2,4-diacetilfloroglucinol (2,4-DAPG), fenazinas (PHZ),
pirrolnitrina (PRN), pioluteorina (PLT), e antibióticos biosurfactantes, que em conjunto são o
tema central dos experimentos descritos nesta tese.
No CÕ aÖ pít×àØ Ú uÛ lÜo Ý 2,
á ß estudou-se a diversidade, relações filogenéticas e a frequência das
bactérias dos gêneros Pseudomonas e Burkholderia capazes de produzir os antibióticos PRN ou
PLT. Primers foram desenvolvidos de sequências conservadas dentro das regiões que codificam
a síntese desses antibióticos e permitiram a detecção de múltiplos isolados dos gêneros
Pseudomonas e Burkholderia spp. que produzem PRN, PLT ou ambos. Análises de RFLP
(Restriction Fragment Length Polymorphisms) de fragmentos amplificados do gene pltC não
mostraram nenhum polimorfismo entre isolados de Pseudomonas spp. capazes de produzir
PLT. Por outro lado, os polimorfismos dentro do fragmento amplificado do gene prnD
permitiram a avaliação da diversidade entre Pseudomonas e Burkholderia spp. capazes de
produzir PRN em um nível semelhante ao obtido por análise de RAPD (Random Amplified
Polymorphic DNA). Análises filogenéticas adicionalmente revelaram que os genes prn de B.
pyrrocinia DSM10685 são mais relacionados aqueles provenientes de espécies de Pseudomonas,
sugerindo uma possível transferência lateral desses genes. Pseudomonas e Burkholderia spp.
produtoras de PRN e PLT não foram detectadas em raízes de trigo cultivado em cinco solos
coletados na Holanda, sendo dois deles naturalmente supressivos ao patógeno do mal-do-pé do
trigo, Gaeumannomyces gramminis var. tritici. Esses resultados sugeriram que essas bactérias

Râ eã sä uå mæ oç
156
não contribuem para a supressividade natural que ocorre nos solos Holandeses ao mal-do-pé
do trigo.
No Cè aé pítêàë ì uå lío ç 3,îðï estudou-se o gene regulador de respostas gacA em espécies de
Pseudomonas. Este gene influencia a produção de antibióticos em bactérias antagonistas do
gênero Pseudomonas. Primers e uma sonda para o gene gacA de espécies de Pseudomonas foram
desenvolvidos. Sequenciou-se o gene gacA de dez espécies de Pseudomonas isoladas de
diferentes combinações planta hospedeira-ambiente. Análise de PCR e Southern hybridization
revelaram que o gene gacA é altamente conservado dentro do gênero Pseudomonas e indicou
que gacA pode ser utilizado como um marcador complementar para o isolamento de
Pseudomonas spp. de amostras ambientais. Relações filogenéticas inferidas das novas
sequências do gene gacA e homólogos desse gene presentes nos bancos de dados genéticos
mostraram que gacA pode também servir como um marcador para estudos filogenéticos não só
de Pseudomonas spp. mas também de outras bactérias Gram-negativas. Estimações das taxas de
substituição não-sinônima para sinônima de nucleotídeos (taxas Ka/Ks) revelaram que uma
seleção purificadora está agindo sobre o gene gacA, indicando que existem pressões de seleção
para evitar substituições de nucleotídeos que levem a mudanças na conformação estrutural e
subsequente perda de função da proteína GacA.
Nñ
o Cç è aé pítêàë ì uå lío ç 4,ò ï estudou-se o papel de Pseudomonas spp. produtoras de antibióticos em
solos Holandeses supressivos ao mal-do-pé do trigo. Solos naturalmente suppressivos ao mal-
do-pé do trigo, referidos como solos com declínio do mal-do-pé (DMP), ocorrem em todo o
mundo. Postula-se que diferentes gêneros microbianos e mecanismos são responsáveis pelo
DMP em solos de diferentes regiões. Baseado em estudos de densidade populacional e uso de
mutantes deficientes na produção de antibióticos, demonstrou-se que Pseudomonas
fluorescentes capazes de produzir o antibiótico 2,4-DAPG desempenham um papel chave na
supressividade natural de dois solos Holandeses ao mal-do-pé do trigo. Nossos resultados
mostraram que apesar dos solos do Estado de Washington (EUA) serem físico-quimicamente
diferentes dos solos Holandeses, Pseudomonas produtoras do antibiótico 2,4-DAPG também são
componentes chaves para a supressividade natural dos solos Holandeses ao mal-do-pé do trigo.
Alem disso, essa é a primeira vez, desde a descrição dos solos Holandeses com DMP, que pelo
menos parte dos mecanismos e microorganismos envolvidos na supressividade são
identificados. Apesar das similaridades em termos de atividade e densidade populacional, as
bacterias do gênero Pseudomonas produtoras de 2,4-DAPG encontradas nos solos Holandeses
com DMP são geneticamente diferentes daquelas encontradas em solos com DMP do Estado
de Washington.

Ró eô sõ uö m÷ oø
157
No Cù aú pítûàü ý uö lþo ø 5,ÿ�� enfocou-se a atividade do antibiótico 2,4-DAPG contra espécies de
Pythium. Os resultados desse estudo mostraram que existe variação na sensitividade ao
antibiótico 2,4-DAPG dentro e entre espécies de Pythium. Vários propágulos de P. ultimum
var. sporangiiferum (Pus), que são parte do ciclo assexual, diferiram consideravelmente quanto a
sua sensitividade a 2,4-DAPG. Micélio foi a estrutura mais resistente, seguida de
zoosporângios, cistos de zoosporos e zoosporos. Estudos de microscopia eletrônica de
transmissão (MET) revelaram que 2,4-DAPG causa diferentes estágios de desorganização em
pontas de hifas de Pus, incluindo alterações (proliferação, retração e ruptura) da membrana
plasmática, vacuolisação e desintegração do conteúdo celular. Adicionalmente, registra-se, pela
primeira vez, que o pH tem um efeito significativo sobre a atividade de 2,4-DAPG, com maior
atividade em pHs mais baixos. Além disso, o número de grupos acetil presentes nas moléculas
dos floroglucinois é também um importante determinante de sua atividade.
N�
o Cø ù aú pítûàü ý uö lþo ø 6,��� concentrou-se no estudo de antibióticos produzidos por Pseudomonas
spp. com atividade de superfície e sua ação contra oomicetos patogênicos. Encontraram-se
isolados de Pseudomonas capazes de produzir biosurfactantes com atividade lítica contra
zoosporos de vários oomicetos patogênicos. A aplicação de suspensões de células do isolado
SS101 de P. fluorescens no solo ou em bulbos de jacinto (Hyacinthus orientalis) resultaram na
proteção dessas plantas contra podridão radicular causada por Pythium intermedium. Cinco
mutantes do isolado SS101, obtidos através de integração ao acaso do transposon Tn5, não
apresentaram habilidade de causar a explosão de zoosporos. Subsequente caracterização
genética das regiões vizinhas ao Tn5 de dois mutantes, 10.24 e 17.18, mostraram que os
transposons estavam inseridos em sequências codificadoras de domínios de condensação de
enzimas responsáveis pela síntese de peptídeos. Quando aplicado em solo, o mutante 10.24
não foi capaz de proteger plantas de jacinto contra podridão radicular causada por P.
intermedium, enquanto que o isolado tipo selvagem SS101 proporcionou um controle
significativo. Cromatografia líquida de alta pressão e fase reversa (RP-HPLC) e cromatografia
líquida conjugada com espectrometria de massa (LC-MS) permitiram a separação de oito
frações diferentes do surfactante, das quais cinco apresentaram capacidade de diminuir a
densidade superficial da água, causar a perda de motilidade e explosão de zoosporos.
Coletivamente, estes resultados indicam que os biosurfactantes produzidos por P. fluorescens
SS101 exercem um papel fundamental nas interações com patógenos capazes de produzir
zoosporos e no controle da podridão radicular do jacinto causada por P. intermedium.
No Cù aú pítûÙü ý uö lþo ø 7,� � os resultados mais importantes foram resumidos e discutidos no
contexto do conhecimento existente.

R� e� s� u� m o
158
Concluindo, os resultados apresentados nesta tese oferecem informações sobre a
distribuição, diversidade e atividade de bactérias do gênero Pseudomonas produtoras de
antibióticos e descrevem a descoberta de antibióticos com atividade de superfície que têm um
grande potencial para o controle de oomicetos patogênicos.

S� A�
M
E�
N�
V�
A�
TT���
I� N� G�
Bacteriën van het genus Pseudomonas zijn potentiële antagonisten van diverse
plantenpathogene schimmels en oömyceten. De productie van antibiotica speelt een
belangrijke rol in de activiteit van diverse Pseudomonas stammen tegen plantenpathogenen.
Ondanks het feit dat er gedurende de laatste decennia veel informatie is verkregen over de
biosynthese en regulatie van antibiotica, is er nog relatief weinig bekend over de verspreiding
en diversiteit van antibiotica-producerende Pseudomonas spp. in natuurlijke milieus. Kennis
over de diversiteit van natuurlijke populaties van antibiotica-producerende Pseudomonas spp.
kan bijdragen aan een verbetering van biologische bestrijding middels de identificatie van
stammen die ecologisch meer competent zijn. De doelstelling van het onderzoek beschreven in
dit proefschrift is het bestuderen van de verspreiding, diversiteit en activiteit van antibiotica-
producerende Pseudomonas spp. in de rhizosfeer.
In H� o� o� f�d�
s� t� uk ��� 1 � wordt een overzicht gegeven van detectie, verspreiding en diversiteit
van antibiotica-producerende Pseudomonas spp. Er wordt specifiek ingegaan op de biosynthese
en regulatie van de antibiotica 2,4-diacetylphloroglucinol (2,4-DAPG), phenazines (PHZ),
pyrrolnitrin (PRN), pyoluteorin (PLT), en oppervlakte-actieve antibiotica, welke onderwerp
zijn van de experimenten beschreven in dit proefschrift.
In H� o� o� f�d�
s� t� uk ��� 2
is de diversiteit, phylogenie en frequentie van Pseudomonas en
Burkholderia spp. die PRN of PLT produceren bestudeerd. Primers werden ontwikkeld op
basis van geconserveerde sequenties in de biosynthetische loci van elk van beide antibiotica.
Deze primers maakten detectie mogelijk van een grote collectie van Pseudomonas en
Burkholderia spp. die PRN of PLT of beide antibiotica produceren. Met behulp van RFLP
(Restriction Fragment Length Polymorphisms) analyse van het geamplificeerde pltC fragment
konden geen polymorfismen worden aangetoond tussen PLT-producerende Pseudomonas
stammen. De polymorfismen in het geamplificeerde prnD fragment maken het echter wel
mogelijk om de diversiteit tussen PRN-producerende Pseudomonas en Burkholderia spp. te
bepalen zelfs tot een vergelijkbaar niveau als verkregen met behulp van RAPD (Random
Amplified Polymorphic DNA) analyse. Tevens tonen de phylogenetische analyses aan dat de
prn genen van B. pyrrocinia DSM10685 meer verwant zijn met die van PRN-producerende
Pseudomonas stammen dan met die van andere Burkholderia stammen, hetgeen mogelijk kan
wijzen op uitwisseling van prn genen. PRN- en PLT-producerende Pseudomonas en
Burkholderia spp. waren niet detecteerbaar op wortels van tarwe geteeld op vijf
landbouwgronden verzameld in Nederland, waarvan twee gronden ziektewerend zijn tegen het

S! a" m# e$ n% v& a" tt'(' i) n% g*
160
pathogeen Gaeumannomyces gramminis var. tritici. Deze resultaten suggereren dat deze
bacteriën waarschijnlijk niet bijdragen aan de natuurlijke ziektewering van deze twee gronden.
Hoo+-,., f/d0
s1 t' uk 2�3 3 4 richt zich op het respons regulator gen gacA in Pseudomonas spp. Dit gen
beïnvloedt de productie van verscheidene antibiotica in antagonistische Pseudomonas spp.
Voor het gacA gen van Pseudomonas spp. werden primers en een probe ontwikkeld en van tien
Pseudomonas stammen, geïsoleerd van verschillende planten, werd vervolgens het gacA gen
geamplificeerd en gesequenced. De PCR analyses en Southern hybridisatie tonen aan dat gacA
in hoge mate geconserveerd is in Pseudomonas en als complementaire genetische marker
gebruikt kan worden voor detectie van Pseudomonas in rhizosfeer monsters. Phylogenetische
verwantschappen, bepaald op basis van de nieuwe gacA sequenties en gacA homologen
aanwezig in de database, toonden aan dat het gacA gen tevens dienst kan doen als
phylogenetische marker voor Pseudomonas spp. en andere Gram-negatieve bacteriën.
Zogenaamde Ka/Ks ratios tonen aan dat het gacA gen onder selective druk staat waardoor
voorkomen wordt dat substituties leiden tot functionele veranderingen in GacA.
In H+ o, o, f/d0
s1 t' uk 2�3 45
werd de rol van antibiotic-producerende Pseudomonas spp. in de
ziektewerendheid van Nederlandse gronden tegen G. gramminis var. tritici bestudeerd.
Ziektewering van bodems tegen G. gramminis var. tritici komt wereldwijd voor en algemeen
wordt aangenomen dat verschillende microbiële genera verantwoordelijk zijn voor de
ziektewering in bodems van diverse geografische oorsprong. Met behulp van populatiestudies,
biotoetsen en het gebruik van antibiotica-deficiënte mutanten werd aangetoond dat
Pseudomonas spp. die 2,4-DAPG produceren een sleutelrol spelen in de ziektewering van twee
Nederlandse gronden. De resultaten laten zien dat ook in bodems die fysisch-chemisch
verschillend zijn van de ziektewerende bodems van Washington State (USA), 2,4-DAPG-
producing fluorescent Pseudomonas spp. een sleutelrol spelen. Het is tevens de eerste keer sinds
de beschrijving van deze Nederlandse ziektewerende gronden dat tenminste een deel van de
mechanismen en microorganismen die een rol spelen in de ziektewering zijn geïdentificeerd.
Ondanks sterke overeenkomsten in populatiedichtheden en activiteit, zijn de populaties van
2,4-DAPG-producerende Pseudomonas spp. in de Nederlandse ziektewerende gronden
genotypisch verschillend van de populaties gevonden in de ziektewerende gronden van
Washington State.
De activiteit van het antibioticum 2,4-DAPG tegen Pythium spp. is in detail
bestudeerd in H+ oo,6, f/d0
s1 t' uk 2�3 57 . De resultaten laten zien dat tussen Pythium spp. variatie bestaat in
gevoeligheid voor 2,4-DAPG. Ook diverse structuren van P. ultimum var. sporangiiferum
(Pus), die deel uitmaken van de asexuale fase van de levenscyclus, verschillen aanzienlijk in hun

S8 a9 m: e; n< v= a9 tt>(> i? n< g@
161
gevoeligheid voor 2,4-DAPG. Mycelium was de meest resistente structuur gevolgd door
zoösporangia, geëncysteerde zoösporen en tot slot zoösporen. Transmissie electronen
microscopie (TEM) toonde aan dat 2,4-DAPG verschillende stadia van desorganisatie
veroorzaakt in hyphen van Pus, inclusief veranderingen (terugtrekking en disruptie) van de
plasmamembraan, vacuolisatie en desintegratie van de inhoud van de cel. Daarnaast, wordt in
dit proefschrift voor de eerste keer beschreven dat de pH een significant effect heeft op de
activiteit van 2,4-DAPG, met een hogere activiteit bij lage pH. Tevens bleek dat de mate van
acetylering van phloroglucinol bepalend is voor de activiteit.
HooA-B.B fCdD
sE t> uk F�G 6H richt zich op oppervlakte-actieve antibiotica geproduceerd door
Pseudomonas spp. en hun activiteit tegen oömyceten. We beschrijven de ontdekking van
Pseudomonas isolaten die biosurfactants produceren met lytische activiteit tegen zoösporen van
meerdere oömycete pathogenen. Toediening van celsuspensies van stam P. fluorescens SS101
aan grond en hyacintenbollen resulteerde in bescherming tegen wortelrot veroorzaakt door
Pythium intermedium. Vijf mutanten van SS101, verkregen met behulp van Tn5 mutagenese,
hebben het vermogen om zoösporen te lyseren verloren. Genetische karakterisatie van de Tn5-
flankerende regios van twee mutanten, 10.24 en 17.18, toonden aan dat de transposons
geïntegreerd waren in condensatie domeinen van peptide synthases. Mutant 10.24 was niet in
staat om hyacint te beschermen tegen Pythium wortelrot terwijl wildtype SS101 aanzienlijke
bescherming bood. Met behulp van reverse-phase high performance liquid chromatography
(RP-HPLC) en liquid chromatography–massa spectrometrie (LC-MS) werden acht fracties
geïsoleerd waarvan vijf oppervlakte-actief waren en lysis van zoosporen veroorzaakten.
Gezamenlijk geven deze resultaten sterke aanwijzingen dat oppervlakte-actieve metabolieten
geproduceerd door P. fluorescens SS101 een belangrijke rol spelen in interacties met zoösporen
van oömyceten en in bestrijding van Pythium wortelrot van hyacint.
In HA oB oB fCdD
sE t> uk F�G 7 I
worden de meest belangrijke resultaten beschreven in dit proefschrift
samengevat en bediscussieerd in de context van bestaande kennis.
Samengevat bevat dit proefschrift nieuwe informatie over de verspreiding, diversiteit
en activiteit van antibiotica-producerende Pseudomonas spp. en beschrijft het de ontdekking
van oppervlakte-actieve antibiotica die van groot belang kunnen zijn voor de bestrijding van
plantenziekten veroorzaakt door oömyceten.

SJ aK mL eM nN vO aK ttP(P iQ nN gR
162

AS
CT KU NV
OW WX
LY EZ
D[ G\ EZ
M]
EZ
NV
T^
S_
I would like to thank CAPES/Brazil (Coordenação de Aperfeiçoamento de Pessoal de
Nível Superior) for the opportunity to come to Wageningen through finantial support.
I am grateful to profs. José da Cruz Machado, Vicente Campos, and Ricardo Magela
from Lavras University for their help and stimulant encouragement in the first and embrionary
stage of this work.
I want to express my gratitude to Prof. Mike Jeger, Aad Thermorshuizen (Biob), M.
Gerlagh (PRI), Ruud van den Bulk (PRI), and Cees Langerak (PRI) for their contribution and
discussions during the second phase of this thesis (the old Sclerotinia project).
I am deeply grateful to Jos Raaijmakers for the opportunity to work in his group, his
guidance, support, and trust. His friendship, creativity, enthusiasm, and optimism (I still think
you are on the top of the list !) were important determinants for the success of this thesis.
Thanks to all people from the Phytopathology department for the nice moments,
collaborations, and sometimes, coffee breaks. Thanks to Willem, Corrie, Mieke, John, Rob,
and Hans for their help in countless occasions. To my promoter Pierre de Wit for his interest
in my work. Thanks to my students Flora, Dennis, Babette, Ana, and George. My roommate
Maria Vlami for her company and nice scientific and other discussions. All other people who
passed in the biological control group, your company and inputs were always appreciated.
Henk-Jan proved to be a excelent cook and also an expert in computers, thank you ! My
already gone friends prof. Dr. Holb and Lucas for their enjoyable company and late hours we
spent in the old building. To my great friend Ioannis for the nice conversations all these years,
you are a very special friend !!!
I appreciated very much the patience of Mieke, Corrie, Grardy, Ali, Ria, Ester, Ciska,
Lutte-Harm, Sander, and Hans for the Dutch lessons. I really learned a lot with you !!!
I want to thank all my Brazilian and international friends and colleagues living in
Wageningen for such wonderful moments togheter. Renatão, I was very fortunate to find a
real friend when I arrived (how cheap things were in the beginning !). Thanks to Arne
(lifizinho), Irene, Denise, Isabela, Eduardo, Pedrão, Mari, Mevrow de Alcantára, Luis, Gilma,
Sandra, Gustavo, Passarinho, Simone, Flavia, Rômulo, Francisco, Joana, Cristiane, Chalfun,
Milza, Manu, Ana, Gerard, Olavo, Marineide, Baro, Suzane, Sara, Raquel, Cristine, Celso,
Wellington, and others …. Thank you all for the nice evenings, for the parties, dinners, beers,
squash games, trips, and other enjoyable activities.

A`
ca kb nc od we lfeg dh gei�g mj eg nc tk sl
164
Thanks to all my friends in Brazil. Leimi, Joãozinho, Arnaldo, Mr. Kléber, Lulu,
Adolfo, Zé Roberto, Boizão, Claudinha, Alan, Roberto, and others … you provided an
important support, thanks a lot !!
I thank the long-distance support of my family in Brazil. My warmest thanks to my
beloved Flora for her comprehension, love, and support…. obrigado, minha flor !!!
Jorge

Am
BOnpo Uq
T Trsr
Ht E u
Am
Uq
Tr
Ht ORowv
Jorge Teodoro de Souza was born on 11th November 1970, in Rio Verde de Mato
Grosso, Mato Grosso do Sul State (MS), Brazil. He obtained his BSc degree in Agronomy in
January 1995 and his MSc degree in 1998 on biological control of nematodes, both at Lavras
University, Minas Gerais State (MG), Brazil. In January 1998 he started his Ph.D. studies at
the laboratory of Phytopathology of the Wageningen University, The Netherlands, on
biological control of soil-borne pathogens with antibiotic-producing Pseudomonas spp.




















