
NOTES Structural andFunctional Analyses ofMutantFurProteins with Impaired Regulatory Function
7
Vol. 176, No. 16 NOTES Structural and Functional Analyses of Mutant Fur Proteins with Impaired Regulatory Function ANNE M. WERTHEIMER, MARCELO E. TOLMASKY, LUIS A. ACTIS, AND JORGE H. CROSA* Department of Molecular Microbiology and Immunology, School of Medicine, Oregon Health Sciences University, Portland, Oregon 97201-3098 Received 23 March 1994/Accepted 8 June 1994 Vibrio anguillarum Fur mutants, 775met9 and 775metll, were characterized. V. anguillarum 775met9 had a change of D to G at position 104 located in the carboxy terminus resulting in impaired Fur activity. Computer analysis predicts perturbation of an alpha-helix in the carboxy terminus which may interfere with Fur protein conformation. Strain 775metll had a change in the start codon resulting in no protein synthesis. The mutants are unstable, and reversion to the wild type occurs frequently. An important virulence factor of the fish pathogen Vibno anguillarum is its plasmid-mediated iron uptake system, com- posed of the siderophore anguibactin and a receptor complex that recognizes ferric-anguibactin (1, 13, 14, 22). In some bacterial species such as Escherichia coli, regulation of the iron-uptake gene expression depends on a single gene, fur (ferric uptake regulator) (20), whose product acts as a classical repressor, blocking transcription in the presence of high con- centrations of iron (4, 21, 36). We recently reported the cloning and characterization of the V anguillarum fiur gene and deter- mined that Fur plays a role in regulation of the synthesis of anguibactin and the outer membrane protein FatA (42, 45). In addition, in V anguillarum there are at least three plasmid- mediated regulators besides the chromosomally mediated Fur (33, 34, 40, 41). The V anguillarum Fur protein shares a high degree of homology with the other Fur protein sequences reported in the literature (5, 23, 24, 28, 37, 42). Fur exerts its regulatory function by binding to the operator of Fur-regulated genes (4, 12, 16, 17). Although indirect, there is evidence implicating the C-terminal portion of the E. coli Fur protein in Fe2+ binding, which may induce a conforma- tional change in the N-terminal region of the protein that allows the binding, possibly as a dimer, to the operator of Fur-regulated genes (12). However, the characteristic helix- turn-helix motif (8, 9) present in many DNA-binding proteins is not found. Therefore, the study of the V anguillarum Fur protein is of great importance not only to understand the regulation of the iron uptake system but also as a general model of regulation and DNA-protein interactions. As part of these studies, we characterized two V. anguillarum fur mutants that we have recently isolated (42), using the MnCl2 resistance selection method (21). In this work we characterize the products of these mutations and investigate the implica- tions of our findings to the structure-function relationships of Fur. (Part of this research was presented at the 94th General * Corresponding author. Mailing address: Department of Molecular Microbiology and Immunology L220, School of Medicine, Oregon Health Sciences University, 3181 SW Sam Jackson Park Rd., Portland, OR 97201-3098. Phone: (503) 494-7583. Fax: (503) 494-6862. Meeting of the American Society for Microbiology, Las Vegas, Nev., 23 to 27 May 1994.) Complementation offur mutants by V. anguillarum wild-type Fur. We have recently isolated two V. anguillarum fur mutants, 775met9 and 775metll (Table 1), by selection in the presence of MnCl2 (42). To determine the nature of these two muta- tions, we performed complementation studies with the wild- type fur gene clone from V anguillarum, pMET67 (45). We conjugated this plasmid together with either the reporter plasmid pSC27.1 or the control plasmid pRT240 (10) into the mutant strains 775met9 and 775metll. Cultures were grown under iron-rich conditions (75 to 100 tLM FeCl3), and conju- gations between V. anguillarum and E. coli were performed as described previously (40). The transconjugants possessed wild-type Fur activity as determined by the P-galactosidase activity (expressed in Miller units [26, 29]) of the reporter constructs: 1.8 for wild-type strain 775; 5.9 and 0.5 for strain 775met9 without and with pMET67, respectively; and 11.7 and 0.4 for strain 775metll without and with pMET67, respec- tively. These values are representative of three independent experiments. Cloning and sequencing of mutantfir genes. Clones from V. anguillarum Fur mutant 775met9 as well as the parent strain V. anguillarum 775 Nal' Rif, carrying the wild-typefur gene, were obtained after PCR with the appropriate primers to amplify the fur open reading frame (18). To detect functional Fur proteins, we used the reporter strain RRJC1 (Table 1). This strain possesses chromosomally encoded ,3-galactosidase un- der the control of a promoter carrying anfhuF Fur box, which, in the presence of a functional Fur, is down regulated under high concentrations of iron, resulting in white colonies on MacConkey plates with 100 liM FeCl3. Therefore, after trans- formation of the PCR clones containing either wild-type fur or mutant 9 fiur inserts into the reporter E. coli RRJC1, we obtained derivatives with mutant phenotypes. In the case of mutant strain 775metll, we amplified by PCR the upstream region as well as the open reading frame to analyze the sequence directly instead of cloning, as discussed in the following section. Three of the clones containing mutation 9 were sequenced. In all three derivatives, we identified a single point mutation, a base change from T to C at position 332, which results in an 5116 JOURNAL OF BACTERIOLOGY, Aug. 1994, p. 5116-5122 0021-9193/94/$04.00+0 Copyright © 1994, American Society for Microbiology on July 7, 2015 by guest http://jb.asm.org/ Downloaded from
Transcript of NOTES Structural andFunctional Analyses ofMutantFurProteins with Impaired Regulatory Function

Vol. 176, No. 16
NOTES
Structural and Functional Analyses of Mutant Fur Proteins withImpaired Regulatory Function
ANNE M. WERTHEIMER, MARCELO E. TOLMASKY, LUIS A. ACTIS, AND JORGE H. CROSA*
Department of Molecular Microbiology and Immunology, School of Medicine,Oregon Health Sciences University, Portland, Oregon 97201-3098
Received 23 March 1994/Accepted 8 June 1994
Vibrio anguillarum Fur mutants, 775met9 and 775metll, were characterized. V. anguillarum 775met9 had achange ofD to G at position 104 located in the carboxy terminus resulting in impaired Fur activity. Computeranalysis predicts perturbation of an alpha-helix in the carboxy terminus which may interfere with Fur proteinconformation. Strain 775metll had a change in the start codon resulting in no protein synthesis. The mutantsare unstable, and reversion to the wild type occurs frequently.
An important virulence factor of the fish pathogen Vibnoanguillarum is its plasmid-mediated iron uptake system, com-posed of the siderophore anguibactin and a receptor complexthat recognizes ferric-anguibactin (1, 13, 14, 22). In somebacterial species such as Escherichia coli, regulation of theiron-uptake gene expression depends on a single gene, fur(ferric uptake regulator) (20), whose product acts as a classicalrepressor, blocking transcription in the presence of high con-centrations of iron (4, 21, 36). We recently reported the cloningand characterization of the V anguillarum fiur gene and deter-mined that Fur plays a role in regulation of the synthesis ofanguibactin and the outer membrane protein FatA (42, 45). Inaddition, in V anguillarum there are at least three plasmid-mediated regulators besides the chromosomally mediated Fur(33, 34, 40, 41). The V anguillarum Fur protein shares a highdegree of homology with the other Fur protein sequencesreported in the literature (5, 23, 24, 28, 37, 42).Fur exerts its regulatory function by binding to the operator
of Fur-regulated genes (4, 12, 16, 17). Although indirect, thereis evidence implicating the C-terminal portion of the E. coliFur protein in Fe2+ binding, which may induce a conforma-tional change in the N-terminal region of the protein thatallows the binding, possibly as a dimer, to the operator ofFur-regulated genes (12). However, the characteristic helix-turn-helix motif (8, 9) present in many DNA-binding proteinsis not found. Therefore, the study of the V anguillarum Furprotein is of great importance not only to understand theregulation of the iron uptake system but also as a generalmodel of regulation and DNA-protein interactions. As partof these studies, we characterized two V. anguillarum furmutants that we have recently isolated (42), using the MnCl2resistance selection method (21). In this work we characterizethe products of these mutations and investigate the implica-tions of our findings to the structure-function relationships ofFur.
(Part of this research was presented at the 94th General
* Corresponding author. Mailing address: Department of MolecularMicrobiology and Immunology L220, School of Medicine, OregonHealth Sciences University, 3181 SW Sam Jackson Park Rd., Portland,OR 97201-3098. Phone: (503) 494-7583. Fax: (503) 494-6862.
Meeting of the American Society for Microbiology, Las Vegas,Nev., 23 to 27 May 1994.)Complementation offur mutants by V. anguillarum wild-type
Fur. We have recently isolated two V. anguillarum fur mutants,775met9 and 775metll (Table 1), by selection in the presenceof MnCl2 (42). To determine the nature of these two muta-tions, we performed complementation studies with the wild-type fur gene clone from V anguillarum, pMET67 (45). Weconjugated this plasmid together with either the reporterplasmid pSC27.1 or the control plasmid pRT240 (10) into themutant strains 775met9 and 775metll. Cultures were grownunder iron-rich conditions (75 to 100 tLM FeCl3), and conju-gations between V. anguillarum and E. coli were performedas described previously (40). The transconjugants possessedwild-type Fur activity as determined by the P-galactosidaseactivity (expressed in Miller units [26, 29]) of the reporterconstructs: 1.8 for wild-type strain 775; 5.9 and 0.5 for strain775met9 without and with pMET67, respectively; and 11.7 and0.4 for strain 775metll without and with pMET67, respec-tively. These values are representative of three independentexperiments.
Cloning and sequencing of mutantfir genes. Clones from V.anguillarum Fur mutant 775met9 as well as the parent strain V.anguillarum 775 Nal' Rif, carrying the wild-typefur gene, wereobtained after PCR with the appropriate primers to amplifythe fur open reading frame (18). To detect functional Furproteins, we used the reporter strain RRJC1 (Table 1). Thisstrain possesses chromosomally encoded ,3-galactosidase un-der the control of a promoter carrying anfhuF Fur box, which,in the presence of a functional Fur, is down regulated underhigh concentrations of iron, resulting in white colonies onMacConkey plates with 100 liM FeCl3. Therefore, after trans-formation of the PCR clones containing either wild-type fur ormutant 9 fiur inserts into the reporter E. coli RRJC1, weobtained derivatives with mutant phenotypes. In the case ofmutant strain 775metll, we amplified by PCR the upstreamregion as well as the open reading frame to analyze thesequence directly instead of cloning, as discussed in thefollowing section.Three of the clones containing mutation 9 were sequenced.
In all three derivatives, we identified a single point mutation, abase change from T to C at position 332, which results in an
5116
JOURNAL OF BACTERIOLOGY, Aug. 1994, p. 5116-51220021-9193/94/$04.00+0Copyright © 1994, American Society for Microbiology
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

NOTES 5117
TABLE 1. Bacterial strains and plasmidsa
Bacterial strain Relevant .ha' Source oror plasmid caractenstc~s1 reference
Bacterial strainsE. coliXL1 Blue supE44 hsdR17 recAl end&l gyrA46 thi relA lac mutant F'[proAB+ lacIq lacZAM15 Stratagene
TnlO(Tetr)]BL21(DE3)(pLysE) entE derivative of AB1515 38RRJC1 BN4020 derivative (E. coli fur mutant) with a f3-galactosidase reporter gene under the 30
control of the fhuF Fur box
V anguillarum775 Nalr Rif' Parent strain from which fur mutants were derived 13775met9 fur mutant isolated in the presence of 10 mM MnCl2 42775metll fur mutant isolated in the presence of 10 mM MnCl2 42775met9(pMET67) pMET67 (wild-type V anguillarum fur) conjugated into 775met9 42775metll(pMET67) pMET67 (wild-type V anguillarum fur) conjugated into 775metl1 42
PlasmidspBluescript SK+ Cloning vector StratagenepT7-5 Expression vector 39pT7-6 Expression vector 39pMET67 V angullarum fur cloned in pVK102 45pMETAW1.8 fur V anguillarum cloned into pBCSK+ 42pAWPCR9.1 Mutant fur from 775met9 subcloned into pBluescript SK+ This workpAWPCR9.IB Mutant fur from 775met9 subcloned into pT7-5 This workpAWPCR9.1M Site-directed mutagenized pAWPCR9.1 restoring wild-type D-104 in fur subcloned This work
into pBluescript SK+pAWPCR11.3 Site-directed mutagenized mutant fur from 775metl1 restoring wild-type AUG This work
initiation codon subcloned into pBluescript SK+pAWPCR11.3M Site-directed mutagenized pAWPCR11.3 creating a G-104 mutation in fur, subcloned This work
into pBluescript SK+pSC27.1 fur reporter gene; 0-galactosidase gene cloned under control of the ompF promoter 10
with a Fur between the promoter and the open reading framepRT240 Control plasmid for fur reporter gene constructs with ompF promoter without Fur box 10pTAW2.1 V anguillarum fur cloned into pT7-5 42a For the DNA procedures, modifications of previously described methods for genomic PCR (3) were used. Genomic DNA was isolated form cultures of V
anguillarum grown overnight at 270C in Trypticase soy broth supplemented with 1% NaCl by using a modification of the method of Birnboim and Doly (6). Alltransformations were performed by the method of Cohen et al. (11).
amino acid change from D to G at position 104. This mutationoccurs in an amino acid conserved in most known Fur proteins.
In the case of mutant 11, sequence analysis revealed a singlepoint mutation of nucleotide G-3 to T-3 resulting in a changeof G to U in the translation initiation codon. This mutationresults in the codon AUU, which is recognized as an initiationcodon in mitochondria (46).We also sequenced the upstream region of the mutant fur
genes to about 200 bp upstream from the Fur start sitecontaining putative -10 and -35 sequences (44, 47). Featuresof this region are depicted in Fig. 1. Comparison of thissequence to those of V vulnificus and V. cholerae shows thatthere is a ca. 100-bp deletion in the V. anguillarum and V
vulnificus regions, while the remainder of the upstream regionremains considerably conserved, including the anticipated ri-bosomal binding site, the predicted -35 and -10 promoterregions, and the predicted transcription start site. There alsois a region resembling a Fur box which overlaps both theputative -10 and + 1 sites of the V. anguillarum fur gene thatis closer to the consensus sequence, compared with the same
region in the fur genes from the other two Vibrio species. Nodifferences were found in the upstream regions of mutant 9 ormutant 11. These results were in agreement with the fact thatapproximately similar levels and sizes offur-specific transcriptswere detected in all samples from V anguillarum 775, 775met9,
and 775metl1 under both iron-rich and iron-limiting condi-tions (data not shown).
Analysis of predicted secondary structure of Fur proteins.By using different approaches, such as marker exchange for V.cholerae (23), V vulnificus (24), and Y pestis (37), and by usingselection in the presence of MnCl2 for Pseudomonas aerugi-nosa (28) and V. anguillarum (42), it was possible to demon-strate that mutations in the fur-like gene in these bacteria alsoled to constitutivity of specific iron-regulated systems. Struc-tural analysis of E. coli Fur by nuclear magnetic resonance andcircular dichroism revealed that approximately 50% of theprotein contains alpha-helix motifs, especially concentrated infour regions comprising residues 15 to 36, 80 to 96, 105 to 123,and 130 to 147 (48). These alpha-helices may interact with eachother or with other regions of the protein upon the binding ofmetal ion and/or DNA (32). However, these helixes, being upto 21 residues in length, do not conform to the classicalhelix-turn-helix motif, which involves a two- or three-turn helixand a four-residue turn followed by a four-turn helix (8, 9).Although extensive work has been carried out on the pheno-type caused byftir mutations and some structural characteriza-tion has been worked out in E. coli Fur, very little is knownabout the structure and function relationships of the Furproteins.To gain information on the structure-function relationships
VOL. 176, 1994
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

5118 NOTES
1 50V.a. TGTTACGCAA TTGTAACTGG TAAGAGCTGT TATATCAGTG ATATTGCGCGV.v. TGTTATTAAA TTGTAACTGG TCAGATGAGC AATATAAGCG ACCTAACGCGV.c. TGTTAAATTA TTGTAACTGG TAAGAGCACT TTGTTCAGTG ATTTGCCTCG
51V.a. GCTTTGTGAC AAATATAAAG TAACCCTTTG AACTTCGTGG TTTAV.v. AATTTACAAA AAGAATATAG TAACCCTTTG AAGTTCGTGG TTV.c. ACTTTGC..A GGTTTCCCAT TAACCTCTTG AAGTTCGTGG TT
101 -10 150V.a. GCAGTGACTC CCC G .ATCGCAATT ATGAATTCTG TT........V.v. TCTCTGGTTA ACCMiAIG .TTCAGAATA TTGAATTCTG TT........V.c. GCGGTGACTC ACC G CAAAGCAACA TTGACTTCTG TTATACCCAA
TfurfgtcoATmATG ATTATCATT AT
151 200V.a. .................... .......... .......... ..........
V.v .......... .......... .......... .......... ..........
V.c. ACGAATTGAG ATGCAGGTAG ACGGCAAGTG AGTGATCCCC ACTACAGCAA
201 250V.a. .................... .......... .......... ..........
V.v . .......... ......... .......... .......... ..........
V.c. CGGTTGAGGT GGGTTTGACA ACGCAGCCAA TACGATTGCC ATTTCATGTA
251 SD + 300V.a..........A ACTGCTGCAG ATQAT CAAGTGTA TGTCAGATAA
V.v..........A ATCGCGGCAG ATCATc AAGTATA TGTCAGACAA
V.c. GAACGGATGT ATCACTGCAG AACATC...,,.G.AAGTATA TGTCAGACAA
301 350V.a. TAACCAAGCG CTCAAGGATG CAGGTCTTAA AGTTACCCTT CCTAGGCTAAV.v. TAACCAAGCG CTAAAGGATG CTGGTCTTAA AGTTACCCTT CCAAGGCTGAV.c. TAACCAAGCG CTAAAGGATG CTGGTCTTAA AGTTACCCTC CCACGGCTTA
FIG. 1. Sequence of the upstream region of V anguillarum fur(V.a.) compared with those of V vulnificus (V.v.) and V cholerae(V.c.), illustrating putative -35 and conserved -10 TATAAT se-
quences, transcriptional start site (J), Shine-Dalgarno (SD), andprotein start site (C). Sequencing of double- and single-stranded DNAor gel-purified PCR fragments was performed by the dideoxy chaintermination method (35) with Sequenase kits (U.S. Biochemical,Cleveland, Ohio).
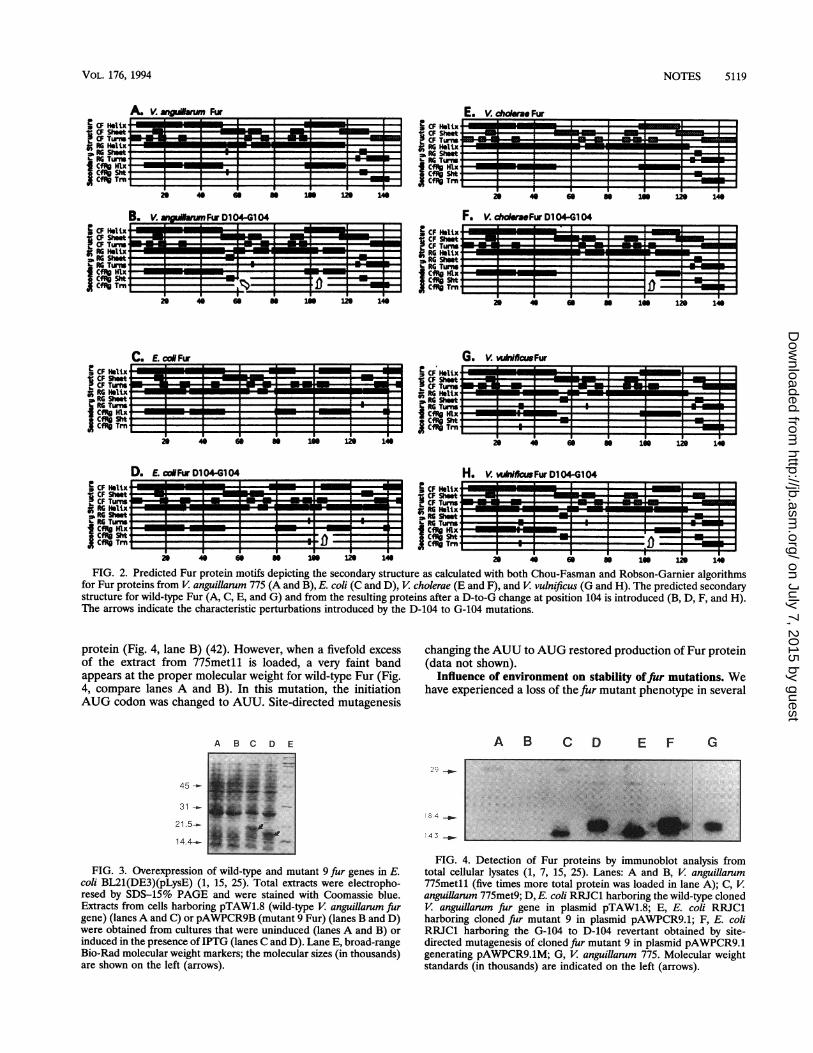
of Fur, we examined our model system, the V. anguillarum Furprotein. We analyzed the computer-predicted secondary struc-ture, as well as the predicted structural changes resulting froma D-104 to G-104 mutation in the amino acid sequence of theV. anguillarum Fur protein, using both the Genetics ComputerGroup (1991) Program Manualfor the GCG Package, version 7,and Mac Vector software (International Biotechnology, Inc.,New Haven, Conn.). Figure 2 shows these computer-predictedsecondary structures, as well as the predicted effect of asimulated D-104 to G-104 mutation in the secondary structureof the Fur proteins from V cholerae (panels E and F), V
vulnificus (panels G and H), and E. coli (panels C and D). Thecomputer analysis confirms a secondary structure for E. coliFur similar to that from the experimental analyses cited above.We found that the general alpha-helical pattern found in E.coli Fur occurs to a similar extent in the wild-type Fur from V.anguillarum, V cholerae, and V vulnificus. The alpha-helixpatterns in the carboxy terminus of V. anguillarum, V cholerae,and V vulnificus Fur proteins are quite similar to each otherbut differ from those in the similar region of the E. coli Furprotein.The D-104 to G-104 mutation results in a critical perturba-
tion in the alpha-helix region found in the carboxy terminus ofthis protein, causing an interruption of the alpha-helix. (Fig.2A and B) (27, 31). As is the case for the V anguillarum Furprotein, introduction of a D-104 to G-104 mutation in V.cholerae and V vulnificus Fur proteins also leads to a disrup-tion in the alpha-helix pattern. However, in these cases, theresult of the mutation is that the initial alpha-helix region ofabout 10 amino acid residues is no longer part of the alpha-helix motif. It appears that this region in the wild-type Furproteins from V cholerae and V vulnificus may be less con-
strained to alpha-helicity than that of the wild-type Fur from V.anguillarum. It is of interest that the E. coli Fur protein alphahelix motif does not appear to be significantly altered by theD-104 to G-104 mutation, a prediction which correlates withthe fact that this alpha-helix motif does not begin until residue105 (48). Thus, on the basis of both our computer model andthe fact that the amino acid flanking this single mutation is aserine in the Vibrid Fur protein sequence, the probability thatthe D-104 to G-104 mutation causes termination of alpha-helicity at this region of the carboxy terminus is greatlyincreased.
Analysis of mutant Fur protein expression. The PCR insertfrom mutant 9 with the D-104 to G-104 mutation in the Vanguillarum Fur protein was subcloned into the expressionvector pT7-5 and then transformed into E. coli BL21(DE3)(pLysE) (39). Using IPTG (isopropyl-p-D-thiogalactopy-ranoside) to induce gene expression, we demonstrated that thismutant fur gene could be expressed in E. coli (Fig. 3, lane D).It can be seen in this figure that the mutant protein migratesmore rapidly than the 17-kDa wild-type protein (comparelanes C and D). To corroborate that our findings are not anartifact of the overexpression of the cloned mutant 9 gene in E.coli, we also analyzed the mobility of the chromosomallyencoded mutant 9 protein synthesized by V. anguillarum775met9 compared with that synthesized by the cloned mutant9 fur gene in E. coli. Figure 4, lanes C and E, shows that bothproteins migrate in the same manner.To further assess that this single point mutation was respon-
sible for the faster migration, as well as for the diminished Furfunction (42), we used pAWPCR9.1, which is the mutant 9gene cloned by using pBluescript SK+ as the vector. Thisrecombinant plasmid was used to perform site-directed mu-tagenesis to revert the mutated fur gene to the wild-typesequence (pAWPCR9.lM) as well as to assess Fur activity.Transformation of pAWPCR9.1M DNA into E. coli RRJC1resulted in white colonies on MacConkey agar containing100 ,uM FeCl3, indicating that the wild-type form of Furwas recovered. As shown by immunoblot analysis (Fig. 4,lanes E and F), concomitant with the reversion from G-104 toD-104, wild-type mobility of the revertant Fur was also recov-ered.
Finally, we also performed site-directed mutagenesis of thecloned wild-type Fur open reading frame reproducing muta-tion 9. This mutated clone encoded a mutant Fur protein thatmigrated with the same mobility as the mutant 9 Fur proteinfrom 775met9 (data not shown). It is also apparent from Fig. 4that similar levels of Fur protein are produced from bothwild-type and mutant 9 fur clones in E. coli, as is the case forwild-type and mutant 9 proteins in V anguillarum.The different migration and diminished activity of Fur
mutant 9 occur when the mutated chromosomal fur gene isexpressed not only in the V. anguillarum cell environment butalso as a cloned gene in an E. coli background, suggesting thatthe D-104 to G-104 mutation results in a change that isintrinsically related to the protein encoded by the mutatedgene, either by changing the spatial conformation or by makingthe protein more susceptible to degradation. The loss of sucha large negatively charged amino acid has previously beendetermined to cause aberrant migration of a protein in sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).Our results showed that mutant 775met11 led to the consti-
tutive expression of both the FatA outer membrane proteinreceptor and catechols under both iron-rich and iron-limitingconditions (2). Inspection of the total proteins with the Furantiserum showed that this mutant did not synthesize any Fur
J. BAcTERiOL.
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
NOTES 5119
A. V. garum Fur
I a
B. V. Mum Fur D104-G104
20 4 G0
16 140
I1iI"n 126
CF Hel x
MG Helixt KF Sheet& K TumeI Cflg HlxCfl" Sht
CfIu Tm
CF Heltx1:CF SimtCF TurnG elitxRG Shet
L. RG TumeI Cal NxCa ShtC Tmdt
140
E. V. dodr Fur
4 i a
F. V.chdraeFurDlO4-Gl04
4 I f i
3I
CF Helix
KG HelixRK ShoetRK TumsefRC NIxRC Shot
YCOR Trn
I in 120 140
G. V. WfiluscFur
CF HelixCF SimtsCF TUmmR HelixRK Shet
L KG TuoeCfR HIxCfR ShtCW Tmn
D. E. ciFur Dl 04-Gl 04
20 40 66 120 146 a 4 60 06 16 120 14
FIG. 2. Predicted Fur protein motifs depicting the secondary structure as calculated with both Chou-Fasman and Robson-Garnier algorithmsfor Fur proteins from V anguillarum 775 (A and B), E. coli (C and D), V cholerae (E and F), and V vulnificus (G and H). The predicted secondarystructure for wild-type Fur (A, C, E, and G) and from the resulting proteins after a D-to-G change at position 104 is introduced (B, D, F, and H).The arrows indicate the characteristic perturbations introduced by the D-104 to G-104 mutations.
protein (Fig. 4, lane B) (42). However, when a fivefold excessof the extract from 775metll is loaded, a very faint bandappears at the proper molecular weight for wild-type Fur (Fig.4, compare lanes A and B). In this mutation, the initiationAUG codon was changed to AUU. Site-directed mutagenesis
A B C D E
45 -
31 -_
21.5-_-
14.4-.
FIG. 3. Overexpression of wild-type and mutant 9 fur genes in E.coli BL21(DE3)(pLysE) (1, 15, 25). Total extracts were electropho-resed by SDS-15% PAGE and were stained with Coomassie blue.Extracts from cells harboring pTAW1.8 (wild-type anguillarum furgene) (lanes A and C) or pAWPCR9B (mutant 9 Fur) (lanes B and D)were obtained from cultures that were uninduced (lanes A and B) orinduced in the presence of IPTG (lanes C and D). Lane E, broad-rangeBio-Rad molecular weight markers; the molecular sizes (in thousands)are shown on the left (arrows).
changing the AUU toAUG restored production of Fur protein(data not shown).
Influence of environment on stability offur mutations. Wehave experienced a loss of the fur mutant phenotype in several
A B C D E F G
29 _
184 _
143 _
FIG. 4. Detection of Fur proteins by immunoblot analysis fromtotal cellular lysates (1, 7, 15, 25). Lanes: A and B, V anguillarum775metll (five times more total protein was loaded in lane A); C, Vanguillarum 775met9; D, E. coli RRJC1 harboring the wild-type clonedV anguillarum fur gene in plasmid pTAW1.8; E, E. coli RRJC1harboring cloned fur mutant 9 in plasmid pAWPCR9.1; F, E. coliRRJC1 harboring the G-104 to D-104 revertant obtained by site-directed mutagenesis of cloned fur mutant 9 in plasmid pAWPCR9.1generating pAWPCR9.1M; G, V anguillarum 775. Molecular weightstandards (in thousands) are indicated on the left (arrows).
CF Helix1:CF ShootCF Turns-G HelixK Sheet
L RG TuraI* fSht,A C ,Tm
CF HelixCF Shoot,CF Turns,AK HelixMG Sheet
L MG TurHlx*fbShtI1Cf Tm,
1 1I* i 14
C. E. cok Fur
26
1N 120 1
I I i 1 I 120 140
VOL. 176, 1994
a
4 i f
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

5120 NOTES
A B C D E F G H J
43
29 _
18.4 _14.3 _..
FIG. 5. Detection of wild-type and mutant Fur proteins from Vanguillarum cells grown in different media. Immunoblot of totalcellular lysates probed with anti-Fur antiserum (1, 7, 15, 25). Furproteins from V anguillarum 775 (lanes A to C) and mutant 775met9(lanes D to I) obtained from respective Trypticase soy broth agarplates and grown in Trypticase soy broth (lanes A and D), in completeminimal medium inoculated from the previous Trypticase soy brothculture (lanes B and E), in iron-rich complete minimal mediuminoculated from the previous complete minimal medium culture (lanesC and F), or in iron-deficient complete minimal medium (lane G). Forlanes F and G, cells were first obtained from the same completeminimal medium culture used to prepare the proteins shown in lane Eand then inoculated into either the iron-rich or iron-deficient completeminimal medium. For lane H, proteins were obtained from cells ofmutant 775met9 grown in complete minimal medium that had beendirectly inoculated from the same Trypticase soy broth agar plate usedto generate the cultures for lanes D to I. Lane I, the proteins from775met9 cells grown in complete minimal medium obtained directlyfrom a frozen stock. Lane J, extract from the nearly null mutant775metll as a control for nonspecific binding. Molecular weightstandards (in thousands) are indicated on the left.
cultures of mutants 775met9 and 775metll. We proceeded toinvestigate this phenomenon with the 775met9 strain. Origi-nally, we observed a decrease in the loss of the constitutivephenotype as determined by the Arnow assay for catechols (2).When total cell extracts were made from these cultures of775met9 (Fig. 5, lane D), two protein bands appeared, one atthe position of the wild-type Fur and one migrating as mutant9 Fur protein. We hypothesize that the presence of bothspecies of proteins may be due to the fact that in the culturethere is a mixture of mutant and revertant cells.
Reversion to wild-type Fur protein mobility was also ob-served for 775met9 extracts when the strain had been passedfrom Trypticase soy agar to Trypticase soy broth, as well asafter serial passage from Trypticase soy agar to Trypticase soybroth (Fig. 5, lane D) and to complete minimal medium (Fig.5, lane E), with a final passage to complete minimal mediumwith added iron (Fig. 5, lane F) and complete minimal mediumcontaining ethylenediamine-di-(o-hydroxyphenyl)acetic acid(EDDHA) (iron limiting) (Fig. 5, lane G). It was of interestthat when the final passage was from complete minimalmedium into iron-limited minimal medium (Fig. 5, lane G), wedetected less wild-type Fur protein. Furthermore, when theinoculum was taken directly from the frozen stock into theiron-limited complete minimal medium (Fig. 5, laneI), the migration of the mutant 775met9 Fur protein wasmaintained with no reversion detected, even after a furtherpassage in this medium. When an inoculum was taken directlyfrom the same Trypticase soy agar plate as that used to initiatethese experiments involving 775met9 and inoculated directlyinto iron-limited complete minimal medium (Fig. 5, laneH), we detected more mutant 9 Fur protein than the wild-type form. Figure 5, lanes A to C, also shows that similarculture manipulations for the 775 Nalr Rif parent strain donot cause any changes in the wild-type mobility of the Furprotein.
Having demonstrated the ability to revert, we attempted todefine a medium which prevented reversion. When cultures
A B C D E29 _
18.414.3_
FIG. 6. Detection of wild-type and mutant Fur 9 proteins grown inthe presence of MnCl2 by immunoblot of total cellular lysates probedwith anti-Fur antiserum (1, 15, 25). Fur proteins from V. anguillarum775 (lane A) and mutant 775met9 (lane B) obtained from culturesgrown in TSBS (Trypticase soy broth supplemented with 1% NaCl)and minimal medium, respectively. Lanes: C to E, proteins from775met9 after frozen inoculum was plated directly onto TSAS supple-mented with 1 mM MnCl2, 2 mM MnCl2, 4 mM MnCl2, respectively.Molecular weight standards (in thousands) are indicated on the left,and the wild-type and mutant Fur 9 are indicated on the right.
were grown on TSAS (Trypticase soy agar supplemented with1% NaCl) medium with increasing MnC12 concentrations, wediscovered the disappearance of the wild-type Fur protein intotal cell extracts run in SDS-PAGE (Fig. 6). Cultures grownfrom frozen inoculum streaked onto TSAS with 4 mM MnCl2typically required 24 to 48 h to grow; only mutant 9 Fur proteinwas detected (Fig. 6, lane E), but at lower concentrations,wild-type Fur protein was present (Fig. 6, lanes C and D). Thewild-type 775 strain, also streaked directly from frozen inocu-lum, consistently failed to grow within 48 h on TSAS platessupplemented with 4 mM MnCl2. An interesting note is thatthese reversions to wild-type mobility of the 775met9 protein inV anguillarum were not seen in any of the clones containingthe mutant 9 fur gene in E. coli.We have consistently seen selection for bacteria carrying the
wild-type Fur proteins under nutrient-rich and/or high-ironconditions. Thus, we compared the growth of 775 and 775met9in iron-rich and iron-limiting minimal media (Table 2) andfound that the mutant Fur strain 775met9 consistently growsmore poorly than the wild type strain. We also demonstratethat while the 775 wild-type strain thrives in iron-rich condi-tions compared with iron-limiting conditions, the mutant strain775met9 grows considerably less voraciously in non-iron-limit-ing conditions, and yet it grows similarly to the wild type iniron-limiting medium. This difference in growth leads us tohypothesize that in these mutant cultures once reversion towild-type Fur begins, the portion of the population containingwild-type Fur will grow more rapidly than the mutant cells inthat population, especially under iron-rich conditions, unlessthere is specific selection pressure in the form of MnCl2 tomaintain the mutant population. We are quite interested inwhy the mutant strain does not thrive in an iron-rich medium.As Fur in other bacteria has been shown to function pleitropi-cally, many possibilities for poor growth exist. Perhaps, a highintracellular iron concentration could be deleterious to thecell, for instance, by catalyzing the production of free radicals,
TABLE 2. Comparison of growth of wild-type V anguillarum 775and mutant strain 775met9
Strain OD6w in minimal medium witha: FeCI3/EDDHA100 ,uM FeCl3 4 ,uM EDDHA ratio
775 2.5 0.94 2.7775met9 1.2 0.75 1.5
a An inoculum was taken directly from frozen cultures diluted into minimalmedium. A sample (100 pl) was inoculated into minimal medium supplementedwith 100 ,LM FeCl3 or 4 ,uM EDDHA. Cultures were grown overnight at 27°C.ODwo, optical density at 600 nm.
J. BA=-1RIOL.
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

NOTES 5121
as in a Haber-Weiss-type reaction (19). Functional Fur proteinmay successfully occupy the metal ion and prevent damage bythe free iron or somehow maintain the intracellular ironbalance.
Understanding the function-structure relationships leadingto the interaction of the Fur protein with iron, DNA, and otherparameters will likely be increased by analysis of these andother mutants that we have isolated. Thus, it has become clearthat we have isolated two very different types of mutationswhich lead to loss of Fur function. Fur mutant 9 gives us theopportunity to examine questions involving the structure-function of this DNA-binding protein, while mutant 11 drawsour attention to a more general role played by Fur within thebacterium.
Special thanks to Richard Brennan for assistance in interpreting ourprotein structure calculations.
This work was supported by NIH grant A119018.
REFERENCES
1. Actis, L. A., S. A. Potter, and J. H. Crosa. 1985. Iron-regulatedouter membrane protein OM2 of Vibrio anguillarum is encoded byvirulence plasmid pJM1. J. Bacteriol. 161:736-742.
2. Arnow, L. 1937. Colorimetric determination of the components of3,4-dihydroxyphenylalanine-tyrosine mixtures. J. Biol. Chem. 118:531-537.
3. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G.Seidman, J. A. Smith, and K. Struhl (ed.). 1990. Current protocolsin molecular biology. John Wiley and Sons, New York.
4. Bagg, A., and J. Neilands. 1987. Ferric uptake regulation proteinacts as a repressor, employing iron(II) as a cofactor to bind theoperator of an iron transport operon in Escherichia coli. Biochem-istry 26:5471-5477.
5. Berish, S. A., S. Subbarao, C.-Y. Chen, D. L. Trees, and S. A.Morse. 1993. Identification and cloning of a fiur homolog fromNeisseria gonorrheae. Infect. Immun. 61:4599-4606.
6. Birnboim, H. C., and J. Doly. 1979. A rapid alkaline extractionprocedure for screening recombinant plasmid DNA. NucleicAcids Res. 7:1513-1523.
7. Bradford, M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of proteins utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:249-254.
8. Brennan, R. 1991. Interactions of the helix-turn-helix bindingdomain. Curr. Opin. Struct. Biol. 1:80-88.
9. Brennan, R. 1993. The winged-helix DNA-binding motif: anotherhelix-turn-helix takeoff. Cell 74:773-776.
10. Calderwood, S. B., and J. Mekalanos. 1988. Confirmation of thefur operator site by insertion of a synthetic oligonucleotide into anoperon fusion plasmid. J. Bacteriol. 170:1015-1017.
11. Cohen, S. N., A. C. Y. Chang, and L. Hsu. 1972. Non-chromosomalantibiotic resistance in bacteria: genetic transformation ofEschenchiacoli by R-factor DNA. Proc. Natl. Acad. Sci. USA 69:2110-2114.
12. Coy, M., and J. Neilands. 1991. Structural dynamic and functionaldomains of the Fur protein. Biochemistry 30:8201-8210.
13. Crosa, J. H. 1980. A plasmid associated with virulence in themarine fish pathogen Vibrio anguillarum specifies an iron-seques-tering system. Nature (London) 284:566-568.
14. Crosa, J. H. 1989. Genetics and molecular biology of siderophore-mediated iron transport in bacteria. Microbiol. Rev. 53:517-530.
15. Crosa, J. H., and L. Hodges. 1981. Outer membrane proteinsinduced under conditions of iron limitation in the marine fishpathogen Vibrio anguillarum 775. Infect. Immun. 31:223-227.
16. de Lorenzo, V. L., M. Herrero, F. Giovannini, and J. B. Neilands.1988. Fur (ferric uptake regulator) protein and CAP (catabolic-activator protein) modulate transcription offur gene in Escherichiacoli. Eur. J. Biochem. 173:537-546.
17. de Lorenzo, V. L., S. Wee, M. Herrero, and J. B. Neilands. 1987.Operator sequences of the aerobactin operon of plasmid pColV-
K30 binding the ferric uptake regulation (fur) repressor. J. Bacte-riol. 169:2624-2630.
18. Gammie, A., M. E. Tolmasky, and J. H. Crosa. 1993. Functionalcharacterization of a replication initiator protein. J. Bacteriol. 175:3563-3569.
19. Haber, F., and J. Weiss. 1934. The catalytic decomposition ofhydrogen peroxyde by iron salts. Proc. R. Soc. Lond. A 147:332-351.
20. Hantke, K. 1984. Cloning of the repressor protein gene of iron-regulated systems in Escherichia coli K12. Mol. Gen. Genet. 197:337-341.
21. Hantke, K. 1987. Selection procedure for deregulated iron trans-port mutants (fur) in Escherichia coli K12. Mol. Gen. Genet.210:135-139.
22. Jalal, M., D. Hossain, J. van der Helm, J. Sanders-Loehr, L. A.Actis, and J. H. Crosa. 1989. Structure of anguibactin, a uniqueplasmid-related bacterial siderophore from the fish pathogenVibrio anguillarum. J. Am. Chem. Soc. 111:292-296.
23. Litwin, C. M., S. A. Boyko, and S. B. Calderwood. 1992. Cloning,sequencing and transcriptional regulation of the Vibrio choleraefurgene. J. Bacteriol. 174:1897-1903.
24. Litwin, C. M., and S. B. Calderwood. 1993. Cloning and geneticanalysis of the Vibrio vulnificus fur gene and construction of afur mutant by in vivo marker exchange. J. Bacteriol. 175:706-715.
25. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
26. Miller, J. H. 1972. Experiments in molecular genetics, p. 352-355.Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
27. Presta, L. G., and G. D. Rose. 1988. Helix signals in proteins.Science 240:1632-1641.
28. Prince, R., C. Cox, and M. Vasil. 1993. Coordinate regulation ofsiderophore and exotoxin A production: molecular cloning andsequencing of the Pseudomonas aeruginosa fur gene. J. Bacteriol.175:2589-2598.
29. Putnam, S., and A. Koch. 1975. Complications in the simplestcellular enzyme assay: lysis of Escherichia coli for the assay of,B-galactosidase. Anal. Biochem. 63:350-360.
30. Read, R., M. Tolmasky, A. Wertheimer, and J. Crosa. Unpublisheddata.
31. Richardson, J. S., and D. Richardson. 1988. Amino acid prefer-ences for specific locations at the ends of a helices. Science240:1648-1652.
32. Saito, T., and J. P. Williams. 1990. The binding of the ferric up-take regulation protein to a DNA fragment. FEBS Lett. 197:43-47.
33. Salinas, P., M. E. Tolmasky, and J. H. Crosa. 1989. Regulation ofthe iron uptake system in Vibrio anguillarum: evidence for acooperative affect between two transcriptional activators. Proc.Natl. Acad. Sci. USA 86:3529-3533.
34. Salinas, P., L. Waldebesser, and J. H. Crosa. 1993. Regulation ofthe expression of bacterial iron transport genes: possible role of anantisense RNA as repressor. Gene 123:33-38.
35. Sanger, F., S. Nicklen, and A. R. Coulson. 1979. DNA sequencingwith chain-termination inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
36. Schaffer, S., K. Hantke, and V. Braun. 1985. Nucleotide sequenceof the iron regulatory gene fiur. Mol. Gen. Genet. 200:110-113.
37. Staggs, T., and R. Perry. 1991. Identification and cloning of a furregulatory gene in Yersinia pestis. J. Bacteriol. 173:417-425.
38. Studier, F., A. Rosenberg, J. Dunn, and J. Dubendorff. 1990. Useof T7 RNA polymerase to direct expression of cloned genes.Methods Enzymol. 185:60-89.
39. Tabor, S., and C. Richardson. 1985. A bacteriophage T7 RNApolymerase/promoter system for controlled exclusive expression ofspecific genes. Proc. Natl. Acad. Sci. USA 82:1074-1078.
40. Tolmasky, M. E., L. A. Actis, and J. H. Crosa. 1988. Geneticanalysis of the iron uptake region of the Vibrio anguillarumplasmid pJM1: molecular cloning of genetic determinants encod-ing a novel trans activator of siderophore biosynthesis. J. Bacteriol.170:1913-1919.
41. Tolmasky, M. E., and J. H. Crosa. 1984. Molecular cloning and
VOL. 176, 1994
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

5122 NOTES
expression of genetic determinants for the iron uptake systemmediated by the Vibrio anguillarum plasmid pJM1. J. Bacteriol.160:860-866.
42. Tolmasky, M. E., A. M. Wertheimer, L. A. Actis, and J. H. Crosa.1994. Characterization of the Vibrio anguillarum fur gene: role inregulation of expression of the FatA outer membrane protein andcatechols. J. Bacteriol. 176:213-220.
43. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
44. Vieira, J., and J. Messing. 1982. The pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencing withsynthetic universal primers. Gene 19:259-268.
45. Waldbeser, L, M. E. Tolmasky, L. A. Actis, and J. H. Crosa. 1993.Mechanisms for negative regulation by iron of the FatA outermembrane protein gene expression in Vibrio anguillanrm 775. J.Biol. Chem. 268:10433-10439.
46. Watson, J. 1987. Molecular biology of the gene, p. 443. BenjaminCummings Publishing Company, Menlo Park, Calif.
47. Weickert, M., and G. Chambliss. 1989. Acid-phenol miniprepsmake excellent sequencing templates. U.S. Biochemical Corp.Editorial Comments 16:5-6.
48. Williams, R. G., M. R. Wormald, R C. Hider, and J. Neilands.1989. Spectroscopic properties of the FUR (Ferric Uptake Regu-lation) protein of Escherichia coli K-12, p. 189-196. In D. Hamerand D. Winge (ed.), Metal ion homeostasis: molecular biology andchemistry. Alan Liss Inc., New York.
J. BACrERIOL.
on July 7, 2015 by guesthttp://jb.asm
.org/D
ownloaded from




















