
Microtrapping characteristics of single and multi-walled carbon nanotubes

Toxicology Letters 178 (2008) 95–102
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
Non-functionalized multi-walled carbon nanotubes alter theparacellular permeability of human airway epithelial cells
Bianca Maria Rotoli a, Ovidio Bussolati a,∗, Massimiliano G. Bianchia, Amelia Barilli a,Chidambara Balasubramanianc, Stefano Bellucci c, Enrico Bergamaschib
a Department of Experimental Medicine, Unit of General and Clinical Pathology, University of Parma, via Volturno 14, 43100 Parma, Italyb Department of Clinical Medicine, Nephrology and Health Sciences, Unit of Occupational Medicine, University of Parma, Parma, Italyc
ble uof Cainglein thlial ee in T
gle-wmet
ring tMorethe T
INFN, Frascati, Rome, Italy
a r t i c l e i n f o
Article history:Received 14 January 2008Received in revised form 26 February 2008Accepted 26 February 2008Available online 4 March 2008
Keywords:Tight junctionsCarbon nanotubesAirway epitheliumFibrosisPermeability
a b s t r a c t
Little information is availastudy the barrier functionexposure to commercial ssition. To assess changesmeasured the trans-epitheCNT caused a large decreasin monolayer viability. Sinthrough the arc dischargewalled CNT were added duresistance was observed.Carbon Black, prevented
cytokine that causes a reversiblmercial multi-walled CNT interin airway epithelial cells.1. Introduction
The potential risks of exposure to nanoparticles (NP) ornanofibers are not predictable a priori from the bulk physicochem-ical characteristics of these materials. Indeed, due to their size andhigh surface/volume ratio, nanostructured materials have specificproperties that increase their industrial value but also point to theneeding for specific toxicological studies on the single nanoma-terial (Oberdorster et al., 2005). To this purpose, due to the highcosts associated to in vivo experiments, in vitro approaches arebeing developed to yield inexpensive and efficient assessments ofbiological effects of nanomaterials.
Among nanomaterials of industrial relevance, carbon nanotubes(CNT) are produced in increasing amounts for several industrial
Abbreviations: AD, arc discharge; CB, Carbon Black; CNT, carbon nanotubes; CVD,chemical vapour deposition; EMEM, Eagle’s minimum essential medium; HUVECs,human endothelial cells from umbilical veins; IL-4, interleukin-4; MWCNT, multi-walled carbon nanotubes; NM, nanomaterials; PBS, phosphate-buffered saline;SWCNT, single-walled carbon nanotubes; TEER, trans-epithelial electrical resistance.
∗ Corresponding author. Tel.: +39 0521 033783; fax: +39 0521 033742.E-mail address: [email protected] (O. Bussolati).
0378-4274/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.toxlet.2008.02.007
pon the effects of carbon nanotubes (CNT) on the airway barrier. Here welu-3 human airway epithelial cells, grown on permeable filters, after the-walled or multi-walled CNT, produced through chemical vapour depo-e paracellular permeability of CNT-treated Calu-3 monolayers, we havelectrical resistance (TEER) and the permeability to mannitol. Multi-walledEER and an increase in mannitol permeability but no substantial alterationalled CNT produced much smaller changes of TEER while CNT, synthesizedhod, and Carbon Black nanoparticles had no effect. If commercial multi-he formation of the tight monolayer, no further increase in trans-epithelial
over, the same nanomaterials, but neither single-walled counterparts norEER recovery observed after the discontinuation of interleukin-4, a Th2e barrier dysfunction in airway epithelia. These findings suggest that com-fere with the formation and the maintenance of tight junctional complexes
© 2008 Elsevier Ireland Ltd. All rights reserved.
applications, because these structures are endowed with extremelyadvantageous chemical and mechanical features. Moreover, it is rel-
atively easy to functionalize CNT with various chemical groups,so as to render them efficient carriers for chemicals and drugs(Banerjee et al., 2003; Bottini et al., 2005, 2006a,c; Donaldson et al.,2006; Lam et al., 2006). At present, CNT production occurs throughthree distinct processes: chemical vapour deposition (CVD), themajor process of industrial importance, arc discharge (AD), andlaser ablation. The two latter processes lead to the production ofCNT less contaminated with metals but are largely restricted tolaboratories for research use. Multi-walled CNT (MWCNT) can beproduced without metals, although the presence of small amountof metal catalysts helps to align the nanotubes; an increase in themetal nanoparticles-to-carbon ratio favors the formation of single-walled CNT (SWCNT) (Iijima and Ichihashi, 1993).The consequences of the interaction of CNT with biologicalstructures are not completely known and their potentially toxiceffects are still object of debate (Donaldson et al., 2006; Lam etal., 2006; Helland et al., 2007). The available information, obtainedmostly in rodents, points to lung toxicity as the major consequenceof exposure to CNT, with the early formation of fibrosis and granu-lomas, hypertrophy of epithelial cells, and the ensuing of functionalimpairment (Lam et al., 2004; Warheit et al., 2004; Muller et

ogy Le
96 B.M. Rotoli et al. / Toxicolal., 2005; Shvedova et al., 2005). Interestingly, these pathologicalchanges are observed with SWCNT but not with Ultrafine Car-bon Black (CB) used at comparable mass doses (Lam et al., 2004;Shvedova et al., 2005).
Although several toxicological studies have investigated CNTeffects on various cell models in vitro (Shvedova et al., 2003; Cuiet al., 2005; Monteiro-Riviere et al., 2005; Bottini et al., 2006b; DeNicola et al., 2007; Zhang et al., 2007), the consequences of CNTexposure on airway cells are only incompletely known. In particu-lar, SWCNT are more toxic than MWCNT for alveolar macrophages(Jia et al., 2005), while, most recently, rat alveolar epithelial cellshave been described to exhibit no cytotoxic effect after exposure toSWCNT (Yacobi et al., 2007).
Airway epithelial cells represent the first body barrier forinhaled particles and constitute a major determinant of the inter-action of the potentially toxic compound with other compartmentsof the organism. The barrier function of these cells is due to the for-mation and maintenance of tight junctional complexes that allowstrictly polarized secretory functions and prevent the entry of xeno-biotics and pathogens, lowering the paracellular permeability of theepithelium to very low levels. Tight junctions are dynamic multi-protein structures, which exhibit a peculiar sensitivity to cytokines,toxins, and pathogens (see for review Schneeberger and Lynch,2004; Aijaz et al., 2006; Shin et al., 2006). On the other hand, alter-ations in the paracellular permeability of airway epithelia havebeen implicated in enhanced sensitivity to pathogens or exagger-ated inflammatory reactions (Reed and Kita, 2004; Landau, 2006).
Airway epithelial cells grown in culture maintain the capabilityto form tight junctional complexes. Therefore, in vitro cell models ofairway epithelium are currently adopted to predict the behaviour ofthe respiratory barrier in vivo (Sakagami, 2006). We have used oneof these models, the human epithelial cell line Calu-3, to assess theconsequences of the exposure to various types of CNT on the para-cellular permeability of the airway epithelial cells. We have found
that commercial MWCNT and, at a much lesser degree, SWCNTimpair the barrier function of airway epithelial cell monolayers.Neither nanotubes synthesized with the arc discharge method (AD-CNT), nor amorphous carbon nanoparticles exhibit the same effects.2. Materials and methods
2.1. Cells and experimental treatments
Calu-3 cells, obtained from a human lung adenocarcinoma and derived fromserous cells of proximal bronchial airways (Finkbeiner et al., 1993), were obtainedfrom the Istituto Zooprofilattico Sperimentale della Lombardia (Brescia, Italy). Cellswere routinely cultured in 10-cm diameter dishes in Eagle’s Minimum EssentialMedium (EMEM) supplemented with 1 mM sodium pyruvate, 10% FBS, strepto-mycin (100 �g/ml) and penicillin (100 U/ml) in a humidified atmosphere of 5%CO2 in air. For the experiments, Calu-3 cells were seeded into cell culture insertswith membrane filters (pore size of 0.4 �m) for Falcon 24-well-multitrays (Cat.N◦ 3095, Becton, Dickinson & Company, Franklin Lakes, NJ, USA), at a density of75 × 103 cells/300 �l. Cells were allowed to grow for 10–14 d, as detailed for thesingle experiments. NM were added in the apical chamber from a 1 mg/ml stocksolution without changing the medium, while basolateral chamber was used forIL-4 supplementation.
THP-1 macrophages were obtained from the Istituto Zooprofilattico Sperimen-tale della Lombardia, cultured in RPMI1640 medium supplemented with 10% FBSand antibiotics, and differentiated for 3 d with 50 ng/ml phorbol-12,13-myristate
Table 1Principal characteristics of CNT preparations
Type Dimensions (diameter × length) Density (g/
MWCNT 110–170 nm × 5–9 �m 1.35SWCNT 0.7–1.2 nm × 2–20 �m 2.25AD-CNTa 10–40 nm × 1–5 �m ND
ND, not determined.a AD-CNT are a mixture of, approximately, 30% SWCNT, 50% MWCNT, and 20% amorpho
tters 178 (2008) 95–102
before use. Human endothelial cells from umbilical veins (HUVECs) were obtainedand cultured in Medium 199 supplemented with 20% FBS, heparin and endothelialcell growth supplement, as previously described (Visigalli et al., 2007).
Images of cells in phase contrast were obtained with a Nikon DS5MC digitalcamera (Nikon Instruments SpA, Firenze, Italy).
2.2. Characterization of CNT
Commercial multi-walled (MWCNT, Aldrich 659258) and single-walled (SWCNT,Aldrich 636797) nanotubes were obtained from Sigma–Aldrich, Milan, Italy. Amixture of single-walled and multi-walled CNT was obtained with the arc dis-charge method (AD-CNT) at the laboratories of Istituto Nazionale di Fisica Nucleare(INFN) Frascati, Rome (Bellucci et al., 2007a,b). Ultrafine Carbon Black nanoparticles(Printex 90TM, 14 nm diameter) were a generous gift of Degussa Italia SpA, AdvancedFillers & Pigments, Ravenna, Italy.
The characteristics of the commercial nanotubes, as declared by the manu-facturer, are as follows: MWCNT, Aldrich 659258 is largely made of multi-wallednanotubes (at least 90%), with residual amorphous carbon (metal contaminants, i.e.mostly iron, amount to less than 0.1%); the purity of commercial single-walled nan-otubes, Aldrich 636797 is only >50%, where the impurities consist approximatelyof a 40% amount of other types of nanotubes, 3% amorphous carbon and traces ofmetals.
In order to obtain a determination of the amount of possible contaminants inthe AD-CNT samples, we carried out their elemental analysis using a JEOL JEM 2010TEM microscope. The results indicate that only a Si contamination was detectable,while the AD-CNT samples were totally metal-free. The main characteristics of theCNT used are summarized in Table 1.
2.3. Exposure to CNT and CB
Before the experiments all the nanomaterials were heated at 220 ◦C for 3 h toeliminate possible contamination from lipopolysaccharide. After cooling at roomtemperature, nanomaterials were dispersed at a concentration of 1 mg/ml in sterilephosphate-buffered saline (PBS) to obtain a stock suspension for a series of experi-ments. Immediately before the single experiments, nanomaterials were extensivelyvortexed, sonicated three times for 15 min in a Bransonic Ultrasonic ultrasound bathand then added to normal growth medium to reach the desired concentration. Nodetergent was used to improve the solubility of nanomaterials in aqueous solutions.After the addition of nanomaterials to culture medium, they rapidly tend to precip-itate and to form more or less expanded aggregates that come into contact with thecell monolayer (see the representative image shown in Fig. 5). Given the volume of
the medium in the apical chamber and the filter surface, Calu-3 monolayers, grownin cell culture inserts for Falcon 24-well-multitrays and incubated with a nominalMWCNT concentration of 100 �g/ml, were exposed to 75 �g of nanomaterials percm2 of monolayer, roughly corresponding to a maximum of 0.3 ng/cell.2.4. Measurements of trans-epithelial resistance and paracellular permeability
Measurements of trans-epithelial electrical resistance (TEER) were made onmonolayers of Calu-3 cells, grown for 10–14 d on permeable supports (Falcon), usingan epithelial voltohmeter (EVOM, World Precision Instruments Inc., Sarasota, FL,USA).
Changes in the paracellular permeability to extracellular solutes were assessedwith measurements of the apical-to-basolateral flux of mannitol, as previouslydescribed (Rotoli et al., 2000).d-[1-14C]-Mannitol was added at a final concentrationof 0.1 mM and at the activity of 5 �Ci/ml to the apical side. At the indicated times,aliquots of 50 �l were sampled from the basolateral compartment to determine theradioactivity with a liquid scintillation spectrometer Wallac Trilux2 (Perkin ElmerSpA, Monza, Italy).
2.5. Cell viability
Cell viability was tested with the resazurin method (O’Brien et al., 2000). Accord-ing to this method, viable cells reduce the non-fluorescent compound resazurin intothe fluorescent resorufin that accumulates into the medium. Since the culture insertsdid not allow direct fluorescence reading from the wells, these experiments wereperformed on cells cultured on standard plasticware. Cells were incubated for 1 h
cm3) Surface area (m2/g) Preparation method
1.3 × 102 CVD1.7 × 103 CVDND Electric arc discharge
us C and fullerene. The dimensions refer to the MWCNT component.

ogy Letters 178 (2008) 95–102 97
B.M. Rotoli et al. / Toxicolwith fresh, serum-free medium supplemented with 44 �M resazurin; fluorescencewas then measured at 572 nm with a fluorimeter (Wallac 1420 Victor2 MultilabelCounter, Perkin Elmer).
2.6. Statistical analysis
Values of TEER of control and NM-treated cultures were compared with a t-testfor unpaired data.
2.7. Materials
Fetal bovine serum and culture media were purchased from Euroclone (Cel-bio, Pero, Milan, Italy). Recombinant human interleukin-4 (rIL-4) was obtainedfrom Vinci-Biochem, Firenze, Italy. [14C]Mannitol (51 mCi/mmol) was obtained fromPerkin Elmer Life and Analytical Sciences, Boston, MA, USA. Sigma–Aldrich (Milan,Italy) was the source of all other chemicals.
3. Results
3.1. Effects of CNT on the paracellular permeability of Calu-3 cellmonolayers
When seeded on permeable filters in a two-chamber culture sys-tem, Calu-3 cells spend 8–10 d to reach confluence. After this periodTEER progressively increases up to 13–15 d and reaches values ashigh as 2000 � cm2. The addition of MWCNT, SWCNT, AD-CNT,or CB, at a concentration of 100 �g/ml, to the apical chamber oftight Calu-3 cell monolayers (TEER ≥ 1000 � cm2) had very differ-ent effects on TEER (Fig. 1, Panel A). AD-CNT and CB were completelyineffective and the TEER of monolayers exposed to these nano-materials (NM) was not significantly different from that of controlmonolayers. In contrast, MWCNT produced a progressive decreasein TEER. The decrease was already detectable after 3 d of incuba-tion and, after 6 d, the TEER value was less than 30% of the controlTEER, determined at the same time in untreated monolayers. Atlater times of treatment (>4 d), also commercial SWCNT produceda statistically significant decrease of TEER, although at a much lesserdegree than MWCNT.
The effect of increasing concentrations of MWCNT on the TEERof tight monolayers was monitored for 4 d (Fig. 1, Panel B). Start-ing from Day 2 of treatment, concentrations as low as 10 �g/mlprevented further increases of TEER, so as to yield TEER values sig-
Fig. 1. Carbon nanotubes lower the trans-epithelial electrical resistance (TEER) of Calu-3filters. At the end of this period, MWCNT, SWCNT, AD-CNT, or CB were added to the apidetermined at the indicated times. Empty symbols, control filters maintained in the abfilters. At the end of this period, different concentrations of MWCNT were added to the aindicated times. For both panels, four filters were used for each condition. The figure showare means ± S.D. *p < 0.05, **p < 0.01 vs. control, untreated cultures.
Fig. 2. MWCNT increase monolayer permeability to mannitol. Calu-3 cells were cul-tured for 10 d on membrane filters. At the end of this period, monolayers wereincubated for 4 d in the absence or in the presence of 100 �g/ml MWCNT. d-[1-14C]-Mannitol (0.1 mM, 5 �Ci/ml) was then added to the apical chamber and50 �l-aliquots of the basolateral medium were sampled at the indicated timesand counted for radioactivity. Three filters were used for each condition. Dataare means ± S.D. Lines represent the best-fit linear regression of the mean val-ues. R2 = 0.909 and 0.908 for control and MWCNT-treated monolayers, respectively.
A representative experiment, performed three times with comparable results, isshown.nificantly lower than control, while even 5 �g/ml were able to lowerTEER at Day 4. At a nominal concentration of 50 �g/ml, the TEERvalue was intermediate between the values measured at 10 and100 �g/ml.
If the decrease in TEER caused by MWCNT were due to theopening of tight junctions, it should correspond to an increasein the paracellular permeability of Calu-3 cell monolayers to cell-excluded solutes. The experiment shown in Fig. 2 demonstrates thatthis is the case. In this experiment, labelled mannitol, an extracellu-lar solute (m.w. 180) that cannot be transported through the plasmamembrane, was added to the apical chamber of Calu-3 monolayerspre-incubated for 4 d in the absence or in the presence of 100 �g/mlMWCNT. The radioactivity in the basolateral chamber was thenmeasured after increasing periods of incubation. The results indi-cate that the paracellular permeability to mannitol increased from11.3 ± 1.03 to 49.6 ± 4.99 cpm/min in control and MWCNT-treatedmonolayers, respectively. In the same cultures, TEER was 57% lowerin MWCNT-treated than in control epithelial monolayers.
cell monolayers. (Panel A) Calu-3 cells were cultured for 10 d on 0.4 �m membranecal chamber of the culture system at a concentration of 100 �g/ml and TEER wassence of CNT. (Panel B) Calu-3 cells were cultured for 10 d on 0.4 �m membranepical chamber of the culture system, as indicated, and TEER was determined at the
s representative experiments repeated three times with comparable results. Data
98 B.M. Rotoli et al. / Toxicology Letters 178 (2008) 95–102
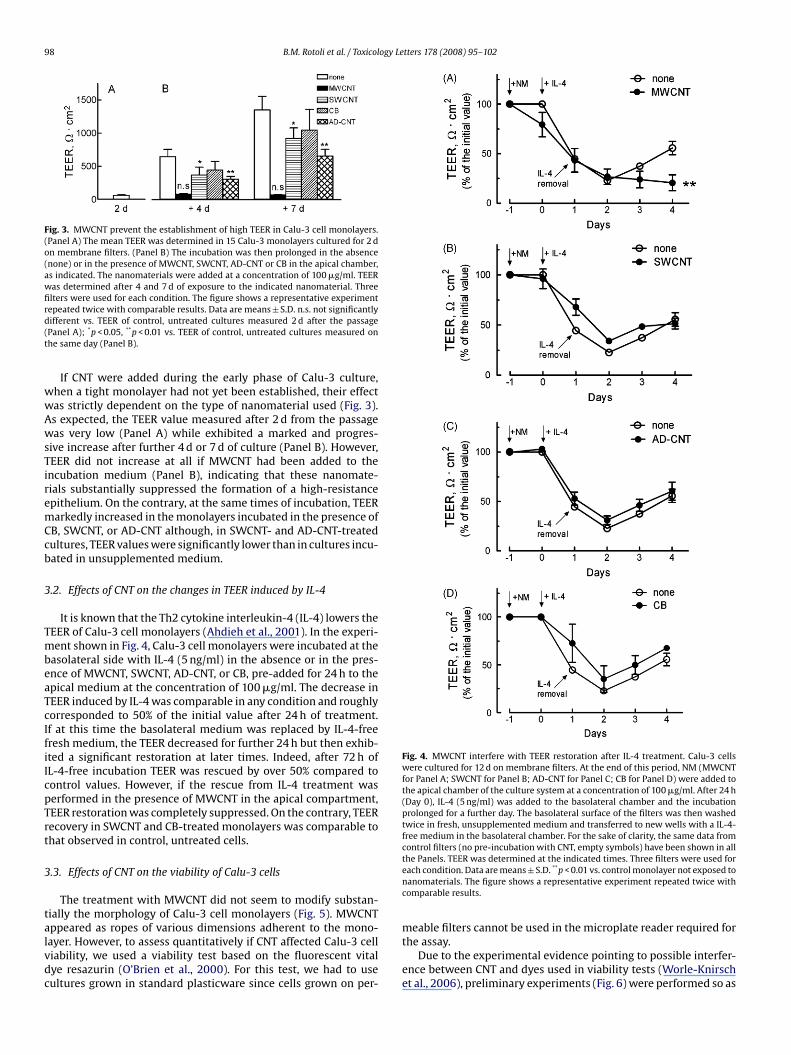
Fig. 3. MWCNT prevent the establishment of high TEER in Calu-3 cell monolayers.(Panel A) The mean TEER was determined in 15 Calu-3 monolayers cultured for 2 don membrane filters. (Panel B) The incubation was then prolonged in the absence(none) or in the presence of MWCNT, SWCNT, AD-CNT or CB in the apical chamber,as indicated. The nanomaterials were added at a concentration of 100 �g/ml. TEERwas determined after 4 and 7 d of exposure to the indicated nanomaterial. Threefilters were used for each condition. The figure shows a representative experimentrepeated twice with comparable results. Data are means ± S.D. n.s. not significantlydifferent vs. TEER of control, untreated cultures measured 2 d after the passage(Panel A); *p < 0.05, **p < 0.01 vs. TEER of control, untreated cultures measured onthe same day (Panel B).
If CNT were added during the early phase of Calu-3 culture,when a tight monolayer had not yet been established, their effectwas strictly dependent on the type of nanomaterial used (Fig. 3).As expected, the TEER value measured after 2 d from the passagewas very low (Panel A) while exhibited a marked and progres-sive increase after further 4 d or 7 d of culture (Panel B). However,TEER did not increase at all if MWCNT had been added to theincubation medium (Panel B), indicating that these nanomate-rials substantially suppressed the formation of a high-resistanceepithelium. On the contrary, at the same times of incubation, TEERmarkedly increased in the monolayers incubated in the presence ofCB, SWCNT, or AD-CNT although, in SWCNT- and AD-CNT-treatedcultures, TEER values were significantly lower than in cultures incu-bated in unsupplemented medium.
3.2. Effects of CNT on the changes in TEER induced by IL-4
It is known that the Th2 cytokine interleukin-4 (IL-4) lowers theTEER of Calu-3 cell monolayers (Ahdieh et al., 2001). In the experi-ment shown in Fig. 4, Calu-3 cell monolayers were incubated at thebasolateral side with IL-4 (5 ng/ml) in the absence or in the pres-ence of MWCNT, SWCNT, AD-CNT, or CB, pre-added for 24 h to theapical medium at the concentration of 100 �g/ml. The decrease in
TEER induced by IL-4 was comparable in any condition and roughlycorresponded to 50% of the initial value after 24 h of treatment.If at this time the basolateral medium was replaced by IL-4-freefresh medium, the TEER decreased for further 24 h but then exhib-ited a significant restoration at later times. Indeed, after 72 h ofIL-4-free incubation TEER was rescued by over 50% compared tocontrol values. However, if the rescue from IL-4 treatment wasperformed in the presence of MWCNT in the apical compartment,TEER restoration was completely suppressed. On the contrary, TEERrecovery in SWCNT and CB-treated monolayers was comparable tothat observed in control, untreated cells.3.3. Effects of CNT on the viability of Calu-3 cells
The treatment with MWCNT did not seem to modify substan-tially the morphology of Calu-3 cell monolayers (Fig. 5). MWCNTappeared as ropes of various dimensions adherent to the mono-layer. However, to assess quantitatively if CNT affected Calu-3 cellviability, we used a viability test based on the fluorescent vitaldye resazurin (O’Brien et al., 2000). For this test, we had to usecultures grown in standard plasticware since cells grown on per-
Fig. 4. MWCNT interfere with TEER restoration after IL-4 treatment. Calu-3 cellswere cultured for 12 d on membrane filters. At the end of this period, NM (MWCNTfor Panel A; SWCNT for Panel B; AD-CNT for Panel C; CB for Panel D) were added tothe apical chamber of the culture system at a concentration of 100 �g/ml. After 24 h(Day 0), IL-4 (5 ng/ml) was added to the basolateral chamber and the incubationprolonged for a further day. The basolateral surface of the filters was then washedtwice in fresh, unsupplemented medium and transferred to new wells with a IL-4-free medium in the basolateral chamber. For the sake of clarity, the same data fromcontrol filters (no pre-incubation with CNT, empty symbols) have been shown in allthe Panels. TEER was determined at the indicated times. Three filters were used foreach condition. Data are means ± S.D. **p < 0.01 vs. control monolayer not exposed tonanomaterials. The figure shows a representative experiment repeated twice withcomparable results.
meable filters cannot be used in the microplate reader required forthe assay.
Due to the experimental evidence pointing to possible interfer-ence between CNT and dyes used in viability tests (Worle-Knirschet al., 2006), preliminary experiments (Fig. 6) were performed so as

B.M. Rotoli et al. / Toxicology Le
Fig. 5. MWCNT do not affect grossly the morphology of Calu-3 cell monolayers.Confluent Calu-3 cell monolayers (TEER ∼ 1200 � cm2) were incubated for 4 d incomplete growth medium in the absence (upper Panel) or in the presence (lowerPanel) of 100 �g/ml MWCNT. Images are microphotographs of representative fields.×100.
to ascertain that the nanomaterials did not interfere with resazurin.The results obtained indicate that the incubation with MWCNT,SWCNT, or CB did not increase the low background level of resazurindetected after 1 h of incubation in the absence of cells (PanelA). Moreover, the addition of CNT or CB immediately before themeasurement did not quench grossly the high cell-dependent flu-orescence, although a significant decrease in the signal (∼15–20%)was detected when CB or MWCNT were present during fluorescencedetection (Panel B).
In Fig. 7 the effect of CNT on cell viability was examined in cul-tures of three different human cells: Calu-3 cells, endothelial cellsderived from umbilical veins (HUVECs), and THP-1 macrophages.
Fig. 6. Validation of resazurin viability test in the presence of CNT. (Panel A) Resazurin wof HUVECs and the incubation prolonged for 1 h at 37 ◦C. Empty bars, incubation in thenanomaterials at a concentration of 100 �g/ml. (Panel B) HUVECs were incubated for 1 h wiwere added at a concentration of 100 �g/ml to the incubation medium. Open bars, no adand data are means ± S.D. *p < 0.05, **p < 0.01 vs. control, NM-untreated cultures. The figur
tters 178 (2008) 95–102 99
The incubation with 100 �g/ml MWCNT markedly affected theviability of THP-1 and, even more markedly, of HUVEC cells. Incontrast, Calu-3 cells were not significantly affected by MWCNT.The different consequences of cell exposure to MWCNT were alsoconfirmed by direct microscopic observation, which showed thesubstantial integrity of the Calu-3 cell monolayer, the heavy dam-age of THP-1 cultures and the almost complete destruction ofthe HUVEC population. At the same concentration of 100 �g/ml,SWCNT had significant effects on the cell viability of HUVEC andTHP-1 cells but no effect on Calu-3 cells. CB produced no signif-icant effect on cell viability in HUVEC and Calu-3 cultures, whileTHP-1 cells exhibited only a moderate viability decrease.
4. Discussion
This study demonstrates for the first time that MWCNT impairthe barrier function of human airway epithelial cells, as indicatedby both a decrease in TEER and an increase of the paracellu-lar permeability of mannitol. The effect of MWCNT is time- anddose-dependent, with significant changes observed at relativelylow doses (5–10 �g/ml) while the exposure to 100 �g/ml MWCNTcaused a very marked TEER decrease.
The changes in paracellular permeability, induced by the expo-sure to MWCNT, are not due to artefacts, referable to detergentsused to solubilize CNT, since no detergent has been used in this
study. LPS contamination of the materials can be excluded, sincea LPS-inactivating protocol has been regularly adopted. Moreover,LPS per se has no significant effect on the TEER of Calu-3 monolay-ers up to a concentration of 1 �g/ml (B.M. Rotoli and O. Bussolati,unpublished observation).Although data on changes in cell viability caused by CNT arehighly variable, some cell types appear severely affected by thesenanomaterials (Cui et al., 2005; Bottini et al., 2006b; Zhang et al.,2007). Moreover, very recent data indicate that, after a few days ofexposure, genotoxic effects are detectable in epithelial cells treatedwith MWCNT (Muller et al., 2008). Hence, the possibility exists thatthe MWCNT effect on the paracellular permeability may be refer-able to toxic effects on Calu-3 cells. However, this does not seem tobe the case, since neither resazurin assay results nor direct observa-tion demonstrate evident changes in viability after a 48 h-exposureto MWCNT, when TEER alteration is already clearly detectable (seeFig. 1B). Rather, Calu-3 cells, cultured on plastic dishes, proved quiteresistant to the direct exposure to MWCNT (see Fig. 7), although itshould be stressed that their behaviour when grown differentiatedon membrane filters may be different. However, when evaluatedwith the same method, other cell types grown on plasticware, such
as added to 24-well dishes in the presence of HUVECs (left bar) or in the absenceabsence of nanomaterials. Solid bars, incubation in the presence of the indicatedth resazurin. Immediately before fluorescence reading, the indicated nanomaterialsdition of nanomaterials. For both Panels, three wells were used for each conditione shows representative experiments repeated twice with comparable results.

100 B.M. Rotoli et al. / Toxicology Letters 178 (2008) 95–102
P-1 hun stan
with rhown.
Fig. 7. Effect of carbon nanoparticles on cell viability. Calu-3 cells (upper Panel), TH(HUVEC, lower Panel) were cultured for 48 h (Calu-3) or 24 h (THP-1 and HUVECs) oconcentration of 100 �g/ml. At the end of the incubation, cell viability was assessedand MWCNT-treated cells (×100), taken just after the resazurin viability test, are scontrol, untreated cultures.
as human macrophages and endothelial cells, exhibit a marked tox-icity upon treatment with MWCNT.
Therefore, MWCNT seem to interfere specifically with theformation of tight junctional complexes, thus preventing the
development of cell polarization and the establishment of a high-resistance epithelial barrier. This hypothesis is consistent with theimpressive effect of MWCNT on the development of tight junc-tions in growing Calu-3 monolayers (see Fig. 3) as well as withthe behaviour exhibited by MWCNT-treated monolayers when tightjunctional efficiency is reversibly hindered by the Th2 cytokine IL-4 (Fig. 4). In this case, while IL-4 withdrawal leads to a fairly rapidrestoration of TEER in control cells, no recovery of resistance occursin monolayers exposed to MWCNT. The barrier dysfunction inducedby the cytokine has been attributed to a marked repression of theexpression of some components of tight junctions, such as ZO-1 andoccludin (Ahdieh et al., 2001). However, examples exist in whichmarked changes in paracellular permeability are not associatedwith evident modifications of tight junction proteins. For instance,asbestos microfibers raise the paracellular permeability of culturedairway epithelial cells in the absence of obvious changes in occludinexpression (Peterson et al., 1993; Peterson and Kirschbaum, 1998).Further experimental work is needed to assess if this is also the casefor MWCNT effect.What are the structural determinants of carbon nanotubesresponsible for their effect on paracellular permeability? As for
man macrophages (middle Panel) and human endothelial cells from umbilical veindard plasticware in the absence or in the presence of CB, SWCNT and MWCNT, at a
esazurin test (see Section 2). At the right of each panel, microphotographs of controlData are means ± S.D. of six independent determinations. **p < 0.01, ***p < 0.001 vs.
many other cases, the paradigm of a relative independence ofthe biological effect on the mass concentration of the material isconfirmed here. Indeed, although used at the same nominal con-centration of 100 �g/ml, SWCNT and AD-CNT had much smaller
effects than MWCNT and the amorphous nanomaterial CB had noeffect at all (Fig. 1A and B). Since several metals can contaminatecommercial CNT synthesized with the CVD method, the pertur-bation of tight junctions might be referable to the presence ofmetals, rather than to CNT themselves. According to the manufac-turer, MWCNT present a low level of metal contamination (<0.1%Fe), while SWCNT, mainly contaminated by Co and Mo (S. Bellucci,unpublished results), have a much lesser effect on paracellular per-meability. These data may point to a peculiar iron effect on tightjunctions that awaits further investigations.Since MWCNT have a much smaller surface/volume ratio thanSWCNT, their greater perturbing effect on paracellular permeabilityshould not be attributed to a wider interaction with the monolayersurface. Rather, they should be also endowed with a much highernumber of structural irregularities that could greatly increase theirchemical reactivity. This hypothesis appears interesting, given thatAD-CNT are characterized by a lower density of defects, comparedto commercial CNT. Indeed, the arc discharge and laser vaporiza-tions processes, which were the most common forms of nanotubeproduction in the past, typically result in nanotubes with lowstructural defects and, hence, outstanding physical properties. CVD

ogy Le
B.M. Rotoli et al. / Toxicolprocesses are more suitable for industrial applications, given thelarger deposition area and mass production involved. However, thisprocess has a major drawback, i.e. the reduced structural integrityof the resulting nanotubes. In particular, it has been pointed out bySalvetat et al. (2006) that MWCNT grown by arc discharge havemuch fewer structural defects and, hence, a better mechanicalbehaviour in terms of tensile modulus, compared to those grownby CVD.
Tight junctions are dynamic structures sensitive to environ-mental factors, such as toxic compounds, mediators, or bacterialproducts. In this contribution, we used the Th2 cytokine IL-4 toperturb reversibly Calu-3 monolayer permeability, as originallydescribed by Ahdieh et al. (2001), and to study the effects of CNTon TEER recovery. With this approach, we found that MWCNT com-pletely suppress TEER rescue, an effect that may have importantimplications. Indeed, it may point to a CNT-dependent increase inthe permeability of the epithelial barrier during the recovery fromenvironmental stress conditions associated with a Th2 response.Interestingly, while Th2 cytokines have been implied in lung fibro-sis (Jakubzick et al., 2004), fibrotic changes have been describedafter the exposure of rat lungs to SWCNT (Shvedova et al., 2005).Moreover, Th2 responses are implicated in the B cell activationand IgE production in allergic diseases. Thus, the results presentedhere may be consistent with the hypothesis, suggested by Murret al. (2005), that a relationship exists between asthma incidenceand indoor exposure to combustion emissions, e.g. natural gas orpropane cooking stoves, where MWCNT aggregates are present athigh number concentrations.
On the other hand, the barrier effects of IL-4 may indicate thatthe activation of a Th2-response during the exposure to MWCNTcould increase the likelihood of extra-pulmonary translocation ofthe nanomaterials from the airways to other body compartments.Nanoparticle translocation has been repeatedly investigated andrecently demonstrated in vivo at least for CB (Shimada et al., 2006).Geys et al. have recently used Calu-3 cell monolayers to assessthe translocation of fluorescent polystyrene particles (Geys et al.,2006, 2007). However, no translocation was observed with cellscultured on 0.4 �m-filters, the condition adopted in the presentstudy, while TEER increases after incubation with the nanomateri-als, possibly as a consequence of the progressive formation of tightjunctional complexes observed also in the present study. On thecontrary, acute and transient changes in TEER have been recentlyobserved in rat alveolar epithelial cells exposed to SWCNT, butnot to polystyrene nanoparticles (Yacobi et al., 2007). The sameauthors observe more evident and sustained changes in TEER after
exposure to chitosan or alginate coated, metal containing quantumdots as well as to ultrafine ambient particulates, which, however,are also rich in carbonaceous aggregates including MWCNT. Thus,the changes in barrier properties of airway cell monolayers seem aspecific effect of particular types of CNT, rather than an alterationgenerically associated to the exposure to carbon nanomaterials.Since the maintenance of the barrier efficiency is an importantfactor for the prevention of respiratory diseases and a determinantof the biological effects of toxic materials, CNT-induced changesmay have important functional consequences. Moreover, the pecu-liar effect of MWCNT on the barrier properties of IL-4-pretreatedepithelium suggests that the biological consequences of the expo-sure to these nanomaterials may depend on the simultaneouspresence of immunological changes, elicited by toxic or infectiousfactors.
Acknowledgments
This research has been funded by the Italian Ministry ofUniversity and Scientific Research (PRIN Grant No. 2006069554
tters 178 (2008) 95–102 101
“Interaction of novel nanoparticulate materials with biological sys-tems: testing models for human health risk assessment”) to S.B.and E.B. We thank Dr. F. Micciulla and Dr. A. Tiberia for the helpin AD-CNT production and characterization, and S. Tardito, for theadaptation of resazurin method. We gratefully acknowledge Prof.V. Dall’Asta and Prof. G.C. Gazzola for helpful suggestions and dis-cussions.
References
Ahdieh, M., Vandenbos, T., Youakim, A., 2001. Lung epithelial barrier function andwound healing are decreased by il-4 and il-13 and enhanced by ifn-gamma. Am.J. Physiol. Cell Physiol. 281, C2029–C2038.
Aijaz, S., Balda, M.S., Matter, K., 2006. Tight junctions: molecular architecture andfunction. Int. Rev. Cytol. 248, 261–298.
Banerjee, S., Kahn, M.G., Wong, S.S., 2003. Rational chemical strategies for carbonnanotube functionalization. Chemistry 9, 1898–1908.
Bellucci, S., Balasubramanian, C., Micciulla, F., Rinaldi, G., 2007a. Cnt composites foraerospace applications. J. Exp. Nanosci. 2, 193–206.
Bellucci, S., Balasubramanian, C., Micciulla, F., Tiberia, A., 2007b. Study of field emis-sion of multiwalled c nanotubes from arc discharge. J. Phys.: Condens. Matter19, 395014 (7 pp.).
Bottini, M., Balasubramanian, C., Dawson, M.I., Bergamaschi, A., Bellucci, S., Mustelin,T., 2006a. Isolation and characterization of fluorescent nanoparticles from pris-tine and oxidized electric arc-produced single-walled carbon nanotubes. J.Phys. Chem. B: Condens. Matter Mater. Surf. Interfaces Biophys. 110, 831–836.
Bottini, M., Bruckner, S., Nika, K., Bottini, N., Bellucci, S., Magrini, A., Bergamaschi,A., Mustelin, T., 2006b. Multi-walled carbon nanotubes induce t lymphocyteapoptosis. Toxicol. Lett. 160, 121–126.
Bottini, M., Magrini, A., Di Venere, A., Bellucci, S., Dawson, M.I., Rosato, N., Bergam-aschi, A., Mustelin, T., 2006c. Synthesis and characterization of supramolecularnanostructures of carbon nanotubes and ruthenium-complex luminophores. J.Nanosci. Nanotechnol. 6, 1381–1386.
Bottini, M., Tautz, L., Huynh, H., Monosov, E., Bottini, N., Dawson, M.I., Bellucci, S.,Mustelin, T., 2005. Covalent decoration of multi-walled carbon nanotubes withsilica nanoparticles. Chem. Commun. (Camb.), 758–760.
Cui, D., Tian, F., Ozkan, C.S., Wang, M., Gao, H., 2005. Effect of single wall carbonnanotubes on human hek293 cells. Toxicol. Lett. 155, 73–85.
De Nicola, M., Mirabile Gattia, D., Bellucci, S., De Bellis, G., Micciulla, F., Pastore, R.,Tiberia, A.C.C., D’Alessio, M., Vittori Antisari, M., et al., 2007. Effect of differentcarbon nanotubes on cell viability and proliferation. J. Phys.: Condens. Matter19, 395013 (7 pp.).
Donaldson, K., Aitken, R., Tran, L., Stone, V., Duffin, R., Forrest, G., Alexander, A.,2006. Carbon nanotubes: a review of their properties in relation to pulmonarytoxicology and workplace safety. Toxicol. Sci. 92, 5–22.
Finkbeiner, W.E., Carrier, S.D., Teresi, C.E., 1993. Reverse transcription-polymerasechain reaction (rt-pcr) phenotypic analysis of cell cultures of human trachealepithelium, tracheobronchial glands, and lung carcinomas. Am. J. Respir. CellMol. Biol. 9, 547–556.
Geys, J., Coenegrachts, L., Vercammen, J., Engelborghs, Y., Nemmar, A., Nemery, B.,Hoet, P.H., 2006. In vitro study of the pulmonary translocation of nanoparticles:a preliminary study. Toxicol. Lett. 160, 218–226.
Geys, J., Nemery, B., Hoet, P.H., 2007. Optimisation of culture conditions to developan in vitro pulmonary permeability model. Toxicol. In Vitro 21, 1215–1219.
Helland, A., Wick, P., Koehler, A., Schmid, K., Som, C., 2007. Reviewing the environ-mental and human health knowledge base of carbon nanotubes. Environ. HealthPerspect. 115, 1125–1131.
Iijima, S., Ichihashi, T., 1993. Single-shell carbon nanotubes of 1-nm diameter. Nature363, 603–605.
Jakubzick, C., Kunkel, S.L., Puri, R.K., Hogaboam, C.M., 2004. Therapeutic target-ing of il-4- and il-13-responsive cells in pulmonary fibrosis. Immunol. Res. 30,339–349.
Jia, G., Wang, H., Yan, L., Wang, X., Pei, R., Yan, T., Zhao, Y., Guo, X., 2005. Cytotoxi-city of carbon nanomaterials: single-wall nanotube, multi-wall nanotube, andfullerene. Environ. Sci. Technol. 39, 1378–1383.
Lam, C.W., James, J.T., McCluskey, R., Arepalli, S., Hunter, R.L., 2006. A review of carbonnanotube toxicity and assessment of potential occupational and environmentalhealth risks. Crit. Rev. Toxicol. 36, 189–217.
Lam, C.W., James, J.T., McCluskey, R., Hunter, R.L., 2004. Pulmonary toxicity of single-wall carbon nanotubes in mice 7 and 90 days after intratracheal instillation.Toxicol. Sci. 77, 126–134.
Landau, D., 2006. Epithelial paracellular proteins in health and disease. Curr. Opin.Nephrol. Hypertens. 15, 425–429.
Monteiro-Riviere, N.A., Nemanich, R.J., Inman, A.O., Wang, Y.Y., Riviere, J.E.,2005. Multi-walled carbon nanotube interactions with human epidermal ker-atinocytes. Toxicol. Lett. 155, 377–384.
Muller, J., Decordier, I., Hoet, P., Lombaert, N., Thomassen, L., Huaux, F., Lison, D.,Kirsch-Volders, M., 2008. Clastogenic and aneugenic effects of multi-wall carbonnanotubes in epithelial cells. Carcinogenesis 29, 427–433.
Muller, J., Huaux, F., Moreau, N., Misson, P., Heilier, J.F., Delos, M., Arras, M., Fonseca,A., Nagy, J.B., Lison, D., 2005. Respiratory toxicity of multi-wall carbon nanotubes.Toxicol. Appl. Pharmacol. 207, 221–231.

ogy Le
102 B.M. Rotoli et al. / ToxicolMurr, L.E., Garza, K.M., Soto, K.F., Carrasco, A., Powell, T.G., Ramirez, D.A., Guerrero,
P.A., Lopez, D.A., Venzor 3rd, J., 2005. Cytotoxicity assessment of some carbonnanotubes and related carbon nanoparticle aggregates and the implications foranthropogenic carbon nanotube aggregates in the environment. Int. J. Environ.Res. Public Health 2, 31–42.O’Brien, J., Wilson, I., Orton, T., Pognan, F., 2000. Investigation of the alamar blue(resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity.Eur. J. Biochem. 267, 5421–5426.
Oberdorster, G., Oberdorster, E., Oberdorster, J., 2005. Nanotoxicology: an emergingdiscipline evolving from studies of ultrafine particles. Environ. Health Perspect.113, 823–839.
Peterson, M.W., Kirschbaum, J., 1998. Asbestos-induced lung epithelial permeability:potential role of nonoxidant pathways. Am. J. Physiol. 275, L262–L268.
Peterson, M.W., Walter, M.E., Gross, T.J., 1993. Asbestos directly increases lungepithelial permeability. Am. J. Physiol. 265, L308–L317.
Reed, C.E., Kita, H., 2004. The role of protease activation of inflammation in allergicrespiratory diseases. J. Allergy Clin. Immunol. 114, 997–1008 (quiz 1009).
Rotoli, B.M., Bussolati, O., Dall’Asta, V., Orlandini, G., Gatti, R., Gazzola, G.C., 2000.Secretin increases the paracellular permeability of capan-1 pancreatic duct cells.Cell Physiol. Biochem. 10, 13–25.
Sakagami, M., 2006. In vivo, in vitro and ex vivo models to assess pulmonary absorp-tion and disposition of inhaled therapeutics for systemic delivery. Adv. DrugDeliv. Rev. 58, 1030–1060.
Salvetat, J.P., Bhattacharyya, S., Pipes, R.B., 2006. Progress on mechanics of carbonnanotubes and derived materials. J. Nanosci. Nanotechnol. 6, 1857–1882.
Schneeberger, E.E., Lynch, R.D., 2004. The tight junction: a multifunctional complex.Am. J. Physiol. Cell Physiol. 286, C1213–C1228.
Shimada, A., Kawamura, N., Okajima, M., Kaewamatawong, T., Inoue, H., Morita,T., 2006. Translocation pathway of the intratracheally instilled ultrafine par-
tters 178 (2008) 95–102
ticles from the lung into the blood circulation in the mouse. Toxicol. Pathol. 34,
949–957.Shin, K., Fogg, V.C., Margolis, B., 2006. Tight junctions and cell polarity. Annu. Rev.Cell Dev. Biol. 22, 207–235.
Shvedova, A.A., Castranova, V., Kisin, E.R., Schwegler-Berry, D., Murray, A.R., Gandels-man, V.Z., Maynard, A., Baron, P., 2003. Exposure to carbon nanotube material:assessment of nanotube cytotoxicity using human keratinocyte cells. J. Toxicol.Environ. Health A 66, 1909–1926.
Shvedova, A.A., Kisin, E.R., Mercer, R., Murray, A.R., Johnson, V.J., Potapovich,A.I., Tyurina, Y.Y., Gorelik, O., Arepalli, S., Schwegler-Berry, D., et al., 2005.Unusual inflammatory and fibrogenic pulmonary responses to single-walledcarbon nanotubes in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 289, L698–L708.
Visigalli, R., Barilli, A., Bussolati, O., Sala, R., Gazzola, G.C., Parolari, A., Tremoli, E.,Simon, A., Closs, E.I., Dall’asta, V., 2007. Rapamycin stimulates arginine influxthrough cat2 transporters in human endothelial cells. Biochim. Biophys Acta1768, 1479–1487.
Warheit, D.B., Laurence, B.R., Reed, K.L., Roach, D.H., Reynolds, G.A., Webb, T.R., 2004.Comparative pulmonary toxicity assessment of single-wall carbon nanotubes inrats. Toxicol. Sci. 77, 117–125.
Worle-Knirsch, J.M., Pulskamp, K., Krug, H.F., 2006. Oops they did it again!Carbon nanotubes hoax scientists in viability assays. Nano Lett. 6, 1261–1268.
Yacobi, N.R., Phuleria, H.C., Demaio, L., Liang, C.H., Peng, C.A., Sioutas, C., Borok, Z.,Kim, K.J., Crandall, E.D., 2007. Nanoparticle effects on rat alveolar epithelial cellmonolayer barrier properties. Toxicol. In Vitro 21, 1373–1381.
Zhang, L.W., Zeng, L., Barron, A.R., Monteiro-Riviere, N.A., 2007. Biological inter-actions of functionalized single-wall carbon nanotubes in human epidermalkeratinocytes. Int. J. Toxicol. 26, 103–113.
Copyright © 2022 FDOKUMEN




















