
Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and...
12
Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures Krisztina O ¨ tvo ¨s 1 , Taras P. Pasternak 2,† , Pa ´ l Miskolczi 2 , Mo ´ nika Domoki 1 , Dulguun Dorjgotov 1 , Attila Sz} ucs 1 , Sa ´ ndor Bottka 2 , De ´ nes Dudits 2 and Attila Fehe ´r 1,* 1 Laboratory of Functional Cell Biology, and 2 Laboratory of Cell Division and Differentiation, Institute of Plant Biology, Biological Research Centre, Hungarian Academy of Sciences, Temesva ´ ri krt. 62, H-6726, Szeged, Hungary Received 3 May 2005; revised 15 June 2005; accepted 17 June 2005. * For correspondence (fax 36 62 433 434; e-mail [email protected]). † Present address: Institute of Biologie II, Ludwig-Albert University, Sonnerster 5, 79104 Freiburg, Germany. Summary It is now well established that nitric oxide (NO) serves as a signaling molecule in plant cells. In this paper experimental data are presented which indicate that NO can stimulate the activation of cell division and embryogenic cell formation in leaf protoplast-derived cells of alfalfa in the presence of auxin. It was found that various NO-releasing compounds promoted auxin-dependent division (as shown by incorporation of bromodeoxyuridine) of leaf protoplast-derived alfalfa cells. In contrast, application of NO scavenger or NO synthesis inhibitor inhibited the same process. Both the promotion and the inhibition of cell cycle activation correlated with the amount and activity of the cognate alfalfa p34 cdc2 protein Medsa;CDKA;1,2. The effect of L-NG-monomethyl-L-arginine (L-NMMA) was transient, and protoplast-derived cells spending more than 3 days in culture become insensitive to the inhibitor as far as cell cycle progression was concerned. L-NMMA had no effect on the cell cycle parameters of cycling suspension-cultured cells, but had a moderate transient inhibitory effect on cells re-entering the cell cycle following phosphate starvation. Cycling cultured cells, however, could respond to NO, as indicated by the sodium nitroprusside (SNP)- and 2-phenyl-4,4,5,5- tetramethylimidazoline-1-oxyl-3-oxide (PTIO)-dependent accumulation of the ferritin protein. Based on these observations, it is hypothesized that L-NMMA-sensitive generation of NO is involved in the activation, but not the progression of the plant cell division cycle. In addition, SNP promoted and L-NMMA delayed the exogenous auxin [2,4-dichlorophenoxyacetic acid (2,4-D)] concentration-dependent formation of embryogenic cell clusters expressing the MsSERK1 gene; this further supports a link between auxin- and NO-dependent signaling pathways in plant cells. Keywords: nitrogen monoxide, bromodeoxyuridine (BrdU) incorporation, Medicago sativa, protoplast, somatic embryogenesis, 2,4-dichlorophenoxyacetic acid. Introduction Nitric oxide (NO) is a small, water- and lipid-soluble free radical known to act as a biological messenger in animals. NO ultimately exerts its biological effects by reacting, either directly or through other reactive nitrogen interme- diates, with a variety of targets such as heme groups, Fe–S or Zn–S clusters, sulfhydryl groups or various other chemical substrates (Cooper, 1999; Wink and Mitchell, 1998). That is why both metal- and thiol-containing proteins are target sites for NO, and as such, are very important cellular vehicles for the orchestration of the biological functions of NO. Among these cellular target proteins are cellular messengers, ion channels, enzymes, receptors and transcription factors (Bogdan, 2001). The diversity of potential targets is reflected by the large number of different cellular systems that use NO as a signal mediator. In mammals, NO was shown to be ª 2005 Blackwell Publishing Ltd 849 The Plant Journal (2005) 43, 849–860 doi: 10.1111/j.1365-313X.2005.02494.x
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and...

Nitric oxide is required for, and promotes auxin-mediatedactivation of, cell division and embryogenic cell formation butdoes not influence cell cycle progression in alfalfa cell cultures
Krisztina Otvos1, Taras P. Pasternak2,†, Pal Miskolczi2, Monika Domoki1, Dulguun Dorjgotov1, Attila Sz}ucs1, Sandor Bottka2,
Denes Dudits2 and Attila Feher1,*
1Laboratory of Functional Cell Biology, and2Laboratory of Cell Division and Differentiation, Institute of Plant Biology, Biological Research Centre, Hungarian Academy of
Sciences, Temesvari krt. 62, H-6726, Szeged, Hungary
Received 3 May 2005; revised 15 June 2005; accepted 17 June 2005.*For correspondence (fax 36 62 433 434; e-mail [email protected]).†Present address: Institute of Biologie II, Ludwig-Albert University, Sonnerster 5, 79104 Freiburg, Germany.
Summary
It is now well established that nitric oxide (NO) serves as a signaling molecule in plant cells. In this paper
experimental data are presented which indicate that NO can stimulate the activation of cell division and
embryogenic cell formation in leaf protoplast-derived cells of alfalfa in the presence of auxin. It was found that
various NO-releasing compounds promoted auxin-dependent division (as shown by incorporation of
bromodeoxyuridine) of leaf protoplast-derived alfalfa cells. In contrast, application of NO scavenger or NO
synthesis inhibitor inhibited the same process. Both the promotion and the inhibition of cell cycle activation
correlated with the amount and activity of the cognate alfalfa p34cdc2 protein Medsa;CDKA;1,2. The effect of
L-NG-monomethyl-L-arginine (L-NMMA) was transient, and protoplast-derived cells spending more than
3 days in culture become insensitive to the inhibitor as far as cell cycle progression was concerned. L-NMMA
had no effect on the cell cycle parameters of cycling suspension-cultured cells, but had a moderate transient
inhibitory effect on cells re-entering the cell cycle following phosphate starvation. Cycling cultured cells,
however, could respond to NO, as indicated by the sodium nitroprusside (SNP)- and 2-phenyl-4,4,5,5-
tetramethylimidazoline-1-oxyl-3-oxide (PTIO)-dependent accumulation of the ferritin protein. Based on these
observations, it is hypothesized that L-NMMA-sensitive generation of NO is involved in the activation, but not
the progression of the plant cell division cycle. In addition, SNP promoted and L-NMMAdelayed the exogenous
auxin [2,4-dichlorophenoxyacetic acid (2,4-D)] concentration-dependent formation of embryogenic cell
clusters expressing the MsSERK1 gene; this further supports a link between auxin- and NO-dependent
signaling pathways in plant cells.
Keywords: nitrogen monoxide, bromodeoxyuridine (BrdU) incorporation, Medicago sativa, protoplast,
somatic embryogenesis, 2,4-dichlorophenoxyacetic acid.
Introduction
Nitric oxide (NO) is a small, water- and lipid-soluble free
radical known to act as a biological messenger in animals.
NO ultimately exerts its biological effects by reacting,
either directly or through other reactive nitrogen interme-
diates, with a variety of targets such as heme groups, Fe–S
or Zn–S clusters, sulfhydryl groups or various other
chemical substrates (Cooper, 1999; Wink and Mitchell,
1998). That is why both metal- and thiol-containing
proteins are target sites for NO, and as such, are very
important cellular vehicles for the orchestration of the
biological functions of NO. Among these cellular target
proteins are cellular messengers, ion channels, enzymes,
receptors and transcription factors (Bogdan, 2001). The
diversity of potential targets is reflected by the large
number of different cellular systems that use NO as a
signal mediator. In mammals, NO was shown to be
ª 2005 Blackwell Publishing Ltd 849
The Plant Journal (2005) 43, 849–860 doi: 10.1111/j.1365-313X.2005.02494.x

involved in signal transduction pathways controlling,
among others, responses to infection, apoptosis, cell pro-
liferation, differentiation and fertilization (Kuo et al., 2000;
Peunova et al., 1996; Schmidt and Walter, 1994).
In mammals, formation of NO mainly relies on the activity
of the enzyme nitric oxide synthase (NOS; EC 1.14.13.39).
NOS enzymes catalyze the formation of NO from L-arginine,
which undergoes oxidation to citrulline. In human cells,
three NOS isoforms have been identified with altered tissue
specificity and Ca2þ dependence (Wang and Marsden, 1995).
Numerous experiments have demonstrated the generation
and presence of NO within plant cells and tissues (Beligni
and Lamattina, 2001; Durner and Klessig, 1999; Lamattina
et al., 2003). Although the Arabidopsis genome does not
contain genes coding for the plant homologs of the mam-
malian NOS enzymes, a plant protein with significant
biochemical similarity to animal NOS has recently been
identified by reverse genetics (Guo et al., 2003). Involvement
of this enzyme in hormonal signaling has also been dem-
onstrated (Guo et al., 2003).
Although NOS-like enzymes have only recently been
discovered in plants, considerable amounts of experimental
data had already been accumulated on the role of NO as a
signaling molecule in plant defense mechanisms (Delle-
donne et al., 1998; Durner and Klessig, 1999) as well as in
abiotic stress responses (Gould et al., 2003; Mackerness
et al., 2001; Mata and Lamattina, 2001). In addition, several
observations indicate that NO is also involved in the
regulation of plant growth and development under normal
growth conditions. It was shown, for example, that NO
affects germination (Beligni and Lamattina, 2000; Leshem
and Pinchasov, 2000; Seregelyes et al., 2003), leaf expansion
(Zhang et al., 2003), senescence (Leshem and Haramaty,
1996), programmed cell death (Clarke et al., 2000; Durzan
and Pedroso, 2002; de Pinto et al., 2002) and organogenesis
(Correa-Aragunde et al., 2004; Pagnussat et al., 2002). Con-
cerning its possible methods of action, NO was demonstra-
ted to modulate signal transduction pathways in plant cells
by affecting the level of secondary messenger molecules
such as cyclic guanosine monophosphate (cGMP), cyclic
ADP-ribose (cADPR) or Ca2þ (Correa-Aragunde et al., 2004;
Durner and Klessig, 1999; Durner et al., 1998; Pagnussat
et al., 2003, 2004) and activating kinase cascades (Pagnussat
et al., 2004).
The interaction of NO-mediated signaling with the regu-
lation of cell division and differentiation in mammalian cells
is well established (Peunova et al., 1996). Nitric oxide has
been shown to interact with cell cycle regulation at various
levels, resulting in blocked cell division (Guo et al., 1998;
Ishida et al., 1997; Pervin et al., 2001; Takagi et al., 1994). In
plant cells, however, there are hardly any data on either the
direct or indirect involvement of NO in signal transduction
events related to the cell cycle or cell division (Pagnussat
et al., 2004).
Here, experiments are reported which indicate that NO
may influence entry into the cell cycle but not cell cycle
progression in cultured plant cells. In addition, the involve-
ment of NO in the formation of embryogenic cells from leaf
protoplasts is also demonstrated.
Results
Frequency of incorporation of bromodeoxyuridine (BrdU)
into cells derived from leaf protoplast is affected by NO
To investigate the possible effects of NO on the division of
protoplast-derived cells, the frequency of incorporation of
bromodeoxyuridine (BrdU) into nuclear DNA was deter-
mined in cultures grown in the presence of NO-releasing
compounds [(�)-(E)-ethyl-2-[(E)-hydroxyimino]-5-nitro-3-
hexeneamide (NOR-3), 3-morpholinosydnonimine (SIN-1),
N-acetyl-3-(nitrosothio)-D-valine (SNAP), sodium nitroprus-
side (SNP), 4-phenyl-3-furoxancarbonitrile (furoxan)], the
NO scavenger 2-phenyl-4,4,5,5-tetramethylimidazoline-1-
oxyl-3-oxide (PTIO) and the NOS inhibitor L-NG-mono-
methyl-L-arginine (L-NMMA). The same concentrations were
used as accepted for the treatment of animal cells. In
Figure 1 it is demonstrated that while the NO-donor com-
pounds increased the number of cells incorporating BrdU
into their nuclei during a 24-h period, both the NOS inhibitor
and the NO scavenger inhibited this process.
It can also be seen that the effect of the NO-releasing
compounds was concentration dependent: 10 lM concen-
trations promoted but 100 lM concentrations inhibited the
increase in S-phase frequency. NOR-3 at a concentration of
100 lM and furoxan at 10 and 100 lM were even deleterious
to the cells (data not shown).
Figure 1. Effect of NO donors (NOR-3, SIN-1, SNAP, SNP), scavenger (PTIO)
and inhibitor (L-NMMA) on relative frequency of incorporation of BrdU into
the nuclei of leaf protoplast-derived alfalfa cells. The cells were grown in the
presence of the indicated concentrations of 2,4-D and drug from the
beginning of culture. BrdU was applied to protoplast-derived cells for 24 h
at the time of entering into the S-phase (checked by flow cytometry). Relative
values in comparison with cells grown in the presence of 1 lM 2,4-D (100%)
are shown as the average of three repeats with standard deviation. As a
positive control we used 10 lM 2,4-D in the medium.
850 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860
Attempts have also been made to determine cellular
NO levels in control and treated cells using a micro-
scopic approach based on the NO-sensitive fluorescent dye
4,5-diaminofluorescein diacetate (DAF-2DA) (Kojima et al.,
1998), allowing the detection of NO levels in individual
cells. DAF2-DA is a cell-permeable molecule that does not
fluoresce until it reacts with NO. Although we could
observe a DAF-2DA-dependent fluorescent signal in a
fraction of protoplast-derived cells, its NO-insensitive ana-
log, 4-aminofluorescein diacetate, exhibited very bright
fluorescence in all cells and this finding brought into
question the specificity of DAF-2DA (Beligni et al., 2002)
(see Figure S1).
The NO-donor sodium nitroprusside (SNP) promotes auxin
concentration-dependent division and embryogenic cell
formation in alfalfa leaf protoplast cultures
As shown in Figure 1, the effect of NO donors on the S-phase
frequency of protoplast-derived cells is very similar to the
effect of increased 2,4-dichlorophenoxyacetic acid (2,4-D)
concentration. In order to gain further insight into the effect
of NO donors and 2,4-D on leaf protoplast-derived cells,
various cellular parameters have been determined on cells
grown in the absence and presence of 10 lM SNP at various
2,4-D concentrations (0, 0.22 or 1 lM). As a control we used
‘inactive’ SNP (iSNP), pre-exposed to direct light for 1 day in
order to ensure complete decomposition preceding its
application to the protoplasts, at the same concentration.
This concentration of iSNP had no significant effect on the
development of protoplast-derived cells in comparison with
untreated cultures (data not shown).
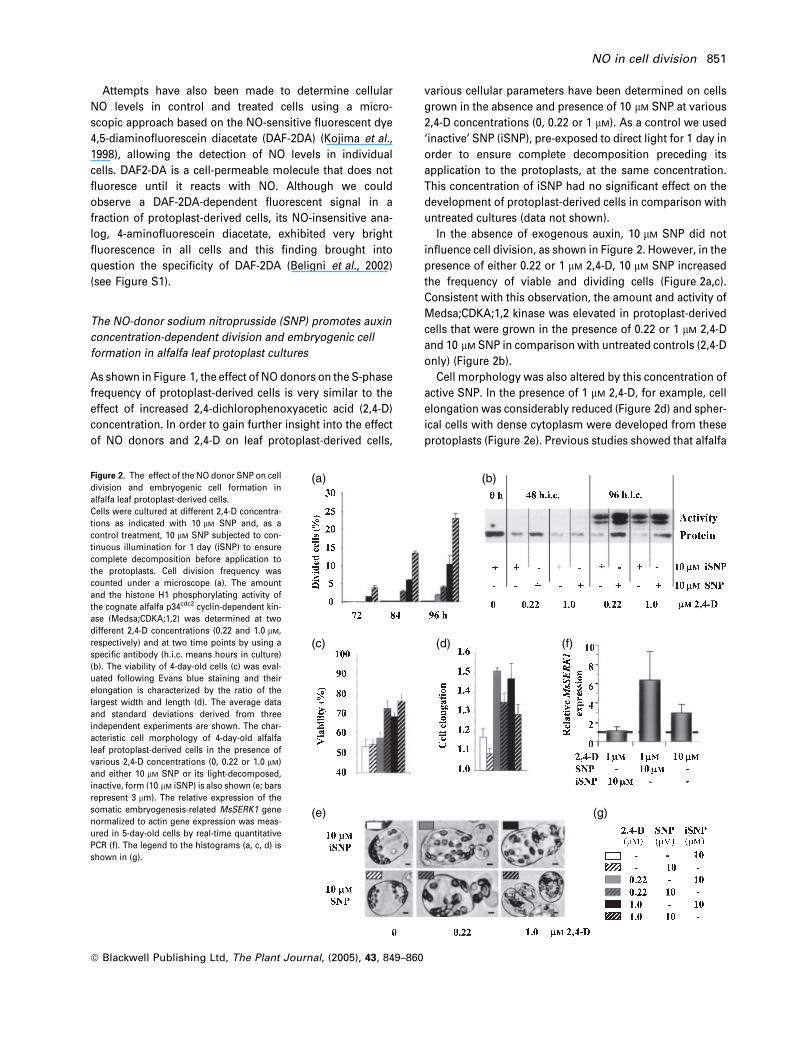
In the absence of exogenous auxin, 10 lM SNP did not
influence cell division, as shown in Figure 2. However, in the
presence of either 0.22 or 1 lM 2,4-D, 10 lM SNP increased
the frequency of viable and dividing cells (Figure 2a,c).
Consistent with this observation, the amount and activity of
Medsa;CDKA;1,2 kinase was elevated in protoplast-derived
cells that were grown in the presence of 0.22 or 1 lM 2,4-D
and 10 lM SNP in comparison with untreated controls (2,4-D
only) (Figure 2b).
Cell morphology was also altered by this concentration of
active SNP. In the presence of 1 lM 2,4-D, for example, cell
elongation was considerably reduced (Figure 2d) and spher-
ical cells with dense cytoplasm were developed from these
protoplasts (Figure 2e). Previous studies showed that alfalfa
(a) (b)
(c) (d)
(e) (g)
(f)
Figure 2. The effect of the NO donor SNP on cell
division and embryogenic cell formation in
alfalfa leaf protoplast-derived cells.
Cells were cultured at different 2,4-D concentra-
tions as indicated with 10 lM SNP and, as a
control treatment, 10 lM SNP subjected to con-
tinuous illumination for 1 day (iSNP) to ensure
complete decomposition before application to
the protoplasts. Cell division frequency was
counted under a microscope (a). The amount
and the histone H1 phosphorylating activity of
the cognate alfalfa p34cdc2 cyclin-dependent kin-
ase (Medsa;CDKA;1,2) was determined at two
different 2,4-D concentrations (0.22 and 1.0 lM,
respectively) and at two time points by using a
specific antibody (h.i.c. means hours in culture)
(b). The viability of 4-day-old cells (c) was eval-
uated following Evans blue staining and their
elongation is characterized by the ratio of the
largest width and length (d). The average data
and standard deviations derived from three
independent experiments are shown. The char-
acteristic cell morphology of 4-day-old alfalfa
leaf protoplast-derived cells in the presence of
various 2,4-D concentrations (0, 0.22 or 1.0 lM)
and either 10 lM SNP or its light-decomposed,
inactive, form (10 lM iSNP) is also shown (e; bars
represent 3 lm). The relative expression of the
somatic embryogenesis-related MsSERK1 gene
normalized to actin gene expression was meas-
ured in 5-day-old cells by real-time quantitative
PCR (f). The legend to the histograms (a, c, d) is
shown in (g).
NO in cell division 851
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

leaf protoplasts treated with a relatively high dose (10 lM) of
the synthetic auxin 2,4-D form small, isodiametric, cytoplas-
mically dense and asymmetrically dividing cells expressing
embryogenic competence (Bogre et al., 1990; Dudits et al.,
1991; Pasternak et al., 2002). As can be seen in Figure 2(f),
consistent with the ‘embryogenic morphology’ of the cell
colonies, the relative expression of the MsSERK1 gene,
coding for the somatic embryogenesis receptor kinase
(SERK) protein, a putative marker of the embryogenic
capability of alfalfa cells (Nolan et al., 2003), was consider-
ably higher in the 10 lM 2,4-D-treated or 1 lM 2,4-D plus
10 lM SNP-treated cells in comparison with control cells
(1 lM 2,4-D-grown). To follow the fate of protoplast-derived
cells, they were cultured in the presence of 1 or 10 lM 2,4-D
with or without 10 lM SNP or 1 mM L-NMMA, respectively,
for 5 days and than washed and embedded in an agarose-
solidified ‘low-auxin’ [0.36 lM indoleacetic acid (IAA)] med-
ium. Under these conditions, the cells initially cultured in the
presence of 10 lM 2,4-D or 1 lM 2,4-D plus 10 lM SNP could
develop further into pro-embryogenic cell clusters consist-
ing of small, isodiametric cells with dense cytoplasm (see
Figure S2). In parallel cultures with cells initially grown in
the presence of 1 lM 2,4-D or 1 lM 2,4-D plus 10 lM iSNP,
viability was considerably decreased and mostly clusters of
enlarged, elongated cells developed from the protoplast-
derived cells (see Figure S2). However, a few embryogenic-
type cell clusters could be observed in these types of
cultures as well (data not shown).
L-NMMA inhibits the activation but not the progression of
the cell division cycle and delays formation of embryogenic
cells from leaf protoplast
To further investigate the possible involvement of NO in the
formation and division of protoplast-derived cells, freshly
isolated and cultured leaf protoplasts were treated in the
presence of 1 lM 2,4-D with PTIO, a NO scavenger, or
L-NMMA, a known inhibitor of NO-generating enzymes in
animal and plant cells. Figure 3 shows that the frequency of
cells incorporating BrdU into their nuclei, as well as the
amount and the histone H1 phosphorylating activity of the
cognate alfalfa p34cdc2 kinase Medsa;CDKA;1,2, was signifi-
cantly decreased during the third day of culture in the pres-
ence of either 200 lM PTIO or 1 mM L-NMMA (Figure 3a).
The inhibitory effect of L-NMMA could be reverted by
parallel application of low (1–10 lM) concentrations of the
NO donor SNP, indicating that the L-NMMA-mediated
decrease in S-phase frequency and cyclin-dependent kinase
(CDK) activity were indeed NO dependent (Figure 3a).
Interestingly, the inhibitory effect of L-NMMA on the accu-
mulation and activity of CDK was restricted to the first 3 days
of culture (Figure 3b). These were reduced at 36 and 72 but
not at 96 h of culture. At 96 h, CDK activity was even slightly
increased in comparison with control cells.
To gain a more detailed view of how the effect of L-NMMA
on CDK activity depends on the culture period, 24-h-shifted
48-h-long pulses of the inhibitor were applied to protoplast-
derived cells over a 7-day period (Figure 3c). As a parameter
of cell cycle progression, the frequency of S-phase cells was
determined by incorporation of BrdU into nuclear DNA
during the second half of the L-NMMA pulse. The activity of
the Medsa;CDKA;1,2 kinase was also determined at the end
of the 48-h period. It could be observed that if L-NMMA was
present in the medium before 72 h of culture, there was a
significant inhibition of BrdU incorporation as well as
Medsa;CDK;A1,2 activity. The presence of the inhibitor in
the medium from 72–120 h resulted in increased activity of
the CDK. This might be the consequence of a release of cells
from blocked cell cycle progression which might result in a
synchronized re-entry to the cell cycle. If the inhibitor was
added to the cells 96 h after protoplast isolation, or later, no
effect could be observed.
It could also be observed that application of L-NMMA
decreased the frequency of protoplast-derived cells comple-
ting their first cell division by the fourth day of culture (to
approximately 25–30% as compared with control; data not
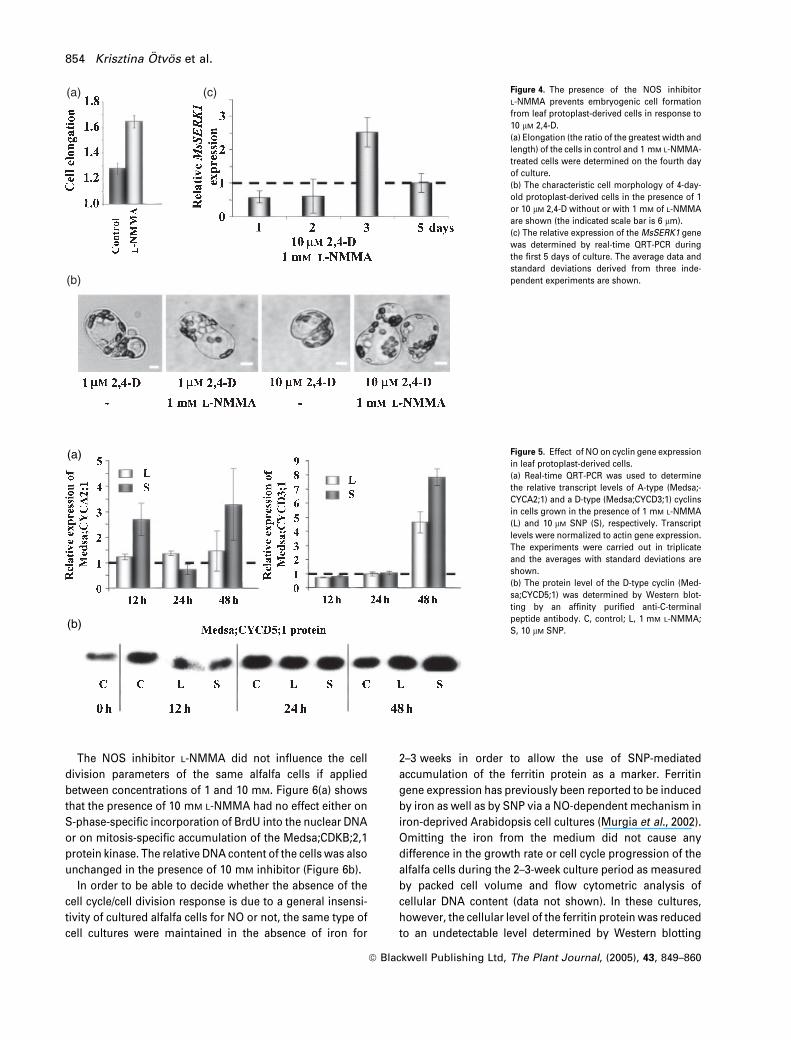
shown), but promoted their elongation (Figure 4a). It was
also found that 1 mM of the NOS inhibitor L-NMMA inhibited
the formation of the embryogenic competent cell type in the
presence of 10 lM 2,4-D (Figure 4b). The effect of L-NMMA on
formation of embryogenic cells was transient, as indicated by
the expression of the MsSERK1 gene. The relative MsSERK1
transcript level was lower in cells treated with L-NMMA than
in control cells during the first 2 days of culture but signifi-
cantly increased on the third day and returned to a normal
level by the fifth day (Figure 4c). In agreement with this
expression pattern, L-NMMA-treated cells could indeed
develop into embryogenic cell clusters in the presence of
10 lM 2,4-D (see Figure S2), but the development of pro-
embryogenic cell clusters was slightly delayed (embryogenic
cell clusters were smaller on average, data not shown).
The effect of NO on cyclin gene expression in
protoplast-derived cells
In order to further study the effect of NO on the cell cycle
machinery, the expression of A- and D-type Medicago cyc-
lins, which have proposed roles during the early phases of
cell cycle entry (G0/G1 transition; Dahl et al., 1995; Meskiene
et al., 1995), have been tested in treated protoplasts during
the first 2 days of culture. As shown in Figure 5(a), the rel-
ative expression of the A-type cyclin Medsa;CYCA2;1 is sig-
nificantly increased by 10 lM SNP at 12 and 48 h of culture,
but drops back at 24 h. L-NMMA at 1 mM had no significant
effect on the expression of the same gene. The transcript
level of Medsa,CYCD3;1 was slightly decreased by both
treatments at 12 h and was returned to a normal level at 24 h
followed with a five- to eightfold increase at 48 h. Using a
852 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

specific antibody, the protein level of Medsa;CYCD5;1, a
strongly interacting cyclin partner of Medsa;CDKA;1,2 in the
yeast two-hybrid system (Meszaros et al., 2000), was also
checked (Figure 5b). Similarly to the Medsa;CYCD3;1 tran-
scription, the level of this cyclin D protein was also reduced
by both treatments at 12 h, reached the control level at 24 h
and increased by 48 h.
NO fails to interfere with cell cycle progression in
exponentially growing alfalfa cell suspensions
As our observations indicated that protoplast-derived cells
became insensitive to the action of L-NMMA on the
third day of culture when the majority of the cells were
progressing through their first cell division cycle, the
responses of continuously dividing de-differentiated cell
cultures to the same compounds were also investigated.
Cells of a fast growing suspension culture from the same
alfalfa genotype used for leaf protoplast isolation were
studied in these experiments.
The NO donor SNP, at concentrations of 10 or 100 lM, had
no effect on frequency of incorporation of BrdU or on the
mitotic accumulation of the protein Medsa;CDKB;2,1 in
exponentially growing suspension-cultured cells (Fig-
ure 6a). It has been previously demonstrated that the
Medsa;CDKB;2,1 protein accumulates preferentially during
the M-phase of the cell cycle (Magyar et al., 1997; Meszaros
et al., 2000).
(a)
(b)
(c)
Figure 3. The effect of the NOS inhibitor L-
NMMA on cell division of alfalfa leaf protoplast-
derived cells.
(a) Incorporation of BrdU and the amount and
activity of Medsa;CDKA;1,2 in 3-day-old proto-
plast-derived cells treated with the NO scavenger
PTIO (200 lM) or the NOS inhibitor L-NMMA
(1 mM) without or together with the NO donor
SNP (1, 10 or 100 lM) from the beginning of the
protoplast culture.
(b) Changes in level and activity of Medsa;CD-
KA;1,2 protein during the first 4 days of proto-
plast culture in the presence of 1 mM L-NMMA.
(c) The effect of culture period-dependent appli-
cation of L-NMMA (1 mM) on cell cycle (S-phase)
progression and Medsa;CDKA;1,2 activity in
alfalfa leaf-protoplast-derived cells. 24-h-shifted
and 48-h-long pulses of 1 mM L-NMMA were
applied to alfalfa leaf protoplast-derived cells as
indicated in the figure. During the second 24-h
period of L-NMMA treatment, 30 lM BrdU was
incorporated into the medium. At the end of the
L-NMMA pulses, frequency of incorporation of
BrdU and histone H1 phosphorylation activity of
the immunoprecipitated Medsa;CDKA;1,2 kinase
complex were determined.
For BrdU incorporation, the average data and
standard deviations derived from three inde-
pendent experiments are shown. Protoplasts
were grown in the presence of 1 lM 2,4-D during
the presented experiments.
NO in cell division 853
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860
The NOS inhibitor L-NMMA did not influence the cell
division parameters of the same alfalfa cells if applied
between concentrations of 1 and 10 mM. Figure 6(a) shows
that the presence of 10 mM L-NMMA had no effect either on
S-phase-specific incorporation of BrdU into the nuclear DNA
or on mitosis-specific accumulation of the Medsa;CDKB;2,1
protein kinase. The relative DNA content of the cells was also
unchanged in the presence of 10 mM inhibitor (Figure 6b).
In order to be able to decide whether the absence of the
cell cycle/cell division response is due to a general insensi-
tivity of cultured alfalfa cells for NO or not, the same type of
cell cultures were maintained in the absence of iron for
2–3 weeks in order to allow the use of SNP-mediated
accumulation of the ferritin protein as a marker. Ferritin
gene expression has previously been reported to be induced
by iron as well as by SNP via a NO-dependent mechanism in
iron-deprived Arabidopsis cell cultures (Murgia et al., 2002).
Omitting the iron from the medium did not cause any
difference in the growth rate or cell cycle progression of the
alfalfa cells during the 2–3-week culture period as measured
by packed cell volume and flow cytometric analysis of
cellular DNA content (data not shown). In these cultures,
however, the cellular level of the ferritin protein was reduced
to an undetectable level determined by Western blotting
(a) (c)
(b)
Figure 4. The presence of the NOS inhibitor
L-NMMA prevents embryogenic cell formation
from leaf protoplast-derived cells in response to
10 lM 2,4-D.
(a) Elongation (the ratio of the greatest width and
length) of the cells in control and 1 mM L-NMMA-
treated cells were determined on the fourth day
of culture.
(b) The characteristic cell morphology of 4-day-
old protoplast-derived cells in the presence of 1
or 10 lM 2,4-D without or with 1 mM of L-NMMA
are shown (the indicated scale bar is 6 lm).
(c) The relative expression of the MsSERK1 gene
was determined by real-time QRT-PCR during
the first 5 days of culture. The average data and
standard deviations derived from three inde-
pendent experiments are shown.
(a)
(b)
Figure 5. Effect of NO on cyclin gene expression
in leaf protoplast-derived cells.
(a) Real-time QRT-PCR was used to determine
the relative transcript levels of A-type (Medsa;-
CYCA2;1) and a D-type (Medsa;CYCD3;1) cyclins
in cells grown in the presence of 1 mM L-NMMA
(L) and 10 lM SNP (S), respectively. Transcript
levels were normalized to actin gene expression.
The experiments were carried out in triplicate
and the averages with standard deviations are
shown.
(b) The protein level of the D-type cyclin (Med-
sa;CYCD5;1) was determined by Western blot-
ting by an affinity purified anti-C-terminal
peptide antibody. C, control; L, 1 mM L-NMMA;
S, 10 lM SNP.
854 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860
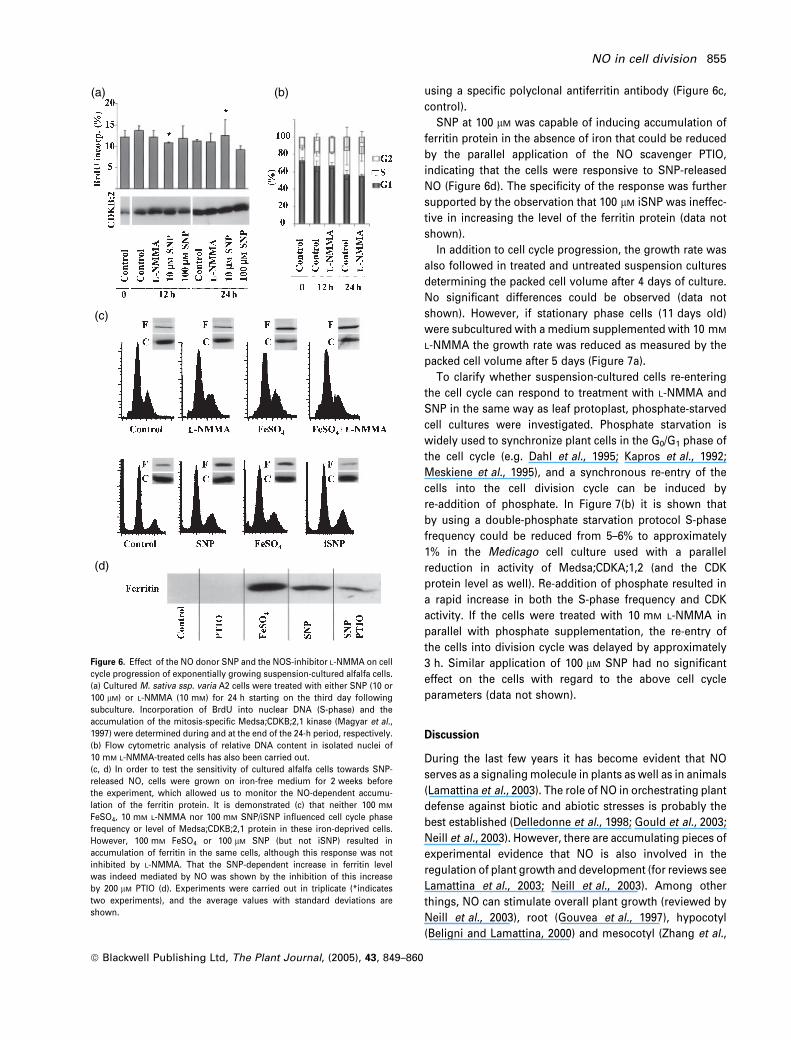
using a specific polyclonal antiferritin antibody (Figure 6c,
control).
SNP at 100 lM was capable of inducing accumulation of
ferritin protein in the absence of iron that could be reduced
by the parallel application of the NO scavenger PTIO,
indicating that the cells were responsive to SNP-released
NO (Figure 6d). The specificity of the response was further
supported by the observation that 100 lM iSNP was ineffec-
tive in increasing the level of the ferritin protein (data not
shown).
In addition to cell cycle progression, the growth rate was
also followed in treated and untreated suspension cultures
determining the packed cell volume after 4 days of culture.
No significant differences could be observed (data not
shown). However, if stationary phase cells (11 days old)
were subcultured with a medium supplemented with 10 mM
L-NMMA the growth rate was reduced as measured by the
packed cell volume after 5 days (Figure 7a).
To clarify whether suspension-cultured cells re-entering
the cell cycle can respond to treatment with L-NMMA and
SNP in the same way as leaf protoplast, phosphate-starved
cell cultures were investigated. Phosphate starvation is
widely used to synchronize plant cells in the G0/G1 phase of
the cell cycle (e.g. Dahl et al., 1995; Kapros et al., 1992;
Meskiene et al., 1995), and a synchronous re-entry of the
cells into the cell division cycle can be induced by
re-addition of phosphate. In Figure 7(b) it is shown that
by using a double-phosphate starvation protocol S-phase
frequency could be reduced from 5–6% to approximately
1% in the Medicago cell culture used with a parallel
reduction in activity of Medsa;CDKA;1,2 (and the CDK
protein level as well). Re-addition of phosphate resulted in
a rapid increase in both the S-phase frequency and CDK
activity. If the cells were treated with 10 mM L-NMMA in
parallel with phosphate supplementation, the re-entry of
the cells into division cycle was delayed by approximately
3 h. Similar application of 100 lM SNP had no significant
effect on the cells with regard to the above cell cycle
parameters (data not shown).
Discussion
During the last few years it has become evident that NO
serves as a signaling molecule in plants as well as in animals
(Lamattina et al., 2003). The role of NO in orchestrating plant
defense against biotic and abiotic stresses is probably the
best established (Delledonne et al., 1998; Gould et al., 2003;
Neill et al., 2003). However, there are accumulating pieces of
experimental evidence that NO is also involved in the
regulation of plant growth and development (for reviews see
Lamattina et al., 2003; Neill et al., 2003). Among other
things, NO can stimulate overall plant growth (reviewed by
Neill et al., 2003), root (Gouvea et al., 1997), hypocotyl
(Beligni and Lamattina, 2000) and mesocotyl (Zhang et al.,
(a)
(c)
(d)
(b)
Figure 6. Effect of the NO donor SNP and the NOS-inhibitor L-NMMA on cell
cycle progression of exponentially growing suspension-cultured alfalfa cells.
(a) Cultured M. sativa ssp. varia A2 cells were treated with either SNP (10 or
100 lM) or L-NMMA (10 mM) for 24 h starting on the third day following
subculture. Incorporation of BrdU into nuclear DNA (S-phase) and the
accumulation of the mitosis-specific Medsa;CDKB;2,1 kinase (Magyar et al.,
1997) were determined during and at the end of the 24-h period, respectively.
(b) Flow cytometric analysis of relative DNA content in isolated nuclei of
10 mM L-NMMA-treated cells has also been carried out.
(c, d) In order to test the sensitivity of cultured alfalfa cells towards SNP-
released NO, cells were grown on iron-free medium for 2 weeks before
the experiment, which allowed us to monitor the NO-dependent accumu-
lation of the ferritin protein. It is demonstrated (c) that neither 100 mM
FeSO4, 10 mM L-NMMA nor 100 mM SNP/iSNP influenced cell cycle phase
frequency or level of Medsa;CDKB;2,1 protein in these iron-deprived cells.
However, 100 mM FeSO4 or 100 lM SNP (but not iSNP) resulted in
accumulation of ferritin in the same cells, although this response was not
inhibited by L-NMMA. That the SNP-dependent increase in ferritin level
was indeed mediated by NO was shown by the inhibition of this increase
by 200 lM PTIO (d). Experiments were carried out in triplicate (*indicates
two experiments), and the average values with standard deviations are
shown.
NO in cell division 855
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

2003) elongation and induce the development of adventi-
tious roots in various plant species (Correa-Aragunde et al.,
2004; Pagnussat et al., 2002, 2003). These observations
indicate that NO may, directly or indirectly, affect cell elon-
gation and division, the two basic processes in plant
morphogenesis. There are no reports, however, on the
involvement of NO in the activation and/or the progression
of the cell division cycle in cultured plant cells.
NO is required for and promotes the activation but not the
progression of the cell division cycle in plants
Individual plant cells without cell walls, i.e. protoplasts,
isolated from leaf tissues and cultured in synthetic liquid
media are probably one of the best in vitro experimental
models for studying the activation of cell division in differ-
entiated plant cells. Although they are isolated using
stressful procedures, experimental evidence demonstrates
that properly handled protoplasts quickly recover in culture
and respond to hormonal as well as stress signals in the
same way as intact plant cells (see, for example, Pasternak
et al., 2002; for reviews Feher et al., 2005; Sheen, 2001).
As the S-phase frequency as well as the division rate of
alfalfa leaf protoplast-derived cells was decreased by the
NOS inhibitor L-NMMA and the NO scavenger PTIO and
increased by the NO donor SNP it is very likely that NO is
involved in the signaling cascade that leads to the de-
differentiation and division of leaf cells. That the observed
effects do indeed depend on NO is strengthened by the
observation that the L-NMMA-mediated inhibition of BrdU
incorporation as well as the activity of Medsa;CDKA;1,2
kinase could be reverted by application of SNP. SNP had a
concentration-dependent effect on the frequency of proto-
plast-derived cells entering the S-phase of the cell cycle. SNP
at concentrations of 1 and 10 lM promoted DNA replication
while SNP at 100 lM inhibited DNA replication. Concentra-
tion-dependent action of NO is well known in both plant and
animal cells. Activation of sea urchin egg cells by NO donors
is only possible in a narrow range of concentrations, and
higher concentrations even inhibit sperm cell-mediated
activation (Kuo et al., 2000). A high level of NO has been
reported to inhibit leaf expansion and plant growth whereas
low levels enhance it (Beligni and Lamattina, 2001; Lamat-
tina et al., 2003; Neill et al., 2003; Seregelyes et al., 2003).
During the process of de-differentiation, the loss of
differentiated functions, recovery from isolation stress, cell
wall synthesis and activation of the cell cycle machinery take
place in a serial, although overlapping, manner in the
protoplast-derived cells (for reviews Feher et al., 2003,
2005). It is not clear at what step NO may accelerate the
process. Shifted pulses of application of L-NMMA and
subsequent measurements of BrdU incorporation and CDK
activity indicate, however, that protoplast-derived cells more
than 3 days old are not responsive to the presence of the
NOS inhibitor. This may indicate that an L-NMMA-sensitive
step operates during the first 3 days of the alfalfa leaf
protoplast culture and this step probably precedes the entry
of the cells into the S-phase of the cell cycle.
In order to address this question, the expression of cyclin
genes previously reported to be activated before the S-
phase entry (Dahl et al., 1995) or even at the G0 to G1
transition (Meskiene et al., 1995), have been tested in the
presence of SNP and L-NMMA. The level of protein of a
further D-type cyclin, the strongest interactor with the
Medsa;CDKA;1,2 kinase in the yeast two-hybrid system
(Meszaros et al., 2000), has also been determined.
Based on the results, specific inhibition of cyclin tran-
scription or accumulation by L-NMMA could not be estab-
lished. The increased level of both cyclins in response to L-
NMMA at 48 h might indicate that the treated cells had
already entered into the division cycle. Interestingly, pro-
toplast-derived cells cultured in the presence of L-NMMA
Pac
ked
cell
volu
me
[ml/l
10m
l cul
ture
]
Con
trol
10 m
M
L-N
MM
A
Med
sa;C
DK
A;1
,2
2.01.81.61.41.21.00.80.6
0.20.0
0.4
201612
8
40
Activity
S phasefrequency
[%]
0
PO4– Control 10 mM
L-NMMA
3h 12h6h 3h 12h6h
Protein
(a) (b)
Figure 7. Effect of L-NMMA on cell division reactivation of suspension-cultured cells.
(a) Eleven-day-old stationary-phase cultures of M. ssp. variaA2 cells were subcultured in the absence (control) and in the presence of 10 mM L-NMMA and the packed
cell volume was measured following 5 days of further culture. The initial packed cell volume is indicated by the dashed line (0.95 ml of cells per 10 ml of culture.
(b) S-phase frequency (determined by flow cytometry) and the amount and histone H1 phosphorylating activity of the Medsa;CDKA;1,2 protein were determined in
suspension-cultured cells following phosphate starvation (see Experimental procedures) and phosphate readdition together with or without 10 mM L-NMMA.
856 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

also exhibited increased CDK activity and SERK gene
expression at around the third day of culture. It is
interesting to note that peaks of amount of endogenous
IAA and S-phase frequency could be detected on the
second and third days of culture, respectively, and embry-
ogenic competence is also established during this period
in these protoplast-derived cells (see Pasternak et al., 2000,
2002, for review Feher et al., 2005). It might be speculated
that L-NMMA does not block but slows down cell activation
that results in the synchronous accumulation of the cells in
the G1/S-phases. The synchrony seems to be lost after the
S-phase as the cells at this stage become insensitive to the
presence of L-NMMA. This may result in transiently higher
transcript/protein/activity levels of the investigated genes/
proteins during a certain period. A similar response could
be seen in the same type of culture when cell activation
was slowed down by the buffering of the pH of
the medium (Pasternak et al., 2002; T. P. Pasternak and
A. Feher, unpublished data).
To validate the above hypotheses as well as to determine
the exact timing and nature of the process(es) sensitive to
L-NMMA during the de-differentiation and division of leaf
cells requires further experimentation.
In agreement with the above findings described for leaf
protoplast-derived cells, L-NMMA did not inhibit cell cycle
progression or the growth rate of continuously dividing
suspension-cultured cells, but delayed re-entry of these cells
to the cell division cycle after phosphate starvation or after
the subculture of 11-day-old stationary phase cells. SNP at
concentrations of 10–100 lM had no effect on cell cycle
progression in suspension-cultured alfalfa cells, although
these cells could respond to NO as indicated by 100 lM SNP-
induced and PTIO-sensitive ferritin protein accumulation
(Murgia et al., 2002). This observation indicates that the
absence of the cell cycle response was not due to a general
insensitivity of suspension-cultured cells towards NO.
Although no response to SNP could be observed on
phosphate-starved or stationary phase cells, this may only
be the consequence of the experimental conditions used
(e.g. very rapid re-entry of phosphate starved cells to the cell
cycle) that did not allow a proper resolution of cell cycle
events.
In contrast to the above observations in plant cells, NO is a
well-known cytostatic agent and a regulator of the balance
between cell proliferation and cell differentiation during
animal development (Kuzin et al., 2000; Peunova et al.,
1996). However, the site where the cell cycle is actually
stopped in response to NO varies in different cell types (Guo
et al., 1998; Ishida et al., 1997; Pervin et al., 2001; Peunova
et al., 1996; Takagi et al., 1994). Our experiments revealed
that SNP-generated NO did not block cell cycle progression
in cultured alfalfa cells. It cannot be excluded, however, that
NO is involved in the regulation of cell division and
differentiation in whole plants or organs.
The observations that neither dividing leaf protoplasts nor
dividing suspension-cultured cells are responsive to
L-NMMA support the hypothesis that the activity of
L-NMMA-sensitive plant enzymes is not required to sustain
cell division activity in plant cells.
NO is involved in 2,4-D-induced formation of embryogenic
cells from alfalfa leaf protoplasts
The presented data indicate that although NO is not required
for and does not influence cell cycle progression in expo-
nentially dividing cultured plant cells, it may interact with
auxins linking the regulation of cell division to differen-
tiation. Plants exhibit a remarkable developmental plasticity
compared with animals. Somatic plant cells can regain the
ability to divide during a process termed ‘de-differentiation’,
and ‘de-differentiated’ plant cells can ‘re-differentiate’ into
whole plants under suitable conditions. Our experiments
with embryogenic leaf protoplast-derived cells support the
view that NO in concert with auxin can play an important
role during these transitions.
The altered response of protoplast-derived cells to exo-
genous auxin concentration in the presence of L-NMMA or
SNP, and the observation that in the absence of auxin SNP
could not promote the division of protoplast-derived cells,
may indicate that NO alters the auxin sensitivity of the cells
and/or is involved in mediation of the action of auxin during
these processes. Auxin and NO have also been suggested to
share common steps in signal transduction pathways lead-
ing to root elongation (Gouvea et al., 1997) and formation of
adventitious roots (Pagnussat et al., 2002, 2003, 2004). In
cucumber explants, treatment with IAA induced the level of
endogenous NO in the region where the new root meristems
developed. The effect of NO on IAA-induced root formation
was shown to be dependent on intracellular levels of cGMP,
although cGMP-independent pathways may also exist
(Pagnussat et al., 2003, 2004). The downstream events of
the putative NO-dependent signaling cascade leading to
mitotic activation by auxin are unknown. Whether the same
NO-affected signal transduction pathways operate during
the formation of dividing embryogenic cells from leaf
protoplasts and the formation of adventitious root meristem
remains to be discovered.
In addition to affecting the frequency of dividing cells,
both L-NMMA and SNP affected the pathway of auxin
concentration-dependent development of leaf protoplast-
derived cells. It has been previously shown that these cells
can develop either to vacuolized, elongated cells or to small,
isodiametric cells with dense cytoplasm exhibiting embryo-
genic competence (Bogre et al., 1990; Pasternak et al., 2002,
for reviews Dudits et al., 1991; Feher et al., 2003, 2005).
These developmental pathways are dependent on the con-
centration of exogenous auxin (2,4-D) or oxidative stress-
inducing agents (Pasternak et al., 2002). The application of
NO in cell division 857
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

the NO donor SNP resulted in the formation of embryogenic-
type cells at a low (1 lM) 2,4-D concentration, although this
type of cell otherwise appears at higher auxin concentra-
tions (5–10 lM 2,4-D). The high-level expression of the
MsSERK1 gene (Nolan et al., 2003) and the further develop-
ment of the cells under culture conditions, allowing somatic
embryo formation, verified that application of SNP could
indeed alter the developmental pathway of the auxin-treated
cells.
The MsSERK1 gene is the alfalfa ortholog of the carrot
(Schmidt et al., 1997) and Arabidopsis (Hecht et al., 2001)
SERK genes implicated both in somatic and zygotic embryo-
genesis. SERK gene expression is frequently used as a
marker of embryogenic competence (for review see Feher
et al., 2003) although its elevated expression was also
associated with auxin-induced root formation (Nolan et al.,
2003) and was suggested to be a morphogenic rather than
only an embryogenic marker.
The application of the NOS inhibitor L-NMMA resulted in
the vacuolization and elongation of the cells at a high (10 lM)
2,4-D level in the medium. Embryogenic cell formation,
however, was not stopped but only delayed in the presence
of L-NMMA, as indicated by SERK gene expression and
further development of the cells. This delay was very similar
to the delay observed in the case of entry of the protoplast-
derived cells into the cell division cycle. It is interesting to
note that delaying cell division activity of the same type of
cells by transiently altering the pH of the medium also
resulted in a parallel inhibition of embryogenic cell forma-
tion (Pasternak et al., 2002).
It is well established that 2,4-D-induced acquisition of
embryogenic competence is associated with endogenous
accumulation of IAA in various types of plant cells including
alfalfa leaf protoplasts (for review Feher et al., 2003). Whe-
ther auxin (in this case 2,4-D) treatment is also associated
with an increase in the level of endogenous NO, in a similar
way to what was observed during root meristem formation
(Pagnussat et al., 2002, 2003), remains an interesting ques-
tion to be answered.
Experimental procedures
Cell cultures and treatments
Protoplasts were isolated from 4–6-week-old alfalfa (Medicagosativa L. ssp. varia; embryogenic genotype A2) plants grown in vitroas described elsewhere (Pasternak et al., 2000, 2002). Purified pro-toplasts were washed twice with W5 solution and resuspended at acell density of 105 protoplasts ml)1 in K75 medium supplementedwith 0.4 M glucose, 0.1 M mannitol, 2.28 lM zeatin and 0.22–10 lM
2,4-dichlorophenoxyacetic acid (2,4-D). The NO donors (�)-(E)-ethyl-2-[(E)-hydroxyimino]-5-nitro-3-hexeneamide (NOR-3), 3-mor-pholinosydnonimine, HCl (SIN-1), N-acetyl-3-(nitrosothio)-D-valine(SNAP) and 4-phenyl-3-furoxancarbonitrile (furoxan) were pur-chased from Calbiochem (Darmstadt, Germany) as the Nitric OxideDonor Set (catalog no. 482675), and were dissolved as indicated
by the supplier. L-NG-monomethyl-L-arginine (L-NMMA; Sigma,St Louis, MO, USA) was dissolved in water and 2-phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (PTIO; Sigma) in DMSO.Solutions were sterilized by filtration and added to the medium atthe appropriate final concentrations. The effects of sodium nitro-prusside (SNP; Sigma) were dependent on culturing the protoplastsin diffuse light to ensure prolonged delivery of NO to the cells.Strong light in the presence of SNP caused considerable cell death(data not shown).
The A2 cell suspension culture derived from cells of the samealfalfa genotype used for leaf protoplast isolation were subcul-tured weekly in liquid MS medium supplemented with 0.2 lM 2,4-D and 1 lM kinetin. All treatments were carried out at least intriplicate.
The suspension culture of M. sativa ssp. varia A2 cultured cellswas maintained and synchronized by double phosphate starvationas described by Kapros et al. (1992).
Determination of frequency and viability of cell division
Cell viability was determined with Evans blue, a non-permeatingdye, that leaks through ruptured plasma membranes and stain thecontents of dead cells (Baker and Mock, 1994). Cell elongation wasdetermined under a light microscope by an ocular micrometer at500· magnification and was expressed as the ratio of the meas-ured length and width of the cells. At least 40 randomly chosencells were measured. The frequency of cells that had alreadydivided was visually determined by inspecting more than 500 cellswith a bright field microscope. For the determination of S-phasefrequency, cells were cultivated in the presence of 15–30 lM ofbromodesoxyuridine (BrdU) (Amersham Biosciences, Vienna,Austria) for 12–24 h. Immunological detection of BrdU in isolatednuclei was carried out using standard protocols (Amersham Bio-sciences), as described previously in detail by Pasternak et al.(2000, 2002).
Flow cytometry
Nuclei were released from protoplasts or protoplast-derived cellsinto a suitable buffer by gentle pipetting as described by Galbraithet al. (1993). In the case of cultured cells a 30 min pre-treatment bycell-wall-degrading enzymes (cellulase YC 3%, pectinase 1%, mac-erozyme 1%, in a solution containing 5.4 g mannitol, 5.4 g sorbitoland 50 mg CaCl2 in 100 ml at pH 5.3) was applied. Nuclei werestained with propidium iodide (5 lg ml)1) and (5–10) · 103 nucleiwere used for flow cytometric determination of the relative DNAcontent with a FACSCalibur flow cytometer from Becton Dickinson(Franklin Lakes, NJ, USA). Cell cycle analysis was carried out by theModFit� software.
Protein extraction, kinase activity assay and Western
blotting
For protein isolation, cells were frozen in liquid nitrogen and storedat )70�C until analysis. Histone H1 phosphorylating activity of thecognate alfalfa p34cdc2 kinase (Medsa;CDKA;1,2) was determined byimmunoprecipitation as described by Magyar et al. (1997). Westernblotting was carried out using standard protocols, exactly as des-cribed elsewhere (Pasternak et al., 2000, 2002). The primary anti-bodies were prepared against a C-terminal peptide of the alfalfaMedsa;CDKA;1,2 and Medsa;CDKB;2,1 proteins (Magyar et al.,1997) as well as the Medsa;CYCD5;1 protein (Meszaros et al., 2000)
858 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

and against the whole purified alfalfa ferritin protein (Deak et al.,1999) and were affinity purified before use.
Gene expression studies
Messenger RNA populations were isolated from approximately thesame number [(200–250) · 105] of protoplast-derived cells using theDynabeads mRNA Direct Kit (Dynal Biotech, Oslo, Norway). ThemRNAs attached to the beads were treated by RNase-free DNase(Promega, Madison, WI, USA) and were thoroughly washed toprevent contamination by genomic DNA. The purity of the mRNAwas verified by PCR using primers for the actin gene (see below) onsevenfold more mRNA that was present in the cDNA mixture usedfor quantitative RT-PCR. No signals could be obtained indicating theabsence of detectable contamination by genomic DNA. First-strandcDNAs were synthesized by RevertAid M-MuLV reverse transcrip-tase in the presence of 40 U of RiboLock ribonuclease inhibitor(Fermentas, Vilnius, Lithuania). The relative expression of theMsSERK1 gene was measured by real-time PCR on the ABI PRISM7700 sequence detection system (Applied Biosystems, Foster City,CA, USA) using the SYBR-green PCR Master Mix from the samesupplier. The MsSERK1 primer sequences were the same as des-cribed by Nolan et al. (2003). The primers to measure the relativetranscript levels of M. sativa cyclins and the actin B gene (TC86577)as endogenous reference were designed by the Primer Expresssoftware (Applied Biosystems): Medsa;CYCA1;2 forward 5¢-TTT-CTTGCACGATGGACATTAGA-3¢, reverse 5¢-GAGGCATAGTGTTCA-AGAGTTGGA-3¢; Medsa;CycD3;1 forward 5¢-TGTTTAAGAAGACC-AAGAATCAAGGA-3¢, reverse 5¢-TGGCAAGAATTCCGACAATG-3¢;actin B forward 5¢-TCCTAGGGCTGTGTTTCCAAGT-3¢, reverse5¢-CATACCGGTGTCATGGTTGG-3¢.
Primer specificity was verified by cloning and sequencing and byend-point dissociation analysis of PCR products. Real time PCRreactions were carried out in triplicate and the results were analyzedusing the comparative threshold cycle (CT) method and averagedaccording to the advices of the manufacturer (Applied Biosystems).The presented data were derived from three independent treat-ments.
Acknowledgements
The presented experiments were partly supported by theHungarian Scientific Research Fund (grants OTKA T34818; OTKAT37910). AF is grateful for the support of the ‘Janos Bolyai’Research Fellowship.
Supplementary Material
The following supplementary material is available for this articleonline:Figure S1. DAF-2DA (a) and 4-AF (b) fluorescence in leaf protoplast-derived alfalfa cells cultured for 3 days under normal cultureconditions (1 lM 2,4-D as exogenous auxin).Figure S2. Effect of SNP (10 lM) or L-NMMA (1 mM) on theformation of embryogenic cell clusters from alfalfa leaf protoplast-derived cells.
References
Baker, C.J. and Mock, N.M. (1994) An improved method for monit-oring cell death in cell suspension and leaf disc assays usingEvans blue. Plant Cell Tissue Organ Cult. 39, 7–12.
Beligni, M.V. and Lamattina, L. (2000) Nitric oxide stimulatesseed germination and de-etiolation, and inhibits hypocotyl
elongation, three light-inducible responses in plants. Planta, 210,215–221.
Beligni, M.V. and Lamattina, L. (2001) Nitric oxide in plants: thehistory is just beginning. Plant Cell Environ. 24, 267–278.
Beligni, M.V., Fath, A., Bethke, P.C., Lamattina, L. and Jones, R.L.
(2002) Nitric oxide acts as an antioxidant and delays programmedcell death in barley aleurone layers. Plant Physiol. 129, 1642–1650.
Bogdan, C. (2001) Nitric oxide and the regulation of gene expres-sion. Trends Cell Biol. 11, 66–75.
Bogre, L., Stefanov, I., Abraham, M., Somogyi, I. and Dudits, D.
(1990) Differences in the responses to 2,4-dichlorophenoxyaceticacid (2,4-D) treatment between embryogenic and non-embryo-genic lines of alfalfa. In Progress in Plant Cellular and MolecularBiology (Nijkamp, H.J.J., van der Plaas, L.H.W. and Van Aartrijk,J., eds). Dordrecht: Kluwer Academic Publishers, pp. 427–436.
Clarke, A., Desikan, R., Hurst, R.D., Hancock, J.T. and Neill, S.J.
(2000) NO way back: nitric oxide and programmed cell deathin Arabidopsis thaliana suspension cultures. Plant J. 24, 667–677.
Cooper, C.E. (1999) Nitric oxide and iron proteins. Biochim. Biophys.Acta, 1411, 290–309.
Correa-Aragunde, N., Graziano, M. and Lamattina, L. (2004) Nitricoxide plays a central role in determining lateral root developmentin tomato. Planta, 218, 900–905.
Dahl, M., Meskiene, I., Bogre, L., Cam Ha, D.T., Swoboda, I., Hub-
mann, R., Hirt, H. and Heberle-Bors, E. (1995) The D-type alfalfacyclin cycMs4 complements G1 cyclin-deficient yeast and isinduced in the G1 phase of the cell cycle. Plant Cell, 7, 1847–1875.
Deak, M., Horvath, G.V., Davletova, S., Torok, K., Sass, L., Vass, I.,
Barna, B., Kiraly, Z. and Dudits, D. (1999) Plants ectopicallyexpressing the iron-binding protein, ferritin, are tolerant to oxi-dative damage and pathogens. Nat. Biotechnol. 17, 192–196.
Delledonne, M., Xia, Y.J., Dixon, R.A. and Lamb, C. (1998) Nitricoxide functions as a signal in plant disease resistance. Nature,394, 585–588.
Dudits, D., Bogre, L. and Gyorgyey, J. (1991) Molecular and cellularapproaches to the analysis of plant embryo development fromsomatic cells in vitro. J. Cell Sci. 99, 475–484.
Durner, J. and Klessig, D.F. (1999) Nitric oxide as a signal in plants.Curr. Opin. Plant Biol. 2, 369–374.
Durner, J., Wendehenne, D. and Klessig, D.F. (1998) Defense geneinduction in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose. Proc. Natl Acad. Sci. USA, 95, 10328–10333.
Durzan, D.J. and Pedroso, M.C. (2002) Nitric oxide and reactivenitrogen oxide species in plants. Biotechnol. Genet. Eng. Rev. 19,293–337.
Feher, A., Pasternak, T.P and Dudits, D. (2003) Transition of somaticplant cells to an embryogenic state. Plant Cell Tissue Organ Cult.74, 201–228.
Feher, A., Pasternak, T., Otvos, K. and Dudits, D. (2005) Plant pro-toplasts: consequences of lost cell walls. In Journey of a SingleCell to a Plant (Murch, S.J. and Saxena, P.K., eds). Enfield, NH,USA: Science Publishers, pp. 59–90.
Galbraith, D.W., Harkins, K.R., Maddox, J.M., Ayres, N.M., Sharma,
D.P. and Firoozabady, E. (1993) Rapid flow cytometric analysis ofthe cell cycle in intact plant tissues. Science, 220, 1049–1051.
Gould, K.S., Lamotte, O., Klinguer, A., Pugin, A. and Wendehenne,
D. (2003) Nitric oxide production in tobacco leaf cells: a general-ized stress response? Plant Cell Environ. 26, 1851–1862.
Gouvea, C.M.C.P., Souza, J.F. and Magelhaes, M.I.S. (1997) NO-releasing substances that induce growth elongation in maize rootsegments. Plant Growth Regul. 21, 183–187.
Guo, K., Andres, V. and Walsh, K. (1998) Nitric oxide-induceddownregulation of cdk2 activity and cyclin A gene transcription invascular smooth muscle cells. Circulation, 97, 2066–2072.
NO in cell division 859
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860

Guo, F.Q., Okamoto, M. and Crawford, N.M. (2003) Identification ofa plant nitric oxide synthase gene involved in hormonal signaling.Science, 302, 100–103.
Hecht, V., Vielle-Calzada, J.P., Hartog, M.V., Schmidt, E.D., Bouti-
lier, K., Grossniklaus, U. and de Vries, S.C. (2001) The Arabidopsissomatic embryogenesis receptor kinase 1 gene is expressed indeveloping ovules and embryos and enhances embryogeniccompetence in culture. Plant Physiol. 127, 803–816.
Ishida, A., Sasaguri, T., Kosaka, C., Nojima, H. and Ogata, J. (1997)Induction of the cyclin-dependent kinase inhibitor p21(Sdi1/Cip1/Waf1) by nitric oxide-generating vasodilator in vascular smoothmuscle cells. J. Biol. Chem. 272, 10050–10057.
Kapros, T., Bogre, L., Nemeth, K., Bako, L., Gyorgyey, J., Wu, S.C.
and Dudits, D. (1992) Differential expression of histone H3 genevariants during cell cycle and somatic embryogenesis in alfalfa.Plant Physiol. 98, 621–625.
Kojima, H., Nakatsubo, N., Kikuchi, K., Kawahara, S., Kirino, Y.,
Nagoshi, H., Hirata, Y. and Nagano, T. (1998) Detection andimaging of nitric oxide with novel fluorescent indicators: diami-nofluoresceins. Anal. Chem. 70, 2446–2453.
Kuo, R.K., Baxter, G.T., Thompson, S.H., Stricker, S.A., Patton, C.,
Bonaventura, J. and Epel, D. (2000) NO is necessary and sufficientfor egg activation at fertilization. Nature, 406, 633–636.
Kuzin, B., Regulski, M., Stasiv, Y., Scheinker, V., Tully, T. and
Enikolopov, G. (2000) Nitric oxide interacts with the retinobla-stoma pathway to control eye development in Drosophila. Curr.Biol. 10, 459–462.
Lamattina, L., Garcia-Mata, C., Graziano, M. and Pagnussat, G.
(2003) Nitric oxide: the versatility of an extensive signal molecule.Annu. Rev. Plant Biol. 54, 109–136.
Leshem, Y.Y. and Haramaty, E. (1996) The characterization andcontrasting effects of the nitric oxide free radical in vegetativestress and senescence of Pisum sativum Linn foliage. J. PlantPhysiol. 148, 258–263.
Leshem, Y.Y. and Pinchasov, Y. (2000) Non-invasive photoacousticspectroscopic determination of relative endogenous nitric oxideand ethylene content stoichiometry during the ripening ofstrawberries Fragaria anannasa (Duch.) and avocados Perseaamericana (Mill.). J. Exp. Bot. 51, 1471–1473.
Mackerness, S.A.H., John, C.F., Jordan, B. and Thomas, B. (2001)Early signaling components in ultraviolet-B responses: distinctroles for different reactive oxygen species and nitric oxide. FEBSLett. 489, 237–242.
Magyar, Z., Meszaros, T., Miskolczi, P. et al. (1997) Cell cycle phasespecificity of putative cyclin-dependent kinase variants in syn-chronized alfalfa cells. Plant Cell, 9, 223–235.
Mata, C.G. and Lamattina, L. (2001) Nitric oxide induces stomatalclosure and enhances the adaptive plant responses againstdrought stress. Plant Physiol. 126, 1196–1204.
Meskiene, I., Bogre, L., Dahl, M., Pirck, M., Cam Ha, D.T., Swoboda,
I., Heberle-Bors, E., Ammerer, G. and Hirt, H. (1995) cycMs3, anovel B-type alfalfa cyclin gene, is induced in the G0-toG1 trans-ition of the cell cycle. Plant Cell, 7, 759–771.
Meszaros, T., Miskolczi, P., Ayaydin, F., Pettko-Szandtner, A., Peres,
A., Magyar, Z., Horvath, V.G, Bako, L., Feher, A. and Dudits, D.
(2000) Multiple cyclin-dependent kinase complexes and phos-phatases control G2/M progression in alfalfa cells. Plant Mol. Biol.43, 595–605.
Murgia, I., Delledonne, M. and Soave, C. (2002) Nitric oxide medi-ates iron-induced ferritin accumulation in Arabidopsis. Plant J.30, 521–528.
Neill, S.J., Desikan, R. and Hancock, J.T. (2003) Nitric oxide signa-ling in plants. New Phytol. 159, 11–35.
Nolan, K.E., Irwanto, R.R. and Rose, R.J. (2003) Auxin up-regulatesMtSERK1 expression in both Medicago truncatula root-formingand embryogenic cultures. Plant Physiol. 133, 218–230.
Pagnussat, G.C., Simontacchi, M., Puntarulo, S. and Lamattina, L.
(2002) Nitric oxide is required for root organogenesis. PlantPhysiol. 129, 954–956.
Pagnussat, G.C., Lanteri, M.L. and Lamattina, L. (2003) Nitricoxide and cyclic GMP are messengers in the indole acetic acid-induced adventitious rooting process. Plant Physiol. 132, 1241–1248.
Pagnussat, G.C., Lanteri, M.L., Lombardo, M.C. and Lamattina, L.
(2004) Nitric oxide mediates the indole acetic acid induction of amitogen-activated protein kinase cascade involved in adventi-tious root development. Plant Physiol. 135, 279–286.
Pasternak, T., Miskolczi, P., Ayaydin, F., Meszaros, T., Dudits, D. and
Feher, A. (2000) Exogenous auxin and cytokinin dependent acti-vation of CDKs and cell division in leaf protoplast-derived cells ofalfalfa. Plant Growth Regul. 32, 129–141.
Pasternak, T., Prinsen, E., Ayaydin, F., Miskolczi, P., Potters, G.,
Asard, H., Van Onckelen, H., Dudits, D. and Feher, A. (2002) Therole of auxin, pH and stress in the activation of embryogenic celldivision in leaf protoplast-derived cells of alfalfa (Medicago sativaL.). Plant Physiol. 129, 1807–1819.
Pervin, S., Singh, R. and Chaudhuri, G. (2001) Nitric oxide-inducedcytostasis and cell cycle arrest of a human breast cancer cell line(MDA-MB-231): potential role of cyclin D1. Proc. Natl Acad. Sci.USA, 98, 3583–3588.
Peunova, N., Kuzin, B., Roberts, I., Okane, C. and Enikolopov, G.
(1996) Nitric oxide, cell multiplication, and cell survival. ColdSpring Harb. Symp. Quant. Biol. 61, 417–426.
de Pinto, M.C., Tommasi, F. and De Gara, L. (2002) Changes in theantioxidant systems as part of the signaling pathway responsiblefor the programmed cell death activated by nitric oxide andreactive oxygen species in tobacco Bright-Yellow 2 cells. PlantPhysiol. 130, 698–708.
Schmidt, H.H. and Walter, U. (1994) NO at work. Cell, 78, 919–925.
Schmidt, E.D., Guzzo, F., Toonen, M.A. and de Vries, S.C. (1997) Aleucine-rich repeat containing receptor-like kinase marks somaticplant cells competent to form embryos. Development, 124, 2049–2062.
Seregelyes, C., Barna, B., Hennig, J., Konopka, D., Pasternak, T.P.,
Lukacs, N., Feher, A., Horvath, G.V. and Dudits, D. (2003) Phyto-globins can interfere with nitric oxide functions during plantgrowth and pathogenic responses: a transgenic approach. PlantSci. 165, 541–550.
Sheen, J. (2001) Signal transduction in maize and Arabidopsismesophyll protoplasts. Plant Physiol. 127, 1466–1475.
Takagi, K., Isobe, Y., Yasukawa, K., Okouchi, E. and Suketa, Y.
(1994) Nitric-oxide blocks the cell-cycle of mouse macrophage-like cells in the early G(2)þM phase. FEBS Lett. 340, 159–162.
Wang, Y. and Marsden, P.A. (1995) Nitric oxide synthases: bio-chemical and molecular regulation. Curr. Opin. Nephrol. Hyper-tens. 4, 12–22.
Wink, D.A. andMitchell, J.B. (1998) Chemical biology of nitric oxide:insights into regulatory, cytotoxic, and cytoprotective mecha-nisms of nitric oxide. Free Radic. Biol. Med. 25, 434–456.
Zhang, M., An, L., Feng, H., Chen, T., Chen, K., Liu, Y., Tang, H.,
Chang, J. and Wang, X. (2003) The cascade mechanismsof nitric oxide as a second messenger of ultraviolet B ininhibiting mesocotyl elongations. Photochem. Photobiol. 77,219–225.
860 Krisztina Otvos et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 43, 849–860




















