NEW SPECIES OF PARAPHIOMYS (RODENTIA, THRYONOMYIDAE) FROM THE LOWER MIOCENE OF AS-SARRAR, SAUDI...
11
NEW SPECIES OF PARAPHIOMYS (RODENTIA, THRYONOMYIDAE) FROM THE LOWER MIOCENE OF AS-SARRAR, SAUDI ARABIA by RAQUEL LO ´ PEZ ANTON ˜ ANZAS and SEVKET SEN UMR 5143 du CNRS, De ´partement Histoire de la Terre, Muse ´um national d’Histoire naturelle, 8 rue Buffon, F-75005 Paris, France; e-mail: [email protected], [email protected] Typescript received 1 July 2003; accepted in revised form 12 December 2003 Abstract: The family Thryonomyidae is represented in the Lower Miocene of Saudi Arabia by a single species, Paraphio- mys knolli sp. nov. This new taxon differs from all other thryonomyids in being small, lower molars having a short metalophulid II and an isolated anterolabial cuspid, and upper molars being antero-posteriorly compressed and pen- talophodont. A cladistic analysis involving all extinct and extant species of thryonomyids is provided. Paraphiomys knolli branches as the sister-species of Paraphiomys pigotti, type species of the genus. Key words: Rodentia, Thryonomyidae, Paraphiomys, phylo- geny, Miocene, Saudi Arabia. The Saudi–French palaeontological Research Project was carried out between 1977 and the early 1980s and resulted in the discovery of several Lower and Middle Miocene mammal localities near the coast of the Ara- bian Gulf, Saudi Arabia. Thomas et al. (1982) published preliminary results of this field work and recognized at least 66 species from the Early Miocene (tentatively cor- related to MN 5) faunas of the As-Sarrar area (Text- fig. 1). Four sites there have yielded rodent remains, but representatives of the family Thryonomyidae are recorded only at localities 8 and 21. Thomas et al. (1982, p. 129) provisionally assigned this material to Paraphiomys sp. They briefly described it as pentalopho- dont with well-developed mesolophs and mesolophids. Flynn et al. (1983) described and illustrated the right lower molar AS8-1100, on the basis of which they announced the presence of a plesiomorphic thryonom- yid in the As-Sarrar fauna. Finally, Winkler (1992, p. 244) considered this taxon to be possibly new, at the species or even genus level. The thryonomyid dental material collected in Saudi Arabia is especially important from a palaeogeographical point of view because it bridges a gap between African and Pakistani representatives. The aim of this paper is to present a detailed study of the thryonomyid dental mater- ial that has been recorded from the Lower Miocene of the Dam Formation in Saudi Arabia and to compare it with all Oligocene and Miocene thryonomyid species known to date. The terminology for tooth morphology follows that of Winkler (1992). Abbreviations: AMNH, American Museum of Natural History, New York, USA; AS, As-Sarrar; CGM, Geological Museum, Cairo, Egypt; GSW, Geological Survey, Windhoek, Namibia; H-GSP, Howard University-Geological Survey of Pakistan; IPS, Instituto de Paleontologı´a, Sabadell, Spain; KNM, National Museums of Kenya, Nairobi, Kenya; MNHN, Muse ´um national d’Histoire naturelle, Paris, France; NMAA, National Museum, Addis Ababa, Ethiopia; UM, Universite ´ des Sciences et Tech- niques du Languedoc, Montpellier, France. SYSTEMATIC PALAEONTOLOGY Order RODENTIA Bowdich, 1821 Family THRYONOMYIDAE Pocock, 1922 Genus PARAPHIOMYS Andrews, 1914 Type species. Paraphiomys pigotti Andrews, 1914 Paraphiomys knolli sp. nov. Text-figure 2A–F Derivation of name. In honour of the palaeontologist Fabien Knoll. Holotype. AS8-1100 (Text-fig. 2A), a right m1-m2. [Palaeontology, Vol. 48, Part 2, 2005, pp. 223–233] ª The Palaeontological Association 223
-
Upload
biochange-lab -
Category
Documents
-
view
1 -
download
0
Transcript of NEW SPECIES OF PARAPHIOMYS (RODENTIA, THRYONOMYIDAE) FROM THE LOWER MIOCENE OF AS-SARRAR, SAUDI...

NEW SPECIES OF PARAPHIOMYS (RODENTIA,
THRYONOMYIDAE) FROM THE LOWER MIOCENE
OF AS-SARRAR, SAUDI ARABIA
by RAQUEL LOPEZ ANTONANZAS and SEVKET SENUMR 5143 du CNRS, Departement Histoire de la Terre, Museum national d’Histoire naturelle, 8 rue Buffon, F-75005 Paris, France; e-mail: [email protected],
Typescript received 1 July 2003; accepted in revised form 12 December 2003
Abstract: The family Thryonomyidae is represented in the
Lower Miocene of Saudi Arabia by a single species, Paraphio-
mys knolli sp. nov. This new taxon differs from all other
thryonomyids in being small, lower molars having a short
metalophulid II and an isolated anterolabial cuspid, and
upper molars being antero-posteriorly compressed and pen-
talophodont. A cladistic analysis involving all extinct and
extant species of thryonomyids is provided. Paraphiomys
knolli branches as the sister-species of Paraphiomys pigotti,
type species of the genus.
Key words: Rodentia, Thryonomyidae, Paraphiomys, phylo-
geny, Miocene, Saudi Arabia.
The Saudi–French palaeontological Research Project
was carried out between 1977 and the early 1980s and
resulted in the discovery of several Lower and Middle
Miocene mammal localities near the coast of the Ara-
bian Gulf, Saudi Arabia. Thomas et al. (1982) published
preliminary results of this field work and recognized at
least 66 species from the Early Miocene (tentatively cor-
related to MN 5) faunas of the As-Sarrar area (Text-
fig. 1). Four sites there have yielded rodent remains,
but representatives of the family Thryonomyidae are
recorded only at localities 8 and 21. Thomas et al.
(1982, p. 129) provisionally assigned this material to
Paraphiomys sp. They briefly described it as pentalopho-
dont with well-developed mesolophs and mesolophids.
Flynn et al. (1983) described and illustrated the right
lower molar AS8-1100, on the basis of which they
announced the presence of a plesiomorphic thryonom-
yid in the As-Sarrar fauna. Finally, Winkler (1992,
p. 244) considered this taxon to be possibly new, at the
species or even genus level.
The thryonomyid dental material collected in Saudi
Arabia is especially important from a palaeogeographical
point of view because it bridges a gap between African
and Pakistani representatives. The aim of this paper is to
present a detailed study of the thryonomyid dental mater-
ial that has been recorded from the Lower Miocene of the
Dam Formation in Saudi Arabia and to compare it with
all Oligocene and Miocene thryonomyid species known to
date.
The terminology for tooth morphology follows that of
Winkler (1992).
Abbreviations: AMNH, American Museum of Natural History,
New York, USA; AS, As-Sarrar; CGM, Geological Museum,
Cairo, Egypt; GSW, Geological Survey, Windhoek, Namibia;
H-GSP, Howard University-Geological Survey of Pakistan; IPS,
Instituto de Paleontologıa, Sabadell, Spain; KNM, National
Museums of Kenya, Nairobi, Kenya; MNHN, Museum national
d’Histoire naturelle, Paris, France; NMAA, National Museum,
Addis Ababa, Ethiopia; UM, Universite des Sciences et Tech-
niques du Languedoc, Montpellier, France.
SYSTEMATIC PALAEONTOLOGY
Order RODENTIA Bowdich, 1821
Family THRYONOMYIDAE Pocock, 1922
Genus PARAPHIOMYS Andrews, 1914
Type species. Paraphiomys pigotti Andrews, 1914
Paraphiomys knolli sp. nov.
Text-figure 2A–F
Derivation of name. In honour of the palaeontologist Fabien
Knoll.
Holotype. AS8-1100 (Text-fig. 2A), a right m1-m2.
[Palaeontology, Vol. 48, Part 2, 2005, pp. 223–233]
ª The Palaeontological Association 223

Paratypes. AS21-1101, left D4 (Text-fig. 2B); AS8-1102 (Text-
fig. 2C), AS8-1103 (Text-fig. 2D), right M1-2; AS8-1104 (Text-
fig. 2E), left M3; AS8-1105, right M3 (Text-fig. 2F). All material
is housed in the collections of the Laboratoire de Paleontologie
(MNHN); size data for each tooth are provided in Table 1.
Localities and type horizon. Loc. 8 (type locality) and Loc. 21,
As-Sarrar area, Saudi Arabia. Dam Formation; late Early Mio-
cene (c. 15–17 Ma).
Diagnosis. Species of Paraphiomys of small size with sem-
ibunodont teeth. Lower molars with three complete lophs,
plus a short metalophulid II, an isolated anterolabial cuspid,
and lacking anterolabial cingulum. Antero-posteriorly
compressed upper molars with five complete lophs and a
labially displaced hypocone.
Differential diagnosis. Paraphiomys knolli differs from
Gaudeamus aegyptius, Neosciuromys africanus, Apodecter
stromeri, P. orangeus, P. renelavocati, P. simonsi, P. ship-
mani, Kochalia geespei, Paraulacodus indicus, P. johanesi,
P. afarensis, P. roessneri, P. australis and the two living
species of Thryonomys notably in having metalophulid II
on the lower molars. It is distinct from Paraphiomys pig-
otti and P. occidentalis in being much smaller than these
and from Epiphiomys coryndoni in having a much shorter
metalophulid II. Paraphiomys knolli differs from P. hop-
woodi in being smaller, having the upper molars more an-
tero-posteriorly compressed and in showing a more
labially displaced hypocone. Finally, this species is distinct
from Sacaresia moyaeponsi in having the teeth semibuno-
dont whereas the latter species is hyperlophodont but
lower crowned with slender lophs.
Description
m1 or m2 (AS8-1100). The single lower molar from As-Sarrar
has a subrectangular occlusal outline. It has three transverse
lophs (metalophid, hypolophid and posterolophid) and a short
metalophulid II. This tooth shows an isolated and transversely
elongated anteroconid. Mesoflexid and metaflexid are approxi-
mately equal in length as well as in depth, and open lingually,
but the valley between the hypolophid and the metalophid is
wider than that between the hypolophid and the posterolophid.
The protoconid is situated slightly external to the hypoconid.
The hypoflexid is deeper than the mesoflexid and the metaflexid,
and it is almost transverse.
D4 (AS21-1101). A single well-preserved tooth (D4) has been
found at locality 21 of As-Sarrar. It is trapezoidal in occlusal
outline. The anterior half of the crown is narrower than the
posterior one. The tooth has an anteroloph, a protoloph, a meta-
loph, a posteroloph and a very short mesoloph. The protoloph
is incomplete and does not reach the labial side of the tooth.
The metaloph is well developed and connects lingually with the
posteroloph. The protoloph and the metaloph are anteriorly
directed. The hypoflexus is nearly transverse.
M1 or M2 (AS8-1102 and AS8-1103). Both specimens are badly
worn and lack their lingual portions. These teeth have an anter-
oloph, a protoloph, a metaloph, a posteroloph and a short,
transverse mesoloph. The anterior half of the crown appears to
be narrower than the posterior one. The paraflexus is longer and
slightly deeper than the metaflexus.
M3 (AS8-1104, AS8-1105). The third upper molars from
As-Sarrar are worn, but all their structures are easily recogniz-
able. The occlusal outline is rounded. The anterior half of the
crown is wider than the posterior one. These teeth have four
TABLE 1 . Measurements (mm) of the teeth of Paraphiomys
knolli sp. nov. from the Arabian Peninsula.
As-Sarrar Length Width
m1 or m2 AS8-1100 1Æ88 1Æ74
D4 AS21-1101 1Æ80 1Æ51
M1 or M2 AS8-1102 1Æ88 –
M1 or M2 AS8-1103 1Æ96 –
M3 AS8-1104 1Æ74 1Æ78
M3 AS8-1105 1Æ77 1Æ86
TEXT -F IG . 1 . Major African, Arabian, and southern Asian
Eocene–Miocene thryonomyid localities. 1, Thymiana (Chios
Island, Greece); 2, Beni Mellal (Morocco); 3, Jebel Zelten
(Libya); 4, Fayum (Egypt); 5, Ch’orora (Ethiopia); 6, Napak and
Bukwa (Uganda); 7, Lothagam and Samburu hills (northern
Kenya); 8, Baringo (central Kenya); 9, Songhor, Rusinga,
Maboko, and Fort Ternan (western Kenya); 10, Otavi
(Namibia); 11, Namib (Namibia); 12, As-Sarrar (Saudi Arabia);
13, Abu Dhabi (United Arab Emirates); 14, Sind Province
(Pakistan); 15, Zinda Pir Dome (Pakistan); 16, Potwar Plateau
(Pakistan). The grey area represents the present day distribution
of the family.
224 P A L A E O N T O L O G Y , V O L U M E 4 8
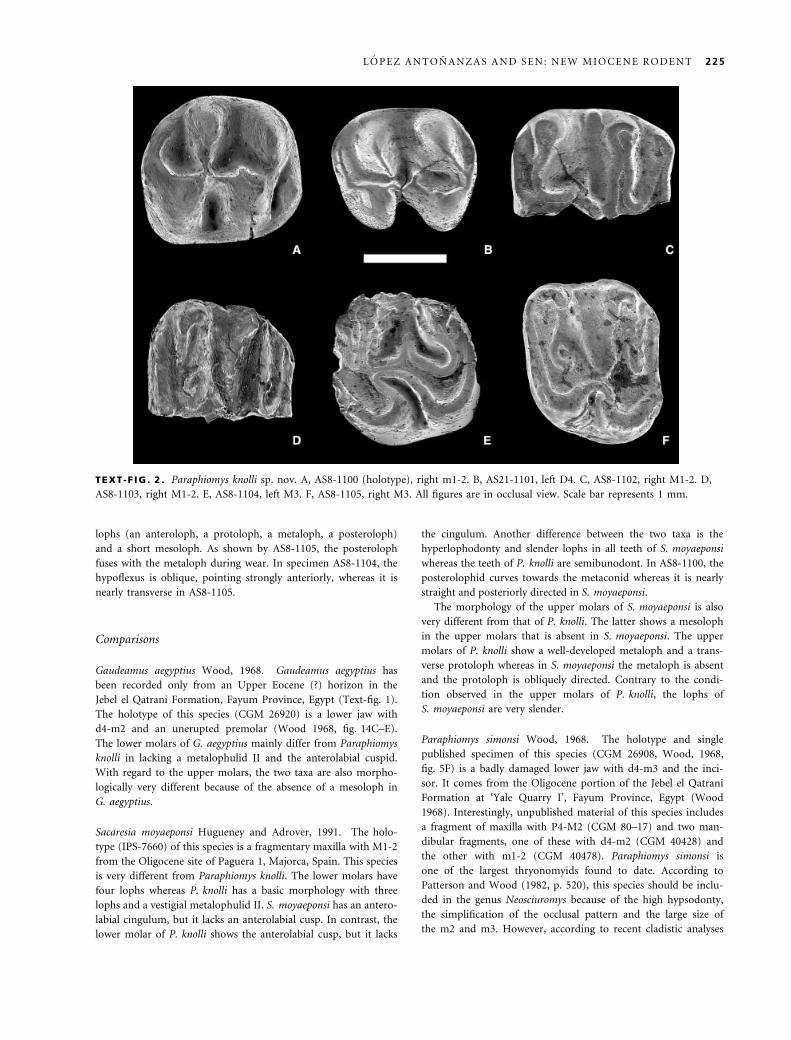
lophs (an anteroloph, a protoloph, a metaloph, a posteroloph)
and a short mesoloph. As shown by AS8-1105, the posteroloph
fuses with the metaloph during wear. In specimen AS8-1104, the
hypoflexus is oblique, pointing strongly anteriorly, whereas it is
nearly transverse in AS8-1105.
Comparisons
Gaudeamus aegyptius Wood, 1968. Gaudeamus aegyptius has
been recorded only from an Upper Eocene (?) horizon in the
Jebel el Qatrani Formation, Fayum Province, Egypt (Text-fig. 1).
The holotype of this species (CGM 26920) is a lower jaw with
d4-m2 and an unerupted premolar (Wood 1968, fig. 14C–E).
The lower molars of G. aegyptius mainly differ from Paraphiomys
knolli in lacking a metalophulid II and the anterolabial cuspid.
With regard to the upper molars, the two taxa are also morpho-
logically very different because of the absence of a mesoloph in
G. aegyptius.
Sacaresia moyaeponsi Hugueney and Adrover, 1991. The holo-
type (IPS-7660) of this species is a fragmentary maxilla with M1-2
from the Oligocene site of Paguera 1, Majorca, Spain. This species
is very different from Paraphiomys knolli. The lower molars have
four lophs whereas P. knolli has a basic morphology with three
lophs and a vestigial metalophulid II. S. moyaeponsi has an antero-
labial cingulum, but it lacks an anterolabial cusp. In contrast, the
lower molar of P. knolli shows the anterolabial cusp, but it lacks
the cingulum. Another difference between the two taxa is the
hyperlophodonty and slender lophs in all teeth of S. moyaeponsi
whereas the teeth of P. knolli are semibunodont. In AS8-1100, the
posterolophid curves towards the metaconid whereas it is nearly
straight and posteriorly directed in S. moyaeponsi.
The morphology of the upper molars of S. moyaeponsi is also
very different from that of P. knolli. The latter shows a mesoloph
in the upper molars that is absent in S. moyaeponsi. The upper
molars of P. knolli show a well-developed metaloph and a trans-
verse protoloph whereas in S. moyaeponsi the metaloph is absent
and the protoloph is obliquely directed. Contrary to the condi-
tion observed in the upper molars of P. knolli, the lophs of
S. moyaeponsi are very slender.
Paraphiomys simonsi Wood, 1968. The holotype and single
published specimen of this species (CGM 26908, Wood, 1968,
fig. 5F) is a badly damaged lower jaw with d4-m3 and the inci-
sor. It comes from the Oligocene portion of the Jebel el Qatrani
Formation at ‘Yale Quarry I’, Fayum Province, Egypt (Wood
1968). Interestingly, unpublished material of this species includes
a fragment of maxilla with P4-M2 (CGM 80–17) and two man-
dibular fragments, one of these with d4-m2 (CGM 40428) and
the other with m1-2 (CGM 40478). Paraphiomys simonsi is
one of the largest thryonomyids found to date. According to
Patterson and Wood (1982, p. 520), this species should be inclu-
ded in the genus Neosciuromys because of the high hypsodonty,
the simplification of the occlusal pattern and the large size of
the m2 and m3. However, according to recent cladistic analyses
TEXT -F IG . 2 . Paraphiomys knolli sp. nov. A, AS8-1100 (holotype), right m1-2. B, AS21-1101, left D4. C, AS8-1102, right M1-2. D,
AS8-1103, right M1-2. E, AS8-1104, left M3. F, AS8-1105, right M3. All figures are in occlusal view. Scale bar represents 1 mm.
L O P E Z A N T O N A N Z A S A N D S E N : N E W M I O C E N E R O D E N T 225

(Lopez-Antonanzas 2004; Lopez-Antonanzas et al. 2004), the
high hypsodonties of Neosciuromys africanus and P. simonsi are
homoplastic. The main differences between P. knolli and
P. simonsi are the hypsodonty and great size of the latter. The
lower molar of P. knolli shows a vestigial metalophulid II whereas
it is absent in P. simonsi. In P. simonsi, a small basal cingulum
lies along the anterior side of the protoconid, rising into a small
cusp, whereas P. knolli the anterolabial cingulum is lacking and
the anterior cusp is larger. Contrary to the upper molars of
P. knolli, those of P. simonsi have a three-lophed morphology.
Paraphiomys pigotti Andrews, 1914. This species was described
and illustrated by Andrews (1914, pp. 177–178) based on a frag-
mentary left mandible with p4-m2 from the Lower Miocene Bed
24 at Kachuku, Karungu area, Kenya. This taxon has been recor-
ded in the Lower Miocene of Rusinga (Lavocat 1973), Moruorot
(Madden 1972; Pickford 1981), and Gumba (Pickford 1981)
Kenya. It is also known in the Lower Miocene of Uganda at
Napak and Bukwa (Pickford 1981; Winkler and Downs 2002),
and it has been reported in the Middle Miocene in the Kenyan
localities of Fort Ternan (Shipman et al. 1981, p. 67) and Kiri-
mon (Pickford 1981; p. 89).
The morphology of the m1–2 of Paraphiomys pigotti (Andrews
1914, pl. 28, fig. 7; Lavocat 1973, pl. 27, fig. 9) is close to that of
the single m1-m2 recorded at As-Sarrar. However, the much
greater size of P. pigotti warrants its systematic distinction from
the As-Sarrar taxon. Furthermore, it shows the hypoflexus posteri-
orly directed whereas it is transverse in the m1–2 from As-Sarrar.
The latter also differs from P. pigotti in having the lingual end of
the posterolophid bent forwards, i.e. towards the lingual end of
the entoconid. Therefore, the metaflexus is more closed than that
of P. pigotti, in which this sinus is entirely open lingually. The m1-2
of P. pigotti (Andrews 1914, pl. 28, fig. 7; Lavocat 1973, pl. 27,
fig. 9), as well as that of P. knolli, show a short metalophulid II
that does not join the metaconid. Both taxa also have an isolated
and transversally elongated anteroconid. The upper molars of
P. pigotti mainly differ from those of P. knolli in their great size.
Lavocat (1973, p. 39) considered Phthinylla fracta Hopwood, 1929
to be a junior synonym of Paraphiomys pigotti, which was acknow-
ledged by Hendey (1978). However, the equally high hypsodonty,
and the discovery of Phthinylla-like maxillary teeth along with
Neosciuromys-like mandibles (P. Mein, pers. comm. 2003) suggest
that the former taxon is a junior synonym of the latter (Lopez-
Antonanzas 2004; Lopez-Antonanzas et al. 2004), as suspected by
Flynn and Winkler (1994) and Winkler (1994).
Only one of the two complete maxillae considered by Lavocat
(1973, p. 37) to be Paraphiomys pigotti corresponds to this taxon
(see Lavocat 1973, pl. 24, fig. 4); the other one (Phthinylla fract-
a-like) pertains to Neosciuromys africanus. The upper molars of
P. pigotti and those from As-Sarrar show a mesoloph, but this is
much less developed in the latter. It can even join the metacone
in P. pigotti. Furthermore, the mesoloph is posteriorly directed
in P. pigotti whereas it is nearly transverse in the specimens from
As-Sarrar.
Neosciuromys africanus Stromer, 1922. Neosciuromys africanus
has been described on the basis of material from the Lower Mio-
cene of Langental and Plant 4 borehole (south of Luderitz Bay,
Namibia). Stromer (1922, 1924, p. 263; 1926, pp. 135–136) des-
cribed and figured as N. africanus two mandibular fragments
with p4-m2 and a fragmentary left maxilla. Interestingly, the
latter specimen is actually a right mandibular fragment with
m2-m3 of the bathyergoidid Bathyergoides neotertiarius (Mein,
pers. comm. 2003). In contrast, the abnormally narrow M1 fig-
ured by Stromer (1926, pl. 42, fig. 24) as cf. Phiomys andrewsi
belongs instead to Neosciuromys africanus. There are obvious
morphological and dimensional differences between Neosciuro-
mys africanus and Paraphiomys knolli.
With regard to the lower molars, AS8-1100 is much smaller
than the m1–2 of N. africanus. In addition, Neosciuromys has
higher-crowned teeth and the pattern of the lower cheek teeth is
reduced to three transverse crests with no trace of metalophulid
II. In the specimen from As-Sarrar, the hypolophid is nearly
transverse whereas it is anteriorly directed in N. africanus. The
antero-posterior difference in width of AS8-1100 is much more
pronounced than in the lower molars of N. africanus. Finally,
the m1-2 from As-Sarrar shows a transverse hypoflexus whereas
it bends antero-posteriorly in N. africanus.
The single D4 from As-Sarrar shows a very short metaloph
whereas it is more developed in N. africanus. AS8-1101 is also
much smaller and less hypsodont than the D4 of N. africanus
and has the protoloph and metaloph anteriorly directed whereas
in N. africanus they are transverse. Contrary to the condition in
N. africanus, in the hypoflexus of AS8-1101 is nearly transverse.
The upper molars from As-Sarrar are very different from
those of N. africanus. As explained above, the maxilla described
by Hopwood (1929, p. 4, fig. 3) as Phthinylla fracta belongs to
N. africanus. The main differences between the As-Sarrar taxon
and N. africanus are the great size and hypsodonty of the latter.
Unfortunately no M3 of N. africanus has been described and
illustrated to date.
Apodecter stromeri Hopwood, 1929. The holotype specimen of
Apodecter stromeri (AMNH 22538) is a right mandibular ramus
with m1-m3, from an unknown Lower Miocene locality of the
Namib desert, south of Luderitz Bay (Hopwood 1929).
Lavocat (1973, p. 42), without justification, reallocated
Apodecter stromeri to the genus Paraphiomys. He noted (p. 44)
that the specimens of P. stromeri found at Rusinga (Kenya,
Lower Miocene) are smaller than those from Songhor (Kenya,
Lower Miocene) and lack the metalophulid II whereas the lower
molars from Songhor frequently show it. Somewhat unusually,
he thought that these differences were insufficient to justify a
specific distinction, but that they are nonetheless reliable indica-
tions of the existence of two subspecies of P. stromeri. Therefore,
he erected Paraphiomys stromeri stromeri to accommodate both
the material from Rusinga and the holotype specimen of Apodec-
ter stromeri, and Paraphiomys stromeri hopwoodi for the material
from Songhor.
We consider that the morphological differences between
these two ‘subspecies’ are sufficient to elevate them to species
rank (Lopez Antonanzas et al. 2004). However, there is no
reason to consider the smaller, more hypsodont and older
holotype of Apodecter stromeri and the remaining specimens of
Paraphiomys stromeri stromeri to be representatives of the same
species. In a very recent systematic study of the Thryonomyidae
226 P A L A E O N T O L O G Y , V O L U M E 4 8

(Lopez-Antonanzas 2004; Lopez-Antonanzas et al. 2004), the
genus Apodecter has been rehabilitated, the material from Ru-
singa has been considered a new species, Paraphiomys renelavo-
cati, and the material from Songhor has been designated as
P. hopwoodi.
The As-Sarrar material differs from Apodecter stromeri in hav-
ing a metalophulid II in the lower molars (absent in Apodecter)
and in the anterior part of the tooth being much wider than the
posterior one (it is only slightly wider in A. stromeri).
Unpublished material from the Namibian locality of Elisabeth
Felt (Mein in prep.) allows a comparison of the upper molars
of P. knolli with those of Apodecter. The basic morphology
of the upper dentition of both taxa is very close. However
the specimens of Apodecter are higher crowned than those from
As-Sarrar.
Paraphiomys hopwoodi Lavocat, 1973. The holotype of Paraph-
iomys hopwoodi (Lavocat 1973, p. 44) is a left mandibular frag-
ment (KNM SO 627) with d4-m2 and an erupting m3 from the
Lower Miocene of Songhor (Text-fig. 1). Although the lower
molars of P. hopwoodi are larger than AS-1008, they are very
similar in morphology. There are slight differences between the
upper molars of the two taxa: those of the As-Sarrar specimen
have a metaloph (absent in P. hopwoodi), a more marked antero-
posterior compression and a hypocone that is more labially
displaced.
Paraphiomys renelavocati Lopez-Antonanzas, Sen and Mein,
2004. The holotype of this species is a mandible with d4-m3
from the Early Miocene of Rusinga figured by Lavocat (1973,
pl. 26, fig. 9). It is housed in the National Museums of Kenya
(Nairobi) where it is available for study under reference number
KNM RU 2208. The dentition of this taxon is very similar in
size to specimens from As-Sarrar, but the morphology is differ-
ent enough in lacking a metalophulid II on the lower molars to
consider the latter to be distinct. Both taxa have a mesoloph on
the upper molars.
Kochalia geespei de Bruijn and Hussain, 1985. The holotype of
this species is a left d4 or p4 (p4 according to de Bruijn and
Hussain 1985, pl. 1, fig. 7a–b) from the middle part of the
Lower Manchar Formation of Pakistan (locality H-GSP 82.24).
The lower molars of Kochalia geespei mainly differ from the
As-Sarrar specimen in lacking any trace of metalophulid II and
in being more lophodont. The upper molars of the two taxa are
very similar. However, K. geespei usually lacks the metaloph
whereas in all of the As-Sarrar specimens it is complete. The
two taxa are comparable in size.
Kochalia sp. De Bruijn and Hussain (1985) also described a sec-
ond species of Kochalia on the basis of first and second right
lower molars from the base of the Nari (Early Miocene) Forma-
tion at the locality H-GSP 81.14A. According to them, this is a
different taxon from K. geespei because of its smaller size, lower
crowned cheek teeth, and the presence of metalophulid II on the
two lower molars. De Bruijn and Hussain (1985) pointed out
the close similarity between these teeth and the lower molar
from As-Sarrar and we concur with them in considering that the
similar size and morphology strongly advocate that all of these
specimens could be assigned to the same species.
Epiphiomys coryndoni Lavocat, 1973. The holotype of this
species (KNM RU 2253) is a left mandibular fragment with
m1-2 from the Lower Miocene of Rusinga, Kenya (Lavocat
1973, p. 46). According to Lavocat (1973), this species has
also been found at Songhor (Kenya) and Napak (Uganda)
(Text-fig. 1).
One of the main differences between Epiphiomys coryndoni
and AS8-1100 is the greater development of the metalophulid II
in the former. With regard to the upper molars, those of E. co-
ryndoni show a mesoloph that is much more developed than in
specimens from As-Sarrar. The molars of E. coryndoni are only
slightly smaller than those of Paraphiomys knolli.
Paraphiomys occidentalis Lavocat, 1961. This species has been
reported from two Middle Miocene sites in Morocco: Beni-
Mellal (Lavocat 1961, p. 45; Jaeger 1977, p. 121) and Azdal (Bena-
mmi et al. 1995) (Text-fig. 1). Little material of Paraphiomys
occidentalis is known to date. The type (UM Ben. Mel. 340) is a
right fragmentary maxilla with P4-M1 (Lavocat 1961, pl. 4,
fig. 1) from Beni-Mellal and the cotype (UM Ben. Mel. 667)
consists of a right m2. The Moroccan site of Azdal has yielded a
single right mandibular ramus (Benammi et al. 1995). The main
difference between P. occidentalis and the specimens from As-
Sarrar is in the size, the former being much larger than any
extinct thryonomyid, except Neosciuromys africanus, Paraphiomys
simonsi and P. pigotti. The lower molars of P. occidentalis and
those of P. knolli have a vestigial metalophulid II and an antero-
labial cuspid, which is larger in the latter. The posterolophid is
more curved in P. knolli than in P. occidentalis.
With regard to the upper molars, P. occidentalis shows an
unreduced mesoloph and lacks the metaloph, whereas those
from As-Sarrar show a mesoloph and a short but distinct
metaloph. The protoloph is transverse in the As-Sarrar speci-
mens whereas it curves posteriorly in P. occidentalis.
Paraphiomys shipmani Denys and Jaeger, 1992. This species
was described by Denys and Jaeger (1992, p. 77) based on five
mandibular rami from the Middle Miocene of Fort Ternan,
Kenya (Text-fig. 1). The holotype (KNM FT 3305) is a right
mandible with d4-m3. Unfortunately, no upper tooth of this
taxon has been found to date. The lower molars of Paraphiomys
shipmani differ from P. knolli in lacking a metalophulid II. Fur-
thermore, the anterior part of the tooth in the latter is wider
than the posterior part, whereas in P. shipmani they are nearly
equal in width. Contrary to the condition in AS8-1100, the
lower molars of P. shipmani show the hypolophid bending back-
wards. The posterolophid is more curved in P. knolli than in
P. shipmani, in which it is nearly transverse.
Paraulacodus indicus Hinton, 1933. Hinton (1933, pp. 621–
622) established the taxon Paraulacodus indicus on the basis of
three specimens (G.S.I. D281, D282, D283) from the Siwalik
‘series’ in Pakistan (then part of northern India). Only a short
diagnosis without illustrations was given, but Black (1972) pro-
vided a thorough systematic revision of the species, including
L O P E Z A N T O N A N Z A S A N D S E N : N E W M I O C E N E R O D E N T 227

original drawings of the three specimens available to Hinton
(1933).
The holotype of this species (GSI D283) is an incomplete
right maxilla with P4-M2 (Black 1972, fig. 2a–c) from an
unknown locality. An isolated upper molar of Paraulacodus aff.
Paraulacodus indicus from the Middle Miocene Chinji Formation
of Pakistan has been reported by Wessels et al. (1982, p. 358,
pl. 2, fig. 14). However, the great difference in size and slight
morphological differences between this tooth and equivalent
teeth of P. indicus suggest that it does not belong to this species
(de Bruijn and Hussain 1985; Lopez-Antonanzas 2004; Lopez-
Antonanzas et al. 2004). De Bruijn and Hussain (1985) designa-
ted it as Paraulacodus sp., an identification that we follow.
De Bruijn and Hussain (1984, 1985) reported specimens of
Paraulacodus cf. P. indicus from the Pakistani locality H-GSP
82.14 (top of the Lower Manchar Formation, Middle Miocene).
Later, the middle Chinji Formation of Pakistan yielded new
specimens of Paraulacodus indicus (Flynn and Winkler 1994).
There are some important differences between the lower
molars from As-Sarrar and those of Paraulacodus indicus. The
m1–2 of P. indicus are much larger than the unique lower molar
recovered from As-Sarrar and show three transverse crests and
no remnant of metalophulid II (whereas Paraphiomys knolli
shows this). In P. knolli , the anterior half of the crown is appre-
ciably wider than the posterior half, whereas in Paraulacodus
indicus the anterior half of the crown is only slightly wider than
the posterior. The upper molars from As-Sarrar are smaller than
those of P. indicus and show a mesoloph, which is absent in the
Pakistani form. Whereas all As-Sarrar specimens show a short
metaloph connecting the metacone with the posteroloph, the
upper molars of P. indicus lack it. The protoloph is transverse in
P. knolli, whereas it bends towards the labial end of the anterol-
oph in the upper teeth of P. indicus.
Paraulacodus johanesi Jaeger, Michaux and Sabatier, 1980. The
holotype of this species (NMAA CHO 1) was described by Jaeger
et al. (1980, p. 367). It is an incomplete maxilla with P4-M2
(Jaeger et al. 1980, fig. 1a) from the Upper Miocene of Chorora,
Ethiopia (Jaeger et al. 1980). Additional material was found later
at the same locality (Geraads 1998, 2001, p. 104) and in the
Upper Miocene of northern Kenya in the lower member of the
Nawata Formation (Winkler 2003) and possibly in the Namu-
rungule Formation (Kawamura and Nakaya 1984, 1987). The
existence of a metalophulid II in AS8-1100 and the much larger
size of Paraulacodus johanesi suggest that there are no close rela-
tionships between them. In addition, contrary to the condition
in P. johanesi, the unique lower molar from As-Sarrar shows a
distinct, isolated anteroexternal cusp. Furthermore, its hypoflex-
us is transverse whereas in P. johanesi it is obliquely directed.
With regard to the upper molars, those of the As-Sarrar form
show a mesoloph, which is absent from those of P. johanesi. In
addition, the posterior half of the crown is relatively wider in
Paraphiomys knolli than in P. johanesi.
Paraphiomys afarensis Geraads, 2001. Geraads (1998, p. 210)
based his new species Paraphiomys chororensis on two upper
molars, two fragmentary upper molars, five fragmentary mandi-
bles, some isolated lower molars and several incisors, all from
the early Late Miocene Ethiopian locality of Chorora (Geraads
et al. 2002). The holotype is an upper molar (unnumbered)
housed in the National Museum of Addis Ababa (Geraads 1998,
pl. 2, fig. 6) and corresponds to the so-called Paraphiomys sp. 2
of Jaeger et al. (1980, p. 371, fig. 3a–b). New material collected
at Chorora has revealed that the holotype of P. chororensis is
actually a P4 of Paraulacodus johanesi. Geraads (2001, p. 103)
coined the name Paraphiomys afarensis to accommodate the
remaining specimens of the hypodigm of P. chororensis. This
species has been recently recorded from the Late Miocene of the
lower member of the Nawata Formation, Lothagam, Kenya
(Winkler 2003).
The morphology of the lower teeth of Paraphiomys afarensis
is very different from that of P. knolli. The three-lophed lower
molars of P. afarensis clearly differ from the four-lophed AS8-
1100. According to Geraads (1998, p. 210), P. afarensis retains
the mesoloph in the upper molars, which is also obvious in the
upper molars from As-Sarrar. With respect to size, the teeth of
P. afarensis are much larger than those of P. knolli.
Paraphiomys roessneri Mein, Pickford and Senut, 2000. The hol-
otype of this species is an upper left molar (GSW Ari 2) from
the Upper Miocene Harasib deposits of Namibia. Paraphiomys
roessneri is distinguished from the As-Sarrar specimens by the
absence of metalophulid II and in lacking a prominent labial
anteroconid on the lower molars. With regard to the upper
molars, one of the more important differences between the two
taxa is the absence of a mesoloph in the teeth of P. roessneri.
Paraphiomys australis Mein, Pickford and Senut, 2000. This
species was erected on the basis of 317 isolated teeth and two
maxillae from the Upper Miocene Harasib deposits of Namibia.
The holotype of this species (GSW Ari 13) is a right p4 (Mein
et al. 2000, fig. 1.13). In addition to the larger size of the teeth of
Paraphiomys australis in comparison with those from As-Sarrar,
this species differs notably in the absence of metalophulid II and
in lacking the anterior cusp on the lower molars. With regard to
the upper molars, one of the differences between the two taxa is
the absence of mesoloph in P. australis.
Paraphiomys orangeus Mein and Pickford, 2003. This species
has been reported from the Early Miocene (17Æ5–17 Ma) Orange
River deposit of Arrisdrift, Namibia (Mein and Pickford 2003,
p. 153). The holotype, AD 408¢97, is a left mandible with i, p4-m3
(Mein and Pickford 2003, fig. 7). This species differs from Para-
phiomys knolli in lacking metalophulid II on the lower molars.
SYSTEMATICS AND PHYLOGENY
McKenna and Bell (1997, pp. 194–195) included many
genera in the Thryonomyidae. However, conflicting opin-
ions have been expressed on the familial attribution of
Paraphiomys, Neosciuromys, Apodecter, Gaudeamus, Epi-
phiomys and Sacaresia. Thus, Paraphiomys is from time to
time seen as a taxon of the extinct family Phiomyidae
(Wood 1968; de Bruijn 1986; Denys and Jaeger 1992;
Geraads 1998). The same is true for Neosciuromys (Wood
228 P A L A E O N T O L O G Y , V O L U M E 4 8

1968), Apodecter (Wood 1968), Gaudeamus (Wood 1968)
and Epiphiomys (de Bruijn 1986). In addition, Sacaresia
has been considered as a taxon close to, but outside,
the thryonomyids by Flynn and Winkler (1994). Our
knowledge of the systematics and phylogeny of the
thryonomyids has recently been significantly improved
(Lopez-Antonanzas 2004; Lopez-Antonanzas et al. 2004).
With the purpose of finding the phylogenetic position
of Paraphiomys knolli within the family Thryonomyidae, a
cladistic analysis has been conducted. In addition to
P. knolli, the taxa involved in this analysis are the rodent
species commonly referred to as thryonomyid representa-
tives whose the validity has been established by Lopez-
Antonanzas (2004) and Lopez-Antonanzas et al. (2004),
namely Apodecter stromeri, Epiphiomys coryndoni, Gaude-
amus aegyptius, Kochalia geespei, Neosciuromys africanus,
Paraphiomys afarensis, P. australis, P. hopwoodi, P. occi-
dentalis, P. orangeus, P. pigotti, P. renelavocati, P. roessneri,
P. shipmani, P. simonsi, Paraulacodus indicus, P. johanesi,
Sacaresia moyaeponsi, Thryonomys gregorianus and
T. swinderianus.
The addition of the non-Thryonomyidae Metaphiomys
schaubi permits us to test the monophyly of the group
constituted by the species listed above. The outgroup
taxon Phiomys andrewsi (Phiomyidae) gives a direction to
the character transformation and an order of reading of
the successive branchings of the tree. Previous cladistic
analyses (Marivaux 2000; Marivaux et al. 2002) showed
that Phiomys is external to a clade comprising Paraphio-
mys and Paraulacodus. Interestingly, Phiomys does not
show a morphological gap with the taxa of the ingroup
and it was selected as outgroup in the phylogenetic analy-
sis of thryonomyid relationships presented by Winkler
(1992) and Flynn and Winkler (1994).
Twenty-two phylogenetically informative dental charac-
ters are coded. Fifteen characters are binary whereas seven
are multistate. Among the latter, characters 5, 12–14, 19
and 22 have two derived states and character 17 has three.
Owing to the lack of a priori information, the weight of
all characters is identical (1), and there is no rationale to
order the multistate characters; hence, all the possible
transformations of one state into another will cost one
step.
The software of phylogenetic reconstruction by parsi-
mony analysis used is PAUP version 3.1.1 (Swofford
1993). The number of terminal taxa and characters has
precluded the exact tree-building method, so a heuristic
search with stepwise addition (‘closest’ option) was per-
formed.
Four most parsimonious trees (80 steps long) have resul-
ted. Their characteristics are CI ¼ 0Æ375, RI ¼ 0Æ580. The
strict consensus tree is identical to that of the majority-rule
with two irresolutions (nodes 43 and 27). The semi-strict
consensus tree (Text-fig. 3) supports 20 internal nodes: it
TEXT -F IG . 3 . Semi-strict consensus tree generated by the cladistic analysis of the Thryonomyidae, showing phylogenetic position of
Paraphiomys knolli sp. nov. Dashed line indicates the node that collapses in the strict consensus tree. Length ¼ 80 steps, Consistency
Index ¼ 0Æ375, Retention Index ¼ 0Æ580.
L O P E Z A N T O N A N Z A S A N D S E N : N E W M I O C E N E R O D E N T 229

only shows basal irresolution with regard to the respective
placement of Metaphiomys schaubi, Sacaresia moyaeponsi,
and the remaining taxa of the ingroup.
One of the most interesting results of the cladistic ana-
lysis presented herein is the evidence of the polyphyletic
nature of the genus Paraphiomys as currently used. In
fact, only a handful of thryonomyid species are narrowly
related to P. pigotti, the type species of the genus, and
therefore could legitimately continue to be attributed to
it. These are P. knolli, P. hopwoodi, P. occidentalis and
P. renelavocati.
The monophyly of the two species of Paraulacodus
(node 26) is only present in 50 per cent of the most parsi-
monius trees. This monophyly is supported by the results
of the phylogenetic analysis of Winkler (1992) and Flynn
and Winkler (1994).
Based on extrinsic (age) considerations the branching
of Gaudeamus aegyptius in the crown group is unexpec-
ted. Lavocat (1973, p. 46) considered Gaudeamus to be
more progressive than Thryonomys. By contrast, Wood
(1968, fig. 17) considered Gaudeamus aegyptius to be a
direct ancestor of the Recent Thryonomys. Incidentally, de
Bruijn and Whybrow (1994) showed a thryonomyid m3
from Abu Dhabi (United Arab Emirates) that morphologi-
cally recalls both Gaudeamus and Thryonomys, and is
intermediate from both size and age (Late Miocene) points
of view. Therefore it turns out that Gaudeamus aegyptius
should be reallocated to the genus Thryonomys (a taxo-
nomic alternative compatible with the present cladogram
topology would be to restore the genus Choeromys for
Thryonomys gregorianus, as initiated by Thomas 1992).
Nevertheless, this part of the cladogram implies that there
is a missing lineage duration of about 30 myr. The posi-
tion of Gaudeamus aegyptius within species of Thryonomys
could be due to convergent evolution towards the same
kind of simplified cheek teeth. In fact, the incisors and
jaws of Gaudeamus and Thryonomys look nothing like
each other and the entire body of the former was about
the size of the head of a Thryonomys. Formally synony-
mizing Gaudeamus with Thryonomys would definitely
place too much confidence in the proposed phylogeny.
CONCLUSION
Winkler (1992, p. 244) suggested that the thryonomyid
rodent from As-Sarrar represents ‘a new species or poss-
ibly a new genus’. Whereas the first possibility has been
confirmed by our study, the existence of a new genus has
been dismissed. In fact, Paraphiomys knolli cannot be dis-
tinguished, in the present state of knowledge, from the
taxon referred to as ‘Kochalia sp.’ by de Bruijn and Huss-
ain (1985). Therefore, we consider provisionally that it is
the same as that from the Early Miocene Nari Formation
in Pakistan. However, it cannot be considered as conge-
neric with Kochalia geespei. Whereas the latter is quite
plesiomorphic (sister-group of the unnamed clade com-
prising ‘Paraphiomys’ simonsi and all other more derived
thryonomyids), P. knolli branches as the sister-species of
P. pigotti, the type species of the genus. Paraphiomys hop-
woodi constitutes the sister-group of (P. pigotti, P. knolli).
As these three species are morphologically fairly close,
there is little doubt that the latter must be considered to
belong to the genus Paraphiomys.
As indicated by de Bruijn and Hussain (1985, p. 162),
the presence of the same species of rodent in the Arabian
Peninsula and the Indian subcontinent provides evidence
for the absence of barriers precluding faunal exchange
between these two distant zones during the first half of the
Miocene. This is supported by the geographical distribution
of the more abundant species of Sayimys during the Early
and Middle Miocene. Paraphiomys represents an important
clade of thryonomyid rodents of this time. Although still
lacking in the Miocene fossil record of some areas such as
Turkey, representatives of this genus were without doubt
widely distributed from western and northern Africa to the
Indian subcontinent before vanishing apparently without
having given rise to any descendants.
Acknowledgements. D. Batten (University of Wales, Aberyst-
wyth), L. Flynn (Harvard University, Cambridge, USA), P. Hol-
royd (University of California, Berkeley), F. Knoll (Staatliches
Museum fur Naturkunde, Stuttgart) and P. Mein (Universite
Claude Bernard-Lyon 1, Villeurbanne) improved various aspects of
this paper through critical reading and ⁄ or discussion. A. Currant
(The Natural History Museum, London) kindly made available
for examination the holotype and paratypes of Paraulacodus
indicus, which are currently housed in his institution. The rodent
remains from Saudi Arabia were collected during field work
organized by H. Thomas (College de France, Paris) with permis-
sion of the Department of Antiquities and Museums in Riyadh.
REFERENCES
A N D R E W S , C. W. 1914. On the Lower Miocene vertebrates
from British East Africa, collected by Dr. Felix Oswald. Quar-
terly Journal of the Geological Society of London, 70, 163–186.
B E N A M M I , M., O R T H , B., V I A N E Y - L I A U D, M.,
CH A I M A N E E , Y., S UT E E T H OR N , V., FE R A U D, G.,
HE R N A N DE Z , J. and J A E G E R , J. J. 1995. Microm-
ammiferes et biochronologie des formations neogenes du flanc
sud du Haut-Atlas Marocain: implications biogeographiques,
stratigraphiques et tectoniques. Africa Geoscience Review, 2,
279–310.
B L A CK , C. C. 1972. Review of fossil rodents from the Neogene
Siwalik Beds of India and Pakistan. Palaeontology, 15, 238–
266.
B R U I J N , H. de 1986. Is the presence of the African family
Thryonomyidae in the Miocene deposits of Pakistan evidence
230 P A L A E O N T O L O G Y , V O L U M E 4 8

for fauna exchange? Koninklijke Nederlandse Akademie Van
Wetenschappen, Proceedings, Series B, 89, 125–134.
—— and H U S S A I N , T. S. 1984. The succession of rodent fau-
nas from the lower Manchar Formation of southern Pakistan
and its relevance for the biostratigraphy of the Mediterranean
Miocene. Paleobiologie Continentale, 14, 191–204.
—— —— 1985. Thryonomyidae from the lower Manchar For-
mation of Sind, Pakistan. Koninklijke Nederlandse Akademie
Van Wetenschappen, Proceedings, Series B, 88, 155–166.
—— and W H Y B R O W , P. J. 1994. A Late Miocene rodent
fauna from the Baynunah Fomation, Emirate of Abu Dhabi,
United Arab Emirates. Koninklijke Nederlandse Akademie van
Wetenschappen, Proceedings, Series B, 97, 407–422.
D E N Y S , C. and J A E GE R , J. J. 1992. Rodents of the Miocene
site Fort Ternan (Kenya), first part: phiomyids, bathyergids,
sciurids and anomalurids. Neues Jahrbuch fur Geologie und
Palaontologie, Abhandlungen, 185, 63–84.
F L Y N N , L. J., J A C OB S , L. L. and S . 1983. La diversite de
Paraulacodus (Thryonomyidae, Rodentia) et des groupes
apparentes pendant le Miocene. Annales de Paleontologie, 69,
355–366.
—— and W I N KL E R , A. J. 1994. Dispersalist implications of
Paraulacodus indicus: a South Asian rodent of African affinity.
Historical Biology, 9, 223–235.
G E R A A D S , D. 1998. Rongeurs du Miocene superieur de
Chorora (Ethiopie): Cricetidae, Rhizomyidae, Phiomyidae,
Thryonomyidae, Sciuridae. Palaeovertebrata, 27, 203–216.
—— 2001. Rongeurs du Miocene superieur de Chorora, Ethio-
pie: Murinae, Dendromurinae et conclusions. Palaeovertebrata,
30, 89–109.
—— Z E R E S E N A Y , A. and HE R V E , B. 2002. The late Mio-
cene mammalian fauna of Chorora, Awash Basin, Ethiopia:
systematics, biochronology and the 40K–40Ar ages of the asso-
ciated volcanics. Tertiary Research, 21, 113–122.
H E N D E Y , Q. B. 1978. Preliminary report on the Miocene ver-
tebrates from Arrisdrift, South West Africa. Annals of the
South African Museum, 76, 1–41.
H I N T O N , M. A. C. 1933. Diagnoses of new genera and species
of rodents from Indian Tertiary deposits. Annals and Maga-
zine of Natural History, 72, 620–622.
H O PW O O D, A. T. 1929. New and little-known mammals
from the Miocene of Africa. American Museum Novitates, 344,
2–9.
H U GU E N E Y , M. and A DR O V E R , R. 1990. Rongeurs (Rod-
entia Mammalia) de l’Oligocene de Sineu (Baleares, Espagne).
Paleontologia i Evolucio, 23, 157–169.
J A E G E R , J. J. 1977. Rongeurs (Mammalia, Rodentia) du
Miocene de Beni-Mellal. Palaeovertebrata, 7, 91–125.
—— M I CH A UX , J. and S A B A T I E R , M. 1980. Premieres
donnees sur les rongeurs de la formation de Ch’orora (Ethio-
pie) d’age Miocene superieur. I: Thryonomyides. Palaeoverteb-
rata, Memoire Jubilaire R. Lavocat, 365–374.
K A W A M UR A , Y and N A K A Y A , H. 1984. Thryonomid
rodent from the Late Miocene Namurungule Formation,
Samburu Hills, northern Kenya. African Study Monographs,
Supplementary Issue, 2, 133–139.
—— —— 1987. Additional materials of the Late Miocene
rodents from the Namurungule Formation in the Samburu
Hills, northern Kenya. African Study Monographs, Supple-
mentary Issue, 5, 131–139.
L A V O C A T , R. 1961. Le gisement de vertebres miocenes de
Beni Mellal (Maroc): etude systematique de la faune de mam-
miferes. Notes et Memoires du Service Geologique du Maroc,
155, 29–77.
—— 1973. Les rongeurs du Miocene d’Afrique orientale.
Memoires et Travaux de l’Institut de Montpellier, 1, 1–284.
L O P E Z - A N T O N A N Z A S , R. 2004. Neogene Ctenodactylidae,
Thryonomyidae, and Zapodidae (Rodentia) from the Middle
East: systematics, phylogeny, biostratigraphy, palaeogeography,
and palaeoecology. Unpublished PhD thesis, Museum national
d’Histoire naturelle, Paris, 323 pp.
—— S E N , S. and M E I N , P. 2004. Systematics and phylogeny
of the cane rats (Rodentia: Thryonomyidae). Zoological Journal
of the Linnean Society, 142, 423–444.
M A DD E N , C. T. 1972. Miocene mammals, stratigraphy and
environment of Muruorot Hill, Kenya. Paleobios, 14, 1–12.
M A R I V A U X , L. 2000. Les Rongeurs de l’Oligocene des Col-
lines Bugti (Balouchistan, Pakistan): nouvelles donnees sur la
phylogenie des Rongeurs paleogenes, implications biochrono-
logiques et paleobiogeographiques. Unpublished PhD thesis,
Universite des Sciences et Techniques du Languedoc, Mont-
pellier, France, 185 pp.
—— V I A N E Y - L I A UD , M., W E L CO M M E , J. L. and
J A E G E R , J. J. 2002. The role of Asia in the origin and diversi-
fication of hystricognathous rodents. Zoologica Scripta, 31,
225–239.
MCKE N N A , M. C. and B E L L , S. K. 1997. Classification of
mammals above the species level. Columbia University Press,
New York, 631 pp.
M E I N , P. and P I C K FO R D, M. 2003. Rodentia (other than
Pedetidae) from the Orange River deposits, Namibia. Memoirs
of the Geological Survey of Namibia, 19, 147–160.
—— and S E N U T , B. 2000. Late Miocene micromammals from
the Harasib karst deposits, Namibia. Part 1- Large muroids
and non-muroid rodents. Communications of the Geological
Survey of Namibia, 12, 375–390.
P A T T E R S ON , B. and W O O D, A. E. 1982. Rodents from the
Deseadan Oligocene of Bolivia and the relationships of the
Caviomorpha. Bulletin of the Museum of Comparative Zoology,
149, 371–543.
P I C KF O R D, M. 1981. Preliminary Miocene mammalian bios-
tratigraphy for western Kenya. Journal of Human Evolution,
10, 73–97.
P OC O CK , R. I. 1922. On the external characters of some hyst-
ricomorph rodents. Proceedings of the Zoological Society of Lon-
don, 1922, 365–427.
S H I P M A N , P., W A L KE R , A., CO U V E R I N G , J. van,
H OO KE R , P. J. and M I L L E R , J. A. 1981. The Fort Ternan
Hominoid Site, Kenya: geology, age, taphonomy, and paleoe-
cology. Journal of Human Evolution, 10, 49–72.
S T R O M E R , E. 1922. Erste mitteilung uber tertiare Wirbeltier-
Reste aus Deutsch-Sudwestafrika. Sitzungsberichte der Bayeris-
chen Akademie der Wissenschaften, 1921, 331–340.
—— 1924. Ergebnisse der Bearbeitung mitteltertiarer Wirbeltier
Reste aus Deutsch-Sudwest-Afrika. Sitzungsberichte der Bayeris-
chen Akademie der Wissenschaften, 1923, 253–270.
L O P E Z A N T O N A N Z A S A N D S E N : N E W M I O C E N E R O D E N T 231

—— 1926. Reste land und Susswasser Bewohnender Wirbeltiere
aus den Diamantfeldern Deutsch Sudwestafrikas. 107–153. In
KA I S E R , E. (ed.). Die Diamantenwuste Sudwestafrika. D. Rei-
mer, Berlin, 535 pp.
S W OF F OR D , D. L. 1993. PAUP, Phylogenetic Analysis Using
Parsimony. Illinois Natural History Survey, Champaign, 178
pp.
T H OM A S , H., S E N , S., KH A N , M., B A T TA I L , B. and
L I G A B UE , G. 1982. The Lower Miocene fauna of As-Sarrar
(Eastern Province, Saudi Arabia). Atlal, 5, 109–136.
T H OM A S , O. 1922. On the animals known as ‘‘Ground-Hogs’’
or ‘‘Cane-Rats’’ in Africa. Annals and Magazine of Natural
History, 9, 389–392.
W E S S E L S , W., BR U I J N , H. de, H US S A I N , S. T. and
L E I N D E R S , J. 1982. Fossil rodents from the Chinji Forma-
tion, Banda Daud Shah, Kohat, Pakistan. Koninklijke Neder-
landse Akademie Van Wetenschappen, Proceedings, Series B, 85,
337–364.
W I N KL E R , A. J. 1992. Systematics and biogeography of Mid-
dle Miocene rodents from the Muruyur beds, Baringo district,
Kenya. Journal of Vertebrate Paleontology, 12, 236–249.
—— 1994. The middle ⁄ upper Miocene dispersal of major
rodent groups between southern Asia and Africa. National Sci-
ence Museum, Monographs, 8, 173–184.
—— 2003. Rodents and lagomorphs from the Miocene and
Pliocene of Lothagam, northern Kenya. 169–198. In
L E A KE Y , M. G. and H A R R I S , J. M. (eds). Lothagam, the
dawn of humanity in eastern Africa. Columbia University
Press, New York.
—— and D OW N S , W. R. 2002. Rodents from the Early
Miocene of Uganda: preliminary results of renewed investiga-
tions. Journal of Vertebrate Paleontology, 22 (Supplement to
No. 3), 120A.
W O O D, A. E. 1968. The African Oligocene Rodentia. Peabody
Museum of Natural History, Yale University, Bulletin, 28,
23–105.
APPENDIX
List of characters used in this analysis.
Fifteen characters are binary whereas seven are multistate. The
plesiomorphic state (0) was obtained by outgroup comparison
with Phiomys andrewsi.
dp4
1. (0) replaced; (1) not replaced.
2. Anterior cingulum: (0) present; (1) weak or absent.
3. Metalophulid II: (0) present; (1) absent.
4. Metalophulid II: (0) transverse; (1) oblique.
5. Hypolophid: (0) posteriorly oblique; (1) transverse; (2) anteri-
orly oblique.
m1)2
6. Anterolabial cuspid: (0) absent; (1) present.
7. Anterolabial cuspid: (0) isolated; (1) partially submerged in
crest.
8. Metalophulid II: (0) present; (1) absent.
9. Mesoflexid: (0) not anteriorly directed; (1) anteriorly directed.
10. Mesoflexid: (0) equal or longer than the metaflexid; (1)
shorter than the metaflexid.
11. Anterolabial cingulum: (0) present; (1) weak or absent.
12. Hypolophid: (0) posteriorly oblique; (1) transverse; (2) ante-
riorly oblique.
13. Length: (0) m1 < m2; (1) m1 ¼ m2; (2) m1 > m2.
14. Width: (0) m1 < m2; (1) m2 ¼ m1; (2). m1 > m2.
m3
15. Length: (0) m3 > m2; (1) m3 < m2.
16. Anterolabial cingulum: (0) present; (1) weak or absent.
I
17. (0) ungrooved upper incisor; (1) one groove, (2) two
grooves, (3) three grooves.
M1)2
18. Mesoloph: (0) present; (1) absent.
19. Length: (0) M1 < M2; (1) M1 ¼ M2; (2) M1 > M2.
20. Metaloph: (0) present; (1) absent.
21. Protoloph: (0) transverse; (1) oblique.
22. Hypsodonty: (0) low; (1) semihypsodonty; (2) high.
Matrix of character codings used in the analysis of
relationships of all species of Thryonomyidae
Characters are listed above. Phiomys andrewsi is used as out-
group. Character codings: 0, postulated plesiomorphic condition
of character; 1, 2 and 3, postulated derived conditions of charac-
ter; –, uncodable character; ?, character state uncertain.
232 P A L A E O N T O L O G Y , V O L U M E 4 8

Taxa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Phiomys andrewsi 0 0 0 0 0 0 - 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Metaphiomys schaubi 0 0 0 1 2 0 - 0 1 0 0 2 0 0 0 0 0 0 0 0 0 0
Gaudeamus aegyptius 0 1 0 0 0 0 - 1 1 1 1 0 0 0 1 1 ? 1 0 1 1 1
Sacaresia moyaeponsi 0 0 0 1 0 0 - 0 0 0 0 0 0 0 1 0 1 1 0 1 1 0
Paraphiomys simonsi ? 1 1 - 2 1 0 1 1 0 1 0 0 0 0 0 0 0 0 1 0 2
Paraphiomys pigotti 1 1 1 - 2 1 0 0 0 1 1 2 0 0 0 1 0 0 1 0 0 1
Neosciuromys africanus 1 1 1 - 2 1 1 1 0 0 1 2 0 0 0 1 ? 0 0 0 1 2
Paraphiomys knolli sp. nov. ? ? ? ? ? 1 0 0 0 0 1 2 ? ? ? 1 ? 0 ? 0 0 0
Apodecter stromeri 1 1 1 - 2 1 1 1 0 0 0 1 2 2 1 1 0 0 0 0 0 1
Paraphiomys orangeus 1 0 1 - 1 1 1 1 1 0 1 1 0 0 1 1 0 0 0 0 0 1
Paraphiomys hopwoodi 1 1 1 - 2 1 0 0 0 0 1 2 1 0 1 1 0 0 ? 1 1 1
Paraphiomys renelavocati 1 1 1 - 2 1 1 1 0 0 0 1 0 1 1 1 0 0 1 1 1 1
Epiphiomys coryndoni ? 0 0 1 2 1 0 0 0 0 0 1 2 0 1 1 ? 0 2 0 0 1
Paraphiomys occidentalis 1 1 1 - 2 1 0 1 0 0 1 1 0 0 0 1 ? 0 2 1 1 1
Paraphiomys shipmani 1 1 1 - 0 1 1 1 1 0 1 0 2 0 1 ? ? ? ? ? ? 0
Kochalia geespei 0 1 1 - 1 1 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 1
Paraulacodus indicus 1 1 1 - 1 1 1 1 0 0 1 1 2 0 1 1 2 1 0 0 1 1
Paraulacodus johanesi 1 1 1 - 1 1 1 1 0 0 1 1 0 0 1 1 2 1 0 0 1 1
Paraphiomys afarensis 1 1 1 - 2 1 1 1 0 0 1 0 0 0 0 1 0 0 ? 0 1 1
Paraphiomys roessneri 1 0 1 - 1 0 - 1 0 0 0 0 0 2 1 1 0 1 2 0 0 1
Paraphiomys australis 1 0 1 - 2 0 - 1 0 0 0 0 2 2 1 0 ? 1 0 0 1 1
Thryonomys swinderianus 1 1 0 1 1 0 - 1 0 0 1 1 0 0 1 1 3 1 0 1 1 1
Thryonomys gregorianus 1 1 0 1 1 0 - 1 0 0 1 0 0 0 1 1 3 1 0 1 1 1
L O P E Z A N T O N A N Z A S A N D S E N : N E W M I O C E N E R O D E N T 233