
Dental evolution in Neophanomys (Rodentia, Octodontidae) from the late Miocene of central Argentina
13
Original article Dental evolution in Neophanomys (Rodentia, Octodontidae) from the late Miocene of central Argentina § Évolution dentaire chez les Neophanomys (Rodentia, Octodontidae) du Miocène tardif d’Argentine centrale Diego H. Verzi a, * , Emma C. Vieytes a , Claudia I. Montalvo b a Sección Mastozoología, Facultad de Ciencias Naturales y Museo de La Plata, CONICET, Paseo del Bosque s/n, 1900 La Plata, Buenos Aires, Argentina b Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina Received 9 August 2010; accepted 11 February 2011 Available online 14 October 2011 Abstract The evolutionary pattern of the molar morphology of the small caviomorph (Octodontidae) Neophanomys from the late Miocene Cerro Azul Formation of central Argentina is analyzed. Two new species (chronomorphs) are recognized, which constitute an anagenetically evolving lineage with a gradual and directional pattern of increasing molar hypsodonty. Dental changes related to increasing hypsodonty are comparable to those of the octodontid lineage Chasichimys also recovered from the Cerro Azul Formation. However, Neophanomys shows comparatively less variation in gross morphology and there are no evidences that this lineage achieved euhypsodonty. In contrast, important changes in enamel microstructure (schmelzmuster) are observed among different populations of Neophanomys, supporting the hypothesis that these changes can occur at least partially independently from modifications in dental gross morphology. The patterns of dental evolution detected in the Neophanomys and Chasichimys-Xenodontomys lineages and the unequivocal polarity of the changes involved, related to increasing hypsodonty, reinforce the hypothesis that chronological differences exist among late Miocene outcroppings of Cerro Azul Formation in central Argentina. # 2011 Elsevier Masson SAS. All rights reserved. Keywords: Octodontidae; Molar evolution; Schmelzmuster; Late Miocene; Biochronology Résumé On analyse le patron d’évolution molaire du petit caviomorphe Neophanomys (Octodontidae) de la Formation Cerro Azul, Miocène tardif, Argentine centrale. Deux nouvelles espèces sont reconnues (chronomorphes), qui constituent une lignée anagénétique avec un patron d’augmentation graduelle de l’hypsodontie molaire. Les changements associés à une augmentation d’hypsodontie sont équivalents à ceux détectés dans la lignée de Chasichimys (Octodontidae), enregistrée aussi dans la Formation Cerro Azul. Toutefois, Neophanomys montre moins de variations dans la morphologie générale des molaires et aucune indication que cette lignée a finalement acquis une euhypsodontie. Au contraire, des variations significatives dans la microstructure de l’émail (schmelzmuster) entre les échantillons de Neophanomys appuient l’hypothèse qu’elles puissent être au moins partiellement indépendantes de celles qui se produisent dans la morphologie générale. Les modifications dentaires détectées dans les lignées de Neophanomys et Chasichimys, et la polarité non équivoque de ces variations, associées à l’augmentation d’hypsodontie, renforcent l’hypothèse selon laquelle des différences chronologiques existent entre les affleurements du Miocène tardif de la Formation Cerro Azul. # 2011 Elsevier Masson SAS. Tous droits réservés. Mots clés : Octodontidae ; Évolution molaire ; Schmelzmuster ; Miocène tardif ; Biochronologie 1. Introduction Octodontoidea are the most widely diversified group of hystricognath rodents from South America. Systematic studies of extinct and living representatives allow recognition of nearly Available online at www.sciencedirect.com Geobios 44 (2011) 621–633 § Corresponding editor: Gilles Escarguel. * Corresponding author. E-mail address: [email protected] (D.H. Verzi). 0016-6995/$ – see front matter # 2011 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.geobios.2011.02.008
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Dental evolution in Neophanomys (Rodentia, Octodontidae) from the late Miocene of central Argentina

Original article
Dental evolution in Neophanomys (Rodentia, Octodontidae)from the late Miocene of central Argentina§
Évolution dentaire chez les Neophanomys (Rodentia, Octodontidae) du Miocènetardif d’Argentine centrale
Diego H. Verzi a,*, Emma C. Vieytes a, Claudia I. Montalvo b
a Sección Mastozoología, Facultad de Ciencias Naturales y Museo de La Plata, CONICET, Paseo del Bosque s/n, 1900 La Plata, Buenos Aires, Argentinab Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina
Received 9 August 2010; accepted 11 February 2011
Available online 14 October 2011
Abstract
The evolutionary pattern of the molar morphology of the small caviomorph (Octodontidae) Neophanomys from the late Miocene Cerro AzulFormation of central Argentina is analyzed. Two new species (chronomorphs) are recognized, which constitute an anagenetically evolving lineagewith a gradual and directional pattern of increasing molar hypsodonty. Dental changes related to increasing hypsodonty are comparable to those ofthe octodontid lineage Chasichimys also recovered from the Cerro Azul Formation. However, Neophanomys shows comparatively less variation ingross morphology and there are no evidences that this lineage achieved euhypsodonty. In contrast, important changes in enamel microstructure(schmelzmuster) are observed among different populations of Neophanomys, supporting the hypothesis that these changes can occur at leastpartially independently from modifications in dental gross morphology. The patterns of dental evolution detected in the Neophanomys andChasichimys-Xenodontomys lineages and the unequivocal polarity of the changes involved, related to increasing hypsodonty, reinforce thehypothesis that chronological differences exist among late Miocene outcroppings of Cerro Azul Formation in central Argentina.# 2011 Elsevier Masson SAS. All rights reserved.
Keywords: Octodontidae; Molar evolution; Schmelzmuster; Late Miocene; Biochronology
Résumé
On analyse le patron d’évolution molaire du petit caviomorphe Neophanomys (Octodontidae) de la Formation Cerro Azul, Miocène tardif,Argentine centrale. Deux nouvelles espèces sont reconnues (chronomorphes), qui constituent une lignée anagénétique avec un patrond’augmentation graduelle de l’hypsodontie molaire. Les changements associés à une augmentation d’hypsodontie sont équivalents à ceuxdétectés dans la lignée de Chasichimys (Octodontidae), enregistrée aussi dans la Formation Cerro Azul. Toutefois, Neophanomys montre moins devariations dans la morphologie générale des molaires et aucune indication que cette lignée a finalement acquis une euhypsodontie. Au contraire,des variations significatives dans la microstructure de l’émail (schmelzmuster) entre les échantillons de Neophanomys appuient l’hypothèsequ’elles puissent être au moins partiellement indépendantes de celles qui se produisent dans la morphologie générale. Les modifications dentairesdétectées dans les lignées de Neophanomys et Chasichimys, et la polarité non équivoque de ces variations, associées à l’augmentationd’hypsodontie, renforcent l’hypothèse selon laquelle des différences chronologiques existent entre les affleurements du Miocène tardif de laFormation Cerro Azul.# 2011 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Octodontidae ; Évolution molaire ; Schmelzmuster ; Miocène tardif ; Biochronologie
Available online at
www.sciencedirect.com
Geobios 44 (2011) 621–633
§ Corresponding editor: Gilles Escarguel.* Corresponding author.
E-mail address: [email protected] (D.H. Verzi).
0016-6995/$ – see front matter # 2011 Elsevier Masson SAS. All rights reserveddoi:10.1016/j.geobios.2011.02.008
1. Introduction
Octodontoidea are the most widely diversified group ofhystricognath rodents from South America. Systematic studiesof extinct and living representatives allow recognition of nearly
.

D.H. Verzi et al. / Geobios 44 (2011) 621–633622
80 genera from this continent (Wood and Patterson, 1959;Patterson and Wood, 1982; McKenna and Bell, 1997;Honeycutt et al., 2003; Frailey and Campbell, 2004; Emmons,2005; Galewski et al., 2005; Woods and Kilpatrick, 2005), mostof which belong to the families Octodontidae (includingCtenomyinae) and Echimyidae. The assignment of octodontoidspecies from the Oligocene-middle Miocene time interval toone or the other of these two families is controversial because ofthe retention of conservative dental morphologies (Frailey andCampbell, 2004; Vucetich and Vieytes, 2006; Vucetich et al.,2010). Since the late Miocene, the global cooling and dryingtrend that led to differentiation of savanna-adapted mammals indifferent continents (Pascual and Ortiz Jaureguizar, 1990; Janis,1993; Leakey et al., 1996; MacFadden and Cerling, 2006;Semprebon et al., 2004; Made et al., 2006) favored thedevelopment of increasing hypsodonty and acquisition ofderived molar morphologies in echimyids and especiallyoctodontids (Pascual, 1967; Verzi, 1999; Verzi et al., 1994);these new morphological and developmental dental pathwaysallow unequivocal recognition of both families in the lateMiocene-Pleistocene record of southern South America (Verzi,2002).
Nearly 650 specimens of octodontoids, among more than20,000 vertebrate remains, collected in different exposures ofthe Huayquerian (upper late Miocene) Cerro Azul Formation incentral Argentina, have allowed recognition of a pattern ofincreasing molar hypsodonty in these rodents (Verzi et al.,2008). Such a trend has been assessed in detail in the phyleticsequence of the octodontids Chasichimys-Xenodontomys(Verzi, 1999; Verzi et al., 2004), and has also been detectedin the echimyid Reigechimys (Verzi et al., 1994; Sostillo, 2009).Because of the fossil richness of the Cerro Azul Formation, newremains of the uncommon octodontid Neophanomys, one of thesmallest caviomorphs, have been recently recovered. Thesample now available supports the hypothesis that anevolutionary pattern towards increasing molar hypsodonty,like that of Chasichimys, also occurs in Neophanomys, aspreliminary advanced by Verzi and Montalvo (2008).
In this paper, the new material of Neophanomys is describedand compared to previously reported material (Rovereto, 1914;Verzi et al., 1999), and a new systematic arrangement of thegenus reflecting its evolutionary history is proposed. Bothontogenetic changes in gross molar morphology and variationin enamel microstructure, accompanying changes in the degreeof hypsodonty of Neophanomys in the Cerro Azul Formation,are revised. The molar evolution of Neophanomys is comparedto that described for the Chasichimys-Xenodontomys lineage,and differences between both patterns are analyzed. Thebiochronological implication of the evolutionary pattern ofNeophanomys in the context of the late Miocene of southernSouth America is discussed.
2. Material and methods
Extinct octodontids studied here are housed in thepaleontological collections of Facultad de Ciencias Exactasy Naturales, Universidad Nacional de La Pampa, Santa Rosa,
Argentina (GHUNLPam); Museo Argentino de CienciasNaturales ‘‘B. Rivadavia’’, Buenos Aires, Argentina (MACN);Museo Municipal de Ciencias Naturales ‘‘L. Scaglia’’, Mar delPlata, Argentina (MMP); Museo de La Plata, Argentina (MLP);Field Museum of Natural History, Chicago, USA (FMNH).Hypsodonty was estimated using the ratio of hypoflexid height/anterior transverse diameter of m1. This index is a goodestimate of hypsodonty in Neophanomys because this flexidoccupies most of the height of the labial side of the crown, and itis taken more directly and with less error than previousestimates of height of the entire molar from x-ray images (Verziet al., 1999). Nevertheless, we also provide an hypsodontyindex based on the latter measurement for materials in which itwas available (Suppl. Table S1). Transverse diameter waschosen because it is more stable than other occlusal diametersthrough ontogeny. To study enamel microstructure, teeth wereembedded in epoxy resin, ground in longitudinal (from thelabial side) and transversal sections with sandpaper andgrinding powder, and etched for three to four seconds with2N HCl to create morphological relief (modified from Flynnand Wahlert, 1978). After rinsing and drying, specimens weresputtercoated and examined with a Jeol JSM-T100 scanningelectron microscope (SEM). Width of the layers of differentenamel types and inclination and number of prisms of theHunter-Schreger bands were quantified using SEM micro-photographs of longitudinal sections. Angles of Hunter-Schreger bands inclination are expressed in relation to an axisperpendicular to the enamel-dentin junction (Martin, 1992: fig.2). Transverse sections were studied searching for schmelzmus-ter differences between the leading edge and the trailing edge ofthe tooth. Nomenclature of gross dental morphology followsWood and Patterson (1959) and Marivaux et al. (2002).Nomenclature of enamel microstructure follows Koenigswaldand Clemens (1992), and Koenigswald and Sander (1997).
3. Systematic paleontology
Order RODENTIA Bowdich, 1821.Suborder HYSTRICOMORPHA Brandt, 1855.Infraorder HISTRICOGNATHI Brandt, 1855.Family OCTODONTIDAE Waterhouse, 1839.Genus Neophanomys Rovereto, 1914.Type species: Neophanomys biplicatus Rovereto, 1914, late
Miocene, northwestern Argentina.Included species: The type species Neophanomys biplica-
tus, Neophanomys pristinus nov. sp., and Neophanomys recensnov. sp. The early Miocene octodontid Acaremys minutissimusAmeghino, 1887 (as Acarechimys in Patterson and Wood, 1982)likely belongs to Neophanomys; nevertheless, a review ofvariation in this species is still needed.
Distribution: Neophanomys has been recorded in upper lateMiocene deposits of western and central Argentina (Fig. 1).
Emended diagnosis: Very small-sized caviomorph, nearly20% smaller than Chasichimys. Masseteric crest of mandiblenot aligned with the notch for the tendon of the M. massetermedialis anterior, with its proximal portion more vertical thanin Chasichimys. Lower incisor high and narrow, suboval in

Fig. 1. Geographic location of the late Miocene deposits of Cerro AzulFormation, Huayquerías de San Carlos, and Valle de Santa María. Solid circlesindicate Neophanomys-bearing deposits.
Fig. 2. Left mandible. a: Neophanomys biplicatus MACN 8253 (holotype); b:‘‘Acarechimys’’ minutissimus MLP 15-188 (lectotype); c: Chasichimys scagliaiMMP 481-M (holotype). M: masseteric crest; N: notch for the tendon of musclemasseter medialis anterior. Scale bar: 5 mm.
D.H. Verzi et al. / Geobios 44 (2011) 621–633 623
cross-section, with curved enamelled anterior face and theenamel extended labially; different from those of Chasichimysand Chasicomys which are wider, subtriangular in cross-section, with a flat anterior enamelled face. Lightly wornmolars at least 15% lower, proportionally to the transversediameter, than in Chasichimys. Molar roots already formedwhen the metaflexid is still open. Dp4 tetralophodont and m1-2trilophodont in juvenile stages, with an infrequent vestigialanterofossetid in the m1. m3 trilophodont, different to that ofChasichimys which is tetralophodont; its intra-alveolar portionis oriented posteriorly over incisor. Molars without flexids orfossettids in older individuals, with bilobed to subquadrangularocclusal morphology. Hypoflexid markedly more persistentthan lingual flexids, almost reaching base of crown.
Neophanomys biplicatus Rovereto, 1914.Figs. 2, 3(g, h) and 41914. Neophanomys biplicatus Rovereto, p. 60, fig. 26.1914. Eumysops parvidens Rovereto, p. 219, fig. 87.1965. Cercomys parvidens Kraglievich, p. 262.Holotype: MACN 8253, left mandibular fragment with
intra-alveolar portion of the incisor and Dp4-m2.Referred material: MACN 8565, right mandibular frag-
ment with Dp4-m2 (holotype of Eumysops parvidens); FMNHP14347, right mandibular fragment with intra-alveolar portionof the incisor and Dp4-m3.
Locality and Horizon: Upper late Miocene deposits ofwestern Argentina (Fig. 1). The holotype MACN 8253 wasfound in the ‘‘Araucanense’’ (Huayquerian, upper lateMiocene) of Valle de Santa María (Catamarca Province,northwestern Argentina), but its precise stratigraphic prove-nance is unknown (Rovereto, 1914). FMNH P14347, referred tothis species (Marshall and Patterson, 1981; Verzi et al., 1999),was found in the same area, in the upper part of the‘‘Araucanense’’ (unit XVIIIa at the Chiquimil locality, below
a tuff of the overlying unit XIX dated at 6.02 Ma; Huayquerian;Marshall and Patterson, 1981). The holotype of Eumysopsparvidens MACN 8565, transferred to N. biplicatus (Verziet al., 1999), comes from the ‘‘Tunuyanense’’ (Huayquerian) ofthe east of the Meseta del Guadal, near the Huayquerías de SanCarlos (Mendoza Province, western Argentina; Rovereto,1914; Marshall et al., 1986; Yrigoyen, 1994).
Emended diagnosis: height of hypoflexid near 90% ofanterior transverse molar diameter in lightly worn m1. Closureof lingual flexids almost synchronous.
Remarks: the m2 of the holotype of N. biplicatus(MACN 8253) has a very small remnant of mesofossettid onthe lingual side of the anterior lobe; it joins the external enamelcrest behind the inflexion corresponding to closure of themesoflexid. In the center of the posterior lobe of this molar, avery small hole corresponds to the bottom of the metafossettid(Fig. 3h). These two vestiges indicate that the closing of bothfossettids was synchronous or nearly so. When the closure ofthese flexids is asynchronous, the traces of the resultantfossettids do not coexist (see below and Petter, 1973: fig. 2).
MACN 8565 and FMNH P14347 are more juvenilespecimens, with higher molar crowns than the holotype ofN. biplicatus. Both in the m1 and m2 of MACN 8565, the meso-and metaflexid have similar height. In FMNH P14347, theclosure of lingual flexids of these molars is nearly synchronous(Figs. 3 and 4; Table 1). According to these characters, thedegree of hypsodonty of MACN 8565 and FMNH P14347 isconsistent with that of the holotype of N. biplicatus, andintermediate between those of N. pristinus and N. recens;accordingly, we keep the name N. biplicatus for these materials(Marshall and Patterson, 1981; Verzi et al., 1999).
Neophanomys pristinus nov. sp.
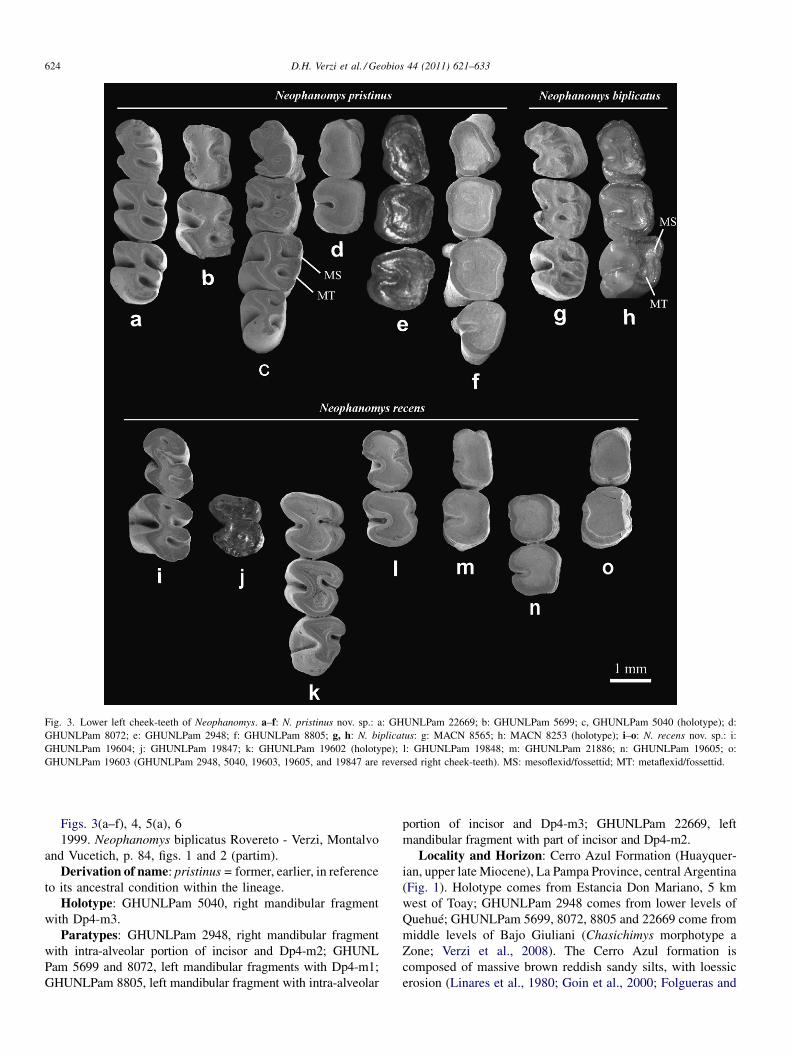
Fig. 3. Lower left cheek-teeth of Neophanomys. a–f: N. pristinus nov. sp.: a: GHUNLPam 22669; b: GHUNLPam 5699; c, GHUNLPam 5040 (holotype); d:GHUNLPam 8072; e: GHUNLPam 2948; f: GHUNLPam 8805; g, h: N. biplicatus: g: MACN 8565; h: MACN 8253 (holotype); i–o: N. recens nov. sp.: i:GHUNLPam 19604; j: GHUNLPam 19847; k: GHUNLPam 19602 (holotype); l: GHUNLPam 19848; m: GHUNLPam 21886; n: GHUNLPam 19605; o:GHUNLPam 19603 (GHUNLPam 2948, 5040, 19603, 19605, and 19847 are reversed right cheek-teeth). MS: mesoflexid/fossettid; MT: metaflexid/fossettid.
D.H. Verzi et al. / Geobios 44 (2011) 621–633624
Figs. 3(a–f), 4, 5(a), 61999. Neophanomys biplicatus Rovereto - Verzi, Montalvo
and Vucetich, p. 84, figs. 1 and 2 (partim).Derivation of name: pristinus = former, earlier, in reference
to its ancestral condition within the lineage.Holotype: GHUNLPam 5040, right mandibular fragment
with Dp4-m3.Paratypes: GHUNLPam 2948, right mandibular fragment
with intra-alveolar portion of incisor and Dp4-m2; GHUNLPam 5699 and 8072, left mandibular fragments with Dp4-m1;GHUNLPam 8805, left mandibular fragment with intra-alveolar
portion of incisor and Dp4-m3; GHUNLPam 22669, leftmandibular fragment with part of incisor and Dp4-m2.
Locality and Horizon: Cerro Azul Formation (Huayquer-ian, upper late Miocene), La Pampa Province, central Argentina(Fig. 1). Holotype comes from Estancia Don Mariano, 5 kmwest of Toay; GHUNLPam 2948 comes from lower levels ofQuehué; GHUNLPam 5699, 8072, 8805 and 22669 come frommiddle levels of Bajo Giuliani (Chasichimys morphotype aZone; Verzi et al., 2008). The Cerro Azul formation iscomposed of massive brown reddish sandy silts, with loessicerosion (Linares et al., 1980; Goin et al., 2000; Folgueras and

Fig. 4. Occlusal morphology and degree of hypsodonty of left m1 of Neophanomys pristinus nov. sp. (grey), Neophanomys biplicatus (white), and Neophanomysrecens nov. sp. (black). GHUNLPam 2948, 5040, 19603, 19605, and 19847 are reversed right cheek-teeth. Holotypes are indicated in bold. BG: Bajo Giuliani; Cl:Caleufú; DM: Don Mariano; QH: Quehué. Bars indicate values of hypsodonty index of m1 (HI; see Material and methods). Occlusal morphology indicatesontogenetic stage.
D.H. Verzi et al. / Geobios 44 (2011) 621–633 625
Zárate, 2009). This formation outcrops in eastern La PampaProvince and western Buenos Aires Province (as ‘‘EpecuénFormation’’ sensu Pascual, 1961), in central Argentina. Fivebiozones, covering the Chasicoan-late Huayquerian time span(late Miocene), have been recognized for this formation, basedmainly on the evolution of octodontoid rodents, especially theanagenetic pattern of change of the Chasichimys-Xenodont-omys lineage (Verzi et al., 2008).
Diagnosis: crown of the lightly worn m1 near 25% lowerthan in N. biplicatus; height of hypoflexid of this molar nearly70% of anterior transverse molar diameter. Closure of lingualflexids almost synchronous.
Description: the hypodigm of Neophanomys pristinus nov.sp. was described by Verzi et al. (1999, as Neophanomysbiplicatus), except for GHNULPam 22669 which was foundrecently. In N. pristinus nov. sp., closure of the mesoflexid and
Table 1Dental measurements (mm) and hypsodonty index (HI) of Neophanomys.
Neophanomys biplicatus Neophanomys pristinus nov. sp.
MACN FMNH GHUNLPam
8253 8565 P14347 5040 2948 5699 8072 8805
Dp4 AP 1.63 1.57 1.41 1.66 1.49 1.74 1.58 1.41Dp4 AW 1.06 1.06 1.04 1.08 1.00 1.08 1.00 1.00Dp4 PW 1.18 1.24 1.15 1.16 1.08 1.24 1.16 1.08m1 AP 1.48 1.66 1.41 1.49 1.41 1.83 1.49 1.58m1 AW 1.39 1.39 1.33 1.41 1.33 1.41 1.41 1.41m1 PW 1.36 1.33 1.26 1.16 1.24 1.41 1.33 1.24m2 AP 1.51 1.69 1.48 1.74 1.49 – – 1.66m2 AW 1.42 1.27 1.37 1.33 1.58 – – 1.49m2 PW 1.21 1.21 1.15 1.33 1.41 – – 1.41m3 AP – – 1.04 1.24 – – – 1.33m3 AW – – 1.04 1.00 – – – 1.33m3 PW – – 0.52 0.66 – – – 1.00m1 HI 0.53 0.91 0.69 0.49 0.00 0.51 0.03 0.00
AP: anteroposterior length; AW: anterior width; D: decidual; PW: posterior width
metaflexid, and formation of corresponding fossettids, is almostsynchronous as in N. biplicatus (Fig. 3). An almost synchronousclosure of these flexids/fossettids is seen in N. pristinus nov. sp.GHUNLPam 5040, 5699 and 22669. However, beyond thesimilarity in the time of closure of flexids, molars of N. pristinusnov. sp. are less hypsodont than those of the holotype ofN. biplicatus and the specimens from western Argentinareferred to this species (FMNH P14347 and MACN 8565; Table1). Molars of GHUNLPam 5040 and 5699 retain flexids andfossettids despite having slightly lower crown than that of theholotype of N. biplicatus in which these structures arepractically absent (Figs. 3 and 4).
The most juvenile specimen GHUNLPam 22669 hastetralophodont Dp4 and trilophodont m1-2. Dp4 has ananterofossettid; a probable remnant of this fossettid is alsopresent in m1, forming a small enamel protuberance near the
Neophanomys recens nov. sp.
GHUNLPam
22669 19602 19603 19604 19605 19847 19848 21886
1.42 – – 1.77 – – 1.6 –
0.94 – – 1.05 – – 1.11 –
0.97 – – 1.14 – – 1.14 –
1.37 1.68 1.42 1.65 1.25 – 1.51 1.42 1.14 1.42 0.97 1.25 1.20 1.59 1.37 0.91 1.08 1.25 1.03 1.11 1.14 1.20 1.31 1.03 1.31 1.48 1.48 – 1.34 1.15 – 1.42 1.03 1.42 – – 1.37 – – 1.20 0.91 1.20 – – 1.20 – – 1.14 – 1.20 – – – – – –
– 1.14 – – – – – –
– 0.74 – – – – – –
0.67 0.81 0.00 1.32 0.00 0.80 0.72 0.35
. Holotypes in bold.

Fig. 5. Lingual (above) and labial (below) view. a. Left Dp4-m2 of Neopha-nomys pristinus nov. sp. GHUNLPam 22669. b. Left Dp4-m1 of Neophanomysrecens nov. sp. GHUNLPam 19604. In all cases forward is to the right, as labialviews are reversed. HP: hypoflexid; MS: mesoflexid; MT: metaflexid.
D.H. Verzi et al. / Geobios 44 (2011) 621–633626
anterior margin of the molar. The mesoflexid and metaflexid areopen in Dp4-m2; in lingual view, the mesoflexid is slightlydeeper than the metaflexid in Dp4-m1, while both flexids aresimilar in depth in m2 (Fig. 5). In labial view, the hypoflexidoccupies almost the entire height of the crown exposed in m1-2;it is separated from the roots by a low basal zone of the crownlacking flexids (see Section 4.1). Relative height of thehypoflexid in m1 is nearly 70% of the anterior transverse widthof the molar (Fig. 4).
In the m1-2 of GHUNLPam 5040 and m1 of 5699, meso-and metaflexid are closing or coming to close; closure of theseflexids is almost synchronous, with the mesoflexid being onlyslightly more persistent (Fig. 3). Height of the m1 hypoflexid ofthese specimens is near 50% of the anterior transverse diameterof molar (Fig. 4).
In the more worn individuals, GHUNLPam 2948, 8072 and8805, molars lack lingual flexids. The m1 of GHUNLPam 2948and 8805 lacks even the hypoflexid; the occlusal surface ofmolars acquires a quadrangular morphology that corresponds tothe basal sector of crown lacking flexids.
The enamel microstructure was analyzed in the m2 ofGHUNLPam 2948 and GHUNLPam 22669 (Figs. 6 and S1). Inboth specimens, the schmelzmuster (i.e., distribution ofdifferent enamel types in a single tooth) varied both alongthe height of the crown and between leading and trailing edges.
In GHUNLPam 2948 (Fig. 6[a]), width of the enamel band is190 mm in the trailing edge and 80 mm in the leading edge. Inthe upper part of the crown, the trailing edge is formed only byradial enamel (RE); this enamel type occupies 40% of thecrown height. In the rest of the crown, down to the base, theenamel band is formed by one external layer of Hunter-Schreger bands (HSB) and one thin layer of inner RE thatoccupies approximately 15% of the whole enamel band. TheHSB are almost horizontal, 58 to 108 in relation to the enamel-dentin junction (EDJ) and 4-5 prisms thick. On the leading edgethere is a very thin layer of inner RE, formed by two or threeprisms, and one layer of HSB that occupies the rest of the widthof the enamel. The HSB are formed by five prisms per band,with an angle of 208 respect to the EDJ. In a low upper portion,there is a thin layer of outer RE, thinning toward the base of thecrown until it disappears. In the trailing edge, the interprismatic
matrix (IPM) of the inner RE is at a right angle, whereas that ofthe HSB is at a very low angle to the prisms. In the leading edge,the IPM is at a high angle to the prisms of the HSB.
In GHUNLPam 22669 (Fig. 6[b]), the enamel band is170 mm wide in the trailing edge and 90 mm wide in the leadingedge. The trailing edge shows a low sector formed only by REin the upper part of the crown. Below this sector, toward thebase of the crown, the enamel has two layers: one of inner RE,variable in thickness between 20 and 50% of total width, andone external of HSB with bands four to five prisms thick and aninclination of 108 to 208. The HSB have a very low decussationangle. The leading edge is three-layered: one thin layer of innerRE that occupies 10% of the enamel, one of HSB, and one ofouter RE that occupies 15%. HSB are 4–6 prisms thick and areinclined 258 to 308. The IPM is arranged as inGHUNLPam 2948.
Neophanomys recens nov. sp.Figs. 3(i–o), 4, 5(b), 7Derivation of name: recens = young, recent, in reference to
its presumed derived and youngest condition within the lineage.Holotype: GHUNLPam 19602, left mandibular fragment
with intra-alveolar portion of incisor and m1-3.Paratypes: GHUNLPam 19603, right mandibular fragment
with intra-alveolar portion of incisor and Dp4-m1; GHUNLPam19605, right mandibular fragment with intra-alveolar portion ofincisor and m1-2; GHUNLPam 19604, left mandibular fragmentwith intra-alveolar portion of incisor and Dp4-m1; GHUNLPam19847, right mandibular fragment with m1; GHUNLPam 19848and 21886, left mandibular fragments with Dp4-m1.
Type locality and Horizon: 10 km southeast of Caleufú(358 410 3700 S, 648 400 800 W), La Pampa Province, centralArgentina; Cerro Azul Formation, Xenodontomys elongatusZone, late Huayquerian, latest Miocene (Verzi et al., 2008)(Fig. 1). A brief description of the geology and paleontology ofthe Caleufú site is given in Verzi et al. (2003), and Verzi andMontalvo (2008). Three levels have been recognized in thissite; all fossils were found in the lowest level.
Diagnosis: molars with higher crown than in otherNeophanomys species; crown of the lightly worn m1 near45% higher than in N. biplicatus. Height of hypoflexid of thismolar near 130% of anterior transverse molar diameter.Metaflexid notably less persistent than mesoflexid in Dp4-m2,resulting in an ontogenetic stage with octodontiform occlusalmorphology.
Description: the new material fits the description ofNeophanomys given by Verzi et al. (1999) except for greaterhypsodonty in molars and related occlusal features. Molars ofN. recens nov. sp. are higher than those of other Neophanomys(Fig. 4; Table 1). This greater hypsodonty is observed clearlyin a comparison of the less worn specimens of N. recens nov.sp., GHUNLPam 19604, and N. pristinus nov. sp.,GHUNLPam 22669, with similar occlusal morphology.
In N. recens nov. sp., the hypoflexid extends along most ofthe crown height and there is a low basal zone without flexids,as in N. pristinus nov. sp. and N. biplicatus. In contrast, depthand consequent ontogenetic persistence of the mesoflexid and

Fig. 6. SEM micrograph of longitudinal section of molars of Neophanomys pristinus nov. sp., showing the schmelzmuster of the leading and trailing edges. a. Rightm2 (GHUNLPam 2948). b. Left m2 (reversed) (GHUNLPam 22669). D: dentine; HSB: Hunter-Schreger bands; RE: radial enamel.
D.H. Verzi et al. / Geobios 44 (2011) 621–633 627
metaflexid vary among species. In N. recens nov. sp., themesoflexid is deeper, whereas the metaflexid is more ephemeralthan in the other species (Fig. 5). This situation results in anontogenetic sequence of occlusal morphology that goes throughan octodontiform stage, with open meso- and hypoflexid, not
detected in N. biplicatus and N. pristinus nov. sp. (Fig. 3). Theclosure sequence of flexids of N. recens nov. sp. is similar tothat of the other protohypsodont octodontids, Chasichimys,Phtoramys and Chasicomys. According to the evidenceprovided by Neophanomys and Chasichimys, the mesoflexid

Fig. 7. SEM micrograph of longitudinal section of right m1 of Neophanomys recens nov. sp. (GHUNLPam 19847), showing the schmelzmuster of the leading andtrailing edges. D: dentine; HSB: Hunter-Schreger bands; RE: radial enamel.
D.H. Verzi et al. / Geobios 44 (2011) 621–633628
becomes progressively more persistent in relation to themetaflexid in the most hypsodont representatives of each genus(Verzi, 1999; Verzi et al., 1999: 88).
GHUNLPam 19604 is the most juvenile specimen ofN. recens nov. sp. Its Dp4 is tetralophodont, with the secondlophid (metalophulid II?) represented by a spur whose lingualend joins to the anterolophid. The anteroflexid and metaflexidare almost closed, whereas the mesoflexid is deeper and inlingual view occupies approximately half of the exposed crown.In labial view, the hypoflexid occupies the whole exposedcrown. The m1 has a small anterofossettid, and the lingualmargin of the metaflexid is almost closing to form ametafossettid (Fig. 5). The height of the hypoflexid is nearly130% of the anterior transverse diameter of this molar. The m1-2 of GHUNLPam 19602 and m1 of GHUNLPam 19847 aremore worn; their metafossettid is small or even absent, whilethe mesoflexid is open (Fig. 3). In the m1 of GHUNLPam19602, the height of the hypoflexid is more than 80% of theanterior transverse diameter of the molar (Fig. 4).
The remaining specimens are more heavily worn. Only theDp4 of GHUNLPam 19848 has the mesoflexid restricted to theapex of crown. In the m1 of this specimen, and the molars ofGHUNLPam 21286, 19605 and 19603, there are no lingualflexids/fossettids; only a shallow fold indicating the place wherethe mesoflexid closed, persists in GHUNLPam 19848. Thehypoflexid is also absent in the m1 of GHUNLPam 19605 and theDp4-m1 of GHUNLPam 19603; the occlusal surface of thesemolars exposes the basal zone of the molar lacking flexids (Fig. 3).
The enamel microstructure was analyzed in the right m1 ofGHUNLPam 19847 (Figs. 7 and S1). Unlike N. pristinus nov.sp., there is no continuous layer of RE on the upper portion ofthe crown, and both trailing and leading edges have an externallayer of RE. The enamel band of the trailing edge is 180 mmwide, whereas that of the leading is 140 mm wide. Both trailingand leading edge are three-layered: two layers of RE, each
occupying 15% of the enamel band, surrounding the HSB. Inthe trailing edge, the HSB are five to six prisms thick and showan inclination of 158 to 208. In the leading edge, the HSB are 4–
5 prisms thick and they are inclined 258 to 308. The IPM isarranged as in N. pristinus nov. sp.
4. Discussion
Since the late Miocene, octodontids are represented by twoclearly defined crown groups, the subfamilies Ctenomyinae andOctodontinae (Reig, 1989; Honeycutt et al., 2003; Verzi, 2008).The differentiation of these subfamilies would have been aconsequence of acquisition of specializations linked to openenvironments, especially euhypsodont molars (Pascual, 1967;Verzi, 1999, 2008). It is part of a faunal change recordedworldwide, triggered by climatic changes that resulted inexpansion of arid environments (Pascual and Ortiz Jaureguizar,1990; Janis, 1993; Leakey et al., 1996; MacFadden and Cerling,2006; Semprebon et al., 2004; Made et al., 2006). The CerroAzul Formation attests to these changes for southern SouthAmerica. In this formation, octodontoids show an evolutionarypattern characterized by parallel trends toward increasing molarhypsodonty. From the oldest exposure in Cerro La Bota to theyoungest one in Caleufú, Octodontidae with rootless molars(crown Octodontinae and Ctenomyinae) increase in numberwhile protohypsodont Octodontidae and Echimyidae decrease(Figs. 1 and 8). This pattern can be studied at the level of asingle phyletic sequence in the octodontids Chasichimys-Xenodontomys. Both gross morphology of the molars andmicrostructure of the enamel of this lineage show patterns ofchange related to hypsodonty which led to the differentiation ofthe basal ctenomyine Xenodontomys (Verzi, 1999; Verzi et al.,2004). A similar evolutionary pattern is now recognized forNeophanomys. However, unlike that of Chasichimys-Xeno-dontomys, there is no evidence that this lineage ever achieved

D.H. Verzi et al. / Geobios 44 (2011) 621–633 629
euhypsodonty, and their relationships with the modernsubfamilies remain uncertain.
4.1. Hypsodonty and ontogenetic variation of molars ofNeophanomys: a comparison with Chasichimys
The most worn molars of adult specimens of N. pristinusnov. sp. and N. recens nov. sp. have the same simplified occlusalmorphology, i.e. without lingual flexids/fossettids, or evenwithout hypoflexid/hypofossettid; but more juvenile stagesoccurring together with adults in the samples of N. pristinusnov. sp. from Bajo Giuliani and N. recens nov. sp. from Caleufúshow that there is important ontogenetic variations in thepersistence of flexids and fossettids related to the differences inhypsodonty between both species (Fig. 3).
This pattern of increasing hypsodonty and related changes ofNeophanomys is comparable to that of the octodontid lineageChasichimys. In both Chasichimys and Neophanomys, themesoflexid is more persistent and the metaflexid moreephemeral in more hypsodont species (chronomorphs). Thehypoflexid is more persistent than the lingual flexids. Never-theless, there is a strong difference between the patterns ofdental evolution of these lineages. In the patterns of crowndevelopment of Neophanomys and Chasichimys, three stagescan be recognized according to the model proposed byKoenigswald (1993): an earliest stage (A), which correspondsto the formation of the apical portion of the crown with itsflexids/fossettids; a later stage (B), which corresponds to theformation of the enamel-covered side walls; finally, the stage ofroot formation (C) (Fig. 9). This is a simplification of the five-phase model proposed by Koenigswald (1993). In bothNeophanomys and Chasichimys, phase A changes during theevolution of hypsodonty, as suggested by modifications inpersistency of flexids. In Neophanomys, phase B does not showany significant change with increasing hypsodonty. Instead,
Fig. 8. Changes in the composition of the octodontoid faunas from the CerroAzul Formation (updated from Verzi and Montalvo, 2008). The relativeabundance of each octodontoid group is expressed as percentage of the totalof octodontoid specimens. Protohypsodont octodontoids are represented byOctodontidae and Echimyidae; euhypsodont octodontoids are the crown Octo-dontidae of the subfamilies Ctenomyinae and Octodontinae.
phase B is extended in Chasichimys, which would be related tothe delay of phase C of root formation; in Chasichimys, thebasal portion of the crown lacking flexids (enamel-covered sidewalls) increases progressively in height until it occupies thewhole crown in its derivative euhypsodont genus Xenodont-omys. In Xenodontomys, the occlusal enamel structures (i.e.flexids/fossettids) have been lost, and roots do not develop. As aresult, euhypsodonty is achieved, leading to the differentiationof the dental pattern of modern ctenomyines (Verzi, 1999). InNeophanomys, the hypoflexid increases its depth withincreasing hypsodonty; hence, the basal zone of the crownlacking flexids remains very low even in the most hypsodontN. recens nov. sp. There is no evidence that this lineage everachieved euhypsodont degree.
According to the available information on muroid rodentsfrom Tummers and Thesleff (2003), the length of phase Bwould depend on a change of the signal regulating the activityof the epithelium loop that surrounds the base of the molar. Thisloop generates the crown until a change in the regulatorysystem modifies its developmental path toward the formation ofroot. Differences in ontogenetic timing of the change of thesignal that regulates activity of the epithelium loop couldexplain differences in hypsodonty (Tummers and Thesleff,2003: fig. 1) such as those detected between Chasichimys-Xenodontomys (Verzi et al., 2004) and Neophanomys.
4.2. Hypsodonty and variation in enamel microstructure
Changes in schmelzmuster associated with increasinghypsodonty have been described for most major clades ofrodents, with characteristic patterns for each group (Koenigs-wald, 2004). Among caviomorphs, the molar enamel of93 species from more than 70 genera representing the four
Fig. 9. Labial view of left m2 of Neophanomys pristinus nov. sp.GHUNLPam 22669 (left) and Chasichimys morphotype a GHUNLPam 2217(right). Comparison shows differences in the extension of molar portionsrepresenting stages of formation of the apical section (A) and enamel-coveredside walls of the crown (B), and roots (C). Not to scale.

D.H. Verzi et al. / Geobios 44 (2011) 621–633630
major clades (Octodontoidea, Chinchilloidea, Cavioidea,Erethizontoidea) shows common patterns of variation of theschmelzmuster associated with hypsodonty, suggesting thatsimilar changes occurred independently in different groups(Vieytes, 2003). In particular, secondary acquisition of REassociated to increasing hypsodonty was detected in cavioids,in the stem group of the modern Caviidae (‘‘Eocardiids’’;Vieytes, 2003), and in the octodontoids related to the origin ofthe Ctenomyinae (Verzi et al., 2004). Such a secondaryacquisition of RE has not been found so far outside ofcaviomorph rodents.
Among octodontoids, the schmelzmuster of species withlow-crowned molars has a layer of RE in a low upper portion ofthe crown, and only HSB (or a thin layer of inner RE and HSB)forming the rest of the enamel band. Such a primitiveschmelzmuster is present in both leading and trailing edges ofthe brachyodont to mesodont molars of the early Miocene basalgenera Caviocricetus, Protadelphomys, Willidewu, Sciamys,and Acaremys (Vieytes, 2003; Verzi et al., 2004). From thispresumed ancestral condition, a loss of the upper RE andsecondary acquisition of layers of RE (internal and/or externalto the HSB) on the whole crown would have occurredindependently in different clades with increasing hypsodonty.The derived enamel microstructure of the Chasichimys-Xenodontomys lineage has been hypothesized to havedifferentiated from a schmelzmuster like that of such putativeancestral genera through the above-mentioned changes (Verziet al., 2004: fig. 8).
Schmelzmuster variation in Neophanomys follows thegeneral pattern of enamel change associated to increasinghypsodonty in caviomorphs (Fig. 10): loss of upper RE andprogressive secondary acquisition of RE in both leading andtrailing edges. It is remarkable that these changes in enamelmicrostructure occur within a context of limited variation ofgross morphology. Moreover, part of these changes takes placeat intraspecific level, within the boundaries of N. pristinus nov.sp. A similar pattern has been detected in the lineageChasichimys-Xenodontomys, in which a change of enamelmicrostructure occurs within the species (chronomorph)
Fig. 10. Right molars showing hypothetical transformation pathway of the schmelzQuehué. c. Neophanomys pristinus nov. sp. from Bajo Giuliani. d. Neophanomys releading edge; TE: trailing edge.
Xenodontomys ellipticus and then persists in the rest of thelineage (Verzi et al., 2004). Variation in molar enamelmicrostructure among species of a single genus is not frequentin caviomorphs (Koenigswald et al., 1994; Vieytes, 2003).Nevertheless, evidences provided by Neophanomys andXenodontomys support the idea that important changes in theenamel microstructure, at least partially independent fromchanges in gross morphology, occurred in lineages withincreasing hypsodonty.
4.3. Evolutionary pattern of Neophanomys andbiochronology
In the standard of the South American stages/ages, the timeinterval between the lower late Miocene Chasicoan and theearly Pliocene Montehermosan is known traditionally asHuayquerian. But the faunas of the Huayquerian stratotype,from the Huayquerías de San Carlos (Mendoza Province,western Argentina), are poorly known. In spite of this, we retainthe term ‘‘Huayquerian’’, commonly used in South Americanstratigraphy and chronology, to designate the latest Miocene(Zárate, 2005). Currently, the best represented fauna of this agein southern South America is that of the Cerro Azul Formation(including those of the ‘‘Epecuén Formation’’; Pascual andBocchino, 1963; Pascual and Bondesio, 1982; Montalvo andCasadío, 1988; Verzi et al., 1994, 1995; Goin et al., 2000;Zárate, 2005). Important differences exist between the faunasrecovered from different outcroppings of this formation (Goinet al., 2000; Urrutia et al., 2008; Verzi et al., 2008). On the basisof evidence provided by octodontoid rodents, at least part ofthis faunal variation among localities has been interpreted asrelated to temporal differences (Verzi, 1999). Since there is nostratigraphic superposition among bearing units, their biochro-nological interpretation was based on the polarity of dentalevolution of octodontoids, especially of the phyletic sequenceChasichimys-Xenodontomys (Verzi, 1999; Verzi et al., 2003,2008). The new evidence of the evolutionary pattern ofincreasing hypsodonty in Neophanomys, independent from thatof Chasichimys-Xenodontomys, supports this interpretation.
muster (not to scale). a. Caviocricetus. b. Neophanomys pristinus nov. sp. fromcens nov. sp. Diagrams of schmelzmuster represent longitudinal sections. LE:

D.H. Verzi et al. / Geobios 44 (2011) 621–633 631
The evolutionary history of Neophanomys and its taxono-mical representation are more complex than previouslyrealized. At least the species from central Argentina,N. pristinus nov. sp. and N. recens nov. sp., constitute ananagenetically evolved lineage with a gradual and directional(Hunt, 2007; Rasskin-Gutman and Esteve-Altava, 2008) patternof dental change. These successive chronomorphs (sensuMartin, 1993) show a change of the ontogenetic pathway ofmolars leading to increasing hypsodonty. Although anageneticchange represents adaptation but not speciation (Futuyma,1987), we recognize discrete, named species because itfacilitates systematic and biostratigraphic analyses.
In the Cerro Azul Formation, the increase of hypsodonty inNeophanomys co-occurs with that in the Chasichimys-Xenodontomys lineage. Although the amplitude and chronol-ogy of the observed changes are not comparable (Fig. 11), thesechanges show the same polarity in both lineages. The record ofthe most hypsodont chronomorph N. recens nov. sp. in Caleufú(Xenodontomys elongatus Zone, late Huayquerian, latestMiocene) supports the younger age of these sediments withrespect to those of Bajo Giuliani (Chasichimys morphotype aZone, Huayquerian, upper late Miocene) where the chron-omorph N. pristinus nov. sp. is recorded. In addition, variationin the enamel of N. pristinus nov. sp., as showed byGHUNLPam 2948 and 22669, suggests that Quehué locality
Fig. 11. Stratigraphic distribution of octodontoid lineages from the late Miocene of cChasichimys-Xenodontomys and Reigechimys follow Verzi et al. (1994, 2004, 200
could be older than Bajo Giuliani. This is consistent with therecord of the echimyid Reigechimys plesiodon in the former,and the presence of a derived representative of the youngestchronomorph of this lineage, R. octodontiformis, in the latter(Fig. 11; Verzi et al., 2008; Sostillo, 2009).
Neophanomys has not been found in localities of the CerroAzul Formation, whose presumed biochronological ages rangebetween those of Bajo Giuliani and Caleufú, i.e. BarrancasColoradas and El Guanaco (Xenodontomys simpsoni Zone,Huayquerian, upper late Miocene; Fig. 11). This could be due totaphonomical reasons; Neophanomys is a rare taxon probablybecause of its small size (Verzi and Montalvo, 2008). BarrancasColoradas and El Guanaco are among the poorest fossilslocalities of the Cerro Azul Formation. Bajo Giuliani andCaleufú, in contrast, bear abundant and taxonomically diversemicromammal assemblages in a reduced area, the genesis ofwhich is interpreted as an accumulation by predators’ activity(Montalvo, 2002, 2004).
The evolutionary pattern of Neophanomys supports theusefulness of the concept of ‘‘stage of evolution’’ in phyleticsequences as a strategy for assigning a relative age to mammalfaunas. Information on the stage of evolution of different coevallineages, such as that provided by octodontoids in Cerro AzulFormation, is important to further support biochronologicalanalyses (Lindsay, 1990). In the particular case of this study,
entral Argentina. Stratigraphic scheme follows Verzi et al. (2008). Sequences of8) and Sostillo (2009).

D.H. Verzi et al. / Geobios 44 (2011) 621–633632
both the simultaneous changes detected in independent lineagesand the unequivocal polarity of these changes related toincreasing hypsodonty (Tummers and Thesleff, 2003: 1055),provide more reliability for the biochronological interpretationsof the Cerro Azul Formation in central Argentina.
Acknowledgements
We thank E. Tonni, M. Reguero, A. Dondas, A. Kramarz,and J. Flynn for granting access to materials under their care.We are grateful to C. Morgan and C. Deschamps for translation,and I. Olivares for assistance with illustrations and tables.Comments from A. Winkler, A. Kramarz, and an anonymousreviewer improved the manuscript. This paper was partiallysupported by ANPCyT PICT 01744, CONICET PIP 0270, andFCEyN-UNLPam 209 grants.
Appendix A. Supplementary data
Supplementary data (Table S1 and Fig. S1) associated withthis article can be found, in the online version, at doi:10.1016/j.geobios.2011.02.008.
References
Emmons, L.H., 2005. A revision of the genera of arboreal Echimyidae(Rodentia: Echimyidae, Echimyinae), with descriptions of two new genera.In: Lacey, E., Myers, P. (Eds.), Mammalian diversification: from chromo-somes to phylogeography (A Celebration of the Career of James L. Patton),133. University of California Press, Berkeley, pp. 247–309.
Frailey, C.D., Campbell Jr., K.E., 2004. Paleogene rodents from AmazonianPeru: the Santa Rosa Local Fauna. In: Campbell, Jr., K.E. (Ed.), ThePaleogene mammalian fauna of Santa Rosa, Amazonian Peru. NaturalHistory Museum of Los Angeles County, Science Series 40, Los Angeles,pp. 71–130.
Flynn, L.J., Wahlert, J.H., 1978. SEM study of rodent incisors: preparation andviewing. Curator 21, 303–310.
Folgueras, A., Zárate, M., 2009. La sedimentación neógena continental en elsector extrandino de Argentina central. Revista de la Asociación GeológicaArgentina 64, 692–712.
Futuyma, D.J., 1987. On the role of species in anagenesis. The AmericanNaturalist 130, 465–473.
Galewski, T., Mauffrey, J.F., Leite, Y.L.R., Patton, J.L., Douzery, E.J.P., 2005.Ecomorphological diversification among South American spiny rats(Rodentia: Echimyidae): a phylogenetic and chronological approach. Mo-lecular Phylogenetics and Evolution 34, 601–615.
Goin, F.J., Montalvo, C.I., Visconti, G., 2000. Los Marsupiales (Mammalia) delMioceno Superior de la Formación Cerro Azul (provincia de La Pampa,Argentina). Estudios Geológicos 56, 101–126.
Honeycutt, R.L., Dowe, D.L., Gallardo, M.H., 2003. Molecular systematics ofthe South American Caviomorph rodents: relationships among species andgenera in the family Octodontidae. Molecular Phylogenetics and Evolution26, 476–489.
Hunt, G., 2007. The relative importance of directional change, random walks,and stasis in the evolution of fossil lineages. Proceedings of the NationalAcademy of Sciences of the United States of America 104, 18404–18408.
Janis, C.M., 1993. Tertiary mammal evolution in the context of changingclimates, vegetation, and tectonic events. Annual Review of Ecology andSystematics 24, 467–500.
Koenigswald, W.v., 1993. Heterochronies in morphology and schmelzmuster ofhypsodont molars in the Muroidea (Rodentia). Quaternary International 19,57–61.
Koenigswald, W.v., 2004. The three basic types of schmelzmuster in fossil andextant rodent molars and their distribution among rodent clades. Palaeonto-graphica Abteilung A 270, 95–132.
Koenigswald, W.v., Clemens, W.A., 1992. Levels of complexity in the micro-structure of mammalian enamel and their application in studies of system-atics. Scanning Microscopy 6, 195–218.
Koenigswald, W.v., Sander, P.M., 1997. Schmelzmuster differentiation inleading and trailing edges, a specific biomechanical adaptation in rodents.In: Koenigswald, W.v., Sander, P. (Eds.), Tooth enamel microstructure.Balkema, Rotterdam, pp. 259–266.
Koenigswald, W.v., Sander, P.M., Leite, M.B., Mörs, T., Santel, W., 1994.Functional symmetries in the schmelzmuster and morphology of rootlessrodent molars. Zoological Journal of the Linnean Society 110, 141–179.
Leakey, M.G., Feibel, C.S., Bernor, R.L., Harris, J.M., Cerling, T.E., Stewart,K.M., Storrs, G.W., Walker, A., Werdelin, L., Winkler, A.J., 1996. Lotha-gam: A record of faunal change in the Late Miocene of East Africa. Journalof Vertebrate Paleontology 16, 556–570.
Linares, E., Llambías, E., Latorre, C., 1980. Geología de la provincia de LaPampa. República Argentina y Geocronología de sus rocas metamórficas yeruptivas. Revista Asociación Geológica Argentina 35, 87–146.
Lindsay, E.H., 1990. The setting. In: Lindsay, E., Fahlbusch, V., Mein, P.(Eds.), European Neogene mammal chronology. Plenum Press, New York,pp. 1–14.
MacFadden, B.J., Cerling, T.E., 2006. Mammalian herbivore communities,ancient feeding ecology, and carbon isotopes: a 10-million-year sequencefrom the Neogene of Florida. Journal of Vertebrate Paleontology 16, 103–
115.Made, J., van der, Morales, J., Montoya, P., 2006. Late Miocene turnover in the
Spanish mammal record in relation to palaeoclimate and the MessinianSalinity Crisis. Palaeogeography Palaeoclimatology Palaeoecology 238,228–246.
Marivaux, L., Vianey-Liaud, M., Welcomme, J.-L., Jaeger, J.-J., 2002. The roleof Asia in the origin and diversification of hystricognathous rodents.Zoologica Scripta 31, 225–239.
Marshall, L.G., Drake, R.E., Curtis, G.H., 1986. 40K-40Ar age calibration ofLate Miocene-Pliocene mammals-bearing Huayquerías and Tunuyan For-mations, Mendoza province, Argentina. Journal of Paleontology 60, 448–
457.Marshall, L.G., Patterson, B., 1981. Geology and geochronology of the
mammal-bearing Tertiary of the Valle de Santa María and Río CorralQuemado, Catamarca Province. Argentina. Fieldiana Geology 9, 1–80.
Martin, R.A., 1993. Patterns of variation and speciation in Quaternary rodents.In: Martin, R., Barnosky, A. (Eds.), Morphological changes in Quaternarymammals in North America. Cambridge University Press, New York,pp. 226–280.
Martin, T., 1992. Schmelzmikrostruktur in den inzisiven alt- und neuweltlicherhystricognather nagetiere. Palaeovertebrata, Mémoire extraordinaire, 1–
168.McKenna, M.C., Bell, S.K., 1997. Classification of mammals above the species
level. Columbia University Press, New York.Montalvo, C.I., 2002. Taphonomic analysis of the Mio-Pliocene micromam-
mal assemblage (Cerro Azul Formation), Caleufú, La Pampa, Argentina.In: De Renzi, M., Pardo Alonso, M., Belinchón, M., Peñalver, E.,Montoya, P., Márquez-Arriaga, A. (Eds.), Current topics on taphonomyand fossilization. TAPHOS, Valencia, pp. 353–359.
Montalvo, C.I., 2004. Paleobiología de la Asociación faunística de Caleufú (LaPampa, Formación Cerro Azul, Mioceno superior-Plioceno inferior), através de análisis tafonómicos. Ph.D. thesis, Universidad Nacional de LaPlata (unpublished).
Montalvo, C.I., Casadío, S., 1988. Presencia del género Palaeoctodon (Roden-tia, Octodontidae) en el Huayqueriense (Mioceno tardío) de la Provincia deLa Pampa. Ameghiniana 25, 111–114.
Pascual, R., 1961. Un nuevo Cardiomyinae (Rodentia, Caviidae) de la For-mación Arroyo Chasicó (Plioceno inferior) de la provincia de Buenos Aires.Ameghiniana 2, 61–64.
Pascual, R., 1967. Los roedores Octodontoidea (Caviomorpha) de la FormaciónArroyo Chasicó (Plioceno inferior) de la Provincia de Buenos Aires. Revistadel Museo de La Plata (Paleontología) 5, 259–282.

D.H. Verzi et al. / Geobios 44 (2011) 621–633 633
Pascual, R., Bocchino, A., 1963. Un nuevo Borhyaeninae (Marsupialia) delPlioceno medio de Hidalgo (La Pampa). Ameghiniana 3, 97–107.
Pascual, R., Bondesio, P., 1982. Un roedor Cardiatheriinae (Hydrochoeridae) dela Edad Huayqueriense (Mioceno tardío) de La Pampa. Sumario de losambientes terrestres en la Argentina durante el Mioceno. Ameghiniana 19,19–35.
Pascual, R., Ortiz Jaureguizar, E., 1990. Evolving climates and mammal faunasin Cenozoic South America. Journal of Human Evolution 19, 23–60.
Patterson, B., Wood, A.E., 1982. Rodents from the Deseadan Oligocene ofBolivia and the relationships of the Caviomorpha. Bulletin of the Museumof Comparative Zoology 149, 371–543.
Petter, F., 1973. Les noms de genre Cercomys, Nelomys, Trichomys et Proe-chimys (Rongeurs, Echimyides). Mammalia 37, 422–426.
Rasskin-Gutman, D., Esteve-Altava, B., 2008. The multiple directions ofevolutionary change. BioEssays 30, 521–525.
Reig, O.A., 1989. Karyotypic repatterning as one triggering factor in cases ofexplosive speciation. In: Fontdevila, A. (Ed.), Evolutionary biology oftransient unstable populations. Springer-Verlag, Berlin, pp. 246–289.
Rovereto, C., 1914. Los estratos araucanos y sus fósiles. Anales del Museo deHistoria Natural de Buenos Aires 25, 1–247.
Semprebon, G., Janis, C., Solounias, N., 2004. The diets of the Dromomer-ycidae (Mammalia: Artiodactyla) and their response to Miocene vegeta-tional change. Journal of Vertebrate Paleontology 24, 427–444.
Sostillo, R., 2009. Análisis de la variabilidad dentaria en Reigechimys (Roden-tia, Echimyidae) en la Formación Cerro Azul (Mioceno tardío, La Pampa).Bachelor’s thesis, Universidad Nacional de La Pampa (unpublished).
Tummers, M., Thesleff, I., 2003. Root or crown: a developmental choiceorchestrated by the differential regulation of the epithelial stem cell nichein the tooth of two rodent species. Development 130, 1049–1057.
Urrutia, J.J., Montalvo, C.I., Scillato Yané, G.J., 2008. Dasypodidae (Xenarthra,Cingulata) de la Formación Cerro Azul (Mioceno tardío) de la provincia deLa Pampa. Argentina. Ameghiniana 45, 289–302.
Verzi, D.H., 1999. The dental evidence on the differentiation of the ctenomyinerodents (Caviomorpha, Octodontidae, Ctenomyinae). Acta Theriologica 44,263–282.
Verzi, D.H., 2002. Patrones de evolución morfológica en Ctenomyinae (Roden-tia, Octodontidae). Mastozoología Neotropical 9, 309–328.
Verzi, D.H., 2008. Phylogeny and adaptive diversity of rodents of the familyCtenomyidae (Caviomorpha): delimiting lineages and genera in the fossilrecord. Journal of Zoology 274, 386–394.
Verzi, D.H., Deschamps, C.M., Montalvo, C.I., 2008. Biostratigraphy andbiochronology of the Late Miocene of central Argentina: evidence fromrodents and taphonomy. Geobios 41, 145–155.
Verzi, D.H., Montalvo, C.I., 2008. The oldest South American Cricetidae(Rodentia) and Mustelidae (Carnivora): late Miocene faunal turnover incentral Argentina and the Great American Biotic Interchange. Palaeogeo-graphy Palaeoclimatology Palaeoecology 267, 284–291.
Verzi, D.H., Montalvo, C.I., Tiranti, S.I., 2003. Un nuevo Xenodontomys(Rodentia, Octodontidae) del Mioceno tardío de La Pampa, Argentina.Patrón evolutivo y bioestratigrafía. Ameghiniana 40, 229–238.
Verzi, D.H., Montalvo, C.I., Vucetich, M.G., 1999. Afinidades y significadoevolutivo de Neophanomys biplicatus (Rodentia, Octodontidae) del Mio-ceno tardío-Plioceno temprano de Argentina. Ameghiniana 36, 83–90.
Verzi, D.H., Vieytes, E.C., Montalvo, C.I., 2004. Dental evolution in Xeno-dontomys and first notice on secondary acquisition of radial enamel inrodents (Rodentia, Caviomorpha, Octodontidae). Geobios 37, 795–806.
Verzi, D.H., Vucetich, M.G., Montalvo, C.I., 1994. Octodontid-like Echimyidae(Rodentia): an Upper Miocene episode in the radiation of the family.Palaeovertebrata 23, 199–210.
Verzi, D.H., Vucetich, M.G., Montalvo, C.I., 1995. Un nuevo Eumysopinae(Rodentia, Echimyidae) del Mioceno tardío de la provincia de La Pampa yconsideraciones sobre la historia de la subfamilia. Ameghiniana 32, 191–
195.Vieytes, E.C., 2003. Microestructura del esmalte de roedores Hystricognathi
sudamericanos fósiles y vivientes. Significado morfofuncional y filogené-tico. Ph.D. thesis, Universidad Nacional de La Plata (unpublished).
Vucetich, M.G., Vieytes, E.C., 2006. A Middle Miocene primitive octodontoidrodent and its bearing on the early evolutionary history of the Octodontoi-dea. Palaeontographica Abteilung A 27, 79–89.
Vucetich, M.G., Vieytes, E.C., Pérez, M.E., Carlini, A.A., 2010. The rodents fromLa Cantera and the early evolution of caviomorphs in South America. In:Madden, R., Carlini, A., Vucetich, M., Kay, R. (Eds.), The paleontology ofGran Barranca: evolution and environmental change through the MiddleCenozoic of Patagonia. Cambridge University Press, Cambridge, pp. 189–201.
Wood, A.E., Patterson, B., 1959. The rodents of the Deseadan Oligocene ofPatagonia and the beginnings of South American rodent evolution. Bulletinof the Museum of Comparative Zoology 120, 281–428.
Woods, C.A., Kilpatrick, C.W., 2005. Infraorder Hystricognathi Brandt, 1855.In: Wilson, D., Reeder, D. (Eds.), Mammal species of the world. JohnsHopkins University Press, Baltimore, pp. 1538–1600.
Yrigoyen, M., 1994. Revisión estratigráfica del Neógeno de las Huayquerías deMendoza septentrional, Argentina. Ameghiniana 31, 125–138.
Zárate, M.A., 2005. El Cenozoico tardío continental de la provincia de BuenosAires. In: de Barrio, R.E., Etcheverry, R.O., Caballé, M.F., Llambías, E.J.(Eds.), Geología y recursos minerales de la provincia de Buenos Aires.Relatorio 168 Congreso Geológico Argentino, La Plata, pp. 139–158.




















