
New Diseases Derived or Associated with the Tight Junction
14
REVIEW ARTICLE New Diseases Derived or Associated with the Tight Junction Marcelino Cereijido, a Rube ´n G. Contreras, a David Flores-Benı ´tez, a Catalina Flores-Maldonado, a Isabel Larre, a Agustı ´n Ruiz, b and Liora Shoshani a a Center For Research & Advanced Studies (CINVESTAV), Mexico City, Mexico b Laboratorio de Medicina, Facultad de Estudios Superioreselztacala y Cuautitla ´n, UNAM, Mexico Received for publication October 18, 2006; accepted February 20, 2007 (ARCMED-D-06-00449). The space between neighboring epithelial cells is sealed by the tight junction (TJ). When this seal is leaky, such as in the proximal tubule of the kidney or the gallbladder, sub- stances may cross the epithelium between the cells (paracellular pathway). Yet, when TJs are really hermetic, as is the case in the epithelium of the urinary bladder or the colon, substances can mainly cross the epithelium through the transcellular pathway. The struc- ture of the TJ involves (so far) some 50-odd protein species. Failure of any of these com- ponents causes a variety of diseases, some of them so serious that fetuses are not viable. A fast-growing number of diseases are recognized to depend or involve alterations in the TJ. These include autoimmune diseases, in which intestinal TJs allow the passage of an- tigens from the intestinal flora, challenging the immune system to produce antibodies that may cross react with proteins in the brain, thyroid gland or pancreas. TJs are also in- volved in cancer development, infections, allergies, etc. The present article does not cat- alogue all TJ diseases known so far, but describes one of each type as illustration. It also depicts the efforts being made to find pharmaceutical agents that would seal faulty TJs or release their grip to allow for the passage of large molecules through the upper respiratory and digestive tracts, such as insulin, thyroid, appetite-regulatory peptide, etc. Ó 2007 IMSS. Published by Elsevier Inc. Key Words: Tight junctions, Epithelia, Claudins, Cancer, Autoimmune diseases, Infections. Introduction Less than a decade ago, biomedicine started to discover a whole family of more than 50 ‘‘new’’ diseases related to the tight junction (TJ). We emphasize ‘‘new’’ because, of course, these diseases have been with us since the dawn of humankind, yet the novelty stems from the fact that they were not suspected to be due to failures of the TJ. With re- spect as to how we forecast that there would be more than 50 new diseases, the explanation is based on a fundamental tenet of evolutionary biology, affirming that natural selec- tion does not incur in overdesign. This means that in spite of having some reserve capacity to deal with occasionally extreme circumstances, every organ or molecule is poten- tially vulnerable when its reserve capacity is exceeded. Ac- cordingly, because today the TJ is known to consist of at least 50-odd different molecular species, each one with the capacity to fail, we may anticipate that there are, at least, an equal number of diseases waiting to be discovered. Unfortunately, a given protein can fail in more than one site, thereby multiplying the number of diseases. Some of these are already well documented. On this basis, the over- all strategy of the present review consists of briefly describ- ing the structure and function of the TJ, discusses what sort of diseases can be forecasted, mentions briefly the group of those diseases already known, and even refers to the phar- maceutical armamentarium that is being developed to fight them. A single cell in the ocean exchanges nutrients and waste products with a constant reservoir that is not exhausted or polluted (Figure 1, upper left). When a cell is instead part of a multicellular organism, e.g., a human brain (Figure 1, upper center), it exchanges with an extremely narrow Address reprint requests to: Dr. Marcelino Cereijido, Center For Re- search & Advanced Studies (CINVESTAV), Avenida Instituto Polite ´cnico Nacional 2508, 07360, Col. San Pedro Zacatenco, Mexico, D.F, Mexico; E-mail: cereijido@fisio.cinvestav.mx 0188-4409/07 $esee front matter. Copyright Ó 2007 IMSS. Published by Elsevier Inc. doi: 10.1016/j.arcmed.2007.02.003 Archives of Medical Research 38 (2007) 465e478
Transcript of New Diseases Derived or Associated with the Tight Junction

Archives of Medical Research 38 (2007) 465e478
REVIEW ARTICLE
New Diseases Derived or Associated with the Tight Junction
Marcelino Cereijido,a Ruben G. Contreras,a David Flores-Benıtez,a Catalina Flores-Maldonado,a
Isabel Larre,a Agustın Ruiz,b and Liora Shoshania
aCenter For Research & Advanced Studies (CINVESTAV), Mexico City, MexicobLaboratorio de Medicina, Facultad de Estudios Superioreselztacala y Cuautitlan, UNAM, Mexico
Received for publication October 18, 2006; accepted February 20, 2007 (ARCMED-D-06-00449).
The space between neighboring epithelial cells is sealed by the tight junction (TJ). Whenthis seal is leaky, such as in the proximal tubule of the kidney or the gallbladder, sub-stances may cross the epithelium between the cells (paracellular pathway). Yet, whenTJs are really hermetic, as is the case in the epithelium of the urinary bladder or the colon,substances can mainly cross the epithelium through the transcellular pathway. The struc-ture of the TJ involves (so far) some 50-odd protein species. Failure of any of these com-ponents causes a variety of diseases, some of them so serious that fetuses are not viable.A fast-growing number of diseases are recognized to depend or involve alterations in theTJ. These include autoimmune diseases, in which intestinal TJs allow the passage of an-tigens from the intestinal flora, challenging the immune system to produce antibodies thatmay cross react with proteins in the brain, thyroid gland or pancreas. TJs are also in-volved in cancer development, infections, allergies, etc. The present article does not cat-alogue all TJ diseases known so far, but describes one of each type as illustration. It alsodepicts the efforts being made to find pharmaceutical agents that would seal faulty TJs orrelease their grip to allow for the passage of large molecules through the upper respiratoryand digestive tracts, such as insulin, thyroid, appetite-regulatory peptide, etc. � 2007IMSS. Published by Elsevier Inc.
Key Words: Tight junctions, Epithelia, Claudins, Cancer, Autoimmune diseases, Infections.
Introduction
Less than a decade ago, biomedicine started to discovera whole family of more than 50 ‘‘new’’ diseases relatedto the tight junction (TJ). We emphasize ‘‘new’’ because,of course, these diseases have been with us since the dawnof humankind, yet the novelty stems from the fact that theywere not suspected to be due to failures of the TJ. With re-spect as to how we forecast that there would be more than50 new diseases, the explanation is based on a fundamentaltenet of evolutionary biology, affirming that natural selec-tion does not incur in overdesign. This means that in spiteof having some reserve capacity to deal with occasionallyextreme circumstances, every organ or molecule is poten-
Address reprint requests to: Dr. Marcelino Cereijido, Center For Re-
search & Advanced Studies (CINVESTAV), Avenida Instituto Politecnico
Nacional 2508, 07360, Col. San Pedro Zacatenco, Mexico, D.F, Mexico;
E-mail: [email protected]
0188-4409/07 $esee front matter. Copyright � 2007 IMSS. Published by Elsevdoi: 10.1016/j.arcmed.2007.02.003
tially vulnerable when its reserve capacity is exceeded. Ac-cordingly, because today the TJ is known to consist of atleast 50-odd different molecular species, each one withthe capacity to fail, we may anticipate that there are, atleast, an equal number of diseases waiting to be discovered.Unfortunately, a given protein can fail in more than onesite, thereby multiplying the number of diseases. Some ofthese are already well documented. On this basis, the over-all strategy of the present review consists of briefly describ-ing the structure and function of the TJ, discusses what sortof diseases can be forecasted, mentions briefly the group ofthose diseases already known, and even refers to the phar-maceutical armamentarium that is being developed to fightthem.
A single cell in the ocean exchanges nutrients and wasteproducts with a constant reservoir that is not exhausted orpolluted (Figure 1, upper left). When a cell is instead partof a multicellular organism, e.g., a human brain (Figure 1,upper center), it exchanges with an extremely narrow
ier Inc.
466 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
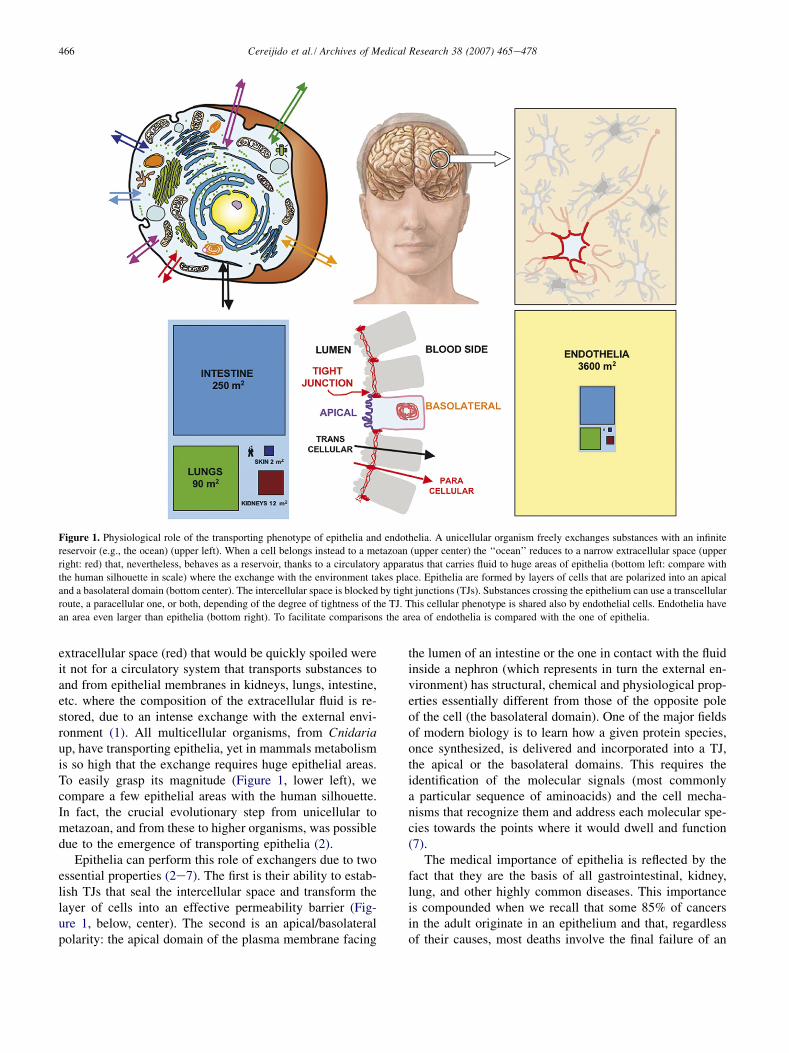
Figure 1. Physiological role of the transporting phenotype of epithelia and endothelia. A unicellular organism freely exchanges substances with an infinite
reservoir (e.g., the ocean) (upper left). When a cell belongs instead to a metazoan (upper center) the ‘‘ocean’’ reduces to a narrow extracellular space (upper
right: red) that, nevertheless, behaves as a reservoir, thanks to a circulatory apparatus that carries fluid to huge areas of epithelia (bottom left: compare with
the human silhouette in scale) where the exchange with the environment takes place. Epithelia are formed by layers of cells that are polarized into an apical
and a basolateral domain (bottom center). The intercellular space is blocked by tight junctions (TJs). Substances crossing the epithelium can use a transcellular
route, a paracellular one, or both, depending of the degree of tightness of the TJ. This cellular phenotype is shared also by endothelial cells. Endothelia have
an area even larger than epithelia (bottom right). To facilitate comparisons the area of endothelia is compared with the one of epithelia.
extracellular space (red) that would be quickly spoiled wereit not for a circulatory system that transports substances toand from epithelial membranes in kidneys, lungs, intestine,etc. where the composition of the extracellular fluid is re-stored, due to an intense exchange with the external envi-ronment (1). All multicellular organisms, from Cnidariaup, have transporting epithelia, yet in mammals metabolismis so high that the exchange requires huge epithelial areas.To easily grasp its magnitude (Figure 1, lower left), wecompare a few epithelial areas with the human silhouette.In fact, the crucial evolutionary step from unicellular tometazoan, and from these to higher organisms, was possibledue to the emergence of transporting epithelia (2).
Epithelia can perform this role of exchangers due to twoessential properties (2e7). The first is their ability to estab-lish TJs that seal the intercellular space and transform thelayer of cells into an effective permeability barrier (Fig-ure 1, below, center). The second is an apical/basolateralpolarity: the apical domain of the plasma membrane facing
the lumen of an intestine or the one in contact with the fluidinside a nephron (which represents in turn the external en-vironment) has structural, chemical and physiological prop-erties essentially different from those of the opposite poleof the cell (the basolateral domain). One of the major fieldsof modern biology is to learn how a given protein species,once synthesized, is delivered and incorporated into a TJ,the apical or the basolateral domains. This requires theidentification of the molecular signals (most commonlya particular sequence of aminoacids) and the cell mecha-nisms that recognize them and address each molecular spe-cies towards the points where it would dwell and function(7).
The medical importance of epithelia is reflected by thefact that they are the basis of all gastrointestinal, kidney,lung, and other highly common diseases. This importanceis compounded when we recall that some 85% of cancersin the adult originate in an epithelium and that, regardlessof their causes, most deaths involve the final failure of an

467Pathologies of the Tight Junction
epithelium, such as those in the kidney, the liver, etc. Thisprompted a great effort to develop techniques to understandthe cell and molecular physiology of epithelia (8,9). Thisinterest was multiplied for three additional reasons. In thefirst place, epithelia share a great deal of similarity, bothat the cell and molecular level, with endothelia. Secondly,a metastasis prospers provided migrating cells can induceendothelial cells to form blood vessels at the site of arrival,otherwise the mass of the tumor cannot be nourished. Third,the intense exchange of substances between biologicalcompartments alluded to above also takes place acrossendothelia. The importance of epithelia and endothelia isdepicted in Figure 1 (below, right) representing the sameillustration of Figure 1 (lower left) but in scale with the areaof endothelia (yellow). The size of these tissues as com-pared with the human silhouette stresses the importanceof understanding the cellular and molecular basis of theepithelial/endothelial phenotype.
Structure and Function of the TJ
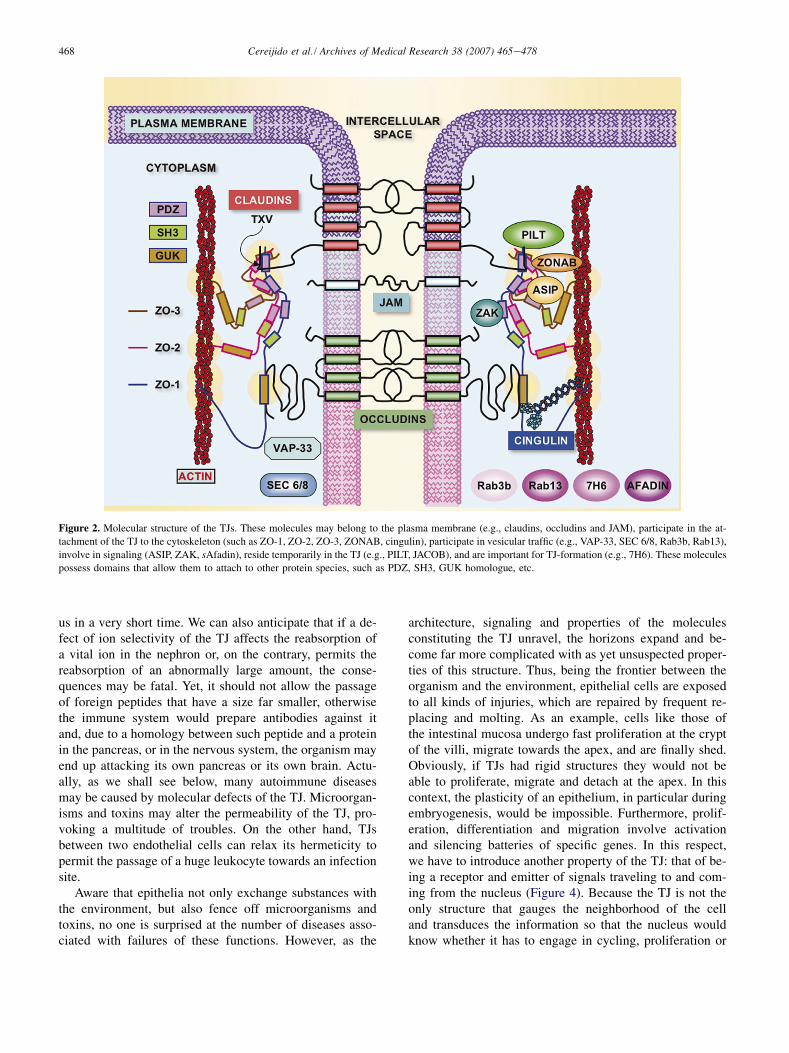
Figure 2 represents molecular details of a TJ located at theoutermost end of the intercellular space (Figure 1, below,center). The scope of this review does not justify digressinginto molecular details or what the name of each protein spe-cies stands for. Nevertheless, it is worthwhile to point outthat some are proteins that cross one [e.g., junctional adhe-sion molecule (JAM)] or four times the plasma membrane(e.g., claudins, occludins) (10,11). Another group of junc-tional proteins does not belong to the membrane, but is inintimate contact with proteins of the first group and anchorsthem to the cytoskeleton [e.g., zona occludens-1, -2, -3(ZO-1, ZO-2, ZO-3), cingulin] (12,13). Other groups aretranscription factors that regulate the expression of proteinsthat determine the proliferation, migration and differentia-tion of epithelial cells (e.g., ZO-1-associated nucleic acidbinding protein (ZONAB), ASIP, ZO-2 (14e16). Thereare members of well-known signaling cascades [short Afa-din (sAfadin), ZO-1 associated kinase (ZAK), guaninenucleotide exchange factor-H1 (GEF-H1)] (17,18), or com-ponents of the machinery involved in membrane polarizedtraffic [secretion protein 6/8 (member of the exocyst SEC6/8), monomeric GTPase of the Ras superfamily, isoform3b (Rab3b), VAMP/synaptobrevin-associated protein of33 kDa (VAP33)] (19,20) and proteins that reside temporar-ily in the TJ [e.g., protein incorporated later into TJs(PILT), JACOB] or are important for TJ formation (e.g.,7H6) (21). The number of molecular species integrated inthe TJ is in fact considerably larger, as we know that thereare many isoforms that perform different tasks.
Until a few years ago we knew in great detail manymembrane functions such as exchange diffusion, actionpotentials, and ion pumping, but ignored the molecularspecies that perform them. Today the situation is totally
reversed, as we know 50-odd molecules of the TJ (Figure 2),their aminoacid sequence, specific molecule�molecule as-sociations, functional domains [e.g., PDZ, Src homology 3(SH3), guanylate kinase (GUK), etc.] but ignore the func-tional role that these proteins play. Of course, the hydro-philic extracellular segments of molecules like claudinssuggest that they may establish homotypic adhesion witha similar protein present in the neighboring cell, presum-ably forming loops that might coil to afford hydrophilicconducts to allow the passage of ions through the paracel-lular route (22e25).
It should be added that TJs constitute promiscuous struc-tures because they can be formed between cells of differentepithelia and even animal species, provided these have theability to form TJs among themselves (23,26,27). This abil-ity sounds logical, otherwise places of transition betweenepithelia of different cellular types would offer a dangerousgap in their barrier function. For comparison, synapses andadherent junctions are highly specific.
A given substance can cross the epithelium through twodifferent routes (Figure 1, bottom, center), the transcellular(black arrow) and the paracellular one (red arrow); this oneis controlled by the degree of tightness, electric conduc-tance (24,25) and specific permeability (28) of the TJ.
The name tight junction was coined a century and a halfago (3), long before it was demonstrated that in some epi-thelia, like the proximal tubule of the kidney and the gall-bladder, the bulk of a given substance crosses through theparacellular permeation route (24,29). The larger the para-cellular route, the lower the value of the transepithelialelectrical resistance (TER). TER varies over several ordersof magnitude, depending on the difference in compositionbetween the two compartments separated by the TJ. Thus,the epithelium of the proximal tubule of the kidney, separat-ing plasma from filtrated plasma, has a TER !10 U$cm2
(Figure 3). On the contrary, the epithelium of the urinarybladder withstands a drastic difference in composition, asit separates plasma from urine and, accordingly, its TERmay be as high as 40,000 U$cm2. Although this correlationmakes sense from the teleological point of view, today weignore how epithelia sense the differences between the twosides, and what is the molecular mechanism involved inproviding 10 or 40,000 U$cm2.
Foreseen Pathologies of the TJ
It is obvious that if a molecular defect causes a colon mu-cosa to have a TER of only 50 U$cm2, instead of havingseveral thousand ohms, the organism will die swiftly of in-toxication by products of its own intestinal flora penetratingthrough the leaky paracellular pathway. Conversely, if in-stead of having a TER of 5e10 U$cm2, a molecular imper-fection in the proximal tubule of the kidney causes it tohave several thousands ohms, the ensuing diuresis will kill
468 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
Figure 2. Molecular structure of the TJs. These molecules may belong to the plasma membrane (e.g., claudins, occludins and JAM), participate in the at-
tachment of the TJ to the cytoskeleton (such as ZO-1, ZO-2, ZO-3, ZONAB, cingulin), participate in vesicular traffic (e.g., VAP-33, SEC 6/8, Rab3b, Rab13),
involve in signaling (ASIP, ZAK, sAfadin), reside temporarily in the TJ (e.g., PILT, JACOB), and are important for TJ-formation (e.g., 7H6). These molecules
possess domains that allow them to attach to other protein species, such as PDZ, SH3, GUK homologue, etc.
us in a very short time. We can also anticipate that if a de-fect of ion selectivity of the TJ affects the reabsorption ofa vital ion in the nephron or, on the contrary, permits thereabsorption of an abnormally large amount, the conse-quences may be fatal. Yet, it should not allow the passageof foreign peptides that have a size far smaller, otherwisethe immune system would prepare antibodies against itand, due to a homology between such peptide and a proteinin the pancreas, or in the nervous system, the organism mayend up attacking its own pancreas or its own brain. Actu-ally, as we shall see below, many autoimmune diseasesmay be caused by molecular defects of the TJ. Microorgan-isms and toxins may alter the permeability of the TJ, pro-voking a multitude of troubles. On the other hand, TJsbetween two endothelial cells can relax its hermeticity topermit the passage of a huge leukocyte towards an infectionsite.
Aware that epithelia not only exchange substances withthe environment, but also fence off microorganisms andtoxins, no one is surprised at the number of diseases asso-ciated with failures of these functions. However, as the
architecture, signaling and properties of the moleculesconstituting the TJ unravel, the horizons expand and be-come far more complicated with as yet unsuspected proper-ties of this structure. Thus, being the frontier between theorganism and the environment, epithelial cells are exposedto all kinds of injuries, which are repaired by frequent re-placing and molting. As an example, cells like those ofthe intestinal mucosa undergo fast proliferation at the cryptof the villi, migrate towards the apex, and are finally shed.Obviously, if TJs had rigid structures they would not beable to proliferate, migrate and detach at the apex. In thiscontext, the plasticity of an epithelium, in particular duringembryogenesis, would be impossible. Furthermore, prolif-eration, differentiation and migration involve activationand silencing batteries of specific genes. In this respect,we have to introduce another property of the TJ: that of be-ing a receptor and emitter of signals traveling to and com-ing from the nucleus (Figure 4). Because the TJ is not theonly structure that gauges the neighborhood of the celland transduces the information so that the nucleus wouldknow whether it has to engage in cycling, proliferation or

469Pathologies of the Tight Junction
differentiation, whether to instruct the formation of a syn-apse, a desmosome or a communicating (gap) junction,the molecules that depart from one of these attaching struc-tures and exchange information with the nucleus receive thename of NACos (nuclear and adhesion complexes) (30).There is no need to emphasize the enormous role that theTJ would therefore play in developmental pathologies.
Diseases Involving TJs
After discussing the structural and physiological back-ground of the TJ, it should not be a surprise that its pathol-ogies conform a highly heterogeneous group of diseases,which at present are difficult to classify. Because a formalclassification is premature, we will group them in disordersthat resemble each other, those caused by mutations inproteins of the TJ, those associated with autoimmunereactions, cancers, and microorganisms, describing onlyparadigmatic diseases in each category.
Figure 3. The hermeticity of TJs varies according to physiological neces-
sities. This is illustrated by the urinary apparatus where different segments
have a progressively higher transepithelial electrical resistance (TER), as
the glomerular filtrate is converted into urine and reaches the bladder.
Figures give the values of TER in U$cm2 (124,125).
Diseases Provoked by Mutations in Proteins of the TJ
Familial Renal Hypomagnesemia with Hypercalciuriaand Nephrocalcinosis
Failure in this illness is clearly due to alteration of paracel-lin-1 (PCLN-1) gene, which results in the absence of thecorresponding protein. In this autosomal recessive disease,patients have a profound renal Mg2þ and Ca2þ wasting,causing renal parenchymal calcification and renal failurethat can only be cured through renal transplantation.PCLN-1, also known as claudin-16, is located in TJs ofthe thick ascending limb of Henle and forms an intercellu-lar pore permitting paracellular passage of Mg2þ and Ca2þ
down their electrochemical gradients (31). Patients withmutations that inactivate a PDZ-domain binding motif ofclaudin-16 present a disabling of the scaffold they formwith protein ZO-1. Mutated claudin-16 no longer localizesto TJs in transfected MDCK cells but accumulates insteadin lysosomes (32). The deletion of a large portion of theC-terminal domain, associated with neonatal hypocalcemicseizures, hypocalcemia and even tetany in early infancy,produces a mutant protein that seems to be properly tar-geted to the plasma membrane but is rapidly endocytosedand delivered to lysosomes. Blocking endocytosis restoresthe expression of functional claudin-16 in the membrane.Therefore, inhibitors of endocytosis may provide noveltherapeutic strategies (33). Interestingly, mutations in clau-din-19 also produce hypomagnesemia, yet this symptom isaccompanied by malfunction of several glands, as well asvisual impairment (34).
Congenital Deafness
In the organ of Corti of the inner ear, TJs of the reticularlamina separate Kþ-rich endolymph from Naþ-rich peri-lymph. In humans, mutations of the gene encoding clau-din-14 protein cause profound congenital deafness (35)because the TJ complex at the apex of the reticular laminarequires this cation-restrictive component of the barrier tomaintain the proper ionic composition of the fluid surround-ing the basolateral surface of outer hair cells (36). Claudin-11�/� adult mice, which lack the basal cell TJs that giverise to the intrastrial compartment, exhibit severe deafnessassociated with low endocochlear potentials (37). It was re-cently shown that four different types of recessive muta-tions of the TJ protein tricellulin cause nonsyndromicdeafness (DFNB49) in humans (38).
Neonatal Ichthyosis and Sclerosing Cholangitis Syndrome(NISCH)
This is a severe chronic condition characterized by inflam-mation and obliterative fibrosis of the intra- and extrahe-patic bile ducts, leading to biliary cirrhosis and,ultimately, to liver failure. A 2-bp deletion (200e201 TT)

470 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
Figure 4. Effect of ouabain on NACos (nuclear-adhesion complexes). Cell junctions have specific molecular components that traverse the plasma membrane
and have a relatively long extracellular domain, which is heavily glycosylated (green). They also have a submembrane scaffold constituted by a series of
proteins with a variety of roles. Some of them (pink) anchor the whole junction to the cytoskeleton (red). Another group (NACos, blue) can shuttle from
the junctional scaffold to the nucleus, according to stimuli acting on the extracellular face of the junction. These stimuli can range from contact with a specific
neighboring cell, hormones, drugs, etc. Once in the nucleus NACos modulate the expression of genes involved in proliferation, migration, differentiation, etc.
Insert: two monolayers of MDCK cells under control condition (above) showing the distribution of b-catenin (green) mainly on the lateral borders. Upon
stimulation with low concentrations of hormone ouabain (below), part of b-catenin delocalizes, and switches to the nucleus, which was stained red with
propidium iodine.
in exon 1 of the claudin-1 gene, yielding a premature stopcodon, was identified in four Moroccan patients. Bile ductinjury may be related to the absence of claudin-1 expres-sion in cholangiocytes (39). The familial hypercholanemia(FHC) in Amish individuals is provoked by mutations ofZO-2 in the N-terminal PDZ domain that disrupt its associ-ation with claudin-1 (40). Moreover, NISCH patients alsosuffer from dry ichthyotic skin and have more compactand thicker stratum corneum (39), a phenotype similar tothe one observed in the claudin-1 knockout mice in whichthe epidermal barrier is severely affected, and animals diewithin 1 day of birth, with severe dehydration due to exces-sive water loss across the skin (41).
Velo-Cardio-Facial Syndrome (VCF)
Claudin-5, also known as transmembrane protein mutatedin VCF, is localized in chromosome 22q11, which is fre-quently deleted in Velo-cardio-facial (VCF)/DiGeorge syn-drome patients (42,43). Claudin-5 is a major component ofTJs of endothelial cells. In the blood-brain barrier (BBB), itrestricts the penetration of molecules with a molecularweight !800 D (44).
TJ Modifications in Immunity-related Diseases
As mentioned above, epithelia hamper the passage into thebody of microorganisms, toxic substances and antigens(3,6). When these barriers are damaged or when TJs are ab-
normally permeable, these antigens can penetrate the bodyand may cause an immune response (45e50). Several anti-gen determinants are similar to host antigens and it is be-lieved that, in vulnerable individuals, these might inducethe generation of antibodies that cross-react with host anti-gens (51).
Inflammatory Bowel Disease
Toll-like receptors (TLR) are involved in both recognitionof normal gut flora (especially probiotics) and increasingbarrier function (52,54). Mice deficient in MyD88, anadapter molecule essential for TLR signaling, are highlysusceptible to experimental inflammatory bowel disease(55). Several cytokines and growth factors known to regu-late the barrier function of the TJ (56) may exacerbatechronic gut inflammation. For example, in healthy individ-uals CD4þ T cells activated by gut antigens in Peyer’spatches migrate to the lamina propia (LP), where they diethrough apoptosis. In a pathological conditions instead, ab-normal epithelial permeability of the TJ increase epithelialpermeability, thus permitting the penetration of antigens toLP, and trigger T-cell activation. Pro-inflammatory cyto-kines, like interferon-g (57) and tumor necrosis factor-a (58), then further increase epithelial permeability, settingup a vicious cycle of chronic inflammation (53).
In Crohn’s disease, as well as in ulcerative colitis, thebarrier function of the TJ is severely affected (59e61). Pa-tients suffer not only from intestinal manifestations, but

471Pathologies of the Tight Junction
present also autoimmune disorders like ankylosing spondy-litis, hemolytic anemia, primary biliary cirrhosis, type I di-abetes, sclerosing cholangitis, arthritis, thyroid disease, etc.Freeze fracture analysis of the TJ reveals both a reducednumber and discontinuous TJ strands in these patients. Nor-mal strands are sketchy and are illustrated in Figure 1. Themolecular analysis of these alterations show that occludinand claudins -5 and -8 are downregulated and redistributedoff the TJ, whereas the pore-forming TJ protein claudin-2 isstrongly upregulated (61).
Type I Diabetes
Although this appears to be in fact a multifactorial disease,one of its most prominent causes is an abnormal gut perme-ability (62). In BioBreeding diabetic-prone rats (an animalmodel of type I diabetes), the onset of diabetes coincideswith an upregulation of zonulin, an endogenous modulatorof intestinal paracellular pathway (63), leading to an abnor-mal permeability of the gut and production of autoanti-bodies against pancreatic b cells. Treatment with anantagonist of the zonulin receptor recovers the gut barrierand reduces diabetes by 70% (62).
Zonulin also seems to participate in the pathogenesis ofceliac disease, an autoimmune enteropathy triggered in sus-ceptible individuals by ingestion of gliadin (ethanol-solublefraction of gluten) (64). It is accompanied by an enhancedintestinal permeability (64,65) and a decrease of TJ strands(66). In vitro studies show that treatment of Caco-2 cellswith gliadin (a component of gluten to which Crohn’s pa-tients are particularly sensitive) downregulates the expres-sion of occludin, claudins-3 and -4, E-cadherin and ZO-1(67).
Multiple Sclerosis
This is a chronic, inflammatory demyelinizing disorder ofthe central nervous system (CNS), characterized by BBBdysfunction that precedes the onset of clinical signs (68).Candidate molecules for autoantigenic involvement in mul-tiple sclerosis include myelin basic proteins, proteolipidprotein, myelin oligodendrocyte glycoprotein and claudin-11 [also known as oligodendrocyte-specific protein(OSP)]. This claudin is robustly expressed in brain and tes-tis, localizing to TJs in myelin sheaths and Sertoli cells(69). It is also differentially expressed at low levels alongthe nephron (70,71). Antibodies that cross-react withclaudin-11 have been detected in the cerebrospinal fluid(CSF) of relapsing-remitting multiple sclerosis patients(72). Moreover, these antibodies not only recognize asequence in claudin-11 that is 71% homologous to proteinsin common pathogens (like variola, EpsteineBarr virus,herpes simplex II, HIV, Coxsackie virus B, E. coli, Pseudo-monas, Klebsiella, Mycobacterium tuberculosis, and Clos-tridium), but can also recognize (bind to) the pathogenproteins (72), which suggests that susceptible individuals
develop antibodies to epitopes that should not be antigenic.Furthermore, in SJL mouse strain (an animal model of au-toimmune encephalomyelitis), injection of short peptidesrepresenting the amino acid sequence of murine claudin-11 trigger autoimmune encephalomyelitis. However, inother mouse strains that are also used as models of autoim-mune encephalomyelitis, the same peptides are ineffective(73). This suggests that genetic background is determinantin the susceptibility to autoimmune diseases development,as is seen in human autoimmune diseases, because themajority have a clear kindred-illness relationship.
Ankylosing Spondylitis
This is a common and highly familiar rheumatic disorderresembling other autoimmune diseases discussed above.In ankylosing spondylitis, there is a link between diseasedevelopment and increased intestinal permeability. Interest-ingly, the anomalous intestinal permeability is a characteris-tic shared among patients and their asymptomatic relatives(74e76).
Allergies
Exogenous perturbants might disrupt airways, increasing ep-ithelial permeability. If allergens promote such disruption, itwould explain why they encounter antigen-presenting cells.The cysteine protease allergen Der p 1 from fecal pellets ofthe house dust mite Dermatophagoides pteronyssinus cleaveoccludin and claudin-1 at its extracellular domains, inducethe reorganization of ZO-1 and cause disruption of TJs(63,77,78). A similar phenomenon is observed with alkalineserine protease Pen ch 13, an allergen from the airborne fun-gus Penicillium chrysegenum that induces cleavage of occlu-din at aminoacids 202 and 211, damaging the airwayepithelium barrier (79). TJ disruption might contribute tosensitization and allergic responses by degrading the func-tion of the epithelial barrier.
TJ Proteins and Cancer
Whereas altered expression of TJ-specific proteins in a mul-titude of cancers is sufficiently documented, their specificrole is still complex and not without controversy. For in-stance, whereas claudin-7 has been found to be decreasedin invasive ductal carcinomas of the breast (80), in ovarian,colon, cervical and gastric cancers it is overexpressed(Table 1). On the other hand, claudins -3, -4 and -7 (81)are frequently elevated in various cancers including breast,ovarian, colon, gastric and prostate. Expression of claudin-3and claudin-4 in human ovarian surface epithelial (HOSE)cells increased cell survival, matrix metalloproteinase-2activity, cell invasion and motility. Conversely, knockdownof claudin-3 and -4 expression in ovarian cancer cell linesreduces invasion (82,83). Breast tumor cells re-expressingclaudin-1 in vitro display elevation in apoptosis, in parallel,
472 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
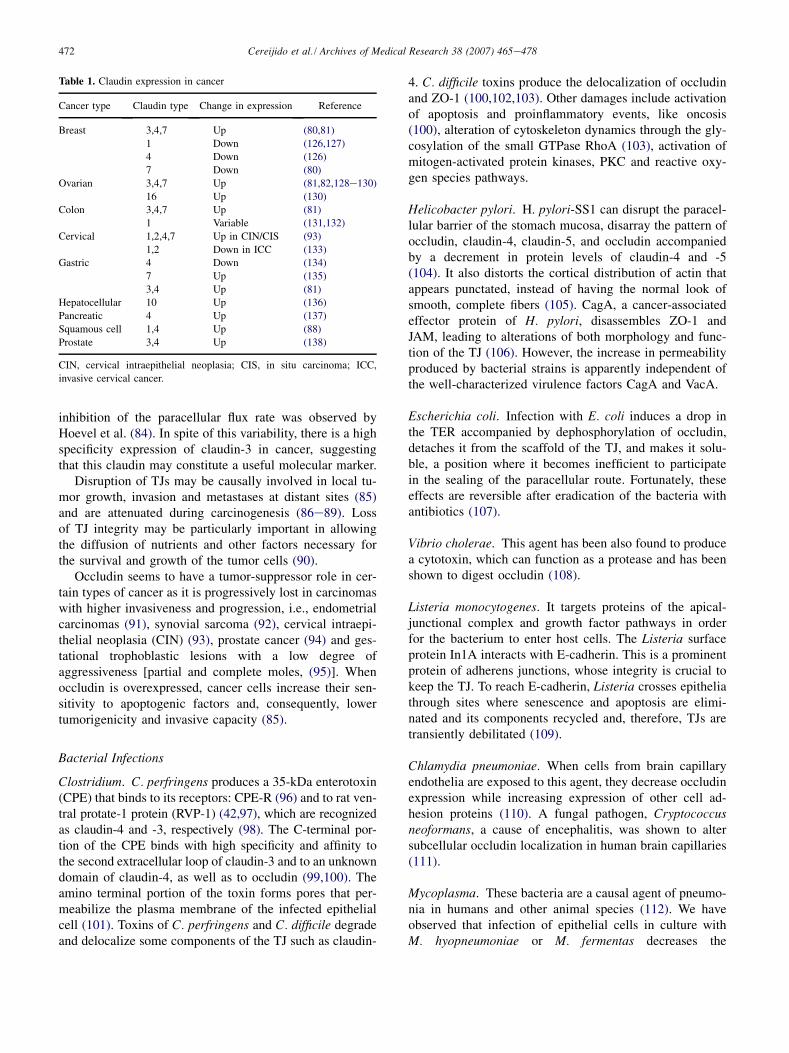
inhibition of the paracellular flux rate was observed byHoevel et al. (84). In spite of this variability, there is a highspecificity expression of claudin-3 in cancer, suggestingthat this claudin may constitute a useful molecular marker.
Disruption of TJs may be causally involved in local tu-mor growth, invasion and metastases at distant sites (85)and are attenuated during carcinogenesis (86e89). Lossof TJ integrity may be particularly important in allowingthe diffusion of nutrients and other factors necessary forthe survival and growth of the tumor cells (90).
Occludin seems to have a tumor-suppressor role in cer-tain types of cancer as it is progressively lost in carcinomaswith higher invasiveness and progression, i.e., endometrialcarcinomas (91), synovial sarcoma (92), cervical intraepi-thelial neoplasia (CIN) (93), prostate cancer (94) and ges-tational trophoblastic lesions with a low degree ofaggressiveness [partial and complete moles, (95)]. Whenoccludin is overexpressed, cancer cells increase their sen-sitivity to apoptogenic factors and, consequently, lowertumorigenicity and invasive capacity (85).
Bacterial Infections
Clostridium. C. perfringens produces a 35-kDa enterotoxin(CPE) that binds to its receptors: CPE-R (96) and to rat ven-tral protate-1 protein (RVP-1) (42,97), which are recognizedas claudin-4 and -3, respectively (98). The C-terminal por-tion of the CPE binds with high specificity and affinity tothe second extracellular loop of claudin-3 and to an unknowndomain of claudin-4, as well as to occludin (99,100). Theamino terminal portion of the toxin forms pores that per-meabilize the plasma membrane of the infected epithelialcell (101). Toxins of C. perfringens and C. difficile degradeand delocalize some components of the TJ such as claudin-
Table 1. Claudin expression in cancer
Cancer type Claudin type Change in expression Reference
Breast 3,4,7 Up (80,81)
1 Down (126,127)
4 Down (126)
7 Down (80)
Ovarian 3,4,7 Up (81,82,128e130)
16 Up (130)
Colon 3,4,7 Up (81)
1 Variable (131,132)
Cervical 1,2,4,7 Up in CIN/CIS (93)
1,2 Down in ICC (133)
Gastric 4 Down (134)
7 Up (135)
3,4 Up (81)
Hepatocellular 10 Up (136)
Pancreatic 4 Up (137)
Squamous cell 1,4 Up (88)
Prostate 3,4 Up (138)
CIN, cervical intraepithelial neoplasia; CIS, in situ carcinoma; ICC,
invasive cervical cancer.
4. C. difficile toxins produce the delocalization of occludinand ZO-1 (100,102,103). Other damages include activationof apoptosis and proinflammatory events, like oncosis(100), alteration of cytoskeleton dynamics through the gly-cosylation of the small GTPase RhoA (103), activation ofmitogen-activated protein kinases, PKC and reactive oxy-gen species pathways.
Helicobacter pylori. H. pylori-SS1 can disrupt the paracel-lular barrier of the stomach mucosa, disarray the pattern ofoccludin, claudin-4, claudin-5, and occludin accompaniedby a decrement in protein levels of claudin-4 and -5(104). It also distorts the cortical distribution of actin thatappears punctated, instead of having the normal look ofsmooth, complete fibers (105). CagA, a cancer-associatedeffector protein of H. pylori, disassembles ZO-1 andJAM, leading to alterations of both morphology and func-tion of the TJ (106). However, the increase in permeabilityproduced by bacterial strains is apparently independent ofthe well-characterized virulence factors CagA and VacA.
Escherichia coli. Infection with E. coli induces a drop inthe TER accompanied by dephosphorylation of occludin,detaches it from the scaffold of the TJ, and makes it solu-ble, a position where it becomes inefficient to participatein the sealing of the paracellular route. Fortunately, theseeffects are reversible after eradication of the bacteria withantibiotics (107).
Vibrio cholerae. This agent has been also found to producea cytotoxin, which can function as a protease and has beenshown to digest occludin (108).
Listeria monocytogenes. It targets proteins of the apical-junctional complex and growth factor pathways in orderfor the bacterium to enter host cells. The Listeria surfaceprotein In1A interacts with E-cadherin. This is a prominentprotein of adherens junctions, whose integrity is crucial tokeep the TJ. To reach E-cadherin, Listeria crosses epitheliathrough sites where senescence and apoptosis are elimi-nated and its components recycled and, therefore, TJs aretransiently debilitated (109).
Chlamydia pneumoniae. When cells from brain capillaryendothelia are exposed to this agent, they decrease occludinexpression while increasing expression of other cell ad-hesion proteins (110). A fungal pathogen, Cryptococcusneoformans, a cause of encephalitis, was shown to altersubcellular occludin localization in human brain capillaries(111).
Mycoplasma. These bacteria are a causal agent of pneumo-nia in humans and other animal species (112). We haveobserved that infection of epithelial cells in culture withM. hyopneumoniae or M. fermentas decreases the

473Pathologies of the Tight Junction
transepithelial electrical resistance and delocalizes and de-grades TJ proteins. Under these conditions, M. hyopneumo-niae binds preferentially to the apical membrane in sites ofcell�cell adhesion, suggesting that TJs may contain a recep-tor for this bacteria (Ruiz A., unpublished data).
Viral Infections
Rotavirus causes TJ leakiness and increased transepithelialpermeability in the absence of any cell death, while provok-ing the redistribution of occludin and claudin-1 in Caco-2cells (113). Disruption of occludin staining patterns ofbloodebrain barrier endothelia is a hallmark of HIV-associated encephalitis (114). Viruses such as reovirus anda-herpesviruses target JAM (Figure 2) and nectin receptors,respectively, and use them for cell entry (115,116). Glyco-protein D, a surface protein of a-herpesviruses, interactswith adherens junctions, specifically with nectin receptorsduring cell entry, disrupts cellecell adhesion in cells ex-pressing nectin-1 and lacking E-cadherin, and interacts withnectin receptors in polarized epithelia (115,117). Rotavirusnormally reaches epithelia through the apical domain.However, given that its receptor is integrin, a componentof the focal contacts located beyond the TJ, the virus pro-vokes a transient opening of this structure, which alsocauses a lateral diffusion of integrins to the apical mem-brane (118). Human immunodeficiency virus (HIV) mustcross the epithelium during transmission because the pri-mary targets for HIV infection, CD4þ cells, are protectedby epithelial lining. It has been proposed that one plausiblemechanism is that HIV utilizes lesions in the mucosal sur-face to invade underlying lymphoid cells. However, a recentstudy (119) showed that claudin-7 increase the susceptibil-ity of CD4� cells to be infected in a gp120-independentmechanism, opening the possibility that HIV can invadeepithelia through direct interaction with TJ proteins.
Ameba
Entamoeba histolytica trophozoites initiate amebiasisthrough invasion into the enteric mucosa, on direct contactwith the apical side of the enteric cells, and caused anincrease in paracellular permeability as evidenced by adecrease of TER associated with an increase in [3H]manni-tol flux. Immunoprecipitation of cocultures revealed de-phosphorylation of ZO-2, loss of ZO-1 from ZO-2, anddegradation of ZO-1 but less so of ZO-2 and none of occlu-din or E-cadherin (120). E. histolytica molecules responsi-ble for these effects are cysteine proteinases, and thepretreatment of trophozoites prevented proteolysis of TJmolecules ZO-1 and ZO-2 and of villin, the major actin-bundling molecule in microvilli (117).
Towards a Pharmacology of the TJ
Today we have no specific agents to treat diseases of the TJ.Faced with an immunological disease produced by passageof a given allergenic substance through the paracellularroute, all one can do is to prevent exposure to such sub-stance. Because TJs have only been known for a centuryand a half, there are no traditional medicines to treat them,and these are likely to come from an intimate understandingof their structure and function. However, some specialiststake advantage of natural ‘‘experiments’’ performed bynature.
Looking for a TER-decreasing Factor
Although rotaviruses bind to intestinal receptors and causediarrhea, they bind to receptors that are not exposed to theintestinal lumen, but hidden beyond the TJ in the basolater-al domain of enterocytes. This situation, by itself, stronglysuggests that in order to gain access to receptors, rotavi-ruses must have a way of opening the TJ. On this basis,Nava et al. (118,121) explored whether the outer capsidprotein VP4 of rotaviruses is capable of opening the para-cellular space sealed by the TJ. Actually, these investigatorscleaved VP4 with trypsin into VP5 and VP8 and found thatonly the last one is capable of diminishing TER, in a dose-dependent and reversible manner. Furthermore, VP8 trig-gers the movement of some basolateral proteins towardsthe apical surface, suggesting that once the fence consti-tuted by the TJ is relaxed, basolateral proteins can diffusein the plane of the membrane.
Now it would be pertinent to ask what would be the ad-vantage of TJ-opening molecules such as VP8. It may beeasy to answer through a concrete example. Treating diabe-tes requires daily injections of insulin because enteric ad-ministration results in thorough digestion and nullificationof this hormone. This is actually what happens with dia-betic rats. Yet, when these rats are instead administered oralinsulin, but in combination with VP8, TJs in the upper partof the digestive tract open, the hormone penetrates into thebody without passing through the stomach, and is able todecrease glucose levels (118). In other words, a substancewith the ability to open the TJ will drastically change thetherapeutic approach to diabetes, as well as the daily lifeof the patient.
A novel TJ-modulating (TJM) peptide was described af-fording a transient, reversible lowering of TER in an in vi-tro model of nasal epithelial tissue. The TJM peptide wastested in the in vitro tissue model for potential to enhancepermeation acetylcholinesterase inhibitor galantamine, aswell as three peptides, salmon calcitonin, parathyroid hor-mone 1e34, and peptide regulator of appetite YY (3,36).In all cases, the TJM peptide afforded a dramatic improve-ment in drug permeation across epithelial tissue. In

474 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
addition, a formulation containing PYY (3e36) and TJMpeptide was dosed intranasally in rabbits, resulting in a dra-matic increase in bioavailability. TJM peptide representsa promising advancement in intranasal formulation devel-opment (112,122,123).
Looking for a TER-increasing Factor
As we have seen above, diseases like multiple sclerosis anddiabetes are likely to be due to a defective sealing of the TJthat allows the passage of foreign peptides and that wouldchallenge the immune system to produce antibodies againstit. These antibodies may be addressed to epitopes also pres-ent in proteins of the brain, thyroid gland, pancreas, etc.The overall consequence is that a permissive TJ ends upcausing an autoimmune disease. In keeping with this inter-pretation, these diseases are not steady, but show periods ofimprovement in which symptoms virtually vanish, alternat-ing with months of aggravation. Unfortunately, remissionsnever reach a total cure, as oscillations progressively dam-age diverse organs. Periods of aggravation are frequentlyassociated with increasing contact with the specific antigen.Therefore a TER-increasing molecular tool would presum-ably prevent the passage of such antigen through the TJ andhelp fight the disease.
In order to search for such a tool, we looked for a normalsituation in which an epithelium progressively increases itsTER. In this respect, the nephron is an obvious candidate(Figure 3). Adult human filters approximately 170 L ofplasma/day, a volume that is constantly reduced as the fil-trate progresses from the glomerulus to the urinary bladder,until only 1e1.5 L of urine is produced. This 150-fold con-centration that converts filtrated plasma into urine ismatched by the value of TER that increases from 5 to 10U/cm2 in the proximal tube, to 5e40,000 U/cm2 in thebladder. We reasoned that a) if the liquid inside the pro-ximal tube contains a TER-increasing factor that is not re-absorbed; b) if this TER-increasing factor acts ina concentration-dependent manner; and (c) it reaches theurinary bladder without losing its activity, we may be ableto collect it and purify it. We did exactly that (124) and dis-covered that dog urine has a TER-increasing factor that actson monolayers of cultured cells, regardless of whether thesecells belong to dog, human, rabbit, cat or rat. Conversely,this factor may be extracted from several animal speciesincluding humans and increase the TER of monolayersof MDCK cells (epithelial, from the dog kidney). We haverecently shown that the factor is epidermal growth factor(EGF) (125), and it changes the value of TER by decreasingthe expression of claudin-2, one of the prominent compo-nents of the TJ (Figure 2).
In summary, we now have several TER-decreasing, andat least one TER-increasing factor.
Conclusions
A few decades ago the TJ was assumed to be an obscureseal at the outermost end of the space between epithelialcells. Today, due to studies with transmission and freeze-fracture electron microscopy, as well as the use of epithelialcell lines cultured as monolayer that resemble natural epi-thelia, the TJ is known to be constituted by more than 50different protein species whose specific role remains to beelucidated. Failure in these molecules results in a multitudeof diseases ranging from those that affect the nervous sys-tem to those that cause diabetes, asthma and allergy. Someof these have been known for centuries, except that theywere not suspected to be caused by defects in the TJ. There-fore, this structure gives rise to a whole new branch of med-icine. Efforts are being made to find substances that wouldincrease or reduce the degree of sealing. The Center for Re-search and Advanced Studies (CINVESTAV) has a solidnumber of specialists in the biology of the TJ, ranging frommolecular to medical and, even pharmacological, aspects.
AcknowledgmentsWe thank Yadira Suarez for her technical and artistic drawings,and the kind and efficient help of Elizabeth del Oso and JazminDe Lorenz. We gratefully acknowledge the economic support ofCONACYT (National Research Council of Mexico).
References1. Cereijido M, Rotunno CA. Introduction to the Study of Biological
Membranes. New York, NY: Gordon and Breach; 1971.
2. Cereijido M, Contreras RG, Shoshani L. Cell adhesion, polarity,
and epithelia in the dawn of metazoans. Physiol Rev 2004;84:
1229e1262.
3. Cereijido M, Anderson J. Tight Junctions. 2nd ed Boca Raton FL:
CRC Press; 2001. p. 772.
4. Cereijido M, Shoshani L, Contreras RG, Larre I. Na,K-ATPase, the
star of epithelial net transports: what do we know about its own
polarized distribution. Physiol Mini-Rev 2005;1:1e10.
5. Cereijido M, Shoshani L, Contreras RG. The polarized distribution of
Naþ, Kþ-ATPase and active transport across epithelia. J Membr Biol
2001;184:299e304.
6. Cereijido M, Shoshani L, Contreras RG. Molecular physiology and
pathophysiology of tight junctions. I. Biogenesis of tight junctions
and epithelial polarity. Am J Physiol Gastrointest Liver Physiol
2000;279:G477eG482.
7. Cereijido M, Contreras RG, Shoshani L, Garcia-Villegas MR.
Membrane targeting. Prog Biophys Mol Biol 2003;81:81e115.
8. Cereijido M, Rotunno CA, Robbins ES, Sabatini DD. Polarized epi-
thelial membranes produced in vitro. In: Hofmann JF, ed. Membrane
Transports Processes. New York: Raven Press; 1978. pp. 434e443.
9. Cereijido M, Robbins ES, Dolan WJ, Rotunno CA, Sabatini DD.
Polarized monolayers formed by epithelial cells on a permeable
and translucent support. J Cell Biol 1978;77:853e880.
10. Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita S,
et al. Occludin: a novel integral membrane protein localizing at tight
junctions. J Cell Biol 1993;123:1777e1788.
11. Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and
-2: novel integral membrane proteins localizing at tight junctions

475Pathologies of the Tight Junction
with no sequence similarity to occludin. J Cell Biol 1998;141:
1539e1550.
12. Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. The tight junc-
tion protein ZO-1 establishes a link between the transmembrane pro-
tein occludin and the actin cytoskeleton. J Biol Chem 1998;273:
29745e29753.
13. Gonzalez-Mariscal L, Islas S, Contreras RG, Garcia-Villegas MR,
Betanzos A, Vega J, et al. Molecular characterization of the tight
junction protein ZO-1 in MDCK cells. Exp Cell Res 1999;248:
97e109.
14. Balda MS, Garrett MD, Matter K. The ZO-1-associated Y-box factor
ZONAB regulates epithelial cell proliferation and cell density. J Cell
Biol 2003;160:423e432.
15. Izumi Y, Hirose T, Tamai Y, Hirai S, Nagashima Y, Fujimoto T, et al.
An atypical PKC directly associates and colocalizes at the epithelial
tight junction with ASIP, a mammalian homologue of Caenorhabditis
elegans polarity protein PAR-3. J Cell Biol 1998;143:95e106.
16. Betanzos A, Huerta M, Lopez-Bayghen E, Azuara E, Amerena J,
Gonzalez-Mariscal L. The tight junction protein ZO-2 associates
with Jun, Fos and C/EBP transcription factors in epithelial cells.
Exp Cell Res 2004;292:51e66.
17. Yamamoto T, Harada N, Kano K, Taya S, Canaani E, Matsuura Y,
et al. The Ras target AF-6 interacts with ZO-1 and serves as a periph-
eral component of tight junctions in epithelial cells. J Cell Biol 1997;
139:785e795.
18. Aijaz S, D’Atri F, Citi S, Balda MS, Matter K. Binding of GEF-H1 to
the tight junction-associated adaptor cingulin results in inhibition of
Rho signaling and G1/S phase transition. Dev Cell 2005;8:777e786.
19. Balda MS, Anderson JM, Matter K. The SH3 domain of the tight
junction protein ZO-1 binds to a serine protein kinase that phosphor-
ylates a region C-terminal to this domain. FEBS Lett 1996;399:
326e332.
20. Yeaman C, Grindstaff KK, Nelson WJ. Mechanism of recruiting
Sec6/8 (exocyst) complex to the apical junctional complex during
polarization of epithelial cells. J Cell Sci 2004;117:559e570.
21. Zhong Y, Saitoh T, Minase T, Sawada N, Enomoto K, Mori M.
Monoclonal antibody 7H6 reacts with a novel tight junction-
associated protein distinct from ZO-1, cingulin and ZO-2. J Cell Biol
1993;120:477e483.
22. Contreras RG, Avila G, Gutierrez C, Bolivar JJ, Gonzalez-
Mariscal L, Darzon A, et al. Repolarization of Naþ-Kþ pumps dur-
ing establishment of epithelial monolayers. Am J Physiol 1989;257:
C896eC905.
23. Gonzalez-Mariscal L, Chavez de Ramirez B, Lazaro A, Cereijido M.
Establishment of tight junctions between cells from different animal
species and different sealing capacities. J Membr Biol 1989;107:
43e56.
24. Stefani E, Cereijido M. Electrical properties of cultured epithelioid
cells (MDCK). J Membr Biol 1983;73:177e184.
25. Cereijido M. Electrical properties of Madin-Darby canine kidney
cells. Fed Proc 1984;43:2230e2235.
26. Contreras RG, Lazaro A, Bolivar JJ, Flores-Maldonado C,
Sanchez SH, Gonzalez-Mariscal L, et al. A novel type of cell-cell co-
operation between epithelial cells. J Membr Biol 1995;145:305e310.
27. Contreras RG, Shoshani L, Flores-Maldonado C, Lazaro A,
Monroy AO, Roldan ML, et al. E-cadherin and tight junctions
between epithelial cells of different animal species. Pflugers Arch
2002;444:467e475.
28. Gonzalez-Mariscal L, Contreras RG, Bolivar JJ, Ponce A, Chavez De
Ramirez B, Cereijido M. Role of calcium in tight junction formation
between epithelial cells. Am J Physiol 1990;259:C978eC986.
29. Fromter E, Diamond J. Route of passive ion permeation in epithelia.
Nat New Biol 1972;235:9e13.
30. Balda MS, Matter K. Epithelial cell adhesion and the regulation
of gene expression. Trends Cell Biol 2003;13:310e318.
31. Simon DB, Lu Y, Choate KA, Velazquez H, Al Sabban E, Praga M,
et al. Paracellin-1, a renal tight junction protein required for paracel-
lular Mg2þ resorption. Science 1999;285:103e106.
32. Muller D, Kausalya PJ, Claverie-Martin F, Meij IC, Eggert P, Garcia-
Nieto V, et al. A novel claudin 16 mutation associated with childhood
hypercalciuria abolishes binding to ZO-1 and results in lysosomal
mistargeting. Am J Hum Genet 2003;73:1293e1301.
33. Muller D, Kausalya PJ, Meij IC, Hunziker W. Familial hypomagnese-
mia with hypercalciuria and nephrocalcinosis: blocking endocytosis
restores surface expression of a novel Claudin-16 mutant that lacks
the entire C-terminal cytosolic tail. Hum Mol Genet 2006;15:
1049e1058.
34. Konrad M, Schaller A, Seelow D, Pandey AV, Waldegger S,
Lesslauer A, et al. Mutations in the tight-junction gene claudin 19
(CLDN19) are associated with renal magnesium wasting, renal fail-
ure, and severe ocular involvement. Am J Hum Genet 2006;79:
949e957.
35. Wilcox ER, Burton QL, Naz S, Riazuddin S, Smith TN, Ploplis B,
et al. Mutations in the gene encoding tight junction claudin-14 cause
autosomal recessive deafness DFNB29. Cell 2001;104:165e172.
36. Ben Yosef T, Belyantseva IA, Saunders TL, Hughes ED,
Kawamoto K, Van Itallie CM, et al. Claudin 14 knockout mice,
a model for autosomal recessive deafness DFNB29, are deaf due to
cochlear hair cell degeneration. Hum Mol Genet 2003;12:
2049e2061.
37. Gow A, Davies C, Southwood CM, Frolenkov G, Chrustowski M,
Ng L, et al. Deafness in Claudin 11-null mice reveals the critical con-
tribution of basal cell tight junctions to stria vascularis function.
J Neurosci 2004;24:7051e7062.
38. Riazuddin S, Ahmed ZM, Fanning AS, Lagziel A, Kitajiri S,
Ramzan K, et al. Tricellulin is a tight-junction protein necessary
for hearing. Am J Hum Genet 2006;79:1040e1051.
39. Hadj-Rabia S, Baala L, Vabres P, Hamel-Teillac D, Jacquemin E,
Fabre M, et al. Claudin-1 gene mutations in neonatal sclerosing chol-
angitis associated with ichthyosis: a tight junction disease. Gastroen-
terology 2004;127:1386e1390.
40. Carlton VE, Harris BZ, Puffenberger EG, Batta AK, Knisely AS,
Robinson DL, et al. Complex inheritance of familial hypercholane-
mia with associated mutations in TJP2 and BAAT. Nat Genet
2003;34:91e96.
41. Furuse M, Hata M, Furuse K, Yoshida Y, Haratake A, Sugitani Y,
et al. Claudin-based tight junctions are crucial for the mammalian
epidermal barrier: a lesson from claudin-1-deficient mice. J Cell Biol
2002;156:1099e1111.
42. Morita K, Furuse M, Fujimoto K, Tsukita S. Claudin multigene fam-
ily encoding four-transmembrane domain protein components of
tight junction strands. Proc Natl Acad Sci USA 1999;96:511e516.
43. Sirotkin H, Morrow B, Saint-Jore B, Puech A, Das GR, Patanjali SR,
et al. Identification, characterization, and precise mapping of a human
gene encoding a novel membrane-spanning protein from the 22q11
region deleted in velo-cardio-facial syndrome. Genomics 1997;42:
245e251.
44. Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, et al. Size-
selective loosening of the blood-brain barrier in claudin-5-deficient
mice. J Cell Biol 2003;161:653e660.
45. Caserta L, de Magistris L, Secondulfo M, Caravelli G, Riegler G,
Cuomo G, et al. Assessment of intestinal permeability and orocecal
transit time in patients with systemic sclerosis: analysis of relation-
ships with epidemiologic and clinical parameters. Rheumatol Int
2003;23:226e230.
46. Di LV, Venturi C, Baragiotta A, Martines D, Floreani A. Gastroduo-
denal and intestinal permeability in primary biliary cirrhosis. Eur J
Gastroenterol Hepatol 2003;15:967e973.
47. Fasano A. Pathological and therapeutical implications of macro-
molecule passage through the tight junction. In: Tight Junctions.

476 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
Cereijido M, Anderson JM, eds. Boca Raton, FL: CRC Press; 2001.
pp. 697e722.
48. Hamilton I, Cobden I, Rothwell J, Axon AT. Intestinal permeability
in coeliac disease: the response to gluten withdrawal and single-dose
gluten challenge. Gut 1982;23:202e210.
49. Kovacs T, Kun L, Schmelczer M, Wagner L, Davin JC, Nagy J. Do
intestinal hyperpermeability and the related food antigens play a role
in the progression of IgA nephropathy? I. Study of intestinal perme-
ability. Am J Nephrol 1996;16:500e505.
50. Meddings JB, Jarand J, Urbanski SJ, Hardin J, Gall DG. Increased
gastrointestinal permeability is an early lesion in the spontaneously
diabetic BB rat. Am J Physiol 1999;276:G951eG957.
51. Christen U, von Herrath MG. Induction, acceleration or prevention of
autoimmunity by molecular mimicry. Mol Immunol 2004;40:
1113e1120.
52. Cario E, Gerken G, Podolsky DK. Toll-like receptor 2 enhances ZO-
1-associated intestinal epithelial barrier integrity via protein kinase
C. Gastroenterology 2004;127:224e238.
53. Macdonald TT, Monteleone G. Immunity, inflammation, and allergy
in the gut. Science 2005;307:1920e1925.
54. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C,
et al. Probiotic bacteria enhance murine and human intestinal epithe-
lial barrier function. Gastroenterology 2001;121:580e591.
55. Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S,
Medzhitov R. Recognition of commensal microflora by toll-like
receptors is required for intestinal homeostasis. Cell 2004;118:
229e241.
56. Shoshani L, Flores-Benıtez D, Gonzalez-Mariscal L, Contreras RG,
Gonzalez-Mariscal L. Regulation of tight junctions’ functional integ-
rity: role of a urinary factor, lipids and ouabain. In: Tight Junctions.
Gonzalez-Mariscal L, ed. New York, NY: Landes Bioscience and
Springer Science þ Business Media, LLC; 2006. pp. 146e163.
57. Wang F, Graham WV, Wang Y, Witkowski ED, Schwarz BT,
Turner JR. Interferon-gamma and tumor necrosis factor-alpha syner-
gize to induce intestinal epithelial barrier dysfunction by up-regulat-
ing myosin light chain kinase expression. Am J Pathol 2005;166:
409e419.
58. Ma TY, Boivin MA, Ye D, Pedram A, Said HM. Mechanism of TNF-
a modulation of Caco-2 intestinal epithelial tight junction barrier:
role of myosin light-chain kinase protein expression. Am J Physiol
Gastrointest Liver Physiol 2005;288:G422eG430.
59. Hollander D, Vadheim CM, Brettholz E, Petersen GM, Delahunty T,
Rotter JI. Increased intestinal permeability in patients with Crohn’s
disease and their relatives. A possible etiologic factor. Ann Intern
Med 1986;105:883e885.
60. Schmitz H, Barmeyer C, Fromm M, Runkel N, Foss HD, Bentzel CJ,
Riecken, et al. Altered tight junction structure contributes to the im-
paired epithelial barrier function in ulcerative colitis. Gastroenterol-
ogy 1999;116:301e309.
61. Zeissig S, Burgel N, Gunzel D, Richter J, Mankertz J,
Wahnschaffe U, et al. Changes in expression and distribution of
claudin-2, -5 and -8 lead to discontinuous tight junctions and barrier
dysfunction in active Crohn’s disease. Gut 2006;56:6e8.
62. Watts T, Berti I, Sapone A, Gerarduzzi T, Not T, Zielke R, Fasano A.
Role of the intestinal tight junction modulator zonulin in the patho-
genesis of type I diabetes in BB diabetic-prone rats. Proc Natl Acad
Sci USA 2005;102:2916e2921.
63. Wang W, Uzzau S, Goldblum SE, Fasano A. Human zonulin, a potential
modulator of intestinal tight junctions. J Cell Sci 2000; 113 Pt 24:
4435e4440.
64. Bjarnason I, Peters TJ. In vitro determination of small intestinal
permeability: demonstration of a persistent defect in patients with
coeliac disease. Gut 1984;25:145e150.
65. Bjarnason I, Peters TJ, Veall N. Intestinal permeability defect in
coeliac disease. Lancet 1983;1:1284e1285.
66. Schulzke JD, Bentzel CJ, Schulzke I, Riecken EO, Fromm M. Epi-
thelial tight junction structure in the jejunum of children with acute
and treated celiac sprue. Pediatr Res 1998;43:435e441.
67. Sander GR, Cummins AG, Henshall T, Powell BC. Rapid disruption
of intestinal barrier function by gliadin involves altered expression of
apical junctional proteins. FEBS Lett 2005;579:4851e4855.
68. Kermode AG, Thompson AJ, Tofts P, MacManus DG, Kendall BE,
Kingsley DP, et al. Breakdown of the blood-brain barrier precedes
symptoms and other MRI signs of new lesions in multiple sclerosis.
Pathogenetic and clinical implications. Brain 1990;113(Pt 5):
1477e1489.
69. Gow A, Southwood CM, Li JS, Pariali M, Riordan GP, Brodie SE,
et al. CNS myelin and Sertoli cell tight junction strands are absent
in Osp/claudin-11 null mice. Cell 1999;99:649e659.
70. Kiuchi-Saishin Y, Gotoh S, Furuse M, Takasuga A, Tano Y,
Tsukita S. Differential expression patterns of claudins, tight junction
membrane proteins, in mouse nephron segments. J Am Soc Nephrol
2002;13:875e886.
71. Shoshani L, Contreras RG, Roldan ML, Moreno J, Lazaro A,
Balda MS, et al. The polarized expression of Naþ,Kþ-ATPase in
epithelia depends on the association between b-subunits located
in neighboring cells. Mol Biol Cell 2005;16:1071e1081.
72. Bronstein JM, Lallone RL, Seitz RS, Ellison GW, Myers LW. A hu-
moral response to oligodendrocyte-specific protein in MS: a potential
molecular mimic. Neurology 1999;53:154e161.
73. Stevens DB, Chen K, Seitz RS, Sercarz EE, Bronstein JM. Oligoden-
drocyte-specific protein peptides induce experimental autoimmune
encephalomyelitis in SJL/J mice. J Immunol 1999;162:7501e7509.
74. Martinez-Gonzalez O, Cantero-Hinojosa J, Paule-Sastre P, Gomez-
Magan JC, Salvatierra-Rios D. Intestinal permeability in patients
with ankylosing spondylitis and their healthy relatives. Br J Rheuma-
tol 1994;33:644e647.
75. Mielants H, De Vos M, Goemaere S, Schelstraete K, Cuvelier C,
Goethals K, et al. Intestinal mucosal permeability in inflammatory
rheumatic diseases. II. Role of disease. J Rheumatol 1991;18:
394e400.
76. Smith MD, Gibson RA, Brooks PM. Abnormal bowel permeability in
ankylosing spondylitis and rheumatoid arthritis. J Rheumatol 1985;
12:299e305.
77. Wan H, Winton HL, Soeller C, Taylor GW, Gruenert DC,
Thompson PJ, et al. The transmembrane protein occludin of epithe-
lial tight junctions is a functional target for serine peptidases from
faecal pellets of Dermatophagoides pteronyssinus. Clin Exp Allergy
2001;31:279e294.
78. Wan H, Winton HL, Soeller C, Tovey ER, Gruenert DC,
Thompson PJ, et al. Der p 1 facilitates transepithelial allergen deliv-
ery by disruption of tight junctions. J Clin Invest 1999;104:123e133.
79. Tai HY, Tam MF, Chou H, Peng HJ, Su SN, Perng DW, et al. Pen ch
13 allergen induces secretion of mediators and degradation of
occludin protein of human lung epithelial cells. Allergy 2006;61:
382e388.
80. Kominsky SL, Argani P, Korz D, Evron E, Raman V, Garrett E, et al.
Loss of the tight junction protein claudin-7 correlates with histolog-
ical grade in both ductal carcinoma in situ and invasive ductal carci-
noma of the breast. Oncogene 2003;22:2021e2033.
81. Hewitt KJ, Agarwal R, Morin PJ. The claudin gene family: expres-
sion in normal and neoplastic tissues. BMC Cancer 2006;6:186.
82. Hough CD, Sherman-Baust CA, Pizer ES, Montz FJ, Im DD,
Rosenshein NB, et al. Large-scale serial analysis of gene expression
reveals genes differentially expressed in ovarian cancer. Cancer Res
2000;60:6281e6287.
83. Agarwal R, D’Souza T, Morin PJ. Claudin-3 and claudin-4 expres-
sion in ovarian epithelial cells enhances invasion and is associated
with increased matrix metalloproteinase-2 activity. Cancer Res
2005;65:7378e7385.

477Pathologies of the Tight Junction
84. Hoevel T, Macek R, Swisshelm K, Kubbies M. Reexpression of the
TJ protein CLDN1 induces apoptosis in breast tumor spheroids. Int J
Cancer 2004;108:374e383.
85. Osanai T, Tsuchiya T, Ogino T, Nakahara K. Long-term prevention
of skeletal complications by pamidronate in a patient with bone me-
tastasis from endometrial carcinoma: a case report. Gynecol Oncol
2006;100:195e197.
86. Morin PJ. Claudin proteins in human cancer: promising new targets
for diagnosis and therapy. Cancer Res 2005;65:9603e9606.
87. Polak-Charcon S, Shoham J, Ben-Shaul Y. Tight junctions in epithe-
lial cells of human fetal hindgut, normal colon, and colon adenocar-
cinoma. J Natl Cancer Inst 1980;65:53e62.
88. Morita K, Tsukita S, Miyachi Y. Tight junction-associated proteins
(occludin, ZO-1, claudin-1, claudin-4) in squamous cell carcinoma
and Bowen’s disease. Br J Dermatol 2004;151:328e334.
89. Kerjaschki D, Krisch K, Sleyter UB, Umrath W, Jakesz R, Depisch D,
et al. The structure of tight junctions in human thyroid tumors. A sys-
tematic freeze-fracture study. Am J Pathol 1979;96:207e225.
90. Mullin JM. Potential interplay between luminal growth factors and
increased tight junction permeability in epithelial carcinogenesis.
J Exp Zool 1997;279:484e489.
91. Tobioka H, Isomura H, Kokai Y, Tokunaga Y, Yamaguchi J,
Sawada N. Occludin expression decreases with the progression of
human endometrial carcinoma. Hum Pathol 2004;35:159e164.
92. Billings SD, Walsh SV, Fisher C, Nusrat A, Weiss SW, Folpe AL.
Aberrant expression of tight junction-related proteins ZO-1, clau-
din-1 and occludin in synovial sarcoma: an immunohistochemical
study with ultrastructural correlation. Mod Pathol 2004;17:141e149.
93. Sobel G, Szabo I, Paska C, Kiss A, Kovalszky I, Kadar A, et al. Changes
of cell adhesion and extracellular matrix (ECM) components in cervi-
cal intraepithelial neoplasia. Pathol Oncol Res 2005;11:26e31.
94. Busch C, Hanssen TA, Wagener C, B OB. Down-regulation of CEA-
CAM1 in human prostate cancer: correlation with loss of cell polar-
ity, increased proliferation rate, and Gleason grade 3 to 4 transition.
Hum Pathol 2002;33:290e298.
95. Marzioni D, Banita M, Felici A, Paradinas FJ, Newlands E, De
NictolisM, et al.Expressionof ZO-1andoccludin in normalhumanpla-
centa and in hydatidiform moles. Mol Hum Reprod 2001;7:279e285.
96. Katahira J, Inoue N, Horiguchi Y, Matsuda M, Sugimoto N. Molec-
ular cloning and functional characterization of the receptor for Clos-
tridium perfringens enterotoxin. J Cell Biol 1997;136:1239e1247.
97. Briehl MM, Miesfeld RL. Isolation and characterization of tran-
scripts induced by androgen withdrawal and apoptotic cell death in
the rat ventral prostate. Mol Endocrinol 1991;5:1381e1388.
98. Furuse M, Tsukita S. Claudins in occluding junctions of humans and
flies. Trends Cell Biol 2006;16:181e188.
99. Fujita K, Katahira J, Horiguchi Y, Sonoda N, Furuse M, Tsukita S.
Clostridium perfringens enterotoxin binds to the second extracellular
loop of claudin-3, a tight junction integral membrane protein. FEBS
Lett 2000;476:258e261.
100. McClane BA, Chakrabarti G. New insights into the cytotoxic mech-
anisms of Clostridium perfringens enterotoxin. Anaerobe 2004;10:
107e114.
101. McClane BA, Singh U. Interactions between Clostridium perfringens
enterotoxin and tight junctions proteins. In: Cereijido M,
Anderson JM, eds. Tight Junctions. 2nd ed. Boca Raton FL: CRC
Press; 2001. pp. 517e532.
102. Chen ML, Pothoulakis C, LaMont JT. Protein kinase C signaling reg-
ulates ZO-1 translocation and increased paracellular flux of T84 co-
lonocytes exposed to Clostridium difficile toxin A. J Biol Chem 2002;
277:4247e4254.
103. Nusrat A, Eichel-Streiber C, Turner JR, Verkade P, Madara JL,
Parkos CA. Clostridium difficile toxins disrupt epithelial barrier func-
tion by altering membrane microdomain localization of tight junction
proteins. Infect Immun 2001;69:1329e1336.
104. Fedwick JP, Lapointe TK, Meddings JB, Sherman PM, Buret AG.
Helicobacter pylori activates myosin light-chain kinase to disrupt
claudin-4 and claudin-5 and increase epithelial permeability. Infect
Immun 2005;73:7844e7852.
105. Suzuki K, Kokai Y, Sawada N, Takakuwa R, Kuwahara K, Isogai E,
et al. SS1 Helicobacter pylori disrupts the paracellular barrier of the
gastric mucosa and leads to neutrophilic gastritis in mice. Virchows
Arch 2002;440:318e324.
106. Amieva MR, Vogelmann R, Covacci A, Tompkins LS, Nelson WJ,
Falkow S. Disruption of the epithelial apical-junctional complex by
Helicobacter pylori CagA. Science 2003;300:1430e1434.
107. Simonovic I, Rosenberg J, Koutsouris A, Hecht G. Enteropathogenic
Escherichia coli dephosphorylates and dissociates occludin from
intestinal epithelial tight junctions. Cell Microbiol 2000;2:305e315.
108. Wu Z, Nybom P, Magnusson KE. Distinct effects of Vibrio cholerae
haemagglutinin/protease on the structure and localization of the tight
junction-associated proteins occludin and ZO-1. Cell Microbiol
2000;2:11e17.
109. Pentecost M, Otto G, Theriot JA, Amieva MR. Listeria monocyto-
genes invades the epithelial junctions at sites of cell extrusion. PloS
Pathog 2006;2:e3.
110. MacIntyre A, Hammond CJ, Little CS, Appelt DM, Balin BJ. Chla-
mydia pneumoniae infection alters the junctional complex proteins of
human brain microvascular endothelial cells. FEMS Microbiol Lett
2002;217:167e172.
111. Chen SH, Stins MF, Huang SH, Chen YH, Kwon-Chung KJ,
Chang Y, et al. Cryptococcus neoformans induces alterations in the
cytoskeleton of human brain microvascular endothelial cells. J Med
Microbiol 2003;52:961e970.
112. Razin S, Yogev D, Naot Y. Molecular biology and pathogenicity of
mycoplasmas. Microbiol Mol Biol Rev 1998;62:1094e1156.
113. Dickman KG, Hempson SJ, Anderson J, Lippe S, Zhao L,
Burakoff R, et al. Rotavirus alters paracellular permeability and en-
ergy metabolism in Caco-2 cells. Am J Physiol Gastrointest Liver
Physiol 2000;279:G757eG766.
114. Dallasta LM, Pisarov LA, Esplen JE, Werley JV, Moses AV,
Nelson JA, et al. Blood-brain barrier tight junction disruption in hu-
man immunodeficiency virus-1 encephalitis. Am J Pathol 1999;155:
1915e1927.
115. Barton ES, Forrest JC, Connolly JL, Chappell JD, Liu Y, Schnell FJ,
et al. Junction adhesion molecule is a receptor for reovirus. Cell
2001;104:441e451.
116. Spear PG, Eisenberg RJ, Cohen GH. Three classes of cell surface
receptors for alphaherpesvirus entry. Virology 2000;275:1e8.
117. Lauwaet T, Oliveira MJ, Callewaert B, De Bruyne G, Mareel M,
Leroy A. Proteinase inhibitors TPCK and TLCK prevent Entamoebahistolytica induced disturbance of tight junctions and microvilli in
enteric cell layers in vitro. Int J Parasitol 2004;34:785e794.
118. Nava P, Lopez S, Arias CF, Islas S, Gonzalez-Mariscal L. The ro-
tavirus surface protein VP8 modulates the gate and fence function
of tight junctions in epithelial cells. J Cell Sci 2004;117:
5509e5519.
119. Zheng J, Xie Y, Campbell R, Song J, Massachi S, Razi M, et al. In-
volvement of claudin-7 in HIV infection of CD4� cells. Retrovirol-
ogy 2005;2:79.
120. Leroy A, Lauwaet T, De Bruyne G, Cornelissen M, Mareel M. Ent-
amoeba histolytica disturbs the tight junction complex in human
enteric T84 cell layers. FASEB J 2000;14:1139e1146.
121. Gonzalez-Mariscal L, Nava P. Tight junctions, from tight intercellu-
lar seals to sophisticated protein complexes involved in drug delivery,
pathogens interaction and cell proliferation. Adv Drug Deliv Rev
2005;57:811e814.
122. Chen SC, Eiting K, Cui K, Leonard AK, Morris D, Li CY, et al. Ther-
apeutic utility of a novel tight junction modulating peptide for en-
hancing intranasal drug delivery. J Pharm Sci 2006;95:1364e1371.

478 Cereijido et al. / Archives of Medical Research 38 (2007) 465e478
123. Sileno AP, Brandt GC, Spann BM, Quay SC. Lower mean weight
after 14 days intravenous administration peptide YY3-36 (PYY3-36)
in rabbits. Int J Obes (Lond) 2006;30:68e72.
124. Gallardo JM, Hernandez JM, Contreras RG, Flores-Maldonado C,
Gonzalez-Mariscal L, Cereijido M. Tight junctions are sensitive
to peptides eliminated in the urine. J Membr Biol 2002;188:
33e42.
125. Flores-Benitez D, Ruiz A, Shoshani L, Cereijido M, Contreras RG.
Control of tight junctional sealing: role of the epidermal growth
factor. Am J Physiol 2007;292:F828eF836.
126. Tokes AM, Kulka J, Paku S, Szik A, Paska C, Novak PK, et al. Clau-
din-1, -3 and -4 proteins and mRNA expression in benign and malig-
nant breast lesions: a research study. Breast Cancer Res 2005;7:
R296eR305.
127. Kramer F, White K, Kubbies M, Swisshelm K, Weber BH. Genomic
organization of claudin-1 and its assessment in hereditary and
sporadic breast cancer. Hum Genet 2000;107:249e256.
128. Hibbs K, Skubitz KM, Pambuccian SE, Casey RC, Burleson KM,
Oegema TR Jr, et al. Differential gene expression in ovarian carci-
noma: identification of potential biomarkers. Am J Pathol 2004;
165:397e414.
129. Santin AD, Zhan F, Bellone S, Palmieri M, Cane S, Bignotti E, et al.
Gene expression profiles in primary ovarian serous papillary tumors
and normal ovarian epithelium: identification of candidate molecular
markers for ovarian cancer diagnosis and therapy. Int J Cancer 2004;
112:14e25.
130. Rangel LB, Agarwal R, D’Souza T, Pizer ES, Alo PL, Lancaster WD,
et al. Tight junction proteins claudin-3 and claudin-4 are frequently
overexpressed in ovarian cancer but not in ovarian cystadenomas.
Clin Cancer Res 2003;9:2567e2575.
131. Resnick MB, Gavilanez M, Newton E, Konkin T, Bhattacharya B,
Britt DE, et al. Claudin expression in gastric adenocarcinomas: a tis-
sue microarray study with prognostic correlation. Hum Pathol 2005;
36:886e892.
132. Dhawan P, Singh AB, Deane NG, No Y, Shiou SR, Schmidt C, et al.
Claudin-1 regulates cellular transformation and metastatic behavior
in colon cancer. J Clin Invest 2005;115:1765e1776.
133. Sobel G, Paska C, Szabo I, Kiss A, Kadar A, Schaff Z. Increased ex-
pression of claudins in cervical squamous intraepithelial neoplasia
and invasive carcinoma. Hum Pathol 2005;36:162e169.
134. Lee SK, Moon J, Park SW, Song SY, Chung JB, Kang JK. Loss of the
tight junction protein claudin 4 correlates with histological growth-
pattern and differentiation in advanced gastric adenocarcinoma.
Oncol Rep 2005;13:193e199.
135. Johnson AH, Frierson HF, Zaika A, Powell SM, Roche J, Crowe S,
et al. Expression of tight-junction protein claudin-7 is an early event
in gastric tumorigenesis. Am J Pathol 2005;167:577e584.
136. Cheung ST, Leung KL, Ip YC, Chen X, Fong DY, Ng IO, et al. Clau-
din-10 expression level is associated with recurrence of primary
hepatocellular carcinoma. Clin Cancer Res 2005;11:551e556.
137. Nichols LS, Ashfaq R, Iacobuzio-Donahue CA. Claudin 4 protein
expression in primary and metastatic pancreatic cancer: support for
use as a therapeutic target. Am J Clin Pathol 2004;121:226e230.
138. Long H, Crean CD, Lee WH, Cummings OW, Gabig TG. Expression
of Clostridium perfringens enterotoxin receptors claudin-3 and clau-
din-4 in prostate cancer epithelium. Cancer Res 2001;61:7878e7881.




















