
Rho Kinase Regulates Tight Junction Function and Is Necessary for Tight Junction Assembly in...
14
Rho Kinase Regulates Tight Junction Function and Is Necessary for Tight Junction Assembly in Polarized Intestinal Epithelia SHAUN V. WALSH,* ANN M. HOPKINS,* JASON CHEN,* SHUH NARUMIYA, ‡ CHARLES A. PARKOS,* and ASMA NUSRAT* *Epithelial Pathobiology Research Unit, Department of Pathology, Emory University School of Medicine, Atlanta, Georgia; and ‡ Department of Pharmacology, Kyoto University Faculty of Medicine, Kyoto, Japan Background & Aims: Tight junctions are crucial determi- nants of barrier function in polarized intestinal epithelia and are regulated by Rho guanosine triphosphatase. Rho kinase (ROCK) is a downstream effector of Rho. Methods: A specific inhibitor of ROCK, Y-27632, was used to examine the role of ROCK in the regulation of tight junctions in model intestinal (T84) cells by electro- physiologic, biochemical, morphologic, and molecular biologic approaches. Results: ROCK inhibition induced reorganization of apical F-actin structures and enhanced paracellular permeability but did not alter the distribu- tion or detergent solubility of tight junction proteins. Confocal microscopy showed colocalization of a subpool of ROCK with the tight junction protein zonula occludens 1. Inhibition of ROCK function by a dominant negative mutant of ROCK also produced reorganization of apical F-actin structures without disruption of tight junctions. ROCK inhibition in calcium switch assays showed that ROCK is necessary for the assembly of tight and adhe- rens junctions. Upon calcium repletion, occludin, zonula occludens 1, and E-cadherin failed to redistribute to the intercellular junctions; assembly of the apical F-actin cytoskeleton was prevented; and barrier function failed to recover. Conclusions: We suggest that ROCK regu- lates intact tight junctions via its effects on the F-actin cytoskeleton. ROCK is also critical for assembly of the apical junctional proteins and the F-actin cytoskeleton organization during junctional formation. T he epithelial lining of the gastrointestinal tract forms a crucial barrier that separates luminal con- tents from the underlying tissue compartments. Paracel- lular permeability across epithelial cells is primarily reg- ulated by the apical-most intercellular junction referred to as the tight junction (TJ). TJs are complex, dynami- cally regulated structures that form a beltlike band en- circling the apical-most region of the lateral plasma membrane. Within the TJ, adjacent plasma membranes are in intimate contact with each other in a series of “membrane kisses.” 1 Such areas of close membrane con- tact appear on freeze fracture replicas as a meshlike series of interconnected strands and grooves. 2 Proteins that constitute the TJ complex include transmembrane pro- teins such as occludin, claudin(s), and junction adhesion molecule. 3–7 These proteins are believed to affiliate with the underlying actin cytoskeleton via linker proteins such as zonula occludens 1 (ZO-1). The C-terminal re- gion of ZO-1 co-sediments with F-actin and thereby provides a mechanism of linkage between TJs and the actin cytoskeleton. 8 F-actin filaments in the apical peri- junctional F-actin ring of epithelial cells associate with TJs and regulate TJ function in diverse physiologic and pathologic states. 9 –11 Intestinal epithelial TJs are also influenced by a variety of endogenous and exogenous agents, including glucose, bacterial antigens, cytokines, and growth factors. 12–15 However, the mechanisms by which these external agents signal to the TJs are not well understood. Recently, the Rho family of small guanosine triphos- phate (GTP)– binding proteins has been shown to be important in the regulation of epithelial TJ structure, function, and assembly. 16 –19 We have previously shown that inactivation of Rho GTPases (with Clostridium bot- ulinum C3 transferase and Clostridium difficile toxins A and B) is associated with reorganization of F-actin in the perijunctional F-actin ring. These effects on the actin cytoskeleton are accompanied by changes in TJ structure and function 16 (Nusrat et al., Infect Immun 2001 [in Abbreviations used in this paper: AFSF, antibiotic-free, serum-free; AJ, adherens junction; BSA, bovine serum albumin; DMEM, Dulbecco modified Eagle medium; EVOM, epithelial voltohmmeter; FD-3, fluo- resceinated dextran, MW 3000; GTP, guanosine triphosphate; HBSS 1 , Hank’s balanced salt solution containing calcium chloride and mag- nesium sulfate; HBSS 2 , Hank’s balanced salt solution without calcium chloride or magnesium sulfate; ROCK, Rho kinase; RT, room temper- ature; SDS, sodium dodecyl sulfate; TER, transepithelial resistance; TJ, tight junction; ZO-1, zonula occludens 1. © 2001 by the American Gastroenterological Association 0016-5085/01/$35.00 doi:10.1053/gast.2001.27060 GASTROENTEROLOGY 2001;121:566 –579
Transcript of Rho Kinase Regulates Tight Junction Function and Is Necessary for Tight Junction Assembly in...

Rho Kinase Regulates Tight Junction Function and IsNecessary for Tight Junction Assembly in PolarizedIntestinal Epithelia
SHAUN V. WALSH,* ANN M. HOPKINS,* JASON CHEN,* SHUH NARUMIYA,‡
CHARLES A. PARKOS,* and ASMA NUSRAT**Epithelial Pathobiology Research Unit, Department of Pathology, Emory University School of Medicine, Atlanta, Georgia; and ‡Department ofPharmacology, Kyoto University Faculty of Medicine, Kyoto, Japan
Background & Aims: Tight junctions are crucial determi-nants of barrier function in polarized intestinal epitheliaand are regulated by Rho guanosine triphosphatase.Rho kinase (ROCK) is a downstream effector of Rho.Methods: A specific inhibitor of ROCK, Y-27632, wasused to examine the role of ROCK in the regulation oftight junctions in model intestinal (T84) cells by electro-physiologic, biochemical, morphologic, and molecularbiologic approaches. Results: ROCK inhibition inducedreorganization of apical F-actin structures and enhancedparacellular permeability but did not alter the distribu-tion or detergent solubility of tight junction proteins.Confocal microscopy showed colocalization of a subpoolof ROCK with the tight junction protein zonula occludens1. Inhibition of ROCK function by a dominant negativemutant of ROCK also produced reorganization of apicalF-actin structures without disruption of tight junctions.ROCK inhibition in calcium switch assays showed thatROCK is necessary for the assembly of tight and adhe-rens junctions. Upon calcium repletion, occludin, zonulaoccludens 1, and E-cadherin failed to redistribute to theintercellular junctions; assembly of the apical F-actincytoskeleton was prevented; and barrier function failedto recover. Conclusions: We suggest that ROCK regu-lates intact tight junctions via its effects on the F-actincytoskeleton. ROCK is also critical for assembly of theapical junctional proteins and the F-actin cytoskeletonorganization during junctional formation.
The epithelial lining of the gastrointestinal tractforms a crucial barrier that separates luminal con-
tents from the underlying tissue compartments. Paracel-lular permeability across epithelial cells is primarily reg-ulated by the apical-most intercellular junction referredto as the tight junction (TJ). TJs are complex, dynami-cally regulated structures that form a beltlike band en-circling the apical-most region of the lateral plasmamembrane. Within the TJ, adjacent plasma membranesare in intimate contact with each other in a series of“membrane kisses.”1 Such areas of close membrane con-
tact appear on freeze fracture replicas as a meshlike seriesof interconnected strands and grooves.2 Proteins thatconstitute the TJ complex include transmembrane pro-teins such as occludin, claudin(s), and junction adhesionmolecule.3–7 These proteins are believed to affiliate withthe underlying actin cytoskeleton via linker proteinssuch as zonula occludens 1 (ZO-1). The C-terminal re-gion of ZO-1 co-sediments with F-actin and therebyprovides a mechanism of linkage between TJs and theactin cytoskeleton.8 F-actin filaments in the apical peri-junctional F-actin ring of epithelial cells associate withTJs and regulate TJ function in diverse physiologic andpathologic states.9–11 Intestinal epithelial TJs are alsoinfluenced by a variety of endogenous and exogenousagents, including glucose, bacterial antigens, cytokines,and growth factors.12–15 However, the mechanisms bywhich these external agents signal to the TJs are not wellunderstood.
Recently, the Rho family of small guanosine triphos-phate (GTP)–binding proteins has been shown to beimportant in the regulation of epithelial TJ structure,function, and assembly.16–19 We have previously shownthat inactivation of Rho GTPases (with Clostridium bot-ulinum C3 transferase and Clostridium difficile toxins Aand B) is associated with reorganization of F-actin in theperijunctional F-actin ring. These effects on the actincytoskeleton are accompanied by changes in TJ structureand function16 (Nusrat et al., Infect Immun 2001 [in
Abbreviations used in this paper: AFSF, antibiotic-free, serum-free;AJ, adherens junction; BSA, bovine serum albumin; DMEM, Dulbeccomodified Eagle medium; EVOM, epithelial voltohmmeter; FD-3, fluo-resceinated dextran, MW 3000; GTP, guanosine triphosphate; HBSS1,Hank’s balanced salt solution containing calcium chloride and mag-nesium sulfate; HBSS2, Hank’s balanced salt solution without calciumchloride or magnesium sulfate; ROCK, Rho kinase; RT, room temper-ature; SDS, sodium dodecyl sulfate; TER, transepithelial resistance; TJ,tight junction; ZO-1, zonula occludens 1.
© 2001 by the American Gastroenterological Association0016-5085/01/$35.00
doi:10.1053/gast.2001.27060
GASTROENTEROLOGY 2001;121:566–579

press]). Additional studies using transfected epithelialcell lines expressing mutants of Rho GTPases have doc-umented a role for these proteins in regulation of TJstructure and function.20,21 In other cell types, such asfibroblasts, Rho GTPases primarily regulate the F-actincytoskeleton in basal stress fibers and are important inlamellipodial and filopodial extensions of migratingcells. Thus, the Rho family of GTP-binding proteins arecentral in regulating actin organization of stationary andmigrating cells.
However, mechanisms by which Rho proteins in-fluence TJs of epithelial cells are not clear. It is knownthat the apical actin cytoskeleton is central in theregulation of TJ function and that F-actin organiza-tion itself is regulated by Rho GTPases. In particular,it is not known if specific downstream effectors of Rhomediate its effects on TJs. It is also unknown whethersuch effectors play a role in Rho-dependent TJ assem-bly. Potential candidate downstream effectors of Rhoinclude the family of serine-threonine kinase isoen-zymes termed p160ROCK (ROKb, ROCK I) and Rhokinase (ROKa /ROCK II).22–26 ROCK is important inmany cellular processes that involve actin cytoskeletalrearrangement, including stress fiber formation, ax-onal growth, tumor cell invasion, and activation ofplatelets.23,27–30 ROCK is a major regulator of cy-toskeletal contractility via its influence on myosinphosphatase and myosin light chain itself.31–35 Acti-vation of this pathway has been associated with actin-myosin contraction.11 Because the actin cytoskeletonis central in regulating epithelial TJs and Rho pro-teins have been documented to influence this function,we determined the role of ROCK in regulation ofTJ structure/function. Using a specific inhibitor ofROCK, Y-27632 [(1)-(R)-trans-4-(1-aminoethyl)-N-(4-pyridyl) cyclohexanecarboxamide], we examinedthe role of ROCK in TJ structure/function.36–38 We furthercorroborated our results using a dominant negative mutantof ROCK. Our results suggest a central role of ROCK inregulation of TJ function. ROCK inhibition is associatedwith dramatic restructuring of the perijunctional F-actincytoskeleton, which has previously been shown to affiliatewith TJs. However, no change in distribution of TJ proteinswas observed, implying that the functional effects of ROCKinhibition on paracellular permeability probably are second-ary to actin cytoskeletal reorganization. Furthermore, ourresults suggest a critical role for ROCK in the assembly ofTJs and adherens junctions (AJs). ROCK inhibition com-pletely abrogated the recovery of barrier function and thetargeting of TJ and AJ proteins to the intercellular junc-tions.
Materials and MethodsCell Culture
T84 cells (American Type Culture Collection, Rockville,MD) were grown in a 1:1 mixture of Dulbecco modified Eaglemedium (DMEM) and Ham F-12 medium supplemented with15 mmol/L HEPES (pH 7.5), 14 mmol/L NaHCO3, 40 mg/mLpenicillin, 8 mg/mL ampicillin, 90 mg/mL streptomycin, and 5%newborn calf serum. Cells were subcultured every 6–8 days with0.1% trypsin and 1.0 mmol/L EDTA in Ca21- and Mg21-freephosphate-buffered saline. For electrophysiologic, immunofluo-rescent, and ultrastructural experiments, T84 cells were grown oncollagen-coated, permeable polycarbonate filters (5 mm pore size)with a surface area of 0.33 cm2 (Costar, Cambridge, MA), aspreviously described.39 For biochemical experiments, T84 cellswere grown on 5-cm2 collagen-coated inserts.40 Caco-2 cells(American Type Culture Collection) were grown in DMEM sup-plemented with 10% fetal bovine serum, 0.292 mg/mL L-glu-tamine, penicillin G, sodium pyruvate, and streptomycin sulfate100 U/mL.
Materials
DC3B toxin (3.2 mg/mL basolaterally for 6 hours, kindgift of Dr. P. Boquet, INSERM, Nice, France) was used toinhibit Rho function in T84 cell monolayers, as previouslydescribed.16,41 Y-27632, a Rho kinase–specific inhibitor, was akind gift of Welfide Corporation, Japan.36,37 All experimentswere performed in antibiotic-free, serum-free media (AFSF),with ultrapure bovine serum albumin (BSA; Sigma, St. Louis,MO) as a carrier protein.
Electrophysiology and ParacellularPermeability
Measurement of transepithelial electrical resistance(TER) was performed using a commercially available epithelialvoltohmmeter (EVOM; World Precision Instruments, FL).Paracellular permeability was determined by measuring apicalto basolateral flux of fluoresceinated dextran (FD-3, MW 3000;Molecular Probes, Eugene, OR) using a modification of pre-viously described methods.42 Briefly, confluent epithelialmonolayers on 0.33-cm2 permeable supports were washedtwice with Hank’s balanced salt solution containing calciumchloride and magnesium sulfate (HBSS1) and maintained at37°C on a shaking, warm plate. FD-3, 1 mg/mL, was addedapically at time 0, and 50-mL samples were removed from thebasolateral compartment at 30-minute intervals from 0 to 120minutes, inclusive. The fluorescence intensity of each samplewas quantitated on a Cytofluor 2350 fluorescence measurementsystem (Millipore, Bedford, MA), and the FD-3 protein con-tent was evaluated against a standard curve of known FD-3concentrations.
Immunofluorescent Localization of ROCK,Junctional Proteins, and F-Actin
Monolayers of human intestinal epithelial cell lines (T84,Caco-2) were grown to confluence on collagen-coated permeable
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 567

supports as described and incubated with DC3B toxin orY-27632 in AFSF. Medium alone was used as control. Cells werewashed in HBSS1 and fixed as described below. To visualizeF-actin microfilaments, monolayers were fixed in 3.7% parafor-maldehyde, permeabilized in 0.5% Triton X100 (TX-100), andincubated with rhodamine phalloidin (Molecular Probes, Eugene,OR) for 40 minutes at room temperature (RT). To visualizeintercellular TJ proteins, AJ proteins, ROCK, and villin, T84 orCaco-2 monolayers were fixed in 100% ethanol at 220°C for 20minutes. Monolayers were incubated in 5% normal goat serum inHBSS1 (1 hour at RT), with primary antibody (1 hour at RT) andfluoresceinated secondary antibodies (1:400 dilution; Jackson Im-munoresearch Laboratory, West Grove, PA; 1 hour at RT). Allmonolayers were mounted in phosphate-buffered saline/glycerol/p-phenylenediamine, 1:1:0.01 (vol/vol/vol) and analyzed by con-focal microscopy (Zeiss LSM 5.10; Zeiss, Thornwood, NY).Monoclonal antibodies used in this study included anti–b-catenin(Transduction Laboratories, Lexington, KY), anti–E-cadherin(Zymed, San Francisco, CA), anti–ZO-1 (Transduction Laborato-ries) (for colocalization experiments), antivillin antibody (AmacInc, Westbrook, ME) and anti-cMyc antibody (Clontech, PaloAlto, CA). Polyclonal antibodies used included antioccludin,anti–ZO-1, anti–claudin-1, and (Zymed) anti-ROCK 20 490(S. Narumiya, Kyoto, Japan).
Differential Detergent Extraction andWestern Blotting
Confluent monolayers (5 cm2) of T84 intestinal epi-thelial cells cultured on collagen-coated permeable supportswere exposed to Y-27632 (100 mmol/L) or AFSF mediumalone for 1 hour. Monolayers were then incubated for 30minutes at 4°C with 1% Triton X-100 (TX-100) extractionbuffer (Relax buffer: 100 mmol/L KCl, 3 mmol/L NaCl, 3.5mmol/L MgCl2 , 10 mmol/L HEPES, pH 7.4, and proteaseinhibitors [phenylmethylsulfonyl fluoride, 50 mg/mL; leupep-tin, 2 mg/mL; pepstatin, 1 mg/mL; aprotinin, 2 mg/mL]). TheTX-100–soluble fraction was removed, subjected to a low-speed centrifugation step (1500g for 10 minutes to remove anycell debris), and added to an equal volume of sodium dodecylsulfate (SDS) sample buffer. The remaining cells constitutingthe TX-100–insoluble fraction were scraped with a rubberpoliceman into an equal volume of SDS sample buffer. Anequal number of cell equivalents of TX-100–soluble and–insoluble fractions were then loaded onto 6%–16% poly-acrylamide gradient gels and analyzed by SDS–polyacrylamidegel electrophoresis (PAGE) and immunoblotting for TJ pro-teins, as previously described.40
Electron Microscopy
Monolayers were treated with Y-27632 (50–100mmol/L) or vehicle alone for 1 hour, fixed with 4% bufferedglutaraldehyde, and cut into 1-mm strips using a microtome.After a buffer wash, the strips were postfixed in 1% osmiumtetroxide, dehydrated through graded alcohols through pro-pylene oxide, then infiltrated with Embed-812 (Electron Mi-croscopy Sciences, Ft. Washington, PA). Strips were embedded
for cross-section orientation. Semithin (0.5 mm) sections werecut and stained with toluidine blue and examined for ade-quacy. Ultrathin sections (900 Å) were cut with a diamondknife, stained with uranyl acetate and lead citrate, and exam-ined with a Philips EM201 electron microscope.
Calcium Switch Assay
T84 monolayers grown on 0.33-cm2 permeable poly-carbonate filters (5 mm pore size) were treated with 2 mmol/LEGTA in Hank’s balanced salt solution without calcium ormagnesium (HBSS2) for 20 minutes (37°C) under sterileconditions to disrupt TJs as previously described.7,39 Afterwashing, monolayers were incubated in sterile cell culturemedium containing 20 mmol/L Y-27632 or medium alone ina 37°C tissue culture incubator. Recovery of TER to passiveion flow was monitored at intervals over 24 hours using anEVOM. Replicate monolayers were fixed before and duringcalcium depletion and after recovery to analyze the immuno-localization of junctional proteins and F-actin.
Transfection
A kinase-defective, Rho binding–defective dominantnegative mutant of p160ROCK (ROCK-KDIA) was used toinvestigate the effects of inhibition of ROCK function on apicalactin in Caco-2 intestinal epithelial cells.43 Briefly, Caco-2 cellswere plated on glass coverslips 24 hours before transfection andcultured in serum-free media. Plasmids to be transfected (ROCK-KDIA: pCAG-myc-ROCK, empty vector control: pCAG-myc)were first purified using the Qiagen (Valencia, CA) maxiplasmidkit. Cells were transfected with plasmid DNA by the applicationof lipofectamine-DNA coprecipitates (Life Technologies Inc.,Carlsbad, CA). At 3 hours, cells were incubated with DMEMsupplemented with 10% FBS, 0.292 mg/mL L-glutamine, peni-cillin G, sodium pyruvate, and streptomycin sulfate, 100 U/mL,for 48 hours. Fixation, permeabilization, and other methods forimmunofluorescence are described above. Cells were first incu-bated with anti-Myc antibody and then stained with rhodaminephalloidin to highlight F-actin or with antibodies to occludin.
Data Analysis
Numeric values for each parameter of control andY-27632–treated monolayers were pooled and expressed asmeans 6 SEM. The Student t test was used to examinestatistical differences between control and test monolayers.Statistical significance was assumed at P values of ,0.05.
ResultsROCK Inhibition Enhances ParacellularPermeability in Epithelial Monolayers.
We used a specific inhibitor of ROCK, Y-27632,for our studies. Control monolayers maintained highTER values of ;1500 V z cm2 in serum-free medium.ROCK inhibition with Y-27632 induced a rapid dose-dependent decrease in T84 monolayer TER (Figure 1). A
568 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3
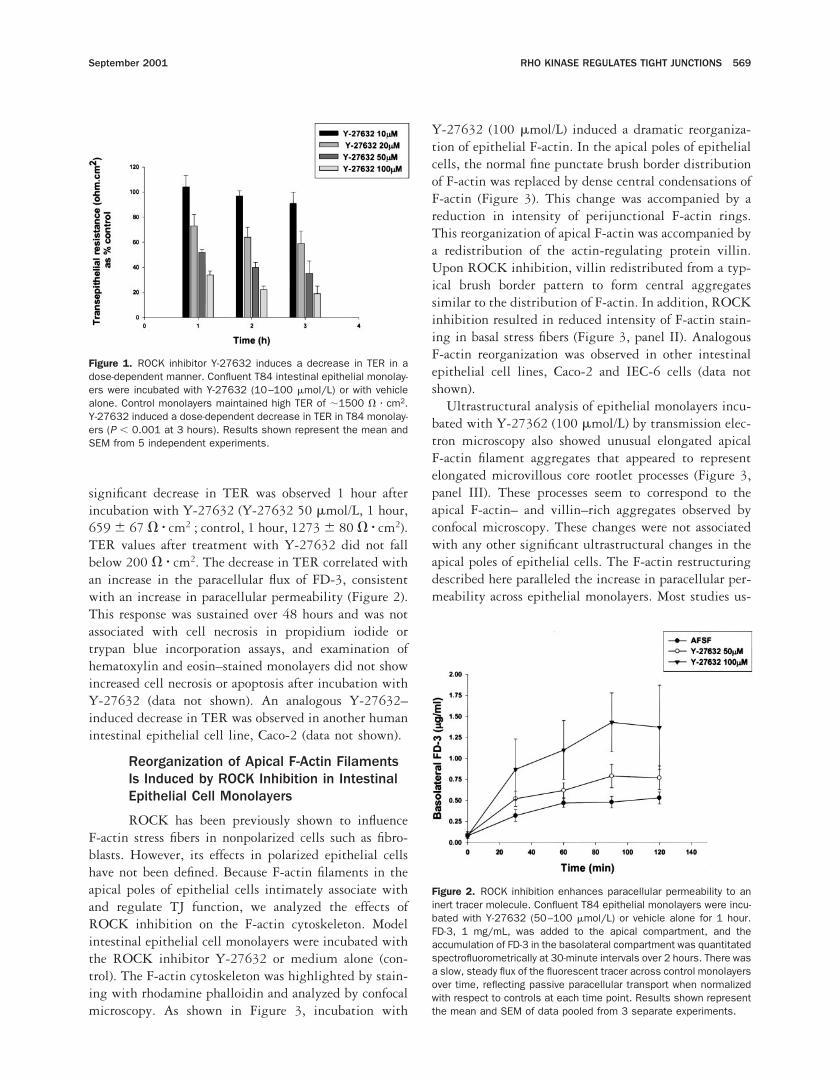
significant decrease in TER was observed 1 hour afterincubation with Y-27632 (Y-27632 50 mmol/L, 1 hour,659 6 67 V z cm2 ; control, 1 hour, 1273 6 80 V z cm2).TER values after treatment with Y-27632 did not fallbelow 200 V z cm2. The decrease in TER correlated withan increase in the paracellular flux of FD-3, consistentwith an increase in paracellular permeability (Figure 2).This response was sustained over 48 hours and was notassociated with cell necrosis in propidium iodide ortrypan blue incorporation assays, and examination ofhematoxylin and eosin–stained monolayers did not showincreased cell necrosis or apoptosis after incubation withY-27632 (data not shown). An analogous Y-27632–induced decrease in TER was observed in another humanintestinal epithelial cell line, Caco-2 (data not shown).
Reorganization of Apical F-Actin FilamentsIs Induced by ROCK Inhibition in IntestinalEpithelial Cell Monolayers
ROCK has been previously shown to influenceF-actin stress fibers in nonpolarized cells such as fibro-blasts. However, its effects in polarized epithelial cellshave not been defined. Because F-actin filaments in theapical poles of epithelial cells intimately associate withand regulate TJ function, we analyzed the effects ofROCK inhibition on the F-actin cytoskeleton. Modelintestinal epithelial cell monolayers were incubated withthe ROCK inhibitor Y-27632 or medium alone (con-trol). The F-actin cytoskeleton was highlighted by stain-ing with rhodamine phalloidin and analyzed by confocalmicroscopy. As shown in Figure 3, incubation with
Y-27632 (100 mmol/L) induced a dramatic reorganiza-tion of epithelial F-actin. In the apical poles of epithelialcells, the normal fine punctate brush border distributionof F-actin was replaced by dense central condensations ofF-actin (Figure 3). This change was accompanied by areduction in intensity of perijunctional F-actin rings.This reorganization of apical F-actin was accompanied bya redistribution of the actin-regulating protein villin.Upon ROCK inhibition, villin redistributed from a typ-ical brush border pattern to form central aggregatessimilar to the distribution of F-actin. In addition, ROCKinhibition resulted in reduced intensity of F-actin stain-ing in basal stress fibers (Figure 3, panel II). AnalogousF-actin reorganization was observed in other intestinalepithelial cell lines, Caco-2 and IEC-6 cells (data notshown).
Ultrastructural analysis of epithelial monolayers incu-bated with Y-27362 (100 mmol/L) by transmission elec-tron microscopy also showed unusual elongated apicalF-actin filament aggregates that appeared to representelongated microvillous core rootlet processes (Figure 3,panel III). These processes seem to correspond to theapical F-actin– and villin–rich aggregates observed byconfocal microscopy. These changes were not associatedwith any other significant ultrastructural changes in theapical poles of epithelial cells. The F-actin restructuringdescribed here paralleled the increase in paracellular per-meability across epithelial monolayers. Most studies us-
Figure 1. ROCK inhibitor Y-27632 induces a decrease in TER in adose-dependent manner. Confluent T84 intestinal epithelial monolay-ers were incubated with Y-27632 (10–100 mmol/L) or with vehiclealone. Control monolayers maintained high TER of ;1500 V z cm2.Y-27632 induced a dose-dependent decrease in TER in T84 monolay-ers (P , 0.001 at 3 hours). Results shown represent the mean andSEM from 5 independent experiments.
Figure 2. ROCK inhibition enhances paracellular permeability to aninert tracer molecule. Confluent T84 epithelial monolayers were incu-bated with Y-27632 (50–100 mmol/L) or vehicle alone for 1 hour.FD-3, 1 mg/mL, was added to the apical compartment, and theaccumulation of FD-3 in the basolateral compartment was quantitatedspectrofluorometrically at 30-minute intervals over 2 hours. There wasa slow, steady flux of the fluorescent tracer across control monolayersover time, reflecting passive paracellular transport when normalizedwith respect to controls at each time point. Results shown representthe mean and SEM of data pooled from 3 separate experiments.
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 569

Figure 3. (Panel I) ROCK inhibition induces reorganization of F-actin filaments and the actin-regulating protein villin in the apical pole of intestinalepithelial cells. Confluent T84 and Caco-2 intestinal epithelial monolayers were incubated for 1 hour with the ROCK inhibitor Y-27632 (100mmol/L) or vehicle alone (CON). Cells were fixed and stained with rhodamine phalloidin (red) or antivillin antibodies (green) and analyzed byconfocal laser scanning microscopy. Shown here are en face (xy) images taken at the level of the apical pole of the epithelial monolayer andcomputer-reconstructed vertical section (xz) images taken through the full thickness of the monolayer (scale bar 5 10 mm). The supporting filterin the xz computer-reconstructed images is shown as a blue line, and the apical and basolateral compartments are annotated (a and bl,respectively). (A) In control T84 cell monolayers, F-actin is organized as a dotlike apical microvillous staining pattern and as prominentperijunctional F-actin rings (arrowheads). (B) ROCK inhibition using Y-27632 (100 mmol/L) induced reorganization of apical F-actin with formationof F-actin condensations in the apical plane (arrows). This reorganization was accompanied by a loss of the normal apical microvillous stainingpattern. In addition, the intensity of the perijunctional F-actin ring was reduced after incubation with Y-27632. In the xz plane, the apical denseaggregates of F-actin (arrows) induced by Y-27632 (B, xz) are seen extending down from the apical membrane through the level of the terminalweb but are not detected in control monolayers (A, xz). (D) Analogous reorganization of F-actin was observed in another intestinal epithelial cellline, Caco-2, after incubation with Y-27632. (C) Normal F-actin organization in the apical region of this cell type. (E, G) Villin distribution in theapical poles of control (E) T84 and (G) Caco-2 monolayers. (F, H) Incubation with Y-27632 (100 mmol/L) induced dramatic changes in villinexpression with loss of the fine brush border pattern and formation of large apical aggregates (arrows). (Panel II) Epithelial cells. Confluent T84intestinal epithelial monolayers were incubated for 1 hour with the ROCK inhibitor Y-27632 (100 mmol/L) or vehicle alone (CON). Cells were fixedand stained with rhodamine phalloidin and analyzed by confocal laser scanning microscopy. Shown here are en face (xy) images taken at the levelof the basal pole of the epithelial monolayer. Incubation with Y-27632 reduced the number of F-actin–rich stress fibers at the basal pole of cells.(Panel III) Ultrastructural examination shows unusual organization of microfilament-rich microvillous core rootlets after ROCK inhibition. ConfluentT84 monolayers, incubated with vehicle alone or with the ROCK inhibitor Y-27632 (100 mmol/L), were examined by transmission electronmicroscopy. (A) Low-power views of control monolayers (CON) showed intact monolayers composed of well-polarized, tall columnar cells. (B) Nochange in cell polarity or length was observed after treatment with Y-27632. (C, E) In control monolayers (CON), F-actin filaments were observedas aggregates in short microvilli and as irregularly dispersed core rootlets (arrow). (D, F) Incubation with Y-27632 induced elongated microvillousF-actin core rootlets that were organized in clusters (arrows) extending downward from the apical region of cells.
570 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3
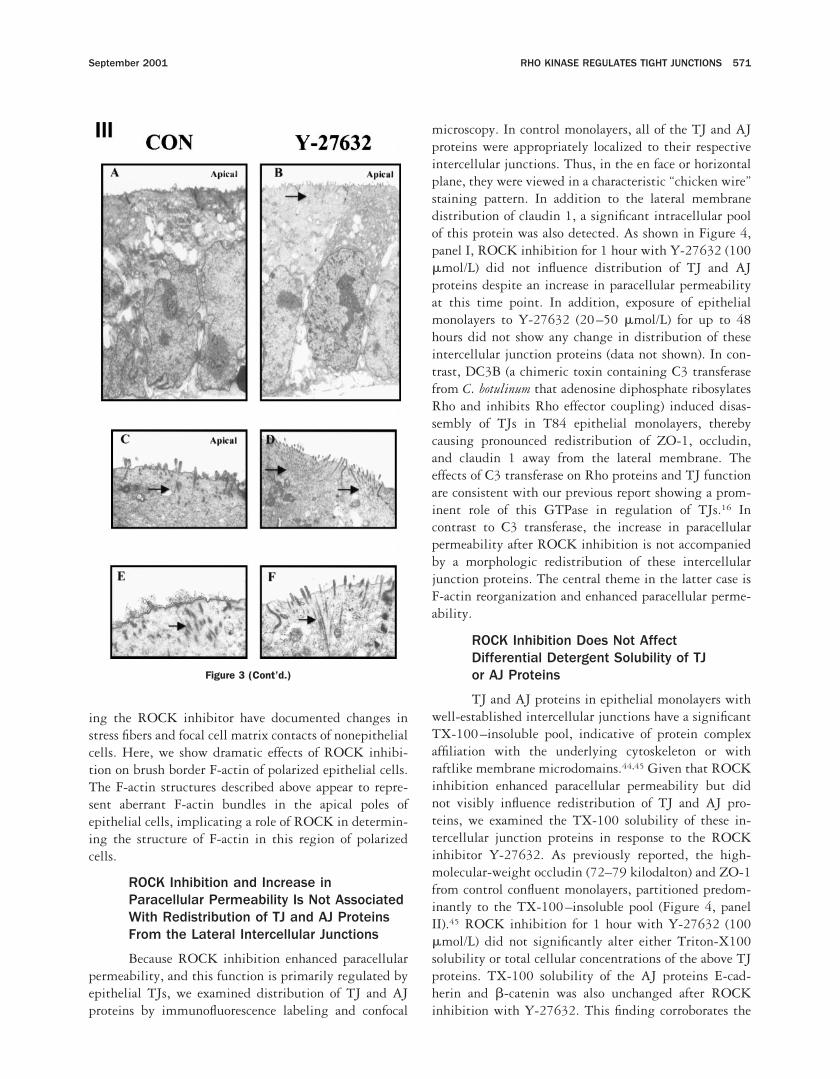
ing the ROCK inhibitor have documented changes instress fibers and focal cell matrix contacts of nonepithelialcells. Here, we show dramatic effects of ROCK inhibi-tion on brush border F-actin of polarized epithelial cells.The F-actin structures described above appear to repre-sent aberrant F-actin bundles in the apical poles ofepithelial cells, implicating a role of ROCK in determin-ing the structure of F-actin in this region of polarizedcells.
ROCK Inhibition and Increase inParacellular Permeability Is Not AssociatedWith Redistribution of TJ and AJ ProteinsFrom the Lateral Intercellular Junctions
Because ROCK inhibition enhanced paracellularpermeability, and this function is primarily regulated byepithelial TJs, we examined distribution of TJ and AJproteins by immunofluorescence labeling and confocal
microscopy. In control monolayers, all of the TJ and AJproteins were appropriately localized to their respectiveintercellular junctions. Thus, in the en face or horizontalplane, they were viewed in a characteristic “chicken wire”staining pattern. In addition to the lateral membranedistribution of claudin 1, a significant intracellular poolof this protein was also detected. As shown in Figure 4,panel I, ROCK inhibition for 1 hour with Y-27632 (100mmol/L) did not influence distribution of TJ and AJproteins despite an increase in paracellular permeabilityat this time point. In addition, exposure of epithelialmonolayers to Y-27632 (20–50 mmol/L) for up to 48hours did not show any change in distribution of theseintercellular junction proteins (data not shown). In con-trast, DC3B (a chimeric toxin containing C3 transferasefrom C. botulinum that adenosine diphosphate ribosylatesRho and inhibits Rho effector coupling) induced disas-sembly of TJs in T84 epithelial monolayers, therebycausing pronounced redistribution of ZO-1, occludin,and claudin 1 away from the lateral membrane. Theeffects of C3 transferase on Rho proteins and TJ functionare consistent with our previous report showing a prom-inent role of this GTPase in regulation of TJs.16 Incontrast to C3 transferase, the increase in paracellularpermeability after ROCK inhibition is not accompaniedby a morphologic redistribution of these intercellularjunction proteins. The central theme in the latter case isF-actin reorganization and enhanced paracellular perme-ability.
ROCK Inhibition Does Not AffectDifferential Detergent Solubility of TJor AJ Proteins
TJ and AJ proteins in epithelial monolayers withwell-established intercellular junctions have a significantTX-100–insoluble pool, indicative of protein complexaffiliation with the underlying cytoskeleton or withraftlike membrane microdomains.44,45 Given that ROCKinhibition enhanced paracellular permeability but didnot visibly influence redistribution of TJ and AJ pro-teins, we examined the TX-100 solubility of these in-tercellular junction proteins in response to the ROCKinhibitor Y-27632. As previously reported, the high-molecular-weight occludin (72–79 kilodalton) and ZO-1from control confluent monolayers, partitioned predom-inantly to the TX-100–insoluble pool (Figure 4, panelII).45 ROCK inhibition for 1 hour with Y-27632 (100mmol/L) did not significantly alter either Triton-X100solubility or total cellular concentrations of the above TJproteins. TX-100 solubility of the AJ proteins E-cad-herin and b-catenin was also unchanged after ROCKinhibition with Y-27632. This finding corroborates the
Figure 3 (Cont’d.)
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 571

Figure 4. (Panel I)
Figure 5. Localization of ROCK in TJs of epithelial monolayers. Confluent T84 epithelial monolayers were immunostained for ROCK (green) andZO-1 (red) and imaged by confocal microscopy. (A–C) En face (xy) confocal images taken in the apical plane of epithelial cells at the level of TJs.Corresponding computer-reconstructed vertical sections in the xz plane are shown in the bottom panels. The supporting filter in these xz imagesis rendered as a white line and the apical and basolateral compartments are annotated (a and bl, respectively). (A) Distribution of ROCK in achicken-wire pattern consistent with its localization in TJs. In addition, the diffuse staining represents an intracytoplasmic pool of ROCK. (B)Distribution of ZO-1 in TJs. (C) Double labeling confirmed colocalization of ZO-1 and ROCK in TJs of epithelial monolayers (yellow).
572 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3

above confocal data documenting normal membrane lo-calization of TJ and AJ proteins in epithelial monolayersdespite enhanced paracellular permeability induced byincubation with Y-27632. The influence of the ROCKinhibitor contrasts with our recent findings with C.difficile toxin showing that inhibition of Rho proteinsenhances paracellular permeability and also induces ashift of TJ proteins (hyperphosphorylated occludin,ZO-1) from a TX-100–insoluble to a TX-100–solublepool (Nusrat et al., Infect Immun, 2001 [in press]).
ROCK Colocalizes With the Epithelial TJProtein ZO-1
Given the profound effect of ROCK inhibitionon TJ function, we localized distribution of ROCK inpolarized epithelial monolayers by immunofluores-cence labeling and confocal microscopy (Figure 5).Confluent epithelial monolayers were double labeled
with antibodies to ROCK and the TJ protein ZO-1.As shown in Figure 5, a major intracellular pool ofROCK was identified within the cytoplasm. In addi-tion, en face images showed a subpool of ROCK thatwas distributed in a chicken wire pattern consistentwith its distribution in TJs of polarized epithelialcells. This pool of ROCK colocalized with ZO-1,further confirming its localization in TJs. These find-ings are further illustrated in reconstructed confocalimages taken in the xz plane.
Transfection With a Dominant NegativeMutant of ROCK Inhibits Formation of thePerijunctional F-Actin Ring in SubconfluentCaco-2 Cells
To confirm the influence of ROCK on apicalF-actin using an independent approach, we transfectedsubconfluent Caco-2 cells with a dominant negative mu-
Figure 4. (Panel II)
Š
Figure 4. (Panel I) ROCK inhibition does not influence distribution of TJ and AJ proteins. Confluent T84 monolayers were incubated with vehiclealone (CON), Y-27632 (100 mmol/L, 1 hour), or the Rho-inactivating toxin, DC3B (3.2 mg/mL basolaterally for 6 hours). The en face (XY) confocalimages shown in this figure were taken at the level of (A–C, F–H, K–M) TJ and (D, E, I, J) AJ. In control monolayers (CON), ZO-1, occludin (OCC),and claudin-1 (CLD-1) were distributed in a chicken-wire pattern consistent with their localization in TJs. A large intracellular pool of CLD-1 wasalso identified. Incubation with Y-27632 did not influence the distribution of TJ proteins. In contrast, Rho inactivation with DC3B toxin completelydisrupted the normal localization of all 3 TJ proteins. In an analogous manner, localization of E-cadherin and b-catenin in AJs was not influencedby incubation with Y-27632. Incubation with Y-27632 (20–50 mg/mL, 1–24 hours) did not change the localization of TJ and AJ proteins. (PanelII) Differential detergent solubility of TJ proteins is not influenced by ROCK inhibition. T84 monolayers were incubated with 100 mmol/L Y-27632for 1 hour or with vehicle alone (CON). Monolayers were subsequently incubated at 4°C with extraction buffer containing 1% TX-100. TheTX-100–soluble and –insoluble fractions were analyzed by SDS-PAGE and immunoblotted for occludin (OCC), ZO-1, claudin-1 (CLD-1), E-cadherin(E-cad), or b-catenin (b-cat). T, S, and I represent proteins in whole-cell lysate, TX-100–soluble, and TX-100–insoluble pools, respectively.High-molecular-weight occludin (72–79 kilodaltons) and ZO-1 were identified predominantly in the TX-100–insoluble pool of cells incubated withvehicle alone. In an analogous manner, a large pool of the AJ proteins E-cadherin and b-catenin were TX-100 insoluble. Incubation with Y-27632(20–50 mmol/L, 1–24 hours) did not change the TX-100 solubility profiles of these TJ and AJ proteins (data not shown).
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 573

tant of ROCK termed ROCK-KDIA. Cells transfectedwith the ROCK mutant (as determined by myc labeling)showed loss of their perijunctional F-actin rings and dis-ruption of the apical brush border compared with surround-ing nontransfected cells (Figure 6). No alteration in apicalF-actin organization was detected in Caco-2 cells transfectedwith an empty vector–only construct. These results furtherconfirm the influence of ROCK on apical F-actin organiza-tion. In cells transfected with the dominant negativeROCK, the myc-tagged mutant localized to the apicallateral junctional region, where it colocalized with occludin.
No loss of occludin staining was detected in transfectedcells, further confirming the inhibitor studies.
ROCK Influences Assembly of TJs inIntestinal Epithelial Cells
Because ROCK inhibition enhanced paracellularpermeability in confluent epithelial monolayers, we de-termined the role of ROCK in assembly of intercellularjunctions. Thus, we examined the influence of Y-27632on transepithelial resistance recovery after reversible dis-ruption of TJs and AJs by calcium depletion. Such
Figure 7. (Panel I)
‹
Figure 7. (Panel I) ROCK is necessary for the recovery of monolayer barrier function after calcium depletion. Confluent epithelial monolayerswere incubated with HBSS2 and EGTA to induce TJ disassembly. Monolayers were allowed to recover in calcium-containing medium in thepresence and absence of Y-27632 (20 mmol/L). Monolayer TER was followed up for 24 hours after calcium repletion. TER of control monolayersrecovered to .1000 V z cm2 within 24 hours. In contrast, in the presence of Y-27632, monolayers failed to recover TER. (Panel II) ROCK isnecessary for the reassembly of TJs, AJs, and the apical actin cytoskeleton in intestinal epithelial cells after calcium depletion. Confluentepithelial monolayers were incubated with HBSS2 and EGTA to induce TJ disassembly. Monolayers were allowed to recover in calcium-containingmedium in the presence and absence of Y-27632 (20 mmol/L). ZO-1, occludin (OCC), E-cadherin (Ecad), and F-actin were immunolocalized byconfocal microscopy before calcium depletion (Cont 0 h), after calcium depletion (EGTA), after 24 hours of recovery in medium alone (Cont,24 h), or after 24 hours in medium containing 20 mmol/L Y-27632 (Y-27632/24 h). The en face (xy) confocal images shown in this figure weretaken in the apical plane of epithelial cells at the level of TJ and AJ. (A, E, M) In control monolayers (CONT 0 h), ZO-1, occludin, and E-cadherinare distributed in the normal chicken-wire pattern, consistent with their localization in TJ and AJ, respectively. (I ) Rhodamine phalloidin labelingof F-actin in the apical plane showed the expected dotlike pattern of microvillous F-actin and its organization in peri-junctional rings. (B, F, J, N)After calcium depletion (EGTA), junctional proteins (occ, ZO-1, E-cad) and F-actin were redistributed to a perinuclear compartment. When allowedto recover in media alone (Control 24 h), (C, G, O) junctional proteins relocalized to the apical aspect of the lateral membrane within 24 hoursand (K) the F-actin staining pattern returned to normal. (D, H, P) In contrast, ZO-1, occludin, and E-cadherin failed to redistribute in theircorresponding intercellular junctions in the presence of Y-27632. ROCK inhibition also prevented the recovery of a normal F-actin stainingpattern. Results shown are representative of 3 independent experiments.
‹
Figure 6. Transfection with a dominant negative mutant of ROCK inhibits formation of the perijunctional F-actin ring and apical actin-richstructures in subconfluent Caco-2 cells. Caco-2 cells were transfected with a dominant negative mutant of ROCK termed ROCK-KDIA and stainedfor cMyc and F-actin or occludin. Colonies of Caco-2 cells showed focal loss of apical F-actin structures (*) including the perijunctional actin ring(arrows) and apical brush border (arrows). (A, C) When examined for expression of the Myc-tagged dominant negative mutant of ROCK (green;arrows), the cells lacking apical F-actin structures strongly expressed the dominant negative mutant ROCK protein. (B, D) Neighboringnontransfected cells clearly maintained apical actin structures serving as excellent internal controls. (C) and (D) show higher-power views ofanother colony. No loss of occludin staining was detected in transfected cells double stained for occludin and Myc. (E) Occludin only (arrows);(F ) occludin and Myc colocalization (arrows).
574 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3

Figure 6.
Figure 7. (Panel II)
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 575

experiments were based on previous studies showing arequirement for calcium in the maintenance of TJ integ-rity.46,47 As shown in Figure 7, panel I, incubation ofT84 monolayers with EGTA resulted in a dramaticdecrease in TER from 1155 6 160 V z cm2 to 159 6 62V z cm2, indicating TJ disruption. Monolayer TER wasthen allowed to recover in the presence of calcium-containing media with or without Y-27632 (20 mmol/L). As shown in Figure 7, panel I, control T84 mono-layers recovered high TER values (.900 V z cm2) within8 hours, whereas ROCK inhibition completely abrogatedmonolayer TER recovery. To further investigate the in-hibitory effect of Y-27632 on TER recovery, we analyzedorganization of the F-actin cytoskeleton and distributionof TJ/AJ proteins under these conditions by immunoflu-orescence labeling and confocal microscopy (Figure 7,panel II). Compared with the normal chicken-wire stain-ing pattern typical of junctional localization, calciumdepletion with EGTA resulted in displacement of ZO-1,occludin, and E-cadherin from lateral intercellular junc-tions to a central perinuclear zone. These findings indi-cated dismantling of TJ and AJ structures and correlatedwith the observed decrease in TER. During the EGTA-induced decrease in TER, the F-actin cytoskeleton wasmarkedly changed. Clumped F-actin filaments were ob-served in a central perinuclear distribution, as previouslyreported.48 Subsequent calcium repletion in controlmonolayers induced redistribution of ZO-1, occludinand E-cadherin to the apical region of the lateral mem-brane. These changes were accompanied by restoration ofa normal apical F-actin staining pattern and recovery ofTER to baseline values. However, in the presence ofY-27632, monolayers were unable to reassemble normalintercellular associations after repletion of extracellularcalcium. Under these conditions, TJ and AJ proteinsfailed to relocalize to their normal distribution in inter-cellular junctions. ROCK inhibition also prevented nor-mal reassembly of the F-actin cytoskeleton, with mono-layers failing to regenerate perijunctional F-actin rings ormicrovillous F-actin cores. These findings suggest a cen-tral role of ROCK in assembly of apical F-actin struc-tures and TJ/AJs in epithelial cells.
DiscussionPrecise regulation of intestinal epithelial TJs is
crucial to maintaining barrier function between the lu-menal milieu and the internal environment. In this re-gard, the small GTPase Rho is known to be an importantregulator of TJ function.16,20,49 Mechanisms by whichRho proteins regulate TJ function are still not com-pletely defined. In particular, whether downstream effec-
tors of Rho are important in this process is not known.ROCK, a 160-kilodalton serine/threonine kinase alsoknown as Rho-associated coiled-coil forming protein ki-nase, has been identified as one of the downstream effec-tors of Rho. In our study, we have examined the role ofthis molecule in the regulation of TJ structure andfunction.
Using a specific inhibitor of ROCK, Y-27632, and adominant negative transfection approach, we show a roleof ROCK in regulation of paracellular permeability.Although the reduction in TER was striking, inhibitionof ROCK did not completely abolish monolayer TER.Such findings would implicate complex mechanisms inregulation of TJ function and would suggest, at least inpart, regulation by ROCK. In contrast to these findings,specific inhibition of its upstream activator Rho is asso-ciated with a dramatic abolition of monolayer TER (,50V z cm2), suggesting that ROCK inhibition accounts foronly a fraction of the enhanced paracellular permeabilityinduced by Rho.
Because TJs are key structural components that regu-late epithelial paracellular permeability, and this func-tion is influenced by ROCK inhibition, we analyzedmechanisms by which ROCK may regulate such prop-erties in epithelial cells. It has previously been docu-mented that the apical perijunctional F-actin ring playsa central role in regulation of TJ function.9 F-actinfilaments in this ring are themselves intimately associ-ated with the actin filaments of the terminal web intowhich the F-actin–rich microvillous core rootlets arelinked in a myosin-dependent manner.50–52 In our study,enhanced permeability induced by inhibition of ROCKfunction was paralleled by profound reorganization of theapical F-actin cytoskeleton. We observed a loss of thenormal fine brush border F-actin pattern and a reductionin the intensity of the perijunctional F-actin rings. Weobserved formation of dense apical condensations of F-actin and the actin-regulating protein villin. Ultrastruc-tural analysis suggested that these aggregates mightrepresent abnormal microvillous core rootlets. These ob-servations were further supported by our transfectionstudies, in which a dominant negative mutant of ROCKinduced loss of the apical F-actin–rich brush border anda reduction in the apical perijunctional F-actin ringwithout diminishing occludin staining. ROCK colocal-ized with the TJ protein ZO-1 in polarized T84 cells.This finding was also corroborated in the transfectionstudies in which a dominant negative, kinase-deficient,and Rho binding–deficient mutant of ROCK also local-ized to the apical junctional region in Caco-2 cells, whereit colocalized with occludin. Therefore, it may be sur-
576 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3

mised that ROCK is pivotal in the maintenance ofnormal apical actin dynamics in epithelial cells. Thisstudy supports the hypothesis that ROCK may influenceepithelial TJ function.
A previous study in low-TER renal epithelial cells53
using a dominant negative inhibitor of ROCK also foundloss of endogenous stress fibers and peripheral actin.However, this study did not address the effects of ROCKon apical F-actin organization. In a different study, RhoAand ROCK have been shown to regulate NHE3, anexchanger that is physiologically located in the apicalmembrane of intestinal epithelia.54 This study lendscredence to the possibility that ROCK may influenceother apical events via its actions on the F-actin cytoskel-eton. However, these experiments were performed innonpolarized AP1 Chinese hamster ovary cells and notpolarized epithelia with intact TJs.
In spite of Y-27632–induced changes in paracellularpermeability, analysis of TJ proteins failed to show re-distribution or disassembly of the TJ protein complex byeither morphologic or biochemical parameters. In con-trast, specific inhibition of Rho with DC3B is associatedwith complete disassembly of the TJ protein complex.These findings suggest that ROCK inhibition alone can-not account for the structural disassembly induced byinhibition of Rho. Because the actin cytoskeleton plays avital role in regulation of TJ function and is dramaticallyreorganized by ROCK inhibition, it can be surmised thatthe increase in paracellular permeability is most likelysecondary to cytoskeletal restructuring in the apical polesof epithelial cells.
Further support for this hypothesis comes from arecent study of cultured endothelial cells. In this study,Hirase et al.55 show that regulation of endothelial TJpermeability and occludin phosphorylation may be in-fluenced by p160ROCK function. Inhibition of ROCKwith Y-27632 prevented increases in paracellular perme-ability and occludin phosphorylation induced by LPAbut not by histamine. However, in untreated controlendothelial cells, inhibition of ROCK function inducedan increase in paracellular permeability that was associ-ated with a subtle reorganization of pericellular actinbundles but no alteration in occludin phosphorylation.These findings in unstimulated endothelial cells are anal-ogous to the findings in our study of similarly unstimu-lated epithelial cells.
Experiments focused on TJ assembly revealed acentral role of ROCK in determining assembly andaffiliation of the TJ protein complex with this inter-cellular junction. ROCK inhibition was associatedwith failure of epithelial monolayers to assemble api-
cal junctional complexes and a lack of recovery ofparacellular barrier function. These findings were as-sociated with failure to recover the normal apicalF-actin cytoskeletal architecture. Failure of monolay-ers to assemble the apical junctional complex, in abackground of a disorganized actin cytoskeleton afterROCK inhibition, could be related to the requirementof normal apical actin dynamics for assembly of theseintercellular associations. Thus, inhibition of ROCKand the consequent apical F-actin disorganizationcould influence targeting of TJ and AJ proteins to thelateral membrane. Alternatively, a direct effect ofROCK on AJ or TJ proteins cannot be excluded.However, it can be surmised that our findings support,for the first time, an essential role of ROCK in theformation of both the F-actin cytoskeleton and junc-tional complexes in the apical poles of polarizingepithelial cells.
References1. Farquhar MG, Palade GE. Junctional complexes in various epithe-
lia. J Cell Biol 1963;17:375–412.2. Staehelin LA. Further observations on the fine structure of freeze-
cleaved tight junctions. J Cell Sci 1973;13:763–786.3. Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita S.
Occludin: a novel integral membrane protein localizing at tightjunctions. J Cell Biol 1993;123:1777–1788.
4. Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and-2: novel integral membrane proteins localizing at tight junctionswith no sequence similarity to occludin. J Cell Biol 1998;141:1539–1550.
5. Fanning AS, Mitic LL, Anderson JM. Transmembrane proteins inthe tight junction barrier. J Am Soc Nephrol 1999;10:1337–1345.
6. Tsukita S, Furuse M. Pores in the wall: claudins constitute tightjunction strands containing aqueous pores. J Cell Biol 2000;149:13–16.
7. Liu Y, Nusrat A, Schnell FJ, Reaves TA, Walsh S, Pochet M,Parkos CA. Human junction adhesion molecule regulates tightjunction resealing in epithelia. J Cell Sci 2000;113:2363–2374.
8. Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. The tightjunction protein ZO-1 establishes a link between the transmem-brane protein occludin and the actin cytoskeleton. J Biol Chem1998;273:29745–29753.
9. Madara JL. Intestinal absorptive cell tight junctions are linked tocytoskeleton. Am J Physiol 1987;253:C171–C175.
10. Madara JL, Barenberg D, Carlson S. Effects of cytochalasin D onoccluding junctions of intestinal absorptive cells: further evi-dence that the cytoskeleton may influence paracellular perme-ability and junctional charge selectivity. J Cell Biol 1986;102:2125–2136.
11. Turner JR, Rill BK, Carlson SL, Carnes D, Kerner R, Mrsny RJ,Madara JL. Physiological regulation of epithelial tight junctions isassociated with myosin light-chain phosphorylation. Am J Physiol1997;273:C1378–C1385.
12. Madara JL, Pappenheimer JR. Structural basis for physiologicalregulation of paracellular pathways in intestinal epithelia. JMembr Biol 1987;100:149–164.
13. Moore R, Pothoulakis C, LaMont JT, Carlson S, Madara JL. C.difficile toxin A increases intestinal permeability and inducesCl-secretion. Am J Physiol 1990;259:G165–G172.
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 577

14. Walsh SV, Hopkins AM, Nusrat A. Modulation of tight junctionstructure and function by cytokines. Adv Drug Deliv Rev 2000;41:303–313.
15. Atisook K, Carlson S, Madara JL. Effects of phlorizin and sodiumon glucose-elicited alterations of cell junctions in intestinal epi-thelia. Am J Physiol Cell Physiol 1990;258:C77–C85.
16. Nusrat A, Giry M, Turner JR, Colgan SP, Parkos CA, Carnes D,Lemichez E, Boquet P, Madara JL. Rho protein regulates tightjunctions and perijunctional actin organization in polarized epi-thelia. Proc Natl Acad Sci USA 1995;92:10629–10633.
17. Braga VM, Machesky LM, Hall A, Hotchin NA. The small GTPasesRho and Rac are required for the establishment of cadherin-dependent cell-cell contacts. J Cell Biol 1997;137:1421–1431.
18. Weber E, Berta G, Tousson A, St. John P, Green MW, Gopalokrish-nan U, Jilling T, Sorscher EJ, Elton TS, Abrahamson DR. Expres-sion and polarized targeting of a rab3 isoform in epithelial cells.J Cell Biol 1994;125:583–594.
19. Hopkins AM, Li D, Mrsny RJ, Walsh SV, Nusrat A. Modulation oftight junction function by G protein-coupled events. Adv Drug DelivRev 2000;41:329–340.
20. Jou TS, Schneeberger EE, Nelson WJ. Structural and functionalregulation of tight junctions by RhoA and Rac1 small GTPases.J Cell Biol 1998;142:101–115.
21. Jou TS, Nelson WJ. Effects of regulated expression of mutantRhoA and Rac1 small GTPases on the development of epithelial(MDCK) cell polarity. J Cell Biol 1998;142:85–100.
22. Leung T, Manser E, Tan L, Lim L. A novel serine/threonine kinasebinding the ras-related RhoA GTPase which translocates the ki-nase to peripheral membranes. J Biol Chem 1995;270:29051–29054.
23. Leung T, Chen XQ, Manser E, Lim L. The p160 RhoA-bindingkinase ROKalpha is a member of a kinase family and is involvedin the reorganization of the cytoskeleton. Mol Cell Biol 1996;16:5313–5327.
24. Ishizaki T, Maekawa M, Fujisawa K, Okawa K, Iwamatsu A, FujitaA, Watanabe N, Saito Y, Kakizuka A, Morii N, Narumiya S. Thesmall GTP-binding protein Rho binds to and activates a 160 kDaSer/Thr protein kinase homologous to myotonic dystrophy ki-nase. EMBO J 1885;15:1885–1893.
25. Matsui T, Amano M, Yamamoto T, Chihara K, Nakafuku M, Ito M,Nakano T, Okawa K, Iwamatsu A, Kaibuchi K. Rho-associatedkinase, a novel serine/threonine kinase, as a putative target forthe small GTP binding protein Rho. EMBO J 1996;15:2208–2216.
26. Aspenstrom P. Effecters for the Rho GTPases. Curr Opin Cell Biol1999;11:95–102.
27. Watanabe N, Kato T, Fujita A, Ishizaki T, Narumiya S. Cooperationbetween mDia1 and ROCK in Rho-induced actin reorganization.Nat Cell Biol 1999;1:136–143.
28. Bito H, Furuyashiki T, Ishihara H, et al. A critical role for aRho-associated kinase, p160ROCK, in determining axon out-growth in mammalian CNS neurons. Neuron 2000;26:431–441.
29. Itoh K, Yoshioka K, Akedo H, Uehata M, Ishizaki T, Narumiya S.An essential part for Rho-associated kinase in the transcellularinvasion of tumor cells. Nat Med 1999;5:221–225.
30. Fujita A, Saito Y, Ishizaki T, Maekawa M, Fujisawa K, Ushikubi F,Narumiya S. Integrin-dependent translocation of p160ROCK tocytoskeletal complex in thrombin-stimulated human platelets.Biochem J 1997;328:769–775.
31. Kimura K, Ito M, Amano M, Chihara K, Fukata Y, Nakafuku M,Yamamori B, Feng J, Nakano T, Okawa K, Iwamatsu A, KaibuchiK. Regulation of myosin phosphatase by Rho and Rho-associatedkinase (Rho-kinase). Science 1996;273:245–248.
32. Nakai K, Suzuki Y, Kihira H, Wada H, Fujioka M, Ito M, Nakano T,Kaibuchi K, Shiku H, Nishikawa M. Regulation of myosin phos-phatase through phosphorylation of the myosin-binding subunit inplatelet activation. Blood 1997;90:3936–3942.
33. Suzuki Y, Yamamoto M, Wada H, Ito M, Nakano T, Sasaki Y,Narumiya S, Shiku H, Nishikawa M. Agonist-induced regulation ofmyosin phosphatase activity in human platelets through activa-tion of Rho-kinase. Blood 1999;93:3408–3417.
34. Amano M, Ito M, Kimura K, Fukata Y, Chihara K, Nakano T,Matsuura Y, Kaibuchi K. Phosphorylation and activation of myo-sin by Rho-associated kinase (Rho-kinase). J Biol Chem 1996;271:20246–20249.
35. Fukata Y, Kimura K, Oshiro N, Saya H, Matsuura Y, Kaibuchi K.Association of the myosin-binding subunit of myosin phospha-tase and moesin: Dual regulation of moesin phosphorylation byRho-associated kinase and myosin phosphatase. J Cell Biol1998;141:409–418.
36. Uehata M, Ishizaki T, Satoh H, Ono T, Kawahara T, Morishita T,Tamakawa H, Yamagami K, Inui J, Maekawa M, Narumiya S.Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature 1997;389:990–994.
37. Ishizaki T, Uehata M, Tamechika I, Keel J, Nonomura K, MaekawaM, Narumiya S. Pharmacological properties of Y-27632, a spe-cific inhibitor of Rho-associated kinases. Mol Pharmacol 2000;57:976–983.
38. Narumiya S, Ishizaki T, Uehata M. Use and properties of ROCK-specific inhibitor Y-27632. Methods Enzymol 2000;325:273–284.
39. Parkos CA, Colgan SP, Bacarra AE, Nusrat A, Delp-Archer C,Carlson S, Su DH, Madara JL. Intestinal epithelia (T84) possessbasolateral ligands for CD11b/CD18-mediated neutrophil adher-ence. Am J Physiol 1995;268:C472–C479.
40. Nusrat A, Parkos CA, Bacarra AE, Godowski PJ, Delp-Archer C,Rosen EM, Madara JL. Hepatocyte growth factor/scatter factoreffects on epithelia. Regulation of intercellular junctions in trans-formed and nontransformed cell lines, basolateral polarization ofc-met receptor in transformed and natural intestinal epithelia,and induction of rapid wound repair in a transformed modelepithelium. J Clin Invest 1994;93:2056–2065.
41. Aullo P, Giry M, Olsnes S, Popoff MR, Kocks C, Boquet P. Achimeric toxin to study the role of the 21 kDa GTP binding proteinrho in the control of actin microfilament assembly. EMBO J 1993;12:921–931.
42. Sanders SE, Madara JL, McGuirk DK, Gelman DS, Colgan SP.Assessment of inflammatory events in epithelial permeability: arapid screening method using fluorescein dextrans. Epith CellBiol 1995;4:25–34.
43. Ishizaki T, Naito M, Fujisawa K, Maekawa M, Watanabe N, SaitoY, Narumiya S. p160rock, a Rho-associated coiled-coil formingprotein kinase, works downstream of Rho and induces focaladhesions. FEBS Lett 1997;404:118–124.
44. Sakakibara A, Furuse M, Saitou M, Ando-Akatsuka Y, Tsukita S.Possible involvement of phosphorylation of occludin in tight junc-tion formation. J Cell Biol 1997;137:1393–1401.
45. Nusrat A, Parkos CA, Verkade P, Foley CS, Liang TW, Innis-Whitehouse W, Eastburn KK, Madara JL. Tight junctions aremembrane microdomains. J Cell Sci 2000;113:1771–1781.
46. Martinez-Palomo A, Meza I, Beaty G, Cereijido M. Experimentalmodulation of occluding junctions in a cultured transporting epi-thelium. J Cell Biol 1980;87:736–745.
47. Balda MS, Gonzalez-Mariscal L, Contreras RG, Macias-Silva M,Torres-Marquez ME, Garcia-Sainz JA, Cereijido M. Assembly andsealing of tight junctions: possible participation of G-proteins,phospholipase C, protein kinase C and calmodulin. J Membr Biol1991;122:193–202.
48. Volberg T, Geiger B, Kartenbeck J, Franke WW. Changes in mem-brane-microfilament interaction in intercellular adherens junc-tions upon removal of extracellular Ca21 ions. J Cell Biol 1986;102:1832–1842.
49. Gopalakrishnan S, Raman N, Atkinson SJ, Marrs JA. Rho GTPase
578 WALSH ET AL. GASTROENTEROLOGY Vol. 121, No. 3

signaling regulates tight junction assembly and protects tightjunctions during ATP depletion. Am J Physiol 1998;275:C798–C809.
50. Temm-Grove C, Helbing D, Wiegand C, Honer B, Jockusch BM.The upright position of brush border-type microvilli depends onmyosin filaments. J Cell Sci 1992;101:599–610.
51. Matsudaira PT, Burgess DR. Structure and function of the brush-border cytoskeleton. Cold Spring Harbor Symp Quant Biol 1982;46:845–854.
52. Hull BE, Staehelin LA. The terminal web. A reevaluation of itsstructure and function. J Cell Biol 1979;81:67–82.
53. Royal I, Lamarche-Vane N, Lamorte L, Kaibuchi K, Park M.Activation of cdc42, rac, PAK, and rho-kinase in response tohepatocyte growth factor differentially regulates epithelial cellcolony spreading and dissociation. Mol Biol Cell 2000;11:1709–1725.
54. Szaszi K, Kurashima K, Kapus A, Paulsen A, Kaibuchi K, Grinstein S,Orlowski J. RhoA and rho kinase regulate the epithelial Na1/H1exchanger NHE3. Role of myosin light chain phosphorylation. J BiolChem 2000;275:28599–28606.
55. Hirase T, Kawashima S, Wong EY, Ueyama T, Rikitake Y, TsukitaS, Yokoyama M, Staddon JM. Regulation of tight junction perme-ability and occludin phosphorylation by RhoA-p160ROCK-depen-dent and -independent mechanisms. J Biol Chem 2001:10423–10431.
Received February 9, 2001. Accepted May 23, 2001.Address requests for reprints to: Asma Nusrat, M.D., Department of
Pathology, Emory University, Room 2335, Woodruff Memorial Re-search Building, 1639 Pierce Drive, Atlanta, Georgia 30322. e-mail:[email protected]; fax: (404) 727-8538.
Supported by grants from the National Institutes of HealthDK 55679, HL 54229, and HL 60540 and by the American Diges-tive Health Foundation Fellowship/Faculty Transition Award (toS.V.W.).
The Rho kinase inhibitor was a kind gift of the Welfide Corporation,Japan. The DC3B toxin was a kind gift of Dr. Patrice Boquet, INSERM,Nice, France. The authors acknowledge the expert technical assis-tance of Christie Vu.
September 2001 RHO KINASE REGULATES TIGHT JUNCTIONS 579




















