
New Bathonian (Middle Jurassic) ammonite assemblages from Kutch, India
24
This article was originally published in a journal published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues that you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial
Transcript of New Bathonian (Middle Jurassic) ammonite assemblages from Kutch, India

This article was originally published in a journal published byElsevier, and the attached copy is provided by Elsevier for the
author’s benefit and for the benefit of the author’s institution, fornon-commercial research and educational use including without
limitation use in instruction at your institution, sending it to specificcolleagues that you know, and providing a copy to your institution’s
administrator.
All other uses, reproduction and distribution, including withoutlimitation commercial reprints, selling or licensing copies or access,
or posting on open internet sites, your personal or institution’swebsite or repository, are prohibited. For exceptions, permission
may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial

Autho
r's
pers
onal
co
py
New Bathonian (Middle Jurassic) ammonite assemblagesfrom Kutch, India
Pinaki Roy, Subhendu Bardhan *, Aparajita Mitra, Sudipta K. Jana
Department of Geological Sciences, Jadavpur University, Kolkata 32, India
Received 1 July 2005; received in revised form 9 October 2006; accepted 5 February 2007
Abstract
The Bathonian ammonite assemblages have been previously poorly recorded in Kutch. The present study has unearthed a rich arrayof ammonite taxa ranging from the Middle to Upper Bathonian. While Oxycerites Rollier (1909) is a new record from Kutch, the oldestoccurrence of Choffatia Siemiradzki (1898) has been found from the Middle Bathonian horizon. Oxycerites cf. orbis (Giebel) is a zonalindex of the Late Bathonian in other areas and thus facilitates interprovincial correlation. Besides, palaeobiogeographic and stratigraphicdistribution of many species have been modified based on new information. For example, macrocephalitin species, i.e., Kamptokephalites
cf. etheridgei Spath (1928), Macrocephalites bifurcatus transient intermedius Spath (1928), M. cf. mantataranus Boehm (1912) were pre-viously known from West Pacific, Indonesia have been now found in Kutch. Gracilisphinctes Buckman (1920) has been previously knownto occur during the Middle Bathonian time, the present work extends its stratigraphic distribution up to the definite Upper Bathonianhorizon. Procerites hians (Waagen) an endemic species in Kutch previously known from the Upper Bathonian beds, its stratigraphicrange has been extended down to the Middle Bathonian.
Detail taxonomy of the newly obtained taxa has been done and in many cases sexual dimorphism has been recognized.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Ammonites; Bathonian; Biostratigraphy; Kutch; Palaeobiogeography
1. Introduction
Kutch in western India is famous over the world for itsrich assemblages of the Callovian–Oxfordian ammonites.Bathonian ammonites were sporadically described mainlyfrom the island areas of Kutch (Singh et al., 1983; Jaitlyand Singh, 1984; Pandey and Agrawal, 1984 Pandeyet al., 1984; Pandey and Westermann, 1988; Pandey andCallomon, 1995). Recent works of Callomon (1993) andKayal and Bardhan (1998) have also convincingly demon-strated the presence of the Late Bathonian ammonite in themainland of Kutch. The present paper describes 10 speciesbelonging to eight genera from the Bathonian sequences inthe mainland of Kutch. They are: Procerites hians
(Waagen, 1875), Sivajiceras congener (Waagen, 1875),
Gracilisphinctes cf. arkelli Collignon (1958), Choffatia sp.,Macrocephalites cf. mantataranus Boehm (1912), M. cf.bifurcatus transient intermedius (Spath, 1928), M. triangularis
Spath (1928), M. cf. etheridgei Spath (1928), Prohecticoceras
manjalense n. sp. and Oxycerites cf. orbis (Giebel). Theammonite assemblages ranging in age from the lower MiddleBathonian to uppermost Late Bathonian. While one speciesis new, some are new record from India. The present studyalso revises the stratigraphic distribution of previouslydescribed Bathonian species from other areas of Kutch.Macrocephalitin genera once believed to be of Early Callo-vian age, have been shown here to range from the MiddleBathonian. Similar Middle Bathonian Macrocephalites
spp. were previously reported from West Pacific, Indonesia(Westermann and Callomon, 1988).
It appears that Bathonian ammonite assemblagesare diverse; they include many time-diagnostic taxa,which help in inter-provincial biostratigraphic correlation.
1367-9120/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jseaes.2007.02.003
* Corresponding author.E-mail address: [email protected] (S. Bardhan).
www.elsevier.com/locate/jaes
Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
Palaeobiogeographic analysis shows immigration of faunasfrom different provinces took place immediately after theopening of the Kutch basin.
2. Stratigraphy
The Jurassic sediments (Bathonian–Tithonian) cover avast area in Kutch including mainland, island and easternKutch (Fig. 1). The Bathonian beds, however, are notexposed in all areas. In mainland the well studied sectionsin Jumara is famous for its Upper Bathonian–Oxfordianammonite assemblages (Datta, 1992). The present paperdescribes Bathonian genera from Jumara as well as twoother areas such as Nara and Jhura within mainland. InJumara, only the Late Bathonian assemblage is foundwhere Nara and Jhura include taxa ranging from the Mid-dle Bathonian time. In Nara, because of the poor exposurecondition for recent erosion and past volcanic activity theUpper Bathonian faunas cannot be found, but Jhura repre-sents a fuller section spanning the Middle to UpperBathonian.
Lithostratigraphic as well as sedimentological works arenow available for all these classical sections of mainland,which facilitates the precise assignation of the newlydescribed present ammonite genera and helps in regionalbiostratigraphic correlation. Bathonian sediments in Kutchare grouped into the Patcham Formation in mainland andin island, its equivalence is the Goradongar Formation(Fursich et al., 1994; Jain et al., 1996). The depositionalenvironment was a carbonate ramp on the epicontinentalshelf and the habitat was widespread throughout the Kutchsea. Limestone and marl are the basic motif of sedimenta-tion intercalated often by oolitic facies (see Fursich andOschmann, 1993).
The Bathonian sediments in Jumara are dominated bythe marl-lime association and are grouped into four beds(1–4 of Bardhan et al., 1994; also see present Fig. 2). These
beds are mainly grey, shelly limestone (packstone/grain-stone) intercalated with greenish to grey shale and whitelimestone (mudstone/wackestone) of varying thickness.Jumara is famous for its Bathonian ammonite assemblages
Fig. 1. Geological map of Kutch, showing the locality of Jumara, Nara and Jhura from where the present collection has been made.
Siva
jice
ras
cong
ener
1
2
3
4
Zone
Upp
er
Bat
honi
an
Tri
angu
lari
sC
hrys
oolit
hicu
s
Mac
roce
phal
ites
tria
ngul
aris
Epi
stre
noce
ras
sp.
Pro
ceri
tes
hian
s
Gra
cili
sphi
ncte
scf
.ark
elli
M.c
f.m
anta
tara
nus
M. c
f.bi
furc
atus
tran
sien
tin
term
ediu
s
Limestone
Golden ooliticlimestone
Coral biostrome
White limestone
Oolitic limestone
Shale
Basic plutons
Bed
No.
Khe
raic
eras
cf.
hann
over
anum
Oxy
ceri
tes
cf.o
rbis
M. m
adag
asca
rien
sis
M. c
hrys
ooli
thic
us
Fig. 2. Stratigraphical column at the Jumara section, illustrating lithos-tratigraphy, biostratigraphy and vertical distribution of importantammonite taxa (not to the scale).
630 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
since the times of Waagen (1875) and Spath (1927–1933).Many recent reports and our additional collection ofnumerous specimens have helped refine and revise thestratigraphic distribution of ammonite species, which isshown in Fig. 2.
Nara is situated 10 km southeast of Jumara and yieldsthe Jurassic beds. The area is extremely disturbed by theDeccan Trap volcanic activity. Previous studies report onlyammonites of Chari Formation (e.g., Spath, 1927–1933 andGhose, 1967). However our recent several field investiga-tions record a thick white limestone-marl alternation whichcontains abundant specimens of oppelid genus Prohecticoc-
eras manjalense n. sp., Procerites hians, Choffatia sp. andKamptokephalites cf. etheridgei (see Fig. 3). The later speciescomes evidently from the Middle Bathonian of Sula Island,Indonesia (Westermann and Callomon, 1988).
Jhura, situated 50 km southeast of Jumara is anotherlocality in mainland where the greater part of the Batho-nian Patcham Formation is exposed (Fig. 4). The oldestbed exposed is a white limestone with shale and grayish yel-low slaby limestone alternations. No ammonite is found sofar from this horizon, only sparse brachiopod taxa havebeen reported (Mukherjee, 2002; Mukherjee et al., 2003).The overlying unit is a massive golden oolite and shalealternation (Bed 2, Fig. 4). K. cf. etheridgei has beendescribed here from this horizon. The next unit is hardslabby grayish yellow limestone with thinner and distantgolden oolite bands and shale alternations (Bed 3,Fig. 4). Jain et al. (1996) have mentioned Gracilisphinctes
sp. from this horizon and Mukherjee et al. (2003) consid-
ered it to be of the Middle Bathonian. Upper part of thisunit (Bed 3) is dominantly made up of grayish yellow lime-stone and marl alternation which has yielded several spec-imens of M. triangularis.
3. Biostratigraphy
The Middle Bathonian ammonite records in Kutch wereso far not well documented. Because of dearth of well time-diagnostic cosmopolitan taxa, all previous attempts torecognize Bathonian assemblage in Kutch were at mosttentative. However, Bardhan et al. (2002) proposed a bio-stratigraphic zonation for the Bathonian and Callovianof Kutch and attempted inter-provincial correlation withother areas. It took a long time by the palaeontologiststo realize that Macrocephalites once was thought to heraldthe Callovian time, already appeared in many provinceswell within the Bathonian (see Callomon, 1993). Evenwhen workers conceived that some older species of
2
1
Zone
Mid
dle
Bat
honi
an
K.c
f.et
heri
dgei
Cho
ffati
asp
.
Pro
hect
icoc
era
sm
anja
lens
en.
sp.
Bed
No.
Eth
erid
geiG
raci
lisp
hinc
tes
cf.a
rkel
li
Pro
ceri
tes
hian
s
Fig. 3. Stratigraphical column at the Nara section, illustrating lithostra-tigraphy, biostratigraphy and vertical distribution of important ammonitetaxa (not to the scale). For legends see Fig. 2.
Bed
No.
4
3
2
1
Zone
Upp
er B
atho
nian
Mid
dle
Bat
honi
an
Mac
roce
phal
ites
tria
ngul
aris
K.c
f.et
heri
dgei
Tri
angu
lari
sC
hrys
oolit
hicu
sE
ther
idge
i
Fig. 4. Stratigraphical column at the Jhura section, illustrating lithostra-tigraphy, biostratigraphy and vertical distribution of important ammonitetaxa (not to the scale) (modified after Jana et al., 2005). For legends seeFig. 2.
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 631

Autho
r's
pers
onal
co
py
Macrocephalites may belong to the Bathonian (Krishnaand Westermann, 1985, 1987; Datta, 1992; Datta et al.,1996), they have confronted with the problem of endemic-ity of Macrocephalites at species level. For example,M. triangularis, which has the Bathonian affinity, isrestricted only to the Indo-Madagascan Faunal Province.In Kutch Epistrenoceras Bentz (1928) co-occurs withM. triangularis (Kayal and Bardhan, 1998). Epistrenoceras
is widely used as an invaluable marker for correlation(Westermann and Callomon, 1988). In Europe, Epistrenoc-
eras comes from the Retrocostatum Zone or the coevalAspidoides Zone (Torrens, 1980; Mangold, 1984), whichhas been recently named as the Orbis Zone (Dietl, 1982;Dietl and Callomon, 1988). It is also known from the LateBathonian of Madagascar (see Westermann and Callomon,1988; Jana, 2002). We, here, have made an attempt toimprove the biostratigraphical scheme, proposed byBardhan et al. (2002) for the Bathonian, based on ourpresent find of many geographically wide ranging taxawith stratigraphic control. They are mainly Oxycerites cf.orbis, M. bifurcatus transient intermedius and K. cf. etherid-
gei. The revised biostratigraphic zonation is shown inTables 1 and 2.
The proposed zones of the Bathonian strata are Etherid-gei, Triangularis and Chrysoolithicus in ascending order.The Chrysoolithicus Zone has been further subdivided intotwo subzones which are Chrysoolithicus and Madagascar-iensis in ascending order. The detailed ammonite faunalassemblages are given below.
arkelli horizon: This is marked by the first appearance ofGracilisphinctes cf. arkelli and Prohecticoceras manjalense
n. sp., Proceritess hians and Choffatia sp.manjalense horizon: P. manjalense is the most abundant
in this horizon, others continue but rare.etheridgei horizon: This is marked by the first appear-
ance of Kamptokephalites cf. etheridgei. Procerites hians.P. manjalense and G. cf. arkelli continue.
hians horizon: P. hians is most common and Macroceph-
alites triangularis appeared in this horizon. Other associ-ated ammonites are M. bifurcatus transient intermedius
and M. cf. mantataranus and Epimorphoceras decorum
(Waagen).triangularis horizon: This horizon is marked by the
abundance of M. triangularis. G. cf. arkelii, M. bifurcatus
transient intermedius and P. hians become rare. Epistrenoc-eras sp. is restricted to this horizon.
Table 1Biostratigraphic zonation of the Bathonain Stage in Kutch and its correlation with the Madagascar and West Pacific (Indonesia)
AGE
KUTCH MADAGASCAR INDONESIA
UPP
ER
BA
TH
ON
IAN
ZONE SUBZONEFAUNALHORIZON
CHRYSOO-LITHICUS
TRIANGU-LARIS
ETHERIDGEI
MADAGAS-CARIENSIS
TRIANGU-LARIS
CHRYSOO-LITHICUS
Macrocephalitesmadagascariensis
Sivajicerascongener
Oxyceritesorbis
M. chrysoolithicus
ZONE
M. triangularis
Procerites hians
TRIANGU-LARIS
FAUNALHORIZON
ETHERIDGEI
K. . etheridgeicf.
Prohecticocerasmanjalense
Gracilisphinctesarkelli
cf.
PROGRACILIS Gracilisphinctesprogracilis
M. madagascariensis
M. triangularis
Epistrenocerashistricoides
Micromphaliteshourcqi
Siemiradzkia sp.
Cadomites sp.
M. bifurcatus
M. b. intermedius
Satoceras satoi
M. apertusmantataranus
FAUNALASSOCIATION
MID
DL
EB
AT
HO
NIA
N
(Present study) (Collignon, 1958)(Westermann &Callomon, 1988)
? ?
Modified after Bardhan et al., 2002.
632 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
pychrysoolithicus horizon: This horizon is marked by the
first appearance of M. chrysoolithicus (Waagen) and Siv-
ajiceras congener and disappearance of M. triangularis.orbis horizon: Oxycerites cf. orbis first appeared in this
horizon. M. chrysoolithicus and S. congener continue.congener horizon: S. congener is the most abundant taxa
in this horizon; M. chrysoolithicus continues. Macrocepha-
lites madagascariensis Lemoine (1911) made firstappearance.
madagascariensis horizon: M. madagascariensis is themost dominant along with S. congener. Khreaiceras cf. han-
noveranum Roemer (1911) first appeared in this horizon.The lowest horizon of Etheridgei Zone described here is
Gracilisphinctes cf. arkelli from Jumara and Nara sectionsin the mainland of Kutch (see Figs. 2 and 3). G. arkelli
has been recorded earlier only from the island of Kutch(Singh et al., 1983 and Jaitly and Singh, 1984; Pandeyand Callomon, 1995) and has been assigned a secure ageof Middle Bathonain. Jaitly and Singh (1984) describedG. cf. arkelli from the Middle Bathonian of Patchamisland. Their specimens were associated with Micrompha-
lites Buckman (1923) which is known to occur from Lowerto Middle Bathonian of Europe (Arkell et al., 1957). But
Collignon (1958) reported Micromphalites (Micromphalites
hourcqi and M. golenkoi) even from the Upper Bathonianof Madagascar. However, Enay et al. (2001) have doubtedthe stratigraphy and believed a still older age. On the otherhand Micromphalites in Britain is known to occur withProcerites of Progracilis Zone (see Pandey and Callomon,1995). Pandey and Callomon (1995) described Procerites
(Gracilisphinctes) arkelli from the Patcham island of Kutchand compared it with G. arkelli from the early part ofProgracilis Zone of Madagascar. In the mainland at NaraG. cf. arkelli is co-occurring with Kamptokephalites cf.etheridgei and Procerites hians. K. cf. etheridgei is alsodescribed from Sula Island, Indonesia where majority ofthe specimens were reported from the Middle Bathonianbeds (Westermann and Callomon, 1988). However, onlya single specimen has been found to be in the Upper Batho-nian assemblage. In Jhura, Jain et al. (1996) have been able totrace the same bed continue from the island, which yieldsdefinite Middle Bathonian association such as Clydoniceras
triangulare Pandey and Agrawal (1984), C. pachchhamense
Pandey and Agrawal (1984), Micromphalites cf. clydocomph-
alus Arkell (1952), G. arkelli Collignon (1958) and Bull-
atimorphites s.s. Buckman (1921) sp. nov. They have also
Table 2Biostratigraphic zonation of the Bathonain Stage in Kutch and its correlation with the zones of France, England and Germany
AGE
KUTCH FRANCE
UP
PE
RB
AT
HO
NIA
N
ZONE SUBZONE FAUNALHORIZON
CHRYSOO-LITHICUS
TRIANGU-LARIS
ETHERIDGEI
MADAGAS-CARIENSIS
TRIANGU-LARIS
CHRYSOO-LITHICUS
Macrocephalitesmadagascariensis
Sivajicerascongener
Oxyceritesorbis
M. chrysoolithicus
ZONE
M. triangularis
Procerites hians
FAUNALHORIZON
ETHERIDGEI
K. c etheridgeif.
Prohecticocerasmanjalense
Gracilisphinctesrkellicf. a PROGRACILIS
Gracilisphinctesprogracilis
Clydonicerasdiscus
Epistrenocerashistricoides
Hemigarantianajulii
MID
DL
EB
AT
HO
NIA
N
(Present study)
?
DISCUS
RETRO-COSTRATUM
BREMERI
SUBCONT-RACTUS
DISCUS
RETRO-COSTRATUM
SUBZONE
BLANAZENSE
MORRISI
SUBCONT-RACTUS
PROGRACILIS
BREMERI
C. hollandi
Prohecticocerasblanazense
Proceritesmirabilis
Cadomitesorbignyi
Wagnericerasfortecostatum
Bullatimorphitesbullatimorphus
Morrisocerasmorrisi
Tulitessubcontractus
ENGLAND
ZONE SUBZONE
DISCUS
DISCUS
HOLLANDI
ORBIS
HODSONI
MORRISI
SUBCONTRACTUS
PROGRACILIS
ZONE SUBZONE
DISCUS
HOLLANDI
DISCUS
GERMANY
ORBIS
HODSONI
MORRISI
SUBCONTRACTUS
PROGRACILIS
Modified after Bardhan et al., 2002.
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 633

Autho
r's
pers
onal
co
py
recorded a Gracilisphinctes specimen from this bed (herebed 3, see Fig. 4) in Jhura (see also Mukherjee et al.,2003). K. cf. etheridgei describes from Jhura underlies theGracilisphinctes bearing bed. Therefore, the faunal associa-tion as well as the stratigraphic position of this zonesecurely places it within the Middle Bathonian.
According to Pandey and Callomon (1995), Procerites
(G.) arkelli of Kutch and Madagascar and Procerites (G.)progracilis of Europe resemble so closely that they may rep-resent geographical subspecies. Hence G. cf. arkelli bearingEtheridgei Zone of Kutch can be correlated with Prograc-ilis Zone of Europe and Madagascar. Correlation of thiszone with contemporary biozonation of other areas hasbeen shown in Tables 1 and 2.
We have also extended the stratigraphic range of thepresent species G. cf. arkelli up to the Upper Bathonian.Singh et al. (1983) described Clydoniceras sp. associatedwith G. cf. arkelli from Middle Bathonian of Patchamisland. Clydoniceras Blake (1905) ranges form Middle toUpper Bathonian (Arkell et al., 1957). Some of our speci-mens of G. cf. arkelli coexist with Macrocephalites triangu-
laris and Epistrenoceras sp. (see Kayal and Bardhan, 1998).Epistrenoceras is a time diagonostic genus of the LateBathonian. Therefore, the present Gracillisphinctes speciesranges evidently up to the Upper Bathonian beds.
In the present study the Upper Bathonian substage hasbeen subdivided into Triangularis and Chrysoolithicuszones in ascending order. Macrocephalites triangularis isthe zonal index fossil for the Late Bathonian of the Indo-Madagascan Province. In Beluchistan and Madagascar ofthe same province, M. triangularis was found to be associ-ated with Clydoniceras Blake, Micromphalites Buckmanand Kheraiceras cf. bullatum (D’Orbigny) of Late Batho-nian age (see Westermann and Callomon, 1988 and refer-ences there in), but in Kutch, the above association ofM. triagularis is totally absent. Based on comparison ofsimilar macrocephalitin ammonoides from Kutch andother parts of the world, workers (Krishna and Wester-mann, 1985, 1987; Callomon, 1993; Datta et al., 1996) sug-gested a Late Bathonian age for M. triangularis. However,no general consensus has been achieved. But the recentrecord of Epistrenoceras Bentz co-occurring with M. tri-
angularis resolves the long-standing problem (Kayal andBardhan, 1998). Epistrenoceras with its near circum globaldistribution, but cryptic appearance in the Late Bathonianmakes it an ideal fleeting genus (sensu Ager, 1984 and alsosee Jana, 2002). Epistrenoceras comes from the Retrocost-atum Zone in Europe (Dietl, 1982). It is also known fromthe Late Bathonian of Madagascar and South America(Westermann and Callomon, 1988 and Sandoval et al.,1990). Other associated ammonoids co-occurring with M.
triangularis are M. cf. mantataranus, M. cf. bifurcatus tran-sient intermedius and Procerites hians. M. cf. mantataranus
is an Upper Bathonian form in Indonesia (see Westermannand Callomon, 1988). In the present endeavour Oxycerites
cf. orbis has been found to occur in the upper part of theChrysoolithicus Subzone. O. orbis is a zonal index of Orbis
Zone of Northwest Europe (see Dietl, 1982; Westermannand Callomon, 1988).
The topmost faunal horizon of the MadagascariensisSubzone, Chyrsoolithicus Zone contains another time-diagnostic ammonite, i.e., Kheraiceras cf. hannoveranum
(Jana et al., 2000; Bardhan et al., 2002). Kheraiceras han-novernum comes from the Late Bathonian of Europe (seealso Callomon, 1993) and is reported from various faunalprovinces of coeval age (Jana et al., 2000; Jana, 2002).
On the basis of above ammonite assemblages within theUpper Bathonian, the Kutch biozones have been correlatedwith those of other provinces (see Tables 1 and 2).
4. Systematic palaeontology
D, shell diameter; U, umbilical diameter; W, width ofthe shell; H, height; all measurements are in millimeter;M, macroconch; m, microconch. All specimens are repos-ited at the Department of Geological Sciences, JadavpurUniversity, Kolkata, India.
Superfamily Perisphinctaceae Steinmann, 1890Family Perisphinctidae Steinmann, 1890Subfamily Zigzagiceratinae Buckman, 1920Genus Procerites Siemiradzki, 1898Type species Ammonites procerus Schloenbach, 1865
(=Perisphinctes schloenbachi Grossouvre, 1907)Procerites hians (Waagen) M & mFigs. 5.1–5.8 and 6.1–6.5
1875 Perisphinctes hians Waagen; p. 153, pl. 57,Figs. 2a–c.
1899 Perisphinctes hians Waagen; Semiradzki,p. 323.
Non 1914 Perisphinctes hians Waagen, Smith, p. 1351.Non 1924 Kinkeliniceras? cf. hians (Waagen) Spath,
p. 23.1931 Procerites hians (Waagen) Spath, pp. 286–287.
4.1. Material
Material include the holotype (Geological Survey ofIndia Type No. 2095) and one phragmocone (JUM/H/3)from Bed 1, Jumara; two other fragments and two bodychamber (JUM/H/1 and JUM/H/4) fragments of macro-conch and two microconchs (JUM/H/5–6) – all are col-lected from Bed 2 of Jumara; one fragmentaryphragmocone (JUM/H/2) from the marly limestone bed(=Bed 1) of Nara.
4.2. Description [M]
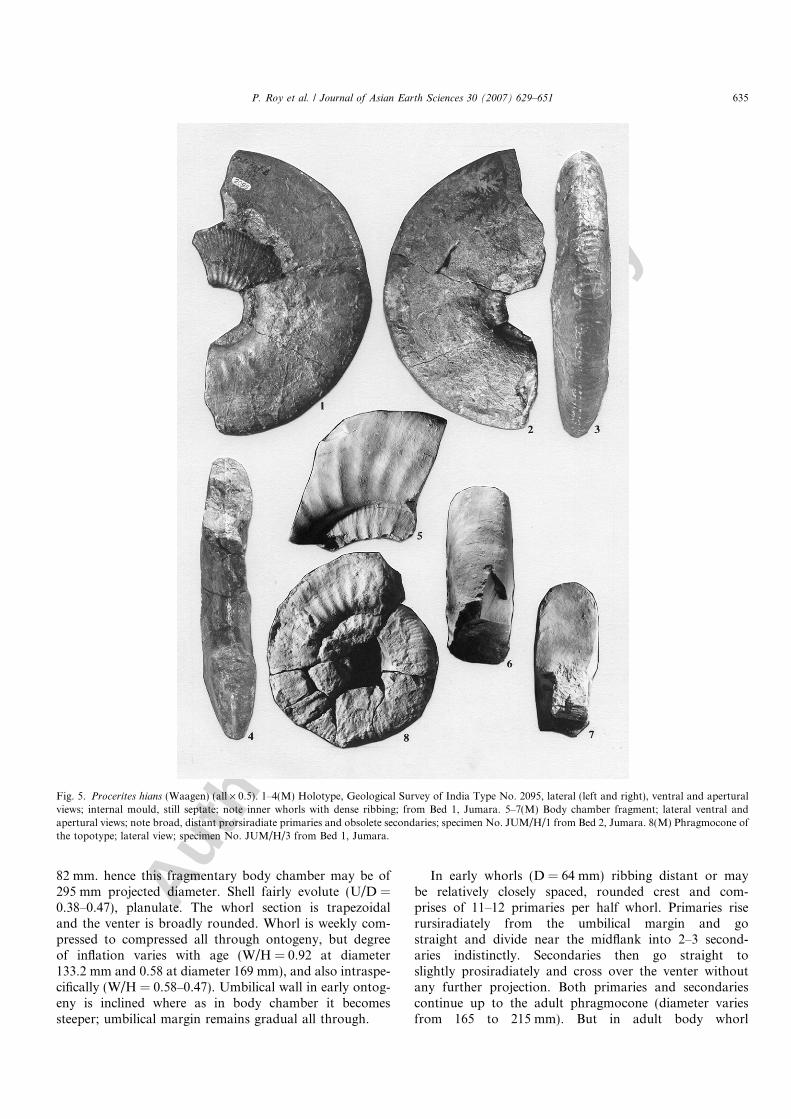
Shell large, maximum septate diameter observed isabout 215 mm. (holotype, Geological Survey of India TypeNo. 2095, here reproduced in Figs. 5.1–5.4). Isolated frag-mentary body chambers are found whose height may attain
634 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651
Autho
r's
pers
onal
co
py
82 mm. hence this fragmentary body chamber may be of295 mm projected diameter. Shell fairly evolute (U/D =0.38–0.47), planulate. The whorl section is trapezoidaland the venter is broadly rounded. Whorl is weekly com-pressed to compressed all through ontogeny, but degreeof inflation varies with age (W/H = 0.92 at diameter133.2 mm and 0.58 at diameter 169 mm), and also intraspe-cifically (W/H = 0.58–0.47). Umbilical wall in early ontog-eny is inclined where as in body chamber it becomessteeper; umbilical margin remains gradual all through.
In early whorls (D = 64 mm) ribbing distant or maybe relatively closely spaced, rounded crest and com-prises of 11–12 primaries per half whorl. Primaries riserursiradiately from the umbilical margin and gostraight and divide near the midflank into 2–3 second-aries indistinctly. Secondaries then go straight toslightly prosiradiately and cross over the venter withoutany further projection. Both primaries and secondariescontinue up to the adult phragmocone (diameter variesfrom 165 to 215 mm). But in adult body whorl
Fig. 5. Procerites hians (Waagen) (all · 0.5). 1–4(M) Holotype, Geological Survey of India Type No. 2095, lateral (left and right), ventral and aperturalviews; internal mould, still septate; note inner whorls with dense ribbing; from Bed 1, Jumara. 5–7(M) Body chamber fragment; lateral ventral andapertural views; note broad, distant prorsiradiate primaries and obsolete secondaries; specimen No. JUM/H/1 from Bed 2, Jumara. 8(M) Phragmocone ofthe topotype; lateral view; specimen No. JUM/H/3 from Bed 1, Jumara.
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 635

Autho
r's
pers
onal
co
py
fragment secondaries disappear or feebly present andprimaries become broad, flat and distant on internalmould.
Septal sutural pattern may be complex, lobes are deeplyincised, highly frilled and saddles are broad with large sec-ondary lobes. Waagen (1875, pl. 57, Fig. 2c.) gave a detailaccount of the sutural pattern of the holotype, which isnow refigured here in Figs. 5.1–5.4.
4.3. Description [m]
Shell represented by only body whorl fragments which iscompressed (W/H = 0.48), planulate to subplanulate.Whorl section subtrapezoidal, umbilicus wide, inclined wallwith gradual margin.
Primaries arise from the umbilical wall, more or lessstraight up to midflank where they divide into 2–3 rel-
Fig. 6. (1–5) Procerites hians (Waagen) (all · 1). 1(M) Incomplete phragmocone; lateral view; specimen No. JUM/H/2 from marly limestone bed, Nara.2–4(m) Lateral, ventral and apertural views of body chamber fragment; specimen No. JUM/H/5 from Bed 2, Jumara. 5(m) Lateral view of body whorlfragment; note modification of ribbing towards the aperture; specimen No. JUM/H/6 from Bed 2, Jumara. (6–13) Sivajiceras congener (Waagen)(6–10 · 0.5, 11 · 0.45, 12–13 · 1). 6–8(M) Lateral, ventral and apertural views of holotype (GSI Type no. 2091); shell still septate; from Bed 1, Jumara.9–10(M) Lateral and ventral views of nearly complete specimen (JUM/GEO/1) from Bed 4, Jumara. 11(M) Lateral view of a fragmentary body chamber;note strong bullae like primaries; specimen No. JUM/GEO/5 from Bed 4, Jumara. 12–13(m) Lateral views of body whorl fragment; specimen Nos.JUM/GEO/7 and JUM/GEO/6, both are from Bed 4, Jumara.
636 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
atively fine secondaries. Primaries are strong, closelyspaced to distant, with rounded crest. There are a fewintercalatories placed irregularly. Secondaries are rectira-diate and straightly cross over the venter till the latepart of the body chamber; but near aperture they showlittle sinuosity which suggests the maturity of theshell.
4.4. Discussion
See discussion part of Sivajiceras congener.
4.5. Dimensions
Specimen No. D U W H
GeologicalSurvey ofIndia TypeNo. 2095 (M)
Phragmocone 215 66 40c 86Phragmocone 163 62 32 68Phragmocone 123.5 35 17 48.5
JUM/H/1 (M) – – 52 82JUM/H/2 (M) Body
chamber128 c c44 29 51
JUM/H/3 (M) Phragmocone 169 81 31.5 54Inner whorl 133.5 55.5 33.8 36.5
JUM/H/4 (M) Inner whorl – – 40.5 72JUM/H/5 (M) Inner whorl – – 32 55JUM/H/7 (M) Phragmocone 17 27JUM/H/6 (m) Body
chamber– – 12 25
Genus Gracilisphinctes Buckman, 1920Type species Ammonites gracilis Buckman, 1844 (nonZieten, 1830) (=Procerites progracilis Cox and Arkell,1950)Gracilisphinctes cf. arkelli CollignonFigs. 8.1–8.8
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0.25 0.35 0.45 0.55U/D
W/H
P. hians
S. congener
Fig. 7. Bivariate analysis involving degree of inflation (width/height) vs.degree of involution (umbilical diameter/shell diameter) of Procerites hians
(Waagen) and Shivajiceras congener (Waagen).
cf. 1958 Gracilisphinctes arkelli Collignon, pl. 6,Figs. 31–33.
cf. 1983 Gracilisphinctes arkelli Collignon, Singh et al.p. 823, Fig. 2.
1984 Gracilisphinctes cf. arkelli Jaitly and Singh,p. 319, Fig. 2.
1995 Procerites (Gracilisphinctes) cf. arkelli Collignon,Pandey and Callomon, p. 129, pl. 1, Fig. 8;pl. 2–5.
4.6. Material
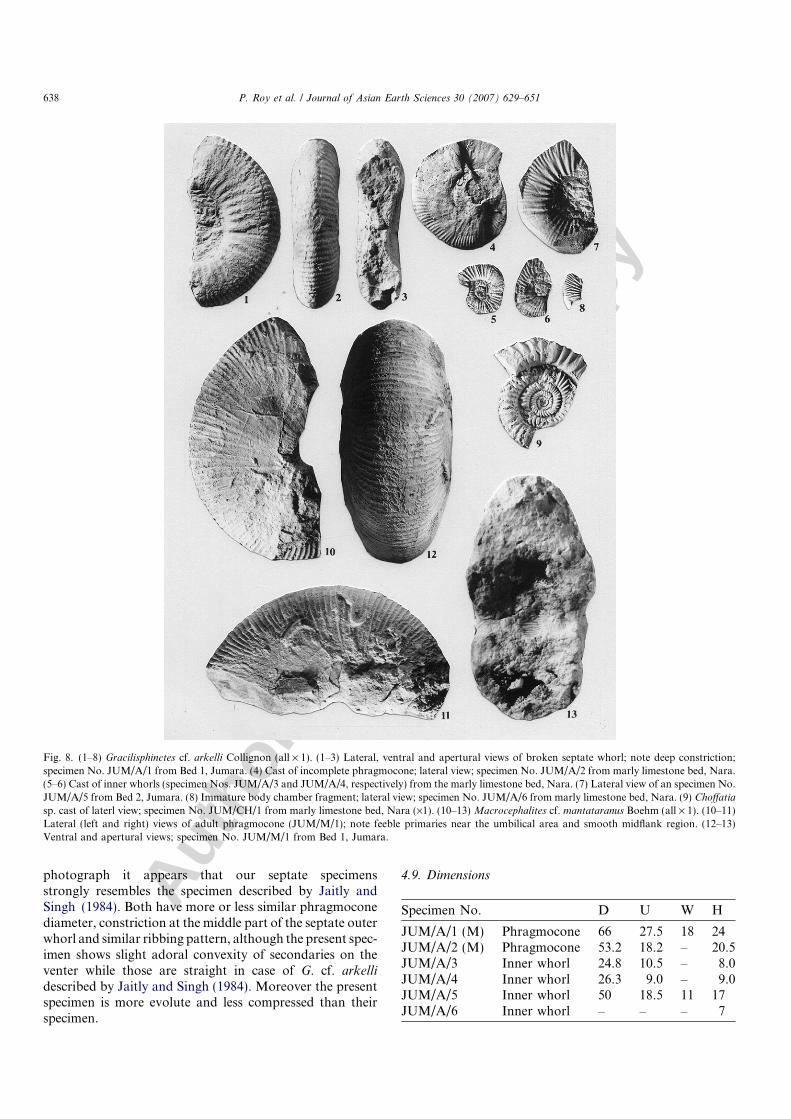
Six specimens; all are immature. JUM/A/1 and JUM/A/5 are from bed 1 and bed 2 of Jumara, respectively. Otherspecimens (JUM/A/2, JUM/A3–4 and JUM/A/6) are frommarly limestone bed of Nara.
4.7. Description
The shell is immature, fully septate, subplanulate, evo-lute (U/D = 0.34–0.43) and compressed (W/H = 0.64–0.75). The shell inflation shows negative allometry duringontogeny (W/H = 1.3 at 38 mm to 0.75 at 66 mm diame-ter). Umbilical wall inclined and umbilical marginrounded. Venter is rounded and narrow. Apertural outlinechanges from subquadrate to ovate during ontogeny.
Dense, thick primary ribs arise from umbilical wall, gostraight up to the midflank and divide into 2–3 secondar-ies. Secondaries cross over the venter with slightly adoralprojection. At the later part of the shell primaries becomestrong and remain rectiradiate. Secondaries are relativelyfiner and sharply crested. There are also occasional soli-tary ribs. The shell is deeply constricted. In fragmentaryspecimen at least two constrictions may be seen per halfwhorl. There are 18 primaries and 40 secondaries per halfwhorl.
4.8. Discussion
Pandey and Callomon (1995) vividly described Proce-
rites (Gracilisphinctes) arkelli based on numerous adultspecimens from the Patcham island of Kutch. Fromtheir description of inner whorls it appears that ourspecimens closely resemble inner whorls of their speci-mens in having equally evolute whorls, straight rectira-diate primaries, number of primaries and secondariesper half whorl and showing negative allometry in degreeof inflation.
The European zonal index, Gracilisphinctes progracilis
(Cox and Arkell) is similar in size with that of Kutch mac-roconch, however G. progracilis is less evolute (in the typespecies U/D is 0.33), relatively strongly ornate (see alsoSingh et al., 1983).
Collignon (1958) reported G. arkelli from Madagascar.Jaitly and Singh (1984) described G. cf. arkelli from thePatcham island of Kutch. From their description and
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 637
Autho
r's
pers
onal
co
py
photograph it appears that our septate specimensstrongly resembles the specimen described by Jaitly andSingh (1984). Both have more or less similar phragmoconediameter, constriction at the middle part of the septate outerwhorl and similar ribbing pattern, although the present spec-imen shows slight adoral convexity of secondaries on theventer while those are straight in case of G. cf. arkelli
described by Jaitly and Singh (1984). Moreover the presentspecimen is more evolute and less compressed than theirspecimen.
4.9. Dimensions
Specimen No. D U W H
JUM/A/1 (M) Phragmocone 66 27.5 18 24JUM/A/2 (M) Phragmocone 53.2 18.2 – 20.5JUM/A/3 Inner whorl 24.8 10.5 – 8.0JUM/A/4 Inner whorl 26.3 9.0 – 9.0JUM/A/5 Inner whorl 50 18.5 11 17JUM/A/6 Inner whorl – – – 7
Fig. 8. (1–8) Gracilisphinctes cf. arkelli Collignon (all · 1). (1–3) Lateral, ventral and apertural views of broken septate whorl; note deep constriction;specimen No. JUM/A/1 from Bed 1, Jumara. (4) Cast of incomplete phragmocone; lateral view; specimen No. JUM/A/2 from marly limestone bed, Nara.(5–6) Cast of inner whorls (specimen Nos. JUM/A/3 and JUM/A/4, respectively) from the marly limestone bed, Nara. (7) Lateral view of an specimen No.JUM/A/5 from Bed 2, Jumara. (8) Immature body chamber fragment; lateral view; specimen No. JUM/A/6 from marly limestone bed, Nara. (9) Choffatia
sp. cast of laterl view; specimen No. JUM/CH/1 from marly limestone bed, Nara (·1). (10–13) Macrocephalites cf. mantataranus Boehm (all · 1). (10–11)Lateral (left and right) views of adult phragmocone (JUM/M/1); note feeble primaries near the umbilical area and smooth midflank region. (12–13)Ventral and apertural views; specimen No. JUM/M/1 from Bed 1, Jumara.
638 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
Subfamily Pseudoperisphinctinae Schindewolf, 1925Genus Sivajiceras Spath, 1928Type species Perisphinctes congener Waagen, 1875Sivajiceras congener (Waagen)Figs. 6.6–6.13
1875 Perisphinctes congener Waagen; p. 171, pl. 56,Fig. 2; pl. 57, Figs. 1a and b.
1899 Perisphinctes congener Waagen, Siemiradzki,p. 310.
1924 Grossouvria (?) congener (Waagen) Spath,pp. 3, 22.
1930 Grossouvria (?) congener (Waagen) Spath,p. 35.
1931 Sivajiceras congener (Waagen) Spath, p. 287.Non 1911 Perisphinctes (Procerites) congener Waagen,
Lemoine, p. 41, pl. v, Fig. 5.1993 Sivajiceras congener (Waagen) Callomon,
p. 236, pl. 23, pl. 24, Figs. 1 and 2.
4.10. Material
Material include the holotype (Geological Survey ofIndia Type No. 2091) from Bed 2, Jumara; one completespecimen (JUM/GEO/1) and two other fragments (JUM/GEO/5 and JUM/GEO/8) from bed 4 of Jumara and 3other fragmentary specimens from bed 2 of Jumara. Twobody chamber fragment of a microconch (JUM/GEO/6–7) from Bed 4 of Jumara.
4.11. Description [M]
Shell large, adult phragmocone diameter may attain170 mm. However smaller adult variant may attain fullgrown size having diameter of 137 mm with adultphragmocone diameter about 82 mm. Shell is evolute(U/D = 0.39–0.45). Shell compressed, degree of inflationdecreases during ontogeny (W/H = 0.64 at D = 85 mmand 0.77 at D = 167 mm). Umbilical wall gradual to stee-ply inclined, umbilical margin rounded in early whorlsand become distinct in adult whorl. Flank is curved in earlyontogeny, becoming flattish subsequently. Venter narrow,convex; ventrolateral margin gradual.
Inner whorls though present destroyed by recrystallisa-tion obliterating the ornament. Primary ribs rising at75 mm diameter from the umbilical wall are strong, distantand are with rounded crest. These are slightly rursiradiateat the umbilical margin and then go straight to slightlyprorsiradiately up to the midflank where they divide into3–4 relatively fine secondaries. Strength of primariesvaries during ontogeny as well as intraspecifically. Inphragmocone primaries are sharp, raised but in adult bodywhorl they may be thicken, very strong to form bullae,which become broad. Secondaries are slightly prorsiradiateand show a slight adoral convexity at the venter. Intercala-tories are also present and irregularly placed. Bodychamber occupies the whole of the last whorl. Ribbing in
the last quarter of the body whorl shows modification,primaries become distant and secondaries show markedadoral convexity; constriction present. In one variantwhere aperture is preserved, peristome appears to be sim-ple. Ribs including both primaries and secondaries con-tinue up to the last part of body chamber, they tend todisappear near the peristome where only dense growthstriae are present.
Septal suture is well discernible in the holotype anddescribed by Waagen (1875, see pl. 46, Fig. 20).
4.12. Description [m]
Two body whorl fragments present. Shell compressed(W/H = 0.65) with distant raised blunt crested primarieswhich originate at umbilical wall, go straight up to the mid-flank and bifurcate. Primaries are thick and bullae like atlater stage. Secondaries go straight over the rounded nar-row venter.
4.13. Discussion
This species is closely comparable with P. hians (Waa-gen) and Callomon (1993) in fact believed that they maybe conspecific, representing two extreme variant of awidely variable biological species. But closer scrutinyreveals that they exhibit many discontinuities in morpho-logical characters and Spath (1931) was right in havingseparated them into two genera. Body size is a morpho-logical character of a species (Jablonsky, 1996). P. hiansis larger than S. congener; phragmocone diameter of theformer exceeds the adult body chamber diameter of thelater. The holotypes of two species are septate and thespecies were previously described on these monotypic hol-otypes. Present collection includes additional materialsome of which having adult body chamber or fragmentsof body whorls. This enables us to compare the ontoge-netic developments in these two species. Inner whorls ofboth of them are indistinguishable with respect to orna-mentation. However, subsequently there is change in nat-ure of ornamentation especially in S. congener in itspenultimate whorl as well as in body chamber; primariesbecome thicken, bullae-like and split relatively at higherflank. Both primaries and secondaries continue till tothe end of adult body chamber without any sign of atten-uation of ribbing. In P. hians however, ribbing becomeprogressively weak in adult phragmocone and body cham-ber. In body chamber the weak to obsolete secondariescompletely disappear from the ventral region. Moreover,bivariate analysis involving degree of inflation and degreeof involution of both species show that S. congener is rel-atively evolute (see Fig. 7).
The difference between these two genera is also evidentin Microconch. Microconch of Procerites is also thor-oughly ribbed, bifurcated and showing modification nearthe aperture but ribs are relatively denser and split on thehigher flank in case of Procerites.
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 639

Autho
r's
pers
onal
co
py
Moreover, these two species have different stratigraphicdistribution with a narrow zone of overlapping. P. hians
being older (Fig. 2), we therefore believe that these two spe-cies are not conspecific and instead form an evolutionaryplexus and as such we support Spath. Sivajiceras evolvedfrom Procerites and an endemic genus which subsequentlydiversified in Callovian and giving rise to other generaObtusicostites Buckman (M)–Hubertoceras Spath (m) (seealso Callomon, 1993). The evolutionary mechanism is envis-aged to be complex heterochronic. Sivajiceras retains earlyornamental features of Procerites (paedomorphosis) andappearance of more advanced features, i.e., bullae-like pri-mary ribs (paramorphosis). This indicates complex inter-play of heterochronic processes. Neotenus descendantoften exhibits novelties (see Bardhan et al., 1994). Rudraet al. (in press) recently demonstrated this kind of mosaicevolution in the Early Cretaceous trigoniid bivalve fromKutch where neotenus retension of shell outline of Opistho-
trigonia spissicostata (Kitchin, 1903) did not hinder the earlyonset of the ribbing pattern in descendant Eselaevitrigonia
cardiniformis (Kitchin, 1903).
4.14. Dimensions
Specimen No. D U W H
Geological Surveyof India TypeNo. 2094 (M)
Phragmocone 167 76.5 43 56.2Inner whorl 140 64.5 29 43
JUM/GEO/1 (M) Body chamber 137 54 32 52Phragmocone 82 42.5 21 31.5
JUM/GEO/ 2 (M) Phragmocone – – 24 25JUM/GEO/3 (M) Phragmocone – – 22 31JUM/GEO/4 (M) Phragmocone – – 39.5 42.5
Inner whorl – – 18 26JUM/GEO/5 (M) Phragmocone – – 24 39JUM/GEO/6 (m) Body chamber – – 17 26JUM/GEO/7 (m) Body chamber – – 23 c7
Genus Choffatia Siemiradzki, 1898Type species Perisphinctes cobra Waagen, 1875,
SD Buckman, 1920Choffatia sp.Fig. 8.9
4.15. Material
One external mould (JUM/CH/1) from marly limestonebed of Nara.
4.16. Description
Shell with partially preserved body chamber; havingreconstructed diameter of 46 mm. Shell is fairly evolute(U/D = 0.58), flank strongly convex. Umbilical wall issteep with rounded margin at the late stage. Inner whorl
densely ribbed, only primaries can be seen in the umbilicussuggesting that furcation takes place higher at the outerflank. Ribs are sharp, straight to slightly forwardly curved;becoming distant, coarse with rounded crest and the lasttwo to three ribs of the preserved specimen show markedforward projection towards the venter indicating perhapsits adulthood. At 39 mm of diameter number of primariesis 38 per half whorl and in the last quarter of the whorlnumber of primaries is 8.
4.17. Discussion
Present specimen resembles some of the Bathonianmicroconchiate forms described by Mangold (1970) fromFrance. Choffatia (Grossouvria) bathonica Mangold (1970,pl. 3, Figs. 3–7) is similarly evolute and has dense ribbingon inner whorls but they are smaller in size and secondariesshows backward projection. Mangold’s Homoeplanulites
(H.) rambertsensis (pl. 3, Fig. 8) may be compared withthe present species in having coarse distant primaries inouter whorl but it can be distinguished by its less evolutecoiling and rursiradiate secondaries. Homoeplanulites (H.)pseudoannularis (Lissajous) (pl. 3, Figs. 11 and 12) stronglyresembles the present Kutch species in having similardegree of involution and nature of ribbing. They appearto be conspecific, but we refrain ourselves from synonymis-ing them because of the incompletely preserved single spec-imen of Kutch.
The present species also resembles Choffatia (Homoe-
planulites) aff. or cf. furcula described by Westermannand Callomon (1988) from the unspecified stratigraphy ofSula island Indonesia. It is a large macroconch. Its innerwhorls are however, ornamented with dense, fine primariessimilar to that of Kutch species, but the former has a char-acteristic straight ribbing without forward projection. It isalso frequently constricted.
The present species has similarities with the Bathonianforms of other areas, e.g., Choffatia boehmi (Steinmann)(m) described by Sandoval et al. (1990, pl. 13, Figs. 6aand b; pl. 14, Figs. 6a and b) from the Steinmanni Zoneof Mexico. Both species have dense, fine primaries in earlywhorls and similarly evolute shell. The Mexican specieshowever, appears to be more flattened with strong radialribbing.
Superfamily Stephanocerataceae Neumayr, 1875Family Sphaeroceratidae Buckman, 1920Subfamily Macrocephalitinae Salfeld, 1921Genus Macrocephalites Zittel, 1884
Type species Ammonites macrocephalus Schlotheim,1813, SD Lemoine, 1910; neotype designated and describedby Callomon, 1971; ICZN opinion 1275 (1984, see Callo-mon, 1980).
Macrocephalites cf. mantataranus BoehmFigs. 8.10–8.13
640 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
cf. 1912 Macrocephalites mantataranus Boehm, p. 159,pl. 35, Figs. 3a and b, Text-Figs. 68a and b.
cf. 1912 Microcephalites keeuwensis sp. nov. Boehm,p. 162, pl. 39, Fig. 4, Text-Fig. 73.
cf. 1978 Macrocephalites formosus, transient triangularis
Spath, 1928; Thierry, p. 287, pl. 24, Figs. 5aand b; ?Text-Fig. 106d.
cf. 1978 Macrocephalites (Macrocephalites) formosus
mantataranus Sato et al., p. 10.cf. 1988 Macrocephalites mantataranus Boehm,
Westermann and Callomon, pl. 10, Figs. 1–5;Text-Figs. 11 and 18.
cf. 1997 Macrocephalites mantataranus Boehm, Enayand Cariou, p. 8.
4.18. Material
One phragmocone (JUM/M/1) from Bed 1, Jumara.
4.19. Description
Shell still septate at 93.5 mm diameter, involute(U/D = 0.1) and shell inflation decrease during ontogeny(W/H = c1.4 at D = 34 mm and 0.95 at D = 93.5 mm).Inflation of the inner whorls also suggests still larger value.Flank slightly curved and converging towards venter. Venterbroad, rounded and the whorl section is elliptical. Umbilicalwall vertical to overhanging with distinct margin.
Primaries originate at the umbilical margin, stronglyrursiradiately, thereafter projected forward near the innerflank, forming a little concavity near the umbilical margin.In early late phragmocone stage dense, sharp crestedprimaries continue and divide at the midflank. Secondariesoriginate at midflank and go straight over the venter. Sec-ondaries become increasingly finer and denser towards theadult phragmocone stage. Primaries in late phragmoconestage die out shortly after forming ‘‘crescentic’’ patternnear the umbilical region rendering the midflank regionsmooth which is the typical ornamental pattern of the spe-cies. This feature along with trace of eggressing body cham-ber indicates that the specimen is an adult phragmocone.
Septal suture is not fully exposed especially near theumbilical region but appears to be quite complex as shownin the holotype and mentioned by Westermann and Callo-mon (1988).
4.20. Discussion
The present species M. mantataranus closely resemblesM. triangularis and Thierry (1978) considered them as con-specific. We support Westermann and Callomon (1988)who correctly pointed out their difference. The juvenilewhorls of both species are moderately evolute and somewhatinflated and coarsely ornate. Their outer whorls are typicallysubtriangular and both have completely smooth adult bodychamber. Westermann and Callomon (1988) described a
small specimen of M. mantataranus (their pl. 10, Figs. 3aand b) as an ornate variant. This is a complete specimenretaining ribs till to the end. We consider this specimen tobe a microconch which is commensurate with the macroce-phalitin microconchiate population of the East Indies. Themain difference between M. mantataranus and M. triangular-is lies in the adult body size and ornamentation. The maxi-mum adult whorl diameter of M. mantataranus is 170 mmwhile that of M. triangularis is 270 mm (see Datta et al.,1996). M. cf. mantataranus has a peculiar ribbing pattern.On the outer whorl primaries die out near midflank whererectiradiate secondaries arise. There remain only distant,crescent primaries on the innermost flank and straight, bluntsecondaries on the ventrolateral and siphonal parts. Thisobliteration of ribbing at midflank is never noticed in anygrowth stage of M. triangularis. Moreover, ribs are sharperand denser in M. triangularis. We believe that M. triangularis
has evolved from M. mantataranus. Krishna and Cariou(1993) also had similar opinion but they did not elaborate.In M. triangularis the obliteration of ribs spread further formmidflank towards umbilical part rendering the whole innerand midflank regions smooth.
4.21. Dimensions
Specimen No. D U W H
JUM/M/1 Phragmocone 93.5 9.6 48.5 51.1Inner whorl 70 – 40 38.5Inner whorl 34 – 21 15
Macrocephalites cf. bifurcatus transient intermedius
(Spath)Figs. 9.1–9.9
cf. 1912 Macrocephalites keeuwensis b Boehm, p. 162,pl. 38, Figs. 3a and b
cf. 1912 Macrocephalites keeuwensis Boehm, p. 162,pl. 40, Figs. 1a and b.
cf. 1928 Eucycloceras intermedium Spath, p. 210 [forBoehm, 1912, Figs. 3a and b].
cf. 1978 Macrocephalites subtrapezinus dimorphemicroconche, morphe intermedius – Thierry,p. 198, pl. 7, Figs. 5–7, Text-Fig. 65.
cf. 1988 Macrocephalites bifurcatus transient intermedius,Westermann and Callomon, p. 54, pl. 4–7;Text-Figs. 15–17.
4.22. Material
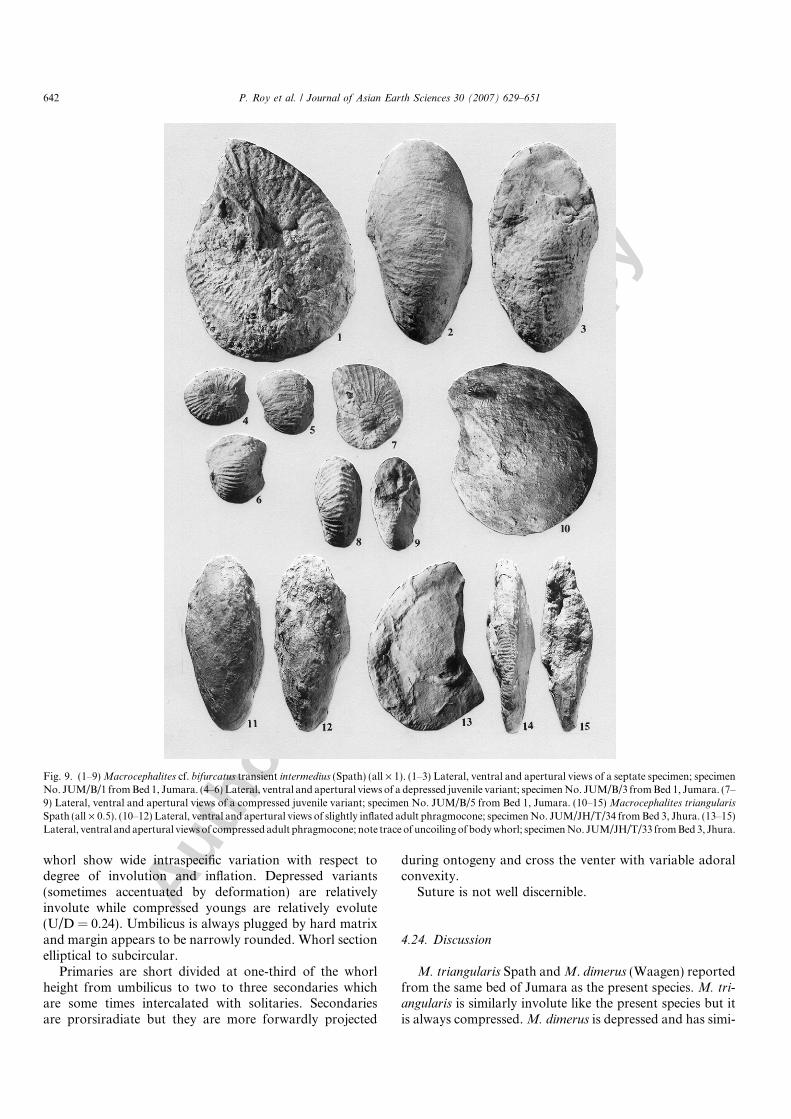
Five phragmocone (JUM/B/1-5) from Bed 1, Jumara.
4.23. Description
Shell septate, maximum diameter observed is 93 mm.Body chamber is not found in any of the specimens. Early
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 641
Autho
r's
pers
onal
co
py
whorl show wide intraspecific variation with respect todegree of involution and inflation. Depressed variants(sometimes accentuated by deformation) are relativelyinvolute while compressed youngs are relatively evolute(U/D = 0.24). Umbilicus is always plugged by hard matrixand margin appears to be narrowly rounded. Whorl sectionelliptical to subcircular.
Primaries are short divided at one-third of the whorlheight from umbilicus to two to three secondaries whichare some times intercalated with solitaries. Secondariesare prorsiradiate but they are more forwardly projected
during ontogeny and cross the venter with variable adoralconvexity.
Suture is not well discernible.
4.24. Discussion
M. triangularis Spath and M. dimerus (Waagen) reportedfrom the same bed of Jumara as the present species. M. tri-angularis is similarly involute like the present species but itis always compressed. M. dimerus is depressed and has simi-
Fig. 9. (1–9) Macrocephalites cf. bifurcatus transient intermedius (Spath) (all · 1). (1–3) Lateral, ventral and apertural views of a septate specimen; specimenNo. JUM/B/1 from Bed 1, Jumara. (4–6) Lateral, ventral and apertural views of a depressed juvenile variant; specimen No. JUM/B/3 from Bed 1, Jumara. (7–9) Lateral, ventral and apertural views of a compressed juvenile variant; specimen No. JUM/B/5 from Bed 1, Jumara. (10–15) Macrocephalites triangularis
Spath (all · 0.5). (10–12) Lateral, ventral and apertural views of slightly inflated adult phragmocone; specimen No. JUM/JH/T/34 from Bed 3, Jhura. (13–15)Lateral, ventral and apertural views of compressed adult phragmocone; note trace of uncoiling of body whorl; specimen No. JUM/JH/T/33 from Bed 3, Jhura.
642 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
lar whorl section like some of the variants of the present spe-cies but it is evolute (see Jana, 2002).
Westermann and Callomon (1988) compared M. bifurcatus
transient intermedius with another Bathonian species M. ethe-
ridgei and they showed that the first species differs in havingfine-ribbing, more involute shell, smaller and smoother outerwhorl; unfortunately this latter aspect can not be seen in theKutch population since all specimens lack body chamber.
4.25. Dimension
Specimen No. D U W H
JUM/B/1 Phragmocone 85 – 39.2 42JUM/B/2 Phragmocone 56 10 33 33JUM/B/3 Phragmocone 27 6 23 16JUM/B/4 Phragmocone 38 – – 17JUM/B/5 Phragmocone 38 10.5 19 23
Macrocephalites triangularis SpathFigs. 9.10–9.15
1924 Macrocephalites aff. chariensis (Waagen);Spath, p. 22 (No. 218).
1928 Macrocephalites triangularis Spath, p. 180,pl. 21, Figs. 1a and b.
1951 Macrocephalites triangularis Spath, Basseand Perrodon, p. 22.
Non 1978 Macrocephalites formosus (J. de C. Sowerby)transient triangularis Spath, Thierry, pl. 24,Figs. 1–5.
Non 1987 Macrocephalites cf. triangularis Spath,Krishna and Westermann, pl. 1, Figs. 2 and 3.
1992 Macrocephalites triangularis Spath, Datta,pl. 1, Figs. 1–4.
1996 Macrocephalites triangularis Spath, Dattaet al., pl. 1, Figs. 1–4; pl. 2, Figs. 1–5;Text-Figs. 3a–d.
2002 Macrocephalites triangularis Spath, Jana,pl. 1, Figs. 1–4; Text-Figs. 8–11.
1997 Macrocephalites triangularis Spath, Enay andCariou, p. 8.
4.26. Material
Three adult phragmocone (JUM/JH/T/33-35) from Bed 3of Jhura.
4.27. Comments
Specimens are not very well preserved, mostly internalmould and lack body chamber. The degree of inflation(W/H) varies from 0.47 to 0.72 and that of involution(U/D) varies from 0.08 to 0.11. This variability falls withinthe range of interspecific variation noted in Jumara basedon large data set of Datta et al. (1996). However the adult
phragmocone diameter shows wider intraspecific variation.One specimen (JUM/JH/T/35) which is a part of adultphragmocone when reconstructed shows approximatediameter to be about 215 mm.
4.28. Dimensions
Specimen No. D U W H
JUM/JH/T/33 Phragmocone 153 12.6 44.8 87JUM/JH/T/34 Phragmocone 179 19 78 109JUM/JH/T/35 Phragmocone – 23.5 56.5 121.4
Genus Kamptokephalites Buckman, 1922Type species Kamptokephalites kamptus BuckmanKamptokephalites cf. etheridgei SpathFigs. 10.1–10.3
cf. 1890 Stephanoceras allied to S. lamellosus Etheridge,p. 175, pl. 29, Fig. 1.
1912 Macrocephalites keeuwensis d var. bifurcata
Boehm, p. 166, pl. 44, Figs. 4a–c.cf. 1928 Kamptokephalites etheridgei Spath, p. 200,
pl. 69, Fig. 3 [for Etheridge, 1890].1988 Macrocephalites? cf. etheridgei (Spath);
Westermann and Wang, p. 319, pl. 21, Figs. 3and 4 [m].
1988 Macrocephalites cf. etheridgei (Spath);Westermann and Callomon, p. 69, pl. 13,Figs. 4 and 5; pl. 14, Figs. 1–4; Text-Figs. 11,13, and 19c.
4.29. Material
The present collection includes only two microconchiatespecimens; one of them (JUM 1000) has been collectedfrom Bed 2 of Jhura and another (JUM/ETH/1) fromwhite limestone bed of Nara.
4.30. Description [m]
Shell depressed, evolute; during ontogeny shell inflationshows negative allometry, while degree of involutionincreases. At inner whorl (D = 15 mm) U/D = 0.33 andW/H = 1.92; at end-phragmocone (D = 72 mm) U/D =0.41 and W/H = 1.67 and at body chamber U/D = 0.35and W/H = 1.1. Whorl section changes from subovate tosemicircular in outline during ontogeny.
Flank short, curved, venter broad and rounded allthrough out ontogeny. Umbilicus fairly wide, umbilicalwall is vertical with a narrowly rounded margin.
Ribbing prominent, distant and are mostly biplicate.Ribbing in phragmocone is blunt crested and relativelycoarser while in body chamber they become sharplycrested. Primaries rise rursiradiately from the umbilicalslope and after crossing the umbilical margin projectforward and go straight up to the midflank where they
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 643

Autho
r's
pers
onal
co
py
bifurcate. Secondaries are also straight and continue overthe venter with a little forward projection. There are 12–13 primaries and 24–25 secondaries per half whorl inend-phragmocone and 14–15 primaries and 29–30 second-aries per half whorl in body chamber.
Aperture is missing. Small size, persistence of strongbiplicate ribs in the adult body chamber suggests that theforms are microconch.
Suture is not discernible.
4.31. Discussion
Kamptokephalites cf. etheridgei was so far reported fromtwo provinces only – the East Indies and the HimalayanTethys (Westermann and Wang, 1988). The present findis the first record of K. cf. etheridgei from the Indo-Mad-agascan Province. The present specimens come from theMiddle Bathonian horizons in two localities. Macrocepha-
lites triangularis is so far considered to be the oldest (Upper
Fig. 10. (1–3) Macrocephalites cf. etheridgei Spath (all · 1). 1–2(m) Lateral and apertural views of adult specimen with incompletely preserved body whorl;specimen No. JUM/ETH/1 from marly limestone bed, Nara. 3(m) Lateral view of adult phragmocone; specimen No. JUM 1000 from Bed 2, Jumara. (4–8)Prohecticoceras manjalense n. sp. (all · 1). 4(M) Cast; lateral view of a phragmocone specimen; note smooth inner whorls and stron falcoid ribbing with ventrolateral bullae; specimen No. JUM/0804/6 from marly limestone bed, Nara. 5(M) Lateral view of adult specimen; note persistence of strong primaries on theearly part of the body chamber and later smoothening; specimen No. JUM/0804/5 from marly limestone bed, Nara. 6A–B(m) Lateral view of the holotype (A)(external mould) and its cast (B) (JUM/0804/8). Note peristome with lappet. 7(m) Cast of Geological Survey of India Type 1919; an adult shell with bodychamber preserved; note strong primaries and ventro lateral nodes; from ‘Patcham Group’ of Waagen (1875), Nara (=Bed 1). 8(m) Lateral view of an adultshell with incompletely preserved body chamber (JUM/0804/12). (9–11) Oxycerites cf. orbis (Giebel) (all · 1). (9–11) Lateral, ventral and apertural views of animmature phragmocone; note sparse, distant secondaries and a mid-lateral ridge sharp venter; specimen No. JUM/O/1 from Bed 4, Jumara.
644 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
Bathonian) macrocephalitin species from Kutch. But thepresent discovery of K. cf. etheridgei extends the lowerrange of macrocephalitins down to the Middle Bathonian –a level from where Macrocephalites including M. cf.etheridgei have also been reported from East Indies(Westermann and Callomon, 1988). Westermann andCallomon (1988, pl. 13, Figs. 4 and 5; pl. 14, Fig. 1)described dimorphic pair within the present species.Macroconch retains ribs up to at least middle part of theadult body chamber – a phenomenon which is almostabsent in Macrocephalites s.st. we therefore follow Spathin describing the species under Kamptokephalites (for detailsee Jana, 2002).
Westermann and Callomon (1988) included Indoceph-alites sphaeroidalis Spath from Kutch in the synonymylist of ‘M. cf. etheridgei’. They considered it as macro-conch. The monotypic holotype of I. sphaeroidalis isfully septate. Although its inner whorls are inflated andcoarsely ornate like that of macroconchiate K. cf. ethe-
ridgei but its outer whorl is typical Indocephalites-likebeing more spheroidal and involute as in I. chrysoolithi-
cus Spath. This was also noticed by Westermann andCallomon (1988) and therefore they considered subse-quently I. sphaeroidalis as a nomen dubium. Moreover,these two species differ in stratigraphy, I. sphaeroidalis
comes from Lower Callovian horizon (see Sapth, 1933;Jana, 2002).
The species like K. misorobensis Spath, ‘Pleurocepha-
lites’ maevaranensis Besairie described by Collignon(1958) from the Bathonian beds (bearing M. triangularis)of Madagascar shows striking resemblance with thepresent species. In fact Westermann and Callomon(1988) synonymies them with the microconch of thepresent species. But K. amboromihatensis of Collignon(1958) from the same stratigraphic level is larger in size,appears to be more close to one Kutch species, i.e.,K. magnumbilicatus (Waagen).
Kamptokephalites lamellosum [m] (Sowerby, 1813)differs from the microconch of present species inbeing less inflated and finely, densely ribbed. K. dime-
rum [m] (Waagen) is less evolute and depressed aswell as finely ribbed than the microconch of the pres-ent species, Moreover, K. lamellosum and K. dimerum
come from stratigraphically higher level (LowerCallovian).
4.32. Dimensions
Specimen No. D U W H
JUM 1000(m)
Innerwhorl 15 5 11.5 6Phragmocone 31 9 21 11Phragmocone 44 16 26 16Phragmocone 55 18 33.5 20
JUM/ETH/1(m)
Body chamber 76.2 26 31.5 28End-phragmocone 57.5 19 – 21.5
Superfamily Haplocerataceae Zittel, 1884Family Oppeliidae Bonarelli, 1894Subfamily Hecticoceratinae Hyatt, 1900Genus Prohecticoceras, Spath, 1928Type species Ammonites retrocostatus Grossouvre, 1888Prohecticoceras manjalense n. sp.Figs. 10.4–10.8
1875 Oppelia (Oecotraustes) cf. serrigera Waagen, p. 57,pl. 10, Fig. 3.
1924 Oecotraustes sp. Spath, p. 19, Table I.1928 Paroecotraustes (Oecotraustes?) sp. Spath, p. 80,
pl. 9, Fig. 3.
4.33. Etymology
The species is named after the locality Manjal fromwhere the specimens have been collected.
4.34. Holotype
JUM/0804/8; collection Jadavpur University, Kolkata.
4.35. Material
The present study includes 12 macroconchs and 4microconchs collected from a marly limestone bed atManjal near Nara village, Kutch. Among the macro-conchs, 2 specimens retain original shell and others areexternal mould. Among the microconchs, 3 specimenshave original shell preserved, while one is an externalmould. Besides, the specimen (Geological Survey of IndiaType No. 1919) described by Waagen (1875) as Oppelia
(Oecotraustes) cf. serrigera has also been included inthe present study.
4.36. Diagnosis
Medium sized, flat flank, relatively evolute; strong pri-mary ribs with bullae at umbilical margin, secondary ribsshow ontogenetic changes being more rursiradiate in laterstage; sexually dimorph, microconchs are more evoluteand have strongly rursiradiate ribbing which continue upto the aperture which is lappeted.
4.37. Description [M]
Shell is evolute (U/D = 0.17–0.32), compressed with flat-tened flank. Medium sized, maximum observed diameterbeing 70 mm. Inner most whorls are smooth. Young whorlsat about diameter 11 mm bear ribs. Primaries rise prorsiradi-ately from the umbilical wall. They are coarse with roundedcrest and may form bullae near the umbilical margin on theadult phragmocone. Secondary ribs originate strongly rurs-iradiately at about one-third of whorl height from the umbil-ical margin and terminate at the ventrolateral margin wherethey form bullae or nodes. During later ontogeny they
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 645

Autho
r's
pers
onal
co
py
become straight to slightly rursiradiate or retain strong earlyrursiradiate pattern. Both primaries and secondaries con-tinue up to the early part of adult body chamber, but after-ward they become weak, distant and finally disappear fromthe last quarter of the body whorl, thus rendering the flanksmooth. Beside secondaries, there may be intercalatoriesand solitaries on outer whorl. Number of primaries and sec-ondaries per half whorl are 10 and 17, respectively.
Beginning of adult body chamber is marked by egres-sion of the whorl. Body chamber length occupies morethan half of the last whorl. Umbilical margin is gradualin phragmocone and prominent in body chamber. Venteris rarely observed, but appears to be wide, flattened withprominent median keel. Suture is not discernible.
4.38. Description [m]
Shell small, maximum diameter being 32 mm, evolute(U/D = 0.27–0.53). Coarsely ornate adult shell resemblesmacroconchiate phragmocone. Egressions of the umbilicalseam on body chamber, prominent ribbing and finally thepresence of lappet at the peristome indicate its adulthood.Body chamber occupies more than half of the last whorl.Both primaries and secondaries are strong; primaries aresharp and secondaries are coarse and blunt. Primaries risestrongly prorsiradiately from the umbilical wall which grad-ually changes from steep to inclined. They form ridge-likebullae on the inner flank and furcate afterwards to twosecondaries. Secondaries are strongly rursiradiate, bluntand end with ventrolateral nodes. Both primaries andsecondaries are sparse on the body chamber (6 and 13,respectively, per half whorl), but persist without showingany sign of attenuation up to the apertural margin. Interca-latories may alternate with secondaries. Peristome lappeted.
4.39. Discussion
Prohecticoceras appeared during the Early Bathonian(Hahn, 1968; Elmi, 1971; Torrens, 1987) and by the MiddleBathonian it spreads to many parts of the World. The old-est species, i.e., P. crassum Elmi (1967) continue from theZigzag Zone of Lower Bathonain up to the Middle Batho-nian Subcontratctus Zone in France where it co-occurswith P. ochraceum Elmi (1967). Prohecticoceras haugi
(Popovici-Hatzeg, 1905) has been reported from theBremeri Zone of the Middle Bathonian in Europe (seeWestermann and Callomon, 1988). Diversity of Prohecti-coceras continued up to the Upper Bathonian and manyspecies have been found through out Europe as well as inChile, Argentina (Riccardi et al., 1988, 1989; Sandovalet al., 1990) and Indonesia (Westermann and Callomon,1988) and now in India. The present species P. manjalense
resembles some species of other provinces.The present species closely resembles Prohecticoceras
retrocostatum (Grossouvre, 1888) from France in ribbingpattern. Both of them have bullae at the ventrolateral mar-gin on the phragmocone. But the present species differs in
having less pronounced chevron-like ribbing pattern andit is found only in some variants on adult body chamberof the macroconch. P. manjalense is also larger and has lessdense ribbing, i.e., about 17 secondaries per half whorl andis relatively more evolute.
The coeval P. ochraceum of France resembles closelyP. manjalense in similar degree of involution and size.Primaries in macroconchs of both species fade away nearthe adult aperture. However, in French species, primariesmay be enormously thick, which never appear in Kutchspecies. Besides, P. ochraceum has strongly angulirursiradi-ate ribs. The upper Bathonian species, P. blanazense Elmi(1967), which has been reported from France and Chile,has similar strong falcoid to chevron ribbing pattern ininner whorls to early adult phragmocone stage, but ribbingstrength falls very sharply since then and body chamber hassparse, blunt, concave secondaries. Moreover, it is large,strongly involute and has midflank ridge.
Prohecticoceras haugi of Europe is a large form withextremely strong rursiradiate secondaries on phragmoconebut body chamber is strongly involute with sparse, falcatesecondaries (Krystyn, 1972; Text-Fig. 9c and pl. 6,Fig. 1). Westermann and Callomon (1988; pl. 2, Figs. 6and 7) reported P. cf. haugi from Sula Island, Indonesiawhich has a phragmocone diameter of about 110 mm. Itdiffers from P. majalense chiefly in having Oxycerites-likesmall umbilicus in phragmocone and feeble primaries, largebody size and subtriangular whorl section.
Paroecotraustes species are generally characterized byevolute shell, thin prorsiradiate primaries on inner whorlsand rursiradiate secondaries which are distant and maycontinue up to the early part of the adult body chamber;secondaries do not form ventrolateral nodes; midflank inmajority of the cases is grooved or has smooth band (seeSapth, 1928; Elmi, 1967; Hahn, 1968). All are lappeted withrounded to squarish adult venter. They chiefly differ fromthe present microconch in lacking strong primaries andtuberculate secondaries. Comparison may be made withsome species. Paroecotraustes serrigerus described by Colli-gnon (1958; pl. 10, Figs. 54 and 54a) from the UpperBathonian of Madagascar resembles the microconch ofP. manjalense in flattened secondaries, fastigate venter,but adult whorl lacking primary ribs, peripheral nodesand has midflank ridge. P. (Nodiferites) sayni Elmi (1967;pl. 13, Fig. 9) from the Subcontractus Zone of Francehas similar chevron pattern of ribbing, but it differs fromthe Kutch microconch by possessing midflank groove.
Oxycerites (Paroecotraustes) aff. parvus [m] describedby Westermann and Callomon (1988, pl. 2, Fig. 4) fromSula Island, Indonesia, resembles P. majalense [m] inhaving similar evolute shell and ribbing pattern on theearly part of adult body chamber. However, it lacksprimaries on the inner whorls and ventrolateral tuber-cules all throughout the ontogeny. They comparedP. aff. parvus with O. (P.) collignoni Stephanov (1966,pl. 7, Figs. 1a and b) from the Upper Bathonian ofMadagascar. The Madagascan species has primaries in
646 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
the inner whorls like the Kutch form but it differs inhaving a mid-lateral groove.
4.40. Dimensions
Since most of the specimens are external moulds andwhorl sections are rarely observed, width of the speci-mens are not given.
Specimen No. D U H
JUM/0804/1 M Phragmocone 48.00 14.00 18.53JUM/0804/2 M Phragmocone 31.00 9.80 15.56JUM/0804/3 M Body chamber 38.77 12.90 16.20JUM/0804/5 M Body chamber 70.00 12 –
End-phragmocone 53.00 13.20 26.5JUM/0804/6 M End-phragmocone 43.21 12.00 19.10JUM/0804/7 M Phragmocone 34.00 12.55 13.49JUM/0804/8 M Partly preserved
body chamber44.00 17.00 14.49
JUM/0804/13 M Phragmocone 38.77 12.89 16.19JUM/0804/14 M Phragmocone 29.01 9.98 12.88JUM/0804/15 M Phragmocone 33.99 11.31 15.21JUM/0804/16 M Phragmocone 18.48 6.01 6.51JUM/0804/9 m Body chamber 22.00 8.00 9.54JUM/0804/10 m Young specimen 16.01 – 6.01JUM/0804/11 m Incompletely
preservedbody chamber
32.00 9.00 14.61
JUM/0804/12 m Incompletelypreservedbody chamber
31.5 9.00 9.34
GeologicalSurveyof India TypeNo. M 1919
Partly preservedbodychamber
43.00 17.00 13.50
Genus Oxycerites Rollier, 1909Type species Ammonites aspidoides Oppel, 1857Oxycerites cf. orbis (Giebel)Figs. 10.9–10.11
1846 Ammonites discus complanatus Quenstedt(Homonym Amm. discus Sowerby, 1813).
1852 Ammonites orbis Giebel.Non 1862 Ammonites aspidoides Oppel (Lectotype of
Rollier, 1911).1967 Oxycerites oppeli Elmi (Junior Synonym
of O. orbis).1982 Oxycerites orbis Giebel. – Dietl, p. 9–11,
pl. 3, Figs. 1–3.
4.41. Material
One internal mould of immature phragmocone (JUM/O/1) from the cream limestone bed of Jumara.
4.42. Description
Shell is fully septate, maximum preserved diameter is64 mm. The shell is discoidal, compressed (W/H = 0.5),highly involute (U/D = 0.13 at diameter 62 mm) with oxy-conic whorl section. Both outer and inner flanks straightto gently convex with maximum width of the whorl liesbelow the midflank. Umbilical margin sharp with steepwall. Venter is extremely narrow, keeled and weaklyshouldered.
Inner flank smooth except dense growth lirae which risesfrom the umbilical wall slightly rursiradiately. Secondariesare weakly concave, rursiradiate, relatively frequent in theinner phragmocone, but their intervals increases duringontogeny. They continue till to the preserved end. Numberof secondaries in the preserved last whorl is 10. A faintridge on the midflank continues till to the end of the pre-served specimen.
4.43. Discussion
Oxycerites orbis (Giebel) is a zonal index for the UpperBathonian of north west Europe and was not previouslyrecorded from the Indo-Madagascan areas. Its closestcounterpart in France is described as O. oppeli Elmi,1967 which both Dietl (1982) and Westermann and Callo-mon (1988) considered as a junior synonym of O. orbis.O. oppeli Elmi (1967, pl. 1, Figs. 1–5) was rightly includedwithin O. orbis by Dietl (1982). The present species has sim-ilar concave sencondaries which become distant in the laterpart of the phragmocone. Both of them have mid-lateralridge.
Oxycerites sulaensis Westermann and Callomon (1988)has similar ornamentation at the same diameter, but it ismore involute (U/D = 0.09) and the mid-lateral ridgeappear at the end of the phragmocone.
4.44. Dimensions
Specimen No. D U W H
JUM/O/1 Phragmocone 62.0 8.0 18.0 32.0
5. Palaeobiogeography
Palaeobiogeographical distributions of ammonites areconstrained by many factors including oceanic surface cur-rent, larval duration, latitudinal gradient etc. The openingup of Kutch basin took place during the?Bajocian-Batho-nian times as a result of the Gondwana fragmentation (Bis-was, 1991). Mainland of Kutch includes older sedimentsextending up to the Middle Bathonian. The older sedi-ments are also reported from further north in island areas.A few records of sporadic occurrences of ammonites werereported from these places which includes Leptosphinctes
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 647

Autho
r's
pers
onal
co
py
from the Bajocian; Micromphalites; Clydoniceras sp. andGracilisphinctes arkelli Collignon (1958) from MiddleBathonian sediments (see Singh et al., 1982; Singh et al.,1983; Jaitly and Singh, 1984). These are all Tethyan formsfound in many areas of Europe. Besides some of them arealso found from Madagascar. Collignon (1958) reportedMicromphalites, Clydoniceras from the different localitiesand mentioned their occurrence with Macrocephalites,but Westermann and Callomon (1988) doubted that theymay be stratigraphically mixed up. Bathonian assemblagesare also found in Beluchistan, Pakistan and it includesChoffatia (Homoeoplanulites) beluchistanensis (Noetling)and Bullatimorphites (Kheraiceras) cf. or aff. bullatus
(D’Orbigny) (=? hannovaranus) (Roemer), (Westermannand Callomon, 1988) and Clydoniceras beluchistanensis
(Spath) (see Noetling, 1897; Spath, 1933 and Arkell,1956). The Bathonian ammonite genera found in differentregions within the Indo-Madagascan Provinces are shownin Table 3a. Therefore, it appears that Indo-Madagascanregions include many Tethyan ammonites during theBathonian but Kutch was most diverse. Close inspectionincluding species level analysis reveals that most of the gen-era have come from different provinces as soon as theKutch basin opened up. K. cf. etheridgei and M. bifuracatus
transient intermedius which were restricted only to the WestPacific Faunal Provinces arrived at the same time whenEuropean Procerites and Choffatia also invaded Kutch.O. cf. orbis and Epistrenoceras sp. reached Kutch duringthe Late Bathonian global marine transgression (Haqet al., 1987, 1988). This indicates the establishment of wellconnected sea routs with other provinces. Like many othertaxa, e.g., corals (see Pandey and Fursich, 2001), brachio-pods (Mukherjee et al., 2003), gastropods (Das et al.,1999; Jaitly et al., 2000; Das, 2003) and nautiloids (Halder,
2000, 2002), ammonites also show species level endemicityonce the immigrant genera made inroads to the Kutchbasin. The evolution of endemic species of Macrocephaliti-nae during the Late Bathonian–Lower Callovian timeconstitutes the characteristic assemblage which is uniqueonly to the Indo-Madagascan Province. Many genera,e.g., Indocephalites Spath, Kamptokephalites evolvedvery rapidly during the Late Bathonian in Kutch (Jana,2002).
Likewise perisphinctid genera underwent similar specia-tion events and developed endemic assemblages, e.g., Pro-
cerites hians whose stratigraphic lower limit has beenextended down to the Middle Bathonian in the presentstudy, resembles several forms of Europe (Callomon,1993). It soon speciated to new genus Sivajiceras whichcontinued and diversified during the Callovian in Kutchand gave rise to a typical endemic assemblage, i.e., Sivajic-
eras–Obtusicostites lineage (Spath, 1927–1933; Callomon,1993). Prohecticoceras also shows similar trend. It althoughappeared in the Early Bathonian, spread to many parts ofthe world including Kutch (personal observation) duringthe Middle Bathonian and became most diverse duringthe Late Bathonian. The Kutch species, Prohecticoceras
manjalense n. sp. is again not only endemic it is the mostevolute form of the genus.
Thus it appears from the stratigraphic and palaeobioge-ographic distribution patterns that immediately after theformation of Kutch basin, immigrant faunas of differentprovinces and latitudes invaded the Kutch sea. West Paci-fic, Indonesia was situated in the same palaeosubtropic(30�S) similar to Kutch and therefore migration of macro-cephalitin taxa faced no temperature gradient during theexcursion. Westermann and Wang (1988) also reportedM. etheridgei from the Himalayan Tethys in Southern
Table 3aBathonian ammonite genera present in different regions of the Indo-Madagascan Province
Kutch (present studyand references herein)
Madagascar (Arkell, 1956;Collignon, 1958; Thierry, 1978and Westermann and Callomon, 1988)
Baluchistan (Arkell,1956; Westermann andCallomon, 1988)
Himalayas (Westermannand Wang, 1988; Enay andCariou, 1997, 1999)
Leptosphinctes x – – –Micromphalites x x – –Clydoniceras x x x –Gracilisphinctes x x – –Macrocephalites x x x xProhecticoceras x – – xOxycerites x x – xBullatimorphites x x – –Kheraiceras x – x –Choffatia x – x xEpistrenoceras x x – –Epimorphoceras x – – –Procerites x – – –Sivajiceras x – – –Indocephalites x x x –Kamptokephalites x x – xCadomites – x – –Wagnericeras – x – x
x, present.
648 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
Tibet which also lay close to the Kutch in the samelatitude. Recently Enay and Cariou (1997, 1999) men-tioned but not described Upper Bathonian assemblagesfrom Nepal. They include Tethyan forms like Prohecti-
coceras, Homoeoplanulites (Parachoffatia) and the latestMiddle Bathonian Wagnericeras. Indonesian Macroceph-
alites bifurcatus and M. apertus mantataranus group ofspecies have been extended westward up to Nepal andwe, in the present endeavour extended some of them(M. cf. bifurcatus and M. manatataranus transient inter-
medius) further up to Kutch. Enay and Cariou (1997,1999) however, mentioned M. triangularis erroneouslyfrom the Early Callovian. M. triangularis is a zonal spe-cies of the Upper Bathonian and co-occurring with Epis-
trenoceras in Kutch and Madagascar.We have made an attempt to establish faunal correlation
at the genus level among different areas using Simpson sim-ilarity coefficient [we have not used Jaccard similarity coef-ficients since faunal diversity among these areas variesgreatly; see also Cecca (2002)]. High Simpson coefficient(see Tables 3a and 3b) suggest the faunal homogeneitywithin the province.
Biogeographic distribution of Bathonian macrocephalit-ids depicts an interesting picture. They are distributed onlyin Tethys and East Pacific Indonesia (see Enay and Cariou,1999, Fig. 4). The eurycephalitin genus Xenocephalites
Spath contrasted macrocephalitin distribution. The oldestmacrocephalitid assemblage had been reported from Indo-nesia where they co-occur with both sphaeroceratids andeurycephalitids. Thierry (1978) believed that macrocepha-litids evolved from eurycephalitids. While Westermannand Callomon (1988) held sphaeroceratids as purportedancestor. Recently Jana (2002) has supported Thierry(1978) based on cladistic analysis. Anyway, there is consen-sus that macrocephalitids originated in Indonesia and latermigrated to Europe, Himalayas, Kutch and Madagascar.Eurycephalitids (Xenocephalites) made excursion at thesametime toward southern part of Gondwana includingNew Zealand and Australia. This indicates ammonitemigration and distribution were not only constrained byoceanographic and other inorganic factors but also byphysiological differences (also see Cecca, 1999).
Acknowledgements
Dr. Fujikawa and the anonymous reviewer had criticallyreviewed the manuscript and provided valuable sugges-tions. We are thankful to G.S. Roy for extending logisticsupport during field work. The help rendered by the local
administration especially the border security outpost ofNara and the local people is also acknowledged. We alsoexpress our thanks to S. Shome for field and laboratoryassistance. Financial support was provided to P.R. byCSIR. S.B. wishes to thank the Centre of Advanced Stud-ies, J.U. for providing financial support.
References
Ager, D.V., 1984. The Nature of the Stratigraphical Record. MacmillanPublication Ltd., Hong Kong, 122p.
Arkell, W.J., 1951–1958. A monograph of English Bathonian Ammonites.Palaeont. Soc., Lond., 104–112 (1–7), 264p.
Arkell, W.J., 1956. Jurassic Geology of the World. Oliver and Boyd Ltd.,London, 757p.
Arkell, W.J., Kummel, B., Wright, C.W., 1957. Mesozoic Ammonoidea.In: Moore, R.C., Teichert, C. (Eds.), Treatise on invertebratepaleontology. Part L. Mollusca. Geological Society of America andUniversity of Kansas Press, Boulder, Colorado and Lawrence, Kansas,pp. L80–L129.
Bardhan, S., Datta, K., Jana, S.K., Pramanik, D., 1994. Dimorphism inKheraiceras Spath from the Callovian Chari Formation, Kutch, India.Journal of Paleontology 68, 287–293.
Bardhan, S., Sardar, S.K., Jana, S.K., 2002. The Middle JurassicKheraiceras Spath 1924 from the Indian Subcontinent. In: Summes-berger, H., Histon, K., Daurer, A. (Eds.), Cepahalopods – Present andPast. Abhandlungen der Geologischen Bundesanstalt, 57, 265–277.
Bentz, A., 1928. Upper Strenoceratina and Garantiana, in particular fromthe middle Dogger of Bielefeld. Jahrb. Preuss. Geol. Landesanst 19,138–206, pls. 14–19.
Biswas, S.K., 1991. Stratigraphy and sedimentary evolution of theMesozoic basin of Kutch, western India. In: Tandon, S.K., Pant,C.C., Casshyap, S.M. (Eds.), Sedimentary Basins of India: TectonicContext. Gyanodoy Prakashan, Nainital, pp. 74–103.
Blake, J.F., 1905. Fauna of the Cornbrash. Monograph of Palaeontolo-gical Society, London, 106p, 9 pls.
Boehm, G., 1912. Contributions to the Geology of Indian Niederland. I,pt. V. Lower Callovian. Palaeontogrphica Supplement 3, 123–179, pls.32–44.
Bonarelli, G., 1894. Hecticoceras novum genus Ammonidarum. Bulletin ofMalacological Society, Italy 18, 73–108.
Buckman, J. 1844. Descriptions of the new species of fossils, In:Murchison, R.I. (Ed.), Outline of the geology of the neighbourhoodof Cheltenham, new ed., pp. 93–106, pls. 1–13.
Buckman, S.S., 1909–1930. Type Ammonites. Wheldon and Wesley,London, 790p.
Callomon, J.H., 1971. On the type species of Macrocephalites Zittel, 1884and the type species of Ammonites macrocephalus Schlotheim, 1813.Palaeontology 14, 114–130.
Callomon, J.H., 1980. Macrocephalites (Stutner MS) Zittel, 1884, andAmmonites macrocephalus Schlotheim 1813. Palaeontology 14, 114–130.
Callomon, J.H., 1993. On Perisphinctes congener Waagen, 1875, andthe age of the Patchman Limestone in the Middle Jurassic ofJumara, Kutch, India. Geologische Blatter fur Nordost-Bayern 43,227–246.
Cecca, F., 1999. Palaeobiogeography of Tethyan ammonite during theTithonian (latest Jurassic). Palaeogeography, Palaeoclimatology,Palaeoecology 147, 1–37.
Cecca, F., 2002. Palaeobiogeography of Marine Fossil InvertebratesConcepts and Methods. Taylor and Francic, pp. 72–103.
Collignon, M., 1958. Atlas des fossiles caracteristiques de Madagascar.Fascicule 1, 2, pls 1–33. Service Geologique Tananarive.
Cox, L.R., Arkell, W.J., 1948–1950. A survey of the Mollusca of the Greatoolite Series, Paleontographical Society (London), 13, 105p.
Table 3bSimpson similarity coefficients of Bathonian ammonite genera betweenarea pairs in different regions of the Indo-Madagascan Province
Kutch Madagascar Baluchistan
Madagascar 0.81Baluchistan 1 0.60Himalayas 0.83 0.66 0.40
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 649

Autho
r's
pers
onal
co
py
Das, S.S., Lahiri, T.C., Bardhan, S., 1999. The Late Bathonian gastropodfauna of Kutch, western India – a new assemblage. PalaeontologicalResearch 3, 268–286.
Das, S.S., 2003. New assemblage of Mesozoic gastropod faunas of Kutch,western India – a study of systematics, palaeobiogeography andevolution. Unpublished Ph.D. thesis, Jadavpur University, Kolkata,India, 161p.
Datta, K., 1992. Facies, fauna and sequence: an integrated approach in theJurassic Patcham and Chari Formations, Kutch, India. UnpublishedPh.D. thesis, Jadavpur University, Kolkata, India, 167p.
Datta, K., Bhaumik, D., Jana, S.K., Bardhan, S., 1996. Age, ontogenyand dimorphism of Macrocephalites triangularis Spath – the oldestmacrocephalitid ammonite from Kutch, India. Journal of theGeological Society of India 47, 447–458.
Dietl, G., 1982. Das wirkliche Fundniveau von Ammonites aspidoides
OPPEL (Ammonoidea, Mittl. Jura) am locus typicus. StuttgartBeitrage zur Naturkunde, Serie B 87, 1–21.
Dietl, G., Callomon, J.H., 1988. Der Orbis-Oolith (Ober-Bathonium,Mittl. Jura) von Sengenthal/Opf., Frank. Alb, und seine Bedeutung furdie Korrelation und Gliederung der Orbis-Zone. Stuttgarter Beitragezur Naturkunde, Series B 142, 1–31.
Elmi, S., 1967. Le Lias superieur et le Jurassique Moyen de L’Ardeche.Documents des Laboratories de Geologie de la Faculte des Sciences deLyon, 845 p.
Elmi, S., 1971. Les faunes a Prohecticoceras (Oppeliidae, Ammonitina) duBathonien inferieur et moyen des confins Algero-marocains – Geobios,4/4, Lyon, 243–264.
Enay, R., Cariou, E., 1997. Ammonite faunas and palaeobiogeography ofthe Himalayan belt during the Jurassic: initiation of a Late Jurassicaustral ammonite fauna. Palaeogeography, Palaeoclimatology andPalaeoecology 134, 1–38.
Enay, R., Gauthier, H., Trevisan, M., Berton, J.-B., Brivet, L., Brodbeck,J.-L., Demaizieres, Donie, J.-F.P., Fourel, A., Trehour, M., 2001. LesMicromphalites (Ammonitina) du Bathonien inferieur de la Nievre(France): installation sur la marge europeenne de la Tethys de formessud-tethysiennes d’origine arabique et description d’un neotype de M.
busqueti (de Gross). Revue de Paleobiologie 20, 503–524.Etheridge, R., 1890. Our present knowledge of the palaeontology of New
Guinea. Records of the Geological Survey of New South Wales 1, 172–179.
Enay, R., Cariou, E., 1999. Jurassic ammonite fauna and their bearing onthe palaeobiogeography of the Himalayan belt. Journal of Asian EarthSciences 17, 829–848.
Fursich, F.T., Oschmann, W., 1993. Shell beds as tools in basin analysis:the Jurassic of Kachchh, Western India. Journal of the GeologicalSociety of London 150, 169–185.
Fursich, F.T., Pandey, D.K., Callomon, J.H., Oschmann, W., Jaitly,A.K., 1994. Contribution to the Jurassic of Kachchh, Western India.II. Bathonian stratigraphy and depositional environment of theSadhara Dome, Patchchham Island. Beringeria 12, 95–125.
Ghose, D.N., 1967. The Jurassic Rhynchonellidae from Jumara, Nara andKeera domes, Kutch western India. Unpublished Ph.D. thesis,Jadavpur University, Kolkata, India, 200 p.
Grossouvre, A.de., 1888. Studies on the Bathonian stage. Bulletin of theGeological Society of France 16 (3), 366–401.
Grossouvre, A.de., 1907. La variabilite de l’espece chez les ammonites.Sixieme Congres de l’Association Franc-Comtoise, tenue a Vesoul1906, 3–10.
Halder, K., 2000. Diversity and biogeographic distribution of Jurassicnautiloids of Kutch, India during fragmentation of Gondwana.Journal of African Earth Sciences 31, 175–185.
Halder, K., 2002. Jurassic nautiloids of Kutch, western India: a study ontheir systematics, palaeotology and evolution. Unpublished Ph.D.thesis, Jadavpur University, Kolkata, India, 187p.
Hahn, W., 1968. Die Oppelidae Bonarelli und Haploceratidae Zittel(Ammonoidea) des Bathoniums (Brauner Jura) im sudwestdeutschenJura. Jahreshefte des Geologischen Landesamtes Baden – Wurttem-berg 10, 7–72.
Haq, B.U., Hardenbol, J., Vail, P.R., 1987. Chronology and fluctuatingsea-levels since the Triassic. Science 235, 1156–1166.
Haq, B.U., Hardenbol, J., Vail, P.R., 1988. Mesozoic Cenozoicchrnostratigraphy and cycles of sea-level change. S.E.P.M. specialpublication, 42, pp. 71–108.
Hyatt, A., 1900. Cephalopoda. In: Zittel, K.A. (Ed.) Textbook ofPalaeontology, 1st English ed. Eastman, pp. 502–604.
Jablonsky, D., 1996. Body size and macroevolution. In: Jablonsky, D.,Erwin, D.H., Lipps, J.H. (Eds.), Evolutionary Palaeontology. TheUniversity of Chicago Press, Chicago, pp. 256–289.
Jain, S., Callomon, J.H., Pandey, D.K., 1996. On the earliest knownoccurrence of the Middle Jurassic ammonite genus Reineckeia in theUpper Bathonian of Jumara, Kachchh. Western India, Palaontolo-gische Zeitschrift 70, 129–143.
Jaitly, A.K., Singh, C.S.P., 1984. On the Bathonian (Middle Jurassic)ammonites Micromphalites Buckman and Gracilisphinctes Buckmanfrom Kachchh, Western India. Geological Magazine (Great Britain)121 (4), 319–321.
Jaitly, A.K., Szabo, J., Fursich, F.T., 2000. Contributions to the Jurassicof Kachchh, Western India VII. The gastropod fauna. Part I.Pleurotomarioidea, Fissurevoidea, Trochoidea and Eucycloidea. Ber-ingeria 27, 31–61.
Jana, S.K., 2002. Macrocephalitinae and Eucycloceratinae of the familySphaeroceratidae (Ammonoidea) and other ancillary taxa from theMiddle Jurassic of Kutch, western India. Unpublished Ph.D. thesis,Jadavpur University, Kolkata, India, 163 p.
Jana, S.K., Bardhan, S., Sardar, S.K., 2000. Kheraiceras Spath(Ammonoidea) – new forms and records from the Middle Jurassicsequence of the Indian Subcontinent. Paleontological Research 4, 205–225.
Jana, S.K., Bardhan, S., Halder, K., 2005. Eucycloceratin ammonitesfrom the Callovian Chari Formation, Kutch, India. Palaeontology 48(4), 1–49.
Kayal, A., Bardhan, S., 1998. Epistrenoceras Bentz (Ammonoidea) fromthe Middle Jurassic of Kutch (western India): a new record and itschronostratigraphic implication. Canadian Journal of Earth Sciences35, 931–935.
Kitchin, F.L., 1903. The Jurassic fauna of Cutch, the Lamellibranchiata;No.1, Genus Trigonia. Memoirs of the Geological Survey of India,Palaeontologia Indica, series 9, 3(2), 1–122.
Krishna, J., Westermann, G.E.G., 1985. Progress report on the MiddleJurassic ammonite Zones of Kachchh, West India. News Stratigraphy14, 1–11.
Krishna, J., Westermann, G.E.G., 1987. Faunal associations of theMiddle Jurassic ammonite genus Macrocephalites in Kachchh,Western India. Canadian Journal of Earth Sciences 24, 1570–1582.
Krishna, J., Cariou, E., 1993. The Tethyan Macrocephalitinae:Evolutionary Environmental and dispersal Strategies. Geobios 15,217–226.
Krystyn, L., 1972. Die Oberbajocium- und Bathonium- Ammoniten derKlaus-Schichten des Steinbruches Neumuhle bei Wien (Ostrreich).Ann. Naturhist. Mus. Wien 76, 195–310.
Lemoine, P., 1910–1911. Paleontologie de Madagascar 8. Ammonites dueJurassique superieur du Cercle d’Analava (Madagascar). Annels dePaleontologie, 5, 1910, 137–168, 5 pls., and 6, 1911, 45–64, 3 pls.
Mangold, C., 1970. Stratigraphie des etages bathonien et callovien du Jurameridional. Documents des Laboratoires de Geologie de la Faculte desSciences de Lyon 41, 1–376.
Mangold, C., 1984. Report of the Bathonian Working Group. In:Michelsen, O., Ziess, A., (Eds.) International Symposium on JurassicStratigraphy, Erlangen (I.U.G.S.), 1, pp. 67–76.
Mukherjee, D., 2002. Jurassic terebratulides and other ancillary taxa fromKutch, Western India – A study of systematics and evolutionarypalaeobiology. Unpublished Ph.D. thesis, Jadavpur University, Kolk-ata, India, 163 p.
Mukherjee, D., Bardhan, S., Datta, K., 2003. The terebratulid Kutch-
ithyris (Brachiopoda) from the Jurassic Sequence of Kutch, WesternIndia Revisited. Palaeontological Research, Japan 7 (2), 11–128.
650 P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651

Autho
r's
pers
onal
co
py
Neumayr, M., 1875. Die Ammoniten der Kreide und die Systematik derAmmonitiden. Zeitschr. Deutsch. Geol. Ges. 77, 854–942.
Noetling, F., 1897. Fauna of the Kellaways of Mazar Drik. Pal. Ind., Ser.16, 1, pp. 22.
Oppel, A., 1854–1858. Die Juraformation England, Frankreichs und dessudwestlichen Deutschlands. Wurttemberg Natuwiss. Jb., Stuttgart,12–14, 857p.
Pandey, D.K., Agrawal, S.K., 1984. On two new species of the MiddleJurassic ammonite genus Clydoniceras Blake from Kachchh, westernIndia. N. Jb fur Geol. and Palaont., Mh. 6, 321–326.
Pandey, D.K., Singh, C.S.P., Agrawal, S.K., 1984. A note on the newfossil finds from Gora Dongar, Pachchham ‘‘Island’’, district Kachchh(Gujarat). The Journal of Scientific Research, Banaras HinduUniversity 34 (1), 299–310.
Pandey, D.K., Westermann, G.E.G., 1988. First record of BathonianBullatimorphites (Jurassic Ammonitina) from Kachchh, India. Journalof Paleontology 62 (1), 148–150.
Pandey, D.K., Callomon, J.H., 1995. Contributions to Jurassic ofKachchh, Western India. III. The Middle Bathonian ammonitefamilies Clydoniceratidae and Perisphinctidae from Pachchham Island.Beringeria 16, 125–145.
Pandey, D.K., Fursich, F.T., 2001. Environmental distribution ofscleractinian corals in the Jurassic of Kachchh, western India. Journalof the Geological Society of India 57, 479–495.
Popovici-Hatzeg, V., 1905. Les cepahalodes du Jurassique moyen duMont Strunga. Memoire Societe Geologie France, Paleontologie, 35,27p.
Riccardi, A.C., Westermann, G.E.G., Elmi, S., 1988. The Bathonian-Callovian ammonite zones of the Argentine-Chilean Andes. In: Rocha,R.B., Soares, A.F. (Eds.), Proceedings of the 2nd InternationalSymposium on Jurassic Stratigraphy, 1987. Instituto Nacional deInvesticao Cientifica, Lisoba, pp. 347–358.
Riccardi, A.C., Westermann, G.E.G., Elmi, S., 1989. The Middle JurassicBathonian-Callovian ammonite zones of the Argentine-Chilean Andes.Geobios 22, 553–597.
Roemer, J., 1911. Die Fauna der Aspidoides-Schichten von Lechstedt beiHildeshein. W. Riemschneider ed., Hannovre, 64p.
Rollier, L., 1909. Phylogenie des principaux genres d’Ammonoides del’Oolithique (Dogger) et de l’Oxfordien. Arch. Sci. Phys. Nat. Geneve27, 611–623.
Rollier, L., 1911. Oolitic facies of the Dogger, Jura and in its vicinity.Memoire of Schnyder, Wartensee Foundation, 5, 352p.
Rudra, P., Bardhan, S., Shome, S., in press. Phylogeny of the three speciesof the existing subgenus Eselaevitrigonia (Bivalvia) from the LateJurassic–Early Cretaceous of Kutch, India, and paleobiogeographicconstraints. Journal of Paleontology.
Salfeld, H., 1921. Kiel-und Furchenbildmg auf der Schlenaussenseite derAmmonoiden in inhrer Bedeutung fur die Systematik und Festlegungvon Biozonen. Centralbulletin fur Mineralogie, Geologie, 11, 343–347.
Sandoval, J., Westermann, G.E.G., Marshall, M.C., 1990. Ammonitefauna, stratigraphy and ecology of the Bathonian-Callovian (Jurassic)
Tecocoyunca Group, South Mexico. Palaeontographica 210 (4–6),93–149.
Schindewolf, O.H., 1925. Systematic description of the Perisphinctids.Neus Jahrbuch for Mineralogy etc. Beil.-Bd. L11, 309–343.
Schloenbach, U.V., 1865. Beitrage zur Palaeontologie der Jura undKreideformation im nordwestlichen Deutschlands. Palaeontographica,Stuttgart 13, 147–192.
Schlotheim, E.F.v., 1813. Beitrage Zur Naturgeschichte der versteinerun-gen in geognostischer Hinsicht. In: Leonhard ‘Taschenbuch fur diegesammte Mineralogie, etc.’ Jahrg 7, Abt., 1, pp. 68.
Siemiradzki, J.V., 1898–1999. Monographic description of the ammonitegroup Perisphinctidae. Palaeontographia, 45, 69–296, pls. 20–25.
Singh, C.S.P., Jaitly, A.K., Pandey, D.K., 1982. First report of someBajocian-Bathonian (Middle Jurassic) ammonoids and the age of theoldest sediments from Kachchh, W. India. Newsletters on Stratigraphy11 (1), 37–40.
Singh, C.S.P., Pandey, D.K., Jaitly, A.K., 1983. Discovery of Clydoniceras
Blake and Gracilisphinctes Buckman (Bathonian-Middle JurassicAmmonites) in Kachchh, Western India. Journal of Paleontology 57(4), 821–824.
Sowerby, J.De.C., 1813. Mineral Conchology. Transaction of theGeological Society London 7, pls. 1–337.
Spath, L.F., 1927–1933. Revision of the Jurassic cephalopod fauna ofKachh (Cutch). Palaeontologia Indicia, Geological Survey of India,New Series 9, Memoir 2, 1–945.
Stephanov, J., 1966. The Serrigerus problem in the Jurassic Ammonoidfamily Oppelidae Bonarelli, 1894. C.R. Acad. Bulgare Sc., Sofia,827–829.
Torrens, H.S., 1980. Bathonian correlation chart. In: Cope, J.C.W. (Ed.)A correlation of Jurassic rocks in the British Isles, part two: Middleand Upper Jurassic, Geological Society of London, Special Report, 15,21–45.
Torrens, H.S., 1987. Ammonites and stratigraphy of the Bathonian rocksin the Digne-Barreme area (South-Eastern France, Dept. Alpes deHaut Provence). Bull. Soc. Paleont. It. 26, 93–108.
Thierry, J., 1978. Le genre Macrocephalites au Callovien inferieur(ammonites Jurassique moyen). Memoire de Geologie de Universitede Dijon, 4, 490p.
Waagen, W., 1875. Jurassic Fauna of Kutch, the Cephalopoda.Palaeontologia Indica, Geological Survey of India, Series 9, Memoir1, pp. 1–247.
Westermann, G.E.G., Callomon, J.H., 1988. The Macrocephalitinae andassociated Bathonian and Early Callovian (Jurassic) Ammonoids ofthe Sula Islands and New Guinea. Palaeontographica, Abteilung A203, 1–90.
Westermann, G.E.G., Wang, Yi-Gang, 1988. Middle Jurassic Ammonitesof Tibet and the age of the Lower Spiti Shales. Palaeontology 1,295–339.
Zieten, K.A.v., 1830–1833. Die Versteinerungen Wurttemberg. Verlag andLithographie der Expedition des Werkes unserer Zeit, 102pp., 72 pls.
Zittel, K.A., 1884. Hand Book of Palaeontology, 1, part-2, 329–522.
P. Roy et al. / Journal of Asian Earth Sciences 30 (2007) 629–651 651




















