
EUCYCLOCERATIN AMMONITES FROM THE CALLOVIAN CHARI FORMATION, KUTCH, INDIA
42
EUCYCLOCERATIN AMMONITES FROM THE CALLOVIAN CHARI FORMATION, KUTCH, INDIA by S. K. JANA* , S. BARDHAN* and K. HALDER *Department of Geological Sciences, Jadavpur University, Kolkata 700032, India; e-mail: [email protected] Department of Geology, Durgapur Government College, Durgapur 713214, India Typescript received 29 January 2002; accepted in revised form 24 June 2004 Abstract: The subfamily Eucycloceratinae Spath, 1928, belonging to the family Sphaeroceratidae, is an important Indo-Madagascan faunal element and is reported here from the Callovian of Kutch, India. Previously the subfamily was considered to consist of 14 morphospecies placed in four morphogenera. The present study is based on re-examina- tion of the type material and more than 350 new speci- mens, collected with secure stratigraphical control, from sections in mainland Kutch. It reveals that the subfamily includes only two highly variable monospecific genera: Eucycloceras Spath, 1924 and Idiocycloceras Spath, 1928. Dimorphism in Eucycloceratinae, which was not previously recognized, is firmly established. Cladograms have been constructed, based on numerous morphological characters, which show relationships among different subfamilies of the Sphaeroceratidae and genera of the Macrocephalitinae and Eucycloceratinae. The palaeobiogeography and evolu- tion of the Eucycloceratinae are discussed in the light of the new data. Key words: Middle Jurassic, Ammonoidea, Eucycloceras, Indo-Madagascan Province, phylogeny, cladistics. Several exhaustive monographic treatments illustra- ting a spectacular range of intraspecific morphological variability are now available for Jurassic (e.g. Westermann and Calloman 1988) and Cretaceous (e.g. Klinger and Kennedy 1989) ammonite species. A direct result of these studies is the drastic reduction of many previously estab- lished morphospecies or morphogenera into a single bio- species. This has helped remove pseudodiversity patterns, and prompted reconsideration of many evolutionary trees. We consider the studies of Callomon (1985) to be an excellent example of such work. Based on his ideas, we have revisited the subfamily Eucycloceratinae, which con- stitutes an important faunal element of the Callovian of the Indo-Madagscan Faunal Province. We have shown that failure to recognize the great intraspecific variability and sexual dimorphism within the eucycloceratin popula- tion led early workers to create a plethora of species. These essentially are morphospecies or morphogenera, and have little evolutionary significance. The present investigation has not only unearthed numerous additional specimens, but also was undertaken within a secure strati- graphical framework. It reveals that many of the previ- ously described species are essentially contemporaneous, being restricted to within a single faunal horizon, or at best a subzone. They show continuous horizontal variation. However, the population may be sharply divi- ded due to sexual dimorphism, an aspect overlooked by early workers who failed to recognize this feature and considered sexual variants as distinct genera. The subfamily Eucycloceratinae of the family Sphaero- ceratidae has not been studied in detail since Spath’s (1928–33) work. He incorporated the four genera Notho- cephalites, Eucycloceras, Subkossmatia and Idiocycloceras within it. They are endemic to the Indo-Madagascan Faunal Province and, as the new stratigraphical data reveal, span a brief period of geological time from the lat- est Early Callovian to early Middle Callovian. In Kutch the appearance of this subfamily is coeval with that of the Reineckeinae, at a time when the ancestral Macrocephalit- inae were on the verge of regional extinction. Spath’s four eucycloceratin genera include 14 species. Most of these species were based on single specimens and the holotypes are mostly fragmentary or preserved with incomplete body-chambers. They are re-examined here along with over 350 additional specimens. Detailed mor- phological and morphometrical analyses reveal that, in many cases, type species fall within the range of variability of a single biospecies. Two of the four existing genera (Eucycloceras and Idiocycloceras) survive close scrutiny and form monospecific assemblages. The former is found in the lower (older) stratigraphical horizon, but has strati- graphical overlap with the latter. Most of the smaller spe- cies of Subkossmatia, which closely resemble the inner [Palaeontology, Vol. 48, Part 4, 2005, pp. 883–924] ª The Palaeontological Association 883
-
Upload
bioinfo-presiuniv -
Category
Documents
-
view
1 -
download
0
Transcript of EUCYCLOCERATIN AMMONITES FROM THE CALLOVIAN CHARI FORMATION, KUTCH, INDIA

EUCYCLOCERATIN AMMONITES FROM THE
CALLOVIAN CHARI FORMATION, KUTCH, INDIA
by S. K. JANA* , S. BARDHAN* and K. HALDER�
*Department of Geological Sciences, Jadavpur University, Kolkata 700032, India; e-mail: [email protected]�Department of Geology, Durgapur Government College, Durgapur 713214, India
Typescript received 29 January 2002; accepted in revised form 24 June 2004
Abstract: The subfamily Eucycloceratinae Spath, 1928,
belonging to the family Sphaeroceratidae, is an important
Indo-Madagascan faunal element and is reported here from
the Callovian of Kutch, India. Previously the subfamily was
considered to consist of 14 morphospecies placed in four
morphogenera. The present study is based on re-examina-
tion of the type material and more than 350 new speci-
mens, collected with secure stratigraphical control, from
sections in mainland Kutch. It reveals that the subfamily
includes only two highly variable monospecific genera:
Eucycloceras Spath, 1924 and Idiocycloceras Spath, 1928.
Dimorphism in Eucycloceratinae, which was not previously
recognized, is firmly established. Cladograms have been
constructed, based on numerous morphological characters,
which show relationships among different subfamilies of
the Sphaeroceratidae and genera of the Macrocephalitinae
and Eucycloceratinae. The palaeobiogeography and evolu-
tion of the Eucycloceratinae are discussed in the light of
the new data.
Key words: Middle Jurassic, Ammonoidea, Eucycloceras,
Indo-Madagascan Province, phylogeny, cladistics.
Several exhaustive monographic treatments illustra-
ting a spectacular range of intraspecific morphological
variability are now available for Jurassic (e.g. Westermann
and Calloman 1988) and Cretaceous (e.g. Klinger and
Kennedy 1989) ammonite species. A direct result of these
studies is the drastic reduction of many previously estab-
lished morphospecies or morphogenera into a single bio-
species. This has helped remove pseudodiversity patterns,
and prompted reconsideration of many evolutionary
trees.
We consider the studies of Callomon (1985) to be an
excellent example of such work. Based on his ideas, we
have revisited the subfamily Eucycloceratinae, which con-
stitutes an important faunal element of the Callovian of
the Indo-Madagscan Faunal Province. We have shown
that failure to recognize the great intraspecific variability
and sexual dimorphism within the eucycloceratin popula-
tion led early workers to create a plethora of species.
These essentially are morphospecies or morphogenera,
and have little evolutionary significance. The present
investigation has not only unearthed numerous additional
specimens, but also was undertaken within a secure strati-
graphical framework. It reveals that many of the previ-
ously described species are essentially contemporaneous,
being restricted to within a single faunal horizon, or
at best a subzone. They show continuous horizontal
variation. However, the population may be sharply divi-
ded due to sexual dimorphism, an aspect overlooked by
early workers who failed to recognize this feature and
considered sexual variants as distinct genera.
The subfamily Eucycloceratinae of the family Sphaero-
ceratidae has not been studied in detail since Spath’s
(1928–33) work. He incorporated the four genera Notho-
cephalites, Eucycloceras, Subkossmatia and Idiocycloceras
within it. They are endemic to the Indo-Madagascan
Faunal Province and, as the new stratigraphical data
reveal, span a brief period of geological time from the lat-
est Early Callovian to early Middle Callovian. In Kutch
the appearance of this subfamily is coeval with that of the
Reineckeinae, at a time when the ancestral Macrocephalit-
inae were on the verge of regional extinction.
Spath’s four eucycloceratin genera include 14 species.
Most of these species were based on single specimens and
the holotypes are mostly fragmentary or preserved with
incomplete body-chambers. They are re-examined here
along with over 350 additional specimens. Detailed mor-
phological and morphometrical analyses reveal that, in
many cases, type species fall within the range of variability
of a single biospecies. Two of the four existing genera
(Eucycloceras and Idiocycloceras) survive close scrutiny and
form monospecific assemblages. The former is found in
the lower (older) stratigraphical horizon, but has strati-
graphical overlap with the latter. Most of the smaller spe-
cies of Subkossmatia, which closely resemble the inner
[Palaeontology, Vol. 48, Part 4, 2005, pp. 883–924]
ª The Palaeontological Association 883

whorls of Eucycloceras, are considered to be microconch
variants of Eucycloceras. The larger Subkossmatia species
have been relegated to members of Idiocycloceras. Idiocyc-
loceras is also found to be sexually dimorphic, although
microconchs were unknown until now. Nothocephalites is
excluded from Eucycloceratinae and placed within Macro-
cephalitinae.
Westermann and Wang (1988) assigned the Himalayan
genus Grayiceras Spath to the Eucycloceratinae. They
reported it from a condensed sequence in Tibet ranging
from the Upper Bathonian to Middle Callovian, although
it was previously described from the Oxfordian Spiti
Shale. We argue against the inclusion of Grayiceras s.s.
within the subfamily Eucycloceratinae and suggest that
the Tibetan forms perhaps belong to different eucyclocer-
atin species.
SUBFAMILY EUCYCLOCERATINAE:A HISTORICAL PERSPECTIVE
Spath (1928) first introduced the subfamily and included
four genera. He described two species of the type genus
Eucycloceras: E. eucyclum (Waagen), the type species, and
E. pilgrimi Spath. Waagen’s only specimen (1875, pl. 35,
fig. 1; here refigured in Pl. 1, fig. 1a–c) of Stephanoceras
eucyclum, which is an adult macroconch with partially
preserved body-chamber, was designated holotype. E. pil-
grimi is represented only by a small but complete adult.
Spath (1928) described six species of Subkossmatia of
which four (S. opis, S. obscura, S. coggin-browni and
S. discoidea) are small. They intergrade both morphologi-
cally and morphometrically (see below) and form a con-
tinuous series with E. pilgrimi. These smaller forms
closely resemble inner and intermediate whorls of E. eucy-
clum, which is a macroconch. The holotype of the latter,
and our recent collection of numerous specimens, indi-
cate that the macroconchs show limited intraspecific vari-
ability, but are marked by strong allometry during
ontogeny. The smaller species of Subkossmatia, as well as
E. pilgrimi, resemble different ontogenetic stages of the
macroconch. They are considered as variants, which
together with the type species, represent the enormously
variable microconch of a single species. Thus in E. eucy-
clum, the macroconch and the different microconchiate
forms are essentially sexual dimorphs of a single biospe-
cies. Interestingly, they are found in the same stratigraph-
ical interval and at the same localities. They thus
represent a widely variable monospecific assemblage (cf.
Callomon 1985). One notable exception from the usual
pattern of dimorphism in Jurassic ammonites is that the
dimorphism itself varies; the different ontogenetic stages
of adult macroconch phragmocones are replicated by the
different microconch adults. Hence, the notion that
dimorphism is a powerful tool for species discrimination
(Callomon 1971) has to be treated with caution. We
know only one other example from the Placenticeras kaff-
rarium population in the Cretaceous where enormous in-
traspecific variability has also been affected by the varying
nature of the dimorphism (see Klinger and Kennedy
1989; Bardhan et al. 2002).
Idiocycloceras of Spath (1928) also suffers from exces-
sive subjective splitting which has resulted in the creation
of several species, namely I. perisphinctoides (the type spe-
cies), I. singulare, I. dubium and I. sp. indet. The type
specimens are monotypic holotypes and represent septate
macroconchs, except that of I. singulare which is an
almost complete adult. They resemble the larger Subkoss-
matia ramosa and S. sp. indet., and many specimens
recently collected by us. All come from the same localities
and coeval levels, and intergrade morphologically. They
appear to be nothing more than different morphospe-
cies ⁄morphogenera, and in fact comprise a continuous
series. They form a monospecific assemblage within a very
narrow stratigraphical interval, representing a wide range
of variability of a single biospecies. The microconchs of
Idiocycloceras were previously unknown, but they are
recognized here on the basis of many new specimens. The
nature of dimorphism closely corresponds with that of
Eucycloceras, but the microconchs of Idiocycloceras show
less variability. The macroconch of Idiocycloceras is the
most evolute of the Sphaeroceratidae.
Eucycloceras and Idiocycloceras share many synapomor-
phies, such as an evolute shell with long primaries on the
outer whorls, the disappearance of secondaries rendering
the venter smooth on the last whorl of the adult macro-
conch (unlike those of macrocephalitins where loss of rib-
bing starts from the inner flanks producing smooth whorl
sides), protracted suture and the nature of dimorphism,
and evolute microconchs with ribbing persistent to the
end. However, both genera have their respective apomor-
phies (see below). For example, Eucycloceras macroconchs
develop adorally concave primaries on the adult body-
chamber.
Nothocephalites Spath appears to be quite different
from the eucycloceratin bauplan. It has a smooth adult
body-chamber, involute shell and short primaries, and
thus shares many plesiomorphic characters with macro-
cephalitins, especially with the endemic and older Macro-
cephalites formosus Spath. However, it has apomorphies
like fine and dense ribs. In the macroconch, both prima-
ries and secondaries suddenly disappear at the beginning
of the adult body-chamber. N. asaphus, the type species,
is considered here as a microconch (maximum diameter
154 mm) and N. semilaevis as a macroconch (maximum
diameter 230 mm). The former is characterized by having
a protracted suture like that of eucycloceratins (Spath
1928; Westermann and Wang 1988). We interpret the
884 PALAEONTOLOGY, VOLUME 48

suture as an evolving feature (see details in ‘Remarks’
section) and, in this respect, Nothocephalites shows an
evolutionary link between Macrocephalitinae and
Eucycloceratinae. Our cladistic analysis also reveals that
Nothocephalites is closer to Macrocephalites and we, fol-
lowing Thierry (1978) and Krishna and Westermann
(1987), retain it within the Macrocephalitinae but as a
distinct genus. In conclusion, Eucycloceratinae comprises
two genera: Eucycloceras Spath, 1924 (type genus) and Idi-
ocycloceras Spath, 1928.
SYSTEMATIC POSITION OFGRAYICERAS
Grayiceras is a problematic Himalayan genus previously
reported from Spiti (Spath 1923, 1924, 1928) and more
recently from southern Tibet (Westermann and Wang
1988). Spath proposed the genus, but did not define it,
and considered it an essentially Oxfordian or younger
form. He described several species of which only two sur-
vived later scrutiny (for detailed history of the genus, see
Arkell 1955 and Westermann and Wang 1988). These two
species are G. nepaulenese (Gray) and G. koeneni (Uhlig);
they are characterized by long primary ribs and protrac-
ted septal suture, i.e. two symplesiomorphic features typ-
ical of the early Mid-Callovian eucycloceratins. The type
specimens of G. nepaulense (see Westermann and Wang
1988, pl. 22, figs 1–3, pl. 23, fig. 1) seem to be micro-
conchs resembling one of the variants (infrasubspecies
obscura) of Eucycloceras. G. nepaulense shows similar
coarse ornamentation to the end without any loss of
strength and egression of the adult umbilical seam, and
inclined umbilical wall. On the other hand, G. koeneni
(Text-fig. 1A–C), which is a large body-chamber fragment
resembling Spath’s ‘Subkossmatia ramosa ⁄ Idiocyclocerassingulare’ (see also Westermann and Wang 1988), is in
fact a macroconch with characteristic coarse, distant, lon-
ger primaries. This ribbing style characterizes typical eu-
cycloceratin macroconchs compared with the short
primaries of the macrocephalitins. The problem is where
to place Grayiceras of the Oxfordian Spiti Shale. Are they
eucycloceratins or heterochronous homeomorphs? In the
evolutionary history of the sphaeroceratids, homeomor-
phism is apparent and widespread. Early Callovian
macrocephalitins have notorious homeomorphs among
Oxfordian mayaitins. Every exterior aspect, including
ornamentation, shell shape and septal suture, and even
interior morphology, of the macrocephalitins has a coun-
terpart among the mayaitins (Callomon in Donovan et al.
1981). Even the short primaries and the radial suture line
are mimicked. It is commonly agreed that the temporal
hiatus (Late Callovian–?Early Oxfordian) between these
two groups marks the distinction at subfamily level. Simi-
larly, the stratigraphically well constrained (near the
Lower ⁄Middle Callovian Substage boundary) eucyclocera-
tins of the Indo-Madagascan Faunal Province are strati-
graphically and geographically separated from Grayiceras
of the Tethyan Himalaya. Recently, it has been shown
that mayaitins include forms that have both radial and
protracted suture lines, e.g. Mayaites with radial, and
Epimayaites, Paryphoceras, Dhosaites and Prograyiceras
with protracted sutures (Westermann and Wang 1988).
This suggests that the suture was rapidly evolving, but
possibly with the gene(s) regulating the radial sutural
patterns remaining suppressed during the Mid-Callovian
A B C
TEXT -F IG . 1 . Grayiceras koeneni (Uhlig) [M]; A, lateral, B, apertural, C, ventral views; GSI 10014, bodywhorl fragment, retention of
secondaries indicate an immature shell; from Spiti Shale, ‘Gieumal’, Spiti; · 0Æ5.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 885

TEXT-F IG . 2 . Map of Kutch showing
important eucycloceratin-bearing
localities; see text-figures 3–6 for details.
TEXT -F IG . 3 . Stratigraphical column of the Jumara section illustrating lithostratigraphy, biostratigraphy and vertical distribution of
eucycloceratins and other important ammonite taxa; see Text-figure 2 for location.
886 PALAEONTOLOGY, VOLUME 48

(cf. Callomon in Donovan et al. 1981). Hence, we believe
that Grayiceras should be included within the subfamily
Mayaitinae, but as a distinct genus retaining the plesio-
morphic long primaries. It should be noted that other
mayaitins are also reported from the same horizons in the
Spiti area. They were described by Uhlig (1910b, pls 77,
81) as ‘Macrocephalites cf. maya’ and ‘Simbirskites n. sp.
ind.’, which according to Spath (1927–33) and Wester-
mann and Wang (1988) correspond well with Epimaya-
ites, especially E. falcoides Spath.
Grayiceras from southern Tibet (Westermann and
Wang 1988) creates further confusion. The area is tecton-
ically disturbed and the succession is interrupted by long
hiatuses. The ancillary taxa within the Grayiceras
assemblage are zonal indices of different ages. For exam-
ple, Macrocephalites cf. etheridgei, claimed by Westermann
and Wang (1988) to be from the Lower Callovian of the
Sula Islands and New Guinea (Indo-Pacific Faunal Prov-
ince), is a Middle Bathonian species, as recognized
by Westermann and Callomon (1988). Macrocephalites
cf. macrocephalus is a European species that comes from
the basal Callovian. Subkossmatia cf. opis, if correctly
identified, denotes a precise time interval straddling the
Early ⁄Middle Callovian boundary in the Indo-Madaga-
scan Faunal Province, especially in Kutch.
Thus it appears that the ferrugenous oolitic horizon
containing Grayiceras in southern Tibet may represent a
condensed sequence, and is certainly not younger than
Mid-Callovian. The species described were based on limi-
ted sample sizes and incomplete preservation. Subkossmatia
TEXT -F IG . 4 . Stratigraphical column of the Jara section illustrating lithostratigraphy, biostratigraphy and vertical distribution of
eucycloceratins and other important ammonite taxa; see Text-figure 2 for location.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 887

cf. opis is based on a few fragmentary specimens only,
one of which was figured (Westermann and Wang 1988,
pl. 23, figs 8–9). From the description and figure it
appears that it may represent a microconch variant of
Eucycloceras. Similar observations can be made on two
body-chamber fragments of G.? nepaulense (their text-
fig. 12A–B). They appear to be close to our larger micro-
conch variant of Eucycloceras, E. opis var. obscura. The
lectotype and paralectotype of their new species G.? waa-
geni significantly come from the Oxfordian Spiti Shale,
i.e. Giumal Sandstone, and are morphologically different
from the Callovian Tibetan specimens (their pl. 24,
figs 1–6). The Spiti specimens are compressed, involute
and relatively densely and finely ribbed in the adult stage.
Moreover, they lack characteristic long primaries, thus
perhaps resembling Epimayaites. On the other hand, Callo-
vian Tibetan forms have fine ribbing with long primaries,
and thus come close to Eucycloceras microconchs. How-
ever, this Tibetan population is more inflated and involute
than the Kutch population. G.? gucuoi sp. nov. (Wester-
mann and Wang 1988, pl. 24, figs 7–14, text-fig. 12C–E)
likewise resembles a microconch of Idiocycloceras, being
evolute (U ⁄D ¼ 0Æ25–0Æ30), depressed (B ⁄H ¼ 1Æ20–1Æ50)and coarsely ornate with biplicate ribs. It is, however, smal-
ler (D �60–70 mm) and relatively less evolute than I. peri-
sphinctoides [m]. We believe that while Grayiceras s.s. of
the Oxfordian belongs to the Mayaitinae, their alleged
Callovian counterparts reported from Tibet should be con-
sidered as a distinct population within the Eucyclocerati-
nae.
TEXT -F IG . 5 . Stratigraphical column of the Keera section illustrating lithostratigraphy, biostratigraphy and vertical distribution of
eucycloceratins and other important ammonite taxa; see Text-figure 2 for location.
888 PALAEONTOLOGY, VOLUME 48

STRATIGRAPHY
The Kutch Basin was initiated during the Middle Jurassic
fragmentation of Gondwana, and soon began to receive a
huge input of sediments due to repeated marine trans-
gressions (Biswas 1977). As sea connections opened up,
the basin received many immigrant faunas, mostly from
the European Tethys. In this new habitat the European
fauna rapidly speciated, producing a characteristic ende-
mic assemblage known as the Indo-Madagascan or Ethio-
pian Fauna. It includes corals (Gregory 1900; Pandey and
Fursich 1993), gastropods (Das et al. 1999; Jaitly et al.
2000), brachiopods (Mukherjee et al. 2002, 2003), nauti-
loids (Halder 2000) and ammonites (many authors
including Waagen 1875 and Spath 1924, 1927–33). While
much of the fauna has been treated monographi-
cally, stratigraphical data have remained subordinate to
taxonomy.
Lithostratigraphy
The major lithostratigraphical units, in ascending order,
are the Patcham, Chari, Katrol and Bhuj formations.
Mesozoic rocks cover nearly half of the area of Kutch,
including both the mainland and three ‘islands’ separated
by an intervening expanse of salt-flat known locally as the
Rann. The regional structure consists of three parallel,
TEXT -F IG . 6 . Stratigraphical column of the Jhura section illustrating lithostratigraphy, biostratigraphy and vertical distribution of
eucycloceratins and other important ammonite taxa; see Text-figure 2 for location.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 889

north-west–south-east-orientated, anticlines. The Jurassic
rocks are best developed in the central anticline (Wynne
1872), situated towards the north of the mainland. These
anticlines are superimposed by a set of zones of culmin-
ation that crop out as topographical domes, as at Jara, Ju-
mara, Keera and Jhura (Text-fig. 2). The domal outcrops
are disturbed by igneous intrusion and their northern
flanks are truncated by an east–west-trending fault. The
older formations (Patcham and Chari) are present as in-
liers and are surrounded by the younger Katrol Forma-
tion.
Ammonites are abundant in the two older formations,
despite the frequent spatiotemporal facies changes (Furs-
ich et al. 1994; Mukherjee et al. 2002; J. H. Callomon,
pers. comm. 2000). The Chari Formation is represented
by a heterolithic facies association consisting of shale,
limestone and sandstone, deposited in a mid-shelf
environment. While the Lower Callovian includes a domi-
nantly shale-limestone (packstone ⁄wackestone) associ-
ation, the Middle Callovian starts with siliciclastics.
Significant sedimentological work and detailed facies
analyses are now available (Datta 1992; Fursich et al.
1992; Fursich and Oschmann 1993). The arenaceous fa-
cies of the Middle Callovian represent a shoaling-upward
phase (Datta 1992; Fursich and Oschmann 1993). This fa-
cies is monotonous in the field, consisting of yellowish
grey sandstone, that is commonly multistoreyed, massive,
with a coarsening-upward sequence, devoid of bioturba-
tion and with large-scale cross-beds. Close study reveals
the presence of three units. They are persistent, forming
west–east ridges on mainland Kutch, with increasing
thickness and decreasing fossil-bearing horizons that
reflect the direction of the palaeoshoreline.
Most of the ammonite specimens are exquisitely pre-
served with their original shell intact and easily recover-
able from the sediments. Field investigations yielded more
than 350 eucycloceratin specimens, which were collected
from four important sections across the mainland
(Jumara, Jara, Keera and Jhura). Detailed lithostratigraph-
ical successions of each section, along with the vertical
distribution of the major ammonite species, is shown in
Text-figures 3–6.
Jumara is the stratotype of the Chari Formation (Text-
fig. 3). It is situated about 100 km north-west of Bhuj,
the district town of Kutch, and is named after the nearby
village. It is an eroded anticlinal dome with an inlier
occupied by the older Patcham Formation. Rajnath
(1932) first studied the Middle Jurassic rocks of Jumara
and mapped the area at a scale of 4 inches to 1 mile. The
Chari Formation, which borders the Patcham Inlier,
occurs as an elliptical outcrop. The beds show radial dip
which are mostly gentle, but may be up to 60 degrees
towards the northern flank. At Jumara the uppermost
Bathonian bed (Bed 4) of the Chari Formation is a lime-
stone (wackestone) 15 m thick. The lowermost bed of the
Lower Callovian (Bed 5) is 10 m thick and comprises
repeated alternations of shale, white limestone (wacke-
stone) and reddish or brownish limestone (pack-
stone ⁄ grainstone). Chevron cross-stratification is
occasionally present in the wackestone but the packstone
and grainstone are relatively massive. Bed 6 is represented
by a shale 16 m thick interbedded with thin, parallel-
sided, dark brown ferruginous limestone. The uppermost
bed of the Lower Callovian (Bed 7) is a thick shale with
coquina bands. This bed is 17 m thick and the first
appearance of the eucycloceratins is recorded from the
top part. The lowermost bed of the Middle Callovian
(Bed 8) is a yellowish grey multistoreyed sandstone 4 m
thick. This unit is overlain by a siltstone 6 m thick (Bed
9) and records the uppermost occurrence of the eucyclo-
ceratins. The immediately younger unit (Bed 10) is a yel-
lowish grey sandstone 15 m thick. The overlying unit
(Bed 11) consists of repeated alternations of greenish grey
shale ⁄ ironstone and white limestone (wackestone). This
bed is 70 m thick and is the uppermost unit of the Mid-
dle Callovian. The lowermost bed of the Upper Callovian
is a ferruginous sandstone 5 m thick (Bed 12) with bored
pebbles of white limestone (wackestone).
Jara is another anticlinal outcrop. It is c. 10 km north-
west of Jumara and named after a village in the vicinity.
Owing to severe weathering the profile of the dome is low
and the contacts between the beds are not always sharp.
However, because of the weathering, it is possible to
recover fossils easily, and the specimens collected are
mostly in an excellent state of preservation. The Callovian
rock sequence of Jara (Text-fig. 4) is similar to that of
Jumara, except that the base of the Lower Callovian is not
exposed. At Jara eucycloceratins are found in Beds 2 and 3.
Keera is an elliptical anticlinal dome situated c. 50 km
north-west of Bhuj. It is named after a hillock composed
of basic rocks in the northern part of the dome. The
older rocks are resistant and occupy high areas at the cen-
tre. They are surrounded by younger beds (for a detailed
lithological map, see Mitra et al. 1979). The lowermost
unit of the Chari Formation (Bed 1) at Keera (Text-
fig. 5) is a grey limestone (bioclastic grainstone) 15 m
thick. This is overlain by a sequence 65 m thick (Bed 2)
of golden coloured oolitic limestone (grainstone and
packstone). It is multistoreyed and cross-stratified (both
chevron and hummocky). There are at least three levels
where bored pebbles (wackestone) are sporadically strewn.
Ooids vary in shape and size, and show various internal
structures. The ooids are heavily impregnated with goeth-
ite, which is responsible for their characteristic golden
lusture. Kheraiceras hannoveranum, a typical Late Batho-
nian ammonite, has been found in the lower part of Bed
2 (Jana et al. 2002), indicating that the Upper Batho-
nian ⁄Lower Callovian boundary passes through this bed.
890 PALAEONTOLOGY, VOLUME 48

The allotype of Eycycloceras opis comes from the upper
part of this bed. The immediately younger bed (Bed 3) is
a grey shale ⁄marl 20 m thick with alternating reddish
brown to grey packstone. The uppermost bed of the
Lower Callovian (Bed 4) is a grey oolitic bioclastic grain-
stone (coquina) 5 m thick. The Middle Callovian litho-
stratigraphical sequences are similar to those of the
Jumara and Jara, and the uppermost level from which
eucycloceratins are recorded at Keera is Bed 6.
Jhura is another elliptical dome, situated c. 25 km
north-west of Bhuj, and named after the nearby village.
The same major lithologies of the western mainland can
be traced here but the individual beds are thicker, with
increasing siliciclastic content. This suggests that Jhura lies
nearer to the palaeoshoreline. Here, resistant and thick
rocks of the Patcham and Chari formations occupy a lar-
ger area and form ranges with hills up to 300 m high with
deep canyons between. The dome, like others, opens
towards the north and is drained by the Kaila River and
many subsidiary streams. Much of the older Patcham For-
mation is exposed at the core. The older beds are only
accessible through some passes that constitute dry streams
flowing towards the north. At Jhura the Callovian rock
sequence (Text-fig. 6) is much thicker than that of the
other areas studied. Here the uppermost Bathonian bed
(Bed 4) is a hard, slabby, white limestone 15 m thick, and
is lithologically similar to Bed 4 of Jumara. The lowermost
unit of the Lower Callovian (Bed 5) is a sequence of shale-
limestone alternations 65 m thick with a hard compact car-
bonate facies that is 1 m thick at the base. The uppermost
unit of the Lower Callovian (Bed 6) is a unit of shale-lime-
stone alternations 96 m thick with coquina bands. The
lowermost unit of the Middle Callovian is a red massive
sandstone bed 31 m thick (Bed 7). This is followed by an
oolitic limestone 69 m thick (Bed 8); eucycloceratins occur
in this unit. The overlying unit (Bed 9) is a 46-m-thick
sandstone that is equivalent to Bed 10 of Jumara, Bed 4 of
Jara and Bed 7 of Keera. The younger units are similar to
those of the other areas studied.
Biostratigraphy
Many stratigraphical schemes encompassing the Callovian
have been proposed and continue to be debated (e.g.
Biswas 1977; Krishna 1984). We, along with others, have
been revising pre-existing schemes as new data emerge
(Bardhan et al. 1994). Recently we have subdivided the
Lower Callovian Substage on the basis of diverse Macro-
cephalites (Bhaumik et al. 1993; Datta et al. 1996) and
Kheraiceras species (Jana et al. 2000; Bardhan et al. 2001),
retaining many of the existing subzones but proposing
several new faunal horizons. A newly proposed general-
ized biozonation based on ammonites for the Middle
Callovian of Kutch, along with a tentative correlation
with the Sub-Mediterranean biostratigraphical schemes, is
presented in Text-figure 7. Systematic collecting of
ammonites has enabled us to refine the position of exist-
ing faunal horizons in addition to introducing many new
ones. These horizons are easily recognizable in the field,
especially in the western part of the mainland (Jara, Ju-
mara, Keera).
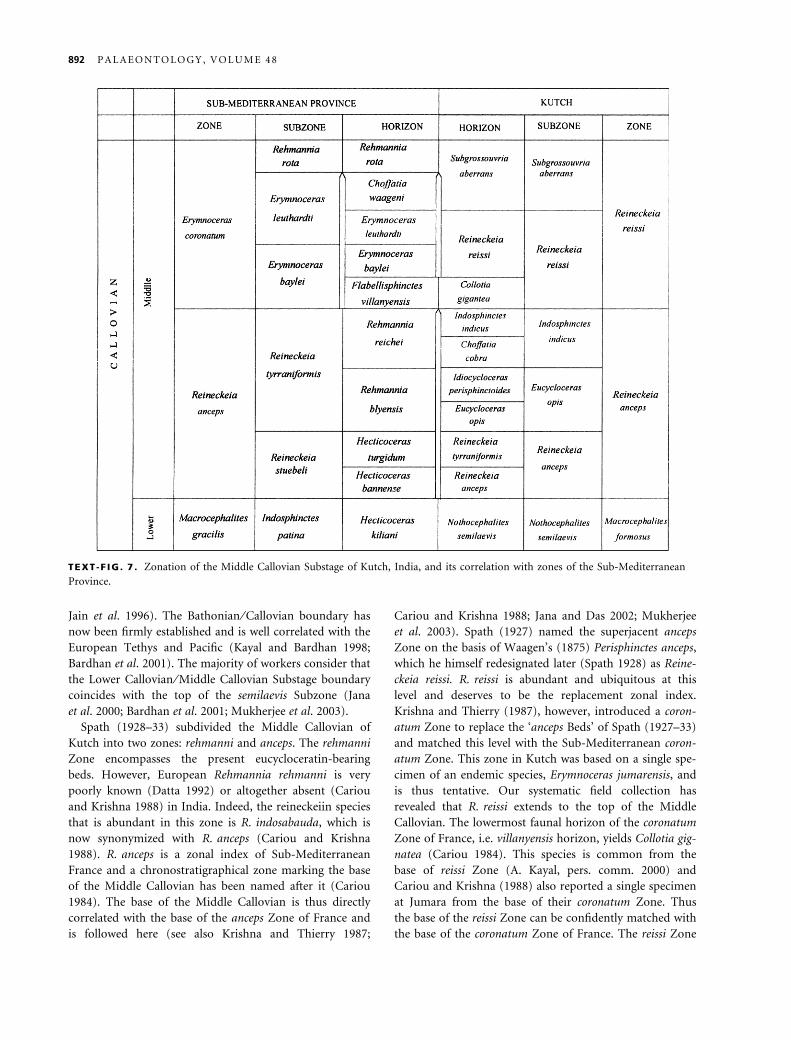
The proposed zones of the Middle Callovian strata are
the anceps and reissi assemblage zones. The former
includes three subzones, anceps, opis and indicus in
ascending order. The reissi Assemblage Zone has been fur-
ther subdivided into two subzones, reissi and aberrans in
ascending order. The detailed ammonite faunal assem-
blages are given below:
anceps horizon. This is marked by the disappearance of
Kheraiceras, Macrocephalites spp., Inodocephalites spp.,
Nothocephalites semilaevis and Kamptokephalites dimerus;
K. lamellosus is very rare; Idiocycloceras makes its first
appearance; Eucycloceras opis, Reineckeia anceps, R. tyr-
anniformis, Collotia oxyptycha, Choffatia recuperoi,
C. cobra and C. pedargatus of the preceding semilaevis
horizon continue.
tyranniformis horizon. This is marked by the disappear-
ance of Macrocephalitinae; Reineckeia tyranniformis is fre-
quent; all other species of the preceding horizon continue.
opis horizon. Eucycloceras opis is dominant; all other
species continue.
perisphinctoides horizon. Idiocycloceras perisphinctoides
is the dominant faunal element; first appearance of the
genus Phlyticeras; all other species continue.
cobra horizon. This is marked by the disappearance of
Eucycloceratinae and the inception of the genus Indo-
sphinctes; reineckeiins, choffatins and Phlycticeras continue.
indicus horizon. Indosphinsctes indicus very abundant;
Phlycticeras is common; all other species continue.
gigantea horizon. Reineckeia anceps, R. tyranniformis
and Indosphinctes disappear; Collotia gigantea, R. reissi,
Erymnoceras jumarensis first appear; Obtusicostites obtusi-
costa and Hubertoceras arcicosta common; choffatins con-
tinue.
reissi horizon. Reineckeia reissi is the most abundant
taxon; Collotia gigantea and choffatins continue.
aberrans horizon: Subgrossouvria aberrans is the most
abundant taxon; R. reissi, C. gigantea and choffatins con-
tinue; first appearance of R. rota; Hecticoceras (Kheraites)
ferrugineum and H. (Putealiceras) trilineatum common.
Chronostratigraphy and correlation with other regions
Demarcation of stage boundaries and correlation of regio-
nal standard zonations with those of stratotypes are two of
the central concerns of Kutch workers (Callomon 1993;
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 891
Jain et al. 1996). The Bathonian ⁄Callovian boundary has
now been firmly established and is well correlated with the
European Tethys and Pacific (Kayal and Bardhan 1998;
Bardhan et al. 2001). The majority of workers consider that
the Lower Callovian ⁄Middle Callovian Substage boundary
coincides with the top of the semilaevis Subzone (Jana
et al. 2000; Bardhan et al. 2001; Mukherjee et al. 2003).
Spath (1928–33) subdivided the Middle Callovian of
Kutch into two zones: rehmanni and anceps. The rehmanni
Zone encompasses the present eucycloceratin-bearing
beds. However, European Rehmannia rehmanni is very
poorly known (Datta 1992) or altogether absent (Cariou
and Krishna 1988) in India. Indeed, the reineckeiin species
that is abundant in this zone is R. indosabauda, which is
now synonymized with R. anceps (Cariou and Krishna
1988). R. anceps is a zonal index of Sub-Mediterranean
France and a chronostratigraphical zone marking the base
of the Middle Callovian has been named after it (Cariou
1984). The base of the Middle Callovian is thus directly
correlated with the base of the anceps Zone of France and
is followed here (see also Krishna and Thierry 1987;
Cariou and Krishna 1988; Jana and Das 2002; Mukherjee
et al. 2003). Spath (1927) named the superjacent anceps
Zone on the basis of Waagen’s (1875) Perisphinctes anceps,
which he himself redesignated later (Spath 1928) as Reine-
ckeia reissi. R. reissi is abundant and ubiquitous at this
level and deserves to be the replacement zonal index.
Krishna and Thierry (1987), however, introduced a coron-
atum Zone to replace the ‘anceps Beds’ of Spath (1927–33)
and matched this level with the Sub-Mediterranean coron-
atum Zone. This zone in Kutch was based on a single spe-
cimen of an endemic species, Erymnoceras jumarensis, and
is thus tentative. Our systematic field collection has
revealed that R. reissi extends to the top of the Middle
Callovian. The lowermost faunal horizon of the coronatum
Zone of France, i.e. villanyensis horizon, yields Collotia gig-
natea (Cariou 1984). This species is common from the
base of reissi Zone (A. Kayal, pers. comm. 2000) and
Cariou and Krishna (1988) also reported a single specimen
at Jumara from the base of their coronatum Zone. Thus
the base of the reissi Zone can be confidently matched with
the base of the coronatum Zone of France. The reissi Zone
TEXT -F IG . 7 . Zonation of the Middle Callovian Substage of Kutch, India, and its correlation with zones of the Sub-Mediterranean
Province.
892 PALAEONTOLOGY, VOLUME 48

of Kutch is subdivided into two subzones, i.e. the lower
reissi Subzone and the upper aberrans Subzone. The aber-
rans Subzone is dominated by Subgrossouvria aberrans.
However, the subzone boundaries of the reissi Zone are
difficult to correlate with those of the contemporaneous
coronatum Zone of France because of the absence of any
cosmopolitan ammonite species.
The anceps Zone of France is subdivided into two sub-
zones: stuebeli and tyranniformis. We have been able to
subdivide the anceps Zone of Kutch into three subzones:
anceps, opis and indicus. Recent study (Kayal, pers. comm.
2000) reveals that R. anceps and R. stuebeli are morpho-
logically very close and have the same biogeographical
and biostratigraphical distributions, and appear to be
conspecific (monospecific assemblage of Callomon 1985).
Thus the anceps Subzone of Kutch matches well the stue-
beli Subzone of France. However, R. tyranniformis occurs
at a higher chronostratigraphical level in France (ranging
from the blyensis horizon, tyranniformis Subzone to baylei
horizon, baylei Subzone of the coronatum Zone) than in
Kutch where it first appears in the semilaevis horizon of
the Lower Callovian and continues to the top of the
anceps Zone. Spath (1928) reported the holotype of
R. tyranniformis from the Golden Oolite of Keera, which
also yields Nothocephalites semilaevis.
Eucycloceratins, however, have different distribution
patterns. The older genus Eucycloceras first appears in the
semilaevis horizon. The holotype of Spath’s (1928) Eucyc-
loceras eucyclum comes from the Golden Oolite of Keera.
He later mentioned the co-occurrence of Eucycloceras
with Nothocephalites semilaevis in the same matrix (Spath
1933, p. 676) and emphasized that none of the eucyclo-
ceratin species passes beyond the ‘anceps Beds’. His table 4
(pp. 716–717) shows the upper limit of the subfamily is
his Lower anceps Beds. Our repeated field investigations
support Spath’s (1933) later view and reconfirm its lower
limit extending back to the semilaevis horizon. To date
we have collected only two specimens of Eucycloceras opis,
one of each sexual variant, from this level. Idiocycloceras
appears only from the basal anceps horizon of the Middle
Callovian and both genera co-occur up to the peri-
sphisctoides horizon before they disappear dramatically.
The two faunal horizons named after them are not always
distinguishable in the field, particularly eastwards on the
mainland.
The substage boundary between the Middle and Upper
Callovian is secured by the discovery of several specimens
of Reineckeia rota (A. Kayal, pers. comm. 2000).
CLADISTIC ANALYSES
Eucycloceratinae is a small clade with two genera that are
restricted to the Lower Callovian ⁄Middle Callovian Sub-
stage boundary. They belong to the family Sphaerocerati-
dae, which is clearly recognizable as a monophyletic
group on the basis of at least three major symplesiomor-
phic characters: non-lappeted microconch, low dimorphic
size ratio and uninterrupted ribbing across the venter.
The phylogenetic relationships among the members of
this family remain poorly understood. For example,
Donovan et al. (1981) questionably retained Eucyclocerat-
inae as a subfamily. The present cladistic analyses were
performed using 25 shell morphology characters to study
phylogenetic relationships among the five existing
subfamilies. The cladograms are based on PAUP, 4.0b 10
version (Swofford 1998). All characters are weighted
equally and unordered. The characters are given in Table 1
and the data matrix used is given in the Appendix.
Otoitidae has been considered as an ancestral outgroup
TABLE 1 . List of characters used in the cladistic analyses of
Sphaeroceratidae subfamilies (see Text-fig. 8A–B).
1. Shell sharply ribbed: present (0); absent (1).
2. Complex septal suture: present (0); absent (1).
3. Dominant first lateral lobe: present (0); absent (1).
4. Well-developed umbilical lobe: present (0); absent (1).
5. Microconch ribbed to the end: present (0); absent (1).
6. Egression of adult body-chamber in microconch: present
(0); absent (1).
7. Non-lappeted microconch: present (0); absent (1).
8. Dimorphic size ratio: high (0); low (1).
9. Ventral discontinuity of ribbing: may be present (0); absent
(1).
10. Adult size of macroconch: small (0); large (1).
11. Variocostation (differentiated ribbing) of microconch: pre-
sent (0); absent (1).
12. Microconch with collar: present (0); absent (1).
13. Lateral tubercle ⁄ bullae in microconch: present (0); absent (1).
14. Terminal constriction in macroconch: absent (0); present (1).
15. Strength of ribbing in microconch: coarse (0); fine to coarse (1).
16. Adult whorl outline: planulate (0); sphaerocone (1); sph-
aerocone to oxycone (2); ovate to subtrapezoidal (3); sph-
aerocone to subelliptical (4).
17. Egression of adult body-chamber in macroconch: absent (0);
present (1).
18. Degree of involution of macroconchiate phragmocone: peri-
sphinctoid evolute (0); obvolute (occluded umbilicus) to
involute (1); involute (2); evolute (3).
19. Depressed inner whorl: present (0); absent (1).
20. Contracted aperture in macroconch: absent (0); present (1).
21. Contracted aperture in microconch: absent (0); present (1).
22. Occluded to strongly involute umbilicus in young micro-
conch: present (0); absent (1).
23. Strength of ribbing in microconch in comparison to that of
macroconch: strong (0); moderate (1); weak (2).
24. Length of primaries in outer whorl: short (0); long (1); both
(2).
25. Order of withdrawal of ribbing in macroconch: primaries
first (0); secondaries first (1).
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 893

(see Donovan et al. 1981) of Sphaeroceratidae. We have
deliberately chosen species that exhibit a reasonable
amount of variation among members of a genus or
higher taxon (such as subfamily and family). The majority
of these species represent type species of a genus and are
marked by asterisks in Table 2. Macrocephalitinae and
Mayaitinae are known to be homeomorphic (Donovan
et al. 1981; Thierry in Dommergues et al. 1989) and
therefore the homoplastic character states, such as radial
to protracted suture line, fine to coarse ribbing in the
macroconch and complete smoothness of body-chamber
of the macroconch, are excluded here. The PAUP search
has been performed using the branch and bound option,
which guarantees finding the shortest trees. Six trees have
been found, of which the strict consensus tree is shown
in Text-figure 8A. From these trees one is selected
(Text-fig. 8B) following Monks (2000, 2002), which
appears to be in agreement with the stratigraphical and
TABLE 2 . Species of outgroup Otoitidae and different subfamilies of Sphaeroceratidae used in the cladistic analyses described
here; *denotes type species. Stratigraphical ranges and palaeogeographical distribution are recorded, with all sources reported in
the text.
Taxa Stratigraphical range Palaeobiogeography
Otoitidae Mascke, 1907 Aalenian–Bajocian Cosmopolitan
Abbasites Buckman, 1921
*A. abbas (Buckman, 1921)
Aalenian Europe
Docidoceras Buckman, 1919
*D. cylindroides Buckman, 1919
Bajocian Europe, North Africa
Emileia Buckman, 1898
*E. brocchii (J. de C.Sowerby, 1818)
Middle Bajocian Cosmopolitan
Zemistephanus McLearn, 1927.
*Z. richardsoni (Whiteaves, 1876)
Middle Bajocian Pacific
Trilobiticeras Buckman, 1919
*T. trilobitoides Buckman, 1919
Middle Bajocian Europe, Western Australia
Otoites Mascke, 1907
*O. sauzei (Orbigny, 1846)
Middle Bajocian Cosmopolitan
Sphaeroceratinae Buckman, 1920. Bajocian Cosmopolitan except Boreal
Sphaeroceras Bayle, 1878
*S. brongniarti (J. de C. Sowerby, 1817)
Bajocian Europe, N. Africa, Persia, S. Alaska
Chondroceras Mascke, 1907
*C. gervillii (Sowerby, 1817)
Bajocian Europe, N. Africa, Caucasus,
New Guinea, America
Preatulites Westermann, 1956
*P. kruizingai Westermann 1956
Bajocian Pacific
Labyrinthoceras Buckman, 1919
*L. perexpansum Buckman, 1919
Middle Bajocian Europe
Frogdenites Buckman, 1921
*F. spiniger Buckman, 1921
Middle Bajocian Europe, Tibet, Canada
Eurycephalitinae Thierry, 1976 Upper Bajocian–
Middle Callovian?
Pacific
Eurycephalites Spath, 1928
*E. vergarensis (Burckhardt, 1903)
E. rotundus (Tornquist)
Lower Callovian East Pacific
Megasphaeroceras Imlay, 1973
M. magnum Riccardi and Westermann
Upper Bajocian East Pacific
Lilloetia Crickmay, 1930
*L. lillotensis Crickmay, 1930
L. steinmanni (Spath, 1928)
Upper Bathonian–
Middle Callovian?
Pacific
Xenocephalites Spath, 1928
*X. neuquenensis (Stehn, 1924)
X. gottschei (Tornquist, 1898)
Upper Bathonian–
Lower Callovian
Pacific
Macrocephalitinae Salfeld, 1921 Middle Bathonian–lower
Middle Callovian
South-west Pacific, Tethyan and
Indo-Madagascan Province
Macrocephalites (Macrocephalites) Zittel, 1884
*M. (M.) macrocephalus (Schlotheim, 1813)
M. (M.) bifurcatus bifurcatus Boehm, 1912
As above Europe, Madagascar, India (Kutch
and Himalayas), Indonesia, New Guinea
894 PALAEONTOLOGY, VOLUME 48

TABLE 2 . Continued.
Taxa Stratigraphical range Palaeobiogeography
M. (M.) bifurcatus intermedius (Spath, 1928)
M. (M.) mantataranaus Boehm, 1912
M. (M.) apertus (Spath, 1928)
M. (M.) keeuwensis Boehm, 1912
M. (M.) triangularis Spath, 1928
M. (M.) madagascariensis Lemoine, 1911
M. (M.) formosus (J. de C. Sowerby, 1840)
M. (M.) jacquoti (Douville, H., 1878)
M. (M.) verus Buckman, 1922
Macrocephalites (Indocephalites) Spath, 1928
*M. (I.) kheraensis Spath, 1928
M. (I.) chrysoolithicus (Waagen, 1875)
M. (I.) diademetus (Waagen, 1875)
Lower Callovian India, Madagascar, France
Macrocephalites (Pleurocephalites) Buckman, 1922
M. (P.) habyensis Spath, 1928
M. (P.) elephantinus (J. de C. Sowerby, 1840)
Lower Callovian Indonesia, India, Madagascar, England
Macrocephalites (Kamptokephalites) Buckman, 1922
M. (K.) lamellosus (J. de C. Sowerby, 1840)
M. (K.) dimerus (Waagen, 1875)
M. (K.) etheridgei Spath, 1928
Middle Bathonian–
lower Middle Callovian
Indonesia, India, Madagascar, France,
England
Macrocephalites (Dollikephalites) Buckman, 1923
M. (D.) subcompressus (Waagen, 1875)
M. (D.) gracilis Spath, 1928
Lower Callovian India, Madagascar, France, England
Nothocephalites Spath, 1928
N. semilaevis (Waagen, 1875)
*N. asaphus Sapth, 1928
upper Lower Callovian India, Madagascar
Eucycloceratinae Spath, 1928 Top of Lower Callovian–
lower Middle Callovian
Indio-Madagascan Province,
Himalayan Tethys
Eucycloceras Spath, 1924
*E. opis (J. de C.Sowerby, 1840)
As above India, Madagascar, Tibet
Idiocycloceras Sapth, 1928
*I. perisphinctoides Spath, 1928
Basal Middle Callovian India, Madagascar, Tibet
Mayaitinae Spath, 1928 Lower Oxfordian–Lower
Kimmeridgian
Indo-Madagascan Province,
Himalayan Tethys, Pacific (South)
Mayaites Spath, 1924
*M. maya (J. de C.Sowerby, 1840)
M. obesus Spath, 1928
M. smeei Spath, 1928
Oxfordian–Lower
Kimmeridgian
India, Madagascar, Indonesia
Mayaites (Araucanites) Westermann and Riccardi, 1975
*M. (A.) stipanicici Westermann and Riccardi, 1975
M. (A) reyesi Westermann and Riccardi, 1975
Oxfordian Argentina, Chile
Epimayaites Spath, 1928
E. polyphemus (Waagen 1875)
*E. transiens (Waagen 1875)
Oxfordian
E. falcoides Spath, 1928 India, Madagascar
Dhosaites Spath, 1924
*D. eliphantoides Spath, 1928
Oxfordian India, Madagascar
Paryphoceras Spath, 1928
*P. badiense Spath, 1928
Oxfordian India, Madagascar
Prograyiceras Spath, 1928
*P. grayi (Spath, 1924)
Oxfordian India, Madagascar
Grayiceras Spath (1923)
*G. nepaulenese (Gray, 1830–32)
G. koeneni (Uhlig, 1910)
Oxfordian India (only Himalayas)
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 895

palaeobiogeographical data (see Table 2). This tree has a
length of 29, consistency index of 0Æ8621 and a retention
index of 0Æ7143. Statistical tests like bootstrap and branch
decay (Bremer 1988) have also been performed using
PAUP and PAST (Hammer et al. 2003). The bootstrap
values for a group is the percentage of replicates support-
ing that group and it tests the robustness of tree topology
(Felsenstein 1985). Bremer support for a clade is the
number of extra steps a tree needs to be lengthened for a
given node to collapse; the greater the number of steps
the more robust the node (Monks 2000). The result of
both of these tests are given in Text-figure 8B.
The systematic position of Nothocephalites has been
debated for a long time. Here we advocate its inclusion
within the Macrocephalitinae. To test our proposed clas-
sification we have performed another cladistic analysis
and constructed a cladogram using 27 morphological
characters (Table 3) to document the geneological pat-
terns among the four genera Macrocephalites, Notho-
cephalites, Eucycloceras and Idiocycloceras. Lower
Callovian Tethyan Macrocephalites has been considered
as an outgroup. A single tree (Text-fig. 8C) has been
derived by the branch and bound option. This tree has
a length of 30, consistency index of 0Æ9333 and a retent-
ion index of 0Æ7143. The data matrix used is given in
the Appendix. Similarly, this cladogram fits perfectly
with stratigraphical and palaeobiogeographical data (see
Table 2). Data sources used here are taken from a range
of literature (Spath 1924, 1927–33; Basse and Perrodon
1951; Westermann 1956; Arkell et al. 1957; Stipanicic
et al. 1975; Thierry 1978; Donovan et al. 1981; Krishna
and Westermann 1987; Westermann and Callomon
1988; Westermann and Hudson 1991; Bhaumik et al.
1993; Datta et al. 1996; Parent 1997; Jana 2002) and
from personal observation.
SYSTEMATIC PALAEONTOLOGY
Abbreviations. Specimen numbers bear the following repository
institutional prefixes: GSI, Geological Survey of India, Kolkata;
JUM, Jadavpur University Museum, Kolkata. [M] and [m] des-
ignate macro- and microconch, respectively. Asterisks (*) against
items in synonymies indicate type specimens. The following let-
ters are used to indicate shell parameters: B, breadth of whorl;
D, diameter of shell; H, height of whorl from umbilical margin;
P, number of primaries per half whorl; S, number of secondaries
per half whorl; U, diameter of umbilicus.
Family SPHAEROCERATIDAE Buckman, 1920
Subfamily EUCYCLOCERATINAE Spath, 1928
Diagnosis. Evolute shell, inner whorls depressed and less
evolute; adult whorls mostly compressed. Strongly dimor-
phic in size and nature of ornamentation. Microconchs
have generally isocostate ribbing which continues to the
EXPLANATION OF PLATE 1
Figs 1–7. Eucycloceras opis (J. de C.Sowerby) [M]. 1a–c, allotype, adult with incompletely preserved body-chamber, GSI 2020, Golden
Oolite (Bed 2), Keera, photographically presented for the first time. 2, whorl section of an adult body-chamber, JUM 506, Bed 8,
Jhura. 3, almost complete adult, JUM 425, Bed 9, Jumara. 4, venter of almost complete adult, JUM 431, Bed 9, Jumara. 5a–b,
wholly septate, JUM 511, Bed 8, Jhura. 6, wholly septate, JUM 565, Bed 8, Jhura. 7, Adult phragmocone with inner whorls
exposed, JUM 428, Bed 6, Keera. Cross-marks indicate the position of adult end-phragmocone. All · 0Æ5.
A
B
C
TEXT -F IG . 8 . Cladograms showing phylogenetic relationships
of different subfamilies of Sphaeroceratidae with Otoitidae as an
outgroup (A–B). Strict consensus of the six most parsimonious
trees is shown in A, and the tree selected as best match with
stratigraphical and palaeobiogeographical data is shown in B.
Percentage bootstrap support (above 50%) and branch decay
values are also plotted (in squares and circles, respectively) on
the latter figure. The genus-level cladistic analysis of
Macrocephalitinae and Eucycloceratinae yields only a single tree,
which is also stratigraphically compatible; it is shown in C.
896 PALAEONTOLOGY, VOLUME 48

aperture and macroconchs have variocostate ribbing.
Body-chamber occupies three-quarters of the last whorl.
Macroconchs are the most evolute sphaeroceratids. Septal
suture protracted. Prosocline primary ribs become
increasingly longer, coarser and blunter during ontogeny.
Secondaries project strongly forward over venter near
1a 1b 1c
2
3 X
4
5a
67 5b
PLATE 1
JANA et al., Eucycloceras
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 897

aperture of microconchs, but continue up to varying dis-
tance with attenuated strength on venter of adult macro-
conch and finally disappear.
Range. Upper Lower Callovian–lower Middle Callovian, distri-
buted mainly in the Indo-Madagascan Faunal Province, with
rare excursions to the Himalayan Tethys.
Genus EUCYCLOCERAS Spath, 1924
Type species. Stephanoceras eucyclum Waagen, 1875; subsequent
designation by Spath, 1924–28; now recognized to be a synonym
of Ammonites opis J. de C. Sowerby (1840).
Eucycloceras opis (J. de C. Sowerby)
Plates 1–4; Plate 5, figures 1–4; Text-figures 9–12
*1840 Ammonites opis J. de C. Sowerby, pl. 23, fig. 9 [m].
1875 Stephanoceras opis (J. de C. Sowerby); Waagen,
p. 141, pl. 36, fig. 1 [m].
*1875 Stephanoceras eucyclum Waagen, p. 142, pl. 35
fig. 1 [M].
non1893 Ammonites (Stephanoceras) opis, Oldham, p. 228
[m].
1902 Ammonites opis J. de C Sowerby; Blake, p. 35 [m].
1910 Kossmatia? eucycla (Waagen); Lemoine, p. 29 [M].
1910 Macrocephalites opis (J. de C. Sowerby); Lemoine,
p. 30 [m]
1910 Macrocephalites opis (Waagen); Uhlig, a, p. 265
[m].
1910 Kossmatia eucycla (Waagen); Uhlig, a, p. 268 [M].
1913 Stephanoceras opis (J. de C. Sowerby); Smith, b and
c, p. 420 [m].
1924 Eucycloceras eucyclum (Waagen); Spath, p. 8 [M].
1924 Subkossmatia opis (J. de C. Sowerby); Spath, p. 11
[m].
1924 Macrocephalites? (Eucycloceras?) subcompressus
(Waagen); Spath, p. 21 (no. 248) [m].
1925 Eucycloceras eucyclum (Waagen); Spath, p. 209,
pl. 23, fig. 4a–b; pl. 25, fig. 4 [M]; pl. 27, fig. 7a–b
[m].
1928 Eucycloceras pilgrimi Spath, p. 209, pl. 27, fig. 6;
pl. 29, fig. 2 [m].
EXPLANATION OF PLATE 2
Figs 1–10. Eucycloceras opis (J. de C. Sowerby). 1–3, M. 1a–b, wholly septate, JUM 248, Bed 9, Jumara. 2, complete adult, JUM 505, Bed
8, Jhura; note dense growth striae near aperture. 3a–b, wholly septate, JUM 509, Bed 8, Jhura. 4–10, m, var A. 4, almost complete
adult with inner whorls exposed, JUM 261, Bed 6, Keera. 5a–b, complete adult, JUM 392, Bed 9, Jumara. 6a–c, paralectotype,
complete adult, JUM 371, Bed 5, Keera. 7a–c, complete adult, JUM 556, Bed 9, Jumara; note sharp umbilical edge and steep wall in
adult body-chamber. 8a–b, almost complete adult, JUM 372, Bed 6, Keera. 9, complete adult, JUM 433, Bed 9, Jumara; note
approximation of ribbing near aperture. 10, complete adult with appoximated ribs at the end, JUM 432, Bed 9, Jumara. All · 0Æ5.
TABLE 3 . List of characters used in the cladistic analyses of
Macrocephalitinae and Eucycloceratinae genera (see Text-fig. 8C).
1. Uncoiling of adult body-chamber: present (0); absent (1).
2. Nucleus whorl densely, finely ribbed, evolute and depressed:
present (0); absent (1).
3. Umbilical wall initially vertical or overhanging, later sloping:
present (0); absent (1).
4. Strongly dimorphic: present (0); absent (1).
5. Macroconch very large: present (0); absent (1).
6. Non-lappeted microconch: present (0); absent (1).
7. Microconch ribbed throughout ontogeny: present (0); absent
(1).
8. Variocostate macroconch, isocostate microconch: present
(0); absent (1).
9. Degree of involution of macroconch: involute (0); evolute
(1); strongly evolute (2).
10. Inflation of adult whorl: compressed to depressed (0); always
compressed (1).
11. Smoothness of body-chamber in adult macroconch: com-
pletely smooth (0); partially smooth (1).
12. Smoothing of outer whorl starts from: inner flank (0); inner
flank to siphonal part (1); siphonal part (2).
13. Egression of umbilical seam starts from: adult body-chamber
(0); phragmocone (1).
14. Suture: radial (0); radial to protracted (i.e. raised umbilical
saddle envelope) (1); protracted (2).
15. Length of primaries on outer whorl: short (0); long (1).
16. Order of withdrawal of ribbing: primaries first (0); primaries
and seconderies simultaneously (1); secondaries first (2).
17. Withdrawal of ribbing starts from: phragmocone (0); begin-
ning of adult body-chamber (1).
18. Nature of primaries in later ontogeny: straight (0); curved (1).
19. Ventral projection of secondaries in macroconch: absent (0);
present (1).
20. Strength of ribbing in macroconch: fine to coarse (0); fine
(1); coarse (2).
21. Intraspecific variability of macroconch: high (0); low (1).
22. Sex ratio in favour of macroconch: present (0); absent (1).
23. Variocostation of macroconch: weak (0); strong (1); moder-
ate (2).
24. Microconch more evolute: absent (0); present (1).
25. Adult body-chamber length: variable (0); more than three-
quarters of last whorl (1); three-quarters of last whorl (2).
26. Shortening of primaries on adult body-chamber: primaries
absent (0); present (1); absent (2).
27. Persistence of secondaries on adult body-chamber in macro-
conch: secondaries absent (0); up to first quarter (1); up to sec-
ond quarter (2).
898 PALAEONTOLOGY, VOLUME 48

1928 Subkossmatia opis (J. de C. Sowerby); Spath, p. 210,
pl. 36, fig. 2; pl. 39, figs 2a–b, 7 [m].
*1928 Subkossmatia obscura Spath, p. 211, pl. 38, fig. 5
[m].
*1928 Subkossmatia coggin-browni Spath, p. 212, pl. 31,
fig. 6; pl. 35, fig. 7; pl. 38, fig. 2; pl. 41, fig. 4a–c
[m].
PLATE 2
JANA et al., Eucycloceras
1a 1b
2
3a 3b
X
4 X5a
X5b
X 6a 6b 6c
7a7b
X
7c8a X98b
X10
X
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 899

*1928 Subkossmatia discoidea Spath, p. 213, pl. 50,
fig. 2a–b [m].
1938 Eucycloceras eucyclum (Waagen); Roman, p. 217,
fig. 29; pl. 19, fig. 204 [M].
1951 Eucycloceras eucyclum (Waagen); Basse and Perro-
don, p. 47, pl. 4, fig. 2a–b [M].
1951 Subkossmatia alfuricoides Basse and Perrodon, p. 49,
pl. 4, fig. 3a–c [m].
1951 Subkossmatia bassei Basse and Perrodon, p. 49,
pl. 3, figs 10a–b, 12; pl. 6, fig. 1a–c [m].
1951 Subkossmatia subfissum Basse and Perrodon, p. 50,
pl. 6, fig. 2a–c [m].
1988 Subkossmatia cf. opis (J. de C. Sowerby); Wester-
mann and Wang, p. 332, pl. 23, figs 8–9 [m].
Nomenclature. Different morphospecies of two previously des-
cribed morphogenera have been combined here. Eucycloceras
retains the generic name, being senior to Subkossmatia in page
priority, but the trivial name opis is the senior objective syno-
nym. Different variants within the microconch are in fact mor-
phospecies in the analytical sense of Linnean taxonomy, but are
here considered to be infrasubspecific units.
Type specimens. The holotype of Ammonite opis Sowerby, 1840
(refigured and designated by Spath 1928, pl. 39, fig. 2a–b as
Subkossmatia opis) from the Callovian of Keera, Kutch, and the
holotype of Stephanoceras eucyclum Waagen, 1875 (GSI Type no.
2020, here refigured in Pl. 1, fig. 1a–c) from the Golden Oolite
of the same locality are considered to be the lectotype and
allotype of the present species, respectively. GSI Type nos. 2023,
16032, 16033, JUM 371 and JUM 505 are designated paralecto-
types.
Diagnosis. Shell compressed except for the inner whorls.
Strongly dimorphic, size ratio c. 2:1, maximum macro-
conch diameter 200 mm, sex ratio c. 1:4 in favour of
microconch. Intraspecific variability is great, especially in
microconchs. Shell with strongly variocostate ribbings,
strong and distant primaries with characteristic adoral
concavity in adult macroconch. Primaries shorten in late
stage in macroconch. Umbilical diameter shows positive
allometry during ontogeny.
A B C D
TEXT -F IG . 9 . Transverse section (body-chamber hatched) of
A, Kamptokephalites lamellosus (Sowerby), B–C, Eucycloceras opis
var. opis, and D, E. opis var. obscura. Note sharp umbilical edge
in inner whorls of both species. All · 0Æ5.
TEXT -F IG . 10 . Growth curves of A, degree of involution and
B, degree of inflation of Eucycloceras opis (J. de C. Sowerby).
TEXT -F IG . 11 . Size distribution of Eucycloceras opis (J. de C.
Sowerby). A, microconch. B, macroconch.
900 PALAEONTOLOGY, VOLUME 48

Range. Upper Lower Callovian–lower Middle Callovian.
Material. The present collection includes 137 microconch and
32 macroconch specimens, mostly shell remains. The micro-
conchs JUM 520, 533, 552, 591 come from Bed 8 of Jhura; JUM
373 from Bed 2, JUM 371, 381, 424, 514–515, 517–519, 522–
525, 532, 534, 541 from bed 5 and JUM 245–247, 251, 253, 256–
257, 261, 271, 273–274, 282, 366–367, 370, 372, 374–380, 382–
383, 417, 435 from Bed 6 of Keera; JUM 415, 429 from Bed 8
and 249, 309–310, 384–414, 419–421, 426–427, 432–433, 436–
439, 530, 538–540, 542–547, 549–551, 553–556, 567, 569–578,
580–581, 586, 592 from Bed 9 of Jumara; JUM 423, 531, 535–
537, 568, 579 from Bed 2 and JUM 252, 368, 416, 418, 434 from
Bed 3 of Jara. The macroconchs JUM 459, 505–513, 529, 560–
566 come from Bed 8 of Jhura; JUM 516, 518, 521 from Bed 5
and JUM 428 from Bed 6 of Keera; JUM 422 from Bed 7, JUM
430, 527–528 from Bed 8 and JUM 248, 425, 431, 548 from Bed
9 of Jumara; JUM 369, 557 from Bed 2 of Jara.
Measurements. Deposited on http://palass.org
Description. The species shows wide intraspecific variability
attributed mainly to genetic and ontogenetic factors. It displays
strong allometry during ontogeny. Early stage is represented by
relatively involute, depressed whorls; dense, fine, sharply crested
ribs with short and straight primaries furcating below mid-flank;
vertical umbilical wall with distinct umbilical margin (Text-
fig. 9). Intermediate stage up to end-phragmocone is typified by
evolute, compressed shell. Primary ribs strong and distant but
still sharp; furcation point shifts to above mid-lateral, i.e. prima-
ries become increasingly longer. Adult body-chamber is also
compressed and marked by strongly forwarded, curved with
adoral concavity at middle, blunt and distant primaries; they
gradually attenuate and finally disappear towards peristome; sec-
ondaries continue up to a little distance on body-chamber, then
disappear; umbilical edge rounded with inclined wall; shell more
evolute due to egression of bodywhorl.
Macroconchs generally cover the full ontogenetic pathways
and thus show low variability (Text-fig. 10). Curiously, micro-
conchs vary greatly and their developmental pathways seem to
be affected by heterochrony, especially paedomorphosis. Five dis-
A B
C
E
G
D
F
H
TEXT -F IG . 12 . Matured septal suture of Eucycloceras opis (J. de C. Sowerby). A–B, [m], E. opis var. obscura, redrawn from Spath
1928, pl. 38, fig. 5 and pl. 41, fig. 4c). C–D, [m], E. var opis, lectotype (C), redrawn from Spath 1928, pl. 36, fig. 2. E, E. opis var A. F,
E. opis var discoidea. G–H, [M] (G) allotype, redrawn from Waagen 1875, pl. 35, fig. 1c. E, external lobe; L, lateral lobe; U, umbilical
lobe. All · 1.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 901

tinct variants (infrasubspecies) have been recognized. All the
infrasubspecies are indistinguishable in early stage, but they rep-
resent in their adulthood different growth stages of the total
spectrum of macroconchiate ontogeny. However, none of the
microconchiate variants retains all the features of the species as
evident in macroconchs. They are generally relatively densely
ornate with retention of secondaries to the end of adult body-
chamber.
Microconch. Nucleus whorls remain depressed (B ⁄H ¼ 1Æ2–1Æ5) up to a diameter of about 18 mm, thereafter shell becomes
compressed. Flanks short and gently curved. Venter broad, mod-
erately arched, ventrolateral margin gently rounded. Whorl sec-
tion subcircular to ovate. Umbilicus relatively narrow (U ⁄D ¼0Æ20–0Æ24) with vertical to slightly overhanging wall, umbilical
margin distinct and sharp. Inner whorls are ornamented with
fine, sharp, densely spaced ribs. Primaries originate rursiradiately
from umbilical wall. They pass over flank straight with slight
forward inclination up to ventrolateral margin and cross venter
with further forward bend. Mostly biplicate ribs; however, trifur-
cating ribs and intercalatories are also present. Numbers of pri-
maries and secondaries per half whorl are 18–19 and 40–51,
respectively.
As the shell grows the furcation point shifts from below mid-
lateral to above. At the same time shell becomes compressed and
evolute, at end-phragmocone B ⁄H ¼ 0Æ69–0Æ80, and U ⁄D ¼0Æ24–0Æ33.
Adult shell is always compressed (B ⁄H ¼ 0Æ71–0Æ98), diameter
67–128 mm (Text-fig. 11), adult phragmocone diameter 45–
82 mm. Adult body-chamber occupies three-quarters of last
whorl. Shell egresses rapidly from the regular spiral before
attainment of adult phragmocone, and gives rise to an elliptic
curve of the umbilical seam and becomes more evolute
(U ⁄D ¼ 0Æ30–0Æ38). Adult shell has broad laterals which are flat
to slightly curved. Venter curved to nearly flat. Apertural outline
subtrapezoidal to subelliptical. Sharp umbilical margin with
steep wall continues up to early part of adult body-chamber but
near the last quarter or so umbilical wall becomes gradually slo-
ping and margin gently rounded. Ribbing pattern remains same
as observed in early whorls. Ribs also remain dense, fine and
sharp on adult body-chamber, but become increasingly promin-
ent and slightly distant or may be approximated towards peris-
tome. Ventral arch of secondaries is accentuated towards
aperture.
There exists a co-variation in shell size, coiling, degree of
inflation and ornamentation among the different microconchiate
variants: generally smaller variants are less evolute, compressed,
densely and finely ornate while large variants are more evolute,
inflated, and distantly and coarsely ornate. But, there are excep-
tions.
External lobe deeply incised; external saddle large, broad trifid.
The first lateral lobe less incised, narrow and trifid. The second
lateral lobe much smaller, trifid. U2 still smaller, asymmetric.
Other auxiliary lobes are well developed. Lobes from the first lat-
eral lobe up to the last auxiliary lobe are tied up in a large sad-
dle envelope which shows a little to a conspicuous rise towards
umbilical margin (Text-fig. 12A–E).
The variants within the microconchs generally differ from
each other on the basis of degree of involution and inflation,
and nature of ornamentation as observed on adult bodywhorl.
The following five variants (infrasubspecies) can be distinguished
and some of them are named according to different eucyclocera-
tin species of Spath (1928).
1. E. opis var. A (Pl. 2, figs 4–10). Smallest variant (D ¼67–83 mm, D at end-phragmocone ¼ 45–54 mm); most
compressed (B ⁄H ¼ 0Æ71–0Æ84); relatively less evolute
(U ⁄D ¼ 0Æ30–0Æ34); very densely, finely ornate until the
end (P ¼ 21–29, S ¼ 46–68); rib-crest sharp, symmetrical
in cross-section; ribs always isocostate and show approxi-
mation towards adult aperture. Umbilical wall remains
more or less steep in adult stage, umbilical margin always
prominent and sharp; abundant (32 specimens) but pre-
viously unknown.
2. E. opis var. opis (Pl. 3, figs 6–10). Small (D ¼88–113 mm, D at end-phragmocone ¼ 63Æ5–67 mm);
moderately compressed (B ⁄H ¼ 0Æ76–0Æ89); evolute
(0Æ31–0Æ35); densely, finely ornate (P ¼ 21–29, S ¼ 49–
66). Ribs isocostate, sharp, symmetrical crest; most abun-
dant (49 specimens).
3. E. opis var. obscura (Pl. 4, figs 1–6). Largest variant (D ¼121–128 mm, D at end-phragmocone ¼ 71–82 mm);
weakly compressed (B ⁄H ¼ 0Æ80–0Æ89); most evolute vari-
ant (U ⁄D ¼ 0Æ32–0Æ38); strongly, distantly ornate (P ¼17–27, S ¼ 40–53); ribs isocostate, high-crested, rounded
and asymmetrical with gentler slope towards aperture;
abundant (33 specimens).
4. E. opis var. discoidea (Pl. 3, figs 1–5). Small (D ¼ 77–
97 mm, D at end-phragmocone ¼ 52–70 mm); moder-
ately compressed (B ⁄H ¼ 0Æ75–0Æ89); evolute (U ⁄D ¼0Æ31–0Æ35); distantly ornate (P ¼ 17–18, S ¼ 36–45); ribs
isocostate with angular crest; rare (five specimens).
5. E. opis var. eucyclum (Pl. 5, figs 1–4). Small to medium-
sized (D ¼ 85–115 mm, D at end-phragmocone ¼73 mm); strongly to moderately compressed (B ⁄H ¼0Æ72–0Æ86); least evolute variant (U ⁄D ¼ 0Æ27–0Æ33);
EXPLANATION OF PLATE 3
Figs 1–10. Eucycloceras opis (J. de C. Sowerby) [m]. 1–5, var. discoidea. 1a–b, paralectotype, adult with partly preserved body-chamber,
GSI 16033, Smith coll., ‘sub-anceps bed ii’ Smith 1912–15, Ler-Hamundra. 2a–c, adult with partly preserved body-chamber, JUM
252, Bed 3, Jara. 3a–c, adult with partly preserved body-chamber, JUM 391, Bed 9, Jumara. 4a–b, complete adult, JUM 415, Bed
8, Jumara. 5, adult with partly preserved body-chamber, JUM 403, Bed 9, Jumara. 6–10, var. opis. 6a–c, almost complete adult
with broken phragmocone, JUM 388, Bed 9, Jumara. 7, complete adult, JUM 389, ibid. 8a–b, complete adult, JUM 416, Bed 3,
Jara. 9a–c, complete adult, JUM 411, Bed 9, Jumara. 10, complete adult, JUM 378, Bed 6, Keera; note egression of whorl takes
place in phragmocone stage. All · 0Æ5.
902 PALAEONTOLOGY, VOLUME 48

primaries broad, blunt, concave forward and fewer in
number (P ¼ 14–20); ribs variocostate, primaries much
stronger than secondaries (S ¼ 49–54) which become
faint to obscure on venter; common (18 specimens).
Macroconch. Shell large, relatively to strongly evolute
(U ⁄D ¼ 0Æ2–0Æ42), compressed (B ⁄H ¼ 0Æ67–0Æ96). Whorl sec-
tion elliptical to subrounded. Maximum adult shell diameter
200 mm, although reconstructed diameter of one specimen
PLATE 3
JANA et al., Eucycloceras
1a1b
X 2a2b 2c
X
3a 3b 3c
X4a
4b
5
X
X6a 6b 6c
7
8a
X
X
9a 9b 9c 10
X
8b
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 903

(JUM 425) appears to be c. 220 mm. Aperture missing; in one
specimen (Pl. 2, fig. 2) presence of dense growth lirae indicates
its almost complete preservation. Adult body-chamber occupies
three-quarters of the last whorl. End-phragmocone diameter
102–135 mm.
Inner whorls up to c. 50 mm in diameter, microconch-like,
barely distinguishable. Laterals gently curved with relatively wide,
broadly arched venter. Shell compressed. Umbilicus relatively
less evolute than microconch at corresponding diameter. Umbil-
ical margin sharp with steep wall. Ribs fine, dense; primaries rise
rursiradiately from umbilical shoulder, straight with forward
projection and furcate below mid-flank; secondaries have the
same forward bend, pass over rounded ventrolateral margin with
a further bend and cross venter with prominent forward projec-
tion. Number of primaries and secondaries on average are 19
and 46, respectively.
After 50 mm diameter up to the end-phragmocone, primaries
become progressively distant, increase in strength but remain
straight and sharp, thus strongly resembling the adult micro-
conch. The furcation point of primaries shifts as observed in the
microconch at a similar growth stage. Umbilical wall becomes
gradually sloping outward, margin narrowly rounded and shell
becomes more evolute. Number of primaries and secondaries
per half penultimate whorl is 16–21 and 51–62, respectively.
From the beginning of adult body-chamber, primaries become
broad, blunt and more distant with pronounced forward concav-
ity whereas secondaries continue up to the first quarter of body-
chamber length before they completely disappear. Subsequently
lengths of primaries shorten as they die out near outer lateral at
about one-third of the flank height from the ventral margin;
thus disappearance of both primaries and secondaries render
ventrolateral and siphonal areas of larger part of adult body-
chamber smooth. Umbilicus appears to be more evolute due to
egression of the seam, thus showing positive allometry and
broadly rounded umbilical margin. Laterals seem to be more
gently curved and venter becomes flattened, as observed in the
largest specimen.
The septal suture of the allotype (¼ Stephanoceras eucyclum
Waagen, 1875 by original designation; refigured here in Text-
fig. 12G), designated here, was described by Waagen (1875) and
reiterated by Spath (1928) and our recent inspection. The septal
suture of the macroconch is similar to that of the microconch;
however, the protracted aspect is less conspicuous in the macro-
conch.
Discussion. The macroconch of Eucycloceras opis has
strongly curved primary ribs on its adult body-chamber, a
feature not shown by Idiocycloceras. The species also dif-
fers in many other characters, as in the comparison
between Eucycloceras and Idiocycloceras that was highligh-
ted in the discussion of the latter.
The microconchs show wide intrasexual variability.
Remarkably, the whole spectrum shows continuous vari-
ation, with only a few exceptions. Failure to recognize
intraspecific variability and sexual dimorphism resulted
in excessive taxonomic splitting by early workers. We
have established that all the small eucycloceratin species
of Spath (1928) are no more than different micro-
conchiate variants of E. opis and most of the infrasub-
species have been named accordingly. Spath’s (1928)
Subkossmatia obscura and S. coggin-browni are so close
that they cannot be differentiated and we have con-
sidered these two species under one infrasubspecies,
obscura.
Callomon (in Donovan et al. 1981) mentioned the
existence of homeomorphism between Macrocephalitinae
and Mayaitinae in every morphological aspect. The pre-
sent eucycloceratin population offers another example of
homeomorphism as in Indonesian macrocephalitin
assemblages (see Westermann and Callomon 1988), but
in this case convergence occurs only in the microconchs.
Basic evolute and compressed shells characterize both
assemblages, thus the Middle Bathonian–Lower Callovian
macrocephalitins of Indonesia are very similar to eucyclo-
ceratins of the Indo-Madagascan Province. Other charac-
ters contributing to wide intraspecific variation within a
species population are remarkably similar. For example,
smaller shells are more compressed and less evolute than
larger ones; compressed shells are finely ornate while
inflated ones have coarse ornamentation; finely and
densely ribbed compressed shells have more projected
secondaries over the venter and a more raised umbilical
suture than their coarsely ornate counterparts. Wester-
mann and Callomon (1988) were confident that individu-
als from two different assemblages or stratigraphic levels
may look alike in isolation, but the overall population
structures vary. In our study, populations are frustratingly
similar, so it is not surprising that earlier workers such as
Spath (1928) and Arkell (1956) did not separate these
heterochronous groups. For example, they synonymized
Indonesian bifurcatus s.s. [m] with Indo-Madagascan Idio-
cycloceras (for details, see Westermann and Callomon
1988). It was Thierry (1978) who returned the Indonesian
forms to Macrocephalites where they were originally
EXPLANATION OF PLATE 4
Figs 1–6. Eucycloceras opis (J. de C. Sowerby) var. obscura [m], all adults with body-chamber. 1a–b, paralectotype, almost complete
with inner whorls exposed, GSI 2023, ‘anceps Zone’, Spath 1927–33, Keera. 2a–b, broken body-chamber with inner whorl
exposed, JUM 401, Bed 9, Jumara. 3a–c, paralectotype, GSI 16032, ‘sub-anceps beds’, Smith Coll. 1912–15, probably from Habye.
4a–b, JUM 545, Bed 9, Jumara. 5a–b, JUM 384, ibid. 6a–c, JUM 402, ibid. All · 0Æ5.
904 PALAEONTOLOGY, VOLUME 48

placed by Boehm (1912). Thierry, however, considered
that many of Spath’s (1928) Indonesian species (e.g.
forms ‘transitional to eucycloceratids’, ‘E. intermedium’
and ‘I. bifurcatum’) were geographical variants of M. sub-
trapezinus (Waagen) (¼ K. lamellosus of Spath 1928)
from Kutch, and Westermann and Callomon (1988) cor-
rectly pointed out differences in their degree of involution
and inflation. Repeated misidentifications have resulted in
1a 1b
X2a 2b
X3a 3b 3c
X 4a 4b
5aX
5b 6a
X6b
6c
JANA et al., Eucycloceras
PLATE 4
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 905

a notion of strong homeomorphism not only between
microconch bifurcatus s.s. and eucycloceratin microconchs
but also between the latter and the microconchiate sub-
compressus group of species. Due to the finite range of
shell coiling and functional constraints (Kennedy and
Cobban 1976) ammonites often developed similar forms
within monophyletic or unrelated groups. The conver-
gence of related groups is compounded if their stratigra-
phy is not adequately known. In their seminal paper
Westermann and Callomon (1988) separated the older
Indonesian forms (Middle Bathonian–Lower Callovian)
from the younger (Middle Callovian) descendant
stock (Eucycloceratinae) using subtle morphological dif-
ferences.
The neotype of M. bifurcatus trans. intermedius
(Westermann and Callomon 1988, pl. 7, fig. 1a–d) is clo-
sely comparable with our E. opis var. obscura [m]. M. keeu-
wensis, especially var. B [m] (Westermann and Callomon
1988, pl. 13, figs 1–2), on the other hand, strongly resem-
bles E. opis var. opis [m]. Westermann and Callomon
(1988) distinguished Indonesian forms, e.g. M. keeuwensis
var. B [m], by their sharp umbilical margin and dense and
sharp ribbing on the body-chamber. Abundant specimens
of the microconch of E. opis are now available. The popu-
lation structure encompasses the whole range of variability
of M. keeuwensis. In E. opis var. A the umbilical edge
remains sharp in the body-chamber. More sharp primary
ribs are present in var. A and var. opis. However, our
microconchs are more evolute and the degree of inflation
shows a wider range of variation. Westermann and Callo-
mon (1988) correctly noted that egression of whorls takes
place in the body-chamber of Indonesian microconchs
whereas it begins early in ‘Subkossmatia’. They wrongly
mentioned, however, that the holotype of ‘S. opis’ is fully
septate and still shows egression. Spath (1928, pl. 39,
fig. 2a) clearly showed the position of the adult end-
phragmocone which suggests that at least two-thirds of the
outer whorl represents the body-chamber. However, egres-
sion still occurs from the septate stage.
Close examination reveals that these two populations,
however, differ quantitatively in some characters. For
example, Eucycloceratinae are more evolute than Indo-
nesian macrocephalitins, as already mentioned, and they
constitute the most evolute forms within the family. Sec-
ondly, they have longer primaries in the outer whorl
and secondaries disappear from the body-chamber
instead of primaries, which are seen in macrocephalitin
macroconchs. Thirdly, egression of the umbilical seam in
Eucycloceratinae takes place from the phragmocone
stage. Finally, the nature of dimorphism completely sep-
arates the two assemblages. Eucycloceratin macroconchs
are quite different in appearance: they are evolute and
dominantly compressed.
The microconchs of Eucycloceras opis, especially var.
A and var. opis, closely resemble Dolikephalites subcom-
pressus. Their nucleus whorls are depressed and barely
distinguishable. E. opis var. A [m] resembles the interme-
diate stage of D. subcompressus in retaining the fine, dense
ribbing while E. opis var. opis resembles the outer whorl.
Morphometrically D. subcompressus and E. opis var. opis
[m] show similar allometric changes during ontogeny in
degree of inflation and involution (personal data based
on numerous topotypes of D. subcompressus); however,
var. opis is more evolute and compressed. Morphologi-
cally fine, dense secondaries with ventral projection char-
acterize both species. D. subcompressus has shorter
primaries throughout ontogeny while this feature is seen
in the early ontogeny of E. opis var. opis. Both species are
characterized by having a prominent umbilical margin
with a steep wall during early stages, which gives way to
sloping wall with rounded margin in adult whorls. E. opis
var. opis retains a sharp umbilical edge for a longer onto-
geny (see Text-fig. 9B–C) and in E. opis var. A it
continues to the end (see Pl. 2, fig. 7c). In Kutch, D. sub-
compressus ranges from the Lower Callovian and E. opis
ranges from the uppermost Lower Callovian–lower Mid-
dle Callovian.
Eucycloceratinae occurs predominantly in the Indo-
Madagascan region and is most abundant in Kutch. There
are, however, reports of their occurrences from the Hima-
layan region and Madagascar. Spath (1928) described
Subkossmatia flemingi (pl. 38, fig. 1a–c) from the doubtful
Callovian horizon of the Salt Range, Pakistan, on the
basis of a fragmented specimen with incompletely pre-
served body-chamber. It is evolute, larger and has coarse
ornamention, and thus may come close to our E. opis
var. obscura. However, based on his description it appears
that it has a smooth venter in the later stage with a roun-
EXPLANATION OF PLATE 5
Figs 1–4. Eucycloceras opis (J. de C. Sowerby) var. eucyclum [m], all adults with body-chamber. 1, JUM 424, Bed 5, Keera. 2a–c, nearly
complete, JUM 423, Bed 2, Jara. 3a–c, complete, JUM 517, Bed 5, Keera. 4a–b, complete, JUM 535, Bed 2, Jara. Note concave
primaries, variocostate ribbing in all lateral views and faint to obsolete secondaries in ventral views.
Figs 5–7. Idiocycloceras perisphinctoides Spath [m]. 5a–b, partly preserved bodywhorl with slightly depressed nucleus whorl, JUM 559,
Bed 3, Jara. 6a–c, complete adult, JUM 296, Bed 9, Jumara. 7, nearly complete adult, JUM 449, Bed 8, Jhura.
All · 0Æ5.
906 PALAEONTOLOGY, VOLUME 48

ded whorl section. Spath did not provide a ventral view
and we cannot determine whether this smoothness is bio-
logical or due to abrasion of the test. Its affinity is there-
fore uncertain.
Basse and Perrodon (1951) described three new spe-
cies of Subkossmatia from Madagascar. Among them, the
affinity of S. bassei and S. alfuricoides with the present
microconch is certain, but the population structure of
JANA et al., Eucycloceras
PLATE 5
X1
2a X 2b 2c
3a X 3b 3c
X4a
4b
5a 5b
6a 6b 6c X7
X
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 907

Madagascar is slightly different. They are relatively coar-
sely and distantly ribbed even in the smaller adult vari-
ant and relatively less evolute. This may be attributed to
geographical variation or small sample size (only three
specimens).
S. bassei (Basse and Perrodon, 1951, pl. 3, figs 10a–b,
12; pl. 6, fig. 6a–c) is a monotypic holotype and is an
almost completely preserved full-grown specimen. Its
sharp, isocostate ribbing, relatively steeper umbilical wall,
degree of inflation and size make it an intermediate
form between the two variants, opis and obscura. The
sharply crested ribs on the body-chamber and degree of
involution bring it close to opis whereas its less densely
ribbed ornamentation and relatively larger size place it
near obscura.
S. alfuricoides (ibid., pl. 4, fig. 3a–c) appears to be a
nearly complete adult shell and matches well with E. opis
var. opis in adult size and ornamentation but it is less
evolute on the body-chamber.
S. subfissum (ibid., pl. 6, fig. 2a–c) is difficult to corre-
late because the measurements provided do not tally with
the magnification provided for the plate. If the text meas-
urements are correct, it is a large form with coarser rib-
bing. It resembles our E. opis var. obscura.
Genus IDIOCYCLOCERAS Spath, 1928
Type species. Idiocycloceras perisphinctoides Spath, 1928.
Idiocycloceras perisphinctoides Spath (1928)
Plate 5, figures 5–7; Plates 6–10; Text-figures 13–18
*1875 Stephanoceras fissum (non Sowerby); Waagen,
p. 134, pl. 37, figs la–b only [M].
1894 Macrocephalites fissus (Waagen); Tornquist, p. 14,
18–19 [M].
1910 Macrocephalites fissus (Waagen); Uhlig, p. 265 [M].
1910 Macrocephalites fissus (Waagen); Lemoine, p. 22,
text-fig. 35, pp. 24, 37 (non p. 29) [M].
1913 Stephanoceras fissum (Waagen); Smith, b, p. 420; c,
p. 423 [M].
1914 Macrocephalites fissus (Waagen); R. Douville,
pp. 362–363 [M].
1928 Subkossmatia ramosa Spath, p. 214, pl. 39, fig. 1;
pl. 41, fig. 1a–b [M].
1928 Subkossmatia sp. ind. Spath, p. 215, pl. 3, fig. 1a–b
[M].
*1928 Idiocycloceras perisphinctoides Spath, p. 215, pl. 36,
fig. 3; pl. 38, fig. 3a–c [M].
*1928 Idiocycloceras singulare Spath, p. 216, pl. 28, fig. 8;
pl. 40, fig. 5; pl. 41, fig. 2 [M].
*1928 Idiocycloceras dubium Spath, p. 217, pl. 39, fig. 6a–
b [M].
1928 Idiocycloceras sp. ind. Spath, p. 218, pl. 35, fig. 6;
pl. 41, fig. 3 [M].
1933 Idiocycloceras singulare Spath, pl. 129, fig. 8; pl. 130,
fig. 8 [M].
non1951 Idiocycloceras Spath sp. ind.?; Basse and Perrodon,
p. 51, pl. 5, fig. 5a–b.
Nomenclature. Spath’s (1928) Idiocycloceras perisphinctoides,
I. singulare and I. dubium are combined here into a single spe-
cies and the name I. perisphinctoides, which is the senior subject-
ive synonym, is retained. Two larger Subkossmatia species of
Spath (1928) (S. ramosa and S. sp. indet.) are also considered to
be conspecific. S. ramosa has page priority over I. perisphincto-
ides, but the more familiar name of the type species is upheld.
Type specimens. Waagen’s (1875) ‘Stephanoceras fissum’ (GSI
Type no. 2029) was considered by Spath (1928) to be the holotype
of his Idiocycloceras singulare. The specimen is an adult shell with
a partly preserved body-chamber. The holotype of I. perisphincto-
ides is an immature specimen. We have, therefore, redesignated
the former as the lectotype of the present species and it is repro-
duced photographically for the first time (Pl. 9, fig. 1a–c). GSI
type nos. 16036, 16038, JUM 327, 489 are considered to be
paralectotypes.
Diagnosis. Shell strongly evolute, macroconch being the
most evolute within the family. Whorl shape variable,
ranging from depressed to compressed in adult whorl.
Shell very large, maximum diameter being 270 mm.
Dimorphic size ratio c. 2Æ5:1 and sex ratio c. 7:1 in favour
of macroconch. Shell coarsely ornate, mostly biplicate and
weakly isocostate. Primaries distant, blunt and straight
on adult body-chamber and finally die out near the
peristome. Secondaries continue up to half of adult body-
chamber length. Degree of involution shows negative
allometry in macroconch which becomes less evolute in
adult stage.
EXPLANATION OF PLATE 6
Figs 1–9. Idiocycloceras perisphinctoides Spath. 1a–c, [m], complete adult, JUM 270, Bed 6, Keera. 2, [m], complete adult, JUM 272,
ibid. 3a–c, [m], adult with partly preserved body-chamber, JUM 260, ibid. 4a–b, [m], adult with partly preserved body-chamber,
JUM 503, Bed 9, Jumara. 5a–c, [M], septate inner whorl with partly preserved body-chamber, JUM 440, Bed 3, Jara. 6, [M],
wholly septate inner whorl, JUM 538, Bed 8, Jumara; note deep crateriform umbilicus in nucleus whorl. 7a–c, [m], inner whorls,
JUM 526, Bed 8, Jhura. 8a–b, [M], wholly septate phragmocone, JUM 295, Bed 9, Jumara. 9a–b, [M], wholly septate
phragmocone, JUM 297, ibid. All · 0Æ5.
908 PALAEONTOLOGY, VOLUME 48
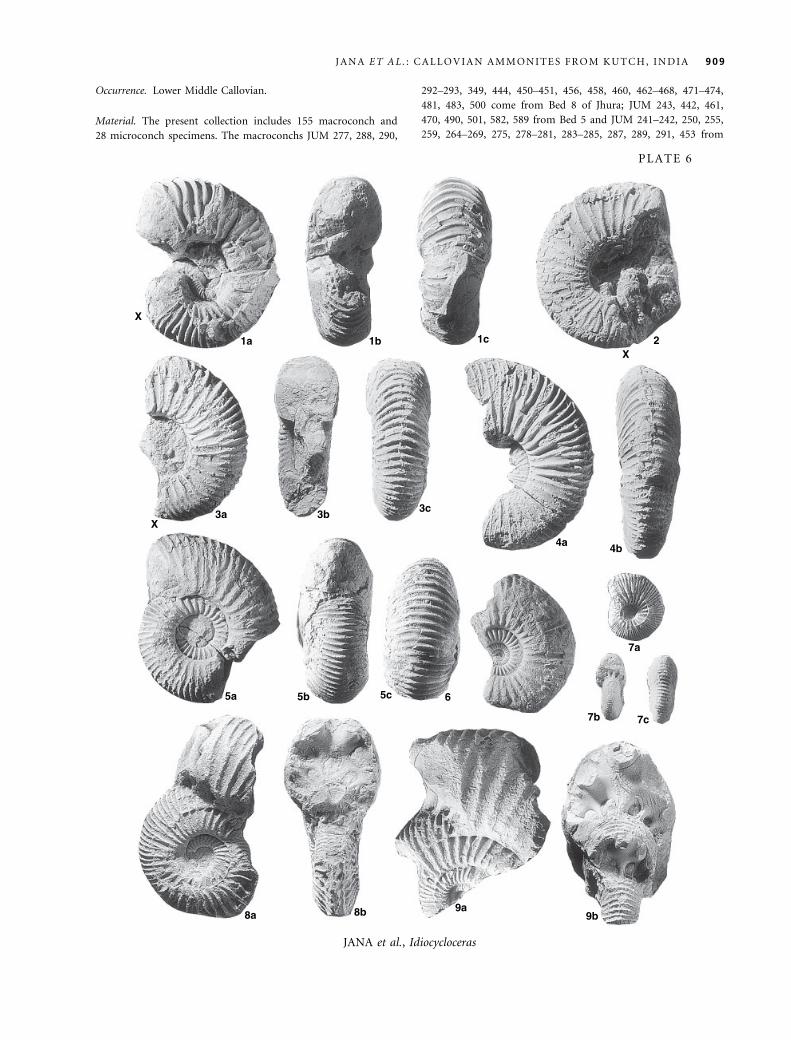
Occurrence. Lower Middle Callovian.
Material. The present collection includes 155 macroconch and
28 microconch specimens. The macroconchs JUM 277, 288, 290,
292–293, 349, 444, 450–451, 456, 458, 460, 462–468, 471–474,
481, 483, 500 come from Bed 8 of Jhura; JUM 243, 442, 461,
470, 490, 501, 582, 589 from Bed 5 and JUM 241–242, 250, 255,
259, 264–269, 275, 278–281, 283–285, 287, 289, 291, 453 from
JANA et al., Idiocycloceras
X
1a 1b 1cX
2
X3a 3b 3c
4a 4b
5a 5b 5c 6
7a
7b 7c
8a 8b 9a9b
PLATE 6
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 909

Bed 6 of Keera; JUM 319, 337, 361, 477, 487, 583 from Bed 8
and JUM 263, 276, 295, 297–305, 307–308, 311–313, 315–318,
320–322, 324–326, 339–342, 344–348, 350–351, 353, 355–360,
365, 454–455, 475–476, 478–480, 482, 485–486, 488–489, 491–
495, 497, 584–586, 590 from Bed 9 of Jumara; JUM 447–448,
452, 469, 496, 498 from Bed 2 and JUM 338, 440–441, 443,
445–446, 457, 484 from Bed 3 of Jara. The microconchs JUM
449, 502, 526 come from Bed 8 of Jhura; JUM 504 from Bed 5
and JUM 244, 254, 258, 260, 262, 270, 272, 323, 354 from Bed 6
of Keera; JUM 286, 294, 296, 306, 314, 327, 343, 352, 362–363,
503, 587 from Bed 9 of Jumara; JUM 558 from Bed 2 and JUM
364, 559 from Bed 3 of Jara.
Measurements. Deposited on http://palass.org
Description
Macroconch. The population shows high intraspecific variability.
Shell moderately to strongly evolute (U ⁄D ¼ 0Æ27–0Æ46), highlycompressed to strongly depressed (B ⁄H ¼ 0Æ74–1Æ84). Whorl out-
line subtrapezoidal to ovate. Shell very large, maximum adult
diameter being 270 mm. One adult body whorl fragment (JUM
365) appears to be a smaller variant with reconstructed diameter
of about 180 mm. The largest specimen (JUM 489, Pl. 12, fig. 1a)
with peristome preserved, shows dense growth striae near aper-
ture. Adult phragmocone diameter ranges from 135 to 192 mm.
Adult body-chamber occupies three-quarters of the last whorl.
Degree of inflation shows strong allometry during ontogeny
(Text-fig. 14). Inner whorls remain depressed at least up to a
diameter of about 50 mm. However, nucleus whorls are very
depressed (at 17 mm, B ⁄H ¼ 1Æ83; at 50 mm, B ⁄H ¼ 1Æ05–1Æ35). Flanks short, strongly curved; wide, gently arched venter
with gradual ventrolateral margin. After 50 mm diameter some
variants develop compressed whorl (B ⁄H ¼ 0Æ74–0Æ95) and
remain compressed to the end. Other variants also develop a
compressed whorl, show maximum compression at end-
phragmocone (B ⁄H ¼ 0Æ83–0Æ97), thereafter again become
depressed (B ⁄H ¼ 1Æ03–1Æ10). However, difficulties lie in separ-
ating them into two morphs because of the fact that there are
specimens that remain depressed throughout. The population
exhibits an overall decrease of B ⁄H ratio during ontogeny (neg-
ative allometry). On outer whorl, flanks may be broad, curved
to flattened. Venter with variable width and curvature. Ventro-
lateral margin gently rounded.
Umbilicus large, perisphinctoid to crateriform. There is a weak
correlation between whorl shape and nature of umbilicus, i.e.
compressed (B ⁄H c. 0Æ9) shells have evolute, perisphinctoid-type
shallow umbilicus (U ⁄D c. 0Æ4) while tumid variants with strongly
curved flanks (B ⁄H c. 1Æ35) are associated with relatively small,
deep, crateriform umbilicus (U ⁄D c. 0Æ28). Again, intermediates
exist. Degree of involution also shows allometry (Text-fig. 14). It
increases up to the end-phragmocone stage, and thereafter decrea-
ses. Umbilical wall is steep in early ontogeny and subsequently
tends to become inclined. Umbilical edge is sharp and distinct in
early stage, becomes gradually rounded during ontogeny.
Shell is strongly ornate, mostly biplicate and weakly varioco-
state. There is also a co-variation between degree of inflation
and strength of ribbing, i.e. ornamentation in inflated variants
is relatively stronger than that in compressed variants. Inner
whorls up to a diameter of about 40 mm are, however, orna-
mented with fine, sharp and moderately dense ribs. Primaries
originate rursiradiately from unbilical wall and are straight with
a slight forward inclination over flanks. Primaries are divided
into two branches. However, the position of furcation changes
during ontogeny. In early whorls, bifurcation takes place below
mid-lateral. At about 75 mm diameter bifurcation takes place
nearly at mid-flank, thereafter primaries become longer and
split at about two-fifths whorl height from the ventral margin.
Secondaries have the same forward projection up to ventrolat-
eral margin from where they are much more projected forward
across venter. Both primaries and secondaries sharply crested,
raised and asymmetric with gentler slope towards aperture up
to end-phragmocone stage. Primaries become progressively dis-
tant and blunt on adult body-chamber and near the last quar-
ter of body whorl they may become curved with slight adoral
TEXT -F IG . 13 . Size distribution of Idiocycloceras
perisphinctoides Spath. A, microconch. B, macroconch.
EXPLANATION OF PLATE 7
Figs 1–4. Idiocycloceras perisphinctoides Spath, [M]. 1a–c, paralectotype, wholly septate phragmocone, GSI 16038, ‘sub-anceps beds’,
Smith Coll. 1912–15, Ler-Hamundra. 2a–c, adult with partly preserved body-chamber, GSI 16035, ‘sub-anceps beds’, Smith Coll.
1912–15, Keera. 3a–c, paralectotype, immature phragmocone with partly preserved body-chamber, GSI 16036, ‘sub-anceps beds’,
Smith Coll. 1912–15, Habye. 4, smooth venter of an adult body-chamber, JUM 450, Bed 8, Jhura. All · 0Æ5.
910 PALAEONTOLOGY, VOLUME 48

concavity. Their strength attenuates, and they become flattened
to obsolete and finally die out leaving the peristome smooth,
but with dense growth striae. Ventral projection of secondaries
increases up to penultimate whorl. Secondaries continue
beyond adult phragmocone end, tend to be blunt and obsolete
first in venter, finally disappearing at about half of body-cham-
ber length rendering the siphonal and ventrolateral region of
the test smooth.
JANA et al., Idiocycloceras
1a1b 1c
2a
X
3a
3b 3c2b
2c 4
PLATE 7
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 911

The general pattern of septal sutures is projected towards
umbilicus with a broad typical saddle envelope of lobes like that
of Eucycloceras (Text-fig. 15). Umbilical projection and degree of
incision are variable. First lateral lobe is relatively larger, strongly
trifid and slightly asymmetric; second lateral lobe and auxiliary
lobes are distinctly developed.
Microconch. Microconchs resemble intermediate stage of
macroconch in many ways. Microconch population shows more
or less similar range of variation in degree of involution
(U ⁄D ¼ 0Æ32–0Æ44) and inflation (B ⁄H ¼ 0Æ8–1Æ18), but they
are less evolute and less depressed than macroconchs. Adult
whorl outline subtrapezoidal to subrounded (Text-fig. 16). Adult
shell diameter ranges from 93 to 131 mm; adult phragmocone
diameter is about 70 mm. Adult body-chamber occupies three-
quarters of the last whorl.
Inner whorls are indistinguishable from those of macroconch;
both are depressed and less evolute. However, in microconch,
depression continues up to 18 mm diameter. Degree of inflation
shows allometry similar to that of macroconch with increase in
shell diameter.
Umbilicus relatively large, shallow to moderately deep. Initial
vertical umbilical wall becomes inclined later, and margin, sharp
and distinct in early stages, becomes rounded in last stage.
Co-variation between degree of involution and degree of infla-
tion as observed in macroconch, however, is not evident, per-
haps because the growth of umbilicus does not replicate the
whole macroconch ontogeny and the degree of involution
resembles late to end-phragmocone stage of macroconch.
Ribbing pattern is similar to that of macroconch. Ribs are
fine, sharp and dense up to a diameter of about 50 mm. Adult
whorl is strongly, distantly ornate. Both primaries and secondar-
ies become blunt, rounded. Adoral convexity of secondaries over
venter becomes pronounced in the last quarter of the body-
whorl. Approximation of ribs takes place near peristome.
Septal sutures not clearly discernible.
Discussion. Idiocycloceras is the most evolute genus of all
the sphaeroceratids. It differs from Eucycloceras in having
a larger shell, more evolute whorl and coarser ornamenta-
tion. In the adult stage of Eucycloceras [M] primaries
become strongly curved, but in the same growth stage of
Idiocycloceras [M] primaries show little curvature. The
adult whorl section of Idiocycloceras is highly variable,
may be depressed; but the adult whorl of Eucycloceras is
always compressed. In the Idiocycloceras population the
sex ratio strongly favours the macroconch (M:m c. 7:1),
whereas the reverse trend is reflected in Eucycloceras pop-
ulation (M:m c. 1:4). The dimorphic size ratio (M:m) in
Idiocycloceras is approximately 2Æ5:1, whereas in Eucycloc-
eras it is 2:1.
Idiocycloceras bifurcatum reported by Spath (1928,
p. 206) from the Bathonian of Sula Island, Indonesia, was
incorrectly identified. Westermann and Callomon (1988)
re-identified it as a microconch of Macrocephalites bifurc-
atus s.s. It closely resembles the microconch of the present
species. They are comparable in degree of involution and
coarse ornamention. The adult size of M. bifurcates [m]
is 70–90 mm whereas that of I. perisphinctoides [m] is
93–131 mm. Moreover, in M. bifurcatus s.s. [m] primaries
bifurcate just below or at mid-flank at all growth stages, a
typical macrocephalitin character, but in I. perisphincto-
ides [m] they bifurcate above mid-flank at a later stage.
The microconch of the present species closely resembles
E. opis var. obscura [m], but I. perisphinctoides [m] is
coarser and is more evolute.
Basse and Perrodon (1951, pl. 5, fig. 5a–b) described a
specimen from Madagascar as Idiocycloceras sp. indet.
From their figure and description the specimen appears
to belong to Kamptokephalites. Collignon (1958, pl. 21,
fig. 86) illustrated, but did not describe, a specimen from
the upper Lower Callovian of Madagascar as Idiocycloceras
rebellyi. The specimen is a small evolute form with
depressed whorls and coarse ornamantion. Westermann
and Wang (1988) were unable to locate this specimen in
the Collignon (Dijon) and Rebelli (Paris) collections.
They doubtfully included it in their synonymy list of
Grayiceras? gucuoi (pl. 24, figs 7–14; text-fig. 12C–E) from
TEXT -F IG . 14 . Growth curves of A, degree of involution and
B, degree of inflation of Idiocycloceras perisphinctoides Spath.
EXPLANATION OF PLATE 8
Figs 1–2. Idiocycloceras perisphinctoides Spath [M], adults with partly preserved body-chamber and broken phragmocone. 1a–b, JUM
333, Bed 9, Jumara. 2a–b, specimen shown in Plate 7, figure 4. All · 0Æ5.
912 PALAEONTOLOGY, VOLUME 48

the Callovian of Tibet, which strongly resembles the
microconch of I. perisphinctoides.
Westermann and Wang (1988) suggested that the holo-
type of Idiocycloceras perisphinctoides is probably a micro-
conch and its supposed macroconch is I. singulare. We
have inspected both of the type specimens and noted
that I. perisphinctoides is a septate macroconch. They also
JANA et al., Idiocycloceras
1a
X
1b
2a
X
2b
PLATE 8
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 913

concluded (p. 331) that Idiocycloceras has a ‘trapezoidal,
rather than elliptical to subovate, and more compressed
section of the inner whorls.’ However, the inner whorls of
Idiocycloceras, which are up to 50 mm in diameter, are
always depressed (Text-fig. 16B–C).
DISCUSSION
Eucycloceratinae occupy an intermediate position between
the Macrocephalitinae and Mayaitinae (Text-fig. 8B).
Most workers consider that they constitute an evolution-
ary plexus, but the fossil record is marked by a great hia-
tus between the Eucycloceratinae and Mayaitinae
(comprising the entire Upper Callovian) (see Donovan
et al. 1981). Eucycloceratinae not only cryptically
A
C
E
B
D
TEXT -F IG . 15 . Septal sutures of Idiocycloceras
perisphinctoides Spath, [M]. A–B and D–E, redrawn
from Spath 1928, pl. 36, fig. 3; pl. 41, fig. 3; pl. 28,
fig. 8; and pl. 41, fig. 2, respectively. C, drawn from a
internal mould of a topotype (JUM 241). All · 1.
A B C
TEXT -F IG . 16 . Transverse sections (body-chamber hatched) of
A, Dolikephalites subcompressus (Waagen) and B–C, microconchs
of Idiocycloceras perisphinctoides Spath. Note sharp umbilical
edge and steep wall of inner whorls of D. subcompressus and
phragmocone of I. perisphinctoides. All · 0.5.
EXPLANATION OF PLATE 9
Figs 1–2. Idiocycloceras perisphinctoides Spath [M], adults with partly preserved body-chamber. 1a–c, lectotype, GSI 2029, Keera,
photographically presented for the first time. 2a–c, paralectotype, GSI 16037, ‘sub-anceps beds II’, Smith Coll. 1912–15, Ler-
Hamundra. All · 0Æ5.
914 PALAEONTOLOGY, VOLUME 48

appeared in the Indo-Madagascan region during the latest
Early Callovian, but flourished and diversified producing
the new genus Idiocycloceras, and then disappeared
equally suddenly during the early Middle Callovian. The
entire history of the subfamily spans less than the dur-
ation of a standard zone. Did they become extinct or
were they ecologically excluded? How did they acquire
novelties such as a protracted suture and an evolute,
compressed shell that their immediate macrocephalitin
ancestor in Kutch lack?
JANA et al., Idiocycloceras
1a
X
1b 1c
X
2a2b
2c
PLATE 9
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 915

The monographic treatment of Macrocephalites assem-
blages of south-west Pacific Indonesia (Westermann and
Callomon 1988) indicates that these oldest macrocephali-
tins show wide interspecific diversity, which was not
revealed by their younger Tethyan counterparts of both
Europe and the Indo-Madagascan region. The macro-
conchiate bauplan, however, shows low diversity and
remains conservative throughout the range of the sub-
family, while the microconchs show great phenotypic vari-
ation with respect to whorl inflation, degree of involution,
nature of ornamentation and, more importantly, septal su-
tural patterns. There may be strong to weak correlations
among these features which result in a plethora of forms
characterizing monospecific or successive assemblages.
Generally microconchiate forms are compressed and evo-
lute. There is a good correlation between forms having
dense, fine ribbing with projected secondaries over the
adult venter and possession of a raised umbilical saddle.
This correlation transcends taxonomic boundary and is
found in many species, e.g. M. bifurcatus [m] and
M. keeuwensis [m], especially var. B. This latter variant is
strongly compressed and evolute with ‘unusually high
(‘‘raised’’) umbilical saddles’ (see Westermann and Callo-
mon 1988, p. 69), thus resembling a ‘Subkossmatia’-like
microconch, e.g. our E. opis var. opis [m]. This homeo-
morphism created confusion for many early workers such
as Spath (1928) and Arkell (1956). Westermann and
Callomon (1988) provided the requisite stratigraphical
data and resolved the issue. These ancestral Indonesian
forms presumably migrated to the Tethys Ocean during
the Late Bathonian, which Callomon (1993) described as a
‘bio-event par excellence’. Upper Bathonian records of
both the European and the Indo-Madagascan areas wit-
nessed the cryptic appearance of Macrocephalites (Dietl
1981; Dietl and Callomon 1988; Datta et al. 1996) and not
at the base of the Callovian as previously thought (e.g.
Donovan et al. 1981). This evolutionary pattern in the
Tethyan region can be linked with the global marine trans-
gression at the start of the Callovian (Hallam 1981; Haq
et al. 1987). We speculate that during this migrational
event not all of the species participated. It may be sheer
chance that only involute, inflated forms with a radial
suture invaded the Tethyan area and subsequently diversi-
fied.
Subsequently, macrocephalitin evolutionary history
continued in two main biogeographical areas: European
Tethys and the Indo-Madagascan Province. It has gener-
ally been considered that they constituted two distinct
provinces, and macrocephalitin species in these had dif-
ferent modes of diversification (cf. Krishna and Cariou
1993), perhaps following vicariance biogeographical mod-
els (see Grande 1996). Occasionally, however, authors
have attempted to compare and synonymize contempora-
neous species of the two areas. For example, Westermann
and Callomon (1988) mentioned their close resemblance
and Krishna and Cariou (1993) found ‘undeniable simi-
larity’ between M. triangularis and M. jaquoti, although
Callomon et al. (1989) and Datta et al. (1996) had reser-
vations. M. jaquoti is the oldest macrocephalitin species
from the Lower Callovian of Europe and is slightly
younger than M. triangularis from the Upper Bathonian
of the Indo-Madagascan region (Datta et al. 1996). The
next youngest species, but still within the Indo-Madaga-
scan Upper Bathonian, is M. madagascariensis. It closely
corresponds to M. verus, which occupies a stratigraphical-
ly higher level than M. jaquoti in the Sub-Mediterranean
Western Tethys (Westermann and Callomon 1988;
Krishna and Cariou 1993). Likewise M. macrocephalus
(Schlotheim) appears to be very similar in grade of
organization (e.g. see Thierry 1978, pl. 9, figs 1–2) to the
contemporaneous M. formosus from the Indo-Madagascan
Faunal Province: both are similar in size and whorl-out-
line, and in having a smooth body-chamber. However,
some variants of the European species (e.g. Thierry 1978,
pl. 8, fig. 2a–b) have traces of weak secondaries restricted
to the adult venter, and the inner whorls of both species
are somewhat different (see also Westermann and Callo-
mon 1988; Bhaumik et al. 1993). Datta et al. (1996) found
similarities between M. formosus and M. cannizaroi. Both
show an early disappearance of primary ribs. Callomon
(1971) reiterated Spath’s (1927–33) view that the M. for-
mosus-M. madagascariensis group is the Indian equivalent
of M. macrocephalus (Schlotheim). The uppermost level
of the Lower Callovian in the Indo-Madagascan region
yields an endemic genus, Nothocephalites, marking the
highest subzone and faunal horizon. Significantly, at
a similar stratigraphical level in Europe, the species
M. gracilis appears, which is closely comparable to Notho-
cephalites (Westermann and Callomon 1988, p. 16). There
are also reports of faunal homogeneity of other species
between Kutch and Sub-Mediterranean France:
Indocephalites transitorius, I. kheraensis, I. chrysoolithicus,
Kamptokephalites lamellosus and K. dimerus are common
elements (Thierry 1978; Cariou 1984). Thus it appears
that the dominant macrocephalitin species of both geo-
graphical areas show a broad morphological homogeneity
EXPLANATION OF PLATE 10
Figs 1–3. Idiocycloceras perisphinctoides Spath [M]. 1a–c, adult with partly preserved body-chamber, JUM 331, Bed 9, Jumara. 2a–b,
adult body-whorl fragment of a small variant showing smooth venter, JUM 365, Bed 9, Jumara. 3a–b, adult with partly preserved
body-chamber, JUM 460, Bed 8, Jhura. All · 0Æ5.
916 PALAEONTOLOGY, VOLUME 48

and ‘parallel’ trends in evolution. Moreover, the micro-
conchiate forms (recognized in the Kamptokephalites-
Dolikephalites group of species) of both Europe and the
Indo-Madagascan region are remarkably similar (Thierry
1978; Westermann and Callomon 1988). Contrary to the
hypothesis of development of independent successions
(cf. Krishna and Cariou 1993) we believe that faunal simi-
larities speak for rapid migrational events and suggest the
JANA et al., Idiocycloceras
PLATE 10
1a
X1b 1c
2a
X
3a 2b 3b
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 917

existence of a well-established sea connection between
these two areas.
The Kamptokephalites-Dolikephalites group of species
has a considerable range in Kutch from the Lower Callo-
vian to the base of the Middle Callovian (Spath 1927–33;
pers. obs.), and thus have stratigraphical overlap with
eucycloceratins. Interestingly this group also extends up
to the lower Middle Callovian in Europe (Cariou 1984).
The younger transients in Kutch, however, show some
interesting developments. For example, ‘Stephanoceras
subtrapezinum’ Waagen (included in the synonymy list of
Kamptokephalites lamellosus by Spath 1928) and ‘Stepha-
noceras subcompressum’ Waagen (¼ Dolikephalites subcom-
pressus of Spath 1928) have a somewhat raised umbilical
saddle, as noted by Waagen (1875, pp. 138–139) and con-
firmed by our recent inspection of the specimens (GSI
nos. 2016 and 2018, respectively). We have already men-
tioned, as evident from the older macrocephalitins from
Indonesia, that there possibly exists a correlation between
a rise in umbilical saddle, fineness of ornamentation and
a compressed, evolute shell. It is possible that these chan-
ges relate to mutation in a pleiotropic gene (i.e. a single
gene that is responsible for a number of different pheno-
typic effects that are apparently unrelated). A mutation in
the younger transients of the K. lamellosus-D. subcompres-
sus group of species may have changed the nature of the
sutural pattern from radial to protracted. Later a similar
mutation may have resulted in complete differentiation of
a new stock, i.e. Eucycloceratinae, which are very similar
to the evolute Indonesian species. It now seems likely that
the eucycloceratin microconchs are heterochronous
homeomorphs of these older Indonesian microconchs (cf.
Westermann and Callomon 1988). Recurrence of homeo-
morphy is widespread in the stratigraphical record of fos-
sils. The innovation of the raised umbilical suture in the
older macrocephalitins of Indonesia during the Mid-
dle Bathonian, its disappearance in the species of late
macrocephalitin phylogeny, i.e. Late Bathonian–Early
Callovian stocks of Europe and Indo-Madagascan area,
and its reappearance in the eucycloceratin descendants
during the Middle Callovian, may be explained by a gen-
etic phenomenon. Like the development of the raised
umbilical suture, the virgatotome ribbing style is another
morphological innovation that appeared time and again
in the phylogeny of the Late Jurassic family Ataxiocerati-
dae, resulting in widespread homeomorphy. Callomon (in
Donovan et al. 1981, p. 127) explained the recurrence of
this feature as ‘the expression of a single gene that oscilla-
ted between dominant and recessive’. Callomon’s specula-
tion is now supported by some recent studies in
developmental biology, especially epigenetics, which sug-
gests that certain ‘sites’ in the genome of the developing
embryo evidently retain a dormant capacity to produce
features believed to have been lost during evolution. It is
now thought that genes are ‘switched off ’ due to altered
epigenetic pathways and may reappear in subsequent phy-
logeny, thus showing parallel and convergent evolution
(Fahræus 1987).
Nothocephalites, which is coeval with the older eucyclo-
ceratins of the late Early Callovian, was perhaps also
affected by a muted gene and is characterized by posses-
sing a raised umbilical saddle envelope (Spath 1928).
Krishna and Westermann (1987), however, mentioned
(also supported by our observations) that this is present
only in N. asaphus. The type specimen of N. semilaevis is
fully septate while that of N. asaphus has an incomplete
body-chamber. These two species are strictly contempora-
neous and were originally based on monotypic holotypes.
We have many adult specimens of both species which
show that N. asaphus is smaller and replicates intermedi-
ate-sized N. semilaevis, and their inner whorls are barely
distinguishable. Thus we strongly believe that N. asaphus
is a microconch (details will be published elsewhere).
However, Nothocephalites was short-lived. The effect of
initial mutation made significant phenotypic changes
since ornamentation became fine and dense and nucleus
whorls are indistinguishable from M. keeuwensis var. B
[m] and our E. opis var. opis [m], as noted by Spath
(1928) and Westermann and Callomon (1988). We have
already mentioned that we prefer to retain Nothocephalites
within the macrocephalitin clade since it is not suffi-
ciently differentiated in the eucycloceratin way. This is
also supported by the cladistic analyses reported herein. It
is worth noting that the raised nature of the umbilical
suture either appeared first, or became more prominent,
in the microconch in macrocephalitin phylogenetic
history, and finally played a major role in the evolution
of the eucycloceratin lineage. Here we must acknowledge
Spath (1924, 1927–33), who correctly envisaged that the
Eucyocloceras-Subkossmatia group owed its origin to the
subcompressus group of Dolikephalites, which is now con-
sidered to be a microconch (Thierry 1978; Westermann
and Callomon 1988).
In Eucycloceratinae, adult micro- and macroconchs
look quite different; in many respects the microconchs
resemble the inner whorls of macroconchs as well as adults
of the K. lamellosus-D. subcompressus group. Eucycloceras
and K. lamellosus-D. subcompressus assemblages have a
slight stratigraphical overlap near the Lower and Middle
Callovian substage boundary. The holotype of ‘Eucycloceras
eucyclum’ and our one microconch specimen of Eucycloc-
eras come from the Golden Oolite of Keera, which is high-
est Lower Callovian. At Jumara we have been able to
collect one macroconch of Eucycloceras from the semilaevis
horizon, which is the same level. On the other hand, sev-
eral specimens of the K. lamellosus-D. subcompressus group
come from the basal Middle Callovian (Spath 1933,
p. 715, Table 4; pers. obs.). Thus the two groups form two
918 PALAEONTOLOGY, VOLUME 48

successive assemblages and are eudemic (cf. Callomon
1985), but the evolution involves cladogenesis. Eucycloceras
is large and its evolution appears to have been induced by
hypermorphosis. Evolutionary novelties, unlike the older
macrocephalitins, include an evolute shell, longer, coarse
primaries with adoral concavity, and disappearance of sec-
ondaries, which makes the adult periphery smooth in the
macroconch. Idiocycloceras, which soon rapidly evolved
from Eucycloceras, is still larger, more evolute and has
coarser ornamentation, and the evolution shows a para-
morphocline trend. Both genera coexisted, flourished in
great abundance for a short time, but suddenly became
extinct (or were ecologically excluded).
The beginning of the Middle Callovian saw rapid diver-
sification of many evolute forms belonging to Reineckeia,
Choffatia and Indosphinctes. Thus the Stephanocerataceae
of the Lower Callovian gave way to the Periphinctaceae in
both Europe and Kutch. All of these forms are large with
an evolute shell. Possibly the eucycloceratins in Kutch
faced stiff competition from the invading groups. Reine-
ckeiins, which flourished in the Early Callovian in Eur-
ope, suddenly invaded the Indo-Madagascan region
(Cariou and Krishna 1988) and in Kutch they are found
in strata below the Lower Callovian ⁄Middle Callovian
boundary. Choffatins, on the other hand, were present
during the Early Callovian in both areas, but diversified
AX B C
TEXT -F IG . 17 . A–C, Idiocycloceras perisphinctoides Spath [M], adult having more than half a whorl body-chamber with inner whorls
exposed, JUM 332, Bed 9, Jumara; · 0Æ25.
A B C
TEXT -F IG . 18 . A–C, Idiocycloceras perisphinctoides Spath [M], adult body-whorl fragment, JUM 489, Bed 9, Jumara; note dense
growth striae at aperture; · 0Æ25.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 919

into many new species in the Middle Callovian in Kutch
(Spath 1928; pers. obs.). Furthermore, there was almost
certainly competition between Eucycloceras and the symp-
atrically derived Idiocycloceras. Sympatric groups generally
try to avoid competition by maintaining geographical seg-
regation or by ‘habitat proliferation’ within the same
region. We have already shown that both genera had the
same biogeographical distribution in Kutch and through-
out the Indo-Madagascan region (see also Basse and
Perrodon 1951; Donovan et al. 1981). However, in
the Subtethyan Himalaya, including Spiti and Tibet,
eucycloceratins have been reported from an assemblage
containing rare reineckeiins and choffatins (Pascoe 1959;
Krishnan 1982; Westermann and Wang 1988). These
might be areas where eucycloceratins migrated to avoid
competitive overlap. Pending studies of functional mor-
phology it is very difficult to know whether these two
genera exploited different ecological niches within the
same environment. However, indirect evidence suggests
that they competed for the same habitat. Eldredge (1974)
showed that while competing for the same ecological
niche two sympatric trilobite species displayed character
displacement. It appears that while Idiocycloceras
remained ‘neutral’, Eucycloceras (especially microconchs)
exhibited a ‘mixed reaction’. They diverged away and
even converged (e.g. E. opis var. obscura) with that of
Idiocycloceras (see ‘Discussion’ under Idiocycloceras peri-
sphinctoides). This perhaps explains the reason behind
the great infrasubspecific variation within the micro-
conchiate population of Eucycloceras. We do not seriously
consider the possibility of hybridization between them,
mainly on the grounds that the macroconchs are quite
distinct. Similarities in the nature of dimorphism between
two related sympatric taxa may complicate the situation
further. Similar size, the perisphinctid nature of coiling,
and bifurcating ribs persistent to the end may have
disrupted the ‘mate recognition’ system. Divergence of
Eucycloceras microconchs might have exaggerated the dif-
ferences in traits that distinguish them for mate recogni-
tion. It is interesting to note that microconchs of
reineckeiins and choffatins also closely resemble eucyclo-
ceratin microconchs with respect to the above features,
although they have lappets. One of the theoretical ecolog-
ical aspects of biological competition within a community
is that the size ratio for sympatric species is about 1:1Æ28(see Clarkson 1999, p. 49). Remarkably the two eucyclo-
ceratin species (including both dimorphs) correspond well
with this figure, as do all the major early Middle Callo-
vian species of Kutch. This suggests that eucycloceratins
successfully coexisted for some time, but were ultimately
excluded after a protracted struggle. Thus, it is possible
that because of the crisis from within, and stiff biological
competition from the reineckeiins and choffatins, the eu-
cycloceratins did not face extinction but were perhaps
ecologically excluded from the Indo-Madagascan region.
Two lines of evidence support this view. Firstly, the phys-
ical environment remained constant, an observation that
is based on the fact that similar lithofacies continue to
the top of anceps Zone (Datta 1992; Fursich et al. 1992),
and reineckeiins and choffatins remained equally diverse
and abundant. Secondly, a sudden faunal break in the eu-
cycloceratin lineage suggests ecological exclusion.
Where did they take refuge? Curently there is no evi-
dence that can shed light on this issue. It has already been
noted, however, that the Himalayan region, where both
macrocephalitin and eucycloceratin populations show
much homogeneity with that of the Kutch region, rarely
yields the perisphinctid group (especially reineckeiins),
which coexisted with eucycloceratins in the Indo-Madaga-
scan area. It is possible that, after being dramatically
excluded from the Indo-Madagascan area, the eucyclo-
ceratins thrived within a restricted geographical range in
the Himalayan region.
The Late Callovian history of Eucycloceratinae
remains unknown. The entire Himalayan region is
marked by a stratigraphical hiatus in which at least the
entire Upper Callovian is missing, although Kutch has a
near-continuous statigraphical record. The Mayaitinae
appeared everywhere during the Oxfordian, possibly rap-
idly, although the exact time is still debated. We suggest
that the Himalayan region is the centre of origin where
an isolated eucycloceratin population gave rise to May-
aitinae, since this area yields the highest diversity of
Mayaitinae, including Grayiceras. The Mayaitinae recall
the diversity of the older Macrocephalitinae of Indonesia
in that forms with radial and protracted sutures are
included. It is not surprising that the species with a
protracted suture, i.e. Paryphoceras badiense, is a homeo-
morph of E. opis var. opis [m], whereas the species with
a radial suture, i.e. Mayaites obesus, is a mayaitin equiv-
alent of Indocephalites diadematus. Soon the Mayaitinae
spread to the Kutch region, and like the Late Bathonian
migration of the Macrocephalitinae, not all of the forms
participated; Grayiceras remained Himalayan. Long-dis-
tance migration to the Pacific (South America) perhaps
resulted in the evolution of an entirely new subgenus,
Araucanites, within the Mayaitinae (Stipanicic et al.
1975).
Acknowledgements. A. Kayal (ONGC) and D. Mukherjee (GSI)
helped at various stages in the field. P. Rudra and P. Roy (JU)
assisted with computer analysis. S.S. Das (ISI) helped with pho-
tography. The Director, Curatorial Division, GSI, granted per-
mission for our study of the type material. Professor J. Hancock,
Dr N. Monks and an anonymous reviewer critically read the
manuscript and provided valuable suggestions. Two of us (SB
and KH) received financial aid from Department of Science and
Technology, India (ESS ⁄ 23 ⁄VES ⁄ 022 ⁄ 98).
920 PALAEONTOLOGY, VOLUME 48

REFERENCES
ARKELL , W. L. 1955. Validation under the plenary powers of
the family-group name ‘Mayaitidae’ Spath (L.F.), 1928 (Class
Cephalopoda, Order Ammoniides). International Commission
on Zoological Nomenclature, 11, 229–231.
—— 1956. Jurassic geology of the world. Oliver and Boyd, Edin-
burgh and London, 806 pp.
—— KUMMEL, B. and WRIGHT, C. W. 1957. Mesozoic
Ammonoidea. L80–L129. In MOORE, R. C. and TEIC-
HERT, C. (eds). Treatise on invertebrate paleontology. Part L.
Mollusca. Geological Society of America, Boulder, and Univer-
sity of Kansas Press, Lawrence, 490 pp.
BARDHAN, S., DATTA, K., JANA, S. K. and PRAMA-
NIK, D. 1994. Dimorphism in Kheraiceras Spath from the
Callovian Chari Formation, Kutch, India. Journal of Paleontol-
ogy, 68, 287–293.
—— GANGOPADHYAY, T. K. and MANDAL, U. 2002.
How far did India drift during the Late Cretaceous? Placentic-
eras kaffrarium Etheridge, 1904 (Ammonoidea) used as a
measuring tape. Sedimentary Geology, 147, 193–217.
—— SARDAR, S. and JANA, S. K. 2001. The Middle Jurassic
Khericeras Spath 1924 from the Indian Subcontinent. 265–277.
In SUMMESBERGER, H., HISTON, K. and DAURER,
A. (eds). Cephalopods – present and past. Abhandlungen der
Geologischen Bundesanstalt, A, 57.
BASSE , E. and PERRODON, M. 1951. Macrocephalitides de
Sud-Ouest de Madagascar Macrocephalitidae, Eucyclocratidae,
Mayaitidae recoltes de la mission E. Basse 1930–31. Memoires
de la Societe Geologique de France, Nouvelle Serie, 30, (3–4),
Memoire, 65, 1–100, 1–7 pls.
BHAUMIK, D., DATTA, K., JANA, S. K. and BARDHAN,
S. 1993. Taxonomy and intraspecfic variation of Macrocepha-
lites formosus (Sowerby) from the Jurassic Chari Formation,
Kutch, western India. Journal of the Geological Society of India,
42, 163–179.
BISWAS, S. K. 1977. Mesozoic rock-stratigraphy of Kutch,
Gujarat. Quarterly Journal of the Geological, Mineralogical, and
Metallurgical Society of India, 49, 1–51.
BLAKE, J. F. 1902. List of the types and figured specimens recog-
nized by C. D. Sherborn in the collection of the Geological Soci-
ety of London. London, 6 pp.
BOEHM, G. 1912. Beitrage zur Geoligie von Niederlandisch-
Indien. I. Abteilung; Die Sudkusten der Sula-Inseln Taliabu
und Mangoli. 4. Abschnitt; Unteres Callovien. Palaeontograph-
ica, Supplement, 4, 124–179.
BREMER, K. 1988. The limits of amino acid sequence data in
angiosperm phylogenetic reconstrution. Evolution, 42, 795–803.
BUCKMAN, S. S. 1909–30. Type ammonites. Wheldon and
Wesley, London, 790 pp.
CALLOMON, J. H. 1971. On the type species of Macrocepha-
lites Zittel, 1884 and the type specimen of Ammonites macro-
cephalus Schlotheim, 1813. Palaeontology, 14, 114–130.
—— 1985. The evolution of the Cardioceratidae. Special Papers
in Palaeontology, 33, 49–90, pls 1–3.
—— 1993. On Perisphinctes congener Waagen, 1875, and the age
of the Patchman Limestone in the Middle Jurassic of Jumara,
Kutch, India. Geologische Blatter fur Nordost-Bayern, 43, 227–
246.
—— DIETL, G. and PAGE, K. N. 1989. On the ammonite
faunal horizons and standard zonations of the Lower Callo-
vian Stage in Europe. In ROCHA, R. B. and ZEISS , A.
(eds). Second International Symposium on Jurassic Stratigraphy,
1987, 1, 359–376.
CARIOU, E. 1984. Biostratigraphic subdivisions of the Callo-
vian Stage in the Subtethyan Province of ammonites, correla-
tions with the Subboreal zonal scale. 315–326. In
MICHELSEN, O. and ZEISS , A. (eds). International Sym-
posium on Jurassic Stratigraphy (IUGS), 1, Erlangen.
—— and KRISHNA, J. 1988. The Tethyan Reineckeiinae of
Kachchh and Jaisalmer (West India): systematic, biostrati-
graphic and biogeographic implications. Palaeontographica,
Abteilung A, 203, 149–170.
CLARKSON, E. N. K. 1999. Invertebrate palaeontology and
evolution. Fourth edition. Blackwell Science, Oxford, xiv + 452
pp.
COLLIGNON, M. 1958. Atlas des fossiles caracteristiques de
Madagascar. Fascicule 1 (2), pls 1–33. Service Geologique
Tananarive.
DAS , S. S., BARDHAN, S. and LAHIRI , T. 1999. The Late
Bathonian gastropod fauna of Kutch, western India – a new
assemblage. Paleontological Research, 3, 268–286.
DATTA, K. 1992. Facies, fauna and sequence: an integrated
approach in the Jurassic Patcham and Chari formations,
Kutch, India. Unpublished PhD thesis, Jadavpur University,
Kolkata, India, 167 pp.
—— BHAUMIK, D., JANA, S. K. and BARDHAN, S.
1996. Age, ontogeny and dimorphism of Macrocephalites
triangularis Spath – the oldest macrocephalitid ammonite
from Kutch, India. Journal of the Geological Society of India,
47, 447–458.
DIETL , G. 1981. Uber Macrocephalites (Ammonoidea) aus dem
Aspidoides – Oolith und die Bathonium ⁄Callovium Grenzs-
chichten der Zollernalb (SW-Deutschland). Stuttgarter Beitrage
zur Naturkunde, Series B, 68, 1–15.
—— and CALLOMON, J. H. 1988. Der Orbis-Oolith (Ober-
Bathonium, Mittl. Jura) von Sengenthal ⁄Opf., Frank. Alb, undseine Bedeutung fur die Korrelation und Gliederung der
Orbis-Zone. Stuttgarter Beitrage zur Naturkunde, Series B, 142,
1–31.
DOMMERGUES, J. L., CARIOU, E., CONTINI , D.,
HANTZPERGUE, P., MARCHAND, D., MEISTER, C.
and THIERRY, J. 1989. Homeomorphies et canalizations
evolutives: le role de l’ontogenese. Quelques examples pres
chez les ammonites du Jurassique. Geobios, 22, 5–48.
DONOVAN, D. T., CALLOMON, J. H. and HOWARTH,
M. K. 1981. Classification of the Jurassic Ammonoidea. In
HOUSE, M. R. and SENIOR, J. R. (eds). The Ammonoidea.
Systematics Association, Special Volume, 18, 101–155.
DOUVILLE , R. 1914. Ammonites remarquables ou peu con-
nues (1ere note). Bulletin of the Geological Society of France,
13, 359–369, pl. 7.
ELDREDGE, N. 1974. Character displacement in evolutionary
time. American Zoologist, 14, 1083–1097.
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 921

FAHRÆUS, L. E. 1987. Darwinian evolution and developmen-
tal biology: a brief review of current ideas. Geoscience Canada,
13, 155–164.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies: an
approach using the bootstrap. Evolution, 39, 783–781.
FURSICH, F. T. and OSCHMANN, W. 1993. Shell beds as
tools in basin analysis: the Jurassic of Kachchh, western India.
Journal of the Geological Society, London, 150, 169–185.
—— OSCHMANN, W., S INGH, I. B. and JAITLY , A. K.
1992. Hardgrounds, reworked concretion levels and condensed
horizons in the Jurassic of western India: their significance for
basin analysis. Journal of the Geological Society, London, 149,
313–331.
—— PANDEY, D. K., CALLOMON, J. H., OSCHMANN,
W. and JAITLY, A. K. 1994. Contribution to the Jurassic of
Kachchh, western India. II. Bathonian stratigraphy and deposi-
tional environment of the Sadhra Dome, Patchchham Island.
Beringeria, 12, 95–125.
GRANDE, L. 1996. Vicariance biogeography. 448–451. In
BRIGGS , D. E. G. and CROWTHER, P. R. (eds). Palaeobi-
ology – a synthesis. Blackwell Science, Oxford, xiii + 583 pp.
GREGORY, J. W. 1900. The corals. Palaeontologia India, Geo-
logical Survey of India, Series 9, Memoir, 2, 12–196.
HALDER, K. 2000. Diversity and biogeographic distribution of
Jurassic nautiloids of Kutch, India during fragmentation of
Gondwana. Journal of African Earth Sciences, 31, 175–185.
HALLAM, A. 1981. Facies interpretation and the stratigraphic
record. Freeman, Oxford, 291 pp.
HAMMER, O., HARPER, D. A. T. and RYAN, P. D. 2003.
PAST – Palaeontological Statistics, version 0.98. (Computer
Software).
HAQ, B. U., HARDENBOL, J. and VAIL , P. R. 1987. Chro-
nology of fluctuating sea levels since the Triassic. Science, 235,
1156–1166.
JAIN, S., CALLOMON, J. H. and PANDEY, D. K. 1996. On
the earliest known occurrence of the Middle Jurassic ammonite
genus Reineckeia in the Upper Bathonian of Jumara, Kachchh,
western India. Palaontologische Zeitschrift, 70, 129–143.
JAITLY, A. K., SZABO, J. and FURSICH, F. T. 2000. Con-
tributions to the Jurassic of Kachchh, western India VII. The
gastropod fauna. Part I. Pleurotomarioidea, Fissurevoidea,
Trochoidea and Eucycloidea. Beringeria, 27, 31–61.
JANA, S. K. 2002. Macrocephalitinae and Eucycloceratinae of
the family Sphaeroceratidae (Ammonoidea) and other ancil-
lary taxa from the Middle Jurassic of Kutch, western India:
systematics, phylogeny and evolution. Unpublished PhD the-
sis, Jadavpur University, Kolkata, 163 pp.
—— and DAS, S. S. 2002. A report of a 157.8 m. y.-old dinosaur
bone from the Jurassic marine Chari Formation, Kutch, Gujarat
and its taphonomic significance. Current Science, 82, 1–4.
—— BARDHAN, S. and SARDAR, S. K. 2000. Kheraiceras
Spath (Ammonoidea) – new forms and records from the Mid-
dle Jurassic sequence of the Indian Subcontinent. Paleontologi-
cal Research, 4, 205–225.
KAYAL, A. and BARDHAN, S. 1998. Epistrenoceras Bentz
(Ammonoidea) from the Middle Jurassic of Kutch: a new
record and its chronostratigraphic implication. Canadian Jour-
nal of Earth Sciences, 35, 931–935.
KENNEDY, W. J. and COBBAN, W. A. 1976. Aspects of
ammonite biology, biogeography and biostratigraphy. Special
Papers in Palaeontology, 17, 94 pp.
KLINGER, H. C. and KENNEDY, W. J. 1989. Cretaceous
faunas from Zululand and Natal, South Africa. The ammonite
subfamily Placenticeratidae Hyatt, 1900; with comments on
the systematic position of the genus Hypengonoceras Spath,
1924. Annals of the South African Museum, 98, 241–408.
KRISHNA, J. 1984. Current status of the Jurassic stratigraphy
of Kachchh, western India. 731–742. In MICHELSEN, O.
and ZEISS , A. (eds). International Symposium on Jurassic
Stratigraphy, 3, Erlangen.
—— and CARIOU, E. 1993. The Tethyan Macrocephalitinae:
evolutionary environmental and dispersal strategies. Geobios,
15, 217–226.
—— and THIERRY, J. 1987. Discovery of the genus Erymnoc-
eras from the Middle Callovian of Kachchh, western India:
paleontological, stratigraphical and paleobiological implica-
tions. Newsletters on Stratigraphy, 17, 71–78.
—— and WESTERMANN, G. E. G. 1987. Faunal associations
of the Middle Jurassic ammonite genus Macrocephalites in
Kachchh, western India. Canadian Journal of Earth Sciences,
24, 1570–1582.
KRISHNAN, M. S. 1982. Geology of India and Burma. CBS
Publishers and Distributors, Delhi, 536 pp.
LEMOINE, P. 1910. Paleontologie de Madagascar. VIII.
Ammonites du Jurassique superieur du Cercle d’Analalava
(Madagascar). Annals of Paleontologie, 5, 137–168, 5 pls.
MITRA, K. C., BARDHAN, S. and BHATTACHARYA, D.
1979. A study of Mesozonic stratigraphy of Kutch, Gujarat
with a special reference of rock-stratigraphy and bio-stratigra-
phy of Keera dome. Bulletin of Indian Geologists’ Association,
12, 129–143.
MONKS, N. 2000. Functional morphology, ecology, and evolu-
tion of the Scaphitaceae Gill, 1871 (cephalopoda). Journal of
Molluscan Studies, 66, 205–216.
—— 2002. Cladistic analysis of a problematic ammonite group:
the Hamitidae (Cretaceous, Albian–Turonian) and proposal
for new cladistic terms. Palaeontology, 45, 689–707.
MUKHERJEE , D., BARDHAN, S., DATTA, K. and
GHOSH, D. N. 2003. The terebratulid Kutchithyris (Brachio-
poda) from the Jurassic sequence of Kutch, western India –
revisited. Palaeontological Research, 7, 111–128.
—— —— and GHOSH, D. N. 2002. Two new species of Cryp-
torhynchia Buckman (Brachiopoda) from the Middle Jurassic
of Kutch, India. Alcheringa, 26, 209–231.
OLDHAM, R. D. 1893. Geology of India (Medlicott and Bland-
ford). Stratigraphical and structural. Geological Survey of India,
Calcutta, xxiii + 543 pp.
PANDEY, D. K. and FURSICH, F. T. 1993. Contributions
to the Jurassic of Kachchh. I. The coral fauna. Beringeria, 8,
3–69.
PARENT, H. 1997. Ontogeny and sexual dimorphism of Eury-
cephalites gottschei (Tornquist) (Ammonoidea) of the Andean
Lower Callovian (Argentine-Chile). Geobios, 30, 407–419.
PASCOE, E. H. 1959. A manual of the geology of India and
Burma. Volume 2. Geological Survey of India, Calcutta, pp.
485–1343.
922 PALAEONTOLOGY, VOLUME 48

RAJNATH. 1932. A contribution to the stratigraphy of Cutch.
Quarterly Journal of the Geological, Mineralogical and Metallur-
gical Society of India, 4, 161–174.
ROMAN, F. 1938. Les Ammonites jurassiques et cretacees. Essai
de genre. Masson, Paris, 554 pp.
SMITH, J. H. 1912a. Notes on the Cutch ammonites. Journal
of the Bombay Natural History Society, 21, 709–715.
—— 1912b. Notes on the Cutch ammonites. II: Samatra. Journal
of the Bombay Natural History Society, 21, 1347–1352.
—— 1913a. Notes on the Cutch ammonites. III: Keera Hill.
Journal of the Bombay Natural History Society, 22, 209–213.
—— 1913b. Notes on the Cutch ammonites. IV: Ler-Hamundra
Ellipse. Journal of the Bombay Natural History Society, 22,
417–421.
—— 1913c. Cutch ammonite beds. Journal of the Bombay Nat-
ural History Society, 22, 421–423.
—— 1914a. Oppeliae teneylobatae (?) in Cutch. Holocosteph-
anus? in Cutch. Journal of the Bombay Natural History Society,
22, 812–814.
—— 1914b. Notes on the Cutch ammonites. V: east of Ler.
Journal of the Bombay Natural History Society, 23, 176–179.
—— 1915. Notes on the Cutch ammonites. VI: the Habye
Hills. Journal of the Bombay Natural History Society, 23, 793–
798.
SOWERBY, J. de C. 1840. Memoir to illustrate a geological
map of Kutch. Transactions of the Geological of Society of Lon-
don, 5, 327–329.
SPATH, L. F. 1923. Appendix on ammonites from New
Zealand. In TRECHMANN, C. T. The Jurassic rocks of New
Zealand. Quarterly Journal of the Geological Society of London,
79, 286–312.
—— 1924. On the Blake collection of ammonites from Kachh
(India). Palaeontologia Indica, Geological Survey of India, New
Series 9, Memoir, 1, 1–129.
—— 1927–33. Revision of the Jurassic cephalopod fauna of
Kachh (Cutch). Palaeontologia Indica, Geological Survey of
India, New Series 9, Memoir, 2, 1–945.
STIPANICIC, P. N., WESTERMANN, G. E. G. and RIC-
CARDI , A. C. 1975. The Indo-Pacific ammonite genus
Mayaites in the Oxfordian of the southern Andes. Revista de
la Asociacion Paleontologica Argentina, 12, 281–305.
SWOFFORD, D. L. 1998. PAUP: Phylogenetic Analysis Using
Parsimony, version 4.0 b. 10. Smithsonian Institute (Compu-
ter Software).
THIERRY, J. 1978. Le genre Macrocephalites au Callovien infe-
rieur (ammonites Jurassique moyen). Memoire de Geologie de
Universite de Dijon, 4 490 pp.
TORNQUIST , A. 1894. Uber Macrocephaliten im Terrain-a-
chailles. Abhandlungen der Schweizerischen Palaontologischen
Gesellschaft, 21, 27, 2 pls.
UHLIG, V. 1910a. The fauna of the Spiti Shales. Palaeontologia
Indica, Series 15, 4, 2, 133–306, 30 pls.
—— 1910b. The fauna of the Spiti Shales. Palaeontologia Indica,
Series 15, 4, 3, 307–395, 30 pls.
WAAGEN, W. 1875. Jurassic fauna of Kutch, the Cephalopoda.
Palaeontologia Indica, Geological Survey of India, Series 9,
Memoir, 1, 1–247.
WESTERMANN, G. E. G. 1956. Monographie der Bajocien –
Gattungen Sphaeroceras und Chondroceras (Ammonoidea).
Beihefte zum Geologischen Jahrbuch, 24, 1–125.
—— and CALLOMON, J. H. 1988. The Macrocephalitinae
and associated Bathonian and Early Callovian (Jurassic)
ammonoids of the Sula Islands and New Guinea. Palaeonto-
graphica, Abteilung A, 203, 1–90.
—— and HUDSON, N. 1991. The first find of Eurycephaliti-
nae (Jurassic Ammonitina) in New Zealand and its biogeo-
graphic implications. Journal of Paleontology, 65, 689–693.
—— and WANG YI-GANG 1988. Middle Jurassic ammonites
of Tibet and the age of the Lower Spiti Shales. Palaeontology,
31, 295–339.
WYNNE, A. B. 1872. Memoir on the geology of Cutch to
accompany a map complied by A. B. Wynne and F. Fedden,
during the seasons 1867–68 and 1868–69. Memoirs of the Geo-
logical Survey of India, 9, 1–29.
APPENDIX
1. Data matrix used to create the cladograms in Text-figure 8A–B
Character numbers refer to the list in Table 1.
Taxa
Characters
1 6 11 16 21
Otoitidae 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Sphaeroceratinae 0 0 0 0 0 0 1 1 1 0 0 0 0 1 1 1 1 1 0 1 1 0 0 0 0
Eurycephalitinae 0 0 0 0 0 0 1 1 1 0 0 1 1 1 0 1 0 1 0 0 0 0 0 0 0
Macrocephalitinae 0 0 0 0 0 0 1 1 1 1 1 1 1 0 1 2 1 2 0 1 0 1 1 0 0
Eucycloceratinae 0 0 0 0 0 0 1 1 1 1 1 1 1 0 1 3 1 3 0 0 0 1 2 1 1
Mayaitinae 0 0 0 0 0 0 1 1 1 1 1 1 1 0 1 4 1 2 0 0 0 1 1 2 0
JANA ET AL . : CALLOVIAN AMMONITES FROM KUTCH, INDIA 923

2. Data matrix used to create the cladogram in Text-figure 8C
Character numbers refer to the list given in Table 3.
Taxa
Characters
1 5 9 13 17 21 25
Macrocephalites 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Nothocephalites 0 0 0 0 0 0 0 0 0 1 0 1 0 1 0 1 1 0 0 1 1 0 0 0 1 0 0
Eucycloceras 0 0 0 0 0 0 0 0 1 1 1 2 1 2 1 2 1 1 1 2 1 1 1 1 2 1 1
Idiocycloceras 0 0 0 0 0 0 0 0 2 0 1 2 1 2 1 2 1 0 1 2 0 0 2 1 2 2 2
924 PALAEONTOLOGY, VOLUME 48




















