
Neuroimaging of cortical development and brain connectivity in human newborns and animal models
11
REVIEW Neuroimaging of cortical development and brain connectivity in human newborns and animal models Gregory A. Lodygensky, 1,2 Lana Vasung, 2,3 Ste ´ phane V. Sizonenko 2 and Petra S. Hu ¨ ppi 2 1 NICU, Department of Pediatrics, University of Geneva, Geneva, Switzerland 2 Division of Development and Growth, Department of Pediatrics, University of Geneva, Switzerland 3 Croatian Institute for Brain Research, School of Medicine, University of Zagreb, Zagreb, Croatia Abstract Significant human brain growth occurs during the third trimester, with a doubling of whole brain volume and a fourfold increase of cortical gray matter volume. This is also the time period during which cortical folding and gyrification take place. Conditions such as intrauterine growth restriction, prematurity and cerebral white matter injury have been shown to affect brain growth including specific structures such as the hippocampus, with subsequent potentially permanent functional consequences. The use of 3D magnetic resonance imaging (MRI) and dedicated postprocessing tools to measure brain tissue volumes (cerebral cortical gray matter, white matter), surface and sulcation index can elucidate phenotypes associated with early behavior development. The use of diffusion tensor imaging can further help in assessing microstructural changes within the cerebral white matter and the establishment of brain connectivity. Finally, the use of functional MRI and resting-state func- tional MRI connectivity allows exploration of the impact of adverse conditions on functional brain connectivity in vivo. Results from studies using these methods have for the first time illustrated the structural impact of antenatal conditions and neonatal intensive care on the functional brain deficits observed after premature birth. In order to study the pathophysiology of these adverse conditions, MRI has also been used in conjunction with histology in animal models of injury in the immature brain. Understanding the histological substrate of brain injury seen on MRI provides new insights into the immature brain, mechanisms of injury and their imag- ing phenotype. Key words DTI; hypoxic-ischemic; inflammatory; MRI; newborn; preterm. Introduction Understanding human brain development is clinically rele- vant as many neurobiological disorders and disabilities have their origin in early structural, functional development and plasticity. With the advent of magnetic resonance imaging (MRI), it has become possible to address the question of where, when and how adverse conditions in fetal and early postnatal life and prematurity relate to the maturation of cortical networks (Ment et al. 2009). MRI has entered the research arena due to its non-invasiveness and potential for high-resolution structural brain imaging. It has allowed for the first time the study of in-vivo brain development in pre- mature and healthy newborn infants providing full 3D imaging data sets.The integration of healthy infants in lon- gitudinal studies has become possible thanks to the absence of irradiation or other side-effects, thus allowing a wide and thorough analysis of brain development. This review presents advanced imaging modalities used to study the immature brain and discusses their potential for the in-vivo assessment of cerebral development in both human as well as animal models established to study the pathophysiology of early adverse conditions for brain development. Conventional magnetic resonance imaging Conventional MRI techniques, including mainly T1- and T2- weighted images, allow the assessment of brain develop- ment in vivo with the highly sensitive assessment of gray and white matter contrast, as well as the differentiation of unmyelinated and myelinated white matter (Battin et al. Correspondence Petra S. Hu ¨ ppi, Division of Development and Growth, Department of Pediatrics, University Hospital of Geneva, Rue Willy Donze ´ 6 1211 Geneva 14, Geneva, Switzerland. E: [email protected] Accepted for publication 12 July 2010 ª 2010 The Authors Journal of Anatomy ª 2010 Anatomical Society of Great Britain and Ireland J. Anat. (2010) 217, pp418–428 doi: 10.1111/j.1469-7580.2010.01280.x Journal of Anatomy
-
Upload
affective-sciences -
Category
Documents
-
view
3 -
download
0
Transcript of Neuroimaging of cortical development and brain connectivity in human newborns and animal models

REVIEW
Neuroimaging of cortical development and brainconnectivity in human newborns and animal modelsGregory A. Lodygensky,1,2 Lana Vasung,2,3 Stephane V. Sizonenko2 and Petra S. Huppi2
1NICU, Department of Pediatrics, University of Geneva, Geneva, Switzerland2Division of Development and Growth, Department of Pediatrics, University of Geneva, Switzerland3Croatian Institute for Brain Research, School of Medicine, University of Zagreb, Zagreb, Croatia
Abstract
Significant human brain growth occurs during the third trimester, with a doubling of whole brain volume and
a fourfold increase of cortical gray matter volume. This is also the time period during which cortical folding
and gyrification take place. Conditions such as intrauterine growth restriction, prematurity and cerebral white
matter injury have been shown to affect brain growth including specific structures such as the hippocampus,
with subsequent potentially permanent functional consequences. The use of 3D magnetic resonance imaging
(MRI) and dedicated postprocessing tools to measure brain tissue volumes (cerebral cortical gray matter, white
matter), surface and sulcation index can elucidate phenotypes associated with early behavior development. The
use of diffusion tensor imaging can further help in assessing microstructural changes within the cerebral white
matter and the establishment of brain connectivity. Finally, the use of functional MRI and resting-state func-
tional MRI connectivity allows exploration of the impact of adverse conditions on functional brain connectivity
in vivo. Results from studies using these methods have for the first time illustrated the structural impact of
antenatal conditions and neonatal intensive care on the functional brain deficits observed after premature
birth. In order to study the pathophysiology of these adverse conditions, MRI has also been used in conjunction
with histology in animal models of injury in the immature brain. Understanding the histological substrate of
brain injury seen on MRI provides new insights into the immature brain, mechanisms of injury and their imag-
ing phenotype.
Key words DTI; hypoxic-ischemic; inflammatory; MRI; newborn; preterm.
Introduction
Understanding human brain development is clinically rele-
vant as many neurobiological disorders and disabilities have
their origin in early structural, functional development and
plasticity. With the advent of magnetic resonance imaging
(MRI), it has become possible to address the question of
where, when and how adverse conditions in fetal and early
postnatal life and prematurity relate to the maturation of
cortical networks (Ment et al. 2009). MRI has entered the
research arena due to its non-invasiveness and potential for
high-resolution structural brain imaging. It has allowed for
the first time the study of in-vivo brain development in pre-
mature and healthy newborn infants providing full 3D
imaging data sets.The integration of healthy infants in lon-
gitudinal studies has become possible thanks to the absence
of irradiation or other side-effects, thus allowing a wide
and thorough analysis of brain development. This review
presents advanced imaging modalities used to study the
immature brain and discusses their potential for the in-vivo
assessment of cerebral development in both human as well
as animal models established to study the pathophysiology
of early adverse conditions for brain development.
Conventional magnetic resonance imaging
Conventional MRI techniques, including mainly T1- and T2-
weighted images, allow the assessment of brain develop-
ment in vivo with the highly sensitive assessment of gray
and white matter contrast, as well as the differentiation of
unmyelinated and myelinated white matter (Battin et al.
Correspondence
Petra S. Huppi, Division of Development and Growth, Department of
Pediatrics, University Hospital of Geneva, Rue Willy Donze 6 1211
Geneva 14, Geneva, Switzerland. E: [email protected]
Accepted for publication 12 July 2010
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
J. Anat. (2010) 217, pp418–428 doi: 10.1111/j.1469-7580.2010.01280.x
Journal of Anatomy

1998; Counsell et al. 2002; Ferrie et al. 1999). The immature
white matter demonstrates a relatively homogenous low
signal on T1-weighted images and a high signal on T2-
weighted images compared with gray matter (Fig. 1A). This
is mainly due to the higher water content of the immature
white matter. With increasing maturation, the signal of the
white matter increases on T1-weighted images, which is
related to multiple concurrent processes occurring in the
developing white matter with decreasing water content, a
change in the water : macromolecule ratio caused by the
arrival of the lipid precursors of myelination, and finally the
process of myelination itself. Myelination is visualized at dif-
ferent rates and times on T1-weighted and T2-weighted
images with evidence of myelination determined by a low
signal on T2-weighted images and a high signal on T1-
weighted images that is already present at 28 weeks of ges-
tation in regions such as the inferior and superior cerebellar
peduncles or the ventrolateral nuclei of the thalamus. Mye-
lination of the posterior limb of the internal capsule is seen
by 36 weeks with a completion of myelination by 2 years of
age (Battin et al. 1998; Counsell et al. 2002).
Brain MRI of developing animals in correlation to histo-
logical findings gives us the opportunity to better define
the substrates of MRI changes during development and
following injury. The possibility of imaging mouse pups,
although technically challenging, opens a new area of
research where specific changes in imaging can be
addressed using genetically modified animals (West et al.
2009). The majority of correlation studies between MRI
and histology in the developing brain have been carried
out on rat pups at 7–14 days of life, an age closer to
infants born at term. Imaging of immature animals gives
the same contrast in conventional imaging (Fig. 1B) with
an inversion of contrast similar to the human as the ani-
mals mature (Lodygensky et al. 2008a). Ultra-high field
MRI scanners are increasingly used in rodent studies and
are now available at up to 14 T, equivalent to 300 000
times the earth’s magnetic field. They provide a sufficient
signal-to-noise ratio to achieve the required high resolu-
tion in immature rodents but bring new challenges such as
the assembly of stable coils at the given magnetic fields,
worsening susceptibility artifacts and magnetic field inho-
mogeneity.
Since the discovery of the transient subplate zone
(Kostovic & Molliver, 1974) and its significance for the nor-
mal development of the human telencephalon (Kostovic &
Rakic, 1990a), there has been an extensive amount of
effort invested in identification of the subplate by MRI
methods (Kostovic et al. 2002; Rados et al. 2006; Huang
et al. 2006; Prayer et al. 2006; Kasprian et al. 2008; Kosto-
vic & Vasung, 2009). The subplate zone can be clearly dis-
tinguished in ex-vivo T1 MRI after 13 weeks of gestation
as an area of hypointensity (Kostovic et al. 2002; Rados
et al. 2006; Kostovic & Vasung, 2009) and on in-vivo T2
MRI as an area of hyperintensity (Prayer et al. 2006)
between the bordering cortical plate and the intermediate
zone (Fig. 2). The presence of this transient subplate zone
is one of the main signs of cortical immaturity (Kostovic &
Vasung, 2009). It represents the major site of the endoge-
nous circuitry reorganization (Kostovic & Jovanov-Milos-
evic, 2006; Volpe, 2009). The subplate is crucial in
developmental processes such as the ingrowth of thalamo-
cortical axons (Kostovic & Jovanov-Milosevic, 2006), reorga-
nization of the fetal white matter and establishment of
cortical layers. The subplate is rich in extracellular matrix
and serves as a ‘waiting’ compartment for the thalamo-
cortical and cortico-cortical connections (Kostovic & Rakic,
1990b). Its neurons are known to contribute, as early as
24 weeks of gestation, to the appearance of the first
evoked potentials (Hrbek et al. 1973), which play an
important role in the functional maturation of the human
brain. Furthermore, after 32 weeks of gestation, in parallel
A B
Fig. 1 (A) Preterm infant born at 29 weeks of gestation scanned at term equivalent, coronal T2-weighted section with cerebral grey matter and
basal ganglia hypointense and non myelinated white matter with a higher water content appearing hyperintense. (B) Rat pup at 9 days of life,
coronal T2-weighted section with the same contrast as the preterm infant with hypointense cerebral grey matter and hyperintense cerebral white
matter. Image acquired at 9.4 Tesla in collaboration with Gregory Lodygensky and Rolf Gruetter (CIBM, Ecole polytechnique federale de Lausanne,
Switzerland).
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al. 419
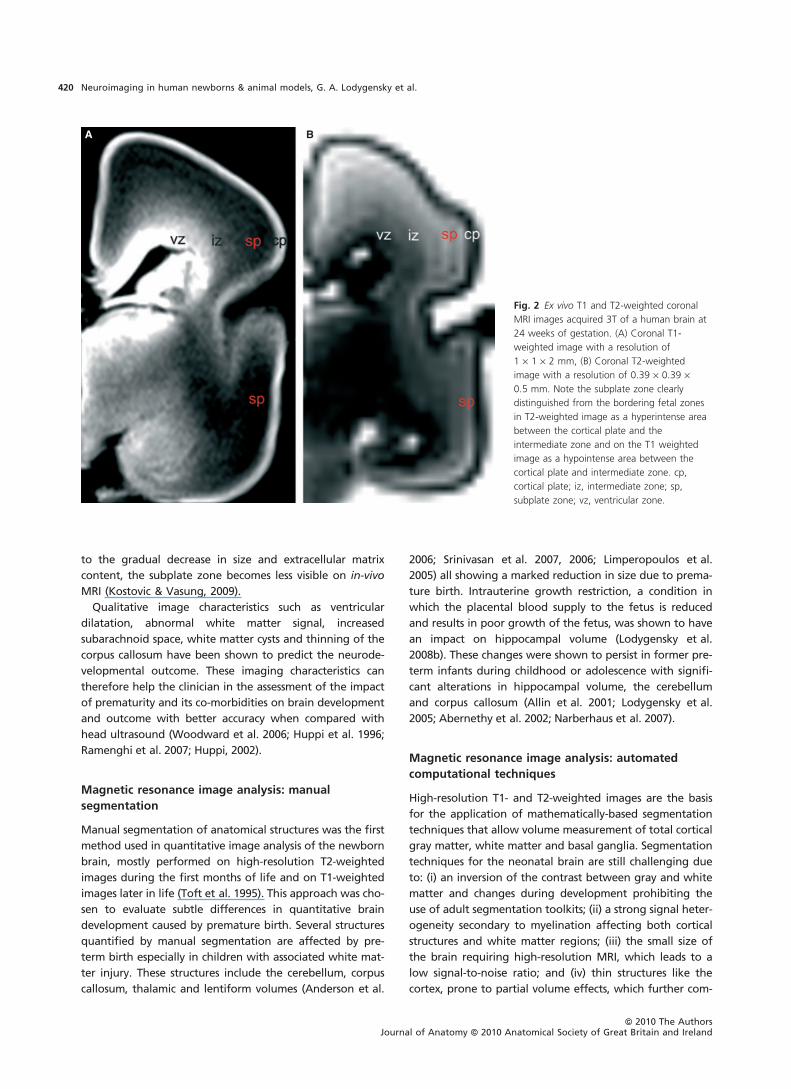
to the gradual decrease in size and extracellular matrix
content, the subplate zone becomes less visible on in-vivo
MRI (Kostovic & Vasung, 2009).
Qualitative image characteristics such as ventricular
dilatation, abnormal white matter signal, increased
subarachnoid space, white matter cysts and thinning of the
corpus callosum have been shown to predict the neurode-
velopmental outcome. These imaging characteristics can
therefore help the clinician in the assessment of the impact
of prematurity and its co-morbidities on brain development
and outcome with better accuracy when compared with
head ultrasound (Woodward et al. 2006; Huppi et al. 1996;
Ramenghi et al. 2007; Huppi, 2002).
Magnetic resonance image analysis: manual
segmentation
Manual segmentation of anatomical structures was the first
method used in quantitative image analysis of the newborn
brain, mostly performed on high-resolution T2-weighted
images during the first months of life and on T1-weighted
images later in life (Toft et al. 1995). This approach was cho-
sen to evaluate subtle differences in quantitative brain
development caused by premature birth. Several structures
quantified by manual segmentation are affected by pre-
term birth especially in children with associated white mat-
ter injury. These structures include the cerebellum, corpus
callosum, thalamic and lentiform volumes (Anderson et al.
2006; Srinivasan et al. 2007, 2006; Limperopoulos et al.
2005) all showing a marked reduction in size due to prema-
ture birth. Intrauterine growth restriction, a condition in
which the placental blood supply to the fetus is reduced
and results in poor growth of the fetus, was shown to have
an impact on hippocampal volume (Lodygensky et al.
2008b). These changes were shown to persist in former pre-
term infants during childhood or adolescence with signifi-
cant alterations in hippocampal volume, the cerebellum
and corpus callosum (Allin et al. 2001; Lodygensky et al.
2005; Abernethy et al. 2002; Narberhaus et al. 2007).
Magnetic resonance image analysis: automated
computational techniques
High-resolution T1- and T2-weighted images are the basis
for the application of mathematically-based segmentation
techniques that allow volume measurement of total cortical
gray matter, white matter and basal ganglia. Segmentation
techniques for the neonatal brain are still challenging due
to: (i) an inversion of the contrast between gray and white
matter and changes during development prohibiting the
use of adult segmentation toolkits; (ii) a strong signal heter-
ogeneity secondary to myelination affecting both cortical
structures and white matter regions; (iii) the small size of
the brain requiring high-resolution MRI, which leads to a
low signal-to-noise ratio; and (iv) thin structures like the
cortex, prone to partial volume effects, which further com-
A B
Fig. 2 Ex vivo T1 and T2-weighted coronal
MRI images acquired 3T of a human brain at
24 weeks of gestation. (A) Coronal T1-
weighted image with a resolution of
1 · 1 · 2 mm, (B) Coronal T2-weighted
image with a resolution of 0.39 · 0.39 ·0.5 mm. Note the subplate zone clearly
distinguished from the bordering fetal zones
in T2-weighted image as a hyperintense area
between the cortical plate and the
intermediate zone and on the T1 weighted
image as a hypointense area between the
cortical plate and intermediate zone. cp,
cortical plate; iz, intermediate zone; sp,
subplate zone; vz, ventricular zone.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al.420

plicate segmentation. The combination of a signal-based
k-means classification with a mathematical morphology
approach for shape recognition is currently the preferred
method for neonatal brain segmentation (Anbeek et al.
2008; Cachia et al. 2003; Weisenfeld & Warfield, 2009).
The recent development of specific newborn brain seg-
mentation tools has allowed the quantification of in-vivo
human brain growth showing a fourfold increase in corti-
cal gray matter, a fivefold increase in myelinated white
matter, a linear steady growth rate of the unmyelinated
white matter (Huppi et al. 1998b) and a 70% growth of
the basal ganglia (Mewes et al. 2006). These major
changes underline the importance of this period in brain
growth and its vulnerability as shown by the toxicity of
dexamethasone on cortical gray matter growth (Murphy
et al. 2001) or intrauterine growth restriction with a
reduction of 30% in cortical gray matter volume (Borra-
dori Tolsa et al. 2004). This approach has shown for the
first time that white matter injury in preterm infants was
associated with a significant impact on cortical gray
matter development (Inder et al. 1999, 2005).
Mathematical morphology: primary cortical folding
One of several possible approaches to investigating cortical
folding in preterm infants is based on a mathematical mor-
phology approach that processes and analyzes shape. The
computational approach quantifies both surface area and
cortical gyration through curvature measurements from 3D
reconstruction of the interface between the developing
cortex and white matter (Dubois et al. 2008a,b). The inner
cortical surface is identified between the cortical gray mat-
ter and white matter. A smooth triangle-based mesh of this
surface is then computed and the global area of this inner
cortical surface can be measured. Finally, the local surface
curvature is estimated from the mesh local geometry; posi-
tive curvatures correspond to the gyrus’s top and negative
curvatures to the fold’s bottom (Cachia et al. 2003). The
sulci are then defined as connected components of nega-
tive curvature. These sophisticated image analysis tools
need high-resolution primary input data with no motion
artifacts.
Cortical folding in preterm infants
In the human brain, the morphology of cortical gyri and
sulci is complex and variable among individuals with an
established asymmetry appearing very early on (Dubois
et al. 2010). A significant amount of cortical folding and
gyrification takes place during the last trimester with a
steep increase of brain surface and degree of sulcation
index (Dubois et al. 2008b). Conditions such as twinning
and intrauterine growth restriction have been shown to
alter sulcation and decrease the sulcation index (Dubois
et al. 2008a) (Fig. 3). Alterations in size and cortical
morphology might reflect abnormal functioning or vice
versa, as shown in correlations between surface, gyrification
index at birth and the Assessment of Preterm Infants’
Behavior score at term equivalent age (Dubois et al. 2008a),
a neonatal neurobehavioral test shown to predict later neu-
rofunctional outcome (Feldman & Eidelman, 2006). So far,
the precise mechanisms responsible for such alterations in
cortical phenotype during intrauterine or postnatal devel-
opment are still poorly understood and therefore studies of
cortical folding in selected animal models will provide fur-
ther insight into the mechanisms underlying cortical folding
(Barnette et al. 2009) (Fig. 4).
Diffusion tensor imaging
The technique of diffusion tensor imaging
Diffusion tensor imaging (DTI) assesses water molecule dis-
placement at a microstructural level with displacement of
water in the order of 10 lm. This technique has been used
in the exploration of the structural basis of white matter
development (Neil et al. 1998; Huppi et al. 1998a; Mukher-
jee et al. 2002) and cortical maturation (McKinstry et al.
2002a).
Fig. 3 3D representation of the inner cortical surface for a singleton
and a twin of equivalent age. SI1 represents the average of the
sulcation index. Note the altered cortical gyrification shown to be
significantly affected in twins when compared to singleton of the
same gestational age. Modified from Dubois et al. (2008a).
A
B
Fig. 4 (A) Cortical surface of ferret brains at 4, 10, 17 days of life and
in an adult. (B) Cortical surface of human brains at 25, 30, 33,
39 weeks of gestation and in an adult. Courtesy of Barnettte et al.
(Barnette et al., 2009).
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al. 421

Based on the tensor model, the main DTI parameters are
the three eigenvalues k1, k2 and k3 representing diffusion
along the three principal axes of an ellipsoid in each voxel.
Eigenvector maps and RGB color-coded maps indicate the
orientation of the major eigenvector providing an indica-
tion of the direction in which water diffusion is highest
(typically parallel to white matter fiber fascicles). The first
eigenvalue, often referred to as axial diffusivity, was shown
to be affected by axonal integrity (Kim et al. 2006). The sec-
ond and third eigenvectors describe diffusivity perpendicu-
lar to the axial diffusivity. Radial diffusivity, defined by the
average of the second and third eigenvectors, was shown
to be affected in adult animals by changes in myelin
ensheathment (Song et al. 2005). From these three eigen-
values is calculated the apparent diffusion coefficient (ADC)
and mathematical measures of anisotropy describing the
degree to which water diffusion is restricted in one direc-
tion relative to all others, referred to as fractional aniso-
tropy (FA) and relative anisotropy (RA) (Fig. 5). FA and RA
are indicators of the degree of water diffusion anisotropy
with a value that is equal to zero for diffusion equal in all
directions and increases with anisotropy.
Fiber tracking: diffusion tensor imaging
Fiber tracking is able to delineate specific cerebral white
matter tracts by following local vector orientation from the
3D vector field (Fig. 6). Different tractography algorithms
have been developed such as the popular streamline deter-
ministic fiber tracking (Mori et al. 1999). The 3D fiber track
is allowed to continue unless it enters a region of FA less
than a predefined value of FA or turns at an angle greater
than a predefined angle between two consecutive voxels.
Fiber crossing causes one of the major downfalls of fiber
tracking. Several algorithms have been proposed to solve
this issue such as the probabilistic index of connectivity
(Parker et al. 2003) or, more recently, the Gibbs tracking
model (Kreher et al. 2008).
Diffusion tensor imaging: analysis strategies
The four main approaches to analyzing DTI data are
region-of-interest quantification, voxel-based approaches,
histogram analyses and tract-based analysis. Manual region-
of-interest selection and quantification is the most fre-
quently used. This method is time-consuming and is charac-
terized by a large inter-rater variability due to a bias in the
selection of the regions of interest. Voxel-based morphome-
try is an alternative method initially designed to quantify
regional changes in volume on conventional imaging
(Ashburner & Friston, 2001; Kesler et al. 2008; Nosarti et al.
2008). Its automated analysis eliminates a-priori knowledge
and user bias but it is not immune to error due to registra-
tion and normalization. Tract-based spatial statistics is an
alternative method (Smith et al. 2006) developed to address
errors in registration by the tools built for conventional
imaging used on DTI data. It is based on the realignment of
the FA maps of all subjects onto a FA map skeleton.
Diffusion tensor imaging studies
During normal brain development, the ADC of the white
matter was shown to decrease together with a steady
increase in anisotropy (Huppi et al. 1998a; Neil et al. 1998;
Dudink et al. 2007). Following a hypoxic–ischemic injury,
ADC also decreases, resulting in values that are much
reduced compared with the age- and region-dependent
normal values (Rutherford et al. 2004; McKinstry et al.
2002b). This ADC restriction is clinically used to detect brain
injury as it appears very early on, preceding conventional
T2 hyperintensity. During white matter development, a
gradual decrease in diffusion is observed principally in k2
and k3 (and much less in k1), which reflect changes in
water diffusion perpendicular to white matter fibers in sin-
gle coherently ordered fiber bundles and may indicate
A B
Fig. 5 (A) Healthy preterm infant born at 29 weeks of gestation
imaged at term equivalent, coronal FA map at the level of the
posterior limb of the internal capsule. (B) P5 live rat pup, coronal FA
map with sufficient in plane resolution to identify major white matter
bundles such as the corpus callosum or the internal capsule. Image
acquired at 11.7 Tesla in collaboration with Gregory Lodygensky and
Jeffrey J Neil (Washington University, St Louis, Missouri).
A B
Fig. 6 (A) 3D representation of the cortical surface of an infant at
term with superimposed fiber tracking through the corpus callosum.
Courtesy Jessica Dubois (CEA/SAC/DSV/DRM/NeuroSpin/Cognitive
Neuroimaging Unit, Gif-sur-Yvette, France) (B) Relative anisotropy map
of a fixed rat brain at 21 days of age with the superimposed fiber
tracking through the corpus callosum. Image acquired at 9.4 Tesla in
collaboration with Gregory Lodygensky and Rolf Gruetter (CIBM, Ecole
polytechnique federale de Lausanne, Switzerland).
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al.422

changes due to premyelination (change of axonal width)
and myelination (Mukherjee et al. 2002; Partridge et al.
2004; Giorgio et al. 2008). The increase in white matter
anisotropy values during development appears to take
place in two stages. The first increase takes place before
the histologic appearance of myelin (Huppi et al. 1998a;
Neil et al. 1998). This increase has been attributed to
changes in white matter structure that accompany the ‘pre-
myelinating state’ (Wimberger et al. 1995). This state is
characterized by an increase in the number of microtubule-
associated proteins in axons, a change in axon caliber, and
the maturation and organization of oligodendrocytes. It is
also associated with changes in the axonal membrane, such
as an increase in conduction velocity and changes in
Na+ ⁄ K+-ATPase activity. The increase in anisotropy associ-
ated with premyelination is notable in that it takes place in
the absence of changes in T1- or T2-weighted imaging as
well as before the histologic appearance of myelin. Regio-
nal anisotropy is not only clearly influenced by myelination
alone, but also by factors such as axon packing, relative
membrane permeability to water, internal axonal structure
and intra-axonal space as outlined in recent studies of ani-
mal spinal cord using AxCaliber, a model of water diffusion
that estimates the axonal diameter distribution within a
nerve bundle (Assaf et al. 2008).
This is different with intracortical maturation, where mat-
uration is associated with a progressive decrease in FA dur-
ing development (Deipolyi et al. 2005), confirmed by
studies in the developing rat brain (Huang et al. 2008;
Sizonenko et al. 2007b) with sufficient resolution to identify
clearly the microstructural organization in the cortex
(Fig. 5). During rat brain development, ADC and FA changes
were detected in cortical layers between postnatal day 3
and 6. A distinct radial organization of the cortical layers
with the eigenvectors perpendicular to the pial surface was
observed at both ages. This organization was most promi-
nent in the external cortical layers 1–3 compared with the
deep layers 4–6. With cortical maturation, ADC was
reduced, whereas FA was decreased only in the deep layers
of the cortex. Histology revealed maturational differences
in the cortical architecture with increased neurodendritic
density and reduction in the radial glia scaffolding
(Sizonenko et al. 2007b). Similar developmental changes in
anisotropy within the cortex have been shown in primates
at different gestational ages. At 90 days of gestation, diffu-
sion exhibits prolate symmetry and high anisotropy, reflect-
ing apical neuronal dendrites and radial glia. Later in
gestation, with maturation of basal neuronal dendrites,
connections with thalamo-cortical afferents, branching of
interneurons and reduction of radial glia, the water diffu-
sion was modified and reflected by a reduction of diffusion
along the principal eigenvalue k1 and an increase in the
eigenvalues k2 and k3 (Kroenke et al. 2005).
The histological correlate of DTI modifications after
cerebral hypoxic–ischemic injury at 3 days of life in rat
pups (equivalent to a preterm infant born at 24–28 weeks
of gestation) showed that DTI was indeed sensitive to
microstructural changes with an alteration of the radial
organization of the cortex (Fig. 7). The decrease in ADC
and FA at 24 and 72 h after injury corresponded to neuro-
nal cell death, astrogliosis, axonal degeneration, and alter-
ation of the radial glia with early transformation into
astrocytes (Sizonenko et al. 2005, 2007b). Long-term assess-
ment with DTI showed an altered cortical structure and FA
reduction in the corpus callosum and external capsule. The
damage resulting from the hypoxic-ischemic injury corre-
lated with functional alteration of the whisker-elicited
somatosensory response (Quairiaux et al. 2010). Lipopoly-
saccharide administration in the developing animal mimics
the inflammation-induced white matter injury seen in pre-
term infants. Intracallosal injection of lipopolysaccharide in
5-day-old rat pups has shown a similar pattern on mag-
netic resonance-DTI to that seen in a hypoxic–ischemic
injury with an initial decrease of the ADC corresponding
to the peak of maximal injury. At 4 days after lipopolysac-
charide exposure, the ADC was increased together with
radial diffusivity corresponding to a decrease in cell den-
sity and an increase in callosal thickness and a diffuse
astrogliosis. The increase in ADC of the white matter at
4 days after injury reflected diffuse gliosis with increased
extracellular space rather than an arrest in development
(Lodygensky et al. 2009). Translating this knowledge to
white matter assessment in preterm infants with a higher
ADC and lower RA worsened by white matter injury
would imply a significant injury rather than a delayed
development. The acute changes seen in different animal
models of preterm brain injury reflect the initial damage
on brain structures. These initial tissue and cellular altera-
tions will influence the subsequent brain development in
the long term in balance with the intrinsic protective
response of the brain (Sizonenko et al. 2007a).
B
A
Fig. 7 Principal eigenvector plot representing a disruption in the
parietal cortex of the radially organized cortical eigenvectors 24 h
after hypoxia-ischemia. (B) Fluoro-Jade B stain showing degenerating
neurons in the same area. Courtesy of Sizonenko et al. (Sizonenko
et al., 2007b).
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al. 423

Preterm infants imaged at term even without evidence
of white matter injury were shown to have a decrease of
the RA in the internal capsule when compared with term
infants and a decrease of FA in the splenium of the corpus
callosum and coronal radiata (Rose et al. 2008). White mat-
ter injury in preterm infants was shown to have an even
stronger impact on the microstructural organization with
an arrest in the normally decreasing ADC and increasing
anisotropy of the white matter together with a decrease of
RA in the internal capsule (Huppi et al. 2001; Miller et al.
2002). Tract-based spatial statistics found significant
changes in regions within the centrum semiovale, frontal
white matter and the genu of the corpus callosum that had
a significantly lower FA in preterm infants imaged at term-
equivalent age compared with term-born controls (Anjari
et al. 2007), thus assessing alterations of brain development
in ex-preterm infants. Further experience-related changes
in diffusion characteristics have been shown in practicing
piano players (Bengtsson et al. 2005) or in working memory
training (Takeuchi et al. 2010) and provide evidence in sup-
port of the experience-based structural plasticity in the
brain. Recently, DTI performed before and after a 2-h task
was able to detect a decrease in ADC in the hippocampus
with an increase in FA in the right amygdala, bilateral
entorhinal cortex and left insula (Tavor et al. 2010). The
recent notion that DTI not only can explore microstructural
foundations of the brain, but also appears to be sensitive
to its functional adaptation opens a new domain of
research.
Functional magnetic resonance imaging andfunctional magnetic resonance imagingconnectivity
Functional MRI (fMRI) is based on the detection of regional
changes in signals that are correlated with brain functional
activity. Neuronal activation is fuelled by an increase in-loco
regional blood flow resulting in a decrease of deoxygen-
ated hemoglobin. As deoxygenated hemoglobin has para-
magnetic properties, a change in its concentration results in
a change in signal intensity that can be quantified. This
effect is known as the blood oxygenation level-dependent
contrast (Ogawa et al. 1990). The high spatial resolution
and relatively fast temporal resolution of fMRI has made it
an excellent technique to study brain function in combina-
tion with structural MRI and diffusion MRI. Functional MRI
connectivity (fcMRI) assesses neural processing (Fox et al.
2005; Schafer et al. 2009) using the spontaneous blood oxy-
genation level-dependent signal intensity time-course’s
coherent oscillations within a neuronal network.
Functional magnetic resonance imaging in infants
Previous studies have demonstrated the feasibility of fMRI
in healthy newborns and infants at different ages using dif-
ferent passive tasks (Seghier & Huppi, 2010; Seghier et al.
2006). These studies have mainly used sensory stimuli in the
visual auditory or sensorimotor domain (Born et al. 1998;
Martin et al. 1999; Yamada et al. 1997; Muramoto et al.
2002). Using passive somatosensorial stimulation, somato-
sensory areas have been identified in the precentral and
postcentral gyri but showed weak hemispheric dominance
as compared with the somatosensory system in adults (Erbe-
rich et al. 2006). Remarkably, speech perception in very
young infants showed prefrontal, temporo-parietal, precu-
neus and angular gyri activations with a significant domi-
nance of the left hemisphere as in the adult brain
(Dehaene-Lambertz et al. 2002). The follow-up of perinatal
brain lesions combining fMRI with DTI has shown remark-
able structural and functional recovery in perinatal stroke
(Seghier et al. 2004, 2005). Unresolved issues of fMRI in
newborns and infants are the equivocal hemodynamic
response influenced by sedation and other mechanisms
controlling the cerebral blood flow response to brain activ-
ity (Seghier et al. 2004; Colonnese et al. 2008).
Impact of prematurity evaluated by functional
magnetic resonance imaging connectivity
Preterm infants scanned at term were shown to have
consistent active resting-state networks during sleep in
the primary visual cortex, bilateral sensorimotor areas,
and bilateral auditory cortex, a network including the
precuneus area, lateral parietal cortex, and cerebellum as
well as an anterior network (Fransson et al. 2007). Serial
resting-state fcMRI every 4–5 weeks was performed on
preterms during their initial hospitalization and compared
with full-term infants (Smyser et al. 2010). Smyser et al.
(2010) identified similar networks and defined their
regional growth and their connections with homotopic
counterparts during development. In addition to the net-
works described by Fransson et al. (2007), they demon-
strated the presence of interhemispheric connectivity as
early as 26 weeks of gestation. The comparison with new-
born infants born at term revealed a reduction of long-
range connectivity worsened by gray and white matter
injury. This study underlines the striking changes occur-
ring during the last trimester of gestation and the
extraordinary susceptibility of white matter in children
born prematurely. More studies are needed to evaluate
the specific impact of prematurity-associated brain injuries
such as periventricular venous infarction or diffuse white
matter injury and to determine its utility in the clinical
arena. Although the acquisition itself is performed in a
very short time and holds a mine of unexploited valuable
information on brain function, it remains a very chal-
lenging imaging technique, as the quantified changes in
signal are close to the noise level and susceptible to a vari-
ety of artifacts such as motion cyclic cardiac and respiratory
artifacts.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al.424

Functional magnetic resonance imaging connectivity
in animal models
The fcMRI has been studied in rats at 4.7 T (Zhang et al.
2010) with a relatively large voxel size of 0.125 · 0.125
· 1 mm limiting its translation to younger animals with
much smaller brains without using a higher field strength
and stronger gradients. The fcMRI was established
between the prefrontal cortex, thalamus, and retrosplenial
cortex and their connectivity with very precise anatomical
regions such as the visual, auditory, motor, and somato-
sensory cortices. Interestingly, fcMRI was studied in an
adult animal model of stroke, showing that it may be con-
sidered as a new approach to quantify brain plasticity.
Indeed, the recovery of the contralateral baseline activa-
tion correlated with the recovery of the sensorimotor
function (van Meer et al. 2010). The fcMRI not only pro-
vides the opportunity to study in detail connectivity
between different parts of the brain, but also provides
information regarding plasticity after injury, thus opening
a new era of research in brain injury too often focused on
the lesion itself.
Conclusion
The use of advanced MRI techniques has revolutionized the
in-vivo visualization and comprehension of human brain
development and function. The advance in microstructural
and functional evaluation techniques such as DTI and fcMRI
will open up ways to study longitudinally important aspects
of brain plasticity during development. The implementation
of imaging on high-field magnetic resonance systems in ani-
mal models of brain development and injury further allows
the translation of the study of mechanisms of development
and disease based on experimental data to human applica-
tions and the testing of emerging hypothesis coming from
human brain magnetic resonance research in animal experi-
ments.
References
Abernethy LJ, Palaniappan M, Cooke RW (2002) Quantitative
magnetic resonance imaging of the brain in survivors of very
low birth weight. Arch Dis Child 87, 279–283.
Allin M, Matsumoto H, Santhouse AM, et al. (2001) Cognitive
and motor function and the size of the cerebellum in
adolescents born very pre-term 3. Brain 124, 60–66.
Anbeek P, Vincken KL, Groenendaal F, et al. (2008) Probabilistic
brain tissue segmentation in neonatal magnetic resonance
imaging. Pediatr Res 63, 158–163.
Anderson NG, Laurent I, Woodward LJ, et al. (2006) Detection
of impaired growth of the corpus callosum in premature
infants. Pediatrics 118, 951–960.
Anjari M, Srinivasan L, Allsop JM, et al. (2007) Diffusion tensor
imaging with tract-based spatial statistics reveals local white
matter abnormalities in preterm infants. Neuroimage 35,
1021–1027.
Ashburner J, Friston KJ (2001) Why voxel-based morphometry
should be used. Neuroimage 14, 1238–1243.
Assaf Y, Blumenfeld-Katzir T, Yovel Y, et al. (2008) AxCaliber: a
method for measuring axon diameter distribution from
diffusion MRI. Magn Reson Med 59, 1347–1354.
Barnette AR, Neil JJ, Kroenke CD, et al. (2009) Characterization
of brain development in the ferret via MRI. Pediatr Res 66,
80–84.
Battin MR, Maalouf EF, Counsell SJ, et al. (1998) Magnetic
resonance imaging of the brain in very preterm infants:
visualization of the germinal matrix, early myelination, and
cortical folding. Pediatrics 101, 957–962.
Bengtsson SL, Nagy Z, Skare S, et al. (2005) Extensive piano
practicing has regionally specific effects on white matter
development. Nat. Neurosci. 8, 1148–1150.
Born P, Leth H, Miranda MJ, et al. (1998) Visual activation in
infants and young children studied by functional magnetic
resonance imaging. Pediatr Res 44, 578–583.
Borradori Tolsa C, Zimine S, Warfield S, et al. (2004) Postnatal
growth and neurodevelopmental outcome of premature
infants born with intrauterine growth restriction. Pediatr Res
55, 411A.
Cachia A, Mangin JF, Riviere D, et al. (2003) A primal sketch of
the cortex mean curvature: a morphogenesis based approach
to study the variability of the folding patterns. IEEE Trans
Med Imaging 22, 754–765.
Colonnese MT, Phillips MA, Constantine-Paton M, et al. (2008)
Development of hemodynamic responses and functional
connectivity in rat somatosensory cortex. Nat Neurosci 11, 72–
79.
Counsell SJ, Maalouf EF, Fletcher AM, et al. (2002) MR imaging
assessment of myelination in the very preterm brain. AJNR Am
J Neuroradiol 23, 872–881.
Dehaene-Lambertz G, Dehaene S, Hertz-Pannier L (2002)
Functional neuroimaging of speech perception in infants.
Science 298, 2013–2015.
Deipolyi AR, Mukherjee P, Gill K, et al. (2005) Comparing
microstructural and macrostructural development of the
cerebral cortex in premature newborns: diffusion tensor
imaging versus cortical gyration. Neuroimage 27, 579–586.
Dubois J, Benders M, Borradori-Tolsa C, et al. (2008a) Primary
cortical folding in the human newborn: an early marker of
later functional development. Brain 131, 2028–2041.
Dubois J, Benders M, Cachia A, et al. (2008b) Mapping the early
cortical folding process in the preterm newborn brain. Cereb
Cortex 18, 1444–1454.
Dubois J, Benders M, Lazeyras F, et al. (2010) Structural
asymmetries of perisylvian regions in the preterm newborn.
Neuroimage 52, 32–42.
Dudink J, Lequin M, van PC, et al. (2007) Fractional anisotropy
in white matter tracts of very-low-birth-weight infants.
Pediatr Radiol 37, 1216–1223.
Erberich SG, Panigrahy A, Friedlich P, et al. (2006) Somatosensory
lateralization in the newborn brain. Neuroimage 29, 155–161.
Feldman R, Eidelman AI (2006) Neonatal state organization,
neuromaturation, mother-infant interaction, and cognitive
development in small-for-gestational-age premature infants.
Pediatrics 118, e869–e878.
Ferrie JC, Barantin L, Saliba E, et al. (1999) MR assessment of the
brain maturation during the perinatal period: quantitative T2
MR study in premature newborns. Magn Reson Imaging 17,
1275–1288.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al. 425

Fox MD, Snyder AZ, Vincent JL, et al. (2005) The human brain is
intrinsically organized into dynamic, anticorrelated functional
networks. Proc Natl Acad Sci USA 102, 9673–9678.
Fransson P, Skiold B, Horsch S, et al. (2007) Resting-state
networks in the infant brain. Proc Natl Acad Sci USA 104,
15531–15536.
Giorgio A, Watkins KE, Douaud G, et al. (2008) Changes in
white matter microstructure during adolescence. Neuroimage
39, 52–61.
Hrbek A, Karlberg P, Olsson T (1973) Development of visual and
somatosensory evoked responses in pre-term newborn infants.
Electroencephalogr Clin Neurophysiol 34, 225–232.
Huang H, Zhang J, Wakana S, et al. (2006) White and gray
matter development in human fetal, newborn and pediatric
brains. Neuroimage 33, 27–38.
Huang H, Yamamoto A, Hossain MA, et al. (2008) Quantitative
cortical mapping of fractional anisotropy in developing rat
brains. J Neurosci 28, 1427–1433.
Huppi PS (2002) Advances in postnatal neuroimaging: relevance
to pathogenesis and treatment of brain injury. Clin Perinatol
29, 827–856.
Huppi PS, Schuknecht B, Boesch C, et al. (1996) Structural and
neurobehavioral delay in postnatal brain development of
preterm infants. Pediatr Res 39, 895–901.
Huppi PS, Maier SE, Peled S, et al. (1998a) Microstructural
development of human newborn cerebral white matter
assessed in vivo by diffusion tensor magnetic resonance
imaging. Pediatr Res 44, 584–590.
Huppi PS, Warfield S, Kikinis R, et al. (1998b) Quantitative
magnetic resonance imaging of brain development in
premature and mature newborns. Ann Neurol 43, 224–235.
Huppi PS, Murphy B, Maier SE, et al. (2001) Microstructural
brain development after perinatal cerebral white matter
injury assessed by diffusion tensor magnetic resonance
imaging. Pediatrics 107, 455–460.
Inder TE, Happi PS, Warfield S, et al. (1999) Periventricular
white matter injury in the premature infant is associated with
a reduction in cerebral cortical gray matter volume at term.
Ann Neurol 46, 755–760.
Inder TE, Warfield SK, Wang H, et al. (2005) Abnormal cerebral
structure is present at term in premature infants. Pediatrics
115, 286–294.
Kasprian G, Brugger PC, Weber M, et al. (2008) In utero
tractography of fetal white matter development. Neuroimage
43, 213–224.
Kesler SR, Reiss AL, Vohr B, et al. (2008) Brain volume
reductions within multiple cognitive systems in male preterm
children at age twelve. J Pediatr 152, 513–520.
Kim JH, Budde MD, Liang HF, et al. (2006) Detecting axon
damage in spinal cord from a mouse model of multiple
sclerosis. Neurobiol Dis 21, 626–632.
Kostovic I, Jovanov-Milosevic N (2006) The development of
cerebral connections during the first 20-45 weeks’ gestation.
Semin Fetal Neonatal Med 11, 415–422.
Kostovic I, Molliver ME (1974) A new interpretation of the
laminar development of the cerebral cortex: synaptogenesis in
different layers of neopallium in the human fetus. American
Association of Anatomists. Eighty seventh annual session.
Anat Rec 178, 395.
Kostovic I, Rakic P (1990a) Developmental history of the
transient subplate zone in the visual and somatosensory
cortex of the macaque monkey and human brain. J Comp
Neurol 297, 441–470.
Kostovic I, Rakic P (1990b) Developmental history of the
transient subplate zone in visual and somatosensory cortex of
the macaque monkey and human brain. J Comp Neurol 297,
441–470.
Kostovic I, Vasung L (2009) Insights from in vitro fetal magnetic
resonance imaging of cerebral development. Semin Perinatol
33, 220–233.
Kostovic I, Judas M, Rados M, et al. (2002) Laminar organization
of the human fetal cerebrum revealed by histochemical
markers and magnetic resonance imaging. Cereb Cortex 12,
536–544.
Kreher BW, Mader I, Kiselev VG (2008) Gibbs tracking: a novel
approach for the reconstruction of neuronal pathways. Magn
Reson Med 60, 953–963.
Kroenke CD, Bretthorst GL, Inder TE, et al. (2005) Diffusion MR
imaging characteristics of the developing primate brain.
Neuroimage 25, 1205–1213.
Limperopoulos C, Soul JS, Gauvreau K, et al. (2005) Late
gestation cerebellar growth is rapid and impeded by
premature birth 1. Pediatrics 115, 688–695.
Lodygensky GA, Rademaker K, Zimine S, et al. (2005) Structural
and functional brain development after hydrocortisone
treatment for neonatal chronic lung disease. Pediatrics 116, 1–
7.
Lodygensky GA, Inder TE, Neil JJ (2008a) Application of
magnetic resonance imaging in animal models of perinatal
hypoxic-ischemic cerebral injury. Int J Dev Neurosci 26, 13–
25.
Lodygensky GA, Seghier ML, Warfield SK, et al. (2008b)
Intrauterine growth restriction affects the preterm infant’s
hippocampus. Pediatr Res 63, 438–443.
Lodygensky G, West T, Stump M, et al. (2009) In vivo MRI
analysis of an inflammatory injury in the developing brain.
Brain Behav Immun 24, 759–767.
Martin E, Joeri P, Loenneker T, et al. (1999) Visual processing in
infants and children studied using functional MRI. Pediatr Res
46, 135–140.
McKinstry RC, Mathur A, Miller JP, et al. (2002a) Radial
organization of developing human cerebral cortex revealed
by non-invasive water diffusion anisotropy MRI. Cereb Cortex
12, 1237–1243.
McKinstry RC, Miller JH, Snyder AZ, et al. (2002b) A prospective,
longitudinal diffusion tensor imaging study of brain injury in
newborns. Neurology 59, 824–833.
van Meer MP, van der Marel K, Wang K, et al. (2010) Recovery
of sensorimotor function after experimental stroke correlates
with restoration of resting-state interhemispheric functional
connectivity. J Neurosci 30, 3964–3972.
Ment LR, Kesler S, Vohr B, et al. (2009) Longitudinal brain
volume changes in preterm and term control subjects
during late childhood and adolescence. Pediatrics 123, 503–
511.
Mewes AU, Huppi PS, Als H, et al. (2006) Regional brain
development in serial magnetic resonance imaging of low-risk
preterm infants. Pediatrics 118, 23–33.
Miller SP, Vigneron DB, Henry RG, et al. (2002) Serial
quantitative diffusion tensor MRI of the premature brain:
development in newborns with and without injury. J Magn
Reson Imaging 16, 621–632.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al.426

Mori S, Crain BJ, Chacko VP, et al. (1999) Three-dimensional
tracking of axonal projections in the brain by magnetic
resonance imaging. Ann Neurol 45, 265–269.
Mukherjee P, Miller JH, Shimony JS, et al. (2002) Diffusion-
tensor MR imaging of gray and white matter development
during normal human brain maturation. AJNR Am J
Neuroradiol 23, 1445–1456.
Muramoto S, Yamada H, Sadato N, et al. (2002) Age dependent
change in metabolic response to photic stimulation of the
primary visual cortex in infants: functional magnetic
resonance imaging study. J Comput Assist Tomogr 26, 894–
901.
Murphy BP, Inder TE, Happi PS, et al. (2001) Impaired cerebral
cortical gray matter growth following treatment with
dexamethasone for neonatal chronic lung disease. Pediatrics
107, 217–221.
Narberhaus A, Segarra D, Caldu X, et al. (2007) Gestational
age at preterm birth in relation to corpus callosum and
general cognitive outcome in adolescents. J Child Neurol 22,
761–765.
Neil JJ, Shiran SI, McKinstry RC, et al. (1998) Normal brain in
human newborns: apparent diffusion coefficient and diffusion
anisotropy measured using diffusion tensor imaging.
Radiology 209, 57–66.
Nosarti C, Giouroukou E, Healy E, et al. (2008) Grey and white
matter distribution in very preterm adolescents mediates
neurodevelopmental outcome. Brain 131, 205–217.
Ogawa SL, Lee TM, Kay AR, et al. (1990) Brain magnetic
resonance imaging with contrast dependent on blood
oxygenation. Proc Natl Acad Sci 87, 9868–9872.
Parker GJ, Haroon HA, Wheeler-Kingshott CA (2003) A
framework for a streamline-based probabilistic index of
connectivity (PICo) using a structural interpretation of MRI
diffusion measurements. J Magn Reson Imaging 18, 242–254.
Partridge SC, Mukherjee P, Henry RG, et al. (2004) Diffusion
tensor imaging: serial quantitation of white matter tract
maturity in premature newborns. Neuroimage 22, 1302–
1314.
Prayer D, Kasprian G, Krampl E, et al. (2006) MRI of normal
fetal brain development. Eur J Radiol 57, 199–216.
Quairiaux C, Sizonenko SV, Megevand P, et al. (2010)
Functional deficit and recovery of developing sensorimotor
networks following neonatal hypoxic-ischemic injury in the
rat. Cereb Cortex, doi:10.1093/cercor/bhp281.
Rados M, Judas M, Kostovic I (2006) In vitro MRI of brain
development. Eur J Radiol 57, 187–198.
Ramenghi LA, Fumagalli M, Righini A, et al. (2007) Magnetic
resonance imaging assessment of brain maturation in preterm
neonates with punctate white matter lesions. Neuroradiology
49, 161–167.
Rose SE, Hatzigeorgiou X, Strudwick MW, et al. (2008)
Altered white matter diffusion anisotropy in normal and
preterm infants at term-equivalent age. Magn Reson Med
60, 761–767.
Rutherford M, Counsell S, Allsop J, et al. (2004) Diffusion-
weighted magnetic resonance imaging in term perinatal brain
injury: a comparison with site of lesion and time from birth.
Pediatrics 114, 1004–1014.
Schafer RJ, Lacadie C, Vohr B, et al. (2009) Alterations in
functional connectivity for language in prematurely born
adolescents. Brain 132, 661–670.
Seghier ML, Huppi PS (2010) The role of functional magnetic
resonance imaging in the study of brain development,
injury, and recovery in the newborn. Semin Perinatol 34,
79–86.
Seghier ML, Lazeyras F, Zimine S, et al. (2004) Combination of
event-related fMRI and diffusion tensor imaging in an infant
with perinatal stroke. Neuroimage 21, 463–472.
Seghier ML, Lazeyras F, Zimine S, et al. (2005) Visual recovery
after perinatal stroke evidenced by functional and diffusion
MRI: case report. BMC Neurol 5, 17.
Seghier ML, Lazeyras F, Huppi PS (2006) Functional MRI of the
newborn. Semin Fetal Neonatal Med 11, 479–488.
Sizonenko SV, Kiss JZ, Inder T, et al. (2005) Distinctive
neuropathologic alterations in the deep layers of the parietal
cortex after moderate ischemic-hypoxic injury in the P3
immature rat brain. Pediatr Res 57, 865–872.
Sizonenko SV, Bednarek N, Gressens P (2007a) Growth factors
and plasticity. Semin Fetal Neonatal Med 12, 241–249.
Sizonenko SV, Camm EJ, Garbow JR, et al. (2007b)
Developmental changes and injury induced disruption of the
radial organization of the cortex in the immature rat brain
revealed by in vivo diffusion tensor MRI. Cereb Cortex 17,
2609–2617.
Smith SM, Jenkinson M, Johansen-Berg H, et al. (2006) Tract-
based spatial statistics: voxelwise analysis of multi-subject
diffusion data. Neuroimage 31, 1487–1505.
Smyser CD, Inder TE, Shimony JS, et al. (2010) Longitudinal
analysis of neural network development in preterm infants.
Cereb Cortex, doi:10.1093/cercor/bhq035.
Song SK, Yoshino J, Le TQ, et al. (2005) Demyelination increases
radial diffusivity in corpus callosum of mouse brain.
Neuroimage 26, 132–140.
Srinivasan L, Allsop J, Counsell SJ, et al. (2006) Smaller cerebellar
volumes in very preterm infants at term-equivalent age are
associated with the presence of supratentorial lesions. AJNR
Am J Neuroradiol 27, 573–579.
Srinivasan L, Dutta R, Counsell SJ, et al. (2007) Quantification of
deep gray matter in preterm infants at term-equivalent age
using manual volumetry of 3-tesla magnetic resonance
images. Pediatrics 119, 759–765.
Takeuchi H, Sekiguchi A, Taki Y, et al. (2010) Training of
working memory impacts structural connectivity. J Neurosci
30, 3297–3303.
Tavor I, Sagi Y, Hostetter S, et al. (2010) Diffusion MRI of Short-
Term Spacial Memory Related Brain Plasticity. Proc Intl Soc
Mag Reson Med 18, 9.
Toft PB, Leth H, Ring PB, et al. (1995) Volumetric analysis of the
normal infant brain and in intrauterine growth retardation.
Early Hum Dev 43, 15–29.
Volpe JJ (2009) Brain injury in premature infants: a complex
amalgam of destructive and developmental disturbances.
Lancet Neurol 8, 110–124.
Weisenfeld NI, Warfield SK (2009) Automatic segmentation of
newborn brain MRI. Neuroimage 47, 564–572.
West T, Stump M, Lodygensky G, et al. (2009) Lack of X-linked
inhibitor of apoptosis protein leads to increased apoptosis
and tissue loss following neonatal brain injury. ASN NEURO 1,
doi:10.1042/AN20090005.
Wimberger DM, Roberts TP, Barkovich AJ, et al. (1995)
Identification of ‘‘premyelination’’ by diffusion-weighted MRI.
J Comptr Assisted Tomography 19, 28–33.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al. 427

Woodward LJ, Anderson PJ, Austin NC, et al. (2006) Neonatal
MRI to predict neurodevelopmental outcomes in preterm
infants. N Engl J Med 355, 685–694.
Yamada H, Sadato N, Konishi Y, et al. (1997) A rapid brain
metabolic change in infants detected by fMRI. Neuroreport 8,
3775–3778.
Zhang N, Rane P, Huang W, et al. (2010) Mapping resting-state
brain networks in conscious animals. J Neurosci Methods 189,
186–196.
ªª 2010 The AuthorsJournal of Anatomy ªª 2010 Anatomical Society of Great Britain and Ireland
Neuroimaging in human newborns & animal models, G. A. Lodygensky et al.428




















