
Cortical auditory processing in preterm newborns: An ERP study
10
Cortical auditory processing in preterm newborns: An ERP study Patrizia Silvia Bisiacchi a,1 , Giovanni Mento a, *, Agnese Suppiej b,2 a Department of General Psychology, University of Padova, Padova, Italy b Child Neurology and Clinical Neurophysiology, Department of Pediatrics, University of Padova, 35128 Padova, Italy 1. Introduction In the last few decades the survival of preterm infants born under 35 gestational weeks (GW) has significantly improved, especially in socio-economically developed countries (Drillien et al., 1980; Robertson et al., 1994; Wilson-Costello et al., 2005). This phenomenon can be considered as the natural consequence of the achievement of medical knowledge about intensive care procedures and introduction of drugs such as antenatal corticos- teroids and surfactants (Rijken et al., 2003), which, in the early 1990s, led to a drastic reduction in the percentage of preterm newborn deaths (Hack and Fanaroff, 2000). As a consequence, the rate of neonatal morbidity, especially for extremely premature newborns (<30 GW), has greatly increased, resulting in several pathologies, increased length of hospitalization, and costs of care (Marlow et al., 2005; Pietz et al., 2004; Wen et al., 2004). These issues are even more significant given that several long- term follow-up studies have demonstrated that survivors without major neurological signs and who are apparently normal during infancy and preschool years show significant neuropsychological and behavioral deficits at school age (Leonard et al., 1997; Pietz et al., 2004; Volpe, 1991). Thus in recent years, the focus of research has changed towards earlier prediction of neurodevelopmental outcome. In 1991, a prognostic tool was proposed, the Neurobio- logical Risk Score (NBRS; Brazy et al., 1991), in order to classify those infants of less than 1500 g as low or high neurological risk. This scale, which takes into account several clinical indices, including some that are not strictly neurological, may be considered to be a good tool for early prognosis of major neurological handicaps, such as cerebral palsy or sensory deficits; however, it is not equally good at predicting minor ones, such as ‘‘soft signs’’, including cognitive, neuropsychological and learning difficulties in healthy or low-risk preterm infants. It has been assumed that one possible cause of these deficits may lie in non- specified neurobiological factors disrupting cortical pathways, especially for newborns of extremely low gestational age (ELGA; Therien et al., 2004). However, despite a number of recent studies, it is not yet understood how the altered neural pattern at birth can result in cognitive deficits in childhood (Inder et al., 2005). In addition to the influence of intra-uterine growth as the main physiological environment, another factor that may play an important role in neonatal cortical pathway development is the sensorial stimulation provided by the extra-uterine environment. Indeed, full-term newborns are able to learn speech sounds while they are sleeping and display a modulation of cortical activity in discriminating sounds (Cheour et al., 2002c), and newborns with more experience of hearing their mother’s voice are more able to recognize it compared to those without this kind of experience, demonstrating the effects of auditory cortical maturation on recognition memory (deRegnier et al., 2002). Nevertheless, there is very little known about the effect of precocious sensory exposure in prematurely born infants, who are known to undergo physiological and behavioral adaptation to the extra-uterine Biological Psychology 82 (2009) 176–185 ARTICLE INFO Article history: Received 9 November 2007 Accepted 1 July 2009 Available online 22 July 2009 Keywords: Preterm newborns Auditory event-related potentials Gestational age Cortical functioning Mismatch response ABSTRACT The aim of this study was to investigate the effects of gestational age on the auditory event-related potentials (AERPs) in preterm newborns of extremely low (23–29 weeks, N = 20) and low gestational age (30–34 weeks, N = 19). All subjects were tested at the same post-conceptional age of 35 weeks. An auditory oddball paradigm was used with frequently occurring ‘‘standard’’ tones at 1000 Hz and rarely occurring ‘‘deviant’’ tones at 2000 Hz. Newborns of less than 30 gestational weeks showed smaller AERPs responses (P1i, N2i and mismatch response) than the older ones. Our results suggest the role of gestational age as the main factor explaining cortical functioning at this early age. ß 2009 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +39 0 498276616. E-mail addresses: [email protected] (P.S. Bisiacchi), [email protected] (G. Mento), [email protected] (A. Suppiej). 1 Tel.: +39 0 498276587/6609. 2 Tel.: +39 0 498213505. Contents lists available at ScienceDirect Biological Psychology journal homepage: www.elsevier.com/locate/biopsycho 0301-0511/$ – see front matter ß 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.biopsycho.2009.07.005
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Cortical auditory processing in preterm newborns: An ERP study

Biological Psychology 82 (2009) 176–185
Cortical auditory processing in preterm newborns: An ERP study
Patrizia Silvia Bisiacchi a,1, Giovanni Mento a,*, Agnese Suppiej b,2
a Department of General Psychology, University of Padova, Padova, Italyb Child Neurology and Clinical Neurophysiology, Department of Pediatrics, University of Padova, 35128 Padova, Italy
A R T I C L E I N F O
Article history:
Received 9 November 2007
Accepted 1 July 2009
Available online 22 July 2009
Keywords:
Preterm newborns
Auditory event-related potentials
Gestational age
Cortical functioning
Mismatch response
A B S T R A C T
The aim of this study was to investigate the effects of gestational age on the auditory event-related
potentials (AERPs) in preterm newborns of extremely low (23–29 weeks, N = 20) and low gestational age
(30–34 weeks, N = 19). All subjects were tested at the same post-conceptional age of 35 weeks. An
auditory oddball paradigm was used with frequently occurring ‘‘standard’’ tones at 1000 Hz and rarely
occurring ‘‘deviant’’ tones at 2000 Hz. Newborns of less than 30 gestational weeks showed smaller AERPs
responses (P1i, N2i and mismatch response) than the older ones. Our results suggest the role of
gestational age as the main factor explaining cortical functioning at this early age.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Biological Psychology
journa l homepage: www.e lsev ier .com/ locate /b iopsycho
1. Introduction
In the last few decades the survival of preterm infants bornunder 35 gestational weeks (GW) has significantly improved,especially in socio-economically developed countries (Drillienet al., 1980; Robertson et al., 1994; Wilson-Costello et al., 2005).This phenomenon can be considered as the natural consequence ofthe achievement of medical knowledge about intensive careprocedures and introduction of drugs such as antenatal corticos-teroids and surfactants (Rijken et al., 2003), which, in the early1990s, led to a drastic reduction in the percentage of pretermnewborn deaths (Hack and Fanaroff, 2000). As a consequence, therate of neonatal morbidity, especially for extremely prematurenewborns (<30 GW), has greatly increased, resulting in severalpathologies, increased length of hospitalization, and costs of care(Marlow et al., 2005; Pietz et al., 2004; Wen et al., 2004).
These issues are even more significant given that several long-term follow-up studies have demonstrated that survivors withoutmajor neurological signs and who are apparently normal duringinfancy and preschool years show significant neuropsychologicaland behavioral deficits at school age (Leonard et al., 1997; Pietzet al., 2004; Volpe, 1991). Thus in recent years, the focus of researchhas changed towards earlier prediction of neurodevelopmental
* Corresponding author. Tel.: +39 0 498276616.
E-mail addresses: [email protected] (P.S. Bisiacchi),
[email protected] (G. Mento), [email protected] (A. Suppiej).1 Tel.: +39 0 498276587/6609.2 Tel.: +39 0 498213505.
0301-0511/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.biopsycho.2009.07.005
outcome. In 1991, a prognostic tool was proposed, the Neurobio-logical Risk Score (NBRS; Brazy et al., 1991), in order to classifythose infants of less than 1500 g as low or high neurological risk.This scale, which takes into account several clinical indices,including some that are not strictly neurological, may beconsidered to be a good tool for early prognosis of majorneurological handicaps, such as cerebral palsy or sensory deficits;however, it is not equally good at predicting minor ones, such as‘‘soft signs’’, including cognitive, neuropsychological and learningdifficulties in healthy or low-risk preterm infants. It has beenassumed that one possible cause of these deficits may lie in non-specified neurobiological factors disrupting cortical pathways,especially for newborns of extremely low gestational age (ELGA;Therien et al., 2004). However, despite a number of recent studies,it is not yet understood how the altered neural pattern at birth canresult in cognitive deficits in childhood (Inder et al., 2005).
In addition to the influence of intra-uterine growth as the mainphysiological environment, another factor that may play animportant role in neonatal cortical pathway development is thesensorial stimulation provided by the extra-uterine environment.Indeed, full-term newborns are able to learn speech sounds whilethey are sleeping and display a modulation of cortical activity indiscriminating sounds (Cheour et al., 2002c), and newborns withmore experience of hearing their mother’s voice are more able torecognize it compared to those without this kind of experience,demonstrating the effects of auditory cortical maturation onrecognition memory (deRegnier et al., 2002). Nevertheless, there isvery little known about the effect of precocious sensory exposurein prematurely born infants, who are known to undergophysiological and behavioral adaptation to the extra-uterine

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185 177
environment very early in their development. In the NeonatalIntensive Care Unit (NICU), newborns of extremely low gestationalage (ELGA, <30 weeks) are exposed to the extra-uterineenvironment for longer than older newborns (>30 weeks) whencompared at the same at term-corrected age. Therefore, theyreceive greater sensory exposure, in terms of both visual andauditory system stimulation, being exposed to more light andfaces and also to noise and voices in an environmental settingmore natural than the physiological low-pass filter of the womb(Vouloumanos and Werker, 2007). Furthermore, they alsoexperience affective and social interaction provided by themother and nurses during feeding and handling. To date, althoughsome authors consider these kinds of experiences as an effectivesource of compensation for the cognitive effects of prematurity(Als et al., 1994; Ostfeld et al., 2000), whether this early experiencecan deprive preterm newborns of necessary stimulation, exposesnewborns to continuous over-stimulation, or lacks a develop-mentally appropriate pattern of stimulation to compensate for theeffects of a premature birth is still a controversial issue (Huppiet al., 1996). Hence, the secondary influence of extra-uterinedevelopment may be an important issue to take into considerationin order to understand how the immature brain of prematurenewborns without any major neurological signs can be modifiedin its functioning when prematurely exposed to the sensorialworld.
These questions have been addressed by many authors, whohave compared preterm newborns at term-corrected-age to full-term newborns using several neurobehavioral measures, andfound significant alterations in motor behavior and highercortically integrated functions in preterm newborns (Aylwardet al., 1984; Brown et al., 2006; Majnemer et al., 1993; Piper et al.,1989). More recently, Dubois et al. (2007, 2009) used a magneticresonance imaging (MRI) methodology enabling the ‘‘in vivo’’cortical organization of the premature brain to be quantified(Huppi et al., 1996; Huppi and Inder, 2001), and investigated theemergence of the cortical folding over a period critical for humanbrain development (26–36 weeks of gestational age). They claimedthat prematurity per se might influence cortical maturation, beingresponsible for the delay observed in appearance of sulci incomparison with post-mortem and fetal studies, as both corticalvolume and surface area of extremely preterm infants imaged atterm-equivalent-age are reduced and less complex than in normalinfants, and this impairment seems to increase with decreasinggestational age (GA) at birth (Ajayi-Obe et al., 2000; Kapellou et al.,2006; Inder et al., 2005). Furthermore, Dubois et al. (2007) alsofound a strong correlation between GA and quantification of thedeveloping cortical and white matter zone volumes, and of theinner cortical surface, whereas Huppi et al. (1996), using bothstructural (MRI) and biochemical data, found significant differ-ences in gray-matter development, stages of myelination, bio-chemical profile, and behavioral testing between preterms andfull-term controls. Nevertheless, other than the neuro-anatomicaland behavioral differences found, no consistent functional datademonstrating a difference in the cortical functioning betweenfull-term and at term-corrected newborns have been reported. Inthis regard, Nunes et al. (1997) analyzed the relationship betweennormal neonatal electroencephalographic (EEG) features andgestational and conceptional age, and evaluated the normalaspects of EEG maturation in preterm compared to full-terminfants, showing that the number of delta brushes and the durationof the interburst interval decrease as gestational and conceptionalage increase. They also found that the percentage of interhemi-spheric synchrony increases with gestational and conceptionalage, suggesting that the extra-uterine life of preterm infants doesnot appear to accelerate EEG maturation. However, other studiesattempting to correlate spontaneous EEG activity to gestational
age have reported controversial findings (Biagioni et al., 2007; Eyreet al., 1988; Ferrari et al., 1992; Nolte and Haas, 1978).
A more direct measure of cortical functioning, both in normaland in pathological neonatal populations, is the measure ofauditory event-related potentials (AERPs). Owing to its hightemporal resolution this method is a powerful tool to investigatethe timing of cognitive processes non-invasively and passively and,for this reason, has been helpful in describing the neurobiologicalunderpinnings of cognitive processing in children and newborns(Fellman and Huotilainen, 2006). The main ERP paradigm used tostudy cognitive functioning in newborns is the auditory ‘‘oddball’’paradigm, an experimental procedure that consists of occasionallyreplacing repetitive ‘‘standard’’ stimuli by a physically ‘‘deviant’’stimulus. The deviant-change discrimination elicits the mismatchnegativity (MMN; Alho et al., 1990; Cheour et al., 1998; Fellmanand Huotilainen, 2006; Naatanen et al., 1978), an automatic andpre-attentive cortical response that reflects auditory discrimina-tion, sensory memory, and automatic attention (Naatanen, 1990,1992), and as such is a reliable measure of cognitive functioning inall age groups (Fellman and Huotilainen, 2006).
From a maturational point of view, the ability to discriminatesounds develops early, according to several reports that providedboth electrophysiological (Alho et al., 1990; Cheour et al., 2002a,2002b; Cheour-Luhtanen et al., 1996; Kushnerenko et al., 2001,2002; Leppanen et al., 1997, 2004; Novitski et al., 2007; Suppiejand Bisiacchi, 1992) and, more recently, electromagnetic (Draga-nova et al., 2007) evidence of auditory-change discrimination innewborns as young as 27 gestational weeks (Holst et al., 2005).However, some authors failed to find an MMN response in all thenewborns tested (Alho et al., 1990; Kurtzberg et al., 1995;Ceponiene et al., 2002; Cheour et al., 1998), whereas in otherstudies MMN was found to be positive in polarity (Dehaene-Lambertz and Dehaene, 1994; Friederici et al., 2002; McIsaac andPolich, 1992; Ruusuvirta et al., 2003). This discrepancy is probablyrelated to the different methods and stimuli used by the authors,who employed speech, vowel, or simple tone stimuli (for a review,see Cheour, 2007). Another underlying cause is the high individualvariability of AERPs in a developmental population, which,especially in preterm newborns, may be related to maturationalissues. In this regard, Kurtzberg et al. (1984) studied thematurational effect on neonatal sound processing in preterminfants, and described five maturational states for the auditory-evoked potentials (AEPs), starting from preterm to term stage,featuring negative responses in medial and lateral scalp sites in lessmature infants and positive responses in medial and lateral scalpsites in more mature infants. They also found that full-term controlnewborns show significantly more mature AEPs to consonant-vowel syllables compared to very low birth weight (meangestational age 31.2 weeks) tested at 40 post-conceptional weeks.However, the stimuli employed by Kurtzberg et al. (1984), in boththe tone (800 Hz tone bursts) and consonant-vowel (‘‘da’’ and ‘‘ta’’)condition, had an equal probability of presentation of 50%, withoutthe deviant condition (that in the oddball paradigm is usually 10–20%). Therefore, the cortical responses here can be regarded asreflecting the primary auditory system processing of sound per seand are not a direct measure of change-detection. More recently,Leppanen et al. (2004), using an oddball paradigm, found asignificant correlation between maturational indexes (gestationalage, heart period, and vagal tone) and difference wave peakamplitude, claiming that the more mature the newborns, the morepositive the difference wave between standard and deviantstimuli. However, they considered only full-term newbornsranging from 37 to 42 gestational weeks.
To date, it is still a controversial issue whether a MMN can beelicited in all newborns, especially in those born prematurely.Furthermore, authors who recorded AERPs in these infants

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185178
evaluated the development of cortical responses only as related togestational development, and not in terms of the influence of anearly extra-uterine exposure. As a consequence, they do notprovide information about the cortical responses in extremely lowgestational age newborns (<30 weeks GA) compared with lesspremature ones (>30 weeks), if recorded after a varying period ofextra-uterine exposure.
The aim of this study was to investigate the effects ofgestational age on the AERPs in preterm newborns of low andextremely low gestational age.
According to one line of reasoning, longer extra-uterine sensoryexposure may exercise a compensatory effect in terms of corticaldevelopment and organization. In this case we would expect nodifferences in AERP amplitudes between low and extremely lowgestational age groups.
Alternatively, given that the volumes of developing cortex andwhite matter zone, the inner cortical surface, and the overallsulcation surface increase dramatically with GA after 30 weeks(Dubois et al., 2007), a significant difference would be expected incortical functioning between newborns of different gestationalages, with longer term newborns showing greater AERP measures.
2. Methods
2.1. Participants
Thirty-nine preterm newborns (aged 23–34 gestational weeks; M = 29;
S.D. = 2.9) were recruited from the NICU of the Children’s Hospital of the University
of Padua. The study was approved by the Hospital’s Ethics Committee. The
participants’ families gave informed consent according to the local ethical
committee and in agreement with the Declaration of Helsinki. Inclusion criteria
were acceptance of the study protocol by the parents and birth before 35 gestational
weeks. Exclusion criteria were craniofacial malformations, genetic syndromes, and
major neurological diseases such as clinical evidence of neonatal encephalopathy,
ultrasound evidence of severe intra-ventricular hemorrhage (III–IV grade by Volpe,
2000), abnormal brainstem auditory-evoked potentials (BAEPs), and administration
of drugs with sedative effects on the central nervous system to the newborn. All
newborns were tested in the same room and exposed to the same environmental
stimulation, including light and noise exposure, during their hospitalization period.
Moreover, they had daily parental care visits including handling and bottle-feeding
by their mother.
The maturational factors considered were gestational age (GA), length, weight,
and cranial circumference, all measured at birth. In order to evaluate the effect of
gestational age on AERPs, we divided the participants into ‘‘extremely low
gestational age’’ (ELGA, 23–29 weeks) and ‘‘low gestational age’’ (LGA, 30–34
weeks) groups. All participants were tested at the same PCA of 35 weeks in order to
have an internal control. This age was chosen as it is usually the mean age at which
newborns are discharged from the NICU. It was computed as the sum of gestational
age at birth and the period of extra-uterine life elapsed from birth to AERP
recording.
2.2. Sleep state
Newborns alternate between two sleep states, called active (AS) and quiet (QS)
sleep, that can easily be detected by behavioral observation of the infant under 36
gestational weeks (Mercuri et al., 1995) as well as with the electroencephalographic
staging as early as 27 weeks (Curzi-Dascalova et al., 1988). Since we found that the
sleep state affects the AERP morphology (Suppiej et al., under revision), with larger
and more reliable responses in AS than in QS, the epochs in which the newborns
were in QS or in a state of drowsiness were excluded from AERP averaging.
According to the criteria proposed by Anders et al. (1971) and adopted by Shepherd
et al. (1999) for behavioral scoring during evoked potential testing, we
characterized AS by closed eyes, irregular respiration, rapid eye movements, and
occasional body movements and, according to Ceponiene et al. (2002), by a mixed
EEG pattern (high voltage in u–d frequency ranges) or a low-voltage irregular (u–branges) continuous pattern. Quiet sleep (QS) was identified on the basis of high
voltage slow or trace-alternant patterns (Stockard-Pope et al., 1992).
2.3. Stimuli and procedure
Recordings were performed when newborns were clinically stable, before
discharge from the NICU, while lying in a comfortable crib in a dimly light,
electrically shielded, and sound attenuated room and between two morning feeds.
No sedation was used. Oddball stimuli consisted of a random series of deviant tones
at 2000 Hz (occurring with a probability 10%) embedded among standard tones at
1000 Hz (probability 90%) delivered binaurally via headphones in an oddball
sequence. We chose these frequency deviants according to Morr et al. (2002), as
they have been demonstrated to elicit extensive responses in newborns. The tone
duration was 100 ms (5 ms rise and 5 ms fall time), the intensity 75 dB nHL. The
inter-stimulus interval (ISI) was randomly varied between 1250 and 1850 ms
(offset-to-onset). The experimental design consisted of three blocks of 300 standard
and 30 deviant stimuli for each infant, for a total of 990 stimuli. The relatively low
number of deviant stimuli presented was mainly due to the necessity of making the
experimental condition as short as possible and to avoid habituation effects which
are strong in newborns and infants (Dehaene-Lambertz, 2000).
2.4. EEG recording
The electroencephalogram (EEG) was recorded at midline electrode sites (Fz, Cz,
and Pz) and referenced to the bilateral linked mastoids, in accordance with the 10–
20 international system (Jasper, 1958), using Ag/AgCl electrodes. Only central sites
were used in order to have as small a number of electrodes to place on the scalp as
possible, given the clinical condition of the prematurely born participants. All
electrode impedances were less than 10 kV and balanced. The ground electrode
was placed on the left side of the forehead (Fp1). Vertical eye movements were
monitored with a bipolar electro-oculogram (EOG) above the canthus of the left and
below the canthus of the right eye. The entire experiment lasted less than 30 min,
including the time taken to place the electrodes. Data were recorded with an
EBNeuro system (EBNeuro/Florence, Italy) and amplified and digitized with a
sampling frequency of 512 Hz. The band-pass filtering was .1–100 Hz.
2.5. Data analysis
EEG and EOG were filtered offline (.5 Hz high pass and 15 Hz low pass). Artefact
rejection for the EEG and EOG was performed with 100 mV as the delta criterion.
Epochs were 1000 ms in duration for both standard and deviant stimulus
conditions, including a 100 ms pre-stimulus baseline interval. AERPs were averaged
separately for standard and deviant tones for each subject. P1i and N2i responses
(where ‘‘i’’ is for ‘‘infant’’ to distinguish them from the adult-like obligatory AEPs)
were measured in both standard and deviant-related waves and defined as the
maxima peaks in the latency range between 100–200 and 150–250 ms,
respectively. These temporal windows were chosen from the Fz electrode site in
the grand average waveform. Difference waves were obtained by subtracting AERPs
elicited by standard trials from those elicited by deviant trials. The main
discriminative component was identified as the main response in the latency
range between 150 and 400 ms in the difference wave, and was generically named
the mismatch response (MMR), regardless of its polarity. Mean AERP amplitudes
were calculated from the data points of �30 ms centered on individual response
peaks for each subject and condition.
2.6. Statistical analysis
In order to verify that no differences were present in PCA between the two
groups, we compared their mean corrected age using a T-test. The effect of
gestational age on AERPs was evaluated using a 2 � 2 mixed repeated measures
analysis of variance (ANOVA) for P1i and N2i responses, with gestational age
(‘‘ELGA’’ versus ‘‘LGA’’) and midline electrode site (Fz and Cz) as factors. The
measurements from the Pz electrode are not reported here because they were found
to be less stable and reliable. For MMR we performed a 2 � 2 mixed repeated
ANOVA, with gestational age (‘‘ELGA’’ versus ‘‘LGA’’) and midline electrode site (Fz
and Cz) as factors. Greenhouse–Geisser adjustment was performed when applicable
(corrected p-values are reported). To confirm the hypothesis of a linear relationship
between maturational indexes (GA, length, weight, and cranial circumference) and
MMR, both a Pearson correlation and a stepwise linear regression using MMR mean
amplitudes separately for Fz and Cz as dependent variables, and sex and
maturational indices (gestational age, cranial circumference, weight and length)
as independent variables, were carried out. The significance was set at p < .05. All
analyses were performed using the Statistical Package for the Social Sciences
(SPSS1 14.0).
3. Results
Table 1 shows the characteristics of 39 newborns (21 males and18 female) in whom AERPs were measured.
3.1. Waveform description
Averaged AERPs in response to standard and deviant stimuli areshown separately in Fig. 1 for the two groups. Fig. 2 displays thegroup comparison of the difference wave (deviant minus standardAERPs). The number of accepted EEG trials after artefacts rejectionfor deviant and standard stimuli was 45 � 15 (range 30–70) and448 � 138 (range 300–600), respectively.
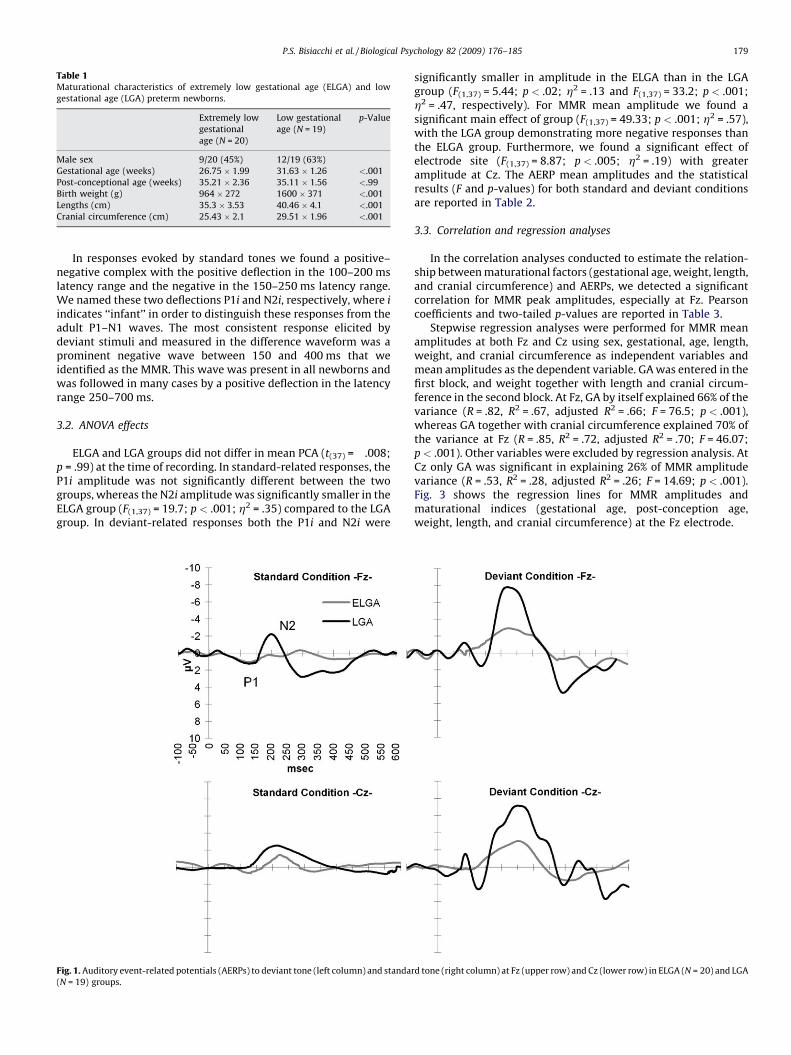
Table 1Maturational characteristics of extremely low gestational age (ELGA) and low
gestational age (LGA) preterm newborns.
Extremely low
gestational
age (N = 20)
Low gestational
age (N = 19)
p-Value
Male sex 9/20 (45%) 12/19 (63%)
Gestational age (weeks) 26.75 � 1.99 31.63 � 1.26 <.001
Post-conceptional age (weeks) 35.21 � 2.36 35.11 � 1.56 <.99
Birth weight (g) 964 � 272 1600 � 371 <.001
Lengths (cm) 35.3 � 3.53 40.46 � 4.1 <.001
Cranial circumference (cm) 25.43 � 2.1 29.51 � 1.96 <.001
P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185 179
In responses evoked by standard tones we found a positive–negative complex with the positive deflection in the 100–200 mslatency range and the negative in the 150–250 ms latency range.We named these two deflections P1i and N2i, respectively, where i
indicates ‘‘infant’’ in order to distinguish these responses from theadult P1–N1 waves. The most consistent response elicited bydeviant stimuli and measured in the difference waveform was aprominent negative wave between 150 and 400 ms that weidentified as the MMR. This wave was present in all newborns andwas followed in many cases by a positive deflection in the latencyrange 250–700 ms.
3.2. ANOVA effects
ELGA and LGA groups did not differ in mean PCA (t(37) = �.008;p = .99) at the time of recording. In standard-related responses, theP1i amplitude was not significantly different between the twogroups, whereas the N2i amplitude was significantly smaller in theELGA group (F(1,37) = 19.7; p < .001; h2 = .35) compared to the LGAgroup. In deviant-related responses both the P1i and N2i were
Fig. 1. Auditory event-related potentials (AERPs) to deviant tone (left column) and standa
(N = 19) groups.
significantly smaller in amplitude in the ELGA than in the LGAgroup (F(1,37) = 5.44; p < .02; h2 = .13 and F(1,37) = 33.2; p < .001;h2 = .47, respectively). For MMR mean amplitude we found asignificant main effect of group (F(1,37) = 49.33; p < .001; h2 = .57),with the LGA group demonstrating more negative responses thanthe ELGA group. Furthermore, we found a significant effect ofelectrode site (F(1,37) = 8.87; p < .005; h2 = .19) with greateramplitude at Cz. The AERP mean amplitudes and the statisticalresults (F and p-values) for both standard and deviant conditionsare reported in Table 2.
3.3. Correlation and regression analyses
In the correlation analyses conducted to estimate the relation-ship between maturational factors (gestational age, weight, length,and cranial circumference) and AERPs, we detected a significantcorrelation for MMR peak amplitudes, especially at Fz. Pearsoncoefficients and two-tailed p-values are reported in Table 3.
Stepwise regression analyses were performed for MMR meanamplitudes at both Fz and Cz using sex, gestational, age, length,weight, and cranial circumference as independent variables andmean amplitudes as the dependent variable. GA was entered in thefirst block, and weight together with length and cranial circum-ference in the second block. At Fz, GA by itself explained 66% of thevariance (R = .82, R2 = .67, adjusted R2 = .66; F = 76.5; p < .001),whereas GA together with cranial circumference explained 70% ofthe variance at Fz (R = .85, R2 = .72, adjusted R2 = .70; F = 46.07;p < .001). Other variables were excluded by regression analysis. AtCz only GA was significant in explaining 26% of MMR amplitudevariance (R = .53, R2 = .28, adjusted R2 = .26; F = 14.69; p < .001).Fig. 3 shows the regression lines for MMR amplitudes andmaturational indices (gestational age, post-conception age,weight, length, and cranial circumference) at the Fz electrode.
rd tone (right column) at Fz (upper row) and Cz (lower row) in ELGA (N = 20) and LGA
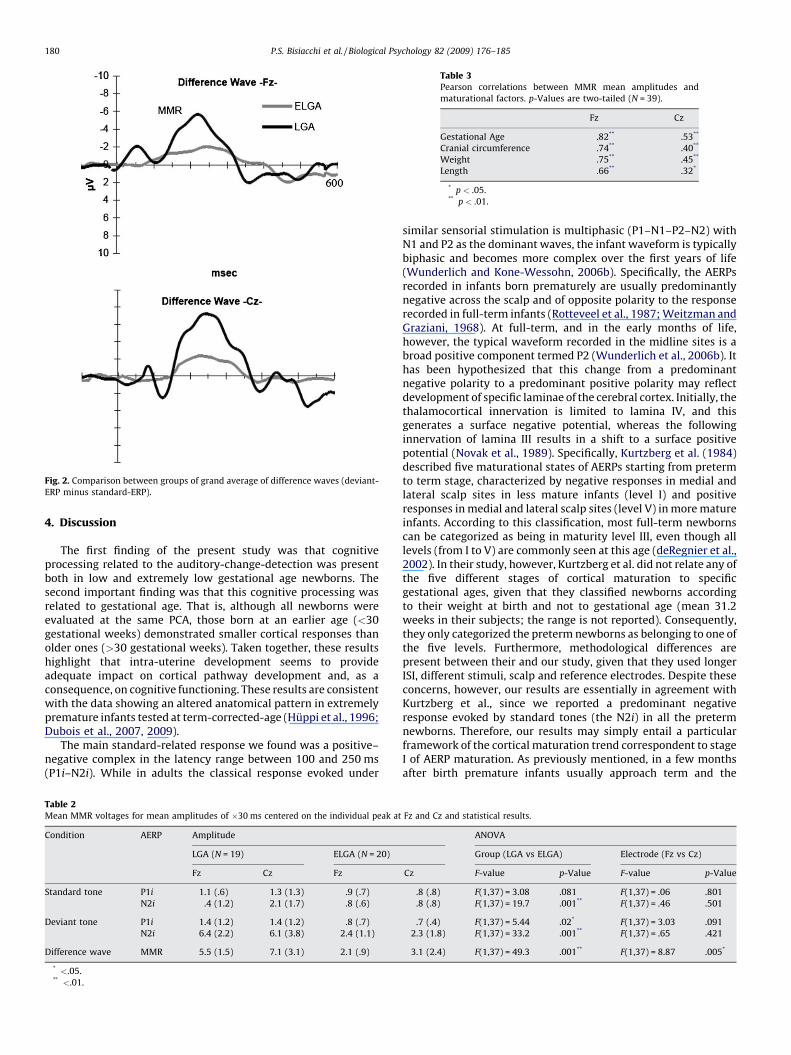
Fig. 2. Comparison between groups of grand average of difference waves (deviant-
ERP minus standard-ERP).
Table 3Pearson correlations between MMR mean amplitudes and
maturational factors. p-Values are two-tailed (N = 39).
Fz Cz
Gestational Age �.82** �.53**
Cranial circumference �.74** �.40**
Weight �.75** �.45**
Length �.66** �.32*
* p < .05.** p < .01.
P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185180
4. Discussion
The first finding of the present study was that cognitiveprocessing related to the auditory-change-detection was presentboth in low and extremely low gestational age newborns. Thesecond important finding was that this cognitive processing wasrelated to gestational age. That is, although all newborns wereevaluated at the same PCA, those born at an earlier age (<30gestational weeks) demonstrated smaller cortical responses thanolder ones (>30 gestational weeks). Taken together, these resultshighlight that intra-uterine development seems to provideadequate impact on cortical pathway development and, as aconsequence, on cognitive functioning. These results are consistentwith the data showing an altered anatomical pattern in extremelypremature infants tested at term-corrected-age (Huppi et al., 1996;Dubois et al., 2007, 2009).
The main standard-related response we found was a positive–negative complex in the latency range between 100 and 250 ms(P1i–N2i). While in adults the classical response evoked under
Table 2Mean MMR voltages for mean amplitudes of �30 ms centered on the individual peak at
Condition AERP Amplitude
LGA (N = 19) ELGA (N = 20)
Fz Cz Fz
Standard tone P1i 1.1 (.6) 1.3 (1.3) .9 (.7)
N2i �.4 (1.2) �2.1 (1.7) �.8 (.6)
Deviant tone P1i 1.4 (1.2) 1.4 (1.2) .8 (.7)
N2i �6.4 (2.2) �6.1 (3.8) �2.4 (1.1)
Difference wave MMR �5.5 (1.5) �7.1 (3.1) �2.1 (.9)
* <.05.** <.01.
similar sensorial stimulation is multiphasic (P1–N1–P2–N2) withN1 and P2 as the dominant waves, the infant waveform is typicallybiphasic and becomes more complex over the first years of life(Wunderlich and Kone-Wessohn, 2006b). Specifically, the AERPsrecorded in infants born prematurely are usually predominantlynegative across the scalp and of opposite polarity to the responserecorded in full-term infants (Rotteveel et al., 1987; Weitzman andGraziani, 1968). At full-term, and in the early months of life,however, the typical waveform recorded in the midline sites is abroad positive component termed P2 (Wunderlich et al., 2006b). Ithas been hypothesized that this change from a predominantnegative polarity to a predominant positive polarity may reflectdevelopment of specific laminae of the cerebral cortex. Initially, thethalamocortical innervation is limited to lamina IV, and thisgenerates a surface negative potential, whereas the followinginnervation of lamina III results in a shift to a surface positivepotential (Novak et al., 1989). Specifically, Kurtzberg et al. (1984)described five maturational states of AERPs starting from pretermto term stage, characterized by negative responses in medial andlateral scalp sites in less mature infants (level I) and positiveresponses in medial and lateral scalp sites (level V) in more matureinfants. According to this classification, most full-term newbornscan be categorized as being in maturity level III, even though alllevels (from I to V) are commonly seen at this age (deRegnier et al.,2002). In their study, however, Kurtzberg et al. did not relate any ofthe five different stages of cortical maturation to specificgestational ages, given that they classified newborns accordingto their weight at birth and not to gestational age (mean 31.2weeks in their subjects; the range is not reported). Consequently,they only categorized the preterm newborns as belonging to one ofthe five levels. Furthermore, methodological differences arepresent between their and our study, given that they used longerISI, different stimuli, scalp and reference electrodes. Despite theseconcerns, however, our results are essentially in agreement withKurtzberg et al., since we reported a predominant negativeresponse evoked by standard tones (the N2i) in all the pretermnewborns. Therefore, our results may simply entail a particularframework of the cortical maturation trend correspondent to stageI of AERP maturation. As previously mentioned, in a few monthsafter birth premature infants usually approach term and the
Fz and Cz and statistical results.
ANOVA
Group (LGA vs ELGA) Electrode (Fz vs Cz)
Cz F-value p-Value F-value p-Value
.8 (.8) F(1,37) = 3.08 .081 F(1,37) = .06 .801
�.8 (.8) F(1,37) = 19.7 .001** F(1,37) = .46 .501
.7 (.4) F(1,37) = 5.44 .02* F(1,37) = 3.03 .091
�2.3 (1.8) F(1,37) = 33.2 .001** F(1,37) = .65 .421
�3.1 (2.4) F(1,37) = 49.3 .001** F(1,37) = 8.87 .005*
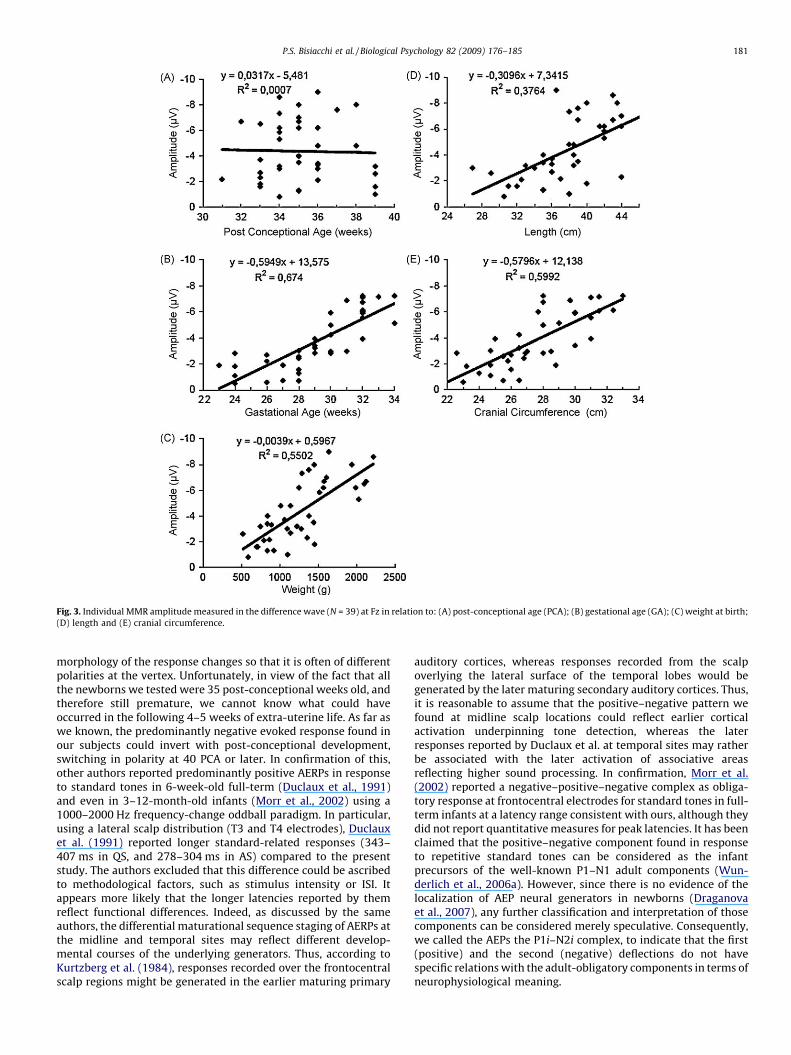
Fig. 3. Individual MMR amplitude measured in the difference wave (N = 39) at Fz in relation to: (A) post-conceptional age (PCA); (B) gestational age (GA); (C) weight at birth;
(D) length and (E) cranial circumference.
P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185 181
morphology of the response changes so that it is often of differentpolarities at the vertex. Unfortunately, in view of the fact that allthe newborns we tested were 35 post-conceptional weeks old, andtherefore still premature, we cannot know what could haveoccurred in the following 4–5 weeks of extra-uterine life. As far aswe known, the predominantly negative evoked response found inour subjects could invert with post-conceptional development,switching in polarity at 40 PCA or later. In confirmation of this,other authors reported predominantly positive AERPs in responseto standard tones in 6-week-old full-term (Duclaux et al., 1991)and even in 3–12-month-old infants (Morr et al., 2002) using a1000–2000 Hz frequency-change oddball paradigm. In particular,using a lateral scalp distribution (T3 and T4 electrodes), Duclauxet al. (1991) reported longer standard-related responses (343–407 ms in QS, and 278–304 ms in AS) compared to the presentstudy. The authors excluded that this difference could be ascribedto methodological factors, such as stimulus intensity or ISI. Itappears more likely that the longer latencies reported by themreflect functional differences. Indeed, as discussed by the sameauthors, the differential maturational sequence staging of AERPs atthe midline and temporal sites may reflect different develop-mental courses of the underlying generators. Thus, according toKurtzberg et al. (1984), responses recorded over the frontocentralscalp regions might be generated in the earlier maturing primary
auditory cortices, whereas responses recorded from the scalpoverlying the lateral surface of the temporal lobes would begenerated by the later maturing secondary auditory cortices. Thus,it is reasonable to assume that the positive–negative pattern wefound at midline scalp locations could reflect earlier corticalactivation underpinning tone detection, whereas the laterresponses reported by Duclaux et al. at temporal sites may ratherbe associated with the later activation of associative areasreflecting higher sound processing. In confirmation, Morr et al.(2002) reported a negative–positive–negative complex as obliga-tory response at frontocentral electrodes for standard tones in full-term infants at a latency range consistent with ours, although theydid not report quantitative measures for peak latencies. It has beenclaimed that the positive–negative component found in responseto repetitive standard tones can be considered as the infantprecursors of the well-known P1–N1 adult components (Wun-derlich et al., 2006a). However, since there is no evidence of thelocalization of AEP neural generators in newborns (Draganovaet al., 2007), any further classification and interpretation of thosecomponents can be considered merely speculative. Consequently,we called the AEPs the P1i–N2i complex, to indicate that the first(positive) and the second (negative) deflections do not havespecific relations with the adult-obligatory components in terms ofneurophysiological meaning.

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185182
The adult P1, whose generators have been identified in theauditory primary cortex (Godey et al., 2001; Liegeois-Chauvelet al., 1994), represents the sensory processing of stimuli, and itsamplitude is related to age (Ponton et al., 2000). The N2, which inthe adult is known to originate bilaterally in the auditory cortex inthe superior temporal lobes with frontal predominance (Cepo-niene et al., 2002; Gomot et al., 2000), has been identified as themain obligatory response in neonatal AERPs (Wunderlich andKone-Wessohn, 2006b) and its amplitude is known to decreasewith age with the emergence of the N1 component in childhood(Fellman and Huotilainen, 2006). Comparing mean amplitudes ofthe P1i response in the standard-related wave, we found nosignificant difference between the two groups but there was asignificant difference in N2i mean amplitude, with the LGA groupdemonstrating greater responses than the ELGA. This could be dueto a more immature primary cortical organization in children bornunder 30 weeks of gestational age (Dubois et al., 2007; Inder et al.,2005). The responses elicited by deviant tones were characterizedby an enhancement of P1i response for the LGA group incomparison with the ELGA group, and this result could reflectdiminished sensorial processing in extremely premature new-borns. Previous findings by Mikkola et al. (2007) reported areduction in amplitude of this wave in five-year-old children bornprematurely compared to full-term controls, although a directcomparison with our results is not possible, since they dividedsubjects into an AGA (appropriate for gestational age), an SGA(small for gestational age) and a control group according to theirbirth weight and not by gestational age as we did here. Thus, themean age of the two experimental groups they tested was thesame (around 27 gestational weeks), precluding the study of theAERP differences related to longer extra-uterine environmentexposure.
The main deviant-related response we found was a prominentnegative wave in the latency range between 150 and 400 ms,which was significantly smaller in ELGA than in LGA. This responsehas been referred to in the infant AERP literature as the precursor ofthe adult MMN (Alho et al., 1990; Ceponiene et al., 1998, 2002;Cheour et al., 1998, 2000, 2002a, 2002b, 2002c; Cheour-Luhtanenet al., 1995, 1996; Kurtzberg et al., 1995). However, with regard tothe P1i–N2i complex, although this response was alwaysnegatively displaced in our recordings, we cannot be certain thatit really corresponds to the adult MMN, whose neural correlateshave been well described (Giard et al., 1994), since there is no wayof knowing where exactly in the cortex this neonatal responsetakes place. Consequently, we considered it as an MMR, regardlessof its polarity. In any case, in the absence of identification of itsneural generators, in newborns it is also assumed to reflect achange-detection mechanism involving both the sensory tracesencoding echoic memory and a matching process with newafferent deviant inputs, reflecting a pre-attentive cognitive processand, as such, a reliable index of cognitive functioning even at thisearly age.
Another possible explanation of the negative deflection foundwithin the latency range in which MMN usually occurs could bethat it does not reflect a context or sensory memory-basedresponse, but rather only an enhanced immature obligatoryresponse, triggered by new afferent sensorial elements of higherfrequency. In this case the large physical difference betweenstandard and deviant stimuli would simply activate a differentneuronal population tuned for the deviant simply becomingresponsive. However, in order to rule out this hypothesis, in aprevious work (Suppiej et al., under revision) we presentedpreterm infants with similar clinical and maturational character-istics with a control condition in which the standard and thedeviant tones (standard 2000 Hz, deviant 1000 Hz) were switched.In this case we also elicited an MMR with comparable amplitude
and latency, demonstrating no difference between the standardand deviant-related ERPs. Therefore, we can reasonably concludethat an MMN-like response is present in preterm newborns.
Our data also show that the MMR was followed in many casesby a positive deflection in the latency range of 250–700 ms with amean peak latency of about 450 ms from stimulus onset and highinter- and intra-subject variability in latency and amplitude. Thefunctional meaning of this wave is still unclear in this neonatalpopulation, although it has been claimed to play a role in stimulusprocessing involving the hypothetical presence of attentivemechanisms at birth (Ceponiene et al., 2002; Fellman et al.,2004; Kushnerenko et al., 2002; Tokioka et al., 1995). This wave hasbeen considered to be analogous to the P3a component thatfollows MMN in adults, where it is thought to reflect an attention-orienting mechanism (Hruby and Marsalek, 2003; Squires et al.,1975), although this hypothesis is not supported by any data innewborn populations, especially those that are preterm. Further-more, P3a in adults has a frontocentral distribution and isconsidered to reflect frontal activity, which is barely developedduring preterm infancy. Yet, this positivity could be related to‘‘actively’’ monitoring the environment during AS, reflecting such akind of automatic orienting reflex as described by Sokolov (1960)in adults rather than to an attentive function in its typical cognitiveprocessing sense. It has also been suggested that this late positivecomponent could partly overlap MMN in some cases (i.e. with alow-discriminatory stimulus deviance), providing a possibleinterpretation of the failure to elicit an MMN or the finding thatit is positively displaced (Morr et al., 2002).
A constraint of the present study is that the central scalpdistribution of electrodes did not allow observation of the possiblepresence of laterality predominance in cortical responses. Thiseffect could be present at this early age, since previous ERP studiesdemonstrated higher responses for speech stimuli over the lefthemisphere in full-term newborns (Cheour et al., 2002b), whereasthe MMN amplitude to tone-frequency change has been found tobe slightly larger over the right hemisphere (Cheour-Luhtanenet al., 1995, 1996). These differences seem to persist withdevelopment, since some authors have reported functionaldifferences between hemispheres in 3-month-old infants usingfMRI (Dehaene-Lambertz et al., 2002). The different functionalorganization of the brain at birth is also sustained by the presenceof structural asymmetries in the temporal lobes (i.e. the planumtemporale) as early as at 29 weeks of gestational age (Wada et al.,1975). Investigating the presence of a laterality effect in pretermnewborns is, in our opinion, an important issue since earlydisruptions in cortical organization development during intra-uterine life, such as gyrification and sulcation, may be related toanatomical and functional anomalies, for instance, altered func-tional asymmetry, underlying impairments in language and socialbehavior (Dubois et al., 2007). Yet, given the clinical condition ofthe participants in the present study, we used a very small numberof electrodes to minimize stress in the newborns.
Nevertheless, as future challenge, it could be interesting toinvestigate, on one side, the influence of gestational developmenton the lateralization of the ERP activity related to central auditoryprocessing, as well as, on the other side, to verify what the MMRwould look like if different deviant stimuli were presented with thesame probability and embedded within a sequence of standardtones but with the same inter-deviant interval. This controlcondition would help us to verify whether a similar mismatchresponse is present, and how it modifies with different kinds ofsensorial deviations. While this could be time-consuming withinthe traditional auditory oddball experimental paradigm and, assuch, not feasible in the case of clinical or developmental groups(such as preterm newborns), innovative multi-feature paradigmsemploying different types of acoustic changes in the same sound

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185 183
sequence (Naatanen et al., 2004) may represent a reliable solutionto gain a wider picture.
The altered cortical functioning in newborns under the criticalage of 30 gestational weeks is probably due to the neurobiologicalproblems caused by the extreme preterm birth (Bhutta and Arand,2002). Indeed, even in the absence of specific and identifiablecentral nervous system insults, preterm birth implies a potentialdisruption to the typical temporal and spatial progression ofcorticogenesis that occurs during the critical periods of the firstweeks of fetal life. Cerebral development processes taking place inthis delicate period, such as the migration phase, organization/differentiation, myelination, and synaptogenesis (Aylward, 2005;Evrard et al., 1992; Geidd, 1997; Gressens et al., 1992; Volpe, 1996)might develop differently in the extra-uterine environment, asseems to occur in extremely preterm newborns. Consistent withthis is the fact that several factors of endogenous neuroprotection,such as neurotrophins and oligotrophins, do not operate wellunder 30 weeks of gestational age, causing as a consequence a highpercentage of cell death. Moreover, the ematho-encephalic barrierdoes not function before 27 weeks of gestational age due to theimmaturity of endothelial and ependymal cells. This allows toxinsto enter the infant’s brain, producing a greater threat to corticalorganization (Aylward, 2005).
One possible interpretation of the failure of the earlyenvironmental sensorial exposure in pairing the effects of thelow gestational age could be addressed by the fact that the earlyauditory experience in the NICU may be harmful at ages 23–25weeks. Indeed, this exposure contains many sinusoidal tones anddifferent kinds of beeps and noise coming from the medicalinstruments. For instance, even though the mother’s voice has beendemonstrated to modify cortical functioning in full-term new-borns, the same kind of stimulation does not have significanteffects in preterm newborns (deRegnier et al., 2002). Therefore, anunstructured auditory stimulation might not be useful for auditorycentral pathway development, resulting in altered ELGA innewborns, as suggested by Graven (2000).
Given the increase in the percentage of preterm births and theevidence in the literature about its relationship with cognitiveimpairments in childhood at school-age, and taking into accountthe relationship between the latter and change-detection corticalresponses reported by previous studies (Kujala and Naatanen,2001; Rinker et al., 2007), this issue, if confirmed by follow-upstudies, could be considered from a prognostic perspective as anew tool for the diagnosis of the cognitive dysfunction at this veryearly stage.
In conclusion, this study stresses the importance of taking intoaccount gestational age when evaluating the risk exposure toneurological and cognitive dysfunctions of premature newborns.Finally, we believe that crucial information will be derived fromthe neurocognitive follow-up of the same groups of LGA and ELGAwhich we are currently carrying out at the same hospital.
Acknowledgments
We thank both the reviewers for their helpful and interestingcomments and suggestions.
References
Ajayi-Obe, M., Saeed, N., Cowan, F.M., Rutherford, M.A., Edwards, A.D., 2000.Reduced development of cerebral cortex in extremely preterm infants. Lancet356 (9236), 1162–1163.
Alho, K., Sainio, K., Sajaniemi, N., Reinikainen, K., Naatanen, R., 1990. Event-relatedbrain potential of human newborns to pitch change of an acoustic stimulus.Electroencephalography and Clinical Neurophysiology 77 (2), 151–155.
Als, H., Lawhon, G., Duffy, F.H., McAnulty, G.B., Gibes-Grossman, R., Blickman, J.G.,1994. Individualized developmental care for the very low-birth-weight preterm
infant. Medical and neurofunctional effects. Journal of the American MedicalAssociation 272 (11), 853–858.
Anders, T., Emde, R., Parmelee, A. (Eds.), 1971. A Manual of Standardized Termi-nology: Techniques and Criteria for Scoring of States of Sleep and Wakefulnessin Newborn Infants. UCLA Brain Information Service/Brain Research Institute,Los Angeles.
Aylward, G.P., Hatcher, R.P., Leavitt, L.A., Rao, V., Bauer, C.R., Brennan, M.J., Gus-tafson, N.F., 1984. Factors affecting neurobehavioral responses of preterminfants at term conceptional age. Child Development 55 (4), 1155–1165.
Aylward, G.P., 2005. Neurodevelopmental outcomes of infants born prematurely.Journal of Developmental and Behavioral Pediatrics 26 (6), 427–440.
Biagioni, E., Frisone, M.F., Laroche, S., Kapetanakis, B.A., Ricci, D., Adeyi-Obe, M.,Lewis, H., Kennea, N., Cioni, G., Cowan, F., Rutherford, M., Azzopardi, D., Mercuri,E., 2007. Maturation of cerebral electrical activity and development of corticalfolding in young vs. preterm infants. Clinical Neurophysiology 118 (1), 53–59.
Bhutta, A.T., Arand, K.J.S., 2002. Vulnerability of the developing brain. Neuronalmechanisms. Clinics in Perinatology 29, 357–372.
Brown, N.C., Doyle, L.W., Bear, M.J., Inder, T.E., 2006. Alterations in neurobehavior atterm reflect differing perinatal exposures in very preterm infants. Pediatrics118 (6), 2461–2471.
Brazy, J.E., Eckerman, C.O., Oehler, J.M., Goldstein, R.F., O’Rand, A.M., 1991. NurseryNeurobiologic Risk Score: important factor in predicting outcome in very lowbirth weight infants. Journal of Pediatrics 118 (5), 783–792.
Ceponiene, R., Cheour, M., Naatanen, R., 1998. Interstimulus interval and auditoryevent-related potentials in children: evidence for multiple generators. Electro-encephalography and Clinical Neurophysiology 108, 345–354.
Ceponiene, R., Kushnerenko, E., Fellman, V., Renlund, M., Suominen, K., Naatanen, R.,2002. Event-related potential features indexing central auditory discriminationby newborns. Brain Research. Cognitive Brain Research 13 (1), 101–113.
Cheour, M., Alho, K., Ceponiene, R., Reinikainen, K., Sainio, K., Pohjavuori, M.,Aaltonen, O., Naatanen, R., 1998. Maturation of mismatch negativity in infants.International Journal of Psychophysiology 29 (2), 217–226.
Cheour, M., Ceponiene, R., Leppanen, P., Alho, K., Kujala, T., Renlund, M., Fellman, V.,Naatanen, R., 2002a. The auditory sensory memory trace decays rapidly innewborns. Scandinivian Journal of Psychology 43 (1), 33–39.
Cheour, M., Kushnerenko, E., Ceponiene, R., Fellman, V., Naatanen, R., 2002b. Electricbrain responses obtained from newborn infants to changes in duration incomplex harmonic tones. Developmental Neuropsychology 22 (2), 471–479.
Cheour, M., Leppanen, P.H., Kraus, N., 2000. Mismatch negativity (MMN) as a tool forinvestigating auditory discrimination and sensory memory in infants andchildren. Clinical Neurophysiology 111, 4–16.
Cheour, M., Martynova, O., Naatanen, R., Erkkola, R., Sillanpaa, M., Kero, P., Raz, A.,Kaipio, M.L., Hiltunen, J., Aaltonen, O., Savela, J., Hamalainen, H., 2002c. Speechsounds learned by sleeping newborns. Nature 415 (6872), 599–600.
Cheour, M., 2007. Development of Mismatch Negativity (MMN). In: deHaan, M.(Ed.), Infant EEG and Event-Related Potentials, 28. Psychology Press, pp. 171–198.
Cheour-Luhtanen, M., Alho, K., Kujala, T., Sainio, K., Reinikainen, K., Renlund, M.,Aaltonen, O., Eerola, O., Naatanen, R., 1995. Mismatch negativity indicatesvowel discrimination in newborns. Hearing Research 82 (1), 53–58.
Cheour-Luhtanen, M., Alho, K., Sainio, K., Rinne, T., Reinikainen, K., Pohjavuori, M.,Renlund, M., Aaltonen, O., Eerola, O., Naatanen, R., 1996. The ontogeneticallyearliest discriminative response of the human brain. Psychophysiology 33 (4),478–481.
Curzi-Dascalova, L., Peirano, P., Morel-Kahn, F., 1988. Development of sleep states innormal premature and full-term newborns. Developmental Psychobiology 21,431–444.
Dehaene-Lambertz, G., Dehaene, S., 1994. Speed and cerebral correlates of syllablediscrimination in infants. Nature 370 (6487), 292–295.
Dehaene-Lambertz, G., 2000. Cerebral specialization for speech and non-speechstimuli in infants. Journal of Cognitive Neuroscience 12 (3), 449–460.
Dehaene-Lambertz, G., Dehaene, S., Hertz-Pannier, L., 2002. Functional neuroima-ging of speech perception in infants. Science 298 (5600), 2013–2015.
deRegnier, R., Wewerka, S., Georgieff, M.K., Mattia, F., Nelson, C.A., 2002. Influencesof postconceptional age and postnatal experience on the development ofauditory recognition memory in the newborn infant. Developmental Psycho-biology 41 (3), 216–225.
Draganova, R., Eswaran, H., Murphy, P., Lowery, C., Preissl, H., 2007. Serial magne-toencephalographic study of fetal and newborn auditory discriminative evokedresponses. Early Human Development 83 (3), 199–207.
Drillien, C.M., Thomson, A.J., Burgoyne, K., 1980. Low-birthweight children at earlyschool-age: a longitudinal study. Developmental Medicine and Child Neurology22 (1), 26–47.
Dubois, J., Benders, M., Cachia, A., Lazeyras, F., Ha-Vinh Leuchter, R., Sizonenko, S.V.,Borradori-Tolsa, C., Mangin, J.F., Huppi, P.S., 2007. Mapping the early corticalfolding process in the preterm newborn brain. Cerebral Cortex 18 (6), 1444–1454.
Dubois, J., Hertz-Pannier, L., Cachia, A., Mangin, J.F., Le Bihan, D., Dehaene-Lambertz,G., 2009. Structural asymmetries in the infant language and sensory–motornetworks. Cerebral Cortex 19 (2), 414–423.
Duclaux, R., Challamel, M.J., Collett, L., Roullet-Solignac, I., Revol, M., 1991. Hemi-spheric asymmetry of late auditory evoked response induced by pitch changesin infants: influence of sleep stages. Brain Research 6 (566 (1–2)), 152–158.
Eyre, J.A., Nanei, S., Wilkinson, A.R., 1988. Quantification of changes in normalneonatal EEGs with gestation from continuous five-day recordings. Develop-mental Medicine and Child Neurology 30 (5), 599–607.

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185184
Evrard, P., Gressens, P., Volpe, J.J., 1992. New concepts to understand the neuro-logical consequences of subcortical lesions in the premature brain. Biology ofthe Neonate 61, 1–3.
Fellman, V., Kushnerenko, E., Mikkola, K., Ceponiene, R., Leipala, J., Naatanen, R.,2004. Atypical auditory event-related potentials in preterm infants during thefirst year of life: a possible sign of cognitive dysfunction? Pediatric Research 56(2), 291–297.
Fellman, V., Huotilainen, M., 2006. Cortical auditory event-related potentials innewborns infants. Seminars in Fetal and Neonatal Medicine 11 (6), 452–458.
Ferrari, F., Torricelli, A., Giustardi, A., Benfatti, A., Balzani, R., Ori, L., Frigieri, G., 1992.Bioelectric brain maturation in full-term infants and in healthy and pathologicalpreterm infants at term post-menstrual age. Early Human Development 28 (1),37–63.
Friederici, A.D., Friedrich, M., Weber, C., 2002. Neural manifestation of cognitive andprecognitive mismatch detection in early infancy. Neuroreport 13 (10), 1251–1254.
Geidd, J.N., 1997. Normal development. Child and Adolescent Clinics of NorthAmerica 6, 265–282.
Giard, M.H., Perrin, F., Echallier, J.F., Thevenet, M., Froment, J.C., Pernier, J., 1994.Dissociation of temporal and frontal components in the human auditory N1wave: a scalp current density and dipole model analysis. Electroencephalogr-phy and Clinical Neurophysiology 92 (3), 238–252.
Godey, B., Schwartz, D., de Graaf, J.B., Chauvel, P., Liegeois-Chauvel, C., 2001.Neuromagnetic source localization of auditory evoked fields and intracerebralevoked potentials: a comparison of data in the same patients. Clinical Neuro-physiology 112 (10), 1850–1859.
Gomot, M., Giard, M.H., Roux, S., Barthelemy, C., Bruneau, N., 2000. Maturation offrontal and temporal components of mismatch negativity (MMN) in children.Neuroreport 11 (14), 3109–3112.
Graven, S.N., 2000. Sound and the developing infant in the NICU: conclusions andrecommendations for care. Journal of Perinatology 20 (8 Pt 2), 88–93.
Gressens, P., Richelme, C., Kadhim, H.J., Gadisseux, J.F., Evrard, P., 1992. Thegerminative zone produces the most cortical astrocytes after neuronal migra-tion in the developing mammalian brain. Biology of the Neonate 61, 4–24.
Hack, M., Fanaroff, A.A., 2000. Outcomes of children of extremely low birthweightand gestational age in the 1990s. Seminars in Neonatology 5 (2), 89–106.
Holst, M., Eswaran, H., Lowery, C., Murphy, P., Norton, J., Preissl, H., 2005. Devel-opment of auditory evoked fields in human fetuses and newborns: a long-itudinal MEG study. Clinical Neurophysiology 116 (8), 1949–1955.
Hruby, T., Marsalek, P., 2003. Event-related potentials—the P3 wave. Acta Neuro-biologiae Experimentalis 63 (1), 55–63.
Huppi, P.S., Schuknecht, B., Boesch, C., Bossi, E., Felblinger, J., Fusch, C., Herschko-witz, N., 1996. Structural and neurobehavioral delay in postnatal brain devel-opment of preterm infants. Pediatrics Research 39 (5), 895–901.
Huppi, P.S., Inder, T.E., 2001. Magnetic resonance techniques in the evaluation of theperinatal brain: recent advances and future directions. Seminars in Neonatol-ogy 6 (2), 195–210.
Inder, T.E., Warfield, S.K., Wang, H., Huppi, P.S., Volpe, J.J., 2005. Abnormal cerebralstructure is present at term in premature infants. Pediatrics 115, 286–294.
Jasper, H.H., 1958. The ten–twenty electrode system of the International Federation.Electroencephalography and Clinical Neurophysiology 10, 371–437.
Kapellou, O., Counsell, S.J., Kennea, N., Dyet, L., Saeed, N., Stark, J., Maalouf, E., Duggan,P., Ajayi-Obe, M., Hajnal, J., Allsop, J.M., Boardman, J., Rutherford, M.A., Cowan, F.,Edwards, A.D., 2006. Abnormal cortical development after premature birthshown by altered allometric scaling of brain growth. PLoS Medicine 3 (8), 265.
Kujala, T., Naatanen, R., 2001. The mismatch negativity in evaluating centralauditory dysfunction in dyslexia. Neuroscience and Biobehavioral Reviews25 (6), 535–543.
Kurtzberg, D., Hilpert, P.L., Kreuzer, J.A., Vaughan Jr., H.G., 1984. Differentialmaturation of cortical auditory evoked potentials to speech sounds in normalfull-term and very low-birthweight infants. Developmental Medicine ChildNeurology 26, 466–475.
Kurtzberg, D., Vaughan, H.G., Kreuzer, J.A., Flieger, K.Z., 1995. Developmentalstudies and clinical application of mismatch negativity: problems and pro-spects. Ear and Hearing 16 (1), 105–117.
Kushnerenko, E., Cheour, M., Ceponiene, R., Fellman, V., Renlund, M., Soininen, K.,Alku, P., Koskinen, M., Sainio, K., Naatanen, R., 2001. Central auditory processingof durational changes in complex speech patterns by newborns: an event-related brain potential study. Developmental Neuropsychology 19 (1), 83–97.
Kushnerenko, E., Ceponiene, R., Balan, P., Fellman, V., Naatanen, R., 2002. Maturationof the auditory change detection response in infants: a longitudinal AERP study.Neuroreport 13 (15), 1843–1848.
Leonard, L.B., Eyer, J.A., Bedore, L.M., Grela, B.G., 1997. Three accounts of thegrammatical morpheme difficulties of English-speaking children with specificlanguage impairment. Journal of Speech, Language, and Hearing Research 40(4), 741–753.
Leppanen, P.H.T., Eklund, K.M., Lyytinen, H., 1997. Event-related brain potentials tochange in rapidly presented acoustic stimuli in newborns. DevelopmentalNeuropsychology 13, 175–204.
Leppanen, P.H.T., Guttorm, T.K., Pihko, E., Takkinen, S., Eklund, K.M., Lyytinen, H.,2004. Maturational effects on newborn AERPs measured in the mismatchnegativity paradigm. Experimental Neurology 1, 91–101.
Liegeois-Chauvel, C., Musolino, A., Badier, J.M., Marquis, P., Chauvel, P., 1994. Evokedpotentials recorded from the auditory cortex in man: evaluation and topogra-phy of the middle latency components. Electroencephalography and ClinicalNeurophysiology 92 (3), 204–214.
Majnemer, A., Rosenblatt, B., Riley, P.S., 1993. Influence of gestational age, birthweight, and asphyxia on neonatal neurobehavioral performance. PediatricNeurology 9 (3), 181–186.
Marlow, N., Wolke, D., Bracewell, M., Samara, M., for the EPICure Study Group, 2005.Neurologic and developmental disability at six years of age after extremelypreterm birth. The New England Journal of Medicine 352 (1), 9–19.
Mercuri, E., von Siebenthal, K., Tutuncuoglu, S., Guzzetta, F., Casaer, P., 1995. Theeffect of behavioural states on visual evoked responses in preterm and full-termnewborns. Neuropediatrics 26 (4), 211–213.
McIsaac, H., Polich, J., 1992. Comparison of infant and adult P300 from auditorystimuli. Journal of Experimental Child Psychology 53 (2), 115–128.
Mikkola, K., Kushnerenko, E., Partanen, E., Serenius-Sirve, S., Leipala, J., Huotilainen,M., Fellman, V., 2007. Auditory event-related potentials and cognitive functionof preterm children at five years of age. Clinical Neurophysiology 118 (7), 1494–1502.
Morr, M.L., Shafer, V.L., Kreuzer, J.A., Kurtzberg, D., 2002. Maturation of mismatchnegativity in typically developing infants and preschool children. Ear andHearing 23 (2), 118–136.
Naatanen, R., Gaillard, A.W., Mantysalo, S., 1978. Early selective-attention effect onevoked potential reintAERPreted. Acta Psychology 42 (4), 313–329.
Naatanen, R., 1990. The role of attention in auditory information processing asrevealed by event-related potentials and other brain measures of cognitivefunction. Behavioural and Brain Sciences 13, 201–288.
Naatanen, R., 1992. Attention and Brain Function. Lawrence Erlbaum, Mahwah, NJ.Naatanen, R., Pakarinen, S., Rinne, T., Takegata, R., 2004. The mismatch negativity
(MMN): towards the optimal paradigm. Clinical Neurophysiology 115, 140–144.
Nolte, R., Haas, G., 1978. A polygraphic study of bioelectrical brain maturation inpreterm infants. Developmental Medicine and Child Neurology 20 (2), 167–182.
Novak, G.P., Kurtzberg, D., Kreuzer, J.A., Vaughan, H.G., 1989. Cortical responses tospeech sounds and their formants in normal infants: maturational sequenceand spatiotemporal analysis. Electroencephalography and Clinical Neurology73, 295–305.
Novitski, N., Huotilainen, M., Tervaniemi, M., Naatanen, R., Fellman, V., 2007.Neonatal frequency discrimination in 250–4000 Hz range: electrophysiologicalevidence. Clinical Neurophysiology 118 (2), 412–419.
Nunes, M.L., Da Costa, J.C., Moura-Ribeiro, M.V., 1997. Polysomnographic quanti-fication of bioelectrical maturation in preterm and full-term newborns atmatched conceptional ages. Electroencephalography and Clinical Neurophy-siology 102 (3), 186–191.
Ostfeld, B.M., Smith, R.H., Hiatt, M., Hegyi, T., 2000. Maternal behavior towardpremature twins: implications for development. Twin Research 3 (4), 234–241.
Pietz, J., Peter, J., Graf, R., Rauterberg-Ruland, I., Rupp, A., Sontheimer, D., Linder-kamp, O., 2004. Physical growth and neurodevelopmental outcome of non-handicapped low-risk children born preterm. Early Human Development 79 (2),131–143.
Piper, M.C., Byrne, P.J., Pinnell, L.E., 1989. Influence of gestational age on earlyneuromotor development in the preterm infant. American Journal of Perina-tology 6 (4), 405–411.
Ponton, C.W., Eggermont, J.J., Kwong, B., Don, M., 2000. Maturation of human centralauditory system activity: evidence from multi-channel evoked potentials.Clinical Neurophysiology 111 (2), 220–236.
Rijken, M., Stoelhorst, G.M., Martens, S.E., van Zwieten, P.H., Brand, R., Wit, J.M.,Veen, S., 2003. Mortality and neurologic, mental, and psychomotor develop-ment at 2 years in infants born less than 27 weeks’ gestation: the Leiden follow-up project on prematurity. Pediatrics 112 (2), 351–358.
Rinker, T., Kohls, G., Richter, C., Maas, V., Schulz, E., Schecker, M., 2007. Abnormalfrequency discrimination in children with SLI as indexed by mismatch nega-tivity (MMN). Neuroscience Letters 413 (2), 99–104.
Robertson, C., Sauve, R.S., Christianson, H.E., 1994. Province-based study of neuro-logic disability among survivors weighing 500 through 1249 grams at birth.Pediatrics 93 (4), 636–640.
Rotteveel, J.J., Colon, E.J., Stegeman, D.F., Visco, Y.M., 1987. The maturation of thecentral auditory conduction in preterm infants until 3 months post-term. IV.Composite group averages of the cortical auditory evoked responses (ACRs).Hearing Research 27, 85–93.
Ruusuvirta, T., Huotilainen, M., Fellman, V., Naatanen, R., 2003. The newborn humanbrain binds sound features together. Neuroreport 14 (16), 2117–2119.
Shepherd, A., Saunders, K., McCulloch, D., 1999. Effect of sleep state on the flashvisual evoked potential. A case study. Documenta Ophthalmologica 98 (3), 247–256.
Sokolov, E.N., 1960. Neuronal models and the orienting reflex. In: Brazier, M.A.B.(Ed.), The Central Nervous System and Behavior. Madison Printing Co., Madison,NJ, pp. 187–276.
Squires, N.K., Squires, K.C., Hillyard, S.A., 1975. Two varieties of long-latencypositive waves evoked by unpredictable auditory stimuli in man. Electroence-phalography and Clinical Neurophysiology 38 (4), 387–401.
Stockard-Pope, J.E., Werner, S.S., Bickford, R.G., 1992. Development of Sleep-WakeStates. Atlas of Neonatal Electroencephalography. Raven Press, New York, pp.93–104.
Suppiej, A., Bisiacchi, P.S., 1992. Potenziali evocati uditivi nel neonato: discrimi-nazione della frequenza tonale. In: Palomba, D. (Ed.), Indici Fisiologici inPsicologia. Cleup, Padova, pp. 239–250.
Therien, J.M., Worwa, C.T., Mattia, F.R., deRegnier, R.A., 2004. Altered pathways forauditory discrimination and recognition memory in preterm infants. Develop-mental Medicine and Child Neurology 46 (12), 816–824.

P.S. Bisiacchi et al. / Biological Psychology 82 (2009) 176–185 185
Tokioka, A.B., Pearce, J.W., Crowell, D.H., 1995. Endogenous event-related potentialsin term and preterm infants. Journal of Clinical Neurophysiology 12 (5), 468–475.
Vouloumanos, A., Werker, J.F., 2007. Listening to language at birth: evidence for abias for speech in neonates. Developmental Science 10 (2), 159–164.
Volpe, J.J., 1991. Cognitive deficits in premature infants. New England Journal ofMedicine 325 (4), 276–278.
Volpe, J.J., 1996. Subplate neurons—missing link in brain injury of the prematureinfant? Pediatrics 97 (1), 112–113.
Volpe, J.J., 2000. Intracranial haemorrhage. In: Volpe, J.J. (Ed.), Neurology of theNewborn. WB Saunders Company, Philadelphia, pp. 428–493.
Wada, J.A., Clarke, R., Hamm, A., 1975. Cerebral hemispheric asymmetry in humans.Cortical speech zones in 100 adults and 100 infant brains. Archives of Neurology32 (4), 239–246.
Weitzman, W.D., Graziani, L.J., 1968. Maturation and topography of the auditoryevoked response of the prematurely born infant. Developmental Psychobiology1, 79–89.
Wen, S.W., Smith, G., Yang, Q., Walker, M., 2004. Epidemiology of preterm birth andneonatal outcome. Seminars in Neonatology 9 (6), 429–435.
Wilson-Costello, D., Friedman, H., Minich, N., Fanaroff, A.A., Hack, M., 2005.Improved survival rates with increased neurodevelopmental disability forextremely low birth weight infants in the 1990s. Pediatrics 115 (4), 997–1003.
Wunderlich, J.L., Cone-Wesson, B.K., Shepherd, R., 2006a. Maturation of the corticalauditory evoked potential in infants and young children. Hearing Research 212(1–2), 185–202.
Wunderlich, J.L., Kone-Wessohn, B.K., 2006b. Maturation of CAEP in infants andchildren: a review. Hearing Research 12, 212–223.




















