
Neural Biases to Covert and Overt Signals of Fear: Dissociation by Trait Anxiety and Depression
14
Neural Biases to Covert and Overt Signals of Fear: Dissociation by Trait Anxiety and Depression Leanne M. Williams 1 , Andrew H. Kemp 1 , Kim Felmingham 1 , Belinda J. Liddell 1,2 , Donna M. Palmer 1,3 , and Richard A. Bryant 1,4 Abstract & Although biases toward signals of fear may be an evolu- tionary adaptation necessary for survival, heightened biases may be maladaptive and associated with anxiety or depression. In this study, event-related potentials (ERPs) were used to ex- amine the time course of neural responses to facial fear stimuli (versus neutral) presented overtly (for 500 msec with con- scious attention) and covertly (for 10 msec with immediate masking to preclude conscious awareness) in 257 nonclinical subjects. We also examined the impact of trait anxiety and depression, assessed using psychometric ratings, on the time course of ERPs. In the total subject group, controlled biases to overtly processed fear were reflected in an enhancement of ERPs associated with structural encoding (120–220 msec) and sustained evaluation persisting from 250 msec and beyond, following a temporo-occipital to frontal topography. By con- trast, covert fear processing elicited automatic biases, reflected in an enhancement of ERPs prior to structural encoding (80– 180 msec) and again in the period associated with automatic orienting and emotion encoding (230–330 msec), which fol- lowed the reverse frontal to temporo-occipital topography. Higher levels of trait anxiety (in the clinical range) were dis- tinguished by a heightened bias to covert fear (speeding of early ERPs), compared to higher depression which was asso- ciated with an opposing bias to overt fear (slowing of later ERPs). Anxiety also heightened early responses to covert fear, and depression to overt fear, with subsequent deficits in emo- tion encoding in each case. These findings are consistent with neural biases to signals of fear which operate automatically and during controlled processing, feasibly supported by paral- lel networks. Heightened automatic biases in anxiety may con- tribute to a cycle of hypervigilance and anxious thoughts, whereas depression may represent a ‘‘burnt out’’ emotional state in which evaluation of fear stimuli is prolonged only when conscious attention is allocated. & INTRODUCTION Vigilance for threat is crucial for survival, and neural sys- tems may have evolved to give precedence to arousing signals of potential danger (Williams, 2006). Both behav- ioral and brain function studies provide evidence of a ‘‘negativity bias’’ toward aversive, emotionally arousing stimuli (e.g., Williams, Palmer, Liddell, Song, & Gordon, 2006; Cacioppo, Gardner, & Berntson, 1999; Taylor, 1991). However, excessive biases toward such stimuli may contribute to affective disorders (Williams, 2006). Consistent with this view, cognitive models propose that there is an exaggerated bias in the automatic processing of fear-related stimuli in anxiety disorder, and a distinc- tive negativity bias involving more controlled processing in depression (Mogg & Bradley, 1998; Williams, Watts, MacLeod, & Mathews, 1997). To date, this proposal has been tested only with behavioral measures. Event-related potentials (ERPs) provide an index of neural activity with millisecond precision (Mangun & Hillyard, 1995), well suited to examining the time course of processing biases. In this study, we used ERPs to investigate the time course of neural biases to fear stimuli presented under condi- tions of both automatic and controlled processing, and whether high trait anxiety is associated with a specific ex- acerbation of these biases relative to both low anxiety and depression. In behavioral studies, preferential attention to nega- tively valenced, particularly fear-related, stimuli has been interpreted as a negativity bias (Cacioppo et al., 1999; Ohman, 1997; Gur et al., 1992; Pratto & John, 1991; Hansen & Hansen, 1988). Enhanced negativity biases in anxiety disorder are reflected in faster reaction times to fear-related stimuli in visual probe tasks using both word and face stimuli (Bryant & Harvey, 1997; Bryant, Harvey, Gordon, & Barry, 1995). These biases may generalize to the greater expectation of aversive events (Smith & Bryant, 2000; Warda & Bryant, 1998). Several findings indicate that biases toward fear stimuli in anxiety may arise from automatic processing, without the need for conscious awareness (Bradley, Mogg, Millar, & White, 1 University of Sydney, Westmead Hospital, Sydney, NSW, Aus- tralia, 2 Brain Resource Co., Ultimo, Sydney, NSW, Australia, 3 University of Sydney, Sydney, NSW, Australia, 4 University of NSW, Randwick, Sydney, NSW, Australia D 2007 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 19:10, pp. 1595–1608
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Neural Biases to Covert and Overt Signals of Fear: Dissociation by Trait Anxiety and Depression

Neural Biases to Covert and Overt Signals of Fear:Dissociation by Trait Anxiety and Depression
Leanne M. Williams1, Andrew H. Kemp1, Kim Felmingham1,Belinda J. Liddell1,2, Donna M. Palmer1,3, and Richard A. Bryant1,4
Abstract
& Although biases toward signals of fear may be an evolu-tionary adaptation necessary for survival, heightened biasesmay be maladaptive and associated with anxiety or depression.In this study, event-related potentials (ERPs) were used to ex-amine the time course of neural responses to facial fear stimuli(versus neutral) presented overtly (for 500 msec with con-scious attention) and covertly (for 10 msec with immediatemasking to preclude conscious awareness) in 257 nonclinicalsubjects. We also examined the impact of trait anxiety anddepression, assessed using psychometric ratings, on the timecourse of ERPs. In the total subject group, controlled biasesto overtly processed fear were reflected in an enhancement ofERPs associated with structural encoding (120–220 msec) andsustained evaluation persisting from 250 msec and beyond,following a temporo-occipital to frontal topography. By con-trast, covert fear processing elicited automatic biases, reflectedin an enhancement of ERPs prior to structural encoding (80–
180 msec) and again in the period associated with automaticorienting and emotion encoding (230–330 msec), which fol-lowed the reverse frontal to temporo-occipital topography.Higher levels of trait anxiety (in the clinical range) were dis-tinguished by a heightened bias to covert fear (speeding ofearly ERPs), compared to higher depression which was asso-ciated with an opposing bias to overt fear (slowing of laterERPs). Anxiety also heightened early responses to covert fear,and depression to overt fear, with subsequent deficits in emo-tion encoding in each case. These findings are consistent withneural biases to signals of fear which operate automaticallyand during controlled processing, feasibly supported by paral-lel networks. Heightened automatic biases in anxiety may con-tribute to a cycle of hypervigilance and anxious thoughts,whereas depression may represent a ‘‘burnt out’’ emotionalstate in which evaluation of fear stimuli is prolonged only whenconscious attention is allocated. &
INTRODUCTION
Vigilance for threat is crucial for survival, and neural sys-tems may have evolved to give precedence to arousingsignals of potential danger (Williams, 2006). Both behav-ioral and brain function studies provide evidence of a‘‘negativity bias’’ toward aversive, emotionally arousingstimuli (e.g., Williams, Palmer, Liddell, Song, & Gordon,2006; Cacioppo, Gardner, & Berntson, 1999; Taylor,1991). However, excessive biases toward such stimulimay contribute to affective disorders (Williams, 2006).Consistent with this view, cognitive models propose thatthere is an exaggerated bias in the automatic processingof fear-related stimuli in anxiety disorder, and a distinc-tive negativity bias involving more controlled processingin depression (Mogg & Bradley, 1998; Williams, Watts,MacLeod, & Mathews, 1997). To date, this proposal hasbeen tested only with behavioral measures. Event-related
potentials (ERPs) provide an index of neural activity withmillisecond precision (Mangun & Hillyard, 1995), wellsuited to examining the time course of processing biases.In this study, we used ERPs to investigate the time courseof neural biases to fear stimuli presented under condi-tions of both automatic and controlled processing, andwhether high trait anxiety is associated with a specific ex-acerbation of these biases relative to both low anxiety anddepression.
In behavioral studies, preferential attention to nega-tively valenced, particularly fear-related, stimuli has beeninterpreted as a negativity bias (Cacioppo et al., 1999;Ohman, 1997; Gur et al., 1992; Pratto & John, 1991;Hansen & Hansen, 1988). Enhanced negativity biases inanxiety disorder are reflected in faster reaction times tofear-related stimuli in visual probe tasks using both wordand face stimuli (Bryant & Harvey, 1997; Bryant, Harvey,Gordon, & Barry, 1995). These biases may generalizeto the greater expectation of aversive events (Smith &Bryant, 2000; Warda & Bryant, 1998). Several findingsindicate that biases toward fear stimuli in anxiety mayarise from automatic processing, without the need forconscious awareness (Bradley, Mogg, Millar, & White,
1University of Sydney, Westmead Hospital, Sydney, NSW, Aus-tralia, 2Brain Resource Co., Ultimo, Sydney, NSW, Australia,3University of Sydney, Sydney, NSW, Australia, 4University ofNSW, Randwick, Sydney, NSW, Australia
D 2007 Massachusetts Institute of Technology Journal of Cognitive Neuroscience 19:10, pp. 1595–1608

1995; Mogg, Bradley, & Williams, 1995; Mogg, Bradley,Williams, & Mathews, 1993). Although contrary find-ings have also been reported (Mathews, Ridgeway, &Williamson, 1996), excessive automatic biases towardfear cues could feasibly contribute to an ongoing cyclein which vigilance for even low probability sources ofdanger increases, and anxious thoughts are intensified(Beck, 1976).
In depression, negativity biases are thought to be ap-parent only when there is sustained stimulus evaluation,or stimuli are particularly salient (Mogg et al., 1995).These biases have been revealed on modified Stroop,visual probe, and facial emotion recognition tasks, whensustained evaluation is promoted by long stimulus du-rations (Gur et al., 1992; Mogg et al., 1993, 1995; Segal,Gemar, Truchon, Guirguis, & Horowitz, 1995; Gotlib &Cane, 1987). Although these studies have focused onstimuli related to sadness, the explicit recognition of fearis also reduced in depressed individuals (Lenti, Giacobbe,& Pegna, 2000). Excessive activation of the amygdala,associated with arousing and aversive stimuli, has alsobeen observed in depressed patients (Siegle, Steinhauer,Thase, Stenger, & Carter, 2002; Drevets, 2001; Shelineet al., 2001). Moreover, depression has been associatedwith particularly sustained amygdala activation, and it hasbeen suggested that the time course of this activity maymediate the negativity biases observed in behavioralstudies (Siegle et al., 2002).
Unlike behavioral measures, event-related brain po-tentials (ERPs) provide a direct window onto the timecourse of neural activity, which may help distinguishautomatic versus controlled biases toward fear stimuli,and whether anxiety is associated with a specific exag-geration of automatic biases versus an exaggeration ofcontrolled biases in depression. Facial expressions offear have been found to enhance ERPs within the first200 msec of stimulus processing, and then again ina more persistent manner after 250 msec, consistentwith biases in automatic orienting, and subsequentsustained evaluation (Williams, Palmer, et al., 2006;Ashley, Vuilleumier, & Swick, 2004). When awareness isrestricted by masking, enhancements in early ERPs remain,but sustained activity is absent (Liddell, Palmer, Gordon,& Williams, under review; Liddell, Williams, Rathjen,Shevrin, & Gordon, 2004; Williams, Liddell, et al., 2004).Masked fearful faces have also been found to elicit en-hancements in the N250 and early P300 associated withautomatic orienting and encoding of significant stimuli(Liddell et al., 2004). The N250 has also been related toencoding emotional expressions and integrating salientstimuli into a gestalt (Han, He, & Woods, 2000; Streit,Wowler, Brinkmeyer, Ihl, & Gaebel, 2000). Functional mag-netic resonance imaging (fMRI) studies, undertaken withequivalent behavioral tasks, suggest that parallel visualpathways to the amygdala may support these responsesto fear at differing levels of awareness: a direct extra-striate pathway for automatic orienting and a slower,
indirect striate occipito-temporal pathway for sustained con-scious evaluation (Williams et al., 2006; Liddell et al., 2005;Morris, Ohman, & Dolan, 1999; Whalen et al., 1998).
ERPs have been used to study negativity biases in anx-iety in only two studies to date. One study focused onanticipatory biases prior to stimulus onset, which wereenhanced to negatively valenced stimuli in participantswith high state (and combined state–trait) anxiety, con-sistent with greater vigilance to these stimuli (Carretieet al., 2004). Another study reported larger and fasterearly ERPs to fear-related face stimuli, in particular, dur-ing an attention-shifting task in 13 ‘‘high anxious’’ com-pared to ‘‘low anxious’’ participants (Bar-Haim, Lamy, &Glickman, 2005). Other techniques provide convergentevidence for an association between higher anxiety andenhancements in early or automatic biases to signalsof fear. Individuals with generalized anxiety show fasterinitial eye movements to fear-related facial expressions(Mogg, Millar, & Bradley, 2000), those with posttraumat-ic stress preferentially allocate their initial fixations totrauma-relevant words (Bryant et al., 1995), and traitanxiety in otherwise healthy subjects predicts amygdalaactivity during covert (nonconscious) perception of fear-ful faces (Etkin et al., 2004).
Lane and Schwartz (1987) have highlighted the impor-tance of considering levels of awareness in integratingcognitive and neuroscientific theories of psychopatholo-gy. Manipulation of awareness provides a means to elicitpreferentially automatic versus controlled processing offear stimuli, yet ERPs have not yet been used with sucha manipulation to examine the impact of trait anxietyrelative to depression on biases to these stimuli. In thisstudy, backward masking was used to define automatic(covert, nonconscious) fear processing, compared to un-masked (overt, conscious) processing. We expected co-vert fear to be distinguished by automatic neural biases(reflected in enhancements in early ERPs and thoseassociated with automatic orienting), and overt fear byneural biases in controlled and sustained evaluation(reflected in enhancements in later ERPs). Drawing onbehavioral findings, we expected higher trait anxiety tobe associated with an augmentation of early ERP biasesto covert fear. It was further expected that these biaseswould distinguish high anxiety not only from low anxietybut also from high trait depression, and its associationwith an exaggeration of later ERP biases to overt fear.
METHODS
Subjects
Subjects comprised 257 healthy individuals, recruited incollaboration with the Brain Resource International Data-base (www.brainresource.com; Gordon, Cooper, Rennie,Hermens, & Williams, 2005; Gordon, 2003a, 2003b). Strictand standardized exclusion criteria were history of mentalillness, brain injury, neurological disorder or other seriousmedical condition, and history of substance abuse. Mean
1596 Journal of Cognitive Neuroscience Volume 19, Number 10

age was 32.49 years (SD = 12.37 years) and sex was rela-tively evenly distributed (124 women, 133 men).
Participants provided written informed consent in ac-cordance with national health and medical research coun-cil guidelines.
These participants included a large subset (n = 156)of the total of 219 subjects who took part in a previousstudy (Williams, Palmer, et al., 2006) which focused onthe independent contrasts and source localization ofERPs to overt fear versus both happiness and neutral.
Psychometric Assessment
All subjects completed the Depression and AnxietyStress Scale (DASS; Lovibond & Lovibond, 1995b) toprovide a measure of trait anxiety and depression. TheDASS is based on a dimensional construct of depressionand anxiety. The DASS Anxiety scale correlates highlywith the Beck Anxiety Inventory and assesses featureswhich correspond to the clinical construct of anxietydisorder, whereas the Depression scale correlates highlywith the Beck Depression Inventory and relates to majordepressive illness (Lovibond & Lovibond, 1995a). Withinits dimensional structure, the DASS yields cutoff scoresto assist in the interpretation of scores within this clini-cal context. Participants in this study spanned the rangeof scores from ‘‘normal’’ to ‘‘severe’’ on each scale, butno individual scored in the extremely severe category.
Behavioral Task
Face stimuli for eight different individuals displayingboth neutral and evoked expressions of fear were se-lected from a standardized set of facial emotion stimuli(Gur et al., 2002). Facial expressions of fear are one ofthe most salient and universally recognized signals ofpotential threat (Ekman, Friesen, & Ellsworth, 1972). Weselected those stimuli rated independently as the mostaccurate representations of fear, and equated the stimuliin terms of size, gray-scale parameters, and central align-ment of the face within the image (with eyes as themidpoint reference). Stimuli were presented under bothcovert and overt conditions, and in each condition therewere eight blocks containing the eight different indi-viduals in pseudorandom order, making a total of 128stimuli. Half of the blocks were fear stimuli and half wereneutral, such that there were 64 stimuli per expression.
In the overt condition, stimuli were presented for500 msec, and it has been observed that even at thisrelatively short duration the subjective experience of theemotion may be elicited (Wild, Erb, & Bartels, 2001).The interstimulus interval was 700 msec, making a totalstimulus onset asynchrony of 1200 msec.
In the covert condition, stimuli were presented ina backward masking protocol. Target stimuli (fear orneutral faces) were presented for 10 msec, followed im-
mediately by a neutral face mask for 150 msec. The mask-ing stimulus was superimposed over the target stimulus,but spatially offset by 1 degree in one of the four diag-onals, randomly allocated. Target stimuli were synchro-nized with the refresh rate (100 Hz) of the stimuluspresentation computer which ensured that each stimu-lus was presented for 1 screen refresh (10 msec) onlyusing a customized stimulus presentation program. Theinterstimulus interval was 1040 msec, ensuring that thetotal stimulus onset asynchrony (1200 msec) was equiv-alent to that in the conscious condition.
Parameters used for covert presentation were verifiedin a series of psychophysical tasks (Williams, Liddell, et al.,2004). In these tasks, subjects indicated whether or nota face stimulus or a blank stimulus was presented priorto the immediate onset of the neutral face mask. Usingsignal detection criteria (Macmillan, 1986), detection ac-curacy was shown to be significantly above chance forstimulus durations of 30, 40, 50 msec, only marginallydifferent to chance at 20 msec but no different fromchance for 10 msec (Williams, Liddell, et al., 2004).
Conditions were presented in the order of covert fol-lowed by overt processing, following previous studies,to avoid the potentially confounding effects of supra-threshold perception on subsequent subthreshold per-ception (Williams, Liddell, et al., 2004; Bernat, Bunce,& Shevrin, 2001; Wong, Shevrin, & Williams, 1994). Sub-jects were instructed to attend to the faces in prepara-tion for posttesting briefings, thereby ensuring activeattention to the stimuli. Posttest behavioral assessmentshowed that participants were able to consistently dis-tinguish fear (83%) and neutral (75%) with well above-chance accuracy, indicating that any differential effectsin ERPs were unlikely to be due to visual processingdifficulties.
ERP Data Acquisition
Participants were seated in a sound- and light-attenuatedroom, with temperature controlled at 248C. They wereasked to refrain from smoking and caffeine for 2 hoursprior to testing, given that these factors can have anacute impact on ERPs. Data were acquired continuouslyat 500 Hz with skin resistance of <5 k! from 32 EEGchannels, with four EOG channels to allow for detec-tion of any eye movement artifacts using a Quikcap andNuAmps system according to the international 10–10electrode system. The focal sites of interest for this studywere those implicated in face processing, and estab-lished in a previous study (Williams, Palmer, et al., 2006):temporal (left, T5; right, T6), occipital (left, O1; right, O2),and medial fronto-central (Fz and Cz). Data were re-corded relative to the virtual ground, and referencedoff-line to linked mastoids. Eye movement correctionwas undertaken off-line using the procedure of Gratton,Coles, and Donchin (1983).
Williams et al. 1597

ERP Data Reduction and Analysis
Average ERPs were calculated for each emotion stimuli.Individual single-trial ERP epochs were filtered with a low-pass Tukey (cosine taper) filter function that attenuatedfrequencies above 25 Hz. Single trials were then averagedto form conventional ERPs, and peak components wereidentified within defined latency windows, validated byvisual inspection across individual subjects for each re-cording site. At the temporal, occipital, and midline re-gions of focal interest, the following components (andlatency windows) were identified: temporal (T5, T6) N80(50–120 msec), P120 (80–180 msec poststimulus), N170(120–220 msec), P230 (180–280 msec), and N250 (230–330 msec); occipital (O1, O2) N80 (50–120 msec), P120(80–180 msec), N170 (120–220 msec), P230 (180–280 msec), and N250 (230–330 msec). The correspondingmedial (Fz, Cz) components were the P80 (40–120 msec;concomitant of the N80), N120 (80–150 msec; concomi-tant of the P120), VPP (120–220 msec; concomitant of theN170), N200 (180–280 msec; concomitant of the P230),early P300 (230–330 msec; concomitant of the N250), andlate P300 (300–450 msec).
ERP components were scored using a baseline to peakmethod, such that peak amplitude and peak latency weredetermined. Outliers were defined as values beyond 2.5standard deviations from the mean, and formed only1.5% of data. These values were excluded and replacedwith age-appropriate (within 1 year) group means. Theexception to this procedure was the early P300, which,by definition, is elicited less frequently than other com-ponents (Squires, Squires, & Hillyard, 1975). In this study,the early P300 was elicited for approximately 50% ofstimuli, and missing values were therefore not replaced,resulting in lower degrees of freedom in focal analyses.
Repeated-measures multivariate analyses of variance(MANOVAs) within a general linear model were under-taken for the amplitude and latency of each ERP compo-nent. Within-subjects factors with two repeated measureseach were condition (overt vs. covert), emotion (fear vs.neutral), and site (Fz, Cz, or T5, T6 or O1, O2). Focal ef-
fects of interest were condition and emotion main effects,and in particular, the interaction of condition and emo-tion. Of secondary interest, we examined instances inwhich condition and/or emotion varied according tosite. The repeated-measures model included planned con-trast to elucidate each of these effects. Given the in-clusion of three regions, we used a corrected alpha levelof .017. We also reported effect size (using eta-squared;h2). The Greenhouse–Geisser epsilon method (relevantto repeated-measures models) was used to ensure thathomogeneity of variance assumptions was met.
Analysis of ERPs with Anxietyand Depression Groups
We formed groups of ‘‘high anxiety’’ and ‘‘high depres-sion,’’ defined by the top quartile of scores on the DASSAnxiety and Depression scales, respectively, followingprevious studies (Bar-Haim et al., 2005). ‘‘Low anxiety’’and ‘‘low depression’’ subjects were those who scoredzero on these respective scales, representing an absenceof negative mood. Table 1 provides the descriptive datafor combined subjects and each of these groups, rela-tive to the DASS categories of ‘‘normal’’ to ‘‘extremelysevere.’’ ANOVA confirmed that ‘‘high anxiety’’ differedsignificantly and substantially from ‘‘low anxiety’’ sub-jects on mean level of anxiety [F(1, 187) = 1599, p <.0001] and ‘‘high depression’’ differed significantly from‘‘low depression’’ subjects on level of depressed mood[F(1, 152) = 934, p < .0001].
Consistent with the comorbidity of anxiety and de-pression, the correlation between DASS depression andanxiety scores in this study was .43 ( p < .0001). Reflect-ing this association, the ‘‘high anxiety’’ and ‘‘high de-pression’’ groups also differed significantly on respectivelevels of depression [F(1, 187) = 31.54, p < .0001] andanxiety [F(1, 152) = 45.58, p < .0001]. Thus, analysis ofcovariance within a general linear model was used tocompare ‘‘high anxiety’’ to ‘‘low anxiety’’ and ‘‘high de-pression’’ to ‘‘low depression,’’ while controlling for
Table 1. Summary of Descriptive Data for Combined Subject Group, and ‘‘High Anxiety,’’ ‘‘High Depression,’’ ‘‘Low Anxiety,’’and ‘‘Low Depression’’ Groups on DASS Anxiety and Depression Scales
DASS Anxiety Scale DASS Depression Scale
Mean SD Range Mean SD Range
Combined subjects (n = 257) 1.94 2.79 0–18 3.88 4.74 0–24
‘‘High anxiety’’ (n = 31) 9.79 2.92 6–18 6.57 5.50 0–20
‘‘Low anxiety’’ (n = 129) 0 0 – 0.38 0.65 0–6
‘‘High depression’’ (n = 57) 3.89 1.80 0–10 11.40 3.68 8–24
‘‘Low depression’’ (n = 97) .45 0.90 0–4 0 0 –
Anxiety scale categories: Normal 0–7; Mild 8–9; Moderate 10–14; Severe 15–19; Extremely severe 20+.
Depression scale categories: Normal 0–9; Mild 10–13; Moderate 14–20; Severe 21–27; Extremely severe 28+.
1598 Journal of Cognitive Neuroscience Volume 19, Number 10

the respective level of depressed mood and anxiety. Todetermine the specificity of effects for fear, these analy-ses were undertaken for fear relative to a neutral base-line for both the overt and covert conditions for ERPcomponent amplitude and latency measures. Given thedifferent numbers in each group, Levene’s Test (relevantto ANOVA models) was applied to ensure that homo-geneity of variance assumptions was met. We also useda corrected alpha level (for 3 regions) of .017 in theseanalyses.
In addition, we undertook a parallel set of ANOVAswhich did not include a covariate, to provide complemen-tary information on the specificity of effects due to anxiety
versus depression, and as a check that covariance analy-sis was not introducing potentially misleading results.
RESULTS
The ERP waveforms elicited during conscious and non-conscious fear (versus) neutral perception are presentedin Figure 1.
Focal MANOVA effects of interest were main effectsfor emotion and condition, and their interaction, as wellas the three-way interactions between these factors andlaterality. These effects are outlined, in turn, in the
Figure 1. ERP waveformselicited in response toovert fear versus neutral(indicated by red and orange,respectively) and in responseto covert fear relative toneutral (indicated bydark blue and light blue,respectively), recorded overmedial fronto-central (Fz andCz), temporal (T5 and T6),and occipital (O1 and O2)sites. Sites indicated by oddnumbers are located in theleft side of each region, andthose indicated by evennumbers are located in theright side. ERP componentsof focal interest are indicatedfor each left-sided site, but theequivalent set of componentswere also scored for theright-sided sites. The y-axispresents microvolts and thex-axis, milliseconds.
Williams et al. 1599

sections below, and effects involving emotion are sum-marized in Table 2.
We first examined condition (covert vs. overt) and emo-tion (fear vs. neutral) main effects.
Main Effects for Condition
Condition effects reflecting enhanced processing in theovert relative to covert condition were revealed acrossregions within the first 150 msec: medial fronto-centralN120 (80–150 msec) [F(1, 256) = 92.72, p < .0001, h2 =.27] (Figure 1) and concomitant temporo-occipital P120[temporal, F(1, 256) = 75.13, p < .0001, h2 = .28; oc-cipital, F(1, 256) = 48.72, p < .0001, h2 = .16] (Figure1). The later P230 (180–280 msec) was also enhancedduring overt processing over the temporal region [F(1,256) = 107.82, p < .0001, h2 = .30].
On the other hand, condition effects representinggreater amplitude in the covert relative to overt conditionwere apparent for 40 msec and beyond. Over the occip-ital region, the N80 (40–120 msec) was enhanced dur-ing overt versus covert processing [F(1, 256) = 10.16,p = .002, h2 = .04]. The temporo-occipital N170 (120–220 msec) was also comparatively enhanced during co-vert processing [temporal, F(1, 256) = 41.46, p < .0001,h2 = .14; occipital, F(1, 256) = 118.75, p < .0001, h2 =.32], as was the concomitant medial fronto-central VPP(120–220 msec) [F(1, 256) = 231.13, p< .0001; h2 = .47].The subsequent temporo-occipital N250 (230–330 msec)[temporal, F(1, 256) = 112.54, p< .0001, h2 = .31; occip-
ital, F(1, 256) = 113.71, p < .0001; h2 = .31] and fronto-central late P300 (300–450 msec) [F(1, 256) = 155.27,p < .0001, h2 = .38] were similarly enhanced for covertrelative to overt processing (Figure 1).
Condition effects for ERP latency revealed significantlyfaster responses to covert processing (regardless of va-lence) across the time course; for the N80 [temporal,F(1, 256) = 9.57, p= .002, h2 = .04; occipital, F(1, 256) =44.89, p < .0001, h2 = .15] and concomitant fronto-central P80 [F(1, 256) = 127.60, p < .0001, h2 = .33],P120 [temporal, F(1, 256) = 144.22, p < .0001, h2 = .31;occipital, F(1, 256) = 182.92, p < .0001, h2 = .42] andconcomitant fronto-central N120 [F(1, 256) = 504.04,p < .0001, h2 = .66], N170 [temporal, F(1, 256) = 94.84,p < .0001, h2 = .27; occipital, F(1, 256) = 104.59, p <.0001, h2 = .29] and concomitant fronto-central VPP[F(1, 256) = 361.12, p < .0001, h2 = .59] and fronto-central early P300 [F(1, 134) = 11.15, p = .001, h2 =.08] and subsequent late P300 [F(1, 256) = 136.14, p <.0001, h2 = .35] (Figure 1).
Main Effects for Emotion
Enhanced responses to fear relative to neutral were re-flected in significant emotion effects for the fronto-centralVPP (120–220 msec) [F(1, 256) = 31.10, p < .0001; h2 =.11] (Table 2; Figures 1 and 2).
There was also a trend toward a main effect for emo-tion for the temporal N250 (230–350 msec) [F(1, 256) =4.53, p = .03, h2 = .02] (Table 2; Figures 1 and 2), and
Table 2. Summary of Focal MANOVA Results (df = 1,256)a for ERP Amplitude, Showing Interactions between Condition (Overtvs. Covert) and Emotion (Fear vs. Neutral) Shaded Blue and Red and Main Effects for Emotion Shaded Yellow and Green
40–100 msec 80–180 msec 120–220 msec 180–280 msec 230–330 msec 300–450 msec
N80/P80 N120/P120 N170/VPP N200/P230 N250/Early P300 Late P300
Overt Fear
Occipital
Temporal
Medial fronto-central
Covert Fear
Occipital
Temporal
Medial fronto-central
Where condition and emotion also interacted with laterality, the right or left side of greatest effect is indicated.
Interaction: Condition-specific increase to fearInteraction & Main effect: Common increase to fear, greater for conscious fearMain effect: Common increase to fear across conditionsInteraction: Condition-specific decrease to fearInteraction & Main effect: Common decrease to fear, greater for conscious fearMain effect: Common decrease to fear across conditions
aThere was a trend toward higher VPP responses to overt versus covert fear ( p= .05), in addition to the main effect for higher VPP responses to fearversus neutral across conditions.
1600 Journal of Cognitive Neuroscience Volume 19, Number 10

concomitant medial fronto-central early P300 [F(1,134) = 6.72, p = .01, h2 = .05], reflecting higher activityfor fear relative to neutral.
By contrast, the temporal P230 (180–280 msec) wasreduced for fear relative to neutral across both aware-ness conditions [F(1, 256) = 11.40, p = .001, h2 = .04](Table 2; Figures 1 and 2).
Condition by Emotion Interaction Effects dueto Modulation by Overt Processing of Fear
ERPs indexing structural encoding and subsequent sus-tained evaluation were particularly enhanced for overt(but not covert) fear, relative to neutral (Table 2; Figure 2).
There was a three-way interaction for the occipitalN170 (120–220 msec), indexing structural face encoding,which was due to an enhancement in responses to overt(but not covert) fear, particularly over the right relativeto left occipital region [F(1, 256) = 10.38, p= .001; h2 =.04] (Table 2; Figures 1 and 2). The corresponding tem-poral N170 (120–220 msec) showed similarly enhancedresponses to overt (but not covert) fear versus neutral[F(1, 256) = 6.78, p = .01; h2 = .03] (Table 2; Figures 1and 2).
There was a parallel trend toward a Condition byEmotion interaction for the medial VPP (120–220 msec)component of the N170/VPP complex associated withfacial emotion encoding [F(1, 256) = 3.86, p = .05; h2 =.02] (Table 2; Figures 1 and 2). Although the VPP wasenhanced for both overt and covert fear (relative toneutral), as indicated by the emotion main effects above,the enhancement was most pronounced in the overtcondition (Figure 1).
The subsequentmedial fronto-central early P300 (230–330 msec), reflecting automatic orienting, also showed aCondition by Emotion interaction [F(1, 134) = 7.02, p =.01; h2 = .05], as did the following fronto-central late
P300 (300–450 msec) [F(1, 256) = 5.98, p= .01; h2 = .04].These interactions were due to similarly greater activityfor overt (but not covert) fear versus neutral (Table 2;Figures 1 and 2).
The specific enhancement of temporal N170 and fronto-central VPP as well as early and late P300 responses toovert fear revealed by these interactions was observedagainst the background of generally reduced activityfor these components in the overt compared to covertcondition, reflected in the above condition main effects(Table 2; Figures 1 and 2).
By contrast, overt perception of fear was also associ-ated with lower ERP activity relative to neutral, for theP120 index of early perceptual processing. The occipitalP120 (80–180 msec) showed a three-way Condition byEmotion by Laterality interaction, due to particularly re-duced right occipital responses to overt (but not covert)fear [F(1, 256) = 9.37, p = .002; h2 = .04] (Table 2;Figures 1 and 2). This interaction was observed in thecontext of a generally enhanced occipital P120 for overtrelative to covert processing, as indicated by the abovecondition main effect.
Condition by Emotion Interaction Effects dueto Modulation by Covert Processing of Fear
In contrast to overt fear, covert fear processing was dis-tinguished by enhancements in early ERPs indexing ini-tial perceptual analysis and in ERPs reflecting automaticorienting.
There was a two-way Condition by Emotion interac-tion for the medial N120 (80–150 msec), due to en-hanced early responses to covert (but not overt) fear[F(1, 256) = 4.99, p = .01; h2 = .02] (Table 2; Figures 1and 2). This interaction was observed in the context ofcomparatively reduced activity for covert perception, asreflected in the above main effect.
Figure 2. Statisticalparameter maps illustrate thespatio-temporal distribution ofneural activity for the contrastof fearful relative to neutralface stimuli for covert andovert conditions. The timecourse of activity is indexedby ERP components N80/P80to the late P300, elicitedin the latency windows of40–120 msec to 300–450 msecpoststimulus. The legendshows the scale of significanteffects; regions colored inthe red spectrum indicatean increase in activity to fear(versus neutral) at an alphathreshold of at least .01, across medial, temporal, or occipital regions, whereas those colored in the blue spectrum indicate a reduction inactivity at an alpha threshold of at least .01 and green indicates no significant difference. Colors between green and red or blue indicatetrend level effects at an alpha level of between .05 and .01.
Williams et al. 1601

Additional Condition by Emotion interactions wereobserved for the N250 (230–350 msec) over both tem-poral [F(1, 256) = 17.18, p < .0001; h2 = .06] andoccipital regions [F(1, 256) = 16.97, p < .0001; h2 =.06] (Table 2; Figures 1 and 2). These interactions weredue to a greater enhancement for responses to covertthan overt fear versus neutral. Although there was atrend main effect for emotion for the temporal N250,the interaction confirms the particularly enhanced re-sponses for covert fear. In this case, these interactionswere present in the context of generally enhanced N250activity in the covert condition, as indicated by the abovecondition main effects.
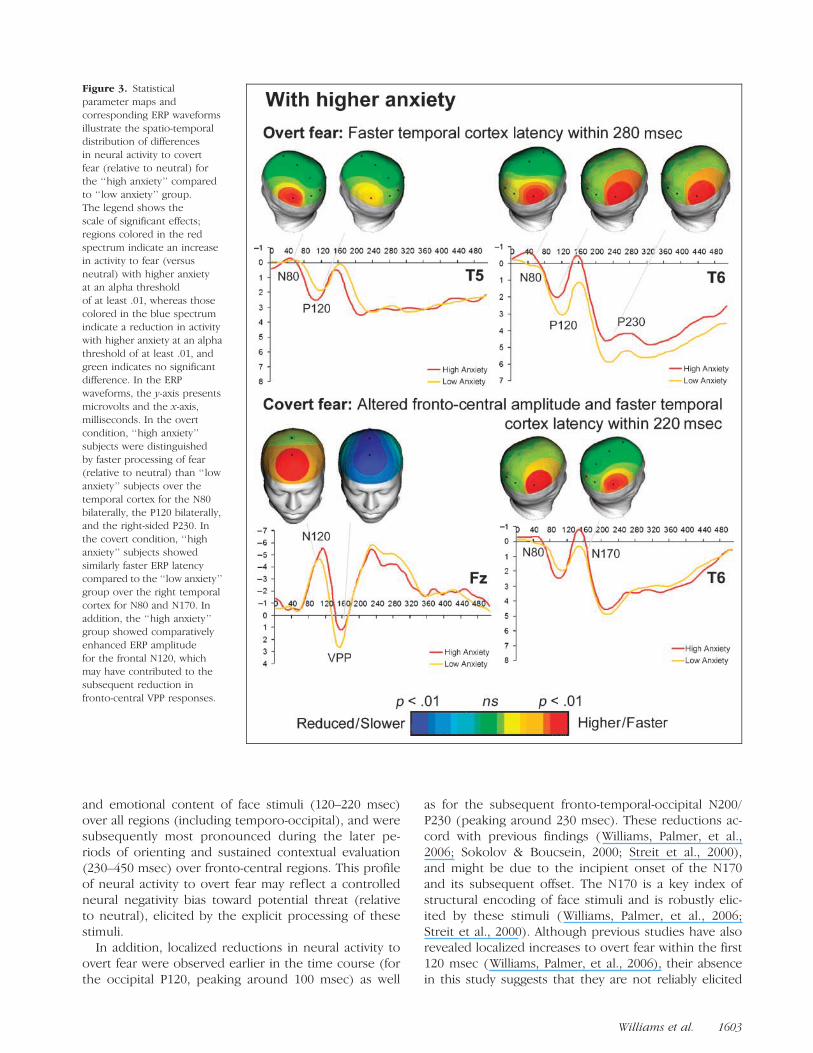
‘‘High Anxiety’’ versus ‘‘Low Anxiety’’ Groupsfor Overt and Covert Processing of Fear
ANCOVA showed that there was a distinctive profile ofneural activity, indexed by ERPs, in the ‘‘high anxiety’’compared to ‘‘low anxiety’’ group, across the time courseof overt and covert fear (relative to neutral) processing.
For overt processing, ‘‘high anxiety’’ subjects showedfaster processing of fear (relative to neutral) than ‘‘lowanxiety’’ subjects. A comparative speeding of ERP laten-cy in the ‘‘high anxiety’’ group was apparent over thetemporal cortex for the N80 bilaterally [left, F(1, 157) =4.30, p= .015, h2 = .04; right, F(1, 157) = 8.43, p= .004,h2 = .05], the P120 bilaterally [left, F(1, 157) = 6.49,p= .012, h2 = .04; right, F(1, 157) = 7.77, p= .006, h2 =.06], and the right-sided P230 [F(1, 157) = 6.25, p =.013, h2 = .04] (Figure 3).
In the covert processing condition, the ‘‘high anxiety’’group showed similarly faster ERP latency compared tothe ‘‘low anxiety’’ group over the right temporal cor-tex for N80 [F(1, 157) = 8.74, p = .004, h2 = .06] andN170 [F(1, 157) = 4.43, p= .013, h2 = .04] (Figure 3). Inaddition, the ‘‘high anxiety’’ group showed compara-tively enhanced ERP amplitude for the frontal N120 [F(1,157) = 4.48, p= .016, h2 = .03], which may have contrib-uted to the subsequent reduction in fronto-central VPPresponses [frontal, F(1, 157) = 4.73, p = .013, h2 = .04;central, F(1, 157) = 6.61, p = .008, h2 = .05] (Figure 3).
In these analyses, depression was a significant covar-iate ( p = .015) only for left temporal (T6) P230 latencyfor covert fear. However, the relationship between de-pression and P230 latency was in the opposite directionto that for anxiety, such that it did not account for theanxiety effect.1
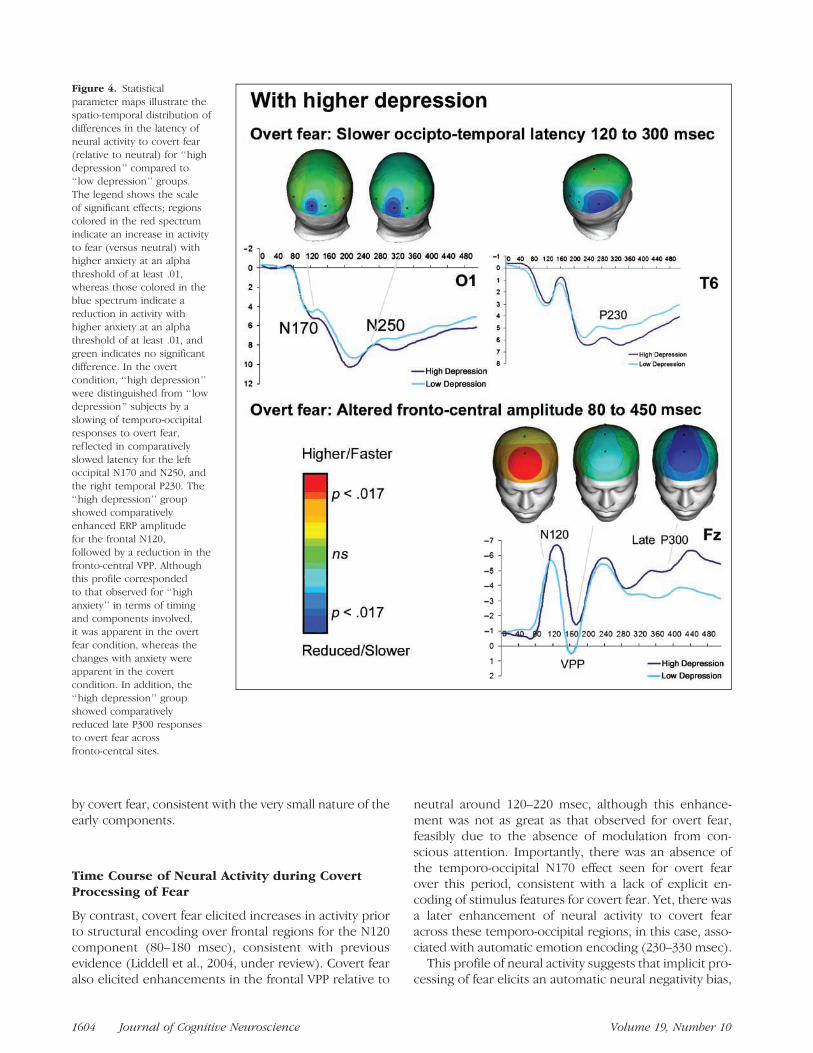
Comparison of ‘‘High Depression’’and ‘‘Low Depression’’ Groups
The specificity of the findings for high anxiety was fur-ther indicated by the findings for comparison of ‘‘highdepression’’ and ‘‘low depression’’ groups. Higher de-pression was found to impact overt processing of fear
(vs. neutral) only, and in an opposing direction to thatfor higher anxiety.
The ‘‘high depression’’ group was distinguished from‘‘low depression’’ by a slowing of temporo-occipital re-sponses to overt fear, reflected in comparatively slowedlatency for the left occipital N170 [F(1, 157) = 4.72, p =.015, h2 = .04] and N250 [F(1, 157) = 4.58, p = .013,h2 = .05] and the right temporal P230 [F(1, 157) = 6.82,p = .010, h2 = .05] (Figure 4).
The ‘‘highdepression’’ group showed comparatively en-hanced ERP amplitude for the frontal N120 [F(1, 157) =5.59, p = .014, h2 = .04], followed by a reduction in thefronto-central VPP [frontal, F(1, 157) = 5.48, p = .012,h2 = .05; central, [F(1, 157) = 5.30, p = .019, h2 = .04](Figure 4). Although this profile corresponded to thatobserved for ‘‘high anxiety’’ in terms of timing and com-ponents involved, it was apparent in the overt fearcondition, whereas the changes with anxiety were appar-ent in the covert condition. In addition, the ‘‘high depres-sion’’ group showed comparatively reduced late P300responses to overt fear across fronto-central regions[frontal, F(1, 157) = 10.88, p = .001, h2 = .07; central,F(1, 157) = 16.01, p < .0001, h2 = .07] (Figure 4).
Anxiety was not found to covary significantly withthese effects of ‘‘high’’ versus ‘‘low’’ depression.2
DISCUSSION
Consistent with the view that neural systems give prece-dence to signals of potential danger, facial signals of fearwere found to elicit increases in neural activity relativeto neutral across the processing time course. The timingand topographical distribution of these enhancementsdiffered with level of awareness. Overt fear elicited globalenhancements around the period of structural and emo-tion encoding (120–200 msec) followed by frontal en-hancements associated with orienting and sustainedcontext evaluation (230–450 msec), whereas covert fearelicited earlier frontal increases (80–180 msec) and a morelocalized later increase in temporo-occipital activity asso-ciated with automatic emotion encoding (230–330 msec).Within this framework, the impact of higher anxiety onneural processing of fear, and the specificity of this impactcompared to depressed mood, was examined. Higheranxiety was characterized by a speeding of neural activityearly in the time course, regardless of awareness, andan early enhancement of responses to covert fear. Thespecificity of this profile was indicated by a slowing andenhancement of responses with higher depression, ap-parent only during overt fear processing.
Time Course of Neural Activity during OvertProcessing of Fear
During overt fear processing, enhancements in neural ac-tivity to fear commenced with encoding of the structure
1602 Journal of Cognitive Neuroscience Volume 19, Number 10
and emotional content of face stimuli (120–220 msec)over all regions (including temporo-occipital), and weresubsequently most pronounced during the later pe-riods of orienting and sustained contextual evaluation(230–450 msec) over fronto-central regions. This profileof neural activity to overt fear may reflect a controlledneural negativity bias toward potential threat (relativeto neutral), elicited by the explicit processing of thesestimuli.
In addition, localized reductions in neural activity toovert fear were observed earlier in the time course (forthe occipital P120, peaking around 100 msec) as well
as for the subsequent fronto-temporal-occipital N200/P230 (peaking around 230 msec). These reductions ac-cord with previous findings (Williams, Palmer, et al.,2006; Sokolov & Boucsein, 2000; Streit et al., 2000),and might be due to the incipient onset of the N170and its subsequent offset. The N170 is a key index ofstructural encoding of face stimuli and is robustly elic-ited by these stimuli (Williams, Palmer, et al., 2006;Streit et al., 2000). Although previous studies have alsorevealed localized increases to overt fear within the first120 msec (Williams, Palmer, et al., 2006), their absencein this study suggests that they are not reliably elicited
Figure 3. Statisticalparameter maps andcorresponding ERP waveformsillustrate the spatio-temporaldistribution of differencesin neural activity to covertfear (relative to neutral) forthe ‘‘high anxiety’’ comparedto ‘‘low anxiety’’ group.The legend shows thescale of significant effects;regions colored in the redspectrum indicate an increasein activity to fear (versusneutral) with higher anxietyat an alpha thresholdof at least .01, whereas thosecolored in the blue spectrumindicate a reduction in activitywith higher anxiety at an alphathreshold of at least .01, andgreen indicates no significantdifference. In the ERPwaveforms, the y-axis presentsmicrovolts and the x-axis,milliseconds. In the overtcondition, ‘‘high anxiety’’subjects were distinguishedby faster processing of fear(relative to neutral) than ‘‘lowanxiety’’ subjects over thetemporal cortex for the N80bilaterally, the P120 bilaterally,and the right-sided P230. Inthe covert condition, ‘‘highanxiety’’ subjects showedsimilarly faster ERP latencycompared to the ‘‘low anxiety’’group over the right temporalcortex for N80 and N170. Inaddition, the ‘‘high anxiety’’group showed comparativelyenhanced ERP amplitudefor the frontal N120, whichmay have contributed to thesubsequent reduction infronto-central VPP responses.
Williams et al. 1603
by covert fear, consistent with the very small nature of theearly components.
Time Course of Neural Activity during CovertProcessing of Fear
By contrast, covert fear elicited increases in activity priorto structural encoding over frontal regions for the N120component (80–180 msec), consistent with previousevidence (Liddell et al., 2004, under review). Covert fearalso elicited enhancements in the frontal VPP relative to
neutral around 120–220 msec, although this enhance-ment was not as great as that observed for overt fear,feasibly due to the absence of modulation from con-scious attention. Importantly, there was an absence ofthe temporo-occipital N170 effect seen for overt fearover this period, consistent with a lack of explicit en-coding of stimulus features for covert fear. Yet, there wasa later enhancement of neural activity to covert fearacross these temporo-occipital regions, in this case, asso-ciated with automatic emotion encoding (230–330 msec).
This profile of neural activity suggests that implicit pro-cessing of fear elicits an automatic neural negativity bias,
Figure 4. Statisticalparameter maps illustrate thespatio-temporal distribution ofdifferences in the latency ofneural activity to covert fear(relative to neutral) for ‘‘highdepression’’ compared to‘‘low depression’’ groups.The legend shows the scaleof significant effects; regionscolored in the red spectrumindicate an increase in activityto fear (versus neutral) withhigher anxiety at an alphathreshold of at least .01,whereas those colored in theblue spectrum indicate areduction in activity withhigher anxiety at an alphathreshold of at least .01, andgreen indicates no significantdifference. In the overtcondition, ‘‘high depression’’were distinguished from ‘‘lowdepression’’ subjects by aslowing of temporo-occipitalresponses to overt fear,ref lected in comparativelyslowed latency for the leftoccipital N170 and N250, andthe right temporal P230. The‘‘high depression’’ groupshowed comparativelyenhanced ERP amplitudefor the frontal N120,followed by a reduction in thefronto-central VPP. Althoughthis profile correspondedto that observed for ‘‘highanxiety’’ in terms of timingand components involved,it was apparent in the overtfear condition, whereas thechanges with anxiety wereapparent in the covertcondition. In addition, the‘‘high depression’’ groupshowed comparativelyreduced late P300 responsesto overt fear acrossfronto-central sites.
1604 Journal of Cognitive Neuroscience Volume 19, Number 10

which arises earlier in the processing sequence thanfor explicit processing. The distinctive spatio-temporalpattern of neural responses to overt versus covert fearaccords with functional neuroimaging evidence that par-allel neural pathways support fear processing at differentlevels of awareness (Williams et al., 2006; Morris et al.,1999). Indeed, there were generally faster responsesacross the time course of covert compared to overt pro-cessing, consistent with a direct pathway for relayingsensory input in the absence of awareness. In contrastto the relatively substantial effect sizes (50–60%) for suchcondition main effects, the effects differentiating overtversus covert processing according to fear versus neutralwere subtle (around 5%), highlighting the value of largersubject numbers to elucidate such effects.
Neural Negativity Biases? Impact of HigherAnxiety on Neural Activity during Overtand Covert Processing of Fear
The distinct time course of neural activity to overt versuscovert fear provided a pertinent framework for under-standing the impact anxiety on this time course, and thespecificity of this impact compared to depressed mood.Moderate correlations between anxiety and depressionin this study accord with previous findings, yet it hasbeen noted that anxiety, nonetheless, represents a sep-arate construct from depression in factor analytic studiesof otherwise healthy individuals (Lovibond & Lovibond,1995a). Features of anxiety indexed by the DASS usedin this study reflect acute fear responses and their linkwith a relatively enduring state of anxiety. Depression,on the other hand, is thought to involve a more chronicand generalized sensitization of responses to potentialthreat (Lovibond & Lovibond, 1995a).
‘‘High anxiety’’ subjects according to the DASS weredistinguished by a speeding of neural responses overthe temporal cortex to both overt and covert fear (vs.neutral), relative to ‘‘low anxiety’’ subjects (Figure 3).For overt fear, this speeding of responses was apparentbilaterally from as early as 40 msec until 180 msec post-stimulus (for the N80 and P120 components), and wasadditionally apparent over the right temporal regionfor 230–330 msec (P230 component). During covert fearprocessing, the speeding of the N80 in the ‘‘high anx-iety’’ group was similarly apparent over the right tem-poral region, as was a speeding of the subsequent N170in the period 120–220 msec. In the covert fear condition,‘‘high anxiety’’ subjects were also distinguished by anexacerbation of the frontal enhancement (80–180 msec,N120 component) observed for covert fear processingin the previous section (Time Course of Neural Activityduring Covert Processing of Fear), which may indicateautomatic hypervigilance to threat in these subjects (Fig-ure 3). The exacerbation of this component may accountfor the subsequent attenuation of fronto-central VPP
responses (120–220 msec) in the ‘‘high anxiety’’ relativeto ‘‘low anxiety’’ group, suggesting that hypervigilancemay lead to deficits in emotion encoding.
Together, these findings point to an exaggerated neu-ral bias toward potential threat with higher anxiety,which operates automatically. Notably, the effect sizesof the results for ‘‘high’’ versus ‘‘low’’ anxiety were small,indicating that subclinical anxiety has a subtle impact onneural fear processing, which might be detected onlywith relatively large subject numbers. This new evidencefor a neural negativity bias in anxiety is compatible withprevious behavioral evidence of an excessive bias towardthreat that is present only under conditions of auto-matic processing (Bradley et al., 1995; Mogg et al.,1993, 1995). It also accords with functional neuroimagingevidence for enhanced temporal cortex amygdala activ-ity with higher trait anxiety during covert processing offear (Etkin et al., 2004), and for a hyperresponsive me-dial prefrontal cortex (including anterior cingulate) andamygdala during covert fear processing in the anxietydisorder of posttraumatic stress disorder (Bryant et al.,2007; Williams, Kemp, et al., 2006). Both behavioral andbrain function markers may reflect aspects of hyper-vigilance in anxiety, which occurs at a physiological levelthat is beyond conscious control. Without control, anongoing cycle of vigilance may contribute to the devel-opment and maintenance of the symptoms of anxietydisorder as well as interfere with the ability to effectivelyencode the emotional content of stimuli.
Specificity of the Impact of Higher Anxiety:A Comparison with the Impact of HigherDepression on Neural Activity duringOvert and Covert Processing of Fear
The specificity of these findings was indicated by thepresence of an opposing pattern of results with higherdepression, which were apparent only for overt fear pro-cessing. In ‘‘high depression’’ relative to ‘‘low depres-sion’’ subjects, neural responses were slowed (ratherthan speeded) over temporo-occipital regions. Slowedresponses to overt fear were apparent later in the timecourse, from structural encoding and beyond (120–330 msec). A pattern of enhanced early responses to fear(80–180 msec) over the frontal region, followed by adeficit in emotion encoding, was also apparent for ‘‘highdepression’’ subjects, but this pattern was present forovert fear processing, and not for covert processing as itwas for higher anxiety. Moreover, higher depression wasalso associated with a marked attention of the fronto-central late P300 to overt fear, suggesting additional def-icits in sustained context evaluation.
These findings provide evidence of a distinctive neuralnegativity bias in depression, which may prolong orient-ing to and evaluation of signals of potential threat whencontrolled processing is engaged. This proposal accords
Williams et al. 1605

with evidence for impaired orienting and impaired fron-tal P150 during working memory updating in a largeinvestigation of subclinical depression (Kemp et al.,2005). Moreover, models of depression based on func-tional neuroimaging evidence also highlight the role ofthe medial prefrontal regions in poor control of emotion(e.g., Mayberg, 1997). The findings are also consistentwith the suggestion from behavioral studies that depres-sion impacts the bias toward fear only when sustainedprocessing is engaged (Mathews et al., 1996; Bradleyet al., 1995; Mogg et al., 1993, 1995), as distinct from thepresence of alterations to covert fear processing withhigher anxiety.
Conclusion and Future Research
Findings from this study indicate that trait anxiety isdistinguished by a speeding of neural activity from theearliest phase of both overt and covert fear processing,in contrast to a more localized slowing of activity dur-ing overt fear processing, commencing later in the timecourse, for depression. In addition, trait anxiety wasfound to enhance initial orienting to covert fear, com-pared to an enhancement to overt fear for depression,which in both cases may disrupt subsequent emotionencoding. These findings accord with our integrativeneuroscience model of emotion processing, in whichthe mechanisms of anxiety involve an excessive auto-matic bias, associated with hypervigilance, to sources ofpotential fear, whereas depression reflects the chronicresult of hypervigilance in which a controlled evaluationof fear stimuli is protracted (Williams, 2006).
These findings also point to several areas of futureinvestigation. Because the present results refer to nor-mal variations in anxiety and depression, it would beimportant to determine whether these effects are pres-ent (or even more pronounced) in clinically diagnosedindividuals. A related issue is whether emotion-relatedneural activity is modulated in similar ways by state aswell as trait variations in mood. ERP studies using moodinduction suggest that state changes in mood bias fa-cial emotion processing within the first 500 msec inways that may be similar to those for trait aspects ofanxiety and depression (Gotlib, Yue, & Joormann, 2005;Esslen, Pascual-Marqui, Hell, Kochi, & Lehman, 2004;Joormann, 2004). A third issue is the question of wheth-er altered biases in anxiety generalize to other emotion-ally salient stimuli, given evidence for biases to angerand significant stimuli which are not explicitly threat-related in posttrauma anxiety conditions (Bryant et al.,2005; Felmingham, Bryant, & Gordon, 2003). Investiga-tion of the role of antidepressants in normalizing neuralnegativity biases is also warranted, given that seroto-nin modulates the neural time course of negative emo-tion processing (Kemp, Gray, Line, Silberstein, & Nathan,2003).
Reprint requests should be sent to Lea Williams, The Brain Dy-namics Centre, Acacia House, Westmead Hospital, Westmead,NSW, 2145, Australia, or via e-mail: [email protected].
Notes
1. The effects for higher anxiety were also largely confirmedin parallel ANOVAs without level of depression as a covariate,providing complementary information to indicate that varia-tion in levels of depression did not account for the effects in‘‘high’’ compared to ‘‘low’’ anxiety. In these analyses, signifi-cance was slightly higher in some cases (covert P120 latency atT5) and slightly lower in others (N80 latency at T5 for bothcovert and overt conditions), but in the remainder it remainedequivalent to ANCOVA results.2. Similarly, results from the ANOVAs (without anxiety as acovariate) paralleled those for the ANCOVAs, with only slightvariations in level of significance for two components (slightlymore significant for overt N250 latency at O1, slightly less forovert P230 latency at T6), and equivalent levels of significancefor the other components.
REFERENCES
Ashley, V., Vuilleumier, P., & Swick, D. (2004). Time courseand specificity of event-related potentials to emotionalexpressions. NeuroReport, 15, 211–216.
Bar-Haim, Y., Lamy, D., & Glickman, S. (2005). Attentionalbias in anxiety: A behavioral and ERP study. Brain andCognition, 59, 11–22.
Beck, A. T. (1976). Cognitive therapy and the emotionaldisorders. New York: Meridian.
Bernat, E., Bunce, S., & Shevrin, H. (2001). Event relatedpotentials differentiate positive and negative moodadjectives during both supraliminal and subliminal visualprocessing. International Journal of Psychophysiology,42, 11–34.
Bradley, B. P., Mogg, K., Millar, N., & White, J. (1995).Selective processing of negative information: Effects ofclinical anxiety, concurrent depression, and awareness.Journal of Abnormal Psychology, 104, 532–536.
Bryant, R. A., Felmingham, K. L., Kemp, A. H., Barton, M.,Peduto, A. S., Rennie, C., et al. (2005). Neural networksof information processing in posttraumatic stress disorder:A functional magnetic resonance imaging study. BiologicalPsychiatry, 58, 111–118.
Bryant, R. A., & Harvey, A. G. (1997). Attentional bias inposttraumatic stress disorder. Journal of TraumaticStress, 10, 635–644.
Bryant, R. A., Harvey, A. G., Gordon, E., & Barry, R. J. (1995).Eye movement and electrodermal responses to threatstimuli in post-traumatic stress disorder. InternationalJournal of Psychophysiology, 20, 209–213.
Bryant, R. A., Kemp, A. H., Felmingham, K. L., Liddell, B.,Olivieri, G., Peduto, A., et al. (2007). Enhanced amygdalaand medial prefrontal activation during nonconsciousprocessing of fear in posttraumatic stress disorder:An fMRI study. Human Brain Mapping (Early view.Published Online: 24 May 2007).
Cacioppo, J. T., Gardner, W. L., & Berntson, G. G. (1999).The affect system has parallel and integrative processingcomponents: Form follows function. Journal ofPersonality and Social Psychology, 76, 839–855.
Carretie, L., Mercado, F., Hinojosa, J. A., Jose, A.,Martin-Loeches, M., & Sotillo, M. (2004). Valence-related
1606 Journal of Cognitive Neuroscience Volume 19, Number 10

vigilance biases in anxiety studied through event-relatedpotentials. Journal of Affective Disorders, 78, 119–130.
Drevets, W. C. (2001). Neuroimaging and neuropathologicalstudies of depression: Implications for thecognitive-emotional features of mood disorders.Current Opinion in Neurobiology, 11, 240–249.
Ekman, P., Friesen, W. V., & Ellsworth, P. (1972). Emotionin the human face. New York: Pergamon.
Esslen, M., Pascual-Marqui, R. D., Hell, D., Kochi, K., &Lehman, D. (2004). Brain areas and time course ofemotional processing. Neuroimage, 21, 1189–1203.
Etkin, A., Klemenhagen, K. C., Dudman, J. T., Rogan, M. T.,Hen, R., Kandel, E. R., et al. (2004). Individual differencesin trait anxiety predict the response of the basolateralamygdala to unconsciously processed fearful faces.Neuron, 44, 1043–1055.
Felmingham, K., Bryant, R. A., & Gordon, E. (2003).Processing angry and neutral faces in post-traumatic stressdisorder: An event-related potentials study. NeuroReport,14, 777–780.
Gordon, E. (2003a). Integrative neuroscience and psychiatry.Neuropsychopharmacology, 28, S2–S8.
Gordon, E. (2003b). Integrative neuroscience in psychiatry:The role of a standardized database. AustralasianPsychiatry, 11, 156–163.
Gordon, E., Cooper, N., Rennie, C., Hermens, D., & Williams,L. M. (2005). Integrative neuroscience: The role of astandardised database. Clinical EEG and Neuroscience,36, 64–75.
Gotlib, I. H., & Cane, D. B. (1987). Construct accessibilityand clinical depression: A longitudinal investigation.Journal of Abnormal Psychology, 96, 199–204.
Gotlib, I. H., Yue, D. N., & Joormann, J. (2005). Selectiveattention in dysphoric individuals: The role of affectiveinterference and inhibition. Cognitive Therapy andResearch, 29, 417–432.
Gratton, G., Coles, M. G., & Donchin, E. (1983). A newmethod for off-line removal of ocular artifact.Electroencephalography and Clinical Neurophysiology,55, 468–484.
Gur, R. C., Erwin, R. J., Gur, R. E., Zwil, A. S., Heimberg, C.,& Kraemer, H. C. (1992). Facial emotion discrimination.II: Behavioral findings in depression. Psychiatry Research,42, 241–251.
Gur, R. C., Sara, R., Hagendoorn, M., Marom, O., Hughett, P.,Macy, L., et al. (2002). A method for obtaining 3-dimensionalfacial expressions and its standardization for use inneurocognitive studies. Journal of NeuroscienceMethods, 115, 137–143.
Han, S., He, H., & Woods, D. L. (2000). Hierarchical processingand level-repetition effect as indexed by early brainpotentials. Psychophysiology, 37, 817–830.
Hansen, C. H., & Hansen, R. D. (1988). Finding the facein the crowd: An anger superiority effect. Journal ofPersonality and Social Psychology, 54, 917–924.
Joormann, J. (2004). Attentional bias in dysphoria: The role ofinhibitory processes. Cognition and Emotion, 18, 125–147.
Kemp, A. H., Gray, M. A., Line, P., Silberstein, R. B., &Nathan, P. J. (2003). Preliminary electrophysiologicalevidence for modulation of the processing of negativeaffect by serotonin. Brain and Cognition, 51, 198–200.
Kemp, A. H., Stephan, B. C., Hopkinson, P., Sumich, A. L.,Paul, R. H., Clark, C. R., et al. (2005). Toward an integratedprofile of depression: Evidence from the brain resourceinternational database. Journal of Integrative Neuroscience,4, 95–106.
Lane, R. D., & Schwartz, G. E. (1987). Levels of emotionalawareness: A cognitive–developmental theory and its
application to psychopathology. American Journal ofPsychiatry, 144, 133–143.
Lenti, C., Giacobbe, A., & Pegna, C. (2000). Recognition ofemotional facial expressions in depressed children andadolescents. Perceptual and Motor Skills, 91, 227–236.
Liddell, B. J., Brown, K. J., Kemp, A. H., Barton, M. J., Das, P.,Peduto, A. S., et al. (2005). A direct brainstem–amygdala–cortical ‘‘alarm’’ system for subliminal signals of fear.Neuroimage, 24, 235–243.
Liddell, B. J., Palmer, D., Gordon, E., & Williams, L. M. (underreview). The complexity of nonconscious emotiondiscrimination: Concurrent evidence from ERPs and fMRI.
Liddell, B. J., Williams, L. M., Rathjen, J., Shevrin, H., & Gordon,E. (2004). A temporal dissociation of subliminal versussupraliminal fear perception: An event-related potentialstudy. Journal of Cognitive Neuroscience, 16, 479–486.
Lovibond, P. F., & Lovibond, S. H. (1995a). The structure ofnegative emotional states: Comparison of the DepressionAnxiety Stress Scales (DASS) with the Beck Depressionand Anxiety Inventories. Behavior Research and Therapy,33, 335–343.
Lovibond, S. H., & Lovibond, P. F. (1995b). Manual forthe Depression Anxiety Stress Scales (2nd ed.). Sydney:Psychology Foundation.
Macmillan, N. (1986). The psychophysics of subliminalperception. Behavioral and Brain Sciences, 9, 38–39.
Mangun, G. R., & Hillyard, S. A. (1995). Mechanismsand models of selective attention. In M. D. Rugg &M. G. H. Coles (Eds.), Electrophysiology of mind(pp. 40–85). Oxford: Oxford University Press.
Mathews, A., Ridgeway, V., & Williamson, D. A. (1996).Evidence for attention to threatening stimuli indepression. Behavior Research and Therapy, 34,695–705.
Mayberg, H. S. (1997): Limbic-cortical dysregulation:A proposed model of depression. Journal of Neuropsychiatryand Clinical Neuroscience, 9, 471–481.
Mogg, K., Bradley, B., & Williams, R. (1995). Attentionalbias in anxiety and depression: The role of awareness.British Journal of Clinical Psychology, 34, 17–36.
Mogg, K., & Bradley, B. P. (1998). A cognitive–motivationalanalysis of anxiety. Behavior Research and Therapy, 36,809–848.
Mogg, K., Bradley, B. P., Williams, R., & Mathews, A.(1993). Subliminal processing of emotional information inanxiety and depression. Journal of Abnormal Psychology,102, 304–311.
Mogg, K., Millar, N., & Bradley, B. P. (2000). Biases in eyemovements to threatening facial expressions in generalizedanxiety disorder and depressive disorder. Journal ofAbnormal Psychology, 109, 695–704.
Morris, J. S., Ohman, A., & Dolan, R. J. (1999). A subcorticalpathway to the right amygdala mediating ‘‘unseen’’ fear.Proceedings of the National Academy of Sciences, U.S.A.,96, 1680–1685.
Ohman, A. (1997). As fast as the blink of an eye: Evolutionarypreparedness for preattentive processing of threat. InP. J. Lang, M. Balaban, & R. F. Simons (Eds.), Attention andorienting: Sensory motivational processes (pp. 165–184).Mahwah, NJ: Erlbaum.
Pratto, F., & John, O. P. (1991). Automatic vigilance: Theattention-grabbing power of negative social information.Journal of Personality and Social Psychology, 61,380–391.
Segal, Z. V., Gemar, M., Truchon, C., Guirguis, M., &Horowitz, L. M. (1995). A priming methodology for studyingself-representation in major depressive disorder. Journalof Abnormal Psychology, 104, 205–213.
Williams et al. 1607

Sheline, Y. I., Barch, D. M., Donnelly, J. M., Ollinger, J. M.,Snyder, A. Z., & Mintun, M. A. (2001). Increased amygdalaresponse to masked emotional faces in depressed subjectsresolves with antidepressant treatment: An fMRI study.Biological Psychiatry, 50, 651–658.
Siegle, G. J., Steinhauer, S. R., Thase, M. E., Stenger, V. A., &Carter, C. S. (2002). Can’t shake that feeling: Event-relatedfMRI assessment of sustained amygdala activity in responseto emotional information in depressed individuals.Biological Psychiatry, 51, 693–707.
Smith, K., & Bryant, R. A. (2000). The generality of cognitivebias in acute stress disorder. Behavior Research andTherapy, 38, 709–715.
Sokolov, E. N., & Boucsein, W. (2000). A psychophysiologicalmodel of emotion space. Integrative Physiological andBehavioral Science, 35, 81–119.
Squires, N. K., Squires, K. C., & Hillyard, S. A. (1975). Two varietiesof long-latency positive waves evoked by unpredictableauditory stimuli in man. Electroencephalography andClinical Neurophysiology, 38, 387–401.
Streit, M., Wowler, W., Brinkmeyer, J., Ihl, R., & Gaebel, W.(2000). Electrophysiological correlates of emotional andstructural face processing in humans. NeuroscienceLetters, 278, 13–16.
Taylor, S. E. (1991). Asymmetrical effects of positive andnegative events: Mobilization-minimization hypothesis.Psychological Bulletin, 110, 67–85.
Warda, G., & Bryant, R. A. (1998). Cognitive bias in acute stressdisorder. Behavior Research and Therapy, 36, 1177–1183.
Whalen, P. J., Rauch, S. L., Etcoff, N. L., McInerney, S. C.,Lee, M. B., & Jenike, M. A. (1998). Masked presentationsof emotional facial expressions modulate amygdala activitywithout explicit knowledge. Journal of Neuroscience, 18,411–418.
Wild, B., Erb, M., & Bartels, M. (2001). Are emotionscontagious? Evoked emotions while viewing emotionallyexpressive faces: Quality, quantity, time course andgender differences. Psychiatry Research, 102, 109–124.
Williams, J. M. G., Mathews, A., & MacLeod, C. (1996). Theemotional Stroop task and psychopathology. PsychologicalBulletin, 120, 3–24.
Williams, J. M. G., Watts, F. N., MacLeod, C., & Mathews, A.(1997). Cognitive psychology and emotional disorders(2nd ed.). Chichester: Wiley.
Williams, L. M. (2006). An integrative neuroscience modelof ‘‘significance’’ processing. Journal of IntegrativeNeuroscience, 5, 1–47.
Williams, L. M., et al. (2006). An amygdala–prefrontaldissociation of subliminal and supraliminal fear. HumanBrain Mapping, 27, 652–661.
Williams, L. M., Brown, K. J., Das, P., Brammer, M. J.,Boucsein, B., Sokolov, E. N., et al. (2004). The dynamicsof cortico-amygdala and autonomic activity over theexperimental time course of fear perception. CognitiveBrain Research, 21, 114–123.
Williams, L. M., Kemp, A. H., Felmingham, K., Barton, M.,Olivieri, G., Peduto, A., et al. (2006). Trauma modulatesamygdala and medial prefrontal responses to consciouslyattended fear. Neuroimage, 29, 347–357.
Williams, L. M., Palmer, D., Liddell, B. J., Song, L., &Gordon, E. (2006). The ‘‘when’’ and ‘‘where’’ of perceivingsignals of threat versus non-threat. Neuroimage, 21,458–467.
Wong, P. S., Shevrin, H., & Williams, W. J. (1994).Conscious and nonconscious processes: An ERP indexof an anticipatory response in a conditioning paradigmusing visually masked stimuli. Psychophysiology, 35,87–101.
1608 Journal of Cognitive Neuroscience Volume 19, Number 10




















