
Natural antibodies of cod ( Gadus morhua L.): Specificity, activity and affinity
8
Natural antibodies of cod (Gadus morhua L.): Specificity, activity and affinity Bergljot Magnadottir ⁎, Sigridur Gudmundsdottir, Bjarnheidur K. Gudmundsdottir, Sigurdur Helgason Institute for Experimental Pathology, University of Iceland, Keldur v. Vesturlandsveg, IS-112 Reykjavik, Iceland abstract article info Article history: Received 1 April 2009 Received in revised form 2 July 2009 Accepted 9 July 2009 Available online 22 July 2009 Keywords: Natural antibodies Acquired antibodies Cod (Gadus morhua L.) IgM Natural antibodies are present in the serum of vertebrates regardless of antigenic stimulation. Characteristic activity is commonly detected against haptenated proteins, single stranded DNA and thyroglobulin. Natural antibodies are believed to provide an instant protection against pathogens of a broad specificity and to participate in homeostasis. Cod is a poor antibody responder but shows a relatively high level of natural antibodies against haptenated proteins. In this project the specificity, activity and affinity of natural antibodies was studied in different groups of cod and the effects of age/size, environmental temperature, immunisation and infection examined. Antigen driven selection of natural antibodies was also studied in one group of cod. The results demonstrated a broad and yet characteristic specificity, primarily directed against haptenated proteins and possible food antigens. The antibody activity increased with increasing age and at higher temperature whereas immunostimulation by immunisation or infection resulted in variable response. The affinity index of natural antibodies of cod generally did not correlate with changes in the antibody activity but it was in the same range as the affinity index of acquired cod antibodies and that of some mammalian monoclonal acquired antibodies. Analysis of antigen driven antibody selection showed that the natural antibody repertoire of individual cod was heterogeneous with respect to its affinity for haptenated protein. © 2009 Elsevier Inc. All rights reserved. 1. Introduction Natural antibodies are antibodies that are present in the serum of vertebrates without any apparent antigenic stimulation. Studies of autoimmune diseases and IgM-deficient mice have revealed certain common characteristics of natural antibodies in mammalian species (Casali and Schettino, 1996; Haury et al., 1997; Carroll and Prodeus, 1998; Boes, 2000): they are mostly of the IgM type, they are secreted by the long-lived, self-renewing B1 subset of B-cells, which are generated during foetal or neonatal development (Boes, 2000; Tung and Herzenberg, 2007), they are polyreactive and commonly show reactivity of low binding affinity with various autoantigens like thyroglobulin, single stranded DNA and heat shock protein but also with haptens like 2,4,6-Trinitrophenyl (TNP) or 2,4-Dinitrophenyl (DNP) (Casali and Schettino, 1996; Pashov et al., 2002). In mammals their primary role is to provide local confinement of infection, to enhance IgG response, to play a protective role in autoimmunity, take part in homeostasis and clearing of cell debris and are believed to form a link between innate and adaptive immunity (Carroll and Prodeus, 1998; Ochsenbein et al., 1999; Boes, 2000; Flajnik and Rumfelt, 2000; Mocci et al., 2000; Lutz, 2007). Natural antibodies have also received some attention in studies of fish immunology both in primitive species like sharks (Leslie and Clem, 1970; Rudikoff et al., 1970; Marchalonis et al., 1993, 1998) and sturgeons (Choudhury and Dick, 1994), and in more advanced teleost species like carp (Vilain et al., 1984; Kachamakova et al., 2006), goldfish (Sinyakov et al., 2002, 2006), snapper (Morrison et al., 2005) and trout (Gonzalez et al., 1989). Gonzalez et al. (1988) studied the natural antibody repertoire of phylogenetically distinct species and found that while the natural antibody of teleosts showed specificity primarily for haptenated protein (TNP-BSA) those of elasmobrachs and chondrosteans (stur- geon) showed specificity for various self and non-self antigens but a lower anti-TNP-BSA titre than the teleosts. Natural antibodies have been shown to take part in the immune defence of rainbow trout and goldfish against both viral and bacterial diseases (Gonzalez et al., 1989; Sinyakov et al., 2002). Sinyakov et al. (2006) showed that there was a negative correlation between the level of natural antibodies in goldfish and the level of acquired antibodies in vaccination trials suggesting that a high level of natural antibodies might inhibit specific antibody response. Teleost B-cells resemble mammalian B-1 cells, expressing mono- meric IgM on their surface, which, as in the mammalian system, acts as a marker for the B-lymphocyte population. However, there are some noticeable dissimilarities, for example, the splicing of the immunoglo- bulin receptor is different (Wilson et al.,1990; Bengten et al.,1991), the secreted IgM is tetrameric rather than pentameric (Pilstrom and Petersson, 1991; Hsu and Steiner, 1992) and commonly composed of different redox forms (Kaattari et al., 1998). Another important feature of the teleost B-cells is their phagocytic activity suggesting a broader Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316 ⁎ Corresponding author. Tel.: +354 5855100; fax: +354 5674714. E-mail address: [email protected] (B. Magnadottir). 1096-4959/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpb.2009.07.005 Contents lists available at ScienceDirect Comparative Biochemistry and Physiology, Part B journal homepage: www.elsevier.com/locate/cbpb
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Natural antibodies of cod ( Gadus morhua L.): Specificity, activity and affinity

Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
Contents lists available at ScienceDirect
Comparative Biochemistry and Physiology, Part B
j ourna l homepage: www.e lsev ie r.com/ locate /cbpb
Natural antibodies of cod (Gadus morhua L.): Specificity, activity and affinity
Bergljot Magnadottir ⁎, Sigridur Gudmundsdottir, Bjarnheidur K. Gudmundsdottir, Sigurdur HelgasonInstitute for Experimental Pathology, University of Iceland, Keldur v. Vesturlandsveg, IS-112 Reykjavik, Iceland
⁎ Corresponding author. Tel.: +354 5855100; fax: +3E-mail address: [email protected] (B. Magnadottir).
1096-4959/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.cbpb.2009.07.005
a b s t r a c t
a r t i c l e i n f oArticle history:Received 1 April 2009Received in revised form 2 July 2009Accepted 9 July 2009Available online 22 July 2009
Keywords:Natural antibodiesAcquired antibodiesCod (Gadus morhua L.)IgM
Natural antibodies are present in the serum of vertebrates regardless of antigenic stimulation. Characteristicactivity is commonly detected against haptenated proteins, single stranded DNA and thyroglobulin. Naturalantibodies are believed to provide an instant protection against pathogens of a broad specificity and toparticipate in homeostasis. Cod is a poor antibody responder but shows a relatively high level of naturalantibodies against haptenated proteins. In this project the specificity, activity and affinity of naturalantibodies was studied in different groups of cod and the effects of age/size, environmental temperature,immunisation and infection examined. Antigen driven selection of natural antibodies was also studied in onegroup of cod. The results demonstrated a broad and yet characteristic specificity, primarily directed againsthaptenated proteins and possible food antigens. The antibody activity increased with increasing age and athigher temperature whereas immunostimulation by immunisation or infection resulted in variable response.The affinity index of natural antibodies of cod generally did not correlate with changes in the antibodyactivity but it was in the same range as the affinity index of acquired cod antibodies and that of somemammalian monoclonal acquired antibodies. Analysis of antigen driven antibody selection showed that thenatural antibody repertoire of individual cod was heterogeneous with respect to its affinity for haptenatedprotein.
© 2009 Elsevier Inc. All rights reserved.
1. Introduction
Natural antibodies are antibodies that are present in the serum ofvertebrates without any apparent antigenic stimulation. Studies ofautoimmune diseases and IgM-deficient mice have revealed certaincommon characteristics of natural antibodies in mammalian species(Casali and Schettino, 1996; Haury et al., 1997; Carroll and Prodeus,1998; Boes, 2000): they are mostly of the IgM type, they are secretedby the long-lived, self-renewing B1 subset of B-cells, which aregenerated during foetal or neonatal development (Boes, 2000; Tungand Herzenberg, 2007), they are polyreactive and commonly showreactivity of low binding affinity with various autoantigens likethyroglobulin, single stranded DNA and heat shock protein but alsowith haptens like 2,4,6-Trinitrophenyl (TNP) or 2,4-Dinitrophenyl(DNP) (Casali and Schettino, 1996; Pashov et al., 2002). In mammalstheir primary role is to provide local confinement of infection, toenhance IgG response, to play a protective role in autoimmunity, takepart in homeostasis and clearing of cell debris and are believed to forma link between innate and adaptive immunity (Carroll and Prodeus,1998; Ochsenbein et al., 1999; Boes, 2000; Flajnik and Rumfelt, 2000;Mocci et al., 2000; Lutz, 2007).
Natural antibodies have also received some attention in studies offish immunology both in primitive species like sharks (Leslie and
54 567 4714.
l rights reserved.
Clem, 1970; Rudikoff et al., 1970; Marchalonis et al., 1993, 1998) andsturgeons (Choudhury and Dick, 1994), and in more advanced teleostspecies like carp (Vilain et al., 1984; Kachamakova et al., 2006),goldfish (Sinyakov et al., 2002, 2006), snapper (Morrison et al., 2005)and trout (Gonzalez et al., 1989).
Gonzalez et al. (1988) studied the natural antibody repertoire ofphylogenetically distinct species and found that while the naturalantibody of teleosts showed specificity primarily for haptenatedprotein (TNP-BSA) those of elasmobrachs and chondrosteans (stur-geon) showed specificity for various self and non-self antigens but alower anti-TNP-BSA titre than the teleosts.
Natural antibodies have been shown to take part in the immunedefence of rainbow trout and goldfish against both viral and bacterialdiseases (Gonzalez et al., 1989; Sinyakov et al., 2002). Sinyakov et al.(2006) showed that there was a negative correlation between thelevel of natural antibodies in goldfish and the level of acquiredantibodies in vaccination trials suggesting that a high level of naturalantibodies might inhibit specific antibody response.
Teleost B-cells resemble mammalian B-1 cells, expressing mono-meric IgM on their surface, which, as in themammalian system, acts asa marker for the B-lymphocyte population. However, there are somenoticeable dissimilarities, for example, the splicing of the immunoglo-bulin receptor is different (Wilson et al.,1990; Bengten et al.,1991), thesecreted IgM is tetrameric rather than pentameric (Pilstrom andPetersson, 1991; Hsu and Steiner, 1992) and commonly composed ofdifferent redox forms (Kaattari et al., 1998). Another important featureof the teleost B-cells is their phagocytic activity suggesting a broader

310 B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
functional activity than that of mammalian B-cells (Li et al., 2006;Randelli et al., 2008). Using a panel of monoclonal antibodies againstIgM and other cell markers, different populations of B-like lympho-cytes have been demonstrated in some fish species (Rombout et al.,1993; Zwollo et al., 2008). Whether these variables include analoguesof mammalian B cell subsets has not been examined. Variable B-cellpopulation in cod has also been demonstrated at our laboratory withthe help of monoclonal anti-IgM antibodies (unpublished data).
A variety of research projects have been carried out at our institutein recent years, dealing with different aspects of cod's immune systemand immune response. The general conclusion has been that cod reliesmore on the quantity than the specificity or quality of its immuneresponse when fighting infection. A case in hand is the antibodyresponse of cod. Immunization and challenge experiments of cod havegenerally resulted in poor acquired antibody response (Espelid et al.,1991; Pilstrom and Petersson, 1991; Schroder et al., 1992; Magnadottiret al., 2001; Pilstrom et al., 2005; Lund et al., 2006). However, theimmunoglobulin (IgM) concentration in serum is relatively high (2–20 mg mL−1) compared to many other fish species like the salmonidsand can be as much as 50–70% of the serum proteins of individual fish(Magnadottir, 1998). Similarly, the activity of natural antibodies, withcharacteristic anti-TNP-BSA activity, is commonly high (Magnadottiret al., 1999a,b, 2001, 2002; Gudmundsdottir et al., 2009).
In this paper a closer look was taken at the specificity, activity andaffinity of the natural antibodies in different groups of cod.
2. Materials and methods
2.1. Cod serum
The serum samples used came from 5 groups of cod (Gadusmorhua; Gadidae) that are listed in Table 1. Group 1 and 2were of wildorigin, whereas groups 3–5 originated from cultured cod reared fromwild gametes fertilized at the Marine Experimental Station, Grindavik,Iceland. All sera were stored at −20 °C or −80 °C. Details of eachgroup can be ascertained from the relevant publications listed inTable 1. Further information about groups 3, 4 and 5 are necessary:
The fish in group 3was immunised three times, inweek 2, 28 and 72,and blood samples collected 10 times from day 0 to week 76, at aninterval of approximately 8 weeks (see insert in Fig. 4a). This was alongitudinal study, the same individuals being sampled throughoutthe duration of the experiment. All three sub-groups, whetherimmunised intra-peritoneal (i.p.) with Freund's complete adjuvant(FCA) alone or TNP-LPH or LPH in FCA showed similar antibodyresponse against TNP-LPH (or other TNP-haptenated proteins), butno activity against the protein (LPH) alone. The conclusion was thatno acquired antibodies were produced but that the immunisation
Table 1The groups of cod used in the present work.
Groups of cod Reference
Group 1: Wild cod of different sizes (ages) sampled off the coast ofIceland during fall
Magnadottir et al.(1999b)
Group 2: Cultured cod, of wild origin, maintained at 1 or 14 °C for12 months
Magnadottir et al.(1999a)
Group 3: Cultured cod immunized three times haptenated (TNP)or non-haptenated Limulus polyphemus hemocyanin (LPH) inFreund’s complete adjuvant or adjuvant alone and sampled overa period of 18 months
Magnadottir et al.(2001)
Group 4: Cultured cod experimentally infected with Aeromonassalmonicida ssp. achromogenes (Asa)
Magnadottir et al.(2002)
Group 5: Cultured cod vaccinated against and challenged withAsa
Unpublished
induced natural antibody response against haptenated proteins(Magnadottir et al., 2001).The fish in group 4 were infected by i.p. or intra muscular (i.m.)inoculation of 6.5×104 or 6.5×105 colony forming units of Aero-monas salmonicida ssp. achromogenes (Asa, strain T-223/9), whichcauses atypical furunculosis in cod, the uninfected cod wereinjected with PBS (Magnadottir et al., 2002).
The fish in group 5 came from a recent vaccination and challengeexperiment at our Institute, not yet published. Briefly: cod werevaccinated against Asa (strain F19/99). An experimental vaccinecontaining formalin killed bacteria and inactivated extra-cellularproducts prepared at our laboratory, was mixed with mineral oiladjuvant, kindly supplied by Dr. Kjersti Gravningen at PHARMAQAS,Norway. The fishwere vaccinated by injection of 0.2 mL i.p. on day 0.Thirteen weeks later, the fish were challenged by bathing for 3 h inseawater containing live bacteria (2×106 CFU mL−1). Two weekslater, when accumulated death was lower than expected in thecontrol group receiving PBS, it was decided to challenge the fishagain, this timewith i.m. injection (5×104 CFUper fish). Four weekslater, accumulated deaths in controls were still unacceptable.However, the bacterium was isolated from 82% of fish dying duringthe challenge period and from 14% of the survivors at the end of theexperiment. Serum samples were collected before vaccination (onday 0), before challenge (at 13 weeks) and from survivors 6 weeksafter the challenges (at 19 weeks).
2.2. Antigens used in dot blot analysis
2.2.1. Non-self antigensAfter some preliminary testing of various protein and carbohydrate
antigens the following seven substances were chosen for furthertesting: Trinitrophenyl haptenated bovine serum albumin (TNP-BSA)prepared at our laboratory (2 mg mL−1) according to Pilstrom andPetersson (1991) and containing 21 TNP residues per BSA molecule,single stranded DNA (ssDNA, Sigma-Aldrich, USA, 0.7 mg mL−1),chitosan (kindly provided by Genis, Iceland, 20 mg mL−1), thyroglo-bulin (Sigma, 20 mg mL−1), lipopolysaccharide from Asa (Asa-LPS,1.6 mgmL−1) prepared at our laboratory using a method described byWestphal and Jann (1965) and Koga et al. (1985), Limulus polyphemushemocyanin (LPH) (Sigma, 2 mg mL−1) and polymannuronic acid(PolyM) from the brown seaweed Durvillea antarctica (Biomar, AS,Norway, Vollstad, kindly provided by Dr. Roy A. Dalmo, Department ofBiotechnology, University of Tromsø, Tromsø, Norway, 20 mg mL−1).The concentration used was the maximum concentration that gave noor limited background reaction with the secondary and conjugatedantibodies used in the dot blot analysis.
2.2.2. Self-antigensProtein solutions were prepared from the following 10 organ tissues
froma100 g cod: gills, heart, liver, spleen, kidney, brain, stomach, pyloriccaecum, and anterior and posterior sections of the gut. A small piece ofeach organ, packed in aluminium foil, was quickly frozen in liquidnitrogen and then stored at −80 °C. A protein solution was thenprepared from each organ as previously described (Magnadottir et al.,2004). Briefly, theorgansamplewaspressed througha stainless steel teastrainer with 1 mL lysis buffer, 50 mM Tris–HCl, pH 7.6, 0.3 M NaCl, 2 %Triton X-100 (Sigma). Protease inhibitor cocktail (Sigma) was added(50 µL mL−1). The suspension was frozen at −80 °C, thawed andcentrifuged at 15,000 g for 5 min. The supernatantwas collected and theprotein content estimated using a kit from Pierce (USA) based on theBradford (1976) method. The protein concentration was adjusted to2 mg mL−1 for each organ sample.

311B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
2.3. Dot blot analysis of the specificity of the natural antibodies
One µL of antigen solution was dotted onto nitrocellulose (NC)membranes (Hybond ECL NC membrane, Amersham PharmaciaBiotech, U.K.), allowed to dry and residual sites blocked in 0.5%semi-skimmed milk powder in 0.1 M Tris buffered saline, pH 7.8containing 0.1% Tween20 (TBS-T), for 1 h at room temperature. Afterwashing in TBS-T, the membrane was incubated in test serum, diluted4×10−3 in TBS-Tovernight at 4 °C or for 1 h at room temperature. Thiswas followed by an incubation with polyclonal anti-cod IgM mouseantibody (prepared at our laboratory) and alkaline phosphataseconjugated goat anti-mouse Ig antibodies (Dako, Denmark), diluted inthe blocking solution, both for 1 h at room temperaturewith extensivewashing between each step. The blot was developed in a substratebuffer of 100 µg mL−1 NBT (nitro blue tetrazolium), 60 µg mL−1 BCIP(5-bromo-4-chloro-3-indonylphosphate) and 4 mM MgCl2 in 0.1 Methanolamine–HCl buffer, pH 9.6. Cod IgM was used as a positivecontrol and other controls included the incubation of blots with TBS-Tin place of cod serum and with TBS-T in place of both cod serum andthe anti-cod IgM antibody.
2.4. Standard ELISA to measure antibody activity
The antibody activity was measured using an indirect ELISA test(Magnadottir et al., 1999a). Briefly, microtrays (MaxiSorp, Nunc,Denmark) were coated with the antigen, diluted in coating buffer(0.05 M carbonate buffer, pH 9.6, Sigma) overnight at 4 °C. Residual siteswere blocked in 0.1% semi-skimmed milk powder in coating buffer for1 h at room temperature and the tray then incubated with cod serumdiluted 10−2 in 0.1 M phosphate buffered saline (PBS), pH 7.2, 0.1%Tween 20 (PBS-T), overnight at 4 °C. Incubations with primary anti-codIgM antibody followed by alkaline phosphatase (AP) conjugated goatanti-mouse Ig antibody (Dako),was for 1 hat 37 °Candcolourdevelopedwith 1 mg mL−1 p-nitrophenyl phosphate (Sigma) in 0.1 M ethanola-mine buffer, 1 mM MaCl2, pH 9.6 and reaction stopped after 30 min atroom temperature with 3 M NaOH. Optical density (OD) was read at405 nm. Extensive washing in PBS-T was between each step.
Natural antibody activity was tested on trays coated with 5 µgmL−1
TNP-BSA and acquired antibody activity of cod immunised or infectedwith Asa was tested on trays coated with 10 µg protein mL−1 ofsonicated whole bacterial suspension (prepared at our laboratory).
All serum samples used in this work were titrated and the criteriafor positive acquired antibody activity was set at exceeding the doublevalue of the blank (normally ODN0.2). For convenience the antibodyactivity was expressed as the OD value of the serum diluted 10−2
rather than as titre since a good correlationwas observed between thetitre obtained and OD values.
2.5. Thiocyanate (SCN)-elution ELISA to measure antibody affinity andaffinity distribution
The antibody affinity i.e. the strength of the antibody–antigenbonding, was measured using a method based on thiocyanate(chaotropic agent) elution (Pullen et al., 1986; Lew et al., 1988;Macdonald et al., 1988; Chapman et al., 2006). Microtrays were coatedwith the relevant antigen and residual sites blocked as describedabove. The cod serum (6 sera tray−1), previously titrated and dilutedto give optical density of 0.8–1.5 on the ascending slope of the titrationcurve, were placed in two columns for each serum, 50 µL well−1, andincubated overnight at 4 °C. After washing in PBS-T, 0.0, 0.05, 0.1, 0.5,1.0, 2.0, 4.0 and 6.0 M ammonium thiocyanate (NH4SCN) in 0.1 Mphosphate buffer, pH 6.0, 50 µL well−1, were added to each of theeight rows and incubated for 15 min at room temperature. The traywas washed and the ELISA test continued, incubating with mouseanti-cod IgM antibody, AP-conjugated goat anti-mouse IgG antibodyand substrate as described above. Themolarity (M) of NH4SCN, which
resulted in 50% elution compared to the buffer control, was defined asthe affinity index.
To test whether the affinity varied depending on the antigenconcentration (affinity distribution) trays were coated with 5 twofoldserial dilutions of the antigen (5.0, 2.5, 1.25, 0.62 and 0.31 µg mL−1
TNP-BSA), one dilution for every 2 columns, two columns were keptfor buffer controls (blanks), coating 3 wells with each antigenconcentration. The thiocyanate elution ELISA was carried out asdescribed above testing one serum sample per tray. The affinity indexwas then calculated for each antigen concentration. Only sera withrelatively high anti-TNP-BSA activity could be tested this way.
2.6. Statistical analysis
The StatView™ analysis system (Abacus Concepts, USA) was usedfor statistical analysis. Unpaired Student's t-test was used forcomparative analysis, the criteria for significance being set atPb0.05 in all instances.
3. Results
3.1. The specificity of natural antibodies of cod
Sera from the various groups of cod were tested in dot blot with 7non-self antigens and 10 self-antigens. Fig. 1a shows the bindingspecificity of representative sera from six individuals with 7 non-selfantigens. Most sera showed relatively strong specificity for TNP-BSA,chitosan and polymannuronic acid. The reaction with LPH varied andthe reaction with ssDNA and thyroglobulin was generally weak orabsent. Sera from wild and cultivate cod, males and females, old andyoung cod gave very similar specificity. The anti-cod IgM antibodyshowed a weak reaction with TNP-BSA and polymannuronic acid.
Fig. 1b shows the binding specificity of four representative serawith the 10 self-antigens. The anti-cod IgM antibody on its own(omitting the serum) showed some reactionwith all the self-antigensindicating the presence of IgM, humoral or a B-cell receptor, in allorgans tested. About half of all the cod sera tested (7 out of 15)showed specific binding with the stomach tissue antigens.
3.2. The activity and affinity of natural antibodies (anti-TNP-BSA) indifferent groups of cod
3.2.1. The effect of cod's age on natural antibody activity and affinitySerum samples from 1–2 year old cod, weighing 500–1000 g, and
from7–8 year old cod,weighing3500–6500 g,were analysed (group 1).Fig. 2a shows the antibody activity and Fig. 2b the antibody affinity(mean and standard errors (S.E.)) of the two age groups.
The natural antibody activity of the younger fish varied from 0.4 to1.7 (mean value 0.78) and of the older cod from 0.9 to 2.4 (mean value1.55). This increase in natural antibody activity with increasing agewas statistically significant (P=0.0003).
The antibody affinity index values of the younger cod varied from1.5 to 2.6 (mean value 2.16) and of the older cod from 1.6 to 3.7 (meanvalue 2.45). This difference in affinity between the age groups was notstatistically significant (P=0.579).
3.2.2. The effect of environmental temperature on natural antibodyactivity and affinity
Serum samples from cod kept at 1 °C or 14 °C for 12 months wereanalysed (group 2). The mean weight of the fish kept at 1 °C was1048 g and of fish kept at 14 °C 2260 g. Fig. 3a shows the antibodyactivity and Fig. 3b the antibody affinity (mean and S.E.) of the twotemperature groups.
In fish kept at 1 °C the natural antibody activity varied from 0.76 to2.03 (mean value 1.21) and in fish kept at 14 °C from 1.25 to 2.6 (mean

Fig. 1. Dot blots showing the specificity of cod serum antibodies with non-self and self antigens. a) Six cod sera (strips 1 – 6) tested with seven non-self antigens as shown. On strip 7serumwas omitted and on strip 8 both serum and anti-cod IgM were omitted. b) Four cod sera (strips 1 – 4) tested with 10 self antigens. On strip 5 serumwas omitted. Cod IgM (b1µl) was used as a positive control.
312 B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
value 1.73). This increase in natural antibody activity with increasingtemperature was statistically significant (P=0.012).
The antibody affinity index of cod kept at 1 °C varied from 1.7 to 3.0(mean value 2.2) and of cod kept at 14 °C from 0.8 to 2.7 (mean value
Fig. 2. The natural antibody activity and affinity of cod serum collected at different ages(group 1). The mean value and standard error (S.E.) are shown, a) shows the antibodyactivity of 10 sera from the young cod and 14 sera from the old cod, b) shows theantibody affinity of 4 sera from the young cod and 5 sera from the old cod. Asterisksindicate a significant difference of Pb0.05.
2.1). This difference in affinity between the two temperature groupswas not statistically significant (P=0.837).
3.2.3. The effect of immunisation on natural antibody activity and affinityThe serum samples analysed were collected from the same six
individuals before immunisation (pre-immunisation) and 40 weekslater or 12 weeks after the second immunisation (post-immunisation),when maximum natural antibody levels were observed (group 3, see
Fig. 3. The natural antibody activity and affinity of serum collected from cod reared at1°C or 14°C for 12 months (group 2). The mean value and S.E. are shown, a) shows theantibody activity of 6 sera from each temperature group, b) shows the antibody af-finity of 6 sera from each temperature group. Asterisks indicate a significant differenceof Pb0.05.

Fig. 4. The natural antibody activity and affinity of serum collected from cod before andafter immunisation (group 3). The mean value and S.E. are shown, a) shows theantibody activity of 6 sera from each time point, b) shows the antibody affinity of 6 serafrom each time point. Asterisks indicate a significant difference of Pb0.05. The insert inFig. 4 a) shows themean natural antibody activity of all fish over the period of 76 weeks,arrows showing the times of immunisation and the shaded squares shows the samplescollected before immunization (1) and post-immunization (2) tested in the presentanalysis.
313B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
insert in Fig. 4a). Themeanmass of the cod at the start of the experimentwas 250 g and this had approximately doubled after 50 weeks(Magnadottir et al., 2001). Fig. 4a shows the natural antibody activityand Fig. 4b the natural antibody affinity (mean and S.E.) before and afterimmunisation. The insert in Fig. 4a shows the mean natural antibodyactivity of all experimental fish over the period of 76 weeks.
The natural antibody activity before immunisation varied from0.27 to 0.59 (mean value 0.41) and after immunisation the activityvaried from 0.47 to 1.11 (mean value 0.86). This increase in antibodyactivity following immunisationwas statistically significant (P=0.001).
The antibody affinity index of cod before immunisation variedfrom 1.60 to 2.05 (mean value 1.77) and after immunisation from 1.2to 1.9 (mean value 1.52). There was no difference in antibody affinity(P=0.171).
Fig. 5. The natural antibody activity and affinity of serum collected from uninfected andinfected cod (group 4). The mean value and S.E. are shown, a) shows the antibodyactivity of seven sera from uninfected and infected cod, b) shows the antibody affinity offour sera from uninfected and infected cod. Asterisks indicate a significant difference ofPb0.05.
3.2.4. The effects of infection on natural antibody activityThe serum samples analysed were collected from uninfected and
infected cod 3 weeks after challengewithAsa (group 4). Themeanmassof the codwas about 42 g. Fig. 5a shows the antibody activity and Fig. 5bthe antibody affinity (mean and S.E.) of uninfected and infected cod.
The natural antibody activity of the uninfected fish varied from 0.96to 2.08 (mean value 1.50) and of the infected fish from 0.98 to 2.75(mean value 2.13). This difference in antibody activity was significant(P=0.0159).
The antibody affinity index of the uninfected fish varied from 1.35to 2.0 (mean value 1.75) and did not differ from that of the infectedfish from 1.45 to 1.8 (mean value 1.66) (P=0.604).
3.3. The acquired and natural antibody activity and affinity of vaccinatedand challenged cod
3.3.1. Natural and acquired antibody activitySerum samples collected at three time points during the vaccina-
tion and challenge trial (group 5) were analysed for natural antibodyactivity against TNP-BSA and for acquired antibody activity against thebacterial (Asa) antigen. The fish weighed about 34 g at the start of theexperiment and between 70 and 90 g at the end of the experiment.Fig. 6a shows the natural antibody activity and Fig. 6b shows theacquired antibody activity (mean and S.E.) at the three samplingpoints.
The natural antibody activity (Fig. 6a) was fairly high at allsampling times regardless of whether the fish was immunised withthe Asa vaccine or PBS. Themean activity was about 0.62 at the start ofthe experiment , 13 weeks after vaccination the mean activity was0.82 and 0.92 in the PBS and the Asa vaccinated fish respectively andfollowing challenge the natural antibody activity was 0.68 and 0.56 inthe PBS and the Asa vaccinated fish respectively. None of thesechanges in natural antibody activity between PBS or Asa vaccinatedfish or between different time points were statistically significant.
At the start of the experiment no antibody activity against Asa wasobserved, the mean value being 0.138 (Fig. 6b). The anti-Asa antibodyactivity remained low in the PBS vaccinated fish, the mean values being0.166 and 0.234 13 weeks after vaccination and 6 weeks after challengerespectively. These changes were not statistically significant.
Acquired anti-Asa antibody activity was observed in the Asavaccinated fish 13 weeks after vaccination and remained high 6 weeks
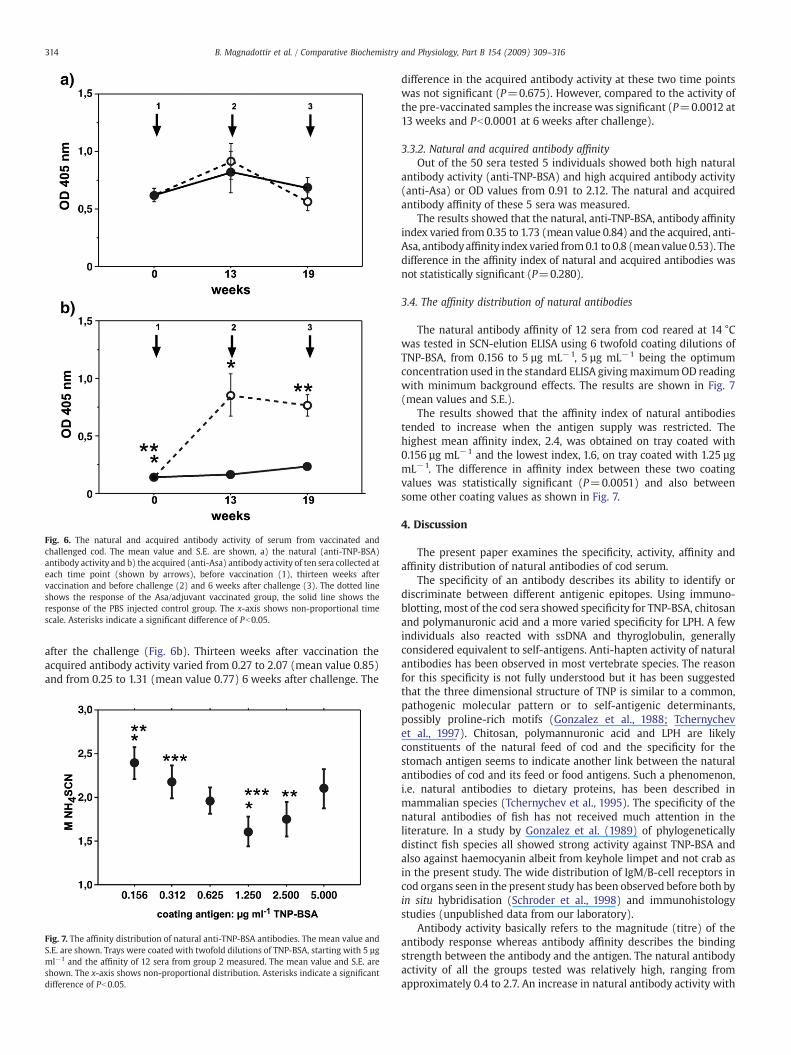
Fig. 6. The natural and acquired antibody activity of serum from vaccinated andchallenged cod. The mean value and S.E. are shown, a) the natural (anti-TNP-BSA)antibody activity and b) the acquired (anti-Asa) antibody activity of ten sera collected ateach time point (shown by arrows), before vaccination (1), thirteen weeks aftervaccination and before challenge (2) and 6 weeks after challenge (3). The dotted lineshows the response of the Asa/adjuvant vaccinated group, the solid line shows theresponse of the PBS injected control group. The x-axis shows non-proportional timescale. Asterisks indicate a significant difference of Pb0.05.
314 B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
after the challenge (Fig. 6b). Thirteen weeks after vaccination theacquired antibody activity varied from 0.27 to 2.07 (mean value 0.85)and from 0.25 to 1.31 (mean value 0.77) 6 weeks after challenge. The
Fig. 7. The affinity distribution of natural anti-TNP-BSA antibodies. The mean value andS.E. are shown. Trays were coated with twofold dilutions of TNP-BSA, starting with 5 µgml−1 and the affinity of 12 sera from group 2 measured. The mean value and S.E. areshown. The x-axis shows non-proportional distribution. Asterisks indicate a significantdifference of Pb0.05.
difference in the acquired antibody activity at these two time pointswas not significant (P=0.675). However, compared to the activity ofthe pre-vaccinated samples the increase was significant (P=0.0012 at13 weeks and Pb0.0001 at 6 weeks after challenge).
3.3.2. Natural and acquired antibody affinityOut of the 50 sera tested 5 individuals showed both high natural
antibody activity (anti-TNP-BSA) and high acquired antibody activity(anti-Asa) or OD values from 0.91 to 2.12. The natural and acquiredantibody affinity of these 5 sera was measured.
The results showed that the natural, anti-TNP-BSA, antibody affinityindex varied from 0.35 to 1.73 (mean value 0.84) and the acquired, anti-Asa, antibodyaffinity index varied from0.1 to 0.8 (meanvalue 0.53). Thedifference in the affinity index of natural and acquired antibodies wasnot statistically significant (P=0.280).
3.4. The affinity distribution of natural antibodies
The natural antibody affinity of 12 sera from cod reared at 14 °Cwas tested in SCN-elution ELISA using 6 twofold coating dilutions ofTNP-BSA, from 0.156 to 5 µg mL−1, 5 µg mL−1 being the optimumconcentration used in the standard ELISA givingmaximumOD readingwith minimum background effects. The results are shown in Fig. 7(mean values and S.E.).
The results showed that the affinity index of natural antibodiestended to increase when the antigen supply was restricted. Thehighest mean affinity index, 2.4, was obtained on tray coated with0.156 µg mL−1 and the lowest index, 1.6, on tray coated with 1.25 µgmL−1. The difference in affinity index between these two coatingvalues was statistically significant (P=0.0051) and also betweensome other coating values as shown in Fig. 7.
4. Discussion
The present paper examines the specificity, activity, affinity andaffinity distribution of natural antibodies of cod serum.
The specificity of an antibody describes its ability to identify ordiscriminate between different antigenic epitopes. Using immuno-blotting, most of the cod sera showed specificity for TNP-BSA, chitosanand polymanuronic acid and a more varied specificity for LPH. A fewindividuals also reacted with ssDNA and thyroglobulin, generallyconsidered equivalent to self-antigens. Anti-hapten activity of naturalantibodies has been observed in most vertebrate species. The reasonfor this specificity is not fully understood but it has been suggestedthat the three dimensional structure of TNP is similar to a common,pathogenic molecular pattern or to self-antigenic determinants,possibly proline-rich motifs (Gonzalez et al., 1988; Tchernychevet al., 1997). Chitosan, polymannuronic acid and LPH are likelyconstituents of the natural feed of cod and the specificity for thestomach antigen seems to indicate another link between the naturalantibodies of cod and its feed or food antigens. Such a phenomenon,i.e. natural antibodies to dietary proteins, has been described inmammalian species (Tchernychev et al., 1995). The specificity of thenatural antibodies of fish has not received much attention in theliterature. In a study by Gonzalez et al. (1989) of phylogeneticallydistinct fish species all showed strong activity against TNP-BSA andalso against haemocyanin albeit from keyhole limpet and not crab asin the present study. The wide distribution of IgM/B-cell receptors incod organs seen in the present study has been observed before both byin situ hybridisation (Schroder et al., 1998) and immunohistologystudies (unpublished data from our laboratory).
Antibody activity basically refers to the magnitude (titre) of theantibody response whereas antibody affinity describes the bindingstrength between the antibody and the antigen. The natural antibodyactivity of all the groups tested was relatively high, ranging fromapproximately 0.4 to 2.7. An increase in natural antibody activity with

315B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
increasing age/size was demonstrated in groups 1 and 2 but relativelyhigh activity values were also seen in groups of smaller (30–40 g) cod,like groups 4 and 5, indicating that other factors apart from size couldbe influencing the normal level of natural antibody activity in cod.
The present analysis emphasised the variable antibody response ofcod depending on both the nature and route of the antigenic stimuliand on the individual cod. While natural antibody activity wasenhanced following immunisation with protein in FCA or FCA alone(group 3) and following infection by i.p. or i.m. inoculation of livebacteria (Asa, group 4) no acquired antibody responsewas detected inthese groups. On the other hand vaccination with experimental Asavaccine in mineral oil (group 5) and subsequent challenge, whilehaving little or no effect on the natural antibody response, resulted inrelatively good acquired antibody response against Asa. As in group 4,no acquired antibody response was seen in the control fish (PBSinjected) 6 weeks after, in this case, infection by both bath treatmentand i.m. inoculation.
Other studies of the immune response of cod have also revealedsimilar variability in cod's response to antigenic stimuli depending onfactors like the bacterial strain used, cod's age and on differencesbetween individual cod (Schroder et al., 2006, 2009; Lund et al., 2007).In a recent vaccination and challenge trial against Vibrio anguillarumboth natural and acquired antibody response was stimulated byvaccination but in both instances stimulation was basically limited to afew but not the same, individuals (Gudmundsdottir et al., 2009).
In the present study thiocyanate elution was used to measureantibody affinity (Pullen et al., 1986; Lew et al., 1988; Macdonald et al.,1988). Thiocyanate is a chaotropic agent that disrupts intra-molecular,non-covalent forces. Themethodwas found to be a relatively simple androbust method for this purpose. Its drawback was, however, that theserum (antibody) samples had to show a linear range of optical densitybetween approximately 0.8 and 1.6 when titrated in ELISA. Hence, serahaving lower OD values had to be omitted from the analysis. For thisreason the affinity data was commonly derived from relatively fewsamples and the statistical significance was therefore limited.
The natural antibody affinity index of the different groups variedfrom 0.35 to 3.7, the mean value of all sera tested being about 1.7.Using thiocyanate elution to determine the affinity of monoclonal(mouse) anti-DNP antibodies Macdonald et al. (1988) obtained anaffinity index varying from 0.2 to 8.0 (average 4.5) for the 6 antibodiestested. Based on this limited data, the affinity of the natural antibodiesof cod appears to be in a comparable range as the affinity of thesemammalian acquired antibodies.
Only 5 sera (from group 5) were available that allowedmeaningfulanalysis of acquired anti-Asa antibody affinity index. The affinity ofthese sera varied from 0.1 to 0.8 with a mean value of 0.53, which islow compared to the range of affinity values determined for thenatural antibodies of cod. Although the difference between the affinityindex of natural and acquired antibodies was not statisticallysignificant when tested on the same set of serum samples it seemsthat the generally assumed view of natural antibodies being of lowaffinity compared to acquired antibodies (Casali and Schettino, 1996;Baumgarth et al., 2005) does not apply to cod.
When the affinity analysis was carried out using variable antigenconcentration in the solid phase it was found that the highestantibody affinity index was obtained when the antigen concentrationwas restricted. This is a phenomenon known as an antigen drivenantibody selection (Kaattari et al., 2002; Bruderer et al., 1992) andshows that the natural antibody repertoire of individual cod isheterogeneous with respect to its affinity for TNP-BSA. However, theaffinity index was not influenced by changes in antibody activity. Forexample, increased antibody activity associated with immunisationand infection did not affect the affinity index. This suggests that thephenomenon of affinity maturation as described by Kaattari et al.(2002) during the acquired antibody response of rainbow trout doesnot apply to the natural antibody response of cod.
The present results have demonstrated that the natural antibodies ofcod have a broad and yet characteristic specificity, primarily directedagainst haptenated proteins and possible food antigens. Such character-istic specificity within a species is well known and reflects the inherentlegacy of that species related to its genetic make-up and environmentalor pathogenic history through evolution (Gonzalez et al., 1988;Avrameas, 1991; Casali and Schettino, 1996; Baumgarth et al., 2005).
The present results have also demonstrated that the naturalantibodies of cod are heterogenous and dynamic immune parameteraffected by both intrinsic and external factors. In addition the naturalantibody affinity is relatively strong and possibly stronger than theaffinity of acquired cod antibodies.
Natural antibodies are commonly described in the literature asbackground antibodies, polyreactive andwith low affinity, which basicfunction is innate pattern recognition, complement activation and toform a link between the innate and adaptive immune system (Boes2000; Flajnik and Rumfelt, 2000). The present findings indicate thatthe natural antibodies of cod, in addition to the functions listed above,may play an important role in the immune defence of cod. Naturalantibodies of cod may thus compensate for the commonly variableand often poor acquired antibody response of cod and warrant furtherstudies especially at the cellular level.
Acknowledgements
Thanks are due to the staff at the Marine Institute's ExperimentalStation at Stadur, Grindavik and at Sandgerdi Research Centre whoprovidedmost of the cod and experimental facilities. Gudmundur LogiNorddahl, Berglind Gisladottir and Ivar Orn Arnason are thanked fortheir technical assistance. Various funding organisations supportedthe original experiments: the Nordic Council of Ministers, Copenha-gen, the Icelandic Research Council, Reykjavik and the Research Fundof the University of Iceland. Special thanks are also due to the IcelandicStudent Innovation Fund for their more recent support.
References
Avrameas, S., 1991. Natural autoantibodies: from “horror autotoxicus” to “gnothi seauton”.Immunol. Today 12, 154–159.
Baumgarth, N., Tung, J.W., Herzenberg, A., 2005. Inherent specificities in naturalantibodies: a key to immune defence against pathogen invasion. Springer Semin.Immunopathol. 26, 347–362.
Bengten, E., Leanderson, T., Pilstrom, L., 1991. Immunoglobulin heavy chain cDNAs fromthe teleost Atlantic cod (Gadus morhua L.): nucleotide sequences of secretory andmembrane form show an unusual splicing pattern. Eur. J. Immunol. 21, 3027–3033.
Boes, M., 2000. Role of natural and immune IgM antibodies in immune responses. Mol.Immunol. 37, 1141–1149.
Bradford, M.M., 1976. A rapid and sensitive method for the quantification of microgramquantities of proteins using the principle of protein–dye binding. Anal. Biochem. 72,248–254.
Bruderer, U., Deusinger, M., Schurch, U., Lang, A.B., 1992. Analyses of affinity distributionswithin polyclonal population of antigen-specific antibodies. Evaluation of theiraccuracy in population detection using monoclonal antibodies. J. Immunol. Methods151, 157–164.
Carroll, M.C., Prodeus, A.P., 1998. Linkages of innate and adaptive immunity. Curr. Opin.Immunol. 10, 36–40.
Casali, P., Schettino, E.M., 1996. Structure and function of natural antibodies. Curr. Top.Microbiol. Immunol. 210, 167–179.
Chapman, M.D., Keir, G., Petzold, A., Thompson, E.J., 2006. Measurement of high affinityantibodies on antigen-immunoblots. J. Immunol. Methods 310, 62–66.
Choudhury, A., Dick, T.A., 1994. Natural anti-phosphorylcholine (PC) antibodies in lakesturgeon, Acipenser fulvescens Rafinesque, 1817 (Chondrostei: Acipenseridae). FishShellfish Immunol. 4, 399–401.
Espelid, S., Rodseth, O.M., Jorgensen, T.O., 1991. Vaccination experiments and studies ofthe humoral immune responses in cod, Gadus morhua L., to four strains ofmonoclonal-defined Vibrio anguillarum. J. Fish Dis. 14, 185–197.
Flajnik, M.F., Rumfelt, L.L., 2000. Early and natural antibodies in non-mammalianvertebrates. Curr. Top. Microbiol. Immunol. 252, 233–240.
Gonzalez, R., Charlemagne, J., Mahana, W., Avrameas, S., 1988. Specificity of naturalserum antibodies present in phylogenetically distinct fish species. Immunology 63,31–36.
Gonzalez, R., Matsiota, P., Torchy, C., De Kinkelin, P., Avrameas, S., 1989. Natural anti-TNPantibodies from rainbow trout interfere with viral infection in vitro. Res. Immunol.140, 675–684.

316 B. Magnadottir et al. / Comparative Biochemistry and Physiology, Part B 154 (2009) 309–316
Gudmundsdottir, S., Magnadottir, B., Bjornsdottir, B., Arnadottir, H., Gudmundsdottir, B.K.,2009. Specific and natural antibody response of cod juveniles vaccinated againstVibrioanguillarum. Fish Shellfish Immunol. 26, 619–624.
Haury, M., Sundblad, A., Grandien, A., Barreau, C., Coutinho, A., Nobrega, A., 1997. Therepertoire of serum IgM in normal mice is largely independent of external antigeniccontact. Eur. J. Immunol. 27, 1557–1563.
Hsu, E., Steiner, L.A., 1992. Primary structure of immunoglobulin through evolutions.Curr. Opin. Struct. Biol. 2, 422–431.
Kaattari, S.L., Evans, D., Klemer, J., 1998. Varied redox forms of teleost IgM: an alternativeto isotypic diversity? Immunol. Rev. 166, 133–142.
Kaattari, S.L., Zhang, H.I., Khor, I.W., Kaattari, I.M., Shapiro, D.A., 2002. Affinitymaturation in trout: clonal dominance of high affinity antibodies late in theimmune response. Dev. Comp. Immunol. 26, 191–200.
Kachamakova, N.M., Irnazarow, I., Parmentier, H.K., Savelkoul, H.F.J., Pilarczyk, A.,Wiegertjes, G.F., 2006. Genetic differences in natural antibody levels in commoncarp (Cyprinus carpio L.). Fish Shellfish Immunol. 21, 404–413.
Koga, T., Nishihara, T., Fujiwara, T., Nisiawa, T., Okahashi, N., Noguchi, T., Hamada, S.,1985. Biochemical and immunobiological properties of lipopolysaccharide (LPS)from Bacteroides gingivalis and comparison with LPS from Escherichia coli. Infect.Immun. 47, 638–647.
Leslie, G.A., Clem, L.W., 1970. Reactivity of normal shark immunoglobulins withnitrophenyl ligands. J. Immunol. 105, 1547–1552.
Lew, A.M., Anders, R.F., Edwards, S.J., Langford, C.J., 1988. Comparison of antibody avidityand titre elicited by peptide as a protein conjugate or as expressed in vaccinia.Immunology 65, 311–314.
Li, J., Barreda, D.R., Zhang, Y.-A., Boshra, H., Gelman, A.E., LaPatra, S., Tort, L., Sunyer, J.O.,2006. B lymphocytes from early vertebrates have potent phagocytic andmicrobicidal abilities. Nat. Immunol. 7, 1116–1124.
Lund, V., Bordal, S., Kjellsen, O., Mikkelsen, H., Schroder, M.B., 2006. Comparison ofantibody responses in Atlantic cod (Gadus morhua L.) to Aeromonas salmonicida andVibrio anguillarum. Dev. Comp. Immunol. 30, 1145–1155.
Lund, V., Bordal, S., Schroder, M.B., 2007. Specificity and durability of antibody responsein Atlantic cod (Gadus morhu L.) immunised with Vibrio anguillarum O2b. FishShellfish Immunol. 23, 906–910.
Lutz, H.U., 2007. Homeostatic roles of naturally occurring antibodies: an overview.J. Autoimmun. 29, 287–294.
Macdonald, R.A., Hosking, C.S., Jones, C.L., 1988. The measurement of relative antibodyaffinity by ELISA using thiocyanate elution. J. Immunol. Methods 106, 191–194.
Magnadottir, B., 1998. Comparison of immunoglobulin (IgM) from four fish species. Icel.Agr. Sci. 12, 47–59.
Magnadottir, B., Jonsdottir, H., Helgason, S., Bjornsson, B., Jorgensen, T., Pilstrom, L.,1999a. Humoral immune parameters in Atlantic cod (Gadus morhua L.) I: the effectsof environmental temperature. Comp. Biochem. Physiol. B 122, 173–180.
Magnadottir, B., Jonsdottir, H., Helgason, S., Bjornsson, B., Jorgensen, T., Pilstrom, L., 1999b.Humoral immune parameters in Atlantic cod (Gadus morhua L.) II: the effects of sizeand gender under different environmental conditions. Comp. Biochem. Physiol. B 122,181–188.
Magnadottir, B., Jonsdottir, H., Helgason, S., Bjornsson, B., Solem, S.T., Pilstrom, L., 2001.Immune parameters of immunized cod (Gadus morhua L.). Fish Shellfish Immunol.11, 75–89.
Magnadottir, B., Bambir, S.H., Gudmundsdottir, B.K., Pilstrom, L., Helgason, S., 2002.Atypical Aeromonas salmonicida infection in naturally and experimentally infectedcod, Gadus morhua L. J. Fish Dis. 25, 583–597.
Magnadottir, B., Lange, S., Steinarsson, A., Gudmundsdottir, S., 2004. The ontogenicdevelopment of innate immune parameters of cod (Gadus morhua L.). Comp.Biochem. Physiol. B 139, 217–224.
Marchalonis, J.J., Hohman, V.S., Thomas, C., Schluter, S.F., 1993. Antibody production insharks and humans: a role for natural antibodies. Dev. Comp. Immunol. 17, 41–53.
Marchalonis, J.J., Schluter, S.F., Bernstein, R.M., Hohman, V.S., 1998. Antibodies of shark:revolution and evolution. Immunol. Rev. 166, 103–122.
Mocci, S., Lafferty, K., Howard, M., 2000. The role of autoantigens in autoimmunedisease. Curr. Opin. Immunol. 12, 725–730.
Morrison, R.N., Lyons, A.B., Nowak, B.F., Hayball, J.D., 2005. Assessment of snapper(Pagrus auratus) natural IgM binding to bromelain treated sheep erythrocytes. FishShellfish Immunol. 18, 91–99.
Ochsenbein, A.F., Fehr, T., Lutz, C., Suter, M., Brombacher, F., Hengartner, H., Zinkernagel,R.M., 1999. Control of early viral and bacterial distribution and disease by naturalantibodies. Science 286, 2156–2159.
Pashov, A., Kenderov, A., Kyurkchiev, S., Kehayov, I., Hristova, S., Lacroix-Desmazes, S.,Giltiay, N., Varamballi, S., Kazatchkine, M.D., Kaveri, S.V., 2002. Autoantibodies to heatchock protein 90 in the human natural antibody repertoire. Int. Immunol.14, 453–461.
Pilstrom, L., Petersson, A., 1991. Isolation and partial characterization of immunoglo-bulin from cod (Gadus morhua L.). Dev. Comp. Immunol. 15, 143–152.
Pilstrom, L., Warr, G.W., Stromberg, S., 2005. Why is the antibody response of Atlanticcod so poor? The search for a genetic explanation. Fish. Sci. 71, 961–971.
Pullen, G.R., Fitzgerald, M.G., Hosking, C.S., 1986. Antibody avidity determination byELISA using thiocyanate elution. J. Immunol. Methods 86, 83–87.
Randelli, E., Buonocore, F., Scapigliati, G., 2008. Cell markers and determinants in fishimmunology. Fish Shellfish Immunol. 25, 326–340.
Rombout, J.H.W.M., Taverne-Thiele, A.J., Villena, M., 1993. The gut-associated lymphoidtissue (GALT) of carp (Cyprinus carpio L.): an immunocytochemical analysis. Dev.Comp. Immunol. 17, 55–66.
Rudikoff, S., Voss, E.W., Sigel, M.M., 1970. Biological and chemical properties of naturalantibodies in the nurse shark. J. Immunol. 105, 1344–1352.
Schroder, M.B., Espelid, S., Jorgensen, T.O., 1992. Two serotypes of Vibrio salmonicidaisolated from diseased cod (Gadus morhua); virulence, immunological studies andvaccination experiments. Fish Shellfish Immunol. 2, 211–221.
Schroder, M.B., Flano, E., Pilstrom, L., Jorgensen, T.O., 1998. Localisation of Ig heavy chainmRNA positive cells in Atlantic cod (Gadus morhua L.) tissues; identified by in situhybridisation. Fish Shellfish Immunol. 8, 565–576.
Schroder, M.B., Mikkelsen, H., Bordal, S., Gravningen, K., Lund, V., 2006. Earlyvaccination and protection of Atlantic cod (Gadus morhua L.) juveniles againstclassical vibrosis. Aquaculture 254, 46–53.
Schroder, M.B., Ellingsen, T., Mikkelsen, H., Norderhus, E.A., Lund, V., 2009. Comparison ofantibody responses inAtlantic cod (Gadusmorhua L.) toVibrio anguillarum,Aeromonassalmonicida and Francisella sp. Fish Shellfish Immunol. 27, 112–119.
Sinyakov, M.S., Dror, M., Zhevelev, H.M., Margel, S., Avtalion, R.R., 2002. Naturalantibodies and their significance in active immunization and protection against adefined pathogen in fish. Vaccine 20, 3668–3674.
Sinyakov, M.S., Dror, M., Lublin-Tennenbaum, T., Salzberg, S., Margel, S., Avtalion, R.R.,2006. Nano- and microparticles as adjuvants in vaccine design: Success and failureis related to host natural antibodies. Vaccine 24, 6534–6541.
Tchernychev, B., Rabinkov, A.,Mirelman,D.,Wilchek,M.,1995.Natural antibodies todietaryproteins: the existence of natural antibodies to alliinase (Alliin lyase) and mannose-specific lectin from garlic (Allium sativum) in human serum. Immunol. Lett. 47, 53–57.
Tchernychev, B., Cabilly, S., Wilchek, M., 1997. The epitopes for natural polyreactiveantibodies are rich in proline. Proc. Natl. Acad. Sci. U.S.A. 94, 6335–6339.
Tung, J.W., Herzenberg, L.A., 2007. Unravelling B-1 progenitors. Curr. Opin. Immunol. 19,150–155.
Vilain, C., Wetzel, M.-C., Du Pasquier, L., Charlemagne, J., 1984. Structural and functionalanalysis of spontaneous anti-nitrophenyl antibodies in three cyprinid fish species:carp (Cyrinus carpio), goldfish (Carassius auratus) and tench (Tinca tinca). Dev.Comp. Immunol. 8, 611–622.
Westphal, O., Jann, K., 1965. Bacterial lipopolysaccharides. Extraction with phenol–waterand further applications of the procedure. Methods Carbohydr. Chem. 5, 83–91.
Wilson, M.R., Marcuz, A., van Ginkel, F., Miller, N.W., Clem, L.W., Middleton, D., Warr, G.W.,1990. The immunoglobulinMheavychain constant region gene of the channel catfish,Ictalurus punctatus: an unusual mRNA splice pattern produces the membrane form ofthe molecule. Nucleic Acids Res. 18, 5227–5233.
Zwollo, P., Haines, A., Rosato, P., Gumulak-Smith, J., 2008. Molecular and cellular analysisof B-cell populations in the rainbow trout using Pax5 and immunoglobulinmarkers. Dev. Comp. Immunol. 32, 1482–1496.




















