
An Integrated Approach to Gene Discovery and Marker Development in Atlantic Cod (Gadus morhua)

Ff
Da
b
a
ARRAA
KPgBARA
1
wewtar
dapcttpcgobm
Uf
0d
International Journal of Biological Macromolecules 49 (2011) 1110– 1116
Contents lists available at SciVerse ScienceDirect
International Journal of Biological Macromolecules
journa l h o me pag e: www.elsev ier .com/ locate / i jb iomac
ree radical scavenging and angiotensin-I converting enzyme inhibitory peptidesrom Pacific cod (Gadus macrocephalus) skin gelatin
ai-Hung Ngoa, BoMi Ryua, Thanh-Sang Voa, S.W.A. Himayaa, Isuru Wijesekaraa, Se-Kwon Kima,b,∗
Department of Chemistry, Pukyong National University, Busan 608-737, Republic of KoreaMarine Bioprocess Research Center, Pukyong National University, Busan 608-737, Republic of Korea
r t i c l e i n f o
rticle history:eceived 21 July 2011eceived in revised form 7 September 2011ccepted 8 September 2011vailable online 16 September 2011
a b s t r a c t
Potent antioxidative peptides were purified from Pacific cod (Gadus macrocephalus) skin gelatin usingalcalase, neutrase, papain, trypsin, pepsin, and �-chymotrypsin. Among them, the papain hydrolysateexhibited the highest antioxidant activity. Therefore, it was further purified and obtained two pep-tides with amino acid sequences of Thr-Cys-Ser-Pro (388 Da) and Thr-Gly-Gly-Gly-Asn-Val (485.5 Da).The antioxidant activity of the purified peptides was performed by electron spin resonance technique.Moreover, their intracellular free radical scavenging activity using 2′,7′-dichlorofluorescin diacetate and
eywords:acific cod (Gadus macrocephalus) skinelatinioactive peptidesntioxidants
the protective effect against oxidation-induced DNA damage were evaluated in mouse macrophages(RAW 264.7 cells). Furthermore, both peptides have shown potential angiotensin-I converting enzymeinhibitory effect. The present study demonstrated that the peptides derived from Pacific cod (G.macrocephalus) skin gelatin could be used in the food industry as functional ingredients with potent
erten
AW 264.7 cellsngiotensin-I converting enzyme inhibitionantioxidative and antihyp
. Introduction
In recent years, cod is one of the important fish-catches of theorld and it is used entirely for the cod fillet production. How-
ver, the cod skin is largely underutilized and discarded as wastes,hich lead to environmental pollution. Recent studies have shown
hat fish skin provides the best source of gelatin because of its highvailability, reducing pollution, no risk of disease transmission, noeligious barriers and possibility of higher yields of collagen [1].
High blood pressure is one of the major risk factors for car-iovascular diseases including coronary heart disease, peripheralrtery disease and stroke. Angiotensin-I converting enzyme (ACE)lays a critical physiological role in regulation of blood pressure byonverting angiotensin-I to angiotensin-II, a potent vasoconstric-or. Therefore, the inhibition of ACE activity is a major target inhe prevention of hypertension [2]. Many natural ACE inhibitoryeptides have been isolated from different food proteins such asheese whey, casein, wakame, fermented soymilk, soybean, cornluten [3], yellowfin sole, Alaska pollack, short-necked clam, pearl
yster, bonito meat and bovine plasma proteins [4] and they coulde applied in the prevention of hypertension and in the initial treat-ent of mildly hypertensive individuals [5].∗ Corresponding author at: Marine Bioprocess Research Center, Pukyong Nationalniversity, Busan 608-737, Republic of Korea. Tel.: +82 51 629 7094;
ax: +82 51 629 7099.E-mail address: [email protected] (S.-K. Kim).
141-8130/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.ijbiomac.2011.09.009
sive benefits.© 2011 Elsevier B.V. All rights reserved.
Antioxidants may have a positive effect on human health sincethey can protect the human body against deterioration by reactiveoxygen species (ROS), including singlet oxygen, hydrogen perox-ide, superoxide and hydroxyl radicals. ROS can result in oxidativedamage to all the important cellular components and have beenlinked with developments of various diseases, such as hyperten-sion, neurodegenerative, inflammation, diabetes, cancer and aging[6].
Recently, several studies have been reported on the utilizationof fish by-products by enzymatic hydrolysis for the recovery ofvarious valuable components [7]. Free radical scavenging activ-ity in gelatin hydrolysates from various fish species such as Niletilapia [8], Alaska pollack [9], hoki fish [10], and cobia [11] has beenpreviously reported. However, there are no studies on the protec-tive effect of peptides from Pacific cod (Gadus macrocephalus) skinagainst free radical-induced oxidative damage. Therefore, the aimof this study was to evaluate the antioxidative activity of peptidesderived from enzymatic hydrolysis of Pacific cod (G. macrocephalus)skin gelatin using mouse macrophage RAW 264.7 cells. In addition,ACE inhibitory activity of the purified peptides was also deter-mined.
2. Experimental
2.1. Materials
Pacific cod (G. macrocephalus) skin was obtained from Jakalchifish market, Busan, South Korea. The fluorescence probes
D.-H. Ngo et al. / International Journal of Biological Macromolecules 49 (2011) 1110– 1116 1111
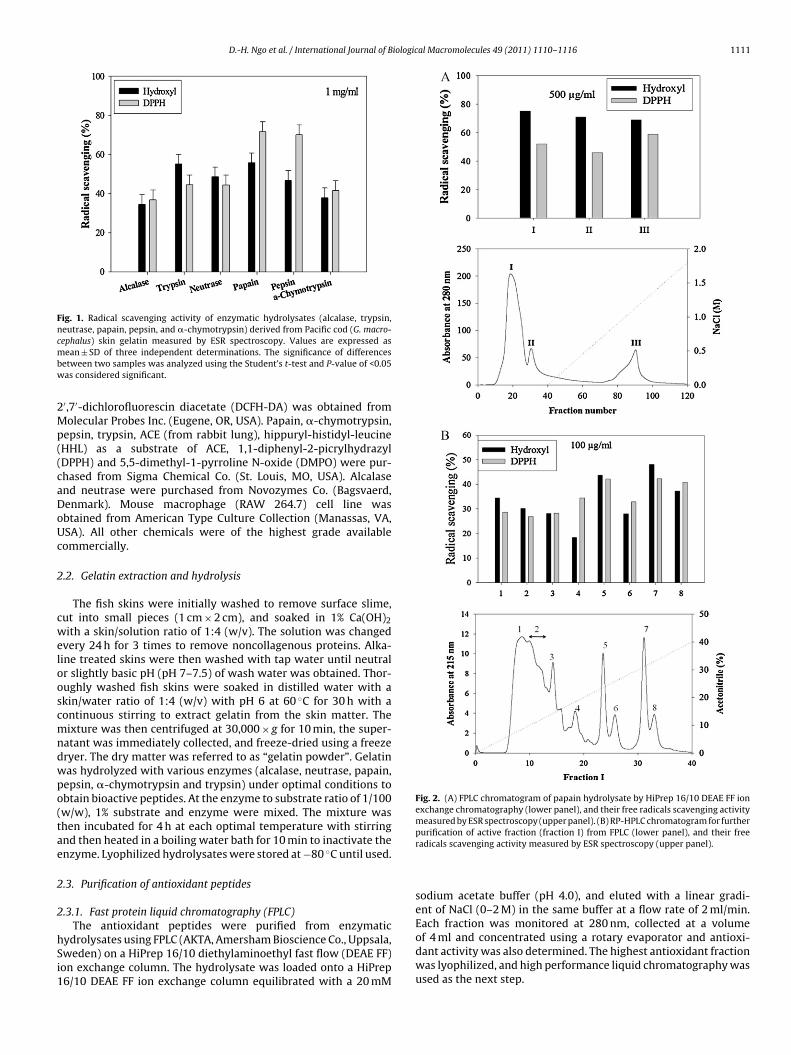
Fig. 1. Radical scavenging activity of enzymatic hydrolysates (alcalase, trypsin,neutrase, papain, pepsin, and �-chymotrypsin) derived from Pacific cod (G. macro-cephalus) skin gelatin measured by ESR spectroscopy. Values are expressed asmbw
2Mp((caDoUc
2
cwelooscmndwpo(tae
2
2
hSi1
Fig. 2. (A) FPLC chromatogram of papain hydrolysate by HiPrep 16/10 DEAE FF ionexchange chromatography (lower panel), and their free radicals scavenging activitymeasured by ESR spectroscopy (upper panel). (B) RP-HPLC chromatogram for furtherpurification of active fraction (fraction I) from FPLC (lower panel), and their free
ean ± SD of three independent determinations. The significance of differencesetween two samples was analyzed using the Student’s t-test and P-value of <0.05as considered significant.
′,7′-dichlorofluorescin diacetate (DCFH-DA) was obtained fromolecular Probes Inc. (Eugene, OR, USA). Papain, �-chymotrypsin,
epsin, trypsin, ACE (from rabbit lung), hippuryl-histidyl-leucineHHL) as a substrate of ACE, 1,1-diphenyl-2-picrylhydrazylDPPH) and 5,5-dimethyl-1-pyrroline N-oxide (DMPO) were pur-hased from Sigma Chemical Co. (St. Louis, MO, USA). Alcalasend neutrase were purchased from Novozymes Co. (Bagsvaerd,enmark). Mouse macrophage (RAW 264.7) cell line wasbtained from American Type Culture Collection (Manassas, VA,SA). All other chemicals were of the highest grade availableommercially.
.2. Gelatin extraction and hydrolysis
The fish skins were initially washed to remove surface slime,ut into small pieces (1 cm × 2 cm), and soaked in 1% Ca(OH)2ith a skin/solution ratio of 1:4 (w/v). The solution was changed
very 24 h for 3 times to remove noncollagenous proteins. Alka-ine treated skins were then washed with tap water until neutralr slightly basic pH (pH 7–7.5) of wash water was obtained. Thor-ughly washed fish skins were soaked in distilled water with akin/water ratio of 1:4 (w/v) with pH 6 at 60 ◦C for 30 h with aontinuous stirring to extract gelatin from the skin matter. Theixture was then centrifuged at 30,000 × g for 10 min, the super-
atant was immediately collected, and freeze-dried using a freezeryer. The dry matter was referred to as “gelatin powder”. Gelatinas hydrolyzed with various enzymes (alcalase, neutrase, papain,epsin, �-chymotrypsin and trypsin) under optimal conditions tobtain bioactive peptides. At the enzyme to substrate ratio of 1/100w/w), 1% substrate and enzyme were mixed. The mixture washen incubated for 4 h at each optimal temperature with stirringnd then heated in a boiling water bath for 10 min to inactivate thenzyme. Lyophilized hydrolysates were stored at −80 ◦C until used.
.3. Purification of antioxidant peptides
.3.1. Fast protein liquid chromatography (FPLC)The antioxidant peptides were purified from enzymatic
ydrolysates using FPLC (AKTA, Amersham Bioscience Co., Uppsala,weden) on a HiPrep 16/10 diethylaminoethyl fast flow (DEAE FF)on exchange column. The hydrolysate was loaded onto a HiPrep6/10 DEAE FF ion exchange column equilibrated with a 20 mM
radicals scavenging activity measured by ESR spectroscopy (upper panel).
sodium acetate buffer (pH 4.0), and eluted with a linear gradi-ent of NaCl (0–2 M) in the same buffer at a flow rate of 2 ml/min.Each fraction was monitored at 280 nm, collected at a volumeof 4 ml and concentrated using a rotary evaporator and antioxi-dant activity was also determined. The highest antioxidant fraction
was lyophilized, and high performance liquid chromatography wasused as the next step.
1112 D.-H. Ngo et al. / International Journal of Biological Macromolecules 49 (2011) 1110– 1116
F ptideI
2
fK(
ig. 3. Identification of molecular mass and amino acid sequence of the purified pe-7 (peptide 7).
.3.2. High performance liquid chromatography (HPLC)
The fraction exhibiting the highest antioxidant activity wasurther purified using reverse-phase HPLC (RP-HPLC, Dionexorea Ltd., Sunnyvale, CA, USA) on a Primesphere 10 C18
10 mm × 250 mm, Phenomenex, Cheshire, England) column with
s using TOF-MS/MS with an ESI source. (A) Fraction I-5 (peptide 5) and (B) fraction
a linear gradient of acetonitrile (0–40% in 40 min) containing 0.1%
trifluoroacetic acid (TFA) at a flow rate of 2 ml/min. Elution peakswere detected at 215 nm, and active peak was concentrated usinga rotary evaporator. Potent peaks were collected, their antioxi-dant activity evaluated, and then lyophilized. The active fraction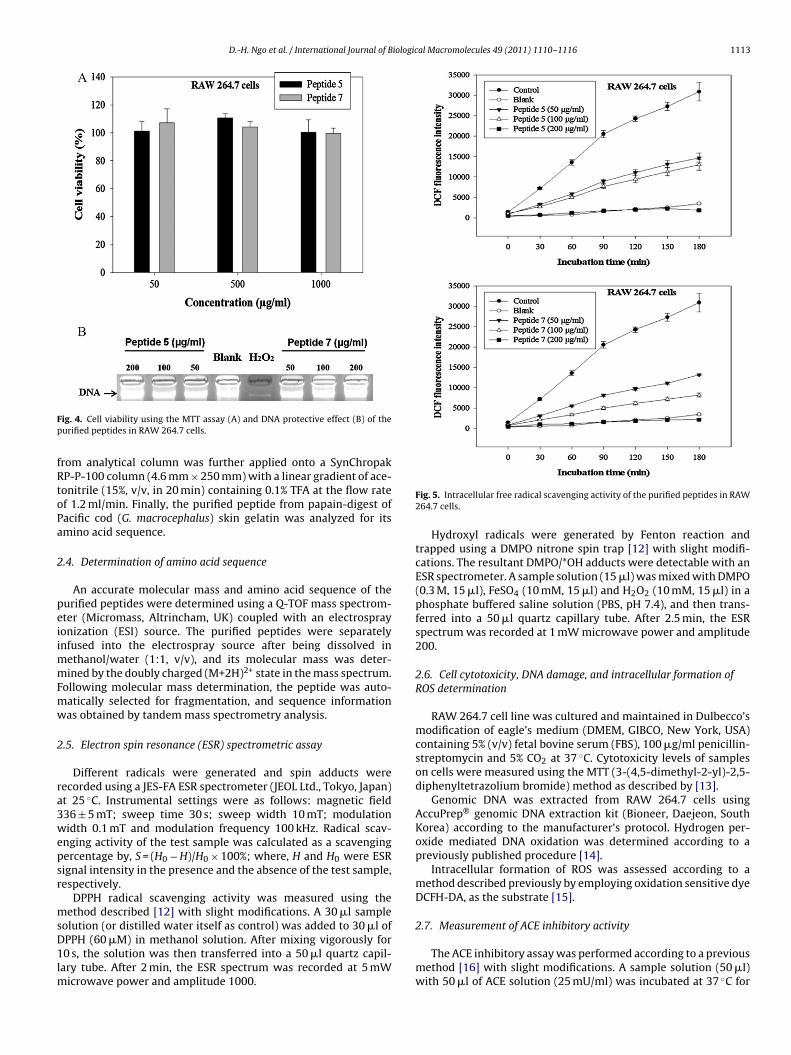
D.-H. Ngo et al. / International Journal of Biological Macromolecules 49 (2011) 1110– 1116 1113
Fp
fRtoPa
2
peiimmFmw
2
ra3wepsr
msD1lm
ig. 4. Cell viability using the MTT assay (A) and DNA protective effect (B) of theurified peptides in RAW 264.7 cells.
rom analytical column was further applied onto a SynChropakP-P-100 column (4.6 mm × 250 mm) with a linear gradient of ace-onitrile (15%, v/v, in 20 min) containing 0.1% TFA at the flow ratef 1.2 ml/min. Finally, the purified peptide from papain-digest ofacific cod (G. macrocephalus) skin gelatin was analyzed for itsmino acid sequence.
.4. Determination of amino acid sequence
An accurate molecular mass and amino acid sequence of theurified peptides were determined using a Q-TOF mass spectrom-ter (Micromass, Altrincham, UK) coupled with an electrosprayonization (ESI) source. The purified peptides were separatelynfused into the electrospray source after being dissolved in
ethanol/water (1:1, v/v), and its molecular mass was deter-ined by the doubly charged (M+2H)2+ state in the mass spectrum.
ollowing molecular mass determination, the peptide was auto-atically selected for fragmentation, and sequence informationas obtained by tandem mass spectrometry analysis.
.5. Electron spin resonance (ESR) spectrometric assay
Different radicals were generated and spin adducts wereecorded using a JES-FA ESR spectrometer (JEOL Ltd., Tokyo, Japan)t 25 ◦C. Instrumental settings were as follows: magnetic field36 ± 5 mT; sweep time 30 s; sweep width 10 mT; modulationidth 0.1 mT and modulation frequency 100 kHz. Radical scav-
nging activity of the test sample was calculated as a scavengingercentage by, S = (H0 − H)/H0 × 100%; where, H and H0 were ESRignal intensity in the presence and the absence of the test sample,espectively.
DPPH radical scavenging activity was measured using theethod described [12] with slight modifications. A 30 �l sample
olution (or distilled water itself as control) was added to 30 �l of
PPH (60 �M) in methanol solution. After mixing vigorously for0 s, the solution was then transferred into a 50 �l quartz capil-ary tube. After 2 min, the ESR spectrum was recorded at 5 mWicrowave power and amplitude 1000.
Fig. 5. Intracellular free radical scavenging activity of the purified peptides in RAW264.7 cells.
Hydroxyl radicals were generated by Fenton reaction andtrapped using a DMPO nitrone spin trap [12] with slight modifi-cations. The resultant DMPO/*OH adducts were detectable with anESR spectrometer. A sample solution (15 �l) was mixed with DMPO(0.3 M, 15 �l), FeSO4 (10 mM, 15 �l) and H2O2 (10 mM, 15 �l) in aphosphate buffered saline solution (PBS, pH 7.4), and then trans-ferred into a 50 �l quartz capillary tube. After 2.5 min, the ESRspectrum was recorded at 1 mW microwave power and amplitude200.
2.6. Cell cytotoxicity, DNA damage, and intracellular formation ofROS determination
RAW 264.7 cell line was cultured and maintained in Dulbecco’smodification of eagle’s medium (DMEM, GIBCO, New York, USA)containing 5% (v/v) fetal bovine serum (FBS), 100 �g/ml penicillin-streptomycin and 5% CO2 at 37 ◦C. Cytotoxicity levels of sampleson cells were measured using the MTT (3-(4,5-dimethyl-2-yl)-2,5-diphenyltetrazolium bromide) method as described by [13].
Genomic DNA was extracted from RAW 264.7 cells usingAccuPrep® genomic DNA extraction kit (Bioneer, Daejeon, SouthKorea) according to the manufacturer’s protocol. Hydrogen per-oxide mediated DNA oxidation was determined according to apreviously published procedure [14].
Intracellular formation of ROS was assessed according to amethod described previously by employing oxidation sensitive dyeDCFH-DA, as the substrate [15].
2.7. Measurement of ACE inhibitory activity
The ACE inhibitory assay was performed according to a previousmethod [16] with slight modifications. A sample solution (50 �l)with 50 �l of ACE solution (25 mU/ml) was incubated at 37 ◦C for

1114 D.-H. Ngo et al. / International Journal of Biological Macromolecules 49 (2011) 1110– 1116
Table 1Antioxidant peptides derived from marine organisms: source and amino acidsequence.
Source Amino acid sequence Refs.
Microalga Val-Glu-Cys-Tyr-Gly-Pro-Asn-Arg-Pro-Gln-Phe [22]Rotifer Leu-Leu-Gly-Pro-Gly-Leu-Thr-Asn-His-Ala [23]
Asp-Leu-Gly-Leu-Gly-Leu-Pro-Gly-Ala-His [23]Hoki Glu-Ser-Thr-Val-Pro-Glu-Arg-Thr-His-Pro-Ala-
Cys-Pro-Asp-Phe-Asn[24]
Tuna Val-Lys-Ala-Gly-Phe-Ala-Trp-Thr-Ala-Asn-Glu-Glu-Leu-Ser
[25]
Jumbo squid Phe-Asp-Ser-Gly-Pro-Ala-Gly-Val-Leu [26]Giant squid Asn-Ala-Asp-Phe-Gly-Leu-Asn-Gly-Leu-Glu- [27]
5(amatiTa(Ar
2
tbPc
3
3p
t
FVia
Table 2ACE inhibitory peptides derived from marine organisms: source and amino acidsequence.
Source Amino acid sequence Refs.
Tuna Gly-Asp-Leu-Gly-Lys-Thr-Thr-Thr-Val-Ser-Asn-Trp-Ser-Pro-Pro-Lys-Tyr-Lys-Asp-Thr-Pro
[30]
Microalga Val-Glu-Cys-Tyr-Gly-Pro-Asn-Arg-Pro-Gln-Phe [33]Sea cucumber Met-Glu-Gly-Ala-Gln-Glu-Ala-Gln-Gly-Asp [34]Rotifer Asp-Asp-Thr-Gly-His-Asp-Phe-Glu-Asp-Thr-
Gly-Glu-Ala-Met[35]
Shrimp Ile-Phe-Val-Pro-Ala-Phe [36]Yellowfin sole Met-Ile-Phe-Pro-Gly-Ala-Gly-Gly-Pro-Glu-Leu [37]Hard clam Tyr-Asn [38]Sea bream Val-Ile-Tyr [39]Alaska pollack Gly-Pro-Leu [40]
Gly-Leu-AlaBlue mussel Phe-Gly-His-Pro-Tyr [28]
min, and the mixture was pre-incubated with 150 �l of substrate8.3 mM HHL in 50 mM sodium borate buffer containing 0.5 M NaClt pH 8.3) for 30 min at the same temperature. The reaction was ter-inated by the addition of 250 �l of 1 M HCl. The resulting hippuric
cid was extracted with 500 �l of ethyl acetate. After centrifuga-ion (30,000 × g, 10 min), 200 �l of the upper layer was transferrednto a glass tube, and evaporated at 60 ◦C for 2 h in a vacuum.he hippuric acid was dissolved in 1 ml of distilled water, and thebsorbance was measured at 228 nm using UV-spectrophotometerGENios microplate reader, Tecan Austria GmbH, Grodig/Salzburg,ustria). The IC50 value defined as the concentration of inhibitorequired to inhibit 50% of the ACE activity.
.8. Statistical analysis
Data were expressed as mean ± standard deviation (SD) ofhree independent determinations. The significance of differencesetween two samples was analyzed using the Student’s t-test and-value of <0.05 was considered as the level of statistical signifi-ance.
. Results and discussion
.1. Preparation of gelatin hydrolysates and their antioxidant
ropertiesThe antioxidative activity of the hydrolysates was evaluated forheir free radical scavenging effect on hydroxyl and DPPH radicals
ig. 6. Angiotensin-I converting enzyme inhibitory activity of the purified peptides.alues are expressed as mean ± SD of three independent determinations. The signif-
cance of differences between two samples was analyzed using the Student’s t-testnd P-value of <0.05 was considered significant.
using electron spin resonance (ESR) technique. As shown in Fig. 1,scavenging effect of the hydrolysates on DPPH radical was moreefficient than that of hydroxyl radical. Among the hydrolysatesresulting from various enzymes, the papain hydrolysate exhib-ited the most potential antioxidant activity than other enzymatichydrolysates. The scavenging effects of papain hydrolysate on DPPHand hydroxyl radicals were found to be about 72% and 56% at1 mg/ml, respectively. Therefore, papain hydrolysate was selectedfor further purification.
3.2. Purification and direct free radical scavenging effect of thepurified peptides
Firstly, papain hydrolysate was dissolved in sodium acetatebuffer (20 mM, pH 4.0) at 20 mg/ml, and loaded onto a HiPrep 16/10DEAE FF ion exchange column using FPLC with a linear gradientof NaCl (0–2.0 M). The elution peaks were monitored at 280 nm,and each fraction was collected as 4 ml and fractionated into threeportions. Each fraction was pooled, lyophilized, and measured forits antioxidant activity in direct free radicals scavenging activity(Fig. 2A). Among all fractions, fraction I had the strongest hydroxylradical scavenging effect by 75% at 500 �g/ml. Therefore, fractionI was further purified on an analytical HPLC column using a lin-ear gradient of acetonitrile (0–40%) and its antioxidant activitiesmeasured using their potency to scavenge free radicals (Fig. 2B).Amino acid sequences of the purified two peptides were deter-mined to be Thr-Gly-Gly-Gly-Asn-Val (485.5 Da, peptide 5) (Fig. 3A)and Thr-Cys-Ser-Pro (388 Da, peptide 7) (Fig. 3B) by Q-TOF ESI massspectroscopy. The structure of peptide 5 was composed of residuesof one threonine, three glycine, one asparagine and one valine. Thestructure of peptide 7 was composed of residues of one threonine,one cysteine, one serine and one proline. Further, in the sequenceof peptide 5 has hydrophobic amino acid such glycine and valine; inthe sequence of peptide 7 has cysteine and hydrophobic amino acidsuch as proline. Sequence analysis studies revealed that the puri-fied peptides are rich in hydrophobic amino acids (glycine, proline),which contribute substantially for scavenging of free radicals act-ing as potent electron donors. Cysteine residues are independentlyimportant for antioxidant activity as they can directly interact withfree radicals. Furthermore, the thiol group of cysteine plays animportant role in the protecting cells and cellular biomoleculesagainst oxidative stress. Threonine is an important component inthe formation of protein, collagen and elastin. Threonine helps theliver and maintains the body’s proteins in balance. Bioactive pep-tides usually contain 3–20 amino acids residues and low molecular
weight peptides are more potent as bioactive peptides than highmolecular weight peptides [17].
iologic
3p
tficCc
cdocaboamw(oiao
3p
patcdciflmiflfipTbpm
3
aptpthiadcimTtA
[[[
[[
[
[[[
[
[
[
[[
[[[[
[[
[
D.-H. Ngo et al. / International Journal of B
.3. Cell viability and DNA protective activity of the purifiedeptides on RAW 264.7 cells
RAW 264.7 cells were treated with different concentrations ofhe purified peptides to determine the cytotoxic effect of the puri-ed peptides. According to the MTT assay, it has shown that noytotoxicity of the purified peptides on RAW 264.7 cells (Fig. 4A).onsequently, we could use the purified peptides for further intra-ellular free radical scavenging experiments.
The highly reactive hydroxyl radicals can easily react with allomponents of the DNA molecule and damage DNA. In addition, thisamage increases with the increment of the free radicals attackedn cellular DNA, which involved in the occurrence of numeroushronic diseases such as inflammation, cancer, mutagenesis andging [18–20]. In this study, DNA oxidation was performed by com-ining effect of 200 �M Fe(II) and 2 mM H2O2 on the integrityf genomic DNA isolated from RAW 264.7 cells. The protectivectivity of the purified peptides against DNA damage was deter-ined by DNA electrophoresis. After the reaction, almost all DNAas degraded in the control group treated with H2O2–Fe(II) alone
Fig. 4B). However, the purified peptides showed a protective effectn free radical-mediated DNA damage by dose-dependently. Evenn low concentration (50 �g/ml) of the purified peptides, DNA dam-ge was inhibited more than 90% determined based on the intensityf DNA bands.
.4. Intracellular free radical scavenging activity of the purifiedeptides on RAW 264.7 cells
The intracellular free radical scavenging effect of the purifiedeptides was carried out on RAW 264.7 cells as these cells areble to release high amounts of ROS following stimulation [21]. Forhis experiment, intracellular ROS was measured using the fluores-ence probe DCFH-DA. During labeling, non-fluorescent DCFH-DAye that easily penetrates into the cells gets hydrolyzed by intra-ellular esterase to become DCFH, and this compound is trappednside the cells and gets oxidized by H2O2. The monitoring of DCFuorescence intensity in every 30 min for 3 h duration that radical-ediated oxidation increased with the incubation time. As shown
n Fig. 5, the treatment with the purified peptides reduced the DCFuorescence intensity in a dose- and time-dependently. The puri-ed two peptides have amino acid residues which are generallyresent in antioxidant peptides derived from marine organisms.he antioxidant activity of marine-derived bioactive peptides haseen summarized in Table 1. These results confirmed that theurified two peptides scavenged free radicals and inhibited radical-ediated oxidation in RAW 264.7 cells.
.5. ACE inhibitory activity of the purified peptides
Bioactive peptides usually contain 3–20 amino acids residuesnd low molecular weight peptides are more potent as bioactiveeptides than high molecular weight peptides [17]. Regarding tohe relationship between activity and structure of ACE inhibitoryeptides, those peptides had tyrosine, proline, or phenylalanine athe C-terminus and isoleucine or valine at the N-terminal showedighly potential inhibitory activity [29]. Most of the reported ACE
nhibitory peptides are usually short peptides with proline residuet the C-terminal end [30,31]. Proline is known to be preventingigestion of enzymes and may pass through the capillary into theirculation of blood in the sequence of short peptides [32]. As shownn Fig. 6, the purified peptide, Thr-Cys-Ser-Pro (388 Da) showed the
ost potent ACE inhibitory activity (81%) than the other peptide,hr-Gly-Gly-Gly-Asn-Val (485.5 Da) (68%) at 500 �g/ml. Therefore,hese peptides can be incorporated into functional foods as novelCE inhibitors. Amino acid residues in the purified two peptides are
[[[[
al Macromolecules 49 (2011) 1110– 1116 1115
generally present in ACE inhibitory peptides derived from marineorganisms. Marine-derived peptides have been shown potent ACEinhibitory activities (Table 2).
4. Conclusion
Marine by-products are natural bioresources that might beuseful in health beneficial products, pharmaceuticals and foodindustries. In this study, two bioactive peptides were purifiedfrom Pacific cod (G. macrocephalus) skin gelatin by papain enzy-matic hydrolysis. Using consecutive chromatographic methods,two antioxidative and antihypertensive peptides, Thr-Cys-Ser-Pro(388 Da) and Thr-Gly-Gly-Gly-Asn-Val (485.5 Da), were shown toexhibit potent radical scavenging and ACE inhibitory activities.In conclusion, these results suggested that the antioxidant andACE inhibitory peptides from Pacific cod (G. macrocephalus) skingelatin could be potential candidates to develop functional foodsor nutraceutical products against free radicals, hypertension andrelated diseases.
Acknowledgement
This research was supported by a grant from Marine BioprocessResearch Center of the Marine Bio 21 Center funded by the Ministryof Land, Transport and Maritime, Republic of Korea.
References
[1] L.S. Senaratne, P.J. Park, S.K. Kim, Bioresour. Technol. 97 (2006) 191–197.[2] I. Wijesekara, S.K. Kim, Mar. Drugs 8 (2010) 1080–1093.[3] K. Suetsuna, T. Nakano, J. Nutr. Biochem. 11 (2000) 450–454.[4] K. Suetsuna, Fisheries Sci. 68 (2002) 233–235.[5] C. Guang, R.D. Phillips, J. Agric. Food Chem. 57 (2009) 5113–5120.[6] D.H. Ngo, I. Wijesekara, T.S. Vo, Q.V. Ta, S.K. Kim, Food Res. Int. 44 (2011)
523–529.[7] J.Y. Je, K.H. Lee, M.H. Lee, C.B. Ahn, Food Res. Int. 42 (2009) 1266–1272.[8] D.H. Ngo, Z.J. Qian, B.M. Ryu, J.W. Park, S.K. Kim, J. Funct. Foods 2 (2010)
107–117.[9] S.K. Kim, Y.T. Kim, H.G. Byun, K.S. Nam, D.S. Joo, F. Shahidi, J. Agric. Food Chem.
49 (2001) 1984–1989.10] E. Mendis, N. Rajapakse, S.K. Kim, J. Agric. Food Chem. 53 (2005) 581–587.11] J.I. Yang, H.Y. Ho, Y.J. Chu, C.J. Chow, Food Chem. 110 (2008) 128–136.12] E. Alvarez-Parrilla, L.A. Rosa, R. Amarowicz, F. Shahidi, J. Agric. Food Chem. 59
(2011) 163–173.13] T.S. Vo, C.S. Kong, S.K. Kim, Carbohydr. Polym. 84 (2011) 649–655.14] N. Rajapakse, M.M. Kim, E. Mendis, S.K. Kim, Bioorg. Med. Chem. 15 (2007)
997–1003.15] Z.J. Qian, W.K. Jung, H.G. Byun, S.K. Kim, Bioresour. Technol. 99 (2008)
3365–3371.16] D.W. Cushman, H.S. Cheung, Biochem. Pharmacol. 20 (1971) 1637–1648.17] L. Pihlanto, Trends Food Sci. Technol. 11 (2000) 347–356.18] T.K. Hazra, A. Das, S. Das, S. Choudhury, Y.W. Kow, R. Roy, DNA Repair 6 (2007)
470–480.19] F. Madia, C. Gattazzo, P. Fabrizio, V.D. Longo, Mech. Ageing Dev. 128 (2007)
45–49.20] L.D. Mello, S. Hernandez, S. Marrazza, M. Mascini, L.T. Kubot, Biosens. Bioelec-
tron. 21 (2006) 1374–1382.21] D.H. Ngo, Z.J. Qian, T.S. Vo, B.M. Ryu, D.N. Ngo, S.K. Kim, Carbohydr. Polym. 84
(2011) 1282–1288.22] I.C. Sheih, T.K. Wu, T.J. Fang, Bioresour. Technol. 100 (2009) 3419–3425.23] H.G. Byun, J.K. Lee, H.G. Park, J.K. Jeon, S.K. Kim, Process Biochem. 44 (2009)
842–846.24] S.Y. Kim, J.Y. Je, S.K. Kim, J. Nutr. Biochem. 18 (2007) 31–38.25] J.Y. Je, Z.J. Qian, H.G. Byun, S.K. Kim, Process Biochem. 42 (2007) 840–846.26] E. Mendis, N. Rajapakse, H.G. Byun, S.K. Kim, Life Sci. 77 (2005) 2166–2178.27] N. Rajapakse, E. Mendis, H.G. Byun, S.K. Kim, J. Nutr. Biochem. 16 (2005)
562–569.28] W.K. Jung, N. Rajapakse, S.K. Kim, Eur. Food Res. Technol. 220 (2005) 535–539.29] R. Kapel, E. Rahhou, D. Lecouturier, D. Guillochon, P. Dhulster, Process Biochem.
41 (2006) 1961–1966.30] S.H. Lee, Z.J. Qian, S.K. Kim, Food Chem. 118 (2010) 96–102.
31] Z.J. Qian, J.Y. Je, S.K. Kim, J. Agric. Food Chem. 55 (2007) 8398–8403.32] H. Korhonen, A. Pihlanto, Int. Dairy J. 16 (2006) 945–960.33] I.C. Sheih, T.J. Fang, T.K. Wu, Food Chem. 115 (2009) 279–284.34] Y. Zhao, L. Bafang, S. Dong, Z. Liu, X. Zhao, J. Wang, M. Zeng, Peptides 30 (2009)1028–1033.

1 iologi
[
[
[
[38] J.S. Tsai, J.L. Chen, B.S. Pan, Process Biochem. 43 (2008) 743–747.
116 D.-H. Ngo et al. / International Journal of B
35] J.K. Lee, S. Hong, J.K. Jeon, S.K. Kim, H.G. Byun, Bioresour. Technol. 100 (2009)
5255–5259.36] H.H. Lun, C.X. Lan, S.C. Yun, Z.Y. Zhong, Z.B. Cheng, J. Pept. Sci. 12 (2006)726–733.
37] W.K. Jung, E. Mendis, J.Y. Je, P.J. Park, B.W. Son, H.C. Kim, Y.K. Choi, S.K. Kim,Food Chem. 94 (2006) 26–32.
[
[
cal Macromolecules 49 (2011) 1110– 1116
39] A. Fahmi, S. Morimura, H.C. Guo, T. Shigematsu, K. Kida, Y. Uemura, ProcessBiochem. 39 (2004) 1195–1200.
40] H.G. Byun, S.K. Kim, Process Biochem. 36 (2001) 1155–1162.
Copyright © 2022 FDOKUMEN




















