
Chapter 3 Effects of Climate Change and Commercial Fishing on Atlantic Cod Gadus morhua
62
Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Advances in Marine Biology, Vol. 56, published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at: http://www.elsevier.com/locate/permissionusematerial From: Nova Mieszkowska, Martin J. Genner, Stephen J. Hawkins, and David W. Sims, Effects of Climate Change and Commercial Fishing on Atlantic Cod Gadus morhua. In D. W. Sims, editor: Advances in Marine Biology, Vol. 56, Burlington: Academic Press, 2009, pp. 213-273. ISBN: 978-0-12-374960-4 © Copyright 2009 Elsevier Ltd. Academic Press.
Transcript of Chapter 3 Effects of Climate Change and Commercial Fishing on Atlantic Cod Gadus morhua

Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Advances in Marine Biology, Vol. 56, published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
From: Nova Mieszkowska, Martin J. Genner, Stephen J. Hawkins, and David W. Sims, Effects of Climate Change and Commercial Fishing on Atlantic Cod Gadus morhua.
In D. W. Sims, editor: Advances in Marine Biology, Vol. 56, Burlington: Academic Press, 2009, pp. 213-273.
ISBN: 978-0-12-374960-4 © Copyright 2009 Elsevier Ltd.
Academic Press.

Author’s personal copy
C H A P T E R T H R E E
A
IS
*
{
{
}
dvance
SN 0
MariUnitSchooColleUniteMarinDrake
Effects of Climate Change and
Commercial Fishing on Atlantic
Cod Gadus morhua
Nova Mieszkowska,* Martin J. Genner,*,† Stephen J. Hawkins,*,‡
and David W. Sims*,§
Contents
1. In
s in
065
ne Bed Kl oge od Ke BCi
troduction
Marine Biology, Volume 56 # 2009
-2881, DOI: 10.1016/S0065-2881(09)56003-8 All rig
iological Association of the United Kingdom, The Laboratory, Citadel Hill, Plymouingdomf Biological Sciences, University of Bristol, Bristol BS8 1UG, United Kingdomf Natural Sciences, Memorial Building, Bangor University, Gwynedd LL57 2UW,ingdomiology and Ecology Research Centre, School of Biological Sciences, University of Plrcus, Plymouth PL4 8AA, United Kingdom
Else
hts
th P
ym
214
1
.1. B asic biology and global distribution 2151
.2. G enetic population structure 2161
.3. T raits in different stocks 2181
.4. M ovement and activity 2192. Im
pacts of Climate Change 2222
.1. B iogeographic changes 2242
.2. P hysiology 2262
.3. M etabolic scope for activity 2292
.4. M aturation and spawning 2302
.5. E arly life stages 2312
.6. R ecruitment 2332
.7. G rowth 2373. Im
pacts of Fishing 2383
.1. N orthwest Atlantic stocks 2383
.2. N ortheast Atlantic stocks 2393
.3. T he fishing versus climate change debate 2444. P
opulation-Level Impacts of Fishing and Climate Change 2454
.1. S tock assessment 2454
.2. S tock evaluation—An example from the North Sea 2464
.3. A llee effects and management plans 2475. M
onitoring Status and Recovery of North Sea Cod: A Case Study 249vier Ltd.
reserved.
L1 2PB,
outh,
213

214 Nova Mieszkowska et al.
Author’s personal copy
6. C
oncluding Remarks 250Ackn
owledgements 252Refe
rences 252Abstract
During the course of the last century, populations of Atlantic cod Gadus
morhua L. have undergone dramatic declines in abundance across their biogeo-
graphic range, leading to debate about the relative roles of climatic warming
and overfishing in driving these changes. In this chapter, we describe the
geographic distributions of this important predator of North Atlantic ecosys-
tems and document extensive evidence for limitations of spatial movement and
local adaptation from population genetic markers and electronic tagging. Taken
together, this evidence demonstrates that knowledge of spatial population
ecology is critical for evaluating the effects of climate change and commercial
harvesting. To explore the possible effects of climate change on cod, we first
describe thermal influences on individual physiology, growth, activity and
maturation. We then evaluate evidence that temperature has influenced popu-
lation-level processes including direct effects on recruitment through enhanced
growth and activity, and indirect effects through changes to larval food
resources. Although thermal regimes clearly define the biogeographic range of
the species, and strongly influencemany aspects of cod biology, the evidence that
population declines across the North Atlantic are strongly linked to fishing activity
is now overwhelming. Although there is considerable concern about low spawning
stock biomasses, high levels of fishing activity continues in many areas. Even with
reduced fishing effort, the potential for recovery from low abundance may be
compromised by unfavourable climate and Allee effects. Current stock assess-
ment and management approaches are reviewed, alongside newly advocated
methods for monitoring stock status and recovery. However, it remains uncertain
whether the rebuilding of cod to historic population sizes and demographic
structures will be possible in a warmer North Atlantic.
1. Introduction
Atlantic cod Gadus morhua Linnaeus, 1758 is one of the most widelystudied marine fishes (Fig. 3.1). The species is a major predator in NorthAtlantic ecosystems as well as being a prey item for larger fishes and piscivo-rous marine mammals. It has been exploited as a human food resource forover 1000 years and forms a key component of major fisheries throughout theNorth Atlantic. Its pivotal ecological role, together with its economic impor-tance, has made it a model system for study among marine fishes. Studiesrange from individual physiology, to population ecology, community inter-actions and responses to environmental change, including climate change andfishing. Here, we review these studies and discuss how key aspects of cod

Figure 3.1 Atlantic cod, Gadus morhua Linnaeus 1758. Photograph courtesy of theMarine Biological Association of the UK.
Climate and Fishing Effects on Cod 215
Author’s personal copy
biology are likely to be influenced by changing environments, such as thoseassociated with changes in fishing pressure and climate. We examine theimpacts of these drivers on the current status and potential for future recoveryof stocks, with a focus on the North Sea and evaluate potential managementstrategies to reverse the current global decline in Atlantic cod.
1.1. Basic biology and global distribution
Atlantic cod is an apex predator of North Atlantic continental shelf waters.It feeds mainly on invertebrates and fish. It grows to a maximum of 2 m intotal length, weighs up to 96 kg, matures at between 2 and 4 years of age(O’Brien et al., 1993) and can live for up to 25 years. Females are typicallyhighly fecund, producing an average of 1 million eggs per individual(Cohen et al., 1990). Spawning takes place between December andJune depending on geographic location, and eggs hatch 2–3 weeks later.The pelagic larvae feed on zooplankton for approximately 2 months beforesettling on demersal nursery grounds.
The biogeographic range of cod, like many marine fish species, isprimarily governed by temperature (Coutant, 1987; Sundby, 2000). In theNorth Atlantic Ocean, it is found between 40 and 80 �N over a tempera-ture gradient of �1 to 20 �C. Northern limits occur in Canada and Iceland,and southern limits are reached around New England in the westernAtlantic, and in the Celtic Sea–English Channel in the eastern Atlantic(Fig. 3.2). Key areas of population abundance are Labrador, Newfoundland,southern Greenland, Iceland, the North Sea, the Baltic Sea and the BarentsSea (Bigg et al., 2008; Sundby, 2000). Its depth range extends from shallowwaters to 200 m, although it has been recorded at depths of over 500 m(Cohen et al., 1990).

90�W 60�W 30�W
90�W
60�N
40�N
60�N
40�N
60�W 30�W 0�
0�
Figure 3.2 The distribution of Atlantic cod in the North Atlantic Ocean (greyshaded area).
216 Nova Mieszkowska et al.
Author’s personal copy
1.2. Genetic population structure
Cod live in a diverse and complex marine landscape with contrasting watertemperatures, depths, salinities, substrates and prey availability. In manymarine species, habitat discontinuities can act as barriers to gene flow byrestrictingmovement of adults or larvae (Hauser andCarvalho, 2008).Geneticdifferences between populations can be further promoted by behaviouraltraits, such as adult philopatry, population-specific migratory behaviour orkin aggregation (Pardini et al., 2001). The formation of genetic substructurewill, however, be counteracted to some extent by the capability that adultfishes have to range over long stretches of coastline and open water, and thepelagic dispersal of eggs and larvae (Hauser and Carvalho, 2008).
Identification of geographic and behavioural barriers to gene flow iscritical for stock identification and management. Cod are very patchilydistributed across their range (�7300 km), and molecular markers havehelped to reveal the extent of migration among populations. Several studieshave revealed a broad spatial trend of isolation-by-distance across the NorthAtlantic. In mitochondrial DNA, despite an apparent lack of nucleotidediversity (Arnason and Palsson, 1996), there are large differences in thefrequency of common haplotypes (Arnason, 2004; Carr and Crutcher,1998). Similar patterns have been revealed using restriction fragments ofnuclear genes (Pogson et al., 1995, 2001), and nuclear microsatellite markers(Hutchinson et al., 2001; O’Leary et al., 2007; Pampoulie et al., 2008;Skarstein et al., 2007). The extent of population differences in microsatelliteallele frequencies are such that cod can be reliably assigned to the Baltic Sea,North Sea and the Northeastern Arctic Ocean on the basis of theirgenotypes alone (Nielsen et al., 2001).

Climate and Fishing Effects on Cod 217
Author’s personal copy
1.2.1. Cod population persistenceNearly all studies making temporal comparisons of cod population structureusing archived genetic material have found remarkable stability (Imslandet al., 2004; Jonsdottir et al., 1999, 2001; Lage et al., 2004; Nielsen et al.,2001; Pampoulie et al., 2008). A major change in genetic diversity was,however, found in the heavily fished Flamborough Head population in theNorth Sea between 1954 and 1970. After the decline observed over thisperiod, diversity subsequently increased, and this has been linked to theincursion of new genotypes as new colonists had proportionally greaterreproductive success (Hutchinson et al., 2003). The general natural stabilityof cod populations, however, implies that the present distribution of geneticdiversity has changed little over recent time. Bigg et al. (2008) concludedthe present distribution of cod dates back more than 100,000 years, but bycontrast, using more rapidly evolving microsatellite markers, Pampoulieet al. (2008) calculated the time scale of divergence dates more closely tothat of the Last Glacial Maximum, approximately 20,000 years ago.Although the accuracy of these time scales will depend on methods usedto calibrate evolutionary rates, together these results suggest that the largerscale distribution of genetic diversity pre-dates the last northern hemisphereglacial cycle.
1.2.2. Regional patterns of genetic diversityUsing microsatellite DNA markers, patterns of isolation-by-distance havebeen revealed over spatial scales of approximately 1000–2000 km (Beachamet al., 2002; O’Leary et al., 2007). At smaller scales (<1000 km), however,the extent of stock structure appears closely linked to environmental para-meters and behavioural differences. For example, genetic structure withinthe Northwest Atlantic has been linked to habitat discontinuities, such aschannels and trenches. Lage et al. (2004) found cod on the southernNantucket shoals to be genetically distinct from those on the neighbouringmore northerly offshore Browns Bank and Georges Bank, but there wereno genetic differences apparent between Browns Bank and Georges Bank,despite these being separated by a stretch of deep water that likely acts as abarrier to movement of adult cod (Lage et al., 2004). Here, it is likely that agyre system leads to retention of eggs and larvae on the two offshore banks,but those spawned on the Nantucket shoals do not enter this gyre and areeither retained locally, or are transported southwest. This evidence iscompatible with both larval dispersal and adult habitat fidelity-determiningspatial patterns of genetic diversity (Ruzzante et al., 1998, 1999).
There are several other examples where substantial genetic differenceshave been identified within regions. Populations in the Canadian Arcticsaltwater lakes at the extreme northwest of the species range are stronglygenetically differentiated from Atlantic populations, and show much lower

218 Nova Mieszkowska et al.
Author’s personal copy
genetic diversity (Hardie et al., 2006). This evidence is consistent with along period of genetic isolation linked to a habitat barrier. Similarly, strongpatterns of genetic differences have been found between populations in theNorth Sea and the Baltic Sea (Nielsen et al., 2003). In this case, a zone ofadmixture is flanked at each side by non-admixed populations (Nielsenet al., 2003, 2006); this pattern is associated with local adaptation fordiffering salinity regimes. Such extreme population structuring is not alwayscommon within regions, and weak genetic structuring, or genetic homo-geneity, are more typical results of studies employing neutral markers atspatial scales around 1000–2000 km (e.g. Beacham et al., 2002). In somecases, genetic differences between populations have been recovered, butthey are not always directly correlated with known environmental condi-tions or geographic distances. On the northern coast of Norway, forexample, coastal cod have clear spatial genetic structure, but no evidenceof isolation-by-distance ( Jorde et al., 2007). In this case, it would appearthat gradual adult or larval dispersal is unlikely, and instead spatial structurehas formed through sporadic colonization waves of genetically similarindividuals.
The existence of co-occurring populations that are geneticallysegregated has also become apparent. In Norway, there are differencesin otolith shapes between the Arctic offshore cod that overwinter inthe warmer, deeper waters, and the coastal inshore cod that overwinter inthe cooler, shallower water, and early genetic work found that theseoffshore and inshore cod possessed adaptive differences in their haemoglo-bin HBI allele frequency (M�ller, 1966). In the North Sea–Skagerrak areathere is also evidence that the migratory-offshore North Sea stock and thenon-migratory-coastal Skagerrak stock are genetically different, but simi-larly co-occur within inshore waters (Case et al., 2005). Genetically differ-ent offshore migratory and inshore overwintering cod are also present inNewfoundland (Ruzzante et al., 1996a,b), and have been found to differ intheir blood ‘antifreeze’ protein levels, with the inshore cod that experiencethe cooler winter temperatures possessing higher antifreeze protein concen-trations. These genetic differences appear to exhibit interannual stability(Ruzzante et al., 1997, 2000).
1.3. Traits in different stocks
There is an increasing body of evidence suggesting that genetically differentcod stocks differ in adaptive life history traits. In the Skagerrak, wherepopulation structure is apparent at a scale of less than 100 km, there isapparent spatial variability in traits, such as juvenile growth rate that corre-sponds with observed genetic variation (Olsen et al., 2008). In support of anevolutionary explanation for the observed pattern, a range of molecular andbreeding studies have revealed evidence for selection on functional genes.

Climate and Fishing Effects on Cod 219
Author’s personal copy
Several genes show variation at the stock level (Hutchinson, 2008), perhapsthe most studied gene in this context is pantophysin (PanI) (Pogson et al.,2001), formerly synaptophysin (Syp I) (Fevolden and Pogson, 1997). Par-ticular life history traits have been associated with this gene, for example,differences in mean weight, length and growth rates have been shown to bedependent on the PanI genotype around Iceland ( Jonsdottir et al., 2002,2003), and temperature and salinity have been shown to influence the PanIgenotype frequency in the Northeast Atlantic (Case et al., 2005). These fieldresults have been supported by ‘common garden’ experiments showingPanI-dependent differences in growth rate and condition factor of indivi-duals reared to 10 weeks. Although these results may be due to genes linkedto the PanI (Case et al., 2006), the pattern nevertheless suggests that stronglocal adaptation of stocks is present in the natural environment.
1.4. Movement and activity
The movement and activity patterns of predatory fish such as Atlantic codcan be considered a major driver of the spatio-temporal dynamics ofpopulations and communities within ecosystems. Movements such asmigration, dispersal and regional philopatry, when played out over longertemporal scales, not only contribute to observed patterns of populationsub-structuring and connectivity, but will also determine how populationdistribution responds to drivers of climatic change or fishing pressure. Moni-toring movements of cod in relation to environment is therefore relevant tounderstanding the effects of these drivers on cod population re-distributions,with a significant role in future adaptive management regimes.
1.4.1. Adult movementsEarly mark-recapture studies, fishery surveys and fishing reports informed ageneral picture of mature cod annual movements, with migration to spawninggrounds followed by spent cod returning to their feeding grounds afterspawning (Harden Jones, 1968). Although this general model is broadlyapplicable to cod, the emerging paradigm is that their movements bygeographic location and by season are complex, with marked differencesin behaviour even within a region such as the North Sea, for example(Hobson et al., 2007). The recent advances in remote telemetry technologyfor tracking fish, particularly the miniaturization of data-loggers, has enabledthese insights (Arnold and Dewar 2001; Sims, 2008) with adult cod move-ments and activity being recorded over long time periods (>1 year) innearly all the main regions within its geographic range (Clark and Green,1990; Cote et al., 2003; Metcalfe, 2006; Neat and Righton, 2006; Neatet al., 2006; Rillahan et al., 2009; Robichaud and Rose, 2001, 2002, 2003,2004; Steinhausen et al., 2006; Wright et al., 2006). Data-logging storagetags have been attached to cod and used to obtain regionally explicit,

220 Nova Mieszkowska et al.
Author’s personal copy
individual-based data on horizontal and vertical movements and thermalhabitat. These studies support the contention that there is not a singleparadigm of extended movement between spawning and foraging areas, asgenerally supposed previously (e.g. Harden Jones, 1968), but rather thatsuch movements vary from individual to individual and from sub-stock tosub-stock (Hobson et al., 2009). Behavioural plasticity is evident for cod inthe extent and timing of migration, in the persistence of spawning orfeeding site fidelity (philopatry), and also in relation to the types of beha-viour displayed on feeding grounds and where and when they occur in thewater column compared with remaining close to the seabed (Hobson et al.,2007, 2009; Neat et al., 2006; Palsson and Thorsteinsson, 2003; Rightonet al., 2001).
Residence and homing behaviour have been shown to be importantfeatures of Atlantic cod behaviour (Hobson et al., 2009). Cod are known toaggregate seasonally to spawn and to feed at particular geographic locations(Metcalfe, 2006). For example, spawning area fidelity shown by aggrega-tions representing more or less distinct groups of fish is a behavioural traitsupported by at least some evidence from genetic and mark-recapturestudies (Metcalfe, 2006). However, the degree to which residence andhoming applies to different populations and to sub-stocks has been foundto vary greatly depending on geographic location. Robichaud and Rose(2004) proposed four categories of populations of Atlantic cod based on thedegree of migration and philopatry. The latter authors identified ‘sedentaryresidents’ that exhibit site fidelity year round, ‘accurate homers’ that returnto spawn in a specific area, ‘inaccurate homers’ that home to a much broaderarea around the original tagging location in the following years, and‘dispersers’ that move and spawn in a more irregular pattern within largegeographical areas (Metcalfe, 2006). It seems coastal areas support residentpopulations more commonly, such as those in the Norwegian fjords, theIcelandic coast and the Canadian east coast (Metcalfe, 2006), whereas theNortheast Atlantic has large subpopulations that home with accuracy com-pared with the Northwest Atlantic that has more inaccurate homers anddispersers (Metcalfe, 2006; Robichaud and Rose, 2004). Nested within thislarger scale complexity, are the variations in individual patterns observedwithin a region and which exemplify the problem with broad categorisationof cod behaviour.
Comprehensive studies of cod movements in the Northeast Atlantichave deployed over 3000 electronic tags in the Barents Sea, the North Sea,the Baltic Sea and on the Icelandic and Faroe Plateau between 2002 and2005, with over 850 tags returned by fishermen, giving more than 130,000days of data (www.codyssey.co.uk). In the North Sea, for example, it ispossible to link horizontal with vertical movement patterns. For individualtracks ranging in duration from 40 to 468 days, cod showed horizontalmovements up to 455 km, however, individuals did not always show signs

Climate and Fishing Effects on Cod 221
Author’s personal copy
of migration during winter months, even displaying continuous localisedresidence for up to 360 days (Hobson et al., 2009). This indicates that cod donot always migrate between feeding and spawning grounds. Vertical move-ments showed even greater flexibility, with a variety of movement patternsseen within both periods of residence and directed horizontal movement.Close association with the seabed was seen during both directed horizontalmovements and residency, while midwater oscillations in swimming depthwere also evident during both horizontal movement types. Therefore,vertical patterns in activity alone could not be used to reliably define periodsof migration or localised residence (Hobson et al., 2009). Taken together,the results from studies that have tracked large numbers of individual cod forlong periods suggest that Atlantic cod behaviour is mediated by complexinteractions between biological and ecological factors that result in diversemovements and activities in relation to changing environment (Hobsonet al., 2009; Righton et al., 2001).
1.4.2. Cod thermal habitsElectronic tagging data show that cod occupy depths from 10 to 860 m, andwater temperatures of�1.5 �C in polar fronts off Iceland and in the BarentsSea, to 21 �C when resting on the seabed in the southern North Sea.In terms of cod distributional responses to thermal habitat, such studiesreport, for example, that northern North Sea populations above 57 �Ndo not intermix with southern populations below 56 �N (Neat andRighton, 2007), thus concurring with previous mark-recapture pro-grammes (Righton et al., 2007; Robichaud and Rose, 2004; Wright et al.,2006) and genetic studies (Hutchinson et al., 2001). Within the northernNorth Sea, west Shetland is the warmest and least variable region (<3 �Cvariation) and no cod movement from here to cooler waters has beenrecorded. By contrast, cod released back into the east Shetland and VikingBank populations moved rapidly into cold fronts and prolonged occupancyof cooler waters was recorded (Neat and Righton, 2007). Within thesouthern sector of the North Sea, the German Bight population experiencesa highly variable thermal environment, with intra-annual variation of�14 �C. The greatest acute fluctuations recorded comprised a 7 �C decreaseover 3 days, and a 7 �C increase over a 2-day period. Individuals from thisregion, and the neighbouring eastern English Channel mostly migrated onlyshort distances, or remained resident, despite experiencing water tempera-tures up to 19 �C during late summer and autumn (Neat et al., 2006;Righton et al., 2001). Notably, adult cod only commenced vertical migra-tion in October–November once surface temperatures began to decline(Righton et al., 2001), and during this period some mature individuals havebeen recorded migrating to spawning grounds in the eastern English Channeland Southern Bight (Daan, 1978).

222 Nova Mieszkowska et al.
Author’s personal copy
Given the complex behaviour of cod, and in relation to thermal changesand other biological and physical factors operating across the broad range oftemperatures in which it is found, predicting how individual cod willrespond to climate-driven changes in sea temperature, for example, remainschallenging.
1.4.3. Larval dispersalGenetic evidence for reproductively isolated stocks that co-occur on groundsduring non-breeding periods, but that often segregate during spawningseasons, demonstrates the importance of knowledge of spawning and nurseryhabitats for appropriate management. Tests of population genetic differencesusing adult individuals have informed us about the general patterns of spatialand temporal stock structure, but such studies convey little about the relativeimportance of spawning grounds within spatial management units. Tradi-tionally, ichthyoplankton surveys and visual identification of larval speciesidentity have been used for identification of spawning grounds. Indeed, thisapproach has allowed broad-scale mapping across its range of cod spawningareas and the general pattern of egg and larval transport (Brander, 1997).Eggs and larvae, however, can be difficult to separate visually from thoseco-occurring gadoids such as whiting (Merlangius merlangus) and haddock(Melanogrammus aeglefinus) (Fox et al., 2005). Recent developments in molec-ular genetic techniques have enabled reliable identification of cod eggs,revealing, for example, considerable overestimation of cod abundance inthe Irish Sea (Fox et al., 2005; Taylor et al., 2002). This approach has alsoallowed the identification of active spawning grounds in the North Sea(Fox et al., 2008). Importantly, these locations, including the Dogger Bank,German Bight and Moray Firth show close corroboration with spawningareas inferred from historical survey data, implying cod have well-defined,active spawning grounds that have been used by multiple generations.However, to date there is very little available information on the extent oflarval dispersal or retention in relation to these spawning grounds, or the cuesemployed for location and settlement on nursery grounds. Hydrodynamicmodels coupled with in situ ichthyoplankton surveys for model verificationoffer considerable promise for revealing mechanisms of larval dispersal orretention over large spatial scales (van der Molen et al., 2007).
2. Impacts of Climate Change
Global climate has warmed to temperatures unprecedented over thelast 1300 years. Anthropogenic inputs into the atmosphere are now recog-nised as the primary driver (IPCC, 2007). The latest model predictionsindicate that global mean surface temperature will increase by a further

Climate and Fishing Effects on Cod 223
Author’s personal copy
1.1–2.9 �C (low emissions B1 scenario), or 2.4–6.4 �C (high emissions A1FIscenario) (IPCC, 2007). Climatic warming has been observed in marineenvironments across the North Atlantic, and appears to be of a greatermagnitude and duration than any periods in recent history (Fig. 3.3)(IPCC, 2001, 2007; Southward, 1963; Southward and Boalch, 1994;Southward et al., 1988). Marine ecosystems are already responding tothese changes in sea temperature, through polewards shifts in biogeographicranges (Beaugrand et al., 2002; Berge et al., 2005; Griffiths, 2003; Hellberget al., 2001; Mieszkowska et al., 2006, 2007; Zacherl et al., 2003), pheno-logical changes (Genner et al., 2009a; Sims et al., 2001, 2004), and throughalterations in the relative abundance of ectothermic species and the struc-turing of the communities they comprise (Barry et al., 1995; Berge et al.,2005; Genner et al., 2004, 2009b; Hellberg et al., 2001; Mieszkowska et al.,2006; Sagarin et al., 1999; Southward, 1995; Southward et al., 1988).Furthermore, there is burgeoning evidence for climate-driven effects onmarine fishes (Graham and Harrod, 2009). In this section, we explore therole of climate drivers on the biology and ecology of Atlantic cod and, inaddition to description and discussion of the known or potential biologicalimpacts, we identify knowledge gaps where new studies will progress ourunderstanding towards prediction of cod responses to changing environments.
Year
Mea
n an
nual
Nor
th A
tlant
ic S
ST
(�C
)
20.2
20.4
20.6
20.8
21
21.2
21.4
1870 1890 1910 1930 1950 1970 1990 2010
Figure 3.3 Mean annual sea surface temperatures of the North Atlantic (weightedaverage 5��5� grid squares from 0 to 70 �N). Data derived from the National Oceanicand Atmospheric Administration Kaplan SST data set, and are available at http://www.cdc.noaa.gov/data/timeseries/AMO/.

224 Nova Mieszkowska et al.
Author’s personal copy
2.1. Biogeographic changes
Distributional limits of marine fish species are governed to a large extent byregional thermal regimes. The observed thermal occupancy or ‘climateenvelope’ of species is the basis for many models that attempt to predictfuture abundance and distributions (Pearson and Dawson, 2003; Waltheret al., 2002, 2005). Where environmental conditions change to fall withinthe physiological tolerance limits of a species, range extensions are predictedas fish are able to colonize new areas of suitable habitat. In practice,however, the range edge may lie some distance inside this fundamentalniche envelope. This is because population distributions are often influ-enced by additional environmental parameters and biological interactionssuch as competition, predation and prey availability, parasitism and otherfactors such as habitat availability and dispersal ability (Brett and Groves,1979; Davis et al., 1997; Kelsch and Neill, 1990). There is still no generalmodel to describe how thermal physiology of ectotherms and climateinteract to determine biogeography (Chown and Gaston, 1999; Clarke,2003). Clearly, development of such models is a significant challenge forunderstanding and predicting the macroecological responses of fish species.
Data from commercial fishing and research vessel surveys have been usedto explore how the biogeography of cod has changed (Blanchard et al.,2005; Daan, 1994). However, use of abundance data from trawl surveys canbe insensitive to short-term individual variations in distribution (Neat andRighton, 2006) which can make range assessments for spatially structuredstocks problematic (Hutchinson et al., 2001; Metcalfe, 2006; Wright et al.,2006). Both migratory behaviour and density-dependent effects linked toprey abundance can affect geographic distributions (Beaugrand et al., 2003;Blanchard et al., 2003; Moyle and Cech, 2004; Roessig et al., 2004; Swain,1999; Swain et al., 2003). Moreover, other environmental variables such assalinity, storminess, cloud cover and precipitation can strongly influence thedistribution and productivity of marine ecosystems (Bakun, 1996; Stensethet al., 2004) and the phenology of production cycles (Edwards andRichardson, 2004). The stochastic nature of these parameters is reflectedin the annual fluctuations in abundance of cod across its range.
Despite the large diversity of factors that can influence distributions,there is compelling evidence that populations have shown responses toclimate-related thermal changes during the twentieth century. The NorthAtlantic warmed at the basin-scale during the 1920s and 1930s, and duringthis warm period the distributional limits of cod were observed to extendsome 1200 km further north from southern Greenland to Disko Island(northwest Greenland), while the Barents Sea population apparently shiftedeastwards (Hansen, 1949; Jensen and Hansen, 1931). Similarly, Icelandiccod were restricted to spawning on southern shelf regions until the1920s, but afterwards spread to the northern shelf (Sæmundsson, 1934;

Climate and Fishing Effects on Cod 225
Author’s personal copy
Vilhjalmsson, 1997). During a cool period during the 1960 and 1970s, thesechanges were seen to reverse. Cod evidently retracted further south in thecolder conditions and disappeared entirely from coastal waters around DiskoIsland (Buch and Hansen, 1988). Furthermore, the population spawning onthe northern Icelandic shelf declined to minimal levels during this period,presumably due to a shift in abundance centred further south. During thelate 1980s and early 1990s, cooler waters were also present in the NorthwestAtlantic from the Labrador Shelf to the Grand Banks, leading to a suddendecline in cod abundance (Atkinson et al., 1997; de Young and Rose, 1993;Drinkwater, 2002; Rose et al., 1994, 2000; Taggart et al., 1993).Whilst fishing activity may also have played a role in establishing this pattern(Hutchings, 1996; Hutchings and Myers, 1994b; Myers et al., 1996),analyses of blood chemistry (Rose et al., 2000) and genetics (Ruzzanteet al., 2001) also support a biogeographic shift of Northwest Atlantic stocksto lower latitudes at this time.
Apparent northward shifts of both the centre of distribution andthe southern range limit of cod in the southern North Sea in recent yearsare possibly a direct response of individuals to increased seawater tempera-ture over the last decade (Hedger et al., 2004; Perry et al., 2005; Rindorf andLewy, 2006). However, there is no direct evidence to suggest that fish haveactively moved to avoid increasing temperatures (Rindorf and Lewy, 2006).Studies supporting a northward shift in cod have based analyses on theassumption that there is single population in the North Sea that is mostabundant at the range centre, and has decreasing numbers of individualstowards range limits. For species that are more or less in a steady state, andare not changing in abundance or distribution rapidly, this pattern can bebroadly accepted. By contrast, where a single dimension of the nichechanges rapidly, and where there is evidence of population subdivisionand local adaptation, responses may not be straightforward. This may bethe case in the North Sea given some evidence that several discrete stocksare present (Hutchinson et al., 2001; Metcalfe, 2006; Wright et al., 2006),and which appear to have distinct habitat preferences during different lifehistory stages (Righton et al., 2007; Robichaud and Rose, 2004; Wrightet al., 2006). An additional consideration is that individual cod can movelarge distances or remain resident, and behaviour shows great flexibilityacross multiple spatio-temporal scales, resulting in complex spatial dynamics(see Section 1.4.1). Thus, there may not necessarily be a long-term locationfor occupation of the most suitable environmental conditions where thehighest abundance of fish can be found. The case of the North Sea is furthercomplicated by a seasonal inversion of the latitudinal temperature gradient,with the southern North Sea being colder in the winter but warmer in thesummer than the northern North Sea. Furthermore, analysis of North Searesearch survey data suggests that cod have responded to a winter bottomtemperature increase of 1.6 �C over 25 years by moving into deeper water at

226 Nova Mieszkowska et al.
Author’s personal copy
an average rate of about 7 m per decade (Dulvy et al., 2008). Together, thisevidence suggests that climate-driven changes to cod distributions may bemore complex than predicted using straightforward ‘climate envelope’approaches.
Observed northward shifts of the southern range limits could also beattributed to local population abundance changes due to fishing pressure,variation in migration between populations (Hedger et al., 2004) or spatialdifferences in the thermal tolerance limits of adult cod leading to localdepletion of stocks (Neat and Righton, 2006; Portner et al., 2008). Thelack of concordant responses to thermal regimes shown by individual cod isconsistent with temperature being only one of the factors determininghabitat choice (Neat and Righton, 2006). Occupancy of space by cod inhistorical habitats has declined from 90% to lower than 50% over the last30 years, and spatial distributions have become increasingly characterized byaggregations in areas of optimal thermal habitat (Blanchard et al., 2005;Horwood et al., 2006; Marshall and Frank, 1994; Myers and Stokes, 1989;Rose and Kulka, 1999). These studies support earlier observations ofdensity-dependent habitat selection in cod (Myers and Stokes, 1989;Swain and Sinclair, 1994), and strongly suggest changing distributionsmay additionally be linked to aggregation behaviour.
2.2. Physiology
Most fish are ectotherms with limited capacity for internal heat regulation(Clarke, 1993). To predict how changes in global climate will affect fishdistributions, it is important to know how physiological functions areinfluenced by temperature variation, and to quantify thermal tolerancethresholds (Guderley, 1990; Portner, 2001, 2002; Portner and Knust,2007; Portner et al., 2001). This individual-level physiological responseextrapolates to population, community and ecosystem-level responses(Roessig et al., 2004). Although temperature is recognised as an importantcontrolling factor for biotic processes, from cellular to ecological levels oforganisation (Fry, 1971), defining thermal optima is a complex process dueto differential effects of temperature on various physiological processes, andon different life history stages. For example, larval fish drifting passivelywithin the plankton may be more vulnerable than juvenile or adult fish thatcan actively move away from unfavourable conditions. Moreover, differentstocks have also been observed to show local adaptation to differing thermalranges (Coutant, 1977, 1987; Daan, 1994; Scott, 1982).
2.2.1. Tolerance limits and thermal preferencesBoth adult and larval cod can tolerate salinities from almost 0% to 35%, butexhibit some preference for 30–35%. They also have a wide thermaltolerance, and have been recorded in waters ranging from �1.5 to 21 �C,

Climate and Fishing Effects on Cod 227
Author’s personal copy
although their temperature range is typically between 0 and 12 �C (Trembleand Sinclair, 1985; Wise, 1961). Despite this apparent broad tolerance, theyare highly sensitive to even slight water temperature variation of �0.3 �C,and individuals seemingly alter their position in the water column tomaintain themselves near their temperature for optimal performance, Topt
(Fig. 3.4) (Herbert and Steffensen, 2005; Rose et al., 1994). Body temperaturehas a significant effect on fitness of cod (Fry, 1947; Huey and Berrigan, 2001),so Topt is expected to correspond to the temperature of maximum fitness,Trmax (Beamish, 1978; Schurmann and Steffensen, 1997). Temperature-fitness curves are asymmetric, however, and therefore body temperaturesgreater than Tmax can lead to stress and rapid fitness decline (Martin andHuey, 2008). The proximity of stressful temperatures to Trmax can result inTopt being lower than Trmax in natural, thermally variable environments. Thismay have important implications for predicting physiological fitness inresponse to future climate warming scenarios.
2.2.2. Sublethal physiological thresholdsIt may not always be possible for a fish to occupy a thermal environmentmatching the temperature of optimal performance, and thermal stress mayresult from exposure to unfavourable temperatures. The first symptoms ofthermal stress are caused by the limited capacity of respiratory systems toprovide sufficient oxygen to body tissue above the pejus temperature Tp
(pejus, meaning getting worse, deleterious) (Frederich and Portner, 2000;Portner, 2001; Portner et al., 2001). This is the threshold beyond which thecardiorespiratory system cannot increase aerobic metabolism and body
0Adult heart rate curve
5
10
15
20
25
Tem
pera
ture
(°C
)
×Highest pp venous O2
×Topt
(≈Tmax)adult cod
Toptg larval cod
Tp range adult cod
Tcrit range adult cod
Tcrit range juvenile cod
Figure 3.4 A schematic diagram of the thermal physiological thresholds of Atlanticcod. See Sections 2.2.1, 2.2.2 and 2.5 for explanations of terms and concepts.

228 Nova Mieszkowska et al.
Author’s personal copy
fluids start to become hypoxic. This has been determined experimentally tobe between 13 and 16 �C in adult cod (Fig. 3.4) (Sartoris et al., 2003), andbeyond this range cardiac arrhythmia, if present, can cause a reduction inblood circulation capacity. As a consequence, this results in lower venousoxygen concentrations, onset of anaerobic mitochondrial metabolism, alter-ation of enzymatic rates (Clarke, 1993; Coutant, 1987; Fry, 1971) and asudden decrease in intracellular pH (Van Dijk et al., 1997). This in turn isaccompanied by energetic collapse in white muscle (Foster et al., 1993;Sartoris et al., 2003), reduced scope for whole organism aerobic activity and,ultimately, death at the critical temperature threshold, Tcrit (Lannig et al.,2004; Portner, 2001; Sartoris et al., 2003). Experimental data suggest thatTcrit ranges between 16.0 and 22.2 �C in adult cod populations (Gollocket al., 2006; Lannig et al., 2004; McKenzie, 1938; Portner et al., 2008;Sartoris et al., 2003) and is lower at 15.5–18.0 �C in juveniles (Perez-Casanova et al., 2008; Yin and Blaxter, 1987), although survival uponacute exposure to 20 �C has been demonstrated in controlled laboratoryconditions (Perez-Casanova et al., 2008). This variation in response may bedue to different experimental methodologies, but it could also reflect stock-specific adaptation of cod (Portner et al., 2008).
The highest partial pressure of venous oxygen in southern North Seacod held in laboratory conditions occurs at approximately 5 �C (Lanniget al., 2004), which relates closely to Topt for adult growth. The frequency ofhaemoglobin genotypes in the population can be affected by environmentaltemperature experienced by parental fish, and underlies thermal preferencesin cod (Andersen et al., 2009). Warmer water preferences are associatedwith the Hb-1 genotype (Petersen and Steffensen, 2003), with highestfrequencies of the Hb-1–1 allele found in the southern North Sea(Husebo et al., 2004; Sick, 1965), a region of some of the warmest bottomtemperatures within the cod biogeographic range (Vaz et al., 2007).Although Topt of all haemoglobin genotypes appears to be centred around14 �C ( Jordan et al., 2006), optimal oxygen extraction rates occur below12 �C (Colosimo et al., 2003), which may explain why many cod popula-tions occur in waters below this temperature.
Thermal adaptation generally optimises whole animal aerobic scope towithin a thermal range or window (Portner, 2001, 2002). However, short-term thermal acclimatisation may also allow occupation of specific thermalregimes, such as the 2–3 �C sea surface isotherm characterising the seasonalmigration highway used by cod in the Northwest Atlantic (Rose, 2004b).Evidence for seasonal acclimatisation in cod is not widespread, but hasbeen provided by laboratory trials demonstrating that the thermal limitbeyond which heart rates begin to decline can be elevated by acclimatisationto warmer temperatures (Lannig et al., 2004). This suggests that cod maybe able to buffer the effects of climate-linked sea temperature warmingsufficiently well in the short term, however, as sea temperatures continue

Climate and Fishing Effects on Cod 229
Author’s personal copy
to rise in the region occupied, shifts in distribution may eventually takeplace that prevent exposure to temperatures beyond the pejus temperature.
2.3. Metabolic scope for activity
Metabolic rate is temperature -dependent (Claireaux et al., 2000; Lanniget al., 2004) and cod inhabiting temperate seas need to acclimatize metabol-ically to seasonal fluctuations in sea temperature. The costs of routinemetabolic activity are lower for individuals experimentally exposed tocolder water. In homogeneous water conditions, voluntary activity, meta-bolic rate and oxygen consumption all increase in response to a 2.5 �C risein temperature, leading to a subsequent decrease in scope for activity(Claireaux et al., 1995). When presented with a thermally stratifiedenvironment, however, behavioural changes are exhibited by cod, withindividuals swimming away from thermally stressful locations (Claireauxet al., 2000).
It appears that both swimming speed (Claireaux et al., 1995) and foragingrate (Peck et al., 2003) are also determined by thermal conditions. BetweenJune and August in the southern North Sea, decreased activity and predom-inantly benthic habitation were recorded from electronically tagged adultcod. When sea temperatures cooled, first nocturnal activity, then almostcontinuous activity took place during the following months (Righton et al.,2001). This pattern corresponds with expectations about energy conserva-tion (Arnold and Walker, 1992). When higher temperatures are encoun-tered it would be expected that they would either be avoided (O’Brienet al., 2000), and/or activity reduced, since occupation of higher tempera-tures will increase standard metabolic rate and reduce scope for aerobicactivity (Soofani and Hawkins, 1982; Soofiani and Priede, 1985). Growthalso decreases under such conditions, with individuals apparently switchingto a ‘translucent’ phase where an opaque band is formed during otolithgrowth (Pilling et al., 2007), indicating exposure to unfavourable tempera-tures (Hussy et al., 2004). Earlier onset and increased duration of thetranslucent zone of growth in recent decades has been attributed toincreased spring/summer water temperatures (Beckman and Wilson,1995), indicating that climate warming may be extending the period ofmetabolic stress (Pilling et al., 2007).
Sea surface temperatures have exceeded Topt for short periods duringrecent summers in the southern North Sea, for example. The subsequentdecrease in aerobic and locomotory performance will theoretically havebeen sufficient to impair activity such as to prevent optimal feeding in adultindividuals (Lannig et al., 2004; Portner, 2001; Portner et al., 2001). Thismay explain the seasonal absence of cod in the southern North Sea regiondespite apparent prey availability (Lannig et al., 2004). From these results ithas been suggested that if the climate continues to warm, seasonal

230 Nova Mieszkowska et al.
Author’s personal copy
disappearance of cod from some areas may increase in frequency andduration. However, these conclusions assume that Topt does not shift as aresult of acclimation to regional temperature regimes, which may not be thecase of course. Differences in compensatory capacity to specific temperaturechallenges are generally expected for populations inhabiting different watermasses or latitudes as a result of acclimation to local thermal regimes (Clarke,1993), so it is unlikely that specific thermal thresholds will be the same for allcod populations across the biogeographic range.
2.4. Maturation and spawning
Significant correlations between mean annual sea temperatures and age atmaturity have been found for many cod stocks ( Jorgensen, 1990, 1992;O’Brien, 1999; Yoneda and Wright, 2004). Together these results indicatea 1-year reduction in maturation age linked to a 2 �C increase in tempera-ture (Drinkwater, 2002). Hence, closer to the southern range edge of thespecies, maturation is predicted to be at a younger age. This pattern hasimplications for future stock success in warmer climates because smaller,younger fish are less fecund and spawn for shorter periods (Kjesbu et al.,1996), and warmer spring seasons may promote earlier maturation.
Climate-linked sea temperature changes may have direct and indirecteffects on cod energy provisioning and maturation processes that precedespawning. Cod build up energy reserves during summer and autumn, andmature during the winter months. Mature female fish in better conditionprior to spawning tend to be more fecund (Kjesbu et al., 1991), and expendless energy and lose less somatic mass during the spawning season (Lambertand Dutil, 2000; Lloret and Ratz, 2000; Ratz and Lloret, 2003). Hence,these fish are at less risk of subsequent natural mortality (Krivobok andTokareva, 1972; Love, 1958). When energy reserves are low, investment inreproduction by females may be maintained, but at a somatic cost, andreproduction may also be reduced or delayed in extreme conditions to limitsomatic loss. Changes in environmental conditions and subsequent ecolog-ical processes that negatively affect food intake will influence the energybudget and ultimately reduce productivity of the stock (Lambert and Dutil,1997). If fishing pressure and climate change act synergistically to reducethe age and condition of the spawning stock, population fecundity maydecline (Ottersen et al., 2006). Faster development of early life stages andsubsequent higher survival under warmer climatic regimes may, however,counteract any such decline in fecundity.
Peak spawning dates have been found to vary among different Norwe-gian coastal cod populations kept in identical environmental conditions(Ottera et al., 2006). This indicates that spawning time is under geneticcontrol, and could be an adaptation related to environmental conditions intheir source location. Consistent with this evidence, survey data show cod

Climate and Fishing Effects on Cod 231
Author’s personal copy
do not rapidly alter spawning time to match the timing of life history eventsin zooplankton prey species (Beaugrand et al., 2003). This suggests thatstock adaptation may result in a limited capacity for the stock to respondrapidly to climate-driven changes in peak abundance of prey. Thus, it canbe hypothesised that weak year classes following high larval mortality maybecome more common with warming sea temperatures.
Contrary to the assumed random mating strategy of many broadcastspawners, there is evidence for male lekking behaviour in cod (Robichaudand Rose, 2004;Windle and Rose, 2007), and for direct female mate choice(Engen and Folstad, 1999; Rowe and Hutchings, 2003; Rowe et al., 2004;Rudolfsen et al., 2005). In laboratory trials, both males and females displayedhigher reproductive success when mating occurred between larger indivi-duals (Rowe et al., 2007). The observed reduction in the size spectra of wildfish is thus likely to be affecting total reproductive output (Bekkevold, 2006;Bekkevold et al., 2002). Although climate will influence fecundity throughtemperature-related effects on maturation and reproductive success, size-targeted fishing mortality (of the largest fish) is one of the most likelydominant factors negatively affecting subsequent year class strength andrecovery of spawning stock biomass (SSB).
2.5. Early life stages
Cod eggs and larvae suffer high mortality (up to 99.9%) via predation(Campana, 1996; Cushing and Horwood, 1994; Houde, 1989; Leggettand Deblois, 1994; Shepherd and Cushing, 1980). The growth-predationhypothesis predicts a direct relationship between mortality rate and growthrate of early life stages of fishes (Anderson, 1988; Hare and Cowen, 1997),with more rapid egg and larval development promoting metamorphosis atan earlier age, thereby decreasing the duration of pre-juvenile stages(Drinkwater, 2005). Support for this theory has come from both experi-mental and simulation studies on early life stages of cod, and togetherthese show how small changes in early growth rates due to increases intemperature can lead to large increases in numbers surviving to recruitment(Chambers and Leggett, 1987; Houde, 1989; Meekan and Fortier, 1996;Miller et al., 1988; Pepin and Myers, 1991).
Cod eggs are found over a wide range of temperatures, from �1.5 �C inthe Northwest Atlantic, to 9 �C in the Celtic Sea (Geffen et al., 2006), andthere is evidence of local adaptation in development rates (Houde, 1989).Cod eggs from North Sea stocks cannot develop at temperatures less than1.5 �C, while larvae from the Baltic Sea can survive exposure to 1 �C, andeggs from the most northern populations will develop below 0 �C (Geffenet al., 2006; Valerio et al., 1992; Wieland and Jarre-Teichmann, 1997). Eggincubation periods can also vary significantly with temperature (Pauly andPullin, 1988; Pepin et al., 1997). Much of the observed seasonal variance in

232 Nova Mieszkowska et al.
Author’s personal copy
egg and larval development times (Pauly and Pullin, 1988) and hatching size(Miller et al., 1988) can also be attributed to thermal conditions, with sizedecreasing as temperature increases. Thermal tolerance experiments,together with knowledge of spawning locations, indicate that it is unlikelythat wild eggs become exposed to lethally high water temperatures imme-diately after release. Egg mortality is probably caused by other processesincluding predation and disease, but egg mortality due to sublethal effects ofincreasing energetic costs at high temperatures may also take place (Nissling,2004).
Optimal temperature for growth (Toptg) undergoes a clear ontogeneticshift in cod (Fig. 3.4) (McCauley, 1977; Reynolds, 1977). Yolk sac larvaehave the lowest Toptg ( Jobling, 1988) while the highest Toptg is for freeswimming larvae and juveniles ( Jobling, 1994; McCauley and Huggins,1979). This ontogenetic difference in Toptg can range from 2 to 11 �Cdepending on the local thermal regime inhabited by the source population(Brander, 1994, 2005; Buckley et al., 2004; Bunn and Fox, 2004; Nissling,2004), but is centred around 7 �C (Buckley et al., 2004). Larval growth ratesincrease as temperature increases between 4 and 14 �C (Caldarone et al.,2003; Laurence, 1978; Otterlei et al., 1999; Steinarsson and Bjornsson,1999) with time to metamorphosis decreasing from 56 days at 4 �C, to23 days at 14 �C (Otterlei et al., 1999). Laboratory estimates of growth ratesmay, however, be dictated in part by indirect thermal effects on foodlimitation, as was observed for Georges Bank cod larvae during an anoma-lously warm period of 1992–1994 (Buckley et al., 2004).
The slowest growing cod larvae are found both in the cold waters of theNortheast Arctic (Otterlei et al., 1999), and in warm water towards thesouthern end of the range (Buckley et al., 2004) including the southernNorth Sea (Pilling et al., 2007). Survival of larvae has been recorded attemperatures as high as 12 �C in the Irish Sea (Geffen et al., 2006). Earlyjuvenile cod from the Irish Sea (Geffen et al., 2006) and those from theNorwegian coastal population exhibit higher Toptg and are heavier at thesame life stage than individuals inhabiting colder waters of the NortheastArctic (G�do and Moksness, 1987; Otterlei et al., 1999). This indicates thatupper pejus (deleterious) temperatures have not been reached, and thattemperature-driven growth relationships seem to be population specific.
Investigations to date indicate that slight warming of the marine climateis likely to enhance egg and larval survival by decreasing the time taken toreach metamorphosis that in turn limits the temporal window of greatestpredation risk. Even stocks close to southern range limits in the centralNorth Sea are likely to show increased survival of early larval stages due towarming of between 1.5 and 4 �C during the twenty-first century (Hulmeet al., 2002). If predator–prey relationships remain unchanged it is possiblethat the survival of young fish may therefore increase. The lagged responsesof cod to climate warming may thus be driven by conditions affecting

Climate and Fishing Effects on Cod 233
Author’s personal copy
survival, growth and food availability during early life stages (O’Brien et al.,2000; Planque and Fredou, 1999; Platt et al., 2003).
2.6. Recruitment
Recruitment success of fish can be dictated by extrinsic stochastic eventssuch as changes in temperature, winds, currents, food availability, preda-tion/parasitism, and intrinsic factors such as variation in adult condition,stock reproductive effort and age structure (Cushing, 1996; Heath andGallego, 1997; Houde, 1989). These factors subsequently affect cod pro-duction, egg viability and survival of the early life stages (Kjesbu et al., 1996;Marshall et al., 1998; Nissling et al., 1998), as well as primary and secondaryproduction of the whole ecosystem (Hooper et al., 2005).
Recent studies indicate that the dominant pattern of recruitment varia-tion may be related to an effect of climate-driven sea temperature changes(Brunel and Boucher, 2007). Such climate-related changes in recruitmentsuccess may occur through one or more of several potential mechanisms,including higher production or survival of pelagic eggs or larvae (Rijnsdorpet al., 2009). Temperature plays a key role in the variation of cod recruit-ment success through a combination of direct and indirect effects (Cushing,1996; Hermann et al., 1965; Ottersen and Loeng, 2000; Sætersdal andLoeng, 1987). Fecundity is lower in mature individuals from northerncod stocks inhabiting colder waters, but given there is no evidence forcompensation through an increase in egg size (Brander, 1994), it appearsthat the cold-induced shift in the energy budget is unfavourable for repro-ductive output (Portner et al., 2001). This is supported by observations ofstrong year classes during warmer years in northern cod stocks (de Youngand Rose, 1993; Drinkwater, 2005; Malmberg and Blindheim, 1994;O’Brien et al., 2000; Ottersen, 1996; Ottersen and Sundby, 1995;Ottersen et al., 1994; Sundby, 2000).
Interannual variation in recruitment success is strongly dependent onseasonal temperatures (Wieland et al., 2000). Temperatures between Februaryand June have most impact on recruitment and subsequent year classstrength in the North Sea. A rise of 0.25 �C has been linked to a 30%reduction in recruitment (Clark et al., 2003). Simulations using the UKHadley Centre HadCM3 climate model reveal an inverse relationshipbetween change in abundance of 1-year-old cod and sea surface tempera-tures the previous spring, implying that climate effects on life stage is key tolater population recruitment success (O’Brien et al., 2000; Planque andFredou, 1999). A broad relationship between temperature regimes andstock success has been proposed by Drinkwater (2005), who suggestedthat recruitment increases as bottom sea temperatures increase until 5 �C,with little subsequent change until 8.5 �C, before a continual decline athigher temperatures. Based on these conclusions, a 2 �C increase in sea

234 Nova Mieszkowska et al.
Author’s personal copy
temperature could result in significant declines in cod abundance in theNorth Sea from recent levels, with local extinctions being more likelybeyond a rise of 3 �C (Drinkwater, 2005). This model assumes that theslope of the temperature–recruitment relationship does not vary betweenstocks, does not account for seasonal variation in temperature and does nottake into account fishing mortality, or water column profile and occupancy.Indeed, such factors may explain why temperature–recruitment relation-ships are often weak (Brander, 2000). O’Brien et al. (2000) found nostatistically significant link between environmental temperature and recruit-ment in Northwest Atlantic stocks during the 1980s and 1990s, and re-analyses of recruitment data sets tend to confirm this result (Frank, 1997;Myers, 1998; Myers and Cadigan, 1995a,b; Myers et al., 1995a). In contrast,predictions of climate change impacts on recruitment and SSB in the NorthSea have been constructed using a model that includes fishing mortality,temperature-dependent growth rates, and a temperature-dependent Rickerstock–recruitment function (Clark et al., 2003). This model indicates thatsea temperature affects population dynamics via recruitment rather thanadult growth. Moreover, the model supports the hypothesis that Februaryto June sea surface temperatures most strongly influence recruitment(Dickson et al., 1974; O’Brien et al., 2000; Planque and Fredou, 1999).Under an unchanging climate scenario, the Clark et al. (2003) modelsuggests that both SSB and recruitment should increase over the next50 years if fishing mortality remains constant. However, even under asmall forcing of the climate by þ0.05 �C per decade, which is much lessthan the 1 �Cwarming that has already occurred in UK coastal seas since themid-1980s (Hawkins et al., 2003), SSB and recruitment are predicted todecrease. Under the A1F high emissions scenario (IPCC, 2001) of þ0.26 �Cper decade, North Sea stocks are predicted to virtually disappear if fishingmortality is not reduced (Clark et al., 2003). However, the model has caveats.It is based on linear models that do not fully capture the relationship betweenrecruitment and temperature, and it also does not account for potential rate-limited recruitment at higher temperatures. There is also a lack of spatialrepresentation of adult distributions, including potential for changes inspawning locations over time (Brander, 1994, 1997; Daan, 1978).
Cod recruitment patterns may also reflect temperature-driven variabilityin availability of food resources at lower trophic levels (Rothschild, 1994).It has been demonstrated that peak spawning date in low biomass codpopulations shows strong associations with temperature (Kjesbu et al.,1994; Wieland et al., 2000). In the Northwest Atlantic, higher temperaturesappear to result in earlier annual spawning dates in stocks occupying higherlatitudes, due to accelerated gonad development (Hutchings and Myers,1994a,b). The most northern stock in the Barents Sea also shows a positiverelationship between recruitment and temperature. The opposite relation-ship was observed in stocks located at lower latitudes towards the southern

Climate and Fishing Effects on Cod 235
Author’s personal copy
limits in the North Sea (Daan, 1994; Dickson et al., 1974; Myers, 1998).These relationships were subsequently re-tested and shown not to hold,emphasizing that studies of recruitment may need to step beyond searchingfor straightforward spawner–recruit and temperature–recruitment relation-ships (Myers, 1998).
In the North Sea, changes in the plankton community have been a keydriver of interannual fluctuations in cod dynamics. Long-term changes inrecruitment co-vary with changes in the abundance and body size of zoo-plankton prey with a 1-year time lag, and show a tighter relationship than thatobserved between cod recruitment and sea surface temperature (Beaugrandet al., 2003; Horwood et al., 2006). The relationship holds for both anoma-lously cool and warm periods, such as the gadoid outburst between the mid1960s and 1980s when cool temperatures were associated with high abun-dances of the boreal copepod Calanus finmarchicus. This copepod is a majordietary component of the cod early life stage, and these high abundancesoccurred in parallel with 12 years of high cod recruitment (Fig. 3.5).Conversely, during the warm period of the late 1990s and early 2000s,anomalously low cod recruitment reflected low biomass of C. finmarchicus.Warmer waters appear to have resulted in unfavourable conditions for theoverwintering stage of this copepod (Greene et al., 2003; Heath et al., 1999)and forced a distributional shift to higher latitudes in the North Sea(Beaugrand and Ibanez, 2004). Higher confidence can be placed on theassumptions of these shifts in biogeographic distribution due to the extensivespatial and temporal coverage of the data, and the use of direct observationaldata rather than extrapolations based on putative centres of distribution.
Direct effects of thermal regimes on physiology of larval fish, andindirect effects on their prey availability, are both likely to be importantdrivers of recruitment strength. However, the exact nature of the
1958
Pla
nkto
n ch
ange
(in b
lack
)
1961
1964
1967
1970
1973
1976
1979
1982
1985
1988
1991
1994
1997
6.1
5.9
5.7
5.5
5.3
Total cod biom
ass1-year lag
(logarithmic scale)
86420
Regimeshift
−2−4−6−8
Figure 3.5 Parallel long-term changes in zooplankton fluctuations (with 1-year lag)and cod recruitment in the North Sea. Adapted from Beaugrand et al. (2003).

236 Nova Mieszkowska et al.
Author’s personal copy
relationships remains unclear. The abundance and size structure of a popula-tion will depend on a range of factors, including the extent of immigrationand emigration from populations with different thermal tolerances (Simset al., 2001, 2004). In the absence of gene flow, adaptation or local acclima-tization may lead to a contraction of the spawning period and spawning area,causing fish such as cod to become more sensitive to changes in environmen-tal conditions (Ottersen et al., 1994). Heavily exploited populations of codmay therefore display amplified responses to climate change (Planque andFredou, 1999). These complications may lead to difficulties when predictingshort-term outcomes of environmental change on populations (Planque et al.,2003; Rothschild, 1994, 1998).
Biological responses may show strong relationships with large-scaleclimate indices such as the North Atlantic oscillation (NAO) (Broitmanet al., 2008; Drinkwater et al., 2003; Fromentin and Planque, 1996;Ottersen et al., 2001; Stenseth et al., 2002; Walther et al., 2002). Suchclimate indices often incorporate multiple variables and the interactionsbetween them, and thus can capture a complex interplay of weather andclimate-induced variations in the natural environment. As a consequence,they can provide useful assessments of climate fluctuations with which toexplore ecosystem change (Namias and Cayan, 1981; Stenseth et al., 2003).The NAO is the main index of winter atmospheric circulation over theNorth Atlantic. During positive NAO years, warmer winters occur andseawater temperatures are warmer around Northwest Europe. When theNAO switches to a negative phase, winter temperatures are colder in theregion (Hurrell and Van Loon, 1997; Van Loon and Rogers, 1978). Overthe last 25 years, the frequency and magnitude of NAO positive-indexevents have increased and winter sea surface temperatures have becomemilder in British coastal waters. The NAO is predicted to remain in a largelypositive phase in the coming decades.
Relationships between the NAO index, the physical environment of theNortheast Atlantic and biological responses within it have been well estab-lished (Broitman et al., 2008; Hurrell and Van Loon, 1997; Reid et al., 2001;Sims et al., 2001, 2004). Importantly, the NAO index has been linked tochanges in recruitment success of most cod stocks in this region (Ottersenet al., 2001), mainly via the direct effects of environmental temperature onsub-adult growth and survival (Attrill and Power, 2002; Dippner, 1997;Ottersen et al., 1994). When all Northeast Atlantic cod stocks are combinedwithin a single recruitment model, a significant geographic relationshipbetween the strength of the NAO and recruitment emerges, with stock-specific trends also apparent (Stige et al., 2006). Environmental conditionsduring a positive NAO index have been shown to have a negative influencefor southern stocks on both seaboards of the Atlantic, but a positive influ-ence on more northerly stocks (Brander and Mohn, 2004).

Climate and Fishing Effects on Cod 237
Author’s personal copy
2.7. Growth
Experimental studies and analyses of catch statistics show that changes inseawater temperature exert a major influence on growth (Bjornsson andSteinarsson, 2002; Bjornsson et al., 2001; Brander, 1995, 2000; Campana,1996; Campana and Hurley, 1989; Jobling, 1988; Solberg and Tilseth, 1987;Steinarsson and Bjornsson, 1999; Taylor, 1958). As such, temperature isconsidered to be a major determinant of production in cod stocks (Dutiland Brander, 2003). Benthic water temperatures have been shown to accountfor 90% of observed variation in growth rates between cod stocks (Brander,1994, 1995) and to drive annual growth fluctuations within them (Brander,1995; Brander et al., 2003; Campana et al., 1994; Clark et al., 2003; deCardenas, 1996; Drinkwater, 2005). This temperature-dependent variabilityin growth rates is mediated by immediate physiological requirements, andtrade offs between growth and reproduction (Portner et al., 2001).
Free swimming cod tend to select temperatures in which growth rate ismaximised (Claireaux et al., 1995; Magnuson et al., 1979), but it is not alwayspossible for individual fish to maintain themselves within thermally optimalhabitats. Low temperatures result in slower growth rates and a reduction in thephysical condition via direct impacts on rate of food assimilation, and indirecteffects on food supply (Otterlei et al., 1999). The resultant physiological andecological impacts are manifested as a small size-at-age, reduced cohort size, adecline in stock biomass, and surplus energy redirected into reproductiveeffort (Brander, 1995; Campana, 1996; Krohn et al., 1996; May et al., 1965;Taylor, 1958). Meta-analyses of cod populations show that body conditionexhibits a significant increase with warmer mean sea bottom temperatures(Drinkwater, 2005; Ratz and Lloret, 2003). The temperature for optimalgrowth, however, decreases with size and age, and ranges between 14.3 and17.0 �C for newly hatched juveniles, to between 5.9 and 10.0 �C for adults(Bjornsson and Steinarsson, 2002; Bjornsson et al., 2001; Brander et al., 2003;Buckley et al., 2004; Portner et al., 2001). Thus, decreasing growth perfor-mance in adult cod is observed with increasing latitude (Brander, 2004;Portner et al., 2001). Genetic differences between stocks are also likely to beat least partly responsible for the observed spatial differences, as populationdifferences in growth rates have been seen under controlled experimentalconditions (Portner et al., 2001). Such population variation in growth rates andbody size may also be a consequence of fishing, as sustained removal of largerindividuals has resulted in evolutionary selection for individuals maturing atearlier ages and sizes (Gadil and Bossert, 1970; Olsen et al., 2004).
Although studies indicate that as seawater temperatures rise, cod inwarmer climates may experience increased growth rates, such increasesmay be counteracted by food availability, which can explain up to 97%of the variance in growth in wild cod (Chabot and Dutil, 1999). Food-dependent growth rates may become particularly apparent because

238 Nova Mieszkowska et al.
Author’s personal copy
exposure to warmer water increases standard metabolic rates. Such negativeeffects on growth may be intensified in conditions of low prey density, asthe temperature of optimal performance can become lowered (Brett andGroves, 1979; Buckley et al., 2004). Since growth and survival of larval fishare implicit to many recruitment hypotheses (Anderson, 1988; Cushing,1990; Ware, 1975), and are affected by climate-driven sea temperaturechanges (Rijnsdorp et al., 2009), predictions of growth rates under futureclimates will clearly require data on prey abundance, and information onprey distribution and sub-adult feeding behaviour (Buckley et al., 2004;Dower et al., 1998; Fiksen and MacKenzie, 2002).
3. Impacts of Fishing
Marine fishing activity in the North Atlantic can be traced back over atleast 1000 years (Barrett et al., 2004, 2008), and there is compelling evidencethat this has severely depleted demersal fish stocks in the region, reflectingglobal trends. Biomass of commercially valuable demersal fish populations isnow estimated to be at only 10% of pre-industrial levels (Worm and Myers,2003), and there has been a concomitant decrease in mean trophic level oflanded fish (Pauly et al., 1998). Atlantic cod abundance in particular hasdeclined dramatically since the onset of commercial fishing. Using historicalrecords of New England cod abundance derived from mid-nineteenthcentury fishing logs, it was estimated that current population biomass nowstands at less than 5% of that in 1852 (Rosenberg et al., 2005). Reconstruc-tions using historical records or proxies have also revealed high levels offisheries exploitation affecting key biological parameters of cod. For exam-ple, by studying cod vertebrae preserved in middens in New England, largedeclines in population body size distributions have been revealed ( Jacksonet al., 2001), changes that have links to the biological effects of commercialfishing (Olsen et al., 2004).
In this section, we describe the impacts of fishing on cod populations ineach of the main regions across its biogeographic range, and bring togetherthese findings with those on climate impacts to evaluate the relative contri-bution of each set of drivers to observed trends.
3.1. Northwest Atlantic stocks
All cod stocks are now generally considered to be either fished at unsustainablelevels, are subject to moratoria following dramatic stock collapses, or haverecovery plans that do not meet ‘precautionary approaches’ advised by theInternational Council for the Exploration of the Seas (ICES) (CEC, 2001;FAO, 2002; Hutchings and Myers, 1994a,b; ICES, 2005, 2006, 2008;

Climate and Fishing Effects on Cod 239
Author’s personal copy
Kuikka et al., 1999;Myers et al., 1997). Arguably themost dramatic collapse ofcod was that seen in the Northwest Atlantic. During 1986 and 1987 recruit-ment was very strong in all cod stocks in this region. As a consequence, fishingmortality (F) increased from 0.5 to>1.0 between 1989 and 1992 for all stocks,except that of the southern Grand Banks (Bishop et al., 1993; Myers et al.,1996). Landings initially increased as SSB was decreasing, a pattern thought tohave been caused by fish at lower population biomass tending to aggregate,perhaps making them easier to capture (Hutchings, 1996; Morgan et al., 1997;Rose and Kulka, 1999). Whatever the mechanism, catches were soon domi-nated by young and small cod (Hutchings and Myers, 1994a,b; Myers et al.,1996). It has been argued that the main factors responsible for the collapseacross many of the cod stocks by 1993 were the ignorance of the relationshipbetween fishing mortality and stock biomass, due to consistent underestima-tion of the proportion of fish harvested annually (Myers et al., 1996), and anunwillingness to cut fishing effort due to the perceived short-term economicconsequences (Rivard and Maguire, 1993; Schiermeier, 2002). Evidenceindicates that incorrect calculations of fishing mortality resulted fromoverestimation of biomass (Steele et al., 1992; Walters and Maguire, 1996),an underestimation of reductions in productivity (Rice and Evans, 1988),overweighting of abundance indices to provide the most optimistic estimatesof SSB (Myers et al., 1996, 1997) and an increase in efficiency of the fishingfleet (Hilborn and Walters, 1992; Walters and Maguire, 1996). No evidencewas found to link abundance of age classes or distributions with water temper-ature, and thus climate change was rejected as a primary driver for the collapseof these cod stocks (Hutchings and Myers, 1994a,b). A decade after themoratorium on cod fishing was introduced in 1992, populations were still athistorically low abundance (Lilly et al., 2003), and even in 2007 the SSBcontinued to have weak representation from all year classes (STECF, 2007).
3.2. Northeast Atlantic stocks
Atlantic cod have a broad distribution on the European continental shelf. Inthis section, however, rather than review stocks in all areas, we focus onthree areas with contrasting habitats and different fates of stocks. First, wedescribe how fishing has impacted cod in the North Sea, a region wheresome of the largest declines have occurred, we then compare this to changesin the Celtic Sea stock, where, in general, cod are found at relatively lowabundance. Finally, we discuss the status of cod in Icelandic waters, wheremanagement structures are different with respect to these other areas.
3.2.1. North SeaThe entire North Sea has been commercially fished since the mid-eigh-teenth century (Cushing, 1988; Smith, 1994), with periods of cessationduring the two World Wars (Beverton and Holt, 1957; Borley, 1923;

240 Nova Mieszkowska et al.
Author’s personal copy
Hardy, 1959; Margetts and Holt, 1948). Catchability of cod was high duringthe late 1940s and 1950s, but began to decline in the 1960s as increasedlevels of exploitation caused dense aggregations to be fished out (Gulland,1964). During the ‘gadoid outburst’ of the 1970s, high recruitment led tomassive increases in juvenile cod abundance, with estimated stock sizes of250,000 tonnes (Brander, 1995; Cushing, 1984; Hislop, 1996; Horwoodet al., 2006). This period was associated with cooler waters and increases inthe abundance of the cold water copepod C. finmarchicus, a primary foodsource for pelagic larval cod (Beaugrand et al., 2003). Landings rose until theearly 1990s, then declined before levelling out at record low levels after2003 (Holden, 1978; ICES, 2007; Jennings et al., 1999; Olsen et al., 2004;Rijnsdorp and van Leeuwen, 1996; Rijnsdorp et al., 1996; Fig. 3.6). Theyear class of 1996 was anomalously strong, but it was removed by fishingbefore reaching maturity (Bannister, 2004), indicating that fishing impactson stock dynamics were far outweighing those of climatic variability. Size ofindividual adults also declined across the twentieth century in response toincreased fishing mortality of larger, older cod (Fig. 3.7).
North Sea cod has been managed under joint management agreementsinvolving annual assessments by the European Council of Ministers and theNorwegian government since 1974 (Reeves and Pastoors, 2007). The definedlimits of theNorth Sea include the Skaggerak and easternEnglishChannel, andall cod are treated as one stock for ease of management despite some geneticevidence for multiple populations within the region (Blanchard et al., 2005;
0
50
100
150
200
250
300
350
400
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Year
Est
imat
ed w
eigh
t (in
thou
sand
s of
tonn
es)
Discards
Landings
Figure 3.6 Total estimated catch (in thousands of tonnes) of cod from the North Seasince 1985 including landings and discards. Data: ICES (2008).

Figure 3.7 High numbers of large cod being landed on the fish market at Aberdeen,Scotland. Photograph ca. 1920.
Climate and Fishing Effects on Cod 241
Author’s personal copy
Hutchinson, 2008; Hutchinson et al., 2001; STECF, 2007). The assessmentand total allowable catch (TAC) advice process is conducted under anICES umbrella, and primarily reflects the needs of the main customer, theCommission of the European Communities (Holden, 1994).
TACs have been restricted for the last two decades to allow an estimated30% increase in spawning biomass, but also whilst falling within 15% of theprevious year’s TAC. Data from both research vessel surveys and fisherieslandings data suggest that TACs have been ineffective. By 1992, only 4% ofNorth Sea cod were surviving to maturity at 4 years old due to intensiveharvesting across the year classes (Cook et al., 1997), and the most com-monly caught age classes over the last two decades were immature 1 and2 year olds (Myers et al., 1996; O’Brien et al., 2000). By 2000, SSB hadfallen to around 40,000 tonnes (ICES, 2000), well below the ICES mini-mum biomass limit of 70,000 tonnes, at which stock production wasconsidered as being severely impaired (Horwood et al., 2006). Conclusionsfrom cod recovery scenarios suggested that even a reduction in fishingmortality to a precautionary F ¼ 0.65 per year provided a low probabilitythat stocks would recover to a precautionary biomass level within a decade(STECF, 2007). Moreover, these models did not take into account climatechange in the projections. A narrow window between ‘potential’ and ‘no’recovery was highlighted (Horwood et al., 2006), which may explain whyreductions in fishing effort close to precautionary levels have failed toincrease stock biomass. Repeated recommendations from ICES of eithertotal closure or reductions in fishing mortality of up to 70% duringthe 2000s have resulted in annual re-calculations of TACs and reductions

242 Nova Mieszkowska et al.
Author’s personal copy
in the number of days at sea for gill-netting vessels and beam trawlers in the70–99mmand>100mmmesh sectors (ICES, 2003, 2004, 2006, 2007, 2008).
A recovery plan involving TACs to allow a 30% increase in biomass wasfinalized in 2004 for the North Sea, Kattegat, West Scotland and Irish Seastocks. A year later, ICES announced that the recovery plan for cod was notprecautionary (ICES, 2004) and by 2006 the North Sea and NorthwestRegional Advisory Committees had found little evidence of recovery, withstocks still at or close to historically low levels (NSRAC, 2006). Althoughthere have been some indications of improved recruitment with a relativelystrong year class in 2005, no increase in SSB has been observed to date.ICES altered their recommendations from ‘lowest possible catch’ in 2002 to‘zero catch’ from 2004 to 2008 (ICES, 2003, 2004, 2006, 2007, 2008).Interestingly, North Sea cod were listed as being ‘harvested sustainably’ byICES in 2008, but these stocks in 2009 have been categorized as ‘overf-ished’, with fishing mortality ‘above target’ and a recommendation of ‘zerocatch’ (ICES, 2008).
3.2.2. Celtic SeaCod have also been targeted by both single- and mixed-species fisheries inthe Celtic Sea for several centuries (Kurlansky, 1997), but intensive com-mercial fishing began relatively recently in comparison with other NorthAtlantic regions (Pinnegar et al., 2002). Recent assessments have valuedlandings made by the combined international fleet to be approximately£10.5 million per year, with maximum landings occurring during wintermonths (Fisheries Science Services, 2008; ICES, 2006).
Dwindling numbers of pelagic and demersal fish, including cod, in thecoastal Celtic Sea and elsewhere around the UK were of concern as early asthe mid-nineteenth century (Sims and Southward, 2006). During theperiod of modern data collection and assessments, fishing mortality in theCeltic Sea has remained at very high levels since the mid-1980s (FisheriesScience Services, 2008; Worm and Myers, 2003) albeit with a slightdecrease in recent years due to a reduction in the section of the fleettargeting cod since 1999 (ICES, 2006). The stock has been well belowsafe minimum limits since 2004, and even with the recent reduction infishing effort, biomass is continually declining (ICES, 2006, 2008). Despitedeclining stock size and low projected recruitment if present fishing effort ismaintained, a successful management strategy has yet to be put into practicein the Celtic Sea region.
The Celtic Sea cod stock is located close to the southern biogeographiclimit of the species in the Northeast Atlantic. The fish have relatively fastgrowth rates and mature at 2–3 years of age (Armstrong et al., 2001; ICES,2003). Recent poor recruitment rates have been purported to be driven inpart by warming seas, but there is little evidence for climate-driven changesin the biogeographic range or abundance of this stock, and thus fishing is

Climate and Fishing Effects on Cod 243
Author’s personal copy
still recognized as a primary factor determining SSB. In addition to species-level impacts, it is notable that significant declines in the mean trophic levelof catches overall have been observed from the region, consistent withfishing-induced changes to the structure of the whole fish community(Pinnegar et al., 2002).
3.2.3. IcelandThe Icelandic cod stock is one of the largest in the North Atlantic, with amaximum sustainable yield (MSY) of 330,000 tonnes providing annualrevenue in excess of £400 million. The stock is managed as a single unit,although there are discrete regional spawning components (Begg andMarteinsdottir, 2000). Stock size declined throughout the twentiethcentury from a peak of 3.3 million tonnes in 1928 to 600,000 tonnes by1993 (Schopka, 1994), accompanied by a decline in SSB to 120,000 tonnes(Marine Research Institute, 2008). A low stock size was recorded in 2007 ascatches have continued to exceed advised levels, although SSB has shown anincrease in recent years to approximately 230,000 tonnes (Marine ResearchInstitute, 2008). Mean weight at age has decreased significantly to an all-time low in 2007 and 2008, attributed to the lack of capelin as a food sourcein recent years. This weight-at-age trend, in combination with recent poorrecruitment, has led to very low productivity of the current stock (MarineResearch Institute, 2008).
Three so-called ‘cod wars’ have taken place in the waters off Icelandduring the latter half of the twentieth century, in 1958, 1972 and 1975.These hostilities arose as a result of extensions in the offshore spatial extentof the Icelandic fishery exclusion zone and a lack of recognition of thisexpanding zone by vessels from European countries already fishing in thesewaters. The first conflict resulted in an extension of Iceland’s territorialwaters to 12 nautical miles from the coast. Further extensions of theIcelandic fishing zone to 50 nautical miles in 1972 and 200 nautical milesin 1975 resulted in more severe international conflicts. The final confronta-tion was resolved when intervention by the North Atlantic Treaty Organi-zation (NATO) succeeded in brokering an agreement that reduced access toBritish vessels within the 200 nautical mile limit, including closure of fourconservation areas to British fishing activities. Various Icelandic govern-ment acts have since been introduced in an attempt to reverse the trend ofdeclining fish stocks within the 200 nautical mile limit, including TheFisheries Act of 1976 (amended in 1983), Individual Effort Restrictions in1977, The Fisheries Management Act 1990, the Harvest Control Rule in1995 and the current system of TACs. Despite these measures, stock size hasshown no signs of recovery to sustainable levels.
In the early decades of the twentieth century, stock size was stronglycorrelated with environmental changes in the Iceland–Greenland region.The coastal current and Atlantic inflow are thought to exert a strong

244 Nova Mieszkowska et al.
Author’s personal copy
influence on annual success of each spawning component via alterations inthe strength of water flow from the main spawning grounds off the north-west coast of Iceland to the main nursery grounds off the north coast (Beggand Marteinsdottir, 2002). Pelagic juveniles are larger and more abundantduring years of stronger current flow and Atlantic inflow, due to bothincreased dispersal and increased abundances of the main prey species ofzooplankton, C. finmarchicus that is brought into the region by the Atlanticinflow (Sundby, 2000). As fishing mortality has steadily increased from 0.16to approximately 1.0 between the 1930s and 2000s, it has exerted anincreasingly dominant influence on the annual stock size of Icelandic cod.Stock models that incorporate both environmental and fishing componentsindicate that substantial reductions in fishing effort are required to permitstock recovery (Baldursson et al., 1996).
3.3. The fishing versus climate change debate
Historically high abundances of North Sea cod during the early 1900s(Eckman, 1953), 1960s and 1970s (Cushing, 1984) have been linked tocooler marine climates, while the recent period of rapid climate warminghas been suggested to have contributed to the observed declines in SSB(Blanchard et al., 2005; Clark et al., 2003; O’Brien et al., 2000; Ottersenet al., 2006; Schiermeier, 2004). During recent decades it is possible thatclimate signals have been overridden by fishing impacts, and recent warmerclimates have exerted additional pressures on already stressed stocks(Graham and Harrod, 2009). However, separating the interactions betweenthe two drivers has proved difficult. Thus, quantifying strengths of compo-nent effects will be challenging, especially between regions that differ incommunity compositions, thermal regimes and habitat structure, and also incod abundance, distributions and behaviour (Neat and Righton 2006;Righton et al., 2001). Despite this, there is a need to more fully understandthe contributive effects of climate change on cod stocks that are already athistorical low abundance due to overexploitation.
Biological responses to environmental changes such as fishing andclimate, for example, can be divided into two categories. Firstly, proximateecological responses that depend upon relationships between physicalfactors and organismal-level processes, population dynamics and commu-nity structure (Harley et al., 2006). Secondly, direct impacts on individualperformance during various life stages through changes in physiology,morphology and behaviour (Mieszkowska et al., 2006, 2007). Theseimpacts lead to population-level responses, which can be additionallyaffected by climate-driven changes in hydrographic processes that affectdispersal of the pelagic larval life stages and recruitment. All of these arelikely to lead to alterations in cod distributions, biodiversity, productivityand micro-evolutionary processes.

Climate and Fishing Effects on Cod 245
Author’s personal copy
4. Population-Level Impacts of Fishing
and Climate Change
Shifts in survival, maturity, fecundity, reproduction, recruitment suc-cess and growth all scale up to the population level. If sufficient numbers ofindividuals display the same response to an external driver, such as climate,then population dynamics are likely to be altered. Such effects of climate arelikely to be observed first within populations located close to the distribu-tional limits, where environmental conditions are most likely to be alreadyapproaching stressful levels (Lewis, 1986, 1996; Orton, 1920; Southward,1995). By contrast, effects of fishing mortality may well affect populationsacross much of their range.
4.1. Stock assessment
SSB is the parameter used to define the size of exploited fish stocks. Bothshort- and long-term variability in SSB may be driven by natural age-structured interactions, including competition between year classes, canni-balism and natural mortality (Bjornstad et al., 1999). The SSB–recruitmentrelationship may be contextualized with respect to underlying environmental,ecological and biological processes that control growth, reproduction andmortality. Supporting this idea, inclusion of age structure data can help toimprove SSB–recruitment models (Marteinsdottir and Thorarinson, 1998).Different age classes may contribute unequally to reproductive output and aspawning stock with a more diverse age structure may exhibit protraction ofthe spawning period due to size or age-dependent factors (Hutchings andMyers, 1994a,b; Marteinsdottir and Petursdottir, 1995). The most commonmethod of determining the spawner–recruitment relationship is the applica-tion of a model such as the Ricker or Beverton–Holt model to a single stock(Myers et al., 2001). Unfortunately, the stock–recruitment curve that isgenerated can be simplistic, and the effects of biological and environmentalfactors can strongly affect this relationship leading to large variations inrecruitment (God�, 2003; Marshall et al., 1998, 2000).
The existence of a relationship between abundance of spawners andnumber of recruits is implicit to SSB calculations (Hilborn and Walters,1992; Myers and Barrowman, 1996), but within-stocks data provide a poorfit to models (Myers and Cadigan, 1995a,b; Myers et al., 1995a,b; Shepherdand Cushing, 1980). Potential reasons for this are pre-recruit mortality(Cushing, 1988; Goodyear and Christensen, 1984; Walters and Collie,1988), model skew caused by outliers (Chen and Paloheimo, 1995) and/orerrors in variablemeasurement (Walters and Ludwig, 1981). There is also thepossibility of high variance in reproductive output among individuals, that

246 Nova Mieszkowska et al.
Author’s personal copy
may in turn result in large disparities between effective and observed popu-lation sizes (Hedgecock, 1994), and even large populations may have fewindividuals that effectively contribute to spawning (Hauser and Carvalho,2008). In theory, a minority of individuals that spawn in favourable oceano-graphic conditions can disproportionately contribute to the next generation(Hedgecock, 1994). Finally, migrations affecting stock connectivity are alsonot typically considered when assessing these relationships (Myers et al.,2001).
4.2. Stock evaluation—An example from the North Sea
The ICES Stock Assessment Working Group uses data from commercialcatches, research vessel surveys and time series to evaluate the status of theNorth Sea stock relative to previous years and defined reference points.Information on catch per unit effort, catch-at-age and natural mortality arecompared to an average between 1980 and 1982, and used to calculate SSB(Beare et al., 2005; Sparre, 1991). These parameters are assumed to remainconstant between years. Forecasts of stock status for the forthcoming yearare made for a range of exploitation rates, and presented as a catch-optiontable of resultant SSB from specific fishing mortalities (Reeves and Pastoors,2007). These projections are then peer-reviewed by the ICES AdvisoryCommittee for Fisheries Management before management decisions aremade at the political level.
The use of stock–recruitment methods in fisheries management has beencriticized due to a lack of concordance between the projected model out-puts of stock assessments, and the observed annual variation in reproductivepotential of stocks. The disparity is likely to be due to interannual stochasticfluctuations, which in turn are likely to be largely driven by natural envi-ronmental changes. This highlights the need for stock assessment analyses toinclude survey-based indices of stock abundance (Marshall et al., 1998).In theory, changes in SSB should be able to provide a clear indication ofwhether fishing mortality or climate change is the dominant factor causingthe continual decline observed during the last two decades. If climate-driven effects are the primary cause of the decrease in population abun-dance, years with high observed SSB would be expected to be preceded byhigh survivorship and recruitment of sub-adults (Myers et al., 1996). How-ever, this is not generally observed. In the North Sea, for example, adultstock biomass has declined from the 1960s until recently (2006–2008),despite numbers of recruits-per-spawner being relatively high, indicatingthat the loss of individuals from populations is occurring from the older ageclasses. By contrast, if fishing is the driving force, proportional mortality dueto harvesting would continue to increase during the decline of the stock(Myers et al., 1996). Figures indicate that such fishing mortality hascontinued to increase, despite severe reductions in the TACs issued
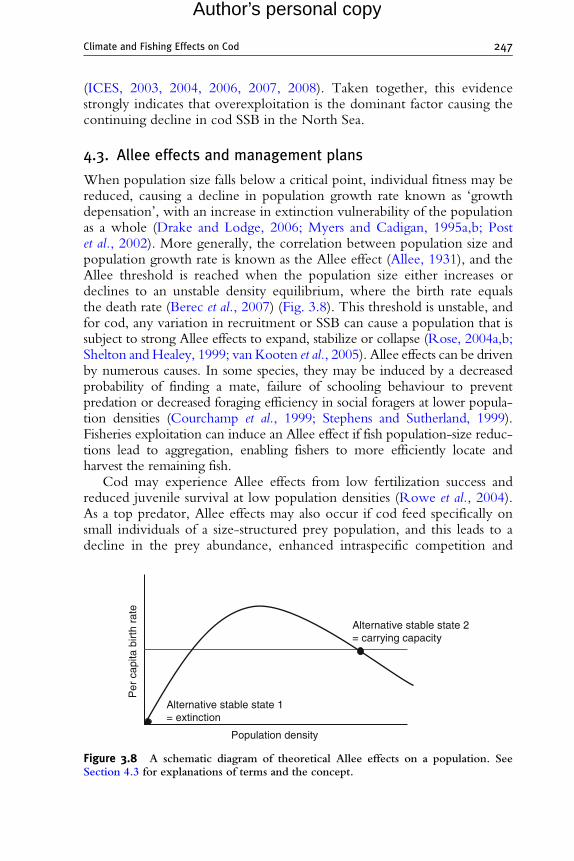
Climate and Fishing Effects on Cod 247
Author’s personal copy
(ICES, 2003, 2004, 2006, 2007, 2008). Taken together, this evidencestrongly indicates that overexploitation is the dominant factor causing thecontinuing decline in cod SSB in the North Sea.
4.3. Allee effects and management plans
When population size falls below a critical point, individual fitness may bereduced, causing a decline in population growth rate known as ‘growthdepensation’, with an increase in extinction vulnerability of the populationas a whole (Drake and Lodge, 2006; Myers and Cadigan, 1995a,b; Postet al., 2002). More generally, the correlation between population size andpopulation growth rate is known as the Allee effect (Allee, 1931), and theAllee threshold is reached when the population size either increases ordeclines to an unstable density equilibrium, where the birth rate equalsthe death rate (Berec et al., 2007) (Fig. 3.8). This threshold is unstable, andfor cod, any variation in recruitment or SSB can cause a population that issubject to strong Allee effects to expand, stabilize or collapse (Rose, 2004a,b;Shelton andHealey, 1999; van Kooten et al., 2005). Allee effects can be drivenby numerous causes. In some species, they may be induced by a decreasedprobability of finding a mate, failure of schooling behaviour to preventpredation or decreased foraging efficiency in social foragers at lower popula-tion densities (Courchamp et al., 1999; Stephens and Sutherland, 1999).Fisheries exploitation can induce an Allee effect if fish population-size reduc-tions lead to aggregation, enabling fishers to more efficiently locate andharvest the remaining fish.
Cod may experience Allee effects from low fertilization success andreduced juvenile survival at low population densities (Rowe et al., 2004).As a top predator, Allee effects may also occur if cod feed specifically onsmall individuals of a size-structured prey population, and this leads to adecline in the prey abundance, enhanced intraspecific competition and
Population density
Per
cap
ita b
irth
rate
Alternative stable state 2= carrying capacity
Alternative stable state 1= extinction
Figure 3.8 A schematic diagram of theoretical Allee effects on a population. SeeSection 4.3 for explanations of terms and the concept.

248 Nova Mieszkowska et al.
Author’s personal copy
ultimately a decline in the adult cod population (de Roos and Persson,2002; de Roos et al., 2003). Equally, Allee effects may act on cod through a‘predator-pit’ scenario. If predators are able to continually maintain codbiomass at low levels, then populations may be unable to produce sufficientrecruits to allow population expansion. Harp seals (Phoca groenlandica) haveconsumed an estimated 88,000 tonnes of cod per annum in the NorthwestAtlantic throughout the second half of the 1990s, many of which are pre-recruits (Stenson et al., 1997). The grey seal (Halichoerus grypus) is alsoreported to be responsible for heavy predation on two Northwest Atlanticstocks (Chouinard et al., 2005; Fu et al., 2001; Trzcinski et al., 2006). NorthSea cod are pre-dated upon by grey seals, but estimated consumption of codonly amounts to approximately 3.7% of the total stock biomass (Hammondand Grellier, 2005). Harbour porpoises (Phocoena phocoena) also feed oncod, and are four times as abundant as harbour seals in the North Sea.Cod populations are therefore exposed to both natural and fisheries-induced depensation.
The debate continues over the existence of fishing-induced Allee effectsin cod. It has been argued that the species does not exhibit any of themechanisms usually associated with the Allee effect (Myers et al., 1995a,b;van Kooten et al., 2005). Instead, it has been suggested that fishing mortalitymay actually enhance juvenile survival due to low spawner densities (Myerset al., 1995a,b). Exclusion of depensation in stock assessment predictions forNorthwest Atlantic stocks gave rise to predictions of an annual growth rateof 19% with zero F and a threefold increase in stock size after 7 years fromthe moratorium in 1993 (Myers et al., 1997). Over a decade on, however,only four stocks have showed any signs of increased abundance, with onlyone experiencing substantial recovery (Shelton et al., 2006).
The potential presence of Allee effects operating on cod populationssuggests that if insufficient information on the life history of the targetspecies is included in management models, the effects of climate changeon fisheries could be vastly underestimated. For populations such as theNorth Sea cod, for example, which have been reduced to levels far belowthe carrying capacity of the system, it is vital that this does not occur. Thecombination of Allee effects and incorrect management could reduce thestock size to a critical level where the population will suddenly crash.Fisheries assessment models generally assume a spawner stock–recruit func-tion whereby recruitment increases with spawner biomass until it reacheseither an asymptote (Beverton–Holt model) or declines (Ricker model).Thus, these models assume that individual reproductive output will actuallyincrease at low fish densities. The inclusion of potential Allee effects intomanagement plans could help to optimise risk management strategies, eventhough Allee thresholds cannot currently be accurately established (Berecet al., 2007). Protection of areas where high densities of cod are presentcould help to prevent Allee effects from occurring.

Climate and Fishing Effects on Cod 249
Author’s personal copy
5. Monitoring Status and Recovery of
North Sea Cod: A Case Study
Monitoring North Sea cod stocks for signs of recovery has provendifficult, not least because catches and bycatches often go unreportedmaking assessment of recovery difficult (ICES, 2004, 2005). The regioncurrently falls within a single ICES management unit, but the existence ofmultiple distinct stocks within the North Sea may obscure the dispropor-tionate decline of some more vulnerable stocks for which separate manage-ment plans may be required (Hutchinson, 2008). Landings may need to besplit into relative contributions from different stocks to allow appropriatemanagement strategies, as recommended by the North Sea Regional Advi-sory Council (NSRAC, 2008). In addition, a range of values of fishingmortality are obtained using multiple research vessel surveys, introducinguncertainty into stock estimations (Blanchard et al., 2005).
Prior to 1988, assessment and forecasting techniques showed largeinterannual variation in performance as the methodology developed, andbetween 1988 and 1995 relatively high performances were obtained. How-ever, since then, there has been a systematic underestimation of fish mortalityand overestimation of the contribution of incoming year classes (Reeves andPastoors, 2007). Similar problems have been encountered in other codstocks (Pastoors, 2005). Under-reporting of landings and the absence ofdiscard data creates uncertainties in catch estimation, although errors in theassessment preceded the problems arising from limited catch data and do notappear to explain the change in model performance. Instead, this may beattributable to the reliance on parameters estimated from earlier timeperiods where the ecosystem may have been in a different state, and/orthat may also have been potentially, at least in part, a consequence of theimpacts of climate change (Beaugrand et al., 2003; Blanchard et al., 2005;O’Brien et al., 2000).
The international aspect and mixed species nature of the North Seafishery have been highlighted as the main factors contributing to the recentpoor state of the North Sea stock (Bannister, 2004), despite efforts to reducefishing mortality at the European scale. A wider ecosystem-based approachis being developed to attempt to account for the mixed species nature ofNorth Sea demersal fisheries (Vinther et al., 2004). However, the process ofdetermining suitable parameters that distinguish relative impacts of fishing,climate change and natural variance is continuing (Blanchard et al., 2005;Rice, 1995, 2000; Rice and Evans, 1988; Rice and Rochet, 2005; Rochetand Trenkel, 2003; Trenkel and Rochet, 2003). Among these approaches issize spectra analysis, a technique that has proven useful for detecting effectsof exploitation on fish stocks, as temporal changes in the index are consistent

250 Nova Mieszkowska et al.
Author’s personal copy
with fishery-mediated changes in community structure (Bianchi et al., 2000;Gislason and Rice, 1998; Jennings and Greenstreet, 1999; Murawski andIdoine, 1992; Rice and Gislason, 1996; Shin et al., 2005; Zwanenburg,2000). Even this approach has difficulties, however, as warmer watersexpected due to climate warming may lead to enhanced recruitment ofsmall species, and concomitant shifts in the size spectra. If such issues can beovercome by allowances for discrimination between forcing factors, thensize-based metrics may become useful management tools (Blanchard et al.,2005; Genner et al., 2009b).
There has been little research into the application and utility of closedareas for fishing, and it appears most closures have been designated withoutclear objectives (SGMOS, 2003). At the geographical scale of the ICESrectangle, the lack of high densities of cod suggest that movement of fishingeffort would save few fish (Blanchard et al., 2005). A 10-week closure ofpart of the North Sea in 2001 was deemed ineffective, and data were ofinsufficient resolution to separate any closure effects from additional factors(Council Regulation (EC) No. 259/2001). It has since been concluded thattemporal closures must be used with other management approaches to havea role in the future maintenance of North Sea cod biomass, and would berequired over several years and across larger spatial scales (Horwood et al.,2006; STECF, 2007).
Emerging management ideas include the introduction of OptimumRestorable Biomass to replace MSY as a target reference point (Ainsworthand Pitcher, 2008). This has been suggested as a suitable cost/benefitapproach to restoration of severely depleted fish stocks (Pitcher, 2008) anduses ‘Back To the Future’ simulation models to establish a restorationtrajectory from the present depleted system to a target of an historical systemafter establishing sustainable fishing (Ainsworth and Pitcher, 2008; Pitcher,2008; Pitcher and Forrest, 2004). Several optimal trajectories can besimulated based on restoration targets and economic profit scenarios. Thismultidisciplinary approach to restorative ecology suggests that rebuildingstocks to defined targets is the main goal for fisheries management (Paulyet al., 1998). In contrast, however, the North Sea Regional AdvisoryCouncil has recently issued advice to the European Commission that thecod recovery plan should be reviewed and targets changed from recoverybased on biomass to that based on fishing mortality (NSRAC, 2008).
6. Concluding Remarks
Most recovery plans for fish stocks are less than two decades old, andquantifying potential recovery is still proving difficult (Caddy and Agnew,2004; Caddy and Surette, 2005). Pelagic stocks appear to be responding

Climate and Fishing Effects on Cod 251
Author’s personal copy
more positively than demersal stocks, and comparatively few stocks havebeen restored. The World Summit of Sustainable Development set a targetdate of 2015 for the management of all fish stocks at the level of MSY.No indication was given, however, as to how multi-species fisheries shouldbe managed to attain such a standard, and it is questionable whetherproposed targets are realistic in rapidly changing environments. Addition-ally, there is debate as to whether the focus of management should besustainability within an already depleted system, or the complete restorationand rebuilding of fish stocks to historic levels (Pitcher and Pauly, 1998).Moreover, reconstruction of past ecosystems may be an impossible policygoal if the past ecosystems existed under different climatic regimes (Pitcherand Forrest, 2004).
It is vitally important that the relative contributions of both fishingmortality and climate change on stock biomass, population structure andabundance changes are quantitatively understood to provide accurate andtimely advice for effective fisheries management (Horwood et al., 2006;Rose, 2004a,b). Predicting the effects of climate change on fish productiv-ity, as discussed in earlier sections of this chapter, is difficult due to incom-plete knowledge of the physiological and ecological mechanisms by whichfishes respond to changes in local and regional environmental conditions.Regime shifts also make predictions of future assemblage states problematicas the drivers force the assemblage into a new stable state which may differgreatly from the previous one (Cury et al., 2008; DeYoung et al., 2004;Mantua, 2004). Time series provide a basis for understanding the effects ofenvironmentally driven fluctuations in abundance and production of stocks,and provide the opportunity for deeper insights of the effects of highexploitation pressure (God�, 2003). However, investigations into themacroecological responses of wide ranging fishes such as Atlantic cod areproblematic due to the lack of uniformity in spatial data collection fromcommercial fisheries. In recent years, the availability of research vessel trawldata and standardized fisheries statistics has gone some way to address thisproblem. It is imperative that these data are now combined with detailedphysiological knowledge of the various life history stages, and an under-standing of interactions between cod and their environment, to allow moreaccurate predictions of future stock structure and abundance. Only then wewill be able to determine to what degree observed changes in cod popula-tions have been due to fishing and warming seas, and have the ability tointroduce effective management plans for mitigation of climate change andfisheries impacts.
The review of the European Common Fisheries Policy (CFP) in 2008acknowledges these factors and a Green Paper on the CFP reform sets out avision for European fisheries by 2020, which includes integration with thenew Integrated Maritime Policy and the Marine Strategy FrameworkDirective (CEC, 2009). The urgent need to review fisheries management

252 Nova Mieszkowska et al.
Author’s personal copy
as part of the wider maritime environment and associated activities isrecognized (Greenstreet and Rogers, 2006), as are the additional impactsof climate change on exploited marine stocks. Fleet over-capacity is stillidentified as the fundamental problem that needs to be addressed. Moreactive stewardship, including appropriate use of the increasing scientificknowledge base, combined with emergent European and internationalmaritime directives is required if we are to redress the decline in Atlanticcod whilst stocks remain to be conserved.
In a rapidly changing world a precautionary approach is required.Climate change interacts with overfishing to increase the risk of decliningstocks, especially for species of boreal biogeographic origin such as Atlanticcod. Occasional large recruitment events can enable a single annual-breed-ing boreal species, which is dependent on the spring/summer temperatureregime, to persist in the face of climatic change. If longevity and size ofspawning fish are reduced, then persistence becomes less likely as theprobability of a major recruitment event during the lifespan of an individualis much reduced. As we describe in this chapter, it seems clear that climatecan act to exacerbate the deleterious effects of overfishing on marineecosystems; however, in this context, climate change effects should notexcuse overfishing. Moreover, given the mosaic structure of cod popula-tions with complex stochastic responses to climate change, the revolution ofmanagement measures must match the scale of these ecological processes.Management strategies with one size and approach, such as the CFP, willnot sufficiently well accommodate the complexities of cod interactions withchanging environments.
ACKNOWLEDGEMENTS
We thank G. Beaugrand for providing Fig. 3.5 and S. Cotterell for providing critical com-ments on an earlier version of this Chapter.We gratefully acknowledge funding support fromthe UK Department for Environment, Food and Rural Affairs (DEFRA), the UK NaturalEnvironment Research Council (NERC) Oceans 2025 Strategic Research Programmethrough Themes 6 (Science for Sustainable Marine Resources) and 10 (Integrating sustainedobservations for marine environmental monitoring) and The Worshipful Company ofFishmongers. MJG was supported by a Great Western Research Fellowship and DWS byan MBA Senior Research Fellowship.
REFERENCES
Ainsworth, C., and Pitcher, T. (2008). Back to the future in Northern British Columbia:Evaluating historic marine ecosystems and optimal restorable biomass as restoration goalsfor the future. Am. Fish. Soc. Symp. 49, 317–329.
Allee, W. C. (1931). ‘‘Animal Aggregations: A Study in General Sociology.’’ The Universityof Chicago Press, Chicago.

Climate and Fishing Effects on Cod 253
Author’s personal copy
Anderson, J. T. (1988). A review of size-dependent survival during pre-recruit stages offishes in relation to recruitment. J. Northwest Atl. Fish. Sci. 8, 55–66.
Andersen, O.,Wetten, O. F., De Rosa, M. C., Andre, C., Alinovi, C. C., Colafranceschi, M.,Brix, O., and Colosimo, A. (2009). Haemoglobin polymorphisms affect the oxygen-binding properties in Atlantic cod populations. Proc. R. Soc. B 276, 833–841.
Armstrong, M. J., Connolly, P., Nash, R. D. M., Pawson, M. G., Alesworth, E.,Coulahan, P. J., Dickey-Collas, M., Milligan, S. P., O’Neill, M. F., Witthames, P. R.,andWoolner, L. (2001). An application of the annual egg production method to estimatethe spawning biomass of cod (Gadus morhua L.), plaice (Pleuronectes platessa L.) and sole(Solea solea L.) in the Irish Sea. ICES J. Mar. Sci. 58, 183–203.
Arnason, E. (2004). Mitochondrial cytochrome bDNA variation in the high fecundity Atlanticcod: Trans-Atlantic clines and shallow gene–genealogy. Genetics 166, 1871–1885.
Arnason, E., and Palsson, S. (1996). Mitochondrial cytochrome b DNA sequence variationof Atlantic cod Gadus morhua, from Norway. Mol. Ecol. 5, 715–724.
Arnold, G., and Dewar, H. (2001). Electronic tagging and tracking in marine fisheriesresearch: a 30-year perspective. In ‘‘Proceedings of the Symposium on Tagging andTracking Marine Fish with Electronic Devices,’’ East-West Center, University ofHawaii. (J. R. Sibert and J. L. Nielsen, eds.), pp. 7–65. Springer-Verlag, New York.
Arnold, G. P., and Walker, M. G. (1992). Vertical movements of cod (Gadus morhua L.) in theopen sea and the hydrostatic function of the swimbladder. ICES J. Mar. Sci. 49, 357–372.
Atkinson, D. B., Rose, G. A., Murphy, E. F., and Bishop, C. A. (1997). Distributionchanges and abundance of northern cod (Gadus morhua), 1981-1993. Can. J. Fish.Aquat. Sci. 54, 132–138.
Attrill, M. J., and Power, M. (2002). Climatic influence on a marine fish assemblage. Nautre417, 275–278.
Bakun, A. (1996). ‘‘Patterns in the Ocean: Ocean Processes and Marine Population Dynam-ics.’’ 323pp. Centro de Investigaciones Biologicas del Noroeste, La Paz, Mexico.
Baldursson, F. M., Danielsson, A., and Stefansson, G. (1996). On the rational utilization ofthe Icelandic cod stock. ICES J. Mar. Sci. 53, 643–658.
Bannister, R. C. A. (2004). The rise and fall of cod (Gadus morhua, L.) in the North Sea.In ‘‘Management of Shared Fish Stocks’’ (C. M. O’Brien and S. I. Rogers, eds.),pp. 316–338. Blackwells, Oxford.
Barrett, J. H., Locker, A. M., and Roberts, C. M. (2004). The origins of intensive marinefishing in medieval Europe: The English evidence. Proc. R. Soc. B 271, 2417–2421.
Barrett, J., Johnstone, C., Harland, J., Van Neer, W., Ervynck, A., Makowiecki, D.,Heinrich, D., Hufthammer, A. K., Enghoff, I. B., Amundsen, C., Christiansen, J. S.,Jones, A. K. G., et al. (2008). Detecting the medieval cod trade: A new method and firstresults. J. Archaeol. Sci. 35, 850–861.
Barry, J. P., Baxter, C. H., Sagarin, R. D., and Gilman, S. E. (1995). Climate-related, long-term faunal changes in a California rocky intertidal community. Science 267, 672–674.
Beacham, T. D., Brattey, J., Miller, K. M., Le, K. D., and Withler, R. E. (2002). Multiplestock structure of Atlantic cod (Gadus morhua) off Newfoundland and Labrador deter-mined from genetic variation. ICES J. Mar. Sci. 59, 650–665.
Beamish, F. W. H. (1978). Swimming capacity. In ‘‘Fish Physiology.’’ vol. 7. (W. S. Hoarand D. J. Randall, eds.), pp. 101–187. London, Academic Press.
Beare, D. J., Needle, C. L., Burns, F., and Reid, D. G. (2005). Using survey data indepen-dently from commercial data in stock assessment: an example using haddock in ICESDivision VIa. ICES J. Mar. Sci. 62, 996–1005.
Beaugrand, G., and Ibanez, F. (2004). Monitoring marine plankton ecosystems. II: Long-term changes in North Sea calanoid copepods in relation to hydro-climatic variability.Mar. Ecol. Prog. Ser. 284, 35–47.

254 Nova Mieszkowska et al.
Author’s personal copy
Beaugrand, G., Reid, P. C., Ibanez, F., Lindley, J. A., and Edwards, M. (2002). Reorgani-zation of North Atlantic marine copeopod biodiversity and climate. Science 296,1692–1694.
Beaugrand, G., Brander, K. M., Lindley, J. A., Souissi, S., and Reid, P. C. (2003). Planktoneffect on cod recruitment in the North Sea. Nature 426, 661–664.
Beckman, D. W., andWilson, C. A. (1995). Seasonal timing of opaque zone formation in fishotoliths. In ‘‘Recent Developments in Fish Otolith Research’’ (D. J. Secor, J. M. Dean andS. E. Campana, eds.), pp. 27–44. University of South Carolina Press, Columbia.
Begg, G. A., and Marteinsdottir, G. (2000). Spawning origins of pelagic juvenile cod Gadusmorhua inferred from spatially explicit age distributions: Potential influences on year-classstrength and recruitment. Mar. Ecol. Prog. Ser. 202, 193–217.
Begg, G. A., and Marteinsdottir, G. (2002). Environmental and stock effects on spawningorigins and recruitment of cod Gadus morhua. Mar. Ecol. Prog. Ser. 229, 245–262.
Bekkevold, D. (2006). Male size composition affects male reproductive variance in Atlantic codGadus morhua L. spawning aggregations. J. Fish Biol. 69, 945–950.
Bekkevold, D., Hansen, M. M., and Loeschcke, V. (2002). Male reproductive competitionin spawning aggregations of cod (Gadus morhua, L.). Mol. Ecol. 11, 91–102.
Berec, L., Angulo, E., and Courchamp, F. (2007). Multiple Allee effects and populationmanagement. Trends Ecol. Evol. 22, 185–191.
Berge, J., Johnsen, G., Gulliksen, B., and Slagstad, D. (2005). Ocean temperature oscillationsenable reappearance of blue mussels Mytilus edulis in Svalbard after a 1000 year absence.Mar. Ecol. Prog. Ser. 303, 167–175.
Beverton, R. J. H., and Holt, S. J. (1957). ‘‘On the Dynamics of Exploited Fish Populations:Fisheries Investigations Series 2 G.B.’’ Ministry for Agriculture Fisheries and Food,London.
Bianchi, G., Gislason, H., Graham, K., Hill, L., Jin, X., Koranteng, K., Manickchand-Heileman, S., Paya, I., Sainsbury, K., Sanchez, F., and Zwanenburg, K. (2000). Impact offishing on size composition and diversity of demersal fish communities. ICES J. Mar. Sci.57, 558–571.
Bigg, G. R., Cunningham, C. W., Ottersen, G., Pogson, G. H., Wadley, M. R., andWilliamson, P. (2008). Ice-age survival of Atlantic cod: Agreement between palaeoecol-ogy models and genetics. Proc. R. Soc. B 275, 163–172.
Bishop, C. A., Murphy, E. F., Davis, M. B., Baird, J. W., and Rose, G. (1993).An assessment of the cod stock in NAFO divisions 2J þ 3KL. In ‘‘ANAFO ScientificCouncil Research Document.’’ Northwest Atlantic Fisheries Centre, St. Johns,Newfoundland.
Bjornsson, B., and Steinarsson, A. (2002). The food-unlimited growth rate of Atlantic cod(Gadus morhua). Can. J. Fish. Aquat. Sci. 59, 494–502.
Bjornsson, B., Steinarsson, A., and Oddgeirsson, M. (2001). Optimal temperature for growthand feed conversion of immature cod (Gadus morhua L.). ICES J. Mar. Sci. 58, 29–38.
Bjornstad, O. N., Fromentin, J. M., Stenseth, N. C., and Gjosaeter, J. (1999). Cycles andtrends in cod populations. Proc. Natl. Acad. Sci. USA 96, 5066–5071.
Blanchard, J. L., Frank, K. T., and Simon, J. E. (2003). Effects of condition on fecundity andtotal egg production of eastern Scotian Shelf haddock (Melanogrammus aeglefinus). Can. J.Fish. Aquat. Sci. 60, 321–332.
Blanchard, J. L., Mills, C., Jennings, S., Fox, C. J., Rackham, B. D., Eastwood, P. D., andO’Brien, C. M. (2005). Distribution–abundance relationships for North Sea Atlantic cod(Gadus morhua): Observation versus theory. Can. J. Fish. Aquat. Sci. 62, 2001–2009.
Borley, J. O. (1923). The plaice fishery and the war. Preliminary report on investigations.Fish. Invest. 2, 56.
Brander, K. M. (1994). Patterns of distribution, spawning, and growth in North Atlanticcod: The utility of inter-regional comparisons ICES Mar. Sci. Symp., Copenhagen.

Climate and Fishing Effects on Cod 255
Author’s personal copy
Brander, K.M. (1995). The effect of temperature on growth of Atlantic cod (Gadus morhua L.).ICES J. Mar. Sci. 52, 1–10.
Brander, K. M. (1997). Effects of climate change on cod (Gadus morhua) stocks.In ‘‘Global Warming: Implications for Freshwater and Marine Fish.’’ (C. M. Woodand D. G. McDonald, eds.), pp. 255–278. Cambridge University Press, Cambridge.
Brander, K. M. (2000). Detecting the effects of environmental change on growth and recruit-ment in cod (Gadus morhua) using a comparative approach.Oceanol. Acta 23, 485–496.
Brander, K. M. (2004). Cod recruitment is strongly affected by climate when stock biomassis low. ‘‘Symposium on Quantitative Ecosystem Indicators for Fisheries Management,’’Paris, pp. 339–343.
Brander, K. M. (2005). Cod recruitment is strongly affected by climate when stock biomassis low. ICES J. Mar. Sci. 62, 339–343.
Brander, K., and Mohn, R. (2004). Effect of the North Atlantic Oscillation on recruitmentof Atlantic cod (Gadus morhua). Can. J. Fish. Aquat. Sci. 61, 1558–1564.
Brander, K., Blom, G., Borges, M. F., Erzini, K., Henderson, G., MacKenzie, B. R.,Mendez, M., Ribiero, J., Santos, A. M. P., and Toresen, R. (2003). Changes in fishdistribution in the eastern north Atlantic: Are we seeing a coherent response to changingtemperature? ICES Mar. Sci. Symp. 219, 261–270.
Brett, J. R., and Groves, T. D. D. (1979). Physiological energetics. In ‘‘Fish Physiology.’’(W. J. Hoar, D. J.Randall, and J. R. Brett, eds.), pp. 279–352. Academic Press, NewYork.
Broitman, B. R., Mieszkowska, N., Helmuth, B., and Blanchette, C. A. (2008). Climate andrecruitment of rocky shore intertidal invertebrates in the eastern North Atlantic. Ecology89, S81–S90.
Brunel, T., and Boucher, J. (2007). Long-term trends in fish recruitment in the north-eastAtlantic related to climate change. Fish. Oceanogr. 16, 336–349.
Buch, E., and Hansen, H. H. (1988). Climate and cod fishery at West Greenland.In ‘‘Longterm Changes in Marine Fish Populations.’’ (T. Wyatt and M. G. Larraneta,eds.), pp. 345–364. Instituto Investigaciones Marinas, Vigo, Spain.
Buckley, L. J., Caldarone, E. M., and Lough, R. G. (2004). Optimum temperature andfood-limited growth of larval Atlantic cod (Gadus morhua) and haddock (Melanogrammusaeglefinus) on Georges Bank. Fish. Oceanogr. 13, 134–140.
Bunn, N., and Fox, C. J. (2004). Spring plankton surveys of the Irish Sea in 2000:Hydrography and the distribution of fish eggs and larvae. Science Series Data Report,No. 41, Centre for Environment, Fisheries and Aquculture Science, Lowestoft, UK.
Caddy, J. F., and Agnew, D. J. (2004). An overview of recent global experience withrecovery plans for depleted marine resources and suggested guidelines for recoveryplanning. Rev. Fish Biol. Fish. 14, 43–112.
Caddy, J., and Surette, T. (2005). In retrospect the assumption of sustainability for Atlanticfisheries has proved an illusion. Rev. Fish Biol. Fish. 15, 313–337.
Caldarone, E. M., St. Onge-Burns, J. M., and Buckley, L. J. (2003). Relationship ofRNA-DNA and temperature to growth in larvace of Atlantic cod Gadus morhua. Mar.Ecol. Prog. Ser. 262, 229–240.
Campana, S. E. (1996). Year-class strength and growth rate in young Atlantic cod Gadusmorhua. Mar. Ecol. Prog. Ser. 135, 21–26.
Campana, S. E., and Hurley, P. C. F. (1989). An age and temperature mediated growthmodel for cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) larvae in the Gulfof Maine. Can. J. Fish. Aquat. Sci. 46, 603–613.
Campana, S. E., Annand, M. C., and McMillan, J. I. (1994). Graphical and statisticalmethods for determining the consistency of age determinations. Trans. Am. Fish. Soc.124, 131–138.
Carr, S. M., and Crutcher, D. C. (1998). Population genetic structure in Atlantic cod (Gadusmorhua) from the North Atlantic and Barents Sea: contrasting or concordant patternsinmtDNA sequence and microsatellite data?. In ‘‘The Implications of Localized Fishery

256 Nova Mieszkowska et al.
Author’s personal copy
Stocks.’’ (I. Hunt von Herbing, I. Kornfield, M. Tupper and J. Wilson, eds.), pp. 91–103.Northeast Regional Agricultural Engineering Service, Ithaca, New York, USA.
Case, R. A. J., Hutchinson, W. F., Hauser, L., Van Oosterhout, C., and Carvalho, G.(2005). Macro- and micro-geographic variation in pantophysin (PanI) allele frequenciesin NE Atlantic cod Gadus morhua. Mar. Ecol. Prog. Ser. 301, 267–278.
Case, R. A. J., Hutchinson, W. F., Hauser, L., Buchler, V., Clemmesen, C., Dahle, G.,Kjesbu, O. S., Moksness, E., Ottera, H., Paulsen, H., Svasand, T., Thorsen, A., et al.(2006). Association between growth and PanI* genotype within Atlantic cod full siblingfamilies. Trans. Am. Fish. Soc. 135, 241–250.
CEC (2001). ‘‘Green Paper on the Future of the Common Fisheries Policy.’’ Commission ofthe European Community, Brussels.
CEC (2009). ‘‘Green Paper on the Reform of the Common Fisheries Policy.’’ Commissionof the European Community, Brussels.
Chabot, D., and Dutil, J. D. (1999). Reduced growth of Atlantic cod in non-lethal hypoxicconditions. J. Fish Biol. 55, 472–491.
Chambers,R.C., andLeggett,W.C. (1987). Size and age atmetamorphosis inmarine fishes.Ananalysis of laboratory-reared winter flounder (Pseudopleutonectes americanus) with a review ofvariation in other species.Can. J. Fish. Aquat. Sci. 44, 1936–1947.
Chen, Y., and Paloheimo, J. E. (1995). A robust regression analysis of recruitment infisheries. Can. J. Fish. Aquat. Sci. 52, 993–1006.
Chouinard, G. A., Swain, D. P., Hammill, M. O., and Poirier, G. A. (2005). Covariationbetween grey seal (Halichoerus grypus) abundance and natural mortality of cod (Gadusmorhua) in the southern Gulf of St. Lawrence. Can. J. Fish. Aquat. Sci. 62, 1991–2000.
Chown, S. L., and Gaston, K. J. (1999). Exploring links between physiology and ecology atmacro–scales: The role of respiratory metabolism in insects. Biol. Rev. 74, 87–120.
Claireaux, G., Webber, D. M., Kerr, S. R., and Boutilier, R. G. (1995). Physiology andbehaviour of free-swimming Atlantic cod (Gadus morhua) facing fluctuating temperatureconditions. J. Exp. Biol. 198, 49–60.
Claireaux, G., Webber, D. M., and Kerr, S. R. (2000). Influence of water temperature andoxygenation on the aerobic metabolic scope of Atlantic cod (Gadus morhua). J. Sea Res.44, 257–265.
Clark, R. A., Fox, C. J., Viner, D., and Livermore, M. (2003). North Sea cod and climatechange—Modelling the effects of temperature on population dynamics. Global ChangeBiol. 9, 1669–1680.
Clark, D. S., and Green, J. M. (1990). Activity and movement patterns of juvenile Atlanticcod, Gadus morhua, in Conception Bay, Newfoundland, as determined by sonic teleme-try. Can. J. Zoolog. 68, 1434–1442.
Clarke, A. (1993). Seasonal acclimatization and latitudinal compensation in metabolism:Do they exist? Funct. Ecol. 7, 139–149.
Clarke, A. (2003). Costs and consequences of evolutionary temperature adaptation. TrendsEcol. Evol. 18, 573–581.
Cohen, D. M., Inada, T., Iwamoto, T., and Scialabba, N. (1990). ‘‘FAO Species CatalogueVolume 10. Gadiform Fishes of the World.’’ p. 442. Food and Agriculture Organizationof the United Nations, Rome.
Colosimo, A., Giuliani, A., Maranghi, F., Brix, O., Thorkildsen, S., Fischer, T., Knust, R.,and Portner, H. O. (2003). Physiological and genetical adaptation to temperature in fishpopulations. Cont. Shelf Res. 23, 1919–1928.
Cook, R. M., Sinclair, A., and Stefansson, G. (1997). Potential collapse of North Sea codstocks. Nature 385, 521–522.
Cote, D., Ollerhead, L. M. N., Scruton, D. A., and McKinley, R. S. (2003). Microhabitatuse of juvenlie Atlantic cod in a coastal area of Newfoundland determined by 2Dtelemetry. Mar. Ecol. Prog. Ser. 265, 227–234.

Climate and Fishing Effects on Cod 257
Author’s personal copy
Courchamp, F., Clutton-Brock, T., and Grenfell, B. (1999). Inverse density dependenceand the Allee effect. Trends Ecol. Evol. 14, 405–410.
Coutant, C. C. (1977). Compilation of Temperature Preference Data. J. Fish Res. BoardCan. 34, 739–745.
Coutant, C. C. (1987). Thermal preference: When does and asset become a liability?Environ. Biol. Fish 18, 161–172.
Cury, P. M., Shin, Y. J., Planque, B., Durant, J. M., Fromentin, J. M., Kramer-Schadt, S.,Stenseth, N. C., Travers, M., and Grimm, V. (2008). Ecosystem oceanography for globalchange in fisheries. Trends Ecol. Evol. 23, 338–346.
Cushing, D. H. (1984). The gadoid outburst in the North Sea. ICES J. Mar. Sci. 41,159–166.
Cushing, D. H. (1988). The northerly wind. In ‘‘Toward a Theory of Biological–PhysicalInteractions in the World Ocean.’’ (B. J. Rothschild, ed.), pp. 235–244. NATOAdvanced Research Workshop, Kluwer, Dordrecht.
Cushing, D. H. (1990). Plankton production and year-class strength in fish populations:An update of the match/mismatch hypothesis. Adv. Mar. Biol. 26, 249–252.
Cushing, D. H. (1996). Towards a science of recruitment in fish populations, p. 155.Ecology Institute. Oldenfof, Germany.
Cushing, D. H., and Horwood, J. W. (1994). The growth and death of fish larvae. J. PlanktonRes. 16, 291–300.
Daan, N. (1978). Changes in cod stocks and cod fisheries in the North Sea. Symposium onNorth Sea Fish Stocks: Recent changes and their causes. Conseil Int. l’Exploration Mer172, 39–57.
Daan, N. (1994). Trends in North Atlantic cod stocks: A critical summary. ICES Mar. Sci.Symp. 198, 269–270.
Davis, A. J., Jenkinson, L. S., Lawton, J. H., Shorrocks, B., and Wood, S. (1997). Makingmistakes when predicting shifts in species range in response to global warming. Nature391, 783–786.
de Cardenas, E. (1996). Some considerations about annual growth rate variations in codstocks. NAFO Sci. Coun. Studies 24, 97–107.
de Roos, A. M., and Persson, L. (2002). Size-dependent life-history traits promotecatastrophic collapses of top predators. Proc. Natl. Acad. Sci. USA 99, 12907–12912.
de Roos, A. M., Persson, L., and Thieme, H. R. (2003). Emergent Allee effects in toppredators feeding on structured prey populations. Proc. R. Soc. B 270, 611–618.
DeYoung, B., Harris, R., Alheit, J., Beaugrand, G., Mantua, N., and Shannon, L. (2004).Detecting regime shifts in the ocean: Data considerations. Prog. Oceanogr. 60, 143–164.
de Young, B., and Rose, G. (1993). On recruitment and distribution of Atlantic cod(Gadus morhua) off Newfoundland. Can. J. Fish. Aquat. Sci. 50, 2729–2741.
Dickson, R. R., Pope, J. G., and Holden, M. J. (1974). Environmental influences on thesurvival of North Sea cod. In ‘‘The Early Life History of Fish.’’ ( J. H. S. Blaxter, ed.),pp. 69–80. Springer-Verlag, Berlin.
Dippner, J. (1997). Recruitment success of different fish stocks in the North Sea in relationto climate variablility. Ocean Dynamics 49, 277–293.
Dower, J. F., Pepin, P., and Leggett, W. C. (1998). Enhanced gut fullness and an apparentshift in size selectivity by radiated shanny (Ulvaria subbifurcata) larvae in response toincreased turbulence. Can. J. Fish. Aquat. Sci. 55, 128–142.
Drake, J., and Lodge, D. (2006). Allee Effects, propagule pressure and the probability ofestablishment: Risk analysis for biological invasions. Biol. Invasions 8, 365–375.
Drinkwater, K. F. (2002). A review of the role of climate variability in the decline ofnorthern cod. Am. Fish. Soc. Symp. 32, 113–130.
Drinkwater, K. F. (2005). The response of Atlantic cod (Gadus morhua) to future climatechange. ICES J. Mar. Sci. 62, 1327–1337.

258 Nova Mieszkowska et al.
Author’s personal copy
Drinkwater, K., Belgrano, A., Borja, A., Conversi, A., Edwards, M., Greene, C., Ottersen, G.,Pershing, A., and Walker, H. (2003). The response of marine ecosystems to climatevariability associated with the North Atlantic Oscillation. In ‘‘The North Atlantic Oscilla-tion: climatic significance and environmental impact’’ ( J. Hurrell, Y. kushnir, G. Ottersenand M. Visbeck, eds.), pp. 211–234. American Geophysical Union, Washington.
Dulvy, N. K., Rogers, S. I., Jennings, S., Stelzenmuller, V., Dye, S. R., and Skjoldal, H. R.(2008). Climate change and deepening of the North Sea fish assemblage: A bioticindicator of warming seas. J. Appl. Ecol. 45, 1029–1039.
Dutil, J.-D., and Brander, K. (2003). Comparing productivity of North Atlantic cod (Gadusmorhua) stocks and limits to growth production. Fish. Oceanogr. 12, 502–512.
Eckman, S. (1953). ‘‘Zoogeography of the Sea.’’ Sidgwick & Jackson, Manchester, NH.Edwards, M., and Richardson, A. J. (2004). Impact of climate change on marine pelagic
phenology and trophic mismatch. Nature 430, 881–884.Engen, F., and Folstad, I. (1999). Cod courtship song: A song at the expense of dance? Can.
J. Zool. 77, 542–550.FAO (2002). ‘‘International Workshop on Factors Contributing to Unsustainability and
Overexploitation in Fisheries.’’ Fodd and Agriculture Organization of the UnitedNations, Rome.
Fevolden, S. E., and Pogson, G. H. (1997). Genetic divergence at the synaptophysin (Syp I)locus among Norwegian coastal and Northeast Arctic populations of Atlantic cod. J. FishBiol. 51, 895–908.
Fiksen, �., and MacKenzie, B. R. (2002). Process-based models of feeding and preyselection in larvae fish. Mar. Ecol. Prog. Ser. 243, 151–164.
Fisheries Science Services (2008). ‘‘The Stock Book.’’ Fisheries and Food Annual Review ofFish Stocks in 2008 with Management Advice for 2009, London. Report to the Ministerfor Agriculture, p. 424.
Foster, A. R., Hall, S. J., and Houlihan, D. F. (1993). The effects of seasonal acclimatizationon correlates of growth rate in juvenile cod, Gadus morhua. J. Fish Biol. 42, 461–464.
Fox, C. J., Taylor, M. I., Pereyra, R., and Rico, C. (2005). Mapping of the spawning groundsof Irish Sea gadoids using genetic identification of planktonic eggs.Mol. Ecol. 14, 879–884.
Fox, C. J., Taylor, M. I., Dickey-Collas, M., Fossum, P., Kraus, G., Rohlf, N., Munk, P.,van Damme, C. J., Bolle, L. J., Maxwell, D. L., and Wright, P. J. (2008). Mapping thespawning grounds of North Sea cod (Gadus morhua) by direct and indirect means. Proc. R.Soc. B 275, 1543–1548.
Frank, K. T. (1997). The utility of early life history studies and the challenges of recruitmentprediction. In ‘‘Early life history and recruitment.’’ (R. C. Chambers and E. A. Trippel,eds.), pp. 495–512.
Frederich, M., and Portner, H. O. (2000). Oxygen limitation of thermal tolerance definedby cardiac and ventilatory performance in spider crab, Maja squinado. Am. J. Physiol.-Reg. I. 279, 1531–1538.
Fromentin, J. M., and Planque, B. (1996). Calanus and environment in the eastern NorthAtlantic. II. Influence of the North Atlantic Oscillation on C. finmarchicus andC. helgolandicus Mar. Ecol. Prog. Ser. 134, 1–3.
Fry, F. E. J. (1947). ‘‘Effects of the Environment on Animal Activity.’’ University ofToronto Press, Toronto.
Fry, F. E. J. (1971). The effect of environmental factors on the physiology of fish. In ‘‘FishPhysiology VI. Environmental Relations and Behavior’’ (W. S. Hoar and D. J. Randall,eds.). Academic Press, New York.
Fu, C., Mohn, R., and Fanning, L. P. (2001). Why the Atlantic cod (Gadus morhua) stock offeastern Nova Scotia has not recovered. Can. J. Fish. Aquat. Sci. 58, 1613–1623.
Gadil, M., and Bossert, W. (1970). Life history consequences of natural selection. Am. Nat.104, 1–24.

Climate and Fishing Effects on Cod 259
Author’s personal copy
Geffen, A. J., Fox, C. J., and Nash, R. D. M. (2006). Temperature-dependent developmentrates of cod Gadus morhua eggs. J. Fish Biol. 69, 1060–1080.
Genner, M. J., Sims, D. W., Wearmouth, V. J., Southall, E. J., Southward, A. J.,Henderson, P. A., and Hawkins, S. J. (2004). Regional climatic warming drives long-term community changes of British marine fish. Proc. R. Soc. B 271, 655–661.
Genner, M. J., Halliday, N. C., Simpson, S. D., Hawkins, S. J., and Sims, D. W. (2009a).Temperature-driven phenological changes in a marine larval fish assemblage. J. PlanktonRes. (in press).
Genner, M. J., Sims, D. W., Southward, A. J., Budd, G. C., Masterson, P., McHugh, M.,Rendle, P., Southall, E. J., Wearmouth, V. J., and Hawkins, S. J. (2009b). Body size-dependent responses of a marine fish assemblage to climate change and fishing over acentury-long scale. Global Change Biol. doi: 10.1111/j.1365-2486.2009.02027.
Gislason, H., and Rice, J. (1998). Modelling the response of size and diversity spectra of fishassemblages to changes in exploitation. ICES J. Mar. Sci. 55, 362–370.
God�, O. R. (2003). Fluctuation in stock properties of Northeast Arctic cod related to long-term environmental changes. Fish Fish. 4, 121–137.
God�, O. R., and Moksness, E. (1987). Growth and maturation of Norwegian coastal codand Northeast Arctic cod under different conditions. Fish. Res. 5, 235–242.
Gollock, M. J., Currie, S., Petersen, L. H., and Gamperl, A. K. (2006). Cardiovascular andhaematological responses of Atlantic cod (Gadus morhua) to acute temperature increase.J. Exp. Biol. 209, 2961–2970.
Goodyear, C. P., and Christensen, S. W. (1984). On the ability to detect the influence ofspawning stock on recruitment. N. Am. J. Fish. Manage. 4, 186–193.
Graham, C. T., and Harrod, C. (2009). Implications of climate change for the fishes of theBritish Isles. J. Fish Biol. 74, 1143–1205.
Greene, C. H., Pershing, A. J., Conversi, A., Planque, B., Hannah, C., Sameoto, D.,Head, E., Smith, P. C., Reid, P. C., Jossi, J., Mountain, D., Benfield, M. C., et al.(2003). Trans-Atlantic responses of Calanus finmarchicus populations to basin-scale forcingassociated with the North Atlantic Oscillation. Prog. Oceanogr. 58, 301–312.
Greenstreet, S. P. R., and Rogers, S. I. (2006). Indicators of the health of the North Sea fishcommunity: Identifying reference levels for an ecosystem approach to management.ICES J. Mar. Sci. 63, 573–593.
Griffiths, S. P. (2003). Rockpool ichthyofaunas of temperate Australia: Species composition,residency and biogeographic patterns. Estuar. Coast. Shelf Sci. 58, 173–186.
Guderley, H. (1990). Functional significance of metabolic responses to thermal acclimationin fish muscle. Am. J. Physiol.-Reg. I. 259, 245–252.
Gulland, J. A. (1964). The reliability of catch per unit of effort as a measure of abundance inNorth Sea trawl fisheries. Rapp. P.-V. Reun. Cons. Perm. 155, 99–102.
Hammond, P. S., and Grellier, K. (2005). Grey seal diet and fish consumption in theNorth Sea.Report of UK Department for Environment, Food and Rural Affairs MF0319, pp. 1–54.
Hansen, P. M. (1949). Studies on the biology of the cod in Greenland waters. Rapp. P.-V.Reun. Cons. Perm. 123, 1–77.
Harden Jones, F. R. (1968). ‘‘Fish Migration.’’ Edward Arnold, London.Hardie, D. C., Gillett, R. M., and Hutchings, J. A. (2006). The effects of isolation and
colonization history on the genetic structure of marine-relict populations of Atlantic cod(Gadus morhua) in the Canadian Arctic. Can. J. Fish. Aquat. Sci. 63, 1830–1839.
Hardy, A. C. (1959). ‘‘The Open Sea: Its Natural History, Part 11.’’ p. 322. Fish andFisheries Series, Collins, London.
Hare, J. A., and Cowen, R. K. (1997). Size, growth, development and survival of theplanktonic larvae of Pomatomus saltatrix (Pisces: Pomatomidae). Ecology 78, 2415–2431.
Harley, C. D. G., Hughes, A. R., Hultgren, K. M., Miner, B. G., Sorte, C. J. B.,Thornber, C. S., Rodriguez, L. F., Tomanek, L., and Williams, S. L. (2006). Theimpacts of climate change in coastal marine systems. Ecol. Lett. 9, 228–241.

260 Nova Mieszkowska et al.
Author’s personal copy
Hauser, L., and Carvalho, G. R. (2008). Paradigm shifts in marine fisheries genetics: Uglyhypotheses slain by beautiful facts. Fish Fish. 9, 333–362.
Hawkins, S. J., Southward, A. J., and Genner, M. J. (2003). Detection of environmentalchange in a marine ecosystem—Evidence from the Western English Channel. Sci. TotalEnviron. 310, 245–256.
Heath, M. R., and Gallego, A. (1997). From the biology of the individual to the dynamics ofthe population: Bridging the gap in fish early life histories. J. Fish Biol. 51, 1–29.
Heath, M. R., Backhaus, J. O., Richardson, K., McKenzie, E., Slagstad, D., Beare, D.,Dunn, J., Fraster, J. G., Gallego, A., Hainbucher, D., Hay, S., Jonasdottir, S., et al.(1999). Climate fluctuations and the spring invasion of the North Sea by Calanusfinmarchicus. Fish. Oceanogr. 8, 163–176.
Hedgecock, D. (1994). Does variance in reproductive success limit effective population sizesof marine organisms? In ‘‘Genetics and the Evolution of Aquatic Organisms.’’(A. R. Beaumont, ed.), pp. 122–134. Chapman and Hall, London.
Hedger, R., McKenzie, E., Heath, M., Wright, P., Scott, B., Gallego, A., and Andrews, J.(2004). Analysis of the spatial distributions of mature cod (Gadus morhua) and haddock(Melanogrammus aeglefinus) abundance in the North Sea (1980–1999) using generalisedadditive models. Fish. Res. 70, 17–25.
Hellberg, M. E., Balch, D. P., and Roy, K. (2001). Climate-driven range expansion andmorphological evolution in a marine gastropod. Science 292, 1707–1710.
Herbert, N., and Steffensen, J. (2005). The response of Atlantic cod, Gadus morhua, toprogressive hypoxia: Fish swimming speed and physiological stress. Mar. Biol. 147,1403–1412.
Hermann, F., Hansen, P. H., and Horsted, S. A. (1965). The effect of temperature andcurrents on the distribution and survival of cod larvae at West Greenland, pp. 389–395,ICNAF Special Publication.
Hilborn, R., and Walters, C. (1992). ‘‘Quantitative Fisheries Stock Assessment and Man-agement: Choice, Dynamics, and Uncertainty,’’ p. 570. Chapman and Hall, New York.
Hislop, J. R. G. (1996). Changes in North Sea gadoid stocks. ICES J. Mar. Sci. 53,1146–1156.
Hobson, V. J., Righton, D., Metcalfe, J. D., and Hays, G. C. (2007). Vertical movements ofNorth Sea cod. Mar. Ecol. Prog. Ser. 347, 101–110.
Hobson, V. J., Righton, D., Metcalfe, J. D., and Hays, G. C. (2009). Link between verticaland horizontal movement patterns of cod in the North Sea. Aquat. Biol. 5, 133–142.
Holden, M. (1994). ‘‘The Common Fisheries Policy. Origin, Evaluation and Future.’’ TheBuckland Foundation, London.
Holden, M. J. (1978). Long-term changes in landings of fish from the North Sea. Rapp.P.-V. Reun. Cons. Perm. 172, 11–26.
Hooper, D. U., Chapin, F. S., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S.,Lawton, J. H., Lodge, D. M., Loreau, M., Naeem, S., Schmid, B., Setala, H., et al.(2005). Effects of biodiversity on ecosystem functioning: A consensus of current knowl-edge. Ecol. Monogr. 75, 3–35.
Horwood, J., O’Brien, C., and Darby, C. (2006). North sea cod recovery? ICES J. Mar. Sci.63, 961–968.
Houde, E. D. (1989). Comparative growth, mortality, and energetics of marine fish larvae:Temperature and latitudinal effects. Fish. Bull. 87, 471–495.
Huey, R. B., and Berrigan, D. (2001). Temperature, demography, and ectotherm fitness.Am. Nat. 158, 204–210.
Hulme, M., Jenkins, G. J., Lu, X., Turnpenny, J. R., Mitchel, l.T. D., Jones, R. G.,Lowe, J., Murphey, J. M., Hassel, l.D., Boorman, P., McDonald, R., and Ed, H. S.(2002). ‘‘Climate Change Scenarios for the United Kingdom: The UKCIP 2002.Scientific Report,’’ 120pp. Tyndall Centre for Climate Change Research, Norwich.

Climate and Fishing Effects on Cod 261
Author’s personal copy
Hurrell, J. W., and Van Loon, H. (1997). Decadal variations in climate associated with theNorth Atlantic Oscillation. Clim. Change 36, 301–326.
Husebo, A., Imsland, A.K., and Navedal, G. (2004). Haemoglobin variation in cod:A description of new variants and their geographical distribution. Sarsia 89, 369–378.
Hussy, K., Mosegaard, H., and Jessen, F. (2004). Effect of age and temperature on amino acidcomposition and the content of different protein types of juvenile Atlantic cod (Gadusmorhua) otoliths. Can. J. Fish. Aquat. Sci. 61, 1012–1020.
Hutchings, J. A. (1996). Spatial and temporal variation in the density of northern cod and areview of hypotheses for the stock’s collapse. Can. J. Fish. Aquat. Sci. 53, 943–962.
Hutchings, J., and Myers, R. (1994a). Timing of cod reproduction: Interannual variabilityand the influence of temperature. Mar. Ecol. Prog. Ser. 108, 21–31.
Hutchings, J., and Myers, R. (1994b). What can be learned from the collapse of a renewableresource? Atlantic Cod, Gadus morhua, off Newfoundland and Labrador. Can. J. Fish.Aquat. Sci. 51, 2126–2146.
Hutchinson, W. F. (2008). The dangers of ignoring stock complexity in fishery manage-ment: The case of the North Sea cod. Biol. Lett. 4, 693–695.
Hutchinson, W. F., Carvalho, G. R., and Rogers, S. I. (2001). Marked genetic structuring inlocalised spawning populations of cod Gadus morhua in the North Sea and adjoiningwaters, as revealed by microsatellites. Mar. Ecol. Prog. Ser. 223, 251–260.
Hutchinson, W. F., van Oosterhout, C., Rogers, S. I., and Carvalho, G. R. (2003).Temporal analysis of archived samples indicates marked genetic changes in decliningNorth Sea cod. Proc. R. Soc. B 270, 2125–2132.
ICES (2000). Report of the North-Western Working Group, p. 338, ICES CM 2000/ACM:15.
ICES (2003). Report of the ICES Advisory Committee on Fishery Management andAdvisory Committee on Ecosystems. ICES Advice.
ICES (2004). Report of the ICES Advisory Committee on Fishery Management andAdvisory Committee on Ecosystems, p. 1544. ICES Advice.
ICES (2005). ICES Fishmap: cod http://www.ices.dk/marineworld/fishmap/ices/default.asp?id ¼ Cod.
ICES (2006). Report of the ICES Advisory Committee on Fishery Management andAdvisory Committee on Ecosystems. ICES Advice.
ICES (2007). Report of the ICES Advisory Committee on the Marine Environment andAdvisory Committee on Ecosystem, p. 249. ICES Advise. Book 6.
ICES (2008). Report of the ICES Advisory Committee on Fishery Management andAdvisory Committee on Ecosystems. ICES Advice, Books 1–10, 1842.
Imsland, A. K., Jonsdottir, O.D. B., and Danıelsdottir, A. K. (2004). Nuclear DNA RFLPvariation among Atlantic cod in south and south-east Icelandic waters. Fish. Res. 67,227–233.
IPCC (2001). Climate change 2001: The scientific basis. In ‘‘Contribution of WorkingGroup I to the Third Assessment Report of the Intergovernmental Panel on ClimateChange.’’ ( J. T. Houghton, Y. Ding, D. J. Griggs, M. Noguer, P. J. van der Linden,X. Dai, K. Maskell, and C. A. Johnson, eds.),873pp. IPCC, Geneva.
IPCC (2007). Climate change 2007 synthesis report. In ‘‘Core Writing Team.’’(R. K. Pachauri and A. Reisinger, eds.),104pp. IPCC, Geneva.
Jackson, J. B. C., Kirby, M. X., Berger, W. H., Bjorndal, K. A., Botsford, L. W.,Bourque, B. J., Bradbury, R. H., Cooke, R., Erlandson, J., Estes, J. A., Hughes, T. P.,Kidwell, S., et al. (2001). Historical overfishing and the recent collapse of coastalecosystems. Science 293, 629–637.
Jennings, S., Greenstreet, S. P. R., and Reynolds, J. D. (1999). Structural change in anexploited fish community: A consequence of differential fishing effects on species withcontrasting life histories. J. Anim. Ecol. 68, 617–627.

262 Nova Mieszkowska et al.
Author’s personal copy
Jensen, A. S., and Hansen, P. M. (1931). Investigations on the Greenland Cod (Gaduscallareias L.) with an introduction on the Greenland cod fisheries. ICES Rapp. P.-V.Reun. 72, 1–41.
Jobling, M. (1988). A review of the physiological and nutritional energetics of cod, Gadusmorhua L., with particular reference to growth under farmed conditions. Aquaculture 70,1–19.
Jobling, M. (1994). Fish Bioenergetics, pp. 309. Chapman & Hall, London.Jonsdottir, O.D. B., Imsland, A. K., Danıelsdottir, A. K., Thorsteinsson, V., and Nævdal, G.
(1999). Genetic differentiation amongAtlantic cod in south and south-east Icelandic waters:Synaptophysin (Syp I) and haemoglobin (HbI) variation. J. Fish Biol. 54, 1259–1274.
Jonsdottir, O. D. B., Danıelsdottir, A. K., and Nævdal, G. (2001). Genetic differentiationamong Atlantic cod (Gadus morhua L.) in Icelandic waters: Temporal stability. ICES J.Mar. Sci. 58, 114–122.
Jonsdottir, O. D. B., lmsland, A. K., Danielsdottir, A. K., and Marteinsdottir, G. (2002).Genetic heterogeneity and growth properties of different genotypes of Atlantic cod(Gadus morhua L.) at two spawning sites off south lceland. Fish. Res. 55, 37–47.
Jorgensen, T. (1990). Long-term changes in age at sexual maturity of Northeast Arctic cod(Gadus morhua L.). ICES J. Mar. Sci. 46, 235–248.
Jorgensen, T. (1992). Long-term changes in growth of North-east Arctic cod (Gadus morhua)and some environmental influences. ICES J. Mar. Sci. 49, 263–278.
Jonsdottir, O. D. B., Imsland, A. K., Atladottir, O. Y., and Danıelsdottir, A. K. (2003).Nuclear DNA RFLP variation of Atlantic cod in the North Atlantic Ocean. Fish. Res.63, 429–436.
Jordan, A. D., Lampe, J. F., Grisdale-Helland, B., Helland, S. J., Shearer, K. D., andSteffansen, J. F. (2006). Growth of Atlantic cod (Gadus morhua L.) with differenthaemoglobin subtypes when kept near their temperature preferenda. Aquaculture 257,44–52.
Jorde, P. E., Knutsen, H., Espeland, S. H., and Stenseth, N. C. (2007). Spatial scale ofgenetic structuring in coastal cod Gadus morhua and geographic extent of local popula-tions. Mar. Ecol. Prog. Ser. 343, 229–237.
Kelsch, S. W., and Neill, W. H. (1990). Temperature preference versus acclimation in fishes:Selection for changing metabolic optima. Trans. Am. Fish. Soc. 119, 601–610.
Kjesbu, O. S., Klungs�yr, J., Kryvi, H., Witthames, P. R., and Greer Walker, M. (1991).Fecundity, atresia, and egg size of captive Atlantic cod (Gadus morhua) in relation toproximate body composition. Can. J. Fish. Aquat. Sci. 48, 2333–2343.
Kjesbu, O. S., Solemdal, P., Bratland, P., and Fonn, M. (1994). Variation in annual eggproduction in individual captive Atlantic cod (Gadus morhua).Can. J. Fish. Aquat. Sci. 53,610–620.
Kjesbu, O. S., Solemdal, P., Bratland, P., and Fonn, M. (1996). Variation in annual eggproduction in individual captive Atlantic cod (Gadus morhua).Can. J. Fish. Aquat. Sci. 53,610–620.
Krivobok, M. N., and Tokareva, G. I. (1972). Dynamics of weight variations of the bodyand individual organs of the Baltic cod during the maturation of gonads. Trans. Ser. Fish.Res. Board Can. 2722, 45–55.
Krohn, M., Reidy, S., and Kerr, S. (1996). Bioenergetic analysis of the effects of temperatureand pray availability on growth and condition of northern cod (Gadus morhua). Can. J.Fish Aquat. Sci. 54, 113–121.
Kuikka, S., Hilden, M., Gislason, H., Hansson, S., Sparholt, H., and Varis, O. (1999).Modeling environmentally driven uncertainties in Baltic cod (Gadus morhua) manage-ment by Bayesian influence diagrams. Can. J. Fish. Aquat. Sci. 56, 629–641.
Kurlansky, M. (1997). ‘‘Cod. A Biography of the Fish that Changed the World,’’ p. 294.Jonathan Cape, London.

Climate and Fishing Effects on Cod 263
Author’s personal copy
Lage, C., Kuhn, K., and Kornfield, I. (2004). Genetic differentiation among Atlantic cod(Gadus morhua) from Browns Bank, Georges Bank, and Nantucket Shoals. Fish. Bull.102, 289–297.
Lambert, Y., and Dutil, J. D. (1997). Condition and energy reserves of Atlantic cod (Gadusmorhua) during the collapse of the northern Gulf of St. Lawrence stock. Can. J. Fish.Aquat. Sci. 54, 2388–2400.
Lambert, Y., and Dutil, J. D. (2000). Energetic consequences of reproduction in Atlantic cod(Gadus morhua) in relation to spawning level of somatic energy reserves. Can. J. Fish.Aquat. Sci. 57, 815–825.
Lannig, G., Bock, C., Sartoris, F. J., and Portner, H. O. (2004). Oxygen limitation ofthermal tolerance in cod, Gadus morhua L., studied by magnetic resonance imaging andon-line venous oxygen monitoring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 287,902–910.
Laurence, G. C. (1978). Comparative growth, respiration and delayed feeding abilities oflarval cod (Gadus morhua) and Haddock (Melanogrammus aeglefinus) as influenced bytemperature during laboratory studies. Mar. Biol. 50, 1–7.
Leggett, W. C., and Deblois, E. (1994). Recruitment in marine fishes. Is it regulated bystarvation and predation in the egg and larval stages? Neth. J. Sea Res. 32, 119–134.
Lewis, J. R. (1986). Latitudinal trends in reproduction, recruitment and population char-acteristics of some rocky littoral molluscs and cirripedes. Hydrobiologia 142, 1–13.
Lewis, J. R. (1996). Coastal benthos and global warming: Strategies and problems.Mar. Poll.Bull. 32, 698–700.
Lilly, G. R., Shelton, P. A., Brattey, J., Cadigan, N. G., Healey, B. P., Murphy, E. F.,Stansbury, D. E., and Chen, N. (2003). An assessment of the cod stock in NAFODivisions2J þ 3KL in February 2003. DFO Can. Sci. Advisory Secretariat Res. Doc. 2003/023.
Lloret, J., and Ratz, H.-J. (2000). Condition of cod (Gadus morhua) off Greenland during1982-1998. Fish. Res. 48, 79–86.
Love, R. M. (1958). Studies on the North Sea cod. III—Effects of starvation. J. Sci. FoodAgric. 9, 617–620.
Magnuson, J. J., Crowder, L. B., and Medvick, P. A. (1979). Temperature as an EcologicalResource. Am. Zool. 19, 331–343.
Malmberg, S. A., and Blindheim, J. (1994). Climate, cod and capelin in northern watersICES Marine Science Symposia. 198, 297–310.
Mantua, N. (2004). Methods for detecting regime shifts in large marine ecosystems:A review with approaches applied to North Pacific data. Prog. Oceanogr. 60, 165–182.
Margetts, A. R., and Holt, S. J. (1948). The effect of the 1939–45War on the English NorthSea trawl fisheries. Rapp. P.-V. Reun. Cons. Perm. 122, 27–46.
Marine Research Institute (2008). ‘‘English Summary of the State of Marine Stocks inIcelandic Waters 2007/2008—Prospects for the Quota Year 2008/2009.’’pp. 169–174. Marine Research Institute, Reykjavik, Iceland.
Marshall, C. T., and Frank, K. T. (1994). Geographic responses of groundfish to variation inabundance: Methods of detection and their interpretation. Can. J. Fish. Aquat. Sci. 51(4),808–816.
Marshall, C. T., Kjesbu, O. S., Yaragina, N. A., Solemdal, P., and Ulltang, O. (1998). Isspawner biomass a sensitive measure of the reproductive and recruitment potential ofNortheast Arctic cod? Can. J. Fish. Aquat. Sci. 55, 1766–1783.
Marshall, C. T., Yaragina, N. A., Adlandsvik, B., and Dolgov, A. V. (2000). Reconstructingthe stock–recruit relationship for Northeast Arctic cod using a bioenergetic index ofreproductive potential. Can. J. Fish. Aquat. Sci. 57, 2433–2442.
Marteinsdottir, G., and Petursdottir, G. (1995). Spatial and temporal variation inreproduction of Icelandic cod at Selvogsbanki and nearby coastal areas. ICES CM 15,1–19.

264 Nova Mieszkowska et al.
Author’s personal copy
Marteinsdottir, G., and Thorarinson, K. (1998). Improving the stock–recruitment relation-ships in Iceland cod (Gadus morhua) by including age diversity of spawners. Can. J. Fish.Aquat. Sci. 55, 1372–1377.
Martin, T. L., and Huey, R. B. (2008). Why ‘‘suboptimal’’ is optimal: Jensen’s inequalityand ectotherm thermal preferences. Am. Nat. 171, E102–E118.
May, A. W., Pinhorn, A. T., Wells, R., and Fleming, A. M. (1965). Cod growth andtemperature in the Newfoundland area. ICNAF Special Publication 6, 545–555.
McCauley, R. W. (1977). Laboratory methods for determining temperature preference.J. Fish. Res. Board Can. 34, 749–752.
McCauley, R. W., and Huggins, N. W. (1979). Ontogenetic and non-thermal seasonaleffects on thermal preferenda of fish. Amer. Zool. 19, 267–271.
McKenzie, R. A. (1938). Cod take smaller bites in ice-cold water. Fish. Res. Board Can.,Atlantic Progress Report 22, 12–14.
Meekan, M. G., and Fortier, L. (1996). Selection for fast growth during the larval life ofAtlantic cod Gadus morhua on the Scotian Shelf. Mar. Ecol. Prog. Ser. 137, 25–37.
Metcalfe, J. D. (2006). Fish population structuring in the North Sea: Understandingprocesses and mechanisms from studies of the movements of adults. J. Fish Biol. 69,48–65.
Mieszkowska, N., Kendall, M. A., Hawkins, S. J., Leaper, R., Williamson, P., Hardman-Mountford, N. J., and Southward, A. J. (2006). Changes in the range of some commonrocky shore species in Britain—A response to climate change? Hydrobiologia 555,241–251.
Mieszkowska, N., Hawkins, S. J., Burrows, M. T., and Kendall, M. A. (2007). Long-termchanges in the geographic distribution and population structures of some near-limitpopulations of Osilinus lineatus in Britain and Ireland. J. Mar. Biol. Assoc. U.K. 87, 1–9.
Miller, T. J., Crowder, L. B., Rice, J. A., and Marschall, E. A. (1988). Larval size andrecruitment mechanism in fishes: Toward a conceptual framework. Can. J. Fish. Aquat.Sci. 45, 1657–1670.
M�ller, D. (1966). Genetic differences between cod groups in the Lofoten area. Nature 212,824.
Morgan, M. J., Deblois, E. M., and Rose, G. A. (1997). An observation on the reaction ofAtlantic cod (Gadus morhua) in a spawning shoal to bottom trawling. Can. J. Fish. Aquat.Sci. 54, 217–223.
Moyle, P. B., and Cech, J. J., Jr. (2004). Fishes: An Introduction to lchthyology, 5. Prentice HallUpper Saddle River, NJ.
Murawski, S. A., and Idoine, J. S. (1992). Multispecies size composition: A conservativeproperty of exploited fishery systems. J. Northwest Atl. Fish. Sci. 14, 79–85.
Myers, R. A. (1998). When do environment–recruit correlations work? Rev. Fish Biol. Fish.8, 285–305.
Myers, R., and Barrowman, N. J. (1996). Is fish recruitment related to spawner abundance?Fish. Bull. US 94, 707–724.
Myers, R. A., and Cadigan, N. G. (1995a). Statistical analysis of catch at age data withcorrelated errors. Can. J. Fish. Aquat. Sci. 52, 1265–1273.
Myers, R. A., and Cadigan, N. G. (1995b). Was an increase in natural mortality responsiblefor the collapse of northern cod? Can. J. Fish. Aquat. Sci. 52, 1274–1285.
Myers, R. A., and Stokes, K. (1989). ‘‘Density-Dependent Habitat Utilization of Groundfishand the Improvement of Research Surveys.’’ International Council for the Explorationof the Sea, Charlottenlund, Denmark. ICES C.M. 1989/D:15.
Myers, R. A., Barrowman, N. J., Hutchings, J. A., and Rosenberg, A. A. (1995a). Popula-tion dynamics of exploited fish stocks at low population levels. Science 269, 1106–1108.
Myers, R. A., Mertz, G., and Barrowman, N. J. (1995b). Spatial scales of variability in codrecruitment in the North Atlantic. Can. J. Fish. Aquat. Sci. 52, 1849–1862.

Climate and Fishing Effects on Cod 265
Author’s personal copy
Myers, R. A., Hutchings, J. A., and Barrowman, N. J. (1996). Hypotheses for the decline ofcod in the North Atlantic. Mar. Ecol. Prog. Ser. 138, 293–308.
Myers, R. A., Hutchings, J. A., and Barrowman, N. J. (1997). Why do fish stocks collapse?The example of cod in Atlantic Canada. Ecol. Appl. 7, 91–106.
Myers, R. A., MacKenzie, B. R., Bowen, K. G., and Barrowman, N. J. (2001). What is thecarrying capacity for fish in the ocean? A meta-analysis of population dynamics of NorthAtlantic cod. Can. J. Fish. Aquat. Sci. 58, 1464–1476.
Namias, J., and Cayan, D. R. (1981). Large-Scale Air-Sea interactions and Short-PeriodClimatic Fluctuatioins. Science 214, 869–876.
Neat, F. C., and Righton, D. (2007). Warm water occupancy by North Sea cod. Proc. R.Soc. B 274, 789–798.
Neat, F. C., Wright, P. J., Zuur, A. F., Gibb, I. M., Gibb, F. M., Tulett, D., Righton, D. A.,and Turner, R. J. (2006). Residency and depth movements of a coastal group of Atlanticcod (Gadus morhua L.). Mar. Biol. 148, 643–654.
Nielsen, E. E., Hansen, M. M., Schmidt, C., Meldrup, D., and Gronk, J. P. (2001).Fisheries: Population of origin of Atlantic cod. Nature 413, 272.
Nielsen, E. E., Hansen, M. M., Ruzzante, D. E., Meldrup, D., and Gronkjaer, P. (2003).Evidence of a hybrid-zone in Atlantic cod (Gadus morhua) in the Baltic and the DanishBelt Sea revealed by individual admixture analysis. Mol. Ecol. 12, 1497–1508.
Nielsen, E. E., Hansen, M. M., and Meldrup, D. (2006). Evidence of microsatellite hitch-hiking selection in Atlantic cod (Gadus morhua L.). Implications for inferring populationstructure in nonmodel organisms. Mol. Ecol. 15, 3219–3229.
Nissling, A. (2004). Effects of temperature on egg and larval survival of cod (Gadus morhua)and sprat (Sprattus sprattus) in the Baltic Sea—Implications for stock development.Hydrobiologia 514, 115–123.
Nissling, A., Larson, R., Vallin, L., and Frohlund, K. (1998). Assessment of egg and larvalviability in cod, Gadus morhua—methods and results from an experimental study. Fish.Res. 38, 169–186.
NSRAC (2006). ‘‘North Sea RAC Position Paper on the Review of Cod RecoveryMeasures.’’ p. 6. The North Sea Regional Advisory Council.
NSRAC (2008). ‘‘North Sea RAC Position Paper on the Review of the Cod RecoveryPlan.’’ p. 6. The North Sea Regional Advisory Council.
O’Brien, L. (1999). Factors influencing the rate of sexual maturity and the effect onspawning stock for Georges Bank and Gulf of Maine Atlantic cod Gadus morhua stocks.J. Northwest Atl. Fish. Sci. 25, 179–203.
O’Brien, L., Burnett, J., and Mayo, R. K. (1993). ‘‘Maturation of Nineteen Species of Finfishoff theNortheast Coast of theUnited States.’’ p. 66. NOAATechnical Report.NMFS 113,Seattle, WA.
O’Brien, C. M., Fox, C. J., Planque, B., and Casey, J. (2000). Fisheries: Climate variabilityand North Sea cod. Nature 404, 142.
O’Leary, D. B., Coughlan, J., Dillane, E., McCarthy, T. V., and Cross, T. F. (2007).Microsatellite variation in cod Gadus morhua throughout its geographic range. J. FishBiol. 70, 310–335.
Olsen, E. M., Heino, M., Lilly, G. R., Morgan, M. J., Brattey, J., Ernande, B., andDieckmann, U. (2004). Maturation trends indicative of rapid evolution preceded thecollapse of northern cod. Nature 428, 932–935.
Olsen, E. M., Knutsen, H., Gj�sæter, J., Jorde, P. E., Knutsen, J. A., and Stenseth, N. C.(2008). Small-scale biocomplexity in coastal Atlantic cod supporting a Darwinian per-spective on fisheries management. Evol. Appl. 1, 524–533.
Orton, J. H. (1920). Sex-phenomena in the common limpet (Patella vulgata).Nature 104, 373.Ottera, H., Agnalt, A.-L., and J�rstad, K. E. (2006). Differences in spawning time of captive
Atlantic cod from four regions of Norway, kept under identical conditions. ICES J. Mar.Sci. 63, 216–223.

266 Nova Mieszkowska et al.
Author’s personal copy
Otterlei, E., Nyhammer, G., Folkvord, A., and Stefansson, S. O. (1999). Temperature- andsize-dependent growth of larval and early juvenile Atlantic cod (Gadus morhua).A comparative study of Norwegian coastal cod and Northeast Arctic cod. Can. J. Fish.Aquat. Sci. 56, 2099–2111.
Ottersen, G. (1996). Environmental impact on variability in recruitment, larval growth anddistribution of Arcto-Norwegian cod, p. 137. University of Bergen.
Ottersen, G., Loeng, H., and Raknes, A. (1994). Influence of temperature variability onrecruitment of cod in the Barents Sea. ICES Mar. Sci. Symp. 198, 471–481.
Ottersen, G., and Sundby, S. (1995). Effects of temperature, wind and spawning stockbiomass on recruitment of Arcto-Norwegian cod. Fish. Oceanogr. 4, 278–292.
Ottersen, G., and Loeng, H. (2000). Covariability in early growth and year-class strength ofBarents Sea cod, haddock, and herring: The environmental link. ICES J. Mar. Sci. 57,339–348.
Ottersen, G., Planque, B., Belgrano, A., Post, E., Reid, P. C., and Stenseth, N. C. (2001).Ecological effects of the North Atlantic Oscillation. Oecologia. 128, 1–14.
Ottersen, G., Hjermann, D., and Stenseth, N. C. (2006). Changes in spawning stockstructure strengthens the link between climate and recruitment in a heavily fished codstock. Fish. Oceanogr. 15, 230–243.
Palsson, O. K., and Thorsteinsson, V. (2003). Migration patterns, ambient temperature, andgrowth of Icelandic cod (Gadus morhua): Evidence from storage tag data. Can. J. Fish.Aquat. Sci. 60, 1409–1423.
Pardini, A. T., Jones, C. S., Noble, L. R., Kreiser, B., Malcolm, H., Bruce, B. D.,Stevens, J. D., Cliff, G., Scholl, M. C., Francis, M., Duffy, C. A., and Martin, A. P.(2001). Sex-biased dispersal of great white sharks. Nature 412, 139–140.
Pastoors, M. (2005). Evaluating fisheries management advice for some North Sea stocks:Is bias inversely related to stock size? ICES Document CM 2005/V 20, 1–14.
Pauly, D., and Pullin, R. S. V. (1988). Hatching time in spherical, pelagic, marine fish eggs inresponse to temperature and egg size. Environ. Biol. Fishes 22, 261–271.
Pauly, D., Christensen, V., Dalsgaard, J., Froese, R., and Torres, F. Jr. (1998). Fishing downmarine food webs. Science 279, 860–863.
Pearson, R. G., and Dawson, T. P. (2003). Predicting the impacts of climate change on thedistribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 12,361–371.
Peck, M. A., Buckley, L. J., Caldarone, E. M., and Bengtson, D. A. (2003). Effects of foodconsumption and temperature on growth rate and biochemical-based indicators of growthin early juvenile Atlantic cod Gadus morhua and haddock Melanogrammus aeglefinus. Mar.Ecol. Prog. Ser. 251, 233–243.
Pepin, P., and Myers, R. (1991). Significance of egg and larval size to recruitment variabilityof temperate marine fish. Can. J. Fish. Aquat. Sci. 48, 1820–1828.
Pepin, P., Orr, D. C., and Anderson, J. T. (1997). Time to hatch and larval size in relation totemperature and egg size in Atlantic cod (Gadus morhua). Can. J. Fish. Aquat. Sci. 54, 2–10.
Perez-Casanova, J. C., Afonso, L. O. B., Johnson, S. C., Currie, S., and Gamperl, A. K.(2008). The stress and metabolic responses of juvenile Atlantic codGadus morhua L. to anacute thermal challenge. J. Fish Biol. 72, 899–916.
Perry, A. L., Low, P. J., Ellis, J. R., and Reynolds, J. D. (2005). Climate change anddistribution shifts in marine fishes. Science 308, 1912–1915.
Petersen, M. F., and Steffensen, J. F. (2003). Preferred temperature of juvenile Atlantic codGadus morhua with different haemoglobin genotypes at normoxia and moderate hypoxia.J. Exp. Biol. 206, 359–364.
Pilling, G. M., Millner, R. S., Easey, M. W., Maxwell, D. L., and Tidd, A. N. (2007).Phenology and North Sea cod Gadus morhua L.: Has climate change affected otolithannulus formation and growth? J. Fish Biol. 70, 584–599.

Climate and Fishing Effects on Cod 267
Author’s personal copy
Pinnegar, J. K., Jennings, S., O’Brien, C. M., and Polunin, N. V. C. (2002). Long-termchanges in the trophic level of the Celtic Sea fish community and fish market pricedistribution. J. Appl. Ecol. 39, 377–390.
Pitcher, T. (2008). The sea ahead: challenges to marine biology from seafood sustainability.Hydrobiologia 606, 161–185.
Pitcher, T., and Forrest, R. (2004). Challenging ecosystem simulation models with climatechange: The ‘perfect storm’. Fish. Centre Res. Rep. 12, 29–38.
Pitcher, T., and Pauly, D. (1998). Rebuilding ecosystems, not sustainability, as the propergoal of fishery management. In ‘‘Reinventing Fisheries Management.’’ (T. J. Pitcher, P. J.B. Hart and D. Pauly, eds.), pp. 311–329. Kluwer Academic Publishers, Dordrecht.
Planque, B., Fox, C. J., Saunders, M. A., and Hamilton, T. (2003). Statistical prediction ofshort term changes in the recruitment of North Sea cod (Gadus Morhua) using tempera-ture forecasts. Sci. Mar. 67, 211–218.
Planque, B., and Fredou, T. (1999). Temperature and the recruitment of Atlantic cod (Gadusmorhua). Can. J. Fish. Aquat. Sci. 56, 2069–2077.
Platt, T., Fuentes-Yaco, C., and Frank, K. T. (2003). Marine ecology: Spring algal bloomand larval fish survival. Nature 423, 398–399.
Pogson, G. H., Mesa, K. A., and Boutilier, R. G. (1995). Genetic population structure andgene flow in the Atlantic codGadus morhua: A comparison of allozyme and nuclear RFLPLoci. Genetics 139, 375–385.
Pogson, G. H., Taggart, C. T., Mesa, K. A., and Boutilier, R. G. (2001). Isolation bydistance in the Atlantic cod, Gadus morhua, at large and small geographic scales. Evolution55, 131–146.
Portner, H. O. (2001). Climate change and temperature-dependent biogeography: Oxygenlimitation of thermal tolerance in animals. Naturwissenschaften 88, 137–146.
Portner, H. O. (2002). Climate variations and the physiological basis of temperaturedependent biogeography: Systemic to molecular hierarchy of thermal tolerance inanimals. Comp. Biochem. Physiol. A 132, 739–761.
Portner, H. O., and Knust, R. (2007). Climate change affects marine fishes through theoxygen limitation of thermal tolerance. Science 315, 95–97.
Portner, H. O., Berdal, B., Blust, R., Brix, O., Colosimo, A., De Wachter, B., Giuliani, A.,Johansen, T., Fischer, T., Knust, R., Lannig, G., Naevdal, G., et al. (2001). Climateinduced temperature effects on growth performance, fecundity and recruitmentin marine fish: Developing a hypothesis for cause and effect relationships in Atlanticcod (Gadus morhua) and common eelpout (Zoarces viviparus). Cont. Shelf Res. 21,1975–1997.
Portner, H. O., Christian, B., Knust, R., Lannig, G., Lucassen, M., Mark, F. C., andSartoris, F. J. (2008). Cod and climate in a latitudinal cline: physiological analyses ofclimate effects in marine fishes. Clim. Res. 37, 253–270.
Post, J. R., Sullivan, M., Cox, S., Lester, N. P., Walters, C. J., Parkinson, E. A., Paul, A. J.,Jackson, L., and Shuter, B. J. (2002). Canada’s recreational fisheries: the invisible collapse?Fish. Bull. US 27, 6–17.
Ratz, H.-J., and Lloret, J. (2003). Variation in fish condition between Atlantic cod (Gadusmorhua) stocks, the effect on their productivity and management implications. Fish. Res.60, 369–380.
Reeves, S. A., and Pastoors, M. A. (2007). Evaluating the science behind the managementadvice for North Sea cod. ICES J. Mar. Sci. 64, 1–8.
Reid, P. C., Holliday, N. P., and Smyth, T. J. (2001). Pulses in the eastern margin currentand warmer water off the north west European shelf linked to North Sea ecosystemchanges. Mar. Ecol. Prog. Ser. 215, 283–287.
Reynolds, W.W. (1977). Temperature as a proximate factor in orientation behavior. J. Fish.Res. Board Can. 34, 734–739.

268 Nova Mieszkowska et al.
Author’s personal copy
Rice, J. (1995). Food web theory, marine food webs, and what climate change may do tonorthern marine fish populations. In ‘‘Climate Change and Northern Fish Populations.’’(R. J. Beamish, ed.), pp. 561–568. Canadian Special Publications on Fisheries andAquatic Sciences.
Rice, J. C. (2000). Evaluating fishery impacts using metrics of community structure. ICES J.Mar. Sci. 57, 682–688.
Rice, J. C., and Evans, G. T. (1988). Tools for embracing uncertainty in the management ofthe cod fishery of NAFO divisions 2J þ 3KL. ICES J. Mar. Sci. 45, 73–81.
Rice, J., and Gislason, H. (1996). Patterns of change in the size spectra of numbers anddiversity of the North Sea fish assemblage, as reflected in surveys and models. ICES J.Mar. Sci. 53, 1214–1225.
Rice, J. C., and Rochet, M. J. (2005). A framework for selecting a suite of indicators forfisheries management. ICES J. Mar. Sci. 62, 516–527.
Righton, D., Metcalfe, J., and Connolly, P. (2001). Fisheries: Different behaviour of cod inthe North and Irish Seas. Nature 411, 156.
Righton, D., Quayle, V. A., Hetherington, S., and Burt, G. (2007). Movements anddistribution of cod (Gadus morhua) in the southern North Sea and English Channel:results from conventional and electronic tagging experiments. J. Mar. Biol. Assoc. UK 87,599–613.
Rijnsdorp, A. D., and van Leeuwen, P. I. (1996). Changes in growth of North Sea plaicesince 1950 in relation to density, eutrophication, beam-trawl effort, and temperature.ICES J. Mar. Sci. 53, 1199–1213.
Rijnsdorp, A. D., van Leeuwen, P. I., Daan, N., and Heessen, H. J. L. (1996). Changes inabundance of demersal fish species in the North Sea between 1906–1909 and 1990–1995.ICES J. Mar. Sci. 53, 1054–1062.
Rijnsdorp, A. D., Peck, M. A., Engelhard, G. H., Mollmann, C., and Pinnegar, J. K. (2009).Resolving the effect of climate change on fish populations. ICES J. Mar. Sci. 66,1570–1583.
Rillahan, C., Chambers, M., Howell, W. H., and Watson Iii, W. H. (2009). A self-contained system for observing and quantifying the behavior of Atantic cod, Gadusmorhua, in an offshore aquaculture cage. Aquaculture 293, 49–56.
Rindorf, A., and Lewy, P. (2006). Warm, windy winters drive cod north and homing ofspawners keeps them there. J. Appl. Ecol. 43, 445–453.
Rivard, D., and Maguire, J. (1993). Reference points for fisheries management: The easternCanadian experience. In ‘‘Risk Evaluation and Biological Reference Points for FisheriesManagement.’’ (S. J. Smith, J. J. Hunt, and D. Rivard, eds.), pp. 21–37. SpecialPublication in Fisheries and Aquatic Science.
Robichaud, D., and Rose, G. A. (2001). Multiyear homing of Atlantic cod to a spawningground. Can. J. Fish. Aquat. Sci. 58, 2325–2329.
Robichaud, D., and Rose, G. A. (2002). The return of cod transplanted from a spawningground in southern Newfoundland. ICES J. Mar. Sci. 59, 1285–1293.
Robichaud, D., and Rose, G. A. (2003). Sex differences in cod residency on a spawningground. Fish. Res. 60, 33–43.
Robichaud, D., and Rose, G. A. (2004). Migratory behaviour and range in Atlantic cod:Inference from a century of tagging. Fish Fish. 5, 185–214.
Rochet, M. J., and Trenkel, V. M. (2003). Which community indicators can measure theimpact of fishing? A review and proposals. Can. J. Fish. Aquat. Sci. 60, 86–99.
Roessig, J. M., Woodley, C. M., Cech, J. J., and Hansen, L. J. (2004). Effects of globalclimate change on marine and estuarine fishes and fisheries. Rev. Fish Biol. Fish. 14,251–275.
Rose, G. (2004a). Reconciling overfishing and climate change with stock dynamics ofAtlantic cod (Gadus morhua) over 500 years. Can. J. Fish. Aquat. Sci. 61, 1553–1557.

Climate and Fishing Effects on Cod 269
Author’s personal copy
Rose, G. A. (2004b). On distributional responses of North Atlantic fish to climate change,In ‘‘ICES Symposium on Influence of Climate Change on North Atlantic Fish Stocks.’’pp. 1360–1374.
Rose, G. A., and Kulka, D. W. (1999). Hyperaggregation of fish and fisheries: How catch-per-unit-effort increased as the northern cod (Gadus morhua) declined.Can. J. Fish.Aquat.Sci. 56, 118–127.
Rose, G. A., Atkinson, B. A., Baird, J., Bishop, C. A., and Kulka, D. W. (1994). Changes indistribution of Atlantic cod and thermal variations in Newfoundland waters, 1980–1992.ICES Mar. Sci. Symp. 198, 542–552.
Rose, G. A., deYoung, B., Kulka, D. W., Goddard, S. V., and Fletcher, G. L. (2000).Distribution shifts and overfishing the northern cod (Gadus morhua). A view from theocean. Can. J. Fish. Aquat. Sci. 57, 644–663.
Rosenberg, A. A., Bolster, W. J., Alexander, K. E., Leavenworth, W. B., Cooper, A. B., andMcKenzie, M. G. (2005). The history of ocean resources: Modeling cod biomass usinghistorical records. Front. Ecol. Environ. 3, 78–84.
Rothschild, B. J. (1994). Decadal transients in biological productivity, with special referenceto the cod populations of the North Atlantic. ICES Mar. Sci. Symp. 198, 333–345.
Rothschild, B. J. (1998). Year class strengths of zooplankton in the North Sea and theirrelation to cod and herring abundance. J. Plankton Res. 20, 1721–1741.
Rowe, S., and Hutchings, J. A. (2003). Mating systems and the conservation of commer-cially exploited marine fish. Trends Ecol. Evol. 18, 567–572.
Rowe, S., Hutchings, J. A., Bekkevold, D., and Rakitin, A. (2004). Depensation, probabilityof fertilization, and the mating system of Atlantic cod (Gadus morhua L.). ICES J. Mar. Sci.61, 1144–1150.
Rowe, S., Hutchings, J. A., and Skjæraasen, J. E. (2007). Non-random mating in a broadcastspawner: Mate size influences reproductive success in Atlantic cod (Gadus morhua). Can.J. Fish. Aquat. Sci. 64, 219–226.
Rudolfsen, G., Figenschou, L., Folstad, I., Nordeide, J. T., and S�reng, E. (2005). Potentialfitness benefits from mate selection in the Atlantic cod (Gadus morhua). J. Evol. Biol. 18,172–179.
Ruzzante, D. E., Taggart, C. T., and Cook, D. (1996a). Spatial and temporal variation in thegenetic composition of a larval cod (Gadus morhua) aggregation: Cohort contribution andgenetic stability. Can. J. Fish. Aquat. Sci. 53, 2695–2705.
Ruzzante, D. E., Taggart, C. T., Cook, D., and Goddard, S. V. (1996b). Genetic differenti-ation between inshore and offshore Atlantic cod (Gadus morhua) off Newfoundland:Microsatellite DNA variation and antifreeze protein levels. Can. J. Fish. Aquat. Sci. 5,634–645.
Ruzzante, D. E., Taggart, C. T., Cook, D., and Goddard, S. V. (1997). Genetic differentia-tion between inshore and offshore Atlantic cod (Gadus morhua) off Newfoundland: A test,and evidence of temporal stability. Can. J. Fish. Aquat. Sci. 5, 2700–2708.
Ruzzante, D. E., Taggart, C. T., and Cook, D. (1998). A nuclear DNA basis for shelf- andbank-scale population structure in Northwest Atlantic cod (Gadus morhua). Labrador toGeorges Bank. Mol. Ecol. 7, 1663–1680.
Ruzzante, D. E., Taggart, C. T., and Cook, D. (1999). A review of the evidence for geneticstructure of cod (Gadus morhua) populations in the Northwest Atlantic and populationaffinities of larval cod off Newfoundland and the Gulf of St. Lawrence. Fish. Res. 43, 79–97.
Ruzzante, D. E., Wroblewskic, J. S., Taggart, C. T., Smedbol, R. K., Cook, D., andGoddard, S. V. (2000). Bay-scale population structure in coastal Atlantic cod in Labradorand Newfoundland, Canada. J. Fish Biol. 56, 431–447.
Ruzzante, D. E., Taggart, C. T., Doyle, R. W., and Cook, D. (2001). Stability in thehistorical pattern of genetic structure of Newfoundland cod (Gadus morhua) despite thecatastrophic decline in population size from 1964 to 1994. Conserv. Genet. 2, 257–269.

270 Nova Mieszkowska et al.
Author’s personal copy
Sæmundsson, B. (1934). Probable influence of changes in temperature on the marine faunaof Iceland. ICES Rapp. P.-V. Reun. 86, 1–16.
Sætersdal, G., and an Loeng, H. (1987). Ecological adaptation of reproduction in northeastArctic cod. Fisheries Research 5(2-3), 253–270.
Sagarin, R. D., Barry, J. P., Gilman, S. E., and Baxter, C. H. (1999). Climate-related changein an intertidal community over short and long time scales. Ecol. Monogr. 69, 465–490.
Sartoris, F. J., Bock, C., Serendero, I., Lannig, G., and Portner, H. O. (2003). Temperature-dependent changes in energy metabolism, intracellular pH and blood oxygen tension inthe Atlantic cod. J. Fish Biol. 62, 1239–1253.
Schiermeier, Q. (2002). How many more fish in the sea? Nature 419, 662–665.Schiermeier, Q. (2004). Climate findings let fishermen off the hook. Nature 428, 4.Schopka, S. A. (1994). Fluctuations in the cod stock off Iceland during the twentieth century
in relation to changes in the fisheries and environment. ICES Mar. Sci. Symp. 198,175–193.
Schurmann, H., and Steffensen, J. F. (1997). Effects of temperature, hypoxia and activity onthe metabolism of juvenile Atlantic cod. J. Fish Biol. 50, 1166–1180.
Scott, J. S. (1982). Depth, temperature and salinity preferences of common fishes of theScotian Shelf. J. Northwest Atl. Fish. Sci. 3, 29–39.
SGMOS (2003). ‘‘Sub-Group on Management Objectives of the STECF.’’ Recovery plansfor southern hake and Norway lobster in ICES areas IXa and VIIIc, Lisbon.
Shelton, P. A., and Healey, B. P. (1999). Should depensation be dismissed as a possibleexplanation for the lack of recovery of the northern cod (Gadus morhua) stock? Can. J.Fish. Aquat. Sci. 56, 1521–1524.
Shelton, P. A., Sinclair, A. F., Chouinard, G. A., Mohn, R., and Duplisea, D. E. (2006).Fishing under low productivity conditions is further delaying recovery of NorthwestAtlantic cod (Gadus morhua). Can. J. Fish. Aquat. Sci. 63, 235–238.
Shepherd, J. G., and Cushing, D. H. (1980). A mechanism for density-dependent survival oflarval fish as the basis of a stock–recruitment relationship. J. Cons. Int. Explor. Mer. 185,255–267.
Shin, Y. J., Rochet, M. J., Jennings, S., Field, J. G., and Gislason, H. (2005). Using size-based indicators to evaluate the ecosystem effects of fishing. ICES J. Mar. Sci. 62,384–396.
Sick, K. (1965). Haemoglobin polymorphism of cod in the Baltic and the Danish Belt Sea.Hereditas 54, 19–48.
Sims, D. W. (2008). Tracking fish with chips: Revealing marine fish movements, behaviourand population structure through technology. J. Mar. Des. Operations 13, 47–58.
Sims, D. W., and Southward, A. J. (2006). Dwindling fish stocks already of concern in 1883.Nature 439, 660.
Sims, D. W., Genner, M. J., Southward, A. J., and Hawkins, S. J. (2001). Timing ofsquid migration reflects North Atlantic climate variability. Proc. R. Soc. B 268,2607–2611.
Sims, D. W., Wearmouth, V. J., Genner, M. J., Southward, A. J., and Hawkins, S. J. (2004).Low-temperature-driven early spawning migration in a temperate marine fish. J. Anim.Ecol. 73, 333–341.
Skarstein, T. H., Westgaard, J. I., and Fevolden, S. E. (2007). Comparing microsatellitevariation in Northeast Atlantic cod (Gadus morhua L.) to genetic structuring as revealed bythe pantophysin (Pan I) locus. J. Fish Biol. 70, 271–290.
Smith, T. D. (1994). ‘‘Scaling Fisheries: The Science of Measuring the Effects of Fishing,1855–1955.’’ p. 404. Cambridge University Press, Cambridge.
Solberg, T. S., and Tilseth, S. (1987).Variations in growth pattern among yolk-sac larvae ofcod (Gadus morhua L.) due to differences in rearing temperature and light regime. 72,pp. 347–349.

Climate and Fishing Effects on Cod 271
Author’s personal copy
Soofiani, N. M., and Hawkins, A. D. (1982). Energetic costs at different feeding levels injuvenile cod, Gadus morhua L. J. Fish Biol. 21, 577–592.
Soofiani, N. M., and Priede, I. G. (1985). Aerobic metabolic scope and swimming perfor-mance in juvenile cod, Gadus morhua L. J. Fish Biol. 26, 127–138.
Southward, A. J. (1963). The distribution of some plankton animals in the English Channeland Western Approaches. III. Theories about long-term biological changes, includingfish. J. Mar. Biol. Assoc. UK 43, 1–29.
Southward, A. J. (1995). The importance of long time-series in understanding the variabilityof natural systems. Helgol. Meeres. 49, 329–333.
Southward, A. J., and Boalch, G. T. (1994). The Effect of Changing Climate onMarine Life:Past Events and Future Predictions. In ‘‘Man and the Maritime Environment.’’ (S. Fisher,ed.), Vol. 9, pp. 101–143. Exeter Maritime Studies. University of Exeter, Exeter.
Southward, A. J., Boalch, G. T., and Maddock, L. (1988). Fluctuations in the herring andpilchard fisheries of Devon and Cornwall linked to change in climate since the 16thcentury. J. Mar. Biol. Assoc. UK 68, 423–445.
Sparre, P. (1991). Introduction to multispecies virtual population analysis. ICES Mar. Sci.Symp. 193, 12–21.
STECF (2007). ‘‘Commissioned Staff Working Paper 23rd Report of the Scientific, Tech-nical and Economic Committee for Fisheries (Spring Plenary Meeting).’’ Commission ofthe European Communities, Brussels.
Steele, D. H., Andersen, R., and Green, J. M. (1992). The managed commercial annihila-tion of northern cod. Newfoundland Stud. 8, 34–68.
Steinarsson, A., and Bjornsson, B. (1999). The effects of temperature and size on growth andmortality of cod larvae. J. Fish Biol. 55, 100–109.
Steinhausen, M. F., Andersen, N. G., and Steffensen, J. F. (2006). The effect of externaldummy transmitters on oxygen consumption and performance of swimming Atlanticcod. J. Fish Biol. 69, 951–956.
Stenseth, N. C., Mysterud, A., Ottersen, G., Hurrell, J. W., Chan, K.-S., and Lima, M.(2002). Ecological Effects of Climate Fluctuations. Science 297, 1292–1296.
Stenseth, N. C., Ottersen, G., Hurrell, J. W., Mysterud, A., Lima, M., Chan, K.-S., andYoccoz, N. G. (2003). Studying climate effects on ecology through the use of climateindices: the North Atlantic Oscillation, El Nio Southern Oscillation and beyong. Proc.Roy. Soc. London B. 270, 2087–2096.
Stenseth, N. C., Ottersen, G., Hurrell, J. W., and Belgrano, A. (2004). ‘‘Marine Ecosystemsand Climate Variation. The North Atlantic: A Comparative Perspective.’’ p. 236. OxfordUniversity Press, Oxford.
Stenson, G. B., Hammill, M. O., and Lawson, J. W. (1997). Predation by Harp Seals inAtlantic Canada: Preliminary Consumption Estimates for Arctic Cod, Capelin andAtlantic Cod. J. Northwest Atl. Fish. Sci. 22, 137–154.
Stephens, P. A., and Sutherland, W. J. (1999). Consequences of the Allee effect forbehaviour, ecology and conservation. Trends Ecol. Evol. 14, 401–405.
Stige, L. C., Ottersen, G., Brander, K., Chan, K. S., and Stenseth, N. C. (2006). Cod andclimate: Effect of the North Atlantic Oscillation on recruitment in the North Atlantic.Mar. Ecol. Prog. Ser. 325, 227–241.
Sundby, S. (2000). Recruitment of Atlantic cod stocks in relation to temperature andadvection of copepod populations. Sarsia 85, 277–298.
Swain, D. P. (1999). Changes in the distribution of Atlantic cod (Gadus morhua) in thesouthern Gulf of St Lawrence; effects of environmental change or change in environ-mental preferences? Fish. Oceanog. 8, 1–17.
Swain, D. P., and Wade, E. J. (1993). Density-Dependent Geographic Distribution ofAtlantic Cod (Gadus morhua) in the Southern Gulf of St. Lawrence. Can. J. Fish.Aquat. Sci. 50, 725–733.

272 Nova Mieszkowska et al.
Author’s personal copy
Swain, D. P., and Sinclair, A. F. (1994). Fish distribution and catchability: What is theappropriate measure of distribution? Can. J. Fish. Aquat. Sci. 51, 1046–1054.
Taggart, C. T., Anderson, J., Bishop, C., Colbourne, E., Hutchings, J., Lilly, G., Morgan, J.,Murphy, E., Myers, R., Rose, G., and Shelton, P. (1993). Overview of cod stocks,biology, and environment in the Northwest Atlantic region of Newfoundland, withemphasis on northern cod. Symposium on Cod and Climate Change, pp. 140–157.
Taylor, C. C. (1958). Cod growth and temperature. J. Conseil. 23, 366–370.Taylor, M. I., Rico, I., Fox, C., and Rico, C. (2002). Species specific Taqman probes
for simultaneous identification of cod (Gadus morhua L.), haddock (Melanogrammusaeglefinus L.) and whiting (Merlangius merlangus L.). Mol. Ecol. Notes 2, 599–601.
Tremble, M. J., and Sinclair, M. (1985). Gulf of St. Lawrence cod: Age specific geographicaldistributions and environmental occurrences from 1971 to 1981. Can. J. Fish. Aquat. Sci.1387, p. 43.
Trenkel, V. M., and Rochet, M. J. (2003). Performance of indicators derived from abun-dance estimates for detecting the impact of fishing on a fish community. Can. J. Fish.Aquat. Sci. 60, 67–85.
Trzcinski, M., Mohn, K., Bowen, R., and Don, W. (2006). Continued decline of an Atlanticcod population: how important is the gray seal predation? Ecol. Appl. 16, 2276–2292.
Valerio, P. F., Goddard, S. V., Kao, M. H., and Fletcher, G. L. (1992). Survival of northernAtlantic cod (Gadus morhua) eggs and larvae when exposed to ice and low temperature.Can. J. Fish. Aquat. Sci. 49(12), 2588–2595.
van der Molen, J., Rogers, S. I., Ellis, J. R., Fox, C. J., and McCloghrie, P. (2007). Dispersalpatterns of the eggs and larvae of spring-spawning fish in the Irish Sea, UK. J. Sea Res. 58,313–330.
Van Dijk, P. L., Hardewig, I., and Portner, H. O. (1997). Temperature-dependent shift ofpHi in fish white muscle: Contributions of passive and active processes. Am. J. Physiol.272, 84–89.
van Kooten, T., de Roos, A. M., and Persson, L. (2005). Bistability and an Allee effect asemergent consequences of stage-specific predation. J. Theor. Biol. 237, 67–74.
Van Loon, H., and Rogers, J. (1978). The seesaw in winter temperature between Greenlandand northern Europe. Part I: General description. Mon. Weather Rev. 106, 296–310.
Vaz, S., Carpentier, A., and Coppin, F. (2007). Eastern English Channel fish assemblages:Measuring the structuring effect of habitats on distinct sub-communities. ICES J. Mar.Sci. 64, 271–287.
Vilhjalmsson, H. (1997). Climatic variations and some examples of their effects on themarine ecology of Icelandic and Greenland waters, in particular during the presentcentury. J. Mar. Res. Inst. 15, 9–29.
Vinther, M., Reeves, S. A., and Patterson, K. R. (2004). From single-species advice tomixed-species management: taking the next step. ICES J. Mar. Sci. 61, 1398–1409.
Walters, C., and Collie, J. (1988). Is research on environmental factors useful to fisheriesmanagement? Can. J. Fish. Aquat. Sci. 45, 1848–1854.
Walters, C., and Maguire, J. J. (1996). Lessons for stock assessment from the northern codcollapse. Rev. Fish Biol. Fish. 6, 125–137.
Walters, G., and Ludwig, D. (1981). Effects of measurement errors on the assessment ofstock–recruitment relationships. Can. J. Fish. Aquat. Sci. 38, 704–710.
Walther, G. R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T. J. C.,Fromentin, J. M., Guldberg, O. H., and Bairlein, F. (2002). Ecological responses torecent climate change. Nature 416, 389–395.
Walther, G. R., Berger, S., and Sykes, M. T. (2005). An ecological ‘footprint’ of climatechange. Proc. R. Soc. B 272, 1427–1432.
Ware, D. M. (1975). Relation between egg size, gorwth and natural mortality of larval fish.J. Fish. Res. Board Can. 32, 2503–2512.

Climate and Fishing Effects on Cod 273
Author’s personal copy
Wieland, K., and Jarre-Teichmann, A. (1997). Prediction of vertical distribution andambient development temperature of Baltic cod, Gadus morhua L., eggs. Fish. Oceanogr.6, 172–187.
Wieland, K., Jarre-Teichmann, A., and Horbowa, K. (2000). Changes in the timing ofspawning of Baltic cod: possible causes and implications for recruitment. ICES J. Mar.Sci. 57, 452–464.
Windle, M. J. S., and Rose, G. A. (2007). Do cod form spawning leks? Evidence from aNewfoundland spawning ground. Mar. Biol. 150, 671–680.
Wise, J. P. (1961). Synopsis of biological data on cod, Gadus Morhua Linnaeus 1758. FAOFish Synopsis. 21, p. 55.
Worm, B., and Myers, R. A. (2003). Meta-analysis of cod-shrimp interactions reveals top-down control in oceanic food webs. Ecology 84, 162–173.
Wright, P. J., Neat, F. C., Gibb, F. M., Gibb, I. M., and Thordarson, H. (2006). Evidencefor metapopulation structuring in cod from the west of Scotland and North Sea. J. FishBiol. 69, 181–199.
Yin, M. C., and Blaxter, J. H. S. (1987). Escape speeds of marine fish larvae during earlydevelopment and starvation. Mar. Bio. 96, 459–468.
Yoneda, M., and Wright, P. J. (2004). Temporal and spatial variation in reproductiveinvestment of Atlantic cod Gadus morhua in the northern North Sea and Scottish westcoast. Mar. Ecol. Prog. Ser. 276, 237–248.
Zacherl, D., Gaines, S. D., and Lonhart, S. I. (2003). The limits to biogeographicaldistributions: Insights from the northward range extension of the marine snail, Kelletiakelletii (Forbes, 1852). J. Biogeogr. 30, 913–924.
Zwanenburg, K. C. T. (2000). The effects of fishing on demersal fish communities of theScotian Shelf. ICES J. Mar. Sci. 57, 503–509.




















