
Vertical movements of North Sea cod
10
MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Vol. 347: 101–110, 2007 doi: 10.3354/meps07047 Published October 11 INTRODUCTION The way in which animals move through their envi- ronment forms a fundamental component of their ecol- ogy and, consequently, there have been efforts to understand animal movements over a range of spatial and temporal scales (Carey et al. 1971, Block 2005). In a marine context, vertical movements often play an important role for a range of animals (Hays 2003, Block 2005, Sims et al. 2006). For example, many members of the zooplankton undergo regular diel vertical move- ments (generally deeper during the day and shallower at night), often seemingly as a predator evasion mech- anism (Hays 2003). Similarly, vertical movements are also important for a range of marine vertebrates. Air- breathing marine vertebrates (Hays 2003) must come to the surface to breathe and so their vertical move- ments (manifested as dives away from the sea surface) occur regularly in association with foraging (Wilson et al. 2002), predator evasion (Heithaus & Frid 2003) or resting (Schreer et al. 2001). Over the last 20 yr, elec- tronic data-loggers and transmitters have revealed the intricacies of these vertical movements and how they vary temporally and spatially. For example, whales, seals, turtles and sharks commonly have diel patterns of resting and foraging (Schreer et al. 2001), with dive depth often varying in association with vertical move- ments of their prey (Sims et al. 2006). Similarly, these groups all show different behaviours during migration, where horizontal travel rather than foraging may be the central objective, and may also show marked spa- tial patterns in diving (Hays et al. 2006). Vertical movements have also been documented in a wide range of fish species. For example, Carey et al. (1971) described the results of deployments of acoustic transmitters that revealed the vertical movements of large pelagic fish such as bluefin tuna Thunnus sp., broadbill swordfish Xiphias gladius and the great © Inter-Research 2007 · www.int-res.com *Email: [email protected] Vertical movements of North Sea cod Victoria J. Hobson 1, 2, *, David Righton 2 , Julian D. Metcalfe 2 , Graeme C. Hays 1 1 Department of Biological Sciences, Institute of Environmental Sustainability, Swansea University, Singleton Park, Swansea SA2 8PP, UK 2 Centre for Environment, Fisheries and Aquaculture Science, Lowestoft Laboratory, Pakefield Road, Lowestoft, Suffolk NR33 0HT, UK ABSTRACT: Various air-breathing marine vertebrates such as seals, turtles and seabirds show dis- tinct patterns of diving behaviour. For fish, the distinction between different vertical behaviours is often less clear-cut, as there are no surface intervals to differentiate between dives. Using data from acoustic tags (n = 23) and archival depth recorders attached to cod Gadus morhua (n = 92) in the southern North Sea, we developed a quantitative method of classifying vertical movements in order to facilitate an objective comparison of the behaviour of different individuals. This method expands the utilisation of data from data storage tags, with the potential for a better understanding of fish behaviour and enhanced individual based behaviour for improved ecosystem modelling. We found that cod were closely associated with the seabed for 90% of the time, although they showed distinct seasonal and spatial patterns in behaviour. For example, cod tagged in the southern North Sea exhib- ited high rates of vertical movement in spring and autumn that were probably associated with migra- tion, while the vertical movements of resident cod in other areas were much less extensive and were probably related to foraging or spawning behaviours. The full reasons underlying spatial and tempo- ral behavioural plasticity by cod in the North Sea warrant further investigation. KEY WORDS: Time-depth recorder · Vertical movement · Satellite tracking · Tuna · Shark · Turtle Resale or republication not permitted without written consent of the publisher
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Vertical movements of North Sea cod

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol. 347: 101–110, 2007doi: 10.3354/meps07047
Published October 11
INTRODUCTION
The way in which animals move through their envi-ronment forms a fundamental component of their ecol-ogy and, consequently, there have been efforts tounderstand animal movements over a range of spatialand temporal scales (Carey et al. 1971, Block 2005). Ina marine context, vertical movements often play animportant role for a range of animals (Hays 2003, Block2005, Sims et al. 2006). For example, many members ofthe zooplankton undergo regular diel vertical move-ments (generally deeper during the day and shallowerat night), often seemingly as a predator evasion mech-anism (Hays 2003). Similarly, vertical movements arealso important for a range of marine vertebrates. Air-breathing marine vertebrates (Hays 2003) must cometo the surface to breathe and so their vertical move-ments (manifested as dives away from the sea surface)occur regularly in association with foraging (Wilson et
al. 2002), predator evasion (Heithaus & Frid 2003) orresting (Schreer et al. 2001). Over the last 20 yr, elec-tronic data-loggers and transmitters have revealed theintricacies of these vertical movements and how theyvary temporally and spatially. For example, whales,seals, turtles and sharks commonly have diel patternsof resting and foraging (Schreer et al. 2001), with divedepth often varying in association with vertical move-ments of their prey (Sims et al. 2006). Similarly, thesegroups all show different behaviours during migration,where horizontal travel rather than foraging may bethe central objective, and may also show marked spa-tial patterns in diving (Hays et al. 2006).
Vertical movements have also been documented in awide range of fish species. For example, Carey et al.(1971) described the results of deployments of acoustictransmitters that revealed the vertical movements oflarge pelagic fish such as bluefin tuna Thunnus sp.,broadbill swordfish Xiphias gladius and the great
© Inter-Research 2007 · www.int-res.com*Email: [email protected]
Vertical movements of North Sea cod
Victoria J. Hobson1, 2,*, David Righton2, Julian D. Metcalfe2, Graeme C. Hays1
1Department of Biological Sciences, Institute of Environmental Sustainability, Swansea University, Singleton Park,Swansea SA2 8PP, UK
2Centre for Environment, Fisheries and Aquaculture Science, Lowestoft Laboratory, Pakefield Road, Lowestoft,Suffolk NR33 0HT, UK
ABSTRACT: Various air-breathing marine vertebrates such as seals, turtles and seabirds show dis-tinct patterns of diving behaviour. For fish, the distinction between different vertical behaviours isoften less clear-cut, as there are no surface intervals to differentiate between dives. Using data fromacoustic tags (n = 23) and archival depth recorders attached to cod Gadus morhua (n = 92) in thesouthern North Sea, we developed a quantitative method of classifying vertical movements in orderto facilitate an objective comparison of the behaviour of different individuals. This method expandsthe utilisation of data from data storage tags, with the potential for a better understanding of fishbehaviour and enhanced individual based behaviour for improved ecosystem modelling. We foundthat cod were closely associated with the seabed for 90% of the time, although they showed distinctseasonal and spatial patterns in behaviour. For example, cod tagged in the southern North Sea exhib-ited high rates of vertical movement in spring and autumn that were probably associated with migra-tion, while the vertical movements of resident cod in other areas were much less extensive and wereprobably related to foraging or spawning behaviours. The full reasons underlying spatial and tempo-ral behavioural plasticity by cod in the North Sea warrant further investigation.
KEY WORDS: Time-depth recorder · Vertical movement · Satellite tracking · Tuna · Shark · Turtle
Resale or republication not permitted without written consent of the publisher

Mar Ecol Prog Ser 347: 101–110, 2007
white shark Carcharodon carcharias. Metcalfe et al.(1990) and Arnold et al. (1994) used acoustic transmit-ters to describe the vertical movements of plaice Pleu-ronectes platessa and cod Gadus morhua. The prohib-itive expense of acoustic tracking of fish on a largescale has led to the development of relatively inexpen-sive electronic data storage tags (DSTs). While theseneed to be physically returned for data download,DSTs have been deployed in large numbers on a vari-ety of fish species (Hunter et al. 2004, 2006, Block 2005,Righton et al. 2006, Sims et al. 2006), particularly thosethat are commercially exploited, to reveal details oftheir habitat utilisation, confirming the large scalemovements of migratory fish (Block 2005).
As with air-breathing vertebrates, patterns of verti-cal movements in fish have been most often asso-ciated with foraging and migration. For example,Metcalfe & Arnold (1997) described a characteris-tic pattern of selective tidal stream transport of aplaice that migrated between spawning and feedinggrounds in the North Sea. Hunter et al. (2004) laterdetailed the vertical movements of plaice in the south-ern North Sea and revealed complex tidal, diel andlunar rhythms in the extent and timing of verticalmovements. Nichol & Somerton (2002) related the dielvertical movements of Atka mackerel Pleurogrammusmonopterygius to foraging and spawning behaviour.Musyl et al. (2003) have described extensive verticalmigrations of pelagic fish species such as big eye tunaThunnus obesus. These vertical movements havebeen related to foraging on prey within the deep scat-tering layer (Holland et al. 1999); this behaviourappears common in other tuna species, such as south-ern bluefin tuna Thunnus maccoyii (Davis & Stanley2002). In addition, some vertical movements seemassociated with behavioural thermoregulation at thesea surface (Carey et al. 1971).
Cod Gadus morhua are currently a species of par-ticular concern with respect to fishery management,given recent rapid declines in certain stocks, such ascod on the Grand Banks (Myers et al. 1997) or from theNorth Sea (Christensen et al. 2003). Over recent yearsthere has been considerable effort to understand thebehaviour of individual cod, for example in Newfound-land (Rose et al. 1995), Iceland (Palsson & Throsteins-son 2003), Barents Sea (Stensholt 2001) and the Northand Irish Seas (Righton et al. 2001, Neat et al. 2006). Asa step towards achieving this goal, we describe the useof acoustic and DSTs to identify the vertical move-ments of cod tagged at a range of sites in the NorthSea. We set out to assess the dominant patterns ofdepth utilisation with respect to the surface and theseabed, to objectively classify behaviour types and toquantify temporal and spatial changes in behaviour inan efficient and automated manner.
MATERIALS AND METHODS
Acoustic tracking. Tracking experiments using codtagged with acoustic transmitters were conducted in1976. Full details of fish capture, tag attachment andtracking can be found in Arnold et al. (1994). In brief,24 cod were caught on rod and line just off the shoreoff Lowestoft, UK (52.77° N, 1.35° E) (Fig. 1) at a maxi-mum depth of 8 m and brought to the surface slowly toprevent damage to the swimbladder. The cod werethen fitted with 300 kHz transmitting acoustic tags andreturned to the seabed in a holding cage to allow themto readapt to ambient pressure over the next 12 to149 h. On release from the cage, estimates of the depthof the fish and seabed were made by elevation scan-ning using a sector scanning sonar (Greer Walker et al.1978) at intervals of 5 min or less (unless the fish wason the seabed, in which case values were estimatedapproximately every 15 min) for periods of up to 48 h.The estimate of depth was subject to an error of ±3 mat the mean tracking range of 175 m. When cod were inmidwater, we calculated distance from the seabed asthe difference between the measured depth of the tagand the measured depth of the seabed.
Data storage tags. In total, 665 cod in 21 batcheswere tagged with electronic DSTs between March1999 and January 2006. The area of release wasbounded by the coordinates 50° N, 3° W to 60° N, 6° E(Fig. 1). Tagged cod were released in 5 general loca-
102
Fig. 1. Southern North Sea, showing the regions (+) wheretagged cod were released

Hobson et al.: Vertical movements of North Sea cod
tions: off Lowestoft (LWT; 279 DSTs), on a feedingground (the Indefatigable Banks) in the southernNorth Sea (IFB; 49 DSTs), off the northeastern coast ofEngland at Flamborough (FLA; 54 DSTs), on the Jut-land Bank (JUT; 100 DSTs), and in the eastern EnglishChannel (CHA; 183 DSTs).
We fitted several different types of DSTs to coddepending upon the size of the individuals (meanlength of cod = 56 cm, SD = 9 cm) or the attachmentmethod. The makes used were Lotek (LTD1200 [279used] and LTD1400 [105]; Lotek Marine Technolo-gies), Star-Oddi (Milli [27], Centi [223]; Star-OddiMarine Device Manufacturing), and CEFAS Technol-ogy (G5 [28]; Cefas Technology). The DSTs wereattached either externally or internally, as describedin Neat et al. (2006) and Righton et al. (2006). DSTswere programmed to collect pressure and tempera-ture every 10 or 20 min during the period of liberty.Depth was measured with a precision of at least0.02 m depending on the type of DST. Sensor driftwas determined by comparing the surface values atthe start and end of deployments. Drift was low (mean= 0.37 m per 100 d, n = 31 DSTs still recording at thetime of recovery) and so did not afffect our dataanalysis. DSTs were returned to Cefas, Lowestaft Lab-oratory, from the commercial fishery via a rewardscheme. On return to the laboratory, the DSTs weredownloaded and the data archived. Measurementsmade at 20 min intervals were then interpolated to10 min intervals to provide the same temporal sam-pling for each individual. At the time of writing, 130DSTs have been returned (Table 1). We then selectedthe records of more than 1 mo duration from eacharea of release for a more detailed analysis of verticalmovement behaviour. The length of each data recordwas highly variable (mean = 120 d at liberty, SD =110 d at liberty). The number of days available foranalysis was not distributed evenly across the seasons(mean d mo–1 = 1050, SD = 435 d); the number of daysused in analysis showed a binomial distribution, peak-ing in April (1924 d), and December (1536 d), and
declining to a minimum of 542 d in October. Tempera-ture measurements were not used in this study.
Data exclusion. The majority of cod showed a periodof re-acclimatisation to their capture depth immedi-ately after release, a consequence of the re-inflation ofthe swimbladder after its rupture or emptying duringthe capture process (Nichol & Chilton 2006). The dura-tion of this post-tagging behaviour is variable, but typ-ically lasts no longer than 7 d, after which normalbehaviour resumes. This behaviour was not observedin the depth records of cod tagged with acoustic trans-mitters because they were re-acclimatised beforetracking began. It was, however, observed frequentlyin the depth series of cod fitted with DSTs, which werenot re-acclimatised before release. We thereforeexcluded the first 14 d from the start of the depth seriesto avoid including this artefact in our analyses. In addi-tion, we excluded the data from the day of recapture toavoid including behaviour or depth changes related tocapture.
RESULTS
Acoustic tracking
Analysis of the acoustic data showed that cod spent alarge proportion of their time in midwater, with indi-viduals visiting the seabed (Fig. 2) at regular intervals(mean 2.07 h, SD = 3.25 h), with a modal interval of 1 h,and a maximum interval of 20 h. The regular fre-quency of these visits to the seabed suggests thatseabed depth can be inferred using data from DSTs;the relationship of the maximum depth fish trackedacoustically (n = 23) recorded per day and the concur-rent seabed depth recorded was highly significant (r2 =0.99, p < 0.0001). Mean depth of individuals rangedbetween 4.6 and 42.3 m, with a standard deviation of8.78 m. Mean distance from the seabed rangedbetween 1.4 and 31.9 m d–1, with a maximum of47 m. The errors associated with the estimated depth
103
Population Release No. of fish No. of tags No. of tags Days at Longestlocation released returned >30 d liberty record (d)
Flamborough 54.51° N, 0.18° E 54 10 9 1002 138Eastern Channel 50.87° N, 0.74° E 183 40 30 3597 456Indefatigable Banks 53.40° N, 2.11° E 49 11 5 958 449Jutland Bank 56.68° N, 7.30° E 100 6 6 1925 540Lowestoft 52.45° N, 1.83° E 279 63 42 3525 449Total 665 130 92 11007
Table 1. Summary statistics for the number of data storage tags (DSTs) deployed on cod in each area (see Fig. 1). Data fromthe DSTs released in Lowestoft and on the Indefatigable Banks were combined for the analysis, due to their close geographic
locations

Mar Ecol Prog Ser 347: 101–110, 2007
measurements of the fish and the seabed are discussedby Arnold et al. (1994).
DST data
Visual inspection of depth time-series from the DSTdata indicated that cod showed a variety of behavioursthroughout their time at liberty. Large descents to-wards the seabed away from the median depth weredescribed by 4.3% of the days (Fig. 3a). For 5.7% ofthe days, cod maintained a steady mean depth abovethe seabed and made continuous ascents into mid-water or descents towards the seabed (Fig. 3b). On21.1% of the days analysed, depth data showedascents away from the seabed. The position of the fishat a fairly static depth near or on the seabed and therise and fall of the tide could be seen, interspersed withintermittent vertical ascents into the water column ofvarying magnitude to shallower water (Fig. 3c).Finally, for the majority of days analysed (68.9%), thecod appeared to be either on or very close to theseabed, as the depth series showed the continuous rise
104
Fig. 2. Gadus morhua. Example data recorded from a fishusing an acoustic tag, with simultaneous seabed measure-
ment, showing regular seabed visits
Fig. 3. Gadus morhua. Examples of the 4 different general behaviour types identified by eye: (a) midwater (MW), (b) continuoussearch (CS), (c) seabed with ascents (SW) and (d) seabed (SB)
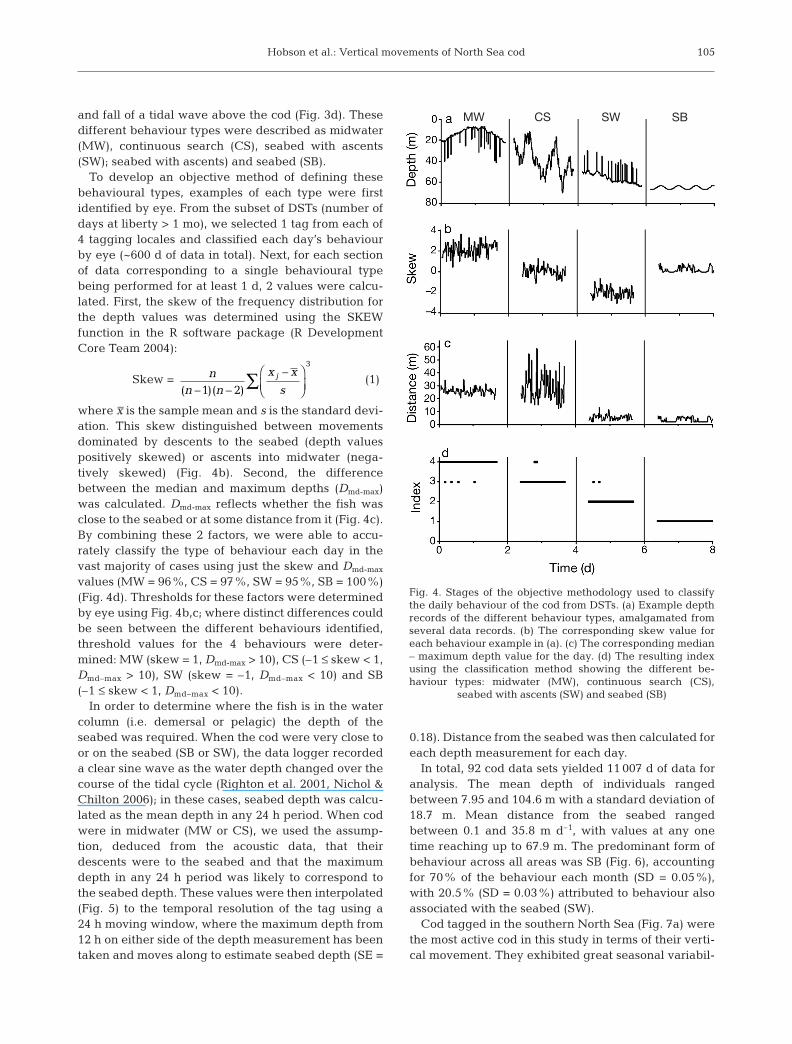
Hobson et al.: Vertical movements of North Sea cod
and fall of a tidal wave above the cod (Fig. 3d). Thesedifferent behaviour types were described as midwater(MW), continuous search (CS), seabed with ascents(SW); seabed with ascents) and seabed (SB).
To develop an objective method of defining thesebehavioural types, examples of each type were firstidentified by eye. From the subset of DSTs (number ofdays at liberty > 1 mo), we selected 1 tag from each of4 tagging locales and classified each day’s behaviourby eye (~600 d of data in total). Next, for each sectionof data corresponding to a single behavioural typebeing performed for at least 1 d, 2 values were calcu-lated. First, the skew of the frequency distribution forthe depth values was determined using the SKEWfunction in the R software package (R DevelopmentCore Team 2004):
Skew = (1)
where x is the sample mean and s is the standard devi-ation. This skew distinguished between movementsdominated by descents to the seabed (depth valuespositively skewed) or ascents into midwater (nega-tively skewed) (Fig. 4b). Second, the differencebetween the median and maximum depths (Dmd-max)was calculated. Dmd-max reflects whether the fish wasclose to the seabed or at some distance from it (Fig. 4c).By combining these 2 factors, we were able to accu-rately classify the type of behaviour each day in thevast majority of cases using just the skew and Dmd-max
values (MW = 96%, CS = 97%, SW = 95%, SB = 100%)(Fig. 4d). Thresholds for these factors were determinedby eye using Fig. 4b,c; where distinct differences couldbe seen between the different behaviours identified,threshold values for the 4 behaviours were deter-mined: MW (skew = 1, Dmd-max > 10), CS (–1 ≤ skew < 1,Dmd–max > 10), SW (skew = –1, Dmd–max < 10) and SB(–1 ≤ skew < 1, Dmd–max < 10).
In order to determine where the fish is in the watercolumn (i.e. demersal or pelagic) the depth of theseabed was required. When the cod were very close toor on the seabed (SB or SW), the data logger recordeda clear sine wave as the water depth changed over thecourse of the tidal cycle (Righton et al. 2001, Nichol &Chilton 2006); in these cases, seabed depth was calcu-lated as the mean depth in any 24 h period. When codwere in midwater (MW or CS), we used the assump-tion, deduced from the acoustic data, that theirdescents were to the seabed and that the maximumdepth in any 24 h period was likely to correspond tothe seabed depth. These values were then interpolated(Fig. 5) to the temporal resolution of the tag using a24 h moving window, where the maximum depth from12 h on either side of the depth measurement has beentaken and moves along to estimate seabed depth (SE =
0.18). Distance from the seabed was then calculated foreach depth measurement for each day.
In total, 92 cod data sets yielded 11 007 d of data foranalysis. The mean depth of individuals rangedbetween 7.95 and 104.6 m with a standard deviation of18.7 m. Mean distance from the seabed rangedbetween 0.1 and 35.8 m d–1, with values at any onetime reaching up to 67.9 m. The predominant form ofbehaviour across all areas was SB (Fig. 6), accountingfor 70% of the behaviour each month (SD = 0.05%),with 20.5% (SD = 0.03%) attributed to behaviour alsoassociated with the seabed (SW).
Cod tagged in the southern North Sea (Fig. 7a) werethe most active cod in this study in terms of their verti-cal movement. They exhibited great seasonal variabil-
nn n
x x
sj
( – )( – )
–
1 2
3⎛⎝⎜
⎞⎠⎟∑
105
Fig. 4. Stages of the objective methodology used to classifythe daily behaviour of the cod from DSTs. (a) Example depthrecords of the different behaviour types, amalgamated fromseveral data records. (b) The corresponding skew value foreach behaviour example in (a). (c) The corresponding median– maximum depth value for the day. (d) The resulting indexusing the classification method showing the different be-haviour types: midwater (MW), continuous search (CS),
seabed with ascents (SW) and seabed (SB)
MW CS SW SB

Mar Ecol Prog Ser 347: 101–110, 2007
ity in vertical movement, with distinct periods whenthey were found high in the water column, either con-tinuously searching or referencing the seabed. These 2behaviours account for more than 25% of their behav-iour in the months from November to March. The restof the year the cod from this region showed a strongassociation with the seabed, and from May to Septem-ber they spent more than 95% of their time very closeto the seabed. Cod tagged off the Danish coast on theJutland Bank (Fig. 7b) showed limited activity
throughout the year. They spent 63.9% of their time atthe seabed, and 35.9% was associated with the seabedwith frequent ascents away from it. Only 0.2% of theJutland fishes’ time could be attributed to midwaterbehaviour.
The behaviour of cod tagged close to Flamborough(Fig. 7c) and in the eastern English Channel (Fig. 7d)showed similar patterns of vertical movement. Analy-sis of the proportion of behaviour types shows that themajority of their time was spent close to the seabed(FLA = 78%, CHA = 80.7%) with an extra 17% (FLA)or 11.8% (CHA) of the time spent on the seabed mak-ing occasional movements into the water column. Thefish spent only 5% (FLA) and 7.5% (CHA) of the timerecorded in midwater. Across all areas, over 55% oftheir time was spent within 5 m of the seabed (Fig. 8).
DISCUSSION
Warming sea temperatures associated with globalclimate change could lead to the loss of cod stocks fromthe North Sea and to the associated socio-economicimplications (Christensen et al. 2003). Knowledge ofthe vertical movements of fish allows an improvedunderstanding of the implications of their vulnerabilityand accessibility to trawls and surveys (Aglen et al.1999, Godo & Michalsen 2000), allowing for improvedstock estimates. In addition, understanding the verticalmovements can enhance our understanding of surveydata of conventional fisheries (Aglen et al. 1999).
A key step in this study was the development of anobjective method for classifying cod behaviour basedon their vertical movements. Objective methods fordefining the vertical movements of marine vertebrateshave been previously documented in studies of seals,turtles and seabirds (Hochscheid & Wilson 1999,Schreer et al. 2001), but to date, the analysis of verticalmovement data for fish has been largely descriptive(e.g. Godo & Michalsen 2000, Righton et al. 2001). Anexception is the recent use of spectral analysis for dis-tinguishing between normal and reverse diel verticalmigration in planktivorous sharks (Graham et al.2006). Various methods for classifying dives for air-breathing vertebrates have been used, often involvingmulti-variant techniques that summarise the duration,depth and shape of dive profiles (Schreer et al. 2001).Each dive type is used to infer a different dive functionor outcome (Schreer et al. 2001), such as prey search-ing, prey capture or sleeping. However, defining thestart and the end of a dive for air-breathing vertebratesis relatively straightforward since each dive starts andends with the animal breathing at the surface. Defin-ing the start of ‘dives’ for fish species is more problem-atic because the movements of many fish have no ob-
106
Fig. 5. Gadus morhua. Depth profile from a fish tagged nearLowestoft in 1999, showing the interpolation of the seabed(solid grey) using the maximum recorded depth of the fish
per day
Fig. 6. Gadus morhua. Proportion of time spent by southernNorth Sea cod (n = 92) in each of the 4 classified behaviours:midwater (MW), continuous search (CS), seabed with ascents(SW) and seabed (SB). Seasonal distribution of behaviour
over the course of a year

Hobson et al.: Vertical movements of North Sea cod
vious start or end points, even though they may bebroadly classed as demersal or pelagic (Godo &Michalsen 2000, Nichol & Chilton 2006). We thereforeused an approach of defining the ‘behavioural state’ ofeach fish on a day-by-day basis rather than attempting
to define when an individual behaviour started orended. Our approach of using simple explanatory sta-tistics (skew and range of median: maximum depth perday) seems robust in that it corroborated the behav-ioural type identified by eye in 97% of occasions.Hence, we have created a methodology that enablesthe dominant behaviour of a fish to be quickly andobjectively defined on a daily basis. Given the hugenumber of data-storage tags currently being employedin various studies with fish (Brill et al. 2002), thisapproach may have wide utility, particularly for dem-ersal fish or those which are known to utilise the wholewater column, such as sharks, where the sea surfacecould be used as a reference point.
Central to our interpretation of DST data was theaxiom that cod regularly visit the seafloor. The devel-opment of this axiom through simultaneous measure-ment of the depth of the fish and seabed depth usingacoustic tracking was therefore central to our work.While direct acoustic tracking from boats is used rela-tively seldom nowadays compared to the use of DSTsand satellite tags, the approach clearly remains impor-tant in being able to reveal detailed aspects of an ani-mal’s immediate environment (e.g. seabed depth) notrevealed by other approaches. Having established that
107
Fig. 7. Gadus morhua. Proportion of time spent by southern North Sea cod in each of the 4 classified behaviours — midwater(MW), continuous search (CS), seabed with ascents (SW) and seabed (SB) — for fish tagged in (a) the southern North Sea,
(b) Jutland Bank, (c) Flamborough, (d) the eastern English Channel
Fig. 8. Gadus morhua. Frequency histogram representing theproportion of time cod fitted with DSTs spent at various
distances from the seabed

Mar Ecol Prog Ser 347: 101–110, 2007
cod regularly visit the seabed, we were then able tomake far more use of the much larger data sets col-lected with DSTs. There may be long-term changes infish behaviour in the North Sea due to fishing pres-sures (Conover & Munch 2002); the behaviour dis-played by fish fitted with DSTs may represent the suc-cessful behavioural characteristics needed to surviveintense fishing efforts. The acoustic data from 1976 isused here to validate vertical movements of cod andconfirm their association with the seabed; however, theextent of behavioural association with the seabed inthe acoustically tagged fish is unknown and may havechanged over time.
The function of the 4 behavioural types we identifiedremains speculative. One of the behavioural states (SB)seems to indicate general inactivity by the fish, withalmost no vertical excursions, although we cannotexclude the possibility that cod were moving horizon-tally across a flat seabed. This depth signal has alsobeen noted in tags attached to plaice (Hunter et al.2004) and thornback rays in the North Sea (Hunter etal. 2006). The other 3 behavioural states (CS, MW, andSW) show greater vertical movement, implying greaterforaging activity. The marked vertical excursions seenin midwater were mostly observed in autumn andspring and were associated with frequent changes inseabed depth (Fig. 5) and occasional sudden changesin temperature. These facts indicate that the individu-als move between different locales during post- or pre-spawning migrations, and the vertical movements mayreflect a foraging or search strategy used by cod asthey transit through unfamiliar locations. Such behav-iour has also been described for some other fish spe-cies. For example, Brill et al. (2002) used acoustictracking of tuna to show that they make use of theentire water column and make frequent (albeit brief)descents to the seabed. Similar dive-like behaviour hasbeen shown in whale sharks (Gunn et al. 1999). Thetemporal resolution of the data collection (once every10 min) means it is likely that fine-scale movementsand behaviours, such as an excursion to capture preyfrom the seabed, are not captured. One of the DSTswas pre-programmed to record at a higher resolution(10 s intervals) for 2 wk during July. This data did notshow any vertical activity, and the fish was associatedwith the seabed for the entire period.
The reasons behind the vertical excursions seen inmidwater and continuous searching behavioural statesare unclear and we can only speculate as to their func-tions. Physicochemical and biological water character-istics often vary much more quickly in the vertical thanin the horizontal dimension. For example, with increas-ing depth, illumination levels and water temperaturegenerally decrease. Similarly, prey and predators maybe concentrated at certain depths. Heithaus et al.
(2002) show tiger sharks undertaking ‘bouncing’ orfast deep dives, followed by a few minutes at depth,then a slow ascent, and suggest that these dives couldbe a foraging tactic to widen opportunities for catchingboth pelagic or surface prey and demersal prey by bal-ancing their time in both areas. Holland et al. (1999)describe a depth record of a tiger shark making large-scale vertical descents when in the open ocean andsuggest that this may be an orientation mechanismthat the shark uses to determine if it is moving into oraway from shallow water. However, Klimley (1993)suggests that hammerhead sharks move up and downthe water column to identify a layer of water with asite-specific olfactant and that the vertical diving oscil-lations were not restricted to thermal boundaries.
Vertical excursions may therefore serve a ‘prospect-ing’ role and allow marine animals to quickly gatherinformation about a wide range of conditions. Forexample, cod have been observed feeding high in thewater column, predominantly at night when pelagicprey are available there (Strand et al. 2005). However,during the day, occasional steep ascents may be per-formed as if cod are prospecting to see if their prey arestill in that area and so might be encountered on thesubsequent night (Godo & Michalsen 2000).
To distinguish between these possibilities, a usefulmeasurement parameter would be the rate of feedingassociated with each behavioural type. Similarly, thereare currently major efforts to measure feeding ratesassociated with dive types for air-breathing verte-brates such as seals (Bowen et al. 2006) and penguins(Wilson et al. 2002) using approaches such as stomachtemperature recorders, video imaging, and mouthopening sensors. Until the technology has been furtherminiaturised to permit the measurement of feedingrate in relatively small vertebrates like cod, our infer-ences about the purpose of vertical movements willremain educated guesses.
Cod of all size classes are found near the seabed;however, despite being regarded as a demersal fish,the species is known to migrate up the water column(Aglen et al. 1999). The activity of cod is also known tovary over seasonal time periods, as seen in studies ofcod in the North and Irish seas (Righton et al. 2001). Akey finding of our study was that, when consideringthe data set as a whole, cod showed a general ten-dency to be closely associated with the seabed andonly 10% of the overall time was spent in midwaterbehavioural states. This finding confirms previouswork with this species in shallow shelf seas (Righton etal. 2001). In contrast to our results, cod living in deeperwater in the Barents Sea and the Icelandic Sea oftenmake large vertical movements to hundreds of metresabove the seabed in order to feed on their main prey,capelin (Godo & Michalsen 2000). In consequence, the
108

Hobson et al.: Vertical movements of North Sea cod
migration patterns and habitat use of cod in the Bar-ents Sea closely match those of capelin (Strand et al.2005). The species can therefore clearly occupy a widerange of depths throughout its range but appears toshape its behaviour to local conditions. For example,despite the fairly shallow water depth throughout theNorth Sea (typically <100 m in the central North Seaand <40 m in the southern North Sea), cod in differentparts of this general area showed markedly differentbehaviours, and these regional differences were alsoaccompanied by seasonal differences. Inherent in theinterpretation of these results is the assumption thatthe bottom topography of the North Sea is relativelysmooth, so the maximum depth in a 24 h period is fairlyconstant. In more topographically variable regions,more detailed information about the seabed may berequired in order to draw conclusions about watercolumn utilisation.
Our primary aim was to develop an objective methodfor classifying cod behaviour from depth utilisationdata. Clearly, now that the technique has been estab-lished, it lends itself to involved statistical analysis ofthe processes driving the spatio-temporal patterns incod behaviour, such as time of day, night versus day,tagging location, month tagged and fish size. However,even at this stage some striking patterns are evident.Those cod tagged in the southern North Sea showedfar more MW behaviour than those in other areas. Theknown spawning grounds of cod tagged in the NorthSea are found to the south (Metcalfe 2006). Conse-quently, this occurrence of MW behaviour fromNovember to March may reflect the migration of thesefish to their spawning grounds from feeding grounds tothe north (Righton et al. 2007) and back again. Thisperiod of migration may be a time when cod are partic-ularly susceptible to mortality in fishing nets becausethey may be away from the shelter of wrecks and otherhard substrates, which provides a sanctuary fromtrawlers. In contrast to the behaviour during theinferred seasonal migration of the fish tagged in thesouthern North Sea, at other times cod showed verylittle activity, spending nearly all their time near or onthe seabed, with only occasional vertical excursions.This relatively inactive behaviour might reflect the factthat meal sizes for cod can be very large (Adlerstein &Welleman 2000) and hence animals may feed onlyoccasionally, being quiescent the rest of the time.Alternatively, this inactivity may relate to the highwater temperatures in the region, where summerwater temperatures can reach around 16°C or higherat the seabed. It is likely that water temperature playsa role in the vertical movements of these fish. Clearly,this linkage between water temperature and codbehaviour is something that needs to be explored fur-ther, particularly given the concern over increasing
water temperatures in the North Sea for the regenera-tion of cod stocks (Neat et al. 2006). Furthermore, incontrast to the preference of cod in the Barents Sea(Floeter & Temming 2003) for sprat Sprattus sprattusand herring Clupea harengus, cod in the North Seahave a much more diverse diet (Adlerstein & Welleman2000), primarily feeding on molluscs, crustaceans,sandeels Ammodytes spp., haddock Melanogrammusaeglefinus, herring Clupea harrengus, and several flat-fish species, which may be associated with the widebehavioural plasticity we noted.
In summary, we have revealed marked differencesin the behaviour of cod in different parts of the NorthSea, which adds further evidence of behavioural plas-ticity in support of previous research (Righton et al.2001, Palsson & Throsteinsson 2003, Neat et al. 2006).These differences probably, at least partly, reflect thelonger seasonal migrations to spawn for fish in certainareas. However, other processes that drive both theregional and seasonal changes in cod behaviourremain enigmatic and warrant further analysis ofbehavioural types along with associated environmen-tal parameters such as water temperature.
Acknowledgements. We are indebted to many colleagueswho contributed to this study and the many skippers and thefishermen who have cooperated with us by helping us to tagcod and also by returning tags. We thank 4 anonymous refer-ees for the constructive criticism of the study. The study wasfunded by EU project CODYSSEY (Q5RT-2002-00813) andDefra (MF0154), and V.J.H. is supported by a NERC CASEstudentship in collaboration with the Centre for Environment,Fisheries and Aquaculture Science, Lowestoft.
LITERATURE CITED
Adlerstein SA, Welleman HC (2000) Diel variation of stomachcontents of North Sea cod (Gadus morhua) during a 24-hfishing survey: an analysis using generalized additivemodels. Can J Fish Aquat Sci 57:2363–2367
Aglen A, Engås A, Huse I, Michalsen K, Stensholt BK (1999)How vertical fish distribution may affect survey results.ICES J Mar Sci 56:345–360
Arnold GP, Greer Walker M, Emerson LS, Holford BH (1994)Movements of cod (Gadus morhua) in relation to the tidalstreams in the southern North Sea. ICES J Mar Sci 51:207–232
Block BA (2005) Physiological ecology in the 21st century:advancements in biologging science. Integr Comp Biol 45:305–320
Bowen WD, Iverson SJ, McMillan JI, Boness DJ (2006) Repro-ductive performance in grey seals: age-related improve-ment and senescence in a capital breeder. J Anim Ecol 75:1340–1351
Brill R, Lutcavage M, Metzger G, Bushnell P, Arendt M, LucyJ, Watson C, Foley D (2002) Horizontal and vertical move-ments of juvenile bluefin tuna (Thunnus thynnus), in rela-tion to oceanographic conditions of the western NorthAtlantic, determined with ultrasonic telemetry. Fish Bull100:155–167
109

Mar Ecol Prog Ser 347: 101–110, 2007
Carey FG, Teal JM, Kanwisher JW, Lawson KD, Beckett JS(1971) Warm-bodied fish. Am Zool 11:137–143
Christensen V, Guenette S, Heymans JJ, Walters CJ, WatsonR, Zeller D, Pauly D (2003) Hundred-year decline of NorthAtlantic predatory fishes. Fish Fish 4:1–24
Conover DO, Munch SB (2002) Sustaining fisheries yieldsover evolutionary time scales. Science 297:94–96
Davis TLO, Stanley CA (2002) Vertical and horizontal move-ments of southern bluefin tuna (Thunnus maccoyii) in theGreat Australian Bight observed with ultrasonic telemetry.Fish Bull 100:448–462
Floeter J, Temming A (2003) Explaining diet composition ofNorth Sea cod (Gadus morhua): prey size preference vs.prey availability. Can J Fish Aquat Sci 60:140–150
Godo OR, Michalsen K (2000) Migratory behaviour of north-east Arctic cod, studied by use of data storage tags. FishRes 48:127–140
Graham RT, Roberts CM, Smart JCR (2006) Diving behaviourof whale sharks in relation to a predictable food pulse. J RSoc Interface 3:109–116
Greer Walker M, Harden Jones FR, Arnold GP (1978) Themovements of plaice (Pleuronectes platessa) tracked inthe open sea. J Cons Int Explor Mer 38:58–86
Gunn JS, Stevens JD, Davis TLO, Norman BM (1999) Obser-vations on the short-term movements and behaviour ofwhale sharks (Rhincodon typus) at Ningaloo Reef, West-ern Australia. Mar Biol 135:553–559
Hays GC (2003) A review of the adaptive significance andecosystem consequences of zooplankton diel verticalmigrations. Hydrobiologia 503:163–170
Hays GC, Hobson VJ, Metcalfe JD, Righton D, Sims DW(2006) Flexible foraging movements of leatherback turtlesacross the north Atlantic Ocean. Ecology 87:2647–2656
Heithaus MR, Frid A (2003) Optimal diving under the risk ofpredation. J Theor Biol 223:79–92
Heithaus MR, Dill LM, Marshall GJ, Buhleier B (2002) Habitatuse and foraging behavior of tiger sharks (Galeocerdocuvier) in a seagrass ecosystem. Mar Biol 140:237–248
Hochscheid S, Wilson RP (1999) A new method for the deter-mination of at-sea activity in sea turtles. Mar Ecol Prog Ser185:293–296
Holland KN, Wetherbee BM, Lowe CG, Meyer CG (1999)Movements of tiger sharks (Galeocerdo cuvier) in coastalHawaiian waters. Mar Biol 134:665–673
Hunter E, Metcalfe JD, O’Brien CM, Arnold GP, Reynolds JD(2004) Vertical activity patterns of free-swimming adultplaice in the southern North Sea. Mar Ecol Prog Ser 279:261–273
Hunter E, Berry F, Buckley AA, Stewart C, Metcalfe JD (2006)Seasonal migration of thornback rays and implications forclosure management. J Appl Ecol 43:710–720
Klimley AP (1993) Highly directional swimming by scallopedhammerhead sharks, Sphyrna lewini, and subsurfaceirradiance, temperature, bathymetry, and geomagneticfield. Mar Biol 117:1–22
Metcalfe JD (2006) Fish population structuring in the NorthSea: understanding processes and mechanisms from
studies of the movements of adults. J Fish Biol 69:48–65Metcalfe JD, Arnold GP (1997) Tracking fish with electronic
tags. Nature 387:665–666Metcalfe JD, Arnold GP, Webb PW (1990) The energetics of
migration by selective tidal stream transport — an analysisfor plaice tracked in the southern North Sea. J Mar BiolAssoc UK 70:149–162
Musyl MK, Brill RW, Boggs CH, Curran DS, Kazama TK, SekiMP (2003) Vertical movements of bigeye tuna (Thunnusobesus) associated with islands, buoys, and seamountsnear the main Hawaiian Islands from archival taggingdata. Fish Oceanogr 12:152–169
Myers RA, Hutchings JA, Barrowman NJ (1997) Why do fishstocks collapse? The example of cod in Atlantic Canada.Ecol Appl 7:91–106
Neat F, Wright P, Zuur A, Gibb I, Gibb F, Tulett D, Righton D,Turner R (2006) Residency and depth movements of acoastal group of Atlantic cod (Gadus morhua). Mar Biol148:643
Nichol DG, Chilton EA (2006) Recuperation and behaviour ofPacific cod after barotrauma. ICES J Mar Sci 63:83–94
Nichol DG, Somerton DA (2002) Diurnal vertical migration ofthe Atka mackerel Pleurogrammus monopterygius asshown by archival tags. Mar Ecol Prog Ser 239:193–207
Palsson OK, Throsteinsson V (2003) Migration patterns, ambi-ent temperature, and growth of Icelandic cod (Gadusmorhua): evidence from storage tag data. Can J FishAquat Sci 60:1409–1423
Righton D, Metcalfe JD, Connolly P (2001) Different behav-iour of North and Irish Sea cod. Nature 411:156
Righton D, Kjesbu OS, Metcalfe JD (2006) A field and experi-mental evaluation of the effect of data storage tags on thegrowth of cod. J Fish Biol 68:385–400
Righton D, Quayle VA, Hetherington S, Burt G (2007) Move-ments and distribution of cod (Gadus morhua) in thesouthern North Sea and English Channel: results fromconventional and electronic tagging experiments. J MarBiol Assoc UK 87:599–613
Rose GA, de Young B, Colbourne E (1995) Cod (Gadusmorhua) migration speeds and transport relative to cur-rents on the north-east Newfoundland Shelf. ICES J MarSci 52:903–913
Schreer JF, Kovacs KM, Hines RJO (2001) Comparative divingpatterns of pinnipeds and seabirds. Ecol Monogr 71:137–162
Sims DW, Wearmouth VJ, Southall EJ, Hill JM and 8 others(2006) Hunt warm, rest cool: bioenergetic strategy under-lying diel vertical migration of a benthic shark. J AnimEcol 75:176–190
Stensholt BK (2001) Cod migration patterns in relation to tem-perature: analysis of storage tag data. ICES J Mar Sci 58:770–793
Strand E, Jorgensen C, Huse G (2005) Modelling buoyancyregulation in fishes with swimbladders: bioenergetics andbehaviour. Ecol Model 185:309–327
Wilson RP, Ropert-Coudert Y, Kato A (2002) Rush and grabstrategies in foraging marine endotherms: the case forhaste in penguins. Anim Behav 63:85–95
110
Editorial responsibility: Rory Wilson (Contributing Editor),Swansea, UK
Submitted: February 21, 2007; Accepted: May 10, 2007Proofs received from author(s): September 17, 2007




















