
Respirable coal dust particles modify cytochrome P4501A1 (CYP1A1) expression in rat alveolar cells
Upload
independentCategory
view
0download
0
Cardiovascular Drugs and Therapy 18 449–459 2004
C© 2005 Springer Science + Business Media, Inc. Manufactured in The Netherlands
BASIC PHARMACOLOGY
N-2-mercaptopropionylglycine, a Scavanger of ReactiveOxygen Species, does not Modify the Early AntiarrhythmicEffect of Ischaemic Preconditioning in Anaesthetised Dogs
Hajnal Agnes, Laszlo Nagy, James R. Parratt,Julius Papp, and Agnes VeghDepartment of Pharmacology and Pharmacotherapy, Universityof Szeged, Albert Szent-Gyorgyi Faculty of Medicine, Dom ter 12,P.O. Box 427, H-6701 Hungary
Summary. Objective: The possible involvement of reac-
tive oxygen species (ROS) in the protective effects of
ischaemic preconditioning (PC) against arrhythmias was
examined in anaesthetised dogs using the ROS scavenger
N-2-mercaptopropionylglycine (MPG).
Methods: PC was induced in 20 chloralose-urethane
anaesthetised dogs by two 5 min occlusions of the left ante-
rior descending (LAD) coronary artery 20 min prior to the
prolonged (25 min) ischaemia/reperfusion (I/R) insult. In
10 of these dogs MPG was infused locally into a small side
branch of the LAD in a dose of 0.15 mg kg−1 min−1, starting
10 min prior to and continuing throughout the entire PC pro-
cedure. In another four dogs subjected to preconditioning
in the absence and then 2h later in the presence of MPG free
radical formation was evaluated by the chemiluminescence
method. Eleven dogs, infused with saline and subjected to a
25 min I/R insult, served as controls. A further 9 dogs, which
were not preconditioned, were given MPG over a period of
60 min prior to occlusion.
Results: Preconditioning markedly reduced the number
of ventricular premature beats (VPBs; 86 ± 34 v. 377 ±78; P < 0.05), the episodes of ventricular tachycardia (VT;
2.0 ± 0.7 v. 13.6 ± 4.5; P < 0.05) and the incidences
of both VT (60% v. 91%) and ventricular fibrillation (0%
v. 82%; P < 0.05) during the prolonged occlusion. Sur-
vival (from the combined ischaemia and reperfusion in-
sult) was significantly increased (40% v. 0%; P < 0.05)
by PC. MPG did not modify the protective effects of PC,
although free radical (mostly superoxide) formation that
occurred following PC was abrogated in the presence of
MPG. Thus, the number of VPBs (111 ± 39), VT episodes
(1.2 ± 0.9) and the incidences of VT (20%) and VF (0%)
during occlusion were similar to the PC dogs. MPG itself
did not significantly modify arrhythmia severity in non-PC
dogs.
Conclusions: We conclude that in our canine model
of ischaemia/reperfusion the generation of ROS does not
play a trigger role in the early PC-induced antiarrhythmic
protection.
Key Words. reactive oxygen species, ventricular ar-
rhythmias, ischaemia, sudden cardiac death, N-2-
mercaptopropionylglycine
Introduction
There is accumulating evidence that oxygen free rad-icals (reactive oxygen species; ROS) are involved inthe delayed cardioprotective effects of ischaemic pre-conditioning [1–4] of exercise [5], an effect secondaryto the opening of mitochondrial KATP channels [6]. In-deed, mediators possibly involved in ischaemic precon-ditioning (bradykinin, α-adrenoceptor agonists, opioidpeptides) also generate free radicals by opening thesechannels [7]. One explanation for this is that the prod-ucts of ROS induce endogenous antioxidant enzymessuch as manganese-superoxide dismutase (MnSOD; re-cently reviewed by [8]).
The situation regarding the role of ROS in the earlyinfarct-limiting effects of ‘classical’ preconditioning, asoriginally defined by Murry et al. [9], remains unclear.For example, whereas some studies [10] have shownthat the infarct-limiting effects of brief periods of coro-nary artery occlusion are attenuated by free radicalscavengers, others [11,12] have failed to demonstrateany significant effect. These differences may be relatedto the dose of the antioxidant used, the route of admin-istration, the method used to evaluate infarct size (es-pecially with the triphenyl tetrazolium chloride (TTC)method; [13]) and the duration and number of the pre-conditioning occlusions [14]. These latters would be par-ticularly important since there is an increasing evidencethat the intensity of the applied preconditioning stim-ulus may determine the signaling pathways that leadto cardioprotection [15,16]. The situation regarding theprofound antiarrhythmic effects of brief precondition-ing occlusions during ischaemia [17–19], and reperfu-sion [20] and the possible role in these effects of ROS, is
Address for correspondence: Prof. Dr. Agnes Vegh, Ph.D., D.Sc.,Department of Pharmacology and Pharmacotherapy, Universityof Szeged, Albert Szent-Gyorgyi Faculty of Medicine, Dom ter12, P.O. Box 427, H-6701 Hungary. Tel.: 36 62 455 673; Fax: 36 63454 565; E-mail: [email protected]
449

450 Agnes et al.
even more confused [12,21]. Thus, the aim of the presentstudy was to examine whether ROS contributes to themarked early antiarrhythmic effect of ischaemic pre-conditioning, induced by two 5 min coronary arteryocclusions. For this purpose we used a highly dif-fusible and low molecular weight sulphydryl contain-ing scavenger N-2-mercaptopropionylglycine (MPG),widely used in previous studies [e.g. 4,10,21]. This hasthe ability to scavenge both intracellular and extracel-lular free radicals; hydroxyl and superoxide radicals[22,23], peroxynitrite [4,21] and hypochlorite [22]. Sincethere is also disagreement over the ability of free radi-cal scavengers to modify ischaemia and reperfusion ar-rhythmias in hearts that have not been preconditioned[24–26] this has also been re-examined.
Material and Methods
Animals and surgical preparation
We used adult, mongrel dogs of either sex, with amean body weight of 26.5 ± 0.9 kg. The origin andmaintenance of these dogs complied with the require-ments of Hungarian law and those of the EuropeanCommission regarding large experimental animals(86/609/ECC). Under light anaesthesia (20 mg kg−1 in-travenous sodium pentobarbitone) the right femoralartery (for the measurement of systolic, mean anddiastolic blood pressure, SABP, MABP, DABP) andvein were catheterised; through this venous cathetera combination of chloralose (60 mg kg−1) and urethane(200 mg kg−1) was given to maintain anaesthesia. Thedogs were ventilated with room air and blood gaseswere monitored and kept within the normal range [19].Temperature was recorded from the rectum or mid-oesophagus and maintained at 37 ± 0.5◦C. Left ventric-ular systolic (LVSP) pressure and end-diastolic pres-sure (LVEDP) and the first derivative of LVSP overtime (LVdP/dt) were measured from a catheter in-serted via the left common carotid artery.
Thoracotomy was performed at the left fourth andfifth intercostal space and the pericardium excised. Theanterior branch of the left coronary artery was pre-pared for occlusion just proximal to the first main di-agonal branch and a side branch of this artery wascatheterised for the intracoronary administration ofsaline or of N-2-mercaptopropionylglycine (MPG). Acomposite electrode [19] was sutured on the surfaceof the potentially ischaemic area, in order to recordthe degree of inhomogeneity of electrical activation.This electrode also contains four unipolar leads bywhich epicardial ST-segment changes is measured.Blood flow was measured on the left circumflex coro-nary artery using an electromagnetic flow probe (Spec-tramed, Hugo Sachs Electronics, Germany). All thedata, together with a limb lead electrocardiogram,were recorded on a Graphtec Thermal Array Recorder(Hugo Sachs) and analysed off-line using the AdvancedCODAS Analysing System and the Windaq Waveform
Browser playback and analysing software (DATAQ In-struments, USA).
Evaluation of ventricular arrhythmias
and area at risk
Ventricular arrhythmias during a 25 min (LAD) coro-nary artery occlusion and following reperfusion wereassessed as outlined in the ‘Lambeth Conventions’ [27]and modified as previously described [19]. During occlu-sion we assessed the total number of ventricular ectopic(premature) beats (VPB’s) as single ectopics and theincidence and number of episodes of ventricular tachy-cardia (VT; defined as a run of four or more VPB’s ata rate faster than the resting sinus rate). We also de-termined the incidence of ventricular fibrillation (VF)during both occlusion and reperfusion. Survival fromthe combined ischaemia-reperfusion insult was definedas those dogs in sinus rhythm 10 min after the end ofreperfusion. Risk area was assessed at the end of eachexperiment by injecting patent blue V dye into the LADat a pressure not greater than the systolic arterial pres-sure in that animal. It was defined as the percentagearea of the left ventricular wall together with the sep-tum served by the occluded artery [28].
Determination of ROS formation during
preconditioning in the presence and in the
absence of MPG
The effect of MPG on free radical production was ex-amined by the lucigenin-enhanced chemiluminescence(CL) assay [29]. Each time two ml blood samples werewithdrawn from the coronary sinus and collected inmicrotubes containing EDTA. The samples were cen-trifuged for 5 min, at 1000 rpm. Plasma aliquots (100 µl)was added to 250 µM lucigenin in DPBS (Dulbeccophosphate-buffered saline) solution (final volume 1 ml).After a 2 min equilibration period in the dark, measure-ments were commenced. CL was measured by a liq-uid scintillation counter (Packard Tri-Carb 2100 Model)with a single photomultiplier tube positioned in out-of-coincidence mode. CL was detected in the presence ofnitro-blue-tetrazolium (NBT; 200 µM). NBT-inhibitedCL was assessed as an index of superoxide genera-tion although NBT, like other superoxide scavengers, isnot entirely specific. Results were expressed in countsmin−1mg−1 protein/100 µl plasma.
In vitro studies for the determination of
superoxide anion generation by canine
polymorphonuclear leucocytes
In four dogs subjected to preconditioning in the ab-sence and then, 2 h later, in the presence of MPGwe examined whether superoxide generated by ca-nine polymorphonuclear leucocytes (PMNs) was af-fected by MPG. Superoxide anion generation was as-sayed by measuring the superoxide-dependent reduc-tion of ferricytochrome-c, as described by Guarnieri andcolleagues [30]. In brief, two ml blood was collected
Preconditioning, Arrhythmias and Reactive Oxygen Species 451
in tubes containing 100 mM EDTA and 100 mM dex-tran and centrifugated at 1000 rpm. PMNs were iso-lated from plasma by Ficoll-Hypaque gradient and sus-pended in DPBS (1.5 × 106 cells/sample/cuvette) andferricytochrome-c (40 µM). The cells were allowed toequilibrate for 2 min at 37◦C, and then stimulated with0.3 µM phorbol myristate acetate (PMA). Superoxideradical production was recorded graphically as the in-crease in absorbance at 550 mm. The effect of increas-ing doses of MPG (1, 5, 10, 20 mM) was expressed as thepercent change in the linear rate of ferricytochrom-c re-duction, using a molar absorption coefficient of 19.1 ×103 M−1 cm−1.
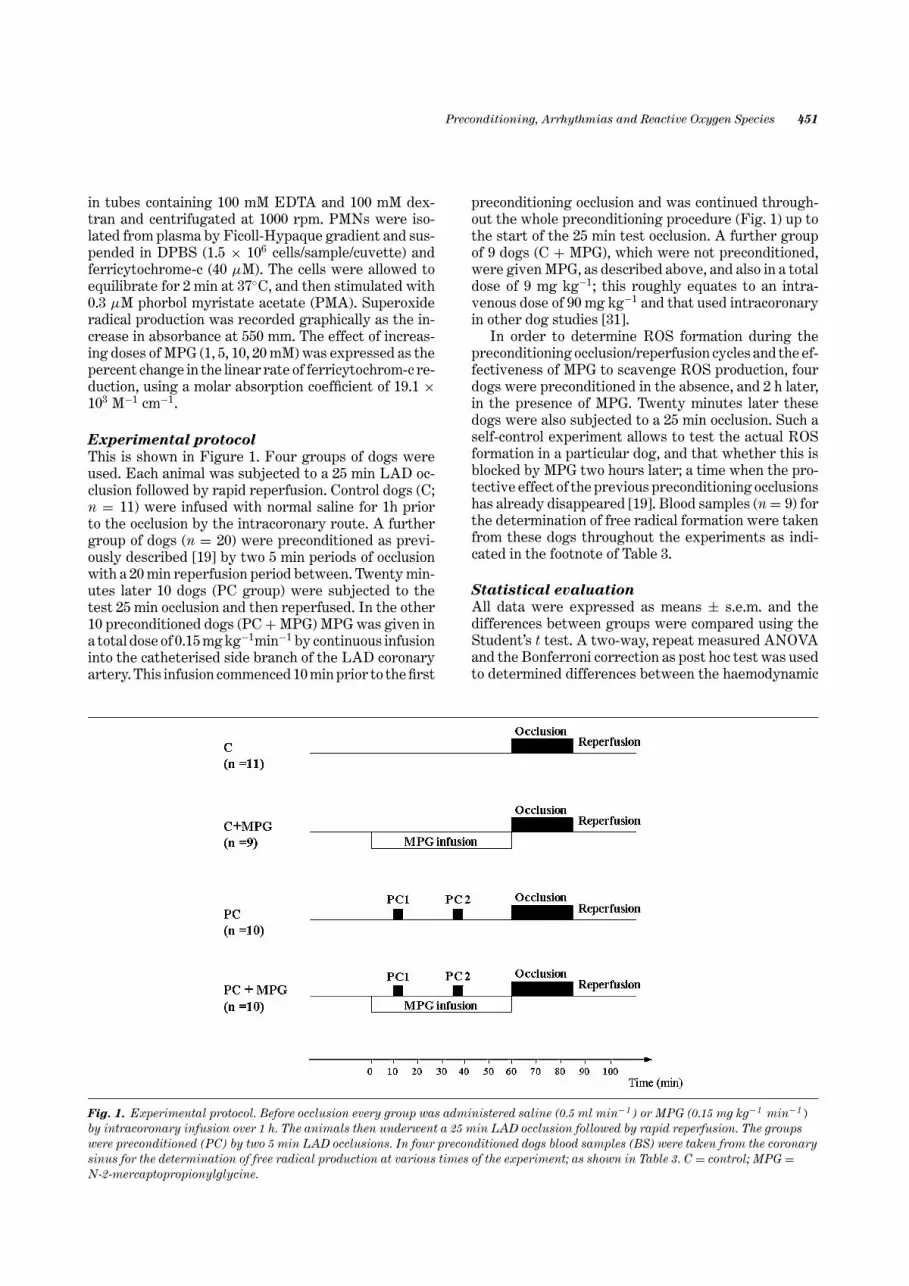
Experimental protocol
This is shown in Figure 1. Four groups of dogs wereused. Each animal was subjected to a 25 min LAD oc-clusion followed by rapid reperfusion. Control dogs (C;n = 11) were infused with normal saline for 1h priorto the occlusion by the intracoronary route. A furthergroup of dogs (n = 20) were preconditioned as previ-ously described [19] by two 5 min periods of occlusionwith a 20 min reperfusion period between. Twenty min-utes later 10 dogs (PC group) were subjected to thetest 25 min occlusion and then reperfused. In the other10 preconditioned dogs (PC + MPG) MPG was given ina total dose of 0.15 mg kg−1min−1 by continuous infusioninto the catheterised side branch of the LAD coronaryartery. This infusion commenced 10 min prior to the first
Fig. 1. Experimental protocol. Before occlusion every group was administered saline (0.5 ml min−1 ) or MPG (0.15 mg kg−1 min−1 )by intracoronary infusion over 1 h. The animals then underwent a 25 min LAD occlusion followed by rapid reperfusion. The groupswere preconditioned (PC) by two 5 min LAD occlusions. In four preconditioned dogs blood samples (BS) were taken from the coronarysinus for the determination of free radical production at various times of the experiment; as shown in Table 3. C = control; MPG =N-2-mercaptopropionylglycine.
preconditioning occlusion and was continued through-out the whole preconditioning procedure (Fig. 1) up tothe start of the 25 min test occlusion. A further groupof 9 dogs (C + MPG), which were not preconditioned,were given MPG, as described above, and also in a totaldose of 9 mg kg−1; this roughly equates to an intra-venous dose of 90 mg kg−1 and that used intracoronaryin other dog studies [31].
In order to determine ROS formation during thepreconditioning occlusion/reperfusion cycles and the ef-fectiveness of MPG to scavenge ROS production, fourdogs were preconditioned in the absence, and 2 h later,in the presence of MPG. Twenty minutes later thesedogs were also subjected to a 25 min occlusion. Such aself-control experiment allows to test the actual ROSformation in a particular dog, and that whether this isblocked by MPG two hours later; a time when the pro-tective effect of the previous preconditioning occlusionshas already disappeared [19]. Blood samples (n = 9) forthe determination of free radical formation were takenfrom these dogs throughout the experiments as indi-cated in the footnote of Table 3.
Statistical evaluation
All data were expressed as means ± s.e.m. and thedifferences between groups were compared using theStudent’s t test. A two-way, repeat measured ANOVAand the Bonferroni correction as post hoc test was usedto determined differences between the haemodynamic

452 Agnes et al.
Table 1. Haemodynamic changes following saline or MPG infusion and during coronary artery occlusion
Max change during Pre occlusion Max changeBaseline saline or MPG infusion value during occlusion
ControlSABP (mmHg) 115 ± 4 −5 ± 0 114 ± 5 −16 ± 4+DABP (mmHg) 80 ± 4 −3 ± 3 77 ± 3 −10 ± 1+MABP(mmHg) 92 ± 4 −3 ± 2 89 ± 4 −12 ± 2+LVEDP (mmHg) 3.5 ± 0.5 0 ± 0 3.8 ± 0.4 9.8 ± 1.1+
+dP/dt (mmHgs−1) 1619 ± 180 −44 ± 44 1610 ± 183 −344 ± 8.3+
−dP/dt (mmHgs−1) 1907 ± 311 169 ± 7 1968 ± 302 −375 ± 5.5+
HR (beats min−1) 160 ± 5 2 ± 0 160 ± 5 3 ± 1C + MPG
SABP (mmHg) 133 ± 4 −17 ± 3∗ 129 ± 6 −15 ± 4+DABP (mmHg) 86 ± 5 −18 ± 3∗ 81 ± 6 −11 ± 2+MABP mmHg) 102 ± 4 −17 ± 3∗ 97 ± 5 −12 ± 3+LVEDP (mmHg) 2.5 ± 0.5 0 ± 0.8 3.5 ± 0.7 8.7 ± 1.1+#
+dP/dt (mmHgs−1) 1747 ± 115 −176 ± 68∗ 1754 ± 134 −261 ± 135−dP/dt (mmHgs−1) 1743 ± 153 −289 ± 70∗ 1580 ± 152 −279 ± 47+
HR (beats min−1) 142 ± 5 2 ± 3 144 ± 7 1 ± 1PC
SABP (mmHg) 121 ± 2 0 ± 2 123 ± 3 −5 ± 1+#
DABP (mmHg) 77 ± 42 1 ± 2 77 ± 5 −9 ± 1MABP (mmHg) 92 ± 42 2 ± 1 94 ± 4 −8 ± 1+LVEDP (mmHg) 3.9 ± 0.5 0.3 ± 0.2 4.1 ± 0.6 5.5 ± 1+#
+dP/dt (mmHgs−1) 1406 ± 115 49 ± 36 1646 ± 144 −294 ± 51+
−dP/dt (mmHgs−1) 1747 ± 148 30 ± 30 1757 ± 186 −150 ± 134HR (beats min−1) 142 ± 6 0 ± 0 138 ± 5 1 ± 1
PC + MPGSABP (mmHg) 130 ± 5 −17 ± 2∗ 124 ± 4 −12 ± 2+DABP (mmHg) 92 ± 6 −16 ± 2∗ 85 ± 5 −13 ± 3+MABP (mmHg) 104 ± 5 −16 ± 2∗ 98 ± 5 −12 ± 3+LVEDP (mmHg) 4.7 ± 0.4 −0.2 ± 0.4 4.2 ± 0.5 5.3 ± 0.6+#
+dP/dt (mmHgs−1) 1478 ± 84 −118 ± 28∗ 1465 ± 92 −306 ± 59+
−dP/dt (mmHgs−1) 1640 ± 173 −151 ± 44 1685 ± 167 −400 ± 88+
HR (beats min−1) 143 ± 5 1 ± 2 144 ± 6 6 ± 3
Values are mean ± s.e.m. Abbreviations are seen in the methods section. ∗ P < 0.05 cp. baseline; + P < 0.05 cp. pre occlusion value; # P < 0.05 cp. controlgroup.
parameters, ST-segment elevation and the degree of in-homogeneity. Ventricular premature beats were com-pared using the Mann-Whitney Rank Sum test and theincidences of VT and VF between the groups usingthe Fisher Exact test. Differences between the groupswere considered significant when P < 0.05.
Results
Haemodynamic effects of MPG and coronary
artery occlusion
These are illustrated in Table 1. Intracoronary infu-sion of MPG resulted in significant reductions in arterialblood pressure and LV dP/dt but these returned to nor-mal by the time of the occlusion. Saline infusion for thesame period of time had no significant haemodynamiceffects.
Occlusion-induced changes were similar in all groupsalthough in preconditioned dogs (PC) these changeswere somewhat less pronounced (Table 1).
Changes in LCX artery coronary blood flow
during LAD artery occlusion
When the LAD coronary artery was occluded there wasan immediate and sustained increase in blood flow inthe adjacent (LCX) coronary artey. This compensatorycoronary blood flow increase was unaffected by MPG(Table 2).
Ventricular arrhythmias during coronary
artery occlusion
The distribution of ventricular premature beats overthe 25 min occlusion period in the four groups is illus-trated in Figure 2. In the control (saline) dogs ectopicactivity appeared in two well differentiated phases (1aand 1b). Phase 1a ectopic activity (but not that duringphase 1b) was suppressed by MPG. In the two precon-ditioned groups there were only a few ectopic beatsduring the 25 min occlusion (86 ± 34 and 111 ± 39in the PC and PC + MPG groups respectively com-pared with 377 ± 78 (P < 0.05) and 244 ± 56 in the

Preconditioning, Arrhythmias and Reactive Oxygen Species 453
Table 2. Changes in LCX artery blood flow (mean; ml · min−1 ) during occlusions of the LAD coronary artery(‘compensatory coronary vasodilatation’)
Preconditioning
Occlusion1 Occlusion2 Prolonged occlusion
Pre Post Pre Post Pre Post
Control (saline) 47.9 ± 5.2 60.2 ± 8.7*Control (MPG) 53.9 ± 2.9 68.7 ± 4.5*Preconditioned (saline) 51.0 ± 6.5 59.6 ± 6.8* 54.4 ± 5.9 62.7 ± 5.8* 53.6 ± 5.9 64.5 ± 7.4*Preconditioned (MPG) 54.4 ± 5.5 59.9 ± 6.5* 51.3 ± 5.0 59.9 ± 6.4* 52.6 ± 5.0 66.1 ± 7.2*
Values as mean ± s.e.m. of 10–11 observations. ∗ P < 0.05 compared with baseline.
Fig. 2. The distribution of ventricular premature beats (VPBs) during 25 min of LAD occlusion. In the control group the two phases ofearly occlusion-induced arrhythmias (1a and 1b) were well marked. Preconditioning reduced arrhythmias throughout the whole25 min occlusion period. MPG did not modify the antiarrhythmic effect of brief preconditioning occlusions although it did itself reducephase Ia arrhythmias. (Abbreviations as in Fig. 1)
relevant controls). Similarly, the incidence and the num-ber of episodes of VT were less in the two precondi-tioned groups (Fig. 3). Thus, VT occurred in 60% of thePC dogs and 20% of the PC + MPG treated dogs (2.0 ±0.7 and 1.2 ± 0.9 episodes of VT) compared to 91% and89% in the C and C + MPG groups (13.6 ± 4.5 and 8.6 ±2.5 VT episodes respectively; all P < 0.05). MPG itselfseemed to reduce VPBs during phase Ia but the totalnumbers over the entire occlusion period was not signif-icantly different from controls (p = 0.165). In the MPGtreated control dogs fatal VF occurred in four dogs outof the nine (44%) during the second half of the occlusionperiod (12 to 25 min) and 3 additional dogs fibrillated onreperfusion (Fig. 3). Survival from this group was 22%.
In the control dogs there was an 82% incidence of VFduring occlusion and this arrhythmia occurred mostlyduring the later phase of the ischaemia. Further, thosedogs in this group that survived the ischaemic periodfibrillated on reperfusion, thus no control dog survivedthe combined ischaemia-reperfusion insult. In contrast,no preconditioned dog fibrillated during the occlusionperiod, whether or not they had been given MPG, butthere was a high incidence of fibrillation on reperfusion.Survival from the combined ischaemia/reperfusion in-sult was 40% in the PC dogs and 30% in the PC +MPG group. The effects on VF during occlusion andreperfusion and survival from the combined ischaemia-reperfusion insult are also shown in Figure 4.

454 Agnes et al.
Fig. 3. The distribution of arrhythmia events (VPBs, VT and VF) in each of the dogs during a 25 min LAD occlusion followed byreperfusion. In the control group, although there was great number of VPBs and VT episodes during the first phase (Ia) of the occlusion,most of the dogs fibrillated during the second half of the occlusion period. In control dogs MPG reduced the incidence and severity ofphase Ia arrhythmias but did not modify the occurrence of VT and VF during phase Ib. Preconditioning reduced the incidence andseverity of these arrhythmia events; an effect not modified by MPG. VF = ventricular fibrillation; VPB = ventricular premature beat;VT = ventricular tachycardia. Other abbreviations as in Figure 1.
Changes in the severity of ischaemia
following coronary artery occlusion
This was examined using both epicardial ST-segmentmapping and the degree of inhomogeneity of electri-cal activation. In control dogs ST-segment elevationwas especially marked during the initial 5 min occlu-sion period and this was maintained throughout the
whole ischaemic period (Fig. 5). MPG given to non-preconditioned dogs did not modify these changes.Preconditioning markedly reduced this index of is-chaemia severity. MPG given to preconditioning dogsdid not modify the reduction in ST-segment eleva-tion that resulted from preconditioning, at least dur-ing the first 5 min of the occlusion. However, later

Preconditioning, Arrhythmias and Reactive Oxygen Species 455
Fig. 4. The incidence of ventricular fibrillation (VF) and survival from the combined 25 min ischaemia-reperfusion insult. In thecontrol group almost all the dogs (82%) died during occlusion; in contrast, no dog died during occlusion in the preconditioned groups;an effect not modified by MPG. ∗P < 0.05 compared with control. (Abbreviations as in Figs. 1 and 3).
Fig. 5. Epicardial ST-segment changes during a 25 min coronary artery occlusion. The maximal elevation had appeared by the first5 min of occlusion and was maintained over the entire ischaemic period. Preconditioning significantly reduced these changes. Inpreconditioned dogs, MPG did not significantly modify ST-segment elevation during the development of ischaemia (first 5 min)although later this did not differ from the controls. Values are means ± s.e.m. ∗ P < 0.05 cp. control. (Abbreviations as in Fig. 1)
in the occlusion (from 8–10 min) on the changes inthe PC dogs given MPG were similar to the controls.Changes in electrical activation are shown in Figure 6;this index of conduction delay within the ischaemicregion was markedly increased during ischaemia andagain, preconditioning reduced this especially early
in ischaemia when suppression of ventricular activ-ity was most pronounced. MPG did not modify thepreconditioning-induced reduction in the degree of in-homogeneity during the first 5 min of occlusion. How-ever, during the later course of ischaemia the degreeof inhomogeneity increased and it was significantly

456 Agnes et al.
Fig. 6. Changes in the degree of inhomogeneity of electrical activation during a 25 min period of LAD occlusion. Preconditioningmarkedly reduced these changes whether or not MPG had been administered. There was only one time point (at 10 min) duringischaemia in the MPG-treated preconditioned dogs, where the degree of inhomogeneity was significantly higher than in the PC dogs notgiven MPG. Values are means ± s.e.m. ∗P < 0.05 cp. control. #P < 0.05 cp. preconditioned dogs. (Abbreviations as in Fig. 1)
higher than in the preconditioned dogs not givenMPG.
Area at risk
There were no significant differences in the area at riskbetween the four groups. These were 32.2 ± 1.3% in thesaline controls, 32.4 ± 1.1% in the controls given MPG,31.4 ± 1.1% in the preconditioned group and 34.2 ± 1.4%in the preconditioned dogs also given MPG.
The effect of MPG on free radical production
by canine granulocytes
MPG concentrations lower than 1 mM did not af-fect ferricytochrome-c reduction. The higher concen-trations of MPG (5, 10 and 20 mM) significantly de-creased ferricytochrome-c reduction by 13, 54 and 96%,respectively.
The effect of MPG on free radical formation
in preconditioned dogs
In four dogs subjected to preconditioning in the pres-ence and in the absence of MPG, free radical formationwas detected by the lucigenin-enhanced chemilumines-cence assay. The results are shown in Table 3. Therewas no detectable free radical formation following thefirst preconditioning occlusion. However, 20 min later,when the second PC occlusion was released there wasa significant, around three-fold, increase in detectableradicals. Two hours later, when the preconditioning pro-cedure was repeated, in the presence of MPG, the occlu-sion/reperfusion cycles did not evoke free radical gen-eration; this was also the case during the prolongedocclusion.
Discussion
In the canine [18,19], as in other species (e.g. rats[12,17,19]), the severity of the ventricular arrhythmiasthat arise when a major coronary artery is occluded ismuch reduced when this is preceeded by one or morebrief (preconditioning) occlusions of that same coronaryartery. There are good reasons for believing that thisis perhaps the most significant cardioprotective effectof ischaemic preconditioning [32,33]. This pronouncedantiarrhythmic effect of ischaemic preconditioning was
Table 3. The effect of MPG on free radical formation in dogssubjected to preconditioning in the presence and in the absenceof MPG
Blood samples � cpm mg−1 protein
BS1 1624 ± 442BS2 1520 ± 258BS3 3758 ± 103∗BS4 1529 ± 88BS5 1326 ± 399BS6 1622 ± 8BS7 1975 ± 495BS8 1068 ± 39BS9 1226 ± 108
100 µl plasma contained 7.714 ± 0.7 mg plasma protein. Abbreviations: BS= blood saples at various time points, such as BS1 = baseline, BS2 and BS3= start of reperfusion of the first and second PC occlusions without MPG,BS4 = baseline value 2 h after the first PC procedure, BS5 = 5 min after thecommencement of MPG infusion, BS6 and BS7 = start of reperfusion of thefirst and second PC occlusions in the presence of MPG infusion, BS8 = endof infusion (pre-occlusion value), BS9 = 20 min of the prolonged occlusion.Values are means ± s.e.m. ∗ P < 0.05 cp.BS1.

Preconditioning, Arrhythmias and Reactive Oxygen Species 457
again clearly demonstrated in the present study; noneof the preconditioned dogs fibrillated during the occlu-sion (VF is a terminal event in this species) comparedto a high incidence (82%) in the controls. The ventricu-lar fibrillation which usually occurs when the ischaemicmyocardium is reperfused at the end of this particularperiod of ischaemia (25 min) was much less influencedby preconditioning; this is again in line with previousexperience in this species [19], and in rats [19,32,33]. Ithas been argued that the less severe arrhythmias dur-ing the ischaemic period following preconditioning isdue in part to the reduction in the severity of ischaemiaas shown by the less pronounced changes in epicardialST-segment elevation [34] and, as in the present study,by less marked changes in the degree of inhomogeneityof electrical activation within the ischaemic area [19].As we have described previously [33], the degree of is-chaemia is only one of several factors that influence ar-rhythmia severity after coronary artery occlusion, andthe precise relationship between ischaemia severity, asrecorded in these ways, and ventricular arrhythmiasduring occlusion is not well defined. Other possibilitiesinclude the size of the ischaemic area, the degree ofcollateral circulation (which is variable in this species),heart rate and the activity of the autonomic nervoussystem [reviewed for example by 35]. Although it isrecognised that the degree of collateral developmentis an important factor in determining the severity ofischaemia-induced arrhythmias in this species [36] pre-vious studies have revealed that the reduction in infarctsize [9] and in arrhythmias [37] following precondition-ing does not depend upon changes in collateral bloodflow. We did not attempt to assess changes in tissueblood flow in all of our studies because the haemody-namic effects of microsphere injection during this vul-nerable, early (25 min) period following the onset ofischaemia might well have modified arrhythmia sever-ity, therefore modifications of the electrocardiographiceffects of coronary artery occlusion were used to as-sess ischaemia severity. Although we are aware thatthe measurement of the epicardial ST-segment and thedegree of inhomogeneity of electrical activation (a mea-sure of conduction delay) cannot be an alternative to themeasurement of tissue blood flow, the rapid changesof these parameters, in particularly during the earlyphase of acute ischaemia, provide an opportunity to es-timate ischaemia severity and its contribution to thegenesis of arrhythmias.
The purpose of the present study was to examinewhether ROS have any part to play in the antiarrhyth-mic effects of brief periods of ischaemia. The dose ofMPG chosen (equivalent to a total intravenous dose of90 mg kg−1) was similar to that used in previous studies[10,12,14] and moreover, was given directly into a sidebranch of the coronary artery that was later occluded.The results show that such locally administered MPGfailed to modify the antiarrhythmic effects of precon-ditioning; the incidences of VF and VT, and the dis-tribution of ventricular ectopic beats were similar in
the preconditioned dogs irrespective of whether or notthe scavenger had been given (Figs. 2 and 3). More-over, MPG given to control dogs (C + MPG) tendedto reduce phase1a arrhythmias (Figs. 2 and 3) a phe-nomenon that deserves further investigation. Never-theless, the total number of VPBs in this group was notsignificantly differed from the saline administered con-trols. Further, the reduction by preconditioning of thetwo measured indices of ischaemia severity, especiallyin the early stages of the occlusion, was not significantlymodified by MPG, at least, during the first 5 to 8 minof ischaemia. We conclude that in this species underin vivo conditions ROS, and in particular superoxide,are not necessary for inducing early protection againstlife-threatening arrhythmias; the protection is presenteven when a highly diffusible and effective scavengerhad been administered. In contrast, this protection isabolished by blockade of bradykinin B2 receptors [38]or by inhibition of nitric oxide synthase [39] or cyclooxy-genase [18].
What are we to make of those studies in species,other than dogs, involving the early (‘classical’, firstwindow) effects of preconditioning in which MPG hadbeen given? The most relevant studies to this discus-sion are those of Richard et al. [12], Tanaka et al. [10]and Baines et al. [14]. Both Tanaka et al. [10] and Baineset al. [14] found, in rabbits, that MPG, in doses lowerthan those used in the present study, given prior toand during a single preconditioning (5 min) occlusion,completely abolished the reduction in infarct size. How-ever, Baines et al. [14] found that MPG (in an evenlarger dose) failed to modify the preconditioning ef-fect of four such 5 min occlusions. They suggested thatother mediators (adenosine, bradykinin) act in concertwith ROS when multiple occlusions are used. Quite whythis should be so is surprising given that one would ex-pect more ROS to be generated by reperfusion aftermany occlusions compared to just one. In rats, Richardet al. [12], who used three preconditioning occlusions,failed to demonstrate any effect of MPG (20 mg kg−1)given intravenously. Further, these authors also mea-sured arrhythmias during and following the 20 min con-trol occlusion period following three 5 min precondition-ing occlusions and found that MPG did not modify thisprotection. They did not distinguish between those ar-rhythmias that developed during the ischaemic periodand those that resulted from reperfusion. Our studiesare in basic agreement with those of Richard et al. [12];treatment with MPG by a route and in a dose one wouldexpect to scavenge ROS failed to modify the protectiveantiarrhythmic effect of ischaemic preconditioning in-duced by two 5 min occlusions. There was also no ev-idence that ROS are involved in arrhythmia severitywhen a coronary artery is occluded without precondi-tioning; the arrhythmias were just as severe after MPGadministration. This also accords with previous expe-rience in this species [26]. These results, of course, donot exclude the opportunity that ROS are involved inthe early cardioprotection when this is induced by other

458 Agnes et al.
stimuli. There is an increasing evidence that the inten-sity of the preconditioning stimulus determines the sig-naling pathway that leads to cardioprotection [15,16].For example, Liam and colleagues [15,16] showed thatin rats preconditioning induced by short (3 min) peri-ods of coronary artery occlusion employs ROS mediatedpathways, whereas the longer (20 min) preconditioningocclusions preferably use adenosine mediated signalingpathways to induce cardioprotection.
In some dogs, where we measured free radical (su-peroxide) formation, we found that following the firstpreconditioning occlusion there was no detectable su-peroxide generation. Whether this means that one5 min occlusion is insufficient to produce superoxideor whether the superoxide produced was effectivelyscavenged by superoxide dismutase, activated by dur-ing this short ischaemia we do not know. There is onlyone free radical that we know is released during theearly phase of ischaemia, and which is involved in boththe early and delayed protection, and that is nitric oxide(NO). This is not scavenged by MPG, although NO inthe presence of superoxide quickly forms peroxinitritewhich can be scavenged by MPG. NO then triggers theprotection possibly involving mito KATP channel open-ing and further free radical formation [40].
We should not, of course, conclude from these studiesthat ROS generated during the preconditioning process(ischaemia and reperfusion) do not contribute to thedelayed effects observed much later. We have not, asyet, examined this possibility in our own models usingcardiac pacing or exercise to induce delayed protectionagainst arrhythmias. However, it should be noted thatmost studies implicating free radicals as triggers for de-layed protection against stunning and infarct size [1–4]have used multiple coronary artery occlusions. Theseare more likely to generate free radicals during thereperfusion process. However, the fact that procedureswhich are unlikely to generate free radicals (e.g. cardiacpacing [28,41]; also result in delayed protection of theheart implies that there are other pathways triggeringdelayed protection.
There is one other effect of coronary artery occlu-sion that was also not modified by MPG. When a coro-nary artery is occluded (e.g. the LAD, as in the presentstudy), there is an immediate increase in blood flow in anadjacent main branch (e.g. the LCX). One explanationfor this phenomenon might be that the compensatoryincrease in flow follows from the increased oxygen de-mands placed on the adjacent essentially normal areaof the left ventricular wall because of the loss of con-tractility in the ischaemic area. Increased oxygen re-quirements then lead to a corresponding, matching in-crease in blood flow. This observation, first shown manyyears ago [42–44] still lacks a convincing explanation,the most likely being the enhanced elaboration of va-sodilator substances by the contracting non-ischaemicmyocardium [45]. Whatever these might be, it is clearthat ROS are not involved.
Acknowledgments
This work was supported by the Hungarian Scientific ResearchFoundation (OTKA; Project number: T037520), the Health Sci-entific Committee of the Hungarian Ministry of Health and bythe National Research and Development Programmes (NKFP;Project number: 1A/0011/2002). Professor Parratt was the recipi-ent of an Albert Szent-Gyorgyi Fellowship of the Hungarian StateGovernment. We are grateful to Dr. Csilla Torday (Department ofExperimental Surgery) for the measurements of the free radicalproduction. We appreciate the technical assistance during theseexperiments of Erika Bako and Maria Gyrffi.
References
1. Sun J-Z, Tang X-L, Park S-W, et al. Evidence for an essen-tial role of reactive oxygen species in the genesis of late pre-conditioning against myocardial stunning in conscious dogs.J Clin Invest 1996;97:562–576.
2. Kaeffer N, Richard V, Thuillez C. Delayed coronary endothe-lial protection 24 hours after preconditioning. Role of freeradicals. Circulation 1997;96:2311–2316.
3. Yamashita N, Hoshida S, Taniguchi N, et al. A “Second Win-dow of Protection” occurs 24 h after ischemic preconditioningin the rat heart. J Mol Cell Cardiol 1998;30:1181–1189.
4. Tang X-L, Takano H, Rizvi A, et al. Oxidant species trig-ger late preconditioning against myocardial stunning in con-scious rabbits. Am J Physiol, 2002;282:H281–H291.
5. Yamashita N, Hoshida S, Otsu K, et al. Exercise pro-vides direct biphasic cardioprotection via manganese su-peroxide dismutase activation. J Exp Med 1999;189:1699–1706.
6. Pain T, Yang X-M, Critz SD, et al. Opening of mitochondrialKATP channels triggers the preconditioned state by gener-ating free radicals. Circ Res 2000;87:460–466.
7. Cohen MV, Yang X-M, Liu GS, et al. Acetylcholine,bradykinin, opioids, and phenylephrine, but not adenosine,trigger preconditioning by generating free radicals andopening mitochondrial KATP channels. Circ Res 2001; 89:273–278.
8. Hoshida S, Yamashita N, Otsu K, Hori M. The importance ofmanganese superoxide dismutase in delayed precondition-ing: Involvement of reactive oxygen species and cytokines.Cardiovasc Res 2002;55:495–505.
9. Murry CE, Jennings RB, Reimer KA. Preconditioning withischemia: A delay of lethal cell injury in ischemic my-ocardium. Circulation 1986;74:1124–1136.
10. Tanaka M, Fujiwara H, Yamasaki K, Sasayama S. Superox-ide dismutase and N-2-mercaptopropionyl glycine attenuateinfarct size limitation effect of ischaemic preconditioning inthe rabbit. Cardiovasc Res 1994;28:980–986.
11. Iwamoto T, Miura T, Adachi T, et al. Myocardial infarct size-limiting effect of ischemic preconditioning was not attenu-ated by oxygen free-radical scavengers in the rabbit. Circu-lation 1991;83:1015–1022.
12. Richard V, Tron C, Thuillez C. Ischaemic preconditioning isnot mediated by oxygen derived free radicals in rats. Car-diovasc Res 1993;27:2016–2021.
13. Shirato C, Miura T, Ooiwa H, et al. Tetrazolium artifac-tually indicates superoxide dismutase-induced salvage inreperfused rabbit heart. J Mol Cell Cardiol 1989;21:1187–1193.

Preconditioning, Arrhythmias and Reactive Oxygen Species 459
14. Baines CP, Goto M., Downey JM. Oxygen radicals re-leased during ischemic preconditioning contribute to cardio-protection in the rabbit myocardium. J Mol Cell Cardiol1997;29:207–216.
15. Liem DA, van den Doel MA, de Zeeuw S, et al. Role of adeno-sine in ischemic preconditioning in rats depends critically onthe duration of the stimulus and involves both A(1) and A(3)receptors. Cardiovasc Res 2001;51:701–708.
16. Liem DA, te Lintel Hekkert M, Mannintveld OC, et al. My-ocardium tolerant to an adenosine-dependent ischemic pre-conditioning stimulus can still be protected by stimuli thatemploy alternative signaling pathways. Am J Physiol HeartCirc Physiol 2004 in press.
17. Komori S, Fukimaki S, Ijili H, et al. Inhibitory effect of is-chaemic preconditioning on ischemic arrhythmias using arat coronary artery ligation model. Jpn J Electrocardiol1990;10:774–782.
18. Vegh A, Szekeres L, Parratt JR. Protective effects of pre-conditioning of the ischaemic myocardium involve cyclo-oxygenase products. Cardiovasc Res 1990;24:1020–1023.
19. Vegh A, Komori S, Szekeres L, Parratt JR. Antiarrhythmiceffects of preconditioning in anaesthetised dogs and rats.Cardiovasc Res 1992;26:487–495.
20. Shiki K, Hearse DJ. Preconditioning of ischemic my-ocardium: Reperfusion-induced arrhythmias. Am J Physiol1987;253:H1470–1476.
21. Altug S, Demiryurek AT, Kane KA, Kanzik I. Evidence forthe involvement of peroxinitrite in ischaemic precondition-ing in rat isolated hearts. Br J Pharmacol 2000;130:125–131.
22. Mitsos SE, Fantone JC, Gallagher KP, et al. Canine myocar-dial reperfusion injury: Protection by a free radical scav-enger, N-2-mercaptopropionyl glicine. J Cardiovasc Phar-macol 1986;8:978–988.
23. Cai M, Ogawa R. Effects of free radical scavengers, methyl-prednisolone, and ulinastatin on acute xanthine and xanthineoxidase-induced lung injury in rats. Circ Shock 1994;43:1–78.
24. Woodward B, Zakaria M. Effect of some free radical scav-angers on reperfusion-induced arrhythmias. J Mol Cell Car-diol 1985;17:485–494.
25. Bernier M, Hearse DJ, Manning S. Reperfusion-induced ar-rhythmias and oxygen derived free radicals: Studies with‘anti-free radical’ interventions and a free radical generatingsystem in the isolated perfused hearts. Circ Res 1986;58:331–340.
26. Wainwright CL, Parratt JR. Failure of allopurinol and a spintrapping agent N-t-butyl-α-phenyl nitrone to modify signif-icantly ischaemia and reperfusion-induced arrhythmias. BrJ Pharmacol 1987;91:49–59.
27. Walker MJA, Curtis MJ, Hearse DJ, et al. The LambethConventions: Guidelines for the study of arrhythmias in is-chaemia, infarction, and reperfusion. Cardiovasc Res 1988;22:447–455.
28. Kaszala K, Vegh A, Papp JG, Parratt JR. Time course of theprotection against ischaemia and reperfusion-induced ven-tricular arrhythmias resulting from brief periods of cardiacpacing. J Mol Cell Cardiol 1996;28:2085–2095.
29. Gyllenhammar H. Lucigenin chemiluminescence in the as-sessement of neutrophil superoxide production. J ImmunMethods 1987;97:209–213.
30. Guarnieri C, Georgountzos A, Caldarera I, et al. Polyaminesstimulate superoxide production in human neutrophils aci-vated by N-Met-Leu-Phe but not by phorbol myristate ac-etate. Biochem Biophys Acta 1987;930:135–139.
31. Bolli R, Jeroudi MO, Patel BS, et al. Marked reduction offree radical generation and contractile dysfunction by an-tioxidant therapy begun at the time of reperfusion. Circ Res1989;65:607–622.
32. Connaughton M, Lawson CS, Hearse DJ. Ischaemic precon-ditioning and arrhythmias induced by ischaemia and reper-fusion. In: Marber MS, Yellon DM, eds. Ischaemia, Precon-ditioning and Adaptation. Oxford: Bios Scientific Publisher,1996;59–84.
33. Parratt JR, Vegh A, Kaszala K, Papp JGY. Suppression oflife-threatening ventricular arrhythmias by brief periods ofischaemia and by cardiac pacing with particular reference todelayed myocardial protection. In: Marber MS, Yellon DM,Eds. Ischaemia, Preconditioning and Adaptation. Oxford:Bios Scientific Publisher, 1996;85–111.
34. Maroko PR, Kjekshus JK, Sobel BE, et al. Factors influ-encing infarct size following experimental coronary arteryocclusions. Circulation 1971;43:67–74.
35. Wit AL, Janse MJ. The Ventricular Arrhythmias of Is-chemia and Infarction. Electrophysiological Mechanisms.Mount Kisco, NY: Futura Publishing Company, 1993.
36. Meesmann W. Early arrhythmias and primary ventricularfibrillation after myocardial ischaemia in relation to preexist-ing coronary collaterals. In: Parratt JR, ed. Early Arrhyth-mias Resulting from Myocardial Ischaemia. New York: Ox-ford University Press, 1982;93–112.
37. Babai L, Szigeti Zs, Parratt JR, Vegh A. Delayed cardio-protective effects of exercise in dogs are aminoguanidinesensitive: Possible involvement of nitric oxide. Clin Sci2002;102:435–445.
38. Vegh A, Papp JGy, Parratt JR. Attenuation of the antiar-rhythmic effects of ischaemic preconditioning by blockadeof bradykinin B2 receptors. Br J Pharmacol 1994;113:1167–1172.
39. Vegh A, Szekeres L, Parratt JR. Preconditioning of the is-chaemic myocardium; involvement of the L-arginine—nitricoxide pathway. Br J Pharmacol 1992;107:648–652.
40. Yellon DM, Downey JM. Preconditioning the myocardium:From cellular physiology to clinical cardiology. Physiol Rev2003;83:113–1151.
41. Vegh A, Szekeres L, Parratt JR. Transient ischaemia in-duced by rapid cardiac pacing results in myocardial precon-ditioning. Cardiovasc Res 1991;25:1051–1053.
42. Herzberg RM, Rubio R, Berne RM. Coronary occlusion andembolisation effect on blood flow in adjacent arteries. Am JPhysiol 1966;210:169–175.
43. Khouri EM, Gregg DE, Lowensohn HS. Flow in the majorbranches of the left coronary artery during experimentalcoronary insufficiency in the unanaesthetised dog. Circ Res1968;23:99–109.
44. Feigl EO. Coronary physiology. Physiol Rev1983;63:1–205.45. Vegh A, Szekeres L, Parratt JR. Inhibition of nitric ox-
ide production in anaesthetised dogs fails to modify work-induced increase in coronary blood flow. J Physiol (Lond.)1992;452:16P.
Copyright © 2022 FDOKUMEN




















