
Anti-inflammatory effects of limb ischaemic preconditioning are mediated by sensory nerve activation...
11
ORIGINAL ARTICLE Anti-inflammatory effects of limb ischaemic preconditioning are mediated by sensory nerve activation in rats Petra Hartmann & Renáta Varga & Zsuzsanna Zobolyák & Júlia Héger & Blanka Csősz & István Németh & Zsolt Rázga & Csaba Vízler & Dénes Garab & Péter Sántha & Gábor Jancsó & Mihály Boros & Andrea Szabó Received: 13 September 2010 / Accepted: 7 December 2010 / Published online: 23 December 2010 # Springer-Verlag 2010 Abstract We have shown that ischaemic preconditioning ameliorates both the local periosteal and the systemic leukocyte activation evoked by limb ischaemia–reperfusion. We hypoth- esized that the activation of chemosensitive afferent nerves by transient ischaemia contributes to the protective mechanisms of ischaemic preconditioning via a calcitonin gene-related peptide (CGRP)-dependent mechanism. In Sprague–Dawley rats, 60- min complete limb ischaemia was followed by 180 min of reperfusion. In further experiments, the CGRP analogue hCGRP (0.3 μg kg -1 ) or ischaemic preconditioning (2×10- min ischaemia/10-min reperfusion) was applied prior to the ischaemia–reperfusion insult. Ischaemic preconditioning was performed in three subgroups in which animals received the CGRP receptor antagonist CGRP 8–37 (30 μg kg -1 h -1 ), the chemosensitive afferent nerve inactivator resiniferatoxin (3× 15 μg kg -1 , sc), or vehicle. The effects of CGRP 8-37 and resiniferatoxin on ischaemia–reperfusion without ischaemic preconditioning were also evaluated. In the tibial periosteum of rats, intravital fluorescence microscopy and immunohisto- chemistry revealed significant attenuations of ischaemia- reperfusion-induced post-ischaemic leukocyte–endothelial interactions (rolling and adherence in the postcapillary venules) and tissue intracellular adhesion molecule expression following ischaemic preconditioning or hCGRP administra- tion. Administration of CGRP 8-37 or pretreatment of animals with resiniferatoxin reversed the anti-inflammatory effects of limb ischaemic preconditioning, but failed to affect the microcirculatory consequences of ischaemia–reperfusion with- out ischaemic preconditioning. The results suggest that activation of the chemo- (capsaicin-) sensitive afferent nerves is involved in the mechanisms of microcirculatory anti- inflammatory protection provided by limb ischaemic precondi- tioning. Controlled activation of chemosensitive C-fibres or the CGRP receptors by the induction of ischaemic preconditioning or other means may furnish therapeutic benefit by ameliorating the periosteal microcirculatory consequences of tourniquet ischaemia. Keywords CGRP . Resiniferatoxin . Preconditioning . Microcirculation . Intravital fluorescence microscopy . ICAM-1 Introduction The benefits of brief periods of ischaemia, generally referred to as ischaemic preconditioning (IPC), against a P. Hartmann : R. Varga : Z. Zobolyák : J. Héger : B. Csősz : D. Garab : M. Boros : A. Szabó (*) Institute of Surgical Research, University of Szeged, Pécsi utca 6, 6720 Szeged, Hungary e-mail: [email protected] R. Varga Department of Oral and Maxillofacial Surgery, University of Szeged, Kálvária sgt. 57, 6725 Szeged, Hungary I. Németh : Z. Rázga Department of Pathology, University of Szeged, Állomás út 2, 6720 Szeged, Hungary C. Vízler Institute of Biochemistry, Biological Research Centre of the Hungarian Academy of Sciences, Dóm tér 9, 6720 Szeged, Hungary P. Sántha : G. Jancsó Department of Physiology, University of Szeged, Dóm tér 10, 6720 Szeged, Hungary Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 DOI 10.1007/s00210-010-0588-4
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Anti-inflammatory effects of limb ischaemic preconditioning are mediated by sensory nerve activation...

ORIGINAL ARTICLE
Anti-inflammatory effects of limb ischaemic preconditioningare mediated by sensory nerve activation in rats
Petra Hartmann & Renáta Varga & Zsuzsanna Zobolyák & Júlia Héger & Blanka Csősz &
István Németh & Zsolt Rázga & Csaba Vízler & Dénes Garab & Péter Sántha &
Gábor Jancsó & Mihály Boros & Andrea Szabó
Received: 13 September 2010 /Accepted: 7 December 2010 /Published online: 23 December 2010# Springer-Verlag 2010
Abstract We have shown that ischaemic preconditioningameliorates both the local periosteal and the systemic leukocyteactivation evoked by limb ischaemia–reperfusion. We hypoth-esized that the activation of chemosensitive afferent nerves bytransient ischaemia contributes to the protective mechanisms ofischaemic preconditioning via a calcitonin gene-related peptide(CGRP)-dependent mechanism. In Sprague–Dawley rats, 60-min complete limb ischaemia was followed by 180 min ofreperfusion. In further experiments, the CGRP analoguehCGRP (0.3 μg kg−1) or ischaemic preconditioning (2×10-min ischaemia/10-min reperfusion) was applied prior to theischaemia–reperfusion insult. Ischaemic preconditioning was
performed in three subgroups in which animals received theCGRP receptor antagonist CGRP8–37 (30 μg kg−1 h−1), thechemosensitive afferent nerve inactivator resiniferatoxin (3×15 μg kg−1, sc), or vehicle. The effects of CGRP8-37 andresiniferatoxin on ischaemia–reperfusion without ischaemicpreconditioning were also evaluated. In the tibial periosteumof rats, intravital fluorescence microscopy and immunohisto-chemistry revealed significant attenuations of ischaemia-reperfusion-induced post-ischaemic leukocyte–endothelialinteractions (rolling and adherence in the postcapillaryvenules) and tissue intracellular adhesion molecule expressionfollowing ischaemic preconditioning or hCGRP administra-tion. Administration of CGRP8-37 or pretreatment of animalswith resiniferatoxin reversed the anti-inflammatory effects oflimb ischaemic preconditioning, but failed to affect themicrocirculatory consequences of ischaemia–reperfusion with-out ischaemic preconditioning. The results suggest thatactivation of the chemo- (capsaicin-) sensitive afferent nervesis involved in the mechanisms of microcirculatory anti-inflammatory protection provided by limb ischaemic precondi-tioning. Controlled activation of chemosensitive C-fibres or theCGRP receptors by the induction of ischaemic preconditioningor other means may furnish therapeutic benefit by amelioratingthe periosteal microcirculatory consequences of tourniquetischaemia.
Keywords CGRP. Resiniferatoxin . Preconditioning .
Microcirculation . Intravital fluorescence microscopy .
ICAM-1
Introduction
The benefits of brief periods of ischaemia, generallyreferred to as ischaemic preconditioning (IPC), against a
P. Hartmann : R. Varga : Z. Zobolyák : J. Héger :B. Csősz :D. Garab :M. Boros :A. Szabó (*)Institute of Surgical Research, University of Szeged,Pécsi utca 6,6720 Szeged, Hungarye-mail: [email protected]
R. VargaDepartment of Oral andMaxillofacial Surgery, University of Szeged,Kálvária sgt. 57,6725 Szeged, Hungary
I. Németh : Z. RázgaDepartment of Pathology, University of Szeged,Állomás út 2,6720 Szeged, Hungary
C. VízlerInstitute of Biochemistry, Biological Research Centreof the Hungarian Academy of Sciences,Dóm tér 9,6720 Szeged, Hungary
P. Sántha :G. JancsóDepartment of Physiology, University of Szeged,Dóm tér 10,6720 Szeged, Hungary
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189DOI 10.1007/s00210-010-0588-4

subsequent ischaemia/reperfusion (IR) injury (Murry et al.1986) have been demonstrated in many organs (Cheung etal. 2006; Gho et al. 1996). It has recently been recognizedthat the protection afforded by IPC involves the action ofseveral endogenous vasodilator mediators including nitricoxide (NO), adenosine and carbon monoxide (for reviews,see Tapuria et al. 2008; Walsch et al. 2007), but the exactmechanisms are still unclear and may involve othersignalling pathways (Yellon and Downey 2003). Oneimportant aspect of IPC is that it is elicited by microcircu-latory reactions with protective consequences in the post-ischaemic vasculature (Koti et al. 2002; Mallick et al. 2005;Starr et al. 2008).
The release of neuropeptides during IPC suggested thatthe activation of sensory and autonomic nerves maycontribute to the protective signalling provided by IPC(Brzozowski et al. 2004; Li and Peng 2002). Calcitoningene-related peptide (CGRP) is one of the most powerfulendogenous vasodilator peptides yet described (Brain et al.1985), and the release of CGRP from capsaicin-sensitiveprimary sensory neurones could be a direct consequence ofhypoxia in hypoperfusion states (Franco-Cereceda 1989).Specifically, the chemosensitive C-fibre afferents express-ing the capsaicin (transient receptor potential vanilloidtype-1 receptor, TRPV1) receptors are activated and/orsensitized by many chemical stimuli released during IR,such as lactate, protons and inflammatory mediators(Caterina et al. 1997; Strecker et al. 2005). Moreover,some of the remote protective effects of IPC might also beattributed to afferent C-fibre-derived mediator release dueto an enhanced NO synthesis (Zhou et al. 2002; Ferdinandyet al. 1997) and the inhibition of tumour necrosis factor-alpha production (Li and Peng 2002).
Although the neurogenic components of IPC are difficultto determine in many organs, the models of limb IPCprovide perhaps the most practical advantages (Hausenloyet al. 2007). Apart from the advantages of the relativesimplicity of the anatomy and accessibility of the majorlimb vessels, the large tissue mass of the limbs producesstrong biochemical signals through the release of consider-able amounts of mediators into well-defined structures andcompartments. It has already been shown that the procedurenot only alleviates the local inflammatory consequences oflimb IR (Szabó et al. 2009), but additionally exertsprotection against remote organ injury (Cheung et al. 2006).
The periosteum is densely innervated by nociceptive C-fibre afferent nerves (Bjurholm et al. 1988; Gajda et al.2005), but their role in IR-induced reactions or IPC-relatedprocesses is as yet unknown. Hence, the main goal of thepresent work was to reveal the possible involvement ofneurogenic factors in the protective effects of limb IPC inthe periosteum, with particular emphasis on the role ofendogenous CGRP in the process. In line with this, the
present study investigated whether CGRP agonist treat-ment, competitive antagonism of the CGRP receptor, orselective depletion of the chemosensitive C-fibres by thecapsaicin analogue resiniferatoxin (RTX) influences IPC-induced microcirculatory reactions in a rat model ofexperimental limb IR. The results demonstrate that theleukocyte-mediated microvascular dysfunctions are signif-icantly reduced by IPC and that this phenomenon iscritically dependent on CGRP in the post-ischaemicperiosteum.
Materials and methods
The experiments were performed in accordance with theNIH Guidelines (Guide for the Care and Use of LaboratoryAnimals), and the study was approved by the AnimalWelfare Committee of the University of Szeged.
Surgical procedure
Male Sprague–Dawley rats (average weight 300±20 g)were anaesthetized with sodium pentobarbital (45 mg kg−1,i.p.) and the trachea cannulated to facilitate respiration. Theright jugular vein and carotid artery were cannulated forfluid and drug administration and for the measurement ofarterial pressure (a Statham P23Db transducer), respective-ly. Further small supplementary doses of pentobarbital(12 mg kg−1) were given intravenously when necessary.The animals were placed in a supine position on a heatingpad to maintain the body temperature between 36°C and37°C, and Ringer’s lactate was infused at a rate of10 ml kg−1 h−1 during the experiments. The left femoralartery was dissected free and the periosteum of the medialsurface of the left tibia was exposed under a Zeiss ×6magnification operating microscope using an atraumaticsurgical technique (Varga et al. 2008). An ultrasonic flowprobe was also placed around the femoral artery for thecontinuous detection of changes in blood flow (TransonicT206, Transonic Systems, Ithaca, NY, USA). Bloodpressure and blood flow signals were recorded with acomputerized data acquisition system (SPEL AdvancedHaemosys 2.72, Experimetria Ltd., Budapest, Hungary).
Experimental protocols
The experiments were performed in two major series wherethe animals were allotted into one or other of the followingexperimental groups. In the first group, the periostealmicrocirculatory responses to 60-min total limb ischaemiafollowed by a 180-min reperfusion period were examined(IR, n=6, Fig. 1). After recording of the baselinemicrocirculatory variables (t=−60 min) with fluorescence
180 Naunyn-Schmied Arch Pharmacol (2011) 383:179–189

intravital videomicroscopy (IVM), complete hindlimbischaemia was induced by placing a tourniquet around theproximal femur, with simultaneous occlusion of the femoralartery with a miniclip. The occlusions were then released(t=0 min) and the periosteal microcirculation observed viaIVM at 60, 120 and 180 min during the reperfusion phase.In further experiments, the CGRP agonist hCGRP(0.3 μg kg−1, i.v., over 5 min, n=6; Shen et al. 2003) orIPC (2×10-min IR) was applied prior to the IR insult. IPCwas performed in three subgroups in which the animalsreceived the CGRP antagonist CGRP8-37 (30 μg kg−1 h−1, i.v., during a 3-h reperfusion; Shen et al. 2003); the capsaicinanalogue RTX, which selectively depletes the chemo-sensitive afferents (15 μg kg−1, s.c., diluted with salinefrom a stock solution dissolved in ethanol; injections wererepeated three times, every second day, ending 14 daysbefore the experiments); or vehicle (saline, n=6–8). Theeffect of the systemic RTX treatment on the sensoryfunctions of the chemosensitive primary sensory neuroneswas confirmed by the abolition of the eye-wipe reaction
induced by the intraconjunctival instillation of 50 μl of0.1% capsaicin solution.
In the second series of experiments, we have investigat-ed the consequences of CGRP8-37 and RTX treatment onthe periosteal inflammatory reactions (n=6–7) after limb IRwithout IPC to determine the direct effects of thecompounds on post-ischaemic microcirculation.
Intravital video microscopy
The left hindlimb with the exposed tibia was positionedhorizontally on an adjustable stage and superfused with37°C saline. The microcirculation of the distal tibia wasvisualized by IVM (Zeiss Axiotech Vario 100HD micro-scope, 100W HBO mercury lamp, Acroplan ×20 waterimmersion objective) using fluorescein isothiocyanate(Sigma Chemicals, St. Louis, MO, USA)-labelled eryth-rocytes (0.2 ml; Ruh et al. 1998) and rhodamine-6Gstaining (Sigma; 0.2%, 0.1 ml, i.v.) for the polymorphonu-clear leukocytes (PMNs). The microscopic images were
Fig. 1 Time sequence of treatments and measurements in series 1 and2. A 60-min total limb ischaemia was followed by 180-minreperfusion (IR ischaemia–reperfusion). In the first series, ischaemicpreconditioning was elicited in two cycles with 10-min ischaemiafollowed by 10-min reperfusion (IPC). In another group of experi-ments, the CGRP receptor agonist hCGRP (0.3 μg kg−1, i.v., over5 min) was applied prior to the IR insult. The capsaicin analoguechemosensitive afferent depletor resiniferatoxin was given in a dose of15 μg kg−1 (s.c.) being repeated three times, every second day, ending14 days before the experiments (RTX). The CGRP receptor antagonist
CGRP8-37 (30 μg kg−1 h−1, i.v.) was infused during the entire 3-h reperfusion period. In the second series of experiments, theconsequences of CGRP8-37 and RTX treatments were investigatedafter limb IR without IPC. Intravital microscopic examinations of theperiosteal microcirculation were performed in the indicated timepoints (IVM) before ischaemia and during reperfusion. At the end ofthe experiments, histology specimen was taken from the periosteumfor immunofluorescence localization of CGRP and TRPV1 andimmunohistochemical determination of ICAM-1, respectively
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 181
recorded with a charge-coupled device video camera (AVTHORN-BC 12) attached to an S-VHS videorecorder(Panasonic AG-MD 830) and a personal computer.
Video analysis
Quantitative assessment of the microcirculatory parameterswas performed off-line by frame-to-frame analysis of thevideotaped images using image analysis software (IVM,Pictron Ltd., Budapest, Hungary). Leukocyte–endothelialcell interactions were analysed within five postcapillaryvenules (diameter between 11 and 20 μm) per animal.Adherent leukocytes (stickers) were defined in each vesselsegment as cells that did not move or detach from theendothelial lining within an observation period of 30 s andare given as the number of cells per square millimetre ofendothelial surface. Rolling leukocytes were defined ascells moving at a velocity <40% that of the erythrocytes inthe centreline of the microvessel passing through theobserved vessel segment within 30 s, and their numberwas given as the number of non-adherent leukocytes persecond per vessel circumference.
Immunohistochemical detection of periosteal ICAM-1expression
For the estimation of endothelial activation and theassessment of the expression changes in an adhesionmolecule playing a role in the adhesion of PMNs to theactivated endothelium, a periosteal tissue specimen wastaken at the end of the experiments. Samples wereprocessed for immunohistochemical detection of tissueintercellular adhesion molecule-1 (ICAM-1) expression,analysed according to a previously described method(Hartmann et al. 2009).
Immunofluorescence detection of the periosteal localizationof TRPV1 and CGRP
TRPV1 receptor and CGRP peptide immunoreactivity wasdemonstrated according to a modified method of Baiou etal. (2007). To detect TRPV1 immunoreactivity, rabbit anti-TRPV1 (Alomone, Jerusalem, Israel; 1:1,000 dilution) andCy3-conjugated donkey anti-rabbit (Jackson, West Grove,USA; 1:500 dilution) antibodies, and to detect CGRPimmunoreactivity, mouse anti-CGRP (Abcam, Cambridge,UK; 1:2,500 dilution) and Daylight 488-conjugated anti-mouse (Jackson; 1:500 dilution) antibodies were used.
Statistical analysis
Data analysis was performed with the SigmaStat statisticalsoftware (Jandel Corporation, San Rafael, CA, USA).
Changes in variables within and between groups wereanalysed by two-way repeated measures ANOVA test,followed by the Holm–Sidak test. All data are expressedas means ± SEM. Values of P<0.05 were consideredstatistically significant.
Results
Macro- and microcirculatory variables
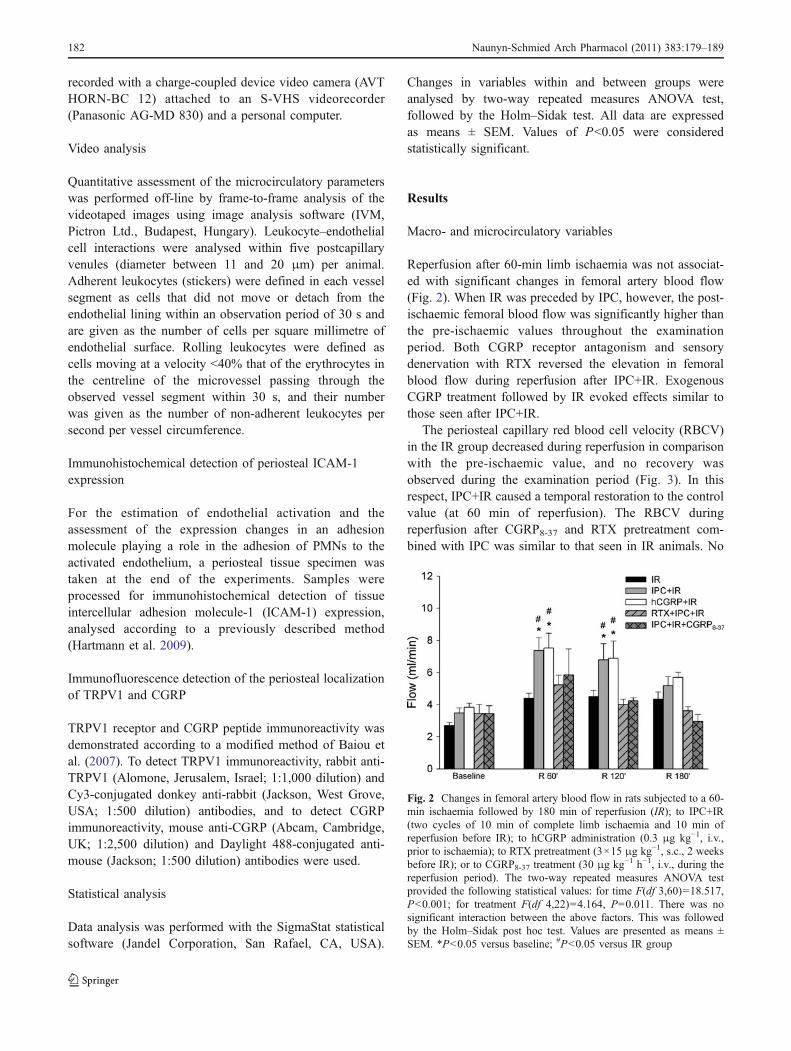
Reperfusion after 60-min limb ischaemia was not associat-ed with significant changes in femoral artery blood flow(Fig. 2). When IR was preceded by IPC, however, the post-ischaemic femoral blood flow was significantly higher thanthe pre-ischaemic values throughout the examinationperiod. Both CGRP receptor antagonism and sensorydenervation with RTX reversed the elevation in femoralblood flow during reperfusion after IPC+IR. ExogenousCGRP treatment followed by IR evoked effects similar tothose seen after IPC+IR.
The periosteal capillary red blood cell velocity (RBCV)in the IR group decreased during reperfusion in comparisonwith the pre-ischaemic value, and no recovery wasobserved during the examination period (Fig. 3). In thisrespect, IPC+IR caused a temporal restoration to the controlvalue (at 60 min of reperfusion). The RBCV duringreperfusion after CGRP8-37 and RTX pretreatment com-bined with IPC was similar to that seen in IR animals. No
Fig. 2 Changes in femoral artery blood flow in rats subjected to a 60-min ischaemia followed by 180 min of reperfusion (IR); to IPC+IR(two cycles of 10 min of complete limb ischaemia and 10 min ofreperfusion before IR); to hCGRP administration (0.3 μg kg−1, i.v.,prior to ischaemia); to RTX pretreatment (3×15 μg kg−1, s.c., 2 weeksbefore IR); or to CGRP8-37 treatment (30 μg kg−1 h−1, i.v., during thereperfusion period). The two-way repeated measures ANOVA testprovided the following statistical values: for time F(df 3,60)=18.517,P<0.001; for treatment F(df 4,22)=4.164, P=0.011. There was nosignificant interaction between the above factors. This was followedby the Holm–Sidak post hoc test. Values are presented as means ±SEM. *P<0.05 versus baseline; #P<0.05 versus IR group
182 Naunyn-Schmied Arch Pharmacol (2011) 383:179–189
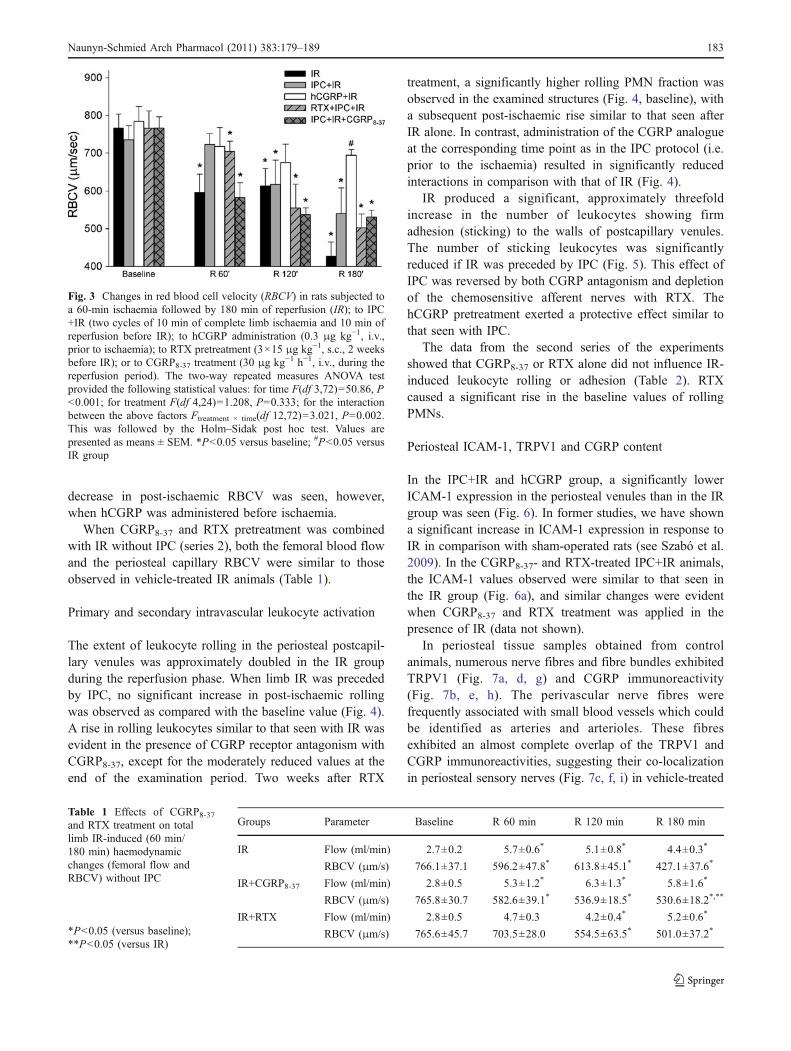
decrease in post-ischaemic RBCV was seen, however,when hCGRP was administered before ischaemia.
When CGRP8-37 and RTX pretreatment was combinedwith IR without IPC (series 2), both the femoral blood flowand the periosteal capillary RBCV were similar to thoseobserved in vehicle-treated IR animals (Table 1).
Primary and secondary intravascular leukocyte activation
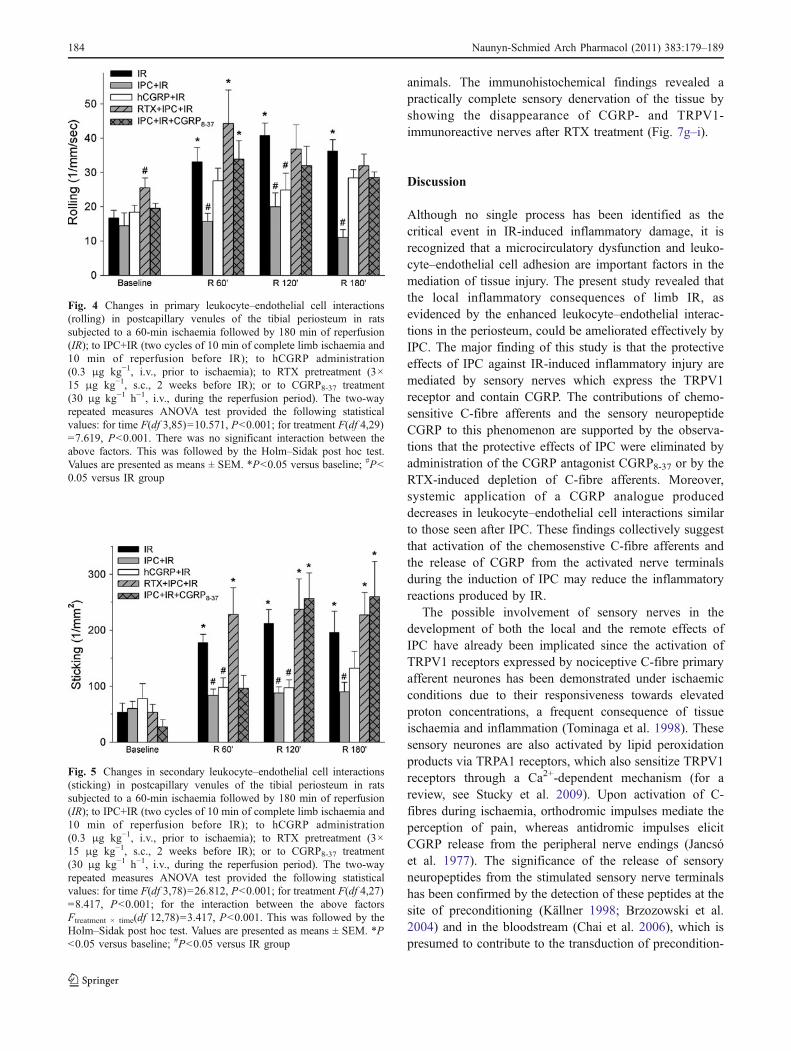
The extent of leukocyte rolling in the periosteal postcapil-lary venules was approximately doubled in the IR groupduring the reperfusion phase. When limb IR was precededby IPC, no significant increase in post-ischaemic rollingwas observed as compared with the baseline value (Fig. 4).A rise in rolling leukocytes similar to that seen with IR wasevident in the presence of CGRP receptor antagonism withCGRP8-37, except for the moderately reduced values at theend of the examination period. Two weeks after RTX
treatment, a significantly higher rolling PMN fraction wasobserved in the examined structures (Fig. 4, baseline), witha subsequent post-ischaemic rise similar to that seen afterIR alone. In contrast, administration of the CGRP analogueat the corresponding time point as in the IPC protocol (i.e.prior to the ischaemia) resulted in significantly reducedinteractions in comparison with that of IR (Fig. 4).
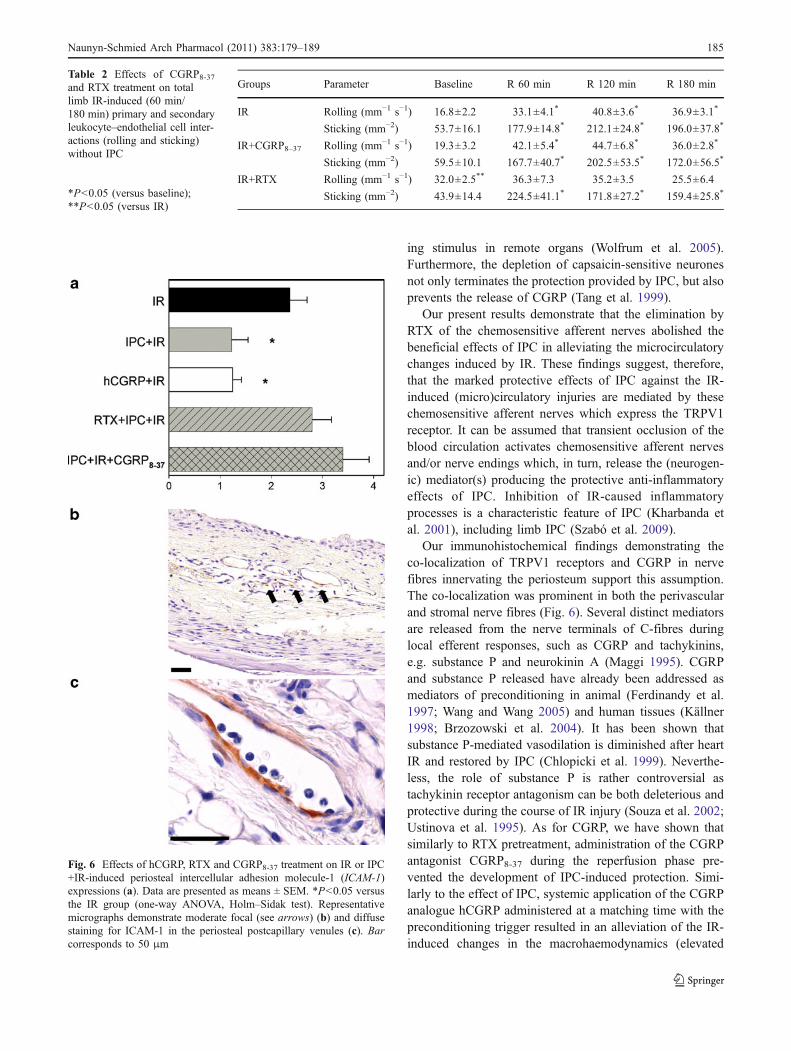
IR produced a significant, approximately threefoldincrease in the number of leukocytes showing firmadhesion (sticking) to the walls of postcapillary venules.The number of sticking leukocytes was significantlyreduced if IR was preceded by IPC (Fig. 5). This effect ofIPC was reversed by both CGRP antagonism and depletionof the chemosensitive afferent nerves with RTX. ThehCGRP pretreatment exerted a protective effect similar tothat seen with IPC.
The data from the second series of the experimentsshowed that CGRP8-37 or RTX alone did not influence IR-induced leukocyte rolling or adhesion (Table 2). RTXcaused a significant rise in the baseline values of rollingPMNs.
Periosteal ICAM-1, TRPV1 and CGRP content
In the IPC+IR and hCGRP group, a significantly lowerICAM-1 expression in the periosteal venules than in the IRgroup was seen (Fig. 6). In former studies, we have showna significant increase in ICAM-1 expression in response toIR in comparison with sham-operated rats (see Szabó et al.2009). In the CGRP8-37- and RTX-treated IPC+IR animals,the ICAM-1 values observed were similar to that seen inthe IR group (Fig. 6a), and similar changes were evidentwhen CGRP8-37 and RTX treatment was applied in thepresence of IR (data not shown).
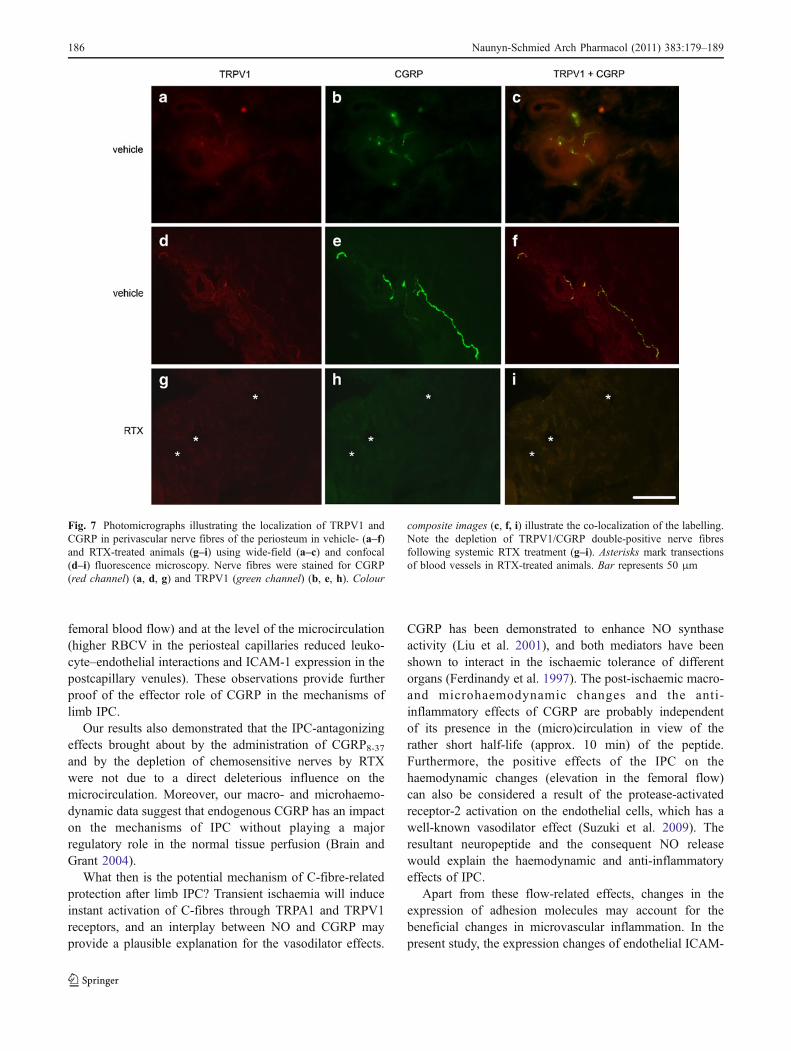
In periosteal tissue samples obtained from controlanimals, numerous nerve fibres and fibre bundles exhibitedTRPV1 (Fig. 7a, d, g) and CGRP immunoreactivity(Fig. 7b, e, h). The perivascular nerve fibres werefrequently associated with small blood vessels which couldbe identified as arteries and arterioles. These fibresexhibited an almost complete overlap of the TRPV1 andCGRP immunoreactivities, suggesting their co-localizationin periosteal sensory nerves (Fig. 7c, f, i) in vehicle-treated
Fig. 3 Changes in red blood cell velocity (RBCV) in rats subjected toa 60-min ischaemia followed by 180 min of reperfusion (IR); to IPC+IR (two cycles of 10 min of complete limb ischaemia and 10 min ofreperfusion before IR); to hCGRP administration (0.3 μg kg−1, i.v.,prior to ischaemia); to RTX pretreatment (3×15 μg kg−1, s.c., 2 weeksbefore IR); or to CGRP8-37 treatment (30 μg kg−1 h−1, i.v., during thereperfusion period). The two-way repeated measures ANOVA testprovided the following statistical values: for time F(df 3,72)=50.86, P<0.001; for treatment F(df 4,24)=1.208, P=0.333; for the interactionbetween the above factors Ftreatment × time(df 12,72)=3.021, P=0.002.This was followed by the Holm–Sidak post hoc test. Values arepresented as means ± SEM. *P<0.05 versus baseline; #P<0.05 versusIR group
Groups Parameter Baseline R 60 min R 120 min R 180 min
IR Flow (ml/min) 2.7±0.2 5.7±0.6* 5.1±0.8* 4.4±0.3*
RBCV (μm/s) 766.1±37.1 596.2±47.8* 613.8±45.1* 427.1±37.6*
IR+CGRP8-37 Flow (ml/min) 2.8±0.5 5.3±1.2* 6.3±1.3* 5.8±1.6*
RBCV (μm/s) 765.8±30.7 582.6±39.1* 536.9±18.5* 530.6±18.2*,**
IR+RTX Flow (ml/min) 2.8±0.5 4.7±0.3 4.2±0.4* 5.2±0.6*
RBCV (μm/s) 765.6±45.7 703.5±28.0 554.5±63.5* 501.0±37.2*
Table 1 Effects of CGRP8-37and RTX treatment on totallimb IR-induced (60 min/180 min) haemodynamicchanges (femoral flow andRBCV) without IPC
*P<0.05 (versus baseline);**P<0.05 (versus IR)
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 183
animals. The immunohistochemical findings revealed apractically complete sensory denervation of the tissue byshowing the disappearance of CGRP- and TRPV1-immunoreactive nerves after RTX treatment (Fig. 7g–i).
Discussion
Although no single process has been identified as thecritical event in IR-induced inflammatory damage, it isrecognized that a microcirculatory dysfunction and leuko-cyte–endothelial cell adhesion are important factors in themediation of tissue injury. The present study revealed thatthe local inflammatory consequences of limb IR, asevidenced by the enhanced leukocyte–endothelial interac-tions in the periosteum, could be ameliorated effectively byIPC. The major finding of this study is that the protectiveeffects of IPC against IR-induced inflammatory injury aremediated by sensory nerves which express the TRPV1receptor and contain CGRP. The contributions of chemo-sensitive C-fibre afferents and the sensory neuropeptideCGRP to this phenomenon are supported by the observa-tions that the protective effects of IPC were eliminated byadministration of the CGRP antagonist CGRP8-37 or by theRTX-induced depletion of C-fibre afferents. Moreover,systemic application of a CGRP analogue produceddecreases in leukocyte–endothelial cell interactions similarto those seen after IPC. These findings collectively suggestthat activation of the chemosenstive C-fibre afferents andthe release of CGRP from the activated nerve terminalsduring the induction of IPC may reduce the inflammatoryreactions produced by IR.
The possible involvement of sensory nerves in thedevelopment of both the local and the remote effects ofIPC have already been implicated since the activation ofTRPV1 receptors expressed by nociceptive C-fibre primaryafferent neurones has been demonstrated under ischaemicconditions due to their responsiveness towards elevatedproton concentrations, a frequent consequence of tissueischaemia and inflammation (Tominaga et al. 1998). Thesesensory neurones are also activated by lipid peroxidationproducts via TRPA1 receptors, which also sensitize TRPV1receptors through a Ca2+-dependent mechanism (for areview, see Stucky et al. 2009). Upon activation of C-fibres during ischaemia, orthodromic impulses mediate theperception of pain, whereas antidromic impulses elicitCGRP release from the peripheral nerve endings (Jancsóet al. 1977). The significance of the release of sensoryneuropeptides from the stimulated sensory nerve terminalshas been confirmed by the detection of these peptides at thesite of preconditioning (Källner 1998; Brzozowski et al.2004) and in the bloodstream (Chai et al. 2006), which ispresumed to contribute to the transduction of precondition-
Fig. 5 Changes in secondary leukocyte–endothelial cell interactions(sticking) in postcapillary venules of the tibial periosteum in ratssubjected to a 60-min ischaemia followed by 180 min of reperfusion(IR); to IPC+IR (two cycles of 10 min of complete limb ischaemia and10 min of reperfusion before IR); to hCGRP administration(0.3 μg kg−1, i.v., prior to ischaemia); to RTX pretreatment (3×15 μg kg−1, s.c., 2 weeks before IR); or to CGRP8-37 treatment(30 μg kg−1 h−1, i.v., during the reperfusion period). The two-wayrepeated measures ANOVA test provided the following statisticalvalues: for time F(df 3,78)=26.812, P<0.001; for treatment F(df 4,27)=8.417, P<0.001; for the interaction between the above factorsFtreatment × time(df 12,78)=3.417, P<0.001. This was followed by theHolm–Sidak post hoc test. Values are presented as means ± SEM. *P<0.05 versus baseline; #P<0.05 versus IR group
Fig. 4 Changes in primary leukocyte–endothelial cell interactions(rolling) in postcapillary venules of the tibial periosteum in ratssubjected to a 60-min ischaemia followed by 180 min of reperfusion(IR); to IPC+IR (two cycles of 10 min of complete limb ischaemia and10 min of reperfusion before IR); to hCGRP administration(0.3 μg kg−1, i.v., prior to ischaemia); to RTX pretreatment (3×15 μg kg−1, s.c., 2 weeks before IR); or to CGRP8-37 treatment(30 μg kg−1 h−1, i.v., during the reperfusion period). The two-wayrepeated measures ANOVA test provided the following statisticalvalues: for time F(df 3,85)=10.571, P<0.001; for treatment F(df 4,29)=7.619, P<0.001. There was no significant interaction between theabove factors. This was followed by the Holm–Sidak post hoc test.Values are presented as means ± SEM. *P<0.05 versus baseline; #P<0.05 versus IR group
184 Naunyn-Schmied Arch Pharmacol (2011) 383:179–189
ing stimulus in remote organs (Wolfrum et al. 2005).Furthermore, the depletion of capsaicin-sensitive neuronesnot only terminates the protection provided by IPC, but alsoprevents the release of CGRP (Tang et al. 1999).
Our present results demonstrate that the elimination byRTX of the chemosensitive afferent nerves abolished thebeneficial effects of IPC in alleviating the microcirculatorychanges induced by IR. These findings suggest, therefore,that the marked protective effects of IPC against the IR-induced (micro)circulatory injuries are mediated by thesechemosensitive afferent nerves which express the TRPV1receptor. It can be assumed that transient occlusion of theblood circulation activates chemosensitive afferent nervesand/or nerve endings which, in turn, release the (neurogen-ic) mediator(s) producing the protective anti-inflammatoryeffects of IPC. Inhibition of IR-caused inflammatoryprocesses is a characteristic feature of IPC (Kharbanda etal. 2001), including limb IPC (Szabó et al. 2009).
Our immunohistochemical findings demonstrating theco-localization of TRPV1 receptors and CGRP in nervefibres innervating the periosteum support this assumption.The co-localization was prominent in both the perivascularand stromal nerve fibres (Fig. 6). Several distinct mediatorsare released from the nerve terminals of C-fibres duringlocal efferent responses, such as CGRP and tachykinins,e.g. substance P and neurokinin A (Maggi 1995). CGRPand substance P released have already been addressed asmediators of preconditioning in animal (Ferdinandy et al.1997; Wang and Wang 2005) and human tissues (Källner1998; Brzozowski et al. 2004). It has been shown thatsubstance P-mediated vasodilation is diminished after heartIR and restored by IPC (Chlopicki et al. 1999). Neverthe-less, the role of substance P is rather controversial astachykinin receptor antagonism can be both deleterious andprotective during the course of IR injury (Souza et al. 2002;Ustinova et al. 1995). As for CGRP, we have shown thatsimilarly to RTX pretreatment, administration of the CGRPantagonist CGRP8-37 during the reperfusion phase pre-vented the development of IPC-induced protection. Simi-larly to the effect of IPC, systemic application of the CGRPanalogue hCGRP administered at a matching time with thepreconditioning trigger resulted in an alleviation of the IR-induced changes in the macrohaemodynamics (elevated
Fig. 6 Effects of hCGRP, RTX and CGRP8-37 treatment on IR or IPC+IR-induced periosteal intercellular adhesion molecule-1 (ICAM-1)expressions (a). Data are presented as means ± SEM. *P<0.05 versusthe IR group (one-way ANOVA, Holm–Sidak test). Representativemicrographs demonstrate moderate focal (see arrows) (b) and diffusestaining for ICAM-1 in the periosteal postcapillary venules (c). Barcorresponds to 50 μm
Groups Parameter Baseline R 60 min R 120 min R 180 min
IR Rolling (mm−1 s−1) 16.8±2.2 33.1±4.1* 40.8±3.6* 36.9±3.1*
Sticking (mm−2) 53.7±16.1 177.9±14.8* 212.1±24.8* 196.0±37.8*
IR+CGRP8–37 Rolling (mm−1 s−1) 19.3±3.2 42.1±5.4* 44.7±6.8* 36.0±2.8*
Sticking (mm−2) 59.5±10.1 167.7±40.7* 202.5±53.5* 172.0±56.5*
IR+RTX Rolling (mm−1 s−1) 32.0±2.5** 36.3±7.3 35.2±3.5 25.5±6.4
Sticking (mm−2) 43.9±14.4 224.5±41.1* 171.8±27.2* 159.4±25.8*
Table 2 Effects of CGRP8-37and RTX treatment on totallimb IR-induced (60 min/180 min) primary and secondaryleukocyte–endothelial cell inter-actions (rolling and sticking)without IPC
*P<0.05 (versus baseline);**P<0.05 (versus IR)
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 185
femoral blood flow) and at the level of the microcirculation(higher RBCV in the periosteal capillaries reduced leuko-cyte–endothelial interactions and ICAM-1 expression in thepostcapillary venules). These observations provide furtherproof of the effector role of CGRP in the mechanisms oflimb IPC.
Our results also demonstrated that the IPC-antagonizingeffects brought about by the administration of CGRP8-37and by the depletion of chemosensitive nerves by RTXwere not due to a direct deleterious influence on themicrocirculation. Moreover, our macro- and microhaemo-dynamic data suggest that endogenous CGRP has an impacton the mechanisms of IPC without playing a majorregulatory role in the normal tissue perfusion (Brain andGrant 2004).
What then is the potential mechanism of C-fibre-relatedprotection after limb IPC? Transient ischaemia will induceinstant activation of C-fibres through TRPA1 and TRPV1receptors, and an interplay between NO and CGRP mayprovide a plausible explanation for the vasodilator effects.
CGRP has been demonstrated to enhance NO synthaseactivity (Liu et al. 2001), and both mediators have beenshown to interact in the ischaemic tolerance of differentorgans (Ferdinandy et al. 1997). The post-ischaemic macro-and microhaemodynamic changes and the anti-inflammatory effects of CGRP are probably independentof its presence in the (micro)circulation in view of therather short half-life (approx. 10 min) of the peptide.Furthermore, the positive effects of the IPC on thehaemodynamic changes (elevation in the femoral flow)can also be considered a result of the protease-activatedreceptor-2 activation on the endothelial cells, which has awell-known vasodilator effect (Suzuki et al. 2009). Theresultant neuropeptide and the consequent NO releasewould explain the haemodynamic and anti-inflammatoryeffects of IPC.
Apart from these flow-related effects, changes in theexpression of adhesion molecules may account for thebeneficial changes in microvascular inflammation. In thepresent study, the expression changes of endothelial ICAM-
Fig. 7 Photomicrographs illustrating the localization of TRPV1 andCGRP in perivascular nerve fibres of the periosteum in vehicle- (a–f)and RTX-treated animals (g–i) using wide-field (a–c) and confocal(d–i) fluorescence microscopy. Nerve fibres were stained for CGRP(red channel) (a, d, g) and TRPV1 (green channel) (b, e, h). Colour
composite images (c, f, i) illustrate the co-localization of the labelling.Note the depletion of TRPV1/CGRP double-positive nerve fibresfollowing systemic RTX treatment (g–i). Asterisks mark transectionsof blood vessels in RTX-treated animals. Bar represents 50 μm
186 Naunyn-Schmied Arch Pharmacol (2011) 383:179–189

1, which is an inducible transmembrane protein of the PMNmigration process, were examined, reflecting endothelialactivation in IR-related events as well (Cowley et al. 1994).To date, no information is available regarding the effects ofsensory nerve activation on adhesion molecule expression,and the reports on related microcirculatory inflammatoryeffects under resting or stimulated circumstances are rathercontroversial. Specifically, endogenous and exogenousCGRP has been shown to be both pro- and anti-inflammatory (Buckley et al. 1991; Jansen et al. 1999;Schneider et al. 2009; Yusof et al. 2007).
The same holds for the adhesion molecule expressions asan increased ICAM-1-dependent PMN adhesion to endo-thelial cells can be demonstrated by CGRP in vitro(Zimmerman et al. 1992) and by capsaicin in vivo inhuman volunteers (Quinlan et al. 1998), suggesting a pro-inflammatory effect on the endothelial cells. On the otherhand, an ameliorating effect of CGRP on the activation ofendothelial cells (Harada et al. 2002), adhesion moleculeCD11b expression (by a cAMP-dependent mechanism;Monneret et al. 2003) and the superoxide production ofPMNs (Tanabe et al. 1996) have been demonstrated invitro. It is conceivable that in the course of the capsaicin-induced neurogenic inflammatory response, the acute andmassive release of neuropeptides produces high local tissueneuropeptide concentrations which most probably exceedthe amount of mediators released by pathophysiologicalstimuli (e.g. IR injury) in vivo. This might provide apossible explanation for the controversial findings men-tioned above. In our study, systemic application ofexogenous hCGRP was highly protective by causing amarked amelioration of the IR-induced PMN responses inthe periosteal microcirculation. This observation supportsother findings demonstrating that exogenous CGRP or theacute release of endogenous CGRP evoked by IPC providespreconditioning-like protection (Chai et al. 2006). Further-more, a decreased tissue ICAM-1 expression also suggesteda protective effect of exogenous CGRP administration onleukocyte–endothelial cell interaction.
Conclusions
This study has demonstrated that chemosensitive afferentnerves which express the TRPV1 receptor and CGRP playdecisive roles in the protective mechanisms exerted by limbIPC. IPC-induced activation of this pathway leads to asignificant amelioration of microcirculatory dysfunctions,including leukocyte–endothelial interactions and endotheli-al activation. These data further suggest that the controlledactivation of chemosensitive afferent nerves and/or theTRPV1 receptor evoked by IPC or by other means, and/oractivation of the peripheral CGRP receptors, may provide
potential therapeutic benefits against IR-induced inflamma-tory injuries.
Acknowledgements This work was supported by research grantsfrom the Hungarian Scientific Research Fund (OTKA K 60752 and K75161).
Conflict of interest The authors state no conflict of interest.
References
Baiou D, Santha P, Avelino A, Charrua A, Bacskai T, Matesz K, CruzF, Nagy I (2007) Neurochemical characterization of insulinreceptor-expressing primary sensory neurons in wild-type andvanilloid type 1 transient receptor potential receptor knockoutmice. J Comp Neurol 503:334–347
Bjurholm A, Kreicbergs A, Brodin E, Schultzberg M (1988)Substance P- and CGRP-immunoreactive nerves in bone.Peptides 9:165–171
Brain SD, Grant AD (2004) Vascular actions of calcitonin gene-relatedpeptide and adrenomedullin. Physiol Rev 84:903–934
Brain SD, Williams TJ, Tippins JR, Morris HR, MacIntyre I (1985)Calcitonin gene-related peptide is a potent vasodilator. Nature313:54–56
Brzozowski T, Konturek PC, Pajdo R, Kwiecień S, Sliwowski Z,Drozdowicz D, Ptak-Belowska A, Pawlik M, Konturek SJ,Pawlik WW, Hahn GG (2004) Importance of brain–gut axis inthe gastroprotection induced by gastric and remote precondition-ing. J Physiol Pharmacol 55:165–177
Buckley TL, Brain SD, Rampart M, Williams TJ (1991) Time-dependent synergistic interactions between the vasodilator neu-ropeptide, calcitonin gene-related peptide (CGRP) and mediatorsof inflammation. Br J Pharmacol 103:1515–1519
Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD,Julius D (1997) The capsaicin receptor: a heat-activated ionchannel in the pain pathway. Nature 389:816–824
Chai W, Mehrotra S, Jan Danser AH, Schoemaker RG (2006) Therole of calcitonin gene-related peptide (CGRP) in ischaemicpreconditioning in isolated rat hearts. Eur J Pharmacol531:246–253
Cheung MM, Kharbanda RK, Konstantinov IE, Shimizu M, FrndovaH, Li J, Holtby HM, Cox PN, Smallhorn JF, Van Arsdell GS,Redington AN (2006) Randomized controlled trial of the effectsof remote ischaemic preconditioning on children undergoingcardiac surgery: first clinical application in humans. J Am CollCardiol 47:2277–2282
Chlopicki S, Lomnicka M, Gryglewski RJ (1999) Reversal of thepostischaemic suppression of coronary function in perfusedguinea pig heart by ischaemic preconditioning. J PhysiolPharmacol 50:605–615
Cowley HC, Heney D, Gearing AJ, Hemingway I, Webster NR (1994)Increased circulating adhesion molecule concentrations inpatients with the systemic inflammatory response syndrome: aprospective cohort study. Crit Care Med 22:651–657
Ferdinandy P, Csont T, Csonka C, Török M, Dux M, Németh J,Horváth LI, Dux L, Szilvássy Z, Jancsó G (1997) Capsaicin-sensitive local sensory innervation is involved in pacing-induced preconditioning in rat hearts: role of nitric oxide andCGRP? Naunyn Schmiedebergs Arch Pharmacol 356:356–363
Franco-Cereceda A (1989) Prostaglandins and CGRP release fromcardiac sensory nerves. Naunyn Schmiedebergs Arch Pharmacol2:180–184
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 187

Gajda M, Litwin JA, Cichocki T, Timmermans JP, Adriaensen D(2005) Development of sensory innervation in rat tibia: co-localization of CGRP and substance P with growth-associatedprotein 43 (GAP-43). J Anat 207:135–144
Gho BC, Schoemaker RG, van den Doel MA, Duncker DJ, VerdouwPD (1996) Myocardial protection by brief ischemia in noncardiactissue. Circulation 94:2193–2200
Harada N, Okajima K, Uchiba M, Katsuragi T (2002) Ischemia/reperfusion-induced increase in the hepatic level of prostacyclinis mainly mediated by activation of capsaicin-sensitive sensoryneurons in rats. J Lab Clin Med 139:218–226
Hartmann P, Szabó A, Eros G, Gurabi D, Horváth G, Németh I,Ghyczy M, Boros M (2009) Anti-inflammatory effects ofphosphatidylcholine in neutrophil leukocyte-dependent acutearthritis in rats. Eur J Pharmacol 622:58–64
Hausenloy DJ, Mwamure PK, Venugopal V, Harris J, Barnard M,Grundy E, Ashley E, Vichare S, Di Salvo C, Kolvekar S,Hayward M, Keogh B, MacAllister RJ, Yellon DM (2007)Effect of remote ischaemic preconditioning on myocardialinjury in patients undergoing coronary artery bypass graftsurgery: a randomised controlled trial. Lancet 370:575–579
Jancsó G, Kiraly E, Jancsó-Gábor A (1977) Pharmacologicallyinduced selective degeneration of chemosensitive primary senso-ry neurones. Nature 270:741–743
Jansen GB, Törkvist L, Löfgren O, Raud J, Lundeberg T (1999)Effects of calcitonin gene-related peptide on tissue survival,blood flow and neutrophil recruitment in experimental skin flaps.Br J Plast Surg 52:299–303
Källner G (1998) Release and effects of calcitonin gene-relatedpeptide in myocardial ischaemia. Scand Cardiovasc J Suppl49:1–35
Kharbanda RK, Peters M, Walton B, Kattenhorn M, Mullen M, KleinN, Vallance P, Deanfield J, MacAllister R (2001) Ischaemicpreconditioning prevents endothelial injury and systemic neutro-phil activation during ischemia–reperfusion in humans in vivo.Circulation 103:1624–1630
Koti RS, Yang W, Dashwood MR, Davidson BR, Seifalian AM (2002)Effect of ischaemic preconditioning on hepatic microcirculationand function in a rat model of ischemia reperfusion injury. LiverTransplant 8:1182–1191
Li YJ, Peng J (2002) The cardioprotection of calcitonin gene-relatedpeptide-mediated preconditioning. Eur J Pharmacol 442:173–177
Liu J, Chen M, Wang X (2001) Calcitonin gene-related peptide-enhanced nitric oxide release and inducible NOS activity andmRNA expression in LPS-stimulated mouse peritoneal macro-phages. Shock 16:64–69
Maggi CA (1995) Tachykinins and calcitonin gene-related peptide(CGRP) as co-transmitters released from peripheral endings ofsensory nerves. Prog Neurobiol 45:1–98
Mallick IH, Yang W, Winslet MC, Seifalian AM (2005) Protectiveeffects of ischaemic preconditioning on the intestinal mucosalmicrocirculation following ischemia–reperfusion of the intestine.Microcirculation 12:615–625
Monneret G, Arpin M, Venet F, Maghni K, Debard AL, Pachot A,Lepape A, Bienvenu J (2003) Calcitonin gene related peptide andN-procalcitonin modulate CD11b upregulation in lipopolysac-charide activated monocytes and neutrophils. Intensive Care Med29:923–928
Murry CE, Jennings RB, Reimer KA (1986) Preconditioning withischemia: a delay of lethal cell injury in ischaemic myocardium.Circulation 74:1124–1136
Quinlan KL, Song IS, Bunnett NW, Letran E, Steinhoff M, Harten B,Olerud JE, Armstrong CA, Wright Caughman S, Ansel JC (1998)Neuropeptide regulation of human dermal microvascular endo-thelial cell ICAM-1 expression and function. Am J Physiol 275:C1580–C1590
Ruh J, Ryschich E, Secchi A, Gebhard MM, Glaser F, Klar E, HerfarthC (1998) Measurement of blood flow in the main arteriole of thevilli in rat small intestine with FITC-labelled erythrocytes.Microvasc Res 56:62–69
Schneider L, Hartwig W, Flemming T, Hackert T, Fortunato F, HeckM, Gebhard MM, Nawroth PP, Bierhaus A, Buchler MW, WernerJ (2009) Protective effects and anti-inflammatory pathways ofexogenous calcitonin gene-related peptide in severe necrotizingpancreatitis. Pancreatology 9:662–669
Shen YT, Mallee JJ, Handt LK, Gilberto DB, Lynch JJ Jr, HargreavesRJ, Koblan KS, Gould RJ, Kane SA (2003) Effects of inhibitionof alpha-CGRP receptors on cardiac and peripheral vasculardynamics in conscious dogs with chronic heart failure. JCardiovasc Pharmacol 42:656–661
Souza DG, Mendonça VA, De A Castro MS, Poole S, Teixeira MM(2002) Role of tachykinin NK receptors on the local and remoteinjuries following ischaemia and reperfusion of the superiormesenteric artery in the rat. Br J Pharmacol 135:303–312
Starr A, Graepel R, Keeble J, Schmidhuber S, Clark N, Grant A, ShahAM, Brain SD (2008) A reactive oxygen species-mediatedcomponent in neurogenic vasodilatation. Cardiovasc Res78:139–147
Strecker T, Messlinger K, Weyand M, Reeh PW (2005) Role ofdifferent proton-sensitive channels in releasing calcitonin gene-related peptide from isolated hearts of mutant mice. CardiovascRes 65:405–410
Stucky CL, Dubin AE, Jeske NA, Malin SA, McKemy DD, Story GM(2009) Roles of transient receptor potential channels in pain.Brain Res Rev 60:2–23
Suzuki H, Motley ED, Eguchi K, Hinoki A, Shirai H, Watts V,Stemmle LN, Fields TA, Eguchi S (2009) Distinct roles ofprotease-activated receptors in signal transduction regulation ofendothelial nitric oxide synthase. Hypertension 53:182–188
Szabó A, Varga R, Keresztes M, Vízler C, Németh I, Rázga Z,Boros M (2009) Ischaemic limb preconditioning downregu-lates systemic inflammatory activation. J Orthop Res 27:897–902
Tanabe T, Otani H, Zeng XT, Mishima K, Ogawa R, Inagaki C (1996)Inhibitory effects of calcitonin gene-related peptide on substance-P-induced superoxide production in human neutrophils. Eur JPharmacol 314:175–183
Tang ZL, Dai W, Li YJ, Deng HW (1999) Involvement of capsaicin-sensitive sensory nerves in early and delayed cardioprotectioninduced by a brief ischaemia of the small intestine. NaunynSchmiedebergs Arch Pharmacol 359:243–247
Tapuria N, Kumar Y, Habib MM, Abu Amara M, Seifalian AM,Davidson BR (2008) Remote ischaemic preconditioning: a novelprotective method from ischemia reperfusion injury—a review. JSurg Res 150:304–330
Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H,Skinner K et al (1998) The cloned capsaicin receptor integratesmultiple pain-producing stimuli. Neuron 21:531–543
Ustinova EE, Bergren D, Schultz HD (1995) Neuropeptide depletionimpairs postischemic recovery of the isolated rat heart: role ofsubstance P. Cardiovasc Res 30:55–63
Varga R, Török L, Szabó A, Kovács F, Keresztes M, Varga G et al(2008) Effects of colloid solutions on ischemia–reperfusion-induced periosteal microcirculatory and inflammatory reactions:comparison of dextran, gelatin, and hydroxyethyl starch. CritCare Med 36:2828–2837
Walsch SR, Tang T, Sadat U, Dutka DP, Gaunt ME (2007)Cardioprotection by remote ischaemic preconditioning. Br JAnaesth 99:611–616
Wang L, Wang DH (2005) TRPV1 gene knockout impairs postische-mic recovery in isolated perfused heart in mice. Circulation112:3617–3623
188 Naunyn-Schmied Arch Pharmacol (2011) 383:179–189

Wolfrum S, Nienstedt J, Heidbreder M, Schneider K, Dominiak P,Dendorfer A (2005) Calcitonin gene related peptide mediatescardioprotection by remote preconditioning. Regul Pept127:217–224
Yellon DM, Downey JM (2003) Preconditioning the myocardium:from cellular physiology to clinical cardiology. Physiol Rev83:1113–1151
Yusof M, Kamada K, Gaskin FS, Korthuis RJ (2007) Angiotensin IImediates postischaemic leukocyte–endothelial interactions: role
of calcitonin gene-related peptide. Am J Physiol 292:H3032–H3037
Zhou ZH, Peng J, Ye F, Li NS, Deng HW, Li YJ (2002) Delayedcardioprotection induced by nitroglycerin is mediated by alpha-calcitonin gene-related peptide. Naunyn Schmiedebergs ArchPharmacol 365:253–259
Zimmerman BJ, Anderson DC, Granger DN (1992) Neuropeptidespromote neutrophil adherence to endothelial cell monolayers. AmJ Physiol 263:G678–G682
Naunyn-Schmied Arch Pharmacol (2011) 383:179–189 189




















