
Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and...
19
Mul t il ocus sequence ana l ys i s r e v ea l s t he gene t i c d i v e r s i t y o f Eur opean frui tt r ee phyt op l asmas and suppor t s t he e xi s t ence o f i n t e r -spec i es r ecomb i na t i on Jean Luc Dane t , 1 Gul nar a Ba l ak i shiy e va, 2 Agne ` sC ime rman, 1 N i col as Sauvi on, 3 Ve ´ ronique Mari e-Jeanne , 3 Ge ´ r ard Labonne , 3 Amparo Lavi n ˇ a, 4 Assumpci o Ba t ll e , 4 Ivana K riz ˇ ana c, 5 Dij ana S ˇ kori c ´ , 6 Paol oE rma cor a, 7 C ¸ i gde m Ul ubas ¸ Se r c ¸ e , 8 Kadriy eC ¸ ag ˘ l ayan, 8 Wol f gang Jar ausch 9 and Xavi e r Foi ssa c 1 Correspondence Xavi er Foissac f oissac @bordeaux.inra.fr Re c eived 7 July 2010 Revised 8 Sept ember 2010 A cc ept ed 10 Sept ember 2010 1 UMR Ge ´ nomique Diversi t e ´ Pouvoir Pat hoge ` ne, I NRA, Universi t e ´ Vi ct or Se ´ gal en Bordeaux 2, 71 Avenue Edouard Bourl aux, BP81, F-33883 Vill enaved’ Ornon Cedex, F ranc e 2 Inst i t ut eof Bot any, Nat i onal A c ademy of Sci enc es of Azerbaij an, Pat amdar shosse 40, AZ-1073, Baku, Azerbaij an 3 UMR B i ol ogi ee t Ge ´ ne ´ t ique des Int era ct i ons Pl ant e-Parasi t e, CIRAD, I NRA, SUPAGRO, Campus Int ernat i onal de Baill argue t, TA A-54/K, F-34398 Mont pe lli er Cedex 5, F ranc e 4 Inst i t ut de Re c erc a i Te cnol ogi a Agroaliment ari es (IRTA), C tra CabrilsKm 2, E-08348 Cabrils, Barc e l ona, Spai n 5 Inst i t ut e f or Pl ant Prot e ct i on, Croat i an Centr e f or Agri cul t ur e, Food and Rural A ff airs, Sve t os ˇ imunska 25, HR-10000 Zagr eb, Croat i a 6 Department of B i ol ogy, Universi ty of Zagr eb, Maruli c ev trg 9a, HR-10000 Zagr eb, Croat i a 7 Di part iment odi B i ol ogi aeProt ezi one de ll ePi ant e, Universi t a ` degli St udi di Udi ne, vi a De ll eSci enze 208, I-33100 Udi ne, It aly 8 Department of Pl ant Prot e ct i on, Must a f a Kemal Universi ty, TK-31034 Ant akya-Hat ay, Turkey 9 Al Pl ant a– Inst i t ut e f or Pl ant Research, RLP AgroSci enc eGmbH, Br e i t enweg 71, D-67435 Neust adt an der We i nstrasse, Germany The gene t i cdiversi ty of t hr ee t emperat e frui ttr ee phyt opl asmas ‘ Candi dat us Phyt opl asma prunorum’ , ‘ Ca. P. mali ’ and ‘ Ca. P. pyri ’ has been est abli shed by mul t il ocus sequence analysi s. Among t he f our gene t i c l oc i used, t he genes i mp and aceF di st i ngui shed 30 and 24 genotypes, r espec t ive ly, and showed t he hi ghest vari abili ty. Per cent age of subst i t ut i on f or i mp ranged from 50 t o 68 % accordi ng t o spec i es. Per cent age of subst i t ut i on vari ed be t ween 9 and 12 % f or aceF , wher eas i t was be t ween 5 and 6 % f or pnp and secY. In t he case of ‘ Ca P. prunorum’ t he t hr ee most pr eval ent aceF genotypes wer e de t ec t ed i n bot hpl ant s and i nsec t vec t ors, conf irmi ng t hat t he pr eval ent i sol at es ar epropagat ed by i nsec t s. The f our i sol at es known t o be hypo-virul ent had t he same aceF sequence , i ndi cat i ng a possi bl e monophyl e t i cori gi n. Hapl otype ne t work r econstruc t ed by eBURST r eveal ed t hat among t he 34 hapl otypes of ‘ Ca. P. prunorum’ , t he f our hypo-virul ent i sol at es al so grouped t oge t her i n t he samec l ade . Genotypi ng of some Spani sh and Azerbaij anese ‘ Ca. P. pyri ’ i sol at es showed t hatt hey shar ed someall e l es wi t h ‘ Ca. P. prunorum’ , support i ng f or t he f irstt ime t o our knowl edge , t he exi st ence of i nt er-spec i es r ecombi nat i on be t ween t hese t wo spec i es. Abbrevi ati ons: AP, appl eproli f erat ion; ESFY, European st one frui t yellows; MLST, mul t ilocus sequenc e typing; PD, pear de cline; SNP, singl e nucl eot ide polymorphism; SSH, suppression subtract ivehybridizat ion. The GenBank/EMBL/DDBJ acc ession numbers f or t he ac eF, pnp, secY and imp gene fragments isol at ed in t hisst udy are list ed in Tabl es 3–6. A suppl ement ary t abl e, showing t he ori gin and haplotype of all phyt opl asma isol at es, isavail abl ewi t h t he online version of t his paper. Mi crobiology ( 2011), 157, 438–450 DO I 10. 1099/ mi c. 0. 043547-0 438 043547 G 2011 SGM Print ed inGre at Bri t ain
Transcript of Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and...

Multilocus sequence analysis reveals the geneticdiversity of European fruit tree phytoplasmas andsupports the existence of inter-speciesrecombinationJean Luc Danet,1 Gulnara Balakishiyeva,2 Agnes C imerman,1
N icolas Sauvion,3 Veronique Marie-Jeanne,3 G erard Labonne,3
Amparo Lavina,4 Assumpcio Batlle,4 Ivana Krizanac,5 Dijana Skoric,6
Paolo Ermacora,7 C igdem Ulubas Serce,8 Kadriye Caglayan,8
Wolfgang Jarausch9 and Xavier Foissac1
C orrespondenceX avier [email protected]
Received 7 July 201 0Revised 8 September 201 0Accepted 10 September 20 10
1UMR G enomique Diversite Pouvoir Pathogene, INRA, Universite Victor Segalen Bordeaux 2,7 1 Avenue Edouard Bourlaux, B P8 1, F-3 38 83 Villenave d’Ornon C edex, France
2Institute of Botany, National Academy of Sciences of Azerbaijan, Patamdar shosse 40, A Z-107 3,B aku, Azerbaijan
3UMR B iologie et G enetique des Interactions Plante-Parasite, CIRAD, INRA, SUPA GRO, CampusInternational de B aillarguet, TA A-5 4 / K , F-34 39 8 Montpellier C edex 5, France
4Institut de Recerca i Tecnologia Agroalimentaries (IRTA), C tra C abrils Km 2, E-0 83 4 8 C abrils,B arcelona, Spain
5Institute for Plant Protection, Croatian C entre for Agriculture, Food and Rural Affairs,Svetosimunska 2 5, HR-10 0 00 Zagreb, Croatia
6Department of B iology, University of Zagreb, Marulicev trg 9a, HR-10 0 00 Zagreb, Croatia7Dipartimento di B iologia e Protezione delle Piante, Universita degli Studi di Udine, via Delle Scienze
2 08, I-33 10 0 Udine, Italy8Department of Plant Protection, Mustafa K emal University, TK-31 03 4 Antakya-H atay, Turkey9 A lPlanta – Institute for Plant Research, RLP AgroScience GmbH , Breitenweg 71, D-6 74 35
N eustadt an der Weinstrasse, G ermany
The genetic diversity of three temperate fruit tree phytoplasmas ‘ C andidatus Phytoplasmaprunorum’, ‘ C a. P. mali’ and ‘ C a. P. pyri’ has been established by multilocus sequence analysis.Among the four genetic loci used, the genes imp and ace F distinguished 3 0 and 2 4 genotypes,respectively, and showed the highest variability. Percentage of substitution for imp ranged from 50to 6 8 % according to species. Percentage of substitution varied between 9 and 1 2 % for ace F ,whereas it was between 5 and 6 % for pnp and secY. In the case of ‘ C a P. prunorum’ the threemost prevalent ace F genotypes were detected in both plants and insect vectors, confirming thatthe prevalent isolates are propagated by insects. The four isolates known to be hypo-virulent hadthe same ace F sequence, indicating a possible monophyletic origin. H aplotype networkreconstructed by e B URST revealed that among the 34 haplotypes of ‘ C a. P. prunorum’, the fourhypo-virulent isolates also grouped together in the same clade. G enotyping of some S panish andAzerbaijanese ‘ C a. P. pyri’ isolates showed that they shared some alleles with ‘ C a. P. prunorum’,supporting for the first time to our knowledge, the existence of inter-species recombinationbetween these two species.
Abbreviations: AP, apple proliferation; ES F Y, European stone fruit yellows; MLST, multilocus sequence typing; PD, pear decline; SNP, single nucleotidepolymorphism; SS H , suppression subtractive hybridization.
The G enBank / EMB L / DD B J accession numbers for the aceF, pnp, secY and imp gene fragments isolated in this study are listed in Tables 3–6.
A supplementary table, showing the origin and haplotype of all phytoplasma isolates, is available with the online version of this paper.
Microbiology (2 01 1), 15 7, 4 3 8–4 5 0 D O I 10.1 09 9 / mic.0.043547-0
438 0 4 35 47 G 2 01 1 SGM Printed in Great Britain

IN TR O D U C TI O NPhytoplasmas are plant-pathogenic bacteria belonging tothe class Mollicutes, a group of wall-less micro-organismsphylogenetically related to low G + C-content, Gram-positive bacteria (Weisburg et al., 1989). They areresponsible for hundreds of crop diseases worldwide (Leeet al., 2000) and are transmitted from plant to plant by sap-sucking hemipteran insects (Weintraub & Beanland, 2006).European temperate fruit trees are affected by diseasesassociated with phytoplasmas which cause importanteconomic losses to the European fruit production.‘Candidatus Phytoplasma prunorum’ is responsible forEuropean stone fruit yellows (ESFY) in trees and shrubs ofthe genus Prunus, ‘Candidatus P. mali’ is the causativeagent of apple proliferation (AP) in those of the genusMalus and ‘Candidatus P. pyri’ is responsible for peardecline (PD) in Pyrus (Seemuller & Schneider, 2004). APaffects European and Turkish apple orchards (Seemuller,1990; Sertkaya et al., 2008) and the highest economicalimpact occurs in Germany and northern Italy. PD has beenreported in Europe, North America and the Middle East(Battle et al., 1999; Blomquist & Kirkpatrick, 2002b;Choueiri et al., 2007; Davies et al., 1992; Lorenz et al.,1995; Malinowski et al., 1996; Sharbatkhari et al., 2008)and also in Taiwan (Liu et al., 2007). ESFY is present in theEuro–Mediterranean basin (Marcone et al., 2010) and kills5% of the apricot trees per year in southern France(Jarausch et al., 2001) due to a severe form of apricotchlorotic leafroll. It also induces plum leptonecrosis onPrunus salicina (Japanese plum) and yellows on most of thepeach accessions in southern Europe. These three phyto-plasmas belong to the same phylogenetic cluster, the group16SrX (Lee et al., 2000), and form three closely relatedsubgroups (Jarausch et al., 1994; Lorenz et al., 1994).
‘Ca. P. mali’, ‘Ca. P. pyri’ and ‘Ca. P. prunorum’ aretransmitted from tree to tree by psyllids of the genusCacopsylla (Carraro et al., 1998; Frisinghelli et al., 2000;Jarausch & Jarausch, 2009; Jensen et al., 1964) and can bepropagated by grafting or multiplication of infectedmaterial in nurseries. In addition, ‘Ca. P. mali’ can alsobe transmitted through rootbridges among neighbouringtrees in an orchard (Ciccotti et al., 2007). Difference invirulence has been shown between isolates of ‘Ca. P.prunorum’ (Kison & Seemuller, 2001) and of ‘Ca. P. mali’(Seemuller & Schneider, 2007) giving the first evidence ofthe biodiversity in this clade of phytoplasmas. In the case of‘Ca. P. prunorum’, hypo-virulent isolates of the samespecies have been reported to occur in France and attemptshave been made to use them for cross protection(Cornaggia et al., 1995; Morvan et al., 1991). The bio-diversity of ‘Ca. P. mali’ was recently investigated by single-strand conformation polymorphism and sequence analysesof the hflB gene (Schneider & Seemuller, 2009) as anoutcome of the ‘Ca. P. mali’ genome sequencing (Kubeet al., 2008). However, little is known about the geneticdiversity of ‘Ca. P. prunorum’ and ‘Ca. P. pyri’.
In the absence of typing tools, it is not possible to trace theroute of propagation of phytoplasma strains. Typing toolsare therefore needed to improve disease control. The mostappropriate method for genotyping prokaryotes is multi-locus sequence typing (MLST) which is widely used inbacterial epidemiology and population genetics (Maiden etal., 1998; Urwin & Maiden, 2003). However, it requires theidentification of gene sequences. For this purpose, andbecause very few genes have been characterized in thisgroup of phytoplasmas, genes of ‘Ca. P. prunorum’ werecloned using a subtractive method. To design a multilocussequence analysis of phytoplasmas of the group 16SrX, fourgenes were targeted: two genes involved in carbon andnucleotide metabolism and two genes encoding the immu-nodominant surface protein Imp and SecY, a componentof the protein secretion machinery. The results of thisinvestigation are presented with special emphasis on thegenetic diversity of ‘Ca. P. prunorum’.
M E T H O D SP l a n t m a t e ri a l a n d i n s e c ts. Reference phytoplasma strains AT,AP15, GSFY2 and PD1 had been transmitted by dodder from fruittrees to periwinkle (Catharanthus roseus cv. Cooler) and since thenhave been maintained in periwinkle by successive graft inoculation(Carraro et al., 1988; Marcone et al., 1999; Marwitz et al., 1974).Samples from infected Prunus, Malus or Pyrus species were collectedfrom different regions of France, Spain, Austria, Germany, Croatia,Hungary, the UK, Bosnia and Herzegovina, Romania, Italy,Switzerland, Turkey, Lebanon and Azerbaijan (Table 1). Cacopsyllapruni psyllids were collected in Spring on Prunus spinosa (wildPrunus) in France and Spain, and on Japanese plum in Italy, byhitting branches above a lying net (Table 1). Plant DNA was tested byECA1-ECA2 PCR for ‘Ca. P. prunorum’ (Jarausch et al., 1998), byAP3-AP4 PCR for ‘Ca. P. mali’ (Jarausch et al., 1994) and by universalphytoplasma 16SrDNA nested-PCR (Gundersen & Lee, 1996)followed by sequencing for ‘Ca. P. pyri’. Only positive samples wereincluded in this study.
D o u b l e s u p pr e ssi o n s u b tr a c tiv e hy b ri d i z a ti o n (S S H) a n dg e n o m e w a l k i n g. Double SSH was performed using the PCR-Select bacterial genome subtraction kit (BD Bioscience) except for theprimers and adaptors which were modified as described previously(Cimerman et al., 2006). Total DNA from healthy and GSFY2-infected periwinkle (2 mg) were digested overnight at 37 uC by RsaI(MBI Fermentas). RsaI-digested healthy plant DNA was used as thedriver. Two populations of tester DNA were produced by ligatingadaptors 1 and 2R to RsaI-digested DNA of infected plants. Aliquotsof the ligation products were hybridized with an excess of driveraccording to the manufacturer’s instructions. These first hybridiza-tion mixtures were then hybridized together in the presence of anadditional amount of driver. Hybrids carrying both adaptors 1 and 2Rwere amplified by nested PCR according to the manufacturer’sinstructions using Taq polymerase (Promega). PCR amplification wasperformed in a 25 ml reaction volume with 0.4 mM primer P1. Thetemplates were first heated for 2 min at 72 uC to fill extremities andthen denatured for 25 s at 94 uC. PCR conditions were 30 cycles (10 sat 94 uC, 30 s at 62 uC, 1 min 30 s at 72 uC) with a single finalextension of 7 min at 72 uC. Twenty identical cycles were used fornested amplification, with the sense and anti-sense primers NP1 andNP2R, and followed by a single final extension of 7 min at 72 uC. Forreverse SSH, DNA from infected plants was used as the driver andDNA from healthy plants was used as the tester.
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 43 9
For double SSH, adaptors 1 and 2R were removed from the SSHproduct by RsaI digestion and replaced by the adaptors 3 and 4Rbefore proceeding to the second round of subtraction. Then, RsaI-digested SSH product ligated to new adaptors NP3 and NP4 was usedas tester and the RsaI reverse SSH product was used as the driver. PCRamplification was carried out with 0.4 mM primer P3. Nestedamplification of the double SSH product was performed with primersNP3 and NP4R (Cimerman et al., 2006). One microlitre of the doubleSSH product was ligated to pGEM-T Easy (Promega) and cloned inEscherichia coli DH10B according to standard procedures (Sambrooket al., 1989).
One of the double SSH clones (SDR47) was extended by genomewalking. Extension of the SDR47 sequence was performed using the
Biosciences Clontech BD GenomeWalker universal kit according tothe manufacturer’s instructions. Libraries for genome walking werecomposed of endonuclease-restricted DNA of periwinkles infectedwith ‘Ca. P. prunorum’ isolate GSFY2.
E xtr a c ti o n o f D N A fr o m p l a n ts a n d i n s e c ts, a n d P C R a m p lifi c a -ti o n . Nucleic acids were extracted from plants and insects asdescribed by Maixner et al. (1995). The aceF, pnp, secY and impgenes were amplified by nested PCR using the primers described inTable 2. First amplifications were performed using an initialdenaturation step at 95 uC for 3 min, followed by 20 cycles consistingof 94 uC for 30 s, 50 uC for 30 s and 66 uC for 45 s, and by a finalextension step at 66 uC for 7 min. Nested amplification was carriedout using 1 ml of the first amplification product with an initialdenaturation step at 95 uC for 3 min followed by 35 cycles consistingof 94 uC for 30 s, 50 uC for 30 s and 66 uC for 45 s, and by a finalextension step at 66 uC for 7 min. Nested PCR products wereseparated on a 1% agarose gel; DNA was stained with ethidiumbromide and visualized under UV light.
S e q u e n cin g a n d s e q u e n c e a n a lysis. Sequencing reactions of theSSH plasmids and MLST PCR products were performed by Cogenicson ABI PRISM capillary sequencing instruments. Raw sequencechromatograms were assembled and edited using GAP4 (Bonfieldet al., 1995) or the PHRED-PHRAP-CONSED package (Ewing & Green,1998; Ewing et al., 1998; Gordon et al., 1998). All MLST PCRproducts were directly sequenced on both strands to achieve a 26coverage. Homologies between SSH clone sequences and knownsequences were detected using the BLASTX algorithms against the non-redundant GenBank database at http://www.ncbi.nlm.nih.gov/blast.Multiple alignments were performed using CLUSTAL W (Thompsonet al., 1994) and the phylogenetic analyses were carried out withMEGA2 (Kumar et al., 2001). Haplotype networks were constructedusing the eBURST programme (Feil et al., 2004)
R E S ULT S
D o u b l e S S H a n d P C R a n a lysis o f p hyt o p l a s m a s
A library was constructed by cloning the RsaI double SSHproduct obtained by double-subtracting the total DNA ofhealthy periwinkle from the total DNA of a periwinkleinfected with ‘Ca. P. prunorum’ isolate GSFY2. Thirtyplasmid inserts with sizes ranging from 536 to 971 bp weresequenced and identified by a BLASTX similarity search asbeing phytoplasma partial gene sequences. Most of thesequences were redundant in the library and finallycorresponded to 11 unique sequences which are listedelsewhere (Danet et al., 2008). The peptides predicted fromSSH sequences shared 53–90% identity with proteinspredicted from the ‘Ca. P. asteris’ genome sequence(Oshima et al., 2004). The sequence downstream of theacoB (insert SDR47) was determined by genome walkingand used to identify the sequence of the neighbouring geneaceF. Among the ‘Ca. P. prunorum’ genes identified, aceFand pnp were chosen as targets for the variability studybecause they presented the lowest conservation level whencompared with the ‘Ca. P. asteris’ homologous genes,which had 41 and 53% identity, respectively, at thepredicted protein level. In addition to these two genesinvolved in sugar and nucleotide metabolism, the twogenes secY and imp were selected to drive the genetic
T a b l e 1. O rigin of the phytoplasma isolates tested
Country Host No. of isolates tested
‘Ca. P.prunorum’
‘Ca. P.pyri’
‘Ca. P.mali’
Azerbaijan Prunus armeniaca 2Prunus myrobolan 1
Croatia Prunus armeniaca 8France Cacopsylla pruni 26
Prunus armeniaca 10Prunus spinosa 9Prunus salicina 3Prunus persica 2Prunus marianaGF8-1
5
Prunus sp. 10Germany Prunus armeniaca 22Greece Prunus armeniaca 1Italy Prunus armeniaca 9
Cacopsylla pruni 4Spain Cacopsylla pruni 2
Prunus armeniaca 9Turkey Prunus armeniaca 3Azerbaijan Pyrus communis 3Croatia Cacopsylla pyri 2
Cacopsyllapyrisuga
1
Pyrus communis 8France Pyrus communis 3Germany Pyrus communis 8Italy Pyrus communis 6Lebanon Pyrus communis 4Spain Pyrus communis 5Turkey Cacopsylla pyri 1
Pyrus communis 2UK Pyrus communis 3Austria Malus domestica 1CzechRepublic
M . domestica 1
France M . domestica 9Germany M . domestica 13Italy M . domestica 13Romania M . domestica 1Switzerland M . domestica 1
J. L. Danet and others
440 Microbiology 1 57

diversity study. secY is involved in protein secretion and itssequence was kindly provided by the ‘Ca. P. mali’ genomesequence consortium (Kube et al., 2008). imp encodes theimmunodominant surface protein and had been sequencedfor the three species members of the 16SrX group (Mortonet al., 2003).
In order to develop a polyvalent PCR test for the entire16SrX group, three nested PCR tests were designed fromaceF, pnp and secY genes with primer pairs selected inregions encoding conserved domains of the proteins. Tocheck the effectiveness of PCR primers, they were challengedagainst periwinkles infected with Italian and German ‘Ca. P.prunorum’ isolates and German ‘Ca. P. mali’ and ‘Ca. P.pyri’ isolates. Each primer pair allowed the direct amplifica-tion of aceF, pnp and secY DNA fragments for allphytoplasma tested provided that annealing and elongationtemperatures were 50 and 66 uC, respectively (data notshown). For imp, due to its important variability among16SrX phytoplasmas (Morton et al., 2003), species-specificprimer pairs had to be designed.
aceF, pnp, secY and imp nested PCR assays were applied toa collection of 197 phytoplasma isolates originating fromFrance, Germany, Italy, Spain, Croatia, Switzerland, theUK, Hungary, Turkey, Greece, Azerbaijan and Lebanon. Allsamples had previously tested positive for 16SrX phyto-plasmas by 16S rDNA PCR diagnosis. Most of the samples(157 of 197) reacted positively for the four nested PCR tests
including all the 32 infected Cacopsylla pruni tested (‘Ca. P.prunorum’ insect vector).
Sequences of the PCR products were determined andcompared by multiple alignments. The presence of species-specific sequence motifs and important genetic distanceswere in agreement with the existence of three distinctphytoplasma species. Sequence alignments indicated thatfour deletions (total 15 bp) and one 3 bp insertion tookplace in the ‘Ca. P. pyri’ aceF sequence by comparison with‘Ca. P. prunorum’, whereas five deletions totalling 48 bpdistinguished the ‘Ca. P. mali’ aceF sequence. Theproportion of substitutions in aceF ranged from 9.6%between ‘Ca. P. pyri’ and ‘Ca. P. mali’ to 12% between ‘Ca.P. pyri’ and ‘Ca. P. prunorum’. No insertion or deletionoccurred in pnp sequences and substitutions affected 5–6%of nucleotide positions depending on the ‘Ca. Phytoplasma’species compared. In the secY gene, sequences divergedabout 5% between species and one 3 bp deletion wasobserved for ‘Ca. P. pyri’.
imp was the most variable gene with 34.3 sequence identitybetween ‘Ca. P. prunorum’ and ‘Ca. P. pyri’, and 32.1%between ‘Ca. P. prunorum’ and ‘Ca. P. mali’, while ‘Ca. P.mali’ and ‘Ca. P. pyri’ imp sequences were 50.8% identical.Two 3 bp deletions took place in imp of ‘Ca. P. prunorum’;one 9 bp deletion and two 3 bp insertions characterize impof ‘Ca. P. mali’; whereas a 3 bp insertion was specific to the‘Ca. P. pyri’ imp gene.
T a b l e 2. O ligonucleotides used for P C R and nested P C R
Gene Length (bp) Purpose Primer Sequence (5§–3§)
aceF 797 PCR AceFf1 TAAAATTCGCTGATGTTGGCGAceFr1 CATCTTTCAATTCATTAAAACTAG
Nested-PCR AceFf2 AGGTATTGAAGAAGGAACTGAceFr2 CAACCGCTTTCATAATAAAAG
pnp 549 PCR Pnpf1 GAAGTTGGTATTACTGCTTTACPnpr1 GATAAATCTATTTGACCGCG
Nested-PCR Pnpf2 TACAATTAGATATTAAAGTTAAAGGPnpr2 ATTAATTTTAATACATTTCGC
secY 664 PCR SecYMalF1 TTAGGACGTAGTATACAAATCCCNTTSecYMalR1 ACAATAATTAAAAATCCTGTNCC
Nested-PCR SecYMalF2 AAGAATGGCGTGAACARGGNGASecYMalR2 GCATCTTGTTTAGATAAATGTTC
imp ‘Ca. P. prunorum’ 673 PCR IMPF1 CAAATGATAAAGCTGATCAAIMPR1 CCTTTAAGACCACATCCTTTG
Nested-PCR IMPF2 CAGTGTATTAAATTAACAATGIMPR2 CCATCATTACAACTAAAACAT
imp ‘Ca. P. pyri’ 452 PCR IMPF2bis GTATTTAGTTACAATTATTTACTAGIMPR1bis CAAGACCTTTAAGGCCACATC
Nested-PCR IMPF3pyr AGGAGAAAAAATAATGGAAGCIMPR4pyrA TTGTGTTCTAAAAGTATCAC
imp ‘Ca. P. mali’ 670 PCR IMPF1 CAAATGATAAAGCTGATCAAIMPR1bis CAAGACCTTTAAGGCCACATC
Nested-PCR IMPF3 GTTTTATGTTATAATAAACAGTGIMPR3mal CAAACTATAGTTAAAATTAAAGC
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 44 1

S e q u e n c i n g a c e F , p n p , s e c Y a n d i m p i d e n tifi e sg e o gr a p h ic a lly r e l a t e d g e n o typ e s
The sequence of the aceF marker was determined for 197isolates of the three Candidatus species. Maximum-parsimony analysis of aceF sequences confirmed theexistence of three phylogenetic clusters corresponding to‘Ca. P. prunorum’, ‘Ca. P. mali’ and ‘Ca. P. pyri’ (Fig. 1,Table 3). For ‘Ca. P. prunorum’, 11 genotypes werediscriminated, most of them differing by one or two singlenucleotide polymorphisms (SNPs). Only genotype A9 [anapricot isolate and a Prunus cerasifera (myrobalan plum)isolate from Azerbaijan] presented 10–12 SNPs comparedwith the other ‘Ca. P. prunorum’ genotypes. The genotypeA3 represented 57% of infection cases. This genotype wasfound in isolates from France, Germany, Spain and Italy(Table 3). Two other genotypes, A8 and A6, accounted for19 and 14% of the isolates, respectively. These genotypeswere detected in isolates from France, Croatia, Germany,Italy, Turkey, Spain, Greece and Azerbaijan (Table 3).Seven aceF genotypes were only found in a single country,for example, genotypes A7, A9 and A17 were only detectedin Croatia, Azerbaijan and Turkey, respectively.Interestingly, the four hypo-virulent isolates of ‘Ca. P.prunorum’ tested (namely PVC-LA8 and PVC-LA9, B7
and E22) clustered together within the same A6 genotype.This monophyletic clustering of hypo-virulent isolatesindicates their possible single origin. Five ‘Ca. P. pru-norum’ genotypes A3, A5, A6, A8 and A21, out of 11genotypes, could be detected in C . pruni insect vectors(Table 3). For ‘Ca. P. mali’, among six genotypes differingin between one and three SNPs, the genotype A13 waspredominant (65%) and was detected in isolates fromFrance, Italy and Germany (Table 3). For ‘Ca. P. pyri’, theseven genotypes showed a genetic divergence higher thanthat of ‘Ca. P. prunorum’ genotypes, in the range of 1 to 24SNPs. Genotype A10 represented 72% of the isolates andwas detected in isolates from eight of the nine countriessurveyed (Table 3). The six other genotypes were eachfound in a single country. Again, countries in the easternpart of the Mediterranean basin, such as Turkey orLebanon, had specific ‘Ca. P. pyri’ genotypes.
Sequences of the pnp and secY markers were determined for172 and 157 isolates of the three Candidatus species,respectively. Maximum-parsimony analyses were con-ducted with the sets of pnp and secY sequences (Fig. 2).Phylogenetic trees obtained for both genes were consistentwith the existence of the three described taxonomic species;all isolates of a given species were clustered on the samebranch supported by high bootstrap values. The diversityof pnp and secY was lower than that of aceF: 15 genotypesfor pnp and 12 genotypes for secY could be discriminated(Tables 4 and 5). For ‘Ca. P. prunorum’, two and threegenotypes could be distinguished with pnp and secY,respectively, a much lower diversity when compared withthe 11 aceF genotypes. The two pnp genotypes were alsodetected in C . pruni psyllids whereas only two of three secYgenotypes were detected in C . pruni (Tables 4 and 5). For‘Ca. P. mali’, five and four genotypes were distinguishedwith pnp and secY respectively, whereas for ‘Ca. P. pyri’,eight and five genotypes were differentiated with the samemarkers. The most prevalent pnp genotype was P1 in ‘Ca.P. prunorum’ isolates originating from plant and insect.For secY, ‘Ca. Phytoplasma’ species-specific prevalenceswere seen: S1 and S2 in ‘Ca. P. prunorum’ isolates, S7 in‘Ca. P. pyri’ isolates and S12 in ‘Ca. P. mali’ isolates.
The sequence of the imp marker was determined for 151isolates of the three Candidatus species. Phylogeneticanalysis using maximum parsimony showed 30 differentgenotypes (Fig. 3a). Clustering was again in agreement withthe description of three Candidatus species; all isolates of agiven species were clustered on the same branch supportedby bootstrap values of 100. Seven, nine and 14 genotypescould be distinguished for ‘Ca. P. mali’, ‘Ca. P. pyri’ and‘Ca. P. prunorum’, respectively. The number of mutationsbetween genotypes of the same species was much higher in‘Ca. P. pyri’ and ‘Ca. P. mali’ than in ‘Ca. P. prunorum’.For ‘Ca. P. prunorum’, the most prevalent genotype,genotype I1, was detected in 56 plant and insect samplescoming from six countries (Table 6). Six genotypes weredetected in two to four countries and eight genotypes werefound in only one country. This was the case for genotypes
F i g . 1. Evolutionary relationships of ace F for isolates of the group16 SrX. The evolutionary history was inferred by using themaximum-parsimony method. The percentage of replicate treesin which the associated taxa clustered together in the bootstraptest (50 0 replicates) is shown next to the branches. B ootstrapvalues below 90 are omitted. The trees are drawn to scale; branchlengths were calculated using the average pathway methodand represent the number of nucleotide changes over thewhole sequence. Major genotypes are indicated (%). H YPV,Hypo-virulent.
J. L. Danet and others
442 Microbiology 1 57
I5 and I26, respectively, which were detected in Azerbaijanand Turkey. Five genotypes were specifically detected inFrance. A gradient in the geographical distributionappeared between western and central Europe (Fig. 3b).Two of the Spanish genotypes (red and pale blue in Fig. 3)were only found in isolates from France and Germany,whereas the prevalent I1 and I2 genotypes (dark blue inFig. 3) were absent in isolates from Spain. Genotypes ofisolates from the eastern part of the Mediterranean basin,such as those detected in isolates from Turkey andAzerbaijan (brown and orange in Fig. 3) were not detectedin central or western Europe.
Id e n tific a ti o n o f p hyt o p l a s m a h a p l o typ e s r e v e a lsn o g e o gr a p h ic a l a ss o c i a t i o n
For all four genes and all three phytoplasma candidatespecies, a total of 73 different haplotypes were detected(Supplementary Table S1, available with the online versionof this paper). In the case of ‘Ca. P. prunorum’, for which118 isolates were fully genotyped, the haplotype networkcombining the four genetic loci aceF, pnp, imp and secY wasbuilt (Fig. 4). Thirty-four haplotypes were organized into acontinuous network with A3-P1-I1-S1 and A3-P1-I9-S1
(Fig. 4, in grey) as founders representing 28% of the isolates.No relationship was found between the haplotype and thegeographical origin of the plant (apricot, peach, Japaneseplum) or psyllid. Interestingly, the four hypo-virulentisolates clustered on the same part of the haplotype network(Fig. 4, in black), again suggesting their common origin.Fifteen of the haplotypes were detected in psyllids and couldpossibly be epidemic if these insects were to be vectors. Itwas also noticeable that the major haplotype A3-P1-I1-S1was only detected once in psyllids, whereas the less abundanthaplotypes A3-P1-I1-S2 and A3-P1-I9-S2 were found twelveand seven times in the insect vector, respectively.
E vi d e n c e f o r ‘ C a . P . pyri’ a n d ‘ C a . P . pru n o ru m ’i n t e r-s p e ci e s r e c o m b i n a ti o n
The most striking result was obtained with three ‘Ca. P.pyri’ isolates collected in Spain and Azerbaijan. Multilocussequence analysis revealed that they possess both ‘Ca. P.pyri’ and ‘Ca. P. prunorum’ genetic markers (Fig. 5). Forexample, isolate Spa3 had aceF and imp genes of ‘Ca. P.prunorum’ while its pnp and secY genes corresponded to a‘Ca. P. pyri’ genotype. Isolate Spa2 also had two genescorresponding to ‘Ca. P. prunorum’ genotypes and two
T a b l e 3. O rigin of phytoplasma isolates with the ace F genotype
Referenceisolate
Geographical origin* EMBLaccession no.
Pathogen aceF genotype No. of plants No. of insects
WJ3703/30 Germany FN598166 ‘Ca. P. prunorum’ A1 1 0Psalor France FN598167 ‘Ca. P. prunorum’ A2 1 0GSFY2 Germany, France, Italy, Spain FN598168 ‘Ca. P. prunorum’ A3 47 24ESFYSpa01 Spain FN598169 ‘Ca. P. prunorum’ A4 3 040-04Cr Croatia, Italy FN598170 ‘Ca. P. prunorum’ A5 1 1B7 France, Croatia, Germany, Italy, Turkey FN598171 ‘Ca. P. prunorum’ A6 15 361-04Cr Croatia FN598172 ‘Ca. P. prunorum’ A7 1 0ESFY Italy, Azerbaijan, Croatia, France,
Germany, Greece, SpainFN598173 ‘Ca. P. prunorum’ A8 21 3
Azer10 Azerbaijan FN598174 ‘Ca. P. prunorum’ A9 2 0Ya15 Turkey FN598175 ‘Ca. P. prunorum’ A17 2 0E60-11B Italy FN598176 ‘Ca. P. prunorum’ A21 0 1PD Germany, Azerbaijan, Croatia,
France, Germany, UK, Italy, TurkeyFN598177 ‘Ca. P. pyri’ A10 24 2
PD121Lib Lebanon FN598178 ‘Ca. P. pyri’ A11 4 0PD2Luca Italy FN598179 ‘Ca. P. pyri’ A12 1 0PP/ZG/1 Croatia FN598180 ‘Ca. P. pyri’ A18 2 0PI/ZG/1 Croatia FN598181 ‘Ca. P. pyri’ A19 0 1PTK7 Turkey FN598182 ‘Ca. P. pyri’ A20 0 1TR/1 Germany FN598183 ‘Ca. P. pyri’ A24 1 0AP-AT Germany, France, Italy FN598184 ‘Ca. P. mali’ A13 22 0AP13 Romania FN598185 ‘Ca. P. mali’ A14 1 0AP032-10 France, Austria, Italy FN598186 ‘Ca. P. mali’ A15 7 0AP1Luca Italy FN598187 ‘Ca. P. mali’ A16 2 0TN/1 Italy FN598188 ‘Ca. P. mali’ A22 1 0NW/2 Germany FN598189 ‘Ca. P. mali’ A23 1 0
*The aceF gene was isolated from samples isolated from the country/countries listed.
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 44 3

genes clustering within the ‘Ca. P. pyri’ branch of thephylogenetic trees, whereas AZ-POI45 had only a ‘Ca. P.prunorum’ pnp gene and three other gene markerscorresponding to ‘Ca. P. pyri’. Altogether, these resultsindicated that these isolates of ‘Ca. P. pyri’ are inter-speciesrecombinants. Surprisingly, for the Spa2 isolate, twodifferent imp sequences could be detected, one corres-ponding to ‘Ca. P. prunorum’ and one clustering with ‘Ca.P. pyri’ imp sequences. This could not be associated with a
mixed infection as only the 16S rDNA sequence of ‘Ca P.pyri’ could be detected.
DIS C U S SI O NAn MLST method has been developed and applied fortyping temperate fruit tree phytoplasmas of the phylogeneticgroup 16SrX. The most variable markers were imp and
F i g . 2. Evolutionary relationships of pnp (a) and secY (b) for isolates of the group 16 SrX. The evolutionary history was inferredby using the maximum-parsimony method. The percentage of replicate trees in which the associated taxa clustered together inthe bootstrap test (5 00 replicates) is shown next to the branches. B ootstrap values below 9 0 are omitted. The trees are drawnto scale; branch lengths were calculated using the average pathway method and represent the number of nucleotide changesover the whole sequence.
T a b l e 4. O rigin of phytoplasma isolates with the pnp genotype
Referenceisolate
Geographical origin* EMBLaccession no.
Pathogen pnpgenotype
No. ofplants
No. ofinsects
WJ3703-30 Germany, Azerbaijan, Croatia, France, Italy, Spain FN598190 ‘Ca. P. prunorum’ P1 75 30ESFY Italy, France, Germany, Azerbaijan FN598191 ‘Ca. P. prunorum’ P2 14 2PD Germany, France, Lebanon, Spain FN598192 ‘Ca. P. pyri’ P3 5 0PD121Lib Lebanon, Azerbaijan, Croatia, UK, Spain FN598193 ‘Ca. P. pyri’ P4 9 1PD84 Italy, Lebanon FN598194 ‘Ca. P. pyri’ P5 7 0PD356-1 France, UK FN598195 ‘Ca. P. pyri’ P6 2 0PD36AZ Azerbaijan FN598196 ‘Ca. P. pyri’ P7 1 0PD2Luca Italy, Spain FN598197 ‘Ca. P. pyri’ P8 2 0PP/ZG/1 Croatia FN598198 ‘Ca. P. pyri’ P14 2 0PI/ZG/1 Croatia FN598199 ‘Ca. P. pyri’ P15 1 1AP-AT Germany, France FN598200 ‘Ca. P. mali’ P9 2 0AP15 Italy, France FN598201 ‘Ca. P. mali’ P10 3 0AP28 Switzerland, France, Italy, Romania FN598202 ‘Ca. P. mali’ P11 6 0AP032-10 France, Austria, Italy FN598203 ‘Ca. P. mali’ P12 4 0AP1Luca Italy FN598204 ‘Ca. P. mali’ P13 2 0
*The pnp gene was isolated from samples isolated from the country/countries listed.
J. L. Danet and others
444 Microbiology 1 57

aceF but should not be solely used, because differentclusterings were obtained with the markers pnp and secY.The efficiency of this typing system was illustrated by thediscrimination of 34 different haplotypes within the ‘Ca. P.prunorum’ species. MLST is particularly useful forepidemiological studies especially to compare the geno-types present in the wild and cultivated compartments, forexample in the case of ‘Ca. P. prunorum’ in Prunus spinosaversus apricot or Japanese plum orchards. MLST of thephytoplasmas detected in the psyllid vector will then showwhether ‘Ca. P. prunorum’ is transmitted from wildPrunus to orchards. Fine genotyping can also be useful totrace the route of propagation by nurseries. The dominanthaplotype A3-P1-I1-S1 that was very rarely detected inpsyllid vectors could have spread by the propagation ofinfected plant material. Phylogenetic studies can also helpto explore the origin of related phytoplasmas living indifferent hosts as has been illustrated with the commonorigin of alder and grapevine phytoplasmas in the 16SrVphylogenetic group (Arnaud et al., 2007). MLST maycontribute to functional studies of the biological traits ofphytoplasmas. For example, the hypo-virulence trait in‘Ca. P. prunorum’ (Kison & Seemuller, 2001) has beenwell characterized in the four French isolates used in thisstudy and all of them had the same aceF genotype, A6, butdifferent pnp, secY and imp genotypes. This shows that theA6 genotype is not carried by a clonal strain but by adiversified cluster of isolates. It is not yet known to whatextent the A6 genotype is linked to the hypo-virulent trait.Are the other ‘Ca. P. prunorum’ isolates with genotype A6also hypo-virulent? Biological characterization of moreisolates is needed to establish a firm correlation betweenthe A6 genotype and the hypo-virulent trait. Two of the‘Ca. P. prunorum’ isolates tested in the present studyinfect sensitive apricot varieties that remained symptom-less for many years. However, these isolates are genotypeA8, showing at least that the hypo-virulent trait does nothave a monophyletic origin. The aceF gene may be directly
responsible for hypo-virulence since the AceF protein isinvolved in glycolysis. Alternatively, a gene geneticallylinked to aceF may determine hypo-virulence. In the A6genotype, a punctual mutation replaces a threonine withan asparagine at position 189 in the E2 component of thepyruvate dehydrogenase. The effect of this mutation isunknown. The possibility that hypo-virulent isolates areimpaired in carbon metabolism can be envisaged.Glycolysis is a pathway essential for phytoplasmas whichhave a reduced physiology as shown by the analysis of theirsmall genome content (Bai et al., 2006; Oshima et al., 2004;Tran-Nguyen et al., 2008). This pathway is only partiallypresent in ‘Ca. P. mali’ strain AT, as deduced from thesequence of its linear chromosome (Kube et al., 2008) eventhough the functional activity of the remaining half of theglycolysis pathway is still unknown.
Due to the low level of similarity between imp sequences ofthe different Candidatus species, it could be stressed thatthey do not correspond to orthologous genes. Despite thelow identity at the nucleotide level, the three imp genes arecertainly orthologues and not paralogues, because identityat the protein level ranges from 43 to 49% between thethree phytoplasma species. In addition, only one copy ofimp exists in the sequenced genome of ‘Ca. P. mali’. It hasrecently been demonstrated that imp genes are highlyvariable and submitted to strong diversifying selection(Kakizawa et al., 2009). Nevertheless we cannot exclude thepossibility of hidden paralogy, in which orthologues wouldhave been duplicated and would have been lost further. Itdoes not, however, affect the results of the genotyping andits analysis.
For the first time to our knowledge, a recombinationbetween phytoplasmas has been shown. We showed that‘Ca. P. pyri’ and ‘Ca P. prunorum’ have undergonerecombination. When the results were first obtained, weenvisaged the possibility of a PCR cross-over contaminationdespite the numerous measures taken in our laboratory to
T a b l e 5. O rigin of phytoplasma isolates with the secY genotype
Referenceisolate
Geographical origin* EMBLaccession no.
Pathogen secY genotype No. ofplants
No. ofinsects
GSFY Germany, France, Italy, Spain FN598205 ‘Ca. P. prunorum’ S1 52 3ESFY Italy, Azerbaijan, Croatia, France, Spain FN598206 ‘Ca. P. prunorum’ S2 29 2914MY France, Croatia, Germany, Spain FN598207 ‘Ca. P. prunorum’ S3 6 0PDSpa2 Spain FN598208 ‘Ca. P. pyri’ S4 1 0PD36Az Azerbaijan FN598209 ‘Ca. P. pyri’ S5 1 0PD356-1 France, UK FN598210 ‘Ca. P. pyri’ S6 2 0PD Germany, France, UK, Italy, Lebanon, Spain FN598211 ‘Ca. P. pyri’ S7 10 0PD2Luca Italy, Spain FN598212 ‘Ca. P. pyri’ S8 2 0AP-AT Germany FN598213 ‘Ca. P. mali’ S9 3 0AP4Luca Italy, Austria FN598214 ‘Ca. P. mali’ S10 2 0AP032-10 France FN598215 ‘Ca. P. mali’ S11 1 0AP15 Italy, France, Romania FN598216 ‘Ca. P. mali’ S12 10 0
*The secY gene was isolated from samples isolated from the country/countries listed.
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 44 5

prevent such artefacts. So, the experiment was repeatedbut the same results were obtained and still the negativePCR controls remained negative. So our results werereproducible and we finally excluded the possibility of aPCR contamination when looking in detail at the result ofthe Azerbaijanese isolate POI45. This isolate is recombin-ant for the pnp gene and its pnp sequence is unique, with
several mutations distinguishing this isolate from theother known pnp genotypes (P1 and P2). If this result wasdue to PCR contamination, the sequence should havebeen that of P1 or P2. For isolate Spa2, in which wedetected two different copies of imp, we could excludemixed infection between ‘Ca. P. pyri’ and ‘Ca. P.prunorum’ after 16SrDNA sequencing, but we could not
F i g . 3. G enetic variability of imp (a) and geographical distribution of ‘ C a. P. prunorum’ imp genotypes (b). (a) Evolutionaryrelationships of imp for isolates of the group 1 6 SrX. The evolutionary history was inferred by using the maximum-parsimonymethod. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (5 0 0 replicates)is shown next to the branches. B ootstrap values below 9 0 are omitted. The trees are drawn to scale; branch lengths werecalculated using the average pathway method and represent the number of nucleotide changes over the whole sequence. (b)D istribution of ‘ C a. P. prunorum’ imp genotypes in the northern Euro-Mediterranean basin. Numbers below the circles indicatethe total number of isolates per country. The colour legend is given in (a).
J. L. Danet and others
446 Microbiology 1 57

reject the possibility of amixed infection between two ‘Ca. P.pyri’ isolates, one isolate being recombinant, the other not.Another possibility could be that an isolate carries two impgenes, one copy having diverged in its function. But thequestion remains about the mechanism of such recombina-tions. Until the publication of the ‘Ca. P. mali’ genome, theability of phytoplasmas to perform recombination was
questionable because they lack the genetic information toperform homologous recombination (Bai et al., 2006;Oshima et al., 2004). Nevertheless, it was discovered that‘Ca. P. mali’ has an extended set of genes for homologousrecombination, including the essential gene recA (Kube et al.,2008). In addition, all members of the 16SrX group have alinear chromosome, a property that in combination with the
T a b l e 6. O rigin of phytoplasma isolates with the imp genotype
Referenceisolate
Geographical origin EMBLaccession no.
Pathogen impgenotype
No. ofplants
No. ofinsects
ESFY Italy, Azerbaijan, Croatia, France, Germany, Greece FN600707 ‘Ca. P. prunorum’ I1 41 15Pru33 France FN600708 ‘Ca. P. prunorum’ I2 1 0WJ3889-42 Germany FN600709 ‘Ca. P. prunorum’ I3 1 040-04Cr Croatia, France, Italy, Hungary FN600710 ‘Ca. P. prunorum’ I4 8 0Azer10 Azerbaijan FN600711 ‘Ca. P. prunorum’ I5 2 0ESFY014-1 France FN600712 ‘Ca. P. prunorum’ I6 1 005MY France FN600713 ‘Ca. P. prunorum’ I7 1 0G32 France FN600714 ‘Ca. P. prunorum’ I8 1 0B7 France, Germany, Spain FN600715 ‘Ca. P. prunorum’ I9 20 1113MY France, Croatia, Italy FN600716 ‘Ca. P. prunorum’ I10 3 3ESFYSpa01 France, Spain FN600717 ‘Ca. P. prunorum’ I11 3 201MY France FN600718 ‘Ca. P. prunorum’ I12 1 0T13 France, Croatia, Germany, Spain FN600719 ‘Ca. P. prunorum’ I13 6 1ETK-Ya15 Turkey FN600720 ‘Ca. P. prunorum’ I26 5 0PD280-4 France, UK FN600721 ‘Ca. P. pyri’ I14 3 0PD20 UK FN600722 ‘Ca. P. pyri’ I15 1 0PD36AZ Azerbaijan, Croatia FN600723 ‘Ca. P. pyri’ I16 2 0PD121Lib Lebanon FN600724 ‘Ca. P. pyri’ I17 3 0PD33Lib Lebanon, Azerbaijan FN600725 ‘Ca. P. pyri’ I18 1 0PD Germany, Croatia, Italy FN600726 ‘Ca. P. pyri’ I19 4 0PDSpa2 Spain, France FN600727 ‘Ca. P. pyri’ I20 1 0KT/1 Croatia FN600728 ‘Ca. P. pyri’ I27 2 1PI/ZG/1 Croatia FN600729 ‘Ca. P. pyri’ I28 0 1AP032-10 France, Austria, Germany, Italy FN600730 ‘Ca. P. mali’ I21 12 0AP13 Romania FN600731 ‘Ca. P. mali’ I22 2 0AP15 Italy, Germany FN600732 ‘Ca. P. mali’ I23 4 0AP-AT Germany, France FN600733 ‘Ca. P. mali’ I24 2 0AP1Luca Italy FN600734 ‘Ca. P. mali’ I25 2 0TN2 Italy FN600735 ‘Ca. P. mali’ I29 1 0HD/1 Germany FN600736 ‘Ca. P. mali’ I30 1 0
*The imp gene was isolated from samples isolated from the country/countries listed.
F i g . 4. H aplotype network of ‘ C a. P. prunorum’isolates. G rey indicates founder haplotypesand black indicates hypo-virulent isolates.
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 44 7

presence of multiple repeated genes such as hflB (Schneider& Seemuller, 2009), a gene known to be present as multiplecopies in all known phytoplasma genomes (Kube et al., 2008;Oshima et al., 2004), can facilitate the exchange of a fullchromosome arm through a single cross-over. Such a simple
recombination event would lead to the exchange of manygene markers at the same time. However, to allowrecombination between two species, they have to share acommon host. ‘Ca. P. pyri’ can be transmitted andmultiplies in peach where it induces the peach yellow leaf
F i g . 5. Evolutionary relationships of ace F (a), pnp (b), secY (c) and imp (d) for recombinant ‘ C a. P. pyri’ and isolates of the group16 SrX. The evolutionary history was inferred by using the maximum-parsimony method. The percentage of replicate trees inwhich the associated taxa clustered together in the bootstrap test (5 00 replicates) is shown next to the branches. B ootstrapvalues below 90 are omitted. The trees are drawn to scale; branch lengths were calculated using the average pathway methodand represent the number of nucleotide changes over the whole sequence. Arrows indicate ‘ C a. P. pyri’ recombinants.
J. L. Danet and others
448 Microbiology 1 57

roll (Blomquist & Kirkpatrick, 2002a; Purcell et al., 1981).Peach is a common host for ‘Ca. P. prunorum’ in the northof the Euro–Mediterranean area; it is therefore probable thatpeach was the host in which the inter-species recombinationoccurred. Recombination might also have occurred in acommon insect vector, such as C . pyri, which is morepolyphagous in autumn and might acquire both phytoplas-mas from their respective host plant. Because none of thethree ‘Ca. P. pyri’ recombinants had the same MLSTgenotype, it is likely that the recombination event is quitefrequent. Indeed, it represented three cases from two distinctgeographical areas over 19 different ‘Ca. P. pyri’ isolatestested. After their appearance, recombinants had beennaturally transmitted back to pear by psyllids, in whichthey were finally detected. One could certainly consider thatif recombination occurs between species it could also occurwithin species. We cannot confirm such a phenomenonbecause, despite incongruence between the phylogenetictrees of the different markers, the bootstrap validity levelswere not high enough to consider it evident.
A C K N O W L E D G E M E N T S
We greatly acknowledge the French Embassy in Baku and FEBS forsupporting G. B. This work was financially supported by grants fromthe French National Research Agency, programme ECOGER and theINRA department of Plant Health and Environment and by theProgramme Sectorial I + D, M.A.P.A., Spain (grant RT2009-00070).This work was partly carried out in the frame of the SEE-ERANETnetwork ‘global epidemiology of phytoplasma diseases of economicalimportance in south-eastern Europe’. We thank M. Kolber and I.Ember (both PHYTOLAB, Budapest, Hungary), P. Gentit (CTIFL,Bergerac, France), and N. Katis and V. Maliogka (both AristotleUniversity of Thessalonoki, Greece) for providing phytoplasmaisolates and H. Bahriz, Michel Yvon, J. Peyre and P. Limon forexcellent technical assistance.
R E F E R E N C E S
Arna ud, G ., Ma le m b ic-Ma h er, S ., S a lar, P., B onn e t, P., Ma ixn er, M.,Marcon e , C ., B oud on-Pad ie u, E . & F o iss a c, X. (2007). Multilocussequence typing confirms the close genetic inter-relatedness betweenthree distinct flavescence doree phytoplasma strain clusters and group16SrV phytoplasmas infecting grapevine and alder in Europe. ApplEnviron M icrobiol 73, 4001–4010.
B a i, X. D ., Z h ang, J. H ., E w ing, A., Miller, S . A., Jancso Ra d e k , A.,S h evch e nk o, D . V., Tsu k erm an, K ., Wa luna s, T., La p idus, A. & oth era uthors (2006). Living with genome instability: the adaptation ofphytoplasmas to diverse environments of their insect and plant hosts.J Bacteriol 188, 3682–3696.
B a ttl e , A ., L a vi n a , A ., M a rt a , F . & M e d i n a , V . (1999). Incidence andepidemiology of pear decline in North-Eastern Spain. First InternetConference of Phytopathogenic Mollicutes.
B lo mqu ist, C . L. & K irk p atric k , B . C . (2002a). Identification ofphytoplasma taxa and insect vectors of peach yellow leaf roll diseasein California. Plant D is 86, 759–763.
B lo mqu ist, C . L. & K irk p atric k , B . C . (2002b). Frequency and seasonaldistribution of pear psylla infected with the pear decline phytoplasmain California pear orchards. Phytopathology 92, 1218–1226.
B onfie ld, J. K ., S m ith, K . F . & S ta d e n, R. (1995). A new DNA sequenceassembly program. Nucleic Acids Res 23, 4992–4999.
C arraro, L., O sler, R., Re fatti, E . & Pogg i-po llini, C . (1988).Transmission of the possible agent of apple proliferation to V incarosea by dodder. Riv Pat V egetale. IV, 43–52.
C arraro, L., O sler, R., Lo i, N ., Erm a cora , P. & Re fatti, E . (1998).Transmission of European stone fruit yellows phytoplasma byCacopsylla pruni. J Plant Pathol 80, 233–239.
C hou e iri, E ., S a lar, P., Jre ijiri, F ., Z a m m ar, S . E ., D an e t, J. L. &F o iss a c, X. (2007). First report and characterization of pear declinephytoplasma on pear in Lebanon. J Plant Pathol 89, S75.
C iccotti, A. M., B ianch e di, P. L., Bra g agna , P., D ero m e di, M., F ilipp i, M.,F orno, F . & Matt e di, L. (2007). Transmission of ‘CandidatusPhytoplasma mali’ by root bridges under natural and experimentalconditions. Bull Insectol 60, 387–388.
C im erm an, A., Arna ud, G . & F o iss a c, X. (2006). Stolbur phytoplasmagenome survey achieved using a suppression subtractive hybridizationapproach with high specificity. Appl Environ M icrobiol 72, 3274–3283.
C orna gg ia , D ., G e ntit, P., B oye , R. & D e svign e s, J. C . (1995). A newphytoplasma disease of apricot tree: the peach vein clearing. ActaHortic 386, 448–453.
D an e t, J. L., B a hriz , H ., C im erm an, A. & F o iss a c, X. (2008). Newmolecular typing tools to monitor fruit tree phytoplasma variabilityin the 16SrX taxonomic group. XXth International symposium on virusand virus-like diseases of temperate fruit crops. Acta Hortic 781, 343–349.
D avie s, D . L., G u is e , C . M., C lark , M. F . & A d a m s, A. N . (1992). Parry’sdisease of pears is similar to pear decline and is associated withmycoplasma-like organisms transmitted by Cacopsylla pyricola. PlantPathol 41, 195–203.
E w ing, B . & Gre e n, P. (1998). Base-calling of automated sequencertraces using phred. II. Error probabilities. Genome Res 8, 186–194.
E w ing, B ., H illier, L., We nd l, M. & Gre e n, P. (1998). Base-calling ofautomated sequencer traces using phred. I. Accuracy assessment.Genome Res 8, 175–185.
F e il, E . J., Li, B . C ., A an e ns e n, D . M., H ana g e , W. P. & S pratt, B . G .(2004). eBURST: inferring patterns of evolutionary descent amongclusters of related bacterial genotypes from multilocus sequencetyping data. J Bacteriol 186, 1518–1530.
Frisingh e lli, C ., D e la iti, L., Grando, M. S ., F orti, D . & Vind im ian, M. E .(2000). Cacopsylla costalis (Flor 1861), as a vector of appleproliferation in Trentino. J Phytopathol 148, 425–431.
G ordon, D ., A b a jian, C . & Gre e n, P. (1998). Consed: a graphical toolfor sequence finishing. Genome Res 8, 195–202.
G und ers e n, D . E . & Le e , I.-M. (1996). Ultrasensitive detection ofphytoplasmas by nested-PCR assays using two universal primer pairs.Phytopathol Mediterr 35, 144–151.
Jara usch, B . & Jara usch, W. (2009). Psyllid vectors and their control.In Phytoplasmas: Genomes, Plant Hosts and Vectors, pp. 250–271.Edited by P. Weintraub & P. Jones. Wallingford: CABI.
Jara usch, W., S a illard, C ., D osb a , F . & B ove , J. M. (1994).Differentiation of mycoplasmalike organisms (MLOs) in Europeanfruit-trees by PCR using specific primers derived from the sequence ofa chromosomal fragment of the apple proliferation MLO. ApplEnviron M icrobiol 60, 2916–2923.
Jara usch, W., Lans a c, M., S a illard, C ., Broqu a ire, J. M. & D osb a , F .(1998). PCR assay for specific detection of European stone fruityellows phytoplasmas and its use for epidemiological studies inFrance. Eur J Plant Pathol 104, 17–27.
Jara usch, W., D an e t, J. L., La bonn e , G ., D osb a , F ., Broqu a ire , J. M.,S a illard, C . & G arnier, M. (2001). Mapping the spread of apricot
G enetic diversity of fruit tree phytoplasmas
http: / / mic.sgmjournals.org 44 9

chlorotic leaf roll (ACLR) in southern France and implication ofCacopsylla pruni as a vector of European stone fruit yellows (ESFY)phytoplasmas. Plant Pathol 50, 782–790.Je ns e n, D . D ., Griggs, W. H ., G on z a le s, C . Q . & S chn e id er, H . (1964).Pear decline virus transmission by pear psylla. Phytopathology 54,1346–1351.
K a k iz a wa , S ., O sh im a , K ., Ish ii, Y., H osh i, A., Ma e jim a , K ., Jung, H . Y.,Y a m a ji, Y. & N a m b a , S . (2009). Cloning of immunodominantmembrane protein genes of phytoplasmas and their in plantaexpression. FEMS M icrobiol Lett 293, 92–101.K ison, H . & S e e m uller, E . (2001). Differences in strain virulence ofthe European stone fruit yellows phytoplasma and susceptibility ofstone fruit trees on various rootstocks to this pathogen. J Phytopathol149, 533–541.
K u b e , M., S chn e id er, B ., K uh l, H ., D and e k ar, T., H e itm ann, K .,Mig d o ll, A. M., Re inh ardt, R. & S e e m uller, E . (2008). The linearchromosome of the plant-pathogenic mycoplasma ‘CandidatusPhytoplasma mali’. BM C Genomics 9, 306.
K u m ar, S ., Ta m ura , K ., Ja k obs e n, I. B . & N e i, M. (2001). MEGA2:Molecular Evolutionary Genetics Analysis (MEGA) software. Bioinfor-matics 17, 1244–1245.
Le e , I. M., D avis, R. E . & G und ers e n-Rind a l, D . E . (2000).Phytoplasma: phytopathogenic mollicutes. Annu Rev M icrobiol 54,221–255.
Liu, H .-L., C h e n, C .- C . & Lin, C .-P. (2007). Detection and identificationof the phytoplasma associated with pear decline in Taiwan. Eur JPlant Pathol 117, 281–291.Lore n z , K .-H ., D osb a , F ., Pog g i-Po llini, C ., Lla c er, G . & S e e m uller, E .(1994). Phytoplasma diseases of Prunus species in Europe are causedby genetically similar organisms. Z Pflanzenkr Pflanzenchutz 101, 567–575.
Lore n z , K .-H ., S chn e id er, B ., A hre ns, U . & S e e m uller, E . (1995).Detection of the apple proliferation and pear decline phytoplasmas byPCR amplification of ribosomal and nonribosomal DNA.Phytopathology 85, 771–776.
Ma id e n, M. C . J., B ygrave s, J. A., F e il, E ., More lli, G ., Russ e ll, J. E .,Urw in, R., Z h ang, Q ., Z h ou, J., Z urth, K . & oth er a uthors (1998).Multilocus sequence typing: a portable approach to the identificationof clones within populations of pathogenic microorganisms. Proc N atlAcad Sci U S A 95, 3140–3145.
Ma ixn er, M., A hre ns, U . & S e e m uller, E . (1995). Detection of theGerman grapevine yellows (Vergilbungskrankheit) MLO in grapevine,alternative hosts and a vector by a specific PCR procedure. Eur J PlantPathol 101, 241–250.Ma linows ki, T., Z and ars k i, J., K o m oro wsk a , B . & Z a w a d z k a , B .(1996). Detection of pear decline phytoplasma in declining pear treesin Poland. Plant D is 80, 464.
Marcon e , C ., H erg e nh a hn, F ., Ra g o z z ino, A. & S e e m uller, E . (1999).Dodder transmission of pear decline, European stone fruit yellows,rubus stunt, Picris echioides yellows and cotton phyllody phytoplas-mas to periwinkle. J Phytopathol 147, 187–192.Marcon e , C ., Jara usch, B . & Jara usch, W. (2010). CandidatusPhytoplasma prunorum, the causal agent of European stone fruityellows: an overview. J Plant Pathol 92, 19–34.Marw itz , R., P e tz o ld, H . & O z e l, M. (1974). Untersuchungen zurubertragbarkeit des moglichen erregers der triebsucht des apfels aufeinen krautigen wirt. J Phytopathol 81, 85–91 (in German).
Morton, A., D avie s, D . L., B lo mqu ist, C . L. & B arb ara , D . J. (2003).Characterization of homologues of the apple proliferation immuno-dominant membrane protein gene from three related phytoplasmas.Mol Plant Pathol 4, 109–114.
Morvan, G ., C a st e la in, C ., C h a st e lli ere , M. G . & A u d ergon, J. M.(1991). An account of the attempts at controling apricot chlorotic leafroll with cross protection. Acta Hortic 293, 555–561.
O shima, K., K akiz awa, S., N ishigaw a, H., Jung, H. Y., We i, W., S uzuki, S.,Arashida, R., Nakata, D., Miyata, S . & other authors (2004). Reductiveevolution suggested from the complete genome sequence of a plant-pathogenic phytoplasma. N at Genet 36, 27–29.
Purc e ll, A. H ., Nyland, G ., Ra ju, B . C . & H ering or, M. R. (1981). Peachyellow leaf roll epidemic in Northern California: effects of peachcultivar, tree age and proximity to pear orchards. Plant D is 65, 365–368.
S a m broo k , J., Fritsch, E . F . & Maniatis, T. (1989). Molecular C loning:a Laboratory Manual, 2nd edn. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory.
S chn e id er, B . & S e e m uller, E . (2009). Strain differentiation ofCandidatus Phytoplasma mali by SSCP and sequence analyses of thehflB gene. J Plant Pathol 91, 103–112.
S e e m uller, E . (1990). Apple proliferation. In Compendium of Appleand Pear D iseases edited by A. L. Jones & H. S. Aldwinkle, pp. 67–68.St Paul, USA: American Phytopathological Society.
S e e m uller, E . & S chn e id er, B . (2004). ‘Candidatus Phytoplasmamali’, ‘Candidatus Phytoplasma pyri’ and ‘Candidatus Phytoplasmaprunorum’, the causal agents of apple proliferation, pear decline andEuropean stone fruit yellows, respectively. Int J Syst Evol M icrobiol 54,1217–1226.
S e e m uller, E . & S chn e id er, B . (2007). Differences in virulence andgenomic features of strains of ‘Candidatus Phytoplasma mali’, theapple proliferation agent. Phytopathology 97, 964–970.
S ertk aya , G ., Martini, M. & O sler, R. (2008). First report of Candidatusphytoplasma mali in Turkey. J Plant Pathol 90, 143.
S h arb atk h ari, M., B a h ar, M. & A hoonm an e sh, A. (2008). Detection ofthe phytoplasmal agent of pear decline in Iran, Isfahan province,using nested-PCR. International Journal of Plant Production 2, 167–173.
Tho m pson, J. D ., H ig g ins, D . G . & G ibson, T. J. (1994). CLUSTAL W:improving the sensitivity of progressive multiple sequence alignmentthrough sequence weighting, position-specific gap penalties andweight matrix choice. Nucleic Acids Res 22, 4673–4680.
Tran-Nguyen, L. T. T., K ub e, M., S chn eider, B ., Reinh ardt, R. & G ibb,K . S . (2008). Comparative genome analysis of ‘‘Candidatus Phytoplasmaaustraliense’’ (subgroup tuf-Australia I; rp-A) and ‘‘Ca. Phytoplasmaasteris’’ strains OY-M and AY-WB. J Bacteriol 190, 3979–3991.
Urw in, R. & Ma id e n, M. C . J. (2003). Multi-locus sequence typing: atool for global epidemiology. Trends M icrobiol 11, 479–487.
We intra ub, P. G . & B e anland, L. (2006). Insect vectors ofphytoplasmas. Annu Rev Entomol 51, 91–111.
We isburg, W. G ., Tu lly, J. G ., Ros e , D . L., P e tz e l, J. P., O ya iz u, H .,Y ang, D ., Mand e lco, L., S e chre st, J., La wre nc e , T. G . & oth er a uth ors(1989). A phylogenetic analysis of the mycoplasmas: basis for theirclassification. J Bacteriol 171, 6455–6467.
Edited by: C . A. Boucher
J. L. Danet and others
450 Microbiology 1 57

Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
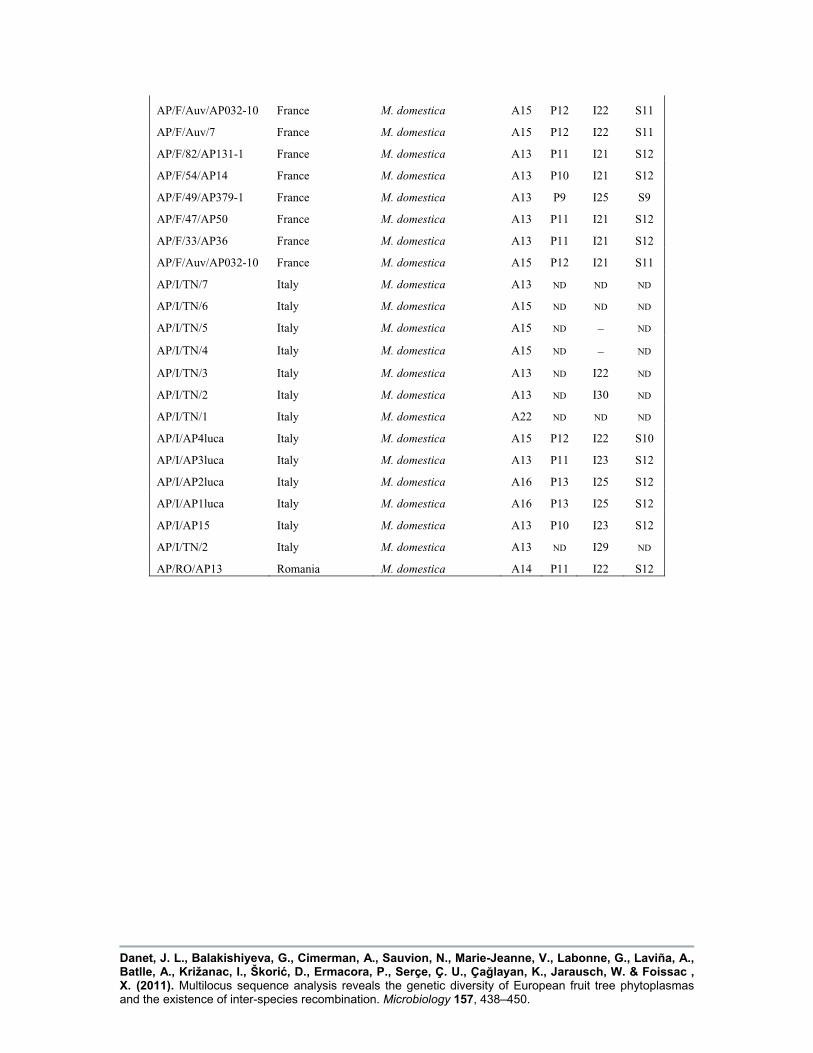
Supplementary Table S1. Origin and haplotype of all phytoplasma isolates
!, Negative amplification; ND, not done.
HaplotypeIsolate code Country of origin Host
AceF Pnp Imp SecY
‘Ca. P. prunorum’
EP/D/WJ3703-30 Germany Prunus armeniaca A1 P1 ND ND
EP/F/82/Psalor France Prunus. salicina A2 P2 I1 S1
EP/D/WJ3677-01 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3677-21 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3677-23 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3680-04 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3680-10 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3901-01 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3901-03 Germany P. armeniaca A3 P1 I1 S1
EP/D/WJ3901-06 Germany P. armeniaca A3 P1 I1 S1
EP/D/GSFY Germany P. armeniaca A3 P2 I1 S1
EI/F/82/B6 France Cacopsylla pruni A3 P2 I1 S1
EP/F/34/02MY France Prunus spinosa A3 P1 I1 S1
EP/F/47/B12 France P. armeniaca A3 P1 I1 S1
EP/F/30/Pru175 France P. armeniaca A3 P1 I1 S1
EP/F/33/GF3 France Prunus sp. A3 P1 I1 S1
EP/F/66/POMY France P. armeniaca A3 P1 I1 S1
EP/I/ESFYCluca Italy P. armeniaca A3 P1 I1 S1
EP/I/LNS1 Italy P. armeniaca A3 P1 I1 S1
EP/I/LNS2 Italy P. armeniaca A3 P1 I1 S1
EP/D/WJ3687-02 Germany P. armeniaca A3 P1 I1 S2
EI/F/03/E160-12 France C. pruni A3 P1 I1 S2
EI/F/03/E160-21 France C. pruni A3 P1 I1 S2
EI/F/03/E160-24 France C. pruni A3 P1 I1 S2
EI/F/09/E92-89 France C. pruni A3 P1 I1 S2
EI/F/12/E155-2 France C. pruni A3 P1 I1 S2
EI/F/03/E160-8 France C. pruni A3 P1 I1 S2
EI/F/34/E148-22 France C. pruni A3 P1 I1 S2
EI/F/43/E159-43 France C. pruni A3 P1 I1 S2
EI/F/66/E49-75 France C. pruni A3 P1 I1 S2
EI/F/78/E163-3 France C. pruni A3 P1 I1 S2
EI/F/89/E168-9 France C. pruni A3 P1 I1 S2

Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
EI/F/86/E80-21 France C. pruni A3 P1 I1 S2
EP/I/WJ3536-28 Italy P. armeniaca A3 P1 I1 S2
EP/F/82/Pru33 France P. salicina A3 P1 I2 S1
EP/F/34/05MY France P. spinosa A3 P1 I7 S1
EI/F/34/18MY France C. pruni A3 P1 I9 S1
EI/F/34/19MY France C. pruni A3 P1 I9 S1
EP/F/34/04MY France P. spinosa A3 P1 I9 S1
EP/D/WJ3677-09 Germany P. armeniaca A3 P1 I9 S1
EP/D/WJ3684-19 Germany P. armeniaca A3 P1 I9 S1
EP/E/ESFYSpa02 Spain P. armeniaca A3 P1 I9 S1
EP/E/ESFYSpa07 Spain P. armeniaca A3 P1 I9 S1
EP/E/ESFYSpa03 Spain P. armeniaca A3 P1 I9 S1
EP/F/49/Pru51 France P. salicina A3 P1 I9 S1
EP/F/34/03MY France P. spinosa A3 P1 I9 S1
EP/F/34/06MY France P. spinosa A3 P1 I9 S1
EP/F/34/07MY France P. spinosa A3 P1 I9 S1
EP/F/34/08MY France P. spinosa A3 P1 I9 S1
EP/F/34/09MY France P. spinosa A3 P1 I9 S1
EP/F/34/10MY France Prunus mariana GF8-1 A3 P1 I9 S1
EP/F/34/11MY France P. mariana GF8-1 A3 P1 I9 S1
EP/F/49/Pru31 France P. salicina A3 P1 I9 S1
EI/F/07/E94-2 France C. pruni A3 P1 I9 S2
EI/F/34/E135-5 France C. pruni A3 P1 I9 S2
EI/F/03/E160-10 France C. pruni A3 P1 I9 S2
EI/F/34/E176-8 France C. pruni A3 P1 I9 S2
EI/F/34/F209-24 France C. pruni A3 P1 I9 S2
EI/F/36/E86-8 France C. pruni A3 P1 I9 S2
EI/F/86/E80-9 France C. pruni A3 P1 I9 S2
EP/E/ESFYSpa04 Spain P. armeniaca A3 P1 I11 S1
EI/F/34/E148-11 France C. pruni A3 P1 I11 S2
EI/E/CA/E18-47 Spain C. pruni A3 P1 I11 S2
EP/F/34/01MY France P. spinosa A3 P1 I12 S1
EP/F/26/ESFY042-3 France P. armeniaca A3 P2 I1 S1
EP/F/26/ESFY021-3 France P. armeniaca A3 P2 I1 S1
EP/F/49/ESFY0293-4 France P. salicina A3 P2 I1 S1
EP/F/84/ECAM200 France P. armeniaca A3 P2 I1 S1
EP/F/84/ESFY017-1 France P. armeniaca A3 P2 I1 S1
EP/F/30/ESFY014-1 France P. armeniaca A3 P2 I6 S1
Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
EP/F/66/G32 France P. armeniaca A3 P2 I8 S1
EP/F/26/ESFY042-1 France P. armeniaca A3 P2 I9 S1
EP/F/30/ESFY014-2 France P. armeniaca A3 P2 I9 S1
EP/E/ESFYSpa10 Spain P. armeniaca A4 P1 I9 S1
EP/E/ESFYSpa01 Spain P. armeniaca A4 P1 I11 S1
EP/E/ESFYSpa09 Spain P. armeniaca A4 P1 ND S1
EP/TR/77/Ya13 Turkey P. armeniaca A17 ND I26 ND
EP/TR/77/Ya15 Turkey P. armeniaca A17 ND I26 ND
EP/HR/40-04Cr Croatia P. armeniaca A5 P1 I4 S2
EI/I/UD/E60-12 Italy C. pruni A5 P1 I10 S2
EP/D/WJ3790-162 Germany P. cerasifera A6 P1 I1 S1
EP/D/WJ3889-42 Germany P. armeniaca A6 P1 I3 S2
EP/F/34/12MY France P. mariana GF8-1 A6 P1 I4 S2
EP/F/84/E22 France P. armeniaca A6 P1 I4 S2
EP/HR/38-05Cr Croatia P. armeniaca A6 P1 I4 S2
EP/I/WJ3532-23 Italy P. armeniaca A6 P1 I4 S1
EP/I/ESFYAluca Italy P. armeniaca A6 P1 I4 S2
EP/I/ESFYDluca Italy P. armeniaca A6 P1 I4 S2
EP/F/84/pru128 France Prunus sp. A6 P1 I9 S2
EI/F/34/16MY France C. pruni A6 P1 I10 S2
EP/F/34/13MY France P. mariana GF8-1 A6 P1 I10 S2
EP/HR/39-05Cr Croatia P. armeniaca A6 P1 I10 S2
EI/I/UD/E60-19 Italy P. armeniaca A6 P1 I10 S2
EP/F/66/PVCLA8 France Prunus persica A6 P1 I10 S2
EP/F/66/PVCLA9 France P. persica A6 P2 I4 S2
EI/F/34/ML6 France C. pruni A6 P2 I9 S2
EP/F/84/B7 France P. armeniaca A6 P2 I9 S2
EP/TR/33/lc19 Turkey P. armeniaca A6 nd I26 nd
EP/HR/61-04Cr Croatia P. armeniaca A7 P1 I1 S2
EP/AZ/Azer4 Azerbaijan P. armeniaca A8 P1 I1 nd
EP/D/WJ3680-01 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3680-03 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3680-09 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3889-35 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3889-41 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3901-11 Germany P. armeniaca A8 P1 I1 S2
EP/D/WJ3687-06 Germany P. armeniaca A8 P1 I1 S2
EP/HR/57-04Cr Croatia P. armeniaca A8 P1 I1 S2

Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
EP/HR/25-04Cr Croatia P. armeniaca A8 P1 I1 S2
EP/HR/26-04Cr Croatia P. armeniaca A8 P1 I1 S2
EI/I/UD/E60-30 Italy P. armeniaca A8 P1 I1 S2
EP/I/ESFY87luca Italy P. armeniaca A8 P1 I1 S2
EP/GR/324GR Greece P. armeniaca A8 ND I1 ND
EI/F/34/17MY France C. pruni A8 P1 I9 S2
EP/E/ESFYSpa05 Spain P. armeniaca A8 P1 I11 S1
EI/E/CA/E12-71 Spain C. pruni A8 P1 I13 S2
EP/D/WJ3889-33 Germany P. armeniaca A8 P1 I13 S3
EP/E/ESFYSpa12 Spain P. armeniaca A8 P1 I13 S3
EP/F/30/14MY France P. armeniaca A8 P1 I13 S3
EP/F/33/T13 France Prunus sp. A8 P1 I13 S3
EP/F/66/15MY France P. armeniaca A8 P1 I13 S3
EP/HR/70-04Cr Croatia P. armeniaca A8 P1 I13 S3
EP/I/ESFY Italy P. armeniaca A8 P2 I1 S2
EP/AZ/Azer10 Azerbaijan P. armeniaca A9 P1 I5 S2
EI/I/UD/E60-11 Italy C. pruni A21 P1 I1 S2
‘Ca. P. pyri’
PP/AZ/PD36AZ Azerbaijan Pyrus communis A10 P7 I16 S5
PP/AZ/POI45 Azerbaijan Pyrus communis A10 P16 I31 S13
PP/D/AW/1 Germany Pyrus communis A10 ND ! ND
PP/D/AW/3 Germany Pyrus communis A10 ND ! ND
PP/D/EMS/2 Germany Pyrus communis A10 ND ! ND
PP/D/PD1 Germany Pyrus communis A10 P3 I19 S7
PP/D/SÜW/1 Germany Pyrus communis ! ND ND ND
PP/D/TR/1 Germany Pyrus communis A24 ND ! ND
PP/E/PD14 Spain Pyrus communis A10 P4 I19 S7
PP/E/PDSpa1 Spain Pyrus communis A3 P1 I9 !
PP/E/PDSpa2 Spain Pyrus communis A3 P8 I20+I9 S4
PP/E/PDSpa3 Spain Pyrus communis A3 P4 I9 S7
PP/E/PDSpa4 Spain Pyrus communis A10 P3 ! S8
PP/F/02/PD280-4 France Pyrus communis A10 P3 I14 S7
PP/F/33/PD356-1 France Pyrus communis A10 P6 ! S6
PP/F/33/PD87 France Pyrus communis A10 P3 ! S7
PP/GB/PD20 UK Pyrus communis A10 P6 I15 S6
PP/GB/PD27 UK Pyrus communis A10 P4 I14 S7
PP/GB/PD29 UK Pyrus communis A10 P4 I14 S7
PI/HR/KT/1 Croatia Cacopsylla pyri A10 P4 I27 ND
Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
PI/HR/OS/1 Croatia Cacopsylla pyri A10 ND ! ND
PI/HR/ZG/1 Croatia Cacopsylla pyrisuga A19 P15 I28 ND
PP/HR/CK/1 Croatia Pyrus communis A18 P15 I16 !
PP/HR/KT/1 Croatia Pyrus communis A10 P4 I27 ND
PP/HR/ST/1 Croatia Pyrus communis A10 P4 ND !
PP/HR/VZ/1 Croatia Pyrus communis A10 P5 I27 ND
PP/HR/ZG/1 Croatia Pyrus communis A18 P14 ! !
PP/HR/ZG/2 Croatia Pyrus communis A10 P14 I19 ND
PP/I/PD1luca Italy Pyrus communis A10 P5 ! !
PP/I/PD2luca Italy Pyrus communis A12 P8 ND S8
PP/I/PD3luca Italy Pyrus communis A10 P5 ! !
PP/I/PD4luca Italy Pyrus communis A10 P5 ND !
PP/I/PD5luca Italy Pyrus communis ! P5 ! !
PP/I/PD84 Italy Pyrus communis A10 P5 I19 S7
PP/LB/PD121Lib Lebanon Pyrus communis A11 P4 I17 S7
PP/LB/PD21Lib Lebanon Pyrus communis A11 P3 I17 !
PP/LB/PD29Lib Lebanon Pyrus communis A11 ! I17 S7
PP/LB/PD33Lib Lebanon Pyrus communis A11 P5 I18 !
PI/TR/PTK7 Turkey Cacopsylla pyri A20 ND ND ND
PP/TR/PTkBu5 Turkey Pyrus communis A10 ND ND ND
PP/TR/PTkBu6 Turkey Pyrus communis A10 ND ND ND
‘Ca. P. mali’
AP/A/AP29 Austria Malus domestica A15 P12 I22 S10
AP/CH/AP28 Switzerland M. domestica A13 P11 ! S9
AP/D/SÜW/1 Germany M. domestica A13 ND I23 ND
AP/D/OG/1 Germany M. domestica A13 ND ND ND
AP/D/NW/2 Germany M. domestica A23 ND I24 ND
AP/D/NW/1 Germany M. domestica A13 ND I22 ND
AP/D/HD/4 Germany M. domestica A13 ND ND ND
AP/D/HD/3 Germany M. domestica A13 ND I24 ND
AP/D/HD/2 Germany M. domestica A13 ND ND ND
AP/D/HD/1 Germany M. domestica A13 ND I31 ND
AP/D/DÜW/2 Germany M. domestica A13 ND I22 ND
AP/D/DÜW/1 Germany M. domestica A13 ND I22 ND
AP/D/AP-AT Germany M. domestica A13 P9 I25 S9
AP/D/HD/1 Germany M. domestica A13 ND I30 ND
AP/D/AP-AT Germany M. domestica A13 P9 I24 S9
AP/F/Ce/AP166-1 France M. domestica A13 P10 I21 S12
Danet, J. L., Balakishiyeva, G., Cimerman, A., Sauvion, N., Marie-Jeanne, V., Labonne, G., Laviña, A., Batlle, A., Kri!anac, I., "kori!, D., Ermacora, P., Serçe, Ç. U., Ça"layan, K., Jarausch, W. & Foissac , X. (2011). Multilocus sequence analysis reveals the genetic diversity of European fruit tree phytoplasmas and the existence of inter-species recombination. Microbiology 157, 438–450.
AP/F/Auv/AP032-10 France M. domestica A15 P12 I22 S11
AP/F/Auv/7 France M. domestica A15 P12 I22 S11
AP/F/82/AP131-1 France M. domestica A13 P11 I21 S12
AP/F/54/AP14 France M. domestica A13 P10 I21 S12
AP/F/49/AP379-1 France M. domestica A13 P9 I25 S9
AP/F/47/AP50 France M. domestica A13 P11 I21 S12
AP/F/33/AP36 France M. domestica A13 P11 I21 S12
AP/F/Auv/AP032-10 France M. domestica A15 P12 I21 S11
AP/I/TN/7 Italy M. domestica A13 ND ND ND
AP/I/TN/6 Italy M. domestica A15 ND ND ND
AP/I/TN/5 Italy M. domestica A15 ND ! ND
AP/I/TN/4 Italy M. domestica A15 ND ! ND
AP/I/TN/3 Italy M. domestica A13 ND I22 ND
AP/I/TN/2 Italy M. domestica A13 ND I30 ND
AP/I/TN/1 Italy M. domestica A22 ND ND ND
AP/I/AP4luca Italy M. domestica A15 P12 I22 S10
AP/I/AP3luca Italy M. domestica A13 P11 I23 S12
AP/I/AP2luca Italy M. domestica A16 P13 I25 S12
AP/I/AP1luca Italy M. domestica A16 P13 I25 S12
AP/I/AP15 Italy M. domestica A13 P10 I23 S12
AP/I/TN/2 Italy M. domestica A13 ND I29 ND
AP/RO/AP13 Romania M. domestica A14 P11 I22 S12




















