
Morphology and wall ultrastructure of the megaspore Lagenicula (Triletes) variabilis (Winslow, 1962)...
11
Morphology and wall ultrastructure of the megaspore Lagenicula (Triletes) mixta (Winslow 1962) comb. nov. from the Carboniferous (Early Mississippian: mid Tournaisian) of Ohio, USA Charles H. Wellman a, ⁎, Caroline Arioli b , Edwin G. Spinner a , Marco Vecoli b a Department of Animal and Plant Sciences, University of Sheffield, Alfred Denny Building, Western Bank, Sheffield S10 2TN, UK b “Géosystèmes”, UMR 8157 CNRS, Université de Lille 1, Sciences de la Terre, 59655 Villeneuve d'Ascq, France abstract article info Article history: Received 14 December 2007 Received in revised form 16 May 2008 Accepted 24 July 2008 Available online 3 August 2008 Keywords: lycopsid megaspore ultrastructure Carboniferous Early Mississippian Tournaisian Megaspores assigned to Lagenicula (Triletes) mixta (Winslow, M., 1962. Plant Spores and Other Microfossils from Upper Devonian and Lower Mississippian Rocks of Ohio. Geol. Surv., Prof. Paper 364, 1–93.) comb. nov., from the Carboniferous (Early Mississippian: mid Tournaisian) of northeastern Ohio, USA, have been analysed using light microscopy (LM), scanning electron microscopy (SEM) and transmission electron microscopy (TEM). These studies provide new information on morphology, gross structure and wall ultrastructure. This taxon has a confused taxonomic history, and the new morphological information allows recognition as a distinct species that can be placed with the genus Lagenicula as a new combination. Morphological/ultrastructural studies confirm the lycopsid affinities of this megaspore and it is suggested that it probably derived from an arborescent lycopsid that belonged with the Lepidocarpaceae. Thus it is an early example of a megaspore derived from an arborescent lycopsid of the type that went on to dominate the Euramerican Coal Measure forests. The Ohio Tournaisian megaspore assemblage is surprisingly diverse revealing an interesting insight into vegetation ecology at this poorly understood time in plant history. © 2008 Elsevier B.V. All rights reserved. 1. Introduction This paper is a continuation of the description of an unusually diverse and well preserved megaspore assemblage of mid Tournai- sian (Early Mississippian: earliest Carboniferous) age from north- eastern Ohio, USA (Arioli et al., 2007). The megaspore assemblage is of particular interest because dispersed megaspores from the early part of the Carboniferous (Tournaisian) are relatively poorly under- stood due to a paucity of described assemblages. Intriguingly, this gap in knowledge masks an important change in megaspore assemblages, with marked differences in morphological composi- tion and diversity between those from the Upper Devonian and later Carboniferous (Viséan-Stephanian) (e.g. Glasspool and Scott, 2005; Arioli et al., 2007). Upper Devonian megaspore assemblages are dominated by relatively small forms, many of which are charac- terised by grapnel-tipped processes (e.g. Allen and Robson, 1981; Candilier et al., 1982; Higgs and Scott, 1982). Later Carboniferous (Viséan-Stephanian) megaspore assemblages lack grapnel-tipped forms and are dominated by larger megaspores that are character- istic of the plants that dominated the Euramerican Coal Measure forests. Interestingly the mid Tournaisian assemblage from Ohio appears to comprise a mixture of forms more typical of the Upper Devonian and forms more typical of the later Carboniferous. A full taxonomic description of the assemblage is currently being prepared by Spinner and Wellman (in prep.). This report focuses on a conspicuous and common element of the assemblage: Lagen- icula (Triletes) mixta (Winslow 1962) comb. nov. L. mixta is an early example of a lageniculate megaspore (i.e. those with a well developed apical prominence/gula) so typical of the later Carboniferous. This taxon has a rather confused taxonomic history. It was originally described as a variety by Winslow (1962): Triletes catenulatus var. mixtus Winslow (1962). Later Dybova-Jachowicz et al. (1979) placed it in synonomy with Sublagenicula (Triletes) variabilis (Winslow) Dybova-Jachowicz et al. (1979) (see also Dybova-Jacho- wicz et al., 1987). Here we regard it as a species in its own right but place it in a new combination with the genus Lagenicula. In order to present a detailed description of its morphology, gross structure and wall ultrastructure we have conducted a thorough investigation using LM, SEM and TEM. The aim is to use this new information to compare this species of Lagenicula with earlier and later megaspores in order to determine its biological affinities and evolutionary relationships and, Review of Palaeobotany and Palynology 156 (2009) 51–61 ⁎ Corresponding author. E-mail address: c.wellman@sheffield.ac.uk (C.H. Wellman). 0034-6667/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.revpalbo.2008.07.008 Contents lists available at ScienceDirect Review of Palaeobotany and Palynology journal homepage: www.elsevier.com/locate/revpalbo
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Morphology and wall ultrastructure of the megaspore Lagenicula (Triletes) variabilis (Winslow, 1962)...

Review of Palaeobotany and Palynology 156 (2009) 51–61
Contents lists available at ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r.com/ locate / revpa lbo
Morphology and wall ultrastructure of the megaspore Lagenicula (Triletes) mixta(Winslow 1962) comb. nov. from the Carboniferous (Early Mississippian: midTournaisian) of Ohio, USA
Charles H. Wellman a,⁎, Caroline Arioli b, Edwin G. Spinner a, Marco Vecoli b
a Department of Animal and Plant Sciences, University of Sheffield, Alfred Denny Building, Western Bank, Sheffield S10 2TN, UKb “Géosystèmes”, UMR 8157 CNRS, Université de Lille 1, Sciences de la Terre, 59655 Villeneuve d'Ascq, France
⁎ Corresponding author.E-mail address: [email protected] (C.H. Wel
0034-6667/$ – see front matter © 2008 Elsevier B.V. Adoi:10.1016/j.revpalbo.2008.07.008
a b s t r a c t
a r t i c l e i n f oArticle history:
Megaspores assigned to Lag Received 14 December 2007Received in revised form 16 May 2008Accepted 24 July 2008Available online 3 August 2008Keywords:lycopsidmegasporeultrastructureCarboniferousEarly MississippianTournaisian
enicula (Triletes) mixta (Winslow, M., 1962. Plant Spores and Other Microfossilsfrom Upper Devonian and Lower Mississippian Rocks of Ohio. Geol. Surv., Prof. Paper 364, 1–93.) comb. nov.,from the Carboniferous (Early Mississippian: mid Tournaisian) of northeastern Ohio, USA, have beenanalysed using light microscopy (LM), scanning electron microscopy (SEM) and transmission electronmicroscopy (TEM). These studies provide new information on morphology, gross structure and wallultrastructure. This taxon has a confused taxonomic history, and the new morphological information allowsrecognition as a distinct species that can be placed with the genus Lagenicula as a new combination.Morphological/ultrastructural studies confirm the lycopsid affinities of this megaspore and it is suggestedthat it probably derived from an arborescent lycopsid that belonged with the Lepidocarpaceae. Thus it is anearly example of a megaspore derived from an arborescent lycopsid of the type that went on to dominate theEuramerican Coal Measure forests. The Ohio Tournaisian megaspore assemblage is surprisingly diverserevealing an interesting insight into vegetation ecology at this poorly understood time in plant history.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
This paper is a continuation of the description of an unusuallydiverse and well preserved megaspore assemblage of mid Tournai-sian (Early Mississippian: earliest Carboniferous) age from north-eastern Ohio, USA (Arioli et al., 2007). The megaspore assemblage isof particular interest because dispersed megaspores from the earlypart of the Carboniferous (Tournaisian) are relatively poorly under-stood due to a paucity of described assemblages. Intriguingly, thisgap in knowledge masks an important change in megasporeassemblages, with marked differences in morphological composi-tion and diversity between those from the Upper Devonian and laterCarboniferous (Viséan-Stephanian) (e.g. Glasspool and Scott, 2005;Arioli et al., 2007). Upper Devonian megaspore assemblages aredominated by relatively small forms, many of which are charac-terised by grapnel-tipped processes (e.g. Allen and Robson, 1981;Candilier et al., 1982; Higgs and Scott, 1982). Later Carboniferous(Viséan-Stephanian) megaspore assemblages lack grapnel-tipped
lman).
ll rights reserved.
forms and are dominated by larger megaspores that are character-istic of the plants that dominated the Euramerican Coal Measureforests. Interestingly the mid Tournaisian assemblage from Ohioappears to comprise a mixture of forms more typical of the UpperDevonian and forms more typical of the later Carboniferous. A fulltaxonomic description of the assemblage is currently beingprepared by Spinner and Wellman (in prep.). This report focuseson a conspicuous and common element of the assemblage: Lagen-icula (Triletes) mixta (Winslow 1962) comb. nov.
L. mixta is an early example of a lageniculate megaspore (i.e. thosewith a well developed apical prominence/gula) so typical of the laterCarboniferous. This taxon has a rather confused taxonomic history. Itwas originally described as a variety by Winslow (1962): Triletescatenulatus var.mixtusWinslow (1962). Later Dybova-Jachowicz et al.(1979) placed it in synonomy with Sublagenicula (Triletes) variabilis(Winslow) Dybova-Jachowicz et al. (1979) (see also Dybova-Jacho-wicz et al., 1987). Here we regard it as a species in its own right butplace it in a new combination with the genus Lagenicula. In order topresent a detailed description of its morphology, gross structure andwall ultrastructure we have conducted a thorough investigation usingLM, SEM and TEM. The aim is to use this new information to comparethis species of Lageniculawith earlier and later megaspores in order todetermine its biological affinities and evolutionary relationships and,

52 C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
hopefully in the process, shed some light on: (i) the nature of theTournaisian vegetation; (ii) the origins of the plants that went on todominate the Euramerican Coal Measure forests.
2. Material and methods
2.1. Locality and geology
Studied material consists of six samples collected from the upperpart of the Cuyahoga Formation (Wooster Member) and overlyinglower part of the Logan Formation (Byer Member) from two localitiesin Wayne County, northeastern Ohio, USA (Fig. 1). The location of thesite and the description of the samples are outlined by Clayton et al.(1998). These deposits are interpreted as accumulating in terrestrial tonearshore marine environments associated with an interdistributarysetting of a deltaic environment. Dispersed miospore assemblagesfrom all six samples belong with the Spelaeotriletes pretiosus–Raistrickia clavata (PC) Miospore Biozone of Western Europe, indicat-ing an early Mississippian age (which corresponds to the midTournaisian inWestern Europe). The geology of the site is summarisedby Coleman and Clayton (1987) and Clayton et al. (1998).
2.2. Preparation and techniques
Samples were macerated using standard HCl-HF-HCl palynologicaltechniques. The residues were sieved using a 120-μm mesh. Themegaspores are very well preserved with little, if any, corrosion andwith delicate spines preserved intact. Because of their low thermalmaturity (megaspores are translucent and yellow-brown in colour)oxidationwas not required. Megaspores were picked directly from theorganic residue using a fine paintbrush. All of the megaspores werefirst mounted on glass slides using glycerine for LM observation andphotography. Selected specimens were later recovered from the slidesand prepared for either SEM or TEM analysis.
For SEM analysis megaspores were mounted on SEM stubs usingdouble-sided sticky carbon tabs and gold coated using a sputter-coater.They were observed using a Philips 501B SEM at 30 kV (University ofSheffield). For TEManalysismegasporeswereplacedonblocks of freshlyprepared agar and sealed into the blocks by covering them in moltenagar which solidifies on cooling. Then themegaspores were dehydrated
Fig. 1. Location
in ethanol and embedded in Spurr resin. The sections made using amicrotome armed with a diamond knife and were stained with uranylacetate followed by Reynold's lead citrate. They were examined using aPhilips CM10 TEM at 80 kV (University of Sheffield). Note that all themegaspores selected for TEM analysis were preserved in lateralcompression and cut perpendicular to the plane of compression, sothat apparent dimensions approximate true thicknesses.
Twenty four specimens of L. mixta were identified and mountedfor examination using LM. Three of these specimens were recoveredfrom LM slides and mounted for SEM analysis (SEM stub CW140a–c).Another three specimens were recovered from LM slides andembedded in individual blocks for TEM analysis (TEM blocks CW086to CW088).
3. Systematic palaeontology
3.1. Terminology
Terminology utilized in descriptions is from Grebe (1971) with theexception of the use of the terms gula and apical prominence (sensuSpinner 1969, 1983) and catenulate (sensuWinslow 1962) (see below).
3.2. Repository of material
All material (sample, residue, slides, stubs, blocks and sections) isstored in the Centre for Palynology of the University of Sheffield.
3.3. Taxonomy
Anteturma SPORITES Potonié, 1893Turma TRILETES (Reinsch 1881) Potonié and Kremp, 1954Subturma LAGENOTRILETES Potonié and Kremp, 1954 emend.
Bhardwaj, 1957Infraturma GULATI Bhardwaj, 1957Genus LAGENICULA (Bennie and Kidston, 1886) Potonié and
Kremp, 1954 emend. Spinner 1969Type species. Lagenicula horrida Zerndt 1934Lagenicula (Triletes) mixta (Winslow 1962) comb. nov.
Triletes catenulatus var. mixtus Winslow 1962, p.33, plate 7, 4
of sections.

Plate I. LM images of Lagenicula mixta. Scale bar=500 µm except for Fig. 6 where scale bar=350 µm.
1. Specimen LM Slide 17. Lateral compression. Specimen removed and remounted as TEM block CW87 for TEM analysis (illustrated in Plate IV).
2. Specimen LM Slide 15. Lateral compression. Specimen removed and remounted as TEM block CW86 for TEM analysis (illustrated in Plate V).
3. Specimen LM Slide 88. Lateral compression.
4. Specimen LM Slide158. Lateral compression. Specimen remounted for SEM analysis as SEM Stub CW140/3 (illustrated in Plate II).
5. Specimen LM Slide 90. Lateral compression.
6. Specimen LM Slide 87. Lateral compression. Specimen remounted for SEM analysis as SEM Stub CW140/1 (illustrated in Plate III).
7. Specimen LM Slide 84. Polar compression.
8. Specimen LM Slide 86b. Lateral compression.
9. Specimen LM Slide 86a. Lateral compression.
53C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61

54 C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
3.3.1. Holotype and type localityTriletes catenulatus var. mixtus Winslow 1962, plate 7, 2, from
cuttings of the Cuyahoga and Black Hand formations (undifferen-tiated), locality 7, maceration 366, slide 2 (R. J. Dalier No. 3 well, R. J.Dallier farm, 1.4 miles southeast of Newcastle, Newcastle Township,Brinkhaven quadrangle, Coshocton County, Ohio, USA) (see Winlow1962, p. 17–19).
3.3.2. RemarksWinslow (1962) described a new species of megaspore: Triletes
catenulatusWinslow 1962with three new varieties: the type variety T.catenulatus var. catenulatus Winslow 1962, T. catenulatus var. margi-natus Winslow 1962 and T. catenulatus var. mixtus Winslow 1962.Winslow allowed very broad morphographic limits in this species,with the three varieties including taxa with different types of apicalprominence (gula) and distal ornamentation of the megaspore body.In all three of her formal named varieties she referred to the presence,to a certain degree, of what she called catenulate ornamentation (seeWinslow 1962). This character/feature was apparently the commonlink between specimens. In this study we were able to recogniseWinslow's three formal varieties, but regard them as being worthy ofspecific rather than varietal taxonomic status, based on evidence ofdifferences in structure, apical prominence (gula) and type ofornament (see below). Furthermore, we consider that all three taxaare better placed with the genus Lagenicula (Bennie and Kidston,1886) Potonié and Kremp (1954) emend. Spinner 1969. Thus Triletescatenulatus var. mixtus Winslow 1962 is upgraded to specific statusand formally transferred to Lagenicula as Lagenicula (Triletes) mixta(Winslow 1962) comb. nov.
Winslow's type variety of Triletes catenulatus (T. catenulatus var.catenulatus) is clearly recognisable by the lageniculate apicalprominence (gula) without any basal constriction (in contrast toLagenicula mixta as described below) and a definite reticulateornament with short pointed spines projecting largely from themurate areas of the reticulum and most clearly seen where theyproject at the megaspore margin. Winslow distinguished varietymarginatus from her type variety (var. catenulatus) on the basis ofsmaller body size, more prominent flange, larger contact areas,shorter spines and less distinct reticulate ornament on the distalsurface. Winslow distinguished varietymixtus on the basis of the lackof a ‘subdued lageniculate type’ of apical prominence (gula) and lessprominent presence of what she described as catenulate ornamenton the distal surface of themegaspore body. However, we believe thatthe preferred lateral compression, the shape of the prominent,basally constricted apical prominence (gula) ornamented with cones(‘conical species’ of Winslow) particularly prominent alongside thetraces of the sutures, together with a discrete baculate to spinoseornament on the distal surface is distinctive and warrant recognitionat specific rather than varietal level. At the margin of the megasporebody the ornamented elements are discrete and generally constant inlength. Some specimens were observed where there was someindication of the ‘catenulation’ as described byWinslow but this maypossibly reflect compression during fossilization.
Dybova-Jachowicz et al. (1979, 1987) regenerated this taxon asa synonym of their Sublagenicula (Triletes) variabilis (Winslow)
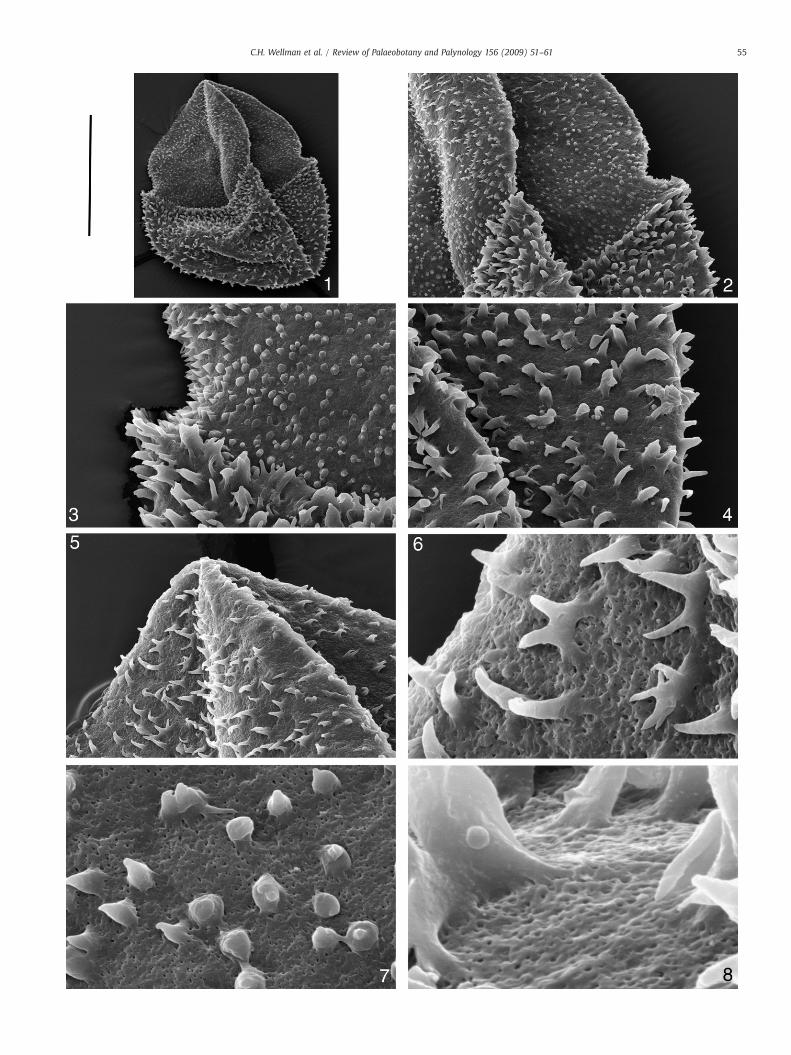
Plate II. SEM images of Lagenicula mixta. SEM stub CW140 (specimen CW140a removed fro
1. Entire specimen. Scale bar=300 μm.
2. Close up of curvaturae at junction between proximal (gula) and distal surface
3. Close up of curvaturae at junction between proximal (gula) and distal surface
4. Close up of ornament on distal surface. Scale bar=65 μm.
5. Close up of ornament on proximal surface towards the top of the gula. Scale b
6. Detailed magnification of ornament on the proximal surface towards the top o
7. Detailed magnification of ornament on the proximal surface towards the base
8. Detailed magnification of ornament on the distal surface towards the top of th
Dybova-Jachowicz 1979 [now Lagenicula (Triletes) variabilis (Win-slow) Arioli et al., 2004] which is rejected here. Although L. variabilishas a similar apical prominence (gula), the ornamentation isdifferent (see Winslow 1962; Dybova-Jachowicz et al., 1987; Arioliet al., 2007).
We consider that Triletes catenulatus var. mixtus Winslow 1962 isbetter placed with the genus Lagenicula because: (i) Triletes has anextremely broad remit and has subsequently been subdivided;(ii) T. catenulatus var. mixtus has a prominent apical thickening/gulaand clearly falls within the remit of the genus Lagenicula ascurrently utilised by certain authors (e.g. Spinner, 1969, 1983; Arioliet al., 2004, 2007). Unfortunately the taxonomic history of the genusLagenicula is highly complex. We prefer to use the genus Lageniculafor gulate megaspores as outlined by Spinner (1969, 1983) (seedetailed discussion in Arioli et al., 2007), although we accept thatfurther work is required to clarify the taxonomy of lageniculatemegaspores.
4. Description of Ohio Lagenicula mixta
Lagenicula (Triletes) mixta (Winslow, 1962) comb. nov.
Triletes catenulatus var. mixtus Winslow (1962), p.33, plate 7, 4Occurrence: Upper Cuyahoga and lower Logan formations.Remarks: Specimens found during the present study agree closely
with Winslow's description in range of overall body size, shape andmorphology of the apical prominence (gula), distal ornamentationand preferred lateral compression direction (perpendicular to thepolar axis). Detailed descriptions are provided below.
4.1. LM description (Plate I)
The 24 specimens examined using LM consist of an originallyspherical body with a well developed apical prominence (gula). Theyare usually compressed laterally or obliquely, but two specimens werepreserved in polar compression (e.g. Plate I, 7). In laterally compressedspecimens the overall length (including apical prominence (gula))ranges from 561(808)1001 μm and maximumwidth of the main bodyranges from 451(648)858 μm.
The gula is basally constricted (e.g. Plate I, 6) and ranges from 289(428)550 μm in height and 385(480)583 μm in width. Occasionallythe gula is ruptured displaying its constituent valves (e.g. Plate I, 1).The gula bears a dense ornament of apiculate elements up to 10 μmtall. These are best observed using SEM and their form and dimensionsare described in the following SEM description.
The contact areas are best observed in rare specimens preserved inpolar compression, where they can be seen to occupy about 3/4 of theproximal surface (e.g. Plate I, 7). The curvaturae perfectae are usuallydistinct, marked by the junction between the larger apiculateelements of the distal surface and the smaller apiculate elements ofthe contact areas and apical prominence (gula).
Outside the contact areas the megaspore body has a denseornament of larger apiculate (baculate to spinose) elements up to30 μm tall. These are best observed using SEM and their form anddimensions are described in the following SEM description.
m LM Slide 87 as illustrated in Plate I, Fig. 6).
. Scale bar=130 μm.
. Scale bar=65 μm.
ar=65 μm.
f the gula. Note the pitting in the wall between ornament elements. Scale bar=16 μm.
of the gula. Note the pitting in the wall between ornament elements. Scale bar=16 μm.
e gula. Note the pitting in the wall between ornament elements. Scale bar=8 μm.
55C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61

56 C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
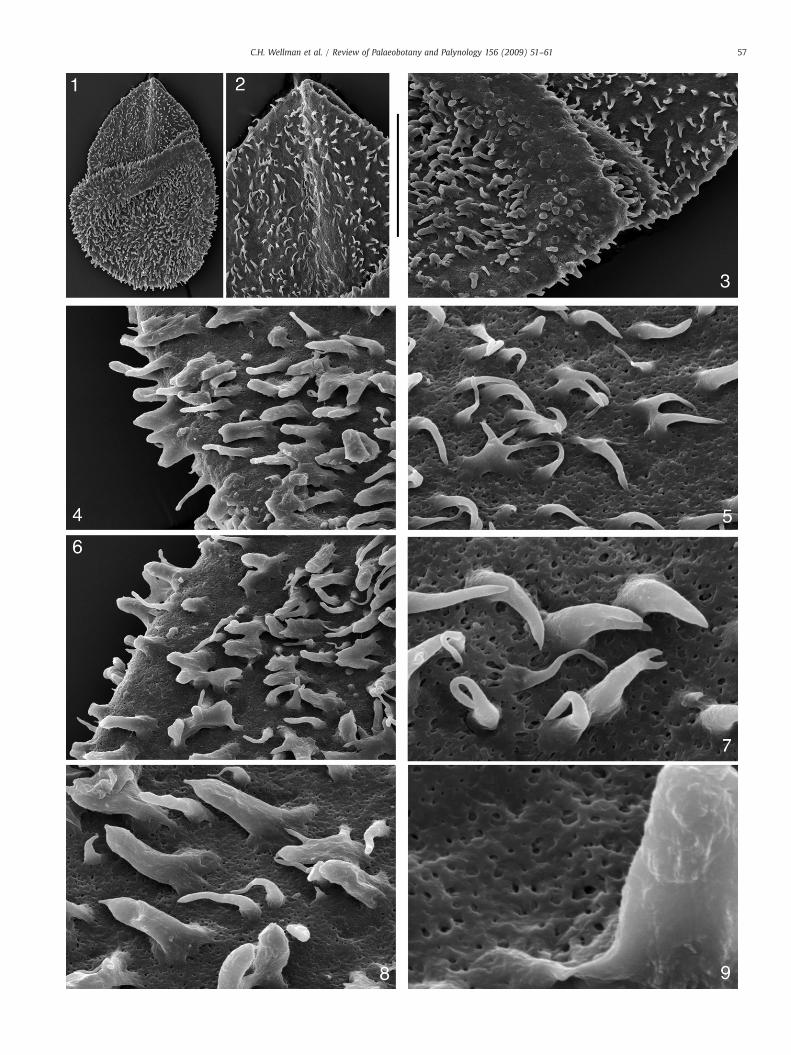
4.2. SEM description (Plates II and III)
SEM images reveal the gross structure of the megaspore with thelarge apical prominence (gula) arising from the megaspore body.Curvaturae are distinct and marked by an abrupt transition from thelarger sculptural elements of the megaspore body and the smallersculptural elements of the contact areas and apical prominence(gula) (Plate II, 1–3). The apical prominence (gula) has a constrictedbase, clearly discernible in the specimen illustrated in Plate II(Plate II, 1–3), but not in others such as the specimen illustrated inPlate III where the base of the apical prominence (gula) and contactareas have collapsed into the megaspore body during compression(Plate III, 1–3).
The contact areas and apical prominence (gula) have a denseornament of apiculate elements, up to 10 μm tall, that are variablein terms of size and shape. They consist predominantly of simplecones and simple spines, but also biform elements, spines that bi-and tri-furcate, and elements that have fused together to form anH-shape. In the specimen illustrated in Plate II it is clear that thereis a general gradation from cones to spines towards the top of thegula (Plate II, 2).
The megaspore body has a dense ornament of larger elements upto 30 μm tall. These are generally discrete, but occasionally combine toform areas of catenulate ornament (sensu Winslow 1962). They arevariable in terms of size and form. They are predominantly baculate tospinose, and often are biform. They sometimes bi- or tri-furcate, andsometimes fuse to form H-shaped structures.
Between the ornament elements thewall is abundantly perforated.Perforations are reasonably regular in size and fairly evenly dis-tributed (Plate II, 6–8; Plate III, 5,7–9). They are similar in terms ofsize, shape and distribution over the entire megaspore. The ornamentelements are entirely smooth, except near the base where sometimesthere are individual strands arising from the perforated wall thatcombine to form the smooth elements (Plate II, 6–8; Plate III, 4–9).This is the buttressed appearance typical of the genus Lagenicula asdiscussed by Glasspool et al. (2000).
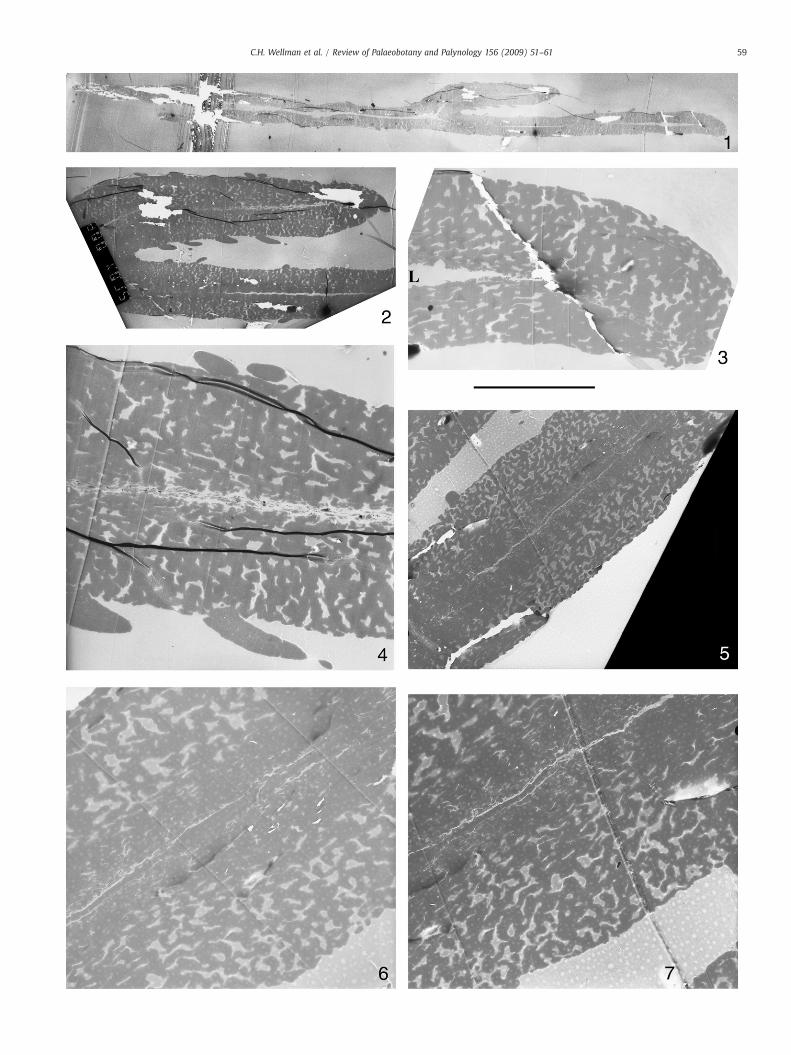
4.3. TEM description (Plates IV and V)
The exospore is similar in all parts of the megaspore [i.e.megaspore body, contact areas and apical prominence (gula)]. It isbetween ca. 50 to 100 μm in width. Two very different layers can bedistinguished in this wall: a thick, spongy outer layer (outer exospore)and a very thin inner layer (inner exospore) also referred to as thebasal lamina or basal layer (Plate IV, 4–7; Plate V, 3–4).
The outer exospore constitutes almost the entire wall thickness. Itconsists of elongate elements that are anastomosing and convolutedforming a spongy layer with many voids (Plate IV, 3–7; Plate V, 1–4).The elements are highly variable in width measuring between 2.5–10 μm. The outer exospore is more compact towards the inside of themegaspore with voids narrower due to closer compaction of thesporopollenin framework. Many of the voids towards the outside ofthe megaspore wall reach the surface (and presumably are respon-sible for the surface perforations observed using SEM).
Plate III. SEM images of Lagenicula mixta. SEM stub CW140 (specimen CW140c removed fr
1. Entire specimen. Scale bar=500 μm.
2. Close up of ornament on proximal surface (gula). Scale bar=200 μm.
3. Close up of junction between proximal (gula) and distal surface. Scale bar=1
4. Close up of ornament on distal surface. Scale bar=65 μm.
5. Close up of ornament on proximal (gula) surface. Scale bar=30 μm.
6. Close up of ornament on distal surface. Scale bar=65 μm.
7. Close up of ornament on proximal (gula) surface. Note the pitting in the wall
8. Close up of ornament on distal surface. Note the pitting in the wall between o
9. Close up of ornament on distal surface. Note the pitting in the wall between o
Sculptural elements are solid consisting of homogeneous spor-opollenin (Plate IV, 2,4; Plate V, 1,2,4). At their base, which coincideswith the megaspore surface, they can be seen to be formed byconvergence of the sporopollenin elements (strands) that comprisethe spongy outer layer of the megaspore wall. It is this feature thatgives the base of the ornament elements their classic buttressedappearance (see p. 8).
The inner layer of the exospore is thin (ca. 2 μm) and consists of aseries of parallel arranged, concentric laminae. Occasionally when amegaspore is not entirely compressed, and the lumen is representedby a gap, the inner layer can be seen to have come loose (e.g. Plate V,3). The junction between the inner laminate layer and the outerspongy layer is difficult to discern, because the spongy layer is morecompressed towards the inside of the megaspore and the elementscomprising it become more aligned to the lamina (e.g. Plate IV, 6–7;Plate V, 3–4).
5. Discussion: affinities and phylogenetic relationships
The population of L. mixtamost likely comprises of predominantly,if not exclusively, freely dispersed mature megaspores. Thus ontoge-netic information is unavailable. It is clear from the descriptions abovethat the population of L. mixta under investigation is very similar interms of morphology, gross structure and wall ultrastructure. There issome variability (e.g. overall dimensions and nature of the ornament),but this almost certainly can be accommodated by the range ofvariation within the species (i.e. intraspecific variation), as has beendescribed in other species of Lagenicula.
Wall ultrastructure has been described in a number of lagenicu-late megaspores (dispersed and in situ forms) (summarized inTaylor, 1990; Glasspool et al., 2000 and Arioli et al., 2007). Thespecimens of L. mixta described herein have wall ultrastructurecomparable with that of most other investigated taxa assigned toLagenicula, and indeed most lageniculate megaspores, in whichthis information is known. That is a narrow, laminate, innerexospore (consisting of either a single basal lamina or multiplelaminae) overlain by a wide, spongy, outer exospore composed ofsmall globular to elongate or laminar units. Only minor differencesoccur between species relating to the number of laminae in theinner layer and orientation, density, size and degree of fusion of theunits in the outer layer. However, a note of caution is advised whenmaking such comparisons, because taphonomic influences (e.g.compression during diagenesis) may alter the appearance of thewall by changing the orientation of the sporopollenin units and thesize, shape and orientation of the voids.
It is clear that wall ultrastructure in the Lower Carboniferous(Tournaisian) L. mixta described herein is similar to that inpreviously described lycopsid lageniculate megaspores fromthroughout the Carboniferous. This supports interpretation of theTournaisian forms as phylogenetically related to later Carboniferousforms. However, it has been noted that wall ultrastructure of La-genicula from the earlier and later Carboniferous differ in two minorrespects: (i) in later forms the spongy outer exospore is denserconsisting of larger sporopollenin units with smaller voids in
om LM Slide 158 as illustrated in Plate I, Fig. 4).
30 μm.
between ornament elements. Scale bar=15 μm.
rnament elements. Scale bar=30 μm.
rnament elements. Scale bar=7.5 μm.
57C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61

58 C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
between (Hemsley et al., 1994); (ii) in earlier forms the laminateinner exospore consists of multiple laminae but this is reduced to asingle lamina in later forms (Scott and Hemsley, 1996). L. mixta fromOhio concurs with both of these observations in that the outer layeris not particularly dense and the inner layer consists of multiplelaminae.
Arioli et al. (2007) provided a detailed discussion of thebiological affinities and phylogenetic relationships of lageniculatemegaspores with respect to Lagenicula variabilis. Exactly the samearguments can be repeated for L. mixta. Studies of in situmegaspores suggest that Lagenicula and related genera from theCarboniferous derive predominantly, possibly exclusively, from theLepidocarpaceae (arborescent lycopsids) (e.g. Balme, 1995). All havevery similar ultrastructure (excepting the minor differences notedabove). Unfortunately megaspores of other lycopsids also have verysimilar wall ultrastructure [e.g. non-lageniculate forms (Glasspoolet al., 2000) and those from Selaginellaceae (Cottnam et al., 2000)].Are the plants that produced lageniculate megaspores phylogeneti-cally closely related or did such megaspores evolve independentlythrough convergence in a number of unrelated plant groups(possibly due to similar ecological demands related to reproductionand/or dispersal)?
Bateman and DiMichele (1994) proposed that heterospory evolvedonly once in the lycopsids in the Selaginellales+Rhizomorphales(lepidodendraleans+isoetaleans) clade (excluding the independentorigin in the barinophytaleans, a derived member of the paraphyleticzosterophylls that probably included the ancestors of the lycopsids). Itis interesting to consider the extent to which megaspore wallultrastructure differs within this clade, and thus if this character isof value in determining biological affinities of the different mega-spore-producers. Arioli et al. (2007) considered this issue andconcurred that heterospory evolved only once and suggested thatthis heterosporous group adopted an ancestral mode of megasporewall formation (and hence wall ultrastructure) from an ancestralhomosporous lycopsid group. Wellman (2002) described wall ultra-structure in Middle Devonian spores with grapnel-tipped processes.These relatively large spores have lycopsid affinities, although it isuncertain if they were truly heterosporous or exhibited incipientheterospory. Nonetheless, they exhibit wall ultrastructure similar tothat in later lycopsid megaspores, and it would appear that themonophyletic heterosporous lycopsids inherited a mode of mega-spore wall formation that evolved through simple modification of thebasic developmental processes of megaspore wall formation in anancestral group of homosporous lycopsids. This basic mode ofmegaspore wall formation persisted relatively unchanged within theheterosporous lycopsids, accounting for the lack of diversity inmegaspore wall ultrastructure in subsequent forms, including livingspecies whose wall ultrastructure and development is particularlywell understood [summarized in Lugardon (1990) and Tryon andLugardon (1991)].
6. Discussion: palaeobotanical considerations
Arioli et al. (2007) discussed the significance of the pronouncedchange in the nature of megaspore assemblages in the Late Devonian–
Plate IV. TEM images of Lagenicula mixta. TEM block CW87 (specimen removed from LM Slicaused by degradation of the coating of the grid.
1. Section through entire megaspore. Scale bar=100 μm.
2. Close up of folded central portion. Scale bar=37.5 μm.
3. Close up of right hand end. Scale bar=14 μm. L=lumen.
4. Close up of Fig. 2. Note the solid spines and spongy wall. Scale bar=10 μm.
5. Close up of wall. Note how the spongy wall ultrastructure becomes denser tow
6. Close up of wall. Note the multiple laminae lining the lumen. Scale bar=6.5 μ
7. Close up of wall. Note the multiple laminae lining the lumen. Scale bar=6.5 μ
Early Carboniferous. They suggested that it reflected importantchanges (evolutionary and ecological) in the vegetation, with theincreasing abundance and diversity of lageniculate forms indicating arise in the importance of forests of arborescent lycopsids belonging tothe Lepidocarpaceae (see also Glasspool and Scott, 2005). The largediversity in the Ohio megaspore assemblage of lageniculate formswith typical lycopsid wall ultrastructure indicates that these forestscomprised a high diversity of species and were of complex ecology.
It is interesting to reflect on why there is such morphologicaldiversity (disparity) among the lageniculate megaspores whilst thewall ultrastructure remains so conserved. Clearly the walls areconstructed in essentially the same way (through similar ontogenyand processes). However, this conserved and widespread method ofconstruction can clearly achieve highly variable morphology. The nextstep is to consider this morphological variation in light of its functionand how it was shaped through natural selection.
It is generally considered that the Lepidocarpaceae formed denseforests of closely spaced trees. This is largely based on evidence fromlater Coal Measures forests (e.g. DiMichele et al., 2007), but findings inthe Early Carboniferous suggest that the forests of this time weresimilar. For example, lycopsid forests preserved as stump horizons ingrowth position have been reported from the Tournaisian of easternCanada by Falcon-Lang (2004) (predominantly Protostigmaria–Lepi-dodendropsis) and Glasspool and Scott (2005) (predominantly Lepi-dodendropsis). These reports suggest dense forests of arborescentlycopsids. Based on consideration of arborescent lycopsids from theCoal Measures (e.g. Phillips and DiMichele, 1992) it is considered thatthe arborescent lycopsids were quick growing and short lived. Juvenileplants on the forest floor developed as an extensive stigmarian systemwith the ‘trunk’ little developed. Once the stigmarian system wassufficiently elaborated the ‘trunk’ would have elongated (probablyrapidly) as an unbranched pole. Pole growth was determinate andprobably dependent on reaching canopy height. Only then would asignificant crown form as lateral branches developed and reproduc-tive structures appeared. This mature stage of the cycle may have beenrather short-lived, possibly with death of the plant ensuing rapidlyafter spore dispersal. Such a lifecycle has interesting implications forthe free-sporing heterosporous reproductive strategies adopted bythese trees. Presumably megaspores would have been produced inlarge quantities once the tree reached maturity and the crown andassociated fertile structures appeared. However, these would not havebeen transported huge distances on release. Their size suggests thatthey would have fallen directly to the ground and settled below theparent tree, with lateral transport occurring only aerially at times ofstrong winds or by overland flow on the forest floor during time offlooding (see discussion of megaspore dispersal in Hemsley et al.,1999). Once settled the megaspores would have lodged and couldremain dormant for extended periods due to their size and hencecapacity to store nutrition. Fertilisation would have been straightfor-ward due to the immense abundance and impressive mobility of themicrospores.
Thediversemorphologyof themegaspores presumably relates to therather simplistic life cycle/reproductive strategy outlined above. Mega-spore functional morphology has been discussed in detail by Hemsleyet al. (1999). It probably relates to a combination of factors involved in
de 17 as illustrated in Plate I, 1). Figs. 5–7 have a mottled appearance that is an artefact
ards the lumen. Scale bar=12 μm.
m.
m.
59C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
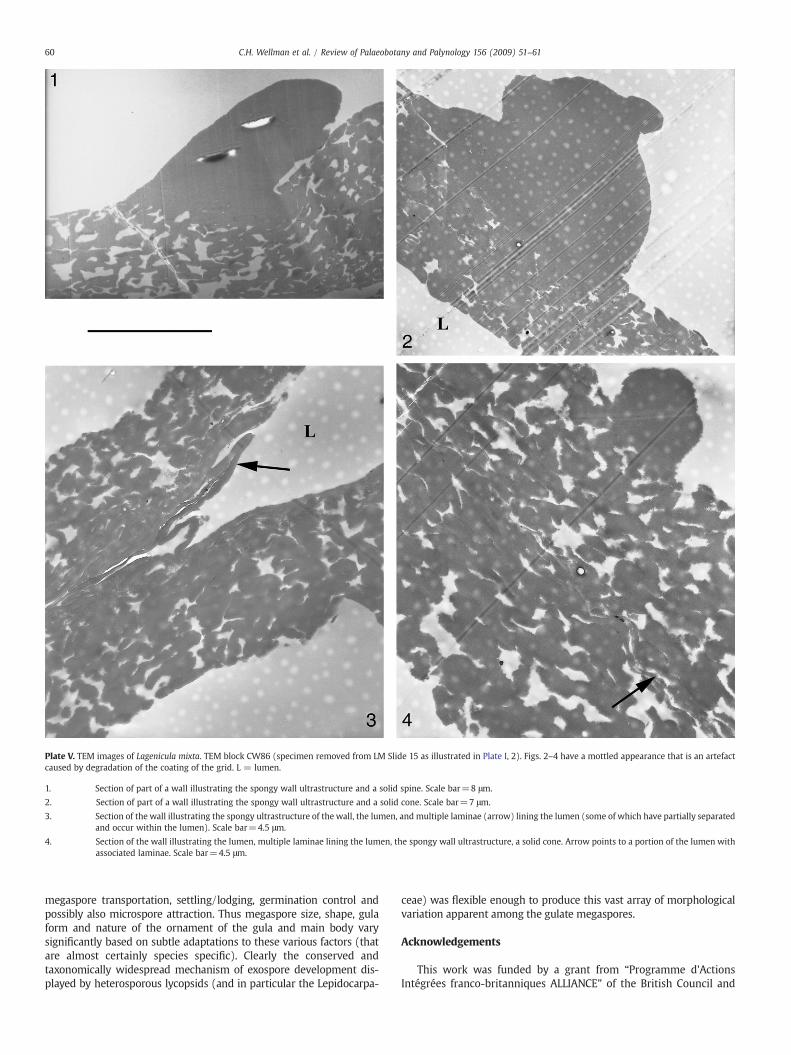
Plate V. TEM images of Lagenicula mixta. TEM block CW86 (specimen removed from LM Slide 15 as illustrated in Plate I, 2). Figs. 2–4 have a mottled appearance that is an artefactcaused by degradation of the coating of the grid. L = lumen.
1. Section of part of a wall illustrating the spongy wall ultrastructure and a solid spine. Scale bar=8 μm.
2. Section of part of a wall illustrating the spongy wall ultrastructure and a solid cone. Scale bar=7 μm.
3. Section of the wall illustrating the spongy ultrastructure of the wall, the lumen, and multiple laminae (arrow) lining the lumen (some of which have partially separatedand occur within the lumen). Scale bar=4.5 μm.
4. Section of the wall illustrating the lumen, multiple laminae lining the lumen, the spongy wall ultrastructure, a solid cone. Arrow points to a portion of the lumen withassociated laminae. Scale bar=4.5 μm.
60 C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
megaspore transportation, settling/lodging, germination control andpossibly also microspore attraction. Thus megaspore size, shape, gulaform and nature of the ornament of the gula and main body varysignificantly based on subtle adaptations to these various factors (thatare almost certainly species specific). Clearly the conserved andtaxonomically widespread mechanism of exospore development dis-played by heterosporous lycopsids (and in particular the Lepidocarpa-
ceae) was flexible enough to produce this vast array of morphologicalvariation apparent among the gulate megaspores.
Acknowledgements
This work was funded by a grant from “Programme d'ActionsIntégrées franco-britanniques ALLIANCE” of the British Council and

61C.H. Wellman et al. / Review of Palaeobotany and Palynology 156 (2009) 51–61
Ministère des Affaires Étrangères. CA's Ph.D. research is funded by anEcole Doctorale SMRE of the University of Lille. CHW's research isfunded by NERC research grants. This study is a contribution to theproject ECLIPSE II: The Terrestrialization process: modelling complexinteractions at the geosphere–biosphere interface. We dedicate thispaper to the fondmemory of Bernard Lugardonwho collaborated withus during an earlier phase of this project and whose kind advisehelped us all gain a better understanding of spore wall ultrastructure.
References
Allen, K.C., Robson, J., 1981. Megaspores with multifurcate and bifurcate processes fromOld Red Sandstone facies of Tournaisian age, from the Taff Gorge, South Glamorgan,Wales. New Phytol. 88, 387–398.
Arioli, C., Servais, T., Wellman, C.H., 2004. Morphology and ultrastructure of a LowerCarboniferous megaspore: Lagenicula variabilis (Winslow 1962) nov. comb. Ann.Soc. Géol. Nord 11, 109–111.
Arioli, C., Wellman, C.H., Lugardon, B., Servais, T., 2007. Morphology and wallultrastructure of the megaspore Lagenicula (Triletes) variabilis (Winslow 1962)Arioli et al. (2004) from the Lower Carboniferous of Ohio, USA. Rev. Palaeobot.Palynol. 144, 231–248.
Balme, B.E., 1995. Fossil in situ spores and pollen grain: an annotated catalogue. Rev.Palaeobot. Palynol. 87, 81–323.
Bateman, R.M., DiMichele, W.A., 1994. Heterospory: the most iterative key innovation inthe evolutionary history of the plant kingdom. Biol. Rev. 69, 345–417.
Bennie, I., Kidston, R., 1886. On the occurrence of spores in the Carboniferous Formationof Scotland. Proc. R. Soc. Edinb. 9, 82–117.
Bhardwaj, C.D., 1957. The spore flora of Velener Schichten (LowerWestphalian D) in theRuhrcoal measures. Palaeontogr., Abt. B 102, 110–138.
Candilier, A.M., Coquel, R., Loboziak, S., 1982. Mégaspores du Dévonien terminal et duCarbonifère inférieur des bassins d'Illizi (Sahara algérien) et de Rhadamès (Libyeoccidentale). Palaeontographica B 183, 83–107.
Clayton, G., Manger, W.L., Owens, B., 1998. Mississippian (Lower Carboniferous)miospores from the Cuyahoga and Logan formations of northeastern Ohio, USA.J. Micropalaeontol. 17, 183–191.
Coleman, U., Clayton, G., 1987. Palynostratigraphy and palynofacies of the UppermostDevonian and Lower Mississippian of Eastern Kentucky (U.S.A.), and correlationwith Western Europe. Cour. Forsch.-Inst. Senckenberg 98, 75–93.
Cottnam, C.F., Hemsley, A.R., Rössler, R., Collinson, M.E., Brain, A.P.R., 2000. Diversity ofexine structure in Upper Carboniferous (Westphalian) selaginellalean megaspores.Rev. Palaeobot. Palynol. 109, 33–44.
DiMichele, W.A., Falcon-Lang, H.J., Nelson, W.J., Brick, S.D., Ames, P.R., 2007. Ecologicalgradients within a Pennsylvanian mire forest. Geology 35, 415–418.
Dybova-Jachowicz, S., Jachowicz, A., Karczewska, J., Lachkar, G., Loboziak, S., Piérart, P.,Turnau, E., Zoldani, Z., 1979. Note préliminaire sur la révision des mégaspores à guladu Carbonifère. Les principes de la classification. Acta Palaeont. Pol. 24, 411–422.
Dybova-Jachowicz, S., Jachowicz, A., Karczewska, J., Lachkar, G., Loboziak, S., Piérart, P.,Turnau, E., Zoldani, Z., 1987. Revision of Carboniferous megaspores with gula(Part three). Pr. Inst. Geol. 121, 1–49.
Falcon-Lang, H.J., 2004. Early Mississipian lycopsid forests in a delta-plain setting atNorton, near Sussex, New Brunswick, Canada. J. Geol. Soc. 161, 969–981.
Glasspool, I.J., Scott, A.C., 2005. An Early Carboniferous (Mississippian), Tournaisian,megaspore assemblage from ThreeMile Plains, Nova Scotia, Canada. Rev. Palaeobot.Palynol. 134, 219–236.
Glasspool, I.J., Hemsley, A.R., Scott, A.C., Golitsyn, A., 2000. Ultrastructure and affinity ofLower Carboniferous megaspores from the Moscow Basin, Russia. Rev. Palaeobot.Palynol. 109, 1–31.
Grebe, H., 1971. A recommended terminology and descriptive method for spores. In:Alpern, B., Neves, R. (Eds.), Microfossiles organiques du Paléozoique. CIMP specialvolume 4, 87 pp.
Hemsley, A.R., Clayton, G., Galtier, J., 1994. Further studies on a late Tournaisian (LowerCarboniferous) flora from Loch Humphrey Burn, Scotland: spore taxonomy andultrastructure. Rev. Palaeobot. Palynol. 81, 213–231.
Hemsley, A.R., Scott, A.C., Collinson, M.E., 1999. The architecture and functional biologyof freely dispersed megaspores. In: Hemsley, A.R., Kurmann, M. (Eds.), PlantArchitecture. Royal Botanic Gardens, Kew, London, UK, pp. 253–277.
Higgs, K., Scott, A.C., 1982. Megaspores from the Uppermost Devonian (Strunian) ofHook Head, County Wexford, Ireland. Palaeontographica B 181, 79–108.
Lugardon, B., 1990. Pteridophyte sporogenesis: a survey of spore wall ontogeny and finestructure in a polyphyletic plant group. In: Blackmore, S., Knox, R.B. (Eds.),Microspores: Evolution and Ontogeny. Academic Press, London, pp. 95–120.
Phillips, T.L., DiMichele, W.A., 1992. Comparative ecology and life-history biology ofarborescent lycopsids in Late Carboniferous swamps of Euramerica. Ann. Mo. Bot.Gard. 79, 560–588.
Potonié, R., 1893. Die Flora des Rotliegenden von Thüringen. Kgl. Preuss. Geol. L.-A.,vol. 9, pp. 1–298.
Potonié, R., Kremp, G., 1954. Die Gattungen der Paläozoischen Sporae dispersae und ihreStratigraphie. Geol. Jb 69, 111–194.
Scott, A.C., Hemsley, A.R., 1996. Chapter 18G. Palaeozoic megaspores. In: Jansonius, J.,McGregor, D.C. (Eds.), Palynology: principles and applications, vol. 2. AmericanAssociation of Stratigraphic Palynologists Foundation, pp. 629–639.
Spinner, E., 1969. Megaspore assemblages from Viséan deposits at Dunbar, East Lothian,Scotland. Palaeontology 12, 441–458.
Spinner, E., 1983. Two new species of Lagenicula from the Upper Carboniferous(Westphalian B) of North Derbyshire, England. Pollen Spores 25, 243–251.
Taylor, W.A., 1990. Comparative analysis of megaspore ultrastructure in PennsylvanianLycophytes. Rev. Palaeobot. Palynol. 62, 65–78.
Tryon, A.F., Lugardon, B., 1991. Spores of the Pteridophyta. Surface, wall structure anddiversity based on Electron Microscope Studies. Springer-Verlag, New-York, p. 648.
Wellman, C.H., 2002. Morphology and wall ultrastructure in Devonian spores withbifurcate-tipped processes. Int. J. Plant Sci. 163, 451–474.
Winslow, M., 1962. Plant Spores and Other Microfossils from Upper Devonian andLower Mississippian Rocks of Ohio. Geol. Surv., Prof. Pap. 364, 1–93.
Zerndt, J., 1934. Les mégaspores du Bassin Houiller Polonais. 1ère partie. Les couchesanticlinales. Acad. Pol. Sc. Lett. Trav. Géol. 1, 1–55.




















