
Fine structure of the fat body and nephrocytes in the life-stages of Dermacentor variabilis
18
. I Experimental & Applied Acarology, 8 ( 1990) 125-142 125 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands Fine Structure of the Fat Body and N ephrocytes in the Life-Stages of Dermacentor variabilis LEWIS B.COONS, WILLIAM J . L'AMOREAUX, ROSEMARIE ROSELL-DAVIS and LINDA STARR-SPIRES De partment of Biology, Memphis State University, Memphis, TN 38152 (U.S.A.) (Accepted 27 October 1988) ABSTRACT Coons, L.B ., L'amoreaux, W.J ., Rosell-Davis, R. and Starr-Spires, L., 1990. Fine structure of the fat body and nephocrytes in the life stages of Dermace ntor variabilis. Exp. Appl. Acarol., 8: 125- 142 The fine structure of the fat body and associated nephrocytes of the American dog tick , Der- macentor variabilis (Say), was described in unfed larvae, unfed nymphs, and in unfed and fed adults of both sexes. The fat body consisted of one type of cell, the trophocyte. Morphological changes that occured in the trophocytes of both sexes were dependent on feeding. The ultra- structure of feeding male trophocytes was distinct from trophocytes of feeding females. In the feeding female , the trophocyte developed an ultrastructure characteristic of cells that produce secretory proteins. A type of scalariform cell junction was found associated with rough endo- plasmic reticulum of the trophocytes. Nephrocytes were closely associated with trophocytes but were not part of the fat body. Nephrocyte ultrastructure was unaltered throughout the life-stages we examined, except at the end of oviposition. Organelles in the nephrocytes were not randomly dist ributed, but were found in distinct regions of the cytoplasm. Slit diaphragms at the surface of the nephrocytes were extracellular specializations that had a periodic ultrastructure. INTRODUCTION In the American dog tick, Dermacentor variabilis (Say), a fat body is present in each life-stage: larva, nymph, and adult (Obenchain and Oliver, 1973). The fat body is located just inside the epidermal layer of cells and between the organs. These two regions are known respectively as the peripheral fat body and the central fat body (Obenchain and Oliver, 1973). In females, the fat body, especially the peripheral fat body, proliferates during and after feeding (Obenchain and Oliver, 1973 ). In the hard ticks Rhipicephalus sanguinius and Dermacentor variabilis both the fat body and the midgut produce the female- specific protein vitellogenin (Coons et al., 1982, 1986, 1988; Tarnowski and Coons, 1988). However, in the soft tick, Ornithodoros moubata, only the fat 0168-8162/90/$03.50 © 1990 Elsevier Science Publishers B.V.
Transcript of Fine structure of the fat body and nephrocytes in the life-stages of Dermacentor variabilis

. I
Experimental & Applied Acarology, 8 ( 1990) 125-142 125 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands
Fine Structure of the Fat Body and N ephrocytes in the Life-Stages of Dermacentor variabilis
LEWIS B.COONS, WILLIAM J . L'AMOREAUX, ROSEMARIE ROSELL-DAVIS and LINDA STARR-SPIRES
Department of Biology, Memphis State University, Memphis, TN 38152 (U.S.A.)
(Accepted 27 October 1988)
ABSTRACT
Coons, L.B ., L'amoreaux, W.J ., Rosell-Davis, R. and Starr-Spires, L., 1990. Fine structure of the fat body and nephocrytes in the life stages of Dermacentor variabilis. Exp. Appl. Acarol., 8: 125-142
The fine structure of the fat body and associated nephrocytes of the American dog tick, Dermacentor variabilis (Say) , was described in unfed larvae, unfed nymphs, and in unfed and fed adults of both sexes. The fat body consisted of one type of cell, the trophocyte. Morphological changes that occured in the trophocytes of both sexes were dependent on feeding. The ultrastructure of feeding male trophocytes was distinct from trophocytes of feeding females. In the feeding female , the trophocyte developed an ultrastructure characteristic of cells that produce secretory proteins. A type of scalariform cell junction was found associated with rough endoplasmic reticulum of the trophocytes. Nephrocytes were closely associated with trophocytes but were not part of the fat body. Nephrocyte ultrastructure was unaltered throughout the life-stages we examined, except at the end of oviposition. Organelles in the nephrocytes were not randomly distributed, but were found in distinct regions of the cytoplasm. Slit diaphragms at the surface of the nephrocytes were extracellular specializations that had a periodic ultrastructure.
INTRODUCTION
In the American dog tick, Dermacentor variabilis (Say), a fat body is present in each life-stage: larva, nymph, and adult (Obenchain and Oliver, 1973). The fat body is located just inside the epidermal layer of cells and between the organs. These two regions are known respectively as the peripheral fat body and the central fat body (Obenchain and Oliver, 1973). In females, the fat body, especially the peripheral fat body, proliferates during and after feeding (Obenchain and Oliver, 1973 ). In the hard ticks Rhipicephalus sanguinius and Dermacentor variabilis both the fat body and the midgut produce the femalespecific protein vitellogenin (Coons et al., 1982, 1986, 1988; Tarnowski and Coons, 1988). However, in the soft tick, Ornithodoros moubata, only the fat
0168-8162/90/$03.50 © 1990 Elsevier Science Publishers B.V.

126 L.B. COONS ET AL.
body produces vitellogenin ( Chinzei and Yano, 1985). These studies used a combination of various immunological techniques and some electron microscopy. Vitellogenic fat body cells in R. sanguinius have large amounts of rough endoplasmic reticulum, Golgi bodies and secretory granules, an ultrastructure characteristic of secretory cells (Coons et al., 1982). N ephrocytes are present in Hyalomma asiaticum (Amosova, 1983). Their ultrastructure is very similar to nephrocytes found in insects (Crossley, 1985 ). Nephrocytes are commonly associated with the fat body and with body organs such as the heart and ovaries. We studied only those nephrocytes associated with the fat body trophocytes.
The objectives of this study were to describe the ultrastructure of the fat body and the closely associated nephrocytes in unfed larva and nymphs, and fed adults, of D. variabilis. We also described a scalariform type of cell junction found in the fat body. Part of this study has been presented at a scientific meeting (Coons et al., 1986).
MATERIALS AND METHODS
Ticks
Ticks used in this study were either from a laboratory colony maintained at Memphis State University, or were collected from dogs at the city animal shelter. We obtained similar results from colony ticks and wild-type ticks. All ticks were maintained in an incubator at 853 r.h., with a dark/light cycle of 12/12h. Ticks were fed on rabbits as previously described by Coons and Kaufman ( 1988). The rabbits had not been previously exposed to tick infestations, and they were used as a host only once. Mated and unmated female ticks were fed in the same way. Ticks were sampled by hand-removing them from the rabbits, or after they had detached from the rabbit as in the case of completely fed mated females. With unmated fed females, no males were present on the rabbits during feeding and we examined the reproductive system of each female for signs of mating. Unmated female ticks were fed for 12 to 13 days, which was as long or longer than was required for mated female ticks to complete feeding and naturally detach. The unmated female ticks did not enter the rapidengorgement phase of feeding and had to be hand-removed from the host. Following removal from the host, ticks were held at room temperature in glass petri dishes until sampled. Some of the unmated females were held in the laboratory until they began to oviposit. We sampled at least three ticks at each feeding and postfeeding period.
Electron microscopy
Tissues were routinely fixed in situ at room temperature (22 °C) for 1-2 h in 2.53 glutaraldehyde in either O.lM cacodylate or 0.2MMillonig's phosphate

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 127
buffer, pH 7 .2 (Millonig, 1961). After fixation, tissues were excised and washed in the appropriate buffer, post-fixed at room temperature (22 ° C) for 1 h in 1 % osmium tetroxide in buffer, dehydrated in ascending acetone series and embedded in Epon-Araldite (Mollenhauer, 1964). Silver to light-gold sections, obtained with a glass knife on a Sorvall Porter-Blum MT-2B ultramicrotome, were stained with uranyl acetate and lead (Venable and Coggeshell, 1965; Sato, 1968). Some tissues were en-bloc stained with uranyl acetate according to karnovsky (1967). Tannie acid preparations (TAG), prepared according to Rodewald and Karnovsky (1974), were employed to increase contrast of the slit diaphragms of the nephrocytes.
RESULTS
Fat body morphology
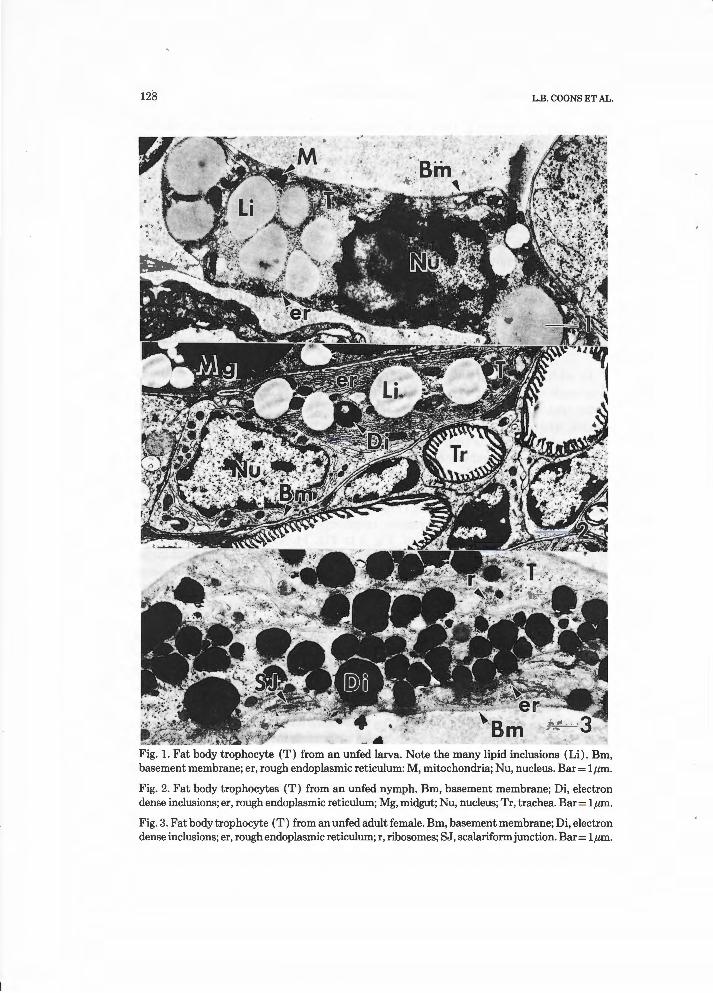
In all life-stages of Dermacentor variabilis that we examined, the fat body was made up of one cell type, the trophocyte. The ultrastructure of the trophocytes was similar in the unfed larvae, unfed nymphs, unfed adults and fed males, whereas the trophocytes in fed females underwent a dramatic change in ultrastructure. The trophocytes were surrounded by a basement membrane in all life-stages. Furthermore, the basement membrane of the trophocyte was distinct from that of the associated nephrocyte. In unfed larvae, trophocytes were characterized by lipid inclusions, sparse rough endoplasmic reticulum, and few scattered mitochondria (Fig. 1). The nucleus occupied a considerable portion of the cell. In unfed nymphs, trophocytes had an increased amount of rough endoplasmic reticulum (cf. Fig. 2 to Fig. 1). Trophocytes in unfed females had sparse rough endoplasmic reticulum, mitochondria and lipid inclusions (Fig. 3). The most common cytoplasmic features were free ribosomes and dense inclusions that sectioned poorly, often causing tearing of the surrounding tissue. The trophocytes of unfed males (not shown) were similar to those in unfed females.
Fat body trophocytes sampled from feeding males had few mitochondria, but did have large glycogen fields and electron-dense inclusions that were distributed throughout the cytoplasm (Fig. 4). Rough endoplasmic reticulum, Golgi bodies, foot processes, and the extracellular spaces associated with foot processes were not nearly as common in the trophocytes from feeding males as they were in the fat body trophocytes of feeding females (cf. Fig. 4 to Fig. 6). Foot processes were located next to the basement membrane and were produced by an infolding of the plasma membrane of the trophocyte which resulted in a partial enclosure of the extracellular space (see Figs. 4, 6, 7, 8, and 9). We did not find lipid inclusions or secretory products in fat body trophocytes sampled from feeding males.
During feeding, female fat body trophocytes underwent considerable changes
128 L.B. COONS ET AL.
~!'i;S~~~J!t!....~ ... --~ ·.
Fig. 1. Fat body trophocyte (T) from an unfed larva. Note the many lipid inclusions (Li) . Bm, basement membrane; er, rough endoplasmic reticulum: M, mitochondria; Nu, nucleus. Bar= lµm.
Fig. 2. Fat body trophocytes (T) from an unfed nymph. Bm, basement membrane; Di, electron dense inclusions; er, rough endoplasmic reticulum; Mg, midgut; Nu, nucleus; Tr, trachea. Bar= lµm.
Fig. 3. Fat body trophocyte (T) from an unfed adult female . Bm, basement membrane; Di, electron dense inclusions; er, rough endoplasmic reticulum; r, ribosomes; SJ, scalariformjunction. Bar= lµm .

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 129
Fig. 4. Fat body trophocyte from a male fed for 3 days. Bm, basement membrane; Di, electron dense inclusions; ES, extracellular space; fp, foot processes; G, glycogen; Nu, nucleus. Bar= 2µm.
Fig. 5. Fat body trophocyte from an unmated female fed for 3 days. Note the swollen cisternae (cs) in the developing rough endoplasmic reticulum. Bm, basement membrane; ES, extracellular space; fp, foot process; G, glycogen; Nu, nucleus. Bar= 1 µm.
Fig. 6. Fat body trophocyte of a mated female sampled just after completion of feeding. Bm, basement membrane; er, rough endoplasmic reticulum; ES, extracellular space; GJ, gap junction; fp , foot processes; GB, Golgi body; Li, lipid inclusion; M, mitochondria; Nu, nucleus; sp, secretory products. Bar= lµm .
130 L.B. COONS ET AL.
~ .... ,,..-'<lop-'' ~~ .,"
~ . "',
~ It ;-; ).'''
-1"$"f .;r,:;.'
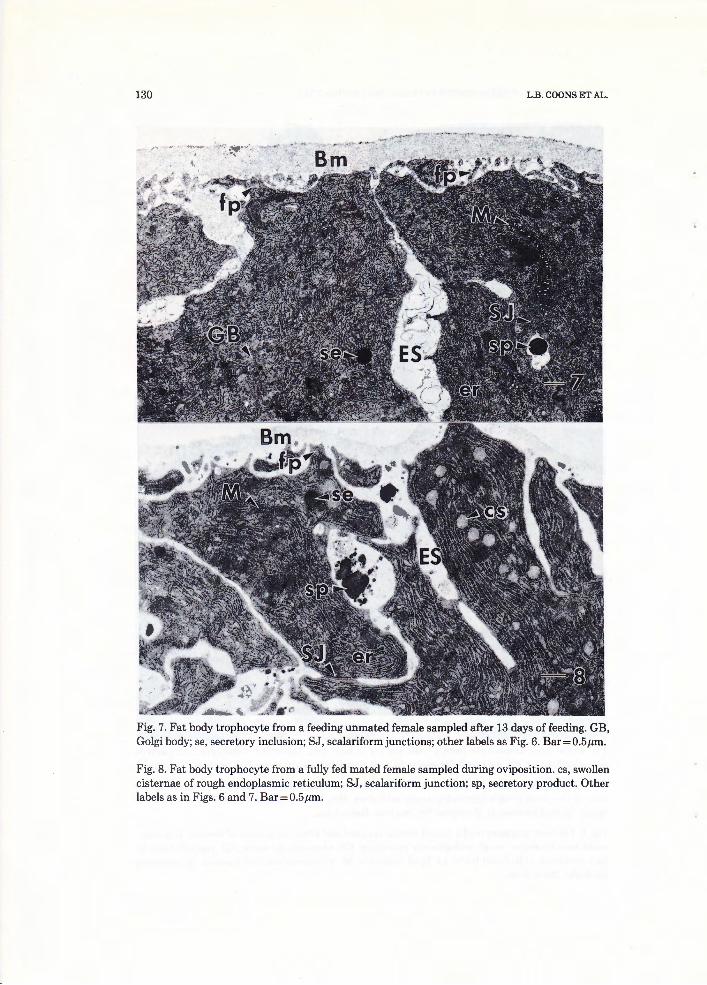
Alt~ ... "~- " " Fig. 7. Fat body trophocyte from a feeding unmated female sampled after 13 days of feeding. GB, Golgi body; se, secretory inclusion; SJ, scalariform junctions; other labels as Fig. 6. Bar= 0.5 µm.
Fig. 8. Fat body trophocyte from a fully fed mated female sampled during oviposition. cs, swollen cisternae of rough endoplasmic reticulum; SJ, scalariform junction; sp, secretory product. Other labels as in Figs. 6 and 7. Bar=0.5µm.

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 131
in their cell structure. Foot processes started to form early in the feeding period (Fig. 5). The dense inclusions found in unfed female trophocytes were not observed in 3-day-fed females (cf. Fig. 3 to Figs. 5 and 6). Rough endoplasmic reticulum proliferated and often had distended cisternae (Fig. 5). Only small amounts of glycogen were found in the cytoplasm (Fig. 5) . By the completion of feeding, fat body trophocytes were much larger and had increased amounts of rough endoplasmic reticulum, extracellular spaces, mitochondria, and Golgi bodies (Fig. 6). The extracellular spaces extended inward between adjacent fat body trophocytes (Fig. 6). Fat body trophocytes sampled from feeding mated and unmated females had developed the same ultrastructure (cf. Fig. 6 to Fig. 7). Nucleolar channels, an unusual organelle, were found in the fat body trophocytes sampled from feeding females (Fig. 11). An electron-dense secretory product was found in the extracellular spaces of trophocytes from fully fed mated females sampled immediately after feeding, in unmated females sampled immediately after being fed 13 days, and in the extracellular spaces of trophocytes from mated and unmated females that had begun to oviposit (Figs. 6, 7, 8 and 9).
By oviposition, fat body trophocytes of both unmated and mated females had developed more extensive extracellular spaces, foot processes and more secretory products than the fat body trophocytes from feeding mated or un -mated females (cf. Figs. 8 and 9 to Figs. 6 and 7). Swollen cisternae in rough endoplasmic reticulum were more common in mated ovipositing females than in feeding, mated, non -ovipositing females (cf. Fig. 8 to Fig. 6). In unmated females, the cisternae of rough endoplasmic reticulum also became distended during oviposition (cf. Fig. 9 to Fig. 7). Golgi bodies were common in trophocytes from females during rapid engorgement (Fig. 12) and in trophocytes from females sampled during oviposition (Fig. 7). Coated pits were common along the plasma membrane of trophocytes (Fig. 13). Near the end of oviposition, the fat body trophocyte had few extracellular spaces or foot processes and mitochondria were often swollen (Fig. 10) .
Two types of cell junctional specializations found between trophocytes were scalariformjunctions and gap junctions (Fig. 14). While scalariform junctions were most common between the trophocytes of females, occasionally these junctions were present between the trophocytes of unfed males (Fig. 15). Fat body trophocytes of unfed females, unfed males, and fed males showed fewer scalariform junctions than fed-female fat body trophocytes. The scalariform junctions lacked mitochondrial associations. Instead, rough endoplasmic reticulum was closely associated with the plasma membrane along the length of the junction (Figs. 14 and 15 ). We measured seven different scalariform junctions. The rough endoplasmic reticular cisternum apposed to the plasma membrane was devoid of ribosomes and exhibited a regular spacing of 4.13 nm between the plasma membrane and the endoplasmic reticulum membrane. The space between the two plasma membranes of the junction was 13.13 nm. Pe-

132 L .B. COONS ET AL.

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 133
Fig. 9. Fat body trophocyte from a unmated female fed 13 days then held until oviposition began 12 days later. cs, swollen cisternae of rough endoplasmic reticulum; se, secretory inclusion; sp, secreted product. Other labels as in Figs. 6 and 7. Bar= 0.5 µm.
Fig. 10. Fat body trophocyte from a mated female sampled near the end of ovipositoin. Bm, basement membrane; Di, electron dense inclusions; er, rough endoplasmic reticulum; M, mitochondria. Bar=0.5µm.
Fig. 11. Fat body trophocyte from a feeding mated female sampled during rapid engorgement. Nucleolar channels (NC) are present within the nucleus (Nu). Bm, basement membrane. Bar=0.2µm.
Fig. 12. Fat body trophocyte from a feeding mated female sampled during rapid engorgement. er, rough endoplasmic reticulum; GB, Golgi body. Bar=O.lµm.
Fig. 13. Fat body trophocyte from an unmated female fed 13 days then held until oviposition began 12 days later. cp, coated pit; cv, coated vesicle; ES, extracellular space; sp, secretory product. Bar=O.lµm .
riodic cross-striations were observed between the two plasma membranes. The center-to-center distance between the pillars of the cross-striations was 19.4 nm. Scalariform junctions were both homocellular and heterocellular. Gap junctions were often found adjacent to the scalariformjunctions. (Fig. 14).
Nephrocytes
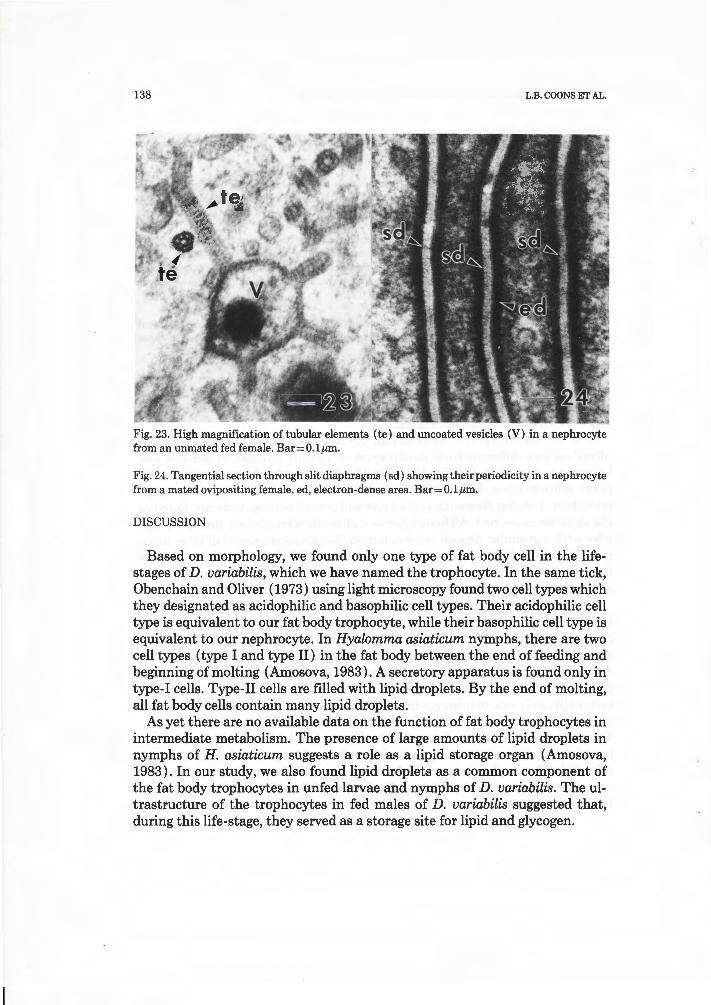
Nephrocytes had a similar ultrastructure during most life-stages examined (Figs. 16, 17 and 18). Nephrocytes underwent a rapid increase in number during the feeding and pre-oviposition stages in females. Organelles were not distributed in a random fashion but were found in distinct regions of the cytoplasm. This was most apparent in the larger nephrocytes of feeding and ovipositing adult females. In nephrocytes from these ticks, the organelles were distributed into three different regions in the cytoplasm (Fig. 18). An outer region, next to the basement membrane, was characterized by slit diaphragms at the surface of extracellular channels. The basement membrane did not continue into these diaphragms. Coated vesicles were common in the cytoplasm in the outer region (Figs. 18 and 21). Coated pits were abundant at the distal ends of the channels (Fig. 21). Tubular elements and uncoated vesicles with an electron-dense core were also present in the outer region (Fig. 18). Often, tubular elements were connected to the uncoated vesicles (Figs. 18 and 23).
The middle region appeared to be a transition zone between the other two regions. This region was characterized by dense membrane-bound inclusions of varying sizes not found in the other regions (Fig. 18). Glycogen fields, lipid inclusions and Golgi bodies were found in both the middle region and the inner

134 L.B. COONS ET AL.
Fig. 14. Scalariformjunction (marked by white arrows) between two fat body trophocytes of a fed mated female. Note the indistinct cross-striations (er) in the extracellular space of the scalariform junction. er, rough endoplasmic reticulum; GJ, gap junction; r, ribosomes. Bar=O.lµm.
Fig. 15. Scalariform junction (SJ) in the fat body trophocyte of an unfed male. Bar=0.2µm.
Fig. 16. Nephrocytes (N) from an unfed larvae. Di, electron dense inclusion; Nu, nucleus; T, fat body trophocyte. Bar=2µm.
Fig .17. Nephrocyte (N) from an unfed nymph. Mg, midgut; Tr, trachea. Other labels as in Fig. 16. Bar=lµm.

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 135
Fig. 18. Nephrocyte from an unmated female fed 6 days. There are three regions in the cell, each with a distinct set of organelles. The outer region (or) next to the basement membrane (Bm) contains slit diaphragms (sd) that are found at the openings of channels. Coated vesicles (cv) and tubular elements (te) are common in the outer region. Uncoated vesicles (V) with a dense region are commonly found in the outer region. Some glycogen deposits ( G) are also present. The middle region (mr) is a transition area characterized by electron-dense inclusions (Di) of varying sizes, Golgi bodies (GB), lipid inclusions, (Li), occasional mitochondria (M), glycogen (G ), and scattered rough (er) and smooth (ser) endoplasmic reticulum. An inner region (ir) is characterized by well-developed fields of glycogen ( G) , rough endoplasmic reticulum (er), smooth endoplasmic reticulum (ser). Mitochondria (M) are common. Bar=0.5µm.

L.B. COONS ET AL.

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 137
Fig. 19. Nephrocyte from a fed mated female at the end of oviposition. Asterisk, uncoated vesicles with granular deposits. Other labels as in Fig. 18. Bar= lµm.
Fig. 20. Nephrocyte (N) and trophocyte (T) from a fully fed mated female. fp, foot processes; GJ, gap junction. Other labels as in Fig. 18. Bar= 1 µm. Inset: Higher resolution of gap junction ( GJ). Bar=0.2µm.
Fig. 21. Outer region of a mated rapid-engorging female nephrocyte showing the various configurations of the silt diaphragms. One (single arrowhead), two, (double arrowhead) or three (triple arrowheads) slit diaphragms are common. Most channels have one slit diaphragm but occasionally multiple slit diaphragms occur. Bm, basement membrane; cp, coated pit; cv, coated vesicle; ed, electron-dense area; te, tubular element. Bar=0.2µm.
Fig. 22. Slit diaphragm (sd) from a fed male showing that it is a modification of the surface coat of the plasma membrane. Tannie acid preparation. Bm, basement membrane; ed, electron-dense area; fp, foot processes. Bar=0.05µm.
region. Isolated glycogen rosettes and mitochondria were common to all three regions. The inner region was characterized by well-organized fields of rough and smooth endoplasmic reticulum.
In ticks near the end of oviposition, the nephrocytes we examined had an ultrastructure different from nephrocytes of feeding females or females just beginning oviposition (cf. Fig. 18 to Fig. 19). Nephrocytes from these ticks had fewer mitochondria, less glycogen, and less rough and smooth endoplasmic reticulum. Tubular elements, coated pits and coated vesicles were not found in the ticks we examined. Although dense inclusions were absent, uncoated vesicles with a granular deposit were common. Nephrocytes found in close association with fat body trophocytes had a portion of their basement membrane fused with the trophocyte basement membrane (Fig. 20). Occasionally, gap junctions were found at the point of fusion (Fig. 20, inset).
Slit diaphragms were surface junctional complexes that readily stained with tannic acid and were located just beneath the basement membrane in nephrocytes (Figs. 18, 21, 22, 24). Slit diaphragms were a modification of the surface coat of the plasma membrane and therefore extracellular in nature (Fig. 22). A band of electron-dense material was located in the cytoplasm adjacent to the plasma membrane on either side of the slit diaphragm (Figs. 21and22). Occasionally, only one diaphragm bridged the opposing plasma membranes, but multiple slit diaphragms were also seen within the same channel (Fig. 21). Tangential sections showed that the slit diaphragms had a periodicity consisting of extracellular cementing material perforated by pores of different sizes (Fig. 24). The diaphragm appeared to be composed of hexagonal particles arranged in a honey-comb fashion, closely resembling those found in insects (Crossley, 1985).
138 L.B. COONS ET AL.
Fig. 23. High magnification of tubular elements (te) and uncoated vesicles (V) in a nephrocyte from an unmated fed female. Bar=O.lµm .
Fig. 24. Tangential section through slit diaphragms (sd ) showing their periodicity in a nephrocyte from a mated oviposit ing ,female. ed, electron-dense area. Bar=O.lµm.
DISCUSSION
Based on morphology, we found only one type of fat body cell in the lifestages of D. variabilis, which we have named the trophocyte. In the same tick, Obenchain and Oliver ( 1973) using light microscopy found two cell types which they designated as acidophilic and basophilic cell types. Their acidophilic cell type is equivalent to our fat body trophocyte, while their basophilic cell type is equivalent to our nephrocyte. In Hyalomma asiaticum nymphs, there are two cell types (type I and type II) in the fat body between the end of feeding and beginning of molting (Amosova, 1983). A secretory apparatus is found only in type-I cells. Type-II cells are filled with lipid droplets. By the end of molting, all fat body cells contain many lipid droplets.
As yet there are no available data on the function of fat body trophocytes in intermediate metabolism. The presence of large amounts of lipid droplets in nymphs of H. asiaticum suggests a role as a lipid storage organ (Amosova, 1983) . In our study, we also found lipid droplets as a common component of the fat body trophocytes in unfed larvae and nymphs of D. variabilis. The ultrastructure of the trophocytes in fed males of D. variabilis suggested that, during this life-stage, they served as a storage site for lipid and glycogen.

FINE STRUCTURE .OF DERMACENTOR FAT BODY AND TROPHOCYTES 139
In both mated females and unmated females, fat body trophocytes underwent a distinct ultrastructural change during feeding, with the resultant cells typical of those manufacturing and exporting secretory proteins ( Palade, 197 5). The change was gradual during the feeding period and continued during the preoviposition phase. By the end of oviposition, the fat body trophocyte resembled an exhausted cell that was near death. We could not distinguish between the ultrastructure of trophocytes in a feeding mated female and a feeding unmated female nor between the trophocytes of an ovipositing mated tick and an ovipositing unmated tick. We observed that, in the trophocytes of ovipositing mated and unmated ticks, there were a greater number of secretory products in the extracellular spaces. Also, the extracellular spaces and foot processes were' larger when compared to trophocytes of feeding mated or unmated ticks.
The nucleolar channels we described in the trophocytes of feeding females are found in human endometrial cells (Terzakis, 1965). The scalariform junction described in this paper is very similar to the scalariform junction in the apterygote insect Petrobius maritimus (Fain-Maurel and Cassier, 1972). In P. maritimus the junction is associated with mitochondria, while in D. variabilis fat body the junction was associated with rough endoplasmic reticulum. It has been suggested by Oschman and Wall (1969) that scalariformjunctions may function to maintain basal infoldings of the cell membrane. Such a role is consistent with the importance of maintaining the integrity of the extracellular channels of the foot processes in vitellogenic trophocytes, thus allowing free passage of the secretory product to the hemolymph. The fat body trophocytes of the fed male had an ultrastructure very different from those of the fed female. The dramatic difference in ultrastructure between the male and female fat body trophocytes in this tick is similar to the trophocytes of the higher t ermites (Han and Bordereau, 1982).
Our study and the study of Obenchain and Oliver (1973) confirm that the fat body proliferates during and after feeding in females. By the onset of oviposition, the peripheral fat body appeared as an anastomosing network oftrophocytes which filled the body cavity between the epidermis and the organs. T his proliferation of the peripheral fat body was also found to a lesser degree in unmated females that were hand-removed and allowed to undergo preoviposition until they started to lay eggs. While feeding resulted in some proliferation of the peripheral fat body, our study found that the greatest proliferation in both mated and unmated females occured during preoviposition, suggesting to us that detachment from the host was a signal that induced this response. This proliferation was never found in fed males that had been removed from the host and held in the laboratory for up to 1 week. Obenchain and Oliver ( 1973) also found that the peripheral fat body in fed males did not proliferate.
Nephrocytes have been described at the electron-microscopic level from Hyalomma asiaticum (Amosova, 1983), and Argas (Persicargas) arboreus (El

140 L.B. COONS ET AL.
Shoura, 1986) and at the light-microscopic level in D. variabilis by Obenchain and Oliver ( 1973). The presence of cell junctions between trophocytes and nephrocytes led Amosova ( 1983) to conclude that both cell types are part of the fat body. However, the nephrocytes of ticks are so similar in ultrastructure to the nephrocytes described in the Insecta (Crossley, 1985) that we have considered them as a separate cell type which is closely associated with the fat body.
In the Insecta, the available evidence supports the conclusion that the slit diaphragm serves as a filter barrier based on size (Crossley, 1985). The slit diaphragm of the tick nephrocyte appeared to closely resemble those found in Calliphora by Crossley ( 1972; 1985) and in H. asiaticum (Amosova, 1983). The electron-dense material that we described associated with the slit diaphragm is also associated with the slit diaphragm of nephrocytes in Calliphora ( Crossley, 1985). We believe that, like insects, the slit diaphragm functions as a filtration barrier based on molecular size. The presence of an array of organelles associated with receptor-mediated endocytosis (coated pits, coated vesicles, tubular elements, and uncoated vesicles) suggested that nephrocytes function to selectively sequester substances from the hemolymph, as in the Insecta (Crossley, 1985 ). Obenchain and Oliver (1973) found that nephrocytes ( = basophilic cell types) incorporated presumptive hemoglobin derivatives from the hemolymph and that they are implicated in purine metabolism.
Nephrocytes in ticks have clusters of glycogen and lipid droplets as do the nephrocytes of insects (Crossley, 1985). The zonation of organelles in nephrocytes of both insects and ticks suggests a similar necessity of metabolic activity and intracellular sorting of materials taken up from the hemolymph and products synthesized by the nephrocytes. The presence of nephrocytes dispersed widely throughout the fat body is not limited to ticks but is also commonly found in the Lepidoptera (Crossley, 1985 ). It should be noted that no one has yet observed degranulation of nephrocytes in ticks as has been seen in some insects (Crossley, 1985).
We found that nephrocytes changed little from larvae to nymphs in the functional states we examined. In feeding and postfeeding females, the nephrocytes proliferated at the same time as the fat body trophocytes. At the end of ovipostion, the reduction of coated vesicles, coated pits or tubular elements in nephrocytes prompted us to conclude that uptake of material from the hemolymph was reduced or lacking.
The function of nephrocytes in ticks is unknown. They may surve as an intermediate in protein metabolism and manufacture of proteins such as lysozyme, which has been postulated in insects (Crossley, 1985). Most probably, they serve to take up selected substances from the hemolymph and act in ticks as they do in insects as an analogue to the vertebrate reticuloendothelial system (Wigglesworth, 1970). Recent evidence indicates that the fat body of D. variabilis produces ecdysteroid in vitro (Schriefer et al., 1987). Steroids are

FINE STRUCTURE OF DERMACENTOR FAT BODY AND TROPHOCYTES 141
thought to be produced in the smooth endoplasmic reticulum (Holtzman and Novikoff, 1984). Smooth endoplasmic reticulum was present in the nephrocytes and it is possible that they produce ecdysteroids.
APPENDIX
List of abbreviations
Bm basement membrane cp coated pit er cross striations cs swollen cisternae CV coated vesicle Di electron dense inclusions ed electron dense area er rough endoplasmic reticulum ES extracellular space fp foot processes G glycogen GB Golgi body GJ gap junction ir inner region Li lipid inclusion M mitochondria Mg midgut mr middle region N nephrocyte NC nucleolar channels Nu nucleus or outer region r ribosomes sd slit diaphragm se secretory inclusion ser smooth endoplasmic reticulum SJ scalariform junction sp secretory product T fat body trophocyte te tubular element Tr trachea v uncoated vesicle * uncoated vesicle with granular deposits
REFERENCES
Amosova, L.I., 1983. Tissues of the internal environment. In: Y. Balashov (Editor), An Atlas of Ixodid Tick Ultrastructure. Entomol. Soc. Am. Spec. Pub!., 8: 147-174.
Chinzei, Y. andYano, I., 1985. Fat body is the site ofvitellogenin sythesis in the soft tick, Ornithodoros moubata. J. Comp. Physiol., B, 155: 671-678.

142 L.B. COONS ET AL.
Coons, L.B. and Kaufman, W.R., 1988. Evidence that developmental changes in type III acini in the tick, Amblyomma hebraeum (Acari: Ixodidae) are initiated by a hemolymph-borne factor. Exp. Appl. Acarol. , 4: 117-139.
Coons, L.B., Lamoreaux, W.J. , Rosell-Davis, R. , and Starr-spires, L., 1986. Ultrastructure of fat body trophocytes in the life stages of Dermacentor variabilis (Say) . Proc. EMSA, 44: 314-315.
Coons, L.B. , Tarnowski, B., and Ourth, D., 1982. Rhipicephalus sanguinius: Localization of vitellogenin synthesis by immunological methods and electron microscopy. Exp. Parasitol., 59: 331-339.
Coons, L.B., lamoreaux, W.J., Rosell-Davis, R. and Tarnowski, B.I., 1989. Onset of vitellogenin production and vitellogenesis, and their relationship to changes in the midgut epithelium and oocytes in the tick Dermacentor variabilis. Exp. Appl. Acarol. , 6: 291-305.
Crossley, A.C., 1972. The ultrastructure and function of pericardia! cells and other nephrocytes in an insect: Calliphora erythrocephala. Tissue Cell, 4: 529-560.
Crossley, A.C., 1985. Nephrocytes and pericardia! cells. In: G.A. Kerkut and L.I. Gilbert (Editors) , Comprehensive Insect Physiology, Biochemistry and Pharmacology, Vol. 3. Integument, Respiration and Circulation. Pergamon Press, New York, pp. 487-515.
El Shoura, S.M., 1986. Fine structure of the hemocytes and nephrocytes of Argas (Persicargas) arboreus (lxodoidea: Argasidae) . J. Morphol., 189: 17-24.
Fain-Maurel, M.A. and Cassier, P ., 1972. Une nouveau type de jonctions: les jonctions scalariformes. Etude ultrastructurale et cytochimique. J. Ultrastruct. Res., 39: 222-238.
Han, S.H. and Bordereau, C., 1982. Ultrastructure of the fat body of the reproductive pair in higher termites. J. Morphol. , 172: 313-322.
Holtzman, E. and Novikoff, A.B., 1984. Cells and Organelles. W.B. Saunders, Philadelphia, pp. 148-152.
Karnovsky, M.J ., 1967. The ultrastructural basis of capillary permeability studied with peroxidase as a tracer. J . Cell Biol., 35: 215-236.
Millonig, G., 1961. Advantages of a phospate buffer for osmium tetroxide solutions in fixation. J. Appl. Physiol. , 32: 1637.
Mollenhauer, H.H., 1964. Plastic embedding mixtures for use in electron microscopy. Stain Technol. , 39: 11.
Obenchain, F.D. and Oliver, J.H., 1973. A qualitative analysis of the form, function and interrelationships of fat body and associated tissues in adult ticks (Acari-Ixodoidea) . J . Exp. Zoo!., 186: 217-236.
Oschman, J .L. and Wall, B.J. , 1969. The structure of the rectal pads of Periplaneta americana L. with regard to fluid transport. J. Morphol., 127: 475-510.
Palade, G., 1975. Intracellular aspects of the process of protein synthesis. Science, 189: 347-358. Rodewald, R. and Karnovsky, M.J. , 1974. Porous structure of the glomerular slit diaphragm in
the rat and mouse. J. Cell Biol., 60: 423-433. Sato, T ., 1968. A modified method for lead staining. J. Electron Microsc., 17: 158:-159. Schriefer, M.E., Berveridge, M. Sonenshine, D.E., Homsher, P.J., Carson, K.A. and Weiman, C.S.,
1987. Evidence of ecdysteroid production by tick (Acari: Ixodidae) fat-body tissues in vitro. J . Med. Entomol., 24: 295-302.
Tarnowski, B.I. and Coons, L.B., 1989. Ultrastructure of the midgut and blood meal digestion in the adult tick Dermacentor variabilis. Exp. Appl. Acarol., 6: 263-289.
Terzakis, J .A., 1965. The nucleolar channel system of human endometrium. J . Cell Biol., 27: 293-304.
Venable, J.J . and Coggeshell, R., 1965. A simplified lead citrate stain for use in electron microscopy. J. Cell Biol., 25: 407-408.
Wigglesworth, V.B., 1970. The pericardia! cells of insects analogue of the reticuloendothelial system. J . Reticuloendothel. Soc., 7: 208-216.




















