
Molecular mechanisms in plant abiotic stress response
267
Ratarstvo i povrtarstvo Field and Vegetable Crops Research 2011: 48 (1) abstracted / indexed in EBSCO Academic Search Complete, CAB Abstracts, CABI Full-Text, FAO Agris, Serbian Citation Index www.nsseme.com/journal.html
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Molecular mechanisms in plant abiotic stress response

Ratarstvo i povrtarstvoField and Vegetable
Crops Research2011: 48 (1)
abstracted / indexed in
EBSCO Academic Search Complete, CAB Abstracts, CABI Full-Text, FAO Agris, Serbian Citation Index
ww
w.ns
sem
e.co
m/jo
urna
l.htm
l


SADRŽAJ / CONTENTS
PREGLEDNI RADOVI / REVIEW ARTICLES
Genetički resursi / Genetic Resources
1-6 Conservation and Breeding of Ornamental Lathyrus Species / Očuvanje i oplemenjivanje ukrasnih vrsta roda Lathyrus
Roger Parsons, Aleksandar Mikić
Genetika i oplemenjivanje / Genetics and Breeding
7-14 Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline: stanje i perspektive / Wheat Breeding for Low Phytic Acid Content: State and Perspectives
Gordana Branković, Desimir Knežević, Dejan Dodig, Vesna Dragičević
Fiziologija stresa / Stress Physiology
15-24 Molecular Mechanisms in Plant Abiotic Stress Response / Molekularni mehanizmi odgovora biljaka na abiotički stres
Palmiro Poltronieri, Stefania Bonsegna, Stefania De Domenico, Angelo Santino
Fitopatologija / Phytopathology
25-30 Crvenilo kukuruza: pola veka kasnije / Corn Reddening: Half a Century Later
Goran Bekavac, Božana Purar, Đorđe Jocković, Sanja Treskić, Bojan Mitrović, Ivica Đalović, Željko Milovac
ORIGINALNI NAUČNI RADOVI / ORIGINAL RESEARCH ARTICLES
Genetički resursi / Genetic Resources
31-36 Genetic Relationships among Wild Lens Mill. Species Revealed by SDS-PAGE / Genetički odnosi između samoniklih vrsta Lens Mill. Otkriveni pomoću SDS-PAGE
Galina Suvorova, Nadezhda Kornienko
Genetika i oplemenjivanje / Genetics and Breeding
Strna žita / Small Grains
37-48 Značaj faze organogeneze formiranja klasića u biologiji prinosa ozimog dvoredog ječma / Importance of Spikelet Formation Phase in the Yield Biology of Winter Barley
Novo Pržulj, Vojislava Momčilović
49-56 Novi konzumni hibridi suncokreta u NS paleti / New Confectionary Sunfl ower Hybrids in NS Palette
Nada Hladni, Vladimir Miklič, Siniša Jocić, Zvonimir Sakač, Velimir Radić, Ilija Radeka
Ana Marjanović-Jeromela, Sreten Terzić, Miroslav Zorić, Radovan Marinković, Jovanka Atlagić, Petar Mitrović, Željko Milovac
67-76
Uljane kulture/ Oil Crops
57-66 Produktivnost NS hibrida suncokreta u mikro-ogledima u Srbiji u 2010. godini / Productivity of NS Sunfl ower Hybrids in Small-Plot Trials in Serbia in 2010
Vladimir Miklič, Igor Balalić, Siniša Jocić, Radovan Marinković, Sandra Cvejić, Nada Hladni, Dragana Miladinović
Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice (Brassica napus L.) / Evaluation of Seed and Oil Yield Stability in NS Rapeseed Cultivars (Brassica napus L.)tte

Kukuruz / Maize
77-82 GGE biplot analiza multilokacijskih ogleda NS hibrida kukuruza / GGE Biplot Analysis of Multi-environment Trials of NS Maize Hybrids
Bojan Mitrović, Dušan Stanisavljević, Sanja Treskić, Milisav Stojaković, Goran Bekavac, Aleksandra Nastasić, Mile Ivanović
Šećerna repa/ Sugar Beet
Kemno bilje / Forage Crops
91-98 Genetički doprinos oplemenjivanju lucerke: prinos eksperimentalnih populacija u odnosu na priznate sorte / Genetic Gain in Alfalfa Breeding: Yield of Experimental Populations Versus Released Cultivars
Slobodan Katić, Dragan Milić, Snežana Katanski, Đura Karagić, Sanja Vasiljević
99-106 Kombinacione sposobnosti i način nasleđivanja debljine stabla F1 hibrida krmnog sirka (Sorghum bicolor L. Moench) / Combining Ability and Mode of Inheritance of Stem Thickness in Forage Sorghum (Sorghum bicolor L. Moench) F1 Hybrids
Imre Pataki
Konoplja / Hemp
107-112 Varijabilnost i međuzavisnost komponenti prinosa konoplje za vlakno / Variability and Interrelation of Yield Components in Fiber Hemp
Vladimir Sikora, Janoš Berenji, Dragana Latković
Molekularna genetika / Molecular Genetics
Strna žita / Small Grains
113-120 Analiza veze marker-svojstvo za vreme klasanja i cvetanja pšenice korišćenjem pojedinačne marker regresije / Marker-Trait Association Analysis for Heading and Flowering Time in Wheat by Single Marker Regression
Dragana Trkulja, Ankica Kondić-Špika, Ljiljana Brbaklić, Borislav Kobiljski
Citogenetika / Cytogenetics
Šećerna repa / Sugar Beet
121-124 Analiza mejoze triploida šećerne repe / Meiose Analysis of Sugar Beet Triploids
Snežana Mezei, Lazar Kovačev, Jovanka Atlagić, Nevena Nagl
Semenarstvo / Seed Science
Uljane kulture / Oil Crops
125-130 Uticaj različitih doza NPK đubriva na kvalitet semena uljane repice/ The Effect of Different Doses of NPK Fertilizers on the Quality of Rapeseed Seeds
Dušica Jovičić, Ana Marjanović-Jeromela, Milka Vujaković, Radovan Marinković, Zvonko Sakač, Zorica Nikolić, Branko Milošević
Krmno bilje / Forage Crops
131-136 Pokazatelji životne sposobnosti semena ozimih grahorica (Vicia spp.) / Indicators of Winter Vetch (Vicia spp.) Seed Vigor
Milka Vujaković, Dušica Jovičić, Đura Karagić, Aleksandar Mikić, Zorica Nikolić, Dragana Petrović, Ksenija Taški-Ajduković
Soja / Soybean
137-142 Prinos i semenski kvalitet soje u zavisnosti od uslova godine / Yield and Quality of Soybean Seeds as Affected by Growing Conditions
Vojin Đukić, Svetlana Balešević-Tubić, Vuk Đorđević, Mladen Tatić, Godana Dozet, Goran Jaćimović, Kristina Petrović
83-86 Nove hibridne sorte šećerne repe Instituta za ratarstvo i povrtarstvo otporne prema rizomaniji / New Sugar Beet Hybrid Varieties Developed at Institute of Field and Vegetable Crops Resistant to Rhizomania
Lazar Kovačev, Nikola Čačić, Snežana Mezei
87-90 Površina korena kao kriterijum za selekciju šećerne repe / Root Surface as Criterion for Selection of Sugar Beet
Dario Danojević, Živko Ćurčić, Nevena Nagl,Lazar Kovačev

Gajenje i agrotehnika / Soil and Crop Management
Šećerna repa / Sugar Beet
143-150 Uticaj rokova setve, đubrenja azotom i načina navodnjavanja na morfološke i fenotipske osobine sadnica šećerne repe / Effects of Planting Dates, Fertilization with N and Irrigation Mode on Morphological and Phenotypic Traits of Sugar Beet Seedlings
Milorad Rajić, Borivoj Pejić
Krmno bilje / Forage Crops
151-154 Yield and Carrying Capacity of Meadows in Albania / Prinos i proizvodnost livada u Albaniji
Luan Hajno, Fiqiri Tahiri
155-160 Uticaj međurednog rastojanja i setvene norme na prinos semena i suve materije crvene deteline (Trifolium pratense L.)/ Effect of Row Spacing and Seeding Rate on Seed and Dry Matter Yield of Red Clover (Trifolium pratense L.)
Rade Barać, Gojko Duronić, Đura Karagić, Sanja Vasiljević, Branko Milošević
Fitopatologija / Phytopathology
161-166 Otpornost topinambura (Helianthus tuberosus L.) prema sivo-mrkoj pegavosti suncokreta u poljskim uslovima / Resistance of Topinambour (Helianthus tuberosus L.)to Sunfl ower Stem Canker in Field Conditions
Sreten Terzić, Boško Dedić, Jovanka Atlagić, Vladimir Miklič
167-172 Sunfl ower Inbred Lines Screening for Tolerance to White Rot on Stalk / Testiranje inbred linija suncokreta na tolerantnost prema beloj truleži stabla
Boško Dedić, Dragana Miladinović, Siniša Jocić, Sreten Terzić, Sonja Tančić, Nenad Dušanić, Vladimir Miklič
173-178
179-182
Združeni usevi/Intercropping
183-188 Effect of Pea and Spring Cereals Intercropping on Grain Yield and Crude Protein Content / Uticaj združene setve stočnog graška i jarih strnih žita na prinos zrna i sadržaj sirovih proteina
Žydrė Kadžiulienė, Lina Šarūnaitė, Irena Deveikytė
Agroekologija / Agroecology
Teški metali / Heavy Metals
189-194 Methods for Assessment of Background Limit of Ni and Cr in Soils of Eastern Serbia /Metode za procenu Background Limita Ni i Cr u zemljištima istočne Srbije
Vesna Mrvić, Ljiljana Kostić-Kravljanac, Mirjana Zdravković, Nikola Koković, Veljko Perović, Dragan Čakmak, Mile Nikoloski
Vodni režim / Water Regime
195-202 Indeks aridnosti kao osnova analize uticaja režima padavina i temperature vazduha na prinos kukuruza u Vojvodini / Aridity Index as a Useful Tool for Describing Impacts of Precipitation and Temperature Regime on Maize Yield in Vojvodina
Borivoj Pejić, Goran Jaćimović, Dragana Latković, Đuro Bošnjak, Branko Marinković, Ksenija Mačkić
Ekonomska analiza / Economic Analysis
203-206 Smanjenje rizika u proizvodnji pšenice primenom vremenske prodajne opcije / Risk Reduction in Wheat Production with Weather Put Option
Todor Marković, Milenko Jovanović
Izolacija i detekcija gljiva Alternaria dauci (Kühn) Groves et Skolk i Alternaria radicina Meier, Drechsler et Eddy iz semena mrkve / Isolation and Detection of the Fungi Alternaria dauci (Kühn) Groves et Skolk and Alternaria radicina Meier, Drechsler et Eddy from Carrot Seeds
Dragana Petrović, Maja Ignjatov, Milka Vujaković, Zorica Nikolić, Ksenija Taški-Ajduković, Dušica Joviči
Detekcija Ustilago nuda (Jensen) Rostrup u semenu ozimog ječma / Detection of Ustilago nuda (Jensen) Rostrup in Winter Barley Seed
Maja Ignjatov, Dragana Petrović, Milka Vujaković, Ksenija Taški-Ajduković, Zorica Nikolić, Dušica Jovičić

207-212 Uticaj količine padavina na prinos pšenice i kukuruza kao proizvodni bazni rizik / Infl uence of Rainfall on Wheat and Corn Yield as a Production-Related Basis Risk
Todor Marković, Milenko Jovanović
213-218 Ekonomski aspekti proizvodnje suncokreta u Srbiji / Economics of Sunfl ower Production in Serbia
Marija Knežević, Rade Popović
Arheobotanika / Archaeobotany
219-226 Pisum & Ervilia Tetovac – Made in Early Iron Age Leskovac. Part One. Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar in Leskovac, South Serbia / Grašak i urov Tetovac – made in ranogvozdenodobni Leskovac. Deo prvi. Dve zalihe ugljenisanih zrnastih mahunarki sa gradinskog naselja Hisar kod Leskovca, južno Pomoravlje
Aleksandar Medović, Aleksandar Mikić, Branko Ćupina, Živko Jovanović, Svetlana Radović, Aleksandra Nikolić, Nemanja Stanisavljević
227-232 Pisum & Ervilia Tetovac – Made in Early Iron Age Leskovac. Part Two. Extraction of the Ancient DNA from Charred Seeds from the site of Hissar in South Serbia / Grašak i urov Tetovac – made in ranogvozdenodobni Leskovac. Deo drugi. Ekstrakcija drevne DNK iz ugljenisanih semena sa nalazišta Hisar u južnoj Srbiji
Živko Jovanović, Nemanja Stanisavljević, Aleksandra Nikolić, Aleksandar Medović, Aleksandar Mikić, Svetlana Radović, Vuk Đorđević
Etnobotanika / Ethnobotany
233-238 Words Denoting Faba Bean (Vicia faba) in European Languages / Reči koje označavaju bob (Vicia faba) u evropskim jezicima
Aleksandar Mikić
PRETHODNA SAOPŠTENJA / RESEARCH NOTES
Genetika i oplemenjivanje / Soil and Crop ManagementUljane kulture / Oil Crops
239-244 Varijabilnost visine biljke i prečnika glave kod novih hibridnih kombinacija suncokreta (Helianthus annuus L.) / Variability of Plant Height and Head Diameter in New Hybrid Combinations of Sunfl ower
Radovan Marinković, Milan Jocković, Siniša Jocić, Mihajlo Ćirić
Gajenje i agrotehnika / Genetics and BreedingKrmno bilje / Forage Crops
245-252 Preliminary Results on Agronomic Performance of Barrel Medic (Medicago truncatula) in Serbia / Prvi rezultati gajenja buretaste lucerke (Medicago truncatula) u Srbiji
Vojislav Mihailović, Aleksandar Mikić, Branko Ćupina, Dragan Milić, Đorđe Krstić, Slobodan Katić, Sanja Vasiljević, Đura Karagić
STAVOVI / VIEWS
253-258 The Legume Manifesto: (Net)workers on Fabaceae, Unite! / Legumistički manifest: (Net)proleteri svih Fabaceae, ujedinite se!
Aleksandar Mikić, Diego Rubiales, Petr Smýkal, Frederick L. Stoddard
PRIKAZI KNJIGA / BOOK REVIEWS
259 Živko Ćurčić: Oplemenjivanje šećerne repe na rizomaniju / Sugar Beet Breeding on Rhizomania
Nevena Nagl
260 Đuro Gvozdenović: Paprika / Pepper (in Serbian) Žarko Ilin

Conservation and Breeding of Ornamental Lathyrus species 1
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6review article / pregledni rad
Conservation and Breeding of Ornamental Lathyrus Species
Roger Parsons · Aleksandar Mikić
received / primljeno: 19.10.2010. accepted / prihvaćeno: 27.11.2010.© 2011 IFVC
Summary: A considerable number of diverse ornamental forms of Lathyrus odoratus exist which require careful conservation if they are not to be lost. Other ornamental Lathyrus species may have potential for increasing their diversity. A seedbank has been established in the United Kingdom to conserve this diversity and make the material readily available to researchers and growers throughout the world. Con-ventional hybridisation provides an opportunity for the existing diversity in L. odoratus to be extended. Opportunities also exist to create improved decorative forms and new colours in some other species through intra-specifi c hybridisation. Inter-specifi c hybridisation within the genus is in its infancy but early results suggest exciting improvements might be introduced by this method. Such work might also provide benefi ts for comparative genomics in the tribe Fabeae.Key words: conservation, hybridisation, Lathyrus, plant breeding, seedbank, seed production, sweet pea
R. ParsonsRoger Parsons Sweet Peas, Primrose Cottage, Clayton Lane, Brackle-sham Bay, Chichester PO20 8JQ United Kingdome-mail: [email protected]. MikićInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Historical Overview
Although Lathyrus species have been grown since ancient times, their cultivation for ornamental purposes can be traced to the sending of the sweet pea, Lathyrus odoratus, from Sicily to England and the Netherlands in 1699. This species quickly became widely distributed for its scent and decorative fl owers. The typical species has maroon and violet bicoloured fl owers but within 20 years a pink and white bicolour cultivar and a white cultivar existed. By 1800, six colours were known and continued to steadily increase but there is no indication that intentional crossing of cultivars was occurring before 1860. Such novel fl ower colours have led to a perception that L. odoratus has a high mutation rate but there is no evidence to support this.
The few decades from 1860 onwards saw several plant breeders actively crossing cultivars to produce new ones, greatly widening the colour range and improving the size and vigour of earlier colours. One of these, Henry Eckford, coined the term ‘Grandifl ora’ for his Sweet
Peas to promote their improved size compared with earlier material. By 1900, dozens of new cultivars were available (Fig. 1). In addition, other Lathyrus species were being grown as part of a wider interest in discovering the world’s fl ora.
Fig. 1. L. odoratus ‘Dorothy Eckford’ has normal fl ower formSlika 1. Sorta mirisnog grahora Doroti Ekford poseduje običnu veličinu cveta
Genetic Resources / Genetički resursiw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Parsons R, Mikić A2
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
In 1900, a new form of L. odoratus was seen that caused a sensation. It was a cultivar with larger, frilly petals and a longer raceme, making it much more attractive to fl orists than the smaller, plain petals of the Grandifl ora cultivars. This fi rst large, frilly cultivar was named ‘Countess Spencer’ and all subsequent cultivars with this fl ower form have been known as Spencer sweet peas (Parsons 2000). This new fl ower form, together with the discovery of Mendel’s research, led to a frenzy of breeding new sweet peas and a considerable increase in growing sweet peas for cut fl owers and by amateur gardeners. By 1910, California alone was growing around 400 ha of sweet pea seeds and had become the main centre of seed production (Wright 1912). The 20th century saw considerable refi nement of Spencer sweet peas by plant breeders to improve fl ower size, purity of colour, raceme length, and vigour. They also introduced new types having early fl owering characteristics, dwarf and intermediate height, non-tendril (multijugate) leaves, and additional petals. As a result of all this work, the vigour of plants and associated raceme length and fl ower size all increased (Parsons 2011).
Conservation of Lathyrus Species and Cultivars
Cultivation of L. odoratus cultivars has taken place in many parts of the world and remains signifi cant. Perhaps the main activity is in cut fl ower production, often to meet local markets. For example, most northern European countries have at least one producer of cut fl ower sweet peas. Although less popular in the UK nowadays because of their short vase life, they remain popular as a winter cut fl ower crop in Japan. By contrast, few sweet peas are grown by amateur gardeners in Japan but they remain a popular fl ower with UK gardeners. Seed production has become more diversifi ed, with New Zealand rising as an important secondary centre of seed production (Parsons 2009).
A problem of fi eld scale production of L. odoratus seed is that the Spencer fl ower form is recessive to normal form. Records of petal size and plant vigour show that Grandifl ora cultivars, with normal fl ower form, have not changed signifi cantly during the last 100 years of seed generations but Spencer cultivars are frequently seen to revert to clamped keels, either with normal fl ower form and size or with an intermediate size and form. These intermediate fl owers have a clamped keel
and frilly petals and have been called Semi-Grandifl ora type (Parsons 2011) (Fig. 2). When cropping a cultivar on a fi eld scale, it is easy to remove any plants with the wrong fl ower colour but reversion in fl ower form, or other faults that arise, are easily overlooked so that commercial stocks of a cultivar deteriorate relatively quickly (Parsons 2011). This problem is made worse because there is no registration authority for Lathyrus cultivars and they are introduced into cultivation without the rigour that applies, for example, to fi eld pea (Pisum sativum) cultivars.
Fig. 2. Flowers forms in sweet pea: Grandifl ora (Lady Grisel Hamilton), Semi-Grandifl ora (Albutt Blue) and Spencer (Millennium)Slika 2. Oblici cveta mirisnog grahora: grandifl ora (sorta Ledi Grizel Hamilton), polu-grandifl ora (sorta Elbat Blu) i spenser (sorta Milenijum)
Commendable effort has gone into conserving the diversity of forage and grain Lathyrus but there was no readily accessible seedbank conserving ornamental Lathyrus. In the UK and elsewhere, sweet pea commercialisation has been characterised by a relatively short cycle of cultivar introduction, deterioration over several generations and replacement by a new cultivar with the same fl ower colour. Such new cultivars may fi ll the same niche in the marketplace for fl ower colour but each cultivar has its own characteristics, for example adaptation to microclimate and different soils. The fi rst-named author became concerned that cultivars were being lost and that the genepool was being reduced. He had been conserving other Lathyrus species since 1991, but in 1998 established a seedbank to conserve L. odoratus cultivars. Fortunately, Lathyrus seed stores very easily when placed in an air-tight container in an ordinary domestic freezer (Hammett 1979).
In 2005, this collection was recognised as a UK National Collection, trading as Roger

Conservation and Breeding of Ornamental Lathyrus species 3
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
Parsons Sweet Peas. It now consists of around 900 L. odoratus cultivars and around 100 taxa from other Lathyrus species. New accessions from around the world are always welcome. Over 100 stocks of annual Lathyrus, mostly L. odoratus cultivars, are grown each year. These are grown in very small quantities, usually between 10 and 60 plants of each cultivar, so that each plant can be assessed for being true to its original qualities. Unlike fi eld cropping, the emphasis is on the quality of the seed stock rather than the quantity of seed produced. It might be thought that fi eld-scale seed producers should do more to maintain the quality of their seed stocks but such neglect has created a niche market for Roger Parsons Sweet Peas. At the end of each season, fresh seed is returned to the seedbank and the surplus is sold. Customers include seed producers who want a true or improved stock of a cultivar that they can then send to be bulked up for fi eld production. Roger Parsons Sweet Peas also retails direct to amateur gardeners since this provides a higher profi t. A limited selection of cultivars and species is advertised on the website www.rpsweetpeas.co.uk and a mail order catalogue is sent each year to previous UK customers. This provides an opportunity to offer people species or colours that they were unaware existed and to encourage them to try something that is currently not popular. However, all this work is a personal domestic effort which makes it vulnerable to being lost.
Opportunities for Lathyrus Breeding
The seedbank provides access to perhaps the world’s most diverse collection of material for this genus. Not content with conserving the fullest diversity, plant breeding provides an opportunity to increase the diversity. This may be from recognising the qualities of mutations that arise casually, but more importantly a small programme of hybridising takes place. Up to now, this has consisted mainly of introducing new colours into the late Spencer type that are so popular with UK gardeners, or improving the qualities of existing colours. In 2010, work began on breeding programmes for dwarf sweet peas and for early-fl owering sweet peas.
This work is possible because the fl owers of L. odoratus cultivars have self-pollinated by the time they are open. This allows cultivars to be grown alongside each other yet remain true from seed. There is a very small risk of some cross-pollination occurring which is why rigorous reselection of
small batches of a cultivar is the best way to conserve it. In order to hybridise two cultivars, the petals of the seed-bearing fl ower must be unfurled while still in bud at a stage when the gynoecium is suffi ciently mature to receive pollen but the anthers have not yet produced pollen (Fig. 3). The anthers are removed and then pollen from the pollen-bearing parent is applied to the stigma. The petals are then closed and the fl ower is allowed to develop normally, having of course been carefully labelled. Subsequent generations are grown and allowed to self-pollinate until single plant selections result in a seed stock that breeds true for the desired characters (Parsons 2011).
Fig. 3. A sweet pea fl ower ready for hybridizationSlika 3. Cvet mirisnog grahora spreman za ukrštanje
There appears to be considerable potential
for other people to engage in breeding new ornamental Lathyrus. There are several types of sweet pea that are not receiving the attention they merit. Very little work is being carried out on early-fl owering cultivars in the UK, which are not as popular there as the summer varieties. However, they have great popularity in other parts of the world and with commercial cut fl ower growers. Japanese cut fl ower growers are quite active with these but are reluctant to allow their cultivars to become available to their commercial competitors. In addition, there is very limited availability of summer-fl owering Multifl ora cultivars and, to the best of our knowledge, nobody is working on these. Some recent interest has been shown in improving dwarf and intermediate height cultivars, which are most popular in the USA, but there is considerable scope for the plant breeder to work on these.
The everlasting pea, L. latifolius, is a popular garden plant in the UK and is grown as a cut fl ower crop in some countries. Another perennial

Parsons R, Mikić A4
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
garden species is the dwarf spring pea, L. vernus. Both of these have a small number of colours that have arisen through mutation and it seems that their position refl ects that of L. odoratus in 1800. However it is not simply the occurrence of mutations that has led to such diversity in L. odoratus but human intervention is needed to identify and secure desirable mutations. It may be that by conventional intra-specifi c hybridisation, other Lathyrus species can be induced to provide a broader range of colours and that other qualities can be improved, for example fl ower size in L. vernus (Fig. 4).
Fig. 4. L. odoratus ‘Frances Kate’ is a novel variety for 2011 and has Spencer fl ower formSlika 4. Sorta mirisnog grahora Franses Kejt je novitet za 2011. godinu i poseduje spenser oblik cveta
New colours are arising in L. odoratus partly as
a result of inter-specifi c hybridisation. Lathyrus species cannot easily be induced to hybridise but cultivars have now become available that arise from a cross between L. belinensis and L. odoratus, using embryo rescue (Hammett et al. 1994). Hybrid material tends to adopt the morphological features of one parent or another after a few generations but may still introduce novel qualities. Material from this particular cross is still being evaluated but the potential for resistance to powdery mildew (Erysiphe polygoni)
has already been reported (Poulter et al. 2003). Personal observations suggest extended vase life may be possible.
Such plant breeding may also benefi t research on other economically important genera of the tribe Fabeae (Smýkal et al. 2010) which, beside the genus Lathyrus, includes Pisum (pea), Vicia (vetch), Lens (lentil) and Vavilovia (vavilovia), among others. For example, a nice confi rmation of the law on homology series in botanically related species, established by N. I. Vavilov nearly a century ago (Vavilov 1920), is the existence of the orthologous genes in sweet pea (t) and fi eld pea (tl), both controlling the development of just the leafl ets (Hofer et al. 2009) into a so-called ‘tendril-less’, ‘acacia’ or ‘multijugate’ leaf type (Fig. 5). This leaf type in fi eld pea may be benefi cial in breeding for increased forage yields (Mikić et al. 2010).
Fig. 5. Phenotypes of two orthologous genes in two close botanical legume relatives: normal leaves in sweet pea (TT) and fi eld pea (TlTl) and tendril-less leaves in sweet pea (tt) and fi eld pea (tltl)Slika 5. Fenotipovi dva ortologna gena dva bliska botanička srodnika: obični tip lista kod mirisnog grahora (TT) i gajenog graška (TlTl) i bagremasti tip lista kod mirisnog grahora (tt) i gajenog graška (tltl)

Conservation and Breeding of Ornamental Lathyrus species 5
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
Conclusions
A considerable number of diverse ornamental forms of L. odoratus exist which require careful conservation if they are not to be lost. Other ornamental Lathyrus species may have potential for increasing their diversity. A seedbank has been established in the UK to conserve this diversity and make the material readily available to researchers and growers throughout the world. Conventional hybridisation provides an opportunity for the existing diversity in L. odoratus to be extended. Opportunities also exist to create improved decorative forms and new colours in some other species through intra-specifi c hybridisation. Inter-specifi c hybridisation within the genus is in its infancy but early results suggest exciting improvements might be introduced by this method. Such work might also provide benefi ts for comparative genomics in the tribe Fabeae.
References
Hammett K R W (1979): Seed Storage. NSPS Annual 1979: 37-38
Hammett K R W, Murray B G, Markham K R, Hallett I C (1994): Interspecifi c hybridisation between Lathyrus odoratus and L. belinensis. Int. J. Plant Sci. 155: 763-771
Hofer J, Turner L, Moreau C, Ambrose M, Isaac P, Butcher S, Weller J, Dupin A, Dalmais M, Le Signor C, Bendahmane A, Ellis N (2009): Tendril-less regulates tendril formation in pea leaves. Plant Cell 21: 420-428
Mikić A, Kosev V, Gantner R, Mihailović V, Smýkal P, Ambrose M (2010): Diversity and agronomic utility of leaf types in pea (Pisum sativum L.). Book of Abstracts, XXIVth Genetic Days, Brno, Czech Republic, 1-3 September 2010, 53
Parsons R (2000): Early History of the Sweet Pea. In Ball C. (ed.) National Sweet Pea Society Centenary Celebration 1900-2000. National Sweet Pea Society, Stockbridge, UK, 5-19
Parsons R (2009): Ornamental Lathyrus species. Grain Legumes 54: 6-7
Parsons R (2011): Sweet Peas – an Essential Guide. Crowood, Marlborough (in press)
Poulter R, Harvey L, Burritt D (2003): Qualitative resistance to powdery mildew in hybrid sweet peas. Euphytica 133: 349-358
Smýkal P, Kenicer G, Mikić A (2010): Molecular taxonomy of the tribe Fabeae (Vicieae). Abstracts, Vth International Con-gress on Legume Genetics and Genomics, Pacifi c Grove, USA, 2-8 July 2010, 122
Vavilov N I (1920): The law of homology series in genetical mutability. Proceedings, All-Russia Congress on Breeding and Seed Production, 1, Saratov, Russia, 4-13 June 1920, 41-56
Wright W P (1912): A Book about Sweet Peas. Headley Broth-ers, London

Parsons R, Mikić A6
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 1-6
Očuvanje i oplemenjivanje ukrasnih vrsta roda Lathyrus
Rodžer Parsons1 · Aleksandar Mikić2
1Mirisni grahor Rodžera Parsonsa, Čičester, Velika Britanija2Institut za ratarstvo i povrtarstvo, Novi Sad, Srbija
Izvod: Ukrasne vrste roda grahor (Lathyrus) odlikuju se visokom učestalošću mutacija, što je dovelo do nastanka raznovrsnih ukrasnih formi od kojih pojedine zahtevaju pažljive mere očuvanja. U Velikoj Britaniji je osnovana ban-ka semena sa ciljem da očuva diverzitet i učini materijal spremnim za korišćenje istraživačima i uzgajivačima širom sveta. Konvencionalno ukrštanje omogućava proširenje postojećeg diverziteta kod mirisnog grahora (L. odoratus). Postoje mogućnosti da se stvore usavršene ukrasne forme i nove boje kod nekih drugih vrsta putem unutarvrsnog ukrštanja. Međuvrsno ukrštanje unutar roda je još uvek u začetku, ali prvi rezultati nagoveštavaju da se ovim putem mogu načiniti izuzetni pomaci, ne samo u oplemenjivanju ovih vrsta, već i u uporednoj genomici tribusa Fabeae.Ključne reči: banka semena, grahor, Lathyrus, mirisni grahor, očuvanje, oplemenjivanje biljaka, semenska proiz-vodnja, ukrštanje

Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline 7
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14pregledni rad / review article
Oplemenjivanje pšenice na nizak sadržajfi tinske kiseline: stanje i perspektive
Gordana Branković · Desimir Knežević · Dejan Dodig · Vesna Dragičević
primljeno / received: 04.10.2010. prerađeno / revised: 15.11.2010. prihvaćeno / accepted: 29.11.2010.© 2011 IFVC
Izvod: Potreba za oplemenjivanjem pšenice na nizak sadržaj fi tinske kiseline proistekla je iz njene uloge antinutritivnog faktora koji vezujući mineralne elemente (Ca, Zn, Fe, Mn, Cu, kao i P) dovodi do njiho-vog nedovoljnog iskorišćavanja. Neiskorišćeni fosfor u kompleksu sa fi tinskom kiselinom se preko lanca ishrane nepreživara (živina, svinje, ribe) izlučuje u spoljašnju sredinu i uzrokuje zagađivanje vodenih ekosistema. Razvijene su brojne indirektne (spektrofotometrijske) i direktne (HPLC - High Performance Liquid Chromatography) metode za brzo i pouzdano utvrđivanje sadržaja fi tinske kiseline u zrnu pšenice. Brojna istraživanja u svetu su pokazala da se sadržaj fi tinske kiseline kreće od jedan do nekoliko pro-cenata suve mase semena i 50% do 85% ukupnog fosfora u semenu. Utvrđena je značajna genetička varijabilnost sadržaja fi tinske kiseline i fi tatnog fosfora u zrnu sorata i linija pšenice u različitim uslovima spoljašnje sredine. Oplemenjivanjem je dobijen i mutant pšenice (Triticum aestivum L.) Js-12-LPA, za osobinu niskog sadržaja fi tinske kiseline (Low phytic acid).Ključne reči: fi tinska kiselina, mutanti, oplemenjivanje, pšenica
G. Branković Poljoprivredni fakultet Univerziteta u Beogradu, Nemanjina 6, 11000, Beograde-mail: [email protected]. KneževićPoljoprivredni fakultet u Lešku, Jelene Anžujske b.b., 38228, Zubin PotokD. Dodig · V. DragičevićInstitut za kukuruz ”Zemun Polje”, Slobodana Bajića 1, 11080 Zemun
Uvod
Rastući interes za istraživanja fi tinske kiseli-ne žitarica i leguminoza rezultat je njene uloge antinutritivnog faktora. Fitinska kiselina kao polivalentni anjon stvara helate sa mineralnim elementima i proteinima, onemogućavajući nji-hovo potpuno iskorišćavanje. Ishrana bogata fi -tinskom kiselinom značajno smanjuje apsorpciju važnih mikronutrijenata (kalcijum, cink, gvožđe, mangan i bakar) koje ljudi i nepreživari (živina, svinje, ribe) u vidu mešovitih soli izlučuju, do-vodeći do njihovog ozbiljnog defi cita, naročito kod siromašnih i zemalja u razvoju (Reicwald & Hatzack 2008, Khan et al. 2007, Guttieri et al. 2004, Lönnerdal 2002, García-Estepa et al. 1999, Tabekhia & Donelly 1982, Nahapetian & Bassiri 1976, Lolas et al. 1976).
U SAD i Evropskoj Uniji ozbiljni napori se ulažu da se smanji zagađivanje vodenih ekosiste-ma fosforom koji se neiskorišćen u kompleksu sa
fi tinskom kiselinom izlučuje životinjskim ekskre-mentima. Usled neiskorišćenosti unešenog fosfo-ra iz žitarica neophodno je dodavati suplemente neorganskog fosfora ili granule enzima fi taze u stočnu hranu, radi postizanja optimalne produk-cije mesa. Granule enzima fi taze se industrijski proizvode korišćenjem gljivičnih gena za fi tazu, što značajno uvećava troškove proizvodnje, dok samo 50% fosfora vezanog u solima fi tinske ki-seline postaje dostupan (Reichwald & Hatzack 2008, Centeno et al. 2003, Raboy 2001a, Wodzin-ski & Ullah 1996, Greiner et al. 1998, Cromwell et al. 1993).
Pšenica doprinosi sa jednom petinom uku-pnom svetskom unosu kalorija ishranom (FAO, 2009). Gaji se na više od 200 milion hektara obra-divih površina od 67º N u Skandinaviji i Rusiji do 45º u Argentini uključujući i visinske delove u tropima i subtropima (Dodig 2010, Knežević et al. 2008a, Feldman 1995). Prosečan prinos u svetu je 2,8 t ha-1 ali je visoko varijabilan po drža-vama i regionima, i u Evropskoj Uniji je u peri-odu 2005-2007. iznosio 5 t ha-1 dok je u Srbiji za isti period iznosio 3,4 t ha-1 (Dodig 2010). Danas
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Rad je rezultat projekta TR20097 „Izučavanje genotipova strnih žita i oplemenjivanje na poboljšanje rodnosti, kvaliteta i adaptivne sposob-nosti“ Ministarstva za nauku i tehnološki razvoj Republike Srbije / This article is a result of the project TR20097 funded by the Mini-stry of Science and Technological Development of the Republic of Serbia

Branković G i sar.8
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
je 95% pšenice koja se gaji u svetu heksaploidna hlebna pšenica, dok preostalih 5% čini tetraplo-idna durum pšenica (Dodig 2010).
Visok sadržaj fi tinske kiseline kod pšenice dovodi najčešće do defi cita mineralnih eleme-nata (Fe, Zn, Ca) i najviše se odražava na rast dece i zdravlje trudnica u zemljama sa niskom kupovnom moći stanovništva. Posledice po zdravlje ljudi koje nastupaju usled nedovoljnog iskorišćavanja ovih mineralnih elemenata su: anemije, kardiovaskularne bolesti, tkivna hi-poksija, poremećaj pažnje i memorije, oslablje-ne motorne sposobnosti (Walter et al. 1997, Prasad 1996).
Pored negativnih antinutritivnih svojstava, fi -tinska kiselina ima i pozitivnih dejstava jer vezuje gvožđe i smanjuje formiranje hidroksilnih radika-la u debelom crevu, pokazujući antioksidativno i antikancerogeno dejstvo (García-Estepa et al. 1999, Harland & Morris 1995, Graf et al. 1987). Pokazano je da fi tinska kiselina povećava vigor klijanaca i usporava razvoj afl atoksina u zrnu pšenice (Ortiz-Monasterio et al. 2007, Tang et al. 2008).
Preporuke SZO i FAO jesu da se tržište u bu-dućnosti snabdeva sortama pšenice sa niskim sa-držajem fi tinske kiseline (low phytic), ali i sortama sa visokim sadržajem fi tinske kiseline (high phytic). Korišćenje oba tipa sorta pšenice će zavisiti od toga da li će služiti za stočnu hranu, za poveća-nje iskorišćavanja mikronutrijenata ili sprečavanje oksidativnog stresa i kancera.
Fitinska kiselina kod pšenice - uloga i fi zička pozicija u semenu
Fitinska kiselina (myo-inositol (1,2,3,4,5,6)-hexakisphosphate (InsP6) predstavlja jedinjenje široko rasprostranjeno u semenu biljaka i najpo-znatija je kao rezervna forma fosfora (P) u seme-nu. Obuhvata jedan do nekoliko procenata suve mase semena i 50% do 85% ukupnog fosfora u semenu pšenice (Raboy 2001a, Guttieri et al. 2004, Hídvégi & Lásztity 2002, Khan et al. 2007, Yenagi & Basarkar 2007, Dintzis et al. 1992, Na-hapetian & Basiri 1976). Količina fosfora sin-tetisana u formi InsP6 predstavlja više od 50% unešenog fosfora u agroekosisteme mineralnim đubrenjem na godišnjem nivou. Najveći deo fi -tinske kiseline je uskladišten u formi soli sa kali-jumom, magnezijumom, kalcijumom, gvožđem i cinkom (fi tini) u mikrovakuolama i proteinskim telima-globoidima (Raboy 2001b, Svečnjak et al. 2007, Williams 1970).
Dokazane su višestruke uloge fi tinske kiseline u obavljanju važnih ćelijskih funkcija: glavni je
izvor inozitola u metaboličkim putevima inozitol fosfata (Safrany et al. 1999); kao ligand sekundar-nih „messenger” molekula (Sasakawa et al. 1995); učešće u popravci dvolančanih DNK prekida (Hanakahi et al. 2000); transportu RNK iz jedra (York et al. 1999); učešće u ATP metabolizmu (Safrany et al. 1999) i fi ziološkom odgovoru sto-minih ćelija na ABA (Lemtiri-Chlieh et al. 2000).
U toku klijanja semena fi tine razlaže enzim endogena fi taza, oslobađajući neorganski fosfor, myo-inozitol i mineralne elemente koji se koriste za rast i razvoj klijanca. InsP6 se akumulira i u drugim biljnim tkivima i organima kao što su po-len, krtole, koren, zbog čega je organski fosfor u zemljištu i najprisutniji u obliku derivata fi tin-ske kiseline. Fitinska kiselina i njeni pirofosfatni derivati imaju ulogu u obnovi ATP molekula ne-ophodnih za održavanje bazalnog metabolizma u najranijim stadijumima klijanja pre formiranja mitohondrijalne membrane (Raboy 2001a, Wi-lliams 1970).
Utvrđeno je na osnovu istraživanja na Triticum vulgare cv. Insignia da se fi tinska kiselina nalazi u klici, skutelumu, aleuronskom omotaču i spo-ljašnjim frakcijama perikarpa, dok se ne nalazi u frakcijama endosperma (Guttieri et al. 2004, Hídvégi & Lásztity 2002, Febles et al. 2001, Williams 1970). Fitinska kiselina se akumulira u aleuronu pšeničnog zrna koje je prošlo nor-malan tok razvoja posle 28 dana od početka kli-janja, i posle 23 dana od početka klijanja kod zrna izloženog stresnim uslovima. Sinteza fi tina je deo reakcije na stres u spoljašnjoj sredini i deo opšteg mehanizma za usporavanje metabolizma pre nastupanja dormantnosti, jer stvaranje he-lata sa metalnim katjonima kontroliše nivo fos-fotransferaza od kojih zavisi ćelijski energetski metabolizam.
In vitro istraživanja su pokazala da fi tinska ki-selina i proteini formiraju komplekse od kojih su mnogi nerastvorljivi i biološki nedostupni u normalnim fi ziološkim uslovima (Knežević et al. 2006, Febles et al. 2001). Proteolitički enzimi su manje efi kasni u razlaganju proteinskih komplek-sa sa fi tinskom kiselinom nego slobodnih protei-na (Cheryan 1980, Fox & Tao 1989).
Metode za utvrđivanje sadržaja fi tinske kiseline
Najšire korišćene metode za utvrđivanje sadr-žaja fi tinske kiseline kod žitarica su modifi kaci-je procedura koje su razvili Heubner & Stadler (1914) i predstavljaju indirektne metode koje uključuju precipitaciju kompleksa sa gvožđem (Fe3+) na niskim pH vrednostima i spektrofoto-

Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline 9
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
metrijsku kvantifi kaciju fosfora, Fe+3, inozitola u precipitatu ili analizu preostalog gvožđa u su-pernatantu (Sandberg 1995, Vaintraub & Lap-teva 1988, Oberleas 1983, García-Villanova et al. 1982, Latta & Eskin 1980, Oberleas 1971). Direktne analitičke metode koje su nezavisne u odnosu na formiranje Fe3+ kompleksa uklju-čuju primenu HPLC metoda (High-performance liqid chromatography) (Larson et al. 2000, Raboy et al. 2000, Dorsch et al. 2003, Graf & Dintzis 1982a), HPLC koja uključuje jonoizmenjivačku proceduru (Dorsch et al. 2002, Graf & Dint-zis 1982b, Tangendjaja et al. 1980, Harland & Oberleas 1977), “capillary isotachophoresis” teh-nike (Blatny et al. 1995, Kikunaga et al. 1985), modifi kovane hromatografi je jonskih parova (Lehrfeld 1989).
Većina metoda za fotometrijsko utvrđivanje fi -tata je zasnovana na indirektnim merenjima. Vrši se ekstrakcija sa kiselinama HCl, H2SO4 ili HCl3 koju sledi precipitacija sa Fe3+. Kisela ekstrakcija je poželjna jer sirovi materijal, kao što je pšeni-ca, može da sadrži visoki nivo endogene fi taze koja razgrađuje fi tin (Oberleas & Harland 1986, Reicwald & Hatzack 2008). Neistaloženi Fe3+ joni se utvrđuju spektrofotometrijski i razlika u kon-centraciji jona istaloženog i neistaloženog gvožđa se koristi za izračunavanje koncentracije fi tinske kiseline. Jedan od najpoznatijih fotometrijskih metoda za određivanje koncentracije fi tinske ki-seline je protokol koji su razvili Haug & Lantzsch (1983) koji koristi 2.2’-bypyridine kao komplek-sirajući agens za kvantifi kaciju Fe3+, ali koji je do sada korišćen samo na kukuruzu. Modifi kaciju metoda Haug & Lantzch za utvrđivanje sadržaja fi tinske kiseline kod pšenice, soje i kukuruza su dali Reichwald & Hatzack (2008) prilagođavajući ekstrakciono vreme na 45 minuta i temperaturu na 100°C i smanjujući količinu toksične tioglikol-ne kiseline sa 1% v/v na 0.13% v/v.
Autori Febles et al. (2002) su primenili kom-pleksometrijsko određivanje sadržaja fi tinske kiseline zasnovano na titraciji Fe3+ sumpor-sali-cilnom kiselinom po metodi Garcia-Villanova et al. (1982).
Metoda HPIC (High performance ion chromatograp-hy) se koristi kao specifi čna za određivanje inozi-tol fosfata (Pontoppodian et al. 2007, Camire & Clydesdale 1982) i kao referentna metoda za po-ređenje rezultata sa indirektnim metodama, kao što je modifi kacija Haug & Lantzsch (1983) (Rei-cwald & Hatzack 2008). Precizno određivanje sa-držaja fi tinske kiseline, na koju ne utiče proteinski sadržaj, omogućeno je ultrafi ltracijom na mem-brani (30kDa) i ispiranjem proteinskih anjona sa kolone pre inozitol fosfata i fi tinske kiseline.
Genetička varijabilnost sadržaja fi tinske kiseline linija i sorata pšenice
Autori Yenagi & Basarkar (2008) su utvrdili da su sorte hlebne pšenice u severnoj Karnataki (In-dija) imale prosečan sadržaj fi tinske kiseline od 2,39 g 100g-1 ± 0,02 g 100g-1, sorte durum pšenice 1,93 g 100g-1 ± 0,002 g 100g-1 i sorte dicoccum pše-nice 1,90 g 100g-1 ± 0,04 g 100g-1. Autori Hídvégi & Lásztity (2002) su dobili da sadržaj fi tinske ki-seline za sorte durum pšenice iznosi 0,72 g 100g-1 i od 0,85 g 100g-1 do 0,93 g 100g-1 za sorte hlebne pšenice. Proučavajući odnos fi tinske kiseline i mi-neralnih elemenata kod 12 sorata hlebne pšenice autori Nahapetian & Bassiri (1976) su utvrdili da je sadržaj fi tatnog fosfora od 109 mg 100g-1 do 318 mg 100g-1. Tabekhia & Donnelly (1982) su za šest sorata durum pšenice utvrdili da je sadržaj fi tinske kiseline izražen kao procenat suve mase u opsegu 0,95% do 1,43%, kao i da sorte pšeni-ce belog zrna imaju niži fi tinski sadržaj od sorata pšenice crvenog zrna, sa čim su u saglasnosti re-zultati Nahapetian & Bassiri (1976).
Lolas et al. (1976) su za 38 sorata i linija hleb-ne i durum pšenice utvrdili da se sadržaj fi tinske kiseline, izražen kao procenat suve mase zrna, kreće od 0,62% do 1,35%. Khan et al. (2007) su analizom 66 mutanata/ sorata hlebne pšenice do-bili da je sadržaj fi tinske kiseline u zrnu od 0,98% do 2,17% izražen kao procenat suve mase zrna. Svečnjak et al. (2007) su utvrdili da prosečan sadržaj fi tatnog fosfora za sorte hlebne pšenice Marija, Žitarka i Renan iznosi 3,84 g kg-1 što čini 82% ukupnog sadržaja fosfora u zrnu pšenice. Sa navedenim rezultatima su u saglasnosti nalazi Syl-tie & Dahnke (1983) i Zebarth et al. (1992), dok su Liu et al. (2006) dobili više vrednosti, u opsegu 5,16-9,87 g kg-1. Tang et al. (2008) su utvrdili da je sadržaj fi tatnog fosfora za 43 sorte pšenice gaje-nih u Jinanu (Kina) iznosio 1.059 mg kg-1. Bassiri & Nahapetian (1977) su utvrdili da sadržaj fi tata u ukupnom fosforu zrna pšenice varira od 38% do 77% za sorte gajene u sušnim uslovima i od 73% do 94% za sorte gajene u uslovima navod-njavanja.
Batten (1986) je u ogledima sa 17 genotipova pšenice, koje su imale različite nivoe ploidnosti, utvrdio da je ukupni fosfor u zrnu 0,14% do 0,71% suve mase zrna, i da je bio najviši kod di-ploidnih genotipova, a najmanji kod heksaploid-nih genotipova (Djukić et al. 2008). Utvrdili su i da je gotovo potpuno korelisan (r = 0.99) sa sadržajem fi tatnog fosfora, dok su Nahapetian & Bassiri (1976) dobili koefi cijent korelacije r = 0.93 i Lolas et al. (1976) su utvrdili da je r = 0.97. Stoga je selekcija za niži sadržaj ukupnog fosfora

Branković G i sar.10
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
predložena kao način da se ostvare niže fi tatne koncentracije u zrnu. Međuzavisnost sadržaja fi tinske kiseline sa agronomskim svojstvima bi bilo poželjno ispitati da bi se utvrdila mogućnost primene indirektne selekcije na sadržaj fi tinske kiseline (Dodig et al. 2008b).
Svečnjak et al. (2007) su utvrdili statistički zna-čajan uticaj genotipa (P < 0.05) na sadržaj fosfora u zrnu kao i statistički značajnu (P < 0.05) kore-laciju (r = - 0.47) između ukupnog fosfora i mase hiljadu zrna. Khan et al. (2007) su zaključili da je sadržaj fi tinske kiseline u zrnu hlebne pšenice statistički značajno korelisan sa širinom semena (P < 0.001), debljinom semena (P < 0.01) i zapre-minom semena (P < 0.05). Analizom varijanse su utvrdili da su genotip, sredina i interakcije geno-tip x sredina visoko statistički značajno uticali na sadržaj fi tinske kiseline u zrnu, sa najvećim utica-jem lokaliteta. Predloženo je da je pravilnu proce-nu sortimenta za sadržaj fi tinske kiseline potreb-no obavljati u višelokacijskom ogledu (Dodig et al. 2007, Dodig et al. 2008a).
Najveći deo fi tata u pšeničnom zrnu se nalazi u aleuronskom omotaču i tokom mlevenja većina aleuronskih ćelija ostaje sa česticama perikarpa, pa je to razlog što je najveći deo fi tata koncentri-san u frakcijama mekinja (Tang et al. 2008) i do 5 puta više nego u brašnu (Zečević et al. 2007). Celo pšenično zrno sadrži 0,3% do 0,4% fi tata dok mekinje sadrže do 5% (O’Dell et al. 1972). García-Estepa et al. (1999) su utvrdili da je sadr-žaj fi tinske kiseline u brašnu dobijenim od hlebne pšenice od 2,94 mg g-1 do 4,04 mg g-1, dok je za durum pšenicu iznosio 9,41 mg g-1. U mekinja-ma je bio 30,34 mg g-1 do 36,42 mg g-1 za hleb-nu pšenicu i 24,96 mg g-1 za durum, dok je griz hlebne pšenice sadržao od 20,18 mg g-1 do 26,73 mg g-1 i 9,87 mg g-1 kod duruma. Autori Febles et al. (2001) su utvrdili da je koncentracija fi tinske kiseline u rafi niranom pšeničnom brašnu iznosila 2-4 mg g-1, dok je za brašno dobijeno od celog zrna bila 6-10 mg g-1. Camire & Clydesdale (1982) su utvrdili da sadržaj fi tinske kiseline kod mekinja dobijenih od durum pšenice crvenog zrna iznosi 68,8 ± 1,85 mg g-1, dok je kod mekinja dobijenih od hlebnih pšenica belog zrna iznosila 50,27 ± 1,45 mg g-1.
Autori Dintzis et al. (1992) su utvrdili da je na sadržaj fi tinske kiseline u frakcijama mekinja 15 sorata meke pšenice prevashodno uticala sredi-na i da je sadržaj fi tinske kiseline u mekinjama korelisan sa parametrima mlinskog kvaliteta: pro-centom ekstrakcije brašna (EXI), endospermnim separacionim indeksom (ESI) i sa drobljivošću.
Na sadržaj fi tinske kiseline mlinskih frakcija pšenice najviše utiče tip frakcije, ali je u ANOVA
utvrđen i statistički značajan (P < 0.01) uticaj lo-kaliteta, genotipa, interakcije genotip x lokalitet i lokalitet x mlinska frakcija (Tabekhia & Donnelly, 1982). Tang et al. (2008) su utvrdili statistički zna-čajan (P < 0.001) uticaj mlinske frakcije i statistič-ki značajan (P < 0.01) uticaj genotipa u ANOVA za sadržaj fi tatnog fosfora.
Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline
S obzirom da je fi tinska kiselina rezervna for-ma fosfora u zrnu, ukupni fosfor i fi tinska kise-lina su u visokoj, pozitivnoj korelaciji. Spoljašnja sredina i genotip su faktori koji redukuju ukupni sadržaj fosfora, dovodeći čak do snižavanja sa-držaja fi tinske kisline (Dragičević et al. 2010, Knežević 2007a, Raboy 2001a). Proučavajući dve populacije ozime pšenice (Triticum aestivum L.) au-tori Raboy et al. (1991) su utvrdili da je sadržaj fi tinske kiseline statistički značajno korelisan sa ukupnim sadržajem fosfora (r = 0.93 i r = 0.96 u populacijama 1 i 2) kao i sa sadržajem proteina (r = 0.65 i r = 0.87). Navedeno ukazuje da će selek-cija na nizak sadržaj fi tinske kiseline dovesti do neželjenog smanjenja ukupnog fosfora i proteina u zrnu. Preveliko sniženje sadržaja fi tinske kiseli-ne ne bi bilo dobro jer je njena uloga kao helatora mineralnih elemenata važna za metaboličku rav-notežu u semenu pre klijanja (Mladenović-Drinić et al. 2009, Perić et al. 2009).
Dobijen je mutant pšenice (Triticum aestivum L.) niskog sadržaja fi tinske kiseline i označen je kao Js-12-LPA (Guttieri et al. 2004). Js-12-LPA ho-mozigoti imaju zrna u kojima je fosfor fi tinske kiseline predstavljen sa 48,2% ukupnog fosfora zrna u poređenju sa nemutantnim (wild type WT) genotipom Js-12-WT, kod koga je sadržaj fi tatnog fosfora u odnosu na ukupni 74,7%. Neorganski fosfor je povećan sa 9,1% u Js-12-WT na 50,1% u Js-12-LPA. Svojstvo niskog sadržaja fi tinske ki-seline je promenilo raspodelu ukupnog fosfora u zrnu, povećavajući njegov sadržaj u centralnom endospermu i smanjujući sadržaj u mekinjama. Sadržaj fi tinske kiseline u mekinjama je povećan za 43%. Praćenje nasleđivanja analiziranjem F2 i F4:6 familija (Knežević et al. 2008b) je utvrdilo da su u nastanak mutantnog fenotipa za nizak sadr-žaj fi tinske kiseline uključena dva gena.
Guttieri et al. (2006a) su izveli oglede 2003. i 2004. na dva lokaliteta koji su uključili WT i LPA genotipove jare pšenice: tvrdih i crvene boje zrna, tvrdih i bele boje zrna i mekih bele boje zrna. LPA genotipovi jarih sorata pšenice sa crvenom bojom zrna su imali zakasneli razvoj i redukovan prinos zrna za 8% do 25% u prinosnijoj sredini,

Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline 11
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
delom zbog redukovane veličine zrna od 3 mg po zrnu. Kod LPA genotipova jare pšenice tvr-dih i bele boje zrna, nisu bile zabeležene razlike u razvoju zrna i prinosu u odnosu na WT geno-tipove, ali su u prinosnijoj sredini LPA genotipo-vi imali manja zrna za 2,0-2,4 mg po zrnu. LPA genotipovi meke pšenice bele boje zrna, su se razvili ranije i prinos im je bio redukovan za 20% do 24% u prinosnijoj sredini, dok su im zrna bila teža i šira u odnosu na WT genotipove. Odsustvo ujednačenih efekata lpa mutacija kroz tri genetič-ki različita materijala vodi zaključku da se štetni efekti genotipova sa svojstvom niskog sadržaja fi tinske kiseline mogu popraviti oplemenjivanjem (Knežević 2007b).
LPA genotipovi tvrde pšenice nisu imali štetne efekte na koncentraciju proteina u brašnu, spo-sobnost razvoja gasova i zapreminu hleba (Gutti-eri et al. 2006b). Utvrđeno je povećanje od 0,93 g kg-1 pepela u brašnu kod LPA genotipova u odnosu na WT genotipove, ali to nije uticalo na boju rezanaca (L* = 86,8-87,5) koja je bila slična boji rezanaca tvrdih WT genotipova bele boje zrna (L* = 86,1-87,9). Kod LPA genotipova jarih sorata pšenice crvene i bele boje zrna, je došlo do preraspodele mineralnih elemenata iz mekinja u endosperm, dovodeći do nepoželjne karakteri-stike kvaliteta brašna-povećanog sadržaja pepela u brašnu (Knežević et al. 2010). LPA genotipovi meke jare pšenice su imali viši sadržaj Na2CO3 i saharoze u brašnu (za 31 g kg-1 i 43 g kg-1) u odnosu na WT genotipove, što je ukazalo da je kod LPA genotipova došlo do većeg oštećenja skroba i pentozana pri mlevenju u odnosu na WT genotipove. Farinografski tolerancioni in-deks i apsorpiona moć su za tvrde sorte hlebne pšenice crvene boje zrna, bili nepromenjeni kod LPA genotipova u odnosu na WT. Promena re-oloških karakteristika nije imala štetne posledice na kvalitet brašna.
Guttieri et al. (2007) su povratnim ukrštanjem uneli mutantni lpa1-1 fenotip u sortu jare tvrde pšenice crvene boje zrna-Grandin. WT i LPA familije srodnika su gajene tokom dve godine i praćene su osobine kvaliteta. Sadržaj neorganskog fosfora (Pi) u LPA zrnima bio je 3,4 puta veći u od-nosu na sadržaj u WT zrnima. Koncentracija fi tin-ske kiseline u LPA zrnima bila je snižena za 65% u odnosu na koncentraciju u WT zrnima. Stoga je i sadržaj Pi u brašnu od LPA genotipova bio 3-4 puta veći, dok je ukupni fosfor bio uvećan za 20% u odnosu na brašno dobijeno od WT genotipova. Koncentracija Mg je u brašnu dobijenom od LPA genotipova bila za 20% veća, dok su koncentracije mineralnih elemenata u mekinjama bile identične i kod LPA i WT genotipova. Povećan sadržaj P i Mg je doveo do porasta udela pepela u brašnu sa 3,86 g kg-1 kod WT genotipova na 4,38 g kg-1 kod LPA genotipova. Koncentracije proteina kod obe familije srodnika su bile slične, ali su farinografsko vreme razvoja i maksimalna konzistencija bili veći kod brašna dobijenom od LPA genotipova.
Zaključak
Istraživanja genetičke varijabilnosti sadržaja fi tinske kiseline pšenice i rad na stvaranju sorata pšenice niskog sadržaja fi tinske kiseline postaju sve aktuelnija. Oplemenjivačke napore je potreb-no usmeriti na optimalno snižavanje sadržaja fi tin-ske kiseline u zrnu da bi se nutritivna svojstva zrna poboljšala i smanjila zagadjenost vodenih ekosi-stema viškom fosfora. Sniženje sadržaja fi tinske kiseline zrna ne bi trebalo da bude preveliko jer fi tinska kiselina pokazuje i pozitivna svojstva kao antioksidans i antikancerogeni agens. Način za ostvarivanje tog cilja je primena mutacija i metoda povratnog ukrštanja, kao i rekurentne selekcije za popravku agronomskih osobina koje se introgre-sijom mutiranih gena delom pogoršavaju.

Branković G i sar.12
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
Literatura
Bassiri A, Nahapetian A (1977): Differences in concentrations and interrelationships of phytate, phosphorus, magnesium, calcium, zinc, and iron in wheat varieties grown under dry land and irrigated conditions. J. Agric. Food Chem. 25: 1118-1122
Batten G D (1986): Phosphorus fractions in the grain of diplo-id, tetraploid and hexaploid wheat grown with contrasting phosphorus supplies. Cereal Chem. 63: 384-387
Blatny P, Kvasnicka F, Kendler E (1995): Determination of phytic acid in cereal grains, legumes and feeds by capillary isotachophoresis. J. Agric. Food Chem. 43: 129-133
Camire A L, Clydesdale F M (1982): Analysis of Phytic Acid in Foods by HPLC. J Food Sci. 47: 575-578
Centeno C, Viveros A, Brenes A, Lozano A, De La Cuadra C (2003): Effect of several germination conditions on total P, phytate P, phytase, acid phosphotase activities and inositol phosphate esters in spring and winter wheat. J. Agric. Sci. 141: 313-321
Cheryan M (1980): Phytic acid interactions in food systems. CRC Critical Reviews in Food Science and Nutrition 13: 296-335
Cromwell G L, Stahly T S, Coffey R D, Monegue H J, Randolph J H (1993): Effi cacy of low-activity, microbial phytase in im-proving the bioavailability of phosphorus in corn-soybean meal diets for pigs. J. Anim. Sci. 71: 1831-1840
Dintzis F R, Lehrfeld J, Nelsen T C, Finney P L (1992): Phytate content of soft wheat brans as related to kernel size, culti-var, location and milling and fl our quality parameters. Cereal Chem. 69: 577-581
Đukic N, Knezevic D, Zečević V (2008): Genetic determina-tion of technological quality in Triticum durum. Periodicum Biologorum 110: 285-289
Dodig D, Zorić M, Knežević D, Dimitrijević B, Šurlan-Momi-rović G (2007): Assessing wheat performance using envi-ronmental information. Genetika 39: 413-425
Dodig D, Zoric M, Knezevic D, King SR, Surlan-Momirovic G (2008a): Genotype x environment interaction for wheat yield in different drought stress conditions and agronomic traits suitable for selection. Aust. J. Agric. Res. 59: 536-545
Dodig D, Zorić M, Mitić N, Nikolić R, Šurlan-Momirović G (2008b): Tissue culture and agronomic traits relationship in wheat. Plant Cell, Tissue and Organ Culture 95: 107-114
Dodig D (2010): Wheat breeding for drought resistance. Druš-tvo genetičara Srbije. Beograd
Dorsch J A, Cook A, Young K A, Anderson J M, Bauman A T, Volkmann C J, Murthy P P N, Raboy V (2003): Seed phosphorus and inositol phosphate phenotype of barley low phytic acid genotypes. Phytochemistry 62: 691-706
Dragičević V, Simić M, Stefanović L, Sredojević S, Dumano-vić Z (2010): The alteration of phytic phosphorus content in maize inbred lines, caused by post-emergence herbicides. Proceedings of 3rd International Scentifi c/Professional Con-ference “Agriculture in Nature and Environment Protection”, 31st May – 2nd June, 2010, Vukovar, Croatia, 59-64
Febles C I, Arias A, Hardisson A, Rodríguez-Alvarez C, Sierra A (2002): Phytic acid level in wheat fl ours. J. Cereal Sci. 36: 19-23
Feldman M (1995): Wheats. In: Smartt J., Simmonds N.W. (eds.) Evolution of Crop Plants. Longman Scientifi c and Techni-cal, Harlow, UK, 185-192
Fox M R S, Tao S H (1989): Antinutritive effects of phytate and other phosphorilated derivatives. Nutrition Toxicology 3: 59-62
García-Villanova R, García-Villanova R J, Ruiz de Lope C (1982): Determination of phytic acid by complexometric titration of excess iron (III). Analyst. 107: 1503-1506
García-Estepa R M, Guera-Hernández E, García-Villanova B (1999): Phytic acid content in milled cereal products and bre-ads. Food Res. Int. 32: 217-221
Graf E, Dintzis F R (1982a): High-performance liquid chro-matographic method for the determination of phytate. Anal. Biochem. 119: 413
Graf E, Dintzis F R (1982b): Determination of Phytic Acid in Foods by High-Performance Liquid Chromatography.J. Agric. Food Chem. 30: 1094-1097
Graf E (1987). Phytic acid: a natural antioxidant. J. Biol. Chem. 262: 11647-11650
Greiner R, Konietzny U, Jany K D (1998): Purifi cation and pro-perties of a phytase from rye. J. Food Biochem. 22: 143-161
Guttieri M, Bowen D, Dorsch J A, Raboy V, Souza E (2004): Identifi cation and characterization of a low phytic acid whe-at. Crop Sci. 44: 418-424
Guttieri M J, Peterson K M, Souza E J (2006a): Agronomic performance of low phytic acid wheat. Crop Sci. 46: 2623-2629
Guttieri M J, Peterson K M, Souza E J (2006b): Milling and baking quality of low phytic acid wheat. Crop Sci. 46: 2403-2408
Guttieri M J, Peterson K M, Souza E J (2007): Nutritional and baking quality of low phytic acid wheat. In: Buck HT et al. (eds.), Wheat production in stressed environments, Springer, 487-493
Hanakahi L A, Bartlet-Jones M, Chappell C, Pappin D, West SC (2000): Binding of inositol phosphate to DNK-PK and stimulation of double-strand break repair. Cell 102: 721-729
Harland B F, Oberleas D (1977): A modifi ed method for phyta-te analysis using an ion-exchange procedure-application to texture vegetable protein. Cereal Chem. 54: 827
Harland B F, Morris E R (1995): Phytate: a good or a bad food component? Nutrition Res. 15: 733-754
Haug W, Lantzsch H J (1983): Sensitive method for the rapid determination of phytate in cereals and cereal products. J. Sci. Food Agric. 34: 1423-1426
Heubner W, Stadler H (1914): On a titration method of deter-ming phytate. Biochem. Z. 64: 422-437
Hídvégi M, Lásztity R (2002): Phytic acid content of cereals and legumes and interaction with proteins. Periodica Polytechni-ca Ser. Chem. Eng. 46: 59-64
Khan A J, Ali A, Azam F I, Zeb A (2007): Identifi cation and isolation of low phytic acid wheat (Triticum aestivum L.) inbred lines / mutants. Pak. J. Bot. 39: 2051-2058
Kikunaga S, Takashi M, Huzisige H (1985): Accurate and sim-ple measurment of phytic acid contents in cereal grains. Plant Cell Physiol. 26: 1323-1330
Knežević D, Mićanović D, Zečević V, Madić M, Paunović A, Djukić N, Šurlan-Momirović G, Dodig D., Urošević D (2006): Oplemenjivanje u funkciji obezbedjenja semena bi-ološki vredne hrane. U: D. Knežević (ured.), Unapredjenje poljoprivredne proizvodnje na Kosovu i Metohiji. Poljopri-vredni fakultet u Prištini, Lešak, 71-87
Knežević D (2007a): Oplemenjivanje pšenice osnova održivog razvoja. U: K. Konstatinov i V. Andjelković (ured.) Nauka - osnova održivog razvoja. Društvo genetičara Srbije, Beo-grad, 241-265
Knežević D (2007b): Osnovne metode u oplemenjivanju i se-menarstvu biljaka. Foto-oko, Novi Sad
Knežević D, Đukić N, Zečević V, Madić M, Paunović A, Dodig D, Knežević J, Branković G (2008a): Varijabilnost osobina semena pšenice (Triticum aestivum L.). Zbornik abstrakata sa Petog naučno-stručnog simpozijuma iz selekcije i semenar-stva Društva selekcionara i semenara Republike Srbije, Vr-njačka Banja, 25-28. maj, 57
Knezevic D, Djukic N, Paunovic A, Madic M (2008b): Variabi-lity of harvest index and yield components in wheat (Triticum aestivum L.). Cereal Res. Commun. 36: 1151-1154

Oplemenjivanje pšenice na nizak sadržaj fi tinske kiseline 13
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
Knežević D, Dodig D, Kandić V, Branković G, Prodanović S, Šurlan-Momirović G. (2010): Tehnološki kvalitet sorti pše-nice Instituta za kukuruz “Zemun polje”. Zbornik izvoda sa VI naučno-stručnog simpozijuma iz selekcije i semenar-stva Društva selekcionara i semenara Republike Srbije, 17-21.05.2010. Vršac, Srbija, 91
Larson S R, Rutger J N, Young K A, Raboy V (2002): Isolation and genetic mapping of a non-lethal rice (Oryza sativa L.) low phytic acid mutation. Crop Sci. 40:1397-1405
Latta M, Eskin M (1980): A simple and rapid colorimetric met-hod for phytine determination. J. Agric. Food Chem. 28: 1308-1311
Lehrfeld J (1989): High-performance liquid chromatography analysis of phytic acid on a pH-stable, macroporus polymer column. Cereal Chem. 66: 510
Lemtiri-Chlieh F, MacRobbie E A C, Brearley C A (2000): Ino-sitol hexakisphosphate is a physiological signal regulating the K-inward rectifying conductance in guard cells. Proc. Natl. Acad. Sci. U.S.A. 97: 8687-8692
Liu Z H, Wang H Y, Wang X E, Zhang G P, Chen P D, Liu D J (2006): Genotypic and spike positional difference in grain phytase activity, phytate, inorganic phosphorus, iron, and zinc contents in wheat (Triticum aestivum L.). J. Cereal Sci. 44: 212-219
Lolas G M, Palamidis N, Markakis P (1976): The phytic acid-total phosphorus relationship in barley, oats, soybeans, and wheat. Cereal Chem. 53: 867-871
Lönnerdal B (2002): Phytic acid-trace element (Zn, Cu, Mn) interactions. International J. Food Sci. Tech. 37: 749-758
Mladenović-Drinić S, Ristić D, Sredojević S, Dragičević V, Micić-Ignjatović D, Delić N (2009): Genetic variation of phytate and ionorganic phosphorus in maize population. Genetika 41: 107-115
Nahapetian A, Bassiri A (1976): Variations in concentrations and interrelationships of phytate, phosphorus, magnesium, calcium, zinc and iron in wheat varieties during two years. J. Agric. Food Chem. 24: 947-950
Oberleas D (1971): The Determination of Phytate and Inositol Phosphates. Methods Biochem. Anal. 20: 87-101
Oberleas D (1983): Phytate content in cereals and legumes and methods of determination. Cereal Foods World 28: 6
Oberleas D, Harland B F (1986): Analytical methods for phyta-te. In: Graf E. (ed) Phytic acid: Chemistry and Applications, Pilatus Press: Minneapolis, MN, 77-78
O’Dell B L, De Boland A R, Koirtyohann S R (1972): Distribu-tion of phytate and nutritionally important elements among morphological components of cereal grains. J. Agric. Food Chem. 20: 718-721
Ortiz-Monasterio I, Palacios-Rojas N, Meng E, Pixle K, Tret-howan R, Pena R J (2007): Enhancing the mineral and vita-min content of wheat and maize through plant breeding. J. Cereal Sci. 46: 293-307
Perić V, Dragičević V, Sredojević S, Srebrić M, Terzić D, Mlade-nović-Drinić S (2009): Genetička varijabilnost sadržaja fos-fora u semenu soje. J. Sci. Agric. Res. 70: 79-84
Pontoppidan K, Petterson D, Sandberg A S (2007): The type of thermal feed treatment infl uences the inositol phosphate composition. Anim. Feed Sci. Technol. 132: 137-147
Prasad A S (1996): Zinc defi ciency in women, infants and chil-dren. J. Am. Coll. Nutr. 15: 113-120
Raboy V, Noaman M M, Taylor G A, Pickett S G (1991): Grain phytic acid and protein are highly correlated in winter wheat. Crop Sci. 31: 631-635
Raboy V (2001a): Progress in breeding low phytate crops. Sym-posium: Plant Breeding: A new tool for fi ghting micronutri-ent malnutrition, April 1st, Orlando, Florida, 503-505
Raboy V (2001b): Seeds for a better future: “low phytate” grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 6: 458-462
Raboy V, Young K A, Dorsch J A, Cook A (2001): Genetics and breeding of seed phosphorus and phytic acid. J. Plant Physiol. 158: 489-497
Reichwald K, Hatzack F (2008): Application of a modifi -ed Haug and Lantzsch method for the rapid and accurate phytate determination in soybean, wheat and maize meals. J. Agri. Food Chem. 56: 2888-2891
Safrany S T, Caffrey J J, Yang X, Shears S B (1999): Diphosp-hoinositol polyphosphates: the fi nal frontier for inositide research. Biol. Chem. 380: 945-951
Sandberg A S (1995): Determination of phytic acid. In: Recent progress in the analysis of dietary fi bre Luxembourg: Euro-pean Commission, Cost 92, 93-103
Sasakawa N, Sharif M, Hanley M R (1995): Metabolism and bi-ological activities of inositol pentakisphosphate and inositol hexakisphosphate. Biochem. Pharmacol. 50: 137-146
Svečnjak Z, Bujan M, Dragojević I V, Vitali D, Čebušnik A, Jenel M (2007): Nitrogen and phosphorus content, hectoliter wei-ght and yield variations of wheat grain as affected by cropping intensity. Agriculturae Conspectus Scientifi cus 72: 251-255
Syltie P W, Dahnke W C (1983): Mineral and protein content, test weight, and yield variations of hard red spring wheat as infl u-enced by fertilization and cultivar. Qual. Plant. 32: 37-49
Tabekhia M M, Donnelly B J (1982): Phytic acid in durum whe-at and its milled products. Cereal Chem. 59: 105-107
Tang J, Zou C, He Z, Shi R, Ortiz-Monasterio I, Qu Y, Zhang Y (2008): Mineral element distributions in milling fractions of Chinese wheats. J. Cereal Sci. 48: 821-828
Tangendjaja B, Buckle K A, Wootton M (1980): Analysis of phytic acid by high-performance liquid chromatography. J. Chromat. 197: 274
Vaintraub L A, Lapteva N A (1988): Colorimetric determina-tion of phytate in unpurifi ed extracts of seeds and the pro-ducts of their processing. Analyt. Biochem. 175: 227-230
Walter T, Peirano P, Ronglacliolo M (1997): Effect of iron de-fi ciency anemia on cognitive skills and neuromaturation in infancy and childhood. In: Fisher, P.W.F, L’Abbé M.R., Coc-kell, K.A., Gibson RS (Eds.), Trace Elements in Man and Animals-9. Proceedings of the Nineth International Sym-posium on Trace Elements in Man and Animals. National Research Council of Canada, Ottawa, 217-219
Williams S G (1970): The role of phytic acid in the wheat grain. Plant Physiol. 45: 376-381
Wodzinski R J, Ullah A H J (1996): Phytase. Adv. Appl. Micro-biol. 42: 263-302
Yenagi S N, Basarkar P W (2008): Antioxidant contents of whole grain cereals of North Karnataka. Karnataka J. Agric. Sci. 21: 602-603
York J D, Odom A R, Murphy R, Ives E B, Wente S R (1999): A phospholipase dependent inositol polyphosphate kinase pathway required for effi cient messenger RNA export. Sci-ence 285: 96-100
Zebarth B J, Warren C J, Sheard R W (1992): Infl uence of the rate of nitrogen fertilization on the mineral content of win-ter wheat in Ontario. J. Agric. Food Chem. 40: 1528-1530
Zečević V, Knežević D, Mićanović D (2007): Variability of te-chnological quality components in winter wheat. Genetika 39: 65-374

Branković G i sar.14
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 7-14
Wheat Breeding for Low Phytic Acid Content: State and Perspectives
Gordana Branković1 · Desimir Knežević2 · Dejan Dodig3 · Vesna Dragičević3
1Faculty of Agriculture, University of Belgrade, Nemanjina 6, 11080 Zemun2Faculty of Agriculture, University of Priština, Jelene Anžujske b.b., 38228 Zubin Potok
3Maize Research Institute ”Zemun Polje”, Slobodana Bajića 1, 11080 Zemun
Summary: Interest in wheat breeding for low phytic acid content arised from its roll as antinutrient factor which chelates mineral elements (Ca, Zn, Fe, Mn, Cu and P), leading to their inadequate use. Excretion of unused P in phytic acid complex through non-ruminant animals such as poultry, swine and fi sh causes water eutrophication. Numerous indirect methods (e.g. spectrophotometric) and direct methods (HPLC - High Performance Liquid Chromatography) were developed for fast and accurate phytic acid determination in wheat. It typically represents 50-85% of seed total phosphorus and one to several percents of dry seed weight. Phytic acid content and phytate phosphorus genetic variability have been determined for wheat cultivars and lines under different environmental conditions. Wheat mutant (Triticum aestivum L.) for low phytic acid content Js-12-LPA was created through breeding efforts.Key words: breeding, mutants, phytic acid, wheat

Molecular Mechanisms in Plant Abiotic Stress Response 15
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24review article / pregledni rad
Molecular Mechanismsin Plant Abiotic Stress Response
Palmiro Poltronieri · Stefania Bonsegna · Stefania De Domenico · Angelo Santino
received / primljeno: 15.04.2010. revised / prerađeno: 02.12.2010. accepted / prihvaćeno: 15.12.2010.© 2011 IFVC
Summary: Improved crop varieties are needed to sustain the food supply, to fi ght climate changes, water scarcity, temperature increase and a high variability of rainfalls. Variability of drought and increase in soil salinity have negative effects on plant growth and abiotic stresses seriously threaten sustainable agricultural production. To overcome the infl uence of abiotic stresses, new tolerant plant varieties and breeding techniques using assisted selection are sought. A deep understanding of the mechanisms that respond to stress and sustain stress resistance is required. Here is presented an overview of several mec-hanisms that interact in the stress response. Localised synthesis of plant hormones, second messengers and local effectors of abiotic stress response and survival, the signalling pathways regulated by plant hormones are today better understood. Metabolic networks in drought stress responses, long distance signalling, cross-talk between plant organs fi nalised to tissue-specifi c expression of abiotic stress relie-ving genes have been at the centre of most recent studies. Key words: abscisic acid (ABA), jasmonate (JA), osmotic adjustment (OA), transcription factor (TF)
P. Poltronieri · S. Bonsegna · S. De Domenico · A. SantinoISPA-CNR, Institute of Sciences of Food Productions, Via Montero-ni 73100 Lecce, Italye-mail: [email protected]
Introduction
Modern agriculture faces abiotic stresses, especially salinity and drought as major factors limiting crop productivity worldwide. Low temperatures, heath shock, heavy metals, UV-radiation and pesticides are also abiotic factors affecting crop yields (Bohnert et al. 1995, Tardieu & Tuberosa 2010). Climate changes with erratic rainfalls and prolonged drought out of the expected season have multiplicative effects on abiotic stress consequences and seriously threaten sustainable agricultural production. High salinity is not only a soil-type characteristic but can be also observed in arable, irrigated areas due to use of low quality water. In addition, soil salinity induces water defi cit that leads to nutrient defi ciencies. Abiotic stress is one primary cause of crop loss worldwide, causing average yield losses of more than 50% for major crops. Plants can resist abiotic stresses through different distinct mechanisms. Tolerance and susceptibility to abiotic stresses are very complex. Plant traits that are associated with resistance mechanisms are multigenic and thus diffi cult to control and engineer.
Drought induces mechanical stress on roots due to soil hardness, osmotic stress because of cell dehydration and removal of water in the extra-cellular space, and oxidative stress by the accumulation of reactive oxygen species (ROS) (Kovtun et al. 2000).
Plants respond to water stress through organ-compartmentalised changes. In roots, there is an enhanced capture of soil moisture (e.g. reaching deep soil moisture with longer roots), while in stem/leaves there is a reduction of water loss (e.g. stomatal closure, reduction of plant size and/or leaf area) retaining tissue hydration via osmotic adjustments (OA). A prolifi c and deep root system confers better growth and yield under terminal drought, especially for varieties showing either deep rooting features and a positive harvest index (number and size of seeds) (Chandra Babu et al. 2001). In the case of water scarcity or drought, plants improve their water use effi ciency either by dehydration avoidance or through dehydration tolerance (Plaut et al. 2004, Thomas & Howarth 2000, Blum 2005).
Reduced growth duration (with early fl owering) is also an important mechanism as the plant generally uses less water and can also avoid the end-of-season (terminal) stress. However, reduced or early fl owering plants may lead to lower production rates and smaller seeds.
Stress Physiology / Fiziologija stresaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Poltronieri P et al.16
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
Stress-Responsive Gene Expression
Under abiotic stress (drought, salinity) plants respond with the activation of synthesis or inhibition of catabolism of osmolytes as sugars (saccharopine, trehalose, raffi nose, galactinol, sorbitol, mannitol, fructans) (Cortina & Culianez-Macia 2005) and sugar alcohols (e.g. polyols), amino acids as branched chain aminoacids (BCA), proline and agmatin, quaternary ammonium compounds and small dipeptides as glycinebetaine (Holmstrom et al. 2000). Polyamines (i.e. putrescine) are involved in a wide range of plant processes including wounding (Groppa & Benavides 2008). Having a role in development and H2O2 production, amine oxidases are involved in the increase of polyamines that mediate the stress responses.
There is a need to provide established or new varieties with genotypes having enhanced or faster induction of expression of genes at the crossroad of permissive growth under stress condition. This group of permissive genes includes aquaporin isoforms able to optimise water fl uxes (Javot et al. 2003). Several genes have been characterised for their role on stress protection. ERECTA is a gene regulating transpiration effi ciency affecting stomatal closure, while the plant is able to maintain biomass production (Masle et al. 2005).
Other important regulatory proteins involved in drought stress are proton antiporters as TNHX1 and a proton pyrophosphatase TVP1 (Brini et al. 2005, Brini et al. 2007a) shown to improve salt and drought stress tolerance in Arabidopsis.
Dehydrin isoforms (LEA/DHN) of sizes 17 kDa, 30 kDa and 40 kDa, function as intracellular chaperones for other proteins and nucleic acids. One dehydrin (DHN-5) was shown to be involved in salt and drought resistant phenotypes of durum wheat (Brini et al. 2007b). Dehydrins accumulate to a high level in the mature embryos. The accumulation of DHN-5 was shown to have a role in preservation of cell integrity during late embryogenesis and desiccation (Brini et al. 2007b). Dehydrins are also known as late embryogenesis abundant (LEA) or early response to dehydration (ERD) proteins (Beck et al. 2007), so called because they also accompany seed formation. It has been shown that LEA/DHNs exert chaperone activity on proteins and also nucleic acids (Kovacs et al. 2008; Hara et al. 2009). DHN/LEAs lack a fi xed three-dimensional structure, remarkably stable even at macromolecular crowding conditions, maintaining a disordered character under conditions (dehydration) in which unfolded states of several globular proteins would tend
to collapse (Mouillon et al. 2008). Several LEA/DHN isoforms are regulated by abscisic acid (ABA) (Jimenez et al. 2008). While the size of induced DHNs was large, ranging between 14 kDa and 74 kDa, drought-tolerant bermudagrass varieties showed to induce preferentially the 31 kDa and 40 kDa isoforms (Hu et al. 2010).
Roots grow and recover their function after environmental stresses through specifi c genes that adapt root development to these restrictive conditions. The recovery of Medicago truncatula roots after a salt stress is mediated by regulatory networks depending on TFIIIA-like transcription factors, involved in the control of root adaptation to salt stress. Those conditions induced synthesis of a novel RNA-binding protein, a small G-protein homologous to ROP9, a receptor-like kinase, two TF IIIA-like and an AP2-like transcription factors (TF), MtZpt2-1, MtZpt2-2 and MtAp2, and a histidine kinase associated with cytokinin transduction pathways (Merchan et al. 2007).
Role of JA and ABA in Stress Signalling
The interplay of several plant hormones regulates the commitment of plants to growth or senescence under abiotic stress. The plant copes with a specifi c stress through complex signals circulating from roots to shoots and leaves (Fig. 1), and delivering hormones and signalling back to the roots, activating the expression of protective proteins or down-regulating unnecessary pathways. Plant hormones sustain signals at short distance, between cells, or in a systemic way in the case of transport systems. In this review, we will focus principally on the roles exerted by jasmonic acid (JA) and abscisic acid (ABA). JA is the fi nal product of the octodecanoic pathway which uses linoleic acid as substrate to produce oxylipins (Hughes et al. 2009). The octodecanoic pathway starts with the oxygenation of a polyunsaturated fatty acid (PUFA) by lipoxygenase (LOX), to form a fatty acid hydroperoxide. In plants, only the type-2 13-LOXs are believed to be associated with JA biosynthesis (De Domenico et al. 2007). In the biosynthesis of JA, the subsequent activities of 13-LOX, allene oxide synthase (AOS), allene oxide cyclase (AOC) lead to formation of cis-(+)-12-oxophytodienoic acid (OPDA), a JA precursor. JA can be methylated by a specifi c JA-methyltransferase. Methyl-Jasmonate (MeJA) is a volatile compound able to cross plasma membranes and exert its action at distance. Another effector in the lipoxygenase biosynthesis pathway is jasmonate-conjugated isoleucin (JA-Ile) that may accumulate stored in organelles and

Molecular Mechanisms in Plant Abiotic Stress Response 17
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
vacuoles. JA-Ile is able to translocate through membranes and move through xylem from roots to leaves and backward.
Once the hormone has arrived at its destination in the target cell (locally or at distance), the priming of jasmonate-inducible genes is regulated by convergent pathways, linked to phosphoinositide- and ABA-dependent signalling components.
1.1 JA Signalling in Root Response to Abiotic Stresses
Plant growth and development are largely dependent on the plant root system, due to its crucial role in water and mineral uptake. Abiotic below-ground stresses (e.g. drought, waterlogging, salt, nutrient soil status, physical soil properties) affect overall plant growth and architecture. This early signalling affects root growth regulation, resource acquisition and root-shoot communication under abiotic stress.
Many plants develop mutualistic interactions with arbuscular mycorrhizal (AM) fungi and with
nitrogen-fi xing bacteria. AM interaction is widely distributed among most of the land plants, while actinorhiza and rhizobia interactions are restricted to species of ten plant families, including the legumes. Fungi of the phylum Glomeromycota exist in a symbiotic association with vascular plants (Schüssler et al. 2006). As for the actinorhiza, the symbiosis occurs between actinobacteria of the
genus Frankia and plants of Fagales, Cucurbitales and Rosales (Pawlowski & Sirrenberg 2003). On the other hand, the legume-rhizobia symbiosis (LRS) occurs between Rhizobium species and legumes (Kistner & Parniske 2002). Both types of nodule symbioses are characterized by the intracellular uptake of bacteria and the formation of specialized organs, the root nodules.
A phytohormone involved in the regulation of interactions between root and bacteria is JA and its conjugated form (MeJA) which cross the membrane barrier functioning as a long-distance signal (Hause & Schaarschmidt 2009). MeJA has been detected in high levels in germinating
Fig. 1. The ABA and JA biosynthesis pathways in plantsSlika 1. Tokovi biosinteze ABA i JA u biljkama
The ABA synthesis involves multiple reactions that occur in the plastid and the cytoplasm. ZEP: zeaxanthin epoxidase; VDE: violaxanthin de-epoxidase; NCED: 9-cis-epoxycarotenoid di-oxygenase. The fi nal steps in ABA synthesis in the cytosol involves a xanthoxin:NAD+ oxidoreductase (mutated in aba2) and the ABA aldehyde oxidase. The JA biosynthesis pathway involves two compartments. Chloroplast route: linolenic acid (18:3) is used as substrate for the sequential action of three plastidial enzymes, lipoxygenase (LOX), allene oxide synthase (AOS) and allene oxide cyclise, which produces 12-oxo-phytodienoic acid (OPDA). Peroxisome route: following OPDA import into peroxisomes, jasmonic acid is produced by oxophytodienoate reductase 3 in three cycles of β-oxidation.

Poltronieri P et al.18
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
soybeans and in root tips (Mason et al. 1992, Oldroyd 2009). Jasmonates directly induce nod gene expression in rhizobia, and indirectly promote bacterial Nod factor production by inducing (iso)fl avonoid biosynthesis genes in the plant (Zhang et al. 2007). As a negative feedback, Nod factor causes Ca2+ spiking in root hairs (Oldroyd 2009).
Regulation of the redistribution of nutrients might be the most important role of jasmonates in AM roots. In plants such as M. truncatula and barley, developing a mutualistic symbiosis which ultimately leads to a promoted growth, jasmonates might help to regulate the nutrient exchange between both partners. However, root JA and Me-Ja are important also in plants devoid of symbiotic relationships.
1.2 JA Receptors and Downstream Signal Transduction in Distant Signalling and Abiotic Stress Response
There is an intense communication between
different organs through the phloem sap, the system of delivery of nutrients and solutes, and through the xylem, the water channels which move soluble compounds from roots to leaves. How these signals move, using the xylem or the phloem way, upward or downward, and which compounds (conjugated phytohormones, proteins, RNAs) is mostly unknown and under investigation, but several fi ndings show a signifi cant role of certain hormones and signals (Lough & Lucas 2006).
JA-related compounds have been identifi ed in plants with mutations in components of the ubiquitin proteasome system (Tiryaki & Staswick 2002, Lorenzo & Solano 2005, Moon et al. 2007). Recent developments include the identifi cation of COI1 as a receptor for jasmonates. COI1 is an F-box protein taking part of an Skp1-Cullin-F-box (SCF) complex, closely related to the auxin signalling TIR1 (TRANSPORT INHIBITOR RESPONSE 1) gene, which encodes the fi rst plant F-box protein to be identifi ed as part of an SCF complex. SCF complex interacts with CORONATINE INSENSITIVE1 (COI1) E3 ubiquitin ligase, that targets proteins to the proteasome degradation pathway. COI1 directly binds (+)-7-iso-JA-l-Ile and thus functions as JA receptor revealed that formation of the ternary complex COI1-JA-Ile-JAZ is an ordered process (Balbi and Devoto 2008). JA-Ile promotes the interaction between the ubiquitin ligase complex SCF(COI1) and JAZ proteins, resulting in their degradation by the 26S proteasome, thereby
liberating AtMYC2 from repression according to the prevailing model.
Two jasmonate-responsive promoter sequences are known. The GCC motif, commonly found in promoters activated synergistically by jasmonate and ethylene, and the G-box, found in promoters activated by jasmonates and repressed by ethylene. Important transcription factors conferring jasmonate-responsive gene expression in Arabidopsis are ORA59 and AtMYC2. ORA59 interacts with the GCC motif and controls the expression of genes that are synergistically induced by jasmonates and ethylene, whereas AtMYC2 interacts with the G-box and related sequences, and controls genes activated by jasmonate alone (Gfeller et al. 2010).
1.3 Early Stress Sensing in Roots and Associated Bacteria and Fungi
Once the soil derived signals (mechanical and
osmotic stress, and signals originated from the stressed ectomycorrhiza) are sensed by root, they are rapidly translated into specifi c signals. The relevance of an early and immediate response in stress tolerant varieties is utmost, while in unresponsive varieties there is a delayed and reduced response (Schachtman & Goodger 2008).
Plant-growth-promoting (PGP) fungi (Pennisi 2006, White & Torres 2010) and plant-growth-promoting rhizobacteria (PGPR) (Liu et al. 2007) augment plant productivity and immunity through an increase in the antioxidative state. PGP endophytes induce root biomass, counteract salt-induced increase in heat effl ux, produce changes in fatty acids composition, reduce lipid peroxidation in leaves, increase antioxidant enzyme activities and enable roots to maintain ascorbate in its reduced state under salt stress (Baltrushat et al. 2008). Certain bacteria promote plant growth through the activity of several genes, coding for putative proteins involved in survival in the rhizosphere (to cope with oxidative stress or uptake of nutrients released by plant roots), root adhesion (pili, adhesion, hemagglutinin, cellulose biosynthesis), colonization/establishment inside the plant (chemiotaxis, fl agella, cellobiose phosphorylase), plant protection against fungal and bacterial infections (siderophore production and synthesis of the antimicrobial compounds 4-hydroxybenzoate and 2-phenylethanol), and improved growth and development through the production of the phytohormones indole acetic acid, acetoin, and 2,3-butanediol (Taghavi et al. 2010).

Molecular Mechanisms in Plant Abiotic Stress Response 19
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
PGPR also elicit the ‘induced systemic tolerance’ to salt and drought. Even in the non-ectosymbiotic population of bacteria, there are positive infl uences on the plant hosts. The Pseudomonas syringae phytotoxin coronatine mimics jasmonoyl-isoleucine (JA-Ile), which is a crucial plant signalling molecule for regulating plant defence responses (Weiler et al. 1994, Bender et al. 1999). During water stress, the nodules sense drought and respond by activating defence mechanisms (Luo et al. 2009). In this way, Ectomycorrhizas (EMs) and rhizobia alleviate and improve also stress tolerance of host plants (Yang et al. 2009). It is worthy to note that arbuscular mycorrhizal fungi host endosymbiotic bacteria providing benefi cial properties such as heat tolerance and protection from pests (Naumann et al. 2010).
Bacteria possess a nitric-oxide-synthase involved in stress resistance. NO is a mobile gaseous second messenger that is sensed at root level. In legumes, globin NO binding proteins, called leghemoglobins, accumulate in symbiosomes (root nodules) of various legumes. Non-symbiotic hb genes are expressed in specifi c plant tissues, and over-expressed in organs of stressed plants. These observations suggest that nsHbs function as additional to O(2)-transporters, such as to modulate levels of ATP and NO (avoiding protein nitrosylation). Non-symbiotic haemoglobins are a key enzymatic system for NO scavenging in plants, indicating that the primordial function of haemoglobins may be linked to the protection against nitrosative stress (Perazzolli et al. 2006, Smagghe et al. 2009).
P. fl uorescens WCS417r bacteria and beta-aminobutyric acid can induce disease resistance in Arabidopsis, priming specifi c defence mechanisms. Both WCS417r and beta-aminobutyric acid prime enhanced deposition of callose-rich papillae after infection by the oomycete Hyaloperonospora arabidopsis (Van der Ent et al. 2009).
2.1 Abscisic Acid in Abiotic Stresses
Abscisic acid (ABA) is a phytohormone involved in senescence, seed dormancy, plant development, drought tolerance and stress response, synthesised in the xylem and in the aerial parts of the plant, where it regulates stomatal movement and the activity of shoot meristems. ABA can fl ow in the root cortex across apoplastic barriers and play an important role in the regulation of signal intensity. The abscisic acid glucose ester (ABA-GE) is a long-distance stress signal, stored in microsomes, and released by activated beta-glucosidases, both in the apoplast and the cytosol
of the mesophyll cells. ABA-GE transporters located on plasma membrane of the xylem parenchyma cells infl uence ABA mobility. When discussing the intensity of the ABA signal in the xylem, internal and external sources of ABA have to be considered. External ABA originates from root exudation and from ABA-producing soil organisms (predominantly fungi), whereas internal ABA comes from its biosynthesis at root shoots level and phloem import (Sauter & Hartung 2000, Sauter et al. 2001). It was shown that conjugated ABA (ABA-GE) also occurs in the soil, often in higher concentrations than ABA. The Casparian bands of the exodermis and endodermis are perfect barriers for ABA-GE. When an exodermis is absent (Fabaceae and hydroponically cultivated plants), external ABA-GE enters into the apoplast of the root cortex. Apoplastic β-glucosidases can cleave the conjugated form and release free ABA, which is distributed to the symplast and/or transported across the endodermis into the xylem (Hartung et al. 2002).
2.2 Timing of ABA Synthesis and Mobilisation
NaCl causes a rapid increase in ABA level in the transpiring portion of growing leaves. Within 10 min following salt addition, ABA levels show a 6-fold increase in the distal portion of the leaf elongation zone whereas it accumulate later in the proximal portion. In the portion of the growing blade, ABA increases 3-fold and remains elevated during the following 20 min. This precedes the decrease in transpiration and stomatal conductivity. Twenty hours following salt addition, ABA concentration returned to the pre-stress level. Leaf elongation velocity was still reduced. An ATP-Binding Cassette (ABC) transporter, AtABCG25, expressed mainly in vascular tissues such as roots and leaf veins, localized in the outer cell membranes (Kuromori et al. 2010) was identifi ed as the gene involved in a plant mutant hypersensitive to ABA at the germination and seedling stages. Plants genetically engineered to over express AtABCG25 had higher leaf temperature compared to normal plants and decreased water loss from isolated leaves. Thus, facilitated ABA movements may affect distant targets such as guard cells.
2.3 Salt Stress and Phosphate Defi ciency Increase ABA Formation in the Roots
Strong ABA synthesis and accumulation in the roots can be observed in plants affected by

Poltronieri P et al.20
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
hemiparasites such as Rhinanthus minor (Jiang et al. 2004). ABA biosynthesis in the roots was 12-fold higher after attack, resulting in 14-fold higher ABA fl ows in the xylem.
Plants regulate inorganic phosphate (Pi) homeostasis to adapt to environmental changes in Pi availability. Some degree of cross-talk between ABA and other signalling pathways was reported in phosphate limitation conditions. This mechanism involves phosphate uptake increase from the soil and phosphate mobilization from the leaf. Upon Pi starvation, upregulated miR399 cleaves its target gene, PHO2, in A. thaliana, an ubiquitin-conjugating E2 enzyme, thereby releasing several protein targets from ubiquitin-pathway dependent degradation and increasing Pi content in the shoots (Franco-Zorrilla et al. 2007).
2.4 Correlation between Xylem ABA Concentration and Leaf Conductance
ABA signalling can be intensifi ed in many crops, for instance grapevine, by applying modern irrigation techniques, such as partial root drying (PRD). PRD is an irrigation technique where water is distributed unevenly to the root system divided into irrigated and dry compartments. Applying PRD to grape cultivation (Stoll et al. 2000), ABA signalling was found to be reduced after prolonged drying of one compartment. To sustain PRD effects, it was necessary to alternate wet and dry parts regularly (PRD-A). PRD-A was shown to intensify ABA signalling with positive consequences for water saving and improvement of the quality of the crops (Dodd et al. 2008). The factors that regulate the intensity of the ABA signal in the xylem, on its route from the root to the target cells, are therefore of particular interest.
2.5 Role of ABA in JA Signalling
Recent fi ndings have suggested an interaction between jasmonic acid and ABA in plants under drought stress (Mahouachi et al. 2007). The pattern of accumulation of JA is compatible with a triggering signal upstream ABA. The phytohormone ABA acts in all adaptive responses
to environmental stresses. The phytohormones gibberellin (GA) and ABA play essential and often antagonistic roles in regulating plant growth, development and stress responses. The transduction of their signal occurs through several types of receptors (Razem et al. 2006). Soil water stress elicits a hydraulic response in the shoot, which precedes ABA signalling and
stomatal closure (Christmann et al. 2007). ABA biosynthesis in leaves is increased signifi cantly only when turgor approaches zero. The location of ABA release and the tissue-specifi c expression of ABA receptor types are signals for that particular tissue (seeds, leaves) to reorganize and reprogram, through epigenetic control, differential gene expression of an ABA-directed cluster of genes (Seki et al. 2007).
2.6 ABA Receptors and Downstream Signal Transduction
The Arabidopsis ABA receptor FLOWERING TIME CONTROL LOCUS A (FCA), the GA receptor GIBBERELLIN INSENSITIVE DWARF1 (GID1) and the nuclear-located auxin receptor show an important activity as plant hormone receptors within the cell nucleus. GID1, FCA and TRANSPORT INHIBITOR RESPONSE1 (TIR1) receptors reveal a novel signalling mechanism that appears to be short, to be independent of relay intermediates, and to involve only protein–protein interactions that are affected by receptor-hormone binding.
FCA is a plant-specifi c RNA-binding protein, possesses two RNA-recognition motifs (RRM) in addition to a tryptophan-tryptophan (WW) protein-interaction domain. FCA shares sequence homology at the C-terminus with the ABA-binding protein (ABAP1) associated with the plasma membranes of barley aleurone cells. FCA ABA receptor affects RNA metabolism by binding directly to an RNA binding protein (Simpson et al. 2003).
Although ABAP1 does not possess the common RRM to bind RNA, it does possess a WW interaction domain that harbours residues homologous to the Prp40 splicing factor and there is substantial evidence to link nuclear WW-containing proteins to both transcription and RNA splicing. ABA–ABAP1 binding results in changes in ABAP1 protein conformation, the subsequent dissociation from an interacting protein in the plasma membrane, and the appearance of ABAP1 in cytosolic and nuclear compartments. In either instance, if the proteins that interact with characterized (FCA) or putative (ABAP1) receptors are restricted in their subcellular localization, then the mobility of ABA hormone receptors might have particular relevance for covering the entire suite of non-genomic and genomic responses encompassed by ABA (Hugouvieux et al. 2001).
PYR/PYL/RCAR proteins function as ABA receptors that, following ABA binding, inhibit

Molecular Mechanisms in Plant Abiotic Stress Response 21
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
the activity of known negative regulators of ABA signalling (the type 2C protein phosphatases (PP2Cs) ABI1, ABI2, HAB1, HAB2 and PP2CA) in both seeds and vegetative tissues. In the presence of ABA, PYR/PYL/RCARs bind to PP2Cs and induce the release of SNRK2 Ser/Thr kinases from PP2Cs, which would otherwise keep SNRK2s in an inactive state. The association of PYR1, ABI1, SRK2E and ABF2 is suffi cient for ABA-triggered ABF2 phosphorylation (Fujii et al. 2009). SNRK2s phosphorylation of downstream substrates, including ABA responsive element-binding protein (AREB) and ABA responsive element-binding factor (ABF) bZIP transcription factors activates ABA-responsive genes and ABA-related responses (Yin et al. 2009).
Another transcription factors activated by ABA signalling is R2R3MYB that translates ABA effects through differential gene expression (Abuqamar et al. 2009).
GPCR-type G proteins (GTG1 and GTG2) bind ABA specifi cally and interact with the Arabidopsis G protein alpha subunit, GPA1, but also have intrinsic GTP-binding and GTPase activity (Pandey et al. 2009).
Epigenetic processes are an integral part of ABA-regulated processes. Epigenetic mechanisms, namely, histone modifi cations and cytosine DNA methylation-induced modifi cation of genome give rise to epigenomes (Chinnusamy et al. 2008). Abiotic stress-induced ABA regulates stomatal response and stress responsive gene expression through HDACs and HOS15-dependent histone deacetylation, as well as through the ATP dependent SWITCH/SUCROSE NONFERMENTING CRC. ABA also probably regulates the abiotic stress response through DNA methylation and short interfering RNA pathways. Further studies on ABA-regulated epigenome will be of immense use to understand the plant development, stress adaptation and stress memory.
2.7 ABA and Abiotic Stress Memory
Abiotic stress-induced genomic changes are necessary for stress memory. UV-C radiation or fl agellin (a plant defence elicitor) stress-induced changes in the genome have been shown to be a dominant trait and transmitted through both the maternal and the paternal crossing partner in Arabidopsis (Molinier et al. 2006). Progeny of tobacco mosaic virus (TMV) infected plants showed enhanced frequency of the somatic and meiotic recombination rates at certain loci due to DNA hypomethylation. Thus, stress memory appears to be inherited through epigenetic changes
(Boyko et al. 2007). ABA may be involved also in the somatic recombination frequency (Boyko et al. 2006) and induces chromatin remodelling, which regulates transcription, recombination, replication and genome organization. It is also likely that ABA might regulate DNA methylation through DML3 and siRNAs. Under abiotic stresses ABA mediates reduction in plant growth. One of the mechanisms of abiotic stress-induced growth arrest is regulated by chromatin remodelling (Mlynarova et al. 2007). Abiotic stress-induced inheritable epigenetic state might have an adaptive advantage but also a negative effect on the yield of crops, as stress memory can inhibit the expression of full potential of crops. There is a great interest to understand the role of ABA in abiotic stress memory to exploit it in agricultural practices.
2.8 Regulatory Metabolic Networks in Drought Stress Responses
Localised delivery of paracrine hormones, long distance signalling and a second wave of tissue specifi c hormone signals regulate the intensity of ABA signals (Jiang & Hartung 2008).
In higher plants, a number of physiological processes are regulated by systemic RNA signalling molecules. This phloem-mediated remote-control system provides specifi c and effi cient regulation to fi ne-tune many plant developmental programs. In phloem, several endogenous RNA-binding proteins have been identifi ed. In addition, viral RNA-binding proteins can counterbalance the plant RNA silencing effectors synthesised to block viral RNA (Lakatos et al. 2004). CmPP16 protein from Cucurbita maxima was shown to possess properties similar to those of viral movement proteins (Xoconostle-Cázares et al. 1999). Messenger RNAs have been shown to travel at long distance in the phloem. Four mRNAs (BEL5, LeT6, KNOTTED1 and GAI) are able to move for long distances in Arabidopsis and potato (Haywood et al. 2005, Banerjee et al. 2006). GAI RNA is the mRNA for gibberellin acid insensitive (GAI), a DELLA protein of GRAS transcription factors (Pysh et al. 1999). DELLA proteins interact and block the activity of GA-responsive transcription factors (Huang & Yu 2009). The RNA–protein complexes are subsequently transported into sieve elements through plasmodesmata (Lucas et al. 2009).
Non-coding RNAs (small RNAs as miRNAs, and long RNAs as antisense RNA, dsRNA, and long RNA species) have many roles, functioning as regulators of other mRNAs, at transcriptional

Poltronieri P et al.22
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
and post-transcriptional level, and controlling protein ubiquitination and degradation. Several microRNAs has been shown to move through the phloem to exert their activity at distance. MIR172 as well as MIR399, which is induced upon phosphate starvation, is present in small RNA libraries made from phloem exudates indicating that they are part of a long distance signalling network (Chuck & O’Connor 2010). Phloem specifi c small RNAs travel to inform the roots of the nutrient status of the shoot, such as miR319 (Zhang et al. 2009) which targets a subset of the TCP family of transcription factors. LOX2 expression may be dependent on TCP4, due to the presence of a promoter motif important for the direct regulation of LOX2 by TCP4 (Schommer et al. 2008).
Conclusions
The molecular analysis of stress-induced signalling pathways that lead to plant adaptation or to cell death constitutes a major research area in the abiotic stress fi eld. The crossroad between stress adaptation and cell death and the determinant events leading to one or the other direction represent a challenging research area in the next future.
References
Abuqamar S, Luo H, Laluk K, Mickelbart M V, Mengiste T (2009): Crosstalk between biotic and abiotic stress respon-ses in tomato is mediated by the AIM1 transcription factor. Plant J. 58: 347-360
Balbi V, Devoto A (2008): Jasmonate signalling network in Ara-bidopsis thaliana: crucial regulatory nodes and new physiologi-cal scenarios. New Phytologist 177: 301-318
Baltruschat H, Fodor J, Harrach B D, Niemczyk E, Barna B, Gullner G, Janeczko A, Kogel K-H, Schäfer P, Schwarczin-ger I, Zuccaro A, Skoczowski A (2008): Salt stress tolerance of barley induced by the root endophyte Piriformospora indi-ca is associated with a strong increase in antioxidants. New Phytol. 180: 501-510
Banerjee A K, Chatterjee M, Yu Y, Suh S G, Miller W A, Ha-nnapel D J (2006): Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 18: 3443-3457
Beck E H, Fettig S, Knake C, Hartig K, Bhattarai T (2007): Spe-cifi c and unspecifi c responses of plants to cold and drought stress. J. Biosci. 32: 501-510
Bender C, Rangaswamy V, Loper J (1999): Polyketide produc-tion by plant-associated pseudomonads. Annu. Rev. Phyto-pathol. 37: 175-196
Blum A (2005): Drought resistance, water-use effi ciency, and yield potential—are they compatible, dissonant, or mutually exclusive? Austral. J. Agric. Res. 56: 1159-1168
Bohnert H J, Nelson D F, Jenson R G (1995): Adaptation to environmental stresses. Plant Cell 7: 1099-1111
Boyko A, Hudson D, Bhomkar P, Kathiria P, Kovalchuk I (2006): Increase of homologous recombination frequency
in vascular tissue of Arabidopsis plants exposed to salt stress. Plant Cell Physiol. 47: 736-742
Boyko A, Kathiria P, Zemp FJ, Yao Y, Pogribny I, Koval-chuk I (2007): Transgenerational changes in the genome stability and methylation in pathogen-infected plants (Vi-rus-induced plant genome instability). Nucleic Acids Res. 35: 1714-1725
Brini F, Gaxiola R, Berkowitz G, Masmoudi K (2005): Cloning and characterization of a wheat vacuolar cation/proton an-tiporter and pyrophosphatase proton pump. Plant Physiol. Biochem. 43: 347-354
Brini F, Hanin M, Mezghanni I, Berkowitz G, Masmoudi K. (2007a): Overexpression of wheat Na+/H+ antiporter TN-HX1 and H+-pyrophosphatase TVP1 improve salt and dro-ught stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 58: 301-308
Brini F, Hanin M, Lumbreras V, Amara I, Khoudi H, Hassairi A, Pagès M, Masmoudi K (2007b): Overexpression of whe-at dehydrin DHN-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 26: 2017-2026
Chandra Babu R, Shashidhar H E, Lilley J M, Thanh N D, Ray J D, Sadasivam S, Sarkarung S, O’Toole J C, Nguyen HT (2001): Variation in root penetration ability, osmotic adjus-tment and dehydration tolerance among accessions of rice adapted to rainfed lowland and upland ecosystems. Plant Breed 120: 233-238
Chinnusamy V, Gong Z, Zhu J-K (2008): Abscisic Acid-medi-ated Epigenetic Processes in Plant Development and Stress Responses. J. Integrative Plant Biol. 50: 1187-1195
Christmann A, Weiler E W, Steudle E, Grill E (2007): A hydrau-lic signal in root-to-shoot signalling of water shortage. Plant J. 52: 167-174
Chuck G, O’Connor D (2010): Small RNAs going the distance during plant development. Curr.Op. Plant Biol. 13: 40-45
Cortina C, Culianez-Macia F A (2005): Tomato abiotic stress enhanced tolerance by trehalose biosynthesis. Plant Sci. 169: 75-82
De Domenico S, Tsesmetzis N, Di Sansebastiano G P, Hughes R K, Casey R, Santino A (2007): Subcellular localisation of Medicago truncatula 9/13-hydroperoxide lyase reveals a new localisation pattern and activation mechanism for CYP74C enzymes. BMC Plant Biol. 7: 58
Dodd I C, Egea G, Davies W J (2008): Abscisic acid signalling when soil moisture is heterogeneous: decreased photoperiod sap fl ow from drying roots limits abscisic acid export to the shoots. Plant Cell Environ. 31: 1263-1274
Franco-Zorrilla J M, Valli A, Todesco M, Mateos I, Puga M I, Rubio-Somoza I, Leyva A, Weigel D, García J A, Paz-Ares J (2007): Target mimicry provides a new mechanism for regu-lation of microRNA activity. Nat. Genet. 39: 1033-1037
Fricke W, Akhiyarova G, Veselov D, Kudoyarova G (2004): Rapid and tissue-specifi c changes in ABA and in growth rate in response to salinity in barley leaves. J. Exp. Bot. 55:1115-1123
Fujii H, Chinnusamy V, Rodrigues A, Rubio S, Antoni R, Park S Y, Cutler S R, Sheen J, Rodriguez P L, Zhu J K (2009): In vitro reconstitution of an abscisic acid signalling pathway. Nature 462: 660-664
Groppa M D, Benavides M P (2008): Polyamines and abiotic stress: recent advances. Amino Acids 34: 35-45
Hara M, Shinoda Y, Tanaka Y, Kuboi T (2009): DNA binding of citrus dehydrin promoted by zinc ion. Plant Cell Environ. 32: 532-541
Hartung W, Sauter A, Hose E (2002): Abscisic acid in the xylem: where does it come from, where does it go to? J. Exp. Bot. 53: 27-32
Hause B, Schaarschmidt S (2009): The role of jasmonates in mutualistic symbioses between plants and soil-born micro-organisms. Phytochem. 70: 1589-1599

Molecular Mechanisms in Plant Abiotic Stress Response 23
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
Haywood V, Yu T S, Huang N C, Lucas W J (2005): Phloem long-distance traffi cking of GIBBERELLIC ACID-INSEN-SITIVE RNA regulates leaf development. Plant J. 42: 49-68
Holmstrom K O, Somersalo S, Mandal A, Palva E T, Welin B (2000): Improved tolerance to salinity and low temperature in transgenic tobacco producing glycine betaine. J. Exp. Bot. 51: 177-185
Hu L, Wang Z, Du H, Huang B (2010): Differential accumu-lation of dehydrins in response to water stress for hybrid and common bermudagrass genotypes differing in drought tolerance. J. Plant Physiol. 167: 103-109
Huang N-C, Yu T-S (2009): The sequences of Arabidopsis GA-INSENSITIVE RNA constitute the motifs that are necessary and suffi cient for RNA long-distance traffi cking. Plant J. 59: 921-929
Hughes R K, De Domenico S, Santino A (2009): Plant cyto-chrome CYP74 family: biochemical features, endocellular localisation, activation mechanism in plant defence and im-provements for industrial applications. ChemBioChem. 10: 1122-1133
Hugouvieux V, Kwak J M, Schroeder J I (2001): An mRNA cap binding protein, ABH1, modulates early abscisic acid signal transduction in Arabidopsis. Cell 106: 477-487
Gfeller A, Liechti R, Farmer E E (2010): Arabidopsis jasmonate signaling pathway. Sci. Signal. 3(109):cm4.
Javot H, Lauvergeat V, Santoni V, Martin-Laurent F, Güçlü J, Vinh J, Heyes J, Franck KI, Schäffner A R, Bouchez D, Ma-urel C (2003): Role of a single aquaporin isoform in root water uptake. Plant Cell 15: 509-522
Jiang F, Hartung W (2008): Long-distance signalling of abscisic acid (ABA): the factors regulating the intensity of the ABA signal. J. Exp. Bot. 59: 37-43
Jiang F, Jeschke W D, Hartung W (2004): Water fl ows in the parasitic association Rhinanthus minor/Hordeum vulgare. J. Exp. Bot. 55: 2323-2329
Jiménez J A, Alonso-Ramírez A, Nicolás C (2008): Two cDNA clones (FsDhn1 and FsClo1) up-regulated by ABA are in-volved in drought responses in Fagus sylvatica L. seeds. J. Plant Physiol. 165: 1798-807
Liu J, Maldonado-Mendoza I, Lopez-Meyer M, Cheung F, Town C D, Harrison M J (2007): Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots. Plant J. 50: 529-544
Kistner C, Parniske M (2002): Evolution of signal transduction in intracellular symbiosis. Trends Plant Sci. 7: 511-518
Kovacs D, Agoston B, Tompa P (2008): Disordered plant LEA proteins as molecular chaperones. Plant Signal Be-hav. 3: 710-713
Kovtun Y, Chiu WL, Tena G, Sheen J (2000): Functional anal-ysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 97: 2940-2945
Kuromori T, Miyaji T, Yabuuchi H, Shimizu H, Sugimoto E, Kamiya A, Moriyama, Y, Shinozaki K (2010): ABC transpor-ter AtABCG25 is involved in abscisic acid transport and res-ponses. Proc. Natl. Acad. Sci. U.S.A. 107: 2361-2366
Lakatos L, Szittya G, Silhavy D, Burgyán J (2004): Molecular mechanism of RNA silencing suppression mediated by p19 protein of tombusviruses. EMBO J. 23: 876-884
Lorenzo O, Solano R (2005): Molecular players regulating the jasmonate signalling network. Curr. Opin. Plant Biol. 8: 532-540
Lough T J, Lucas W J (2006): Integrative plant biology: role of phloem long-distance macromolecular traffi cking. Annu. Rev. Plant Biol. 57: 203-232
Lucas W J, Ham B K, Kim J Y (2009): Plasmodesmata - bridg-ing the gap between neighboring plant cells. Trends Cell Biol. 19: 495-503
Luo Z-B, Janz D, Jiang X, Gobel C, Wildhagen H, Tan Y, Rennenberg H, Feussner I, Polle A (2009): Upgrading root physiology for stress tolerance by ectomycorrhizas: insights from metabolite and transcriptional profi ling into reprogramming for stress anticipation. Plant Physiol. 151: 1902–1917
Mahouachi, J, Arbona V, Gomez-Cadenas A (2007): Hormonal changes in papaya seedlings subjected to progressive water stress and re-watering. Plant Growth Regul. 53: 43-51
Masle J, Gilmore S R, Farquhar GD (2005): The ERECTA gene regulates plant transpiration effi ciency in Arabidopsis. Nature 436: 866-870
Mason H S, DeWald D B, Creelman R A, Mullet J E (1992): Coregulation of soybean vegetative storage protein gene expression by methyl jasmonate and soluble sugars. Plant Physiol. 98: 859-867
Merchan F, de Lorenzo L, Rizzo S G, Niebel A, Manyani H, Frugier F, Sousa C, Crespi M (2007): Identifi cation of regu-latory pathways involved in the reacquisition of root growth after salt stress in Medicago truncatula. Plant J. 51 :1-17
Mlynáriková L, Nap J P, Bisseling T (2007): The SWI/SNF chromatin-remodeling gene AtCHR12 mediates temporary growth arrest in Arabidopsis thaliana upon perceiving envi-ronmental stress. Plant J. 51: 874-885
Molinier J, Ries G, Zipfel C, Hohn B (2006): Transgeneration memory of stress in plants. Nature 442: 1046-1049
Moon J, Zhao Y, Dai X, Zhang W, Gray W M, Huq E, Estelle M (2007): A new CULLIN 1 mutant has altered responses to hormones and light in Arabidopsis. Plant Physiol. 143: 684-96
Mouillon J M, Eriksson S K, Harryson P (2008): Mimicking the plant cell interior under water stress by macromolecular crowding: disordered dehydrin proteins are highly resistant to structural collapse. Plant Physiol. 148: 1925-1937
Naumann M, Schüßler A, Bonfante P (2010): The obligate en-dobacteria of arbuscular mycorrhizal fungi are ancient heri-table components related to the Mollicutes. ISME J. [Epub ahead of print].
Oldroyd GE (2007): Plant Science. Nodules and hormones. Science 315: 52-53
Pandey S, Nelson D C, Assmann S M (2009): Two novel GP-CR-type G proteins are abscisic acid receptors in Arabidop-sis. Cell 136: 136-148
Pawlowski K, Sirrenberg A (2003): Symbiosis between Frankia and actinorhizal plants: root nodules of non-legumes. Indian J. Exp. Biol. 41: 1165-1183
Pennisi E (2006): Fungi Helps Grass Beat the Heat. Science 302: 774-775
Perazzolli M, Romero-Puertas M C, Delledonne M (2006): Mo-dulation of nitric oxide bioactivity by plant haemoglobins. J. Exp. Bot. 57: 479-488
Plaut Z, Butow B J, Blumenthal C S, Wrigley C W (2004): Tran-sport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water defi cit and elevated temperature. Field Crops Res. 86: 185-198
Pysh L D, Wysocka-Diller J W, Camilleri C, Bouchez D, Benfey P N (1999): The GRAS gene family in Arabidopsis: sequ-ence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 18: 111-119
Razem F A, Baron K, Hill R D (2006): Turning on gibbe-rellin and abscisic acid signaling. Curr. Opin. Plant Biol. 9: 454-459
Sauter A, Hartung W (2000): Radial transport of abscisic acid conjugates in maize roots: its implication for long distance stress signals. J. Exp. Bot. 51: 929-935
Sauter A, Davies W J, Hartung W (2001): The long-distance abscisic acid signal in the droughted plant: the fate of the hormone on its way from root to shoot. J. Exp. Bot. 52: 1991-1997

Poltronieri P et al.24
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 15-24
Schachtman D P, Goodger J Q D (2008): Chemical root to sho-ot signalling under drought. Trends Plant Sci. 13: 281-287
Schommer C, Palatnik J, Aggarwal P, Chételat A, Cubas P, Far-mer E E, Nath U, Weigel D (2008): Control of jasmonate biosynthesis and senescence by miR319 targets. PLoS Biol. 6: e230
Seki M, Umezawa T, Urano K, Shinozaki K (2007): Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 10: 296-302
Schüssler A, Martin H, Cohen D, Fitz M, Wipf D (2006): Cha-racterization of a carbohydrate transporter from symbiotic glomeromycotan fungi. Nature 444: 933-6
Simpson G G, Dijkwel PP, Quesada V, Henderson I, Dean C (2003): FY is an RNA 3′ end-processing factor that interacts with FCA to control the Arabidopsis fl oral transition. Cell 113: 777-787
Smagghe B J, Hoy J A, Percifi eld R, Kundu S, Hargrove MS, Sarath G, Hilbert JL, Watts RA, Dennis ES, Peacock WJ, Dewilde S, Moens L, Blouin GC, Olson JS, Appleby CA (2009): Review: correlations between oxygen affi nity and sequence classifi cations of plant hemoglobins. Biopolymers 1: 1083-96
Stoll M, Loveys B, Dry P (2000): Hormonal changes induced by partial rootzone drying of irrigated grapevine. J. Exp. Bot. 51: 1627-1634
Taghavi S, van der Lelie D, Hoffman A, Zhang Y-B, Walla M D, Vangronsveld J, Newman L, Monchy S (2010) Genome sequence of the plant growth promoting endophytic bacteri-um Enterobacter sp. 638. PLoS Genet. 6: e1000943
Tardieu F, Tuberosa R (2010): Dissection and modelling of abiotic stress tolerance in plants. Curr. Opin. Plant Biol. 13: 206-212
Thomas H, Howarth C J (2000): Five ways to stay green. J. Exp. Bot. 51: 329-337
Tiryaki I, Staswick P E (2002): An Arabidopsis mutant defective in jasmonate response is allelic to the auxin-signaling mutant axr1. Plant Physiol. 130: 887-894
Van der Ent S, Van Hulten M, Pozo M J, Czechowski T, Ud-vardi M K, Pieterse C M, Ton J (2009): Priming of plant innate immunity by rhizobacteria and beta-aminobutyric acid: differences and similarities in regulation. New Phytol. 183: 419-431
Weiler E W, Kutchan T M, Gorba T, Brodschelm W, Niesel U, Bublitz F (1994): The Pseudomonas phytotoxin coronatine mimics octadecanoid signalling molecules of higher plants. FEBS Lett. 345: 9-13
White J F, Torres M S (2010): Is plant endophyte-mediated de-fensive mutualism the result of oxidative stress protection? Physiol. Plant. 138: 440-446
Xoconostle-Cázares B, Xiang Y, Ruiz-Medrano R, Wang HL, Monzer J, Yoo BC, McFarland KC, Franceschi VR, Lucas WJ (1999): Plant paralog to viral movement protein that potentiates transport of mRNA into the phloem. Science 283: 94-98
Yang J, Kloepper J W, Ryu C M (2009): Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 14: 1-4
Yin P, Fan H, Hao Q, Yuan X, Wu D, Pang Y, Yan C, Li W, Wang J, Yan N (2009): Structural insights into the mechanism of abscisic acid signaling by PYL proteins. Proc. Natl. Acad. Sci. U.S.A. 16: 1230-1236
Zhang J, Subramanian S, Zhang Y, Yu O (2007): Flavone syn-thases from Medicago truncatula are fl avanone-2-hydroxylases and are important for nodulation. Plant Physiol. 144: 741-751
Zhang S, Sun L, Kragler F (2009): The phloem-delivered RNA pool contains small noncoding RNAs and interferes with translation. Plant Physiol. 150: 378-387
Molekularni mehanizmi odgovora biljaka na abiotički stres
Palmiro Poltronijeri · Stefanija Bonsenja · Stefanija De Domeniko · Anđelo Santino
ISPA-CNR, Institut za prehrambenu tehnologiju, Via Monteroni 73100 Leće, Italija
Izvod: Poboljšane sorte ratarskih useva su potrebne za održavanje zaliha hrane, borbu protiv klimatskih promena, nedostatka vode, povećanja temperature i visoke varijabilnost padavina. Varijabilnost suše i porast zaslanjenosti zemljišta imaju nepovoljan uticaj na rast biljaka, a abiotički stres ozbiljno ugrožava održivu poljoprivrednu proizvodnju. Kako bi se prevazišao uticaj abiotičkog stresa, javlja se potreba za novim, tolerantnim sortama i tehnikama oplemenjivanja uz asistiranu selekciju. Neophodno je dublje razumevanje mehanizama koji reaguju na stres i održavaju otpornost na stres. Ovde je prikazan pregled nekoliko mehanizama koji međusobno deluju pri odgovoru na stres. Danas se bolje razumeju lokalizovana sinteza biljnih hormona, sekundarni mesendžeri i lokalni efektori odgovora na abiotički stres i opstanak, kao i signalni putevi regulisani biljnim hormonima. U žiži mnogih novijih istraživanja su metaboličke mreže u odgovoru na sušu, signalizacije na velika rastojanja i komunikacija između biljnih organa pripremljenih za ekspresiju gena protiv abiotičkog stresa osobenu za pojedina tkiva. Ključne reči: abscisinska kiselina (ABA), jasmonat (JA), osmotsko prilagođavanje (OP), transkripcioni faktor (TF)

Crvenilo kukuruza: pola veka kasnije 25
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30pregledni rad / review article
Crvenilo kukuruza: pola veka kasnije
Goran Bekavac · Božana Purar · Đorđe Jocković · Sanja Treskić ·Bojan Mitrović · Ivica Đalović · Željko Milovac
primljeno / received: 15. 11. 2010. prihvaćeno / accepted: 08.12.2010© 2011 IFVC
Izvod: U prvoj deceniji 21. veka, crvenilo kukuruza se javilo u značajnijem obimu ne samo u regionu Banata, nego i u drugim krajevima Srbije. Rezultati dosadašnjih istraživanja upućuju na zaključak o fi toplazmama kao potencijalnim prouzrokovačima, cikadama kao vektorima i snažnom uticaju faktora spoljašnje sredine. Osim promena u sistemima ratarenja, plodoredu i primeni pesticida, u proizvodnju je neophodno uvesti odgovarajući sortiment. U okviru oplemenjivačkih programa identifi kovana je popu-lacija kukuruza izrazito tolerantna prema crvenilu. Sproveden je jedan ciklus kombinovane S1-HS reku-rentne selekcije sa ciljem popravke karakteristika populacije per se i njenih kombinacionih sposobnosti. Gajenjem hibrida tolerantnih prema crvenilu može se postići zadovoljavajuće i ekonomski prihvatljivo rešenje. Ključne reči: crvenilo, hibrid, kukuruz
G. Bekavac · B. Purar · Đ. Jocković · S. Treskić · B. Mitrović · I. Đalović · Ž. MilovacInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Crvenilo kukuruza (CR) i dalje zaokuplja pa-žnju istraživača i proizvođača kukuruza u Srbiji. Od 1957. godine kada je prvi put registrovano u južnom Banatu (Marić & Kosovac 1959), gotovo četiri decenije se sporadično javljalo, uglavnom u istom regionu, ne prouzrokujući ekonomski zna-čajnije štete. Crvenilo kukuruza je zabeleženo u Rumuniji 1965. godine (A. Hulea, cit. prema Iva-nović i sar. 2002) i Bugarskoj 1966. godine (M. Markov & A. Popov, cit. prema Ivanović i sar. 2002). U periodu 2002-2006. godina slična pojava primećena je i u Mađarskoj, ali je prouzrokovač crvenjenja biljaka bio MDMV virus (Kalman & Toth 2006 – lična komunikacija; cit. prema Purar et al. 2009a), što nije slučaj sa crvenilom kukuru-za u Srbiji.
Ubrzo je postalo jasno da ova pojava ne pred-stavlja samo enigmu za istraživače, nego i oz-biljnu pretnju proizvodnji kukuruza, posebno u Banatu. Tokom 2002-2003. intenzitet crvenila je varirao ne samo od lokaliteta do lokaliteta, nego i između hibrida, parcela, rokova setve, itd. Iako su postojale razlike između komercijalnih
hibrida gajenih u regionu, svi su bili manje ili više osetljivi prema crvenilu bez obzira na du-žinu vegetacije i instituciju odnosno kompaniju u kojoj su selekcionisani. Na nekim parcelama je registrovano čak do 90% biljaka sa tipičnim simptomima crvenila, što je izazvalo brojne dis-kusije i sumnju u ekonomičnost proizvodnje ku-kuruza u regionu Banata.
Simptomi
Iako ime crvenilo kukuruza možda ne opisu-je najadekvatnije sve promene koje se javljaju na biljci, u potpunosti odgovara najuočljivijem simptomu – pojavi crvene boje na vegetativnim delovima biljke. Promena boje uglavnom počinje krajem jula ili početkom avgusta dostižući svoj maksimum sredinom avgusta do prve polovine septembra. Biljke zahvaćene crvenilom rastu normalno i ni po čemu se ne razlikuju od ostalih biljaka sve do generativne faze. Prvi simptomi, promena svetlo zelene boje centralnog nerva u crvenu odnosno crveno-ljubičastu, mogu se uo-čiti na listovima iz čijih se rukavaca javlja klip, odnosno listovima iznad klipa. Od centralnog nerva, crvena boja se širi prema osnovi lista i marginama, odnosno vrhu lista. Slični simptomi se vrlo brzo mogu uočiti na susednim listovima i lisnim rukavcima, a zatim na svim delovima biljke (Bekavac et al. 2007). Biljke kod kojih se
Fitopatologija / Phytopathologyw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Bekavac G i sar.26
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
crvenilo proširilo na sve delove biljke ubrzano venu i uginjavaju. Simptomi venjenja i uginja-vanja su posebno izraženi ukoliko je ovaj pro-ces praćen sušom i visokim temperaturama. Sa venjenjem dolazi do gubitka crvene boje tkiva i to od vrha prema osnovi. Na kraju biljke dobi-jaju specifi čnu tamno-smeđu, gotovo crnu boju po čemu se na kraju vegetacije jasno razlikuju od biljaka koje su normalno fi ziološki sazrele. Osim promena na vegetativnim delovima bilja-ka, promene se uočavaju i na klipovima. Klipovi biljaka zahvaćenih crvenilom su gumaste forme, nerazvijeni sa sitnim, klimavim i nenalivenim zr-nima. Kao posledica napred navedenih prome-na, dolazi do značajnog smanjenja prinosa zrna kukuruza.
Postoji širok spektar variranja u formi kli-pa, ozrnjenosti i nalivenosti zrna kod biljaka sa simptomima crvenila (Sl. 1). Kod biljaka sa navedenim simptomima na vegetativnim delovi-ma, tipični simptomi se uglavnom mogu uočiti i na klipu. Ipak, neke biljke kod kojih se simp-tomi uočavaju na vegetativnim delovima mogu imati potpuno normalan klip sa kompletnim se-tom dobro nalivenih zrna (Bekavac et al. 2007). Posebna specifi čnost crvenila kukuruza u nas je što se promena normalne zelene boje biljnog tkiva u crvenu dešava samo na delovima biljke izloženim uticajima spoljašnje sredine. Delovi stabla pokriveni lisnim rukavcima ostaju zeleni do kraja vegetacije tako da biljke izgledaju kao da su bile izložene uticaju toplih i suvih vetrova (efekat fena).
Priroda crvenila kukuruza
Dosadašnja istraživanja nisu u potpunosti dala odgovor na pitanje šta je prouzrokovač
crvenila kukuruza. Po nekim autorima (Rosić i sar. 1973, Kerečki 2003) crvenilo kukuruza je posledica ekstremnih agroekoloških uslova, dok je po drugima (Marić & Kosovac 1959, Marić 1971, Šutić 1987, Šutić i sar. 2002, 2003) crveni-lo kukuruza prouzrokovano patogenim organiz-mima. U poslednjih nekoliko godina značajan doprinos rasvetljavanju ovog problema dali su domaći istraživači. Proučavajući pojavu crvenila kukuruza u regionu Banata, korišćenjem mole-kularnih tehnika Duduk & Bertaccini (2006) su ustanovili prisustvo stolbur fi otplazme (podgru-pa 16SrXII-A) u uzorcima biljaka koje su poka-zivale simptome crvenila. S obzirom da je stol-bur fi toplazma detektovana samo na biljkama sa simptomima, autori su zaključili da je pomenuta
fi toplazma verovatni prouzrokovač crvenila ku-kuruza. Ipak, autori se ograđuju ističući da je ovo prvi publikovan rad u svetu o prisustvu stolbur fi toplazme na kukuruzu i da su neophodna do-datna biološka istraživanja kojima bi se potvrdi-li dobijeni rezultati, odnosno pomoću kojih bi se zadovoljili Kohovi postulati. Fitoplazme su prokariotski organizmi koji nastanjuju fl oem ve-likog broja divljih i gajenih biljaka i prisutne su praktično u celom svetu. Simptomi zaraženosti biljaka fi toplazmama se generalno mogu opisa-ti kao žućenje, dekoloracija, pojava neprirod-no zelenih listova, i pojava kržljavih, odnosno žbunastih biljaka (Duduk & Bertaccini 2006). Verovatno je ovo bio glavni razlog što se dugo vremena nije sumnjalo na fi toplazme kao prou-zrokovače crvenila, jer se navedeni simptomi ne javljaju na biljkama kukuruza u nas. Poteškoće u radu sa ovim organizmima predstavlja činjenica da se ne mogu gajiti u kulturi i da moraju biti preneti sa zaražene na zdravu biljku insektima, kalemljenjem ili parazitnim biljkama (Weintraub
Slika 1. Različiti efekti na klipu biljaka kukuruza zahvaćenih crvenilomFig. 1 Different effects of corn reddening on the ears of affected plants

Crvenilo kukuruza: pola veka kasnije 27
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
& Beanland 2006). Proučavajući potencijalne prenosioce stolbur fi toplazme na kukuruz, Jović et al. (2007, 2009) su identifi kovali 33 vrste, ali su samo populacije cikade Reptalus panzeri Löw pokazivale karakteristike glavnog vektora. Pre-ma istim autorima, faktori spoljašnje sredine imaju značajnu ulogu u pojavi i intenzitetu crve-nila i ovom fenomenu posebno pogoduju ranije setve kukuruza, topla i suva leta. Međutim, od ovog pravila takođe postoje odstupanja. Tako se 1959. navodi kao izuzetno povoljna godina za proizvodnju kukuruza sa aspekta količine i rasporeda padavina, ali i godina sa značajnom pojavom crvenila (Marić 1971). Slično se može reći i za 2010. U mnogim regionima Srbije bilo je više nego dovoljno padavina, dobro raspo-ređenih tokom vegetacije, ali se i ova godina može okarakterisati kao godina sa intenzivnom pojavom crvenila.
Imajući u vidu izuzetnu složenost biotičkih i abiotičkih faktora, Purar et al. (2009a, 2009b) su sproveli seriju istraživanja sa ciljem utvrđivanja uticaja biotičkih, odnosno abiotičkih faktora na crvenilo kukuruza. Ogledi sa insekticidima za tretiranje semena i folijarnu primenu tokom vegetacije su izvođeni sa ciljem da se utvrdi veza između insekata, kao direktnih ili mogućih vektora bioloških agenasa i crvenila kukuruza. Kako svaki živi organizam jednog agroekosiste-ma ima specifi čan biološki ciklus, postavljeni su ogledi sa ciljem utvrđivanja veze između razli-čitih rokova setve i pojave crvenila kukuruza u smislu podudaranja fenofaza razvoja kukuruza i prisustva određenih insekata. U cilju utvrđivanja biološkog porekla crvenila kukuruza, testirano je prisusto Corn Stunt Spiroplazme i nematoda u biljkama sa tipičnim simptomima crvenila. S obzirom da nedostatak nekih makro i mikro elemenata može prouzrokovati pojavu crvene boje na pojedinim delovima biljke kukuruza, određena je pH vrednost i izvršena analiza sadr-žaja N, P, K, Ca i humusa u zemljištu. Na kraju, ispitivan je kvalitet semena biljaka kod kojih su ispoljeni tipični simptomi i biljaka bez simpto-ma. Ustanovljeno je da corn stunt spiroplasma i nematode ne izazivaju crvenilo, te da ne postoji jasna veza između crvenila kukuruza i datuma setve, odnosno plodnosti zemljišta. Nije bilo ra-zlike ni u klijavosti, odnosno energiji klijanja se-mena dobijenog sa biljaka zahvaćenih crvenilom u odnosu na normalne biljke, niti je primećena inferiornost istih u poljskim uslovima. Ipak, na parcelama tretiranim insekticidima uočena je slabija pojava crvenila nego na kontrolnim va-rijantama, što je ukazivalo na moguće biološko poreklo ove pojave.
U biljkama infi ciranim mikroorganizmima dolazi do poremećaja brojnih biohemijskih i fi zioloških procesa kao posledica kompleksnih odnosa biljka domaćin - patogen. Iako malo-brojni, dosadašnji rezultati su potvrdili pretpo-stavku da infekcija fi toplazmama može dovesti do promena nivoa određenih produkata me-tabolizma u biljnom tkivu. Povećanje sadržaja proteina, redukujućih šećera i fenolnih jedinje-nja, uz istovremeno smanjenje sadržaja hlorofi -la, ukazuje na mogućnost da fi toplazme remete normalne fi ziološke procese kukuruza i ubrza-vaju starenje biljnog tkiva. Kod biljaka zaraženih fi toplazmom žbunaste patuljavosti kukuruza (MBS phytoplasma - maize bushy stunt phytoplasma) ustanovljeno je povećanje sadržaja šečera u li-stu, a smanjenje u korenu, što ukazuje na pore-mećaje u transportu fotosintata fl oemom usled blokiranja istog metabolitima patogena ili biljke domaćina (Junqueira et al. 2005). Pored pojave crvene boje listova, tipični simptomi koje poka-zuju biljke infi cirane fi toplazmama jesu reduko-vana visina, povećana proliferacija klipa i izra-žena kržljavost. Sve ovo nije tipično za crvenilo kukuruza u nas. Pošto različiti patogeni mogu izazivati kako tipične, tako i netipične simptome (Nault 1980), s pravom se postavlja pitanje – da li je crvenilo kukuruza u Srbiji prouzrokovano samo stolbur fi toplazmom ili je u pitanju mnogo kompleksniji problem?
Kako dalje ?
U traženju odgovora na ovo pitanje raz-matraju se brojne opcije. S obzirom da nema pravilnosti u smislu uticaja faktora spoljašnje sredine, kao i da biotička priroda ovog feno-mena nije u potpunosti rasvetljena, intenzivna istraživanja se nastavljaju. Šta god da je prou-zrokovač crvenila kukuruza, interesantan izvor otpornosti (populacija NS 1-257 CRS) identi-fi kovan je u genetskoj kolekciji Instituta za ra-tarstvo i povrtarstvo, Novi Sad (Bekavac et al. 2007). NS 1-257 CRS je srednje rana populacija kukuruza (FAO450) stvorena ukrštanjem linija B14, 103/II, i NS 257. Samooplodna linija B14 je izvedena iz sintetika Iowa Stiff Stalk Synt-hetic (BSSS C0), linija 103/II je domaća lini-ja selekcionisana iz populacije šire genetičke osnove SG 77-A, a linija NS 257 je izvedena iz lokalne populacije Rumski zuban. Samoo-plodna potomstva populacije NS 1-257 CRS su pokazala izuzetnu tolerantnost prema crvenilu kukuruza. U dvogodišnjem ispitivanju (2004-2005) među S1 familijama nije identifi kovana ni jedna biljka sa simptomima crvenila (Sl. 2).

Bekavac G i sar.28
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
S1 – samooplodna potomstva populacije NS 1-257 CRS / selfed progenies of the population NS 1-257 CRS
TC – test-cross potomstva populacije NS 1-257 CRS / test-cross progenies of the population NS 1-257 CRS
M St – srednja vrednost korišćenih standarda / mean value of the used check hybrids
W St - srednja vrednost dva najosetljivija stan-darda / mean value of two the most susceptible check hybrids
Naknadna ispitivanja, sprovedena u periodu 2006-2010. potvrdila su potpunu tolerantnost ove populacije prema crvenilu. Nažalost, to je do sada jedini izvor tolerantnosti u genetskoj kolek-ciji Instituta za ratarstvo i povrtarstvo.
Oplemenjivanje kao mera borbe
U rešavanju brojnih problema u poljoprivre-di, stvaranje genotipova tolerantnih prema bio-tičkim i abiotičkim faktorima pokazalo se kao efi kasno, relativno jeftino i održivo rešenje. Sa praktičnog stanovišta u jedan hibrid je teško ugraditi visoku tolerantnost prema stresnim faktorima i istovremeno visoku produktivnost. Purar et al. (2009a) ističu da su neki visokopri-nosni hibridi pokazivali najvišu osetljivot prema crvenilu i shodno tome najznačajniju redukciju prinosa (Sl. 3).
Loc. N – lokaliteti bez pojave crvenila / locati-ons without CR occurrence
Loc. R – lokaliteti sa pojavom crvenila / locati-ons with CR occurrence
0,00
2,00
4,00
6,00
8,00
10,00
12,00
% C
R b
iljak
a
20042005
2004 0,00 0,32 6,71 10,80
2005 0,00 0,30 5,84 8,73
S1 TC M St W St
Slika 2. Srednje vrednosti crvenila kukuruza Fig. 2. Mean values of corn reddening
6,00
7,00
8,00
9,00
10,00
11,00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Prin
os (t
/ha)
0
2
4
6
8
10
12
% C
R b
iljak
a
Prinos zrna, Loc. N Prinos zrna, Loc. R CR, Loc. R
Slika 3. Prosečne vrednosti za prinos zrna (t/ha) i pojavu crvenila (%) hibrida kukuruza (1-15)Fig. 3. Mean values for grain yield (t/ha) and CR occurrence (%) of corn hybrids (1-15)

Crvenilo kukuruza: pola veka kasnije 29
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
Iz tih razloga, koncipiran je oplemenjivački program sa ciljem stvaranja hibrida kukuruza tolerantnih prema crvenilu. Objektivni problem predstavlja činjenica da se pomenuti izvor ot-pornosti (populacija NS 1-257 CRS) ne može direktno koristiti u komercijalnom oplemenjiva-nju jer su mu kako svojstva per se tako i kombi-nacione sposobnosti na prilično niskom nivou. Nepostojanje tolerantnosti u grupi materijala koji bi mogli poslužiti kao heterotični par za po-pulaciju NS 1-257 CRS dodatno otežava stvara-nje otpornih hibrida. U cilju popravke osnovne populacije sprovodi se program rekurentne se-lekcije na najznačajnija agronomska svojstva uz istovremeno zadržavanje nepromenjenog nivoa tolerantnosti prema crvenilu. Osim toga, ukršta-njem sa samooplodnim linijama iste heterotične grupe pokušava se stvoriti novi materijal pobolj-šanih kombinacionih svojstava i visoke toleran-tnosti prema crvenilu. Do sada je kompletiran jedan ciklus kombinovane S1-HS rekurentne selekcije. HS potomstva dobijena ukrštanjem sa inbred-testerom (linija NS 78-83) bila su oset-ljivija prema CR nego samooplodna potomstva populacije NS 1-257 CRS (Bekavac i sar. 2010), ali je tolerantnost istih bila značajno viša od ko-mercijalnih hibrida koji se gaje u širokoj proi-zvodnji. Osim toga, vrše se intenzivna testiranja velikog broja populacija i sintetika, potencijalnih heterotičnih parova, sa ciljem identifi kacije onih sa najvišim nivoom tolerantnosti. Ukrštanjem takvih materijala sa linijama izvedenim iz popu-lacije NS 1-257 CRS postigao bi se postavljeni cilj - hibrid visoke tolerantnosti prema crvenilu kukuruza i odgovarajućih agronomskih perfor-mansi.
Agrotehničke mere borbe
Za iznalaženje efi kasnih mera borbe neophod-no je poznavati prirodu crvenila kukuruza (pro-uzrokovače, vektore, faktore spoljašnje sredine, itd). Otkriće da pored tatule, poponca i divljeg sirka, kukuruz i pšenica mogu biti rezervoari
stolbur fi toplazme i ključni domaćini vektora (R. panzeri), suštinski menja pogled na ovu pojavu i nameće neka nova rešenja. U sprovođenju agro-tehničkih mera, akcenat treba staviti na efi kasnije suzbijanje korovskih vrsta, pre svega višegodiš-njih, koji predstavljaju prirodne rezervoare stol-bur fi toplazme. Imajući u vidu zakorovljenost naših parcela, ova mera bi mogla dati značajan doprinos u borbi protiv crvenila kukuruza. Po-sebna pažnja mora se obratiti na rubne delove parcela, poljske puteve i kanale koji su po pravilu izuzetno zakorovljeni. Druga veoma važna činje-nica odnosi se na naš uobičajeni plodored kuku-ruz-pšenica. Kako su obe biljne vrste domaćini stolbur fi toplazme, plodosmena kukuruz-pšenica može povećati probleme vezane za crvenilo ku-kuruza preko povećanja brojnosti populacija R. panzeri. Bez ozime pšenice, na čijem korenu se tokom zimskog perioda hrane nimfe R. panzeri, brojnost vektora crvenila kukuruza bi bila zna-čajno smanjena jer bi njihovo preživljavanje bilo vezano samo za ishranu na korenovom sitemu vi-šegodišnjih korova (Jović et al. 2009). Uvođenje tropoljnog ili višepoljnog plodoreda bi verovatno ublažilo probleme vezane za crvenilo kukuruza, pa su dodatna istraživanja na ovom polju više nego neophodna.
Zaključak
Uprkos rezultatima dosadašnjih istraživanja, čini se da enigma o tome šta je prouzrokovač crvenila kukuruza i dalje stoji. Otkriće stolbur fi toplazme kao mogućeg prouzrokovača, identi-fi kacija prirodnih rezervoara među korovskim i drvenastim vrstama, poznavanje njihove biologi-je kao i biologije vektora, doprinela je kompletni-jem sagledavanju ovog fenomena. Osim promena u sistemima ratarenja, plodoredu i primeni pesti-cida, rad na stvaranju hibrida kukruza tolerantnih prema crvenilu je od suštinskog značaja. Jedino multidisciplinaran pristup ovako kompleksnom problemu može povećati verovatnoću iznalaže-nja zadovoljavajućih rešenja.

Bekavac G i sar.30
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 25-30
Literatura
Bekavac G, Purar B, Jocković, Đ (2007): Corn reddening: the di-sease and breeding for resistance. J. Plant Pathol. 89: 397-404
Bekavac G, Purar B, Jocković Đ, Stojaković M, Ivanović M, Malidža M, Treskić S., Nastasić A, Dolapčev S, Stanisavljević D, Mitrović B (2010): Kombinovana familijska selekcija na tolerantnost prema crvenilu kukuruza. Ratar. Povrt. / Field Veg. Crop Res. 47: 103-108
Duduk B, Bertaccini A (2006): Corn with symptoms of red-dening: New host of stolbur phytoplasma. Plant Dis. 90: 1313-1319
Ivanović D, Lević J, Marić A, Penčić V (2002): Bolesti kukuruza i njihovo suzbijanje (Corn Diseases and Corn Protection). In: Bolesti, štetočine i korovi kukuruza i njihovo suzbijanje (Diseases, Pests, and Weeds of Maize and Their Control). Institut za kukuruz-Zemun Polje, Školska knjiga, Novi Sad, Serbia, 15-262
Jović J, Cvrković T, Mitrović M, Krnjajić S, Redinbaugh M G, Pratt R C, Gingery R E, Hogenhout S A, Toševski I (2007): Roles of stolbur phytoplasma and Reptalus panzeri (Ciicinae, Auchenorrhyncha) in the epidemiology of maize redness in Serbia. Eur. J. Plant Pathol. 118: 85-89
Jović J, Cvrković T, Mitrović M, Krnjajić S, Petrović A, Redin-baugh M G, Pratt R C, Hogenhout S A, Toševski I (2009): Stolbur phytoplasma transmission to maize by Reptalus pan-zeri and the disease cycle of maizer in Serbia. Phytopathol. 99: 1053-1061
Junqueira A, Bedendo I, Pascholati S (2005): Biochemical chan-ges in corn plants infected by the maize bushy stunt phyto-plasma. Physiol. Mol. Plant Pathol. 65: 181-185
Kerečki B (2003): Uticaj suše i visokih temperatura na fi ziološke promene u biljci kukuruza (Drought and high temperature effects on physiological changes in maize plants). Zbornik
referata XXXVII Seminara agronoma, Institut za ratarstvo i povrtarstvo, Novi Sad 99-106
Marić A (1971): Bolesti kukuruza , U: Kolektiv autora, Zašti-ta kukuruza od štetočina, bolesti i korova. Zadružna knjiga, Beograd
Marić A, Kosovac Z (1959): Proučavanje uzroka i štetnosti cr-venila kukuruza u Vojvodini (Study of causes and harmful-ness of corn reddening in Vojvodina). Savrem. Poljopr. 12: 1028-1043
Nault L R (1980): Maize bushy stunt and corn stunt: A compa-rison of disease symptoms, pathogen host ranges and vec-tors. Phytopathol. 70: 659-662
Purar B, Bekavac G, Jocković Đ, Toldi Toth E, Kalman L, Ra-spudić E, Dimitrijević M (2009a): Corn reddening: Occur-rence, symptoms and fi eld observations. Cereal Res. Com-mun. 37: 121-129
Purar B, Bekavac G, Jocković Đ, Sekulić P, Vasin J, Jasnič S, Raspudić E (2009b): Proučavanje crvenila kukuruza u Ba-natu. Zbornik radova Instituta za ratarstvo i povrtarstvo 46: 213-220
Rosić K, Kerečki B, Mišović M (1973): Selekcija hibrida kuku-ruza otpornih prema suši (Corn breeding for drought tole-rance). Arhiv za poljoprivredne nauke 26: 59-65
Šutić D (1987): Anatomija i fi ziologija bolesnih biljaka (Ana-tomy and Physiology of Diseased Plants). Nolit, Belgrad, Serbia
Šutić D, Tošić M, Starović M, Stanković R, Tomić T (2002): O crvenilu kukuruza (Redness of Maize). Zaštita bilja 53: 240-241
Šutić D, Starović M, Tošić M, Stojanović S, Kuzmanović S (2003): Novija proučavanja crvenila kukuruza u nas (New studies of corn reddening). Zbornik rezimea VI savetovanja o zaštiti bilja, 42
Weintraub P G, Beanland L (2006): Insect vectors of phytopla-smas. Annu. Rev. Entomol. 51: 91-111
Corn Reddening: Half a Century Later
Goran Bekavac · Božana Purar · Đorđe Jocković · Sanja Treskić ·Bojan Mitrović · Ivica Đalović · Željko Milovac
Institute of Field and Vegetable crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: In the fi rst decade of the 21st century, corn reddening occured on a larger scale not just in Banat, but also in some other regions of Serbia. The results of previous studies suggest phytoplasmas as potential causes, ci-ixides as vectors and strong infl uence of environmental factors. Besides the changes in corn growing practices, crop rotation, and the use of pesticides, it is necessary to introduce appropriate hybrids into corn production. Within corn breeding programs, a highly tolerant corn reddening population has been identifi ed. One cycle of combined S1-HS recurrent selection has been conducted with the goal to improve the characteristic of the population per se as well as its combining ability. Growing corn hybrids tolerant to reddening can provide satisfactorily and economically acceptable solution.Key words: corn, hybrid, reddening

Wild Lens Mill. Species Revealed by SDS –PAGE 31
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36original research article / originalni naučni rad
Genetic Relationships Among Wild Lens Mill. Species Revealed by SDS –PAGE
Galina Suvorova · Nadezhda Kornienko
received / primljeno: 17.11.2010. accepted / prihvaćeno: 30.11.2010.© 2011 IFVC
Summary: Seed storage proteins of 18 lentil accessions of the following Lens taxa: L. culinaris, L. orien-talis, L. odemensis, L. nigricans, L. ervoides, L. lamottei, L. tomentosus and L. lenticula, were analyzed by sodium dodecyl sulfate polyacrilamide gel electrophoresis (SDS–PAGE). All visible polypeptide bands were scored for presence or absence and data were used for dendrogram construction by UPGMA method. Three clusters were formed at the dendrogram: the fi rst one integrated accessions of L. orientalis and L. odemensis; the second cluster joined L. culinaris и L. tomentosus accessions, and related to them L. lamottei species, the third cluster included L. nigricans and L. ervoides accessions. Species affi liation of accessions L. orientalis к-2861, L. nigricans к-2859 and L. nigricans к-2860 was confi rmed. Taxonomic position of L. lenticula k-2858 accession was not determined exactly.Key words: lentil, SDS-PAGE, seed storage protein, wild species
G. Suvorova · N. KornienkoThe All-Russia Research Institute of Legumes and Groat Crops, P/b Streletskoye, 302502 Orel, Russiae-mail: [email protected]
Introduction
Lentil, as an important legume crop, needs further improvement in order to increase plant productivity, disease resistance and adaptability to machinery cultivation. Wild lentil species can serve as an additional source of genetic diversity for creation of new lentil cultivars.
Taxonomy of genus Lens Mill. is still a topic open to discussions. The classifi cation suggested by Barulina (1930) includes 5 species, where one cultivated species is Lens esculenta Moench. (a synonym is L. culinaris Medik) and four are wild species L. lenticula (Schreb) Alef., L. nigricans (M.B.) Godr., L. kotschyana (Boiss) Alef., L. orientalis (Boiss) Hand.-Mazz.
Czefranova (1971) made a critical study of Lens species and suggested the system including 9 taxa: L. montbretii (Fish.et May.) Davis et Peltm, L. penduncularis (Nabel) Czefr., L. culinaris Medik., L. orientalis (Boiss) Schmalh., L. cyanea (Boiss. et Hohen.), L. nigricans (Bieb.) Webl et Berth., L. lamottei Czefr., L. ervoides (Brign) Grande, L. unifl ora (Ten.) Schur. The species L. montbretii is a synonim of L. kotschyana species, and L. ervoides is the one for L. lenticula. As a result there was re-
established authors’ priority for L. orientalis and L. nigricans, renamed and described L. lamottei, restored 2 species L. unifl ora and L. cyanea.
Studying wild Lens species and making interspecifi c crossings, Ladizinsky (1984) suggested his own system of Lens genus. Due to an interspecifi c incompatibility and hybrid sterility, different Lens accessions were grouped in 2 botanic species: L. culinaris and L. nigricans. L. culinaris and contained 3 subspecies: ssp. culinaris, ssp. orientalis, ssp. odemensis, while L. nigricans included ssp. nigricans and ssp. ervoides. It deserves mentioning that ssp. odemensis was excluded by Ladizinsky from L. nigricans on the bases of morphological differences of stipules and incompatibility with other forms of L. nigricans.
Summing up 15 years of research in wild species collecting, studying and hybridization, Ladizinsky (1993) came to a conclusion that in Lens genus there should be isolated such species as L. culinaris ssp. culinaris, L. culinaris ssp. orientalis, L. odemensis, L. ervoides, L. nigricans. Taxon L. montbretii should be returned to Vicia genus as Vicia montbretii. This species is different not only morphologically, but in the number of chromosomes (2n = 12) whereas the Lens genus chromosomes number is 2n = 14.
Genetic Resources / Genetički resursiw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Acknowledgements: The researchers were supported by the Ru-ssian Foundation for Basic Research (09-04-97516).

Suvorova G, Kornienko N32
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
Two more species were later added to Lens genus (Oss et al. 1997). The researches admitted L .lamottei Czefr. species identifi ed by Czefranova (1971) in herbarium samples of L. nigricans. Some known populations of this species came from France, Spain, and Morocco. Ladizinsky isolated L. tomentosus Ladiz. species, that was known earlier as a L. orientalis variant and was different from the latter by its pubescent pods.
Though there was an opinion that L. tomentosus and L. odemensis should be reduced to sub-species of L. culinaris (Ferguson et al. 2000), data of molecular and cytological analysis (Galasso 2003, Sonnante et al. 2003, Duran 2004) confi rmed isolation of L. lamottei and L. tomentosus to independent species.
So, most contemporary researches recognized the next seven taxa of genus Lens: L. culinaris Medik. (L. esculenta Moench.), L. orientalis (Boiss.) Hand.-Mazz., L. odemensis Ladiz., L. nigricans (M.B.) Godr., L. ervoides (Brign.) Grande, L. lamottei Czefr., L. tomentosus Ladiz. Only L. culinaris is a cultivated species, and all the rest are wild ones. In this study we used species defi nitions for Lens culinaris and Lens orientalis (Boiss.) Schmalh, accepted by Russian botanist Czefranova (1971).
The objective of this study was to establish phylogenetic relations between the species of genus Lens and to clarify systematic position of some lentil accessions on the base of seed storage proteins SDS-PAGE analysis.
Materials and Methods
Materials for research were 18 Lens accessions of different origins, covering all up to date known taxa (Tab. 1). Lentil accessions were obtained from N. I. Vavilov Research Institute of Plant Industry (VIR), where part of them that originally belong to VIR collection was marked with letter k, and accessions fi rstly received from the International Center for Agricultural Research in the Dry Areas (ICARDA) retained their original code. Seed storage proteins were analyzed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) in gel of 12.5% concentration (Konarev et al. 2000). Electrophoretic spectra were analyzed on the base of presence or absence of all visible protein bands. Dissimilarity value (1-F) was estimated according to Nei and Li (1978) and dendrogram was constructed by UPGMA method.
Table 1. List of lentil accessions used in SDS-PAGE analysisTabela 1. Spisak akcesija sočiva korišćenih u SDS-PAGE analizi
NumberBroj
SpeciesVrsta
Accession nameNaziv akcesije
Country of originZemlja porekla
1 L. orientalis ILWL 7 Turkey2 L. orientalis ILWL 11 Syria3 L. orientalis k-2861 Syria4 L. lenticula k-2858 Turkey5 L. tomentosus ILWL 90 Turkey6 L. tomentosus ILWL 120 Syria7 L. odemensis ILWL 21 Palestine8 L. odemensis ILWL 164 Syria9 L. lamottei ILWL 29 Spain10 L. lamottei ILWL 428 Spain11 L. ervoides ILWL 43 Croatia12 L. ervoides ILWL 54 Palestine13 L. nigricans ILWL 30 Spain14 L. nigricans k-2859 Italy15 L. nigricans k-2860 Turkey16 L. culinaris Vekhovskaya Russia17 L. culinaris Obraztsov Chifl ik 7 Bulgaria 18 L. culinaris Svetlaya Russia

Wild Lens Mill. Species Revealed by SDS –PAGE 33
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
Results and Discussion
The Electrophoregram revealed 75 polypeptide bands of different insensitivity; most of them were polymorphic (Fig. 1). The greatest protein diversity was observed in area of 30-65 kDa. Dendrogram, created on the base of seed storage protein electrophoretic spectra, showed that Lens
species were quite different from each other and accessions of every species formed independent groups except of L tomentosus samples (Fig. 2).
The intraspecifi c variation of lentil in seed storage protein spectra was signifi cantly lower than the interspecifi c one. Nevertheless, high diversity was observed inside of L. orientalis, L.
Figure 1. Protein profi les of Lens species generated by SDS-PAGE. M - marker of molecular weight (soybean protein). Lanes: 1, 2, 3 - L. orientalis; 4 – L. lenticula; 5,6 - L. tomentosus; 7,8 - L. odemensis; 9,10 - L. lamottei; 11,12 - L. ervoides; 13,14,15 - L. nigricans; 16,17,18 - L. culinaris (numeration corresponds to Table 1)Slika 1. Proteinski profi li vrsta roda Lens species stvoreni pomoću SDS-PAGE. M - marker molekularne mase (protein soje). Trake: 1, 2, 3 - L. orientalis; 4 – L. lenticula; 5, 6 - L. tomentosus; 7, 8 - L. odemensis; 9, 10 - L. lamottei; 11, 12 - L. ervoides; 13, 14, 15 - L. nigricans; 16, 17, 18 - L. culinaris (brojevi odgovaraju onima u tabeli 1)
Figure 2. Dendrogram constructed on the base of Nei and Li distance. 1, 2, 3 - L. orientalis; 4 – L. lenticula; 5,6 - L. tomentosus; 7,8 - L. odemensis; 9,10 - L. lamottei; 11,12 - L. ervoides; 13,14,15 - L. nigricans; 16,17,18 - L. culinaris (numeration corresponds to Table 1)Slika 2. Dendrogram konstruisan na osnovu udaljenosti Neija i Lija. 1, 2, 3 - L. orientalis; 4 – L. lenticula; 5, 6 - L. tomentosus; 7, 8 - L. odemensis; 9, 10 - L. lamottei; 11, 12 - L. ervoides; 13, 14, 15 - L. nigricans; 16, 17, 18 - L. culinaris (brojevi odgovaraju onima u tabeli 1)

Suvorova G, Kornienko N34
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
nigricans, L. culinaris species. The accessions of L. odemensis and L. ervoides were less distinguished and L. lamottei samples were characterized by absolutely identical spectra. It was interesting that L. orientalis k-2861 accession from VIR collection was quite closely connected to L. orientalis ILWL11 accession from ICARDA, L. nigricans k-2859 and k-2860 accessions were close to each other and belonged to L. nigricans ILWL 30 group that proved their species affi liation. Accession k-2858 known as L. lenticula appeared to be in one group with L. orientalis, though not very close.
However, in spite of obvious relation of L. lenticula k-2858 accession to L. orientalis, at the moment there is no sound evidence to determine it as L. orientalis species. At the same time, those accessions were located at the dendrogram far enough from L. ervoides, species which was supposed to be a synonym of L. lenticula by Chefranova (1971). Dissimilarity value between к-2858 and L. ervoides accessions was high and means 0.723 and 0.745. On the whole, L. lenticula was quite distant from all analyzed species and its dissimilarity coeffi cient between the nearest L. orientalis ILWL7 accession was also at high level (0.422). Therefore, the described situation did not clarify the systematic position of L. lenticula.
Presumably three clusters can be isolated on the dendrogram. The fi rst cluster integrates accessions of L. orientalis and L. odemensis species. The second one joins L. culinaris and L. tomentosus accessions, and related to them L. lamottei species. The third cluster includes L. nigricans and L. ervoides species. According to electrophoretic spectra of seed storage proteins, it was revealed that L. orientalis appeared to be closely related to L. odemensis than to other species; L. culinaris was found to be closely relative to L. tomentosus; L. nigricans was supposed to be closely allied to L. ervoides. Though L. lamottei was grouped into one cluster with L. culinaris and L. tomentosus, dissimilarity value between L. lamottei and other species was not less than 0.522.
SDS-PAGE analysis was applied for seed proteins of wild lentil relative by other researchers (Sammour 1994, Ahmad et al. 1997, Zimmniak-Przybylska et al. 2001). Studying fi ve Lens taxa, Sammour (1994) and Ahmad et al. (1997) reached a conclusion that L. orientalis and L. odemensis appeared to be the wild progenitors
of L. culinaris. A complete set of taxa including recently recognized L. tomentosus and L. lamottei was used by Zimmniak-Przybylska et al. (2001). As for relationships among studied taxa the most closely relative species were L. culinaris and L. odemensis from the one side, and L. orientalis and L. tomentosus from the other side.
Our results are to some extent in contradiction with the above mentioned regarding the nearest relatives of L. culinaris. This discrepancy among our result and data of Zimmniak-Przybylska (2001) revealed by the same method can be explained by different Lens accessions used in analysis.
In contrast, Duran & Peres de la Vega (2004) using RAPD and ISSR markers found that three taxa L. culinaris, L. orientalis and L. tomentosus grouped into one cluster, with L. tomentosus being the closest species to cultivated lentil.
Applying multi-target fl uorescence in situ hybridization (FISH) on seven Lens taxa I. Galasso (2003) concluded that the most similar karyotype to cultivated lentil was that of L. orientalis, whereas L. nigricans and L. tomentosus were two species that showed the most divergent FISH patterns. At the same time, Galasso suggested that two L. tomentosus accessions (ILWL 149 and ILWL 120) were probably misclassifi ed based on the fact that their chromosome morphology and FISH karyotypes were more similar to L. culinaris than to L. tomentosus.
One of the L. tomentosus accessions used at the present study was ILWL 120. If the hypothesis of misclassifi cation can be accepted, it explains the fact of close similarity among L. tomentosus and L. culinaris cultivars revealed in our experiment.
Conclusions
Having performed the SDS-PAGE analysis of lentil seed storage protein of 8 taxa of Lens genus, we ascertained genetic relations among the species and reached the conclusion that L. tomentosus ILWL 120 is the closest accession to cultivated lentil and confi rmed specifi c affi liation of L. orientalis k-2861 and L. nigricans k-2859 and k-2860 samples of the collection of N. I. Vavilov Research Institute of Plant Industry. The obtained results can be useful in taxonomic defi nition of various accessions, interspecifi c hybridization and breeding programs.

Wild Lens Mill. Species Revealed by SDS –PAGE 35
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
References
Ahmad M, Fautrier A G, Burritt D J, McNell D L (1997): Ge-netic diversity and genetic relationships in Lens species and their interspecifi c hybrids as determined by SDS-PAGE. New Zealand J. Crop Hort. Sci. 25: 99-108
Barulina E (1930): Chechevitsa SSSR i drugih stran. Nauka, Leningrad
Chefranova Z (1971): Obzor vidov roda Lens Mill. Novosti si-stematiki vysshikh rastenii 8: 184-191
Duran Y, Peres de la Vega M (2004): Assessment of genetic va-riation and species relationships in a collection of Lens using RAPD and ISSR. Spanish J. Agric. Res. 2: 538-544
Ferguson M E, Maxted N, Van Slageren M, Robertson L D (2000): A re-assessment of the taxonomy of Lens Mill. (Le-guminose, Papilionoideae, Vicieae). Bot. J. Linnean Society 133: 41-59
Galasso I (2003): Distribution of highly repeated DNA sequ-ences in species of the genus Lens Mill. Genome 46: 1118-1124
Ladizinsky G (1993): Wild Lentils. Crit. Rev. Plant Sci. 12: 169-184
Ladizinsky G, Braun D, Goshen D, Meuhlbauer F (1984): The biological species of the genus Lens L. Bot. Gaz. 145: 253-261
Oss H, Aron Y, Ladizinsky G (1997): Chloroplast DNA va-riation and evolution in the genus Lens Mill. Theor. Appl. Genet. 94: 452-457
Sammour R H (1994): Species relationships in genus Lens as indicated by electrophoresis. Feders Repert. 105: 283-286
Sonante G, Galasso I, Pignone D (2003): ITS sequence analysis and phylogenetic inference in genus Lens Mill. Ann. Bot. 91: 49-54
Zimniak-Przybylska Z, Przybylska J, Krajewski P (2001): Elec-trophoretic seed globulin patterns and species relationships in the genus Lens Miller. J. Appl. Genet. 42: 435-447

Suvorova G, Kornienko N36
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 31-36
Genetički odnosi između samoniklih vrsta Lens Mill. otkriveni pomoću SDS –PAGE
Galina Suvorova · Nadežda Kornjijenko
Sveruski istraživački institut za mahunarke i strna žita, Arjol, Rusija
Izvod: Izvršena je elektroforeza natrijum-dodecil-sulfatnim poliakrilamidnim gelom (SDS-PAGE) rezervnih pro-teina semena 18 akcesija sledećih taksona roda Lens: L. culinaris, L. orientalis, L. odemensis, L. nigricans, L. ervoides, L. lamottei, L. tomentosus i L. lenticula. Ispitano je prisustvo svih vidljivih polipeptidnih traka, a podaci su poslužili za pravljenje dendrograma UPGMA metodom. Dendrogram su činila tri klastera: prvi sa združenim akcesijama L. orientalis i L. odemensis, drugi sa združenim akcesijama L. culinaris i L. tomentosus i njima srodnom vrstom L. lamottei i treći koji je uključio akcesije L. nigricans i L. ervoides. Bila je potvrđena srodnost akcesija L. orientalis k-2861, L. nigricans k-2859 i L. nigricans k-2860. Taksonomski položaj akcesije L. lenticula k-2858 nije bilo moguće precizno odrediti.Ključne reči: rezervni proteini semena, samonikle vrste, sočivo, SDS-PAGE

Organogeneza i prinos dvoredog ječma 37
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48originalni naučni rad / original research article
Značaj faze organogeneze formiranje klasića u biologiji prinosa ozimog dvoredog ječma
Novo Pržulj · Vojislava Momčilović
received / primljeno: 23.11.2010. accepted / prihvaćeno: 28.12.2010.© 2011 IFVC
Izvod: Biljke prolaze kroz tačno defi nisane faze razvića koje mogu biti predstavljene na osnovu prome-na koje se dešavaju na fenotipu biljke ili na osnovu promena u morfologiji apikalnog meristema. Dife-rencijacija apikalnog meristema odvija se kroz faze koje se nalaze u određenom hronološkom redosledu. U radu je analiziran značaj dužine formiranja primordija klasića, faza dvostrukog grebena (DR) i njene genotipske i fenotipske povezanosti sa ostalim fazama razvića i komponentama prinosa. Tokom pet sezona (od 2002-2003. do 2006-2007) proučavano je razviće apikalnog meristema kod tri sorte ozimog dvoredog ječma Novosadski 525, Sonate i Monaco. DR faza je bila najkraća u sezonama kada su vladali povoljni vremenski uslovi tokom odvijanja ove faze, a najduža kada su uslovi bili nepovoljni, pre svega visoke temperature. Najkraći DR period imala je ranozrela sorta Novosadski 525, a najduži srednje rana sorta Sonate. Utvrđene su slične genotipske i fenotipske korelacije između dužine DR i dužine trajanja ostalih faza razvića i komponenti prinosa. Dužina DR perioda bila je u pozitivnoj korelaciji sa dužinom perioda formiranja primordija listova, a u negativnoj korelaciji sa ostalim fazama razvića, periodom na-livanja zrna, brojem listova i klasova i žetvenim indeksom. Formiranje primordija listova i klasića kao i razviće cvetova i klasića nalazi se pod najvećom kontrolom genotipa, gde je učešće varijanse sorte preko 80% i godine do 10%. Dobijeni rezultati mogu imati praktičan značaj u oplemenjivanju ozimog dvore-dog ječma. Za potvrđivanje ovih rezultata neophodna su detaljnija istraživanja sa većim brojem sorti.Ključne reči: faza dvostrukog grebena, genotipske i fenotipske korelacije, ječam (Hordeum vulgare L.), organogeneza
N. Pržulj · V. Momčilović Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Postoji tačno određen i predvidljiv redosled faza razvića strnih žita. Mehanizmi koji leže u osnovi ovih procesa nisu u potpunosti rasvetljeni i uglavnom su empirijski (McMaster 2005). Faze razvića biljaka mogu biti predstavljene na dva načina: (i) promenama koje se dešavaju unutra apikalnog meristema (unutrašnje promene), i (ii) spoljašnjim ili fenotipskim promenama (Zadoks skala, Feekes skala, Haun skala). U literaturi po-znate skale razvića žita zasnivaju se samo na jed-noj od dve vrste promena, unutrašnjim ili spoljaš-njim, dok ne postoji zvanična skala koja predstav-lja odnose između promena apikalnog meristema i fenotipa biljke (Landes & Porter 1989, Hay & Kirby 1991). Kod ječma i većine vrsta iz porodi-ce Poaceae vegetacioni meristem stvara primordije listova i klasića u hronološkom redosledu, tako
da su sve primordije listova formirane kada se zametne primordija prvog klasića, odnosno za-metanje prve primordije klasića označava prekid daljeg formiranja primordija listova. U periodu dok apikalni meristem formira primordije klasi-ća, primordije listova se izdužuju i formiraju se listovi.
Diferencijacija apikalnog meristema odvija se kroz faze u sledećem redosledu: formiranje pri-mordija listova, formiranje primordija klasića, diferencijacija klasića i cvetova, formiranje pri-mordije terminalnog klasića i odvajanje klasa od osnove biljke. Na početku formiranja primordija, primordije klasića ne mogu se razlikovati od pri-mordija listova, i većina istraživača smatra da jako proširenje tačke rasta predstavlja početak for-miranja primordija klasića (George 1982). Prvi vidljiv signal da su klasići zametnuti je formira-nje dvostrukog grebena, ali je tada već formira-
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )Ova istraživanja su sprovedena u okviru projekta TR 031066 koji fi -nansira Ministarstvo za nauku i tehnološki razvoj Republike Srbije. / This research has been conducted within the project TR 031066 fun-ded by the Ministry of Science and Technological Development of the Republic of Serbia.

Pržulj N, Momčilović V38
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
no pola primordija klasića. Formiranje klasića u velikoj meri zavisi od genotipa i sredine (Baker & Gallagher 1983). Kod ječma je teško odrediti završetak formiranja primordija klasića, dok se poslednje primordije ne diferenciraju u klasiće i abortiraju (Arisnabarreta & Miralles 2006), što znači da se prekid aktivnosti apikalnog meriste-ma ne podudara sa zametanjem poslednjeg klasi-ća koji je vidljiv u klasanju.
Broj formiranih primordija klasa nalazi se u li-nearnoj regresiji sa sumom aktivnih temperatura (GDD) jer (a) postoji visoka podudarnost linije regresije sa eksperimentalnim podacima, (b) nije predložen drugi statistički model koji u većoj meri odražava biološki prirodu formiranja broja primordija klasića i (c) izračunata vrednost stope formiranja primordija omogućava poređenje iz-među različitih sorti ili eksperimentalnih tretma-na (Gallagher et al. 1976).
Sve primordije ne formiraju klasiće i svi for-mirani klasići ne donose zrno, jer dolazi do nji-hovog odumiranja. Broj odumrlih klasića teško je odrediti jer se mora raditi precizna disekcija i mora postojati vešt istraživač, gde ocena neće biti podložna subjektivnosti. Broj izumrlih klasi-ća određuje se na osnovu razlike između maksi-malnog broja formiranih klasića i fertilnih klasića (McMaster et al. 1992). Autori Gallagher et al. (1976) smatraju da je 58% formiranih klasića fer-tilno u fazi cvetanja kod primarne vlati i 53% kod primarnog bokora. Konkurencija za hranljivim materijama, unutar klasa i između klasa i drugih organa biljke, može biti uzrok odumiranja klasića pre cvetanja (Kirby 1988).
Određivanje veze između faza fenotipa biljke i morfologije apikalnog meristema doprinosi bo-ljem upoznavanju koordiniranosti procesa razvića i daje osnovu u modeliranju razvića biljke. Proce-njivanje faze razvića apikalnog meristema važno ja za daljnji napredak u oplemenjivanju, za tehnologi-ju gajenja i izradu modela rastenja (Kirby 1994).
U većini slučajeva skale fenotipskog razvića ne nalaze se u jakim vezama sa promenama api-kalnog meristema, čija se faza mora odrediti di-sekcijom i mikroskopiranjem (Landes & Porter 1989). Međutim, razviće biljaka strnih žita nalazi se u visokoj koordinaciji sa procesima pojavljiva-nja listova, odnosno razviće apikalnog meristema i izduživanje listova su međuzavisne, integrisane osobine (Kirby 1990, Arduini et al. 2010), što znači da se faza razviće apikalnog meristema može odrediti na osnovu broja formiranih listo-va, bez disekcije (Kirby et al. 1985).
Cilj ovih istraživanja je bio da se odredi (a) uti-caj sorte i godine na dužinu perioda formiranja broja klasića, (b) međuzavisnost dužine trajanja
faze formiranja primordija klasića i drugih faza razvića, te (c) odnos dužine trajanja faze formira-nja klasića i komponente prinosa.
Materijal i metod rada
Eksperiment je izveden u periodu od 2002-2003. do 2006-2007. sezone u Institutu za ratar-stvo i povrtarstvo na Rimskim Šančevima. Ogled je postavljen po potpuno randomiziranom blok sistemu u tri ponavljanja sa tri sorte ozimog dvo-redog ječma: Novosadski 525 (rana sorta), Sonate (srednje-rana sorta) i Monaco (kasna sorta). Setva je obavljena sledećih datuma 09.10.2002, 15.10. 2003, 22.10.2004, 18.10.2005. i 12.10.2006, a pre-dusev u svim godinama bila je soja. Veličina osnov-ne parcelice bila je 12 m2, sa 6 redova i razmakom između redova od 20 cm, a sklop 400 biljaka po m2. Meteorološki podaci uzeti su sa Meteorološke stanice Rimski Šančevi, koja je udaljena 500 m od mesta izvođenja ogleda. U cilju sprečavanja nega-tivnog efekta bolesti i štetočina na dužinu trajanja pojedinih faza i donošenja pogrešnih zaključaka, ogled je tretiran fungicidom Tilt 250 EC u Zadoks fazi 34, sa fungicidom Artea plus u Zadoks fazi 64 i sa insekticidom Karate zeon protiv leme.
Prvi i šesti red, kao i 30 cm od početka redova sa obe strane parcelice služili su kao zaštita radi sprečavanja rubnog efekta, pa sa te površine nisu uzimane biljke ili uzorci za analizu. Za određi-vanje faza razvića apikalnog meristema - dužine vegetativnog perioda (GDD), dužine perioda dvostrukog grebena (GDD), dužine perioda pojava zastavičara na glavnom stablu -cvetanje (GDD), dužine perioda nalivanja zrna (GDD) i broja listova po biljci uzimane su po tri biljke sa korenom jednom nedeljno iz drugog i trećeg reda po ponavljanju, u periodu od nicanja do lista zastavičara na glavnom stablu. Biljke su stavljane u plastične kese i odmah prenete u laboratoriju, a zemlja je isprana pod mlazom vode. Biljke su sušene na sobnoj temperaturi i određen je broj li-stova na glavnom stablu prema Haun skali (Haun 1973). Disekcija je rađena prema metodologiji Kirby & Appleyard (1984) na svim biljkama, a posmatranje i fotografi sanje pod binokularnim mikroskopom (C. Zeiss, Stemi 2000-C) radi utvr-đivanja faze razvića. Visina biljke (cm), računajući od površine zemljišta do vrha klasa isključujući dužinu osja, određena je dve nedelje posle cveta-nja na tri biljke iz 3. i 4. reda po ponavljanju. Masa biljke u cvetanju (g), masa nadzemnog vegetativ-nog dela biljke u zrenju (g), broj klasova po biljci, masa prosečnog prinosa po klasu (g) i prosečan broj zrna po klasu određeni su na tri biljke iz 3. i 4. reda. Žetveni indeks (%) određen je na osnovu

Organogeneza i prinos dvoredog ječma 39
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
snopa ječma požnjevenog (do površine zemljišta) sa 4. i 5. reda sa 0,5 m2 u svakom ponavljanju (dužina dva reda od 125 cm), a prinos na osnovu ukupnog prinosa 4. i 5. reda, uključujući i seme od analize HI, sa dužine 9,4m i ukupne površine 3,76 m2 preračunato na kg ha-1. Na uzorcima za obračun prinosa određena je hektolitarska masa (kg hl-1) i masa hiljadu zrna (g). Određivanje mase vegetativnih delova biljaka i zrna rađeno je nakon sušenja uzoraka u sušnici u periodu od 48 časova na 70°C.
Za početak formiranja klasića uzet je mo-menat formiranja dvostrukog grebena, a za završetak faze formiranja primordija klasića momenat pojava osja na prvim formiranim klasićima, jer ječam ima indeterminantno cve-tanje i na vrhu klasa ne formira terminalni kla-sić. Prva primordija klasića formirana je u 2,5 fazi Haun skale a poslednja primordija klasića u 7,0 fazi Haun skale, što je odgovaralo pojavi prvog nodusa iznad površine zemlje (Arduini et al. 2010).
Kao vremenska skala trajanja faza korišćena je suma aktivnih temperatura (GDD), jer pojava li-stova i razviće apikalnog meristema kod žita zavi-si od temperature i varira u zavisnosti od sezone (Martin et al. 1998, Pržulj 2001). Prednost ove skale u odnosu na kalendarsku skalu je u anulira-nju temperaturnih odstupanja u odnosu na pro-sečne tokom pojedinih faza razvića i rastenja. Za baznu temperaturu uzeta je temperatura od 0°C, jer ispod te vrednosti temperatura nema efekat na procese u biljkama. Za početak svake faze uzet je momenat kada je 50% biljaka bilo u toj fazi.
Statistička analiza
Analiza je urađena u programu Statistica 9. Sor-te su tretirane kao fi ksni, a godine kao random faktori. Značajnost sorte testirana je greškom, a godine interakcijom (Steel & Torrie 1980). Heri-tabilnost je izračunata iz podataka ANOVA po formuli h2=σ2
g/σ2f, gde σ2
g predstavlja genotip-sku, a σ2
f fenotipsku varijansu (Falconer 1989).
Slika 1. Srednje dekadne temperature i ukupne padavine u sezonama 2002/03-2006/07. Fig. 1 Mean temperature and precipitation per 10-day periods during growing seasons 2002/03-2006/07

Pržulj N, Momčilović V40
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
Vremenski uslovi
Prosečne temperature na nivou višegodišnjih i sufi cit vode u oktobru u sezoni 2002-2003. omo-gućili su brzo nicanje i prolazak vegetativne faze (Sl. 1). U periodu generativne faze u novembru prosečne temperature bile su na nivou letnjih i defi cit vode. Mirovanje useva počelo je sredinom prve dekade decembra i trajalo je do kraja februa-ra. Ekstremno sušni uslovi i niske temperature bili su u periodu nastavka generativne faze u proleće, a tokom perioda nalivanja zrna sušni uslovi i viso-ke temperature. Višestruko veće količine padavina u odnosu na višegodišnji prosek i prosečne tem-perature bile su u fazi nicanja 2003-2004. sezone. Tokom vegetativne faze krajem oktobra i počet-kom novembra temperature su bile nešto niže, a tokom početka faze dvostrukog grebena krajem novembra izuzetno visoke i uz smanjene padavi-ne. Zimsko mirovanje proteklo je u normalnim uslovima. Završetak generativne faze u proleće i nalivanje zrna bili su u normalnim uslovima. U sezoni 2004-2005. temperaturni i vodni uslovi bili su povoljni tokom vegetativne faze. Karakteristika ove sezone je da su u jesen povoljni vremenski uslovi vladali duže tri nedelje u odnosu na proseč-nu godinu, što je omogućilo odvijanje generativne faze. Veoma hladno vreme u februaru i prvoj po-lovini marta odložili su nastavak vegetacije. Nasta-
vak vegetacije, vlatanje i nalivanje zrna odvijalo se u povoljnim uslovima. Nicanje i vegetativna faza ječma u sezoni 2005-2006. odvijali su se u suš-nim uslovima, a generativna faza u novembru i decembru u povoljnim uslovima. Nastavak gene-rativne faze u proleće odvijalo se u uslovima jakih temperaturnih kolebanja, dok su tokom vlatanja bile visoke temperature i defi cit vode. Prva polo-vina nalivanja zrna bila je u optimalnim uslovima, a druga u uslovima hladnog i vlažnog vremena. Visoke temperature i defi cit vode u oktobru i no-vembru u sezoni 2006-2007. sprečavale su nicanje i odvijanje vegetativne faze ozimog ječma. Povolj-ni vodni i temperaturni uslovi tokom decembra i zimskih meseci omogućili su da biljke akumuliraju potrebne sume efektivnih temperatura i nado-knade zaostatak u razviću. Završetak generativne faze, vlatanje i nalivanje odvijalo se u periodu vi-sokih temperatura i sufi cita vode.
Rezultati i diskusija
Podela životnog ciklusa rastenja i razvića prak-tična je kako u svrhu istraživanja tako i u svrhu gajenja biljaka (McMaster et al. 2005). U ovome radu proučavan je značaj perioda dvostrukog gre-bena, tokom kojeg se formiraju primordije klasi-ća na apikalnom meristemu, a koji su osnova za potencijalnu plodnost klasa i prinos.
Slika 2. Početak i kraj faze formiranja klasića kod tri sorte ozimog dvoredog ječma (VP - vegetativni period, DR - dvostruki greben, VL - period vlatanja, ZC - pojava zastavičara - cvetanje, GF - period nalivanja zrna)Fig. 2. Beginning and end of double ridge phase in three winter barley cultivars (VP - vegetative period, DR - double ridge phase, VL - jointing, ZC - fl ag leaf emergence-fl owering, GF - grain fi lling period)

Organogeneza i prinos dvoredog ječma 41
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
U odnosu na promene koje se dešavaju u api-kalnom meristemu, period do cvetanja može se podeliti u tri faze: zametanje listova (vegetativna faza - VP), zametanje klasića (rana reproduktivna
faza - DR) i porast klasa tokom vlatanja (kasna reproduktivna faza) prema autorima Slafer & Whitechurch (2001) (Sl. 2). Pojava dvostrukog grebena predstavlja tranziciju iz vegetativne faze
OsobineTrait
Genotip (G)Genotype
Godina (Y)Year G x Y Greška
Pooled errorVP 631,7** 673,9* 137,5 102,6DR 6184,8* 4254,9* 1078,6 1170,5VL 100306,2** 193210,0** 8009,4** 1421,9ZC 9610,8** 18119,8* 3041,7** 361,8GF 885,4 90575,3** 5970,4** 593,0BL 18,4** 3,8* 0,9 0,4VI 509,6** 1243,0** 19,9 13,2
VMC 321,2** 138,3 82,5** 0,9VMZ 4,7** 41,3** 3,5** 0,6PZK 0,040** 0,134** 0,017 0,006BZK 7.0* 32.8 10.1* 1.3BK 103402,2** 1188441,1** 108324,4** 10558,9HI 0,009 0,015 0,010** 0,003HM 4,1** 45,6** 3,7** 0,4
MHZ 6,0* 85,6* 20,9** 1,4PR 3469862,2** 27961075,6* 4517728,9** 149570,8
% komponenti varijanse i heritabilnost % of variance components and heritability
2Gs
2Ys 2
GYs2Es 2
bhVP 82,6 6,0 1,2 10,3 13,8DR 84,8 3,5 0,0 11,7 24,1VL 86,4 11,6 1,2 0,8 7,5ZC 78,9 12,1 6,4 2,6 6,5GF 0,0 79,8 15,2 5,0 0,0BL 97,0 1,1 0,5 1,5 92,7VI 84,4 14,1 0,2 1,4 6,0
VMC 92,1 1,4 6,3 0,2 64,1VMZ 25,3 54,0 12,5 8,2 0,5PZK 62,1 21,9 6,6 9,4 2,8BZK 0,0 33,8 23,1 43,1 0,0BK 0,0 73,6 20,0 6,5 0,0HI 0,0 8,7 39,2 52,0 0,0HM 10,3 67,8 16,1 5,9 0,2
MHZ 0,0 47,7 43,2 9,1 0,0PR 0,0 61,9 34,6 3,6 0,0
Tabela 1. Sredine kvadrata, procentualni udeo komponenti varijanse i heritabilnost za ispitivane osobine ozimog dvoredog ječmaTable 1. Mean squares, percentages of variance components and heritability for the investigated traits of two-rowed winter barley
VP-vegetativni period/vegetative period, DR- dvostruki greben/double ridge phase, VL-period vlatanja/jointing, ZC- pojava zastavičara–cvetanje /fl ag leaf emergence-fl owering, GF-periodnalivanja zrna/grain fi lling period, BL- broj listova na glavnom stablu/number of leaves on main culm, VI-visina/plant height, VMC- vegetativna masa u cvetanju/aboveground vegetative plant parts weight at fl owering, VMZ- vegetativna masa u zrenju/ aboveground vegetative plant parts weight at ripening, PZK-prinos zrna po klasu/grain yield per spike, BZK- broj zrna po klasu/grain number per spike, BK-broj klasova po biljci/number of spikes per plant, HI-žetveni indeks/harvest index, HM-hektolitarska masa/test weight, MHZ-masa hiljadu zrna/thousand-kernel weight, PR-prinos zrna/grain yield

Pržulj N, Momčilović V42
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
u generativnu fazu. Završetak vegetativne faze kod ječma fenotipski se podudara sa pojavom li-gule drugog lista (Gallagher et al. 1976) ili u Haun fazi 2,4-2,5 (Arduini et al. 2010), a kod ovsa u Haun fazi 3,6 (Sonego et al. 2000). Tokom vege-tativne faze formira se 11-12 primordija lista (Ar-duini et al. 2010). Autori Banerjee & Wienhues (1965) utvdili su da ne postoji morfološka razlika u obliku vegetacione kupe između pšenice, ječ-ma i raži do faze početka diferencijacije klasića. Inače, formiranje dvostrukog grebena, kada su primordije listova i klasova prisutne na apikal-nom meristemu, jeste drugi jasan indikator da je počela tranzicija u reproduktivnu fazu (McMaster 1997). Mi smo utvrdili pozitivnu korelaciju izme-đu razvijenost vegetacione kupe i ranozrelošću, odnosno najranija sorta Novosadski 525, prva ulazi u DR fazu a najkasnija sorta Monaco naj-kasnije (Sl. 2).
Kada se formira poslednja primordija lista (primordija lista zastavičara) kod biljke počinje reproduktivna faza (Robertson et al. 1996). Is-punjavanje zahteva za niskim temperaturama za potrebe jarovizacije takođe je jedan od indikato-ra tačke tranzicije od vegetativne ka generativnoj fazi (Robertson et al. 1996). Kod ozimog ječma jarovizacija se obavi u periodu od 28 do 35 dana (Mahfoozi et al. 2001).
Komponente varijanse
Iz praktičnog iskustva u gajenju ozimih žita poznato je da nicanje veoma zavisi prvenstveno od vlažnosti zemljišta, s obzirom da su temperature u ovom delu godine uglavnom uvek povoljne za ovu fenofazu, na osnovu čega se može doneti zaključak da su faze razvića u jesenjem delu vegetacije pod velikim uticajem negenetičkih faktora. Međutim, rezultati naših istraživanja pokazuju da je dužina perioda formiranja primordija listova i dužina perioda formiranja primordija klasića (period dvostrukog grebena - DR) uglavnom pod uticajem genotipa, gde je više od 80% variranja nastalo pod uticajem sorte (Tab. 1). Autori Arduini et al. (2010) utvrdili su da se kod sorti ozimog dvoredog ječma formira ukupno 35 primordija, od kojih su prvih 10-11 primordije listova, a ostalih 24-25 primordije klasića. Na osnovu logističke krive kojom se predstavlja formiranje primordija listova i klasića dobija se podatak da su već formirane prosečno tri primordije listova na glavnoj vegetacionoj kupi kod nicanja, dok je maksimalan broj primordija kod pojave desetog lista. Formiranje poslednje primordije odvija se u Haun fazi 6,6-7,0 kod ječma (Arduini et al. 2010) i 5,3 kod ovsa (Sonego et al. 2000).
Visok značaj genotipa za morfološke osobine koje su povezane sa brojem listova takođe je dobijen u našim ranijim istraživanjima (Pržulj & Momčilović 2002). Dužina trajanja VP bila je najkraća u sezoni 2005-2006. kada su oktobar i novembar bili nepovoljni za nicanje, a najduža u povoljnoj sezoni 2004-2005. (Tab. 2). Rana sorta Novosadski 525 u proseku imala je najkraći VP a srednje rana sorta Sonate najduži VP (Tab. 2). Najkraći period formiranja klasića bio je u 2003-2004. sezoni kada su vladali uglavnom povoljni vremenski uslovi za odvijanje životnih procesa, a najduži u sezonama 2005-2006. i 2006-2007. koje su se odlikovale visokim temperaturama u drugom delu DR. Iz navedene analize vremenskih uslova i dužine trajanja DR teško je izvesti bilo kakvu zakonomernost o uticaju temperatura i padavina na dužinu trajanja formiranja primordija klasića. Za dobijanje takve informacije potrebno je analizirati veći broj sorti, sa mnogo češćim uzimanjem uzoraka. U odnosu na genetsko variranje ne može se utvrditi zakonomernost u dužini trajanja DR; najkraći period formiranja primordija klasića bio je kod kasnozrele sorte Monaco, a najduži kod srednje rane sorte Sonate (Tab. 2 i Sl. 2).
Period vlatanja (VL) tokom kojeg se odvija diferencijacija cvetova u klasićima bio je pod kontrolom sve tri komponente varijanse, genotipa, godine i interakcije, mada je učešće genotipa bilo najveće (Tab. 1). Najduži period vlatanja za sve sorte bio je u sezoni 2006-2007. kada su bile visoke temperature i defi cit vode, a od sorti najduži period vlatanja imala je kasnozrela sorta Monaco (Tab. 2). Godina je u najvećoj meri određivala dužinu perioda nalivanja zrna (GF), dok se uticaj genotipa manifestovao samo kroz interakciju GxY (Tab. 1). Nalivanje zrna je bilo najduže u povoljnim uslovima poput sezone 2004-2005., a u nepovoljnim uslovima najkraće (Tab 2).
Mnogi faktori sredine u interakciji sa genotipom utiču na fenologiju biljke. Pošto se na osnovu nijednog pojedinačnog faktora sredine ne može predvideti razviće, temperatura i povremeno fotoperiod su najvažniji faktori koji određuje dužinu trajanja različitih faza razvića i porasta (McMaster 2005). Dostupnost vode takođe može uticati na fenologiju, posebno smanjujući sumu aktivnih temperatura u kasnijim fazama rastenja, kao što je skraćenje perioda nalivanja zrna (McMaster & Wilhelm 2003). Iako je analiziran mali broj sorti, preliminarno se može zaključiti da su nepovoljni uslovi produžavali ranije faze razvića, a skraćivali kasnije.

Organogeneza i prinos dvoredog ječma 43
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
OsobinaTrait
SortaCultivar (B)
Godina/Growing period (A) Prosek/
2002/03 2003/04 2004/05 2005/06 2006/07 Average LSD A B AB
VP
Novosadski 525 227,2 226,0 243,1 217,3 230,0 228,7 0,05 9,8 7,6 17,0
Sonate 243,3 245,2 258,4 226,8 233,3 241,4 0,01 13,2 10,2 22,9
Monaco 227,4 235,4 239,4 226,6 247,3 235,2 CV 4,4
Prosek/average 232,6 235,5 247,0 223,5 236,9 235,1
DR
Novosadski 525 357,6 308,3 346,5 340,7 394,7 349,5 0,05 33,0 25,6 57,2
Sonate 354,4 339,0 343,7 396,6 393,2 365,4 0,01 44,6 34,5 77,2
Monaco 332,8 298,3 320,6 353,4 320,4 325,1 CV 9,5
Prosek/average 348,2 315,2 336,9 363,6 369,4 346,7
VL
Novosadski 525 613,3 453,3 454,5 535,4 621,3 555,6 0,05 36,4 28,2 63,1
Sonate 638,3 459,2 529,9 498,3 733,1 591,8 0,01 49,1 38,1 85,1
Monaco 704,7 621,3 592,4 607,6 833,0 711,8 CV 5,9
Prosek/Average 652,1 511,3 525,6 547,1 862,5 619,7
ZC
Novosadski 525 59,0 178,3 103,3 67,9 122,2 106,2 0,05 18,4 14,2 31,8
Sonate 57,2 129,7 53,8 62,5 108,9 82,4 0,01 24,8 19,2 42,9
Monaco 129,2 196,2 72,8 126,7 90,2 133,0 CV 17,2
Prosek/Average 81,8 184,7 76,6 85,7 107,1 107,2
GF
Novosadski 525 855,8 882,0 1004,3 955,2 798,6 899,2 0,05 23,5 18,2 40,7
Sonate 854,7 849,0 1040,9 973,1 760,6 895,7 0,01 31,7 24,6 54,9
Monaco 869,3 756,2 1066,5 1002,2 857,7 910,4 CV 2,4
Prosek/Average 859,9 829,1 1037,2 976,8 805,6 901,7
BL
Novosadski 525 11,3 11,3 10,3 11,7 11,0 11,13 0,05 0,6 0,5 1,1
Sonate 12,7 12,3 11,3 11,0 12,7 12,00 0,01 0,9 0,7 1,5
Monaco 14,3 13,7 11,7 13,3 13,7 13,33 CV 5,5
Prosek/Average 12,78 12,44 11,11 12,00 12,44 12,16
VI
Novosadski 525 77,7 103,3 101,7 89,3 105,0 95,4 0,05 3,6 2,7 6,1
Sonate 77,0 104,3 98,3 86,0 103,3 93,8 0,01 4,7 3,7 8,2
Monaco 86,7 119,0 104,7 100,7 112,0 104,6 CV 3,7
Prosek/Average 80,4 108,9 101,6 92,0 106,8 97,9
Tabela 2. Uticaj sorte i godine na ispitivane osobine ozimog dvoredog ječmaTable 2. Effects of cultivar and year on the investigated traits of two-rowed winter barley
VP-vegetativni period/vegetative period, DR- dvostruki greben/double ridge phase, VL-period vlatanja/ jointing, ZC- pojava zastavičara–cvetanje /fl ag leaf emergence-fl owering, GF-periodnalivanja zrna/grain fi lling period, BL- broj listova na glavnom stablu/number of leaves on main culm, VI- visina biljke/
plant height

Pržulj N, Momčilović V44
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
Osobina Trait
SortaCultivar (B)
Godina/Growing period (A) Prosek
2002/03 2003/04 2004/05 2005/06 2006/07 Average LSD A B AB
VM
C Novosadski 525 2,75 4,55 5,79 8,90 7,89 5,98 0,05 0,9 0,7 1,6
Sonate 8,15 6,84 11,31 8,37 8,55 8,65 0,01 1,2 1,0 2,2Monaco 8,51 8,95 30,71 9,00 17,77 14,99 CV 9,7Prosek/Average 6,47 6,78 15,94 8,76 11,40 9,87
VM
Z
Novosadski 525 3,68 5,71 5,40 8,68 7,49 5,59 0,05 0,8 0,6 1,3Sonate 5,75 7,72 5,69 6,68 8,61 6,29 0,01 1,0 0,8 1,8Monaco 4,04 7,56 5,31 6,25 5,74 5,18 CV 14,0Prosek/Average 4,49 7,00 5,47 7,20 7,28 5,69
PZK
Novosadski 525 1,42 1,25 1,37 1,09 1,11 1,25 0,05 0,07 0,06 0,12Sonate 1,33 1,36 1,58 1,22 1,23 1,34 0,01 0,10 0,07 0,17Monaco 1,26 1,40 1,36 1,17 1,14 1,26 CV 5,7Prosek/Average 1,33 1,34 1,44 1,16 1,16 1,29
BZK
Novosadski 525 24.7 26.7 27.0 24.0 22.3 24.9 0,05 1.60 1.24 2.77Sonate 22.7 25.0 29.0 27.3 25.0 25.8 0,01 2.16 1.67 3.73Monaco 22.7 25.0 25.3 27.3 22.7 24.6 CV 6.6Prosek/Average 23.3 25.6 27.1 26.3 23.3 25.11
BK
Novosadski 525 730 1253,3 1633 967 1147 1146 0,05 99,2 76,9 171,9Sonate 1260 1260,0 1970 897 973 1272 0,01 133,9 103,7 231,8Monaco 1407 1180,0 1980 967 980 1303 CV 8,9Prosek/Average 1132 1231 1861 943 1033 1240
HI
Novosadski 525 0,454 0,354 0,411 0,447 0,406 0,414 0,05 0,054 0,042 0,094Sonate 0,357 0,371 0,496 0,463 0,355 0,408 0,01 0,073 0,056 0,126Monaco 0,521 0,467 0,530 0,386 0,357 0,452 CV 13,2Prosek/Average 0,444 0,397 0,479 0,432 0,373 0,425
HM
Novosadski 525 74,1 71,4 70,0 76,2 74,8 73,3 0,05 0,6 0,5 1,1Sonate 74,3 71,1 69,9 77,3 72,7 73,0 0,01 0,8 0,6 1,4Monaco 71,9 72,2 70,7 75,1 71,5 72,3 CV 0,9Prosek/average 73,4 71,6 70,2 76,2 73,0 72,9
MH
Z
Novosadski 525 47,4 41,5 49,5 47,1 47,8 46,7 0,05 1,1 0,9 2,0Sonate 45,3 44,5 53,0 47,7 44,1 46,9 0,01 1,5 1,2 2,6Monaco 44,7 46,7 52,7 43,5 41,0 45,7 CV 3,3Prosek/Average 45,8 44,2 51,7 46,1 44,3 46,4
PR
Novosadski 525 5600 6313,3 7600,0 9907 5813 7047 0,05 373,5 289,3 646,8Sonate 10453 6173,3 8146,7 9667 5440 7976 0,01 503,8 390,2 872,6Monaco 9240 6000,0 7653,3 9240 4360 7299 CV 6,4Prosek/Average 8431 6162 7800 9604 5204 7440
Tabela 3. Uticaj sorte i godine na ispitivane osobine ozimog dvoredog ječmaTable 3. Effects of cultivar and year on the investigated traits of two-rowed winter barley
VMC- vegetativna masa u cvetanju/aboveground vegetative plant parts weight at fl owering, VMZ- vegetativna masa u zrenju/ aboveground vegetative plant parts weight at ripening, PZK-prinos zrna po klasu/grain yield per spike, BZK- broj zrna po klasu/grain number per spike, BK-broj klasova po biljci/number of spikes per plant, HI-žetveni indeks/harvest index, HM-hektolitarska masa/test weight, MHZ-masa hiljadu zrna/thousand-kernel weight, PR-prinos zrna/grain yield

Organogeneza i prinos dvoredog ječma 45
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
Broj listova se nalazi pod najjačom kontrolom genotipa, sa najvećom heritabilnošću i u pozitiv-nom odnosu sa dužinom perioda do pojave lista zastavičara (Tab. 1). Formiranje vegetativne mase do cvetanja uglavnom je određeno sortom (Tab. 1) gde kasnozrele sorte imaju veću masu, dok je ukupna vegetativna masa u zrenju pod uticajem sve tri komponente varijabilnosti, gde godina ima najveće učešće (Tab. 1).
Udeo varijansi kod pojedinih komponenti prinosa je bio različit (Tab. 1). Godina je imala najveći uticaj na broj klasova, koji se kretao od 943 (2005-2006) do 1.861 (2004-2005) (Tab. 3). Mali broj klasova u sezoni 2005-2006. sigurno je posledica nepovoljnih uslova u fenofazi bokore-nja, odnosno veliki broj klasova u sezoni 2004-2005. rezultat je veoma povoljnih uslova, kada je jesenji deo sezone bio duži tri nedelje u odnosu na prosečnu godinu. Kasnozrela sorta Monaco, koja je najkasnije cvetala, imala je u proseku naj-veći broj klasova. Nepovoljni vremenski uslovi u sezonama 2002-2003. i 2006-2007. uslovili su smanjenje broja zrna po klasu (Tab. 3). Johnson & Kanemasu (1984) su utvrdili da visoke tempe-rature smanjuju broj klasića i broj zrna u svakom klasiću. Između sorti nije postojala statistički zna-čajna razlika u broju zrna po klasu. Prinos zrna po klasu bio je pod najjačim uticajem genotipa, mada je heritabilnost bila niska zbog visoke greš-ke. Najveći prinos po klasu imala je srednje kasna sorta Sonate (Tab. 3).
Godina i interakcija GxY imaju najveći uticaj na variranje mase hiljadu zrna i hektolitarske mase (Tab. 1). Uz dužinu perioda nalivanja, fi -zičke osobine zrna imaju najmanji koefi cijent varijabilnosti (Tab. 3). Najveća masa hiljadu zrna bila je u povoljnoj sezoni 2004-2005. a najmanja u 2006-2007. kada su tokom perioda nalivanja zrna vladale visoke temperature i bio sufi cit vode (Sl. 1 i Tab. 3). Između sorti nije bilo statistički značajne razlike u masi hiljadu zrna. Najmanja hektolitarska masa bila je u sezoni 2004-2005, a najveća u sezoni 2005-2006. (Tab. 3). Podaci koji se odnose na vegetativni biljni materijal, npr. masa vegetativnog dela u zrenju i žetveni indeks, imaju visoke vrednosti koefi cijenata varijacije, što ukazuje na neophodnost analiziranja metode uzi-manja uzoraka.
Različite faze od setve do cvetanja mogle bi biti pod različitom genetičkom kontrolom, što omo-gućava njihovo kombinovanje oplemenjivanjem bez promene ukupnog vremena do cvetanja (Sla-fer at al. 2001). Iako se prinos formira kontinui-rano od setve do zrenja, period zametanja klasića je važan u defi nisanju potencijalnog broja zrna, a period izduživanja stabljike za determinisanje
broja fertilnih cvetova u cvetanju, od kojeg zavisi konačan broj zrna (Miralles & Slafer 2007).
Genotipske i fenotipske korelacije
Poznavanje odnosa između parova kvantita-tivnih osobina i stepena njihove povezanosti ili korelacije može olakšati tumačenje dobijenih re-zultata u oplemenjivanju i doprineti uspešnijem praktičnom oplemenjivanju. Postojanje jake ge-netske korelacione veze između jedne osobine niske heritabilnosti, npr. prinos, sa drugom oso-binom visoke heritabilnosti može biti iskorišćeno za indirektno oplemenjivanje na osobinu niske heritabilnosti, putem oplemenjivanja na osobinu visoke heritabilnosti. U ovom slučaju posebno je važan nivo i tip korelacije, odnosno kovarijanse za dve odabrane osobine.
U našim istraživanjima genotipske i fenotipske korelacije između dužine perioda formiranja pri-mordija klasića i drugih ispitivanih osobina uglav-nom su slične, izuzev sa dužinom perioda vlata-nja i sa prinosom (Tab. 4). Dužina DR perioda bila je u pozitivnoj korelaciji sa dužinom perioda formiranja primordija listova, a u negativnoj ko-relaciji sa svim ostalim fazama razvića, periodom nalivanja zrna, kao sa brojem listova i klasova, vi-sinom biljke i žetvenim indeksom. Duži DR pe-riod bio je u negativnoj korelaciji sa masom cele biljke u cvetanju, što je i očekivano, s obzirom da se najveća masa biljke formira u periodu vlatanja, gde duži DR period znači kraći period vlatanja. U odnosu na komponente prinosa DR period bio je u negativnoj korelaciji sa brojem klasova po jedinici površine, gde je u stvari duži DR period značio i kraći period bokorenja. Duži DR period nalazio se u pozitivnoj genotipskoj i fenotipskoj korelaciji sa prinosom zrna po klasu, hektolitar-skom masom, masom hiljadu zrna i prinosom. Ove pozitivne korelacione veze posebno genet-ske, pokazatelj su u kom pravcu treba usmeriti selekciju.
Vrednosti genotipskih i fenotipskih korelacije slične su za većinu ispitivanih osobina (Tab. 4). Kod aditivnog modela fenotipske korelacije sa-stoje se od genotipske komponente i komponen-te sredine i njihovi odnosi određuju se relativnim značajem naslednih i nenaslednih efekata. Ako je fenotipsko variranje rezultat sličnog delovanja genetičkih i ekoloških faktora, genetske i ekološ-ke korelacije moraju biti slične, imajući u vidu da odnosi između osobina u razviću predstavljaju i korelacije. Pošto fenotipske korelacije označavaju sumu genotipskih komponenti i komponenti sre-dine, genotipske i fenotipske korelacije biće slič-ne, bez obzira na vrednost heritabilnosti. Razlike

Pržulj N, Momčilović V46
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
Osobina Godina/Growing period
Trait 2002/03-07 2002/03 2003/04 2004/05 2005/06 2006/07
VP rg 0,44 0,51 0,92 0,58 3,49 -3,94
rf 0,35 0,22 0,53 0,57 0,14 -0,59
VL rg -0,81 -1,11 -0,68 -0,92 -0,35 -1,00*
rf -0,20 -0,82 -0,65 -0,85 -0,76 -0,88
ZC rg -0,99* -1,80 -0,97* 0,21 -1,14 1,13
rf -0,60 -1,36 -0,88 0,25 0,01 0,63
GF rg -0,99* -1,54 0,50 -3,66 -0,07 -1,12
rf -0,68 -0,69 0,45 -3,03 0,12 -0,66
BL rg -0,69 -1,12 -0,24 -0,93 -0,60 -0,97*
rf -0,70 -0,69 -0,41 -0,59 -0,60 -0,58
VI rg -0,97* -1,02 -0,78 -1,18 -1,00* -1,01*
rf -0,93 -0,94 -0,45 -0,59 -0,28 -0,95
VMC rg -0,77 -0,73 -0,16 -1,07 -1,77 -1,09
rf -0,75 -0,51 -0,30 -0,87 -0,58 -0,89
VMZ rg 0,96* 0,43 0,35 1,24 -0,81 1,17
rf 0,86 0,04 0,33 0,10 -0,46 0,57
PZK rg 0,68 1,12 -0,17 0,46 1,58 0,02
rf 0,64 0,99 -0,08 0,44 0,75 0,24
BZK rg 0.94 0.89 -0.59 -4.29 1.04 0.51
rf 0.89 0.91 0.01 8.46 0.53 0.25
BK rg -0,30 -0,82 1,94 -0,98* -2,39 0,57
rf -0,07 -0,65 0,20 -0,24 -0,32 0,38
HI rg -0,98* -1,02* -1,14 -1,03* 0,20 0,89
rf -0,81 -0,29 -0,51 -0,63 0,13 0,96
HM rg 0,81 1,06* -1,40 -2,27 1,44 0,93
rf 0,81 0,89 -0,51 -0,61 0,44 0,62
MHZ rg 0,98* 0,84 -0,17 -0,64 0,85 0,94
rf 0,89 0,63 -0,13 -0,38 0,24 0,72
PR rg 0,61 -0,43 1,58 0,25 0,92 1,23
rf 0,02 -0,42 -0,43 0,42 -0,37 0,64
Tabela 4. Genotipski (rg) i fenotipski (rf) koefi cijenti korelacije između DR i ostalih ispitivanih osobina ozimog dvoredog ječmaTable 4. Genotypic (rg) and phenotypic (rf) correlations between double ridge stage and the investigated traits of two-rowed winter barley
VP-vegetativni period/vegetative period, DR- dvostruki greben/double ridge phase, VL-period vlatanja/jointing, ZC- pojava zastavičara–cvetanje /fl ag leaf emergence-fl owering, GF-periodnalivanja zrna/grain fi lling period, BL- broj listova na glavnom stablu/number of leaves on main culm, VI-visina/plant height, VMC- vegetativna masa u cvetanju/aboveground vegetative plant parts weight at fl owering, VMZ- vegetativna masa u zrenju/ aboveground vegetative plant parts weight at ripening, PZK-prinos zrna po klasu/grain yield per spike, BZK- broj zrna po klasu/grain number per spike, BK-broj klasova po biljci/number of spikes per plant, HI-žetveni indeks/harvest index, HM-hektolitarska masa/test weight, MHZ-masa hiljadu zrna/thousand-kernel weight, PR-prinos zrna/grain yield

Organogeneza i prinos dvoredog ječma 47
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
između genotipskih i fenotipskih korelacija mogu nastati usled razdvajanja načina delovanja ekološ-kih i genetičkih efekata na fenotip i/ili usled greš-ke kod uzimanja uzoraka (Cheverud 1988).
Hill & Thompson (1978) su odredili verovat-noću dobijanja negativne genetičke varijanse i genetskih korelacija, gde su proste ili parcijalne korelacije >1 ili <-1, na osnovu čega se dobila i negativna heritabilnost. Verovatnoća dobijanja ovakvih, teoretski nemogućih genetskih kovari-jansi, značajno se povećava sa smanjenjem veliči-ne uzorka i smanjenjem heritabilnosti ispitivanih osobina. Ovim se tumače i neuobičajeno visoke vrednosti fenotipskih i genotipskih koefi cijenata korelacije po godinama u našim istraživanjima (Tab. 4), zbog čega se vrednosti po godinama ne mogu smatrati relevantnim. Naime, osnovni me-todološki nedostatak ovih istraživanja je mali broj sorti, te je testiranje t-testom urađeno za 1 stepen slobode. Iz tog razloga i podaci o rg i rf za petogo-dišnji prosek nemaju statističke značajnosti, bez obzira na visoke apsolutne vrednosti. Smoček (1986) citira autore koji su dobili veliko variranje u vrednosti koefi cijenta korelacije između istog para osobina, ne samo u vrednosti nego i u pred-znaku, što su i potvrdili i naši rezultati (Tab. 3).
Vrednost heritabilnosti potencijalno utiče na sličnost fenotipske i genotipske korelacije na dva načina. Visoka heritabilnost povećava proporcio-nalni doprinos genetike fenotipskoj korelaciji, od-nosno doprinosi većoj sličnosti ove dve korelacije (Cheverud 1988). U našim istraživanjima herita-bilnost za broj listova iznosila je 92,7% (Tab. 3), a razlika između rg i rf samo 0,01 (Tab. 4). Kod niske heritabilnosti, kao što je slučaj za prinos, razlika između genotipskih i fenotipskih korelaci-ja je visoka (Tab. 3). Kada je heritabilnost visoka očekuje se mala razlika između visine vrednosti i tipa genotipskih i fenotipskih korelacija, mada u našim istraživanjima, postoje i izuzeci kao što je npr. hektolitarska masa i masa hiljadu zrna, gde su razlike između rg i rf male, iako su niske vred-nosti heritabilnosti.
Zaključak
Nepovoljni vremenski uslovi, pre svega viso-ke temperature i nedostatak vode, uticali su na produžavanje trajanja pojedinih faza razvića do cvetanja, dok su skraćivali period nalivanja zrna. Formiranje primordija listova i klasića, kao i ra-zviće cvetova i klasića nalazi se pod najvećom kontrolom genotipa, gde je učešće varijanse sor-te preko 80% a godine do 10%. Značajno uče-šće varijanse genotipa u pojedinim fazama treba iskoristiti u procesu defi nisanja genotipa i fenoti-
pa ozimog ječma za uslove jugoistočne Evrope. Poznavanje povezanost pojedinih faza može se iskoristiti za indirektnu selekciju. Kao i kod dru-gih autora i istraživanja, rezultati ovog istraživanja istakli su značaj velikog uzorka u kvantitativno-genetskim istraživanjima.
Literatura
Arduini I, Ercoli L, Mariotti M, Masoni A (2010): Coordination between plant and apex development in Hordeum vulgare ssp. distichum. Comptes Rendus Biologies 333: 454-460
Arisnabarreta S, Miralles D J (2006): Floret development and grain setting in near isogenic two- and six-rowed barley lines (Hordeum vulgare L.). Field Crop Res. 96: 466-476
Baker C K, Gallagher J N (1983): The development of winter wheat in the fi eld. 1. Relation between apical development and plant morphology within and between seasons. J. Agric. Sci. Camb. 101: 327-335
Benerjee S, Wienhues F (1965): Comparative Studies of the Development of the Spike in Wheat, Barley and Rye. Z. Pfl anzenzüchtung 54: 130-142
Cheverud J M (1988): A comparison of genetic and phenotypic correlations. Evolution 42(5): 958-968
Falconer D S (1989): Introduction to Quantitative Genetics. 3rd Ed. Logman Scientifi c & Technical, Logman House, Burnt Mill, Harlow, Essex, England
Gallagher J N, Biscoe P V, Scott R K (1976): Barley and its en-vironment. IV. Growth and development in relation to yield. J. Appl. Ecology 13: 563-583
George D W (1982): The growing point of fall-sown wheat: a useful measure of physiological development. Crop Sci. 22: 235-239
Haun J R (1973): Visual quantifi cation of wheat development. Agronomy Journal 65: 116-119.
Hay R K M, Kirby E J M (1991): Convergence and synchrony – a review of the coordination of development in wheat. Aust. J. Agric. Res. 42: 661-700
Hill W, Thompson R (1978): Probabilities of non-positive defi -nite between-group of genetic covariance matrices. Biome-trics 34: 429-439.
Johnson R C, Kanemasu E T (1984): Sensitivity of wheat pha-ses development to high temperatures. Aust. J. Plant Physiol. 10: 393-426
Kirby E J M, Appleyard M (1984): Cereal Development Gu-ide 2nd Edition. Arable unit, National Agricultural Centre, Conventry, UK, 95
Kirby E J M, Appleyard M, Fellowes G (1985): Variation in development of wheat and barley in response to sowing date and variety. J. Agric. Sci. Camb. 104: 383-360
Kirby E J M (1990): Co-ordination of leaf emergence and leaf and spikelet primordium initiation in wheat. Field Crop Res. 25: 253-264
Kirby E J M (1994): Identifi cation and prediction of stages of wheat development for management decisions. Home-Grown Cereals Authority, London, Project Report 90, 39
Landes A, Porter J R (1989): Comparision of scales used for ca-tegorising the development wheat, barley, rye and oats. Ann. Appl. Biol. 115: 343-360
Mahfoozi S, Limin A E, Fowler D B (2001): Infl uence of Ver-nalization and Photoperiod Responses on Cold Hardiness in Winter Cereals. Crop Science 41: 1006-1011.
Martin R J, Sinton S M, Jamieson P D, Sonego M (1998): The effect of temperature on leaf appearance rate in drummond oats. Proceedings of the Ninth Australian Agronomical Conference, 534-544

Pržulj N, Momčilović V48
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 37-48
McMaster G S, Wilhelm W W, Morgan J A (1992): Simulation winter wheat shoot apex phenology. J. Agric. Sci. 119: 1-12
McMaster G S (1997): Phenology, development, and growth (Triticum aestivum L.) shoot apex: A review. Adv. Agron. 59: 63-118
McMaster G S, Wilhelm W W (2003): Phenological responses of wheat and barley to water and temperature: improving simulation models. Journal of Agricultural Science, Cam-bridge 119: 1-12
McMaster G S (2005): Phytomers, phyllochrons, phenology and temperate cereal development. Journal of Agricultural Science 143: 137-150
Miralles D J, Slafer G A (2007): Sink limitations to yield in whe-at: how could it be reduced? J. Agric. Sci. 145: 139-149
Pržulj N (2001): Cultivar and year effect on grain fi lling of win-ter barley. Plant Breed. Seed Sci. 45: 45-58
Pržulj N, Momčilović V (2002): Mainstem Leaf Development during Tillering in Spring Barley. EUCARPIA, Cereal Sec-tion Meeting, Salsomaggiore, Italy, 21-25 November 2002, Book of Abstracts, 93
Robertson M J, Brooking I R, Ritchie J T (1996): Temperature response of vernalization in wheat: Modeling the effect of the fi nal number of main stem leaves. Ann. Bot. 78: 371-381
Slafer G A, Abeledo L G, Miralles D J, Gonzalez F G, Whi-techurch E M (2001): Photoperiod sensitivity during stem elongation as an avenue to raise potential yield in wheat. Euphytica 119: 191-197
Slafer G A, Whitechurch E M (2001): Manipulating wheat de-velopment to imptove adaptation and to search for alterna-tive opportunities to increase yield potential. In: Reynolds, M.P. et al. (eds) Application of Physiology in Wheat Bree-ding. CIMMYT, Mexico, DF, 160-170
Smoček J (1968): Phenotypic, Genotypic and Environmental Correlations of Quantitative Characters in Winter Wheat. Biologia Plantarum 10: 146-155
Sonego M, Moot D J, Jamieson P D, Martin R J, Scott W R (2000): Apical development in oats predicted by leaf stage. Field Crop Res. 65: 79-86
Steel R D, Torrie J H (1980): Principles and procedures of stati-stics. 2nd ed. McGraw-Hill Book Co., New York
Importance of Spikelet Formation Phasein the Yield Biology of Winter Barley
Novo Pržulj · Vojislava Momčilović
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Plants evolve through strictly defi ned stages of development, which can be identifi ed by changes in plant phenotype or by changes in the morphology of apical meristem. Differentiation of apical meristem passes through phases that proceed in a precise chronological order. This paper analyzes the signifi cance of the length of the period of spikelet primordia formation, i.e. the double ridge (DR) phase, and its genotypic and phenotypic correlation with the other growth phases and yield components. In the course of fi ve growing seasons (2002-03 to 2006-07) we studied the development of apical meristem in three two-rowed winter barley cultivars - Novi Sad 525, Sonate and Monaco. DR stage was shortest in the seasons with favorable weather conditions during that stage. It was longest when conditions were unfavorable, especially when temperatures were high. The shortest DR stage was found in the early cultivar Novosadski 525, the longest in the medium early cultivar Sonate. The cultivars had similar genotypic and phenotypic correlations between the length of DR stage on one side and the duration of the other development stages and yield components on the other side. DR period length was positively correlated with the duration of the formation of leaf primordia and negatively correlated with the other development stages, i.e. grain fi lling period, numbers of leaves and spikes and harvest index. The obtained results may fi nd practical application in winter barley breeding. Further more detailed study involving a number of cultivars is required in order to confi rm the above results.Key words: barley (Hordeum vulgare L.), double ridge stage, genotypic and phenotypic correlations, organogenesis

Novi konzumni hibridi suncokreta u NS paleti 49
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56originalni naučni rad / original research article
Novi konzumni hibridi suncokretau NS paleti
Nada Hladni · Vladimir Miklič · Siniša Jocić ·Zvonimir Sakač · Velimir Radić · Ilija Radeka
received / primljeno: 02.11.2010. prerađeno / revised: 24.11.2010. accepted / prihvaćeno: 01.12.2010.© 2011 IFVC
Izvod: Rad na oplemenjivanju konzumnog suncokreta usmeren je na povećanje genetskog potencijala za prinos, stabilnosti prinosa, zdravstvene bezbednosti i nutritivnog kvaliteta uz povećanje ekonomič-nosti proizvodnje. U skladu sa zahtevima srpskog tržišta u Institutu za ratarstvo i povrtarstvo se radi na oplemenjivanju visoko produktivnih hibrida za posebne namene konzumnog tipa. U ovom programu posebna pažnja je posvećena povećanju sadržaja i kvaliteta proteina uz smanjenje sadržaja ulja u semenu, povećanju mase 1000 semena i smanjenju udela ljuske. Novi NS-konzumni hibridi NS-H-6316, NS-H-6318 i NS-H-6320 pogodni su za ishranu, ljuštenje i proizvodnju jezgra a pripadaju grupi srednje ranih hibrida. Genetski potencijal za prinos semena im je preko 4,5 t ha-1 a sadržaj ulja u semenu je manji od 40%. Sadržaj proteina u jezgru je 23-27%, masa 1000 semena iznad 95 g, dok je udeo ljuske nizak. Stva-ranjem nove palete konzumnih NS hibrida suncokreta za različite namene i uslove proizvodnje dobija se kvalitetnija sirovina za potrebe uljarske, prehrambene i pekarske industrije. Ključne reči: konzumni tip, kvalitet semena, prinos, visokoproteinski suncokret
N. Hladni · V. Miklič · S. Jocić · Z. Sakač · V. Radić · I. RadekaInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Danas se u svetu gaje dva glavna tipa sunco-kreta, uljani i neuljani (konzumni tip). Neuljani tip suncokreta Helianthus annuus L. var. macrocarpus (DC.) Ckll. naziva se još konzumni, proteinski, gricko ili krupnoplodni suncokret. Seme niskou-ljanog suncokreta je obično crno sa belim pruga-ma ili šareno, znatno krupnije od semena uljanog tipa, sa debljom ljuskom labavo vezanom za jez-gro. Ljuska se lako odvaja od jezgra i omogućava kompletno ljuštenje semena (Gonzalez-Perez & Vereijken 2007). Konzumni tip suncokreta odli-kuje se velikim udelom ljuske, najčešće od 40% do 50% (Jovanović 2001) i visokom masom 1000 semena koja je često viša od 100g (Dozet & Jova-nović 1997). Rad na oplemenjivanju suncokreta usmeren je na povećanje: genetskog potencijala za prinos, stabilnosti prinosa, zdravstvene bez-bednosti i nutritivnog kvaliteta uz povećanje eko-nomičnosti proizvodnje (Hladni 2010). Visoko-proteinski hibridi suncokreta za konzumnu upo-trebu našli su svoje mesto u proizvodnji SAD-a, Mađarske, Slovačke, Francuske, Španije, Turske, Ukrajine, Rusije, Kine, Srbije i mnogih drugih
zemalja. Tome je doprinela sve veća briga sta-novništva o zdravlju i želja da poboljšaju kvalitet života unoseći visoko nutritivno vredne namirni-ce. Redovno korišćenje suncokretovog semena značajno podiže odbrambene snage organizma i zbog toga ono spada u funkcionalne namirnice. Seme suncokreta bogato je proteinima, uljem, vitaminima (E, B1 i B5), mineralnim materijama (mangan, magnezijum, bakar, selen, cink, gvož-đe, kalcijum), folnom kiselinom i prehrambenim vlaknima. Antioksidansi, vitamin E i selen koji su prisutni u semenu suncokreta imaju sposobnost neutralizacije štetnih slobodnih radikala i na taj način štite organizam od brojnih bolesti današnji-ce, poput kardiovaskularnih i malignih oboljenja. Sve ovo je uticalo da konzumni suncokret, bilo u svom neprerađenom ili prerađenom obliku, sve više nalazi svoje mesto u ljudskoj ishrani, pre-hrambenoj i konditorskoj industriji.
U skladu sa zahtevima srpskog i svetskog tr-žišta, u Institutu za ratarstvo i povrtarstvo u Novom Sadu radi se na oplemenjivanju sun-cokreta za posebne namene konzumnog tipa. Oplemenjivanje konzumnih hibrida usmereno je u pravcu povećanja kvaliteta i sadržaja proteina (>25%), stabilnosti ulja uz smanjenja sadržaja u semenu (<40%), mase 1000 semena (>100 g), hektolitarske mase, povećanje udela jezgra uz smanjenje udela ljuske, uniformnosti u veličini i boji semena, ljuštivosti kao i tolerantnosti na
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Hladni N i sar.50
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
dominantne bolesti u regionu gde se gaje (Hladni i sar. 2009a). Prema Miklič i sar. (2009) kontinu-iran rad na stvaranju novih konzumnih hibrida visokog i stabilnog prinosa semena značajan je za proizvođače suncokreta i prehrambenu in-dustriju. Dva najznačajnija kriterijuma za uvo-đenje konzumnih (visoko proteinskih) hibrida u proizvodnju su visok prinos semena i proteina (Hladni i sar. 2009b). Sadržaj proteina u semenu je jedan od pokazatelja kvaliteta semena sunco-kreta, kvantitativno je svojstvo determinisano većim brojem gena (minor geni ili poligeni). Pre-ma autorima Jovanović & Stanojević (1996) i Jovanović et al. (1996) sadržaj proteina u semenu varira u od 13% do 28%. Sa povećanjem jezgra povećava se i količina proteina u semenu, tako da oplemenjivanje na povećanje količine pro-teina u semenu treba da prati izbor genotipova sa što krupnijim jezgrom (Hladni i sar. 2009a). Masa 1000 semena spada u glavne komponente prinosa, oplemenjivanje na povećanje mase 1000 semena dovodi do povećanja prinosa semena, te se ovo svojstvo koristi kao selekcioni kriterijum kod stvaranja hibrida suncokreta (Hladni et al. 2008). Ustanovljena je visoka pozitivna korela-cija između mase 1000 semena i prinosa semena (Dušanić et al. 2004, Hladni et al. 2008).
Dobro poznati NS-konzumni hibridi Vranac i Cepko i dalje su prisutni na tržištu Srbije, EU, Ru-sije i Ukrajine. Povećanje potražnje u industrijskoj proizvodnji za proteinskim proizvodima na bazi suncokreta (seme, jezgro, proteinsko brašno) po-stavilo je nove zahteve pri stvaranju konzumnih hibrida. Potvrda uspešnog oplemenjivačkog rada na stvaranju NS-konzumnih hibrida jeste pri-znavanje novog konzumnog hibrida Mahelan (NS-H-6318) u Republici Slovačkoj (EU) 2010. Zahvaljujući kvalitetu i dobroj prilagođenosti ra-zličitim uslovima spoljne sredine NS-hibridi us-pevaju da budu registrovani u zemljama EU i po-lako ulaze na tržište EU gde postoji potražnja za semenom suncokreta visoke hranjive vrednosti. Velik uticaj na ostvareni prinos konzumnih hibri-da suncokreta imaju genetski potencijal rodnosti hibrida, vremenske prilike (količina i raspored pa-davina) u semiaridnom rejonu u kome se nalazi Srbija i primenjena tehnologija gajenja (Hladni i sar. 2009d).
Cilj ovog rada je ocena novih visokoprotein-skih hibrida suncokreta za konzumnu upotrebu: Mahelan (NS-H-6318), NS-H-6316 i NS-H-6320 na osnovu rezultata dobijenih iz Sortne komisije Srbije i Slovačke u 2009. kao i ocena proizvodnih karakteristika i stepen adaptabilnosti novih hi-brida u poređenju sa dobro poznatim hibridima Vranac i Cepko na osnovu dvogodišnjih i trogo-
dišnjih rezultata dobijenih u regionima Vojvodine i centralne Srbije.
Materijal i metod
Analizirani su novi visokoproteinski dvolinij-ski hibridi konzumnog tipa (NS-H-6316, NS-H-6318, NS-H-6320) stvoreni ukrštanjem citoplaz-matsko muško sterilne linije majke i linije oca sa genom za restauraciju fertilnosti. U radu će se prikazati deo rezultata dobijenih od Sortne komi-sije Ministarstva poljoprivrede Republike Srbije i Republike Slovačke, kao i rezultati dobijeni ispi-tivanjem ovih hibrida u uslovima regiona Vojvo-dine i regionu centralne Srbije. NS konzumni hi-bridi su ispitivani na tri lokaliteta Sortne komisije Srbije u poređenju sa dva standarda Proteinac 94 i Cepko, te na tri lokaliteta Slovačke sortne komi-sije u poređenju sa hibridima drugih semenskih kompanija. Prikazaće se deo ocene UPOV-a hi-brida NS-H-6318 (Mahelan) priznatog u Sortnoj komisiji Slovačke. Prinos semena obračunat je na 9% vlage.
Konzumni hibridi su ispitivani na dva lokaliteta Kula i Srbobran u toku 2007. i Kula i Rimski Šančevi u toku 2008. i 2009. radi ocene proi-zvodnih karakteristika i stepena adaptabilnosti. Njihove proizvodne karakteristike (prinos se-mena i sadržaj ulja) su poređene sa standardnim konzumnim hibridima Cepko (Kongo) i Vranac (Rio) koji su priznati u Sortnoj komisiji Srbije i Sortnoj komisiji Slovačke. U dvogodišnjim ogle-dima na lokalitetu Rimski Šančevi (2008-2009) ispitivan je prinos semena, proteina i ulja, kao i kvalitet semena: masa 1000 semena, sadržaj ulja u semenu, sadržaj proteina u semenu, sa-držaj proteina u jezgru. Ogledi su postavljeni po slučajnom blok sistemu u tri ponavljanja sa osnovnom parcelicom od 28 m2 (4 reda). U toku vegetacije su rađena fenološka opažanja, a u fazi fi ziološkog zrenja ocenjena je otpornost na do-minantne bolesti. Za berbu su korišćene biljke iz dva srednja reda. Prinos semena je izmeren na vagi i preračunat na 9% sadržaja vlage u se-menu. Određen je sadržaj ulja u semenu (%) iz apsolutno čistog i vazdušno suvog semena na NMR-analizatoru. Sadržaj proteina u semenu i jezgru određen je po metodi Kjeldahla. Masa 1000 semena (g) izmerena je na slučajnom uzor-ku apsolutno čistog i vazdušno suvog semena. Utvrđen je prinos proteina iz prinosa semena i sadržaja proteina u semenu (t ha-1).
Statistička obrada podataka je urađena u pro-gramu Excel i Statistica 7. Aritmetička sredina kao pokazatelj centralne tendencije i analiza vari-janse urađeni su prema Hadživukoviću (1991).

Novi konzumni hibridi suncokreta u NS paleti 51
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
Rezultati i diskusija
Stabilan prinos i kvalitet semena su najvažnija svojstva NS konzumnih hibrida suncokreta. Vi-sina prinosa zavisi od genetskog potencijala za prinos, a stabilnost prinosa zavisi od sposobno-sti reagovanja hibrida na uslove sredine (Hladni i sar. 2009d). Zato se genetski potencijal za prinos hibrida suncokreta razlikuje od poljoprivrednog prinosa. Poljoprivredni prinos je pod velikim uticajem ekoloških faktora, što dovodi do velike varijabilnosti u prinosu u zavisnosti od lokaliteta i godine, zato je važno uvoditi u proizvodnju sta-bilne hibride suncokreta koji postižu visoke pri-nose u različitim uslovima sredine (Miklič i sar.
2010). Rezultati dobijeni u Sortnoj komisiji Srbije, Sortnoj komisiji Republike Slovačke u 2009. i tro-godišnji i dvogodišnji ogledi Instituta za ratarstvo i povrtarstvo omogućiće pravu ocenu vrednosti novih konzumnih hibrida. Dobijeni rezultati za 2009. iz Sortne komisije Srbije za prinos semena, masu 1000 semena i sadržaj ulja pokazali su da NS konzumni hibridi imaju veći prosečan prinos semena i masu 1000 semena, a manji prosečan sadržaj ulja od standarda Proteinac 94 i Cepko (Tab. 1).
Najviši prosečan prinos semena ostvario je hi-brid NS-H-6318 od 3,97 t ha-1. Prosečna masa 1000 semena novih NS konzumnih hibrida bila je iznad 100 g, što je znatno više od oba standar-
Tabela 1. Prosečan prinos semena (t ha-1), masa 1000 semena (g) i sadržaj ulja u semenu (%) NSkonzumnih hibrida na 3 lokaliteta u toku 2009. u ogledima državne Sortne komisije SrbijeTable 1. Average seed yield (t ha-1), 1000 seed weight (g) and seed oil content (%) of the NS confectionary hybrids at 3 locations during 2009 in the offi cial trials by the Varietal Release Committee of Serbia HibridiHybrids
Prinos semena Seed yield (t ha-1)
Masa 1000 semena1000 seed weight (g)
Sadržaj ulja u semenuSeed oil content (%)
Lokaliteti Locations Lokaliteti Locations Lokaliteti LocationsN S P S PH/AH R N S P S PH/AH R N S P S PH/AH R
NSH 6320 3,81 3,18 3,60 3,53 3 118,5 98,7 86,4 101,2 3 37,7 37,4 38,2 37,8 4NSH 6318 4,17 3,56 4,18 3,97 1 113,0 97,4 100,4 103,8 2 40,4 38,3 39,6 39,4 3NSH 6316 3,76 3,29 3,58 3,54 2 122,5 104,4 113,9 113,6 1 35,0 34,0 35,3 34,8 5P/A 3,91 3,34 3,79 3,68 118,2 100,2 100,2 106,2 37,7 36,6 37,7 37,3Proteinac 94 4,32 2,52 3,10 3,31 5 98,1 81,5 71,0 83,5 4 44,3 43,5 43,9 43,9 1Cepko 3,94 2,96 3,17 3,36 4 81,7 70,0 56,5 69,4 5 42,0 41,9 43,0 42,3 2
LSD Hibrid LokalitetHybrid Location
Hibrid LokalitetHybrid Location
Hibrid LokalitetHybrid Location
LSD 0,05 0,69 11,85 1,00LSD 0,01 1,04 17,94 1,51
Lokaliteti/ location: NS-Novi Sad, P-Pančevo, S-Sombor; P-prosek lokaliteta/A-average of one location; PH-prosek hibrida /AH-average of hybridsR-rang/rank
Tabela 2. Prosečan prinos (t ha-1) NS konzumnih hibrida na 3 lokaliteta u toku 2009. u ogledima državne Sortne komisije SlovačkeTable 2. Average yield (t ha-1) of the NS confectionary hybrids at 3 locations during 2009 in the offi cial trials by the Varietal Release Committee of Slovakia
HibridiHybrids
Prinos semena / Seed yield (t ha-1) Lokaliteti / LocationsBAH VME VRV PH/AH R
H1 4,17 2,95 3,85 3,66 7H2 3,97 2,90 4,62 3,83 4H3 4,0 3,46 4,33 3,93 3
NS-H-6318 4,72 3,73 4,58 4,34 1NS-H-6316 4,40 3,37 4,62 4,13 2NS-H-6320 4,47 3,05 3,61 3,71 6
H4 4,04 3,05 4,21 3,77 5P/A 4,25 3,22 4,26 3,91
LSD Hibrid Lokalitet/Hybrid LocationLSD 0,05 0,530LSD 0,01 0,754
* H1-H4 su hibridi drugih kompanija koji su se nalazili u postupku registracije; P-prosek lokaliteta/A-average of one location; PH-prosek hibrida /AH-average of hybridsR rang/rank

Hladni N i sar.52
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
da (83,5 g; 69,4 g). Kod sva tri ispitivana hibri-da prosečan sadržaj ulja je bio niži od 40%, dok su Proteinac 94 i Cepko imali sadržaj ulja iznad 40% (43,9%; 42,3%). Najviša masa 1000 semena (113,6 g), a najniži sadržaj ulja (34,8%) ustanov-ljen je kod hibrida NS-H-6316 (Tab. 1).
Ispitivanja Slovačke sortne komisije su poka-zala da su hibridi NS-H-6318(4,34 t ha-1), NS-H-6316(4,13 t ha-1) i NS-H-6320(3,71 t ha-1) ostvarili visoke prosečne prinose semena (Tab. 2).
Prosečni prinos NS konzumnih hibrida (4,06 t ha-1) bio je znatno viši od prosečnog prinosa svih ispitivanih hibrida (3,91 t ha-1). Od svih ispitiva-nih hibrida najviši prosečan prinos postigao je hi-brid NS-H-6318 ostvarivši znatno viši prinos od prinosa ispitivanih hibrida iz drugih semenskih kompanija (Tab. 2). Merenjem mase 1000 semena u Sortnoj komisiji Srbije i Slovačke dobijen je op-šti prosek veći od 100 g za sva tri hibrida. Najvišu prosečnu masu 1000 semena imao je hibrid NS-
H-6316 (117,8 g) (Tab. 3). Opšti prosek prinosa NS konzumnih hibrida na tri lokaliteta Sortne komisije Srbije i Slovačke je bio 3,87 t ha-1.
Najviši opšti prosek prinosa ostvario je hibrid NS-H-6318 (4,16 t ha-1). Ostvareni prosečan pri-nos semena i masa 1000 semena su bili viši u Slo-vačkoj sortnoj komisiji (4,06 t ha-1; 115,0 g) nego u Sortnoj komisiji Srbije (3,68 t ha-1; 106,2 g). U uslovima Srbije i Slovačke novi konzumni hibridi postigli su visoke prinose potvrđujući svoju pla-stičnost, tj. mogućnost gajenja u različitim agroe-kološkim uslovima (Tab. 3).
Prema autorima Vranceanu (2000) i Škorić i sar. (2005, 2006) pravilna slika o vrednosti hi-brida dobija se nakon višegodišnjeg ispitivanja. Stoga je važno videti uticaj agroekoloških uslo-va na ostvarenje genetskog potencijala hibrida uključivši u ispitivanje osim lokaliteta i godine kao faktora (Miklič i sar. 2010). Da bi se videla prilagođenost novih konzumnih hibrida našim
Tabela 3. Prosečan prinos semena (t ha-1) i masa 1000 semena NS konzumnih hibrida na 3 lokaliteta u toku 2009. u ogledima Sortne komisije Srbije i Slovačke Table 3. Average seed yield (t ha-1) and 1000 seed weight (g) of the NS confectionary hybrids at 3 locations during 2009 in the offi cial trials by the Varietal Release Committees of Serbia and Slovakia
Hibridi Hybrids
Prinos semena / Seed yield (t ha-1) Masa 1000 semena / 1000 seed weight (g)Srbija Slovačka PH/AH R Srbija Slovačka PH/AH R
NSH 6320 3,53 3,71 3,62 3 101,2 111,1 106,2 3NSH 6318 3,97 4,34 4,16 1 103,8 111,9 107,9 2NSH 6316 3,54 4,13 3,84 2 113,6 121,9 117,8 1
P/A 3,68 4,06 3,87 106,2 115,0 110,6
P-prosek lokaliteta za prinos semena i masu 1000 semena /A-average location for seed yield and 1000 seed weight;PH-sredina proseka hibrida za prinos semena i masu 1000 semena /AH-average hybrid for seed yield and 1000 seed weight; R rang/rank
Tabela 4. Prosečni prinos semena i sadržaj ulja NS konzumnih hibrida ostvarenih na dva lokaliteta u toku tri godine (2007-2009) na ogledima Instituta za ratarstvo i povrtarstvoTable 4. Average seed yield and seed oil content of the NS hybrids acquired at two locations in three-year trials (2007-2009) at the Institute of Field and Vegetable Crops Hibridi Hybrids Prinos semena / Seed yield (t ha-1) Sadržaj ulja u semenu / Seed oli content (%)
07 08 09 SP/MA R 07 08 09 SP/MA RNS-H-6316 4,84 3,88 4,40 4,37 2 30,3 31,0 27,9 29,7 5NS-H-6318 4,94 3,99 4,45 4,46 1 36,5 35,4 34,3 35,4 3NS-H-6320 4,75 3,94 4,04 4,24 3 35,3 33,9 34,7 34,7 4
P/A 4,85 3,94 4,30 4,36 34,0 33,4 32,3 33,2Vranac 4,32 3,45 3,74 3,84 4 44,6 44,1 41,6 43,8 1Cepko 3,83 3,65 3,83 3,77 5 40,9 40,9 39,8 40,5 2
LSD Hibrid Godina/Hybrid Year Hibrid Godina/Hybrid YearLSD 0,05 0,404 1,503LSD 0,01 0,612 2,277
P-prosek /A-average;SP-sredina proseka /MA-mean average;R rang/rank

Novi konzumni hibridi suncokreta u NS paleti 53
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
proizvodnim uslovima, oni su ispitivani tokom 2007-2009. na dva lokaliteta u ogledima Instituta za ratarstvo i povrtarstvo u poređenju sa standar-dima Vranac i Cepko koji već imaju svoje mesto u proizvodnji. Agroekološki uslovi u sve tri godi-ne ispitivanja (2007, 2008. i 2009) bili su različiti ali su pogodovali gajenju suncokreta koji dobro podnosi visoke temperature praćene sušom. Vi-soke prosečne prinose ostvarili su konzumni hi-bridi NS-H-6318 (4,46 t ha-1), NS-H-6316 (4,37 t ha-1) i NS-H-6320 (4,24 t ha-1) u trogodišnjim ispitivanjima na dva lokaliteta Kuli (region cen-tralne Srbije) i Srbobran i Rimski Šančevi (region Vojvodine) u odnosu na kontrole Vranac i Cepko (3,84 t ha-1; 3,77 t ha-1). Prosečan sadržaj ulja kod sva tri hibrida NS-H-6316 (29,7%), NS-H-6318 (35,4%), NS-H-6320 (34,7%) bio je znatno niži od standarda (43,8; 40,5%) (Tab. 4).
Analiza varijanse je pokazala da postoji razlika u varijabilnosti u ekspresiji prinosa semena i sadržaja ulja u semenu. Sredina kvadrata genotipa, godine i lokaliteta je visoko značajna i ukazuje na postoja-nje različitog efekta svih uspitivanih godina, diver-gentnost analiziranih hibrida kao i divergentnost lokaliteta, što potvrđuje i visoko značajna interak-cija genotip/ godina/ lokalitet (Tab. 5).
Sve veća potražnja za konzumnim hibridima zahteva detaljnije ispitivanje kvaliteta semena koji pored genetskog faktora zavisi i od uslova spoljne sredine. Analiza kvaliteta semena prikazana u ovom radu omogućiće proizvođačima uvid o kvalitetu konzumnih hibrida koji se već nalaze u proizvod-nji, kao i hibridima koji treba svojim osobinama da dokažu svoju vrednost i da nađu svoje mesto u proizvodnji. Novi konzumni hibridi u dvogo-dišnjim ispitivanjima na lokalitetu Rimski Šančevi
Tabela 5. Trofaktorijalna analiza varijanse za prinos semena i sadržaj ulja u semenuTable 5. Trifactorial analysis of variance for seed yield and seed oil content
Izvori varijacije Sources of variation DFPrinos semena
Seed yieldSadržaj ulja u semenu
Seed oil contentMS F test MS F test
Ponavljanje - Replication 2Genotip - Genotype (G) 4 1,867 11,670** 562,1 14052**Godina - Year (Y) 2 4,278 26,739** 50,8 1269**Lokalitet - Location (L) 1 63,908 399,424** 0,1 2**Genotip/Godina - Genotype/Year 8 0,220* 1,377* 5,6 139**Genotip/Lokalitet - Genotype/Location 4 0,625 3,906** 5,8 145**Godina/Lokalitet - Year /Location 2 8,429 52,681** 14,9 374**G/L/G G/L/Y 8 0,256 1,597** 3,0 74**Greška – Error (E) 60 0,160 0,04Ukupno - TOTAL 89
DF-stepeni slobode (Degrees of freedom); MS-sredina kvadrata (Mean Squares)
Tabela 6. Prosečne vrednosti mase 1000 semena, sadržaja proteina u semenu i jezgru, prosečni prinosi semena, proteina i ulja ostvareni u ogledima Instituta za ratarstvo i povrtarstvo u 2008-2009. Table 6. Average 1000 seed weight, seed oil content, seed and kernel protein content and average yields of seed, protein and oil from the trails performed at the Institute of Field and Vegetable Crops during 2008-2009
HibridiHybrids
MHSTSW R SUS
SOC R SPSSPC R SPJ
KPC R PSSY R PU
OY R PPPY R
g % % % tha-1 tha-1 tha-1
NS-H-6316 120,1 1 30,7 5 13,6 5 24,9 1 5,14 2 1,578 5 0,699 4NS-H-6318 117,4 2 35,5 3 14,6 4 23,6 2 5,15 1 1,827 1 0,767 2NS-H-6320 103,7 3 34,6 4 15,0 3 22,4 4 5,07 3 1,753 3 0,775 1
P/A 113,7 33,6 14,4 23,6 5,12 1,719 0,752Vranac 73,4 5 43,2 1 15,3 2 21,9 5 3,85 5 1,664 4 0,589 5Cepko 79,3 4 39,9 2 15,8 1 23,5 3 4,45 4 1,811 2 0,703 3
Masa 1000 semena MHS/1000 seed weight TSW; Sadržaj ulja u semenu SUJ/Seed oil content SOC; Sadržaj proteina u semenu SPS/Seed protein content SPC; Sadržaj proteina u jezgru SPJ/Kernel protein content KPC; Prinos semena PS/Seed yield SY; Prinos ulja PU/Oil yield OY; Prinos proteina PP/Protein yield PY; P-prosek /A-average ;R rang/rank

Hladni N i sar.54
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
imali su znatno višu prosečnu vrednost mase 1000 semena (103,7 g; 117,4 g; 120,1 g) u poređenju sa standardima (73,4 g; 79,3 g). Najveću masu 1000 semena imao je hibrid NS-H-6316 (120,1 g) što je u saglasnosti sa rezultatima dobijenim iz Sortne komisije Srbije (Tab. 1. i Tab. 6).
Ustanovljen je znatno niži prosečni sadržaj ulja u semenu novih konzumnih hibrida (30,7%; 34,6%; 35,5%) u odnosu na sadržaj ulja kod hibrida Cepko i Vranac (43,2%; 39,9%). Novi konzumni hibridi ostvarili su nešto niži prosečan sadržaj proteina u semenu od standarda, dok je prosečan sadržaj proteina u jezgru bio na nivou boljeg standarda ili nešto viši (Tab. 6). Ostvareni prinos proteina kod svih ispitivanih hibrida NS-H-6320 (0,775 t ha-1), NS-H-6318 (0,767 t ha-1) i NS-H-6316 (0,699 t ha-1) je bio na nivou boljeg standarda ili nešto viši (0,703 t ha-1). Na osnovu ispitivanja Sortne komisije Srbije, Sortne komi-sije Slovačke i ispitivanja u ogledima Instituta za ratarstvo i povrtarstvo posebno se istakao novi hibrid NS-H-6318 po ostvarenom prosečnom prinosu semena. Postižući dobre rezultate kako u uslovima Slovačke tako i u uslovima Srbije po-kazuje svoju plastičnost, tj. mogućnost gajenja u različitim agroekološkim uslovima. Prema re-zultatima Slovačke sortne komisije hibrid Ma-helan (NS-H-6318) poseduje dobre tehničko-tehnološke karakteristike za hibrid konzumnog tipa, što mu je omogučilo priznavanje u 2010. Hibrid Mahelan je u dvogodišnjim ispitivanjima Slovačke sortne komisije (2008-2009) ostvario opšti prosek prinosa od 4,89 t ha-1, masu 1000 semena od 113 g, sadržaj ulja u semenu 38,8% i sadržaj proteina u semenu koji se kretao od 15,2% do 16,6%. Na osnovu DUS testa urađe-nog u Slovačkoj sortnoj komisiji hibrid Mahe-lan (NS-H-6318) poseduje potrebnu različitost, ujednačenost, postojanost, a odlikuje se seme-nom crne boje sa sivim šarama izraženim u sre-dini i na krajevima (Tab. 7).
Novi NS-konzumni hibridi suncokreta (NS-H-6316, NS-H-6318 i NS-H-6320) ostvarili su veće prinose od standarda Sortne komisije Srbije i u odnosu na hibride drugih kompanija prijavlje-nih u Slovačkoj sortnoj komisiji. Pripadaju grupi srednje ranih hibrida pogodnih za ishranu, ljušte-nje i proizvodnju jezgra. Genetski potencijal za prinos im je preko 4,5 t ha-1. Sadržaj ulja u se-menu je manji od 40%, sadržaj proteina u jezgru od 23% do 27%, masa 1000 semena iznad 95 g, a udeo ljuske je nizak. To su novi genotipovi sa većim potencijalom za ekonomičniju i stabilniju proizvodnju koji po ostvarenom prinosu semena i proteina kao i po agronomskim osobinama za-služuju pažnju proizvođača konzumnih hibrida.
Tabe
la 7.
Deo
rezu
ltata
dob
ijen
DU
S m
etod
om is
pitiv
anje
m h
ibrid
a M
ahel
an (N
S-H
-631
8); p
rimen
jena
met
oda
DU
ST T
EST
(TP/
81/1
), Br
atisl
ava
(Sta
nica
za
ispiti
vanj
e Ve
liki M
eder
); Ta
ble 7
. Par
tial r
esul
ts of
DU
S re
sear
ch m
etho
d of
Mah
elan
(NS-
H-6
318)
hyb
rid; m
etho
d ap
plied
DU
ST T
EST
(TP/
81/1
), Br
atisl
ava (
Rese
arch
stat
ion
Velik
i Med
er)
Svoj
stvo
/ Ch
arac
teris
ticSt
epen
eks
pres
ijeE
xpre
sion
Oce
naN
ote
3Li
st /
Lea
fve
ličin
a /
size
sred
nje
velik
/ m
ediu
m la
rge
64
zele
na b
oja
/ gr
een
colo
rsr
ednj
e sv
etla
/ m
ediu
m li
ght
419
Jeziča
sti c
veto
vi /
Ray
fl or
ets
boja
/ co
lor
sred
nje
žuta
/ m
ediu
m y
ello
w3
20Tr
ubas
i cve
tovi
/ D
isk fl
ower
boja
/ co
lor
nara
ndža
sta
/ or
ange
228
Biljk
a /
Plan
tpr
irodn
a vi
sina
/ na
tura
l hei
ght
viso
k /
tall
733
Glav
a /
Hea
dve
ličin
a /
size
sred
nja
/ m
ediu
m5
35Se
me
/ Se
ed
velič
ina
/ siz
ekr
upno
do
veom
a kr
upno
/ la
rge
to v
ery
large
836
oblik
/ sh
ape
usko
jajo
lik /
nar
row
ovo
id2
37de
bljin
a /
thick
ness
relat
ive
to w
idth
sred
nja
/ m
ediu
m5
38os
novn
a bo
ja /
main
col
orcr
na /
blac
k7
39ša
re n
a kr
ajevi
ma
/ st
ripes
on
mar
gin
jako
izra
žene
/ st
rong
ly e
xpre
ssed
340
šare
na
sred
ini /
strip
es b
etw
een
mar
gins
jako
izra
žene
/ st
rong
ly e
xpre
ssed
341
boja
šare
/ c
olor
of
strip
essiv
a /
grey
2

Novi konzumni hibridi suncokreta u NS paleti 55
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
Zaključak
Dobijeni rezultati iz Sortne komisije Srbije i Slo-vačke, trogodišnjih i dvogodišnjih ogleda Instituta za ratarstvo i povrtarstvo, Novi Sad potvrđuju da su u novim NS-konzumnim hibridima suncokreta (NS-H-6316, NS-H-6318 i NS-H-6320) uspešno kombinovani geni odgovorni za visok genetski potencijal za prinos i dobra tehničko tehnološka svojstva semena. Uvođenjem u proizvodnju no-vih konzumnih NS hibrida suncokreta koji su po-godni za ishranu, ljuštenje i proizvodnju jezgra, ratarima se pruža veća mogućnost izbora najbo-ljih genotipova za različite namene i uslove pro-izvodnje, a time i kvalitetnije sirovine za potrebe uljarske i prehrambene industrije.
Literatura
Dušanić N, Miklič V, Joksimović J, Atlagić J (2004): Path co-effi cient analysis of some yield components of sunfl ower. Proc. 16th Int. Sunf. Conf., Fargo, North Dakota, USA, II: 531-537
Dozet B, Jovanović D (1997): Kombinacione sposobnosti i način nasleđivanja dimenzije semena u F1 generaciji konzu-mnog suncokreta (Helianthus annuus var. macrocarpus Dc. Ckll.) Sel. Semen. 3-4
Gonzalez-Perez S, Vereijken J M (2007): Sunfl ower proteins: overview of their physicochemical, structural and functional properties. J. Sci. Food Agric. 87: 2173-2191
Jovanović D, Stanojević D, Golubović M (1996): Variability of protein and husk content in seed of selffertilization sun-fl ower lines. Proc. 14th Int. Sunf. Conf., Beijing/Shenyang, China, 151-155
Jovanović D, Stanojević D (1996): Oplemenjivanje suncokreta na povećani sadržaj proteina u semenu. Zbornik radova, 37. Savetovanje: Proizvodnja i prerada uljarica, Budva, 223-241
Jovanović D (2001) Mogučnost korišćenja suncokreta i ople-menjivanje za posebne namene. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 35: 209-221
Hadživuković S (1991): Statistički metodi s primenom u po-ljoprivrednim i biološkim istraživanjima. Poljoprivredni fa-kultet, Institut za ekonomiku poljoprivrede i sociologiju sela, Novi Sad
Hladni N, Jocić S, Miklič V, Mijić A, Saftić Panković D (2008): Direct and indirect effects of morphophysiological traits on seed yield of sunfl ower (Helianthus annuus L.) Proc. 17th Int. Sunf. Conf., Cordoba, Spain, I: 393-397
Hladni N, Jocić S, Miklič V, Dušanić N, Saftić-Panković D, Ra-deka I, Lečić N (2009a): Rezultati ispitivanja novih eksperi-mentalnih hibrida suncokreta konzumnog tipa u toku 2007. i 2008. godine. Zbornik radova Instituta za ratarstvo i povr-tarstvo, Novi Sad 46: 385-392
Hladni N, Jocić S, Miklič V, Dušanić N, Saftić-Panković D, Ra-deka I, Lečić N (2009b): Ocena vrednosti novih konzumnih hibrida suncokreta Zbornik radova 50. Savetovanja industri-je ulja, Herceg Novi, 57-61
Hladni N, Jocić S, Miklič V, Radić V, Radeka I, Lečić N (2009d): Borneo - Novi NS hibrid suncokreta konzumnog tipa. Sel. Semen. 15: 63-73
Hladni N (2010): Geni i prinos suncokreta. Monografi ja. Za-dužbina Andrejević, Beograd, 116
Miklič V, Balalić I, Jocić S, Marinković R, Hladni N, Gvozdeno-vić S, Stojšin V (2009): Produktivnost NS hibrida suncokreta u multilokacijskim ogledima i preporuka sortimenta za setvu u 2009. godini. Zbornik radova Instituta za ratarstvo i povr-tarstvo, Novi Sad 46: 293-310
Miklič V, Balalić I, Jocić S, Marinković R, Cvejić S, Hladni N, Miladinović D (2010): Ocena stabilnosti prinosa semena i ulja NS hibrida suncokreta u mikro-ogledima i preporuka sortimenta za setvu u 2010. godini. Ratar. Povrt. / Field Veg. Crops Res. 47: 131-146
Škorić D, Joksimović J, Jocić S, Jovanović D, Marinković R, Hladni N, Gvozdenović S (2005): Ocena vrednosti pro-duktivnih svojstava NS-hibrida suncokreta. Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad41: 21-33
Škorić D, Jocić S, Joksimović J, Jovanović D, Hladni N, Marinković R, Atlagić J, Panković D, Vasić D, Miladi-nović F, Gvozdenović S, Terzić S, Sakač Z (2006): Do-stignuća u oplemenjivanju suncokreta. Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 42: 131-171
Vranceanu AV (2000): Floarea-sourelni hibrida. Editura Ceres, Bucharest

Hladni N i sar.56
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 49-56
New Confectionary Sunfl ower Hybrids in NS Palette
Nada Hladni · Vladimir Miklič · Siniša Jocić ·Zvonimir Sakač · Velimir Radić · Ilija Radeka
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Sunfl ower breeding is directed towards the increase of the genetic potential for yield, stability of the yield, health care, and the quality of nutrition along with the increase of the production economy. In accordance with the Serbian market demands there have been efforts in breeding highly productive hybrids for special uses of the confectionary type at the Institute of Field and Vegetable Crops in Novi Sad. In this program, special attention has been given to the increase of the protein content and quality while decreasing the seed oil content, to the incre-ase of 1000 seed weight and the decrease of the hull ratio. New NS confectionary hybrids NS-H-6316, NS-H-6318 and NS-H-6320 are adequate for nutrition, dehulling and core production. They belong to the group of mid-early hybrids while their genetic potential for seed yield is above 4.5 t ha-1 and seed oil content is less than 40%. Protein content in the core is 23-27%, mass of 1000 seeds is above 95 g, and the hull ratio is low. With the creation of the new palette of NS sunfl ower hybrids for various production purposes and conditions, we have gained higher quality material for the needs of the oil, food and baking industry. Key words: confectionary type, high-protein sunfl ower, seed quality, yield

Produktivnost NS hibrida suncokreta u mikro-ogledima 57
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66originalni naučni rad / original research article
Produktivnost NS hibrida suncokreta u mikro-ogledima u Srbiji u 2010. godini
Vladimir Miklič · Igor Balalić · Siniša Jocić · Radovan Marinković · Sandra Cvejić · Nada Hladni · Dragana Miladinović
primljeno / received: 16.11.2010. prihvaćeno / accepted: 29.11.2010.© 2011 IFVC
Izvod: Cilj istraživanja je bio da se ocene NS hibridi suncokreta na osnovu rezultata koje su ostva-rili za prinos semena, sadržaj ulja u semenu i prinos ulja u mreži mikro-ogleda u Vojvodini i cen-tralnoj Srbiji tokom 2010. godine, kao i da se da preporuka sortimenta za 2011. U mikro-ogledima je ispitivano 13 hibrida na 10 lokaliteta u Vojvodini i 3 u centralnoj Srbiji. Prinos semena, sadržaj ulja u semenu i prinos ulja su se značajno razlikovali u mreži mikro-ogleda u ispitivanim regionima. U Vojvodini, najveći prosečan prinos semena ostvarili su hibridi Duško (3,08 t ha-1), Sremac (2,98 t ha-1), Orfej (2,97 t ha-1), Velja (2,93 t ha-1) i NS-H-111 (2,92 t ha-1). U regionu centralne Srbije najviši prosečan prinos semena su postigli NS-H-111 (2,94 t ha-1) i Duško (2,87 t ha-1). Najveći prosečan sadržaj ulja u semenu ostvario je hibrid Baća, kako u regionu Vojvodine (48,54%) tako i u regionu centralne Srbije (48,80%). Najveći prosečan prinos ulja (preko 1,30 t ha-1) u regionu Vojvodine postigli su Duško, Bačvanin i NS-H-111, a u centralnoj Srbiji najveći prosečan prinos ulja (preko 1,30 t ha-1) ostvarili su hidridi NS-H-111, Oskar i Novosađanin. Na osnovu rezultata trogodišnjih ispitivanja u Vojvodini, četiri hibrida (Duško, Sremac, Velja i NS-H-111) su dali prinos semena iznad opšteg proseka. Najveći prosečan prinos semena za tri godine postigao je hibrid Duško (3,54 t ha-1). Ključne reči: lokalitet, mikro-ogledi, prinos semena, prinos ulja, sadržaj ulja u semenu, suncokret
V. Miklič · I. Balalić · S. Jocić · R. Marinković · S. Cvejić · N. Hladni · D. Miladinović Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Suncokret (Helianthus annuus L.) se u svetu gaji na preko 23 miliona hektara sa prosečnim prino-som od 1,3 t ha-1 (FAOSTAT 2009). U Srbiji je suncokret najznačajnija uljana kultura, koja već godinama postiže stabilne proizvodne rezultate. Velika potražnja proizvođača u Srbiji i na svet-skom tržištu za što kvalitetnijim i rodnijim hibri-dima zahteva brzu izmenu sortimenta sa boljim, produktivnijim i stabilnijim hibridima suncokre-ta. Visina i stabilnost prinosa hibrida su osobine od najvećeg interesa kako za oplemenjivače, tako i za proizvođače. Stvaranje rodnijih hibrida, kao i primena odgovarajućih agrotehničkih mera koje smanjuju uticaj limitirajućih faktora u proizvod-nji, doprinose povećanju prinosa suncokreta (Mi-klič i sar. 2010).
Uslovi za proizvodnju hibrida suncokreta va-riraju u zavisnosti od godine. U godinama kada
su povoljni uslovi za gajenje suncokreta postižu se visoki prinosi semena, međutim česte su go-dine kada jedan ili više stresnih perioda rezulti-ra smanjenjem prinosa (Škorić 1988). Protekla 2010. bila je veoma specifi čna godina za proi-zvodnju suncokreta zbog većih količina padavi-na za naše agroekološke uslove i čestih olujnih naleta što se odrazilo na prinose semena. Velike količine padavina u prolećnom periodu (u perio-du januar-jun palo je 534 mm, Republički hidro-meteorološki zavod – lokalitet Rimski Šančevi) rezultirale su stvaranje vodoleža i lošije nicanje suncokreta, a velika temperaturna kolebanja u tom periodu prouzrokovala su stres kod bilja-ka, što se kasnije odrazilo na prinos semena. U periodu cvetanja i nalivanja semena olujni naleti izazvali su poleganje, pa čak i lomljenje stabala, što je dodatno negativno uticalo na formiranje prinosa. Vrednosti hibrida ispituju se i godina-ma lošijim za proizvodnju suncokreta, kako bi se dobili potpuniji podaci o njihovoj adaptabil-nosti i stabilnosti.
Cilj rada je ocena novosadskih hibrida sunco-kreta na osnovu rezultata koje su postigli za pri-nos semena, sadržaj ulja u semenu i prinos ulja
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Miklič V i sar.58
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
u mreži mikro ogleda, postavljenih na različitim lokalitetima u Vojvodini i centralnoj Srbiji tokom 2010. godine, kao i davanje preporuke sortimenta za gajenje u 2011. godini. Bolja ocena vrednosti pojedinih hibrida izvršena je poređenjem prino-sa semena 9 hibrida suncokreta koji su gajeni u mikro-ogledima u Vojvodini tokom trogodišnjeg perioda (2008-2010).
Materijal i metod
U istraživanja je uključeno 13 hibrida sunco-kreta stvorenih u Institutu za ratarstvo i povrtar-stvo iz Novog Sada, od kojih je 10 hibrida već rašireno u proizvodnji (Duško, Novosađanin, Sremac, NS-H-111, Vladimir, Baća, Branko, Ve-lja, Bačvanin i Šumadinac) i tri nova perspektiv-na hibrida (Oskar, Orfej i Orion). Tokom 2010. u Vojvodini su postavljeni mikro-ogledi na 10 lokaliteta (Vrbas, Senta, Šupljak, Sombor, Alek-sa Šantić, Pančevo, Vršac, Kuzmin, Krčedin i Neštin). U centralnoj Srbiji su mikro-ogledi po-
stavljeni na tri lokaliteta (Kula-Vitovnica, Nego-tin i Zaječar).
Veličina osnovne parcelice iznosila je 28 m2
. Dva srednja reda (isključujući rubne biljke) korišćena su za berbu. Veličina neto parcele iznosila je 13,3 m2 (0,7 x 0,25 x 76). Ogledi su postavljeni po slučajnom blok sistemu u 4 po-navljanja, uz primenu optimalne agrotehnike. Tokom vegetacije vršena su fenološka opaža-nja i merenja. U radu su analizirana tri glavna parametra produktivnosti: prinos semena (t ha-1), sadržaj ulja u semenu (%) i prinos ulja (t ha-1). Dobijeni prinos semena suncokreta je preračunat u t ha-1 sa 11% vlage. Sadržaj ulja u čistom semenu određen je metodom NMR (nu-klearno-magnetna rezonanca) prema Granlund & Zimmerman (1975). Prinos ulja izračunat je kao proizvod prinosa semena i sadržaja ulja. Rezultati su statistički obrađeni kao specifi čan dvofaktorijalni dizajn Lokalitet x Genotipovi gde se u izačunavanju LSD vrednosti koriste sume kvadrata pogreški iz (jednodimenzional-
Tabela 1. Prinos semena (t ha-1) hibrida suncokreta u mreži mikro-ogleda u Vojvodini 2010. Table 1. Seed yield (t ha-1) of sunfl ower hybrids obtained in a network of small-plot trials in Vojvodina in 2010
HibridHybrid
Lokalitet / Location
Vrba
s
Sent
a
Šupl
jak
Som
bor
Ale
ksa
Šant
ić
Panč
evo
Vrša
c
Krč
edin
Neš
tin
Kuz
min Prosek
MeanRangRank
1 DUŠKO 2,78 3,17 3,20 3,32 2,86 3,32 2,80 2,97 3,85 2,58 3,08 1
2 NOVOSAĐANIN 2,36 2,43 2,94 3,06 2,64 2,75 1,98 2,47 2,99 3,17 2,68 10
3 SREMAC 2,61 3,18 3,16 3,00 2,98 3,04 2,49 2,75 3,42 3,15 2,98 2
4 NS-H-111 2,80 3,20 2,21 3,22 2,91 2,86 2,47 2,69 3,50 3,39 2,92 5
5 VLADIMIR 2,24 2,13 2,83 3,02 2,98 2,50 1,92 2,59 3,34 2,97 2,65 11
6 BAĆA 2,17 2,37 2,55 3,01 3,04 3,09 2,71 2,81 2,91 2,54 2,72 9
7 BRANKO 1,99 2,49 2,62 2,87 2,67 2,55 2,14 2,41 2,61 2,63 2,50 13
8 VELJA 2,82 3,14 1,68 3,01 2,80 3,20 2,55 3,26 3,23 3,63 2,93 4
9 BAČVANIN 2,13 2,67 2,77 2,96 3,05 2,59 2,78 2,98 2,97 3,30 2,82 6
10 ŠUMADINAC 2,41 2,91 3,07 2,70 2,51 3,07 2,28 2,61 2,90 2,89 2,74 7
11 OSKAR 2,10 2,99 2,60 2,60 2,94 2,47 1,82 2,50 3,00 3,00 2,60 12
12 ORFEJ 2,26 2,80 3,47 3,14 2,81 3,03 2,29 3,11 3,25 3,57 2,97 3
13 ORION 2,40 2,51 2,48 2,59 2,88 2,68 2,27 2,83 3,08 3,62 2,73 8Prosek lokalitetaMean over location
2,39 2,77 2,74 2,96 2,85 2,86 2,34 2,77 3,16 3,11 2,79
LSD Hibridi/ Hybrids Lokaliteti/ Locations Hibrid x lokalitet /Hybrid x location
0,05 0,18 0,12 0,55
0,01 0,23 0,15 0,72

Produktivnost NS hibrida suncokreta u mikro-ogledima 59
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
ne) ANOVE za svaki lokalitet ponaosob u pro-gramu Statistica 8.
Rezultati i diskusija
Prinos semena suncokreta
Visina, kao i stabilnost prinosa semena hibri-da suncokreta su od najvećeg interesa, kako za oplemenjivače tako i za proizvođače. Prinos se-mena suncokreta je proizvod tri osnovne kom-ponente: broja biljaka po jedinici površine, broja semena po biljci i mase semena (Merrien 1992). U prinosu semena postoje značajne razlike iz-među pojedinih hibrida, lokaliteta i godina ispi-tivanja (Crnobarac & Dušanić 1996, Schoeman 2003).
Na osnovu ostvarenih rezultata može se kon-statovati da su prinosi semena većine ispitanih hibrida bili visoki s obzirom na vremenske prilike. Prosečan prinos semena suncokreta u mikro-ogledima postavljenim u Vojvodini, uzi-majući u obzir sve hibride i lokalitete, iznosio
je 2,79 t ha-1 (Tab. 1). Prinos semena u regionu centralne Srbije na tri lokaliteta iznosio je 2,73 t ha-1 (Tab. 4).
Najveći prosečan prinos semena u Vojvodini postigli su hibridi Duško (3,08 t ha-1), Sremac (2,98 t ha-1), Orfej (2,97 t ha-1), Velja (2,93 t ha-1) i NS-H-111 (2,92 t ha-1), s tim da su prva tri navedena imali statistički značajno viši prinos od opšteg proseka. Hibrid Duško postigao je najviše prinose na 4 lokaliteta (Sombor, Panče-vo, Vršac i Neštin) (Tab. 1). U regionu centralne Srbije najveći prosečan prinos semena postigao je hibrid NS-H-111 (2,94 t ha-1), a statistički zna-čajno više prinose od opšteg proseka imali su hibridi Duško, Novosađanin i Orfej (Tab. 4).
U Vojvodini je tokom 2010. najveći prinos se-mena suncokreta postignut na lokalitetu Neštin (3,16 t ha-1). Takođe, na lokalitetu Kuzmin prinos semena je bio značajno veći u odnosu na ostale lokalitete i iznosio je 3,11 t ha-1. Na pojedinim lo-kalitetima neki hibridi su ostvarili izuzetno visoke prinose. Tako su npr. većina hibrida na lokalitetu Neštin dali prinos preko 3 t ha-1, što ukazuje na
Tabela 2. Sadržaj ulja u semenu (%) hibrida suncokreta u mreži mikroogleda u Vojvodini 2010. Table 2. Seed oil content (%) of sunfl ower hybrids obtained in a network of small-plot trials in Vojvodina in 2010
HibridHybrid
Lokalitet / Location
Vrba
s
Sent
a
Šupl
jak
Som
bor
Ale
ksa
Šant
ić
Panč
evo
Vrša
c
Krč
edin
Neš
tin
Kuz
min Prosek
MeanRangRank
1 DUŠKO 42,62 39,17 46,00 42,77 43,86 42,76 42,76 46,05 42,83 39,62 42,44 9
2 NOVOSAĐANIN 43,94 41,00 47,37 43,75 46,10 42,02 44,48 46,76 44,28 43,26 42,42 10
3 SREMAC 40,40 36,67 42,31 40,43 42,53 41,69 41,07 45,24 39,87 37,94 40,74 13
4 NS-H-111 43,31 42,77 46,10 45,82 45,75 44,19 42,95 47,83 44,49 41,86 44,79 4
5 VLADIMIR 42,78 39,61 47,57 44,08 43,35 41,40 40,32 46,68 43,47 41,76 43,09 8
6 BAĆA 42,91 43,11 50,75 49,02 48,26 44,90 45,13 51,88 46,71 44,86 48,54 1
7 BRANKO 44,67 37,83 45,31 43,68 46,16 44,11 42,14 46,68 44,66 42,07 43,16 7
8 VELJA 40,85 38,36 42,81 42,87 42,40 43,33 40,68 46,81 41,26 41,56 41,82 11
9 BAČVANIN 43,91 42,29 48,93 47,07 47,37 43,84 43,79 49,82 46,13 47,13 46,64 2
10 ŠUMADINAC 43,67 41,56 50,58 46,01 46,12 42,31 42,37 47,85 44,81 43,81 43,54 6
11 OSKAR 45,49 43,44 49,66 45,94 47,09 44,41 43,96 48,92 46,52 45,13 45,11 3
12 ORFEJ 42,35 37,82 46,37 44,11 43,99 41,76 41,81 45,48 42,65 40,86 41,64 12
13 ORION 44,75 43,25 48,48 47,30 45,19 44,36 43,80 48,66 45,90 44,50 44,10 5Prosek lokalitetaMean over location
43,20 40,53 47,09 44,83 45,24 43,16 42,71 47,59 44,12 42,64 43,69
LSD Hibridi/ Hybrids Lokaliteti/ Locations Hibrid x lokalitet /Hybrid x location
0,05 0,63 0,43 1,96
0,01 0,82 0,67 2,58

Miklič V i sar.60
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
visoki genetski potencijal NS hibrida suncokre-ta. Duško je apsolutni rekorder u mikro-ogledi-ma postavljenim tokom 2010. pošto je postigao najviši prinos semena u odnosu druge hibride, a na najviše lokaliteta je bio najrodniji. Po visokim prinosima semena izdvajaju se i hibridi Sremac, NS-H-111, Velja i Orfej (Tab. 1).
Prinos semena u mikro-ogledima postavljenim na lokalitetima u centralnoj Srbiji utvrđen je za 13 hibrida i 3 lokaliteta (Tab. 4). Najveći prinosi semena postignuti su na lokalitetima Kula-Vitov-nica (3,08 t ha-1) i Zaječar (2,92 t ha-1), dok su na lokalitetu Negotin postignuti značajno manji prinosi semena. Najviši prosečan prinos semena na nivou cele centralne Srbije ostvario je hibrid NS-H-111 sa 2,94 t ha-1. Isti hibrid zabeležio je i rekordan prinos od 3,52 t ha-1 na lokalitetu Kula-Vitovnica. Visoke prinose imali su i hidridi Duš-ko, Novosađanin i Orfej (Tab. 4).
Sadržaj ulja u semenu suncokreta
Značajan uticaj na sadržaj ulja u semenu ima-ju genotip, zemljišno-klimatski uslovi i nivo pri-
menjene agrotehnike (Balalić i sar. 2008). Od zemljišno-klimatskih uslova važnu ulogu imaju temperatura vazduha i količina raspoložive vlage u zemljištu u fazi nalivanja semena (Škorić 1988, Marinković i sar. 2003). Poznavanjem edafskih i klimatskih karakteristika rejona gajenja sunco-kreta, uz primenu odgovarajućih agrotehničkih mera, može se uticati na povećanje sadržaja ulja u semenu, a time i na ukupnu proizvodnju ulja (Marinković i sar. 2003).
Prosečan sadržaj ulja u semenu za 13 hibrida suncokreta u mikro-ogledima postavljenim na 10 lokaliteta u regionu Vojvodine tokom 2010. iznosio je 43,69% (Tab. 2), što je značajno niže u odnosu na rezultate mikro-ogleda 2009. u ko-joj je prosečan sadržaj ulja u semenu 19 hibrida na 11 lokaliteta u Vojvodini iznosio 46,56% (Mi-klič i sar. 2010). Najveći prosečan sadržaj ulja na lokalitetima u Vojvodini ostvario je hibrid Baća (48,54%). Statistički značajno viši sadržaj ulja u odnosu na opšti prosek ispitivanih hibrida u re-gionu Vojvodine pokazali su i hibridi Bačvanin, Oskar i NS-H-111. Hibrid Baća je bio najuljaniji na 7 lokaliteta (Šupljak, Sombor, Aleksa Šantić,
Tabela 3. Prinos ulja (t ha-1) hibrida suncokreta u mreži mikroogleda u Vojvodini 2010. Table 3. Seed oil yield (t ha-1) of sunfl ower hybrids obtained in a network of small-plot trials in Vojvodina in 2010
HibridHybrid
Lokalitet / Location
Vrba
s
Sent
a
Šupl
jak
Som
bor
Ale
ksa
Šant
ić
Panč
evo
Vrša
c
Krč
edin
Neš
tin
Kuz
min Prosek
MeanRangRank
1 DUŠKO 1,19 1,24 1,45 1,42 1,26 1,42 1,20 1,37 1,65 1,02 1,32 1
2 NOVOSAĐANIN 1,04 1,00 1,40 1,34 1,22 1,15 0,88 1,16 1,33 1,38 1,19 11
3 SREMAC 1,06 1,17 1,33 1,21 1,26 1,27 1,02 1,25 1,36 1,20 1,21 9
4 NS-H-111 1,21 1,37 1,01 1,47 1,33 1,26 1,06 1,28 1,56 1,42 1,30 3
5 VLADIMIR 0,96 0,85 1,35 1,33 1,29 1,04 0,78 1,21 1,45 1,24 1,15 12
6 BAĆA 0,94 1,02 1,29 1,47 1,47 1,38 1,22 1,46 1,36 1,14 1,28 4
7 BRANKO 0,89 0,95 1,19 1,25 1,23 1,12 0,90 1,13 1,17 1,11 1,09 13
8 VELJA 1,15 1,21 0,72 1,29 1,19 1,38 1,03 1,52 1,33 1,51 1,23 7
9 BAČVANIN 0,94 1,13 1,36 1,39 1,44 1,13 1,22 1,48 1,37 1,55 1,30 2
10 ŠUMADINAC 1,05 1,21 1,55 1,24 1,16 1,30 0,97 1,25 1,30 1,27 1,23 8
11 OSKAR 0,96 1,30 1,29 1,20 1,38 1,10 0,80 1,22 1,40 1,35 1,20 10
12 ORFEJ 0,96 1,06 1,61 1,39 1,23 1,26 0,96 1,41 1,38 1,46 1,27 5
13 ORION 1,07 1,09 1,20 1,23 1,30 1,18 0,99 1,37 1,42 1,62 1,25 6Prosek lokalitetaMean over location
1,03 1,12 1,29 1,33 1,29 1,23 1,00 1,32 1,39 1,33 1,23
LSD Hibridi/ Hybrids Lokaliteti/ Locations Hibrid x lokalitet /Hybrid x location
0,05 0,08 0,06 0,25
0,01 0,10 0,07 0,33
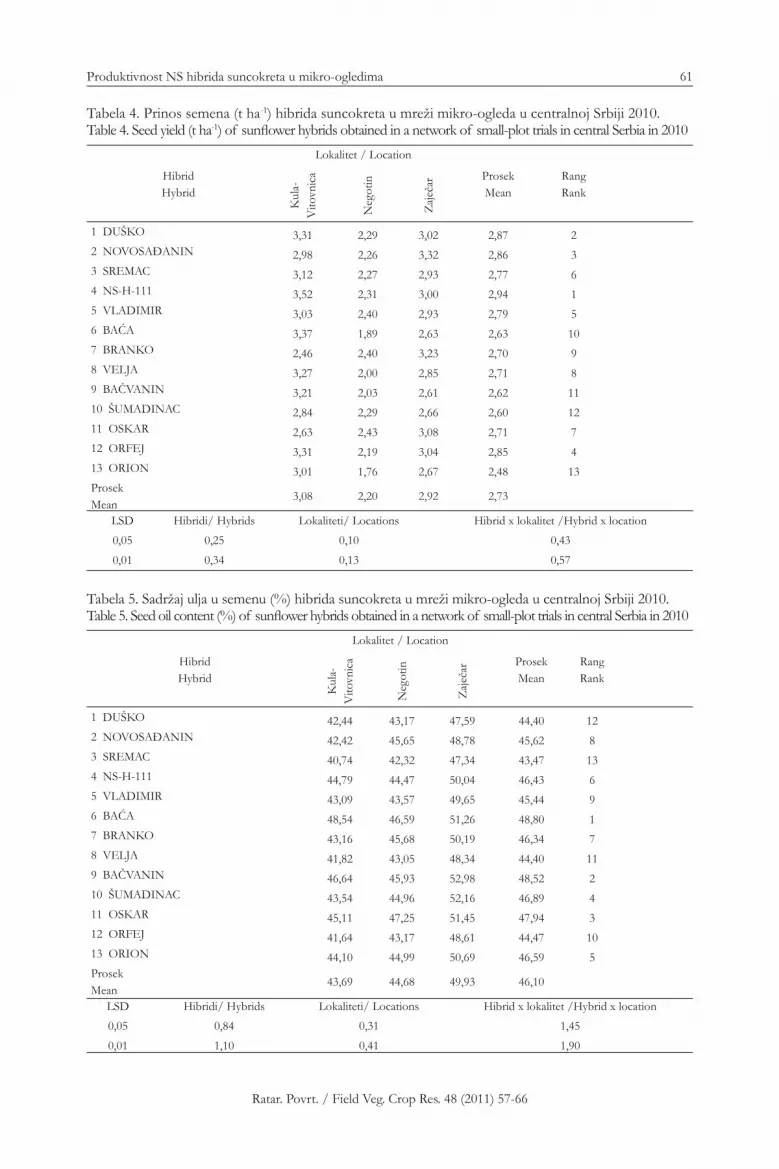
Produktivnost NS hibrida suncokreta u mikro-ogledima 61
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Tabela 5. Sadržaj ulja u semenu (%) hibrida suncokreta u mreži mikro-ogleda u centralnoj Srbiji 2010. Table 5. Seed oil content (%) of sunfl ower hybrids obtained in a network of small-plot trials in central Serbia in 2010
Hibrid Hybrid
Lokalitet / Location
ProsekMean
RangRank
Kul
a-V
itovn
ica
Neg
otin
Zaječa
r
1 DUŠKO 42,44 43,17 47,59 44,40 122 NOVOSAĐANIN 42,42 45,65 48,78 45,62 83 SREMAC 40,74 42,32 47,34 43,47 134 NS-H-111 44,79 44,47 50,04 46,43 65 VLADIMIR 43,09 43,57 49,65 45,44 96 BAĆA 48,54 46,59 51,26 48,80 17 BRANKO 43,16 45,68 50,19 46,34 78 VELJA 41,82 43,05 48,34 44,40 119 BAČVANIN 46,64 45,93 52,98 48,52 210 ŠUMADINAC 43,54 44,96 52,16 46,89 411 OSKAR 45,11 47,25 51,45 47,94 312 ORFEJ 41,64 43,17 48,61 44,47 1013 ORION 44,10 44,99 50,69 46,59 5ProsekMean
43,69 44,68 49,93 46,10
LSD Hibridi/ Hybrids Lokaliteti/ Locations Hibrid x lokalitet /Hybrid x location
0,05 0,84 0,31 1,45
0,01 1,10 0,41 1,90
Tabela 4. Prinos semena (t ha-1) hibrida suncokreta u mreži mikro-ogleda u centralnoj Srbiji 2010. Table 4. Seed yield (t ha-1) of sunfl ower hybrids obtained in a network of small-plot trials in central Serbia in 2010
Hibrid Hybrid
Lokalitet / Location
ProsekMean
RangRank
Kul
a-V
itovn
ica
Neg
otin
Zaječa
r
1 DUŠKO 3,31 2,29 3,02 2,87 22 NOVOSAĐANIN 2,98 2,26 3,32 2,86 33 SREMAC 3,12 2,27 2,93 2,77 64 NS-H-111 3,52 2,31 3,00 2,94 15 VLADIMIR 3,03 2,40 2,93 2,79 56 BAĆA 3,37 1,89 2,63 2,63 107 BRANKO 2,46 2,40 3,23 2,70 98 VELJA 3,27 2,00 2,85 2,71 89 BAČVANIN 3,21 2,03 2,61 2,62 1110 ŠUMADINAC 2,84 2,29 2,66 2,60 1211 OSKAR 2,63 2,43 3,08 2,71 712 ORFEJ 3,31 2,19 3,04 2,85 413 ORION 3,01 1,76 2,67 2,48 13ProsekMean
3,08 2,20 2,92 2,73
LSD Hibridi/ Hybrids Lokaliteti/ Locations Hibrid x lokalitet /Hybrid x location
0,05 0,25 0,10 0,43
0,01 0,34 0,13 0,57

Miklič V i sar.62
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Pančevo, Vršac, Krčedin i Neštin), a rekordan sa-držaj ulja u semenu od 51,88% imao je na lokali-tetu Krčedin (Tab. 2). U regionu centralne Srbije prosečan sadržaj ulja u semenu za 13 hibrida i 3 lokaliteta iznosio je 46,10% a najviši je imao hibrid Baća (48,80%), kao i u Vojvodini. Hibrid Baća je postigao najveću vrednost sadržaja ulja u semenu u regionu Zaječar (51,26%) (Tab. 5). Na istom lokalitetu visok sadržaj ulja imao je i hibrid Bačvanin (52,98%). U mikro-ogledima 2008. i 2009. hibrid Baća je takođe bio među najuljanijima (Miklič i sar. 2009, Miklič i sar. 2010). Prema podacima koje iznose Kondić & Mijanović (2008) na osnovu trogodišnjih is-pitivanja u regiji Banja Luka, sadržaj ulja kod hibrida suncokreta stvorenih u Institutu za ra-tarstvo i povrtarstvo iz Novog Sada se kretao u intervalu od 33,0% kod hibrida Labud do 49,7% kod hibrida Baća.
Kao veoma povoljni lokaliteti za sintezu ulja u regionu Vojvodine u 2010. pokazali su se Krče-din, Šuljak i Aleksa Šantić, sa prosekom sadržaja ulja u semenu preko 45%. Na lokalitetu Som-bor je takođe bio statistički značajno viši sadržaj ulja u odnosu na opšti prosek ogleda. Zapaža se da je hibrid Baća na 7 lokaliteta imao najveću prosečnu vrednost sadržaja ulja u semenu (Tab.
2). Između prosečnih vrednosti za sadržaj ulja u regionu centralna Srbija, ustanovljene su vi-soko značajne razlike. Pri tome je lokalitet Za-ječar imao značajno veći sadržaj ulja u odnosu na lokalitete Kula-Vitovnica i Negotin. Sadržaj ulja u semenu preko 50% konstatovan je samo na lokalitetu Zaječar i to kod sedam ispitivanih hibrida.
Prinos ulja suncokreta
Prinos ulja je glavni pokazatelj produktivnosti svakog hibrida (Škorić i sar. 2005) i zavisi od prino-sa semena i sadržaja ulja u semenu. Do smanjenja sadržaja ulja dovode visoke temperature (>25°C) u periodu cvetanja, što dovodi do smanjenja pri-nosa ulja, kako navode Miklič i sar. (2009).
Prosečan prinos ulja u Vojvodini u 2010. iznosio je 1,23 t ha-1 (Tab. 3) i nije se značajno razlikovao u odnosu na lokalitete regiona centralne Srbije, gde je prosečan prinos ulja bio 1,26 t ha-1 (Tab. 6).
Na lokalitetima u Vojvodini najbolji prosečan prinos ulja među 13 ispitivanih hibrida imao je hibrid Duško (1,32 t ha-1). Statistički značajno viši prinos od opšteg proseka imao je i hibrid Ba-čvanin, a visoki prinosi ulja zabeleženi su i kod hibrida NS-H-111, Baća i Orfej. Hibridi Duško,
Tabela 6. Prinos ulja (t ha-1) hibrida suncokreta u mreži mikro-ogleda u centralnoj Srbiji 2010. Table 6. Seed oil yield (t ha-1) of sunfl ower hybrids obtained in a network of small-plot trials in central Serbia in 2010
Hibrid Hybrid
Lokalitet / Location
ProsekMean
RangRank
Kul
a-V
itovn
ica
Neg
otin
Zaječa
r
1 DUŠKO 1,41 0,99 1,44 1,28 52 NOVOSAĐANIN 1,26 1,03 1,62 1,31 33 SREMAC 1,27 0,96 1,39 1,21 114 NS-H-111 1,58 1,03 1,50 1,37 15 VLADIMIR 1,31 1,05 1,45 1,27 76 BAĆA 1,63 0,88 1,35 1,29 47 BRANKO 1,06 1,10 1,62 1,26 98 VELJA 1,37 0,86 1,38 1,20 129 BAČVANIN 1,50 0,93 1,38 1,27 610 ŠUMADINAC 1,24 1,03 1,39 1,22 1011 OSKAR 1,19 1,15 1,59 1,31 212 ORFEJ 1,38 0,95 1,48 1,27 813 ORION 1,33 0,79 1,35 1,16 13ProsekMean
1,35 0,98 1,46 1,26
LSD Hibridi/ Hybrids Lokaliteti/ LocationsHibrid x lokalitet /Hybrid x
location0,05 0,12 0,04 0,20
0,01 0,16 0,06 0,26

Produktivnost NS hibrida suncokreta u mikro-ogledima 63
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Tabela 7. Prinos semena (t ha-1) hibrida suncokreta u mreži mikro-ogleda u Vojvodini u periodu 2008-2010. Table 7. Seed yield (t ha-1) of sunfl ower hybrids obtained in a network of small-plot trials in Vojvodina in the period 2008-2010
HibridHybrid
2008 2009 2010Prosek 2008-2010Mean 2008-2010
ProsekMean
RangRank
ProsekMean
RangRank
ProsekMean
RangRank
ProsekMean
RangRank
BAČVANIN 3,40 7 3,38 9 2,82 5 3,20 8
NS-H-111 3,49 6 3,57 4 2,92 4 3,33 4
VELJA 3,67 2 3,46 8 2,93 3 3,35 3
BAĆA 3,60 4 3,52 6 2,72 6 3,28 5
SREMAC 3,62 3 3,62 3 2,98 2 3,41 2
DUŠKO 3,79 1 3,75 1 3,08 1 3,54 1
BRANKO 3,35 9 3,49 7 2,50 9 3,11 9
NOVOSAĐANIN 3,38 8 3,74 2 2,68 7 3,26 6
VLADIMIR 3,59 5 3,53 5 2,65 8 3,26 7ProsekMean
3,54 3,56 2,81 3,30
NS-H-111 i Baća ostvarili su najviše prinose ulja na po tri lokaliteta, dok je Orion bio najuljaniji na dva lokaliteta. Rekordan prinos ulja zabeležen je na loka-litetu Neštin od 1,65 t ha-1 (Duško) (Tab. 3). Hibridi NS-H-111, Novosađanin i Oskar dali su prinos ulja preko 1,3 t ha-1 u regionu centralna Srbija. Rekordan prinos ulja dali su hibridi Novosađanin i Branko, 1,62 t ha-1 na lokalitetu Zaječar (Tab. 6).
Analizom prinosa ulja po lokalitetima u Voj-vodini u 2010. po značajno višim vrednostima od opšteg proseka izdvajaju se Neštin, Kuz-min, Sombor i Krčedin gde je prosečan prinos ulja preko 1,30 t ha-1. Ostali lokaliteti su imali prinos ulja iznad tone. Lokalitet Neštin odliku-je se najvećim prinosom ulja u mikro-ogledima koji su izvedeni u Vojvodini u 2010. Na tom lokalitetu prinos ulja preko 1,5 t ha-1 dali su hibridi Duško i NS-H-111. Na lokalitetu Kuz-min prinos ulja preko 1,5 t ha-1 dali su hibridi Bačvanin i Orion (Tab. 3). U regionu centralna Srbija, na lokalitetu Zaječar zabeležen je naj-veći prinos ulja (1,46 t ha-1) dok je nešto manji prinos ulja postignut na lokalitetu Kula-Vitov-nica (1,35 t ha-1) (Tab. 6). Vrednost preko 1,5 t ha-1 ostvarena je kod četiri hibrida na lokalitetu Zaječar i kod tri hibrida na lokalitetu Kula-Vi-tovnica (Tab. 6).
Rezultati višegodišnjih ispitivanja NShibrida suncokreta
U cilju što bolje ocene vrednosti pojedinih hibrida, analizirani su rezultati trogodišnjih
ispitivanja prinosa semena u mikro-ogledima suncokreta, koji su izvedeni u regionu Vojvodi-ne (Tab. 7). Opšti prosek prinosa za trogodišnji period (2008-2010) iznosio je 3,30 t ha-1. Pri-nos semena iznad opšteg proseka dalo je četiri hibrida (Duško, Sremac, Velja i NS-H-111), pri čemu je hibrid Duško imao najveći prinos (3,54 t ha-1). Ukoliko se posmatraju godine ispitiva-nja, uočava se da je prosečan prinos semena u 2008. i 2009. bio podjednak, sa većim pro-sečnim vrednostima u odnosu na 2010. Niže vrednosti prinosa u 2010. mogu se objasniti lo-šijim vremenskim prilikama u odnosu na dru-ge dve ispitivane godine, koje su bile povoljne za razvoj suncokreta. U proleće 2010. bilo je obilnih padavina, što je dovelo do stvaranja vodoleža i lošijeg nicanja, ali i ukorenjavanje u površinskom delu zemljišta, što je omogućilo intenziviranje poleganja useva. U maju i junu bile su česte promene temperature vazduha što je uticalo stresno na biljke u fazi butonizacije. Olujni naleti u avgustu napravili su velike štete u usevu suncokreta izazivajući poleganje i lo-mljenje biljaka.
S obzirom na činjenicu da nepovoljni uslovi spoljašnje sredine predstavljaju ograničavaju-ći faktor u proizvodnji suncokreta (Vrancea-nu 2000, Škorić i sar. 2006, Miklič i sar. 2007, Miklič i sar. 2008), neophodno je u ispitivanja, osim lokaliteta uključiti kao faktor i godine u cilju što boljeg sagledavanja uticaja agroekološ-kih uslova na realizaciju genetskog potencijala hibrida.

Miklič V i sar.64
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Izbor sortimenta NS hibrida suncokreta za setvu u 2011. godini
Prema rezultatima koji su postignuti u širokoj proizvodnji, kao i u mikro-ogledima u toku ove i prethodnih godina, za setvu u 2011. godini pred-lažu se sledeći hibridi:
Dominantni hibridi na parcelama gde je uo-A. čeno prisustvo volovoda (Orobanche cumana) treba da budu: Baća, Bačvanin, Šumadinac, Perun, Branko i NovosađaninTakođe, na ovim parcelama se uspešno mogu • gajiti i IMI-hibridi uz obaveznu primenu her-bicida Pulsar-40 (a. m. imazamox), a to su hi-bridi Rimi i Vitalko, kao i najnoviji hibrid Rimi PR koji je genetski otporan i na plamenjačuZa setvu na parcelama gde nije prisutan vo-B. lovod najbolji izbor su hibridi: Sremac, Duš-ko, NS-H-111, Velja, Kazanova, NS-H-45 i Pobednik. Na ovim parcelama preporučuje se i novi hibrid Duško koji je postigao odlične rezultate u mreži mikro-ogleda u periodu od 2008-2010. godine. Ovaj hibrid je takođe ge-netski otporan i na plamenjačuOd hibrida specijalne namene preporučuje-C. mo:Za proizvodnju hladno ceđenog ulja i biodize-• la hibride Oliva i OlivkoZa proizvodnju proizvoda od jezgre sunco-• kreta hibride Vranac i CepkoZa setvu krajem maja i u prvoj polovini juna D. meseca treba sejati rani hibrid DukatU grupi dekorativnih suncokreta preporučuje E. se Neoplanta, koja se odlikuje bordo bojom jezičastih cvetova
Zaključak
Na osnovu rezultata postignutih u mikro-ogle-dima postavljenih u regionu Vojvodine i central-ne Srbije, mogu se doneti sledeći zaključci: • Tokom 2010. u mreži mikro-ogleda prosečan
prinos semena u regionu Vojvodine (10 lo-kaliteta) posmatrajući sve hibride i lokalitete iznosio je 2,79 t ha-1, dok je u centralnoj Srbiji, gde su mikroogledi postavljeni na tri lokalite-ta, prinos semena bio 2,73 t ha-1. U Vojvodini, najveći prosečan prinos semena ostvarili su hi-bridi Duško (3,08 t ha-1), Sremac (2,98 t ha-1), Orfej (2,97 t ha-1), Velja (2,93 t ha-1), i NS-H-111 (2,92 t ha-1). U regionu centralne Srbije najveći prosečan prinos semena postigao je hibrid NS-H-111 (2,94 t ha-1).
• U 2010. prosečan sadržaj ulja u semenu za 13 ispitivanih hibrida na 10 lokaliteta u regionu Vojvodine iznosio je 43,69%. Visoko značaj-no najveći sadržaj ulja u odnosu na ostale is-pitivane hibride u regionu Vojvodine pokazali su hibridi Baća, Bačvanin i Oskar. U regionu centralne Srbije najveći prosečan sadržaj ulja iznad 48% imali su hibridi Baća i Bačvanin.
• Za sve hibride i lokalitete u Vojvodini opšti prosek prinosa ulja tokom 2010. iznosio je 1,23 t ha-1. Najveći prinosi ulja postignuti su kod hibrida Duško, Bačvanin, NS-H-111 i Baća na lokalitetima regiona Vojvodine. Pri-nos ulja u proseku za 13 hibrida i tri lokaliteta u centralnoj Srbiji iznosio je 1,26 t ha-1. Lokali-tet Zaječar u regionu centralna Srbija proizveo je najuljanije hibride među kojima se ističu NS-H-111, Oskar i Novosađanin sa proseč-nim prinosom ulja preko 1,3 t ha-1. Lokalitet Zaječar proizveo je najuljanije hibride.
• Opšti prosek prinosa za trogodišnji period (2008-2010) u regionu Vojvodine iznosio je 3,30 t ha-1. Prinos iznad opšteg proseka dala su četiri hibrida (Duško, Sremac, Velja i NS-H-111), pri čemu je hibrid Duško imao najveći prinos (3,54 t ha-1). Uočava se da je prosečan prinos semena u 2008. i 2009. bio podjednak, sa većim prosečnim vrednostima u odnosu na 2010. Niže vrednosti prinosa u 2010. mogu se objasniti lošijim vremenskim prilikama u odnosu na druge dve ispitivane godine, koje su bile povoljne za razvoj suncokreta. Ovi re-zultati ukazuju na plastičnost i stabilnost na-vedenih hibrida, jer su agroekološki uslovi u ispitivanim godinama bili različiti.
• Rezultati ovog rada su od značaja pri odlučiva-nju vezanom za izbor hibrida suncokreta viso-kih i stabilnih prinosa.

Produktivnost NS hibrida suncokreta u mikro-ogledima 65
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Literatura
Balalić I, Miklič V, Jocić S, Jovanović D, Hladni N, Marinković R, Gvozdenović S (2008): Rezultati mikroogleda NS hibrida suncokreta i preporuka sortimenta za 2008. godinu. Zbor-nik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 45: 111-124
Crnobarac J, Dušanić N (1996): Uticaj gustine useva na prinos i kvalitet suncokreta. Zbornik 30. savetovanja o unapređenju uljarstva Jugoslavije, Budva, 490-49.
Granlund M, Zimmerman D C (1975): Effect of drying con-ditions on oil contents of sunfl ower (Helianthus annuus L.) seed determined by wide-line Nuclear Magnetic Resonance (NMR). North Dakota Acad Sci Proc. 27: 128-132
FAOSTAT (2009): [Elektronski izvor] Dostupno na www.fao-stat.org (citirano: 10.11.2010.)
Kondić J, Mijanović K (2008): Selection of sunfl ower hybrids for Banja Luka area in Bosnia and Herzegovina. Proc. 17th International Sunfl ower Conference, Cordoba, Spain 495-497
Marinković R, Dozet B, Vasić D (2003): Oplemenjivanje sunco-kreta. Monografi ja. Školska knjiga, Novi Sad, 1-367
Merrien A (1992): Some aspects of sunfl ower crop physiolo-gy, Proc. of 13th International Sunfl ower Conference, Pisa, Italy, 1: 481-498
Miklič V, Škorić D, Balalić I, Jocić S, Jovanović D, Hladni N, Marinković R, Joksimović J, Gvozdenović S (2007): Rezultati ispitivanja NS hibrida suncokreta u ogledima i preporuka za setvu u 2007. godini. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 43: 115-128
Miklič V, Jocić S, Miladinović D, Dusanić N, Hladni N (2008): Changes in seed oil content of sunfl ower (Helianthus annus L.), as affected by harvesting date. Proc 17th International Sunfl ower Conference, Cordoba, Spain 1:281-285
Miklič V, Balalić I, Jocić S, Marinković R, Hladni N, Gvozdeno-vić S, Stojšin V (2009): Produktivnost NS hibrida suncokreta u multilokacijskim ogledima i preporuka sortimenta za setvu u 2009. godini. Zbornik radova Instituta za ratarstvo i povr-tarstvo, Novi Sad 46: 293-310
Miklič V, Balalić I, Jocić S, Marinković R, Cvejić S, Hladni N, Miladinović D (2010): Ocena stabilnosti prinosa semena i ulja NS hibrida suncokreta u mikro-ogledima i preporuka sortimenta za setvu u 2010. godini. Ratar. Povrt. / Field Veg. Crop Res. 47: 131-146
Schoeman L J (2003): Genotype × environment interaction in sunfl ower (Helianthus annuus) in South Africa. PhD thesis. University of the Free State, Bloemfontein, South Africa
Statistica 8. Škorić D (1988): Sunfl ower breeding. J. Edible Oil Industry
„Uljarstvo“ 25: 1-90Škorić D, Joksimović J, Jocić S, Jovanović D, Marinković R,
Hladni N, Gvozdenović S (2005): Ocena vrednosti pro-duktivnih svojstava NS-hibrida suncokreta. Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 41: 21-33
Škorić D, Joksimović J, Jocić S, Jovanović D, Marinković R, Hladni N, Gvozdenović S (2006): Rezultati dvogodišnjih is-pitivanja novosadskih hibrida suncokreta u mikroogledima. Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 42: 61-74
Vranceanu A V (2000): Floarea-sourelni hibrida. Editura Ceres, Bucharest

Miklič V i sar.66
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 57-66
Productivity of NS Sunfl ower Hybrids in Small-Plot Trials in Serbia in 2010
Vladimir Miklič · Igor Balalić · Siniša Jocić · Radovan Marinković · Sandra Cvejić · Nada Hladni · Dragana Miladinović
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: The objective of this study was the evaluation of NS sunfl ower hybrids on the basis of the results of seed yield, seed oil content and oil yield obtained in a network of small-plot trials in Vojvodina and central Serbia in 2010, as well as recommendations for the sowing season 2011. Seed yield, seed oil content and oil yield were signifi cantly different in small-plot trials in the investigated regions. In Vojvodina, the highest average seed yield was produced by Duško (3.08 t ha-1), Sremac (2.98 t ha-1), Orfej (2.97 t ha-1), Velja (2.93 t ha-1) and NS-H-111 (2.92 t ha-1). In the region of central Serbia, the highest seed yield was achieved by the hybrids NS-H-111 (2.97 t ha-1) and Duško (2.87 t ha-1). Hybrid Baća showed the highest average seed oil content both in the region of Vojvodina (48.54%) and central Serbia (48.80%). The highest average oil yield (over 1.30 t ha-1) was produced by Duško, Ba-čvanin and NS-H-111 in the Vojvodina region, and in central Serbia oil yield (over 1.30 t ha-1) by NS-H-111, Oskar and Novosađanin. In the three-year trials in Vojvodina four hybrids (Duško, Sremac, Velja and NS-H-111) showed seed yield over general mean. The highest seed yield was reached by hybrid Duško (3.54 t ha-1). Key words: location, oil yield, seed oil content, seed yield, small-plot trials, sunfl ower

Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice 67
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76originalni naučni rad / original research article
Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice (Brassica napus L.)
Ana Marjanović-Jeromela · Sreten Terzić · Miroslav Zorić · Radovan Marinković · Jovanka Atlagić · Petar Mitrović · Željko Milovac
primljeno / received: 03.11.2010. prerađeno / revised: 23.11.2010. prihvaćeno / accepted: 25.11.2010.© 2011 IFVC
Izvod: U cilju ocene stabilnosti prinosa semena i ulja NS sorti uljane repice u promenljivim klimatskim uslovima severnog dela Srbije, urađena je analiza ogleda izvedenih u periodu 2007-2010. Analiziran je prinos semena, sadržaj i prinos ulja 40 sorti ozime i 9 sorti jare repice. Utvrđeno je da su godine imale najznačajniji uticaj na prinos semena i ulja. Interakcija genotip (sorta) × spoljašnja sredina (godina) dalje je analizirana primenom AMMI (aditivni glavni efekti i višestruka interakcija) modela. U uslovima koji su vladali u godinama ispitivanja na lokalitetu Rimski Šančevi, prema analizi regresionog koefi cijenta (bi) i ekovalence (Wi) i AMMI modela, za prinos semena kao najstabilnije i visokoprinosne su se izdvojile ozime sorte Nena i NS-L-102, a za prinos ulja i hibridna sorta NS-H-R-3. Ove sorte se preporučuju za uključivanje u dalje oplemenjivačke programe i za proizvodnju u uslovima navedenim u radu. Ključne reči: interakcija sa izmenom ranga, prinos semena, prinos ulja, stabilnost
A. Marjanović-Jeromela · S. Terzić · R. Marinković · J. Atlagić · P. Mitrović · Ž. MilovacInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, SrbijaM. Zorić Tehnološki fakultet, Bulevar Cara Lazara 1, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Uljana repica (Brassica napus L.) je u svetskim razmerama na trećem mestu kao izvor biljnih ulja i obezmašćene sačme (www.faostat.org). Ozime forme se uglavnom gaje u umerenom klimatskom pojasu, dok se jare sorte gaje u hlad-nijim klimatskim područjima. Stare sorte uljane repice su u ulju imale čak do 50% eruka kise-line (C22:1) što je masna kiselina bez hranljive vrednosti, štetna po zdravlje jer oštećuje krvne sudove i izaziva hemolitsku anemiju. Intenziv-nim oplemenjivačkim radom stvorene su sorte sa manje od 2% eruka kiseline u ukupnom sadr-žaju ulja („0“ sorte). Ovakve sorte se mogu bez zdravstvenih ograničenja koristiti u prehrambe-ne svrhe.
Uljane pogače, koje ostaju nakon ekstrakcije ulja su visokokvalitetno proteinska komponen-ta u koncentrovanoj hrani za domaće životinje. Ograničavajući činilac korišćenja sačme u ishrani bili su glukozinolati i alkaloidi koji se u organima za varenje životinja razlažu na vrlo toksična jedi-
njenja. Njihov sadržaj je kod tradicionalnih sorti iznosio i do 450 mmol g-1. Savremene sorte uljane repice sadrže svega 15-20 mmol g-1 glukozinola-ta što se smatra bezopasnom količinom u ishra-ni domaćih životinja („00“) (Friedt & Snowdon 2009).
Danas se često uljane repice sa niskim sadrža-jem štetnih materija u semenu nazivaju canola, a ulje dobiveno od njih kanola ulje. Etimološki reč canola je skraćenica od sledećih reči: Canadian Oil Low Acid.
Stаlno povećаnje potrebа zа hrаnom i sirovinаmа zа prerаdu, nа koje morа dа odgovori poljoprivrednа proizvodnjа, rаsprostrаnjenost i znаčаj uljаnih biljnih vrstа, kаo rаzlozi zа poveća-nje prinosа u Srbiji zahtevaju stalno proučаvаnje genetike i njenu primenu u oplemenjivаnju uljаne repice. Nepovoljаn uticаj fаktorа spoljne sredine nа prinos i kvаlitet uljаne repice znаčаjno može redukovаti efekte rаdа oplemenjivаčа u reаlizаciji perspektivnih sorti.
Nove sorte testiraju se u različitim uslovima spoljašnjih sredina (godina, lokalitet) u cilju iz-
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( ) Ovo istraživanje je rezultat projekta TR31025 „Razvoj novih sorti i poboljšanje tehnologija proizvodnje uljanih biljnih vrsta za različite namene“ Ministarstva za nauku i tehnološki razvoj Republike Srbije / This research results from project TR31025 „Development of new varieties and production technology improvement of oil crops for di-fferent purposes“ funded by Ministry of Science and Technological Development of the Republic of Serbia.

Marjanović-Jeromela A i sar.68
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
bora sorte i davanja preporuke proizvođačima za setvu. Interakcija genotipa i spoljašnje sre-dine komplikuje agronomska i oplemenjivačka istraživanja, jer se vrednost neke osobine ne može tumačiti samo na osnovu glavnih efekata: genotipa i spoljašnje sredine. Interakcija otežava preporuku sortimenta za neki region, odnosno specifi čnu spoljašnju sredinu (Ebdon & Gauch 2002). Sa povećanjem uticaja interakcije, teže je izabrati najbolje genotipove. Ukoliko se zbog interakcije rang genotipova u različitim sredi-nama menja, genotipovi odabrani kao najbo-lji u jednoj sredini mogu u drugoj biti izrazito slabi. Interakcija koja uzrokuje promene ranga genotipova zove se kvalitativna interakcija ili in-terakcija sa izmenom ranga (Baker 1988) i od velikog je značaja za oplemenjivanje biljaka čime se ističe važnost analize frekvencije i magnitude ovog tipa interakcije u oplemenjivačkim ogledi-ma (Singh et al. 1999).
Sorte koje imaju manji doprinos interakciji manje su osetljive na promene uslova sredine, pa se vrednosti ispitivanih osobina neće mno-go menjati sa promenom uslova sredine. Takve sorte su stabilne. Jedna od najznačajnijih i najvi-še upotrebljavanih multivarijacionih statističkih modela je AMMI (engl. Additive Main Effects and Multiplicative Interaction) prema Gauch & Zobel (1996). Isti autori ga smatraju inicijalnim mode-lom u analizi podataka iz višegodišnjih ili višelo-kacijskih oplemenjvačkih ogleda, jer omogućuje analitički pristup u dijagnozi drugih pogodnijih statističkih modela za analizu podataka. Naro-čito je pogodan za razumevanje kompleksnosti odnosa genotipa i spoljašnje sredine (Zobel et al. 1988, Crossa et al. 1990). Autori Shafi i & Pri-ce (1998) ističu prednost AMMI modela u situ-aciji značajne interakcije, a ne značajnih glavnih efekata ili kada na strukturu interakcije utiče jedna ili više ekstremnih vrednosti. Ovom se analizom otkriva visoko značajna komponenta interakcije koja ima odgovarajuće agronomsko značenje. Veličina interakcije pokazuje uticaj spoljašnje sredine na adaptabilnost i stabilnost, koja je poželjna osobina samo ukoliko je u vezi sa prinosom iznad proseka (Yan & Hunt 2003). Uprkos širokoj primeni ovog modela u literaturi jedan od značajnih nedostataka je nemogućnost razlikovanja interakcija sa izmenom ranga od in-terakcija bez izmene ranga.
Cilj istraživanja u ovom radu je bio da se oce-ni stabilnost NS sorti uljane repice u varijabilnim klimatskim uslovima severnog dela Srbije. Ovakvi rezultati su veoma značajni za razvoj i preporuku najboljih sorti za specifi čna područja gajenja kao i za odabir kriterijuma u selekciji.
Materijal i metode rada
Eksperimentalni materijal u ogledu vrlo je vari-jabilan i obuhvata sortu Banaćanka, koja je stan-dard za sorte ozime uljane repice u Komisiji za priznavanje sorti uljanih biljaka u Republici Srbi-ji, zatim registovane ozime sorte (Slavica, Nena, Kata, Branka, Zlatna, Nevena, Jasna, Zorica), sorte u priznavanju (Ilia, Elena, Marija), kao i hibridne sorte u postupku registracije (NS-H-1, NS-H-2, NS-H-3), a takođe i jare sorte i to regi-strovane u Srbiji (Jovana, Mira) i 7 sorti u postup-ku testiranja. Jare sorte uključene su u ispitivanje kako bi se testirala njihova pripadnost fakultativ-nom tipu sorti. Analiziran je prinos semena i ulja kod 49 sorti uljane repice iz selekcionog programa Odeljenja za uljane kulture Instituta za ratarstvo i povrtarstvo Novi Sad. Ova istraživanja predstav-ljaju deo kontinuiranog rada na ispitivanju sorti uljane repice u našim agroekološkim uslovima i u radu je prikazan jedan deo rezultata višegodišnjih mikro-ogleda koji je izveden na oglednom polju Instituta za ratarstvo i povrtarstvo na Rimskim Šančevima (SGD: 19° 51’; IGŠ: 45° 20’; nmv: 84 m) na zemljištu tipa černozem. Ogled je postav-ljen po slučajnom blok sistemu u tri ponavljanja tokom vegetacionih sezona 2007-2008, 2008-2009. i 2009-2010. Setva je obavljena mašinski u prvoj polovini septembra na međuredni razmak 25 cm, a razmak unutar reda od 5 cm dobijen je proređivanjem u fazi F4-F5. Veličina ogledne parcele je 5 m2. Tokom vegetacije primenjena je uobičajena agrotehnika za ovu kulturu, a žetva je izvršena u optimalnom roku u prvoj i trećoj go-dini ogleda, dok je u drugoj godini kasnila zbog obilnih i učestalih padavina.
Prinos semena u tehnološkoj zrelosti biljaka meren je nakon žetve i obračunat na 9% vlage. Sadržaj ulja određen je NMR (nuklearno-magnet-na rezonanca) metodom u Hemijskoj laboratoriji Odeljenja za uljane kulture Instituta za ratarstvo i povrtarstvo Novi Sad. Prinos ulja izračunat je kao proizvod prinosa semena i sadržaja ulja.
Inicijalna obrada podataka izvršena je prime-nom analize varijanse faktorijalnog ogleda pre-ma modelu slučajnog blok sistema, gde su svi efekti u modelu tretirani kao fi ksni. S obzirom na činjenicu da je ogled izveden tokom trogo-dišnjeg perioda, upotrebljena je i kombinovana analiza varijanse sa dva faktora u izračunavanju glavnih efekata i odgovarajućih interakcija prvog reda. Na osnovu vrednosti sredina kvadrata po-greške, izračunati su LSD testovi za poređenje razlika između godina i sorti. Statistička obrada podataka urađena je primenom programa Stati-stica 8.0.

Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice 69
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Ocena stabilnosti i adaptabilnosti je izvršena koristeći dva parametra: ekovalencu (Wi), i regre-sioni koefi cijent (bi) (Wricke, 1962). Rezultati su interpretirani po modelu Haufe & Geidel (1978; cit. prema Becker & Leon 1988).
Suma kvadrata interakcije sorte i godine ana-lizirana je primenom AMMI modela (Gauch & Zobel 1996), a fi nalni rezultati modela su prika-zani grafi čki. Broj adekvatnih komponenata za aproksimaciju interakcije odredjen je postdiktiv-nim F testom (Gauch & Zobel 1988). Statistička značajnost i broj interakcija sa izmenom ranga je analiziran primenom parametrijskog Azzali-ni–Cox testa (Azzalini & Cox 1984, Baker 1988). Za pretpostavljeni broj sorti i godina ukupni broj mogućih 2 × 2 kombinacija je izračunat prema sledećoj formuli: a(a–1)b(b–1)/4 gde je a – uku-pan broj sorti u ogledu i b – ukupan broj godina u kojima su sorte testirane. Nulta hipoteza o po-stojanju interakcija sa izmenom ranga se prihvata ukoliko za bilo koji par sorti i godina sredina ap-solutne vrednosti sorti prelaze kritičnu vrednost u obe spoljašnje sredine i imaju suprotan pred-znak razlika od jedne do druge spoljne sredine.
Rezultati i diskusija
Radi lakšeg tumačenja rezultata razdvojene su ozime i jare sorte. Kod ozimih sorti prinos seme-na je varirao od 0,68 t ha-1 (NS-L-46 u 2008/09) do 4,88 t ha-1 (NS-L-44 u 2007/2008); sadržaj ulja od 37,5% (NS-L-51 u 2009/10) do 47,2% (NS-L-102 u 2007/08); prinos ulja od 0,25 t ha-1
(NS-L-46 u 2008/09) do 2,20 t ha-1 (NS-L-44 u 2007/08). Značajna variranja zapažena su i kod jarih sorti, tako je prinos semena varirao od 0,12 t ha-1 (JR-NS-9 u 2008/09) do 1,32 t ha-1 (JR-NS-6 u 2007/2008); sadržaj ulja od 37,1% (JR-NS-26 u 2008/09) do 49,2% (JR-NS-11 u 2007/08); pri-nos ulja od 0,05 t ha-1 (JR-NS-9 u 2008/09) do 1,42 t ha-1 (JR-NS-11 u 2007/08). Najviše pro-sečne vrednosti ovih svojstava u trogodišnjem periodu bile su kod ozimih sorti NS-L-47 za pri-nos semena (2,83 t ha-1), NS-L-102 za sadržaj ulja (44%), NS-L-102 i NS-L-47 za prinos ulja (1,17 t ha-1). Kod jarih sorti izdvojile su se po visini proseka JR-NS-6 za prinos semena (1,93 t ha-1), JR-NS– 11 za sadržaj ulja (45,1%), JR-NS-6 za prinos ulja (0,82 t ha-1) (Graf. 1 i 2).
Grafi kon 1. Prinos semena kroz tri godine zа 49 sorti uljаne repice, Rimski Šаnčevi Graph. 1. Seed yield in three years for 49 rapeseed cultivars, Rimski Šаnčevi
Ozime sorte / Winter cultivars: 1. NS-H-R 1, 2. NS-H-R 2, 3. NS-H-R-3, 4. Banaćanka, 5. Slavica, 6. NS-L-51, 7. NS-L-110, 8. Zlatna, 9. NS-L-74, 10. Branka, 11. NS-L-24, 12. NS-L-7, 13. Nevena, 14. NS-L-23, 15. Ilia, 16. Kata, 17. Nena, 18 .NS-L-31, 19. NS-L-126, 20. NS-L-33, 21. NS-L-128, 22. NS-L-129, 23. Jasna, 24. NS-L- 101, 25. Zorica, 26. NS-L-102, 27. NS-L-134, 28. NS-L-32, 29. NS-L-136, 30. NS-L-137, 31 NS-L-138, 32. NS-L-251, 33. NS-L-210, 34. NS-L- 44, 35. NS-L- 45, 36. NS-L- 46, 37. NS-L- 47, 38. NS-L- 52, 39. Forward, 40. Maidan; Jare sorte / Spring cultivars: 41. JR-NS-6, 42. Mira, 43. Jovana, 44. JR-NS-26, 45. JR-NS-28, 46. JR-NS-36, 47. JR-NS-7, 48. JR-NS- 9, 49. JR-NS– 11

Marjanović-Jeromela A i sar.70
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Na osnovu LSD testa uočljiva je visoko značaj-na razlika između godina. U klimatski veoma po-voljnoj 2007/08. prosečan prinos semena bio je 3,75 t ha-1, odnosno kod ozimih 3,99 t ha-1, a kod jarih 2,65 t ha-1, dok je u sledećoj, izrazito nepo-
voljnoj godini 2008/09. prosečan prinos bio 1,06 t ha-1, odnosno kod 40 ozimih sorti bio je samo 1,17 t ha-1, a kod devet jarih 0,56 t ha-1. Treća go-dina 2009/10. imala je takođe niže prinose od prosečnih u ovom regionu (Marjanović-Jeromela
Grafi kon 2. Prinos ulja kroz tri godine zа 49 sorti uljаne repice, Rimski Šаnčevi Graph 2. Three year oil yield for 49 rapeseed cultivars, Rimski Šаnčevi
Ozime sorte / Winter cultivars: 1. NS-H-R 1, 2. NS-H-R 2, 3. NS-H-R-3, 4. Banaćanka, 5. Slavica, 6. NS-L-51, 7. NS-L-110, 8. Zlatna, 9. NS-L-74, 10. Branka, 11. NS-L-24, 12. NS-L-7, 13. Nevena, 14. NS-L-23, 15. Ilia, 16. Kata, 17. Nena, 18 .NS-L-31, 19. NS-L-126, 20. NS-L-33, 21. NS-L-128, 22. NS-L-129, 23. Jasna, 24. NS-L- 101, 25. Zorica, 26. NS-L-102, 27. NS-L-134, 28. NS-L-32, 29. NS-L-136, 30. NS-L-137, 31 NS-L-138, 32. NS-L-251, 33. NS-L-210, 34. NS-L- 44, 35. NS-L- 45, 36. NS-L- 46, 37. NS-L- 47, 38. NS-L- 52, 39. Forward, 40. Maidan; Jare sorte / Spring cultivars: 41. JR-NS-6, 42. Mira, 43. Jovana, 44. JR-NS-26, 45. JR-NS-28, 46. JR-NS-36, 47. JR-NS-7, 48. JR-NS- 9, 49. JR-NS– 11
Tabela 1. Deskriptivna statistika zа prinos semena i ulja kroz tri godine zа 49 sorti uljаne repice, Rimski Šаnčevi Table 1. Descriptive statistics for three year seed and oil yield for 49 rapeseed cultivars, Rimski Sancevi
GodinaYear
Prosek Average Min. Max. CV (%)
Prinos semena (t ha-1)Seed yield ( t ha-1)
2007/2008 3,75 2,35 4,88 22,42008/2009 1,06 0,12 1,82 44,62009/2010 2,03 0,47 2,72 40,9
LSD 1% 0,11 5% 0,44
Prinos ulja ( t ha-1)Oil yield (t ha-1)
2007/2008 1,63 1,05 2,20 21,92008/2009 0,43 0,05 0,58 45,12009/2010 0,73 0,20 1,12 41,9
LSD 1% 0,05 5% 0,18

Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice 71
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
et al. 2008a, Marjanović-Jeromela i sar. 2008b), prvenstveno zbog problema sa sušom u vreme klijanja i nicanja, duge i hladne zime i nepovolj-nog rasporeda padavina. Prosečan prinos u ovoj godini bio je 2,03 t ha-1, kod ozimih 2,02 t ha-1, a kod jarih 0,84 t ha-1. Sličan trend može se uočiti kod sadržaja ulja, jer se nepovoljni klimatski uslo-vi odražavaju i na metabolizam biljke pri sintezi ulja, kao i na prinos ulja, kao proizvod vrednosti dva prethodno navedena svojstva (Tab. 1).
Ovakva pravilnost odražava se i na koefi cijent varijacije ovih svojstava. Posmatrajući sumarne rezultate za oba tipa sorti koefi cijent varijacije je u prvoj godini za prinos semena 22,4% i prinos ulja 21,9%, dok su u drugoj i trećoj vegetacionoj sezoni, ove vrednosti značajno veće i iznose 44, 6% i 45,1% u 2008/09, odnosno 40,9% i 41,9% u 2009/10. (Tab. 1).
Analizom varijanse uočava se da je godina sa preko 80% učestvovala u variranju ispitivanih osobina (81,0% za prinos semena, 83,6% za pri-nos ulja), zatim je uticaj sorte (13,1% i 11,3%) i
njihove interakcije (5,8% i 5,1%) (Tab. 2). U svo-jim istraživanjima na uljanoj repici Marjanović-Je-romela (2005) takođe ističe značajnost razlika za interakciju sorta×godina za prinos i komponente prinosa semena uljane repice.
U tabeli 3. su prikazani rezultati analize interak-cija sa izmenom ranga primenom Azzalini-Cox (1984) testa za oba proučavana svojstva. Pored toga, ista analiza je ponovljena izdvojeno za ozi-me, odnosno jare sorte. Kao što je i očekivano, kod prinosa semena je detektovan nešto veći broj značajnih interakcija sa izmenom ranga (254 od ukupno mogućih 3528, odnosno 7,2%) u odno-su na prinos ulja (241 od ukupno mogućih 3528, odnosno 6,8%) (Tab. 3), što se može objasniti većom genetskom kompleksnošću prinosa seme-na kao i većim uticajem klimatskih faktora na ek-spresiju ovoga svojstva, od kojih se ističe prolećni nedostatak vlage (Wricke 1962).
Analizom interakcija po tipu sorti ustanovljen je minimalan broj značajnih interakcija kod jarih sorti u odnosu na ozime. Na osnovu rezultata
Tabela 2. Učešće u sumi kvаdrаtа (SS) i sredinа kvаdrаtа (MS) аnаlize vаrijаnse zа AMMI model zа prinos semena i ulja kroz tri godine zа 49 sorti uljаne repice, Rimski Šаnčevi Table 2. Partitioning Sums of Squares (SS) and Mean Square (MS) effect variance analysis in AMMI model for seed and oil yield in three years for 49 rapeseed cultivars, Rimski Šаnčevi
Izvor varijacijeVariation source df
SS (%) MSa SS (%) MSa
Prinos semenaSeed yield
Prinos uljaOil yield
Sorta/Cultivar 48 13.1 0.69 11.3 0.12Godina/Year 2 81.0 101.9 83.6 20.8Interakcija/Interaction 96 5.8 0.15 5.1 0.03 AMMI-1 49 52.9 0.16 55.9 0.029 AMMI-2 47 47.1 0.15 44.1 0.023Pogreška/Error 288 - 0.08 - 0.014a svi efekti u modelu su visoko značajni (P < 0.01) / all effects in the model are highly signifi cant (P < 0.01)
Tabela 3. Analiza interakcija sa izmenom ranga primenom Azzalini–Cox testaTable 3. Interaction analysis with rank change using Azzalini–Cox test
SortaCultivar
Br. kombinacija/Br. interakcija sa izmenom rangaNo. of combinations/No. of
rank change interactions*
% značajnosti% of
signifi cance
Br. kombinacija/Br. interakcija sa izmenom ranga
No. of combinations/No. ofrank change interactions*
%značajnosti% of
signifi cance
Prinos semena / Seed yield Prinos ulja / Oil yieldSve sor.All cul. 3528 / 254 7,2 3528 / 241 6,8Ozimi Winter 2340 / 226 9,7 2340 / 198 8,5JariSpring 108 / 1 0,9 0 / 108 0,0*P < 0.05

Marjanović-Jeromela A i sar.72
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
ovih istraživanja jare sorte su bile otpornije na stresne uslove godine gajenja u odnosu na ozime što se može objasniti njihovim poreklom. Jare sorte u ogledu nastale su ukrštanjem sorti i linija poreklom iz Kanade i Rusije, a ozime imaju za-jednički pedigre sa sortama iz područja zapadne i jugoistočne Evrope.
Daljim raščlanjavanjem interakcije dobija se vrednost za AMMI-1 koja objašnjava 52,9% sume interakcije sorta×godina za prinos seme-na, odnosno 55,9% za prinos ulja (Tab. 2). Prva komponenta AMMI analize obuhvata 49 stepeni slobode što je 51% ukupnog broja stepeni slo-bode interakcije, tako da je AMMI model veoma efi kasan u analizi interakcije u ovim istraživanji-ma. S obzirom da je postdiktivni F test ukazao na statističku značajnost obe interakcijske kom-ponente procena realne strukture interakcije sa-držane u sumu kvadrata interakcije (Tab. 2) izvr-šena je primenom jednostavnog postupka opi-sanog od strane Gauch & Zobel (1996). Prema ovom metodu, vrednost realne strukture inte-rakcije za prinos semena iznosi 52,9%, dok je za prinos ulja ta vrednost 55,9%. Ovakav rezultat nam ukazuje da je za adekvatan opis interakci-je kod oba svojstva potrebno koristiti AMMI–1 model, odnosno da u drugoj interakcijskoj kom-ponenti dominira šum ili nesistematska varijacija koja je nastala kao proizvod interakcije sorti sa uslovima godine. Autori Crossa et al. (1990) na-vode da faktori kao što su biljna vrsta, genetska divergentnost ispitivanih sorti i variranje uslova spoljašnje sredine uslovljavaju kompleksnost in-terakcije, a time i izbor “najboljeg” modela za opis interakcije. U ovim istraživanjima u grafi č-kom prikazu koristili smo samo AMMI-1 mo-del, što je u saglasnosti sa saopštenjem autora Gauch & Zobel (1996) koji na osnovu analize velikog broja radova o primeni AMMI modela u oceni interakcije genotip×spoljašnja sredina (G×E) zaključuju da je u najvećem broju sluča-jeva (70%) AMMI model sa značajnom prvom glavnom komponentom (IPC1) bio najbolji mo-del za adekvatan opis interakcije.
Grafi kon za AMMI-1 analizu se konstruiše tako da se na apscisu nanose vrednosti glavnih efekata (sorta i godina), a na ordinatu vrednosti za prvu interakcijsku osu (IPC1). Svakoj sorti i godini tada pripada tačka čija je prva koordina-ta vrednost glavnog efekta, a druga koordinata interakcijska vrednost. Na ovaj način poređenje sorti, odnosno godine po horizontali ukazuje na razlike u glavnim efektima, dok poređenje po vertikali ukazuje na razlike u interakcijskom efektu. Sorte i godine sa visokom vrednošću IPC1 komponente imaju veliki efekat interakci-
je. Godine sa IPC1 blizu nulte vrednosti s obzi-rom da imaju mali efekat interakcije su pogod-ne za sve sorte. Niska vrednost IPC1 ose sorti ukazuje da su oni pod manjim uticajem uslova godine (Zobel et al. 1988). Sa agronomskog sta-novišta poželjne su sorte čije su vrednosti IPC1 bliske nuli, a da su prosečni prinosi oko i iznad opšteg proseka. Sorte grupisane na grafi konu imaju sličnu adaptabilnost, dok grupisane spo-ljašnje sredine utiču na sorte na sličan način (Ba-lalić 2009). Blizina tačaka sorti i godina na gra-fi konu ne ukazuje na visok nivo adaptabilnosti sorte uslovima kakvi su bili u toj godini. Njihov položaj može se objasniti time da sorte i godi-ne sa istim predznakom (+/-) IPC1 ose, imaju pozitivnu interakciju, odnosno te sorte će imati viši prinos u takvim uslovima. Obrnuto, sorte koje imaju različit predznak IPC1 sa godinama sa njima imaju negativnu interakciju (Crossa et al. 1990).
Na osnovu rezultata AMMI-1 grafi kona, koji na slikovit način omogućava predstavljanje i razumevanje interakcije sorta x godina uoča-va se da je u 2009/10. bilo najmanje variranje sorti u prinosu semena jer se ona nalazi najbli-že liniji stabilnosti. Prva i druga godina imaju daleko veći doprinos interakciji, s tim što je prinos u prvoj godini bio značajno veći (Graf. 3). Uočljivo je razdvajanje sorti u okviru grafi -kona na jare i ozime forme. Jare sorte JR-NS-7 i JR-NS–11 bile su najstabilnije i sa visokim prinosom semena. Različito reagovanje sorti u pogledu prinosa na promenu uslova godine može se delimično objasniti i njihovim pore-klom, tj. kombinacijama roditelja i metodama oplemenjivanja korišćenim u njihovom stva-ranju (Shafi i et al. 1992, Marjanović-Jeromela et al. 2008a). Kod ozimih sorti po stabilnosti i visokom prinosu semena su se izdvojile sor-te Nena i NS-L-102. Nešto nižeg prinosa, ali izrazito stabilna je i hibridna sorta NS-H-R-3 i sorte NS-L-32, NS-L-251, NS-L-52. Na osno-vu navedenih kriterijuma navedene sorte mogu se uključiti u oglede komisija za registraciju i uvođenje sorti u proizvodnju.
Na grafi konu 4. prikazana je AMMI-1 analiza za prinos ulja. Uočava se sličan raspored u pogle-du godina kao i za prinos semena. Treća godina ima zanemarljiv doprinos interakciji, što znači da sorte korišćene u istraživanjima u klimatskim uslovima, kakvi su bili u 2009/10. imaju ujedna-čenije vrednosti prinosa ulja u odnosu na klimat-ske uslove kakvi su bili u prvoj i drugoj godini. Pri tome, na osnovu položaja sorti u grafi konu uočava se da sa prvom godinom većina ozimih sorti ima izrazitu pozitivnu interakciju. Raspored

Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice 73
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Grafi kon 3. AMMI-1 grafi kon za prinos semena 49 sorti uljane repice, Rimski ŠančeviGraph 3. AMMI-1 seed yield graph for 49 rapeseed cultivars, Rimski Šančevi
Grafi kon 4. AMMI-1 grafi kon za prinos ulja 49 sorti uljane repice, Rimski ŠančeviGraph 4. AMMI-1 oil yield graph for 49 rapeseed cultivars, Rimski Šančevi

Marjanović-Jeromela A i sar.74
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Grafi kon 5. Stabilnost i adaptabilnost prinosa semena za 49 sorti uljane repice na Rimskim ŠančevimaGraph 5. Stability and adaptability of seed yields for 49 rapeseed cultivars in Rimski Šančevi
Grafi kon 6. Stabilnost i adaptabilnost prinosa ulja za 49 sorti uljane repice na Rimskim ŠančevimaGraph 6. Stability and adaptability of oil yields for 49 rapeseed cultivars in Rimski Šančevi

Ocena stabilnosti prinosa semena i ulja NS sorti uljane repice 75
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Zaključak
Na osnovu rezultata višegodišnjeg ogleda utvr-đeno je da su variranja prinosa semena, sadržaja i prinosa ulja u najvećem procentu uzrokovana uti-cajem godine, odnosno različitih klimatskih uslo-va. Sorta je takođe bila značajan faktor variranja. Jare sorte imaju niži prinos semena i ulja od ozi-mih, a prosečan sadržaj ulja nije se značajno ra-zlikovao. Interakcija sorta x godina je imala nešto manji, ali značajan udeo u variranju svojstva kod 49 analiziranih sorti. Jare sorte gotovo da ne me-njaju rang između godina, tj. vrednosti svojstva ostaju slične u različitim godinama, dok je kod ozimih sorti uočena obrnuta pravilnost. AMMI-1 grafi čka analiza je omogućila vizuelnu ocenu adaptabilnosti i stabilnosti sorti u različitim go-dinama. Na osnovu dobijenih rezultata izdvojene su stabilne sorte čije vrednosti za prinos semena i ulja najmanje variraju pod uticajem promena u spoljašnjoj sredini (godini). Kod ozimih sorti kao visokoprinosne i stabilne izdvojile su se sorte Nena i NS-L-102 i hibridna sorta NS-H-R-3.
sorti kod jarih ističe visok prinos, ali izrazitu ni-sku stabilnost sorte JR-NS-6, dok najstabilnija sorta JR-NS-26 ima prinos ulja oko proseka za ovaj tip sorti. Kod ozimih sorti kao visokoprino-sne i stabilne izdvojile su se sorte Nena i NS-L-102 i hibridna sorta NS-H-R-3. Osim njih u dalji program oplemenjivanja, testiranja i registracije treba uključiti i sorte koje se nisu izdvojile po vi-sokom prinosu, ali su izrazito stabilne, jer bi u višegodišnjim multilokacijskim ogledima mogle ostvariti značajno bolje prosečne prinose. Autori Piepho (1988) i Ceccareli (1994) navode da poljo-privredni proizvođači stabilnost prinosa smatraju najznačajnijim socio-ekonomskim ciljem u biljnoj proizvodnji i to naročito u ekstremnim uslovima spoljašnje sredine i zato je važno u proizvodnji koristiti stabilne sorte uljane repice, koje daju do-bre prosečne prinose u različitim agroekološkim uslovima.
Pri analizi fenotipske stabilnosti preporučljivo je da se koriste u kombinaciji regresioni i AMMI modeli (Ferreira et al. 2006). Nakon analize re-gresionog koefi cijenta (bi) i ekovalence (Wi) usta-novljeno je da se analizirane sorte razlikuju po prinosu semena ali ih karakterišu bi ≈ 1 i niska Wi. U ovom istraživanju to je bilo najprimetnije kod linija NS-L-24 i NS-L-134, kao i kod jedne registrovane sorte Nena, ali i linije NS-L-102 koja se dobro pokazala i u AMM-1 analizi za prinos semena (Graf. 3. i 5).
Slični rezultati su dobijeni za prinos ulja, sa bi ≈ 1 i najnižim Wi kod linija NS-L-134, NS-L-129 i registrovane sorte Banaćanka. AMM-1 analiza za prinos ulja je pokazala kao najstabilniju i najpri-nosniju hibridnu sortu NS-H-R 3 koja se i po re-gresionom koefi cijentu i ekovalenci nalazi u vrhu ispitivanog materijala (Graf. 4. i 6).

Marjanović-Jeromela A i sar.76
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 67-76
Literatura
Azzalini A, Cox D R (1984): Two new tests associated with analysis of variance. J. R. Statist. Soc. Series B 46: 335-343
Baker R J (1988): Tests for crossover genotype–environmental interactions. Can. J. Plant Sci. 68: 405-410
Balalić I (2009): Multivarijaciona analiza uticaja interakcije hibri-da i rokova setve na sadržaj ulja, prinos i komponente prino-sa suncokreta. Doktorska disertacija, Poljoprivredni fakultet, Univerzitet u Novom Sadu
Becker H C, Leon J (1988): Stability analysis in plant breeding. Plant Breed. 101: 1-23
Ceccareli S (1994): Specifi c adaptation and breeding for margi-nal conditions. Euphytica 77: 205-219
Crossa J, Gauch H G, Zobel R W (1990): Additive main effects and multiplicative interaction analysis of two international maize cultivar trials. Crop Sci. 30: 493-500
Ebdon J S, Gauch H G (2002): Additive main effect and mul-tiplicative interaction analysis of national turfgrass perfor-mance trials. II Cultivar recommendations. Crop Sci. 42: 497-506
Ferreira D F, Demetrio C G B, Manly B F J, Machado A A, Vencovsky R (2006): Statistical models in agriculture: Bio-metrical methods for evaluating phenotypic stability in plant breeding. Cerne, Lavras, 12: 373-388
Friedt W, Snowdon R J (2009): Oilseed Rape. In: Johann Vo-llman, Istvan Rajcan (eds), Oil Crops, Springer Dordrecht Heidelberg London New York, 91-127
Gauch H G, Zobel R W (1988): Predictive and postdictive su-ccess of statistical analysis of yield trials. Theor. Appl. Ge-net. 76: 1-10
Gauch H G, Zobel R W (1996): AMMI analysis of yield trials. In: M.S. Kang and H.G. Gauch (ed.) Genotype-by-envi-ronment interaction. CRC Press, Boca Raton, FL, 85-122
Marjanović-Jeromela A (2005): Genetička divergentnost i vari-
jabilnost komponenti prinosa semena uljane repice (Brassica napus L.). Doktorska disertacija, Poljoprivredni fakultet, Uni-verzitet u Novom Sadu
Marjanović-Jeromela A, Marinković R, Mijić A, Jankulovska M, Zdunić Z, Nagl N (2008a): Oil Yield Stability of Winter Rapeseed (Brassica napus L.) Genotypes. Agric. Conspec. Sci. 73: 217-220
Marjanović-Jeromela A, Marinković R, Mitrović P, Miklič V (2008b): Proizvodne vrednosti novih genotipova uljane re-pice (Brassica napus L.). Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 45: 103-110
Piepho H P (1998): Method for comparing the yield stability of cropping systems. J. Agron. Crop Sci. 180: 193-213
Shafi i B, Mahler K A, Price W J, Auld D L (1992): Genotype x environment interaction effects on winter rapeseed yield and oil content. Crop Sci. 32: 922-927
Shafi i B, Price W J (1998): Analysis of genotype-by-envi-ronment interaction using the additive main effects and mul-tiplicative interaction model and stability estimates. J. Agric. Biol. Environ. Stat. 3: 335-345
Singh M, Ceccarelli S, Grando S (1999): Genotype × envi-ronment interaction of crossover type: detecting its presen-ce and estimating the crossover point. Theor. Appl. Genet. 99: 988-995
Wricke G (1962): Über eine methode zur erfassung der ökolo-gischen streubreite in feldversuchen. Z. Pfl anzenzüchtg 47: 92-96
Yan W, Hunt L A (2003): Biplot analysis of multi-environment trial data. In: M. S. Kang (ed.) „Quantitative Genetics, Geno-mics, and Plant Breeding“, CAB International, Wallingford, Oxon, UK, 289-303
Zobel R W, Wright M J, Gauch H G (1988): Statistical analysis of a yield trial. Agron. J. 80: 388-393
Evaluation of Seed and Oil Yield Stability in NS Rapeseed Cultivars(Brassica napus L.)
Ana Marjanović-Jeromela1 · Sreten Terzić1 · Miroslav Zorić2 · Radovan Marinković1 · Jovanka Atlagić1 · Petar Mitrović1 · Željko Milovac1
1Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia2Faculty of Technology, University of Novi Sad, Bulevar Cara Lazara 1, 21000 Novi Sad, Serbia
Summary: Rapeseed trials performed in the period 2007-2010 were analyzed to evaluate seed and oil yield stability in NS rapeseed cultivars in changeable environmental conditions of northern Serbia. Seed yield, oil content and yield for 40 winter and 9 spring rapeseed cultivars were analyzed. It was found that the infl uence of year on seed and oil yield was most signifi cant. Genotype (cultivars) by environment (year) interaction was further analyzed using AMMI (Additive Main Effects and Multiplicative Interaction) model. In the environmental conditions present during the trial in Rimski Sancevi, according to the analysis of regression coeffi cient (bi) and ecovalence (Wi) and AMMI model, winter cultivars Nena and NS-L-102 were the most stable and high seed yielding, and hybrid cultivar NS-H-R-3 was singled out for oil yield. These cultivars are recommended for implementation in future breeding programs and for growing in the conditions described in this paper.Key words: oil yield, rank change interactions, seed yield, stability

GGE biplot analiza multilokacijskih ogleda NS hibrida kukuruza 77
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82originalni naučni rad / original research article
GGE biplot analiza multilokacijskihogleda NS hibrida kukuruza
Bojan Mitrović · Dušan Stanisavljević · Sanja Treskić · Milisav Stojaković · Goran Bekavac · Aleksandra Nastasić · Mile Ivanović
received / primljeno: 05.11.2010. accepted / prihvaćeno: 29.12.2010.© 2011 IFVC
Izvod: Multilokacijski ogledi za prinos zrna izvode se svake godine kako bi se ocenila reakcija hibri-da kukuruza na različite agroekološke uslove. Osnovni problem ovog načina ispitivanja hibrida jeste prisustvo interakcije genotip-spoljna sredina. Uzimajući u obzir interakciju, GGE biplot analiza pruža vizuelni prikaz ponašanja hibrida u različitim sredinama. Dvadeset hibrida kukuruza je ispitivano u toku 2006. i 2007. na pet lokaliteta po godini. Rezultati su predstavljeni preko GGE biplotova u cilju grupi-sanja sredina u tzv. megasredine i izdvajanja idealne sredine. Značaj izdvajanja megasredine ogleda se u odsustvu unakrsne interakcije koja remeti njihov rang, pa se hibridi mogu odabrati na osnovu prosečnih vrednosti određene osobine. Određivanje tzv. idealne sredine može biti od velikog značaja za izbor naj-reprezentativnijeg lokaliteta za testiranje, koji takođe ima i najveću moć diskriminacije hibrida.Ključne reči: GE interakcija, GGE biplot, hibrid, kukuruz, megasredina, prinos zrna
B. Mitrović · D. Stanisavljević · S. Treskić · M. Stojaković · G. Bekavac · A. Nastasić · M. IvanovićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Ispitivanje hibrida u različitim agroekološkim uslovima predstavlja jednu od najvažnijih faza u oplemenjivanju kukuruza. Na ovaj način dobijaju se veoma važne informacije o tome kako hibridi reaguju na različite uslove spoljne sredine. Me-đutim, izdvajanje najsuperiornijih hibrida (npr. za prinos zrna) znatno je otežano usled prisu-stva interakcije genotip-spoljna sredina (GEI). Interakcija genotip-spoljna sredina podrazume-va različitu reakciju genotipova, sorti ili hibrida na uslove sredine u kojima se gaje (Kang 1998, 2004). Prisustvo interakcije komplikuje proces selekcije i smanjuje međuzavisnost genotipskih i fenotipskih vrednosti, čime se usporava napre-dak selekcije (Comstock & Moll 1963). Razume-vanje uzroka interakcije (GEI) je veoma važno jer može doprineti određivanju cilja oplemenjivanja, identifi kaciji idealnih uslova testiranja i preporuci hibrida ili sorti koje se odlikuju boljom adaptabil-nošću (Yan et al. 2000).
Razvijene su brojne metode kojima je moguće proceniti interakciju genotip-spoljna sredina. Ove
metode mogu biti zasnovane na analizi varijanse, linearnoj regresiji, nelinearnoj analizi, multivarija-cionoj analizi, biplotima, kao i na metodama ne-parametrijske statistike (Balestre et al. 2009).
Analiza varijanse kao aditivni model objašnjava samo glavne efekte i pruža informacije da li je interakcija genotip-spoljna sredina značajan izvor varijacije. Međutim, ona ne pruža uvid u pojedi-načne genotipove i lokacije koji učestvuju u inte-rakciji (Samonte et al. 2005).
Jedan od modela multivarijacione analize koji se često koristi za analizu interakcije genotip-spoljna sredina je AMMI model (Additive Main Effects and Multiplicative Interactions) (Gauch & Zobel 1996). AMMI model kombinuje analizu varijanse za objašnjenje glavnih efekata genotipa i spoljne sredine i analizu glavnih komponenata za objašnjenje interakcije (GE). Za interpretaciju rezultata AMMI analize koristi se biplot koji stav-lja u odnos srednje vrednosti svojstva genotipova i prvu, odnosno neku od glavnih komponenata (PC) GE interakcije.
Detaljnije objašnjenje interakcije genotip-spolj-na sredina obezbeđuje vizuelni metod po nazivu GGE biplot (Yan et al. 2000) koji na neki način predstavlja modifi kaciju AMMI modela. Ovaj metod je zasnovan na dva koncepta, biplot kon-ceptu (Gabriel 1971) i GGE konceptu (Yan et al. 2000), a korišćen je za vizuelnu analizu podataka
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Mitrović B i sar.78
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
multilokacijskih ogleda. Ovaj metod koristi biplot za prikaz dva faktora, genotipa i interakcije geno-tip-spoljna sredina (G i GE), koji mogu biti od velikog značaja za evaluaciju hibrida kukuruza u različitim agroekološkim uslovima. GGE biplot prikazuje prve dve glavne komponente (PC1 i PC2) koje takođe predstavljaju dva osnovna izvora varijacije (Yan et al. 2000, 2001). Glavne komponente (PC1 i PC2) dobijaju su dekom-pozicijom singularnih vrednosti GGE podataka (Alwala et al. 2010).
Osnovni cilj ovog rada je da se uz pomoć me-toda GGE biplot analize geografski različiti lo-kaliteti koji pokazuju sličnost svrstaju u određe-ne grupe formiranjem takozvanih megasredina. Navedeni model pruža mogućnost da se izdvoji „idealni lokalitet“ tj. lokalitet koji ima najveću moć diskriminacije genotipova, koji je i najrepre-zentativniji.
Materijal i metod
U radu su korišćeni podaci iz dva jednogodiš-nja multilokacijska ogleda sa komercijalnim i po-tencijalno komercijalnim NS hibridima kukuruza FAO grupe zrenja 600-700. Za istraživanje u obe godine odabrano je 20 NS hibrida kukuruza. Oba ogleda posejana su po modelu slučajnog blok si-
stema sa tri ponavljanja, na pet lokaliteta: Rim-ski Šančevi (RS), Srbobran (SR), Pančevo (PA), Sombor (SO) i Sremska Mitrovica (SM). Površina elementarne parcele iznosila je 9,75 m2 za svaki genotip (dva reda, svaki dužine 6,5 m) sa rastoja-njem od 0,75 m između redova i 0,22 m između biljaka u redu. Gustina useva iznosila je 60.606 biljaka ha-1. Setva i berba ogleda obavljene su mašinski, pri čemu su od svakog genotipa sejana četiri reda, a brana i merena su dva središnja reda. Primenjena je uobičajena tehnologija gajenja koja je prilagođena agroekološkim uslovima lokaliteta. U radu su analizirani rezultati za prinos zrna (t ha-1 sa 14% vlage).
Za proučavanje interakcije hibrida kukuruza i lokaliteta odabrane su 2006. i 2007. kao godi-ne veoma različite po svojim klimatskim karak-teristikama. U 2006. klimatski pokazatelji su se uglavnom zadržali blizu višegodišnjeg proseka, mada je na skoro celoj teritoriji Srbije zabele-ženo pozitivno odstupanje srednjih godišnjih temperatura vazduha za 0,2ºC do 1ºC. Ukupne godišnje sume padavina bile su nešto veće od prosečnih količina, ali su ostale u opsegu pro-sečnih vrednosti. Godina 2007. je za razliku od prethodne bila jedna od najnepovoljnijih godi-na za proizvodnju kukuruza posle drugog svet-skog rata (Ivanović i sar. 2008). U letnjem peri-
Tabela 1. Prosečan prinos hibrida kukuruza u 2006. (t ha-1)Table 1. Average grain yield of maize hybrids in 2006 (t ha-1)
HibridiHybrids RS PA SM SO SR
ProsekAverage
1 13,40 12,93 10,00 10,88 11,25 11,692 12,03 12,00 9,70 10,36 11,72 11,163 12,60 12,50 9,72 13,40 12,55 12,154 12,16 11,85 11,03 9,11 12,25 11,285 12,79 13,10 10,16 14,39 13,62 12,816 13,41 13,58 10,02 12,33 14,16 12,707 12,05 10,36 10,68 12,69 12,16 11,598 13,17 12,28 9,87 13,48 14,43 12,659 11,75 12,69 8,03 14,08 13,33 11,9810 12,68 12,60 13,36 14,41 13,00 13,2111 12,42 11,57 11,16 12,37 11,71 11,8512 13,47 13,69 10,10 11,08 13,88 12,4413 12,72 13,60 7,74 13,34 13,69 12,2214 13,66 12,90 11,67 12,21 12,60 12,6115 12,07 11,14 9,11 11,43 12,26 11,2016 13,29 12,11 7,21 12,44 13,22 11,6517 10,38 7,96 9,49 10,90 8,63 9,4718 11,94 12,72 10,89 12,65 11,47 11,9319 13,08 11,24 11,15 13,09 13,65 12,4420 11,29 11,97 10,66 14,45 11,73 12,02
ProsekAverage 12,52 12,14 10,09 12,45 12,57 11,95
RS, PA, SM, SO, SR lokaliteti / locations

GGE biplot analiza multilokacijskih ogleda NS hibrida kukuruza 79
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
odu je zabeležen dvostruko veći broj tropskih dana tj. dana kada je maksimalna temperatura veća ili jednaka 30ºC, što se izuzetno nepovolj-no odrazilo na cvetanje i oplodnju kukuruza. Padavine su bile neravnomerno raspoređene tokom godine. U aprilu je bio izuzetno mali priliv padavina u odnosu na uobičajen. U ne-kim mestima nije uopšte bilo padavina tokom celog meseca, pa je suša ugrožavala većinu ra-tarskih kultura (Lalić 2008). Evidentne razlike u klimatskim pokazateljima između 2006. i 2007. odrazile su se i na prinos kukuruza. Op-šti prosek hibrida u ogledima 2006. bio je veći za više od 2,5 t ha-1 u odnosu na opšti prosek 2007. (Tab. 1 i 2).
Analiza varijanse izračunata je preko modela:
Yij = μ + Gi + Ej + GEij
gde je Yij odgovarajuća promenljiva i-tog genoti-pa na j-tom okruženju (lokalitetu); μ predstavlja opšti prosek; Gi je glavni efekat genotipa i; Ej je glavni efekat okruženja j; GEij je efekat interakci-je genotip-spoljna sredina.
GGE biplotovi (Yan et al. 2000) konstruisani su preko osnovnog modela:
Yij – μ – Ej = λ1εi1η1j + λ2εi2η2j + eij
gde je Yij odgovarajuća promenljiva i-tog genoti-pa na j-tom okruženju (lokalitetu); μ predstavlja opšti prosek; Ej je glavni efekat j-tog okruženja; λ1 i λ2 singularne vrednosti glavnih komponenti PC1 i PC2; εi1 i εi2 su eigenvektori (karakteristič-ne vrednosti) i-tog genotipa za PC1 i PC2; η1j i η2j su eigenvektori j-tog okruženja (lokaliteta) za PC1 i PC2; eij je opšti ostatak genotipa i i okruženja j.
Dekompozicija singularnih vrednosti u odgova-rajuće karakteristične vrednosti genotipa i okruže-nja može se predstaviti sledećom formulom:
gi1 = λ1 f lεi1 i e1j = λ1
1-f lη1j
Za konstruisanje GGE biplota vrednosti glav-nih komponenti za i-ti genotip i e-to okruženje iskazane su na sledeći način:
Yij – μ – Ej = gi1e1j + gi2e2j + eij
gde je Yij odgovarajuća promenljiva i-tog genoti-pa na j-tom okruženju (lokalitetu); μ predstavlja opšti prosek; gi1 i gi2 su PC vrednosti genotipa i; e1j i e2j su PC vrednosti okruženja j.
Tabela 2. Prosečan prinos hibrida kukuruza u 2007. (t ha-1)Table 2. Average grain yield of maize hybrids in 2007 (t ha-1)
HibridiHybrids RS PA SM SO SR
ProsekAverage
1 12,56 10,76 6,55 7,27 9,27 9,282 12,43 8,82 10,67 7,12 9,40 9,693 12,08 9,15 10,25 6,93 10,37 9,764 12,84 7,65 8,32 9,06 8,32 9,245 11,12 8,48 7,17 5,97 8,06 8,166 13,74 9,97 9,97 9,67 9,40 10,557 11,48 9,78 9,21 7,18 9,38 9,408 11,11 8,67 6,60 7,25 9,28 8,589 11,76 9,48 9,84 6,79 9,10 9,3910 12,37 10,39 11,24 6,81 9,85 10,1311 12,17 8,53 9,62 8,69 9,21 9,6412 11,78 7,94 11,51 7,80 8,98 9,6013 11,33 9,17 8,89 7,71 9,85 9,3914 10,51 7,47 8,76 8,99 9,55 9,0615 9,59 8,38 9,62 4,67 9,27 8,3116 12,13 10,44 10,65 6,71 10,32 10,0517 13,33 10,49 11,29 8,54 10,92 10,9118 11,92 7,93 10,29 7,19 7,72 9,0119 12,30 8,67 11,90 8,78 9,94 10,3220 13,72 9,97 10,99 8,93 7,93 10,31
ProsekAverage 11,88 9,11 9,67 7,60 9,31 9,54
RS, PA, SM, SO, SR lokaliteti / locations

Mitrović B i sar.80
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
U ovom radu su svi biplotovi generisani meto-dom skaliranja prema okruženju „enviroment focused scalling“. Analiza podataka urađena je u programu GGEbiplot Version 6.3. koji je u punoj verzi-ji dobijen ljubaznošću dr Weikai Yan, saradnika Univerziteta u Guelfu, Ontario, Kanada.
Rezultati i diskusija Iz tabele analize varijanse (Tab. 3) uočava se
da su sredine kvadrata oba faktora (hibrida i lo-kaliteta) kao i njihove interakcije bile značajne (P<0,01) za prinos zrna kukuruza u obe po-smatrane godine. Zajednički efekat genotipa i interakcije (G+GE) dalje je razložen metodom GGE biplot analize pri čemu su prve dve glavne komponente bile značajne objašnjavajući 71,2% (46,6% PC1 i 24,6% PC2) u 2006. godini, odno-sno 70,9% (45,8% PC1 i 25,2% PC2) GGE sume kvadrata u 2007. godini (Sl. 1 i 2).
Za multilokacijske oglede je uobičajena pojava unakrsnog (crossover) i ne unakrsnog (non-crossover) tipa interakcije (GE). U ovom radu različiti hibridi postizali su najviše prinose u različitim lokaliteti-ma. Tako je, na primer, u 2006. hibrid 14 postigao najviši prinos na Rimskim Šančevima, hibrid 12 u Pančevu, a hibrid 8 u Srbobranu (Tab. 1). U 2007. je hibrid 6 bio najprinosniji na Rimskim Šančevi-ma, hibrid 1 u Pančevu, a hibrid 17 u Srbobranu (Tab. 2). Ovakvi različiti rangovi hibrida ukazuju na prisustvo unakrsne (crossover) interakcije.
Grafi čki prikaz tzv. „gde je ko najbolji“ („which-one-where“) modela na osnovu podataka multilo-
kacijskih ogleda važan je za izdvajanje mogućih megasredina u regionu (Gauch & Zobel 1997, Yan et al. 2000, 2001). Genotipovi koji su naj-udaljeniji od koordinatnog početka povezani su pravim linijama obrazujući na taj način mnogo-ugao. Linije koje polaze iz koordinatnog početka i normalne su na stranice mnogougla dele isti na određeni broj sektora. Lokaliteti koji se nalaze unutar jednog sektora jesu lokaliteti u kojima je dati genotip postigao najviši prinos i mogu se smatrati megasredinama za taj genotip. Na slici 1. mogu se izdvojiti dve grupe sredina. Jednu grupu čine lokaliteti: Rimski Šančevi, Pančevo i Srbo-bran, dok u drugu grupu spadaju Sremska Mi-trovica i Sombor. Primećeno je slično grupisanje i u ranijim istraživanjima. Ivanović i sar. (2008) ukazuju na veliku sličnost po pitanju performansi hibrida u okruženjima Rimski Šančevi i Panče-vo. Ovo grupisanje odnosi se na 2006. koja se po svojim meteorološko-klimatološkim karakteristi-kama može svrstati u prosečnu godinu. Potpuno drugačije grupisanje karakteriše 2007. koja je bila veoma sušna i izrazito nepovoljna za gajenje ku-kuruza. Na slici 2. sredine Rimski Šančevi i Som-bor pripadaju jednoj grupi, Pančevo i Sremska Mitrovica drugoj, dok Srbobran nije svrstan ni u jednu grupu. Ovakve razlike u grupisanju sredina između 2006. i 2007. mogu se objasniti velikom razlikom u klimatskim uslovima koja je uslovila drugačiju interakciju. Za preciznije podatke o postojanju megasredina potrebna su višegodišnja ispitivanja hibrida kukuruza u multilokacijskim ogledima (Yan et al. 2000).
Tabela 3. Analiza varijanse prinosa hibrida kukuruza u 2006. i 2007. Table 3. Analysis of variance of maize hybrids grain yield in 2006 and 2007
1 genotip/genotype ** p<0.012 sredina/environment
GodinaYear
Izvor varijacijeSource of variation df SS MS F %SS
G1 19 187.01 9.84 9.58** 19.5E2 4 267.85 66.96 65.2** 27.9
2006 GxE 76 285.07 3.75 3.65** 29.8ponavljanje/replications 2 14.17 7.09 6.9** 1.5
ostatak/residue 198 203.36 1.03 - 21.3total 299 957.48 - - 100G1 19 172.65 9.09 5.67** 12.9E2 4 623.08 155.77 97.23** 46.5
2007 GxE 76 224.01 2.95 1.84** 16.8ponavljanje/replications 2 1.51 0.75 0.47 0.1
ostatak/residue 198 317.19 1.60 - 23.7total 299 1338.43 - - 100

GGE biplot analiza multilokacijskih ogleda NS hibrida kukuruza 81
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
GGE biplot analiza pruža mogućnost izdva-janja tzv. „idealnih“ sredina. Idealnom sredinom se smatra sredina koja ima visoku PC1 vrednost, odnosno najveću moć diskriminacije genotipo-va i nisku PC2 vrednost, odnosno najreprezen-tativnije je od svih sredina. Iako ovakva sredina najčešće ne postoji u realnosti, ona može poslu-žiti za vrednovanje ostalih sredina. Što je sre-dina bliža „idealnoj“ to je poželjnije. Da bi se olakšala ocena sredine u odnosu na „idealnu“, tačka u kome se ona nalazi predstavlja centar iz koga polaze koncentrični krugovi (Yan et al. 2000). U 2006. godini najbliža „idealnoj“, a ujedno i najpoželjnija sredina je bila Sombor, a
zatim Pančevo, Srbobran, Rimski Šančevi. Naj-udaljenija, a samim tim najnepoželjnija sredina bila je Sremska Mitrovica (Sl. 3). Na slici 4. koja predstavlja 2007. vidljive su razlike u odnosu na prethodnu godinu. Kao najpoželjnije okru-ženje mogu se izdvojiti Rimski Šančevi, zatim Sremska Mitrovica, Sombor, Pančevo, dok je najnepoželjnije okruženje Srbobran. Ovakva različitost između rezultata u 2006. i 2007. nije iznenađujuća, jer se radi o dve godine izuzetno različite po klimatskim karakteristikama, kao što je opisano ranije.
Slika 1. GGE biplot sa prikazom megasredina u 2006. Fig. 1. GGE biplot showing the megaenviro-ments in 2006
Slika 2. GGE biplot sa prikazom megasredina u 2007. Fig. 2. GGE biplot showing the megaenviroments in 2007
Slika 3. GGE biplot sa prikazom poređenja okruženja sa „idealnim“ okruženjem u 2006. Fig. 3. GGE biplot showing comparison of the environments to ideal environment in 2006
Slika 4. GGE biplot sa prikazom poređenja okruženja sa „idealnim“ okruženjem u 2007. Fig. 4. GGE biplot showing comparison of the environments to ideal environment in 2007

Mitrović B i sar.82
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 77-82
Zaključak
GGE biplot analiza je metoda koja se koristi za identifi kaciju megasredina, tj. grupisanje, razdvaja-nje lokaliteta koji se ispituju u zajedničke megasre-dine, a potom evaluaciju genotipova u okviru tih megasredina i odabir najdiskriminativnije i najrepre-zentativnije lokacije u okviru jedne megasredine.
U ovom radu analizirano je dvadeset hibrida kukuruza na pet lokaliteta u toku 2006. i 2007. Analizom megasredina u prvoj godini veća slič-nost javila se između lokaliteta Rimski Šančevi, Pančevo i Srbobran tako da su oni svrstani u jednu grupu, potencijalnu megasredinu, dok su Sremska Mitrovica i Sombor svrstani u drugu grupu. U 2007. je grupisanje bilo drugačije tako da su jednu megasredinu činili Rimski Šančevi i Sombor, dru-gu Sremska Mitrovica i Pančevo dok se Srbobran izdvojio iz obe grupe. Nepodudarnost obrazova-nja mega-sredina može se objasniti različitim kli-matskim uslovima u dve posmatrane godine, što je potvrđeno i analizom idealne sredine gde su se ta-kođe javile nepodudarnosti između dve posmatra-ne godine. Dobijeni rezultati u ovom radu ukazuju da se pouzdaniji podaci o formiranju megasredina mogu dobiti višegodišnjim proučavanjem odgova-rajućih genotipova na većem broju lokaliteta.
Literatura
Alwala S, Kwolek T, McPferson M, Pellov J, Meyer D (2010): A comprehensive comparison between Eberhart and Russell
joint regression and GGE biplot analysis to identify stable and high yielding maize hybrids. Field Crops Res.199: 225-230
Balestre M, Candido de Souza J, Garcia von Pinho R, Lunzzo de Oliveira R, Muro Valente Paes J (2009): Yield stability and adaptability of maize hybrids based on GGE biplot analysis characteristics. Crop Breed. Appl. Biotechnol. 9: 219-228
Comstock R E, Moll R H (1963): Genotype-environment in-teraction. In: Statistical genetics and plant breeding. NAS NRC, 982, 164-196
Gabriel K R (1971): The biplot graphic display of matrices with applications to principal component analysis. Biometrics 58: 453-467
Gauch H G, Zobel R W (1996): AMMI analysis of yield trials. In: Kang MS, Gouch HG (eds.), Genotype by environment interaction. CRC Press, Boca Raton, FL, 85-122
Gauch H G, Zobel R W (1997): Interpreting mega-envrio-nments and targeting genotypes. Crop Sci. 37: 311-326
Lalić B (2008): Agrometeorološka analiza za period januar-jun 2007. godine u Vojvodini [Elektronski izvor]. [5 str] dostu-pno na adresi
cmep-serbia.df.pmf.uns.ac.rs/agrmet/analiza/Agm_an_300607.pdf
Ivanović M, Čapelja V, Radojčić S, Popov R, Nastasić A (2008): Kukuruz u 2007. godini: pouke sušne godine. Zbornik rado-va Instituta za ratarstvo i povrtarstvo, Novi Sad 45: 61-65
Kang M S (1998): Using genotype-by-environment interaction for crop cultivar development. Adv. Agron. 62: 199-252
Kang M S (2004): Genotype-by-environment interaction. Encyc-lopedia of Plant and Crop Science, Macel Dekker Inc, 218-221
Samonte S O P B, Wilson L T, McClung A M, Medley J C (2005): Targeting cultivars onto rize growing environments using AMMI and SREG GGE biplot analysis. Crop Sci. 45: 2414-2424
Yan W, Corneluis P L, Crossa J, Hunt L A (2001): Two types of GGE biplots for analyzing multi-envorinment trial data. Crop Sci. 41: 656-663
Yan W, Hunt L A, Sheng Q, Szlavnics Z (2000): Cultivar eva-luation and mega-environment investigation based on the GGE biplot. Crop Sci. 40: 597-605
GGE Biplot Analysis of Multi-Environment Trials of NS Maize Hybrids
Bojan Mitrović · Dušan Stanisavljević · Sanja Treskić · Milisav Stojaković · Goran Bekavac · Aleksandra Nastasić · Mile Ivanović
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: GGE biplot analysis is a method used to identify mega-environments (i.e. grouping, division of locati-ons, which are examined in a joint mega-environment), evaluate the genotypes within these mega-environments and select the most discriminating and the most representative locations within a single mega-environment. This paper analyses twenty hybrids at fi ve locations in 2006 and 2007. For the fi rst year, mega-environment analysis showed a greater similarity between locations Rimski Šančevi, Pančevo and Srbobran classifying them into one group (the potential mega-environment), whereas Sombor and Sremska Mitrovica were classifi ed into another group. In 2007, grouping was different from the previous in that Rimski Šančevi and Sombor comprised one mega-envi-ronment, whereas Sremska Mitrovica and Pančevo comprised the other, while Srbobran was separated from both groups. Discrepancies in formation of the mega-environments can be explained by different climatic conditions in two research years. This was confi rmed by analyzing the ideal environment where the discrepancies between those two years are also reported. The results in this study suggest that more reliable data on the mega-environments formation should be sought in multi-year studies of the certain genotypes in a number of locations.Keywords: GE interaction, GGE biplot, grain yield, hybrid, maize, mega-environment

Nove hibridne sorte šećerne repe otporne prema rizomaniji 83
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 83-86
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 83-86originalni naučni rad / original research article
Nove hibridne sorte šećerne repeInstituta za ratarstvo
i povrtarstvo otporne prema rizomaniji
Lazar Kovačev · Nikola Čačić · Snežana Mezei
primljeno / received: 20.10.2010. accepted / prihvaćeno: 08.12.2010.© 2011 IFVC
Izvod: Rizomanija je jedno od najdestruktivnijih obolenja koje je do sada identifi kovano i opisano na šećernoj repi. Obolenje je vrlo kompleksnog karaktera, jer rizomaniju izaziva virus nekrotičnog žutila nerava šećerne repe (VNŽNŠR), a prenosilac virusa je parazitna gljiva Polymyxa betae. Smanjenje prinosa korena i sadržaja šećera kod osetljivih hibrida na zaraženom zemljištu može biti i preko 80%. U ovom momentu najefi kasnija i ekonomski najprihvatljivija mera suzbijanja je stvaranje i gajenje tolerantnih ili otpornih hibridnih sorti. Koristeći konvencionalne metode oplemenjivanja i različite izvore otpornosti prema ovom obolenju u Institutu za ratarstvo i povrtarstvo u 2009. godini priznate su dve diploidne hibridne sorte, Nora i Vera. Obe hibridne sorte se po kvantitativnim i genetičkim osobinama razlikuju, a zbog postojanja dva izvora i tipa otpornosti – tolerantnosti mogu se gajiti i na zemljištima sa vrlo viso-kim nivoom inokuluma. Obe hibridne sorte poseduju visoki genetički potencijal rodnosti za najvažnije proizvodne osobine. Ključne reči: hibridne sorte, kvantitativna svojstva, otpornost, rizomanija, tolerantnost
L. Kovačev · N. Čačić · S. MezeiInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Na povećanje prinosa u određenim agroekološ-kim uslovima direktno utiču dve grupe faktora: sorta (genotip) i tehnologija proizvodnje. Značaj sorte kao jednog od najcelishodnijeg i najekono-mičnijeg načina povećanja prinosa davno je poznat i dokazan (Kovačev i sar. 1998). Rizomanija, koju izaziva virus nekrotičnog žutila nerava šećerne repe (VNŽNŠR), kod šećerne repe je najdestruktivnije obolenje koje je do sada opisano, te je gajenje tole-rantnih ili otpornih sorti jedino ekonomski prihvat-ljivo rešenje koje još više ističe ulogu genotipa kao presudnog faktora visoke i stabilne proizvodnje.
Kao potvrda kompleksnosti i destruktivnosti rizomanije može da posluži i činjenica da je od momenta identifi kacije i opisa obolenja pa do stvaranja prve visokotolerantne hibridne sorte prošlo više od trideset godina rada velikog broja istraživača genetičara, oplemenjivača, fi topatolo-ga, virologa itd. (De Biaggi 2005).
Identifi kacija i stvaranje otpornosti prema ri-zomaniji u Odeljenju za šećenu repu Instituta za ratarstvo i povrtarstvo u Novom Sadu počelo je u ranim osamdesetim godinama prošlog veka, a
prva tolerantna sorta prema rizomaniji priznata je 2000. godine (Kovačev i sar. 2001, 2003).
Rezultat višegodišnjeg intenzivnog, kontinuira-nog i vrlo komplikovanog rada na stvaranju otpor-nih hibridnih sorti šećerne repe prema rizomaniji je priznavanje osam sorti od strane Odeljenja za priznavanje sorti Ministarstva poljoprivrede, šu-marstva i vodoprivrede Republike Srbije.
Cilj ovog rada je da se prikažu kvantitativna svojstva i prednosti novopriznatih otpornih sorti šećerne repe (Nora i Vera) u odnosu na standard hibridnu sortu Rama koja je u godinama ispitiva-nja bila najraširenija otporna sorta u Evropi.
Materijal i metod
U radu su korišteni rezultati ogleda Komisije za priznavanje sorti Republike Srbije sprovedenih tokom 2007. i 2008. Hibridne sorte Nora i Vera testirane su na lokalitetima Sombor, Kikinda, Novi Sad, Sremska Mitrovica i Pančevo. Kod obe sorte standard je bila sorta Rama. Ogledi su bili postav-ljeni po slučajnom blok sistemu u četiri ponavlja-nja, a veličina osnovne parcelice za obračun bila je 9,6 m2. Određivanje najvažnijih kvantitativih svojstava (prinos korena, sadržaj šećera i prinos šećera) koja će u radu biti predstavljena urađena je uobičajenim metodama za ispitivanje šećerne repe. Statistička obrada podataka obavljena je analizom varijanse pomoću MSTATC programa.
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Kovačev L i sar.84
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 83-86
Rezultati i diskusija
Poreklo hibridnih sortiMonogermna diploidna (2n=2x=18 hromozo-
ma) hibridna sorta Nora nastala je ukrštanjem ci-toplazmatsko nuklearno muške sterilne linije NS-MS-071R sa multigermnom diploidnom popula-cijom uske genetičke osnove NS-2nMM071R.
Muško sterilna majčinska komponenta i njen fertilni analog (održivač citoplazmatsko nuklearne muške sterilnosti «o» tip) dobijena je konvergen-tnim oplemenjivanjem ukrštanjem između komer-cijalne monogermne linije NS 6 sa monogermnim održivačem citoplazmatsko nuklearne muške ste-rilnosti koji nije imao komercijalnu vrednost, ali je posedovao «Holly» gen otpornosti prema rizo-maniji. Holi gen otpornosti vodi poreklo iz SAD (Lewellen 1987, 1995). Da bi se zadržala dobra kombinaciona sposobnost i komercijalna vred-nost linije izvršena su četiri povratna ukrštanja sa rekurentnim roditeljem koristeći boju hipokotila i nuklearnu mušku sterilnost kao marker gene. To-kom povratnih ukrštanja kontinuirano je ispitiva-no prisustvo holi gena otpornosti prema rizoma-niji, gajenjem genotipova na zemljištu sa visokim nivoom inokuluma. U svakom ciklusu su vršena povratna ukrštanja sa citoplazmatsko–nuklearno muško sterilnim izvorom u cilju dobijanja steril-nog analoga. Multigermna diploidna populacija 2nMM071R poseduje rizor gene tolerantnosti prema rizomaniji. Nastala je prostim ukrštanjem domaće multigermne diploidne populacije sa populacijom koja je posedovala rizor gene tole-rantnosti. Na populaciji su urađena tri ciklusa po-vratne selekcije na otpornost prema rizomaniji i specifi čnu kombinacionu sposobnost.
Hibridna sorta šećerne repe Vera pripada še-ćernatom (Z) tipu, a nastala je ukrštanjem mono-germne diploidne muške sterilne linije NS-88 MS i multigermne diploidne populacije NS2nMM MTRBrek.
Majčinska komponenta sorte Vera, monoger-mna diploidna muškosterilna linija NS-88 MS na-stala je iz monogermnog materijala šećerne repe poreklom iz Nemačke. U toku stvaranja ove linije (sterilne linije i njenog fertilnog analoga) procesom samooplodnje fi ksirani su u homozigotnom stanju recesivni x i z geni koji kontrolišu mušku sterilnost i recesivni m gen koji kontroliše monogermnost. Primenom metoda povratnog ukrštanja (back-cross method) u ovu liniju je unet Holly gen koji kontroliše otpornost prema rizomaniji. Monogermna lini-ja NS-88 MS se odlikuje dobrim kombinacionim sposobnostima na sadržaj šećera.
Očeva komponenta sorte Vera, multigermna dioploidna populacija NS-2nMM MTRBrek, na-
stala je metodom rekurentne selekcije na posebne kombinacione sposobnosti iz očeve komponente sorte Lara, a kao tester poslužila je monogermna linija NS-88 MS. U toku procesa stvaranja kori-šćena je i vegetativna propagacija (kultura tkiva), kako bi se svaki genotip zadržao u nepromenje-nom stanju do utvrđivanja njegovih kombinaci-onih sposobnosti. Nakon utvrđivanja kombina-cionih sposobnosti od superiornih genotipova formirana je populacija NS-2nMM MTRBrek. Ova populacija poseduje rizor gene tolerantno-sti (otpornosti) prema rizomaniji i veoma dobre konbinacione sposobnosti za sadržaj šećera i teh-nološke karakteristike.
Prinos korena Pored sadržaja šećera, prinos korena je osnovni
pokazatelj vrednosti hibridnih sorti šećerne repe. U dvogodišnjim sortnim ispitivanjima na pet loka-liteta državne Sortne komisije hibridna sorta Nora imala je skoro identičan prinos korena u odnosu na standardnu sortu Rama (Tab. 1). Posmatrajući po lokalitetima, utvrđene su kako visoko značaj-no više, a takođe i značajno niže vrednosti prinosa korena u odnosu na standard. Koefi cijent varijaci-je je iznosio 6,16 % što ukazuje da su ogledi vrlo kvalitetno izvedeni. Što se tiče hibridne sorte Vera, ona je po prinosu korena bila lošija od standarda, ali razlika nije bila statistički opravdana. I pored toga što po prinosu korena statistički nisu preva-zišle standard ukazuje na to da prinos korena sam za sebe nije i najvažniji pokazatelj produktivnosti određenog genotipa (Kovačev i sar. 2003 ).
Sadržaj šećeraOsnovni pokazatelj tehnološke vrednosti odre-
đenog genotipa je sadržaj šećera. Pošto je sadržaj šećera tipično kvantitativno svojstvo na njegovo ispoljavanje utiče čitav niz faktora, a presudnu ulogu ima genotip. Obe hibridne sorte u svim lo-kalitetima i u proseku imale su visoko značajno viši sadržaj šećera u odnosu na standard Rama (Tab. 2). Pošto se u Srbiji šećerna repa plaća u zavisnosti od sadržaja šećera, ovakve hibridne sorte šećerne repe sa naglašenim sadržajem še-ćera interesantne su kako za proizvođače tako i za prerađivače.
Prinos kristalnog šećeraOstvareni prinos kristalnog šećera po jedinici
površine predstavlja osnovni cilj gajenja šećerne repe i glavni je pokazatelj ekonomske vrednosti gajenog genotipa. Obe ispitivane hibridne sorte še-ćerne repe u proseku za dve godine i na pet repre-zentativnih lokaliteta po prinosu kristalnog šećera (Tab. 3) bile su bolje od standarda i to za 400 kg

Nove hibridne sorte šećerne repe otporne prema rizomaniji 85
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 83-86
ha-1 (sorta Nora) tj. 200 kg ha-1 (sorta Vera). Upra-vo ova činjenica ukazuje da hibridne sorte šećerne repe Nora i Vera imaju izbalansiran odnos između prinosa korena i sadržaja šećera, te da imaju dobru adaptabilnost i stabilnost prinosa pošto su ispitiva-ne na pet različitih agroekoloških lokaliteta.
Pored dobrih kvantitativnih svojstava, hibridne sorte šećerne repe Nora i Vera imaju visoku toleran-tnost prema obolenjima lista kao što su pegavost lista koju izaziva Cercospora beticola Sacc. zonalnoj pegavosti koju izaziva Phoma betae Frank i sivoj pegavosti lista koju izaziva Ramularia beticola Faurt & Lamb.
Tabela 1. Prinos korena hibridnih sorti šećerne repe Nora i Vera u odnosu na standard Rama u ogledima komisije za priznavanje sorti Republike SrbijeTable 1. Root yield of hybrid varieties of sugar beet Nora and Vera compared with standard variety Rama in the trials of Varietal Release Committee of the Republic of Serbia
LokalitetLocation
Prinos korena / Root yield (t ha-1)
Nora Vera RamaKikinda 76,9 74,3 76,0Novi Sad 79,5++ 72,1 72,3Pančevo 74,2- 70,8-- 79,7Sremska Mitrovica 69,9- 76,1 75,1Sombor 76,8 74,9 78,0X 76,2 73,7 76,3
0,01 3.95 LSD
0,05 3.21 CV% 6.16
Tabela 2. Sadržaj šećera hibridnih sorti šećerne repe Nora i Vera u odnosu na standard Rama u ogledima komisije za priznavanje sorti Republike SrbijeTable 2. Sugar content of hybrid varieties of sugar beet Nora and Vera compared with standard variety Rama in the trials of Varietal Release Committee of the Republic of Serbia
LokalitetLocation
Sadržaj šećera / Sugar content (%)
Nora Vera RamaKikinda 16,2++ 16,7++ 15,2Novi Sad 16,1++ 16,4++ 15,1Pančevo 16,9++ 16,4++ 15,6Sremska Mitrovica 16,9++ 15,8++ 15,9Sombor 15,4+ 16,0++ 15,1X 16,0++ 16,3++ 15,4
0,01 0,27 LSD
0,05 0,31 CV% 2,1
Tabela 3. Prinos kristalnog šećera hibridnih sorti šećerne repe Nora i Vera u odnosu na standard Rama u ogledima komisije za priznavanje sorti Republike SrbijeTable 3. Sugar yield of hybrid varieties of sugar beet Nora and Vera compared with standard variety Rama in the trials of Varietal Release Committee of the Republic of Serbia
LokalitetLocation
Prinos kristalnog šećera / Sugar yield (t ha-1)
Nora Vera RamaKikinda 10,7 10,6 9,6Novi Sad 11,5 10,6 9,8Pančevo 10,0 10,0 10,7Sremska Mitrovica 10,2 10,3 11,0Sombor 10,4 10,5 10,3X 10,6 10,4 10,2

Kovačev L i sar.86
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 83-86
Zaključci
Na osnovu rezultata dvogodišnjih ispitivanja na pet lokaliteta koja su izvedena u organizaciji Odseka za priznavanje sorti poljoprivrednog bilja Ministarstva poljoprivrede, šumarstva i vodopri-vrede Republike Srbije može se zaključiti slede-će:
U prinosu korena hibridne sorte Nora i Vera - bile su statistički na nivou standardaObe hibridne sorte u sadržaju šećera bile su - statistički vrlo značajno bolje u proseku za dve godine istraživanjaŠto se prinosa kristalnog šećera tiče, hibridna - sorta Vera bila je bolja od standarda za 200 kg ha-1, a hibridna sorta Nora za 400 kg ha-1
S obzirom da obe hibridne sorte imaju gene-tičku otpornost prema rizomaniji i dobre tehno-loške karakteristike, one se preporučuju za rane i srednje rokove vađenja u svim rejonima gajenja šećerne repe u Republici Srbiji.
Literatura
De Biaggi M (2005): Resistance to Parasites, Rhizomania In: Genetics and breeding of sugar beet, 80-85
Kovačev L, Čačić N, Mezei S, Dokić P, Sklenar P ( 1998): Forty years of sugar beet breeding at the Institute of fi eld and veg-etable crops in Novi Sad, Proc 2nd Balkan Symposium Field Crops, Novi Sad, 1: 123-127
Kovačev L, Čačić N, Sklenar P, Mezei S (2001): Karakteristike novosadskih hibridnih sorti šećerne repe priznatih u 2000-oj godini. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 35: 271-278
Kovačev L, Mezei S, Čačić N, Sklenar P, Nagl N (2003): Rizo-manija – novija saznanja i proizvodna svojstva NS hibridnih sorti šećerne repe tolerantnih prema ovom obolenju. Zbor-nik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 38: 281-290
Lewellen R T (1987): Breeding sugarbeet for resistance to rhi-zomania: evaluation of host plant reactions and selection for and inheritance of ressistance, Proc. 50th IIRB winter con-gress, Bruxelles, Palias des Congress, 139-156
Lewellen R T (1995): Performance of near-isolines of sugar-beet with resistance to
rhizomania from different sources. Proc 58th IIRB Congress, 83-89
New Sugar Beet Hybrid Varieties Developed at Institute of Fieldand Vegetable Crops Resistant to Rhizomania
Lazar Kovačev · Nikola Čačić · Snežana Mezei
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Rhizomania is one of the most destructive sugar beet diseases that has been found and described so far. The disease has a complex character - it is caused by beet necrotic yellow veins virus (BNYVV) transmitted by a parasitic fungus Polymyxa betae. When susceptible hybrids are grown on infected soil, their root yield and sugar con-tent can reduce by over 80%. Presently, the most effi cient and economically most acceptable measure of controlling the disease is to develop and cultivate tolerant or resistant hybrid varieties. Using conventional breeding methods and different sources of disease resistance, breeders of Institute of Field and Vegetable Crops have developed two diploid hybrid varieties of sugar beet (Nora and Vera) which were released by Varietal Release Committee of the Republic of Serbia in 2009. These varieties differ in quantitative genetic traits and depending on the source and type of resistance/tolerance, they can be cultivated on soils containing a very high level of rhizomania inoculum. Both hybrid cultivars have high genetic potentials for most important production traits.Key words: hybrid varieties, rhizomania, resistance, sugar beet

Površina korena kao kriterijum za selekciju šećerne repe 87
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 87-90
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 87-90originalni naučni rad / original research article
Površina korena kao kriterijumza selekciju šećerne repe
Dario Danojević · Živko Ćurčić · Nevena Nagl · Lazar Kovačev
received / primljeno: 18.10.2010. accepted / prihvaćeno: 08.11.2010.© 2011 IFVC
Izvod: U ovom istraživanju je analizirano 10 monogermnih i 10 multigermnih genotipova šećerne repe koji se nalaze u kolekciji Instituta za ratarstvo i povrtarstvo u Novom Sadu. Ispitivana je površina korena šećerne repe kao svojstvo koje ima uticaj na iznošenje zemljišta pri vađenju, kao i pranju korena tokom procesa prerade. Primenjeno je neparametarsko testiranje sa Kruskal-Wallisovim testom i analiza grupisanja kao metod multivarijacione analize. Na osnovu dobijenih rezultata, najmanju ocenu površine korena (najmanje brazdi na korenu) imao je genotip 12 koji će se koristiti za poboljšanje ovog svojstva kod drugih genotipova.Ključne reči: multivarijaciona analiza, površina korena, selekcija, šećerna repa
D. Danojević · Ž. Ćurčić · N. Nagl · L. KovačevInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Transport i odlaganje zemljišta su neizbežni procesi pri preradi korena šećerne repe, ali i ne-poželjni s obzirom na velike troškove pri mani-pulaciji i prevozu šećerne repe do šećerana. Koli-čina zemljišta koja ostaje na korenu šećerne repe prilikom prevoza do šećerana prvenstveno zavisi od tipa i vlažnosti zemljišta (Koch 2002), mada i površina korena igra veoma važnu ulogu. Zemlji-šte koje ostaje na korenu nalazi se u brazdama ko-rena, kao i na manje glatkim površinama (Gram & Joergensen 2002). Koren repe koji ima duboke brazde teže je očistiti od čestica zemljišta kako u polju tako i prilikom prerade, naročito ako je repa gajena na težim zemljištima (Mesken 1987). Koren glatke površine se stoga smatra poželjnom osobinom kod šećerne repe zato što onemogu-ćava lepljenje čestica zemljišta za površinu ko-rena pri vađenju, što rezultira u nižem sadržaju nečistoća, nižim troškovima prevoza i smanje-nom širenju zemljišnih patogena (Theurer 1993). Glatki genotipovi (smooth root) nastali ukrštanjem šećerne repe i cvekle mogu smanjiti iznošenje zemljišta i do 50% zavisno od vrste i vlažnosti zemljišta (Panella & Lewellen 2007). Ocenjivanje korena na osnovu izgleda površine je u korelaciji sa količinom zemljišta koje je zaostalo na površi-
ni korena šećerne repe nakon vađenja (Gram & Joergensen 2002). U poslednjih deset godina je postignut značajan napredak u oplemenjivanju na ovo svojstvo, tako da je registrovano već nekoli-ko linija koje imaju gladak koren (Saunders et al. 1999, 2000a, 2000b, McGrath 2003, McGrath & Lewellen 2004).
Cilj ovog istraživanja je bio da se na delu selek-cionog materijala različitog porekla ustanovi po-stojanje varijabilnosti za površinu korena šećerne repe i izdvoje genotipovi koji se odlikuju glatkim korenom u cilju korištenja za dalji oplemenjivački rad.
Materijal i metod
Ispitivano je 10 monogermnih (mm) inbred li-nija - održivača citoplazmatske muške sterilnosti (genotipovi 1-10) i 10 multigermnih (MM) popu-lacija uske genetičke osnove i različitog porekla (genotipovi 11-20).
Ogled je izveden tokom 2008. na oglednim poljima Instituta za ratarstvo i povrtarstvo (Rim-ski Šančevi), Novi Sad. Genotipovi su posejani u prvoj dekadi marta po slučajnom blok sistemu
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Ovo istraživanje je deo projekta broj TR 20020: Poboljšanje germplaz-me šećerne repe u cilju povećanja prinosa i smanjenja gubitaka nastalih uticajem biotskih i abiotskih faktora (01. 04. 2008- 31. 12. 2010) Mini-starstva za nauku i tehnološki razvoj Republike Srbije / This research results from project 20020: „Improvement of sugar beet germplasm with aim to increase yield and reduce losses caused by effect of bio-tic and abiotic factors“ (01. 04. 2008- 31. 12. 2010) fi nanced by the Ministry of Science and Technological Development of the Republic of Serbia

Danojević D i sar.88
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 87-90
u tri ponavljanja. Tokom vegetacije su prime-njene redovne agrotehničke mere za šećernu repu. Veličina osnovne parcele je iznosila 16 m2. Razmak između biljaka je nakon ručnog proređivanja iznosio 50 x 20 cm. Da bi se uma-njilo oštećenje korena, vađenje je vršeno ručno u oktobru. Osnovni uzorak se sastojao od 10 biljaka po ponavljanju. Površina korena je vi-zuelno ocenjena od 1-9 (1-gladak, 9-izbrazdan koren) prema deskriptoru za rod Beta (IBPGR/CGN 1991).
Pomoću programa Statistica for Windows ver. 8, StatSoft. Inc., izvršeno je neparametarsko te-stiranje sa Kruskal-Wallisovim testom. Izvršena je analiza grupisanja, kao metod multivarijacione analize, koristeći Euklidske distance odstojanjа.
Rezultati i diskusija
Za svih 20 genotipova srednje vrednosti - me-dijane kretale su se u intervalu od 2 do 9. Na osnovu Kruskal-Wallisovog testa ustanovljene su značajne razlike između pojedinih genotipova. Najmanju srednju vrednost 2 imao je genotip 12, kod kojeg su pojedini korenovi imali ocene od 1 do 4 (Graf. 1), (Sl. 1). Pored genotipova 1 i 3 sa srednjom vrednošću ocene 5 se odlikuje genotip 4 (Slika 2). Najveću srednju vrednost 9 za povr-šinu korena imao je genotip 17, kod kojeg nije ustanovljeno variranje za ovo ispitivano svojstvo (Sl. 3). Najveća varijabilnost ocene za površinu korena je zabeležena kod dva multigermna geno-tipa 11 i 14 čije su se ocene kretale u intervalu
Grafi kon 1. Medijane, min. i maks. za površinu korena šećerne repeGraph 1. Medians, min. and max values for sugar beet root surface
Slika 1. Genotip 12 Slika 2. Genotip 4 Fig. 1. Genotype 12 Fig. 2. Genotype 4

Površina korena kao kriterijum za selekciju šećerne repe 89
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 87-90
od 2 do 7. U poređenju sa monogermnim, mul-tigermni genotipovi su iskazali veće variranje za ovo ispitivano svojstvo. Pošto većina roditeljskih linija nisu duboko u inbredingu, a posebno linije oprašivači (populacije), genetička varijabilnost se više pojavljuje kod hibrida šećerne repe nego kod hibridnog kukuruza, ili samooplodnih biljaka kao što su pšenica i soja (Panella & Lewellen 2007).
Pored mase korena i sadržaja šećera kao najzna-čajnijih svojstava za koje se vrši selekcija matičnih
korenova, u okviru programa oplemenjivanja še-ćerne repe u Institutu za ratarstvo i povrtarstvo se odabiraju oni korenovi koji su manje izbrazdani, tj. imaju gladak koren. Selekcijom je relativno lako smanjiti dubinu brazde na korenu eliminacijom nepoželjnih matičnih korenova, ali je mnogo teže poboljšati oblik korena zbog varijabilnosti zemlji-šta i klimatskih činilaca (Biancardi 1999).
Kao rezultat analize grupisanja, dendrogram za površinu korena čine pet grupa genotipova. U prvoj grupi se nalaze tri monogermna genotipa: 1, 3 i 4, čija srednja vrednost ocene iznosi 5. U okviru prve grupe na nešto višem hijerarhijskom nivou se pridružuje genotip 11 sa srednjom vred-nošću ocene 4 (Graf. 2).
Genotipovi 2, 6, 13, 19 i 20 se karakterišu sred-njom vrednošću 3 i oni se nalaze u drugoj grupi. U trećoj grupi se izdvajaju monogermni genoti-povi: 5, 7, 8, 9 i 10. Njima se pridružuju multiger-mni genotipovi: 14, 15, 16 i 18 koji se karakterišu nešto većim variranjem za ovo ispitivano svoj-stvo. Ukupna srednja vrednost ocene genotipova iz treće grupe iznosi 4. U okviru dendrograma se posebno izdvajaju genotip 12, koji se karakteriše najmanjom srednjom vrednošću 2 za površinu korena i genotip 17 sa najvećom srednjom vred-nošću 9 (Graf. 2). Specifi čnost genotipa 17 je što se odlikuje odsustvom varijabilnosti za ispitivano svojstvo, a ujedno poseduje veliki udeo germplaz-me divljeg srodnika šećerne repe Beta maritima, te se pretpostavlja da je to jedan od glavnih razloga visoke vrednosti za izgled površine korena ovog genotipa.
Slika 3. Genotip 17Fig. 3. Genotype 17
Grafi kon 2. Dendrogram ispitivanih genotipova za površinu korena šećerne repeGraph 2. Dendrogram of evaluated sugar beet genotypes for root surface

Danojević D i sar.90
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 87-90
Zaključci
Genotip 12 je iskazao najmanju srednju vred-nost ocene za površinu korena od svih ispitivanih genotipova, a zabeleženo je i prisustvo koreno-va sa ocenom 1. Odabiranjem takvih korenova postigla bi se još niža ocena za površinu korena nakon selekcije, a time i manje čestica zemljišta na njima. Genotip 12 predstavlja izvor za pobolj-šanje drugih genotipova koji nemaju nisku ocenu za površinu korena, a pored ovoga se odlikuje i drugim dobrim agronomskim svojstvima.
U našoj zemlji se do sada nije značajnije ba-vilo istraživanjem površine korena šećerne repe. Zbog interesovanja šećerana, kao i proizvođača za gajenje glatkih genotipova, u Institutu za ratar-stvo i povrtarstvo u Novom Sadu se odnedavno obraća veća pažnja na površinu korena. Počelo je odabiranje u okviru postojećeg selekcionog ma-terijala, kao i prikupljanje materijala iz inostranih kolekcija koje će se uključiti u program opleme-njivanja u cilju dobijanja hibrida šećerne repe sa manjim sadržajem zemljišta na korenu.
Literatura
Biancardi E (1999): Miglioramento genetico. In: Casarini B, Biancardi E, Ranalli P: La barbabietola negli ambientimedi-terranei. Edagricole, Bologna, Italy, 57-145
Gram N H, Joergensen A B (2002) : Reduction of soil tare through breeding techniques. Proc. 65th IIRB Congress: 99-106
International Board for Plant Genetic Resources (1991): Des-criptors for Beta. IBPGR/CGN, Rome
Koch H J (2002): Einfl uss der Anbaugestaltung auf den Erdan-hang von Zuckerrüben. Proc. 65th IIRB Congress: 107-116
McGrath J M (2003): Registration of SR96 and SR97 Smoth-Rooth Sugarbeet Germplasm with High Sucrose. Crop Sci. 43: 2314-2315
McGrath J M, Lewellen R T (2004): Registration of EL0204 Sugarbeet Germplasm with Smooth-Root and Resistance to Rhizomania. Crop Sci. 44: 1032-1033
Mesken M (1987): Mass selection for crown height in sugar beets (Beta vulgaris L.), Divergent selection in diploids. Eup-hytica 36: 129-145
Panella L, Lewellen R T (2007): Broadening the genetic base of sugar beet: introgression from wild relatives. Euphytica 154: 383-400
Saunders J W, McGrath J M, Halloin J M, Theurer J C (1999): Registration of SR94 sugarbeet germplasm with smooth root. Crop Sci. 39: 297
Saunders J W, McGrath J M, Halloin J M, Theurer J C (2000a): Registration of SR95 sugarbeet germplasm with smooth root. Crop Sci. 40: 1205-1206
Saunders J W, McGrath J M, Theurer J C, Halloin J M (2000b): Registration of SR87 sugarbeet germplasm with low soil tare and resistances to Cercospora and Aphanomyces. Crop Sci. 40: 1833-1834
Theurer J C (1993): Pre-breeding to change sugarbeet root arc-hitecture. J. Sugar Beet Res. 30: 221-239
Root Surface as Criterion for Selection of Sugar Beet
Dario Danojević · Živko Ćurčić · Nevena Nagl · Lazar Kovačev
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: In this study we analyzed 10 monogerm and 10 multigerm sugar beet genotypes from the collections of the Institute of Field and Vegetable Crops in Novi Sad. The appearance of sugar beet root surface was evaluated as a property that has an impact on the soil removal and root washing in the sugar factory. The genotypes were tested with nonparametric Kruskal-Wallis’s test and cluster analysis as a method of multivariate analysis. Based on these results, genotype 12 had the lowest values for the root surface (the least furrow of the root), which will further be used in sugar beet breeding programs.Key words: multivariate analysis, root surface, selection, sugar beet

Genetički doprinos oplemenjivanju lucerke 91
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98originalni naučni rad / original research article
Genetički doprinos oplemenjivanju lucerke: prinos eksperimentalnih populacija
u odnosu na priznate sorte
Slobodan Katić · Dragan Milić · Snežana Katanski · Đura Karagić · Sanja Vasiljević
primljeno / received: 29.10.2010. prihvaćeno / accepted: 16.11.2010.© 2011 IFVC
Izvod: Oplemenjivanje lucerke u svetu je započeto početkom prošlog veka, a u Srbiji sredinom prošlog veka. Kontradiktorni rezultati o genetičkom doprinosu oplemenjivanja na prinos lucerke u svetu i kod nas su posledica analize prinosa iz različitih ogleda, na različitim lokacijama, kroz različite godine, za-nemarujući interakciju germplazma x godina (starost lucerišta) i germplazma x okolina (ekološki uslovi vezani za lokalitet ili uslove u godini). U želji da se proceni napredak u oplemenjivanju lucerke na prinos u našoj zemlji upoređene su nove eksperimentalne populacije sa novim i starim sortama lucerke stvo-renim u Institutu za ratarstvo i povrtarstvo u Novom Sadu. Ogled je zasnovan 2008. i praćen tokom 2008-2010. Razlike u prinosu između sorti i eksperimentalnih populacija lucerke nisu značajne. Nove programe oplemenjivanja lucerke treba fokusirati na prinos per se, primenom novih saznanja o genetičkoj kontroli prinosa, adaptacijom postojećih ili kreiranjem novih procedura oplemenjivanja.Ključne reči: genetički doprinos, lucerka, oplemenjivanje, prinos, sorta
S. Katić · D. Milić · S. Katanski · Đ. Karagić · S. VasiljevićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Lucerka je stara kultura koja se gaji radi proi-zvodnje kabaste stočne hrane. Raširena je širom sveta na preko 30 miliona hektara (Michaud et al. 1988), što je izazvalo potrebu za oplemenjiva-njem i stvaranjem sorti koje se odlikuju različitim osobinama. Oplemenjivanje lucerke u svetu je za-počelo početkom prošlog veka (Lamb et al. 2006) a u našoj zemlji sredinom prošlog veka (Mihailo-vić i sar. 2008).
S obzirom na cilj gajenja, glavni pravac ople-menjivanja lucerke bio je i ostaje prinos nadze-mne vegetativne mase (krme). Za oplemenjivanje lucerke koriste se jednostavne metode selekcije (masovna, individualna, fenotipska, rekurentna selekcija i dr.). Međutim, napredak u oplemenji-vanju lucerke je spor, pre svega primenom ovih metoda selekcije. Glavni rezultat primene aktu-elnih oplemenjivačkih metoda kod lucerke go-vori da efi kasnost oplemenjivanja snažno zavisi od populacije sa kojom oplemenjivač radi i spe-cifi čnih uslova istraživanja, a da uspeh selekcije
(genetska dobit) ne može biti sigurna (Rowe & Hill 1999).
Od sredine prošlog veka počinju da se rade analize procene genetskog doprinosa opleme-njivanju lucerke na prinos. Jednu od prvih ana-liza načinili su Hill et al. (1988) koji ukazuju da je napredak bio < 1% godišnje za period 1971-1981. Sveobuhvatniju analizu su uradili autori Bingham et al. (1994) posmatrajući sorte kroz tri perioda stvaranja 1910-1940, 1953-1979 i 1979-1985, te zaključuju da prinos raste od prvog ka trećem periodu. Međutim, autori Wiersma et al. (1997) porede stvorene sorte u periodu 1907-1998. i dobijaju manji pad u prinosu kod novijih sorti. Različiti rezultati o genetičkom napretku i promeni prinosa sorti lucerke stvorenih u prošlom veku doveli su do diskusije o realnoj mogućnosti oplemenjivanja lucerke na prinos (Bingham et al. 1994, Brummer 1999, Volonec et al. 2002, Lamb et al. 2006). Ističu se neki faktori koji doprinose sporom napretku ople-menjivanja lucerke na prinos: višegodišnji život biljaka što zahteva višegodinje proučavanje pre bilo koje odluke u selekciji, zatim preživljavanje ekoloških i biotičkih faktora, entomofi lno opra-šivanje, sitan, hermafroditan cvet, polisomično nasleđivanje. Pošto se koristi ceo nadzemni deo biljke nemoguće je preusmeravanje biljnih
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Katić S i sar.92
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
asimilativa da bi se povećao žetveni indeks, kao što se to čini kod zrnastih kultura (Hill et al. 1988).
Spor napredak u oplemenjivanju lucerke na prinos doveo je do toga da su selekcioneri lu-cerke fokusirali oplemenjivačke programe na druge ciljeve, kao što su kvalitet, hranljiva vred-nost krme, otpornost na bolesti, štetočine i dr. Međutim, otpornost sorti prema bolestima ima prednost u uslovima povoljnim za tu bolest, ali u uslovima bez rizika za pojavu bolesti nema razlike u prinosu. Oplemenjivanjem na otpor-nost prema drugim osobinama ne povećava se frekvencija poželjnih gena za prinos (Lamb et al. 2006).
U našoj zemlji nije analiziran genetički dopri-nos oplemenjivanja na prinos, ali su analizirane i upoređivane domaće i strane sorte (Katić et al. 2005), poređene su domaće sorte po ciklusima i prema ciljevima oplemenjivanja (Katić i sar. 2008). Analize ukazuju da se sorte razlikuju iz ogleda u ogled i da su relativno male razlike u pri-nosu sorti, posebno kad se u ogledu nalazi manji broj sorti (Volonec et al. 2002, Katić i sar. 2008, Katić i sar. 2010).
Kontradiktorni rezultati o genetičkom dopri-nosu oplemenjivanja na prinos lucerke u svetu i kod nas su posledica analize prinosa iz različitih ogleda, na različitim lokacijama, kroz različite godine zanemarujući interakciju germplazma x godina (starost lucerišta) i germplazma x okoli-na (ekološki uslovi vezani za lokalitet ili uslove u godini).
Oplemenjivanje lucerke na prinos je pono-vo u žiži interesovanja mnogih oplemenjivača lucerke (Brumner 1999, Volonec et al. 2002, Lamb et al. 2006). Predlaže se primena novih metoda selekcije oplemenjivanja u cilju iskori-šćavanja (zahvatanja) neaditivnog dejstva gena (Riday et al. 2002).
U želji da se proceni napredak u oplemenji-vanju lucerke na prinos u našoj zemlji upore-đene su nove eksperimentalne populacije sa novim i starim sortama lucerke stvorenim u Institutu za ratarstvo i povrtarstvo u Novom Sadu.
Materijal i metod rada
U ogled su uključene četiri eksperimentalne populacije (80407; SKV 120607; SRB 80607; i MSS x MSV) i četiri sorte Instituta za ratarstvo i povrtarstvo (Banat VS, NS Mediana ZMS V, NS Alfa, Nijagara).
Poreklo i nastanak eksperimentalnih populacija: po-sle jednokratne masovne selekcije osam doma-
ćih populacija lucerke nastala je populacija SRB 80607; primenom individualne selekcije i umno-žavanjem u polycross-u najboljih genotipova iz ukrštanja sorti ♀ Live x ♂ Mediana ; ♀ Cheronia x ♂ Zuzana; ♀ La rocca x ♂ Vertibenda i ♀ Do-lichi x ♂ NS Banat ZMS II nastala je populacija 80407; iz ukrštanja sorti ♀ Europe x ♂ Jogeva 118, setvom F1 potomstava u redove i umnožava-njem u polycross-u nakon dve generacije nastala je populacija MSS x MSV, i primenom jednokratne individualne selekcije i izborom najboljih bilja-ka iz srpskih, bugarskih, francuskih američkih, čeških i španskih sorti nastala je populacija SKV 120607.
Poljski ogled zasnovan je u proleće 2008. na slabo-karbonatnom tipu zemljišta na eksperimen-talnom polju Instituta za ratarstvo i povrtarstvo na Rimskim Šančevima. Ogled je postavljen po slučajnom blok sistemu u pet ponavljanja. Veliči-na osnovne parcele bila je 5 m2, razmak između parcela 40 cm, a između blokova 100 cm. Tokom 2008. dobijena su dva otkosa, u 2009. tri otkosa, a u 2010. četiri otkosa. Prinos zelene krme odre-đen je merenjem pokošene nadzemne mase kom-bajnom CIBUS direktno na parceli i preračunat u t ha-1. Uzet je uzorak (0,3 kg) za određivanje sadržaja suve materije iz svake parcelice iz čega je obračunat prinos suve materije (t ha-1). Visi-na biljaka izmerena je u prvom i drugom otkosu 2010. merenjem najviših biljaka u tri ponavljanja u svakoj parcelici.
Rezultati su predstavljeni po godinama (2008, 2009. i 2010) i obrađeni analizom varijanse za dvofaktorijalni ogled, gde su faktor A ekspe-rimentalne populacije i sorte a faktor B otkos i predstavljeni po faktoru A. Podaci za period 2009-2010. su predstavljeni kao suma 7 otkosa tokom dve godine i obrađeni analizom varijanse za jednofaktorijalni ogled. Razlike u prinosu i visini biljaka ocenjene su NZR testom, a varira-nje je izraženo koefi cijentom varijacije.
Rezultati i diskusija
U godini setve (2008) prinos zelene krme se na godišnjem nivou kretao od 19,8 t ha-1 (MS Me-diana ZMS V), do 22,4 t ha-1 (Nijagara), a prinos suve materije od 7,6 t ha-1 (SRB 80607) do 9,6 t ha-1 (Banat VS). Razlike u prinosu između pro-seka sorti i proseka populacija nisu bile značajne (Tab. 1).
U drugoj godini istraživanja (2009) na godiš-njem nivou (suma tri otkosa) najveći prinos je ostvaren kod sorte Nijagara (56,4 t ha-1 zelene krme i 16,5 t ha-1 suve materije), a najmanji kod populacije SKV 120607 (48,3 t ha-1 zelene krme

Genetički doprinos oplemenjivanju lucerke 93
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
i 11,4 t ha-1 suve materije). Nema statistički zna-čajnih razlika u prinosu između proseka ekspe-rimentalnih populacija i proseka sorti lucerke. Međutim, dobijene su značajne razlike u prinosu između eksperimentalne populacije SKV120607 i sorte Nijagara (Tab. 2).
Razlike u prinosu zelene krme i suve materije u trećoj godini života biljaka lucerke (2010) nisu statistički značajne (Tab. 3). Najveći prinos ze-lene krme kod sorti ostvario je Banat VS (82,3 t ha-1), a suve materije sorta NS Alfa (17,8 t ha-1). Najmanje prinose zelene krme i suve materije
Tabela 1. Prinos zelene krme i suve materije sorti/populacija lucerke u 2008. Table 1. Green forage and dry matter yield of alfalfa cultivars/populations in 2008
Godina priznavanja
Year of release
Sorta/populacijaCultivar/population
Prinos zelene krmeGreen forage yield
(t ha-1)
Prinos suve materijeDry matter yield
(t ha-1)
1980 (2000)* NS Mediana ZMS V 19,8 8,42005 Banat VS 21,3 9,62005 NS Alfa 21,0 8,62007 Nijagara 22,4 8,6
Prosek sorti - mean 21,1 8,8eks. pop. 80407 20,0 8,4eks. pop. SKV 120607 21,4 9,6eks. pop. SRB80607 20,4 7,6eks. pop. MSS x MSV 20,6 9,0
Prosek pop. - mean 20,6 8,7Prosek ogleda - mean 20,9 8,7
CV 13,1 13,10,05
LSD 0,012,43,2
1,01,4
* godina repriznavanja; year of release
Tabela 2. Prinos zelene krme i suve materije sorti/populacija lucerke u 2009. Table 2. Green forage and dry matter yield of alfalfa cultivars/populations in 2009
Godina priznavanja
Year of release
Sorta/populacijaCultivar/population
Prinos zelene krmeGreen forage yield
(t ha-1)
Prinos suve materije Dry matter yield
(t ha-1)1980 (2000)* NS Mediana ZMS V 52,2 12,3
2005 Banat VS 51,8 12,62005 NS Alfa 52,2 12,92007 Nijagara 56,4 13,5
Prosek sorti - mean 53,1 12,9eks. pop. 80407 53,7 13,2eks. pop. SKV 120607 48,3 11,4eks. pop. SRB80607 51,0 12,6eks. pop. MSS x MSV 52,2 12,9
Prosek pop. - mean 51,3 12,6Prosek ogleda - mean 52,2 12,6
CV 18,6 20,10,05
LSD0,016,99,3
1,82,4
* godina repriznavanja; year of release

Katić S i sar.94
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
ostvarila je eksperimentalna populacija SKV 120607 (73,6 t ha-1 i 15,2 t ha-1). Razlike u pro-sečnoj visini biljaka bile su relativno male (CV = 4,9), a nije bilo statistički značajnih razlika izme-đu priznatih sorti i eksperimentalnih populacija (Tab. 3)
Najviše biljke (značajno više) od svih sorti i eksperimentalnih populacija ima populacija SKV 120607 (91,1 cm). Najniže biljke na nivou ogleda imala je populacija 80407 (81,8 cm).
Na osnovu sumarnih rezultata prinosa zelene krme i suve materije (prinos 7 otkosa), u dru-goj i trećoj godini ispitivanja (2009-2010), a to su godine pune eksploatacije lucerišta, uočeno je da između proseka eksperimentalnih populacija i priznatih sorti nema značajnih razlika u prinosu (Tab. 4).
U periodu 2009-2010. najveći prinos zelene krme na nivou ogleda imaju sorta Nijagara i ek-sperimentalna populacija 80407 (137,2 t ha-1), a suve materije sorta NS Alfa i eksperimentalna populacija 80407 sa istim prinosom (30,8 t ha-1). Između prosečnog prinosa sorti i prosečnog prinosa eksperimentalnih populacija nisu zabe-ležene značajne razlike (Tab. 4). Rezultati istra-živanja ukazuju na značajan genetski potencijal prinosa zelene krme i suve materije kako pri-znatih sorti tako i eksperimentalnih populacija lucerke.
Na osnovu dobijenih rezultata može se reći da nema značajnih korelacija između prinosa i visi-ne biljaka. Međutim, veći broj radova ukazuje na značajnu vezu visine i prinosa kod lucerke (Rotili et al. 1999, Riday and Brummer 2002). Razlike u našem eksperimentu u odnosu na druge možda
leže i u načinu merenja visine biljaka. Naime, u našem radu visina biljaka je registrovana na tri mesta merenjem najviših biljaka, a u drugim istra-živanjima određivana je visina (dužina) 20 slučaj-no izabranih izdanaka (Katić i sar. 2008, Riday & Brummer 2002).
Male razlike u prinosu između eksperimental-nih populacija i sorti u skladu su sa ranijim rezul-tatima koji navode da je primenom jednostavnih metoda oplemenjivanja lucerke (klasične masov-ne i individualne selekcije) moguć mali napredak (Bingham et al. 1994, Brummer 1999, Volonec et al. 2002, Lamb et al. 2006). Takođe, u oplemenji-vanju lucerke se ne može zanemariti interakcija genotip x spoljašni uslovi, odnosno uticaj godine života biljaka kao i ekološki uslovi koji su pri-sutni u godinama ispitivanja (Lamb et al. 2006). Naši rezultati govore o smenjivanju sorti u visini prinosa zavisno od godine života biljaka, ali i od ekoloških uslova u godini kao i različite plodnosti zemljišta.
Genetički efekti utiču da su razlike između sor-ti i eksperimentalnih populacija male. Stvaranjem
Tabela 3. Prinos zelene krme, suve materije i visina sorti/populacija lucerke u 2010. Table 3. Green forage, dry matter yield and plant height of alfalfa cultivars/populations in 2010
Godina priznavanja
Year of release
Sorta/populacijaCultivar/population
Prinos zelene krmeGreen forage yield
(t ha-1)
Prinos suve materijeDry matter yield
(t ha-1)
VisinaHeight(cm)
1980 (2000)* NS Mediana ZMS V 82,1 17,3 82,12005 Banat VS 82,3 17,7 82,32005 NS Alfa 82,0 17,8 84,72007 Nijagara 80,8 16,9 82,8
Prosek sorti - mean 81,8 17,4 83,0eks. pop. 80407 83,1 17,4 84,4eks. pop. SKV 120607 73,6 15,2 91,1eks. pop. SRB80607 76,0 16,6 81,8eks. pop. MSS x MSV 79,1 16,2 84,3
Prosek pop. - mean 78,0 16,4 85,4Prosek ogleda - mean 79,9 16,9 84,2
CV 12,0 15,7 4,90,05
LSD 0,016,07,9
1,72,2
3,74,5
* godina repriznavanja; year of release

Genetički doprinos oplemenjivanju lucerke 95
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
i umnožavanjem eksperimentalnih populacija, gde su prvi koraci rad sa jednom ili nekoliko do-brih (prinosnih) roditeljskih biljaka, moguća je pojava inbridinga i inbred depresije, koja brzo dovodi da potomstvo najprinosnijih biljaka ne bude prinosnije od priznatih široko rasprostra-njenih sorti. Gajenjem priznatih sorti lucerke u cilju proizvodnje semena na većem broju loka-liteta daje mogućnost izbora najboljih biljaka sa tih lokaliteta i setvu odabranih biljaka u širi po-likros, čime se može izbeći inbred depresija koja se javlja pri inicijalnom razvoju sorte. Ovakav pristup semenarstvu i održavanju sorti dovodi do ispoljavanja brojnih rekombinacija, ili čak do akumulacije poželjnih gena, ne javlja se problem inbred depresije i odabiraju se ekološkim uslo-vima najprilagođeniji genotipovi, što priznatim i široko rasprostranjenim sortama daje izuzetnu prednost.
Suočeni sa ovakvim problemima, oplemenjiva-či lucerke u svetu i Srbiji fokusiraju svoj rad na popravljanje drugih osobina: otpornost prema bolestima, veći kvalitet, intenzitet kosidbe i dru-go (Volonec et al. 2002, Lamb et al. 2006, Katić i sar. 2008). Analizirajući sorte stvorene u Institutu za ratarstvo i povrtarstvo u Novom Sadu važno je reći da je sorta Banat VS proizišla iz sorte NS Banat ZMS II (priznata 1964. godine), veoma otporna na niske temperature, sušu, da podnosi intenzivnu kosidbu i daje veći prinos suve mate-
rije (Katić i sar. 2008). Sorta NS Mediana ZMS V stvorena je ukrštanjem sa žutom lucerkom i ističe se pogodnošću za gajenje na lošijim zemljištima kao i kvalitetom. Sa istim ciljem stvorena je i sor-ta Nijagara koja takođe podnosi lošije zemljišne uslove, otporna je na dominantne bolesti i odlič-nog kvaliteta, NS Alfa je stvorena u cilju poprav-ljanja otpornosti prema poleganju i povećanja kvaliteta (Mihailović i sar. 2008).
Ukrštanjem stare francuske sorte Evropa (M. sativa ssp. sativa L.) i estonske sorte Jogeva 118 (M. sativa ssp. varia L.) stvorena je populacija MSS x MSV u cilju da se kombinuju uspravan i brz porast (Evropa) i otpornost prema ekološki stre-snim uslovima (Jogeva 118). Cilj stvaranja popu-lacije 80407 je bio da se stvori sorta pogodna za teža hidromorfna zemljišta. Kod populacije SRB 80607 pokušano je da se homogenizuje domaći materijal i akomuliraju poželjni geni iz većine do-maćih sorti. Cilj stvaranja populacije SKV 120607 je bio da se objedini dosta heterogen materijal iz različitih gen centara u cilju stvaranja ranog sin-tetika lucerke prilagođenog intenzivnom sistemu kosidbe. Niži prinos ove eksperimentalne popu-lacije može biti posledica široke genetičke osno-ve ove populacije, ali i to što ekološki uslovi nisu pogodovali primeni petootkosnog sistema, već je primenjen ekstezivan sistem kosidbe (tri i četiri otkosa u drugoj i trećoj godini), pa genetski po-tencijal ove sorte nije došao do izražaja.
Tabela 4. Prinos zelene krme i suve materije sorti/populacija lucerke tokom 2009-2010.Table 4. Green forage and dry matter yield of alfalfa cultivars/populations during 2009-2010
Godina priznavanja
Year of release
Sorta/populacijaCultivar/population
Prinos zelene krmeGreen forage yield
(t ha-1)
Prinos suve materijeDry matter yield
(t ha-1)
1980 (2000)* NS Mediana ZMS V 134,4 30,12005 Banat VS 133,7 30,12005 NS Alfa 134,4 30,82007 Nijagara 137,2 30,1
Prosek sorti - mean 135,1 30,1eks. pop. 80407 137,2 30,8eks. pop. SKV 120607 121,8 26,6eks. pop. SRB80607 126,7 29,4eks. pop. MSS x MSV 131,6 28,7
Prosek pop. - mean 129,5 28,7Prosek ogleda - mean 132,1 29,6
CV 8,1 7,70,05
LSD 0,0114,018,2
2,84,2
* godina repriznavanja; year of release

Katić S i sar.96
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
Saznanja o malom genetičkom unapređenju lu-cerke u svetu i kod nas dovelo je do preispitivanja postojećih metoda u oplemenjivanju lucerke na prinos per se. U više centara (Brummer 1999, Ro-tili et al. 1999) se istovremeno došlo do ideje o mogućnosti delimičnog iskorišćavanja heterozisa kod lucerke konceptom proizvodnje semihibrida lucerke. Ovaj pristup podrazumeva poboljšanja unutar odvojenih populacija, identifi kaciju i odr-žavanje heterotičnih grupa (u geografski udalje-nim centrima ili u jednom centru u prostornoj izolaciji) i ukrštanje tih populacija u cilju proi-zvodnje semihibridnog semena. Ideja o semihi-bridnom pristupu oplemenjivanja lucerke prihva-ćena je i intezivno se radi i u Institutu za ratarstvo i povrtarstvo u Novom Sadu, te se u budućnosti očekuju prvi rezultati primene ovakvog pristupa u oplemenjivanju lucerke na prinos.
Zaključak
Razlike u prinosu između sorti i eksperimen-talnih populacija lucerke nisu statistički značajne. Primenom jednostavnih metoda selekcije nema genetičkog napretka u oplemenjivanju lucerke na prinos, ili je napredak veoma mali.
Nove programe oplemenjivanja lucerke treba fokusirati na prinos per se, primenom novih sa-znanja o genetičkoj kontroli prinosa, adaptacijom postojećih ili kreiranjem novih procedura ople-menjivanja.
Lucerka je najznačajnija krmna biljka kod nas i u svetu, pa inteziviranje rada na poboljšanju pri-nosa per se ima smisla i predstavlja jak motiv za dalji rad na oplemenjivanju lucerke.

Genetički doprinos oplemenjivanju lucerke 97
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
Literatura
Bingham E T, Groose R W, Woodfi eld D R, Kidwell K K (1994): Complementary gene interactions in alfalfa are greater in autotetraploids than diploids. Crop Sci. 34: 823-829
Hill R R Jr, Shenk J S, Barnes R F (1988): Breeding for yield and quality. In: Hanson, A.A., Barnes D.K. and Hill R.R. Jr. (ed.). Alfalfa and Alfalfa Impro-vement. ASA-CSSA-SSSA, Madison, Wisconsin, USA, 809-825
Katic S, Milic D, Vasiljevic S (2005): Variability of dry matter yield and quality of lucerne genotypes de-pending on geographic origin. EGF, Grassland Sci. Eur. 10: 537-540
Katić S, Mihailović V, Milić D, Vasiljević S, Karagić Đ (2008): Osobine sorti lucerke kao rezultat različitih ciljeva oplemenjivanja. Zbornik radova Instituta za ratarstvo i povrtarstvo Novi Sad 45: 163-170
Katić S, Milić D, Mihailović V, Karagić Đ, Pojić M (2010): Prinos, komponente prinosa i svarljivost suve materije eksperimentalnih populacija lucerke. Ratar. Povrt. / Field Veg. Crop Res. 47: 209-216
Lamb F S J, Sheaffer C C, Rhodes H L, Sulc R M, Un-dersander J D, Brummer E C (2006): Five Deca-des of Alfalfa Cultivar Improvement: Impact on Forage Yield, Persistence, and Nutritive Value. Crop Sci. 46: 902-909
Michaud R, Lehnan W F, Rumbaugh M D (1988): World distribution and historical development. In: Han-
son, A.A., Barnes D.K. and Hill R.R. Jr. (eds.), Al-falfa and Alfalfa Improvement. ASA-CSSA-SSSA, Madison, Wisconsin, USA, 25-124
Mihailović V, Katić S, Ćupina B, Vasiljević S, Karagić Đ, Pataki I, Mikić A, Milić D (2008): Rezultati u ople-menjivanju, agrotehnici i semenarstvu krmnih bi-ljaka u Institutu za ratarstvo i povrtarstvo. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 45: 81-101
Riday H, Brummer E C, Moore K (2002): Heterosis of forage quality in alfalfa. Crop Sci. 42: 1088-1093
Riday H, Brummer E C (2002): Heterosis of agronomic traits in alfalfa. Crop Sci. 42: 1081-1087
Rotili P, Gnocchi G, Scotti C, Zannone L (1999): Some aspects of breeding methodology in alfalfa. The Alfalfa Genome. [Elektronski izvor] Dostupno na http://www.naaic.org/TAG/TAGpapers/rotili/rotilipapers.html
Rowe D E, Hill R R Jr (1999): Breeding Theory and the Development of Alfalfa. The Alfalfa Genome. [Elektronski izvor] Dostupno na www.naaic.org/TAG/TAGpapers/RoweAbs.html
Volenec J J, Cunningham S M, Haagenson D M, Berg W K, Joern B C, Wiersma D W (2002): Physiological genetics of alfalfa improvement: past failures, fu-ture prospects. Field Crops Res. 75: 97-110
Wiersma D W, Undersander D J, Lauer J G (1997): Lack of alfalfa yield progress in the Midwest. p. 36. In Central Alfalfa Improvement Conf. Abstracts. La-Crosse, 16–18 July 1997.

Katić S i sar.98
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 91-98
Genetic Gain in Alfalfa Breeding: Yield of Experimental Populations Versus Released Cultivars
Slobodan Katić · Dragan Milić · Snežana Katanski · Đura Karagić · Sanja Vasiljević
IInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Alfalfa breeding programs in the world started at the beginning of the last century, while in Serbia they started in the middle of last century. The contradictory results on the genetic gain in alfalfa breeding for yield in the world and in our country had as a consequence the yield analysis from different experiments, in different locations, through different years, neglecting the interaction germplasm x year (age of alfalfa fi elds), and germplasm by envi-ronment (ecological conditions related to site or conditions in the year). In order to assess the progress in breeding alfalfa for yield in our country, new experimental populations were compared with old and new alfalfa cultivars developed in the Institute of Field and Vegetable Crops, Novi Sad, Serbia. Yield differences between cultivars and experimental populations of alfalfa were not signifi cant. New alfalfa breeding programs should focus on yield per se, with the use of new knowledge on the genetic control of yield and with adaptation of existing or creating new breeding procedure.Key words: alfalfa, breeding, cultivar, genetic gain, yield

Način nasleđivanja debljine stabla F1 hibrida krmnog sirka 99
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106originalni naučni rad / original research article
Kombinacione sposobnosti i način nasleđivanjadebljine stabla F1 hibrida krmnog sirka
(Sorghum bicolor L. Moench)
Imre Pataki
primljeno / received: 04.11.2010. prerađeno / revised: 06.12.2010. prihvaćeno / accepted: 29.12.2010.© 2011 IFVCIzvod: Cilj ispitivanja je bilo utvrđivanje načina nasleđivanja, efekta gena, komponenti genetske vari-janse, kombinacionih sposobnosti, prosečnog doprinosa linija i testera i njihove interakcije u ekspresiji debljine stabla u F1 generaciji krmnog sirka. Primenjen je metod linija x tester. Materijal je obuhvatao osam genetski divergentnih A-inbred linija sirka za zrno, tri R linije-testera sudanske trave i dvadeset četiri F1 hibrida dobijenih njihovim ukrštanjem. Između ispitivanih genotipova ustanovljene su značajne razlike u srednjim vrednostima za debljinu stabla. Rezultati analize varijanse kombinacionih sposobnosti pokazuju da postoje visoko značajne razlike za opšte kombinacione sposobnosti (OKS) i posebne kom-binacione sposobnosti (PKS). Neaditivna komponenta genetske varijanse (dominacija i epistaza) ima veći udeo u ukupnoj genetskoj varijansi za debljinu stabla. U prvoj godini ispitivanja najveći doprinos u ekspresiji debljine stabla F1 hibrida na oba lokaliteta imala je interakcija inbred linija majki sa testerima, u drugoj godini ispitivanja na lokalitetu Rimski Šančevi najveći doprinos pripada linijama, dok su na lokalitetu Mačvanski Prnjavor najveći doprinos imali testeri. Ocena kombinacionih sposobnosti ukazuje na opravdanost korišćenja sledećih inbred linija sirka za zrno u svojstvu majki: SS-1 646, SS-1 688 i S-8 682 u programima oplemenjivanja krmnog sirka na veću debljinu stabla. Perspektivni hibridi krmnog sirka na osnovu analize PKS su S-8 682 x ST-R lin H, P-21 656 x C-198. Ovo istraživanje može biti od značaja za stvaranje novih visoko prinosnih hibrida krmnog sirka.Ključne reči: debljina stabla, kombinacione sposobnosti, komponente genetske varijanse, krmni sirak, način nasleđivanja debljine stabla, sudanska trava
I. Pataki Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
U poređenju sa drugim krmnim biljkama koje se gaje za biomasu, sirak ima tu prednost što podnosi ispašu i dobro podnosi sušu jer troši malo vode za stvaranje suve materije. Sirak ima razvijen korenov sistem jake usisne moći, te ko-risti iz zemljišta vodu i hranljive materije koje su za ostale gajene kulture nepristupačne ili teško pristupačne, a dobro reaguje na đubrenje i na-vodnjavanje. Posle kosidbe iz pupoljaka u prize-mnom delu stabla sirak se ponovo regeneriše. U ishrani se sirak koristi u mlađim fazama, kada je sadržaj celuloze manji.
Krmni sirak se gaji zbog dobijanja zelene mase, a glavni parametar za procenu prinosa je prinos zelene mase po biljci. U zelenoj masi biljke najveći udeo čini stablo. U mlađim fazama po-rasta sirak ima stabljiku sa puno soka, a od faze
porasta i brzine isušivanja stabla genotipovi se ra-zlikuju. Zbog kombinovanog iskorišćavanja sir-ka (zelena masa ili silaža) poželjno je da stabljika ostane sočna do kraja vegetacije, bez obzira na vremenske prilike. Suvo stablo ili stabljika sa ma-nje vlage je determinisana dominantnim genima, dok je sočna stabljika recesivno svojstvo (Swan-son & Parker 1931).
Debljinu stabla kod krmnog sirka najbolje je meriti od 3. do 6. Internodije, a sa stabljike treba odstraniti lisnu dršku. Patuljasti tipovi sirka ima-ju veći prečnik stabla u odnosu na visoke tipove, kako navode Barabas et al. (1985). Prečnik sta-bla kod hibrida krmnog sirka ima direktan uticaj na prinos zelene mase po biljci, što su ustanovi-li u svojim istraživanjima Zhan QuinWen et al. (2004), i on se nasleđuje parcijalno dominantno. U dialelnom ukrštanju 10 divergentnih genotipo-va sirka autori Tiwari et al. (2003) nisu ustanovili heterozis za debljinu stabla.
U proučavanju heterozisa 45 hibrida sirka nastalih ukrštanjem 10 homozigotnih linija u po-ludialelnom ukrštanju, autori Ravindrababu et al. (2002) su konstatovali mali heterozis za debljinu
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Pataki I100
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
stabla. Hibridna snaga se manifestuje u većini ekonomski važnih svojstava sirka i osnovni je pre-duslov za dobijanje hibrida s većom produktiv-nošću i uniformnošću. Po visokom proizvodnom potencijalu naročito se ističu hibridi tipa sirak za zrno x sudanska trava. Ovi hibridi su naročito pogodni za proizvodnju zelene krme, silažiranje i pravljenje sena (Berenji 1988). Na osnovu zapa-žanja i analize podataka iz ogleda oplemenjivač odabira inbred linije sa dobrim kombinacionim sposobnostima za poželjna svojstva, kako bi se u što kraćem vremenu izdvojili i umnožili najbolji F1 hibridi.
Cilj ovog rada je bio da se ispita način nasleđi-vanje debljine stabla, opšte kombinacione sposob-nosti (OKS), posebne kombinacione sposobnosti (PKS), komponente genetičke varijanse kao poka-zatelja aditivne i neaditivne varijanse zatim propor-cionalni doprinos linija, testera i njihove interakcije u ekspresiji debljina stabla inbred linija testera i F1 hibrida tipa sirak za zrno x sudanska trava.
Materijal i metod
Materijal korišćen u ovom radu obuhvata osam divergentnih citoplazmatsko muško ste-rilnih A-linija sirka za zrno: SS-1 646, SS-1 648, SS-1 688, S-8 682, P-21 656, P-74 633, P-170 630, P-174 621, tri R-restorer linije sudanske trave koje su korišćene kao testeri dobrih kombinacio-nih sposobnosti: C-198, ST-R lin H, ST-310 i 24 hibrida F1 generacije.
Materijal je selekcionisan u Odeljenju za kr-mno bilje Instituta za ratarstvo i povrtarstvo u Novom Sadu na oglednom polju Rimski Šančevi. Ukrštanje i proizvodnja semena hibrida F1 ge-neracije i inbred linija izvršeni su 2001. na od-vojenim parcelama, sve cms linije su posejane sa jednom R-linijom. Parcele su bile udaljene više od 150 m. Ogled sa roditeljima i F1 hibridima bio je postavljen 2002. i 2003. na dva lokaliteta, na oglednom polju Instituta za ratarstvo i povrtar-stvo u Novom Sadu i u Zemljoradničkoj zadruzi Mačvanski Prnjavor. Materijal je posejan u jed-noredne parcele dužine 5 m u tri ponavljanja po slučajnom blok sistemu. Međuredni razmak je iznosio 70 cm, a između biljaka unutar reda 15 cm. U toku vegetacije primenjene su uobičajene agrotehničke mere. Osnovni uzorak za analizu sastojao se od 30 biljaka (10 biljaka po ponavlja-nju) bez rubnih biljaka. Debljina stabla je merena od 3. do 6. internodije i izražena je u cm. Mor-fološka analiza je rađena nakon cvetanja kada su biljke postigle punu visinu.
Od osnovnih statističkih parametara je kori-šćena aritmetička sredina, standardna greška ari- G
enot
ip
Gen
otyp
eD
eblji
na st
abla
/ St
em th
ickne
ss
Rim
ski Š
anče
vi 2
002
Mač
vans
ki P
rnjav
or 2
002
Rim
ski Š
anče
vi 2
003
Mač
vans
ki P
rnjav
or 2
003
x
± s
x
S
V x
± s
x
S
V
x
± s
x
S
V
x
± s
x
S
V
SS-1
646
SS-1
648
SS-1
688
S-8
682
P-21
656
P-74
633
P-17
0 63
0P-
174
621
2.63
± 0
.03
0
.19
7.2
82.
12±
0.03
0.2
0 9
.23
2.32
±0.
04
0
.21
9.1
42.
27±
0.03
0.1
5 6
.66
2.42
±0.
02
0
.12
5.1
12.
30±
0.02
0.1
3 5
.64
2.01
±0.
02
0
.11
5.6
02.
31±
0.03
0.1
4 6
.23
1.20
±0.
02
0
.14
11.
441.
30±
0.03
0.1
4 1
0.94
1.57
±0.
03
0
.15
9.
961.
87±
0.03
0.1
6
8.59
1.10
±0.
02
0
.14
12.
701.
49±
0.02
0.1
3
8.60
1.60
±0.
02
0.1
2 7
.52
1.47
±0.
03
0
.15
10.1
3
1.24
±0.
03
0
.14
11.
511.
39±
0.03
0.1
6 11
.32
1.60
±0.
03
0
.18
11.4
01.
92±
0.03
0.1
9 9
.82
1.16
±0.
03
0
.16
13.7
51.
54±
0.03
0.1
4 9
.27
1.68
±0.
03
0
.15
9.0
31.
53±
0.03
0.1
9 12
.39
2.53
±0.
02
0
.13
4.9
82.
10±
0.02
0.1
3 6
.20
2.40
±0.
02
0
.12
5.1
82.
57±
0.01
0.0
7 2
.92
2.40
±0.
02
0
.12
5.1
92.
30±
0.01
0.0
8 3
.61
2.00
±0.
01
0
.08
4.1
52.
31±
0.02
0.1
2 5
.13
C-19
8ST
-Rlin
HST
-310
0.72
±0.
01
0
.07
9.9
3 0.
82±
0.02
0.10
11.
80
0.8
0±0.
01
0
.08
9.8
50.
92±
0.02
0.
14 1
5.18
0.8
1±0.
03
0.15
18
.75
0
.90±
0.02
0.1
3 1
4.58
1.
40±
0.01
0.0
7 5
.14
0.90
±0.
01
0
.08
8.7
91.
00±
0.01
0.0
8 7
.88
1.01
±0.
01
0
.07
6.8
70.
91±
0.01
0.0
7 7
.63
0.72
±0.
02
0
.09
12.3
1
Tabe
la 1.
Sre
dnje
vre
dnos
ti i p
okaz
atel
ji va
rijab
ilnos
ti za
deb
ljinu
stab
la (c
m)
Tabl
e 1.
Mea
n va
lues
and
indi
cato
rs o
f va
riabi
lity
for s
tem
thick
ness
(cm
)

Način nasleđivanja debljine stabla F1 hibrida krmnog sirka 101
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
SS-1
646
x C
-198
SS-1
648
x C
-198
SS-1
688
x C
-198
S-8
682
x C-
198
P-21
656
x C
-198
P-74
633
x C
-198
P-17
0 63
0 x
C-19
8P-
174
621
x C-
198
SS-1
646
x S
T-R
lin H
SS-1
648
x S
T-R
lin H
SS-1
688
x S
T-R
lin H
S-8
682
x ST
-R li
n H
P-21
656
x S
T-R
lin H
P-74
633
x S
T-R
lin H
P-17
0 63
0xST
-R li
n H
1.44
±0.
02 p
d- 0
.13
8.8
51.
58±
0.02
pd+
0.
13
7.9
7 1.4
3±0.
02
pd-
0.13
9.
211.
50±
0.02
i
0.1
3 8
.68
1.69
±0.
02 p
d+ 0
.12
7.3
51.
40±
0.02
i
0.1
3 9
.38
1.37
±0.
02 i
0.10
7
.63 1
.29±
0.02
pd
- 0.
13
9.72
1.31
±0.
02 p
d- 0
.13
10.4
81.
40±
0.02
i
0.1
2 8
.87
1.41
±0.
02 p
d- 0
.13
8.9
91.
49±
0.02
i
0.1
2 7
.75
1.27
±0.
03 p
d- 0
.14
11.4
21.
46±
0.03
i
0.1
4 9
.91
1.22
±0.
02 p
d- 0
.12
10.1
4
1.53
±0.
03 h
+
0.17
11.
401.
34±
0.02
h+
0.
12
8.8
91.
39±
0.02
pd+
0.1
4 9
.97
1.40
±0.
02 i
0.1
3 9
.38
1.43
±0.
02 h
+
0.13
9.
401.
40±
0.02
d+
0.
13
8.9
91.
36±
0.02
pd+
0.0
9 6.
801.
20±
0.01
i
0.08
6.
391.
21±
0.02
d+
0
.13
10.4
21.
32±
0.02
d+
0
.13
9.6
21.
42±
0.02
pd+
0.
13
9.13
1.38
±0.
03 i
0.1
4 10
.24
1.01
±0.
02 i
0.0
9 9
.25
1.22
±0.
02 i
0.1
3 10
.37
1.21
±0.
02 i
0.1
3 1
0.65
2.33
±0.
02 h
+
0.1
0 4
.39
2.00
±0.
02 h
+
0.1
3 6
.56
2.62
±0.
02 h
+
0.1
2 4
.61
1.98
±0.
02 h
+
0.1
3 6
.36
2.33
±0.
02 h
+
0.1
0 3
.90
2.02
±0.
02 h
+
0.1
0 4
.76
1.90
±0.
02 h
+
0.1
3 7
.05
1.79
±0.
02 h
+
0.1
3 7
.45
1.93
±0.
02 h
+
0.1
2 6
.26
1.72
±0.
02 h
+
0.1
0 6
.03
2.31
±0.
02 h
+
0.1
1 4
.95
1.89
±0.
02 h
+
0.1
2 6
.35
2.00
±0.
02 h
+
0.1
3 6
.37
1.78
±0.
02 h
+
0.1
2 6
.92
1.84
±0.
02 h
+
0.1
0 5
.64
1.59
±0.
01 p
d-
0.07
4.
481.
59±
0.01
i
0.
07
4.34
1.
50±
0.01
pd-
0.
07
4.47
1.59
±0.
01 p
d-
0.07
4.
161.
61±
0.02
i
0.
13
8.16
1.39
±0.
01 p
d-
0.0
6 4
.19
1.71
±0.
02 p
d+
0.11
6.69
1.10
±0.
01 d
- 0
.07
6.0
61.
31±
0.01
pd-
0
.06
4.46
1.01
±0.
01 d
- 0
.05
5.1
71.
20±
0.01
pd-
0
.07
5.5
61.
51±
0.01
pd-
0
.05
3.4
61.
10±
0.01
d -
0.0
6 5
.04
1.00
±0.
01 d
- 0
.06
6.1
71.
30±
0.01
pd-
0
.07
5.7
1
Gen
otip
G
enot
ype
Deb
ljina
stab
la /
Stem
thick
ness
Rim
ski Š
anče
vi 2
002
Mač
vans
ki P
rnjav
or 2
002
Rim
ski Š
anče
vi 2
003
Mač
vans
ki P
rnjav
or 2
003
x
± s
x
S
V
x
± s
x
S
V
x
± s
x
S
V
x
± s
x
S
V
P-17
4 62
1xST
-R li
n H
SS-1
646
x S
T-31
0 SS
-1 6
48 x
ST-
310
SS-1
688
x S
T-31
0S-
8 68
2 x
ST-3
10P-
21 6
56 x
ST-
310
P-74
633
x S
T-31
0P-
170
630
x ST
-310
P-17
4 62
1 x
ST-3
10
1.26
±0.
02 p
d-
0.09
7.
731.
50±
0.02
pd-
0.
12
7.91
1.32
±0.
02 p
d-
0.12
9.
371.
38±
0.02
pd-
0.
13
9.40
1.40
±0.
02 p
d-
0.12
8.
671.
02±
0.02
pd-
0.
13 1
2.71
1.40
±0.
02 p
d-
0.13
9.
261.
31±
0.02
i
0.0
9 7
.13
1.
38±
0.02
pd-
0.
14
9.88
1.21
±0.
02 i
0
.13
10.
101.
31±
0.02
h+
0.
14 1
0.67
1.20
±0.
02 d
+
0.12
10.
031.
19±
0.02
i
0.1
4 1
1.42
1.30
±0.
02 i
0
.13
10.
021.
09±
0.02
i
0.1
3 1
1.50
1.04
±0.
02 p
d-
0.13
12.
251.
33±
0.03
i
0.1
6 1
2.03
1.22
±0.
02 i
0
.13
11.
02
1.86
±0.
02 h
+
0.1
0 5
.42
1.87
±0.
02 h
+
0.1
2 6
.56
1.50
±0.
02 h
+
0.1
1 7
.43
1.90
±0.
02 h
+
0.1
2 6
.18
1.91
±0.
02 d
+
0.1
1 5
.78
2.11
±0.
02 h
+
0.1
3 6
.35
1.51
±0.
02 d
+
0.1
1 7
.45
2.19
±0.
02 h
+
0.1
2 5
.54
1.97
±0.
02 h
+
0.1
2 6
.23
1.06
±0.
01 d
- 0
.08
7.
261.
30±
0.01
pd-
0.0
7
5.16
1.40
±0.
01 i
0
.07
5.
451.
50±
0.01
i
0.0
7
4.78
1.63
±0.
02 i
0
.10
6.
031.
54±
0.02
i
0.1
0
6.05
1.41
±0.
02 i
0
.12
8.
191.
33±
0.02
i
0.1
0
7.19
1.43
±0.
02 i
0
.10
6.
62
L
SD 0
.05
0.0
10.
030.
040.
040.
050.
020.
030.
020.
03

Pataki I102
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
tmetičke sredine (SE), standardna devijacija (S) i koefi cijent varijacije (V) prema Hadživukoviću (1991). Za ocenjivanje načina nasleđivanja prime-njen je test značajnosti (t-test) srednjih vrednosti hibrida u odnosu na roditeljski prosek. Za ocenu kombinacionih sposobnosti stranooplodnih vrsta korišćen je metod linija x tester, koji je predložio Kempthorne (1957), a primenili Singh & Chaud-hary (1976). Metod linija x tester rađen je u progra-mu Excel. Analizom varijanse dobijene su infor-macije o značajnosti razlika između izvora varija-cije, opštim kombinacionim sposobnostima linija, posebnim kombinacionim sposobnostima hibrida, komponentama genetičke varijanse, prosečnom doprinosu linija, testera i njihove interakcije.
Rezultati i diskusija
Kod ispitivanih inbred linija sirka za zrno i sudanske trave i njihovih hibrida ustanovljene su značajne razlike u srednjim vrednostima za de-bljinu stabla, što ukazuje na postojanje genetičkih razlika između genotipova (Tab. 1).
Srednje vrednosti za debljinu stabla kod linija u prvoj godini ispitivanja bile su značajno veće na lokalitetu Rimski Šančevi u odnosu na lokalitet Mačvanski Prnjavor, a u drugoj godini ispitivanja bile su veće na lokalitetu Mačvanski Prnjavor. Srednje vrednosti linija u prvoj godini ispitivanja na lokalitetu Rimski Šančevi, i srednje vrednosti u drugoj godini ispitivanja na lokalitetu Mačvanski Prnjavor su približne ili su iste (SS-1 648, P-21 656, P-74 633, P-170 630, P-174 621). Srednje vrednosti linija (SS-1 646, SS-1 688, S-8 682, P-21 656, P-74 633 i dr.) u prvoj godini ispi-tivanja na lokalitetu Mačvanski Prnjavor, i srednje vrednosti u drugoj godini ispitivanja na lokalitetu Rimski Šančevi su takođe slične (Tab. 1). Najveća srednja vrednost za ovo svojstvo je izmerena kod linije S-1 646 (2,63 cm) u prvoj godini ispitivanja na lokalitetu Rimski Šančevi, a najniža kod linije P-21 656 (1,10 cm) na lokalitetu Mačvanski Pr-njavor u istoj godini ispitivanja (Tab. 1).
Srednje vrednosti testera za debljinu stabla su bile veće na lokalitetu Mačvanski Prnjavor u pr-voj, a na lokalitetu Rimski Šančevi u drugoj godi-ni ispitivanja. Najveća debljina stabla je izmerena kod testera C-198 (1,40 cm) u drugoj godini ispi-tivanja na lokalitatu Rimski Šančevi, dok su naj-manju srednju vrednost imali testeri C-198 (0,72 cm) u prvoj godini ispitivanja na lokalitetu Rim-ski Šančevi i ST-310 u drugoj godini ispitivanja na lokalitetu Mačvanski Prnjavor. Tester ST- R lin H na oba lokaliteta i u svim godinama ispitivanja imao je srednje vrednosti od 0,81 cm do 0,91 cm, što pokazuje najveću stabilnost za debljinu stabla.
Testeri za debljinu stabla imaju niže vrednosti u odnosu na linije, a razlike u pojedinim godinama i lokalitetima su manje (Tab. 1), što je u saglasnosti sa rezultatima Barabas et al. (1985), Tiwari et al. (2003), Zhan Quin Wen i sar. (2004).
Debljina stabla hibridnih kombinacija u prvoj i u drugoj godini ispitivanja je veća na lokalite-tu Rimski Šančevi, s tim da su razlike u srednjim vrednostima manje po lokalitetima u prvoj godini merenja. Najveća srednja vrednost u prvoj godi-ni ispitivanja je zabeležena kod hibrida P-21 656 x C-198 (1,69 cm) na lokalitetu Rimski Šančevi, dok je najmanja zabeležena kod hibrida P-21 656 x ST R lin H (1,01 cm) na lokalitetu Mačvanski Prnjavor. U drugoj godini ispitivanja najveća sred-nja vrednost (2,62 cm) zabeležena je kod hibrida SS-1 688 x C-198 na lokalitetu Rimski Šančevi, a najmanja (1,00 cm) kod hibrida P 74 633 x ST- R lin H u lokalitetu Mačvanski Prnjavor (Tab. 1).
Najveća vrednost standardne devijacije je izračunata kod linije SS-1 688 (0,21) u prvoj go-dini ispitivanja na lokalitetu Rimski Šančevi. Ma-nje vrednosti (0,05) ovog obeležja su izračunate u drugoj godini ispitivanja na lokalitetu Mačvan-ski Prnjavor kod testera i hibridnih kombinacija (Tab. 1). Koefi cijent varijacije kao relativni poka-zatelj varijacije bio je najveći kod testera ST-R lin H (V=18,75%) u prvoj godini ispitivanja na loka-litetu Mačvanski Prnjavor, a najmanji kod linije S-8 682 (V=2,92%) u drugoj godini ispitivanja na lokalitetu Mačvanski Prnjavor (Tab. 1).
U prvoj godini ispitivanja na lokalitetu Ma-čvanski Prnjavor u F1 generaciji u nasleđivanju debljine stabla kod petnaest ukrštanja ispoljila se parcijalna dominacija roditelja manje srednje vrednosti, kod sedam intermedijarnost a kod dve hibridne kombinacije parcijalna dominacija (Tab. 1). U prvoj godini ispitivanja na lokalitetu Ma-čvanski Prnjavor u nasleđivanju debljine stabla ispoljila se parcijalna dominacija roditelja manje srednje vrednosti kod jednog, intermedijarnost kod dvanaest, parcijalna dominacija roditelja veće srednje vrednosti kod tri, dominacija roditelja veće srednje vrednosti kod četiri i superdomina-cija kod četiri kombinacije ukrštanja (Tab. 1).
U drugoj godini ispitivanja na lokalitetu Rim-ski Šančevi u F1 generaciji dominacija roditelja veće srednje vrednosti ispoljila se kod dve, a su-perdominacija kod dvadesetdve hibridne kom-binacije. Na lokalitetu Mačvanski Prnjavor u F1 generaciji u nasleđivanju ovog svojstva ispoljila se dominacija roditelja manje srednje vrednosti kod pet, parcijalna dominacija roditelja manje srednje vrednosti kod devet, intermedijarnost kod devet i parcijalna dominacija roditelja veće srednje vred-nosti kod jedne hibridne kombinacije (Tab. 1).

Način nasleđivanja debljine stabla F1 hibrida krmnog sirka 103
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
Analiza varijanse linija x tester za debljinu sta-bla pokazuje, da su sredine kvadrata za sve izvore varijacije visoko značajne, osim za ponavljanja na oba lokaliteta i u obe godine ispitivanja (Tab. 2).
Analiza opštih kombinacionih sposobnosti za debljinu stabla pokazala je visoko značajne razlike između inbred linija majki, izuzev linije SS-1 648 u prvoj godini ispitivanja na lokalitetu Mačvanski Prnjavor i linije P-170 630 u drugoj godini ispiti-vanja na lokalitetu Rimski Šančevi (Tab. 3). Treba napomenuti da je selekcija na bazi OKS za svoj-stvo debljina stabla krmnog sirka orijentisana na genotipove sa što većim pozitivnim efektom. Ne-gativne vrednosti OKS u ispitivanim godinama i
Izvo
r var
ijaci
je
Sour
ce o
f va
riatio
nSt
epen
iSl
obod
e D
egre
es
of fr
eedo
m
Ana
liza
varij
anse
za
debl
jinu
stab
la /
Ana
lysis
of
varia
nce
for s
tem
thick
ness
Rim
ski Š
anče
vi 2
002
Mač
. Prn
javor
200
2Ri
msk
i Šanče
vi 2
003
Mač
. Prn
javor
200
3Sr
edin
a
Fe
kvad
rata
M
ean
squa
res
Sred
ina
F
ekv
adra
ta
Mea
n sq
uare
s
Sred
ina
F
ekv
adra
ta
Mea
n sq
uare
s
Sred
ina
F
ekv
adra
ta
Mea
n sq
uare
sPo
navl
janja
/ Re
petit
ions
Tr
etm
ani /
Tre
atm
ents
Rodi
telji
/ P
aren
tsRo
d. p
rem
a uk
ršt.
/ Pa
rent
s:cro
ssin
gsU
kršt
anja
/ Cr
ossin
gsLi
nije
/ L
ines
Test
eri /
Tes
ters
Lini
je x
test
eri /
Lin
es x
Tes
ters
Pogr
eška
/ E
rror
2 34 10 1 23 7 2 14 68
0.00
09
1.95
0.66
60
1436
.56
**1.
5815
34
11.3
4 **
5.62
94
1
2132
.33
**0.
0521
1
12.4
8 **
0.03
65
7
8.70
**
0.10
81
233
.28
**0.
0519
1
12.1
1 **
0.00
04
0.00
01
0
.19
0.13
44
268
.32
**0.
3431
6
84.7
6 **
0.00
52
10
.34
**0.
0493
98.4
8 **
0.04
26
84
.99
**0.
1921
3
83.3
8 **
0.03
23
64
.52
**0.
0005
0.00
01
0.47
0.
4314
16
64.7
3 **
0.27
94
1078
.23
**7.
4324
297
29.6
0 **
0.19
32
745
.34
**0.
2975
11
47.9
7 **
0.42
41
1636
.47
**0.
1080
4
16.7
2 **
0.00
025
0.00
04
2.1
00.
7264
3
544.
27 *
*1.
4656
7
150.
63 *
*1.
0273
5
136.
50 *
*0.
1362
664.
81 *
*0.
1224
597.
36 *
*0.
7216
3
520.
59 *
*0.
0595
290.
56 *
*0.
0002
Tota
l10
4
Tabe
la 2
. A
naliz
a va
rijan
se li
nije
x te
ster
i za
debl
jinu
stab
laTa
ble
2. A
naly
sis o
f va
rianc
e lin
es x
test
ers f
or st
em th
ickne
ss
Gen
otip
Gen
otyp
eO
KS
za d
eblji
nu st
abla
/ G
CA fo
r ste
m th
ickne
ssRi
msk
i Š. 2
002
Mač
v. P.
200
2Ri
msk
i Š. 2
003
Mač
v. P.
2003
SS-1
646
SS-1
648
SS-1
688
S-8
682
P-21
656
P-74
633
P-17
0 63
0P-
174
621
0.03
4 **
0.04
9 **
0.02
1 *
0.07
8 **
-
0.05
9 **
0.03
7 **
-
0.08
8 **
-
0.07
4 **
0.0
69 *
*
0
.006
0
.054
**
0.0
80 *
*
-0.
100
**
-0.
060
**
0.
021
*
-0.
071
**
0.07
5 **
-
0.22
8 **
0.30
5 **
-
0.03
9 **
0.17
4 **
-
0.19
8 **
0.
007
-
0.09
7 **
0.02
0 **
-
0.04
4 **
0.02
3 **
0.19
6 **
0.03
9 **
-
0.12
1 **
0.06
9 **
-
0.18
1 **
C-19
8ST
-Rlin
HST
-310
0.07
7 **
-
0.03
3 **
-
0.04
4 **
0.10
1 **
-
0.03
2 **
-0.
069
**
0.15
1 **
-
0.05
3 **
-
0.09
8 **
0.13
2 **
-
0.19
6 **
0.06
4 **
SE O
KS
lin.
SE(G
i-Gj)l
in.
SE O
KS
test
.SE
(Gi-G
j) te
st.
0.02
1
0.
010
0.03
5
0.
006
0.02
2
0.
010
0.03
6
0.
006
0.01
6
0.
007
0.02
6
0.
004
0.01
4
0.
005
0.02
3
0.
004
LSD
lin
0
.05
0
.01
LSD
tes
t. 0.
05
0.0
1
0.02
0
0.
027
0.01
2
0.
016
0.02
1
0.
028
0.01
3
0.
017
0.01
5
0.
020
0.00
9
0.
012
0.01
0
0.
013
0.00
8
0.
011
Tabe
la 3.
Vre
dnos
ti O
KS
inbr
ed li
nija
za d
eblji
nu st
abla
Tabl
e 3.
Inbr
ed li
nes G
CA v
alues
for s
tem
thick
ness

Pataki I104
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
lokalitetima imala je linija P-174 621. Negativne vrednosti OKS na oba lokaliteta u prvoj godini imala je linija P-21 656, a u drugoj godini ispitiva-nja linije SS-1 648 i P-74 633. Pozitivne i negativ-ne vrednosti OKS u prvoj godini ispitivanja na različitim lokalitetima imala je linija P-74 633, a u drugoj linija S-8 682. Pozitivne vrednosti OKS u prvoj godini na oba lokaliteta imala je linija SS-1 648, a u drugoj linije P-21 656 i P-170 630. Pozi-tivne vrednosti OKS izračunate su za obe godine i oba lokaliteta za linije SS-1 646 i SS-1 688, što znači da su one za selekciju na svojstvo debljina stabla najbolje.
U godinama ispitivanja i lokalitetima teste-ri su pokazali visoko značajne razlike za OKS. Negativne vrednosti OKS imali su testeri ST-R lin H i ST-310. Pozitivne vrednosti OKS po go-dinama i lokalitetima imao je tester C-198, te se može smatrati najboljim opštim kombinatorom od ispitivanih testera za svojstvo debljina stabla (Tab. 3).
Ispitivanje PKS linija u velikoj većini pokazu-je visoko značajne razlike sa pozitivnim i nega-
tivnim predznakom (Tab. 4). Pozitivne vrednosti PKS u svim godinama i lokalitetima ispitivanja imale su hibridne kombinacije P-21 656 x C-198 gde je linija u prvoj godini na oba lokaliteta imala negativan OKS, a u drugoj godini pozitivan OKS na oba lokaliteta i tester sa pozitivnom OKS, hi-brid P-174 621 x ST-310 gde su linija i tester sa negativnom OKS, zatim hibrid S-8 682 x ST-R lin H gde je linija sa pozitivnom OKS i testerom sa negativnom OKS. Interesantno je da su sedam hibridnih kombinacija koje su u toku dve godine ispitivanja i na dva lokaliteta posmatranja imale tri pozitivne vrednosti PKS i jednu sa negativnim predznakom. Pet hibrida su sa roditeljima sa po-zitivnom i negativnom vrednostima OKS, jedan sa roditeljima sa pozitivnom OKS i jedan hibrid sa roditeljima sa negativnom OKS.
Kod tri hibridne kombinacije S-8 682 x C-198, P-174 621 x C-198 i P-21 656 x ST-R lin H izra-čunate su negativne PKS u toku dve godine ispi-tivanja na dva lokaliteta (Tab. 4), od kojih jedan hibrid ima roditelje sa pozitivnim vrednostima OKS, jedan hibrid ima roditelje sa pozitivnim i
Genotip Genotype
PKS za debljinu stabla / SCA for stem thicknessRimski Š. 2002 Mačv. P. 2002 Rimski Š. 2003 Mačv. P. 2003
SS-1 646 x C-198SS-1 648 x C-198SS-1 688 x C-198S-8 682 x C-198P-21 656 x C-198P-74 633 x C-198P-170 630 x C-198P-174 621 x C-198SS-1 646xST-R lin HSS-1 648xST-R lin HSS-1 688xST-R lin HS-8 682xST-R lin HP-21 656xST-R lin HP-74 633xST-R lin HP170 630xST-R lin HP174 621xST-R lin HSS-1 646 x ST-310SS-1 648 x ST-310SS-1 688 x ST-310S-8 682 x ST-310P-21 656 x ST-310P-74 633 x ST-310P-170 630 x ST-310P-174 621 x ST-310
-0.053 ** 0.072 ** -0.054 ** -0.044 * 0.287 ** -0.099 **
-0.014 -0.095 ** -0.776 **
0.002 0.036 * 0.059 **
-0.026 0.074 ** -0.047 **
-0.022 0.129 ** -0.073 **
0.018-0.015
-0.261 **0.025
0.061 ** 0.117 **
0.080 **-0.050 **-0.048 **-0.061 ** 0.152 ** 0.079 **
-0.039 * -0.113 ** -0.110 ** 0.067 ** 0.119 ** 0.055 ** -0.134 **
0.029 -0.062 ** 0.037 *
0.030 -0.017
-0.071 ** 0.005 -0.018
-0.108 ** 0.101 ** 0.077 **
0.134 ** 0.108 ** 0.191 ** -0.098 ** 0.032 * 0.098 ** -0.228 ** -0.236** -0.061 ** 0.035 ** 0.085 **
0.016 -0.094 ** 0.065 ** -0.084 ** 0.037 ** -0.073 ** -0.143 ** -0.276 ** 0.082 ** 0.062 ** -0.163 ** 0.311 ** 0.199 **
0.060 ** 0.127 ** -0.036 ** -0.116 ** 0.058 **
0.005 0.128 ** -0.225 ** 0.105 ** -0.131 **
-0.001 0.129 ** -0.117 ** -0.094 ** 0.049 ** 0.060 ** -0.165 ** 0.044 ** 0.038 **
-0.012 0.059 ** 0.089 ** -0.178 ** 0.165 **
SE PKS (SCA)SE (Sij-Ski)
0.0120.017
0.013 0.018
0.009 0.013
0.008 0.012
LSD 0.05 0.01
0.0350.046
0.036 0.048
0.026 0.035
0.024 0.032
Tabela 4. Vrednosti PKS hibrida za debljinu stablaTable 4. Hybrid GCA values for stem thickness

Način nasleđivanja debljine stabla F1 hibrida krmnog sirka 105
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
negativnim OKS i jedan hibrid sa oba roditelja sa negativnim OKS.
Analiza komponenata genetičke varijanse po-kazuje da u nasleđivanju debljine stabla kod F1 hibridnih kombinacija krmnog sirka u ispitivanim godinama i lokalitetima, važniju ulogu imaju geni sa neaditivnim delovanjem, koja uključuje do-minaciju i epistazu, a što potvrđuje i izračunata vrednost odnosa OKS/PKS (Tab. 5). Ovi rezul-tati su u saglasnosti sa rezultatima autora Zhan et al. (2004) i Grewal et al. (2003).
U drugoj godini ispitivanja izračunate vred-nosti aditivne varijanse su nešto veće u odnosu na prvu godinu, ali je aditivna varijansa oko dva puta manja u odnosu na dominantnu varijansu.
U prvoj godini ispitivanja najveći doprinos u ekspresiji debljine stabla F1 hibrida na oba lokaliteta imala je interakcija inbred linija majki sa testerima, a doprinos linija i testera po loka-litetima je različit. Na lokalitetu Rimski Šančevi doprinos linija bio je veći od doprinosa testera, dok su na lokalitetu Mačvanski Prnjavor nešto veći doprinos imali testeri. U drugoj godini ispi-tivanja na lokalitetu Rimski Šančevi najveći do-prinos pripada linijama, pa njihovim interakcija-ma sa testerima, a najmanji testerima, dok su na lokalitetu Mačvanski Prnjavor najveći doprinos imali testeri. Linije i interakcija linije x testeri imali su jednak doprinos u ekspresiji debljine stabla (Tab. 6).
Zaključak
Na osnovu dobijenih rezultata istraživanja u ovom radu mogu se doneti sledeći zaključci:
Između inbred linija (majki i testera) i hibrida F1 generacije postojale su značajne razlike u sred-njim vrednostima za debljinu stabla.
Za debljinu stabla ustanovljeni su sledeći na-čini nasleđivanja: parcijalna dominacija roditelja niže srednje vrednosti, intermedijarnost, parcijal-na dominacija i dominacija roditelja veće srednje vrednosti i superdominacija.
Rezultati ispitivanja kombinacionih sposob-nosti pokazuju da je glavnu ulogu u nasleđivanju debljine stabla imala neaditivna komponenta, što se vidi iz analize komponenti genetske varijanse. To potvrđuju odnosi OKS/PKS u F1 generaciji koji su manji od jedan.
Na osnovu vrednosti opštih kombinacionih sposobnosti odabrane su linije sa najboljim OKS za debljinu stabla SS-1 646, SS-1 688 i S-8 682.
U toku ispitivanja pozitivne značajne vredno-sti PKS imali su hibridi: S-8 682 x ST-R lin H i P-21 656 x C-198.
U prvoj godini ispitivanja najveći doprinos u ekspresiji debljine stabla F1 hibrida na oba loka-liteta imala je interakcija inbred linija majki sa te-sterima, a u drugoj godini ispitivanja na lokalitetu Rimski Šančevi linije, dok na lokalitetu Mačvan-ski Prnjavor testeri.
Tabela 5. Komponente genetičke varijanse za debljinu stablaTable 5. Components of genetic variance for stem thickness
Rimski Š. 2002 Mačv. P. 2002 Rimski Š. 2003 Mačv. P. 2003
Aditivna varijansa / Additive variance
Va (F=0)Va (F=1)OKS / GCA
0.00060.00080.0004
0.00690.00350.0017
0.02980.01490.0074
0.02900.01450.0072
Dominantna varijansa / Dominant variance
Vd (F=0)Vd (F=1)PKS / SCA
0.05150.01290.0129
0.03180.00790.0079
0.10770.02690.0269
0.05930.01480.0148
OKS/PKSGCA/SCA
0.0320 0.2185 0.2764 0.4883
Tabela 6. Doprinos linija, testera i njihova interakcija u ekspresiji debljine stabla (%)Table 6. Contribution of lines, testers and their interaction in expression of stem thickness (%)
Prosečan doprinos Average contribution
Ekspresija debljina stabla / Expression of stem thickness
Rimski Š. 2002 Mačv. P. 2002 Rimski Š. 2003 Mačv.P. 2003
Linija / LineTester / TesterLinija x tester / Line x tester
21.2918.0460.67
26.2733.8539.88
46.8819.0934.03
27.3546.0526.60

Pataki I106
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 99-106
Literatura
Barabás Z, Bányai L (1985): A cirok és a szudáni fű Sorghum bicolor (L.) Moench, S. sudanense (Piper) Stapf. Magyarország kulturfl órája, Akadémiai kiadó, Budapest
Berenji J (1988): Produkciona sposobnost hibrida tipa sirak x sudanska trava. VI. Jugoslovenski simpozij o krmnom bilju, Osijek 151-159
Grewal R P, Rajesh S Y, Pahuja S K (2003): Inheritance of fod-der yield and its components in multicut forage sorghum (Sorghum bicolor (L.) Moench). Forage Res. 29: 142-144
Hadživuković S (1991): Statistički metodi s primenom u poljopri-vrednim i biološkim istraživanjima. Poljoprivredni fakultet, In-stitut za ekonomiku poljoprivrede i sociologiju sela, Novi Sad
Kempthorne O (1957): An introduction to genetic statistics, John Wiley and sons, Inc. New York.
Ravindrababu Y, Pathak A R, Tank C J (2002): Studies on heterosis over environments in sorghum. Crop Res. 24: 90-92
Singh R K, Chaudhary B D (1976): Biometrical methods in quantitative genetic analysis. Kalyani Publishhers, Ludhiana, New Delhi
Swanson A F, Parker J H (1931): Inheritance of smut resistance and juicness of stalk in the sorghum cross red amber x fete-rita. J. Hered. 22: 51-56
Tiwari D K, Gupta R S, Mishra R (2003): Study of heritability and genetic advance in grain sorghum (Sorghum bicolor (L.) Moench), Plant Archives 3: 181-182
Zhan Q W, Qian Z Q (2004): Heterosis utilization of hybrid between sorghum (Sorghum bicolor (L.) Moench) and Sudan-grass (Sorghum sudanense (Piper) Stapf). Acta Agron. Sinica 30: 73-77
Combining Ability and Mode of Inheritance of Stem Thickness in Forage Sorghum (Sorghum Bicolor L. Moench) F1 Hybrids
Imre Pataki
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Aim of this research was determination of mode of inheritance, gene effects, components of genetic variance, combining abilities, average contribution of lines and testers and their interactions in expression of stem thickness in forage sorghum F1 generation. Method line x tester was applied. Material comprised of eight geneti-cally divergent A-inbred lines of grain sorghum, three R lines-testers of Sudan grass and twenty-four F1 hybrids obtained by crossing lines with testers. Among tested genotypes there were signifi cant differences in mean values of stem thickness. Analysis of variance of combining abilities showed that there were highly signifi cant differences for general combining abilities (GCA) and specifi c combining abilities (SCA); non-additive component of genetic variance (dominance and epistasis) had greater portion in total genetic variance for stem thickness. During the fi rst research year, interaction between inbred maternal line with testers had the largest contribution in expression of stem thickness of F1 hybrid at both locations, while in the second year at location Rimski Šančevi the largest contribution belongs to lines and at location Mačvanski Prnjavor the largest contribution belongs to testers. Asse-ssment of combining abilities showed that these inbred lines of grain sorghum can be used as mothers: SS-1 646, SS-1 688 and S-8 682 in breeding forage sorghum for thicker stem. According to SCA, promising forage sorghum hybrids are S-8 682 x ST-R lin H and P-21 656 x C-198. This research can be of importance for developing new high-yielding forage sorghum hybrids.Key words: combining abilities, components of genetic variance, forage sorghum, mode of inheritance of stem thickness, stem thickness, Sudan grass

Komponente prinosa konoplje za vlakno 107
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112originalni naučni rad / original research article
Varijabilnost i međuzavisnost komponentiprinosa konoplje za vlakno
Vladimir Sikora · Janoš Berenji · Dragana Latković
primljeno / received: 28.10.2010. prihvaćeno / accepted: 27.12.2010.© 2011 IFVC
Izvod: U poljskim ogledima je ispitivana varijabilnost i međuzavisnost komponenti prinosa konoplje za vlakno. U istraživanja je bilo uključeno 20 sorti koje se danas komercijalno gaje u Evropi. Varijabilnost visine biljke, prinosa stabla, sadržaja vlakna i prinosa vlakna je u ispitivanom materijalu značajna i on može poslužiti kao dobra osnova za dalji rad na oplemenjivanju i unapređenju proizvodnje konoplje za vlakno. Istraživanja su pokazala da je debljina stabla u prvom redu determinisana uslovima spoljne sredi-ne a da je genetski faktor od manjeg uticaja na ekspresiju ovog svojstva. Kod ostalih analiziranih osobina izraženiji je značaj nasledne osnove u odnosu na uslove sredine. Korelacije između analiziranih osobina ukazuju da se veći prinosi stabla i vlakna postižu u porastima veće visine i debljeg stabla. Sa povećanjem debljine stabla povećava se i sadržaj vlakna u stablu.Ključne reči: komponente prinosa, konoplja za vlakno, međuzavisnost
V. Sikora · J. BerenjiInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, SrbijaD. LatkovićPoljoprivredni fakultet, Univerzitet u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Konoplja kao sirovina za proizvodnju prirod-nih vlakana spada među najstarije gajene biljne vrste. Tokom šezdesetih godina prošlog veka površine pod konopljom u svetu su se značajno smanjile, pre svega zbog pojave oštre konkuren-cije u vidu pamučnog vlakna i sintetičkih vlakana. Pored toga, kao posledica borbe protiv korišćenja marihuane u većini zapadnih država zabranjeno je gajenje i industrijske konoplje. Poslednjih go-dina je u okviru potrage za alternativnim kultu-rama obnovljen interes za konopljom iz nekoliko razloga: relativno manje potrebe za hemijskim inputima u poređenju sa intenzivnim kulturama, solidna rezistentnost prema patogenima i pove-ćanje javne tražnje za prirodnim vlaknima (Mon-tford & Small 1999). Tehnologija proizvodnje i industrijska primena konoplje su u zadnje vreme postali predmet istraživačkih projekata širom sve-ta (Cromack1998, Ranalli 2002, Amaducci 2003, Müssig & Martens 2003, Karus & Vogt 2004).
Sadašnji sortiment industrijske konoplje gajene za vlakno obuhvata populacije slobodne oplod-nje, selekcionisane populacije, kao i F1 i F2 hibri-de širokog stepena genetičke varijabilnosti (Bócsa & Karus 1998).
Istraživanje je bilo usmereno na ispitivanje vari-jabilnosti i međuzavisnosti komponenti prinosa u kolekciji konoplje za vlakno, sa ciljem utvrđivanja širine genetičke osnove za dalji rad na oplemenji-vanju i unapređenju njene proizvodnje.
Materijal i metod rada
Poljski ogledi su zasnovani 2005. godine u Bačkom Petrovcu. Zemljište na kom je ogled postavljen je srednjeduboki karbonatno oglejeni černozem na lesu i lesolikim sedimentima. Sadr-žaj organske materije u zemljištu je bio 2,85%, pH u H2O 8,66, P2O5 46,4 mg na 100g a K2O 57,0 mg na 100g. Pre setve su aplikovana azotna mineralna đubriva u količini od 60 kg ha-1, što uz postojeći sadržaj ukupnog azota od 0,204% pred-stavlja optimalan nivo makroelemenata za gaje-nje konoplje u oblasti u kojoj je ogled postavljen (Starčević 1996, Amaducci et al. 2002a).
Ogled je obuhvatao dvadeset sorti industrijske konoplje (Tab. 1) koje se danas nalaze u proizvod-nji. Pojedine sorte su posejane po slučajnom blok rasporedu u pet ponavljanja. Ogled je zasnovan po preporuci za gajenje industrijske konoplje za
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Sikora V i sar.108
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
vlakno (Struik et al. 2000). Setva je izvršena me-haničkom sejačicom 12. aprila. Pri setvenoj nor-mi od 60 kg ha-1 i međurednom razmaku od 12,5 cm postignut je sklop od 240 biljaka m-2. Elemen-tarna parcela je bila veličine 5 m2 a žetva je vršena sukcesivno tokom avgusta u fazi punog cvetanja ženskih odnosno jednodomih biljaka (Mediavilla et al. 2001). Nakon što su pokošene 3 cm iznad zemlje, biljke sa pojedinih parcela su radi sušenja ostavljene u krstovima na parceli.
Sa svake parcele je odabrano 20 prosečnih bi-ljaka na kojima je merena debljina (na visini od 30 cm) i ukupna visina stabla.
Nakon merenja prinosa vazdušno suvog stabla na pokretnoj elektronskoj vagi, sa svake elemen-
tarne parcele je uzet uzorak od 30 stabala za ana-lizu sadržaja vlakna. Uzorkovanje je vršeno na taj način što je nakon odstranjivanja donjih 25 cm stabala odsečeno sledećih 20 cm što je predstav-ljalo laboratorijski uzorak.
Odvajanje vlakana od drvenastog dela stabla je vršeno u laboratoriji po metodi Bredemana (1942). Uzorci su najpre izmereni na elektronskoj vagi, a zatim kuvani tokom 1,5 h u 0,35% rastvoru NaOH da bi se omogućilo lako ručno odvajanje vlakna. Odvojena vlakna su isprana tekućom vodom i osušena u sušnici na 105oC. Na elektronskoj vagi je izmerena težina suvih vlakana. Iz odnosa težine kompletnog uzorka i težine vlakana izračunat je procentualni sadržaj vlakna u stablu. Prinos vlakna
Tabela 1. Poreklo kolekcije konoplje za vlaknoTable 1. Origin of fi ber hemp collection
Poreklo / Origin Sorta / VarietyMađarska / Hungary Tiborszálasi, Kompolti sárgaszárűFrancuska / France Fédora, Futura 75, Felina 34, Futura žuta Italija / Italy Chameleon, Dioica 88, Epsilon 68, Carmagnola, CS, Silesia, Fibrinova Rumunija / Romania Lovrin 110, SecuieniRusija / Russia YUSO 11Poljska / Poland Beniko, BialobrzeskiaSrbija / Serbia Novosadska, Novosadska plus
Tabela 2. Srednje vrednosti komponenti prinosa konoplje za vlaknoTable 2. Mean values for yield components in fi ber hemp
Sorta / Variety Visina biljke /Plant height(cm)
Debljina stabla /Stem diameter(cm)
Prinos stabla /Stemyield(t ha-1)
Sadržaj vlakna /Fiber content(%)
Prinos vlakna /Fiber yield(t ha-1)
Fédora 177 5,5 8,8 25,2 2,2Tiborszállasi 225 5,5 14,1 23,2 3,3Chameleon 211 5,6 9,4 34,6 3,3Dioica 88 270 5,3 17,4 31,2 5,4Futura 75 218 5,4 12,4 32,2 3,9Epsilon 68 219 5,0 12,5 27,8 3,5Carmagnola 275 5,2 16,4 21,3 3,5CS 269 5,4 18,1 22,8 4,1Felina 34 187 5,5 9,9 29,7 2,9Fibrinova 260 5,5 19,5 29,5 5,7Futura žuta 207 5,6 10,0 26,6 2,6Kompolti sárgaszárű 225 5,6 12,9 27,2 3,5Lovrin 110 206 5,5 14,3 23,4 3,4YUSO 11 195 5,5 10,6 26,5 2,8Silesia 166 5,5 8,8 30,6 2,7Secuieni 191 5,0 13,4 24,6 3,3Beniko 197 5,1 12,1 31,4 3,8Bialobrzeskia 195 5,4 10,5 30,8 3,2Novosadska 246 5,5 15,2 28,0 4,2Novosadska plus 230 5,5 16,3 37,8 6,1Prosek / Average 218 5,4 13,1 28,2 3,7LSD 0,05 24 0,5 1,5 5,0 0,7 0,01 32 0,6 1,9 6,7 0,9

Komponente prinosa konoplje za vlakno 109
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
po jedinici površine je izračunat množenjem pri-nosa sa sadržajem vlakna (Sankari 2000).
Za obradu podataka je primenjen GEN stati-stički paket, pri čemu su pored analize varijanse izračunati: genotipska, ekološka i fenotipska va-rijansa, koefi cijenti genotipske i fenotipske vari-jacije, heritabilnost u širem smislu i genotipske i fenotipske korelacije između ispitivanih osobina.
Rezultati i diskusija
Varijabilnost visine stabla, prinosa stabla, sadr-žaja vlakna i prinosa vlakna je u ispitivanom mate-rijalu značajna, dok se kod debljine stabla (prosek 5,4 cm) ona kreće u okvirima najmanje značajne razlike (Tab. 2). Selekcija superiornih genotipova u pogledu sadržaja i prinosa vlakna je jedan od najznačajnijih agronomskih faktora unapređenja proizvodnja konoplje za vlakno (Amaducci et al. 2008), te stoga ovakva kolekcija može poslužiti kao dobra osnova za dalji rad na oplemenjivanju i unapređenju njene proizvodnje. Radi proširenja genetske osnove za analizirana, kao i za druga kvantitativna i kvalitativna svojstva, kolekciju bi trebalo obogatiti lokalnim populacijama.
Prosečna visina biljke analiziranog materijala iznosi 218 cm. U tom pogledu su se sa vredno-stima od preko 250 cm istakle italijanske sorte Carmagnola (275 cm), Dioica 88 (270 cm) i Fi-brinova (260 cm). Najveći prinos stabla (preko 15 t ha-1) i vlakna (preko 4 t ha-1) imaju italijanske sorte Fibrinova (19,5 t ha-1 i 5,7 t ha-1), CS (18,1 t ha-1 i 4,1 t ha-1) i Dioica 88 (17,4 t ha-1 i 5,4 t ha-1), kao i Novosadska plus (16,3 t ha-1 i 6,1 t ha-1). Prosečan prinos stabla iznosi 13,1 t ha-1 a vlakna 3,7 t ha-1. Sadržaj vlakna u stablu se kreće
u rasponu od 21,3 % (Carmagnola) do 37,8 % (Novosadska plus) uz prosek od 28,2.
Varijabilnost kvantitativnih svojstava je uslov-ljena naslednom osnovom i faktorima spoljne sredine. Udeo ovih komponenti u ukupnoj varija-ciji je različit za pojedine osobine (Tab. 3). Vred-nosti komponenti fenotipske varijacije pokazuju da je ekspresija većine komponenti prinosa deter-minisana pre svega naslednom osnovom, dok je uticaj faktora spoljne sredine slabiji. Izraženiji uti-caj spoljne sredine u odnosu na naslednu osnovu genotipova je zabeležen jedino kod debljine sta-bla, gde je u ukupnoj fenotipskoj varijansi (σ2
p = 0,16) ekološka varijansa (σ2
e = 0,15) znatno veća u odnosu na genotipsku (σ2
g = 0,01). Fenotipska i genotipska varijansa visine biljke (σ2
p = 5339,01 i σ2
g = 4978,28) su veće od varijansi prinosa stabla (σ2
p = 53,04 i σ2g = 51,71) i sadržaja vlakna (σ2
p = 93,61 i σ2
g = 77,67). Vrednosti varijansi za prinos vlakna su izrazito male (σ2
p = 1,31 i σ2g = 1,01), ali
i kod ove osobine svedoče o izraženijem uticaju genetske osnove na njenu ekspresiju
Genotipski koefi cijent varijacije (Tab. 3) se kre-će od 1,85 % za debljinu stabla do 54,73 % za prinos stabla, dok fenotipski varira u rasponu od 7,41 % za debljinu stabla do 55,43 % za prinos stabla. Kod većine svojstava je razlika između genotipskog i fenotipskog koefi cijenta varijacije mala, što potvrđuje značaj genetske konstitucije genotipova u fenotipskoj manifestaciji ispitivanih svojstava. O relativno malom uticaju genotipa na debljinu stabla konoplje govori značajna razlika između fenotipskog (7,41 %) i genotipskog (1,85 %) koefi cijenta varijacije za ovu osobinu.
Izrazito niska vrednost heritabilnosti u širem smislu za debljinu stabla (6 %) potvrđuje mali
Tabela 3. Genotipska (σ2g), ekološka (σ2
e) i fenotipska (σ2p) varijansa; genotipski (CVg) i fenotipski (CVp)
koefi cijent varijacije i heritabilnost (h2) komponenti prinosa konoplje za vlaknoTable 3. Genotypic (σ2
g), ecologic (σ2e) and phenotypic (σ2
p) variance, genotypic (CVg) and phenotypic (CVp) variation coeffi cient and heritability (h2) for yield components in fi ber hemp
Osobina / Trait
σ2g σ2
e σ2p CVg
(%)CVp(%)
h2
(%)
Visina biljke /Plant height
4978,28 360,73 5339,01 32,37 33,52 0,93
Debljina stabla /Stem diameter
0,01 0,15 0,16 1,85 7,41 0,06
Prinos stabla /Stem yield
51,71 1,33 53,04 54,73 55,43 0,97
Sadržaj vlakna /Fiber content
77,67 15,94 93,61 31,25 34,31 0,83
Prinos vlakna /Fiber yield
1,01 0,30 1,31 27,31 31,10 0,77

Sikora V i sar.110
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
udeo genotipske u ukupnoj fenotipskoj varijabil-nosti (Tab. 3). Kod ostalih komponenti prinosa vrednosti heritabilnosti u širem smislu su izrazito velike i kreću se od 77 % za prinos vlakna do 97 % za prinos stabla.
Za uspešno oplemenjivanje je pored pozna-vanja genetičkog porekla, heritabilnosti, struk-ture genotipske i fenotipske varijabilnosti ma-terijala potrebno poznavanje i međuzavisnosti funkcionalno povezanih svojstava. Zbog toga poseban značaj zauzimaju genotipske korela-cije koje ukazuju na pravac mogućih promena
pod uticajem primenjenih metoda selekcije. Prema prethodnim istraživanjima neki agro-nomski faktori koji nisu bili obuhvaćeni našim ogledima, poput gustine porasta i vremena žetve imaju odlučujuću ulogu u determinaciji proizvodnje vlakna konoplje u smislu kvantite-ta i kvaliteta (Van der Werf et al. 1995, Struik et al. 2000, Mediavilla et al. 2001, Amaducci et al. 2002b) i industrijske primene (Keller et al. 2001).
Iz vrednosti koefi cijenata korelacije se vidi da su u većini slučajeva genotipske korelacije (iznad dijagonale) nešto veće u odnosu na fenotipske (ispod dijagonale) (Tab. 4). Manje vrednosti ge-notipske u poređenju sa fenotipskom korelaci-jom su između prinosa stabla i sadržaja vlakna (rg = -0,032 i rp = -0,034) i prinosa i sadržaja vla-kna (rg = 0,182 i rp = 0,309). Redosled ostalih koefi cijenata fenotipske je sličan redosledu ko-efi cijenata genotipske korelacije i ukazuje na to da je prinos vlakna u najvećoj meri determinisan
visinom biljke (rg = 0,309 i rp=0,273) i prinosom stabla (rg=0,363 i rp=0,350), dok prinos stabla zavisi od visine biljke (rg=0,176 i rp=0,174). De-bljina stabla je u negativnoj korelaciji sa visinom biljke (rg= -0,118 i rp=-0,090), prinosom stabla (rg= -0,209 i rp=-0,148) i prinosom vlakna (rg= -0,200 i rp=-0,109), što u praksi znači da se veći prinosi stabla i vlakna postižu u porastima veće visine i debljeg stabla. Sa povećanjem debljine stabla povećava se i sadržaj vlakna u stablu (rg= 0,193 i rp=0,108).
Zaključak
Varijabilnost visine stabla, prinosa stabla, sadr-žaja vlakna i prinosa vlakna je značajna u ispiti-vanom materijalu konoplje za vlakno i ona može poslužiti kao dobra osnova za dalji rad na ople-menjivanju i unapređenju proizvodnje, s tim da bi postojeću kolekciju trebalo dopuniti lokalnim populacijama.
Na ekspresiju visine stabla, prinosa stabla, sa-držaja vlakna i prinosa vlakna značajniji uticaj ima genetski faktor, dok je debljina stabla u prvom redu determinisana uslovima spoljne sredine.
Veći prinosi stabla i vlakna se postižu u pora-stima veće visine i debljeg stabla. Sa povećanjem debljine stabla takođe se povećava sadržaj vlakna u stablu.
U daljem radu treba obratiti pažnju na uticaj agronomskih faktora poput vremena žetve i gu-stine porasta na komponente prinosa i kvaliteta konoplje za vlakno.
Tabela 4. Genotipske (iznad dijagonale) i fenotipske (ispod dijagonale) korelacije komponenti prinosa konoplje za vlaknoTable 4. Genotypic (above diagonal) and phenotypic (bellow diagonal) correlation for yield components in fi ber hemp
Osobina / Trait
Visina biljke /Plant height
Debljina stabla /Stem diameter
Prinos stabla /Stem yield
Sadržaj vlakna /Fiber content
Prinos vlakna /Fiber yield
Visina biljke /Plant height
-0,118 0,176 -0,035 0,309
Debljina stabla /Stem diameter
-0,090 -0,209 0,193 -0,200
Prinos stabla /Stem yield
0,174 -0,148 -0,032 0,363
Sadržaj vlakna /Fiber content
-0,034 0,108 -0,034 0,182
Prinos vlakna /Fiber yield
0,273 -0,109 0,350 0,309

Komponente prinosa konoplje za vlakno 111
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
Literatura
Amaducci S (2002a): Response of hemp to plant population and nitrogen fertilization. Ital. J. Agron. 6: 103-111
Amaducci S, Errani M, Venturi G (2002b): Plant population effect on fi bre hemp morphology and production. J. Ind. Hemp 7: 33-60
Amaducci S (2003): HEMP-SYS: design, development and up-scaling of a sustainable production system for HEMP tex-tiles - an integrated quality SYStem approach. J. Ind. Hemp 8: 79-83
Amaducci S, Zatta A, Pelatti F, Venturi G (2008): Infl uence of agronomic factors on yield and quality of hemp fi bre and implication for innovative production system. Field Crop Res. 107: 161-169
Bócsa I, Karus M (1998): The hemp cultivation. Botany, varie-ties, cultivation and harvesting. Hemptech, Sebastopol, CA
Bredeman G (1942): Die bestimmung des fasergehaltes bei massenuntersuchungen von hanf, fl achs, fasernesseln und anderen bastfaserpfl anzen. Faserforschung 16: 14-39
Cromack H T H (1998): The effect of cultivation and seed den-sity on the production and fi bre content of Cannabis sativa in southern England. Ind. Crops Prod. 7: 205-210
Karus M, Vogt D (2004): European hemp industry: cultivation, processing and product lines. Euphytica 140: 7-12
Keller A, Leupin M, Mediavilla V, Wintermantel E (2001): In-fl uence of the growth stage of industrial hemp on chemical and physical properties of the fi bres. Ind. Crop Prod. 13: 35-48
Mediavilla V, Leupin M, Keller A (2001): Infl uence of the growth stage of industrial hemp on the yield formation in relation to certain fi ber quality traits. Ind. Crops Prod. 13: 49-56
Montford S, Small E (1999): Measuring harm and benefi t: The biodiversity friendliness of Cannabis sativa. Global Biodiver-sity 8: 2-13
Müssig J, Martens R (2003): Quality aspects in hemp fi bre pro-duction – infl uence of cultivation, harvesting and retting. J. Ind. Hemp 8: 11-32
Ranalli P (2002): Hemp in Italy: a new research project. J. Ind. Hemp 7: 139-141
Sankari H (2000): Comparison of bast fi bre yield and mechani-cal fi bre properties of hemp cultivars. Ind. Crops Prod. 11: 73-84
Starčević Lj (1996): Tehnologija gajenja konoplje za vlakno. Zbornik radova Instituta za ratarstvo i povrtarstvo 26: 39-53
Struik PC, Amaducci S, Bullard MJ, Stutterheim NC, Venturi G, Cromack HTH (2000): Agronomy of fi bre hem in Europe. Ind. Crops Prod. 11: 107-118
Van der Werf H M G, Wijlhuizen M, de Schutter J A A (1995): Plant density and self-thinning yield and quality of fi ber hemp. Field Crop Res. 40: 153-164

Sikora V i sar.112
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 107-112
Variability and Interrelation of Yield Components in Fiber Hemp
Vladimir Sikora1 · Janoš Berenji1 · Dragana Latković2
1Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia2Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
Summary: Variability and interrelation of yield components in fi ber hemp were analysed in fi eld experiments. This research included 20 commercial varieties currently being cultivated in Europe. Signifi cant variability was determined for plant height, stem yield, fi ber content and fi ber yield, so these materials can be useful as a good base for future fi ber hemp breeding and production improvement. Stem diameter was predominantly determined by ecological factors, and genetic background of examined varieties had small infl uence on expression of this trait. Regarding expression of other yield components, infl uence of genetic factors was more important than en-vironmental conditions. Correlation between analyzed traits shows that high stem and fi ber yield are achieved in higher populations of fi ber hemp with thicker stem. Fiber content increased with stem thickness increase.Key words: fi ber hemp, interrelation, yield components

Veza marker-svojstvo kod pšenice 113
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120originalni naučni rad / original research article
Analiza veze marker – svojstvo za vremeklasanja i cvetanja
pšenice korišćenjem pojedinačne marker regresije
Dragana Trkulja · Ankica Kondić-Špika · Ljiljana Brbaklić · Borislav Kobiljski
primljeno / received: 28.10.2010. prihvaćeno / accepted: 25.11.2010.© 2011 IFVC
Izvod: Dihaploidne mapirajuće populacije predstavljaju pogodan materijal za genetičku anal-izu lokusa koji utiču na ekspresiju kvantitativnih svojstava (QTL - Quantitative Trait Loci). U cilju detekcije lokusa koji su u vezi sa vremenom klasanja i cvetanja u našem agro-klimatskom regionu, u radu je analizirana alelna varijabilnost pet mikrosatelitskih lokusa (Xgwm18, Xg-wm194, Xgwm261, Xpsp3071 i Xpsp3200) kod 177 linija dihaploidne populacije Savana/Rene-sansa. Pored toga, na oglednim poljima Instituta za ratarstvo i povrtarstvo u Novom Sadu u periodu 2003-2008. rađena je fenotipska evaluacija ove mapirajuće populacije za navedene os-obine. Metodom pojedinačne marker regresije detektovan je QTL u blizini lokusa Xgwm261, pomoću kog se moglo objasniti od 20,2% do 30,7% fenotipske varijabilnosti za vreme klas-anja i 13,6% do 28,8% za vreme cvetanja, u svim analiziranim godinama.Ključne reči: cvetanje, dihaploidi, klasanje, mikrosateliti, pšenica, QTL
D. Trkulja · A. Kondić-Špika · Lj. Brbaklić · B. KobiljskiInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Kod klasičnih metoda oplemenjivanja bilja-ka, efi kasnost selekcije u velikoj meri zavisi od mogućnosti da se predvidi genotipska vrednost individua, na osnovu fenotipske analize i opaža-nja svojstava od agronomskog značaja. Imajući u vidu da se većina svojstava od značaja za opleme-njivače nalazi pod kontrolom velikog broja gena (tzv. kvantitativna svojstva), identifi kacija QTL-ova, bazirana isključivo na fenotipskoj selekciji, smatra se nemogućom (Eathington et al. 2007), naročito ako se uzme u obzir i uvek prisutna in-terakcija genotip/spoljna sredina, a koja je kod svojstava uslovljenih poligenima uvek karakteri-sana i različitim vrednostima heritabilnosti, tač-nije različitim udelom genotipske u ukupnoj fe-notipskoj varijansi. Sa pojavom tehnika moleku-larnih markera osamdesetih godina prošlog veka, stvorene su mogućnosti za preciznu genetičku analizu i detekciju lokusa koji kontrolišu osobi-ne, koje su deteminisane većim brojem gena. U odnosu na druge tehnike molekularnih markera,
mikrosateliti pokazuju znatno viši nivo polimor-fnosti i informativnosti pri proučavanju genoma heksaploidne pšenice (Ganal & Röder 2007).
Dihaploidne populacije imaju veliki potencijal za identifi kaciju QTL-ova jer je usled visokog procenta homozigotnosti i jednostavnog održa-vanja omogućena njihova evaluacija u različitim godinama i različitim uslovima spoljašnje sredine. Istraživanja bazirana na kompjuterskim simulaci-jama demonstrirala su da dihaploidne populacije povećavaju efi kasnost marker asistirane selekcije (MAS – Marker Assisted Selection), kroz strategiju kombinovanja velikog broja gena sa minimalnim brojem marker testova (Howes et al. 1998).
Vreme klasanja i cvetanja predstavlja glavnu odrednicu regionalne i sezonske adaptacije sor-ti pšenice. Prilagođenost se ogleda u izbegavanju niskih temperatura za vreme cvetanja (koje mogu da prouzrokuju mušku sterilnost), kao i visokih temperatura i suše u vreme nalivanja zrna. Stoga je razumevanje genetičke osnove vremena klasanja i cvetanja neophodan preduslov za stvaranje sorti
Molekularna genetika / Molecular Genetics w
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Ovo istraživanje je deo projekta broj TR-20138: Povećanje genetič-kih i proizvodnih potencijala strnih žita primenom klasične i moderne biotehnologije (2008-2010) Ministarstva za nauku i tehnološki razvoj Republike Srbije / This research results from the project TR-20138 funded by the Ministry of Science and Technological Development of the Republic of Serbia

Trkulja D i sar.114
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
koje odlikuje visok potencijal za prinos (Zhang et al. 2009).
Cilj rada je bio da se na osnovu analize rezultata molekularne i fenotipske evaluacije dihaploidne mapirajuće populacije pšenice utvrdi potencijalna veza između odabranih mikrosatelitskih markera i vremena klasanja i cvetanja u našem agroklimat-skom regionu.
Materijal i metod rada
Dihaploidna mapirajuća populacija Savana/Re-nesansa nastala je ukrštanjem dve sorte heksaplo-idne pšenice visokog potencijala za prinos, od kojih je jedna severnoevropska (Savannah, UK), a druga južnoevropska (Renesansa, SRB). Ona je stvorena 2000. u kompaniji Advanta Seeds (Velika Britanija) u saradnji sa John Innes Centrom (Norič, Velika Britanija) i Institutom za ratarstvo i povrtarstvo (Novi Sad, Srbija) u cilju determinacije lokusa za kvantitativna svojstva, koji utiču na ekspresiju pri-nosa u evropskom agroklimatskom regionu, čini je 177 linija, a dobijena je metodom eliminacije hro-mozoma polenskog roditelja u ukrštanju pšenice i kukuruza (Kondić-Špika & Kobiljski 2005).
Poljski ogled bio je postavljen po slučajnom blok sistemu u tri ponavljanja, na oglednim polji-ma Instituta za ratarstvo i povrtarstvo u Novom Sadu. Površina elementarne parcelice iznosila je 1,2 m2. Svaka je imala po 6 redova, a sejano je 60 zrna po redu. Fenotipska evaluacija se odvijala u periodu od šest godina (2003-2008) za vreme klasanja, odnosno pet godina za vreme cvetanja (gde je analiza izostala u 2006. godini). Navedena svojstva ocenjena su prema UPOV deskriptorima za pšenicu, na sledeći način:
• vreme klasanja – izraženo kao broj dana od 1. januara do datuma klasanja (50% klasalih biljaka/eksperimentalnoj jedinici)
• vreme cvetanja – izraženo kao broj dana od 1. januara do datuma cvetanja (50% biljaka u cveta-nju/eksperimentalnoj jedinici).
Za evaluaciju varijabilnosti mapirajuće popu-lacije Savana/Renesansa, odabran je set od pet mikrosatelitskih markera (GWM18, GWM 194, GWM 261, PSP3071 i PSP3200.), za koje je u ranijim istraživanjima utvrđena značajna pove-zanost sa agronomski važnim svojstvima pšenice (Quarrie et al. 2003, Kobiljski et al. 2007, Kumar et al. 2007, Chu et al. 2008).
DNK je izolovana iz svežeg mladog biljnog tkiva prema CTAB metodu (Doyle & Doyle 1990). Lan-čana reakcija polimeraze (PCR – Polymerase Chain Reaction) odvijala se prema modifi kovanom proto-kolu Röder et al. (1998). Reakciona smeša, ukupne zapremine 20 μl, sadržala je 25 ng genomske DNK,
1x pufer za Taq polimerazu, 2 mM MgCl2, 0,2 mM svakog dinukleotida (dATP, dCTP, dGTP, dTTP), 10 pmol svakog od prajmera i 2 jedinice Taq poli-meraze. Nakon 5 minuta inicijalne denaturacije na 94oC, sledilo je 47 ciklusa, pri čemu se svaki sastojao od sledećih koraka:
denaturacija na 94- oC u trajanju od 30 sekundihibridizacija prajmer sekvenci za matričnu -
DNK u trajanju od 30 sekundi na 52oC (GWM18 i GWM194) ili 62oC (GWM261, PSP3071 i PSP3200)
ekstenzija 30 sekundi na 72- oC.Posle toga sledila je fi nalna ekstenzija 10 mi-
nuta na 72oC.Analiza PCR produkata, dobijenih umnožava-
njem ispitivanih mikrosatelitskih lokusa, odvijala se pomoću automatskog laserskog DNK sekven-cera, model ABI Genetic Analyzer 3130 (Appli-ed Biosystems). Po završetku analize, podaci su obrađivani primenom GeneMapper softvera, verzija 4.0 (Applied Biosystems).
Rezultati fenotipske evaluacije obrađeni su anali-zom varijanse, primenom programa Statistica 8.0.
Za utvrđivanje veze marker-svojstvo korišćen je metod pojedinačne marker regresije (SMR - Single Marker Regression), primenom softve-ra QGene 4.0. Analizirani su sledeći parametri: aditivni efekat (Add effect), F vrednost za aditivni efekat [F (add)], LOD (logarithm of odds) i regresija (R2). Parametar aditivni efekat služi za procenu efekta alela. U ovom radu, pozitivna vrednost aditivnog efekta ukazuje na veći doprinos alela poreklom od Renesanse i obrnuto. Veza marker-svojstvo utvrđena je na bazi LOD vrednosti, koja predstavlja odnos verovatnoće da su dva lokusa vezana sa određenom rekombinacionom vredno-šću i verovatnoće da dati lokusi nisu vezani (Rish 1992). Vrednost R2 je izražena u procentima, a ukazuje na to koji deo ukupne fenotipske varija-bilnosti određenog svojstva je objašnjen pomoću datog markera. Imajući u vidu broj ispitivanih godina i heritabilnost analiziranih svojstava, veza marker-svojstvo smatrana je postojanom ukoliko se javljala u svim ispitivanim godinama.
Rezultati i diskusija
Na osnovu rezultata molekularne evaluacije, u svakom od pet ispitivanih mikrosatelitskih lo-kusa (Xgwm18, Xgwm194, Xgwm261, Xpsp3071 i Xpsp3200), pored alela koji su detektovani kod sorti roditelja, Savane i Renesanse (Tab. 1.), u mapirajućoj populaciji je utvrđeno prisustvo i do-datnih alela, ali je njihova zastupljenost bila ispod 10% (Sl. 1). Ovakvi rezultati mogu biti posledica eksperimentalne greške ili stranooplodnje.

Veza marker-svojstvo kod pšenice 115
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Analizom varijanse, između 177 linija dihaplo-idne mapirajuće populacije Savana/Renesansa, ustanovljene su statistički veoma značajne razli-ke u pogledu vremena klasanja i cvetanja, pa se
može pretpostaviti da se radi o genetičkom ma-terijalu koji odlikuje visok procenat rekombinaci-ja, što ovu populaciju čini pogodnom za analizu veze marker-svojstvo (Tab. 2.).
0.0%
10.0%
20.0%
30.0%
40.0%
50.0%
60.0%
70.0%
80.0%
nulti
186
188
190
nulti
163
165
175
193
nulti
110
130
110/
130
nulti
148
153
161
163
167
nulti
168
171
174
168/
171
Xgwm18 Xgwm261 Xgwm194 Xpsp3071 Xpsp3200
Frek
venc
ija a
lela
Fr
eque
ncy
of a
llele
s
Tabela 1. Aleli detektovani u ispitivanim mikrosatelitskim lokusima kod sorti roditeljaTable 1. Alleles detected at analysed microsatellite loci in parent varieties
LokusLoci
HromozomChromosome
Alel (bp)Allele (bp)
Savana RenesansaXgwm18 1B 190 186Xgwm261 2D 175 193Xgwm194 4D 110 130Xpsp3071 6A 161 153Xpsp3200 6D 168 171
Slika 1. Frekvencija alela u ispitivanim mikrosatelitskim lokusima kod dihaploidne populacije Savana/RenesansaFig. 1. Frequency of alleles at analysed microsatellite loci in doubled haploid population Savana/Renesansa
Tabela 2. Analiza varijanse vremena klasanja i cvetanja kod ispitivanih dihaploidnih linijaTable 2. Analysis of variance of the heading and fl owering of the analysed doubled haploid lines
Izvori varijacijeSource of variation
Suma kvadrata SS
Stepeni slobode
df
Sredina kvadrata
MSF p
Klasanje Heading
GenotipGenotype 20468 176 116 3.5** 0.00
PogreškaError 29252 885 33
CvetanjeFlowering
GenotipGenotype 15631 176 89 2.1** 0.00
PogreškaError 29400 708 42

Trkulja D i sar.116
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Analizom veze marker-svojstvo, u svim ispi-tivanim godinama, detektovan je QTL u blizini markera GWM261, pomoću kog se moglo obja-sniti od 20,2% do 30,7% fenotipske varijabilno-sti za vreme klasanja i 13,6% do 28,8% za vre-me cvetanja. Za vreme klasanja, LOD vrednost za dati marker, kretala se od 8,672 do 14,102, a za vreme cvetanja od 5,610 do 13,029. Negativ-
ne vrednosti za aditivni efekat, pri F(add) većoj od 15,0, ukazuju da je alel od 193 bp, poreklom od Renesanse, imao redukujući efekat na vreme klasanja i vreme cvetanja, odnosno determinisao ranije sazrevanje. Na osnovu rezultata ovog rada, nisu ustanovljene statistički značajne veze između preostala četiri ispitivana mikrosatelitska lokusa i vremena klasanja i cvetanja (Tab. 3. i Tab. 4).
Tabela 3. Pojedinačna marker regresija za vreme klasanja za šest ispitivanih godina (LOD > 3,0; F(add) > 15,0)Table 3. Single Marker Regression for heading time for six evaluated years (LOD > 3,0; F(add) > 15,0)
2003Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,472 2,111 0,461 1,22D Xgwm261 -1,963 44,292 8,672 20,24D Xgwm194 -0,447 1,549 0,339 0,96A Xpsp3071 -0,019 0,003 0,001 0,06D Xpsp3200 0,777 6,104 1,318 3,4
2004Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,651 2,058 0,449 1,22D Xgwm261 -3,212 67,152 12,483 27,74D Xgwm194 -0,558 1,237 0,271 0,76A Xpsp3071 0,173 0,135 0,030 0,16D Xpsp3200 1,212 7,683 1,651 4,2
2005Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,208 0,484 0,106 0,32D Xgwm261 -1,971 56,201 10,704 24,34D Xgwm194 -0,196 0,354 0,078 0,26A Xpsp3071 0,282 0,841 0,184 0,56D Xpsp3200 0,692 5,772 1,247 3,2
2006Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,234 0,658 0,144 0,42D Xgwm261 -2,007 64,797 12,107 27,04D Xgwm194 -0,054 0,029 0,006 0,06A Xpsp3071 0,436 2,169 0,474 1,26D Xpsp3200 0,486 3,000 0,653 1,7
2007Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,420 0,886 0,194 0,52D Xgwm261 -3,314 77,568 14,102 30,74D Xgwm194 -0,961 3,868 0,840 2,26A Xpsp3071 0,532 1,341 0,293 0,86D Xpsp3200 1,101 6,555 1,413 3,6
2008Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,336 0,881 0,193 0,52D Xgwm261 -2,606 73,418 13,465 29,64D Xgwm194 -0,311 0,621 0,136 0,46A Xpsp3071 0,346 0,881 0,193 0,56D Xpsp3200 0,820 5,624 1,216 3,1

Veza marker-svojstvo kod pšenice 117
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Marker GWM261 je jedan od najviše prouča-vanih mikrosatelitskih markera, jer se najčešće nasleđuje zajedno sa jednim od gena reduktora visine stabljike, Rht8 genom (Rht – Reducing hei-ght). Nalazi se 0,6 cM distalno od Rht8 gena na kratkom kraku 2D hromozoma (Korzun et al. 1998). Analizom preko 800 sorti pšenice pore-klom iz 20 različitih zemalja, utvrđeno je da su u lokusu Xgwm261, alelne varijante od 165, 174 i 192 bp, bile zastupljene sa preko 90% (Wor-land et al. 2001). Kobiljski et al. (2006) su eva-luacijom varijabilnosti mikrosatelitskog lokusa Xgwm261 kod 269 sorti i linija stvorenih u ople-
menjivačkim centrima u Srbiji, utvrdili da 73,6% genotipova poseduje alel od 192 bp, 14,9% alel od 174 bp, 7% alel od 165 bp i 4,56% novi fra-gment veličine oko 200 bp. Kod sorte Renesan-sa detektovan je alel od 192 bp. U ovom radu kod sorte Savana u lokusu Xgwm261 utvrđeno je prisustvo alela od 175 bp, a kod Renesanse od 193 bp (Tab. 1). Ovi aleli bili bi ekvivalentni fragmentima od 174 bp i 192 bp prethodno na-vedenih autora (Worland et al. 2001, Kobiljski et al. 2006), a razlike od 1 bp posledica su primene različitih sistema pri analizi fragmenata i kasnijoj obradi dobijenih rezultata.
Tabela 4. Pojedinačna marker regresija za vreme cvetanja za pet ispitivanih godina (LOD > 3,0; F(add) > 15,0)Table 4. Single Marker Regression for fl owering time for fi ve evaluated years (LOD > 3,0; F(add) > 15,0)
2003Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,601 3,437 0,748 1,92D Xgwm261 -1,613 27,501 5,610 13,64D Xgwm194 -0,577 2,585 0,564 1,56A Xpsp3071 -0,167 0,244 0,054 0,16D Xpsp3200 0,845 7,229 1,556 4,0
2004Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,637 2,310 0,504 1,32D Xgwm261 -3,014 69,947 12,924 28,64D Xgwm194 -0,494 1,133 0,248 0,66A Xpsp3071 0,167 0,147 0,032 0,16D Xpsp3200 1,101 7,403 1,592 4,1
2005Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,279 0,891 0,195 0,52D Xgwm261 -1,977 57,895 10,985 24,94D Xgwm194 -0,251 0,591 0,129 0,36A Xpsp3071 0,301 0,976 0,214 0,66D Xpsp3200 0,690 5,828 1,259 3,2
2007Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,586 2,312 0,504 1,32D Xgwm261 -2,774 70,175 12,960 28,64D Xgwm194 -0,964 5,215 1,129 2,96A Xpsp3071 0,442 1,232 0,270 0,76D Xpsp3200 1,022 7,546 1,623 4,1
2008Hromozom
ChromosomeLokusLoci Add effect F(add) LOD R^2 (%)
1B Xgwm18 -0,494 2,003 0,437 1,12D Xgwm261 -2,517 70,617 13,029 28,84D Xgwm194 -0,279 0,520 0,114 0,36A Xpsp3071 0,359 0,988 0,216 0,66D Xpsp3200 0,739 4,744 1,028 2,6

Trkulja D i sar.118
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Statistički značajna asocijacija između mar-kera GWM261 i vremena klasanja i cvetanja, ustanovljena u ovom radu, može biti rezultat veze ovog markera sa Ppd-D1 genom, koji pre-ma Scarth and Law (1983) ima značajnu ulogu u determinisanju vremena klasanja kao odgovor na dužinu trajanja dana. Primenom RFLP mar-kera, Ppd-D1 gen je lociran na kratkom kraku hromozoma 2D, 20,9 cM proksimalno od Rht8 gena (Worland et al. 1998). Xu et al. (2005) su, takođe na 2DS hromozomu, identifi kovali jedan QTL za vreme klasanja, 12,1 cM proksimalno od mikrosatelitskog lokusa Xgwm261, koji je objašnjavao oko 40% fenotipske varijabilnosti u tri ispitivane godine. Narasimhamoorthy et al. (2006) su pomoću metoda Single Marker Analysis ustanovili statistički značajnu asocijaciju izme-đu markera GWM261 i vremena klasanja, a na osnovu ovog mikrosatelita moglo se objasniti 17% fenotipske varijabilnosti. Primenom inter-valnog mapiranja veza je takođe bila statistički signifi kantna, ali se njime moglo objasniti 6,6% fenotipske varijabilnosti.
Ispitujući mogućnosti primene MAS pri oplemenjivanju pšenice na uslove suše, Quarrie et al. (2003) su utvrdili značajnu vezu između alela mikrosatelitskog lokusa Xpsp3200 na 6DS hromozomu i vremena cvetanja. U ovom radu, gde je dihaploidna populacija Savana/Rene-sansa gajena uz primenu uobičajene agroteh-
nike, nije ustanovljena statistički značajna veza između markera PSP3200 i vremena cvetanja (Tab. 4).
Imajući u vidu da su rezultati ovog rada dobi-jeni analizom jedne biparentalne populacije, de-tektovani QTL-ovi su validni samo u odnosu na dve alelne forme koje nose roditelji, te se lokus Xgwm261 može posmatrati samo kao kandidat marker za MAS. Sledeća faza istraživanja po-drazumevala bi validaciju dobijenih rezultata na materijalu koji je relevantan za oplemenjivanje u našem agroklimatskom regionu.
Zaključak
Utvrđeno je da se linije dihaploidne mapirajuće populacije Savana/Renesansa statistički veoma značajno razlikuju u pogledu vremena klasanja i cvetanja. Može se pretpostaviti da se radi o gene-tičkom materijalu koji odlikuje visok procenat re-kombinacija, što ovu populaciju čini pogodnom za analizu veze marker-svojstvo. Primenom me-toda pojedinačne marker regresije, ustanovljene su statistički značajne veze između mikrosatelit-skog markera GWM261 na kratkom kraku hro-mozoma 2D i oba posmatrana svojstva. Ova veza bila je postojana u svim analiziranim godinama. Preostala četiri mikrosatelitska lokusa nisu bila u statistički značajnoj asocijaciji sa vremenom kla-sanja i cvetanja.

Veza marker-svojstvo kod pšenice 119
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Literatura
Chu C G, Xu S S, Friesen T L, Faris J D (2008): Whole genome mapping in a wheat doubled haploid po-pulation using SSRs and TRAPs and the identifi -cation of QTL for agronomic traits. Mol. Breed. 22: 251-266
Doyle J J, Doyle J L (1990): Isolation of plant DNA from fresh tissue. Focus 12: 13-15
Eathington S R, Crosbie T M, Edvards M D, Reiter R S, Bull J K (2007): Molecular markers in a commerci-al breeding program. Crop Sci. 47(S3): S154-S163
Ganal M W and Röder M S (2007): Microsatellite and SNP markers in wheat breeding. In: Varshney R K and Tuberosa R (eds.), Genomics assisted crop improvement: genomics applications in crops, Springer, The Netherlands, 2: 1-24
Howes N K, Woods S M, Townley-Smith T F (1998): Simulations and practical problems of applying multiple marker assisted selection and doubled haploids to wheat breeding programs. Euphytica 100: 225–230
Kobiljski B, Denčić S, Hristov N, Mladenov N, Quarrie S, Stephenson P, Kirby J (2007): Potential uses of microsatellites in marker-assisted selection for improved grain yield in wheat. In: Buck H T et al. (eds), Wheat Production in Stressed Envi-ronments, Springer, The Netherlands, 729-736
Kobiljski B, Denčić S, Pilipović J (2006): Molecular screening of domestic germplasm for allelic va-riants at the dwarfi ng gene Rht8 locus in wheat. Genetika 38: 67-74
Kondić-Špika, A i Kobiljski, B (2005): Uticaj roditeljskih genotipova na formiranje haploidnih embriona u ukrštanjima pšenice i kukuruza. Sel. Semen. 11: 7-12
Korzun V, Röder M S, Ganal M W, Worland A J, Law C N (1998): Genetic analysis of the dwarfi ng gene (Rht8) in wheat. Part I. Molecular mapping of
Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.) Theor. Appl. Genet. 96: 1104-1109
Kumar N, Kulwal P L, Balyan H S, Gupta P K (2007): QTL mapping for yield and yield contributing tra-its in two mapping populations of bread wheat. Mol. Breed. 19: 163-177
Narasimhamoorthy B, Gill B S, Fritz A K, Nelson J C, Brown-Guedira G L (2006): Advanced backcross QTL analysis of a hard winter wheat x synthetic wheat population. Theor. Appl. Genet. 112: 787-796
Quarrie S A, Dodig D, Pekić S, Kirby J, Kobiljski B (2003): Prospects for marker-assisted selection of improved drought responses in wheat. Bulg. J. Plant Physiol. Special Issue. 83-95
Risch N (1992): Genetic linkage: Interpreting LOD sco-res. Science 255: 803-804
Röder M S, Korzun V, Wendehake K, Plaschke J, Tixier M-H, Leroy P, Ganal M W (1998): A Microsatellite Map of Wheat. Genetics 149: 2007-2023
Scarth R, Law C N (1983): The control of day-length response in wheat by the group 2 chromosome 2B of wheat. Heredity 51: 607-619
Worland A J, Börner A, Korzun V, Li W M, Petrović S, Sayers S J (1998): The infl uence of photoperi-od genes on the adaptability of European winter wheat. Euphytica 100: 385-394
Worland A J, Sayers E J, Korzun V (2001): Allelic vari-ation at the dwarfi ng gene Rht8 locus and its si-gnifi cance in international breeding programmes. Euphytica 119: 155-159
Xu X, Bai G, Carver B F, Shaner G E (2005): A QTL for early heading in wheat cultivar Suwon 92. Euphyti-ca 146: 233-237
Zhang K, Tian J, Zhao L, Liu B, Chen G (2009): De-tection of quantitative trait loci for heading date based on the doubled haploid progeny of two elite Chinese wheat cultivars. Genetica 135: 257-265

Trkulja D i sar.120
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 113-120
Marker-Trait Association Analysis for Heading and Flowering Time in Wheat by Single Marker Regression
Dragana Trkulja · Ankica Kondić-Špika · Ljiljana Brbaklić · Borislav Kobiljski
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Doubled haploid (DH) mapping populations are suitable material for genetic analysis of quantitative trait loci (QTL). In order to detect loci associated with heading and fl owering time in our agro-climate region, 177 lines of DH population Savana/Renesansa were analysed with fi ve microsa-tellite markers (GWM18, GWM194, GWM261, PSP3071 and PSP3200). These lines were also evaluated for heading and fl owering at the experimental fi eld of the Institute of Field and Vegetable Crops in Novi Sad. The experiment was conducted from 2003 to 2008. According to Single Marker Regression analysis, the marker GWM261 was associated with QTL which explained from 20.2% to 30.7% of the total variance for heading and from 13.6% to 28.8% of the total variance for fl owering time, in all analysed years.Key words: doubled haploids, fl owering, heading, microsatellites, QTL, wheat

Analiza mejoze triploida šećerne repe 121
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 121-124
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 121-124originalni naučni rad / original research article
Analiza mejoze triploida šećerne repe
Snežana Mezei · Lazar Kovačev · Jovanka Atlagić · Nevena Nagl
primljeno / received: 08.07.2010. prerađeno / revised: 22.11.2010. prihvaćeno / accepted: 30.11.2010.© 2011 IFVC
Izvod: U semenskoj proizvodnji triploida šećerne repe u toku žetve dolazi do osipanja semena. Zbog toga u narednih nekoliko godina niču triploidne i tetraploidne biljke. Cilj rada je bio da se uradi cito-genetska analiza nepravilnosti u mejozi kod triploidne hibridne sorte šećerne repe. Triploidi šećerne repe u Metafazi I imaju 9 univalenata i 9 bivalenata i mogu da budu izvor aneuploidnih gameta. Pojava aneuploida kao i njihova posledica, delimična ili potpuna sterilnost, jesu problemi koji prate semensku proizvodnju šećerne repe. Acetokarmin squash metodom pregledano je ukupno 2124 mejocita, kod kojih su u 14,64% ustanovljene različite nepravilnosti. Konstatovane su nepravilnosti tipa dislociranih hromo-zoma, ubrzanih (izbeglih) i usporenih (izostalih), kao i hromozomski mostovi.Ključne reči: citogenetska analiza, mejoza, šećerna repa, triploidi
S. Mezei · L. Kovačev · J. Atlagić · N. NaglInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Triploidne hibridne sorte šećerne repe (2n = 27) imaju intermedijarne morfološke karakteristi-ke u odnosu na roditelje, poseduju bolja kvanti-tativna svojstva i višu otpornost prema bolesti-ma, kada je otpornost uslovljena većim brojem minor gena (Nakamura et al. 1992) kao što je to kod otpornosti na Rhizoctonia (Hecker & Ruppel 1976) i otpornosti na Cercosporu (Skaracis & Smi-th 1987). Pojedina istraživanja imala su za cilj da uporede F1 hibride iste genetičke konstitucije na diploidnom, triploidnom i tetraploidnom nivou, da bi se ustanovio optimalni nivo ploidnosti u F1 generaciji. Unošenjem 9 ili 18 hromozoma pro-cesom ukrštanja u F1 hibrid nije bilo ni korisnih, ali ni štetnih efekata za prinos korena i sadržaj šećera (Hecker et al. 1969). Većina triploida koji su bili bolji od roditelja, dobijeni su sa tetraploid-ne majčinske komponente. Triploidi dobijeni sa tetraploida u proseku su za 17 % bolji od reci-pročnih hibrida (Smith et al. 1979).
U komercijalnoj proizvodnji triploidnih hibrida sa tetraploidne majčinske komponente ograniča-vajući faktor je prinos semena, koji iznosi svega 50% u poređenju sa triploidnim hibridima dobi-jenim sa diploidne majčinske komponente. Ra-zlog za ovo povećanje produktivnosti triploida se
ne zna sa sigurnošću. Jedna od pretpostavki je 4:3 odnos perisperma i embrija triploidnog semena sa tetraploida, u poređenju sa 2:3 odnosom kod recipročnog ukrštanja. Ovim se objašnjava brže rano razviće triploida sa tetraploidne majčinske komponente, mada se smatra da je ovo možda najmanji razlog za njihov bolji prinos korena (Bo-semark 1977). Prema rezultatima McFarlane et al. (1972) prinos korena triploidnih hibrida je samo 10% bolji od genetički identičnih diploidnih hi-brida. Stvaranje visoko produktivnih triploidnih hibrida zahteva širok program ispitivanja kombi-nacionih sposobnosti muško sterilnih (MS) linija i oprašivača. Triploidne hibridne sorte šećerne repe komercijalno su interesantne jer su potpuno ili parcijalno sterilne obzirom na dodati „n“ broj hromozoma, te je iz triploida teško koristiti odre-đena pozitivna svojstva za stvaranje nove genetič-ke varijabilnosti (Kovačev i sar. 2005, Kovačev et al. 2008). Pored ovih saznanja u procesu parenja hromozoma u dijakinezi triploida prosek trivale-nata je 7,8 % a frekvencija pojavljivanja je 3 do 9 po mejociti. Gamete triploida mogu da sadrže broj hromozoma koji varira od 9 do 18. Više od
Citogenetika / Cytogeneticsw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Ovo istraživanje je deo projekta broj TR20020: „Poboljšanje germ-plazme šećerne repe u cilju povećanja prinosa i smanjenja gubitaka nastalih uticajem biotskih i abiotskih faktora“ (01.04.2008-31.12.2010) Ministarstva za nauku i tehnološki razvoj Republike Srbije / This re-search results from project TR20020: „Improvement of sugar beet germplasm with aim to increase yield and reduce losses caused by biotic and abiotic factors„ (01.04.2008-31.12.2010) fi nanced by the Ministry of Science and Technological Development of the Republic of Serbia.

Mezei S i sar.122
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 121-124
70% potomstva iz ukrštanja triploida sa triploi-dima ili diploidima sadrži 18 ili 19 hromozoma, a većina klijalog semena je dobijena sa biljaka sa 18, 19, 20, 26 i 27 hromozoma (Skaracis 2005). Srbija je vrlo pogodno agroekološko područje za semensku proizvodnju šećerne repe. Pojava osi-panja semena u proizvodnji triploidnih hibrida ili sa biljke koja je triploidna proraslica, dovela bi do kontaminacije zemljišta i ono u narednih desetak godina ne bi bilo pogodno za proizvodnju se-menskog useva. Seme triploida zadržava klijavost i sukcesivno niče tokom dužeg vremenskog peri-oda, te se ova činjenica ne sme zanemariti. Ako bi se na ovim površinama posejala semenska šećer-na repa došlo bi do nekontrolisane oplodnje i na-rušavanja genetičke konstitucije hibridne sorte.
Obzirom da su abnormalnosti u mejotskom ciklusu uzrok parcijalne sterilnosti, cilj rada je bio da se utvrde nepravilnosti u mejozi triploida. Takođe, bolje poznavanje karakteristika mejoze triploida imaće značaj u daljem radu na opleme-njivanju šećerne repe.
Materijal i metod
Seme triploida šećerne repe (sorte Delta) pose-jano je u kadice u staklaru. Biljke su gajene u sta-kleniku u uslovima dugog dana, do faze desetak razvijenih listova. Jarovizacija sadnog materijala je obavljena u hladnoj komori na +4°C. Fototer-malna indukcija je trajala četiri meseca, a zatim
su biljke u rano proleće rasađene u polje. Sađeno je po pet biljaka u 10 redova na jednu parcelu, u izolaciju postignutu usevom konoplje.
Za citogenetske analize su uzete antere iz mla-dih cvetnih pupoljaka (vrhovi cvasti u dužini od oko 5 mm). Sa svake biljke skinuto je po 10 uzo-raka i fi ksirano 24 sata u smeši Carnoy I (3:1). Ukupno je analizirano pet stotina uzoraka. Do početka citogenetskih analiza uzorci su čuvani u 70% alkoholu, a predtretman je urađen sa 4% gvožđe amonijum sulfatom. Pravljeni su privre-meni preparati, a bojenje je vršeno acetokarmi-nom po metodi Snow (1963). Analizirano je 2124 mejocite na više stotina preparata. Analize su ura-đene na Laboval 4, Carl Zeiss Jena mikroskopu, uz korišćenje imerzionog objektiva HI 100/1,25.
Dobijeni rezultati su predstavljeni tabelarno kao procenat pojave nepravilnosti za analizirane faze mejoze, a najčešće zabeležene nepravilnosti u me-jozi su prikazane mikrosnimcima koji su napravljeni na mikroskopu Amplival 30-G048c Carl Zeiss Jena, uz korisćenje video kamere Sony DXC151AP.
Rezultati i diskusija
Triploidne hibridne sorte šećerne repe po svo-jim kvantitativnim svojstvima prevazilaze rodi-teljske komponente od kojih su nastale. Razlog ovog povećanja se ne zna, iako je poznato da se kod triploida ispoljavaju dva efekta, poliploidija i heterozis.
FazaPhase
Karakteristike deobe / Characteristics of cell division
Pravilna / Regular
Nepravilna / Irregular
Ubrzani (izbegli) hromozomi
Fast chromosomes
Usporeni (izostali) hromozomi
Lagging chromosomes
Hromozomski mostChromosome bridge
Broj mejocita
No. meiocytes
% mejocita
%meiocytes
Broj mejocita
No. meiocytes
% mejocita
%meiocytes
Broj mejocita
No. meiocytes
% mejocita
%meiocytes
Broj mejocita
No. meiocytes
% mejocita
%meiocytes
Meta I 291 78,44 80 21,56Ana I 593 84,96 79 11,32 26 3,72Telo I 422 100Meta II 170 75,89 54 24,11Ana II 276 79,31 53 15,23 19 5,46Telo II 61 100
UkupnoTotal
1813 134 132 45311
2124
Tabela 1. Analiza mejoze triploidne hibridne sorte šećerne repe DeltaTable 1. Meiosis of triploid hybrid sugar beet variety Delta

Analiza mejoze triploida šećerne repe 123
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 121-124
Poznato je da je mejoza triploidnih hibrida ne-pravilna i da nastaju gamete sa različitim brojem hromozoma. Iz tih razloga kod triploidne hibrid-ne sorte Delta analizirana je mejoza po fazama. U rezultatima je naveden broj pregledanih mejocita i procenat pravilnosti ili odstupanja od normalne redukcione deobe. Citogenetskom analizom me-jotskog ciklusa analizirane su pojave dislociranih hromozoma i hromozomskih mostova u prvoj i drugoj fazi mejoze. Rane faze mejoze nisu anali-zirane (Sl. 1a). Pahiten i diploten u Profazi I nisu pogodni za analizu hromozomske konfi guracije kod šećerne repe, jer su hromozomi ispreplete-ni u vidu dugih tankih niti (Yu 1977, Tsuchiya & Nakamura 1979).
Od 2124 analizirane mejocite u 85,36% ćelija redukciona deoba je proticala normalno. Faze mejoze su bile nepravilne kod 14,64% analizi-ranih ćelija (Tab. 1). Najčešće su uočene ćelije u Metafazi I, Anafazi I, kao i ćelije u Metafazi II i Anafazi II. Takođe je detektovana asinhrona de-oba hromozoma u drugoj redukcionoj deobi, a što se javlja i kod analize tetraploidnih populacija šećerne repe (Mezei et al. 2005).
Od ukupno 291 analizirane ćelije u Metafazi I, 78,88% ćelija nije imalo nepravilnosti u deobi.
Kod 21,56% mejocita nađeni su ubrzani (izbegli) hromozomi. Najčešće su izbegla jedan ili dva hro-mozoma. U Anafazi I analizirano je 593 ćelije od kojih je 84,96 % imalo pravilnu deobu (Sl. 1b), dok je 11,32 % mejocita bilo sa usporenim (izostalim) hromozomima (Sl. 1c). Hromozomski mostovi su zapaženi kod 3,72% analiziranih ćelija (Sl. 1d). Pregledom mejocita u Telofazi I ustanovljeno je da je mejotička deoba kod 422 analizirane mejocite proticala bez nepravilnosti (Sl. 1e) (Tab. 1).
U Metafazi II od analiziranih 170 mejocita 75,89% je imalo pravilan mejotski ciklus, dok je 24,11% mejocita bilo sa ubrzanim (izbeglim) hromozomima (Sl. 1f). Analizom 276 mejocita u Anafazi II 79,31 % ćelija je prolazilo kroz pra-vilnu deobu, 15,23 % ćelija je bilo sa usporenim (izostalim) hromozomima (Sl. 1g), a 5,46 % sa hromozomskim mostom (Sl. 1h) (Tab. 1). U ovoj fazi se uočavaju formacije „v“ tipa rasporeda hromozoma, kao rezultat zadržavanja mostova iz Anafaze I. Pojava hromozomskih mostova uka-zuje na postojanje hromozomskih aberacija tipa paracentričnih inverzija, koje je takođe detekto-vao Yu (1978) u Profazi I mejoze. U Telofazi I kao i u Telofazi II (Sl. 1i) nisu ustanovljene ne-pravilnosti u deobi.
Slika 1. Faze mejoze triploidnog hibrida šećerne repea) Dijakineza, b) Anafaza I pravilna, c) Anafaza I sa usporenim (izostalim) hromozomom, d) Anafaza I sa hromozomskim mostom, e) Telofaza I pravilna, f) Metafaza II sa ubrzanim (izbeglim) hromozomom, g) Anafaza II sa usporenim (izostalim) hromozomima, h) Anafaza II sa hromozomskim mostom, i) Telofaza II pravilna. Fig. 1. Phases of meiosis of triploid hybrid sugar beeta) Diakinesis, b) Anaphase I normal, c) Anaphase I with lagging chromosome, d) Anaphase I with chro-mosome bridge, e) Telophase I normal, f) Metaphase II with fast chromosome, g) Anaphase II with lagging chromosomes, h) Anaphase II with chromosome bridge, i) Telophase II normal

Mezei S i sar.124
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 121-124
Zaključci
Kod triploidne hibridne sorte šećerne repe Delta analizirane su mejocite u različitim faza-ma redukcione deobe primenom acetokarmin metode. U 85,36% mejocita mejoza je proticala normalno.
Od nepravilnosti koje su se pojavljivale u 14,64% analiziranih mejocita najveći procenat se odnosio na pojavu neuključenih hromozoma, izbeglih (ubrzanih) u Metafazi I i Metafazi II, za-tim izostalih (usporenih) u Anafazi I i Anafazi II, kao i pojava hromozomskih mostova u Anafazi I i Anafazi II.
Zbog pojave navedenih nepravilnosti u mejozi očekuje se veliki procenat gameta sa različitim bro-jem hromozoma, pojava aneuploida i delimične odnosno potpune sterilnosti. Ova saznanja su ve-oma važna u semenskoj proizvodnji šećerne repe jer mogu da budu izvor potencijalnih problema.
Literatura
Bosemark N O (1977): Use of tetraploid monogerm male steri-les in triploid hybrid seed production. Proceeding Inst. Sugar Beet Res. 271-287
Hecker R J, Ruppel E G (1976): Polyploid and maternal effects on Rhizoctonia root rot resistance in sugarbeet. Euphytica 25: 419-423
Hecker R J, Staford R E, Helmeriich R H, Maag G W (1969): Comparasion of the same sugar beets F1 hybrids as diploids, triploids and tetraploids. J. Amer. Soc. Sugar Beet Tech. 16: 106-116
Kovačev L, Čačić N, Mezei S, Nagl N (2005): Značaj genetike, oplemenjivanja i genetičkog inženjeringa u povećanju poten-cijala rodnosti kod šećerne repe i produktivnost NS hibrid-nih sorti. Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 41: 35-50
Kovačev L, Čačić N, Stančić I, Mezei S (2008): Sugar beet bree-ding in Serbia. International Conference «Conventional and Molecular Breeding of Field and Vegetable Crops», Novem-ber 24-27, Novi Sad, Serbia, 513-517
Mc Farlane J S, Skoyen I O, Lewellen R T (1972): Performance of sugar beet hybrids as diploids and triploids. Crop Sci. 12: 118-129
Mezei S, Atlagić J, Kovačev L (2005): Pollen viability and meio-sis in tetraploid populations of sugar beet (Beta vulgaris L.). J. Genet. Breed. 59: 157-164
Nakamura C, Skaracis N G, Romagosa I (1992): Cytogenetics and breeding in sugar beet. In: Chromosome engineering in plants: Genetics, Breeding, Evolution. Elsevier, Amsterdam, Netherlands, 295-314
Skaracis N G, Smith G A (1987): Ploidy level effect on resi-stance to cercospora leaf spot in sugar beet. Proc. 24th Meet. ASSBT, 24-26
Skaracis N G (2005): Cytology and cytogenetics 19-52. In: Bi-ancardi E, Campbell L G, Skaracis N G, De Biaggi M (eds.), Genetics and breeding of sugar beet, Science Publishers Inc., Enfi elds, NH, USA, 367
Smith G A, Hecker R J, Martin S S (1979): Effect of ploidy level on the components of sucrose yield and quality in sugarbeet. Crop Sci. 19: 319-323
Snow R (1963): Alcoholic hydrochloric acid-carmine as stain for chromosomes in squash preparations. Stain Technol. 38: 9-13
Tsuchiya T, Nakamura C (1979): Acetocarmine squash method for observing sugar beet chromosomes. Euphytica 28: 149-156
Yu M H (1977): Preliminary study of pachytene morphology in a homologous line of sugar beet. Crop Sci. 17: 833-836
Yu M H (1978): Meiotic behavior of a disomic nematode resi-stant sugarbeet. Crop Sci. 18: 615-618
Meiose Analyses of Sugar Beet Triploids
Snežana Mezei · Lazar Kovačev · Jovanka Atlagić · Nevena Nagl
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: During the seed crop growing there can be also problems with uncontrolled pollination if in the previo-us years there was seed shedding during the harvest, which can cause growth and fl owering of diploid, triploid and tetraploid plants and that can present serious problem for the next sugar beet seed crop. Meiosis of triploid hybrids is irregular and can result in development of gametes with different chromosome number. Sugar beet triploids in metaphase I have 9 univalents and 9 bivalents and therefore can be source of aneuploid gametes. After cytological analysis that was performed on 2124 meiocytes, in 14.64% different types of irregularities were detected, such as dislocated chromosomes and chromosome bridges.Key words: chromosome, meiosis, seed, sugar beet, triploid

Uticaj različitih doza NPK đubriva na kvalitet semena uljane repice 125
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130originalni naučni rad / original research article
Uticaj različitih doza NPK đubrivana kvalitet semena uljane repice
Dušica Jovičić · Ana Marjanović-Jeromela · Milka Vujaković ·Radovan Marinković · Zvonko Sakač · Zorica Nikolić · Branko Milošević
received / primljeno: 01.11.2010. accepted / prihvaćeno: 07.12.2010.© 2011 IFVC
Izvod: Uljana repica (Brassica napus L.) nalazi veliku primenu u proizvodnji ulja, ishrani domaćih životi-nja i proizvodnji biodizela. Kvalitet semena uljane repice zavisi od genotipa, meteoroloških i zemljišnih uslova, đubrenja i ostalih agrotehničkih mera. Veoma je važno da se zemljište obezbedi dovoljnim ko-ličinama hraniva pred setvu (osnovno đubrenje), kako bi mlade biljke mogle da formiraju lisnu rozetu i pripreme se za period niskih zimskih temperatura. Ispitivanja su izvedena sa četiri komercijalne sorte uljane repice (Banaćanka, Slavica, Express i Valeska) proizvedene u vegetacionoj sezoni 2008-2009. Ispi-tivani su sledeći parametri: klijavost semena, masa 1000 semena, sadržaj ulja i masnih kiselina. Klijavost semena, masa 1000 semena i sadržaj ulja zavisio je od sorte, ali ne i od primenjene doze NPK đubriva. Sadržaj masnih kiselina zavisio je i od sorte i od količine primenjene doze đubriva.Ključne reči: klijavost, masa 1000 semena, sadržaj masnih kiselina, sadržaj ulja, uljana repica
D. Jovičić · A. Marjanović-Jeromela · M. Vujaković · R. Marinković · Z. Sakač · Z. Nikolić · B. MiloševićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Uljana repica (Brassica napus L.) je najvažnija uljana biljka umerenog klimata i po površina-ma na kojima se gaji zauzima treće mesto među uljaricama u svetu (FAO 2009). Gaji se pre sve-ga zbog semena koje sadrži veliku količinu ulja (40% do 48%) (Marinković i sar. 2009). Nakon ekstrakcije ulja ostaje nusproizvod sačma koja sa-drži 25% do 40% proteina (Enami 2011) i 8% ulja, te predstavlja kvalitetnu komponentu stočne hrane. Ova uljarica postaje posebno aktuelna sa aspekta proizvodnje biodizela, koji je važan izvor bioobnovljive energije.
Ulje dobijeno iz semena uljane repice sadrži više masnih kiselina. Sadržaj nezasićenih masnih kiselina (oleinska i linolna) daje kvalitet ulju, dok su neke manje poželjne (linoelinska) ili čak nepo-željne (eruka). Sorte uljane repice sa sadržajem eruka kiseline u ulju ispod 2% pripadaju tzv. “0“ tipu. Daljim oplemenjivanjem ove kulture dobi-jene su tzv. “00“ sorte koje karakteriše smanjen sadržaj glukozionolata u semenu, koji nakon
ceđenja ostaju u sačmi i toksični su za domaće životinje (Marjanović-Jeromela i sar. 2008a). Po-slednjih godina u Evropi je otpočelo gajenje tzv. “000“ sorte sa još povoljnijim karakteristikama ulja. Upotreba ulja uljane repice zavisi od odnosa pojedinih masnih kiselina. Visok sadržaj oleinske kiseline i nizak sadržaj zasićenih masnih kiselina veoma je važna karakteristika ulja uljane repice, koja zajedno sa prisustvom linolne i linolenske ki-seline daje ovom ulju jedinstvenu nutritivnu vred-nost za prevenciju kardiovaskularnih oboljenja (Massaro et al. 2006). Visokokvalitetno prirodno rafi nisano ulje bogato je nezasićenim masnim kiselinama koje mu daju izuzetnu biološku vred-nost i posebno mesto u ljudskoj ishrani. Zato je neophodno utvrditi koji su parametri značajni za variranje sadržaja i sastava ulja u semenu, odno-sno utvrditi faktore spoljašnje sredine koji utiču na ova svojstva.
Kvalitet semena uljane repice zavisi od genoti-pa, meteoroloških i zemljišnih uslova, đubrenja i ostalih agrotehničkih mera (Laaniste et al. 2004). Veoma je važno da se zemljište obezbedi dovolj-
Semenarstvo / Seed Sciencew
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Ovo istraživanje je rezultat projekta TR31025 „Razvoj novih sorti i poboljšanje tehnologija proizvodnje uljanih biljnih vrsta za različite namene“ Ministarstva za nauku i tehnološki razvoj Republike Srbije / This research results from project TR31025 „Development of new varieties and production technology improvement of oil crops for dif-ferent purposes“ funded by the Ministry of Science and Technological Development of the Republic of Serbia

Jovičić D i sar.126
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
nim količinama hraniva pred setvu (osnovno đu-brenje), kako bi mlade biljke mogle da formiraju lisnu rozetu sa 6 do 8 pari listova i koren dubine 15 cm do 20 cm, gde će se u korenovom vratu nakupiti dovoljno hranljivih materija i izvršti ne-ophodno kaljenje biljaka za period niskih zimskih temperatura. Azot je veoma važan elemenat ne-opohodan za normalan rast i razvoj biljaka, ula-zi u sastav proteina, hlorofi la, pospešuje proces fotosinteze i sintezu mnogih metabolita. Fosfor povoljno utiče na razvoj korenovog sistema i re-produktivnih organa, dok kalijum pospešuje na-kupljanje hranljivih materija u plodu i povećava otpornost biljaka na niske temperature i bolesti. Preporučene optimalne količine hraniva za posti-zanje visokih prinosa semena dobrog kvaliteta su sledeće: 210 kg N ha-1, 75 kg P2O5 ha-1 i 300 kg K2O ha-1 (Marinković i sar. 2006). Obezbeđenost useva uljane repice dovoljnom količinom azota je posebno važno pre svega zbog činjenice da je efi -kasnost korišćenja ovog elementa niska (Hocking et al. 1997). Autori Nemeth et al. (2007) smatraju da azot predstavlja odlučujući faktor za prinos se-mena uljane repice.
Cilj rada je bio da se utvrdi uticaj različitih doza NPK đubriva, primenjenih pri osnovnom đubre-nju, na kvalitet semena uljane repice: klijavost se-mena, masu 1000 semena, sadržaj ulja i masnih kiselina odabranih sorti uljane repice.
Materijal i metod
Ispitivanja su izvedena sa četiri komercijalne sorte uljane repice (Banaćanka, Slavica, Express i Valeska) gajene tokom vegetacione sezone 2008-2009. Ogled je postavljen na oglednom polju In-stituta ratarstvo i povrtarstvo u Novom Sadu na zemljištu tipa černozem. Pred setvu je izvršeno đubrenje NPK đubrivom (15:15:15) u različitim dozama: (i) 0 kg N ha-1, 0 kg P2O5 ha-1, 0 kg K2O ha-1 (kontrolna varijanta), (ii) 20 kg N ha-1, 20 kg
P2O5 ha-1, 20 kg K2O ha-1 i (iii) 40 kg N ha-1, 40 kg P2O5 ha-1, 40 kg K2O ha-1. Ispitivani su sledeći parametri: klijavost semena, masa 1000 semena, sadržaj ulja i masnih kiselina.
Klijavost semena i masa 1000 semena određe-ni su standardnom metodom prema Pravilniku o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87). Naklijavanje je izvršeno u četiri po-navljanja po sto semena u Petri posudama. Inku-bacioni period trajao je sedam dana na tempera-turi 20°C do 30°C.
Sadržaj ulja određen je NMR metodom u He-mijskoj laboratoriji Odeljenja za uljane kulture Instituta za ratarstvo i povrtarstvo u Novom Sadu, a sadržaj i sastav masnih kiselina GC hro-matografi jom sa FID detektorom.
Rezultati su obrađeni u statističkom programu Statistica 7, metodom analize varijanse i korelaci-onom analizom. Značajnost razlika između sredi-na utvrđena je Dankanovim testom (p<0,05).
Rezultati i diskusija
Klijavost semena uljane repice kretala se od 80% (Valeska, kontrolna varijanta) do 95% (Ba-naćanka, 40 kg ha-1) (Tab. 1). Kod svih ispitivanih genotipova (osim kod sorte Slavica) sa poveća-njem količine đubrenja povećavala se i klijavost semena, ali razlika nije bila statistički značajna. Sorta Valeska je pri svim primenjenim količinama đubriva imala statistički značajno nižu klijavost u odnosu na sve ostale ispitivane sorte. Zavisnost klijavosti semena od sorti utvrdili su takođe Vuja-ković i sar. (2008).
Masa 1000 semena nije zavisila od primenjene količine đubriva (Tab. 2). Sorta Valeska imala je najveću masu 1000 semena pri svim primenjenim količinama đubriva. Autori Danesh-Shahraki et al. (2008) navode zavisnost mase 1000 semena od količine primenjenog azota, što se može obja-sniti činjenicom njegovog povoljnog dejstva na
Tabela 1. Uticaj različitih doza N15P15K15 đubriva na klijavost semenaTable 1. The effect of different doses of N15P15K15 fertilizers on seed germination
Doze đubrivaDoses of fertilizers Banaćanka Slavica Express Valeska
Prosek (doze)Mean (dose)
0 kg ha-1 91a 93a 91a 80b 88,75a
20 kg ha-1 94a 91a 92a 82b 89,75a
40 kg ha-1 95a 91a 92a 84b 90,5a
Prosek (sorte)Mean (genotype) 93,33a 91,67a 91,67a 82,00b

Uticaj različitih doza NPK đubriva na kvalitet semena uljane repice 127
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
intenzitet fotosinteze, pa samim tim i na akumu-laciju hranljivih materija u zrnu. Međutim, ovo nije pokazano ovim istraživanjima, iako je kod najveće doze uneta veća količina azota (267 kg ha-1) od količine koju preopručuje Marinković i sar. (2006) (210 kg ha-1). Sorta Valeska je imala statistički najveću srednju vrednost mase 1000 semena (4,07 g) dok je najmanju imala sorta Sla-vica (3,31 g). Zavisnost ovog parametra od sorte pokazali su autori Sana et al. (2003), El-Nakhlawy & Bakhashwain (2009) i Vujaković i sar. (2010).
Popravka prirodne plodnosti zemljišta NPK đubrivima pred osnovnu obradu nije imala zna-čajan uticaj na sadržaj ulja u ovom istraživanju (Tab. 3) što se poklapa sa rezultatima Marjano-vić-Jeromela i sar. (1999). Samo je sorta Express pokazivala smanjenje sadržaja ulja sa poveća-njem količine hraniva u zemljištu, ali razlika nije bila statistički značajna. Ova osobina u većoj meri zavisila je od same sorte, tako da je najma-
nji sadržaj zabeležen kod sorte Valeska (38,7% kontrolna varijanta), a najveći kod sorte Express (43,8% kontrolna varijanta). Autori Brennan i sar. (2000) su utvrdili da se sa povećanjem ko-ličine azota u zemljištu smanjuje sadržaj ulja. U proseku za ceo ogled sadržaj ulja je bio niži od prosečnih vrednosti za ovaj lokalitet, što se može objasniti nepovoljnim uslovima za fotosintezu i sintezu ulja u vreme nalivanja zrna (Marjano-vić-Jeromela i sar. 2008b) u vegetacionoj sezoni 2008-2009. Iako su temperature bile optimalne, nepravilan raspored padavina između perioda nalivanja zrna (oko 86 mm) i perioda sazrevanja zrna (oko 130 mm) se odrazio na smanjen sadr-žaj ulja u semenu. Autori Rehman et al. (1987) smatraju da pored suše i visoke temperature u periodu nakon cvetanja bitno utiču na veličinu semena, sadržaj ulja i prinos.
Sastav masnih kiselina koje dominiraju (pal-mitinska, stearinska, oleinska, linolna, linolenska
Tabela 2. Uticaj različitih doza N15P15K15 đubriva na masu 1000 semenaTable 2. The effect of different doses of N15P15K15 fertilizers on 1000-seeds weight
Doze đubrivaDoses of fertilizers Banaćanka Slavica Express Valeska
Prosek (doze)Mean (dose)
0 kg ha-1 3,75abcd 3,31e 3,45de 4,08a 3,65a
20 kg ha-1 3,69bcd 3,39de 3,51cde 4,09a 3,67a
40 kg ha-1 3,80abc 3,42de 3,41de 4,04ab 3,67a
Prosek (sorte)Mean (genotype) 3,75b 3,37c 3,46c 4,07a
Tabela 3. Uticaj različitih doza N15P15K15 đubriva na sadržaj ulja i masnih kiselina u semenu uljane repice Table 3. The effect of different doses of fertilizers N15P15K15 on oil content and fatty acids in rapeseed
Đubrenje / Fertilization
Sorta / Genotype
Sadržaj ulja / Oil content
(%)
Sadržaj masnih kiselina / Fatty acid content (%)Palmitinska
kis. Palmitic
acid 16:0
Stearinska kis. Stearic acid 18:0
Oleinska kis. Oleic
acid 18:1
Linolna kis.
Linoleic acid 18:2
Linolenska kis.
Linolenic acid 18:3
Eruka kis. Erucic acid
22:1
0 kg ha-1
Banaćanka 42.29ab 4.44cdef 2.02a 63.21bc 18.33ab 9.33a 0.19c
Slavica 42.10abcd 4.97a 2.06a 62.92bcd 18.42ab 9.40a 0.12c
Express 43.80a 4.13fg 2.38a 65.58a 16.06c 8.12bc 0.70bc
Valeska 38.70bcd 4.76abc 1.77a 61.72cd 19.46a 7.62c 1.29b
20 kg ha-1
Banaćanka 43.12a 4.36def 1.86a 63.59b 17.88b 9.15a 0.54c
Slavica 41.48abcd 4.92ab 2.26a 62.07bcd 18.39ab 9.78a 0.12c
Express 43.63a 4.18efg 2.28a 65.86a 16.03c 8.05bc 0.82bc
Valeska 39.68cd 4.92ab 2.41a 59.25e 18.39ab 8.13bc 2.47a
40 kg ha-1
Banaćanka 42.23abc 4.26defg 2.19a 63.60bc 18.53ab 8.89ab 0.22c
Slavica 42.16abcd 4.59bcd 2.33a 63.68b 17.68b 9.09a 0.17c
Express 43.53a 3.93g 2.21a 67.07a 15.67c 7.76c 0.66bc
Valeska 39.62d 4.52cde 1.96a 61.17d 18.66ab 7.63c 2.09a

Jovičić D i sar.128
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
i eruka) kod četiri sorte uključene u istraživanje odgovara rezultatima koje u svom radu navode Pospišil i sar. (2007) na osnovu višegodišnjeg ogleda u regionu jugoistočne Evrope. Autori Baux et al. (2008) navode da je sastav ulja uljane repice moguće indirektno povezati sa đubrenjem azotom preko dužine trajanja fenofaza i razvoja biljaka, kao i meteoroloških parametara tokom vegetacije, posebno u vreme sinteze masnih kise-lina. U našem istraživanju nije utvrđen značajan uticaj dodatih makroelemenata na sastav masnih kiselina, ali je evidentno da sa povećanjem doze đubriva postoji izvesno povećanje sadržaja ole-inske kod svih sorti osim kod sorte Valeska, a smanjenje sadržaja linolne kiseline kod sorti Sla-vica i Express (Tab. 3). Osim toga, konstatovan je negativan koefi cijent korelacije r=- 0,7592 iz-među sadržaja oleinske i linolne kiseline (Sl. 1). Dobijena negativna korelacija veoma je značajan pokazatelj oplemenjivačima, kao i proizvođačima semena uljane repice pri izboru metoda selekcije i agromera u proizvodnji semena uljane repice u zavisnost od njegove namene. Pozitivna korelaci-ja oleinske (r=0,4281) i negativne eruka kiseline (r=-0,5669) sa klijavošću semena uljane repice upućuje na složenost mehanizama koji regulišu semenske kvalitete uljane repice. Sadržaj stearin-ske kiseline nije zavisio ni od doze primenjenog đubriva, kao ni od sorte. Sadržaj linolenske kiseli-ne kod sorti Banaćanka i Express smanjivao se sa porastom količine NPK đubriva.
Zaključci
Na osnovu dobijenih rezultata mogu se doneti sledeći zaključci:
Sa povećanjem količine đubrenja došlo je do povećanja klijavosti semena, ali to povećanje nije bilo statistički značajno. Statistički značajne razli-ke u klijavosti semena su se javile između ispiti-vanih sorti i najniže vrednosti su dobijene kod sorte Valeska.
Masa 1000 semena je bila statistički značajno najviša kod sorte Valeska, dok ispitivani para-metar nije zavisio od primenjene doze đubri-va.
Sadržaj ulja u semenu u većoj meri je zavi-sio od same sorte. Najmanji sadržaj ulja imala je sorta Valeska (38,7% kontrolna varijanta), a najveći sorta Express (43,8% kontrolna vari-janta).
Kod svih sorti (osim sorte Valeska) sa pove-ćanjem doze đubriva zabeleženo je izvesno po-većanje sadržaja oleinske kiseline. Sorte Slavica i Express pokazale su smanjenje sadržaja linolne kiseline sa povećanjem doze đubriva, a koefi ci-jent korelacije između sadržaja oleinske i linolne kiseline bio je negativan.
Pozitivan korelacioni odnos između oleinske kiseline i negativan odnos između eruka kiseline sa klijavošću semena uljane repice dobijene anali-zom rezultata upućuje na složenost mehanizama koji regulišu semenske kvalitete uljane repice.
Slika 1. Korelacija između sadržaja oleinske i linolne kiselineFig. 1. The correlation between the content of oleic and linoleic acid

Uticaj različitih doza NPK đubriva na kvalitet semena uljane repice 129
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
Literatura
Brennan R F, Mason M G, Walton G H (2000): Effect of ni-trogen fertilizer on the concentrations of oil and protein in canola (Brassica napus) seed. J. Plant Nutr. 23: 339-348
Baux A, Hebeisen T, Pellet D (2008): Effects of minimal tem-peratures on low-linolenic rapeseed oil fatty-acid compositi-on. Europ. J. Agron. 29: 102-107
Danesh - Shahraki A, Nadian H, Bakhshandeh A, Fathi G, Ala-misaied K, Gharineh M (2008): Optimization of Irrigati-on and Nitrogen Regimes for Rapeseed Production Under Drought Stress. J. Argon. 7: 321-326
El-Nakhlawy F, Bakhashwain A (2009): Performance of Ca-nola (Brassica napas L.) Seed Yield, Yield Components and Seed Quality under the Effects of Four Genotypes and Nitrogen Fertilizer Rates. Met., Env. Arid Land Agric. Sci. 20: 33-37
Enami, H R (2011): A review of using canola/rapeseed meal in aquaculture feeding. J. Fish Aquat. Sci. 6: 22-36
FAOSTAT (2009): [Elektronski izvor] dostupno na adresi http://faostat.fao.org.
Hocking P J, Kirkegaard J A, Angus J F, Gibson A H, Koetz E A (1997): Comparison of canola, Indian mustard and Linola in two contrasting environments. I. Effects of nitrogen fer-tilizer on dry-matter production, seed yield and seed quality. Field Crops Res. 49: 107-125
Laaniste P, Joudu J, Eremeev V. (2004): Oil content of spring oilseed rape seeds according to fertilization. Agron. Res. 2: 83-8
Marinković R, Marjanović-Jeromela A, Sekulić R, Mitrović P (2006): Tehnologija proizvodnje ozime uljane repice. Nauč-ni institut za ratarstvo i povrtarstvo, Novi Sad
Marinković R, Marjanović-Jeromela A, Mitrović P (2009): Oso-benosti proizvodnje ozime uljane repice (Brassica napus L.) pregledni rad. Zbornik radova Instituta za ratarstvo i povr-tarstvo, Novi Sad, 46: 33-43
Marjanović-Jeromela A, Marinković R, Crnobarac J (1999): Uti-caj rokova setve i đubrenja na komponente prinosa uljane
repice (Brassica napus L.). Proizvodnja i prerada uljarica, 40. Savetovanje industrije ulja – zbornik radova: 243-254
Marjanović-Jeromela A, Marinković R, Atlagić A, Saftić-Pan-ković D, Miladinović D, Mitrović P, Miklič V (2008a): Do-stignuća u oplemenjivanju uljane repice (Brassica napus L.) u Institutu za ratarstvo i povrtarstvo. Zbornik radova Instituta za ratarsvo i povrtarstvo, Novi Sad 45: 131-144
Marjanović-Jeromela A, Marinković R, Mitrović P, Miklič V (2008b): Proizvodne vrednosti novih genotipova uljane re-pice (Brassica napus L.). Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 45: 103-110
Massaro M, Scoditti E, Annunziatta Carluccio M, De Caterina R (2006): Epidemiology of Olive Oil and Cardiovascular Disease. In: J. L. Quiles, M. C. Ramirez-Tortosa, P. Yaqoob (eds), Olive oil & health. CAB International, Oxfordshire, 152-171
Nemeth T, Mathe-Gaspar G, Radimszky L, Gyori Z (2007): Effect of nitrogen fertilizer on the nitrogen, sulphur and carbon contents of canola (Brassica napus L.) grown on a cal-careous chernozem soil. Cereal Res. Commun. 35: 837-840
Pospišil M, Škevin D, Mustapić Z, Neđeral Nakić S, Butorac J, Matijević D (2007): Fatty Acid Composition in Oil of Re-cent Rapeseed Hybrids and 00-Cultivars. Agric. Conspec. Sci. Vol. 72: 187-193
Službeni list SFRJ (1987): Pravilnik o kvalitetu semena poljopri-vrednog bilja, 47/87
Sana M A, Ali M, Malik A, Saleem M F, Rafi q M (2003): Com-parative yield potential and oil contents of different canola cultivars (Brassica napus L.). Pak. J. Agron. 2: 1-7
Vujaković M, Marjanović-Jeromela A, Marinković R, Nikolić Z, Radić V, Tatić M (2008): Genotype specifi city of rape-seed (Brassica napus L.) seed quality. Conference Proceedin-gs. International Conference “Conventional and Molecular Breeding of Field and Vegetable Crops”. 24-27 November, Novi Sad: 483-486
Vujaković M, Marjanović-Jeromela A, Jovičić D, Marinković R, Nikolić Z, Crnobarac J, Taški-Ajduković K (2010): Uticaj prihrane na prinos i komponente kvaliteta semena uljane re-pice. Ratar. Povrt. / Field Veg. Crop Res. 47: 539-544

Jovičić D i sar.130
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 125-130
The Effect of Different Doses of NPK Fertilizerson the Quality of Rapeseed Seeds
Dušica Jovičić · Ana Marjanović-Jeromela · Milka Vujaković ·Radovan Marinković · Zvonko Sakač · Zorica Nikolić · Branko Milošević
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Rapeseed (Brassica napus L.) is widely used in the production of oil, animal feed and biodiesel. Quality of oilseed rape depends on the genotype, meteorological conditions and crop management. It is important to provide suffi cient quantities of soil nutrients before planting (basal fertilization), so that young plants may form a leaf rosette and prepare for a period of low winter temperature. This research was performed on four commercial genotypes of rapeseed (Banaćanka, Slavica, Express and Valeska) produced in the growing season 2008-2009. The following parameters were determined: germination, 1000-seed weight, oil content and fatty acids. Seed germinati-on, 1000-seed weight and oil content depended on the genotype, but not on the applied NPK dose. The fatty acids content depended on the genotype and the amount of fertilizer applied.Key words: fatty acids content, germination, oil content, rapeseed, 1000-seed weight

Životna sposobnost semena ozimih grahorica 131
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136originalni naučni rad / original research article
Pokazatelji životne sposobnosti semenaozimih grahorica (Vicia spp.)
Milka Vujaković · Dušica Jovičić · Đura Karagić · Aleksandar Mikić ·Zorica Nikolić· Dragana Petrović · Ksenija Taški-Ajduković
primljeno / received: 30.10.2010. prihvaćeno / accepted: 30.11.2010.© 2011 IFVC
Izvod: U proizvodnom području Srbije najviše se gaji obična grahorica (Vicia sativa L), zatim maljava grahorica (Vicia villosa Roth), pa panonska grahorica (Vicia pannonica Crantz). Najznačajniji problem u proizvodnji semena grahorice predstavlja poleganje. Prinos i kvalitet semena u poleglom usevu mogu biti značajno smanjeni. Zbog toga je vrlo bitno precizno utvrditi kvalitet proizvedenog semena, odno-sno njegovu životnu sposobnost. Ispitivanje životne sposobnosti semena izvršeno je na običnoj, malja-voj i panonskoj grahorici primenom standardnog laboratorijskog testa, hladnog testa, Hiltner testa i tri varijante testa ubrzanog starenja (39oC,72 h; 39oC, 96 h; 45oC, 48 h). Obična grahorica je imala najvišu vrednost klijavosti semena (90%) primenom standardnog laboratorijskog testa, dok su nepovoljni uslovi primenjeni u vigor testovima uticali na smanjenje klijavosti semena. Parametri porasta nadzemnog dela ponika su bili najniži kod primene Hiltner testa. Dužina i masa korenovog sistema je bila najniža pri-menom testa ubrzanog starenja. Kod maljave i panonske grahorice primenom Hiltner testa dobijene su najniže vrednosti za sve ispitivane parametre. Primena različitih varijanti testa ubrzanog starenja ukazuje da ispitivani parametri ne zavise od primenjene temperature, dok dužina trajanja temperature utiče na smanjenje životne sposobnosti semena grahorica.Ključne reči: grahorica, klijavost semena, porast ponika, vigor testovi
M. Vujakovic · D. Jovičić · Đ. Karagić · A. Mikić · Z. Nikolić· D. Petrović · K. Taški-AjdukovićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
U Srbiji se komercijalno gaje tri vrste ozime grahorice. Po površini koju zauzima na prvom mestu se nalazi obična grahorica (Vicia sativa L), zatim maljava grahorica (Vicia villosa Roth), pa pa-nonska grahorica (Vicia pannonica Crantz). Površi-ne koje zauzima se kreću od 3.000 ha do 7.000 ha (Mikić et al. 2006). Grahorice se kod nas najčešće gaje za proizvodnju kabaste stočne hrane, mada se takođe koriste za proizvodnju zrna i kao zele-nišno đubrivo. Pri proizvodnji semena poleganje predstavlja značajan problem. Usev počinje da poleže već u fazi cvetanja, prizemni deo biljaka truli, broj formiranih mahuna je mali, nalivanje semena u mahunama je usporeno, a prinos i kva-litet semena vrlo su niski (Karagić et al. 2008). Pored poleganja, nepovoljni vremenski uslovi u vreme nalivanja semena i žetve mogu značajno smanjiti prinos i kvalitet semena.
Utvrđivanje kvaliteta semena i njegove život-ne sposobnosti ukazuje na to koje partije semena mogu da se nađu na tržištu, te je iz tog razloga potrebno dobro razmotriti metode i testove koji se upotrebljavaju prilikom ispitivanja kvaliteta semena, kvaliteta partije semena, odnosno utvr-đivanja životne sposobnosti semena, tj. vigora semena (Milošević i sar. 2010). Uslovi u kojima se seme nalazi za vreme ispitivanja često su u direktnoj suprotnosti sa uslovima koji se obično nalaze u polju. Nicanje u polju zavisi od vlažnosti i temperature zemljišta, kvaliteta setve, a poseb-no od životne sposobnosti semena. Termin ži-votna sposobnost (vigor) se generalno koristi za opisivanje fi zioloških karakteristika semena koji kontrolišu njegovu sposobnost da brzo klija u ze-mljištu i da toleriše razne činioce spoljne sredine, uglavnom negativne.
Ispitivanje životne sposobnosti semena pu-tem različitih vigor testova je veoma značajno,
Semenarstvo / Seed Sciencew
ww.
nsse
me.
com
/jour
nal.h
tml
( )Ovo istraživanje je rezultat projekta TR20090: Ministarstva za nauku i tehnološki razvoj Republike Srbije / This research results from project TR20090 funded by Ministry of Science and Technological Develo-pment of the Republic of Serbia

Vujaković M i sar.132
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
s obzirom da vigor testovi daju rezultate koji su često u boljim korelacionim odnosima sa rezul-tatima poljskog nicanja u nepovoljnim uslovima sredine od rezultata standardne laboratorijske klijavosti (Johansen & Wax 1978). Standardni test klijavosti je dobar pokazatelj kvaliteta i ži-votne sposobnosti semena, kojim se može pred-viditi poljsko nicanje, ukoliko su uslovi zemljišta skoro idealni (Duurant & Gummerson 1990).
U našim agroekološkim uslovima, u vreme se-tve ozimih grahorica, javljaju se nepovoljni uslovi za klijanje i nicanje što se odražava na ujednače-nost nicanja, postizanje optimalnog sklopa bilja-ka, a na kraju i na sam prinos. Zbog toga je cilj ovoga rada bio da se utvrdi životna sposobnost semena kod različitih vrsta iz roda Vicia i da se utvrde vrste koje bolje tolerišu nepovoljne uslove u vreme klijanja i nicanja.
Materijal i metod rada
Ispitivanje životne sposobnosti semena izvr-šeno je 2009. u Laboratoriji za ispitivanje seme-na Instituta za ratarstvo i povrtarstvo u Novom Sadu kod obične, maljave i panonske grahorice. Za ispitivanje je korišćeno seme sorti Neoplanta (obična grahorica), NS Viloza (maljava grahori-ca) i Panonka (panonska grahorica). Sve sorte stvorene su u Institutu za ratarstvo i povrtar-stvo, Novi Sad. Ispitivanje životne sposobnosti
semena izvršeno je primenom standardnog la-boratorijskog metoda i vigor testova.
Standardni laboratorijski metod je primenjen na 4 x 100 semena (ISTA 2009), za podlogu je korišćen pesak. Kao predtretman za ispitivanje semena kori-šćen je metod izlaganja semena niskoj temperaturi (5–10oC) u period od 5 dana. Nakon tog perioda održavana je temperatura od 20oC. Inkubacioni pe-riod trajao je 14 dana, nakon čega je utvrđena kli-javost semena i parametri porasta ponika (dužina i masa nadzemnog dela i korenovog sistema). Ovi parametri određeni su i kod vigor testova.
Od vigor testova primenjeni su hladni test, Hiltner test i test ubrzanog starenja (TUS). Me-todom hladnog testa ispitano je 4 x 50 semena. Seme je stavljeno u mešavinu zemlje i peska u od-nosu 2:1 (Hampton & TeKrony 1995). Seme je izloženo 7 dana temperaturi od 10oC, a potom 4 dana temperaturi od 20oC. Kod Hiltner testa kao podloga se koristi vlažan pesak na koji se stavlja seme koje se pokriva slojem lomljene cigle de-bljine 3 cm (Hampton & TeKrony 1995). Seme je 6 dana izloženo niskoj temperaturi 5–10oC, a potom 14 dana temperaturi od 20oC.
Test ubrzanog starenja je izvršen prema Hamp-ton & TeKrony (1995) u vodenom kupatilu gde je relativna vlažnost vazduha 100%. Test je pri-menjen u tri varijante: (i) temperatura od 39oC u periodu od 72 h, (ii) temperatura od 39oC u peri-odu od 96 h, (iii) temperatura od 45oC u periodu
Tabela 1. Klijavost semena i parametri porasta ponika kod obične grahorice (Vicia sativa L.) primenom različitih vigor testovaTable 1. Seed germination and seedling growth parameters in common vetch (Vicia sativa L.) using different vigor tests
TestoviTests
Klijavost semenaSeed
germination (%)
Nadzemni deo ponikaAbove ground part of seedling
Korenov sistemRoot system
DužinaLenght(mm)
MasaMass(g)
DužinaLenght(mm)
MasaMass(g)
Standardni laboratorijski testStandard laboratory test 90 172,0 1,54 142,8 0,63Hladni testCold test 80 126,8 1,15 136,8 0,52Hiltner testHiltner test 77 65,0 0,69 77,7 0,42TUS (39oC, 72h)AA (39oC, 72h) 87 98,5 0,87 73,2 0,32TUS (39oC, 96h)AA (39oC, 96h) 76 91,5 0,81 67,7 0,30TUS (45oC, 48h)AA (45oC, 48h) 87 99,7 0,87 74,0 0,34NZR 0,05LSD 0,05 3,158 8,648 0,065 9,653 0,035

Životna sposobnost semena ozimih grahorica 133
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
od 48 h. Nakon navedenih tretmana u vodenom kupatilu, seme je stavljeno u vlažan pesak, na temperaturu 20oC u vremenu od 14 dana.
Dobijeni rezultati su statistički obrađeni prime-nom analize varijanse u statističkom programu MSTAT. Značajnost razlika utvrđena je za prag značajnosti 0,05. Izračunat je i koefi cijent korela-cije između dužine nadzemnog dela i korenovog sistema i njihove mase.
Rezultati i diskusija
Klijavost semena obične grahorice dobijene primenom standardnog laboratorijskog metoda iznosila je 90%, dok su primenom vigor testova dobijene statistički značajno niže vrednosti (Tab. 1). Primenom testa ubrzanog starenja klijavost semena se kretala od 76% (varijanta 39oC za 96 h) do 87% (varijanta 39oC za 72 h i 45oC za 48 h). Razlika između ove dve vrednosti statistički je značajna. Dužina i masa nadzemnog dela ponika je zavisila od primenjenog testa. Statistički zna-čajno više vrednosti su dobijene primenom stan-dardnog laboratorijskog testa (172,0 mm/1,54 g) u odnosu na druge testove (Tab. 1). Različite va-rijante testa ubrzanog starenja su uticale na ispiti-vani parametar i najniža vrednost je utvrđena pri varijanti 39oC za 96h (91,5 mm/0,81 g). Utvrđen je i pozitivan koefi cijent korelacije između ova
dva parametra r = 0,96. Dužina korenovog siste-ma je bila statistički značajno viša kod primena standardnog laboratorijskog testa (142,8 mm) i hladnog testa (136,8 mm) u odnosu na druge pri-menjene vigor testove (Tab. 1). Primenom stan-dardnog laboratorijskog testa dobijene su stati-stički značajno više vrednosti mase korenovog sistema (0,63 g), a najniže vrednosti su dobijene primenom testa ubrzanog starenja u svim varijan-tama (0,30–0,34 g). Kao i kod nadzemnog dela, tako i kod korenovog sistema je dobijen pozitivan koefi cijent korelacije za dužinu i masu r = 0,914.
Maljava grahorica je imala statistički značajno najvišu klijavost semena primenom standardnog laboratorijskog testa (93%), a statistički značajno najniža vrednost je dobijena primenom Hiltner testa (72%) (Tab. 2). Primenom različitih varijan-ti testa ubrzanog starenja, varijanta 39oC za 96h (79%) imala je statistički značajno nižu klijavost semena u odnosu na druge dve primenjene va-rijante (87% i 89%). Dužina i masa nadzemnog dela ponika je bila statistički značajno najniža pri-menom Hiltner testa (64,5 mm/0,44 g) (Tab. 2). Kao i kod obične grahorice utvrđen je pozitivan koefi cijent korelacije između ova dva parametra r = 0,917. Primenom Hiltner testa dužina (63 mm) i masa korenovog sistema (0,18 g) su bile statistič-ki značajno niže u odnosu na druge primenjene testove (Tab. 2).
Tabela 2. Klijavost semena i parametri porasta ponika kod maljave grahorice (Vicia villosa Roth) primenom različitih vigor testovaTable 2. Seed germination and seedling growth parameters in hairy vetch (Vicia villosa Roth) using different vigor tests
TestoviTests
Klijavost semena
Seed germination
(%)
Nadzemni deo ponikaAbove ground part of seedling
Korenov sistemRoot system
DužinaLenght(mm)
MasaMass(g)
DužinaLenght(mm)
MasaMass(g)
Standardni laboratorijski testStandard laboratory test 93 164,0 1,20 150,3 0,41Hladni testCold test 84 163,8 1,42 111,3 0,29Hiltner testHiltner test 72 64,5 0,44 63,0 0,18TUS (39oC, 72h)AA (39oC, 72h) 87 156,0 1,10 141,0 0,38TUS (39oC, 96h)AA (39oC, 96h) 79 151,3 1,09 130,8 0,36TUS (45oC, 48h)AA (45oC, 48h) 89 158,3 1,10 147,8 0,39NZR 0,05LSD 0,05 2,410 5,248 0,082 5,682 0,035

Vujaković M i sar.134
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
Panonska grahorica je imala nižu klijavost u odnosu na druge dve ispitivane vrste. Klijavost se kretala od 68% (TUS, varijanta 39oC za 96 h) do 75% (Hiltner test (Tab. 3). Dužina i masa nadze-mnog dela (39 mm/0,47 g) i korenovog sistema (35,2 mm/0,18 g) je bila statistički značajno naj-niža kod primene Hiltner testa.
Primena niske temperature u hladnom testu nije negativno uticala na životnu sposobnost grahorice, što je utvrđeno i prethodnim ispiti-vanjima Vujaković et al. (2008). AOSA (2002) navodi da se primenom hladnog testa može proceniti fi ziološko oštećenje semena koje je uzrokovano produženim i neuslovnim skladi-štenjem, kao i oštećenjima od mraza i suše.
Primena sloja lomljene cigle u Hiltner testu delovala je negativno na porast i masu nadze-mnog dela i korenovog sistema. Sloj lomljene ci-gle predstavlja fi zičku prepreku prilikom porasta ponika, pa samo seme koje poseduje visok vigor formira tipičan i snažan ponik (Hampton & Te-Krony 1995). Brzina kojom seme klija i brzina porasta ponika može da zavisi od genetičkih ra-zlika u veličini semena, osobini semenjače i he-mijskom sastavu (Ferguson 1995). Analizu para-
metara porasta ponika treba vršiti samo unutar jedne sorte, a ne između sorti, jer sorte mogu da imaju nasledno različite stope rasta ponika.
Primenom testa ubrzanog starenja korišće-ne su tri varijante gde je menjana temperatura i dužina izlaganja semena nepovoljnim uslovima. McDonald (2004, cit. prema Milivojević 2005) je
mišljenja da treba primenjivati niže temperature za TUS, a produžiti period dejstva temperature, jer na 45oC dolazi znatno brže do denaturacije proteina, pa uslovi starenja sve manje odgovaraju uslovima prirodnog starenja. Treba voditi računa o variranju temperature i ono ne sme biti veće od ± 0,3oC (TeKrony 2003). U našem eksperimentu temperatura nije uticala na klijavost semena, ali je klijavost značajno zavisila od dužine tretmana. Sa povećanjem dužine tretmana došlo je do značaj-nog smanjenja klijavosti semena kod sve tri vrste grahorice. Do ovakvih rezultata je došao i Sama-rah (2006) u svojim istraživanjima. Test ubrzanog starenja omogućava da se utvrdi da li određena partija semena ima manju ili veću sposobnost očuvanja klijavosti i da relativno precizno odre-di period bezbednog čuvanja semena u skladištu (Milošević et al. 2010).
Tabela 3. Klijavost semena i parametri porasta ponika kod panonske grahorice (Vicia pannonica Crantz) primenom različitih vigor testovaTable 3. Seed germination and seedling growth parameters in Hungarian vetch (Vicia pannonica Crantz.) using different vigor tests
TestoviTests
Klijavost semena
Seed germination
(%)
Nadzemni deo ponikaAbove ground part of
seedling
Korenov sistemRoot system
DužinaLenght(mm)
MasaMass(g)
DužinaLenght(mm)
MasaMass(g)
Standardni laboratorijski testStandard laboratory test 74 102,8 0,88 76,2 0,39
Hladni testCold test 69 99,5 0,86 73,0 0,37
Hiltner testHiltner test 75 39,0 0,47 35,2 0,18
TUS (39oC, 72h)AA (39oC, 72h) 70 97,7 0,87 77,7 0,39
TUS (39oC, 96h)AA (39oC, 96h) 68 89,5 0,76 69,7 0,34
TUS (45oC, 48h)AA (45oC, 48h) 74 98,5 0,86 76,7 0,38
NZR 0,05LSD 0,05 3,589 4,923 0,043 5,053 0,035

Životna sposobnost semena ozimih grahorica 135
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
Zaključci
Obična grahorica je imala najvišu vrednost klijavosti semena (90%) primenom standardnog laboratorijskog testa, dok su nepovoljni uslovi primenjeni u vigor testovima uticali na smanjenje klijavosti semena. Parametri porasta nadzemnog dela ponika su bili najniži kod primene Hiltner testa. Dužina i masa korenovog sistema je bila najniža primenom testa ubrzanog starenja. Kod maljave i panonske grahorice primenom Hiltner testa dobijene su najniže vrednosti za sve ispiti-vane parametre. Primena različitih varijanti testa ubrzanog starenja ukazuje da ispitivani parametri ne zavise od primenjene temperature, dok dužina trajanja temperature utiče na smanjenje životne sposobnosti semena grahorica.
Literatura
AOSA (2002): Seed Vigour Testing Handdbook, Contri-bution No. 32 to the Handbook of Seed Testing Association of Offi cial Seed Analysts, NE, USA
Durrant M J, Gummerson R J (1990): Factors associ-ated with germination of sugarbeet seed in the standard test establishment in the fi eld. Seed Sci. Technol. 18: 1-10
Ferguson J (1995): An introduction to seed vigour te-sting. In: Van de Vanter H. A. (ed.) ISTA Seed Vigor Testing Seminar, ISTA 1-10
Hempton J G, TeKrony D M (1995): Handbook of Vigour Test Methods. International Seed Testing Association
ISTA (2009): International Rules for Seed Testing. Inter-national Seed Testing Association, Switzerland
Johansen R R, Wax L M (1978): Relationship of soybean germination and vigour test to fi eld performance. Agron. J. 70: 273-276
Karagić Đ, Katić S, Mihailović V, Vasiljević S, Mikić A, Milić D (2008): What a seed producer needs from a plant breeder – the example of Novi Sad (NS) forage legumes varieties. Book of Abstracts of the Second Grain Legumes Technology Transfer Platform (GL-TTP) Workshop. Novi Sad, Serbia, 27-28 November, 56
Milivojević M (2005): Primena vigor testova u ocenjiva-nju kvaliteta semena kukuruza (Zea mays L.). Magi-starska teza, Univerzitet u Novom Sadu, Poljopri-vredni fakultet
Mikić A, Ćupina B, Katić S, Karagić Đ (2006): Impor-tance of annual forage legumes in supplying plant proteins. Zbornik radova Instituta za ratarstvo i povrtarstvo 42: 91-103
Milošević M, Vujaković M, Karagić Đ (2010): Vigour tests as Indicators of seed viability. Genetika 42: 103-118
Samarah H N (2006): Effect of air-drying immature seeds in harvested pods on seed quality of com-mon vetch (Vicia sativa L.). New Zealand J. Agric. Res. 49: 331-339
TeKroni D M (2003): Precision is an essential compo-nent in seed vigour testing. Seed Sci. Technol. 31: 435-447
Vujaković M, Nikolić Z, Milošević M, Mikić A, Ignjatov M (2008): Germination and vigour of fodder pea and vetch seed. Book of abstracts 43rd Croatian & 3rd Internacional symposium on agriculture, 18-21 February, Opatija, 327

Vujaković M i sar.136
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 131-136
Indicators of Winter Vetch (Vicia spp.) Seed Vigor
Milka Vujaković · Dušica Jovičić · Đura Karagić · Aleksandar Mikić ·Zorica Nikolić· Dragana Petrović · Ksenija Taški-Ajduković
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Common vetch (Vicia sativa L), hairy vetch (Vicia villosa Roth), and Pannonian vetch (Vicia pannonica Crantz) are the most often grown varieties in the production area of Serbia. The most important problem in the production of vetch seed is lodging. Yield and seed quality in lodged crops can be signifi cantly reduced. Therefore, it is very important to accurately estimate the quality, and viability of produced seed. Common vetch, hairy vetch and Pannonian vetch were submitted to vigor testing using standard laboratory test, cold test, Hiltner test, and three variants of accelerated aging tests (39oC,72 h; 39oC, 96 h; 45oC, 48 h). When the standard laboratory test was appli-ed, the common vetch achieved the highest seed germination value of 90%, while decreased rate of seed germina-tion was due to unfavorable conditions applied in vigor tests. Above ground seedling growth parameters were the lowest when Hiltner test was applied. Length and mass of the root system were the lowest when accelerated aging test was applied. Hairy vetch obtained the lowest values for all tested parameters when Hiltner test was applied. Application of different variants of accelerated aging tests indicated that tested parameters were not dependent on the applied temperature, while reduction in legumes vigor was under the infl uence of temperature duration. Key words: seed germination, seedling growth, vetch, vigor tests

Prinos i semenski kvalitet soje u zavisnosti od uslova godine 137
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142originalni naučni rad / original research article
Prinos i semenski kvalitet sojeu zavisnosti od uslova godine
Vojin Đukić · Svetlana Balešević-Tubić · Vuk Đorđević · Mladen Tatić · Gordana Dozet · Goran Jaćimović · Kristina Petrović
primljeno / received: 29.10.2010. prerađeno / revised: 30.11.2010. prihvaćeno / accepted: 09.12.2010.© 2011 IFVC
Izvod: U dvogodišnjem istraživanju ispitivan je uticaj agrometeoroloških činilaca (temperatura i padavi-na) na prinos i semenske kvalitete zrna soje. Ogled je postavljen u četiri ponavljanja na oglednim parcela-ma Instituta za ratarstvo i povrtarstvo na Rimskim Šančevima u 2009. i 2010. Vremenski uslovi značajno su uticali na ispitivana svojstva, odnosno u godini sa povoljnim meteorološkim uslovima (2010) ostvaren je veći prinos i kvalitet semena ispitivanih sorti soje. Uočena je umerena pozitivna korelacija između prinosa i klijavosti semena.Ključne reči: klijavost, masa hiljadu zrna, padavine, prinos, soja, temperatura
V. Đukić · S. Balešević-Tubić · V. Đorđević · M. Tatić · K. PetrovićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]. DozetFakultet za biofarming Megatrend Univerziteta, Maršala Tita 39, 24300 Bačka Topola, SrbijaG. Jaćimović Poljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbija
Uvod
Soja je biljna vrsta koja intenzivno reaguje na agrometeorološke uslove u toku vegetacije. Cen-tar porekla soje su suptropski severoistočni delo-vi Kine, odnosno regioni sa dosta padavina i po-višenom vlagom u vazduhu. Za visoke i stabilne prinose soje, neophodno je obezbediti dovoljno vlage u zemljištu, naročito u periodu cvetanja, formiranja mahuna i zrna, kao i u periodu nali-vanja zrna. Najbolji pokazatelj povoljnosti uslova jedne godine ili rejona za proizvodnju soje, po-red ostvarenih prinosa takođe je masa 1000 zrna (Dozet 2006). Nedostatak vlage u kritičnim faza-ma razvoja značajno umanjuje prinos i smanjuje masu hiljadu zrna. U trogodišnjim istraživanjima Dozet (2009), Đukić (2009) i Đukić i sar. (2010) ukazuju na značajan uticaj godine na prinos i masu hiljadu zrna soje. Ovi autori zapažaju po-većanje prinosa i mase hiljadu zrna u godinama sa većim količinama padavina, dok su u najsuš-
nijim godinama oni bili najniži. Pored smanjenja mase hiljadu zrna, što se negativno odražava i na prinos, u godinama sa pojavom suše u fazama formiranja i nalivanja zrna dolazi do značajnog umanjenja klijavosti semena soje.
Cilj ovih istraživanja bio je da se ispita uticaj godine, odnosno padavina i temperatura na pri-nos, masu hiljadu zrna i klijavost semena soje u uslovima proizvodnje bez navodnjavanja.
Materijal i metod
U Institutu za ratarstvo i povrtarstvo na Rim-skim Šančevima kontinurano se izvodi ogled sa sortama soje koje su široko zastupljene u pro-izvodnji, a u ovom radu su prikazani rezultati iz 2009. i 2010. Ogled se postavlja u četiri po-navljanja po randomiziranom blok sistemu sa ukupno 8 sorti soje različitih grupa zrenja. Od veoma ranih sorti (00 grupe) u ogledu su bile zastupljene Gracia i Merkur, od ranih (0 grupa) Galina i Valjevka, srednjestasnih (I grupa) Sava i Victoria, a od kasnih sorti (II grupa zrenja) Ve-nera i Trijumf.
Površina osnovne parcelice iznosila je 15 m2 (5 m x 3 m). Oko ogleda posejana su četiri reda kao zaštitna zona. Gustina setve za veoma rane sorte soje iznosila je 50 x 3,6 cm (550.000 biljaka ha-1), za rane sorte 50 x 4 cm (500.000 biljaka ha-1), srednje-stasne 50 x 4,4 cm (450.000 biljaka ha-1) i za kasne
Semenarstvo / Seed Sciencew
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Đukić V i sar.138
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
sorte druge grupe zrenja 50 x 5 cm (400.000 bi-ljaka ha-1). U obe godine ispitivanja primenjene su standardne agrotehničke mere za proizvodnju soje. Setva je obavljana mašinski, a žetva četiri središnja reda obavljena je kombajnom za mikrooglede. Na-kon žetve izmerena je masa i vlažnost semena, te obračunat prinos (kg ha-1) sa 14% vlage. Takođe je određena masa hiljadu zrna i klijavost semena soje po metodici ISTA (1999).
Rezultati istraživanja obrađeni su statistički, analizom varijanse specifi čnog jednofaktorijalnog ogleda (isti ogled izveden u više godina) (Had-živuković 1973), pomoću statističkog softvera Mstatc (procedura One factor randomized complete block design combined over years), a značajnost razli-ka testirana je LSD testom. Međuzavisnost pri-nosa i klijavosti semena ispitana je korelacionom analizom, a rezultati rada prikazani su tabelarno i grafi čki.
Rezultati i diskusija
Godine u kojima je obavljeno istraživanje ve-oma su se razlikovale po vremenskim uslovima (Tab. 1). Tako su april, maj i avgust 2009. bili znatno topliji u odnosu na 2010. i višegodišnji prosek, dok su u junu i julu veće vrednosti sred-njih mesečnih temperatura zabeležene u 2010.
Na početku vegetacionog perioda u 2009. (april) bilo je značajno manje padavina u odnosu na višegodišnji prosek (palo je svega 3,6 mm kiše; odnosno svega nešto iznad 7% padavina od više-godišnjeg aprilskog proseka). Početkom jula iste godine bilo je značajnijih padavina, nakon čega se
javio defi cit koji se produžio sve do kraja vegeta-cionog perioda soje.
Visoke temperature vazduha tokom aprila 2009. (za 3,2°C više od višegodišnjeg proseka) i nedostatak padavina uticali su na isušivanje povr-šinskog sloja zemljišta, što je imalo za posledicu zaostajanje soje u porastu i razvoju. Pojava viso-kih temperatura i suše zabeležena je i u većem delu maja, što je dodatno ugrozilo razvoj soje. Tek je krajem maja došlo do pada temperatura na prosečne majske vrednosti i pojave prvih kiša, što je donekle ublažilo posledice dugotrajne suše. Početak juna obeležilo je svežije vreme sa čestim i obilnim padavinama, što je znatno ubrzalo razvoj soje u narednom periodu. Period gotovo svakod-nevnih padavina nastavljen je sve do polovine jula meseca. Zalihe vlage su znatno povećane zahva-ljujući ovim padavinama, tako da je soja u tom periodu imala dovoljno pristupačne vlage. Sredi-nom jula nastupilo je toplo i sunčano vreme, što je pogodovalo intenziviranju razvoja soje, no suv i topao period nastavljen je i početkom avgusta, što je zalihe vlage u zemljištu ponovo spustilo na minimum. U drugoj dekadi avgusta došlo je do značajnijeg osveženja sa padavinama, koje su de-limično popravile stanje vlažnosti zemljišta, što je uz povoljne temperature omogućilo dobre uslove za formiranje i nalivanje zrna. Do kraja avgusta
bili su uglavnom povoljni toplotni uslovi, koji su u završnim fazama razvoja soje bili važniji od sta-nja vlage u zemljištu.
U 2010. tokom vegetacionog perioda soje za-beleženo je 683,6 mm padavina, što je 87,6% više u odnosu na višegodišnji prosek i 152% više u
Tabela 1. Vremenski uslovi u godinama istraživanja (2009-2010)Table1. Meteorological conditions during research years (2009-2010)
Mesec
Month
Srednja mesečna
temperatura
Average monthly temperature (oC)
Višegodišnji
prosek
Long-term average
Padavine
Rainfall
(mm)
Višegodišnji
prosek
Long-term average
2009 2010 1964-2004 2009 2010 1964-2004April 14,6 12,3 11,4 3,6 63,7 48,8Maj 18,6 16,9 16,8 50,4 113,7 59,5Jun 19,6 20,2 19,9 127,2 171,8 85,9Jul 22,8 23,1 21,4 58,1 99,0 68,2
Avgust 22,9 21,9 21,0 19,1 167,7 56,9Septembar 19,2 16,1 16,8 13,1 67,7 45,1Veg. period
Vegetation period
19,6 18,4 17,9 271,5 683,6 364,4

Prinos i semenski kvalitet soje u zavisnosti od uslova godine 139
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
odnosu na 2009. Vremenski uslovi u vegetacio-nom periodu u 2010. povoljno su delovali na rast i razviće biljaka, što se odrazilo kako na prinos, tako i na semenski kvalitet zrna soje.
Maj i jun 2010. karakterisalo je nešto toplije vreme sa znatno više padavina u poređenju sa višegodišnjim mesečnim vrednostima. Prvu po-lovinu jula takođe su obeležile svakodnevne plju-skovite padavine, često praćene nepogodama. Tek se krajem prve i u drugoj dekadi jula pojavilo veoma toplo, sunčano i suvo vreme, što je pogo-dovalo razvoju soje.
Povoljniji klimatski uslovi, odnosno više me-sečne temperature u periodu april-avgust 2010.
uz istovremeno znatno obilnije padavine u odno-su na višegodišnje mesečne proseke, uticale su na znatno bolji razvoj soje, što se konačno odrazilo i na značajno veće prinose kod svih ispitivanih sorti (Tab. 2).
Posmatrajući prosečne vrednosti prinosa soje po godinama istraživanja, uočen je statistički ve-oma značajno niži prinos u 2009. (3.400 kg ha-1)
u odnosu na drugu godinu istraživanja (4.526 kg ha-1). Povećanje prinosa u 2010. kod ispitivanih sorti kretalo se u rasponu od 16,1% kod sorte Gracia do 61,3% kod sorte Valjevka.
Razlike prinosa u posmatranim godinama posledica su različite količine padavina i tempe-ratura vazduha u vegetacionom periodu, što je u saglasnosti sa rezultatima koje iznosi Dozet (2006). Pre neposrednog uvođenja u proizvod-nju novostvorene sorte soje se testiraju duži niz godina na više lokaliteta, kako bi se ustanovila njihova stabilnost i otpornost na različite stresne uslove. U cilju postizanja što stabilnijeg prinosa i u stresnim uslovima, kakvi se javljaju u godinama
sličnim 2009. selekcioneri primenjuju sve savr-šenije metode u oplemenjivanju soje (Đorđević i sar. 2008).
U nepovoljnoj godini (2009) statistički veoma značajno niži prinos uočen je kod sorte Gracia (2.800 kg ha-1) u odnosu na ostale ispitivane ge-notipove soje (Tab. 2). Najviši prinos u ovoj go-dini imala je sorta Trijumf (4.012 kg ha-1), što je
Tabela 2. Prinos (kg ha-1), masa hiljadu zrna (g) i klijavost semena soje (%) u 2009. i 2010. Table 2. Soybean yield (kg ha-1), thousand seed mass (g) and seed germination (%) in 2009 and 2010
SortaVariety
Prinos Yield (kg ha-1)
Masa 1000 zrnaSeed weight (g)
KlijavostGermination (%)
2009 2010Δ
(2010-2009)
ProsekAverage 2009 2010
Δ (2010-2009)
ProsekAverage 2009 2010
Δ (2010-2009)
ProsekAverage
Gracia 2800 3253 453 3026 170,2 222,4 52,2 196,3 86,0 91,0 5,0 88,5
Merkur 3307 4414 1107 3861 152,0 163,0 11,0 157,5 86,8 91,0 4,2 88,9
Galina 3357 4556 1199 3957 147,2 159,2 12,0 153,2 94,0 96,0 2,0 95,0
Valjevka 3006 4850 1844 3928 121,1 138,5 17,4 129,8 89,8 90,3 0,5 90,0
Sava 3749 4705 956 4227 171,3 198,0 26,7 184,7 88,0 95,3 7,3 91,6
Victoria 3428 4856 1428 4142 147,3 162,3 15,0 154,8 90,0 97,3 7,3 93,6
Venera 3538 4508 970 4023 137,8 151,8 14,0 144,8 95,3 95,0 -0,3 95,1
Trijumf 4012 5070 1058 4541 128,1 151,0 22,9 139,5 90,0 96,3 6,3 93,1
Prosek Average 3400 4526 1126 3963 146,9 168,3 21,4 157,6 90,0 94,0 4,0 92,0
Faktor / FactorPrinos
Yield (kg ha-1)Masa 1000 zrna Seed weight (g)
Klijavost Germination (%)
LSD0,05 LSD0,01 LSD0,05 LSD0,01 LSD0,05 LSD0,01
Godina Year 35 54 1,5 2,3 1,5 2,3Sorta Variety 65 88 2,3 3,0 2,2 2,9God. x Sorta Year x Variety 93 124 3,2 4,3 3,1 4,1

Đukić V i sar.140
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
statistički veoma značajno više u odnosu na sorte Gracia, Merkur, Galina, Valjevka, Victoria i Ve-nara. Kod sorte Gracia je takođe u 2010. zabele-žen veoma značajno niži prinos (3.253 kg ha-1) u odnosu na ostale sorte soje zastupljene u ogledu. Najviši prinos 2010, kao i 2009. zabeležen je kod sorte Trijumf (5.070 kg ha-1) sa statistički značaj-nom razlikom u odnosu na sorte Gracia, Merkur, Galina, Venera i Sava. Može se uočiti da je razlika po sortama skoro ista u obe godine ispitivanja, što ukazuje na činjenicu da uslovi uspevanja to-kom vegetacije imaju veći uticaj na prinos, nego što je uticaj sorte (Vidić i sar. 2009).
Posmatrajući sorte po grupama zrenja (Tab. 3.), uočava se da su najveći prinos zrna u 2009. (3.775 kg ha-1) ostvarile sorte II grupe zrenja (Ve-nera i Trijumf), dok su u 2010. sorte II, I i nulte grupe formirale gotovo identične prinose (4.789 kg ha-1, 4.781 kg ha-1, odnosno 4.703 kg ha-1). Ovo takođe ukazuje na postojanje značajnih razlika iz-među ispitivanih godina. Naime, sorte različitih grupa zrenja su u 2009. različito odreagovale na
nepovoljne vremenske uslove, dok su povoljni klimatski uslovi u proizvodnoj 2010. podjedna-ko favorizovali sorte svih grupa zrenja do gotovo potpunog iskorišćenja genetskog potencijala.
Najveću reakciju na povoljne klimatske uslove u 2010. (u poređenju sa 2009) pokazale su sorte nulte grupe zrenja reagujući povećanjem prinosa zrna od prosečno 1.522 kg ha-1 (47,8%), zatim slede sorte I i II grupe zrenja (33,2% i 26,9%), dok su najranije sorte najslabije odreagovale ap-solutnim povećanjem prinosa (780 kg ha-1), iako ovo povećanje posmatrano u procentima (25,5%) nije bilo nimalo zanemarljivo. Adaptibilnost, mo-gućnost gajenja u konkretnom regionu se ocenju-je na osnovu sortnih ogleda i za naše područje je utvrđeno da su najpogodnije sorte iz grupe zre-nja 0, I i II. U proizvodnji soje najbolje je odlučiti se za sorte različite dužine vegetacije, jer moguća pojava stresnih uslova neće zateći sve sorte u istoj
fazi razvoja, od čega zavisi otpornost biljke, čime će se umanjiti ili čak izbeći negativni efekat stresa (Crnobarac i sar. 2008).
Masa 1000 zrna je svojstvo koje najvećim de-lom zavisi od genetske osnove sorte, ali takođe od spoljnih faktora. To je svojstvo koje direktno utiče na prinos, jer predstavlja jednu od tri osnov-ne komponente prinosa (Đukić i sar. 2009). Pro-sečna vrednost mase hiljadu zrna u 2009. (146,9 g) bila je značajno manja u odnosu na vrednost dobijenu u 2010. (168,3 g). Povećanje mase hi-ljadu zrna u 2010. u odnosu na 2009. bilo je u intrvalu od 7,2% (Merkur) do 30,7% (Gracia).
Najveću masu hiljadu zrna u 2009. godini imala je sorta Gracia (170,2 g) što je statistički veoma značajno više u odnosu na ostale ispitivane sorte. Najniže vrednosti za masu hiljadu zrna zabeleže-ne su kod sorti Valjevka (121,1 g) i Trijumf (128,1 g) što je značajno niže u odnosu na ostale sorte soje. U 2010. sorta Gracia je takođe imala značaj-no veću masu hiljadu zrna (222,4 g) u odnosu na ostale sorte, dok je sorta Valjevka (138,5 g) imala
najmanju masu hiljadu zrna, što je bilo statistički značajno niže u odnosu na ostale genotipove u ogledu (Tab. 2).
Masa hiljadu zrna i prinos soje u direktnoj su vezi sa količinom padavina i temperaturom vaz-duha u vegetacionom periodu soje. Setva semena soje visokog kvaliteta jedan je od veoma važnih agronomskih zahteva (Balešević-Tubić i sar. 2004) zbog čega se i preporučuje da se semenski usevi soje gaje u sistemima za navodnjavanje, kako bi se izbegli stresni uslovi koji ograničavaju prinos i smanjuju kvalitet samog semena. Analizirajući rezultate makroogleda, Vidić i sar. (2009) konsta-tuju da je količina padavina u julu i prvoj polovini avgusta imala veliki uticaj na visinu ostvarenog prinosa.
Kako za prinos i masu hiljadu zrna, tako je i za klijavost semena utvrđena statistički značajno niža vrednost u 2009. (90%) u odnosu na 2010.
Tabela 3. Prinos zrna (kg ha-1) sorti soje po grupama zrenja u 2009. i 2010. Table 3. Soybean grain yield (kg ha-1) at different maturity groups in 2009 and 2010
Grupa zrenja Maturity group 2009 2010
Δ (2010-2009) ProsekAveragekg ha-1 %
00 3054 3834 780 25,5 34440 3182 4703 1522 47,8 3942I 3589 4781 1192 33,2 4185II 3775 4789 1014 26,9 4282

Prinos i semenski kvalitet soje u zavisnosti od uslova godine 141
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
(94%). Razlika u klijavosti semena između ispiti-vanih godina po sortama kretala se od 2% (Ga-lina) do 7% (Sava i Victoria). Najviša klijavost semena u 2009. zabeležena je kod sorte Venera (95%), što je bilo statistički značajno više u od-nosu na sorte Gracia, Merkur, Sava i Trijumf, dok je klijavost semena sorte Gracia bila najniža u odnosu na ostale genotipove. Najvišu klijavost
semena u 2010. imala je sorta Victoria (97%) sa statistički veoma značajnom razlikom u odnosu na sorte Gracia, Merkur i Valjevka. Sorta Valjev-ka imala je visoko značajno nižu klijavost semena (90%) u odnosu na sve ispitivane sorte, osim u odnosu na sortu Merkur.
Nedostatak padavina u 2009. u kritičnom peri-odu vegetacije soje (tokom nalivanja zrna) praćen visokom temperaturom, rezultirao je smanjenjem klijavosti požnjevenog semena, naročito ranih sorti soje Gracia (86%) i Merkur (87%). Viso-ka temperatura i nedostatak padavina u letnjim mesecima dovode do značajnog pada klijavosti u odnosu na seme proizvedeno u uslovima dovolj-ne vlažnosti i srednje dnevne temperature ispod 22oC (Balešević-Tubić i sar. 2001).
Posmatrajući međuzavisnosti prinosa, mase 1000 zrna i klijavosti semena, jedina i to visoko značajna korelaciona zavisnost (r = 0,66**) uo-čena je između prinosa i klijavosti semena (Sl. 1). Iako je masa semena takođe jedna od bitnih kom-poneti prinosa, vezu između ove osobine i prino-sa neophodno je posmatrati u kontekstu uticaja spoljnih faktora (Đukić i sar. 2009). Takođe, od-sustvo veze između klijavosti i mase 1000 zrna ukazuje da je oplemenjivanjem moguće selekci-
Slika 1. Međuzavisnost prinosa i klijavosti semena sojeFig. 1. Interdependence of soybean yield and seed germination
onisati sorte krupnijeg, odnosno sitnijeg zrna, u zavisnosti od zahteva prerađivačke industrije, bez rizika da će takve sorte otežavati proizvodnju semena. Veza između klijavosti semena i prinosa bila je umerena, što pokazuje da ekološki faktori slično utiču na ove dve osobine soje, ali da ipak postoje variranja u zavisnosti od ostalih činilaca tokom vegetacije.
Zaključak
Na osnovu rezultata dvogodišnjih istraživanja uticaja padavina i temperatura na prinos, masu hiljadu zrna i klijavost semena soje, može se za-ključiti sledeće:
Količina i raspored padavina, temperaturni uslovi tokom vegetacionog perioda, vreme poja-ve, trajanje kao i intenzitet suše, značajno odre-đuju visinu prinosa, masu hiljadu zrna i klijavost semena soje.
Povoljniji klimatski uslovi, odnosno više me-sečne temperature u periodu april-avgust 2010. uz istovremeno znatno obilnije padavine u odno-su na višegodišnje mesečne proseke, uticale su na znatno bolji razvoj soje, što se konačno odrazilo i na značajno veće prinose i klijavost semena kod svih ispitivanih sorti.
Na visinu i stabilnost prinosa u proizvodnji soje, kao i na kvalitet semena može se uticati pra-vilnim izborom sorti, odnosno optimalnom za-stupljenošću različitih grupa zrenja.
Uočena je umerena pozitivna koralaciona zavi-snost između prinosa i klijavosti semena.
y = 0,0032x + 79,393
r = 0,66**
84
86
88
90
92
94
96
98
100
2500 3000 3500 4000 4500 5000 5500
Prinos - Yield (kg ha-1)
Klij
avos
t sem
ena
- Ger
min
atio
n(%
)

Đukić V i sar.142
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 137-142
Literatura
Balešević-Tubić S, Hrustić M, Milošević M, Tatić M, Vujaković M (2001): Uticaj suše na kvalitet i prinos semena soje. Zbor-nik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad, 35: 383-390
Balešević-Tubić S, Tatić M, Vujaković M, Miladinović J (2004): Kvalitet semena soje u zavisnosti od lokaliteta gajenja. Zbor-nik radova III međunarodne ECO-konferencije, Novi Sad, 22-25. septembar 2004., 31-35
Crnobarac J, Đukić V, Marinković B (2008): Agrotehnika soje, U: Miladinović, J., Hrustić, Milica, Vidić, M. (red.): Soja. In-stitut za ratarstvo i povrtarstvo, Novi Sad, Sojaprotein, Bečej: 289-322
Dozet G (2006): Prinos i kvalitet soje u zavisnosti od među-rednog razmaka i grupe zrenja u uslovima navodnjavanja, Magistarska teza, Univerzitet u Novom Sadu, poljoprivredni fakultet, Novi Sad, 73
Dozet G (2009): Uticaj đubrenja predkulture azotom i primene Co i Mo na prinos i osobine zrna soje, Doktorska disertacija, Megatrend Univerzitet Beograd, Fakultet za biofarming Bač-ka Topola, Bačka Topola, 154
Yield and Quality of Soybean Seeds as Affected by Growing Conditions
Vojin Đukić1 · Svetlana Balešević-Tubić1 · Vuk Đorđević1 · Mladen Tatić1 · Gordana Dozet2 · Goran Jaćimović3 · Kristina Petrović1
1Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia2 Faculty of Biofarming, University Megatrend, Maršala Tita 39, 24300 Bačka Topola, Serbia
3 Faculty of Agriculture University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
Summary: Effect of environmental conditions (temperatures and precipitation) on soybean yield and seed quality was examined during two years. Trials with four replications were located at experimental fi elds of Institute of Field and Vegetable Crops at Rimski Šančevi. Environmental conditions had signifi cant infl uence on examined traits, and higher yield and seed quality was achieved during benefi cial year. There was a moderate positive relationship between yield and seed germination.Key words: germination, precipitation, soybean, temperature, yield, 1000-seed weight
Đordjevic V, Miladinovic J, Balesevic-Tubic S, Djukic V (2008): Future perspective in soybean breedinig, Second GL-TTP workshop, Novi Sad, Serbia: 27-28 novembar, 2008, Book of abstracts 18
Đukić V (2009): Morfološke i proizvodne osobine soje ispi-tivane u plodoredu sa pšenicom i kukuruzom, Doktorska disertacija, Univerzitet u Beogradu, Poljoprivredni fakultet Zemun, 127
Đukić V, Balešević-Tubić S, Tatić M, Ilić A (2009): Uticaj azota na prinos i masu hiljadu zrna soje. Sel Semen. 15: 73-80
Đukić V, Đorđević V, Popović V, Balešević-Tubić S, Petrović K, Jakšić S, Dozet G (2010): Efekat azota i nitragina na prinos soje i sadržaj proteina. Ratar. Povrt./Field Veg. Crop Res. 47: 187-192
Hadživuković S (1973): Statistički metodi s primenom u poljo-privrednim i biološkim istrazivanjima. Radnički univerzitet »Radivoj Ćirpanov«, Novi Sad, 301-304
ISTA (1999) International Rules for Seed Testing. International Seed Testing Association, Switzerland
Vidić M, Hrustić M, Miladinović J, Đukić V, Đorđević V (2009): Sortni ogledi soje u 2008. godini. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 46: 261-270

Uticaj rokova setve na osobine sadnica šećerne repe 143
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150originalni naučni rad / original research article
Uticaj rokova setve, đubrenja azotom i načinanavodnjavanja na morfološke i fenotipske
osobine sadnica šećerne repe
Milorad Rajić · Borivoj Pejić
primljeno / received: 09.11.2010. prerađeno / revised: 01.12.2010. prihvaćeno / accepted: 23.12.2010.© 2011 IFVC
Izvod: Ispitivan je uticaj dva različita vremenska roka setve, đubrenje azotom i bez njega, kao i dva ra-zličita načina navodnjavanja na morfološke i fenotipske osobine sadnica šećerne repe. Pod dobro razvi-jenim sadnicama semenske šećerne repe podrazumevamo pravilno usaglašene zahteve gajene biljke sa agroekološkim uslovima regiona. Ispitivan je broj biljaka po ha, prečnik korena, sveža masa korena, broj listova, sveža masa nadzemnog dela, masa suve materije nadzemnog dela i masa suve materije korena. Analizom varijanse trofaktorijalnog ogleda u tri godine ispitivanja pokazao se značajan uticaj prvog roka setve na sve posmatrane morfološke i fenotipske osobine sadnica semenske šećerne repe. Predsetve-nim đubrenjem azotom ostvarena je opravdano veća razlika u odnosu na kontrolu, sa svežom masom korena, svežom masom nadzemnog dela i većim prečnikom korena. Različitim načinima navodnjavanja ostvaren je značajno veći broj biljka i sveža masa korena.Ključne reči: đubrenje azotom, navodnjavanje, rokovi setve, sadnice semenske šećerne repe
M. Rajić Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, SrbijaB. PejićPoljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Pod sadnicom semenske šećerne repe podrazumeva se biljka semenske šećerne repe proizvedena od osnovnog semena sa masom od nekoliko stotina grama i 10-15 stalnih listova. Za uspešan porast sadnica semenske šećerne repe u prvim fazama porasta značajnu ulogu ima voda, čijim se ograničavanjem značajno smanjenje masa biljaka (Geerts & Raes 2009). U našim agroekološkim uslovima setva se izvodi u julu i avgustu kada su temperature najveće a padavine najmanje, što znači da uspešna proizvodnja sadnica mora biti u uslovima navodnjavanja. Ako bi se u laboratorijskim uslovima odstranio perikarp semena šećerne repe količina vode za klijanje bi se višestruko smanjila (Akeson et al. 1980). Đubrenje šećerne repe azotom treba bazirati na
rezultatima N-min metode i rasporedu mineralnog azota po dubini (Marinković & Crnobarac 2000).
Posle nicanja mladog ponika đubrenje azotom značajno smanjuje vezanu vodu u lišću šećerne repe, što za posledicu ima smanjenje potrošnje vode u prvim fazama porasta mladog ponika šećerne repe (Pečenov 1988).
Cilj ovoga rada bio je da se u našim agroekološkim uslovima utvrdi uticaj vremena setve, đubrenja azotom i primene načina navodnjavanja na morfološke i fenotipske osobine sadnica semenske šećerne repe.
Materijal i metod
Proučavanja iznete problematike izvršena su na proizvodnim parcelama u Bačkom Petrovcu tokom 2006, 2007. i 2008. Kao eksperimentalni materijal korišćene su roditeljske komponente hibridne sorte Lare. Trofaktorijalni ogled AxBxC sa nivoima (2x2x2) postavljen je po metodi slučajnog blok sistema u četiri ponavljanja. Veličina osnovne parcele bila je 1 m2.Đubrenje azotom je obavljeno predsetveno
sa 70 kg N ha-1. Setva je bila na 50 cm x 12 cm, odnosno isejano je 168.000 semena ha-1. Navodnjavanje je bilo izvođeno kišnim krilima: kod C1 varijante pre setve sa zalivnom normom od 50 mm, a kod varijante C2 posle setve sa zalivnom normom od 10 mm u pet puta.
Gajenje i agrotehnika / Soil and Crop Management w
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Rajić M, Pejić B144
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
Faktor A đubrenjeA1 kontrolaA2 đubrenje, 70 kg N ha-1
Faktor B rokovi setveB1 prvi rok setve 15.07B2 drugi rok setve 15.08Faktor C navodnjavanjeC1 navodnjavanje pre setveC2 navodnjavanje posle setve
Morfološka merenja biljaka rađena su u periodu 05-10. novembar, a uključivala su broj biljaka, prečnik korena, svežu masu korena, broj listova, svežu masu nadzemnog dela, sadržaj suve materije nadzemnog dela i sadržaj suve materije korena.
Rezultati istraživanja i diskusija
Tokom leta poljska klijavost semenske šećerne repe primarno zavisi od vlažnosti zemljišta, kvaliteta predsetvene obrade zemljišta, kao i od kvaliteta semena (Rajić 1992). U toku letnje setve temperature su visoke, a površinski sloj zemljišta brzo se suši. U takvim agroekološkim uslovima šećerna repa koristi malo vode za početni rast, najveći utrošak vode u ovom periodu odlazi na
evaporaciju (Dragović 1987). Analizom varijanse trofaktorijalnog ogleda u prve dve godine ispitivanja utvrđeno je da navodnjavanje posle setve ostvaruje značajno veći broj biljaka u odnosu na navodnjavanje pre setve (Tab. 1), što znači da sukcesivna navodnjavanja posle setve sa manjim zalivnim normama imaju prednost u odnosu na veliku zalivnu normu pre setve. U trećoj godini ispitivanja nije bilo opravdanih razlika u načinu navodnjavanja. U prvom roku setve u prvoj i trećoj godini ispitivanja ostvaren je značajno veći broj biljaka u odnosu na drugi rok setve. Razlike u broju biljaka u drugoj godini ispitivanja bile su takođe značajne u korist drugog roka setve. Prvi rokovi setve u letnjem periodu mogu ostvariti čak do 280.000 biljaka ha-1 sa setvom semenske šećerne repe po jednogodišnjem načinu gajenja (Gorelov et al. 1978). Imeđu kontrolne varijante bez đubriva i varijante sa đubrenjem 70 kg N ha-1 nije bilo značajnih razlika u broju poniklih biljka u sve tri godine ispitivanja.
Primenom đubrenja sa 70 kg N ha-1 ostvaren je značajno veći prečnik korena u drugoj i trećoj godini ispitivanja u odnosu na kontrolu, dok je prvoj godini kontrolna varijanta bez đubriva imala značajno veći prečnik korena prema đubrenju sa 70 kg N ha-1. Prvim rokom setve u sve tri godine
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 117 110 80 84 146 162 85 89 107 111 109 84 98 1202007 88 85 118 119 119 124 118 118 11 112 104 118 102 1202008 145 145 76 75 152 153 82 85 114 114 149 79 113 118
Tabela 1. Broj poniklih biljaka posle setve (1000 ha-1)Table 1. Number of germinated plants after planting (1000 ha-1)
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 20 27 8 11 10 14B 20 27 8 11 10 14C 20 27 8 11 10 14
AB 28 38 11 15 14 19AC 28 38 11 15 14 19BC 28 38 11 15 14 19
ABC 39 53 16 21 20 27

Uticaj rokova setve na osobine sadnica šećerne repe 145
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
ispitivanja ostvaren je visoko značajno veći prečnik korena u odnosu na drugi rok setve (Tab. 2). Neka ispitivanja pokazuju da primenom srednjih količina azota od 90 kg N ha-1 može se ostvariti značajan porast korena šećerne repe u početku vegetacije
(Vamerali et al. 2009). Prečnik korena u našim ispitivanjima bio je značajno veći navodnjavanjem posle setve u sve tri godine ispitivanja.
Masa korena u toku sve tri godine ispitivanja bila je veća sa đubrenjem azotom u odnosu na
Tabela 2. Prečnik korena (mm)Table 2. Root diameter (mm)
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 3 5 3 5 2 2B 3 5 3 5 2 2C 3 5 3 5 2 2
AB 5 6 5 7 2 3AC 5 6 5 7 2 3BC 5 6 5 7 2 3
ABC 7 9 7 9 3 4
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 31,6 41,2 13,2 12,2 40,4 51,0 12,2 13,2 24,4 20,4 41,0 12,7 24,6 29,22007 38,9 44,0 15,3 17,0 38,9 48,7 14,6 20,3 26,9 32,5 42,6 17,4 28,8 30,62008 31,6 36,0 7,6 8,4 32,8 38,5 9,0 10,6 20,2 23,4 34,7 8,9 20,9 22,7
Tabela 3. Masa korena (g/biljci)Table 3. Root weight (g per plant)
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 116 169 12 10 194 218 11 13 83 102 174 12 77 1092007 153 167 17 20 172 199 16 30 90 104 173 21 89 1042008 76 84 11 13 81 95 12 22 45 54 84 15 46 53
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 30 40 40 50 20 20B 30 40 40 50 20 20C 30 40 40 50 20 20
AB 40 60 50 70 30 30AC 40 60 50 70 30 30BC 40 60 50 70 30 30
ABC 60 80 70 100 40 50

Rajić M, Pejić B146
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
kontrolu, što je potvrđeno ispitivanjima Garter & Traveler (1981) da uneta đubriva azota pre setve uvek ostvaruju brži porast biljaka. Prvi rok setve u odnosu na drugi rok setve u sve tri godine ispitivanja ostvario je značajnu razliku sa masom korena (Tab. 3). Slični rezultati ostvareni su u Bugarskoj sa rokovima setve. Prvi rok setve (30.07) imao je masu korena od 60 g/biljci, dok je kasnom setvom (10.09) ostvarena masa korena od 10 g/biljci (Slavov 1983). Navodnjavanje posle setve je samo u prvoj godini ostvarilo značajne razlike u odnosu na navodnjavanje pre osnovne obrade. Đubrenje azotom i prvi rok setve imali su
značajan uticaj na povećanje broja listova u odnosu na kontrolu bez đubrenja i drugi rok setve tokom sve tri godine ispitivanja. Navodnjavanje posle setve je ostvarilo značajan uticaj u drugoj i trećoj godini ispitivanja (Tab. 4). Slični rezultati ostvareni su u Bugarskoj: setvom 30.07. formirano je osam listova, dok je setvom 10.09. formirano 22 lista (Slavov 1983).
Masa nadzemnog dela može biti znatno veća u odnosu na masu korena tokom letnjeg i jesenjeg perioda. Prema rezultatima Rajića (1992) formira se osam puta veća masa nadzemnog dela, a prema Stefanoviću (1987) deset puta veća masa nadzemnog dela. U ovom ogledu obrazovana masa nadzemnog dela bila je tri puta veća od mase korena. Đubrenje azotom je značajno uticalo na
masu nadzemnog dela u sve tri godine ispitivanja. Značajnog uticaja na masu nadzemnog dela imao je prvi rok setve u odnosu na drugi rok setve. U prvoj godini ispitivanja zalivanje posle setve imalo je značajnu razliku u odnosu na drugi rok setve. U drugoj godini ispitivanja razlika je bila obrnuta, dok u trećoj godini ispitivanja između načina navodnjavanja nije bilo opravdanih razlika (Tab. 5). Đubrenje azotom nije uticalo na sadržaj suve
materije u korenu, u sve tri godine ispitivanja (Tab. 6). Međutim, autori Smolen & Sady (2009) pokazuju da đubrenje azotom ima značajan uticaj na povećanje suve materije u korenu šećerne repe u odnosu na kontrolu bez đubrenja. Treba uzeti u obzir da je ovaj ogled bio u uslovima navodnjavanja i da su sadnice semenske šećerne repe bile sa malom masom korena. Prednost prvog roka setve od 30 dana je duža vegetacija u odnosu na drugi rok setve, zbog čega je sadržaj suve materije bio značajno veći u sve tri godine ispitivanja. Slične rezultate ostvario je Korženko (1984) sa više rokova setve u letnjem periodu.
Na sadržaj suve materije u nadzemnom delu nije uticalo đubrenje azotom u prve dve godine ispitivanja (Tab. 7). U trećoj godini ispitivanja procenat suve materuje bio je veći na varijanti đubrenja sa azotom u odnosu na kontrolu. U drugim istraživanjima đubrenje azotom u toku
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 18 18 8 10 16 18 10 12 13 15 18 10 14 142007 22 24 14 14 26 30 14 14 19 21 26 14 19 212008 22 22 14 14 20 24 14 18 18 20 22 15 18 19
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 1 2 2 2 1 2B 1 2 2 2 1 2C 1 2 2 2 1 2
AB 2 3 3 4 2 3AC 2 3 3 4 2 3BC 2 3 3 4 2 3
ABC 3 4 4 5 3 4
Tabela 4. Broj listova po biljciTable 4. Leaves per plant
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing

Uticaj rokova setve na osobine sadnica šećerne repe 147
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
leta uticalo je na veći procenat suve materije u lišću i veću količinu hlorofi la u odnosu na kontrolu (Malnou et al. 2008). Prvi rok setve ostvario je značajno veći procenat suve materije od drugog roka setve u prve dve godine ispitivanja.
Rokovima setve mogu se ostvariti razlike od 4% suve materije u listu prema istraživanju (Dolja & Golubovskij 1971). Između načina navodnjavanja nije bilo opravdanih razlika u sadržaju suve materije nadzemnog dela.
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 286 397 56 59 514 700 62 77 230 302 474 64 271 3382007 420 520 115 129 567 675 122 184 306 377 546 138 546 3872008 280 340 23 36 239 377 41 49 146 201 309 37 170 177
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 40 60 50 60 30 40B 40 60 50 60 30 40C 40 60 50 60 30 40
AB 60 80 70 90 40 50AC 60 80 70 90 40 50BC 60 80 70 90 40 50
ABC 90 120 90 130 60 80
GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 10,4 11,5 3,4 7,77 11,2 13,2 4,8 5,7 7,5 9,5 11,5 5.4 8,2 8,72007 14,12 13,2 11,6 9,7 11,3 13,6 11,3 11,7 12,1 12,1 13,1 11,1 12,2 11,92008 12,2 14,3 6,9 8,2 15,4 12,0 9,7 7,9 11,1 10.6 13,5 8,2 10,4 11,3
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 2 3 2 2 2 3B 2 3 2 2 2 3C 2 3 2 2 2 3
AB 3 4 2 3 3 4AC 3 4 2 3 3 4BC 3 4 2 3 3 4
ABC 4 5 3 4 4 6
Tabela 5. Masa nadzemnog dela (g/biljci)Table 5. Aboveground weight (g per plant)
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing
Tabela 6. Procenat suve materije u korenuTable 6. Root dry matter (%)
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing

Rajić M, Pejić B148
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
Zaključak
U cilju proizvodnje sadnica semenske šećerne repe sa dobrim morfološkim, fenotipskim i kvalitetnim osobinama potrebno je u našim agroekološkim uslovima primenuti sledeću agrotehniku, koja je u ovim istraživanjima pokazala prednost:
Neophodno je izvršiti predsetveno đubrenje 1. sa azotom.
Navodnjavanje posle setve treba da ima 2. prednost u odnosu na navodnjavanje pre oranja.
Raniji rok setve (15.07) ima prednost u 3. odnosu na kasniji rok setve(15.08).
Ova preporuka je naročito važna usled kasne setve, koja se sada primenjuje u praksi. Kasnom setvom preko sadnica ne može se proizvesti kvalitetno seme šećerne repe.
Tabela 7. Procenat suve materije nadzemnog delaTable 7. Aboveground dry matter (%)GodinaYear
C1 C2 Prosek / AverageB1 B2 B1 B2
A1 A2 A1 A2 A1 A2 A1 A2 A1 A2 B1 B2 C1 C2
2006 10,8 13 8,5 6,1 13,4 15,6 5,6 8,6 9,6 10,8 13,2 7,2 9,6 10,82007 13,4 12,6 11,2 10,6 11,5 12,1 9,6 10,5 11,4 11,5 12,5 10,5 12,0 10,92008 12,8 15,1 15,5 13,8 14,3 13,4 15,2 15,2 11,5 14,4 13,9 14,9 14,3 14,5
Godina / Year2006 2007 2008
LSD 0,05 0,01 0,05 0,01 0,05 0,01A 2 2 2 2 2 3B 2 2 2 2 2 3C 2 2 2 2 2 3
AB 2 3 2 3 3 5AC 2 3 2 3 3 5BC 2 3 2 3 3 5
ABC 3 4 3 5 5 7
A1 kontrola bez đubrenja / control without fertilizers A đubrenje / fertilizationA2 đubrenje / fertilization B1 prvi rok setve / fi rst planting date B setva / planting dateB2 drugi rok setve / second planting date C1 navodnjavanje pre oranja / irrigation before plowing C navodnjavanje / irrigationC2 navodnjavanje posle setve / irrigation after plowing

Uticaj rokova setve na osobine sadnica šećerne repe 149
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
Literatura
Akeson W R, Henson M A, Freutag A H, Westfall D G (1980): Sugar beet Fruit Germination and Emergence under Moi-sture and Temperature Stress, Crop Sci. 20: 735-739
Dolja V S, Golubovskij R A (1971): Rolj doz form i sposobov unesenija u udobrenija v povišenim vrožaja i vlučšeniji ka-čestva semjan saharnoj svekli, Semenenovodstvo Saharnoj svekli, Moskva, 125-131
Dragović S (1987): Navodnjavanje ratarskih kultura potrebe u vodi i efekat navodnjavanja, Poljoprivredne Aktuelnosti 28: 221-223
Garter J N, Traveler D J (1981) Effect of Time and Amount of Nitrogen Uptake an Sugar beet Growth and Yield, Agron. J. 73: 665-670
Geerts S, Raes D (2009): Defi cit irrigation as an on-farm stra-tegy to maximize crop water productivity in dry areas, Agric. Water Manag. 96: 1275-1284
Gorelov E P, Batirov H, Sajdavutov M S (1978): Viraščivanije semjan saharnoj svekli bezvisadočnim spasob, Selekcija i Se-menarstvo Moskva 3: 48-49
Korženko N P (1984): Viraščivanie semjan eliti saharnoj svekli bezvisadočnim sposobom, Selekcija i Semenovodstvo, Mo-skva 5: 44-45
Malnou C S, Jaggard K W, Sparkes D L (2008): Nitrogen fertili-zer and the effi ciency of the sugar beet crop in late summer 28: 47-56
Marinković B, Crnobarac J (2000): Zavisnost kvaliteta i prinosa šećerne repe od primene NPK hraniva, Acta Periodica Te-chnologica 31: 345-350
Pečenov V A (1988): Vlijanija mineralnogo pitanija na rast i vodobmen semenoj saharnoj svekli, Fiziološki-biohemiski osnovi povišenija produktivnosti rasta, Zaščita ih am boleznoj i neblagoprijatnah faktorov, Frunze, 3-14
Rajić M (1992): Uticaj rokova setve načina đubrenja i navod-njavanja na prinos i kvalitet semena šećerne repe, Doktorska disertacija, Univerzitet Novi Sad, Poljoprivredni fakultet, 69-73
Smolen S, Sady W (2009): The effect of various nitrogen fer-tilization and foliar nutrition regimes on the concentrati-ons of nitrates, ammonium ions dry matter and N-total in carrot (Daucus carota) roots, Scientia Horticulturae 119: 219-231
Slavov K (1983): Prauchvane vahrkhy njakoi ot uslovijata za prezimuvane na zakharnoto sveklo za semeproizvodstvo po bezrazsadnija, Rast Nauki, Sofi ja, 71-77
Stefanović D (1987): Prinos i poljoprivredna vrednost semena šećerne repe u zavisnosti od sklopa useva roka žetve i desi-kacije, Doktorska disertacija, Univerzitet Novi Sad, Poljopri-vredni fakultet, 37-39
Vamerali T, Guarise M, Ganis A, Mosca G (2009): Effects of water and nitrogen management on fi brous root dis-tribution and turnover in sugar beet, Europ. J. Agron. 31: 69-76

Rajić M, Pejić B150
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 143-150
Effects of Planting Dates, Fertilization with N and Irrigation Mode on Morphological and Phenotypic Traits of Sugar Beet Seedlings
Milorad Rajić1 · Borivoj Pejić2
1Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia2Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
Summary: In this study we tested the effects of two planting dates, fertilization with N and lack of fertilization, as well as two different irrigation modes on morphological and phenotypic traits of sugar beet seedlings. Well-devel-oped sugar beet seedlings are thought to be cultivated plants whose requests comply with agroecological conditions of the region. We tested number of plants per ha, root diameter, root fresh weight, number of leaves, aboveground fresh weight, aboveground dry matter weight, and root dry matter weight. Analysis of variance of a trifactorial trial in three research years showed signifi cant impact of the fi rst planting date on all observed morphological and phenotypic traits of sugar beet seedlings. N fertilization applied before planting provided justifi ed higher difference when compared to control, regarding root fresh weight, aboveground fresh weight and larger root diameter. Differ-ent irrigation modes provided signifi cantly higher number of plants and root fresh weight.Key words: irrigation, N fertilization, planting dates, sugar beet seedlings

Yield and Carrying Capacity of Meadows in Albania 151
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 151-154
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 151-154original research article / originalni naučni rad
Yield and Carrying Capacity of Meadows in Albania
Luan Hajno · Fiqiri Tahiri
received / primljeno: 20.08.2010. revised / prerađeno: 06.12.2010. accepted / prihvaćeno: 27.12.2010.© 2011 IFVC
Summary: This study was carried out during 2003-2005 and included total meadow surface of 32,000 ha from seven regions throughout Albania (summer meadows of Shishtavec, Gramoz, Çajup, Lenie, Progo-nat, and winter meadows of Heremec-Delvine and Dukaj). The results of the study showed that average dry matter yield of the summer meadows is low (about 0.6±0.14 t ha-1) ranging from 0.33±0.07 t ha-1 to 0.53±0.10 t ha-1 at poor meadows (Progonat, Cajup, Lenie) and from 0.9±0.17 t ha-1 to 0.95±0.18 t ha-1 at rich meadows (Gramoz, Shistavec). Dry matter yield of the winter meadows ranged from 0.45±0.17 t ha-1 (Dukaj) to 0.57±0.13 t ha-1 (Heremec-Delvine). Regarding the rate of herbaceous biomass utilization in general, the plots closer to the villages were overstocked, which put high pressure on meadows resulting in their degradation. The coeffi cient of herbaceous phytomass utilization was about 94-95% at Lenia’s mea-dows and a few other meadows close to villages. At a few plots of meadows (Lenia, Gramoz, Shishtavec) si-tuated away from villages and at high sea level, the coeffi cient of meadow utilization reached 50-60%. Ener-getic value expressed by starch unit (SU) depends on the fl ora and environmental conditions. According to laboratory analyses, the meadows of Dukaj had the highest energetic values (about 1.16±0.22 SU kg-1 dry matter) while Lenia’ meadows had the lowest energetic values (about 0.84 ±0.16 SU kg-1 dry matter).Keywords: capacity of meadow, sample, small ruminants, plot, region, dry matter yield
L. Hajno · F. Tahiri Center for Agricultural Technology Transfer, Fushe-Kruje, Albaniae-mail: [email protected]
Introduction
Albania has a total surface area of 2,844,800 ha. According to the offi cial agricultural statistical data (2005), a large part (36%) of this area is classifi ed as forests, which includes not only high and coppice forests, but also shrub lands. The second major land use are arable lands (24%) where herbaceous crops and trees are grown. The third major land use are pastures, which are largely uncultivated areas covered mainly by herbaceous vegetation and small shrubs.
Pastures and meadows are distributed throughout the country and divided in two groups depending on the season of grazing: winter and summer ones. The latter include all the pastures found in the sub-alpine and alpine zones and amount to about 61% of the whole area (Shundi & Buzi 1991).
Dominant herbaceous species in the winter pastures are several annual grasses and legumes as well as perennials such as Dactylis glomerata, Festuca ovina, Poa bulbosa and Lolium perenne; while in the summer pastures only perennial species can be found, such as Poa pratensis, Phleum montanum, Nardus stricta, Trifolium
alpestre, T. repens, etc. The productivity of pastures depends on several factors including the ecological zone they are found in, the physical characteristics and the grazing history. In these meadows, the phytomass production in a favourable year reaches 4 t ha-1 and in an unfavourable year (without suffi cient rain) it decreases to 2 t ha-1. Although the provisions of the Pastures and Meadows’ Law are technically and scientifi cally correct and constitute a sound basis for rational management of the pasture resources, very few of these provisions have actually been implemented.
Practically, there is no control over grazing management and very few state pastures and meadows have been rented to individuals or companies. As a result, the condition of pastures and meadows is poor to a large extent due to (a) large amount of unpalatable plant species or weeds, both herbaceous (e.g. Asphodelus microcarpus, Urginea maritima, etc) and woody (e.g. Phlomis fruticosa, Rubus sp., etc); (b) a large portion of bare soil on pastures and (c) a conspicuous accelerated erosion. The latter phenomenon is widespread in the pastures of the whole country, often accompanied by landslides, especially in the areas of fl ysch or marls.
Soil and Crop Management / Gajenje i agrotehnikaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )Acknowledgements: Authors would like to thank NSRBA which su-pported this study.

Hajno L, Tahiri F152
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 151-154
Gilles et al. (1981) reported that one of the causes of desertifi cation is overgrazing by domestic animals. Many range managers have also argued that the lack of privately owned pastures is a major cause of overgrazing.
Carrying capacity in its most basic defi nition determines the maximum livestock that an ecosystem can support on a sustainable basis. Livestock carrying capacity is based on the idea of “ecological carrying capacity” under which populations grow until restricted feed supplies decrease birth rates and increase death rates. This point is defi ned as “K” the asymptote of the population growth curve based on the available resources. The dominant range model based on the carrying capacity concept developed assumes that pastoralists occupy potentially equilibrial grazing systems. Destabilization of these systems is caused by overstocking. Carrying capacity is calculated from an estimate of annual production of consumable vegetation, linked to animal requirements for feed and nutrients. Annual primary production can be estimated (i) by the measurement of standing biomass (normally at the end of the growing season) or (ii) by remote sensing or (iii) predicted by using empirical relationships of primary productivity to rainfall or (iv) from more complex relationships of primary productivity to rainfall or (v) from more complex relationship between soils, soil water, fertility and rainfall. These estimates are normally corrected for the available or useful biomass, which is the proportion of the vegetation actually useable by livestock (grazing effi ciency). Different types of livestock have different dietary preferences, grazing habits and effi ciency. “Proper use” will vary according to growing conditions, species composition and the type of herbivore (Bartels et al. 1993, Sinsin 1993, Kussela et al. 2002, Macdonald et al. 2008).
Steenenkamp et al. (1995) reported that the integrated system for plant dynamics (ISPD) can serve as a basis for range condition and grazing capacity assessments.
The defi nition of carrying capacity as the maximum stocking rate possible without including damage to vegetation or related resources is clear and unambiguous. It is the range manager’s task to balance the regeneration of the plants and the grazing pressure (Hobbs et al. 1985, Roe 1997).
Meadows constitute the main feed base for small ruminants, and in fact the meadows meet 75% of small ruminants’ requirements in Albania. The evaluation of meadow yield is one of the key elements for the assignment of grazing capacity and stocking
rate in order to provide sustainable management. Such assessments could help to establish a proper balance among meadow yield, livestock and land.
The objectives of this study were to assess average dry matter yield (t ha-1), coeffi cient of phytomass utilization, dry matter content and energetic value (SU) of herbaceous mass, growth rate of standing biomass and optimum period of grazing (from beginning to the end of grazing).
Materials and Methods
Winter and summer meadows amount to 32,000 ha and these were divided into 334 plots (the size 100 ha per plot) in order to evaluate the yield. Winter meadows (Heremec-Delvine, Dukaj) are located on the lowlands and are grazed during winter (also including spring and autumn). Summer pastures (Shishtavec, Gramoz, Çajup, Lenie, Progonat) are located on the mountains and are grazed during summer for 4-6 months (May-October). Total of 677 samples randomly chosen from 334 plots served as tested surfaces (the size of tested surface was 4 m2), which means there were 2.1 tested surfaces for each plot. These tested surfaces were harvested for three consecutive years in order to evaluate their dry matter yield (t ha-1) and energetic values (expressed in starch unit).
Number of samples of residues after grazing was 677, which were used to calculate the coeffi cient of the herbaceous mass utilization. The coeffi cient of phytomass utilization by livestock was calculated in this manner: yield minus residues after livestock grazing. For this purpose, harvesting of residues after grazing was carried out close to each of the tested surfaces. Dry matter yield was estimated per useful surface, which is actually meadow surface covered with herbaceous mass and available for grazing by livestock.
Total of 45 plots for every 100 m of altitude were used to defi ne cycles of grazing and to estimate herbages growth rate for three years (growth rate = fi nal phytomass (kg) - initial phytomass (kg) / time interval (days)). Every year, 800 samples were analyzed for each plot in order to estimate dry matter intake and energetic value SU kg-1 of dry matter. Dry mater content and energetic value of meadow phytomass were estimated by the Laboratory of Livestock Research Institute (IKZ), Tirana-Albania. Data were processed by employing ANOVA.
Results and Discussions
Results of the study showed that average yield of summer meadows was 0.6t±0.14 t ha-1 which is

Yield and Carrying Capacity of Meadows in Albania 153
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 151-154
generally considered low (Tab. 1). Dry matter yields ranged from 0.33 to 0.53 t ha-1 for poor meadows (Proganat, Çajup, Lenie) and 0.90 to 0.95 t ha-1 on average for rich meadows (Gramoz, Shishtavec).
The rate of herbaceous biomass utilization by livestock for all the regions is shown in table 2, together with the assessment of yield and energetic value of meadows biomass. The data showed that plots close to villages were overstocked and had the highest consumption of fresh grass by livestock per unit surface. This high pressure on meadows has caused their degradation.
Plots no. 16 and 17 of Lenia region and plots no. 46 and 47 of Gramoz region that were close
to villages had a coeffi cient of herbaceous biomass utilization of 94-95% while plots no. 37 and 38 in Lenia, no. 10 and 11 in Gramoz, no. 6, 7, 9, 10 in Shishtavec that were situated at higher altitudes and further away from villages had a rate of phytomass utilization about 50-60%. Similar results were reported by Bartles et al. (1993) and Roe (1997).
There was no sound management of meadows in relation to the techniques of grazing. The grazing of some meadows was delayed, meaning that livestock came to graze at the time when grass has begun to fl ower and some meadow plants have reached the phase of low eating or they were inedible. In springtime, absence of proper ratio between intensive growth of meadow plants and ample grazing livestock was observed. The same phenomenon, caused by incomplete utilization of herbaceous biomass, was reported by Sinsin (1993) and within our study this occurred in some plots of Hemerec-Delvine, Dukaj and Gramoz meadows.
As for energetic value content of herbages, the meadows of Dukaj had the highest energetic values (1.16±0.22 SU kg-1 dry matter) and Lenia’ meadows had the lowest energetic values (0.84 ±0.16 SU kg-1 dry matter).
In some cases, low yield of meadows is as a result of earlier grazing by livestock before plants reach
Table 1. Meadows dry matter yield (t ha-1) for the whole surfaceTabela 1. Prinos suve materije livada (t ha-1) za ukupnu površinu
RegionsRegioni
Whole surface / Ukupna površina
ha
total dry matter yield
ukupni prinos suve materije (t)
t ha-1 CV(%)
Shishtavec 2,902 2756.90 0.95±0.18 18.9Gramoz 4,822 4339.80 0.90±0.14 15.5Çajup 3,608 1804.00 0.50±0.13 26.0Lenie 5,038 2670.14 0.53±0.10 18.8Progonat 5,846 1929.18 0.33±0.07 21.2Heremec-Delvine 5,470 3227.30 0.59±0.11 16.9Dukaj 3,200 1536.00 0.48±0.88 18.3
RegionsRegioni
Useful surface / Korisne površine SU* kg-1 dry matter
suve materijeha t ha-1 rate of phytomass
utilization stopa iskorišćenja fi tomase (%)
Shishtavec 2,584 1.06 82 0.94Gramoz 3,723 1.16 77 0.93Çajup 2,013 0.89 83 1.04Lenie 3,124 0.85 80 0,84Progonat 2,474 0.78 83 0,98Heremec-Delvine 2,020 1.59 83 0,96Dukaj 2,144 0.71 67 1.15
Table 2. Useful surface dry matter yield (t ha-1), phytomass utilization and energetic valueTabela 2. Prinos suve materije korisnih površina (t ha-1), iskorišćenje fi tomase i energetska vrednost
*SU = 1420 calories / kalorija

Hajno L, Tahiri F154
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 151-154
optimum vegetation and the technical maturity for grazing (8-10 cm for sheep and 12-15 cm for cattle). As a consequence, useful plants of meadows are highly thinned out. The results of the study show that the fi rst grazing should begin between May 25 and June 10 for summer pastures, i.e. between March 20 and April 20 for winter pastures.
Conclusions
Albanian meadows are characterized by low yield per unit surface, different cycles of plant growth and seasonal vegetation of meadow plants.
Some meadows are not completely utilized, which is related to the techniques of grazing.
Complex measures such as establishment of the cultivated pasture, cleaning from solid materials, stones, scrubs and fertilization, sowing seeds in uncropped portions of meadows, irrigation, etc. must be implemented in order to increase the yield of meadows and their carrying capacities, as well as the lasting of grazing cycles.
Effi cient and sustainable use of meadows could be achieved by a rational grazing by livestock. Two schemes of meadow utilization should be applied during the spring time while yield of meadows is abundant - harvesting one part of meadows and grazing the other part. If harvest is not possible, then meadows should only be grazed.
It is necessary to construct shelters and puddles on meadows to provide timely water supply to grazing livestock.
References
Bartels G B, Norton B E, Perrier G K (1993): An examination of the carrying capacity concept. In: R.H. Behnke, I. Scoones and C. Kerven (eds.): Range ecology at disequilibrium, overseas.
De Leeuw P N, Macdonald I A W (1994): Historical changes in stocking rates of domestic livestock as a measure of semi-arid and arid rangeland degradation in the Cape province, South Africa. J. Arid Environ. 26: 281-298
Gilles J, Jamtgaard K (1981): Overgrazing in pastoral areas: the commons reconsidered. Sociologia Ruralis 21: 129-141
Herve D, Calagua D, Poupon H, Fernandes R (1989): Utilisati-on agropastorale du sol par les communautes du Haut Cane-te. Les Cahiers de la Recherche Development 24: 1-14
Hobbs N T, Swift D M (1985): Estimates of habitat carrying capacity incorporating explicit nutritional constraints. J. Wil-dl. Manage. 49: 814-822
Kuusela E, Khalili H (2002): Effect of grazing method and her-bage allowance on the grazing effi ciency of milk production in organic farming. Animal Feed Sci Technol. 98: 87-101
Lebaron A, Bond L K, Aitken P S, Michaelsen L (1979): An explanation of the Bolivian highlands grazing-erosion syn-drome. J. of Range Management 32: 201-208
Macdonald K A, Penno J W, Lancaster J A S, Roche J R (2008): Effect of stocking rate on pasture production, milk pro-duction and reproduction of dairy cows in pasture-based systems. J. of Dairy Sci. 91: 2151-2163
Potvin F, Huot J (1983): Estimating carrying capacity of a white-tai-led deer wintering area in Quebec. J. Wildl. Manage. 47: 463-475
Roe E M (1997): Viewpoint: On rangeland carrying capacity. Journal of range management 50: 467-472
Sinsin B (1993): Phytosociologie, ecologie, valeur pastorale, production et capacite de charge des paturages naturels du perimeter Nikki-Kalale au Nord-Beni; These presentee en vue de l’obtention du grade de docteur en Sciences Agrono-miques; Universite libre de Bruxelles, Bruxelles
Steenekamp S J, Bosch O J H (1995): Construction and evalua-tion of condition assessment and grazing capacity models for use in grazing management in the Eastern mixed bushveld of Southern Africa. Journal of Arid Environments 30: 351-360
Shundi A, Buzi T (1991): Experimental results in improvement of mountainous pastures in Albania. Herbe 4: 50-52
Prinos i proizvodnost livada u Albaniji
Luan Hajno · Fikiri Tahiri
Centar za transfer poljoprivredne tehnologije u Fuše-Kruji, Albanija
Izvod: Ovo istraživanje je sprovedeno u periodu 2003-2005 na ukupnoj površini livada od 32.000 ha u sedam regiona Albanije (letnje livade Šištavec, Gramoz, Čajup, Lenije i Progonat i zimske livade Heremec-Delvine i Dukaj). Rezultati istraživanja su pokazali da je prosečan prinos suve materije letnjih livada nizak (oko 0,6 ± 0,14 t ha-1) u rasponu od 0,33 ± 0,07 t ha-1 do 0.53 ± 0.10 t ha-1 za siromašne livade (Progonat, Čajup, Lenije) i od 0,9 ± 0,17 t ha-1 do 0,95 ± 0,18 t ha-1 za bogate livade (Gramoz, Šištavec). Prinos suve materije zimskih livada kretao se u rasponu od 0,45 ± 0,17 t ha-1 (Dukaj) do 0,57 ± 0,13 t ha-1 (Heremec-Delvine). Što se uopšteno tiče stope iskorišćenja travnate biomase, parcele bliže selima bile su preopterećene prevelikim brojem grla stoke, što je rezultiralo njihovom degradacijom. Koefi cijent iskorišćenja travnate biomase je bio oko 94-95% na livadama regiona Lenija i nekoliko drugih livada u blizini sela. Na nekoliko livadskih parcela (Lenia, Gramoz, Šištavec) koje su udaljene od sela i na velikoj nadmorskoj visini, koefi cijent iskorišćenja livada je dostigao 50-60%. Energetska vrednost izražena u jedinicama skroba (SU) zavisi od fl ore i uslova životne sredine. Prema laborato-rijskim analizama, livade regiona Dukaj imale su najveće energetske vrednosti (oko 1,16 ± 0,22 SU kg-1 suve materije) dok su livade regiona Lenija imale najmanje energetske vrednosti (oko 0,84 ± 0,16 SU kg-1 suve materije).Ključne reči: parcela, prinos suve materije, proizvodnost livada, region, sitni preživari, uzorak

Prinos semena i suve materije crvene deteline 155
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160originalni naučni rad / original research article
Uticaj međurednog rastojanja i setvene norme na prinos semena i suve materije
crvene deteline (Trifolium pratense L.)
Rade Barać · Gojko Duronić · Đura Karagić · Sanja Vasiljević · Branko Milošević
primljeno / received: 29.10.2010. prihvaćeno / accepted: 10.11.2010.© 2011 IFVC
Izvod: U radu je proučavan uticaj međurednog rastojanja i setvene norme na prinos semena i suve materije crvene deteline u sistemu proizvodnje za kombinovano iskorišćavanje. Dvogodišnja poljska istraživanja (2006. i 2007) obavljena su u Poljoprivrednoj školi u Futogu. Usev crvene deteline je zasno-van na tri međuredna rastojanja: 12,5 cm , 25 cm i 50 cm, sa tri setvene norme: 2,5 kg ha-1, 5 kg ha-1 i 10 kg ha-1. Najveći prinos semena u sistemu proizvodnje crvene deteline za kombinovano iskorišćavanje (seme-krma) tokom obe posmatrane godine je dobijen setvom na 25 cm međurednog razmaka i sa ko-ličinom semena 10 kg ha-1, dok je najveći prinos suve materije ostvaren sa istom setvenom normom na 12,5 cm međurednog razmaka.Ključne reči: crvena detelina (Trifolium pratense L), međuredno rastojanje, setvena norma, prinos seme-na, prinos suve materije
Đ. Karagić · S. Vasiljević · B. MiloševićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]. BaraćPoljoprivredna škola sa domom učenika, Carice Milice 2, 21410 Futog, SrbijaG. DuronićD.O.O. COPEX, Novi Sad, Srbija
Uvod
Gajenje i agrotehnika / Soil and Crop Management w
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Crvena detelina je posle lucerke druga po značaju višegodišnja krmna leguminoza u Srbiji. Nasuprot velikom značaju crvene deteline, kako s privrednog tako i sa agroekološkog aspekta, evidentno je smanjenje površina pod ovom kr-mnom leguminozom kako u svetu tako i u našoj zemlji. Tek u skorije vreme saglasno savreme-nim trendovima održive i organske poljoprivre-de, prevashodno u Kanadi, SAD i pojedinim zemljama Evrope (Švedska, Velika Britanija, Holandija, Danska, Nemačka, Austrija) crvena detelina opet dobija na značaju, naročito kao komponenta travno-detelinskih smeša, kao i u proizvodnji zdravstveno-bezbedne stočne hra-ne i lekovite herbe (Kovačević & Kundaković 2005).
Semenska proizvodnja crvene deteline u Re-publici Srbiji, koja bi pravilnom rejonizacijom
trebala da se organizuje u severozapadnom delu Republike Srbije, već duži niz godina ne zadovo-ljava domaće potrebe za setvu proizvodnih po-vršina (Karagić et al. 2010). Najveći deo semena ove biljne vrste se još uvek uvozi, iako za to nema nikakvog opravdanja, imajući u vidu slabiju pri-lagođenost inostranih sorti našim agroekološkim uslovima, kao i manju produkciju krme u odnosu na domaći sortiment (Vasiljević i sar. 2010).
Lugić i sar. (1996) navode da je u Republici Srbiji uobičajena i veoma perspektivna kombino-vana proizvodnja krme i semena crvene dateline. Pri kombinovanom korišćenju detelišta na terito-riji Republike Srbije u godini zasnivanja najčešće se prvi otkos ostavlja za seme. Crvena detelina iz prvog otkosa sazreva tokom jula i avgusta. Dru-ge godine drugi otkos se obično ostavlja za seme (Duronić 2010). U tom slučaju, košenje prvog otkosa se vrši nešto ranije, tj. u fazi butonizacije ili samog početka cvetanja. Lugić i sar. (1999) isti-ču da je proizvodnja semena crvene deteline pod izuzetno velikim uticajem klimatskih faktora, pa je stoga neophodno da se sve mere, pa i mome-nat kosidbe, prilagode mikroklimatu područja u kome se ona gaji za seme.
Cilj naših istraživanja je bio da se ispita uticaj međurednog rastojanja i setvene norme na prinos semena i suve materije crvene deteline u sistemu proizvodnje za kombinovano iskorišćavanje.

Barać R i sar.156
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
Materijal i metod
Poljski ogled je postavljen 2006. u Poljopri-vrednoj školi u Futogu na zemljištu tipa degradi-rani černozem lesne terase. Za setvu se koristila domaća sorta crvene deteline Una selekcionisana u Institutu za ratarstvo i povrtarstvo u Novom Sadu. Postavljen je dvofaktorijalni ogled u četiri ponavljanja, sa elementarnom parcelom veličine 10 m2. Predusev za crvenu detelinu je bio ozimi ječam. Osnovna obrada zemljišta obavljena je u jesen na dubinu od 30 cm. Setva je obavljena ruč-no 2. aprila 2006.
Prinos semena je utvrđen žetvom ukupne količi-ne semena sa svake parcele i preračunat je na prinos semena po hektaru i to u prvoj godini istraživanja iz prvog otkosa, a u drugoj godini iz drugog otkosa. Košenje krme u godini zasnivanja crvene deteline iz drugog otkosa, a u 2007. godini iz prvog otkosa,
vršeno je ručno u početku cvetanja kada je 10% do 15% biljaka cvetalo, nakon čega je na uzorku od 2 x 100 g utvrđen sadržaj suve materije.
Ispitivani faktori su bili: 1. međuredno rasto-janje (A): (i) 12,5 cm između redova, (ii) 25 cm između redova i (iii) 50 cm između redova, te 2. količina semena po hektaru (B): (v) 10 kg ha-1, (vi) 5 kg ha-1 i (vii) 2,5 kg ha-1.
Vremenski uslovi
Oblast Bačke ima odlike umereno kontinental-ne klime sa hladnim zimama i toplim letima izme-đu kojih su prelazna godišnja doba, svežije jeseni i vlažnija proleća.
Crvena detelina za proizvodnju semena zahteva veće količine padavina, od oko 750 mm godišnje (Miladinović 2001), pri čemu je raspored padavina tokom letnjeg perioda od naročitog značaja za posti-
Slika 1. Klimadijagram po Walter-u za Futog (2006)Fig. 1. Climatograph by Walter for Futog (2006)
Slika 2. Klimadijagram po Walter-u za Futog (2007)Fig. 2. Climatograph by Walter for Futog (2007)
-5
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8 9 10 11 12
meseci
tem
pera
ture
0
20
40
60
80
100
120
140
160
pada
vine temp.°C
pad.mm
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10 11 12
meseci
tem
pera
tura
0
20
40
60
80
100
120
pada
vine temp.°C
pad.mm

Prinos semena i suve materije crvene deteline 157
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
zanje visokih prinosa semena crvene deteline. Sušu i visoku temperaturu slabije podnosi, što značajno ograničava semensku proizvodnju ove kulture na rejone umerenog klimata. Povoljni rejoni za semen-sku proizvodnju crvene deteline treba da obezbede dovoljno vlage pre početka cvetanja, a suvo i toplo vreme tokom cvetanja i dozrevanja semena.
Višegodišnji prosek padavina za period 1999-2008. za lokalitet Futog iznosi 632 mm, odnosno u vegetacionom periodu 384 mm, što ukazuje na defi cit padavina. Bolji uvid u vremenske prilike i njihov uticaj na useve se dobija stavljanjem u odnos padavina i temperatura vazduha po mese-cima grafi čkim prikazivanjem, klima dijagramom po Walter-u (Sl. 1. i 2.)
Toplo vreme u aprilu u godini zasnivanja crve-ne deteline sa dosta padavina uticalo je povoljno na nicanje i početni porast biljaka, a time i na pri-nos semena u 2006.
U poređenju sa višegodišnjim prosekom uoča-va se da je 2006. bila prosečna dok je 2007. bila godina toplija od proseka. U toku 2006. registro-vano je 656 mm padavina, odnosno 24 mm više od višegodišnjeg proseka. U vegetacionom peri-odu je palo 445 mm što je više u proseku za 61 mm. Suma mesečnih junskih i julskih padavina je bila veća od proseka. Avgust je imao više pada-vina od višegodišnjeg proseka, dok je septembar imao znatno manje padavina od proseka.
Za razliku od prethodne godine, 2007. je bila nešto povoljnija po pitanju padavina. Ukupna ko-ličina vodenog taloga iznosila je 763 mm, što je za 129 mm više od višegodišnjeg proseka. Među-
tim, letnji meseci u 2007. su bili izuzetno topli, uz česte pojave vazdušne i zemljišnje suše, praćene suvim vetrovima i vrlo visokim temperaturama koje su na momente dosezale 42°C.
Rezultati i diskusija
Prinos semena
Najveći prinos semena, u prvoj godini od 482,4 kg ha-1 dobijen je setvom 10 kg ha-1 semena na me-đurednom rastojanju od 25 cm, dok je najmanji prinos u prvoj godini iznosio 294,7 kg ha-1, a dobi-jen je setvom 2,5 kg ha-1 semena na 12,5 cm (Tab. 1). Setvom na međuredno rastojanje od 25 cm ostvaren je prosečan prinos semena za 12% veći od prosečnog prinosa dobijenog setvom na 12,5 cm, a za 6% veći od prosečnog prinosa dobijenog setvom na 50 cm. Nasuprot dobijenim podacima u ovom radu, Vasiljevic et al. (2010) su u istom agroekološkom regionu (južnobački) u godini za-snivanja crvene deteline za period 2008. i 2009. ostvarili najveći prinos semena crvene deteline (686,2 kg ha-1 i 680,7 kg ha-1) setvom na 60 cm i primenom 4,2 kg ha-1 semena, odnosno setvom na 40 cm i primenom 3,5 kg ha-1 semena, što je u saglasnosti sa rezultatima ruskih istrašivača (Ar-temenko & Kuljka 1988, Kišenjkov & Putnjikov 1983, Perepravo & Akatišev 1989). Prema njima, širokoredna setva sa niskim setvenim normama u uslovima aridnog klimata osigurava optimalan broj biljaka, koji ne dovodi do ¨zagušenja¨, te se na taj način može osigurati visoka proizvodnja semena.
Tabela 1. Uticaj međurednog rastojanja i setvene norme na prinos semena (kg ha-1) crvene deteline u prvom otkosu 2006. Table 1. Effect of row spacing and seeding rate on seed yield (kg ha-1) of red clover in the fi rst cut of 2006
Prinos semena ( kg ha -1 ) crvene deteline u prvom otkosu 2006. Seed yield ( kg ha-1 ) of red clover in the fi rst cut of 2006
Međuredno rastojanje / Row spacing (cm) ( A )
Setvena norma / Seeding rate (kg ha-1) ( B )
2,5 ( B 1 ) 5 ( B 2 ) 10 ( B 3 ) Prosek AAverage A
12,5 ( A 1 ) 294,7 387,1 435,2 372,3
25 ( A 2 ) 351,4 420,7 482,4 418,1
50 ( A 3 ) 318,2 402,5 465,3 395,3
Prosek B – Average B 321,5 403,4 460,9 395,2
L S D
NIVO A** B** AB
5% 14,99 19,23 31,00
1% 22,72 26,34 43,39

Barać R i sar.158
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
Korošec (1972) je dobio visok prinos semena cr-vene deteline u godini setve pri međurednom ra-stojanju od 20 cm, a u drugoj godini 40 cm.
U drugoj godini istraživanja najveći prinos se-mena od 546,2 kg ha-1 ostvaren je takođe setvom 10 kg ha-1 semena na međuredno rastojanje od 25 cm, a najmanji prinos (364,6 kg ha-1) dobijen je setvom 2,5 kg ha-1 semena na 12,5 cm (Tab. 2). Razlike u prinosu semena po hektaru tokom
obe posmatrane godine u odnosu na ispitivane faktore (način setve, količina semena) statistički su visoko značajne (Tab. 1 i Tab. 2).
Ispitujući uticaj načina i gustine setve na prinos semena crvene deteline u agroekološkim uslovi-ma zapadnog Srema, autori Vučković et al. (1997) su ostvarili maksimalni prinos semena crvene de-teline (554,8 kg ha-1) setvom na 20 cm i prime-nom 10 kg ha -1 semena.
Tabela 2. Uticaj međurednog rastojanja i setvene norme na prinos semena (kg ha-1) crvene deteline u drugom otkosu 2007. Table 2. Effect of row spacing and seeding rate on seed yield (kg ha-1) of red clover in the second cut of 2007
Prinos semena ( kg ha -1 ) crvene deteline u drugom otkosu 2007. godineSeed yield ( kg ha-1 ) of red clover in the second cut of 2007
Međuredno rastojanje / Row spacing (cm) ( A )
Setvena norma / Seeding rate (kg ha-1) ( B )
2,5 ( B 1 ) 5 ( B 2 ) 10 ( B 3 ) Prosek AAverage A
12,5 ( A 1 ) 364,6 442,1 511,7 439,5
25 ( A 2 ) 425,8 485,3 546,2 485,7
50 ( A 3 ) 397,4 469,1 532,6 466,3
Prosek B – Average B 395,9 465,5 530,1 463,8
L S D
NIVO A** B** AB
5% 21,69 18,64 34,05
1% 32,87 25,53 48,46
Tabela 3. Uticaj međurednog rastojanja i setvene norme na prinos suve materije (t ha-1) crvene deteline u drugom otkosu 2006. Table 3. Effect of row spacing and seeding rate on dry matter yield (t ha-1) of red clover in the second cut of 2006
Prinos suve materije (t ha -1) crvene deteline u drugom otkosu 2006. Dry matter yield (t ha-1) of red clover in the second cut of 2006
Međuredno rastojanje / Row spacing (cm) ( A )
Setvena norma / Seeding rate (kg ha-1) ( B )
2,5 ( B 1 ) 5 ( B 2 ) 10 ( B 3 ) Prosek AAverage A
12,5 ( A 1 ) 0,97 1,16 1,35 1,16
25 ( A 2 ) 0,95 1,10 1,29 1,10
50 ( A 3 ) 0,90 1,01 1,01 0,99
Prosek B – Average B 0,94 1,09 1,22 1,08
L S D
NIVO A B* AB
5% 0,19 0,15 0,28
1% 0,29 0,28 0,41

Prinos semena i suve materije crvene deteline 159
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
Stjepanović i sar. (1990) navode da crvena de-telina daje prinose semena od 200 kg ha-1 do 800 kg ha-1 u zavisnosti od uslova gajenja. Proizvod-ni potencijal crvene deteline je do 1000 kg ha-1 semena, ali u savremenim uslovima proizvodnje prinosi od 500 kg ha-1 mogu se smatrati vrlo do-brim i realnim.
Prinos suve materije
Prinos suve materije iz drugog otkosa u godini zasnivanja crvene deteline za kombinovano isko-rišćavanje (Tab. 3) bio je znatno niži u odnosu na prinos suve materije ostvaren iz prvog otkosa druge godine istraživanja (Tab. 4). Tokom obe posmatrane godine razlike u prinosu suve mate-rije između različitih načina setve nisu bile stati-stički značajne, dok su razlike dobijene setvom različitih količina semena statistički značajne.
Najveći prinos suve materije crvene deteline iz prvog otkosa u drugoj godini istraživanja od 6,76 t ha-1 dobijen je setvom 10 kg ha-1 semena na me-đuredno rastojanje od 12,5 cm, dok je najmanji prinos od 3,55 t/ha dobijen setvom 2,5 kg ha-1 semena na 50 cm (Tab. 4).
Proučavajući uticaj načina setve u zasnivanju crvene deteline za kombinovanu proizvodnju (seno-seme) na zemljištu tipa parapodzol, loka-litet Kruševac, Lugić i sar. (1996) preporučuju setvenu normu od 14 kg ha1 sa međurednim ra-stojanjem 12,5 cm i 25 cm. Prethodna istraživanja na istom lokalitetu (Miladinović 1978) sa fi ksnom setvenom normom od 20 kg ha-1 pokazala su da je
prinos vegetativne mase kako po godinama tako i ukupno bio najveći pri međurednom rastojanju od 20 cm, dok je najviši prinos semena osvaren pri međurednom razmaku od 40 cm.
U istočnim krajevima SAD crvena detelina za kombinovano iskorišćavanje seje se setvenom normom 10 kg ha-1 do 15 kg ha-1 (Rincker & Ramptom 1985).
Zaključci
Na osnovu rezultata istraživanja uticaja načina i gustine setve na prinos semena i suve materije crvene deteline u sistemu proizvodnje za kom-binovano iskorišćavanje, mogu se izvesti sledeći zaključci:
Vremenski uslovi na lokalitetu Futog (južno-bački region) su povoljni za proizvodnju semena u sistemu proizvodnje crvene deteline za kombi-novano iskorišćavanje.
Adekvatnim izborom optimalnog sklopa bilja-ka mogu se ostvariti visoki prinosi semena crvene deteline. Prosečno najveći prinos semena u obe godine istraživanja (482,4 kg ha-1, odnosno 546,2 kg ha-1) je postignut setvom 10 kg ha-1 semena na međuredno rastojanje od 25 cm.
Prosečno najveći prinos suve materije u obe godine istraživanja postignut je setvom 10 kg semena na međuredno rastojanje od 12,5 cm. U prvoj godini istraživanja iz drugog otkosa postignut je prosečan prinos suve materije od samo 1,08 t ha-1 što je posledica nedostatka vlage.
Tabela 4. Uticaj međurednog rastojanja i setvene norme na prinos suve materije (t ha-1) crvene deteline u prvom otkosu 2007. Table 4. Effect of row spacing and seeding rate on dry matter yield (t ha-1) of red clover in the fi rst cut of 2007
Prinos suve materije (t ha -1 ) crvene deteline u prvom otkosu 2007. Dry matter yield (t ha-1 ) of red clover in the fi rst cut of 2007
Međuredno rastojanje / Row spacing (cm) ( A )
Setvena norma / Seeding rate (kg ha-1) ( B )
2,5 ( B 1 ) 5 ( B 2 ) 10 ( B 3 ) Prosek AAverage A
12,5 ( A 1 ) 4,37 5,95 6,76 5,69
25 ( A 2 ) 3,94 5,28 6,39 5,20
50 ( A 3 ) 3,55 4,73 5,88 4,76
Prosek B – Average B 3,94 5,22 6,34 5,22
LSD
NIVO A B* AB
5 % 0,99 1,26 2,03
1 % 1,50 2,50 2,85

Barać R i sar.160
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 155-160
Literatura
Artemenko M I, Kuljka L S (1988): Polučenije semjan klevera lugovogo v god poseva. Selekcija i semenovodstvo 5: 55-56
Duronić G (2010): Uticaj načina setve i količine semena na pri-nos i kvalitet semena crvene deteline (Trifolium pratense L.). Magistarski rad. Univerzitet u Beogradu, Poljoprivredni fa-kultet
Karagić Đ, Jevtić G, Terzić D (2010): Forage legumes seed pro-duction in Serbia. Biotechnology in animal husbandry 26: 133-149
Kišenjkov F G, Putnjikov L I (1983): Opit proizvodstva semjan klevera lugavogo. Selekcija i semenovodstvo 3: 35-36
Korošec J (1972): Vpliv nekatorih agrotehničkih ukrepov na pridelek črne detelje. I Jugoslovenski simpozium o krmnom bilju, Kruševac
Kovačević N, Kundaković T (2005): Biljni proizvodi u preven-ciji i terapiji blažih oblika simptoma menopauze. Arhiv za farmaciju 55: 197-208
Lugić Z, Krstić O, Tomić Z, Radović J (1996): Uticaj načina setve na produkciju suve mase i semena crvene deteline (Trifolium pratense L.), Zbornik radova Instituta za ratarstvo i povrtarstvo 26: 259-264
Lugić Z, Radović J, Terzić D, Tomić Z, Spasić R (1999): Seme-narstvo višegodišnjih leguminoza u Centru za krmno bilje Kruševac. Sel. Semen. 4: 27-32
Miladinović M (1978): Uticaj načina setve na prinos semena i vegetativne mase crvene deteline. Savremena poljoprivreda 7-8: 69-74
Miladinović M (2001): Proizvodnja semena krmnog bilja. Fe-ljton, Novi Sad,75-96
Perepravo N I, Akatišev P M (1989): Formirovanije semenih travostoev s optimaljnoj gustotoj stojanjija rastenjij. Selekcija i semenovodstvo 6: 36-37
Rincker C M, Rampton H H (1985): Seed production, In: N.L. Taylor (ed). Clover Science and Technology. Madison WI. ASA-CSSA-SSSA Mono. 25: 417-443
Stjepanović M, Bošnjak D, Popović S (1990): Stanje i perspec-tive proizvodnje sjemena krmnih leguminoza i trava. Poljo-privredne aktuelnosti 37: 499-505
Vasiljević S, Mihailović V, Katić S, Mikić A, Karagić Đ (2010): Potencijal rodnosti sorti crvene deteline (Trifolium pratense L.). Ratar. Povrt. / Field Veg. Crop Res. 47: 217-223
Vasiljević S, Karagić Đ, Mihailović V, Pataki I, Milošević B, Pejić B (2010): Effect of sowing method and seeding rate on yield components and seed yields in red clover (Trifolium pratense L.). Biotechnology in animal husbandry 26: 159-165
Vučković S, Pavešić-Popović J, Nedić M, Glamočlija Đ, Zarić D, Radović J (1997): Infl uence of Row Spacing and Sowing rate on Red Clover (Trifolium pratense L.) Seed Yield and Qu-ality. J. Sci. Agric. Res. 58: 75-85
Effect of Row Spacing and Seeding Rate on Seed and Dry Matter Yield of Red Clover (Trifolium pratense L)
Rade Barać1 · Gojko Duronić2 · Đura Karagić3 · Sanja Vasiljević3 · Branko Milošević3
1 Agricultural Highschool, Futog, Serbia2D.O.O. COPEX, Novi Sad, Serbia
3Institute of Field and Vegetable Crops, Novi Sad, Serbia
Summary: A two-year study (2006 and 2007) was carried out to examine the effect of row spacing and seeding rate on seed and dry matter yield of red clover in a production system for mixed utilization. Field experiments were conducted at the fi eld of Agricultural Highschool in Futog, Serbia. Red clover crop was established in three row distances: 12.5 cm, 25 cm and 50 cm, and three seeding rates: 2.5 kg ha-1, 5 kg ha-1 and 10 kg ha-1. Studies have shown that in the location of Futog (region of south Bačka), sowing 10 kg ha-1 with row distance of 25 cm provides red clover seed yield of 482.4 kg ha-1 in the fi rst year of growing and 546.2 kg ha-1 in the second year. The highest yield of dry matter grown in the system of production for mixed use (seed-forage) in Futog was achieved with the row distance of 12.5 cm and seeds rate of 10 kg ha-1.Key words: dry matter yield, red clover (Trifolium pratense L), row spacing, seed yield, seeding rate

Otpornost topinambura prema sivo-mrkoj pegavosti suncokreta 161
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166originalni naučni rad / original research article
Otpornost topinambura (Helianthus tuberosus L.) prema sivo-mrkoj pegavosti suncokreta
u poljskim uslovima
Sreten Terzić · Boško Dedić · Jovanka Atlagić · Vladimir Miklič
received / primljeno: 27.09.2010. accepted / prihvaćeno: 19.11.2010.© 2011 IFVC
Izvod: Topinambur je višegodišnja vrsta suncokreta poreklom iz centralnog dela Severne Amerike. Ova vrsta je prepoznata kao potencijalni izvor otpornosti na neke od ekonomski značajnih bolesti koje se javljaju kod gajenog suncokreta kao što je sivo-mrka pegavost stabla. U ogledu je korišćena ukupno 141 populacija topinambura na karantinskoj parceli Instituta za ratarstvo i povrtarstvo u Novom Sadu. Inokulisane su biljke u stalnom zasadu i u oglednoj parceli na kojoj su sađene krtole izolatima Phomopsis helianthi sa Rimskih Šančeva. Nakon sedam dana gajenja na PDA (Potato-Dextrose Agar) podlozi, isečci micelije gljive Phomopsis helianthi su stavljani na vrh lista i potom prekrivani folijom. Napredovanje bole-sti je mereno u pravilnim vremenskim intervalima. Praćen je intenzitet prirodne infekcije i zabeležena pojava ostalih bolesti. Utvrđene su značajne razlike u brzini napredovanja bolesti između pojedinih populacija. Populacije TUB2046, TUB2062, TUB CG 65 su pokazale najveću otpornost.Ključne reči: otpornost, Phomopsis helianthi, topinambur
S. Terzić · B. Dedić · J. Atlagić · V. MikličInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Topinambur je višegodišnja vrsta suncokreta poreklom iz centralnog dela Severne Amerike koja je početkom XVII veka prenešena u Evro-pu. On pripada obimnom i polimorfnom rodu Helianthus L. u okviru kojeg su Schilling i Heiser (1981) opisali 49 vrsta. U odnosu na većinu gaje-nih biljaka, topinambur je domaćin za mali broj bolesti i štetočina koje ga mogu ugroziti. Najveće gubitke u prinosu uzrokuju gljivična ili bakterijska oboljenja. Od gljiva značajne štete mogu uzroko-vati Puccinia helianthi Schw. i Sclerotium rolfsii Sacc. Sclerotinia sclerotiorum (Lib) de Bary, koja je široko rasprostranjena i napada stablo, koren i krtole to-pinambura. Može da uzrokuje rano uginuće bilj-ke, ali šire zaraze nisu česte (Kays & Nottingham 2007).
Genetski resursi su od značaja za svaki opleme-njivački program. Topinambur je prepoznat kao potencijalni izvor otpornosti na neke od ekonom-ski značajnih bolesti koje se javljaju kod gajenog suncokreta, kao što su plamenjača i bela trulež
cvasti, ali i volovod kao parazitna cvetnica. Zbog toga se vrše međuvrsna ukrštanja sa gajenim sun-cokretom radi prenosa poželjnih osobina u vidu otpornosti na bolesti ili insekte (Seiler 1992). Topinambur je testiran i kao izvor otpornosti na sivo-mrku pegavost stabla suncokreta (prouzro-kovač Phomopsis (Diaporthe) helianthi) i pokazalo se da postoje populacije koje bi mogle da se iskori-ste za oplemenjivanje gajenog suncokreta (Dozet et al. 1992). Pojava ove gljive i značajne štete su prvi put zabeleženi u Srbiji početkom osamdese-tih godina (Mihaljčević i sar. 1980).
Komercijalni hibridi sa visokim nivoom tole-rantnosti prema P. helianthi dostupni su od kada je Škorić (1985) došao do visoko tolerantnih inbred linija dobijenih ukrštanjem gajenog suncokreta sa vrstom H. tuberosus, vrste H. argophyllus sa popu-lacijom H. annuus Armavirski 9345 i jedne linije koja potiče od lokalne populacije iz Maroka. Na bazi tih inbred linija, on je razvio visoko toleran-tne hibride: NS-H-43, NS-H-44 i NS-H-45 (Ško-rić 1985).
Danas je P. helianthi rasprostranjen u mnogim zemljama i odlikuje se velikom varijabilnošću izolata u pogledu fenotipa i agresivnosti (Viguie et al. 1999). Studije sprovedene u Francuskoj na izolatima ove gljive iz poslednje epidemije 1997, korišćenjem AFLP markera (Amplifi ed Fragment
Fitopatologija / Phytopathologyw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Terzić S i sar.162
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Length Polymorphism) potvrdile su pretpostavke o genetskoj varijabilnosti na osnovu čega su au-tori istraživanja izneli pretpostavku o mogućem smanjenju osetljivosti pojedinih populacija prema fungicidima i prevazilaženje postojećeg nivoa ot-pornosti (Says-Lesage et al. 2002). Pri poređenju izolata Yu 4 i Yu 12 od kojih je Yu 12 poreklom iz epidemije 1997. Jocić i sar. (2004) su ustanovi-li da između njih nema značajnih razlika i da je epidemija iz 1997. u dolini reke Dunav rezultat povoljnih klimatskih uslova za razvoj bolesti.
Pošto postojeći izvor otpornosti na P. helianthi funkcioniše već 25 godina i postoji realna opa-snost da bude prevaziđen, cilj ovog rada je prona-laženje novih izvora otpornosti prema P. helianthi. Zbog izražene genetske varijabilnosti unutar vrste (Atlagić i sar. 2006) kao i činjenice da je potvrđe-no postojanje otpornosti u ranijim istraživanjima (Škorić 1985) testirane su populacije topinambu-ra na otpornost prema P. helianthi.
Materijal i metod
Ogled je izveden na lokalitetu Rimski Šančevi, na karantinskoj parceli Odeljenja za uljane kultu-re Instituta za ratarstvo i povrtarstvo u Novom Sadu. U ogledu je korišćena ukupno 141 popula-cija topinambura. Među njima je 27 gajenih sorti (TUB BP), 111 divljih populacija koje su uklju-čene u kolekciju putem sakupljačkih ekspedicija u Sjedinjenim Američkim Državama (38) i Crnoj Gori (73) i tri lokalne populacije (Tab. 1. i 2). Bilj-ke su gajene 2007. godine na dva načina, u ogled-noj parceli i u stalnom zasadu. U oglednoj par-celi je svaka populacija gajena u po jednom redu dužine 7,5 m sa po 15 biljaka. Setva je izvršena presađivanjem krtola u deo karantinske parcele agrotehnički pripremljene za gajeni suncokret, ukupne površine 1,2 ha. Prihrana zemljišta je iz-vršena sa 300 kg ha-1 NPK đubriva tipa 15:15:15. U stalnom zasadu su populacije gajene u parceli-cama širine 0,5 m i dužine 3m bez kontrole broja biljaka unutar parcelice, u okviru kolekcije divljih vrsta suncokreta. Prihrana zemljišta je izvršena sa 300 kg ha-1 NPK đubriva tipa 15:15:15 rasutog po površini. Uređivanje parcelica je vršeno u proleće pre pojave izdanaka, a navodnjavanje po potrebi sistemom za navodnjavanje.
Infekcija je vršena po metodi Tourvieille de La-brouhe et al. (1988). Inokulisano je po dva lista na po pet biljaka za svaku ispitivanu populaciju. Inokulisani su listovi koji se nalaze na polovini ukupne visine stabla.
Kao inokulum je korišćena micelija gljive P. he-lianthi na PDA podlozi stara 4 dana. Zbog razlike u fazama vegetacije između populacija topinam-
bura, listovi testiranih biljaka su inokulisani suk-cesivno u fazi neposredno pre početka cvetanja.
Intenzitet infekcije je meren 14 dana nakon inokulacije određivanjem napretka gljive preko glavnog lisnog nerva ka lisnoj dršci i izražavan je u milimetrima, čime je dobijena apsolutna ocena intenziteta infekcije. Dužina liske je iskorišćena za dobijanje relativne ocene otpornosti kao procenta liske zahvaćene gljivom/pegom. Na istoj parceli su kao kontrole korišćene osetljiva inbred linija gajenog suncokreta NSL1 i tolerantna inbred li-nija NSL2. Ocena biljaka koje nisu inokulisane je vršena prema skali od 0 do 10 (Jocić i sar. 2004) na po 5 biljaka po populaciji.
Zabeleženo je i prisustvo ostalih bolesti u toku vegetacije na svim biljkama radi ocene poljske ot-pornosti za oba tipa gajenja.
Rezultati i diskusija
Količina padavina 2007. godine je bila u nivou proseka ili veća kao u februaru, maju i periodu od avgusta do novembra, ali je april bio bez pa-davina. Temperature su se kretale oko prosečnih vrednosti. Apsolutni temperaturni maksimum na nivou mesečne dekade od 42˚C je zabeležen u julu. Klimatski uslovi što se tiče razvoja P. heli-anthi su bili relativno nepovoljni zbog perioda sa povišenim temperaturama.
Na linijama koje su korišćene kao kontrole za-beležen je različit intenzitet infekcije, koji je na osetljivoj kontroli NSL1 je posle 14 dana iznosio 62 mm, a na tolerantnoj NSL2 34 mm.
Iako su rezultati istraživanja pokazali da je došlo do infekcije kod svih testiranih populacija, evidentno je postojanje visokog nivoa toleran-tnosti. Nakon 14 dana od inokulacije, samo kod 17 od 141 testirane populacije procenat liske sa simptomima bolesti je iznosio više od 50% (Tab. 1. i 2).
Na kontrolnom uzorku populacija, deo liske sa simptomima se povećao za 15% nakon 28 dana od inokulacije, što je bilo dvostruko manje od prosečnog napretka za prvih 14 dana (37%). To ukazuje na usporenje napredovanja patogena što objašnjava i krajnji rezultat da do kraja vegetacije nije došlo do pojave simptoma na stablu. Rezul-tati posle 28 dana su bili nekompletni jer su po-jedine populacije zbog granatosti ranije odbacile listove sa centralnog stabla zbog čega ovi rezultati nisu prikazani.
U ogledu je zabeleženo ukupno dvadeset od testirane 141 populacije sa manje od 25% liske zahvaćenih simptomima bolesti, od čega je 13 od 73 poreklom iz Crne Gore (Tab. 1), 6 od 38 iz SAD-a i 1 od 27 gajenih sorti (Tab. 2).

Otpornost topinambura prema sivo-mrkoj pegavosti suncokreta 163
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Tabela 1. Reakcija populacija topinambura poreklom iz Crne Gore na Phomopsis helianthi Munt.-Cvet.Table 1. Reaction of topinambour populations from Montenegro to Phomopsis helianthi Munt.-Cvet.
PopulacijaPopulation
intenzitetinfekcijeinfection intensity
(mm)
% liske sa simptomima
bolesti% of leaf
with disease symptoms
PopulacijaPopulation
intenzitetinfekcijeinfection intensity
(mm)
% liske sa simptomima
bolesti% of leaf
with disease symptoms
TUB CG 3 62 27 TUB CG 41 86 41TUB CG 4 64 38 TUB CG 42 44 21TUB CG 5 44 21 TUB CG 43 75 37TUB CG 6 54 25 TUB CG 44 73 34TUB CG 7 60 30 TUB CG 45 81 47TUB CG 8 43 19 TUB CG 46 91 42TUB CG 9 79 59 TUB CG 47 88 45TUB CG 10 58 28 TUB CG 48 68 39TUB CG 11 67 30 TUB CG 49 75 35TUB CG 12 58 28 TUB CG 50 71 40TUB CG 13 46 22 TUB CG 51 81 38TUB CG 14 36 24 TUB CG 52 79 35TUB CG 15 46 22 TUB CG 53 93 45TUB CG 16 68 31 TUB CG 54 78 39TUB CG 17 62 31 TUB CG 55 36 21TUB CG 18 68 32 TUB CG 56 72 43TUB CG 19 106 75 TUB CG 57 88 40TUB CG 20 35 14 TUB CG 58 69 36TUB CG 21 39 21 TUB CG 59 74 45TUB CG 22 54 27 TUB CG 60 76 35TUB CG 23 52 26 TUB CG 61 61 35TUB CG 24 65 32 TUB CG 62 39 20TUB CG 25 43 22 TUB CG 63 70 31TUB CG 26 63 36 TUB CG 65 19 10TUB CG 27 65 44 TUB CG 66 73 36TUB CG 28 56 30 TUB CG 67 64 29TUB CG 69 64 32 TUB CG 69 80 36TUB CG 70 77 34 TUB CG 70 51 25TUB CG 72 51 27 TUB CG 72 57 28TUB CG 75 70 30 TUB CG 75 52 25TUB CG 33 68 32 TUB CG 76 86 42TUB CG 34 78 37 TUB CG 77 84 48TUB CG 36 81 38 TUB CG 78 37 19TUB CG 37 82 37 TUB CG 79 56 28TUB CG 38 79 38 TUB CG 80 59 33TUB CG 39 79 37 NSL1 62 37TUB CG 40 81 40 NSL2 34 20

Terzić S i sar.164
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Tabela 2. Reakcija divljih populacija iz SAD-a i gajenih sorti topinambura na Phomopsis helianthi Munt.-Cvet.Table 2. Reaction of wild populations from USA and topinambour cultivars to Phomopsis helianthi Munt.-Cvet.
PopulacijaPopulation
intenzitetinfekcijeinfection intensity
(mm)
% liske sa simptomima
bolesti% of leaf
with disease symptoms
PopulacijaPopulation
intenzitetinfekcijeinfection intensity
(mm)
% liske sa simptomima
bolesti% of leaf
with disease symptoms
TUB 6 76 39 TUB 2080 58 28TUB 7 58 26 TUB 2089 59 32TUB 8 70 35 TUB 2189 126 68TUB 15 56 34 TUB BP 1 127 72TUB 16 25 14 TUB BP 2 56 30TUB 20 55 31 TUB BP 3 86 48TUB 26 33 17 TUB BP 4 69 41TUB 675 50 29 TUB BP 5 64 35TUB 1540 41 20 TUB BP 6 34 18TUB 1625 79 40 TUB BP 7 101 46TUB 1628 57 31 TUB BP 8 75 40TUB 1698 117 61 TUB BP 9 131 74TUB 1699 75 40 TUB BP 10 60 31TUB 1700 118 70 TUB BP 11 62 39TUB 1701 83 46 TUB BP 12 167 82TUB 1702 73 37 TUB BP 13 66 37TUB 1703 69 40 TUB BP 14 116 64TUB 1704 84 52 TUB BP 15 76 47TUB 1705 53 27 TUB BP 16 75 50TUB 1945 70 40 TUB BP 17 73 40TUB 1959 86 53 TUB BP 19 55 38TUB 2024 54 33 TUB BP 20 61 35TUB 2045 77 38 TUB BP 21 70 35TUB 2046 20 10 TUB BP 22 63 48TUB 2047 76 40 TUB BP 23 121 66TUB 2050 63 34 TUB BP 24 60 36TUB 2051 61 23 TUB BP 25 65 39TUB 2052 80 54 TUB BP 26 62 37TUB 2061 62 30 TUB BP 27 63 35TUB 2062 18 10 TUB Č 189 100TUB 2066 89 42 TUB NS 1 64 25TUB 2067 137 61 TUB NS 2 51 30TUB 2069 73 42 NSL1 62 37TUB 2070 135 67 NSL2 34 20TUB 2071 48 25

Otpornost topinambura prema sivo-mrkoj pegavosti suncokreta 165
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Ukupno šest populacija je imalo intenzitet in-fekcije isti ili manji od tolerantne kontrole NSL2. Od tih šest populacija četiri su bile poreklom iz SAD-a. Veći ili isti intenzitet infekcije u odnosu na osetljivu kontrolu je imalo 92 populacije, od čega je 45 od 73 poreklom iz Crne Gore, 22 od 38 iz SAD-a, 20 od 27 gajenih sorti i 2 od 3 lo-kalne populacije. Ako bi se po poreklu uporedio udeo populacija osetljivijih od osetljive kontrole populacije poreklom iz Crne Gore i SAD-a su približno iste sa 58% i 61% dok kod gajenih sorti taj udeo iznosi 74%.
Testom otpornosti na sivo-mrku pegavost sta-bla populacija jednogodišnje vrste suncokreta H. petiolaris, autori Caseres et al. (2007) su ustanovili pojavu infekcije na svim inokulisanim biljkama sa prosečnim intenzitetom infekcije od 22 mm, a označili su kao visoko tolerantne populacije sa in-tenzitetom infekcije do 17 mm, odnosno one kod kojih je do 23% liske imalo simptome bolesti. Samo tri populacije (TUB2046, TUB2062, TUB CG 65) su imale intenzitet infekcije manji od 22 mm (Tab. 1. i 2). Broj populacija sa procentom li-ske sa simptomima manjim od 23% je bio veći jer su liske vrste H. tuberosus skoro dvostruko veće od liski vrste H. petiolaris (Heiser et al. 1969).
Od ostalih bolesti su na osetljivoj kontroli zabe-ležene: mrka pegavost lišća (prouzrokovač Alter-naria helianthi), crna pegavost stabla (prouzroko-vač Phoma macdonaldii) i pepelnica (prouzrokovač Erysiphe chichoracearum). Ispitivanje poljske otpor-nosti je uključeno u ovaj rad zbog povećanog in-teresa za topinambur kao gajenu biljku (Seiler et al. 2006; Kays & Nottingham 2007) i malog broja literaturnih izvora koji opisuju bolesti i štetočine koje bi ga mogle ugroziti u agroekeološkim uslo-vima gajenja na teritoriji Srbije.
U oceni poljske otpornosti topinambura kon-statovani su simptomi pepelnice koja se pojavila pri kraju faze cvetanja na 99 od 141 populacije u oglednoj parceli i 71 od 95 opažanih u stalnom zasadu. Od ostalih patogena najčešća je bila bela trulež korena koja se javila samo kod biljaka u oglednoj parceli, a zabeležena je na ukupno 16 populacija sa učestalošću od 7% do 30% biljaka
po populaciji. Bela trulež korena se javila na poje-dinim biljkama, kako se inače i javlja na parcelama sa topinamburom, gde retko zahvata veću povr-šinu (Kays & Nottingham 2007). Crna pegavost stabla je zabeležena na 4 populacije na oglednoj parceli sa učestalošću od 7% do 40%.
U oglednoj parceli se osim pepelnice pojavila sivo-mrka pegavost lista na 6 populacija sa uče-stalošću od 7% do 27% i sivo-mrka pegavost stabla na populaciji TUB BP 12. U stalnom za-sadu su simptomi iste bolesti zabeleženi samo na populaciji TUB1625. Manja učestalost poja-ve sivo-mrke pegavosti putem prirodne infekcije može se objasniti nedostatkom prouzrokovača bolesti, ali i relativno nepovoljnim uslovima za njegov razvoj, pogotovo u parcelicama stalnog zasada gde je relativna vlažnost vazduha bila niža. Istraživanjima na gajenom suncokretu je potvrđeno da su agrotehničke mere takođe važne za suzbijanje P. helianthi jer gustina setve pozitivno utiče na povećanu relativnu vlažnost vazduha u usevu, a samim tim i na povećanu učestalost infekcija lista i lezija stabla (Debaeke & Moinard 2010). Pored toga, najbolji način za kontrolu ove gljive jeste gajenje otpornih hibri-da (Mihaljčević i sar. 1982).
Zaključak
Velika genetska varijabilnost unutar kolekcije topinambura se iskazala kroz značajne razlike u intenzitetu napredovanja bolesti između poje-dinih populacija. Ustanovljeno je da 18 od 141 testirane populacije spada u grupu visoko tole-rantnih prema sivo-mrkoj pegavosti i da bi se kao takve mogle upotrebiti u oplemenjivanju gajenog suncokreta.
Zbog većeg udela visoko tolerantnih popula-cija u grupi iz SAD-a ona bi se mogla označiti kao interesantnija u odnosu na ostale grupe za dalji rad na traženju novih izvora otpornosti uz populacije poreklom iz Crne Gore. Populacije TUB2046, TUB2062 i TUB CG 65 su pokazale najveću tolerantnost prema prouzrokovaču sivo-mrke pegavosti suncokreta P. helianthi.

Terzić S i sar.166
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 161-166
Literatura
Atlagić J, Terzić S, Škorić D, Marinković R, Vasiljević Lj, Panko-vić D (2006): The wild sunfl ower species collection in Novi Sad. Helia 44: 55-64
Caseres C, Castano F, Rodriguez R, Ridao A, Salaberry T, Eche-veria M, Colabelli M (2007): Phomopsis resistance on leaves and stems of Helianthus petiolaris. Helia, 47: 213-218
Debaeke P, Moinard J (2010): Effect of crop management on epidemics of phomopsis stem canker (Diaporthe helianthi) for susceptible and tolerant sunfl ower cultivars. Field Crops Res. 115: 50–60
De Labrouhe T, Vear F, Pelletier C (1988): Use of two myceli-um tests in breeding sunfl ower resistant to Phomopsis. 110-114. In Proc. 12th Int. Sunfl ower Conf., Novi Sad, Serbia. 25-29 July. Int. Sunfl ower Assoc., Paris, France
Dozet B M, Atlagic J, Skoric D (1992): Sources of resistance to Diaporthe/Phomopsis helianthi and their use in sunfl ower bree-ding applying the in vitro embryo culture. p. 1449-1454. In Proc. 13th Int. Sunfl ower Conf., Pisa, Italy. 7-11 September. Int. Sunfl ower Assoc., Paris, France
Gulya T J, Maširević S (1991): Common names for plant disea-ses: sunfl ower (Helianthus annuus L.) and Jerusalem artichoke (Helianthus tuberosus L.). Plant Dis.75: 230
Heiser Ch B, Smith D M, Clevenger S B, Martin W C (1969): The North American Sunfl owers (Helianthus). Mem. Torrey Bot. Club, 22: 219
Jocić S, Lačok N, Miklič V, Škorić D, Griveau Y (2004): Testing two isolates of Diaporthe/Phomopsis helianthi in a populati-on of sunfl ower recombinant inbred lines. Helia 41: 129-136
Kays S J, Nottingham S F (2007): Biology and chemistry of Jerusalem artichoke (Helianthus tuberosus L.). CRC press, Boca Raton, FL, 478
Mihaljčević M, Petrov M, Muntanola-Cvetković M (1980): Pho-mopsis sp. - a new parasite of sunfl ower in Yugoslavia, Savre-mena polj. 11-12: 531-540
Mihaljčević M, Muntanola-Cvetković M, Petrov M (1982): Further studies on the sunfl ower disease caused by Dia-porthe (Phomopsis) helianthi and possibilities of breeding for resistance. in: Int. Sunfl ower Conference (10th), Sur-fers Paradise, Australia, Paris: Int. Sunfl ower Association, 157-159
Says-Lesage V, Roeckel-Drevet P, Viguie A, Tourvieille J, Nico-las P, Tourvieille de Labrouhe D (2002): Molecular variability within Diaporthe/Phomopsis helianthi from France. Phytopat-hol. 92: 308-313
Schiling E E, Heiser Ch B (1981): Infrageneric classifi cation of Helianthus (Compositae). Taxon 30: 393-403.
Seiler G J (1992): Utilization of wild sunfl ower species for the improvement of cultivated sunfl ower. Field Crops Res. 30: 195-230
Seiler G J, Campbell L G (2006): Genetic variability for mineral concentration in the forage of Jerusalem artichoke cultivars. Euphytica 150: 281-288
Škorić D (1985): Sunfl ower breeding for resistance to Diaport-he/Phomopsis helianthi Munt. -Cvet. et al. Helia 8: 21-24
Viguie A, Vear F, Tourvieille de Labrouhe D (1999): Interacti-ons between French isolates of Phomopsis/Diaporthe helianthi Munt.-Cvet. et al. and sunfl ower (Helianthus annuus L.) ge-notypes, Europ. Plant Patho. 105: 693-702
Resistance of Topinambour (Helianthus tuberosus L.) to Sunfl ower Stem Canker in Field Conditions
Sreten Terzić · Boško Dedić · Jovanka Atlagić · Vladimir Miklič
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Topinambour is a perennial species originating from central part of North America. This species is known to be a potential source of resistance to some diseases of cultivated sunfl ower like stem canker. In this re-search we evaluated 141 populations of Helianthus tuberosus from the collection of Institute of Field and Vegetable Crops in Novi Sad. Plants in the collection fi eld and in the experimental fi eld were artifi cially inoculated using Pho-mopsis helianthi isolate from Rimski Šančevi. After seven days of growing on PDA (Potato-Dextrose Agar) medium, mycelial plugs were placed on leaf tip and wrapped with aluminum foil. Disease progress was measured in regular intervals. Occurrence of natural infections was also evaluated and all diseases were noted. As a result of this survey signifi cant differences were detected among populations regarding disease intensity. Out of all assessed populations those labeled as TUB2046, TUB2062 and TUB CG 65 were most resistant.Keywords: Phomopsis helianthi, resistance, topinambour

Sunfl ower Inbred Lines Screening for Tolerance to White Rot on Stalk 167
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172original research article / originalni naučni rad
Sunfl ower Inbred Lines Screeningfor Tolerance to White Rot on Stalk
Boško Dedić · Dragana Miladinović · Siniša Jocić · Sreten Terzić ·Sonja Tančić · Nenad Dušanić · Vladimir Miklič
received / primljeno: 02.11.2010. accepted / prihvaćeno: 14.12.2010.© 2011 IFVC
Summary: Seventy sunfl ower (Helianthus annuus L.) inbred lines were screened for tolerance to white rot on stalk. Plants were inoculated at the budding stage, with 4-day old Sclerotinia mycelium grown on PDA medium. Mycelium was placed on the leaf top and covered with tin foil, and the leaf was put into transparent nylon bag in order to maintain high humidity. Spot length on leaf was measured and plant tolerance was determined at the full fl owering stage. Obtained results were analyzed by analysis of va-riance. Tested lines showed signifi cant variability in response to disease. Eleven lines had lesion length less than 50% compared to susceptible control, so they could be considered tolerant and potentially interesting for breeding program.Key words: Sclerotinia sclerotiorum, sunfl ower, tolerance, white rot
B. Dedić · D. Miladinović · S. Jocić · S. Terzić · S. Tančić · N. Dušanić · V. MikličInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbiae-mail: [email protected]
Introduction
Sclerotinia sclerotiorum (Lib.) De Bary is omnivorus fungi and the causal agent for diseases on more than 400 plant species (Boland & Hall 1994). The fungus had been fi rst identifi ed on sunfl ower in 1861 (Purdy 1979) while in Serbia it was fi rst described in 1948 (Aćimović 1998). It is common and widespread on sunfl ower and can attack all parts of plant. Midstalk rot is one of several forms of disease on sunfl ower and it is incited by the airborne ascospores. These spores are produced in apotecia developed on sclerotia in surface layer of soil. Dependence of apotecial formation on high humidity in the fi rst few centimeters of soil for a certain period of time greatly limits regular appearance of disease, particularly in regions with drier climate. Disease could appear from seedling to maturity, but most commonly after period of fl owering when the most devastating symptom is logging (Gulya et al. 1997). In high S. sclerotiorum infested sunfl ower fi elds under favourable conditions number of infected plants can reach up to 100% (Maširević & Gulya 1992, Rashid 1993).
Complete resistance has not yet been observed and partial resistance is under quantitative control (Bert et al. 2002, Mestries et al. 1998). There has been a great effort in searching for tolerance to midstalk rot both in cultivated sunfl ower and wild sunfl ower species. Hence, several methods of artifi cial inoculation have been developed (Degener et al. 1998, Castano et al. 1992b, Maširević & Gulya 1992, Grauert et al. 1980, Castano et al. 2002, Vasić et al. 2004). To this date, a number of sunfl ower lines and hybrids with various levels of tolerance have been reported (Reimonte & Castano 2008, Rönicke et al. 2004, Vear 2004), but level of tolerance is not yet considered adequate for control of the disease (Fick & Miller 1997).
The aim of this study was to determine variability in response to S. sclerotiorum infection and to fi nd tolerance within selected sunfl ower inbred lines.
Materials and Methods
Sunfl ower Inbred LinesSeventy sunflower (Helianthus annuus L.)
inbred lines were screened for tolerance to midstalk rot (Tab. 1). Tested inbred lines were created at Institute of Field and Vegetable Crops, Novi Sad, Serbia and have diverse origin. There was no prior information about their response to S. sclerotiorum infection of
Phytopathology / Fitopatologijaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Dedić B i sar.168
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
stem. Inbred line NSL 21, which was found to be highly susceptible to midstalk rot in field conditions (data not shown), was used as susceptible control.
Experimental PlanThe experiment was conducted on
experimental fi eld of Institute of Field and Vegetable Crops at Rimski Šančevi. The experiment was designed as completely randomized block system with three replications. Each experimental unit consisted of two 3.6 m length rows. Row spacing was
0.7 m and space between plants in the row was 0.25 m. Seeds of inbred lines were sown manually, three seeds per spot, in the fi rst ten-day period of April and thinned when plants fully developed fi rst true pair of leaves.
Inoculum and InoculationFour-day old Sclerotinia mycelium grown on
PDA medium was used for the inoculation. The fungal isolate used in this study was collected from infected stems of the plants grown at the same fi eld where the test was performed. In total, 36 plants per line were inoculated
GenotypeGenotip
InoculationInokulacija1
Reaction of inbred lines
Reakcija inbred linija2
GenotypeGenotip
InoculationInokulacija1
Reaction of inbred lines
Reakcija inbred linija2
NSL1 + MS NSL37 N SNSL2 + MS NSL38 + SNSL3 + MS NSL39 + MSNSL4 + MS NSL40 N SNSL5 + MS NSL41 + SNSL6 + MS NSL42 + SNSL7 + S NSL43 + MSNSL8 + S NSL44 N SNSL9 + MS NSL45 + MSNSL10 + T NSL46 + MSNSL11 + T NSL47 + MSNSL12 + MS NSL48 + SNSL13 + S NSL49 + SNSL14 + T NSL50 + SNSL15 + MS NSL51 + SNSL16 + MS NSL52 + TNSL17 N S NSL53 + SNSL18 + MS NSL54 + HSNSL19 N S NSL55 + SNSL20 N S NSL56 + SNSL22 N S NSL57 + TNSL23 N S NSL58 + SNSL24 + MS NSL59 N SNSL25 + T NSL60 + SNSL26 + T NSL61 + MSNSL27 + T NSL62 + SNSL28 + MS NSL63 N SNSL29 + MS NSL64 + TNSL30 N S NSL65 + SNSL31 + MS NSL66 + MSNSL32 N S NSL67 + MSNSL33 + T NSL68 + MSNSL34 + MS NSL69 + MSNSL35 N S NSL70 + MSNSL36 + T NSL213 + S
Table 1. Successfully inoculated genotypes and their reaction to leaf inoculation with S. sclerotiorum myceliumTabela 1. Uspešnost inokulacije i reakcija genotipova posle inokulacije sa micelijom gljive S. sclerotiorum

Sunfl ower Inbred Lines Screening for Tolerance to White Rot on Stalk 169
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
at the budding stage, with diameter of bud approximately 5 cm (Mestries et al. 1998). Mycelium plugs from the edge of colony were placed on the leaf top and covered with tin foil, and the leaf was put into transparent nylon bag in order to achieve high humidity (Degener et al. 1998). In addition to this, experimental fi eld was irrigated using sprinklers three times a week with average rate of 15 mm of water per week.
Data Recording and Statistical AnalysisLesion length on leaf was measured
periodically after inoculation and plant tolerance was determined at the full fl owering stage, while percentage of plants without symptoms was recorded. Line NSL21 was used as a susceptible control. Statistical analysis was done using software Statistica 9.0.
Results and Discussion
Inoculation procedure was successful in 57 out of 70 tested inbred lines including control line, while other 13 lines had very low infection rate (Tab. 1). The lines with low infection rate were excluded from the experiment.
1 + - successful inoculation, N - unsuccessful inoculation; 2 T – tolerant genotype, MS – medium susceptible genotype, S – susceptible genotype; 3 susceptible control
1 + - uspešna inokulacija, N – neuspešna inokulacija; 2 T – tolerantni genotipe, MS – srednje osetljiv genotip, S – osetljiv genotip; 3 osetljiva kontrola
Group of 11 lines (NSL52, NSL25, NSL10, NSL27, NSL36, NSL64, NSL57, NSL14, NSL11, NSL26, NSL33) had average relative length of lesion less than 50% compared to lesion length of susceptible control. Eight lines (NSL 61 to NSL45) had relative lesion length very close to lower limit of 50% (46.4% to 56.6%). These eight lines together with lines NSL 5 to NSL 68 could be considered as moderately susceptible, because their reaction to S. sclerotiorum was stronger in terms of lesion length than tolerant lines, although they had more than 25% shorter relative lesion length compared to susceptible control. Group of nine susceptible lines (NSL4, NSL58, NSL13, NSL48, NSL41, NSL42, NSL51, NSL53, NSL7) had relative length of lesion less than susceptible control (80.3% to 97.9%), and group of eight lines (NSL60, NSL8, NSL50, NSL38, NSL62, NSL65, NSL49, NSL54) had the most severe symptoms of white rot (100.4% to 126.8%) (Fig. 1. and Tab. 1).
Average length of lesion on leaves was 6.25 cm for the most tolerant line and 29.13 cm for the most susceptible line (Fig. 2). The box-whiskers plots for lines NSL52, (being the most tolerant one), NSL57, NSL24, NSL12, NSL16, NSL4, NSL53, and NSL50 show high value of standard error. This means that data varies in that extent that certainty for conclusion about tolerance/susceptibility reaction is greatly reduced. Furthermore, great number of outliers in case of lines NSL66, NSL67, NSL41, NSL21, NSL38, NSL49 also makes it diffi cult to reach a conclusion on reaction of the line in question (Fig. 2). These lines should be retested in order to achieve more coherent results.
Fig. 1. Relative length of lesion along mid-vein showed as percentage of lesion length of tested line compared to lesion of susceptible controlSlika 1. Reletivna dužina pega duž srednjeg nerva prikazana kao odnos dužine lezije kod testirane linije i osetljive kontrole

Dedić B i sar.170
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
The existence of variability between different genotypes marked as moderately susceptible and susceptible could be seen from both fi gures. This type of reaction to S. sclerotiorum could be
explained by inheritance of tolerance to this fungus. Studies have shown that tolerance is partial and polygenic, and levels of tolerance vary continually (Vear & Tourvieille 1988, Achbani et al. 1996).
Breeding sunfl ower for tolerance to S. sclerotiorum is connected to various diffi culties. The fungus causes rot on roots, stem, terminal bud and head, and all these are considered as separate diseases. Tolerance is mainly controlled by additive genes and screening for tolerance relies on various methods of artifi cial inoculation.
Suitable methods for artifi cial inoculation were developed in order to successfully test breeding material for disease tolerance in most cases. A good artifi cial inoculation test should produce high infection rate, be easy to handle and imitate natural disease incidence (Degener
et al. 1998). Necessity of artifi cial inoculation of sunfl ower with S. sclerotinia is due to its high dependence on the environmental conditions for natural infection. Reliance on natural attack
of fungus often results in inhomogeneous infection and could produce unreliable results (Serre et al. 2004). Natural attack is highly dependent on high humidity when disease progress is rapid and destructive, whereas dry period immediately after infection could completely stop the disease (Tourvieille et al. 1992). In our study there was complete absence of natural attack in the experimental fi eld, presumably due to high temperatures and low RH during period when sunfl ower plants are most susceptible to mid-stalk rot. This indicates possible escape mechanism when there is source of inoculum but environmental factors are limiting appearance of the disease. Contrary to this, disease development was recorded on artifi cially inoculated leaves.
For the statistical analysis of lesion length we used only data from successfully inoculated plants.
Fig. 2. Mean, standard error, 1.96 SE and outliers for length of lesion along mid-vein on leaf Slika 2. Prosek, standardna greška, 1,96 SE i vrednosti van intervala obuhvaćenog standardnom greškom dužine pege duž srednjeg nerva na listu

Sunfl ower Inbred Lines Screening for Tolerance to White Rot on Stalk 171
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
Leaf lesion is a reliable trait for screening against Sclerotinia and it is relatively simple compared with tests using ascospores on capitula (Castano et al. 1992b, Becelaere & Miller 2004), and tests using mycelium on capitula in growth chamber (Vear & Guillman 1977). However, reliability of this trait was not confi rmed in the study of Degener et al. (1998) who found measurement of stem lesion more adequate.
Although different authors measured leaf lesions 4 to 7 days after inoculation (Degener et al. 1998, Castano et al. 1992a, Castano et al. 1992b, Caceres et al. 2006), in our case it was not possible due to slow progression of leaf lesions. One of the reasons for measuring leaf lesions later than 7 days after inoculation might be environmental conditions, especially high temperatures in the period immediately after artifi cial inoculation (Hahn 2002). However, disease progress was constant and symptoms were similar with the description of this disease. Small number of lines had lesion length lower than 50% compared to susceptible control. Lines that have less than one-half of the infection percentage of highly susceptible lines could be considered as the tolerant ones according to Miller (1992) and Castano et al. (1993). Taking into consideration signifi cant correlation between lesion length and percentage of resistant plants, these lines can be considered tolerant to Sclerotinia midstalk rot.
It is known that responses to foliar and stem inoculations with mycelium are independent (Castaño et al. 1993). However, method of leaf inoculation is much closer to natural infection process than methods used for direct inoculation of stem. Moreover, methods used for direct inoculation of stem often imply artifi cial wounding in which case conclusion of some levels of tolerance might be lost.
Nevertheless, use of different methods might be needed as it is in the case with studying of white rot of head where results produced using inoculation with ascospores and mycelium proved to be complementary (Castano et al. 1993). Eleven tolerant lines were selected based on this study using method for leaf inoculation. Therefore, in order to narrow the selected lines and confirm these results, additional study using methods other than artificial inoculation of leaf will be done on all lines used in this study, with special consideration to those lines which proved to be tolerant in this study, for these lines can be used in breeding programs.
References
Achbani H, Tourvieille de Labrouche D, Vear F (1996): To-lerance of sunfl ower (Helianthus annuus L.) to terminal bud attack by Sclerotinia sclerotiorum (Lib.) de Bary. Europ. J. Plant Pathol. 102: 421-429
Aćimović M (1998): Bolesti suncokreta. Naučni institut za ra-tarstvo i povrtarstvo, Novi Sad
Becelaere G, Miller J F (2004): Methods of inoculation of sun-fl ower heads with Sclerotinia sclerotiorum. Helia 27: 137-142
Bert P F, Jouan I, Tourvieille de Labrouche D, Serre F, Nicolas P, Vear F (2002): Comparative genetic analysis of quantitati-ve traits in sunfl ower (Helianthus annuus L.) .1. QTL involved in tolerance to Sclerotinia sclerotiorum and Diaporthe helianthi. Theor. Appl. Genet. 105: 985-993
Boland G J, Hall R (1994): Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 16: 93-108
Caceres C, Castano F, Rodriguez R, Ridao A, Salaberry T, Eche-verria M, Colabelli M (2006): Variability of Sclerotinia respon-ses in Helianthus petiolaris. Helia 29: 43-48
Castano F, Vear F, Tourvieille de Labrouche D (1992a): Rela-tions between the reaction of sunfl ower genotypes to tests of tolerance to root and leaf attacks by Sclerotinia sclerotiorum. Proc 13th Int. Sunfl ower Conf., Piza, Italy, 1011-1016
Castano F M, Hemery-Tardin M C, Tourvieille de Labrouhe D, Vear F (1992b): The inheritance and biochemistry of to-lerance to Sclerotinia sclerotiorum leaf infections in sunfl ower (Helianthus annuus L.). Euphytica 58: 209-219
Castano F, Vear F, Tourvieille D (1993): Tolerance of sunfl ower inbred lines to various forms of attack by Sclerotinia sclero-tiorum and relations with some morphological characters. Euphytica 68: 85-98
Castano F, Baldini M, Re J, Vanozzi G P, Rodriguez R, Turi M, Tahmasebi-Enferadi S (2002): Genotypic responses and diallel analysis for an early tolerance test to Sclerotinia scle-rotiorum in sunfl ower. Oléagineux, Corps Gras, Lipides 9: 474-478
Degener J, Melchinger E A, Gumber K R, Hahn V (1998): Breeding for Sclerotinia tolerance in sunfl ower: A modifi ed screening test and assessment of genetic variation in current germplasm. Plant Breed. 117: 367-372
Fick G N, Miller J F (1997): Sunfl ower breeding. In: A. Schnei-ter (ed.), Sunfl ower Technology and Production, ASA-CS-SA-SSSA, Madison, Wisconsin, USA, 395-440
Grauert P, Schlösser E, Schuster W (1980): Untersuchungen über die Anfälligkeit von Sorten und Linien der Sonnenblu-me (Helianthus annuus L.) gegenüber Sclerotinia sclerotiorum (Lib.) de Bary. Angew. Botanik 54: 349-364
Gulya T, Rashid K, Maširević S (1997): Sunfl ower diseases. In: A. Schneiter (ed.), Sunfl ower Technology and Production, ASA-CSSA-SSSA, Madison, Wisconsin, USA, 263-379
Hahn (2002): Genetic variation for resistance to Sclerotinia head rot in sunfl ower inbred lines. Field Crop. Res. 77: 153-159
Maširević S, Gulya T (1992): Sclerotinia and Phomopsis - two deva-stating sunfl ower pathogens. Field Crop. Res. 30: 271-300
Mestries E, Gentzbittel L, Tourvieille de Labrouche D, Nicolas P, Vear F (1998): Analyses of quantitative trait loci associ-ated with tolerance to Sclerotinia sclerotiorum in sunfl owers (Helianthus annuus L.) using molecular markers. Mol. Breed. 4: 215-226
Miller J F (1992): Breeding for Sclerotinia tolerance. Proc Sun-fl ower Res. Workshop, Fargo, ND, USA, 10
Purdy L H (1979): Sclerorinia sclerotiorum: history, diseases and symptomatology, host range, geographic distribution and impact. Phytopathol. 69: 875-880
Rashid K Y (1993): Sclerotinia head rot and mid-stem infection of sunfl ower in Manitoba. Proc 15th Sunfl ower Res. Wor-kshop, Fargo, ND, USA

Dedić B i sar.172
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 167-172
Reimonte G, Castano F (2008): Susceptibility of sunfl ower (Helianthus annuus) hybrids to mid stem rot and broken stem caused by Sclerotinia sclerotiorum. Cien. Inv. Agr. 35: 21-28
Rönicke S, Hahn V, Horn R, Gröne I, Brahm L, Schnabl H, Fri-edt W (2004): Interspecifi c hybrids of sunfl ower as a source of Sclerotinia sclerotiorum. Plant Breed. 123: 152-157
Serre F, Walser P, Tourvieille de Labrouche D, Vear F (2004): Sclerotinia sclerotiorum capitulum tolerance tests using ascospo-res: results over the period 1991-2003. Proc 16th Int. Sun-fl ower Conf., Fargo, North Dakota, USA 129-133
Tourvieille D, Vear F, Achbani E H (1992): Attack of sunfl ower terminal buds by Sclerotinia sclerotiorumi in sunfl owers. Proc13-th Sunfl ower Conference, Pisa, Italy, 859-864
Vasić D, Jocić S, Malenčić Đ, Pajević S, Miladinović F, Škorić
D (2004): Testiranje na otpornost prema Sclerotinia različitih genotipova suncokreta. Zbornik radova Instituta za ratarstvo i povrtarstvo, Novi Sad 40: 291-300
Vear F (2004): Breeding for durable tolerance to main diseases of sunfl ower. Proc 16th Int. Sunfl ower Conf., Fargo, North Dakota, USA, 15-28
Vear F, Guillaumin J J (1977): Development of a test useable in breeding for tolerance to sunfl ower capitulum attack by Scle-rotinia sclerotiorum. Proc 7th Int. Sunfl ower Conf., Krasnodar, USSR, 205-212
Vear F, Tourvieille D (1988): Heredity of tolerance to Scleroti-nia sclerotiorum in sunfl ower. II Study of capitulum tolerance to natural and artifi cial ascospore infections. Agronomie 8: 503-508
Testiranje inbred linija suncokreta na tolerantnost prema beloj truleži stabla
Boško Dedić · Dragana Miladinović · Siniša Jocić · Sreten Terzić ·Sonja Tančić · Nenad Dušanić · Vladimir Miklič
Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbija
Izvod: Sedamdeset inbred linija suncokreta (Helianthus annuus L.) je testirano na tolerantnost prema prouzrokovaču bele truleži stabla. Biljke su inokulisane u fazi butonizacije sa 4 dana starom kolonijom gljive gajenom na PDA pod-lozi. Delovi podloge sa micelijom su stavljeni na vrh lista, prekriveni aluminijumskom folijom i potom providnom najlonskom kesom da bi se obezbedio visok nivo vlažnosti. Merena je dužina pega i nivo tolerantnosti u fazi punog cvetanja suncokreta. Testirane linije su pokazale značajnu varijabilnost u reakciji prema patogenu, a 11 linija čija je prosečna relativna dužina pega iznosila ispod 50% u odnosu na osetljivu kontrolu su označene kao tolerantne i potencijalno interesantne za uključivanje u proces oplemenjivanja.Ključne reči: bela trulež stabla, Sclerotinia sclerotiorum, suncokret, tolerantnost

Izolacija i detekcija gljiva A. dauci i A. radicina iz semena mrkve 173
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178originalni naučni rad / original research article
Izolacija i detekcija gljiva Alternaria dauci (Kühn) Groves et Skolk i Alternaria radicina Meier,
Drechsler et Eddy iz semena mrkve
Dragana Petrović · Maja Ignjatov · Milka Vujaković · Zorica Nikolić · Ksenija Taški-Ajduković · Dušica Jovičić
received / primljeno: 01.11.2010. accepted / prihvaćeno: 07.12.2010.© 2011 IFVC
Izvod: Mrkva spada u veoma značajnu povrtarsku biljnu vrstu zbog svog sastava i sadržaja vitamina. Gljive koje pripadaju rodu Alternaria, naročito A. dauci (Kühn) Groves et Skolk, prouzrokovač lisne pegavosti i sušenja lista, i A. radicina Meier, Drechsler et Eddy, prouzrokovač crne truleži korena mrkve, mogu da ugroze celokupnu proizvodnju mrkve. Obe vrste se mogu u visokom procentu naći na seme-nu, ne samo mrkve, već i peršuna i celera. U cilju njihove detekcije preporučuje se metod fi lter papira i metod izolacije na hranljivoj podlozi (ISTA Rules). Ispitivanje prisustva A. dauci i A. radicina vršeno je na 12 uzoraka potencijalno zaraženog semena mrkve poreklom iz Francuske kao dela laboratorijskih kružnih testova. Procenat prisustva A. dauci metodom na fi lter papiru kretao se od 0% do 21%, dok je A. radicina bila prisutna od 1% do 30%. Na hranljivoj podlozi procenat prisustva A. dauci se kretao od 0% do 23% a A. radicina od 1% do 31%.Ključne reči: A. dauci, A. radicina, fi lter papir metod, metod izolacije na hranljivoj podlozi, mrkva, seme
D. Petrović · M. Ignjatov · M. Vujaković · Z. Nikolić · K. Taški-Ajduković · D. JovičićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Mrkva (Daucus carota L.) spada u veoma zna-čajnu povrtarsku biljnu vrstu zbog svog sastava i sadržaja vitamina. Po sadržaju A vitamina, tj. beta karotina – provitamina A, izdvaja se od svih drugih vrsta povrća zauzimajući prvo mesto. Po-vršine na kojima se gaji, kao i ekonomski značaj, u stalnom su porastu u svetu.
Zabeleženo je da postoji više od deset patoge-na koji se prenose semenom mrkve (Richardson 1990). Gljive koje pripadaju rodu Alternaria, na-ročito A. dauci (Kühn) Groves et Skolk, prouzro-kovač lisne pegavosti i sušenja lista, i A. radicina Meier, Drechsler et Eddy, prouzrokovač crne truleži korena mrkve, mogu da ugroze celokupnu proizvodnju mrkve. Obe vrste se mogu u viso-kom procentu naći na semenu, ne samo mrkve, već i peršuna i celera. Gljiva u vidu micelije ili ko-nidija locirana je na površini perikarpa i semenog omotača (Ivanović & Mijatović 2003). Pored se-mena kao primarni izvor inokuluma brojni autori
navode biljne ostatke, korove i samonikle biljke mrkve, a vetar i česte padavine doprinose bržem širenju patogena (Strandberg 1977).
Među prouzrokovačima oboljenja mrkve, A. dauci jedna je od najznačajnijih. Ima izuzetan ekonomski značaj i predstavlja vodeću bolest ove biljne vrste (Pryor & Strandberg 2002). Jav-lja se na listu u vidu brojnih sitnih nekrotičnih pega koje vremenom zahvataju celu lisku, dok je na lisnim drškama uočljiva crtičasta nekroza koja prethodi njihovom sušenju. Crnu trulež mrkve (A. radicina) karakteriše pojava crnih ugnutih pega na korenu mrkve, a čest simptom je i sužavanje vrha korena (Bulajić & Krstić 2007).
Sve do 2004. u našoj zemlji nisu izvođena istra-živanja o vrstama koje parazitiraju gajene biljke iz fam. Apiaceae. Tada je problem koje Alternaria vrste izazivaju skrenuo pažnju, pošto je došlo do jake pojave pegavosti i sušenja listova mrkve i peršuna u nekoliko lokaliteta u Srbiji, a uzročnici su bile vrste iz roda Alternaria (Bulajić 2007).
Obzirom da su A. dauci i A. radicina gljive koje se prenose semenom, veoma je važna tač-na identifi kacija patogena. Pravilna identifi kacija prisustva A. dauci i A. radicina može se obaviti primenom konvencionalnih, ali i savremenih molekularnih metoda detekcije (Konstantinova
Fitopatologija / Phytopathologyw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Petrović D i sar.174
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
et. al. 2002, Pryor & Gilbertson 2002). U cilju identifi kacije A. dauci i A. radicina preporučuje se metod fi lter papira i metod izolacije na hran-ljivoj podlozi (ISTA Rules 2010). Cilj rada je utvrđivanje procentualne zaraženosti semena ispitivanih uzoraka gljivama A. dauci i A. radicina navedenim metodama.
Materijal i metod
Ispitivanje prisustva A. dauci i A. radicina na se-menu mrkve kao i određivanje procenta zaraže-nosti semena vršeno je na 12 uzoraka potencijal-no zaraženog semena mrkve poreklom iz Francu-ske, kao deo laboratorijskih kružnih testova koje organizuje ISTA. Za detekciju gljiva korišćen je fi lter papir metod i metod izolacije na hranljivoj podlozi (ISTA Rules 2010).
Izolacija patogena iz semena mrkve
Filter papir metod (ISTA 7-001a, 7-002a). Za ispitivanje zaraženosti semena mrkve metodom fi lter papira korišćeno je po 100 semena iz svakog uzorka. Set od tri sterilisana fi lter papira prečnika 9 cm natapan je u posudi sa sterilnom vodom i tako natopljen postavljen u Petri posudu. U sva-ku Petri posudu, postavljeno je po deset semena na rastojanju od 20 mm. Inkubacija je vršena na temperaturi od 20ºC u mraku tokom tri dana. Nakon isteka tog vremena posude sa uzorcima su izložene temperaturi od -20ºC u trajanju od 24 h. Potom su premeštene u termostat na 20ºC 6 dana uz smenu svetla (NUV) i mraka svakih 12 h (Gambogi 1987).
Metod hranljive podloge (ISTA 7-001b, 7-002b). Za determinaciju gljive na hranljivoj podlozi preporučuje se Malt agar (MA). Seme je pažljivo isprano u 1% rastvoru NaOCl u trajanju
od 10 minuta, isprano u sterilnoj vodi i osuše-no na sterilnom fi lter papiru. Podloga je nakon sterilizacije i hlađenja do 50ºC razlivena u Petri posude prečnika 9 cm. U svaku Petri posudu po-stavljeno je po deset semena na rastojanju od 20 mm. Inkubacija je obavljena na 20ºC uz smenu svetla (NUV) i mraka svakih 12 sati u trajanju od 10 dana.
Nakon isteka inkubacionog perioda, svako seme je mikroskopski pregledano na uveličanju od 40 x. Determinacija vrste A. dauci i A. radici-na je izvršena na osnovu podataka koje navode Mathur & Kongsdal (2003) i prema ISTA Rules (2010). Na osnovu utvrđenog procenta zaraze dobijen je uvid u zdravstveno stanje ispitivanih uzoraka mrkve.
Rezultati i diskusija
Na osnovu ispitivanja zdravstvenog stanja se-mena mrkve korišćenjem metode na fi lter papi-ru i metode izolacije na hranljivoj podlozi, utvr-đeno je prisustvo gljiva A. dauci i A. radicina. Na svih dvanaest testiranih uzoraka konstatovano je prisustvo bar jednog patogena. Po procentu zaraženosti uzoraka A. radicina je bila zastuplje-nija. Procenat zaraze kretao se od 1% do 30% na fi lter papiru, dok je na hranljivoj podlozi taj procenat iznosio od 1% do 31% (Sl. 1). Na fi l-ter papiru A. dauci je bila zastupljena od 0% do 21%, a na hranljivoj podlozi od 0% do 23% (Sl. 2).
Na fi lter papiru A. dauci obrazovala je tamno-braon do crnu vazdušastu miceliju. Konidiofore su jednostavne ili blago razgranate, pojedinačne ili u malim grupama. Mikroskopskim pregledom konstatovane su pojedinačne konidije, sužene na jednom kraju, sa repom koji je i do tri puta duži od ostatka tela konidije, veličine 100-450 x
468
10121416182022242628303234
% Filter papir metod
Metod hranljive podloge
02468
10121416182022242628303234
1 2 3 4 5 6 7 8 9 10 11 12
%
Šifra uzoraka
Filter papir metodMetod hranljive podloge
Slika 1. Procenat pojave A. radicina u 12 uzoraka semena mrkve utvrđeno metodom fi lter papira i izola-cijom na hranljivu podloguFig 1. The percentage of A. radicina in 12 seed sampes on carrot by blotter method and agar plate method.

Izolacija i detekcija gljiva A. dauci i A. radicina iz semena mrkve 175
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
16-25 μm, što je saglasno sa navodima literatu-re koje daju Mathur & Kongsdal (2003) i ISTA rules (2010). Boja konidija je okeržuta ili svetlo-mrka (Sl. 3a).
Na hranljivoj podlozi kolonije gljive su masli-nastomrke boje koje sa starenjem postaju tamno-mrke (Sl. 3b). Gljiva produkuje mrki pigment u podlozi. Konidije su pojedinačne, a po obliku i
dimenzijama iste kao i konidije konstatovane na fi lter papiru.
Pored konidija gljive A. dauci, mikroskopskim pregledom su uočene i konidiofore, jednostavne ili blago razgranate, u malim grupama. One for-miraju na domaćinu površinsku, sivu vazdušastu miceliju. Konidije su takođe pojedinačne ili u kratkim lancima (dve-tri konidije), elipsoidnog ili
468
101214161820222426
%
Filter papir metodMetod hranljive podloge
02468
101214161820222426
1 2 3 4 5 6 7 8 9 10 11 12
%
Šifra uzorka
Filter papir metodMetod hranljive podloge
Slika 2. Procenat pojave A. dauci u 12 uzoraka semena mrkve metodom fi lter papira i izolacijom na hran-ljivu podloguFig. 2. The percentage of A. dauci in 12 seed sampes on carrot by blotter method and agar plate method
Slika 3a. Konidija gljive A. dauci Fig. 3a. Conidia of A. dauci
Slika 3b. Micelija gljive A. dauci na MA podloziFig. 3b. Mycelium of A. dauci on MA medium
Slika 4a. Konidija gljive A. radicina Fig. 4a. Conidia of A. radicina
Slika 4b. Micelija gljive A. radicina na MA podlozi Fig. 4b. Mycelium of A. radicina on MA medium

Petrović D i sar.176
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
buretastog oblika, maslinasto mrke boje, veličine 27-57 μm x 9-27 μm što nam je na osnovu lite-raturnih podataka koje daju Mathur & Kongsdal (2003) i ISTA rules (2010) ukazivalo na vrstu A. radicina (Sl. 4a).
Na hranljivoj podlozi gljiva formira koncentrič-no raspoređenu koloniju sa bogatom maslinasto sivom micelijom (Sl. 4b). U podlozi se obrazuje plavocrni pigment. Izgled i veličina konidija od-govaraju dimenzijama konidija konstatovanih na fi lter papiru.
Infekcija semena biljakа fam. Apiаceаe sa Alternаriа spp. jedаn je od nаjvаžnijih nаčinа zа opstаnаk i širenje ovih gljivа u prirodi (Kwаsnа 1992, Agrios 2005). Infi cirano seme može dа dovede do smаnjenjа klijаvosti (Tilkovskа 1992) i prinosa i može doprineti prenošenju Alternаriа spp. u nove proizvodne oblаsti (Prior i sаr 1997).
Izolacija i identifi kacija gljiva sa semena ratar-sko povrtarskih biljnih vrsta predstavlja važan segment u proizvodnji semena. Veliki broj pato-gena izolovan je sa semena korišćenjem metoda izolacije na hranljive podloge (Milošević i sar. 2008, Petrović i sar. 2010, Ignjatov i sar. 2010). Metod fi lter papira je najčešće korišćen metod u identifi kaciji gljiva roda Alternaria. Tako su autori Wan & Suaresh (2006) koristili ovaj metod za de-tekciju Alternata spp. U tim istraživanjima od pet testiranih uzoraka semena mrkve A. alternata i A. radicina su detektovane na svim uzorcima u viso-kom procentu od 25,8% do 70,5% i od 37,5% do 63,5% dok je A. dauci detektovana na četiri uzor-ka sa nivoom zaraze od 0,5% do 7,7%. Bulajić i sar. (2009) su u Srbiji primenom metode zamrza-
vanja na fi lter papiru detektovali prisustvo četiri fi topatogene vrste iz roda Alternaria: A. dauci, A. radicina, A. petroselini i A. alternata koje su izazvale smanjeno nicanje mrkve, peršuna, paštrnaka i ce-lera. Postoji još nekoliko izveštaja o visokoj infek-ciji semena mrkve sa A. radicina čije su se vredno-sti kretale od 0,5% do 82% (Tylkowska 1992); od 0,5% do 7,5% (Wan & Suaresh 2006) i od 0,25% do 30% (Maude 1966). Infekcija sa A. radicina bila je detektovana čak i u semenu mrkve tretirane sa iprodionom od 37,5% do 63,5% (Wan & Sua-resh 2006). Infekcija semena mrkve gljivom A. dauci kreće se od 0,3% do 8% (Tylkowska 1992) i 13,2% (Netzer & Kenneth 1969). Takođe je u ne-kim slučajevima konstatovana i mešana infekcija A. radicina i A. dauci (Strandberg 2002). Ovako visok procent zaraze sa A. dauci može dovesti do jake epidemije koje mogu smanjiti prinos mrkve od 40% do 60% (Farrar et al. 2004). A. radicina ta-kođe ima izuzetan ekonomski značaj, pa su tako u Australiji zabeleženi gubici prinosa čak do 88% (Pryor 2002).
Zaključci
S obzirom da je glavni izvor inokuluma za na-rednu sezonu zaraženo seme, potrebno je koristi-ti zdravo seme za setvu. Stoga je potrebno vršiti ispitivanja zdravstvenog stanja semena mrkve i utvrđivanje prisustva ova dva patogena. Na osno-vu razvoja micelije i kolonije izolovanih gljiva možemo zaključiti da su metoda fi lter papira i metoda izolacije na hranljivu podlogu podjedna-ko efi kasne u cilju identifi kacije gljiva A. dauci i A. radicina.

Izolacija i detekcija gljiva A. dauci i A. radicina iz semena mrkve 177
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
Literatura
Agrios G N (2005): Plant Pathology, Elsevier Academic Press, Burlington, San Diego, London 4-922
Bulajić A (2007): Identifi kacija i taksonomski međuodnos vrsta roda Alternaria Ness patogenih za povrtarske i začinske biljke familije Apiaceae u Srbiji. Doktorska disertacija, Poljoprivred-ni fakultet, Beograd
Bulajić A, Krstić B (2007): Patogeni mrkve (Alternaria radicina, A. dauci). Biljni lekar 6: 595-604
Bulajić A, Đekić I, Lakić N, Krstić B (2009): The presence of Alternaria spp. on the seed of apiaceae plants and their infl u-ence on seed emergence. Arch. Biol. Sci. 61: 871-881
Farrar J, Pryor B A, Davis R M (2004): Alternaria diseases of carrot. Plant Dis. 88: 776-784
Gambogi P (1987): Carrot. Leaf blight, damping off. Daucus carota, Alternaria dauci (Kühn) Groves & Skolko. ISTA Han-dbook on Seed Health Testing. 2nd Edition, Working Sheet 4: 1-5
Ignjatov M, Gašić K, Ivanović M, Šević M, Obradović A, Milo-šević M (2010): Characterization of Xanthomonas euvesicatoria strains pathogens of pepper in Serbia. Pesticidi i fi tomedici-na 25: 139-149
International Rules for Seed Testing (2010): Annexe to Chapter 7 Seed Health Testing Methods
Ivanović M, Mijatović M (2003): Patogene gljive semena povr-ća. Biljni lekar 6: 595-603
Konstantinova P, Bonants P J M, van Gent-Pelzer M P E, van der Zouwen P, van den Bulk R (2002): Development of spe-cifi c primers for detection and identifi cation of Alternaria spp. in carrot material by PCR and comparison with blotter and plating assays. Mycological Res. 106: 23-33
Kwasna H (1992): Occurrence of Alternaria species in Poland, In: J. Chelkowski and A. Visconti (eds.), Alternaria Biology, Plant Diseases, and Metabolites, Topics in Secondary Me-tabolism, 3. Elsevier Science Publishers B. V. Amsterdam, 301-336
Mathurm S B and Kongsdal O (2003): Common Laboratory Seed Health Testing Methods for Detecting Fungi. ISTA, Zürich, Switzerland
Maude R B (1966): Studies on etiology of black rot, Stemphilium radicinum (Meier, Drechsl. and Eddy) Neerg., and leaf blight, Alternaria dauci (Kuhn) Groves et Skolko, on car rot crops; and on fungicide control of their seed-borne infection pha-ses. Ann. Appl. Biol. 57: 83-93
Milošević M, Ignjatov M, Vujaković M, Petrović D, Nikolić Z, Dokić V (2008): Svojstva gljiva roda Fusarium sp.i moguć-nost dugoročnog čuvanja metodom liofi lizacije. Arhiv za poljoprivredne nauke 69: 89-95
Netzer D, Kenneth R G (1969): Persistence and transmis sion of Alternaria dauci (Kuhn) Groves et Skolko in the semi-arid conditions of Israel. Ann. Appl. Biol. 63: 289-294
Petrović D, Ignjatov M, Vujaković M, Taški-Ajduković K, Ni-kolić Z, Milošević M, Jovičić D (2010): Mikopopulacija se-mena kukuruza (2006-2008). Ratar. Povrt. / Field Veg. Crop Res. 47: 561-566
Pryor B M, Davis R M, Gilbertson R L (1997): Detection of soil-borne A. radicina and its occurrence in California carrot fi elds. Plant Dis. 82: 891-895
Pryor B M, Gilbertson R L (2002): Relationships and taxono-mic status of Alternaria radicma, A. carotiincultae and A. petro-selini based upon morphological, biochemical and molecular characteristics. Mycologia 94: 49-61
Pryor B M and Strandberg J O (2002): Alternaria leaf blight of carrot, In: R. M. Davis and R. N. Raid (eds.), Compendium of Umbelliferous Crop Diseases The American Phytopat-hological Society, St. Paul, Minnesota, 15-16
Richardson M J (1990): An Annotated List of Seed-Borne Di-seases. Fourth edition. International Seed Testing Associati-on, Zurich, Switzerland
Strandberg J O (1977): Spore production and dispersal of Alter-naria dauci. Phytopathol. 67: 1262-1266
Strandberg J O (2002): A selective medium for the detection of Alternaria dauci and Alternaria radicina. Phytoparasitica 30: 269
Tylkowska K (1992): Carrot seed-borne diseases caused by Alternaria species. In: J. Chelkowski and A. Visconti (eds.). Alternaria Biology, Plant Diseases, and Metabolites, Topics in Secondary Metabolism, 3. Elsevier Science Publishers B. V. Amsterdam, 337-352
Wan G K, Suaresh B M (2006): Detection of Alternaria spp. in carrot seeds and effect of the fungy on seed germination and seedling growth of carrot. Plant Pathol. J. 22: 11-15

Petrović D i sar.178
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 173-178
Isolation and Detection of the Fungi Alternaria dauci (Kühn) Groves et Skolk and Alternaria radicina Meier, Drechsler et Eddy from Carrot Seeds
Dragana Petrović · Maja Ignjatov · Milka Vujaković · Zorica Nikolić · Ksenija Taški-Ajduković · Dušica Jovičić
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Carrot is among the most important vegetable crops because of its composition and vitamin content. The fungi belonging to genus Alternaria, especially A. dauci (Kühn) Groves et Skolk, causal agent of leaf spot and blight of carrots, and A. radicina Meier, Drechsler et Eddy, the agent of black root rot, can jeopardize the entire production of carrots. Both types can be found in a high percentage of seeds, carrots, parsley and celery. The method of detection fi lter paper (blotter method) and the method of isolation on malt agar were recommended by ISTA Rules. The presence of A. dauci (causal agent of leaf spot and blight of carrots) and A. radicina was per-formed on 12 samples of potentially infected carrot seeds originating from France, as part of laboratory ring tests. The presence of A. dauci and A. radicina using the fi lter paper method in 12 tested samples ranged from 0% to 21% and from 1% to 30% respectively. On MA medium the respective percentages ranged from 0% to 23% and from 1% to 31%.Key words: A. dauci, A. radicina, agar plate method, blotter method, carrot, seed

Detekcija Ustilago nuda (Jensen) Rostrup u semenu ozimog ječma 179
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 179-182
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 179-182originalni naučni rad / original research article
Detekcija Ustilago nuda (Jensen) Rostrupu semenu ozimog ječma
Maja Ignjatov · Dragana Petrović · Milka Vujaković · Ksenija Taški-Ajduković ·Zorica Nikolić · Dušica Jovičić
received / primljeno: 01.11.2010. accepted / prihvaćeno: 07.12.2010.© 2011 IFVC
Izvod: Ječam spada u najvažnija strna žita koja se gaje u našim agroekološkim uslovima. Prouzrokovač otkrivene gari ječma Ustilago nuda (Jensen) Rostrup javlja se često, jer su u proizvodnji zastupljene sorte osetljive prema ovom patogenu. Simptomi bolesti se manifestuju na klasu. Parazit se prenosi semenom, a održava se u embrionu zaraženog semena u vidu mirujuće micelije. Prema preporuci International Seed Testing Association (ISTA) za detekciju U. nuda korišćen je metod brojanja zaraženih embriona. U ispitivanjima je korišteno 9 uzoraka prirodno zaraženog semena ječma (oko 1000 semena u zavisnosti od apsolutne mase semena). Posmatranje, ocena i fotografi sanje zaraženih embriona obavljeno je pomoću Zeiss mikroskopa pri uveličanju od x 40. Konstatovana je micelija gljive debljine oko 3 μ, zlatno-mrke boje koja je vidljiva bez bojenja. Procenat zaraženosti embriona ispitivanih uzoraka semena ječma kretao se od 0,8% do 5,2%. Ključne reči: ozimi ječam, seme, Ustilago nuda
M. Ignjatov · D. Petrović · M. Vujaković · K. Taški-Ajduković ·Z. Nikolić · D. JovičićInstitut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Pored pšenice, ječam spada u najvažnija strna žita koja se gaje u našim agroekološkim uslovima. Značaj ječma proizilazi iz njegove raznovrsne upo-trebe u ljudskoj i stočnoj ishrani. Ječam ima veliki značaj u proizvodnji i industriji piva i slada, a tako-đe se koristi u tekstilnoj i farmaceutskoj industriji, u poslastičarstvu, pekarstvu, u proizvodnji sladnog mleka, alkohola, kvasca i dr. Jedan od patogena koji može u znatnoj meri uticati na prinos i kvalitet ječ-ma je Ustilago nuda (Jensen) Rostrup prouzrokovač otkrivene gari ječma. U. nuda se pojavljuje svake godine, a razlog tome je zastupljenost sorata ječ-ma osetljivih prema ovom patogenu u proizvodnji (Jevtić i sar. 2003). Areal rasprostranjenja U. nuda je veoma širok. Sreće se u Evropi (Austrija, Rusija, Italija, Rumunija, Švajcarska, Ukrajina i dr.), Aziji (Kina, Indija, Turska, Iran), Severnoj Americi (Ka-nada), Južnoj Americi (Argentina) i Africi (Južna Afrika) (CABI 2004). U Bugarskoj je ovaj patogen uzrokovao velike štete (1970-1980) kada se na ne-kim parcelama intenzitet zaraze kretao od 15% do 20% (Nakov et al. 1994). Kod nas se otkrivena gar ječma često javlja na parcelama privatnih proizvo-
đača koji ne koriste deklarisano seme, ali podaci o velikim štetama nisu zabeleženi (Stojanović 2004). Veliki značaj U. nuda može da ima u proizvodnji certifi kovanog semena ječma viših kategorija.
Parazit se održava u kotiledonu zaraženog se-mena u vidu mirujuće micelije. Proces infekcije semena započinje teleutosporama sistemično zaraženih biljaka ječma koje se šire i infi ciraju zdrave biljke. Teleutospore klijaju u promiceliju, a infektivna hifa prodire kroz zidove ovariju-ma, zatim kroz nukleus i aleurone stiže do me-ristema embriona pretvarajući se u dormantnu miceliju koja će u narednoj godini pratiti tačku porasta biljke. Simptomi otkrivene gari se uoča-vaju na klasu koji je potpuno preobraćen u crnu masu teleutospora. Oboleli klasovi se pojavljuju istovremeno sa zdravim, teleutospore se šire na zdrave biljke, infi ciraju ih i ciklus se ponavlja. U zaraženom klasu formiraju se teleutospore koje se vetrom raznose na zdrave biljke (Ivanović 1992). Neergard (1977) navodi da je optimalni momenat za infekciju u vreme oprašivanja, kao i jedan do dva dana nakon oprašivanja biljaka. Vetrovito vreme je takođe pogodno za širenje teleutospora u toku vegetacije (Tusa et al. 1988). Seme tretirano fungicidima daje manji procenat zaraženih biljaka (1%) u odnosu na netretirano seme koje daje i do 7% zaraženih biljaka u polju, koje predstavljaju primarni inokulum za infekci-ju (Reeves et al. 1994).
Fitopatologija / Phytopathologyw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Ignjatov M i sar.180
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 179-182
S obzirom da se Ustilago nuda prenosi semenom i da simptomi na semenu nisu vidljivi uobičaje-nim metodama, za ispitivanja zdravstvenog stanja semena radi identifi kacije ovog patogena prepo-ručuje se metod brojanja zaraženih embriona (ISTA Rules 2008). Cilj rada je bio utvrđivanje procentualne zaraženosti semena ispitivanih uzo-raka gljivom U. nuda navedenom metodom.
Materijal i metod
Za detekciju U. nuda korišćen je metod broja-nja zaraženih embriona bez prethodnog bojenja (ISTA Rules method 7-013). U ispitivanjima je korišćeno 9 uzoraka prirodno zaraženog semena ječma (oko 1000 semena u zavisnosti od apsolutne mase semena), dobijenih od ISTA-e (International Seed Testing Asociation) u okviru kružnog testa akre-ditovanih laboratorija. Ispitivani uzorci obeleženi su brojevima od 1 do 9. Postupak ocene zaraže-nosti embriona sastoji se u prethodnom odvajanju embriona od ostatka semena pomoću 5% rastvo-ra NaOH. Svaki uzorak, ponaosob potapan je u jedan litar sveže pripremljenog rastvora NaOH. Inkubacija potopljenih uzoraka izvršena je na tem-peraturi od 20ºC tokom 24 časa. Uzorci su nakon inkubacije prebačeni u odgovarajuću posudu i ispi-rani toplom vodom radi razdvajanja embriona od perikarpa semena. Sakupljanje odvojenih embri-ona izvršeno je pomoću sita promera perforacija od 1 mm. Radi dodatnog prečišćavanja i odvajanja embriona od perikarpa semena, embrioni su pre-bačeni u prethodno pripremljen rastvor glicerola i vode u odnosu 1:1. Nakon toga izvršeno je zagre-vanje prečišćenih embriona u rastvoru bezvodnog
laktofenola do tačke ključanja, u trajanju od 30 se-kundi. Nakon ključanja, radi lakšeg mikroskopira-nja embrioni su potapani u svež rastvor glicerola. Nakon toga su pincetom pojedinačno postavljani na mikroskopsku pločicu i mikroskopirani. Po-smatranje, brojanje i fotografi sanje zaraženih em-briona obavljeno je pomoću Zeiss mikroskopa pri uveličanju od x 40. Iz svakog uzorka pregledano je po 1000 embriona na prisustvo micelije gljive U. nuda a rezultati su izraženi u procentima.
Rezultati i diskusija
Na osnovu ispitivanja i posmatranja embriona proučavanih uzoraka semena ječma ustanovljeno je prisustvo gljive Ustilago nuda u svim ispitivanim uzorcima. Zapaženo je da gljiva formira karakteri-stičnu razgranatu miceliju zlatno-mrke boje. Mice-lija je septirana, a oblik, veličina i boja micelije gljive najuočljivije su u skutelumu zaraženog embriona (Sl. 1). Nezaražen skutelum je potpuno proziran bez prisustva razgranate micelije (Sl. 2). Procenat infekcije ispitivanih uzoraka semena ječma kretao se od 0,8% do 5,2% (Sl. 3). Najveći procenat za-raze od 5,2% zabeležen je u uzorku pod brojem 3, dok je najniži procenat infekcije od 0,8% utvrđen u prvom uzorku. S obzirom da se procenat zara-ze ocenjuje na osnovu ukupnog broja zaraženih embriona bez obzira na količinu prisutne micelije u embrionu, zapaženo je da prožetost skuteluma hifama i micelijom nije bio ujednačen. Pored pro-centa infekcije embriona, prisustvo micelije gljive u skutelumu i intenzitet infekcije ocenjen je kao slab (prisustvo retkih, kratkih hifa gljive), srednji (pro-žetost skuteluma micelijom do 1/3) i jak ukoliko je
Slika 1. Izgled micelije u skutelumu infi ciranog embriona gljivom Ustilago nuda Fig. 1. Infected embryo, smut mycelium of Ustilago nuda in scutellum

Detekcija Ustilago nuda (Jensen) Rostrup u semenu ozimog ječma 181
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 179-182
prožetost skuteluma bila veća od 1/3 do celog sku-teluma. Rezultati ocene u uzorcima 2 i 4 ukazuju da je prožetost skuteluma bila slaba sa prisustvom retkih, kratkih hifa. Kod uzoraka pod brojevima 1, 3, 5 i 9 prisustvo micelije u skutelumu bilo je oce-njeno kao srednje, dok je u uzorcima pod brojevi-ma 6, 7 i 8 intenzitet zaraze micelijom gljive U. nuda bio najveći uz jaku prožetost većeg dela skuteluma. Dobijeni rezultati o procentualnoj prirodnoj zara-ženosti semena gljivom U. nuda su u saglasnosti sa podacima o prosečnoj zaraženosti semena u pri-rodnim uslovima tokom vegetacije ozimog ječma koju navode Girsch & Weinhappel (1996). Ispiti-vanjem semena ječma u Velikoj Britaniji korišće-njem metode brojanja zaraženih embriona, autori Cockerell & Rennie (1996) su ustanovili da je 25% ispitivanih uzoraka ozimog ječma bilo infi cirano
gljivom U. nuda u procentu većem od 0,5 dok su Jevtić i sar. (1997) korišćenjem iste metode utvrdili da je 10% semena ječma bilo zaraženo patogenom U. nuda kod nas. Jones (1999) je primenom ove metode u svojim istraživanjima detektovao 24,2% semena ozime sorte ječma (Panda) prirodno zara-ženog gljivom U. nuda. Hewett & Damgaci (1986) su metodom brojanja embriona detektovali veoma nizak procenat zaraze od 0,1% i ocenili ovaj metod kao pouzdan, dovoljno osetljiv i pogodan za rutin-ske analize radi detekcije U. nuda. Postoje podaci o korišćenju seroloških testova (ELISA) i molekular-nih metoda (PCR) u detekciji ove gljive. Međutim, Eibel et al. (2005) su u svojim istraživanjima zaklju-čili da je ELISA test manje pouzdan i osetljiv me-tod za rutinske analize identifi kacije ovog patogena u odnosu na metod brojanja embriona.
0 8
1,5
5,2
2,83,3
1,11,5
2,3
4,1
2
3
4
5
6%
0,8
1,5
5,2
2,83,3
1,11,5
2,3
4,1
0
1
2
3
4
5
6
1 2 3 4 5 6 7 8 9
%
Broj uzorka
Slika 2. Izgled zdravog embriona semena ječmaFig. 2. Healthy barly seed embryo (non-infected)
Slika 3. Procenat infekcije embriona ispitivanih uzoraka semena ječma Fig. 3. Percentage of infected embryos investigated in samples of barley seed

Ignjatov M i sar.182
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 179-182
Zaključci
Na osnovu dobijenih rezultata ispitivanih uzo-raka konstatovano je pristustvo gljive U. nuda u svim uzorcima metodom brojanja embriona. Procenat zaraze ispitivanih uzoraka kretao se od 0,8% do 5,2% uz neujednačen intenzitet infek-cije skuteluma. Slabu infekciju karakteriše prisu-stvo retkih, kratkih hifa dok je kod nekih uzo-raka konstatovana potpuna prožetost skuteluma hifama gljive U. nuda. S obzirom na širok areal rasprostranjenja i mogućnost širenja novih rasa patogena semenom, ovaj metod detekcije U. nuda u semenu ječma pruža zadovoljavajuću pouzda-nost.
Literatura
CABI (2004): Crop protection compendium CAB Internatio-nal, CD-ROM
Cockerell V, Rennie W (1996): Survey of seed-borne pathogens in certifi ed and farm-saved cereal seed in Britain between 1992 and 1994. HGCA Project Rep. 124: 40
Eibel P, Wolf G, Koch E (2005): Development and evaluation of an enzyme-linked immunosorbent assay (ELISA) for the detection of loose smut of barley (Ustilago nuda). Europ. J. Plant Pathol. 111: 113-124
Girsch L, Weinhappel M (1996): Health testing of seed - basis for determining a targeted seed treatment. Förderungsdienst 44:416-417
Hewett P, Damgaci E (1986): New procedure to detect a low incidence of Ustilago nuda in seed barley. Plant Pathol. 35: 377-379
ISTA Rules (2008): Handbook on Seed Health.. Zurich, Swit-zerland: International Seed Testing Association
Ivanović M (1992): Mikoze biljaka. Nauka, Beograd Jevtić R, Stojanović S, Dopuđa M, Matijević D, Milošević M
(1997): Glavnica i gari ponovo aktuelne bolesti strnih žita u Srbiji. Rezime radova sa XIX Seminara iz zaštite bilja, 13-14, Beograd
Jevtić R, Marić A, Stojanović S (2003): Paraziti semena strnih žita. Biljni lekar 6: 557-564
Jones P (1999): Control of loose smut (Ustilago nuda and U. tritici) infections in barley and wheat by foliar applications of systemic fungicides. Europ. J. Plant Pathol. 105: 729-732
Nakov B, Karov S, Popov A, Nešev G (1994): Specijalna fi topa-tologija. Akademika Press, Plovdiv
Neergaard P (1977): Seed Pathology. Volume 1. London, UK: Macmillan Press Ltd., 839
Reeves J C, Wray M W, Martin T (1994): Seed treatment: pro-gress and prospects: Proceedings of a Symposium held at the University of Kent, Canterbury, 5-7 January Monograp-hy 57: 37-46
Stojanović S (2004): Poljoprivredna fi topatologija. Srpsko bio-loško društvo „Stevan Jakovljević“, Kragujevac
Tusa C, Miclaus D, Damian V, Constantin L, Paulian F (1988): Capacity for spread of Ustilago nuda (Jens.) Rostr. on some winter barley cultivars. Probleme de Protectia Plantelor, 16: 175-193
Detection of Ustilago nuda (Jensen) Rostrup in Winter Barley Seed
Maja Ignjatov · Dragana Petrović · Milka Vujaković · Ksenija Taški-Ajduković ·Zorica Nikolić · Dušica Jovičić
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Barley is one of the most important cereals grown in our agroecological conditions. The causal agent of loose smut on barley Ustilago nuda (Jensen) Rostrup occurs frequently as varieties susceptible to this pathogen are present in the production. Disease symptoms are manifested on barley head (spike). Parasite is transmitted by seed (seedborne) and stays in the embryo tissue of the infected kernel as dormant mycelium. Recommended method for detection of U. nuda is given by ISTA Rules (method 7-013). In tests, nine samples (weighing 120 g each) of naturally infected barley seed (about 1000 seeds, depending on the absolute mass of seed) were examined, observed and described using a Zeiss microscope with sub stage illumination with magnifi cation range x 40 or higher. Myce-lium of the fungus approximately 3 μ thick, golden brown in colour was detected and visible without a stain. The percentage of infected embryos in the examined samples of barley seeds ranged from 0.8% to 5.2%. Key words: seed, Ustilago nuda, winter barley

Pea and Spring Cereals Intercropping 183
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188original research article / originalni naučni rad
Effect of Pea and Spring Cereals Intercropping on Grain Yield and Crude Protein Content
Žydrė Kadžiulienė · Lina Šarūnaitė · Irena Deveikytė
received / primljeno: 31.05.2010. accepted / prihvaćeno: 08.10.2010.© 2011 IFVC
Summary: The advantage of intercropping is benefi cial biological interactions between crops and their different use of growth resources. Our experiment aimed at investigating the effects of pea intercrop-ping with cereals on the improvement of cereal supply with nitrogen and protein content in grain on a loamy soil in organic farming conditions. It was carried out during 2007-2009 at the Lithuanian Institute of Agriculture in Dotnuva. Pea (Pisum sativum L.) and spring wheat (Triticum aestivum L.), spring barley (Hordeum vulgare L.), oats (Avena sativa L.), and triticale (x Triticosecale Wittm.) were sown as intercrops 50:50 or as a sole crop. The results obtained during the experimental years showed that the productivity and quality of spring cereal sole crops or intercrops depended on the species of cereals and varied between different year’s cultivation conditions. Grain productivity in some cases was estimated to be higher in the intercrops, however it was not stable in three experimental years. The protein content and yield in the grain of cereals grown in intercrops was positively affected by pea intercrops. Key words: intercrop, pea, protein, spring cereal, yield
Ž. Kadžiulienė · L. Šarūnaitė · I. DeveikytėInstitute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, Instituto al. 1, Akademija, Kedainiai LT-58344, Lithu-ania e-mail: [email protected]
Introduction
The inclusion of legumes in crop rotations
and intercrops can provide increased protein-rich yields and a more sustainable source of nitrogen, while on the other hand it saves cost by reducing the requirement for mineral nitrogen application (Anil et al. 1998, Crews & Peoples 2004). In many countries, legume plants have recently been receiving increasing attention from researchers and growers (Hauggaard-Nielsen et al. 2009, Mariotti et al. 2009, Stoddard et al. 2009, Mikić et al. 2010). Pea (Pisum sativum L.) is the most common legume in crop rotations and quite productive in temperate conditions, as well as in Lithuania (Auskalnis 2001).
Being one of the most popular plants in the organic cropping system, pea provides high quality forage, rich in crude protein and mineral elements (Corre-Hellou & Crozat 2005, Lauk & Lauk 2008). However, it is not less important for its ability to accumulate atmospheric nitrogen, on which depends the state of the whole crop rotation and agroecosystem, and
possible farming benefi t. Pea is productive as main crops, but its vines could also be useful as a nitrogen source (Tripolskaja et al. 2008). Various soils are suitable for pea cultivation, but the crop yields best on moderately heavy loam (Dovydaitis 1992). However, due to its biological characteristics, pea cultivation is not so simple. Although of more recent cultivars, pea grown as a sole crop has some disadvantages – it is prone to severe lodging, which aggravates its harvesting and weed control. As a result, pea productivity is more dependent on proper soil and crop management (Auškalnis & Dovydaitis 1998, Auškalnis & Dovydaitis 1999, Maikštėnienė & Satkus 2000, Auškalnis 2001). When grown in a mixture, e.g. with spring barley (Hordeum vulgare L.), the crop does not lodge and in the conventional cropping system produces a grain yield of nearly 5 t ha-1 (Auškalnis & Dovydaitis 2000). Intercropping of cereals and legumes is widely used in low-input agriculture because the mixture of nitrogen N-fi xing and non-N-fi xing crop species provides complementarities in the utilization of resources, reduces weed pressure and sustains plant health (Hauggaard-Nielsen et al. 2003, Jensen et al. 2006). Mixing species in cropping systems may lead to a range of benefi ts that are expressed on various space and time scales, from a short-term increase in crop yield and quality, to long-term agroecosystem
Intercropping / Združeni useviw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Kadžiulienė Ž et al.184
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
sustainability, up to societal and ecological benefi ts (Malezieux et al. 2009).
Our experiment aimed at investigating the effects of pea intercropping with cereals on the improvement of cereal supply with nitrogen and protein content in grain on a loamy soil in organic farming conditions.
Materials and Methods
The fi eld experiment was carried out on a loamy Endocalcari-Epihypogleyic Cambisol during 2007-2009 at the Lithuanian Institute of Agriculture in Dotnuva (55o24’N, 23o50 E). The soil pH varied at about 7.5, humus content was 2.3%, available P 74-79 mg kg-1 and K 135-140 mg kg-1. Wheat (Triticum aestivum L.), barley, oats (Avena sativa L.) and triticale (x Triticosecale Wittm.) were grown as sole crops and intercrops
with fi eld pea. The experimental plots were laid out in a complete one-factor randomised block design in three replicates. The intercrop design was based on the proportional replacement principle, with mixed pea grain and spring cereals grain at the same depth in the same rows at relative frequencies of 50:50. Wheat seeds rates were calculated on the basis of 5.5, barley 4.7, oat 6.0, triticale 4.5 and pea 1.0 million seeds ha-1 for sole crop. The crops were cultivated according to organic management practices.
The crops were harvested at complete maturity stage. Grain yield components were determined from plant samples taken before harvest. Grain dry matter (DM) yield was determined after threshing. Nitrogen was determined by Kjeldahl, while crude protein content was
calculated according to the amount of nitrogen, by multiplying with 6.25 for pea and 5.7 for spring cereals. The LER values were calculated as follows (Mead & Willey 1980):
Lc = cerealSC
cerealIC
YY
; Lp =peaSC
peaIC
YY
and LER = Lc+Lp. Ycereal IC and YpeaIC are the intercrop yields and YcerealSC and YpeaSC are the sole crop yields of cereal and pea respectively. A LER value above one indicates a benefi t of intercropping.
The experimental data were processed by analysis of variance (ANOVA) applying Least Signifi cant Difference (LSD) test using the computer software Selekcija (Tarakanovas & Raudonius 2003).
The precipitation sums and the temperature means during experimental period are shown in table 1.
Results and Discussion
The number of the productive stems was signifi cantly lower in the intercrops than in sole peas or cereals (Tab. 2) in all three years of experiment and naturally fl uctuated between years. Number of pea grain in intercrops was also lower, therefore number of cereals grain per ear was higher in intercrops and number of wheat and triticale was signifi cantly higher. In previous research on pea productivity on light loam soil, it was noticed that a stand density of 80-90 plants m-2 was suffi cient to produce a rather high yield (Auškalnis & Dovydaitis 1998).
monthmesec
mean air temperature / srednja temperatura vazduha (oC)
1924- 2009
precipitation per month / mesečna suma padavina (mm)
1924 -2009
2007 2008 2009 2007 2008 2009 January 1.0 0.5 -2.8 -4.8 88.3 68.8 41.0 30.2February -7.4 2.2 -3.5 -4.5 25.7 31.1 18.7 25.3March 5.2 2.6 0.9 -0.8 23.6 53.4 53.9 28.5April 6.9 8.8 8.8 5.8 15.8 38.7 13.1 36.9May 13.5 12.2 12.7 12.2 98.2 13.2 26.7 51.8June 17.6 16.1 14.6 15.6 61.5 49.2 168.6 62.4July 17.2 18.2 18.1 17.6 118.1 47.6 90.0 73.4August 18.7 18.0 16.8 16.7 50.8 90.8 67.1 73.7September 12.8 12.1 13.9 12.0 49.1 16.0 48.2 51.3October 7.7 8.9 5.2 6.8 48.7 80.4 95.4 50.2
Table 1. Temperature and precipitation during the winter and growing periodTabela 1. Temperatura i padavine tokom zime i vegetacionog perioda
data by Dotnuva meteorology station / podaci meteorološke stanice Dotnuva

Pea and Spring Cereals Intercropping 185
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
The results obtained during the tree experimental years showed that the productivity of spring cereal sole crops or cereal and pea intercrops depended on the species of cereals and cultivation conditions (Tab. 3).
The productivity of spring wheat and barley intercrops in the 2007 was signifi cantly higher compared with that of the other intercrops and cereal sole crops, except for spring wheat (Tab. 3). The fact that intercropping of legumes and cereals produced higher yields than sole cereal crops without nitrogen fertilization was noticed
by several researchers (Jensen 1996, Lauk & Lauk 2005). The cultivation conditions were different in experimental years, therefore the results varied. In the year 2008, yields of cereal intercrops were lower than in the 2007, because the climatic conditions were droughty in the summer of 2008. Oat and pea intercrop grain yield was the highest and the productivity of barley intercrop was insignifi cantly lower compared with the other intercrops. In 2009, yields of cereal intercrops were lower than in the 2007, however higher than in 2008. In 2009, the intercrops were more
treatmentvarijanta
number of productive stems broj produktivnih stabala (m-2)
grain number per ear/legume stem broj zrna po klasu/stablu (m-2)
thousand grain weight masa 1000 zrna(g)
2007 2008 2009 2007 2008 2009 2007 2008 2009pea + wheat
grašak + pšenicapeagrašak 48 50 46 10 9 7 219 221 196wheatpšenica 367 227 300 39 42 40 39 40 28
pea + barleygrašak + ječam
peagrašak 38 50 32 8 9 6 111 108 213barleyječam 305 215 357 23 19 24 52 48 44
pea + oatgrašak + ovas
peagrašak 57 64 52 9 11 5 199 215 195oat 311 198 266 48 48 60 28 36 33
pea + triticalegrašak + tritikale
peagrašak 46 42 56 13 8 8 227 219 223triticaletritikale 279 197 239 44 44 49 38 40 40
sole crops / pojedinačni usevipeagrašak 126 99 102 20 13 15 239 220 233wheatpšenica 584 327 522 33 36 37 36 38 31barleyječam 462 338 395 21 21 25 45 50 43oatovas 499 373 454 44 40 46 28 39 34triticaletritikale 415 307 382 38 38 46 36 37 37
LSD0.05peagrašak 10.6 9.86 11.8 1.74 2.52 2.05 0.59 1.92 20.4spring cerealjara strna žita 42.2 43.7 63.3 4.51 4.65 3.24 1.60 0.19 0.26
Table 2. The grain yield components in pea and spring cereals crop structureTabela 2. Komponente prinosa zrna u strukturi useva stočnog graška i jarih strnih žita

Kadžiulienė Ž et al.186
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
productive than sole cereal crops, except pea and oat intercrop and the most productive was pea and triticale intercrop. Some researchers noted that the yield of the intercrops was more stable than that of sole crops (Corre-Hellon et al. 2006). In our experiment, pea and barley intercrop in two years out of three produced higher yield compared with the sole barley crop, however we could not conclude that this intercrop yielded more stably.
Even in organic agriculture it is diffi cult to obtain comparative values for protein contents of different cereals. Analysis of data showed
that crude protein concentration of the intercropped cereals was improved compared to sole cereal crops in all experimental years, but was estimated lower than in sole pea (Tab. 3). Differences in protein content in different cereals were recorded. The highest positive infl uence was achieved for intercropped triticale grain. In all three experimental years, protein concentration was signifi cantly higher than that in sole triticale. The lowest infl uence was observed in intercropped oat and the protein concentration was insignifi cantly higher than that in sole oat.
treatmentvarijanta
grain yieldprinos zrna(kg ha-1)
crude protein contentsadržaj sirovih proteina(% )
crude protein yieldprinos sirovi proteini(kg ha-1)
2007 2008 2009 2007 2008 2009 2007 2008 2009peas+wheatgrašak+pšenica 4491 2139 2225 16.1 14.5 14.3 689 311 319
peasgrašak 722 559 369 22.9 23.1 23.7 165 130 88wheat
pšenica 3769 1580 1854 13.9 11.6 12.4 524 181 231pea+ barleygrašak+ječam 3951 1851 2448 16.7 15.7 12.3 611 291 302
peasgrašak 649 783 265 23.1 22.4 23.6 150 175 63barleyječam 3302 1068 2183 14.0 10.8 11.0 461 116 239
pea + oatgrašak+ ovas 3010 2379 2277 15.2 13.8 11.9 423 332 271
peasgrašak 634 514 188 23.4 23.6 22.8 150 121 43
oatovas 2376 1865 2089 11.5 11.4 10.9 275 211 228
pea+triticalegrašak+tritikale 3567 2089 2549 18.1 13.2 14.6 545 275 374
peasgrašak 838 414 310 23.0 22.9 23.3 193 96 72
triticaletritikale 2729 1675 2239 12.9 10.9 13.5 352 179 302
sole crops / pojedinačni usevipeagrašak 4232 2342 2236 22.7 23.3 23.6 960 549 530wheatpšenica 4650 2210 2149 11.3 10.1 11.6 527 247 249barleyječam 3304 2114 2332 12.8 9.50 10.4 421 181 242oatovas 3241 2924 2526 11.1 10.6 10.0 359 268 253triticaletritikale 3773 2341 2039 11.6 9.80 12.0 439 257 245
*LSD0.05 373.7 565.8 275.4 1.04 0.94 0.96 61.5 125.0 70.6
Table 3. Grain yield and crude protein content and yield in intercrops and in sole pea or spring cerealsTabela 3. Prinos zrna i sadržaj i prinos sirovih proteina u združenom i čistom usevu stočnog graška i jarih strnih žita
*for total yield / za ukupni prinos

Pea and Spring Cereals Intercropping 187
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
The yield of crude protein in the intercrops varied between experimental years. Although the protein yield depended greatly on grain yield of crops, the yield of crude protein was determined to be lower in crops in 2008 and 2009 than in 2007. However, the yield of crude protein was increased in pea and cereal intercrops compared with cereal sole crops. Jensen et al. (2006) indicated that intercropping proved especially valuable for the production of protein on land with weed problems and for enhancing the grain protein concentration in cereals to levels, which were only likely to be obtained with high levels of animal manure.
The effi ciency of resource use in intercropping relative to sole crop from interropping expose LER values (Fig. 1) based on grain yield. Total LERs above 1.00 were found in case of barley in 2007 and 2009, of wheat in 2009 and of triticale in 2009 intercrops, which shows the advantage of these mixtures over sole crops. Other cases in terms of total LER showed some disadvantage of intercrops. LERs calculations based on crude protein yield strengthen demonstrate that intercrops produce more per area than its sole cereal crops.
Conclusions
The productivity of spring cereal sole crops or pea and cereal intercrops depended on the species of cereals and varied depending on different year’s cultivation conditions. Grain productivity in some cases was estimated to be higher in intercrops, however it could not be concluded that pea/cereals intercrops yielded stably in three experimental years. However, protein content in the grain of cereals grown in intercrops was positively affected by pea and cereal intercrops. Pea-cereal intercrops can increase land use productivity.
References
Anil L, Park J, Phipps R H, Miller F A (1998): Temperate inter-cropping of cereals for forage: a review of the potential for growth and utilization with particular reference to the UK. Grass Forage Sci. 53: 301-317
Auškalnis A, Dovydaitis V (1999): Productivity of different pea varieties (in Lithuanian with English abstract). Zemdirbyste-Agriculture 68: 90-103
Auskalnis A (2001): Infl uence of sowing depth on producti-vity of peas on sandy loam soil (in Lithuanian with English abstract). Zemdirbyste-Agriculture 74: 105-111
Auškalnis A, Dovydait is V (1998): The dependence of pea crop density and productivity on seed rate and sowing time on the light loam (in Lithuanian with English abstract). Žemdirbystė-Agriculture 63: 143-155
Auškalnis A, Dovydait is V (2000): Productivity of pea, barley and their mixtures and the relationship between the grain protein content and nitrogen fertiliser and nitragine (in Lithuanian with English abstract). Zemdirbystė-Agriculture 69: 33-55
Cor re -He l lou G, Croza t Y (2005): N2 fi xation and N su-pply in organic pea (Pisum sativum L.) cropping systems as affected by weeds and peaweevil (Sitona Lineatus L.). Eur. J. Agron. 22: 449-458
Corre-Hellou G, Fustec J, Crozat Y (2006): Interspecifi c com-petition for soil N and its interactions with N2 fi xation, leaf expansion and crop growth in pea-barley intercrops. Plant Soil 282: 195-208
Crews T E, Peoples M B (2004): Legume versus fertilizer so-urces of nitrogen: ecological tradeoffs and human needs. Agric. Ecosyst. Environ. 102: 279-297
Dahlmann C, Fragstein N P (2006): Infl uence of different seed rates, sowing techniques and N supply on grain yield and quality parameters in intercropping systems. Proceedings of the European Joint Organic Congress, Denmark, 256-257
Dovydait is V (1992): Baltymingi pašariniai augalai. Vilnius, 189
Hauggaard-Nielsen H, Ambus P, Jensen E S (2003): The com-parison of nitrogen use and leaching in sole cropped versus intercropped pea and barley. Nutr. Cycl. Agroecosyst. 65: 289-300
Hauggaard-Nielsen H, Knidsen M T, Jørgensen J R, Jensen E S (2006): Intercropping wheat with pea for improved wheat baking quality. Proceedings of the European Joint Organic Congress, Denmark, 268-269
Hauggaard-Nielsen H, Mundus S, Jensen E S (2009): Nitro-gen dynamics following grain legumes and subsequent catch crop and the effects on succeeding cereal crops. Nutr. Cycl. Agroecosyst. 84: 281-291
c
abb
abababbaa
cabc
00,20,40,60,8
11,21,4
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
2007 2008 2009
LER
(tot
al g
rain
yie
ld)
Pea Cereal
b
aaa
ababbababca
cabc
0,00,20,40,60,81,01,21,41,6
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
pea+
whe
at
pea+
bar
ley
pea
+ oa
t
pea+
tritic
ale
2007 2008 2009
LER
(tot
al C
P y
ield
)
Pea Cereal
Fig. 1. Land equivalent ratio (LER) for pea-cereal intercrops based on grain and crude protein yieldsSlika 1. Površinski ekvivalentni odnos (LER) za združeni usev graška i pšenice zasnovan na prinosu sirovih proteina

Kadžiulienė Ž et al.188
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 183-188
Jensen E S (1996): Grain yield, symbiotic N2 fi xation and in-terspecifi c competition for inorganic N in pea-barley inter-crops. Plant Soil 182: 25-38
Jensen E S, Ambus N, Bellostas N, Boisen S, Brisson N, Corre-Hellou G, Crozat Y, Dahlmann C, Dibet A, Fragstein F, Gooding M, Hauggaard-Nielsen H, Kasyanova E, Launay M, Monti M, Pristeri A (2006): Intercropping of cereals and grain legumes for increased production, weed control, im-proved product quality and prevention of N-losses in Euro-pean organic farming systems. Proceedings of the European Joint Organic Congress, Denmark, 180-181
Lauk R, Lauk E (2005): The yields of legume-cereal mixes in years with hight – precipitation vegetation periods. Latvian J. Agron. 8: 281-285
Lauk R., Lauk E. (2008): Pea-oat intercrops are superior to pea-wheat and pea-barley intercrops. Acta Agric. Scand. Sec-tion B: Plant Soil Sci. 58: 139-144
Malezieux E, Crozat Y, Dupraz C, Laurans M, Makowski D, Ozier-Lafontane H, Rapidel B, de Tourdonnet S, Valantin-Morison M (2009): Mixing plant species in cropping sy-stems: concepts, tools and models. A review. Agron. Sustain. Dev. 29: 43-62
Maikštėnienė S, Satkus A (2000): Priešsėjinio dirvos dir-bimo būdai ir sėjos gylis žirniams sunkiame priemolyje, Žemdirbystė-Agriculture 69: 71-84
Mariotti M, Masoni A, Ercoli L, Arduini I (2009): Above- and below-ground competition between barley, wheat, lupin and vetch in a cereal and legume intercropping system. Grass Fo-rage Sci. 64: 401-412
Mead R, Willey R W (1980): The concept of ‘Land Equivalent Ration’ and advantages in yields from intercropping. Exp. Agric. 16: 217-228
Mikić A, Mihailović V, Ćupina B, Đordević V, Stoddard F L (2010): Introduction of novel legume crops in Serbia – Whi-te lupin (Lupinus albus). Ratar. Povrt. / Field Veg. Crop Res. 47: 21-26
Petersen R G (1994): Agricultural fi eld experiments: design and analysis. New York, USA, 407
Stoddard F L, Hovinen S, Konnturi M, Lindström K, Nykanen A (2009): Legumes in Finnish agriculture: history, present status and future prospects. Agric. Food Sci. 18: 191-205
Tarakanovas P, Raudonius S (2003): Agronominių tyrimų duo-menų statistinė analizė taikant kompiuterines programas ANOVA, STAT, SPLIT-PLOT iš paketo SELEKCIJA ir IRRISTAT. Akademija, 56
Tripolskaja L, Romanovskaja D, Slepetiene A (2008): The effect of various soil and crop management practices on the hu-mus status in sandy loam Haplic Luvisol (in Lithuanian with English abstract). Zemdirbyste-Agriculture 95: 3-18
Uticaj združene setve stočnog graška i jarih strnih žita na prinos zrna i sadržaj sirovih proteina
Židre Kadžijuljene · Lina Šarunajte · Irena Devejkite
Poljoprivredni institut, Litvanski istraživački centar za poljoprivredu i šumarstvo,Instituto al. 1, Akademija, Kedainiai LT-58344, Litvanija
Izvod: Prednost združenih useva je u povoljnim biološkim odnosima između useva i njihovog različitog iskorišćenja raspoloživih resursa za rast. Ovaj eksperiment je imao za cilj da istraži uticaj stočnog graška kao združenog useva sa žitaricama na poboljšanje dostupnosti azota žitaricama i sadržaja proteina u zrnu na ilovastom zemljištu u uslovima organske poljoprivrede. Eksperiment je izveden u periodu 2007-2009. u Poljoprivrednom institutu Litvanije u gradu Dotnuva (55o24’N, 23o50 E). Grašak (Pisum sativum L.) i jara pšenica (Triticum aestivum L.), jari ječam (Hordeum vulgare L.), ovas (Avena sativa L.) i tritikale (x Triticosecale Wittm.) bili su posejani kao združeni usevi 50:50 ili kao čisti usevi. Dobijeni trogodišnji rezultati pokazuju da su produktivnost i kvalitet jarih strnih žita kao čisti ili kao združeni usev zavisili od vrste žita i varirali su u zavisnosti od agrotehničkih uslova u različitim godinama. Produktivnost zrna u nekim slučajevima je procenjena kao viša u združenom usevu, međutim nije bila stabilna u tri godine eksperimenta. Kao združeni usev, grašak je imao pozitivan uticaj na koncentraciju i sadržaj proteina u zrnu žita. Ključne reči: grašak, jara strna žita, prinos, proteini, združeni usevi

Background Limit of Ni and Cr in Soils 189
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194original research article / originalni naučni rad
Methods for Assessment of Background Limit of Ni and Cr in Soils of Eastern Serbia
Vesna Mrvić · Ljiljana Kostić-Kravljanac · Mirjana Zdravković ·Nikola Koković · Veljko Perović · Dragan Čakmak · Mile Nikoloski
received / primljeno: 31.08.2010. accepted / prihvaćeno: 27.10.2010.© 2011 IFVC
Summary: The primary objective of the present study was to point out different approaches to bac-kground limits assessment. The content of selected trace elements is relatively low, and elements have right-skewed asymmetrical distribution, with high dispersion. Background for Cr and Ni (60 mg kg -1) which are obtained by graphical methods cumulative probability plot (CDF) and box-plot are similar. Natural and log-transform data are used for empirical methods. Results from antilogarithmic values are signifi cantly higher than from natural ones. Background limits obtained by empirical methods are different. Maps show that the largest part of territory has a relatively low concentration of investigated elements, whose background limits calculated by [Median+2MAD] methods are the most suitable. On the part of area with elevated elements content, where natural origin of elements had previously been proven, background limits are determined with box-plot method.Key words: background limit, Cr, Ni, soil
V. Mrvić · Lj. Kostić-Kravljanac · M. Zdravković · N. Koković · V. Perović · D. Čakmak · M. NikoloskiInstitute of Soil Science, Teodora Drajzera 7, 11000 Belgrade, Serbiae-mail: [email protected]
Introduction
Content of trace elements depends on pedogeological and anthropogenic factors. Therefore, it is advisable to determine the background concentration, the content in trace elements in soils without the infl uence of human activity, although it is very hard, almost impossible to fi nd territory without any infl uences of that kind (Sekulić et al. 2004, Čuvardić et al. 2004, Mrvić et al. 2009). Risk assessment of trace elements in soils is a key to many environmental protection measures and requires prediction of site specifi c background limit (Reimann & Garrett 2005). Determination of a unique background concentration for a large area (on a regional scale) does not take into account spatial variation of soil type and properties, so background limits for large area (e.g. for a whole state or a region) may cause either overestimation or underestimation of trace elements contamination and the associated risk for a particular soil (Diez et al. 2009). Thus, background limits evaluation is necessary on a local level. This approach enables development
of monitoring system, risk assessment and determination of trace elements levels, which require protection, remediation and sanation measures.
Several methods for assessing background limits have been used so far: classical [Mean ± 2Sd], [Median ± 2MAD], Tukey-s boxplot and cumulative probability plot - CDF graph (Gil et al. 2004, Galan 2008, Mico et al. 2008, Pérez-Sirvent et al. 2009) (Sd-standard deviation, MAD- median of the absolute deviations from the data′s median).
Results from classical method are not always satisfying for unequal data sets, and methods [Median ± 2MAD] and Tukey-s boxplot give better results (Reimann et al. 2005, Diez et al. 2009). Very often, geochemical data are right-skewed, thus besides natural values, logtransform values are used in order to get symmetric normal distribution (Reimann et al. 2005, Diez et al. 2009). On the other hand, graphical methods, EDA (histogram, and especially box plot and CDF) enable much better insight into data structure, infl ection points and possible background limits (Bech et al. 2005, 2008).
There are signifi cant differences between results obtained by different methods. Results or limits obtained by selected procedures and subsequent
Heavy Metals / Teški metali w
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Mrvić V et al.190
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
application of those limits on geochemical maps enable clearer insight in spatial distribution of trace elements and enable more accurate assessment of background limits. Although some methods have an obvious advantage, neither of them can clearly be distinguished as satisfying for all conditions.
This paper presents results from investigation of Eastern Serbian soil, which has very heterogenic properties. The primary objective of the present study was to point out different approaches for determination of background limits. Due to high anthropogenic infl uences at the investigated area, background limits for Cr and Ni have approximate values.
Materials and Methods Soil sampling was performed in 2005-2006.
Total of 979 surface (0-25cm) soil samples were taken by grid system at each 3.3 x 3.3 km (Fig. 1).
Fig. 1. Geographical characteristics of Eastern SerbiaSlika 1. Geografske karakteristike istočne Srbije
Analytical methods included determination of content of the hot acid extractable forms (“pseudototals”) of trace elements Cr, Hg, Ni and Zn, extracted with HNO3 and H2O2 with AAS.SPSS.10.0 was used for statistical analysis. Background values were determined by several methods, described in Reimann et al. (2005): classic [Mean ± 2Sd], [Median ± 2MAD], boxplot - upper whisker (Tukey 1977) and graphics - cumulative probability plot (CDF) and boxplot (Tukey 1977). For background assessment natural and logtransformed date were used. The software used for mapping was ESRI′ Arc View 8.3.
Results and Discussion
Contents and Distribution of Trace ElementsCompared to the interquaternary values,
the content of the selected trace elements is relatively low (Tab. 1). The values of Ni in about 93% of samples are below 50 mg kg-1, which is the maximal allowed concentration (MAC) for agricultural soils (SG RS 1994, BBodSchV 1999), while the values of Cr are below the critical (100 mg kg-1) in about 99% of samples.
Statistical analyses for all trace elements show that Skewness coeffi cient has a positive value (above zero) which means that elements have right-skewed asymmetrical distributions. There are positive Kurtosis coeffi cients, which means that the observations cluster has longer tails then those in the normal distribution. These elements show similar distribution patterns.
Graphical Methods
Cumulative probability plot (CDF) and box-plot methods provide a valuable tool for describing elements distribution, infl ection points and can be used for setting up background limits (Reimann et al. 2005).
CDF graphs are suitable because points at which line trends are changing, i.e. infl ection
Table 1. Statistical summary of selected soil properties and Ni and Cr concentrations (mg kg-1)Tabela 1. Statistički pokazatelji karakteristika zemljišta i sadržaja Ni i Cr (mg kg-1)
Para
met
ers
Mea
n
Std.
Dev
.
CV %
Skew
ness
Kur
tosis Min Max
Percentiles
5 25 50 75 95 98
Cr 23.75 19.20 80.8 2.81 13.92 0.20 185.79 2.8 11.8 21.0 31.1 52.6 74.1Ni 24.34 22.60 92.9 3.06 14.25 0.06 208.19 3.6 9.8 19.8 31.6 59.0 110.0

Background Limit of Ni and Cr in Soils 191
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
points, are clearly visible. At the same time, background limits also point at which core data separated from outliners can be accessed. Values of infl ection points from CDF graph are similar for Ni and Cr. First point is approximately at 40 mg kg-1, and second is at 60 mg kg-1, which is background limit at the same time (Fig. 2).
Since assessment of background limits from CDF graph is to a large extent based on subjective assessment, obtained values are not entirely reliable, thus it is advisable to use CDF graph in combination with other EDA graphs (box-plot, histograms, one-dimensional scattergram). Box-plot was used in this effort. For every element box-plot shows statistical
values (mathematically calculated): minimum, lower whisker, lower hinge (25%), median (50%), upper hinge (75%), upper whisker, outliers, far outliers (extreme) and maximum. Combination of these methods enables better assessment of background limits and percentage of samples which are above this values which belong to outliners and extremes. Application of both of these methods on realistic data sets gives equal background limits for Cr and Ni (60 mg kg-1).
Empirical (Calculated Methods)Besides graphical assessment of background
limits, empirical (calculated) method may also
be used. Three methods were used in this effort: classic [Mean+2Sdev], [Median+2MAD] and boxplot – mathematically calculated value upper whiskers.
Classic [Mean+2Sdev] method is commonly used, but it is greatly infl uenced by outliners and extremes and thus results in unrealistically high background limits for geochemical data, and it is not suitable for calculating action levels or clean-up goals in environmental legislation (Reimann et al., 2005).
Infl uences of extremes are much smaller in [Median+2MAD] method. MAD is a robust measure of the variability of a univariate sample of quantitative data. For a univariate data set X1, X2, ..., Xn, the MAD is defi ned as the median of
the absolute deviations from the data’s median: MAD = Median i (|Xi - Medianj (Xj)|).
Besides these methods, box-plot is also useful for the identifi cations of extreme values. Two parameters are calculated: upper inner fence (UIF), which is lower bounder of outliners appearance, and upper whisker, which is background limit. UIF is defi ned as the box extended by 1.5 times the length of the box towards the maximum: UIF = upper hinge (x) +1.5*HW(x). Upper whisker = max (x [x<UIF]), where HW (hinge with) is the difference between the hinges (upper hinge - lower hinge).
Fig. 2. Graphical description of Cr and Ni contents through box-plot and CDF diagram Slika 2. Grafi čki prikaz sadržaja Cr i Ni (box-plot i CDF dijagram)

Mrvić V et al.192
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
Assessment of background limits by the above mentioned method with natural data is possible because they have small variability. If variability is higher, logtransformation of data is required. According to Reimann et al. (2005) if the CV is between 70% and 100%, the use of logarithmical scale will likely be informative. If the CV>100% logarithmical scale should be used. This rule is applied on our data sets, since they are highly diverse (CV> 70%), as shown in table 2.
In natural simulations values from the [Median+2MAD] procedure results in the lowest background limits, followed by the boxplot. The
classical [Mean+2Sdev] rule presents the highest background limit, duo to large standard deviation. Similar results are obtained by other researchers (Reimann et al. 2005, Galan 2008). Results from logtransformed values are signifi cantly higher then from natural values. Background limits from the [Median+2MAD] procedure remain the lowest, while the highest is for boxplot. Logtransformed background limit for [Median+2MAD] methods is similar to background values from graph.
Since background limits are different for different methods, results should be checked on maps. Only after this checking, it is possible to
Ele
men
ts
Mean Sdev Median MAD [Mean+2Sd] [Median+2MAD]
Upper whisker
98th
per
cent
ile
Out
er
limit
-gr
ap
hic
s m
etho
ds
Nat
ural
Log1
0
Nat
ural
Log1
0
Nat
ural
Log1
0
Nat
ural
Log1
0
Nat
ural
Ant
i-log
Nat
ural
Ant
i-log
Nat
ural
Ant
i-log
Cr 23.7 1.23 19.2 0.41 21.0 1.32 9.5 0.19 62.1 111.2 40.0 50.9 58.9 131.8 74.1 60
Ni 24.3 1.23 22.6 0.41 19.8 1.30 10.6 0.24 69.5 113.2 41.0 60.8 64.0 184.1 110.0 60
Table 2. Background concentrations of Ni and Cr determined with different methods (mg kg-1)Tabela 2. Background koncentracije Ni i Cr u zemljištu određene različitim metodama (mg kg-1
Fig. 3. and Fig. 4. Spatial distribution of Cr and Ni in soils of Eastern Serbia (mg kg-1)Slika 3. i Slika 4. Prostorni raspored Cr i Ni u zemljištu istočne Srbije (mg kg-1)

Background Limit of Ni and Cr in Soils 193
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
propose which methods are the most suitable for local characteristics.
MapsBorder values of maps present infl ection
points from CDF graphs and logtransformed background values obtained by three methods (Fig. 3 and Fig. 4).
Spatial distribution of Ni shows that the highest levels of nickel are in alluvial of Velika Morava, and on slopes of Deli Jovan. For these areas previous investigation showed that Ni is of natural origin (Kalenić et al. 1976, Jakovljević et al. 1997, Antić-Mladenović 2003). On these areas proposed background concentrations are above 60 mg kg-1. For this region the method which best corresponds with local condition is box-plot, which in relation to other methods gives the highest values of background limits. On the other areas background limits are smaller, below 60 mg kg-1. For this area the best methods are CDF and [Median+2MAD] method. Similar to Ni, naturally elevated concentration of Cr content is on the slopes of Deli Jovan and for this area background is set by box-plot, which is 130 mg kg-1. In Velika Morava valley, content of Cr is smaller compared to those from Deli Jovan (40 mg kg-1 to 110 mg kg-1), so for this valley the most suitable method is [Mean+2Sdev].
Conclusions
This paper presents results from investigation of Eastern Serbian soil, which is of very hetero-genic properties. The primary objective of the present study was to point out at different ap-proaches for assessment background limits. Due to anthropogenic infl uences in this region, cal-culated values have orientation values. Statistical analyses show that content of the selected trace elements is relatively low, and they have right-skewed asymmetrical distributions, with high dis-persion. According to graphical methods, CDF and box-plot, background limits for Cr and Ni are equal (60 mg kg-1). Natural and logtransformed data were used in empirical methods. In natural simulations, values from the [Median+2MAD] procedure result in the lowest background limits, followed by the boxplot. Results from logtrans-formed values are signifi cantly higher than from natural and the values of background limits from the [Median+2MAD] procedure remain the low-est, while the highest are for box-plot.
Border values of maps present infl ection points from CDF graphs and antilogarithmic background values obtained by three methods. Maps show that the larger part of territory has relatively low concentrations of investigated elements whose background limits correspondent with limits obtained by [Median+2MAD] methods. On those parts of territory with elevated content, where there was previously established natural origin, background limits correspond with methods which give the highest values, i.e. box-plot.

Mrvić V et al.194
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 189-194
References
Antić-Mladenović S, Jakovljević M, Kostić N, Tomić Z (2005): Content and distribution of Ni and Cr in particle-size and density soil fraction, Zbornik abstrakta, XI Kongres društva za proučavanje zemljišta Srbije i Crne Gore, Budva, 63
Bech J, Tume P, Longan L, Reverter F (2005): Baseline concen-trations of trace elements in surface soils of the Torrelles and Sant Climent municipal districts (Catalonia, Spain). En-viron. Monit. Assess. 108: 309-322
Bech J, Tume P, Sokolovska M, Reverter F, Sanchez P, Longan L, Bech J, Puerte A, Oliver T (2008): Pedogeochemical ma-pping of Cr, Ni, and Cu in soils of the Barcelona Province (Catalonia, Spain): Relationships with soil physico-chemical characteristics. J. Geochem. Explor. 96: 106-116
Bundes-Bodenschutz und Altlasten verordnung (BBodSchV) (1999): [Elektronski izvor] [1 str] dostupno na adresi: http://bundesrecht.jurs.de/bundesrecht/bbodschv/gesamt.pdf. (citirano 8.3.2010., verifi kovano 29.8.2010.), Bundes-Boden-schutz und Altlastenverordnung, Deuchland
Čuvardić M, Hadžić V, Sekulić P, Kastori R, Belić M, Gove-darica M, Nešić Lj, Pucarević M, Vasin J (2004): Kontrola kvaliteta poljoprivrednog zemljišta i vode za navodnjavanje Vojvodine. Zbornik radova Instituta za ratarstvo i povrtar-stvo 40: 115-127
Díez M, Simón M, Martín F, Dorronsoro C, García C, Van Gestel I (2009): Ambient trace element background concen-trations in soils and their use in risk assessment. Sci. Total Environ. 407: 4622-4632
Galán E, Fernández-Caliani J C, González I, Aparicio P, Rome-ro A (2008): Infl uence of geological setting on geochemical baselines of trace elements in soils. Application to soils of South–West Spain. J. Geochem. Explor. 98: 89-106
Gil C, Boluda R, Ramos J (2004): Determination and evaluation of cadmium, lead and nickel in greenhouse of Almería (Spa-in). Chemosphere 55: 27-34.
Jakovljević M, Kostić N, Stevanović D, Blagojević S, Wil-son M, Martinović Lj (1997): Factors Infl uencing the Distribution of Heavy Metals in the Alluvial Soils of the Velika Morava River Valley, Serbia. Appl. Geo-chem.12: 637-642
Kalenić M, Hadži-Vuković M, Dolić D, Lončarević Č, Rakić M (1973): Tumač za list Kučevo L 34-128. Osnovna geološka karta 1:100 000. Zavod za geološka i geofi zička istraživanja. Beograd.
Micó C, Peris M, Recatalá L, Sánchez J (2007): Baseline values for heavy metals in agricultural soils in an Eu-ropean Mediterranean region. Sci. Total Environ. 378: 13-17
Mrvić V, Zdravković M, Sikirić B, Čakmak D, Kostić-Kravlja-nac Lj (2009): Sadržaj štetnih i opasnih elemenata. U: Mrvić V, Antonović G, Martinović Lj (ured.): Plodnost i sadržaj opasnih i štetnih materija u zemljištima centralne Srbije, In-stitut za zemljište, Beograd 75-145
Pérez-Sirvent C, Martínez-Sánchez M J, García-Lorenzo M L, Molina J, Tudela ML (2009): Geochemical background levels of zinc, cadmium and mercury in athropogenically infl uenced soils located in a semi-arid zone (SE, Spain). Ge-oderma 148: 307-317
Reimann C, Filzmoser P, Garrett R G (2005): Background and threshold: critical comparison of methods of determination. Sci. Total Environ. 346: 1-16
Reimann C, Garrett R G (2005): Geochemical background concept and reality. Sci. Total Environ. 350: 12 -27
Sekulić P, Hadžić V, Belić M, Pucarević M, Nešić Lj, Vasin J, Ze-remski-Škorić T, Ralev J (2004): Utvrđivanje stanja zagađe-nosti zemljišta na teritoriji opštine Novi Sad. Zbornik radova Instituta za ratarstvo i povrtarstvo 40: 65-72
SG RS 23/ 94 (1994): Pravilnik o maksimalno dozvoljenim koncentracijama štetnih elemenata u poljoprivrednom ze-mljištu, Službeni glasnik Republike Srbije, 23
Tukey J W (1977): Exploratory Data Analysis. Addison-Wesley, Reading
Metode za procenu Background Limita Ni i Cr u zemljištima istočne Srbije
Vesna Mrvić · Ljiljana Kostić-Kravljanac · Mirjana Zdravković ·Nikola Koković · Veljko Perović · Dragan Čakmak · Mile Nikoloski
Institut za zemljište, Teodora Drajzera 7, 11000 Beograd, Srbija
Izvod: U radu su prikazane različite metode određivanja background limita. Sadržaj ispitivanih elemenata u tragovima je relativno nizak, a distribucija elemenata je desno asimetrična, sa visokom disperzijom. Primenom grafi čkih me-toda (kumulativna kriva - CDF i box-plot) dobijene su slične vrednosti background limita za Cr i Ni (60 mg kg-1). Za empirijske metode korišćeni su prirodni i log-transformisani podaci. Vrednosti log-transformisanih podataka su znatno veće nego prirodne. Background limiti dobijeni empirijskim metodama su različiti. Karte pokazuju da najveći deo teritorije ima relativno niske koncentracije ispitivanih elemenata čiji background limiti najviše odgovaraju vrednostima izračunatim metodom [Median+2MAD]. Na delovima teritorije sa povećanim sadržajima, uslovljenim prvenstveno geohemijskim sastavom, background limit odgovara metodi sa najvećim vrednostima - box-plot. Ključne reči: background limit, Cr, Ni, zemljište

Indeks aridnosti i prinos kukuruza u Vojvodini 195
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202originalni naučni rad / original research article
Indeks aridnosti kao osnova analize uticaja režima padavina i temperature
vazduha na prinos kukuruza u Vojvodini
Borivoj Pejić · Goran Jaćimović · Dragana Latković · Đuro Bošnjak · Branko Marinković · Ksenija Mačkić
primljeno / received: 27.10.2010. prihvaćeno / accepted: 03.12.2010© 2011 IFVC
Izvod: Obračunom indeksa aridnosti (AI) analiziran je uticaj režima padavina i temperature vazduha na prinos kukuruza u Vojvodini. Eksperimentalna istraživanja proizvodnje kukuruza u uslovima navodnja-vanja obavljena su na oglednom polju Instituta za ratarstvo i povrtarstvo iz Novog Sada, na Rimskim Šančevima, na zemljištu tipa karbonatni černozem lesne terase u periodu 1997-2007. Prosečni prinosi kukuruza u Vojvodini za ispitivani period preuzeti su iz Statističkog godišnjaka Srbije. AI je obračunat korišćenjem standardizovanih vrednosti srednje mesečnih temperatura vazduha (T`) i mesečnih suma padavina (P`) u junu, julu i avgustu. Utvrđena je visoko signifi kantna korelacija (r = 0,86**) između AI i prosečnih prinosa kukuruza u Vojvodini. Korelacije između AI i prinosa kukuruza dobijenih u ogledima sa navodnjavanjem (r = -0,24) i bez navodnjavanja (r = 0,40) nisu bile statistički značajne. Velike razlike u vrednostima koefi cijenta korelacije između AI i prinosa kukuruza dobijenih u ogledima u uslovima prirodne obezbeđenosti biljaka vodom i prosečnih prinosa kukuruza u Vojvodini ukazuju na velike mo-gućnosti unapređenja tehnologije proizvodnje ove biljne vrste. Ostvareni efekti navodnjavanja na prinos kukuruza od oko 20% (2,04 t ha-1) ukazuju na opravdanost proizvodnje ove biljne vrste u navodnjavanju u klimatskim uslovima Vojvodine. Ključne reči: indeks aridnosti (AI), kukuruz, padavine, temperatura vazduha
B. Pejić · G. Jaćimović · D. Latković · Đ. Bošnjak · B. Marinković · K. MačkićPoljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Kukuruz (Zea mays L.) je jedna od najznačajnijih ratarskih biljaka u svetu. Po površinama na koji-ma se gajio u svetu u periodu 1995-2007. (oko 140 miliona ha), zauzimao je treće mesto, iza pšenice i pirinča. Međutim, po proizvodnji zrna od oko 609 miliona tona bio je na prvom mestu ispred pirin-ča i pšenice. Kukuruz se u istom periodu u svetu gajio na približno 24% površina pod žitaricama, sa učešćem oko 30% u proizvodnji zrna i proseč-nim prinosom od 4,33 t ha-1 (FAOSTAT 2007). Još veći značaj kukuruz ima za našu zemlju, jer se u istom periodu gajio na oko 38% oraničnih površi-na, a naročito u Vojvodini, gde je njegovo učešće u strukturi setve bilo na 41% oranica, ili drugim rečima, prosečne površine za dugi niz godina izno-sile su oko 650.000 ha (Starčević & Latković 2005). Prosečan prinos u periodu 1997-2007. iznosio je
5,0 t ha-1 varirajući u širokom rasponu od 2,94 t ha-1 do 6,44 t ha-1 (Republički zavod za statistiku 2007) i bio je u korelaciji, pre svega, sa količinom i rasporedom padavina.
Klimatsko područje Vojvodine odlikuje se pro-menljivošću meteoroloških uslova u pojedinim godinama, što se posebno odnosi na padavine, koje po godinama variraju kako po količini tako i po rasporedu. Ako sušu defi nišemo kao nedo-voljno snabdevanje biljaka vodom, kada je po-tencijalna evapotranspiracija veća od stvarne, bez obzira na uzroke toj pojavi, onda je očigledno da je suša u našim klimatskim uslovima redovna po-java, javlja se svake godine i ostavlja manje ili veće posledice na prinose gajenih biljaka.
Obično se smatra da je kukuruz otporan pre-ma suši, te da ekonomično troši vodu. Međutim, pošto stvara veliku vegetativnu masu, daje visoke prinose, ima dugačak vegetacioni period u toku prolećno-letnjih, toplih meseci, kukuruz troši ve-like ukupne količine vode (Bošnjak & Pejić 1994, Pejić 1996, Bošnjak & Pejić 1997). U nedostatku vode kukuruz uspešno prebrodi sušu, ali daje niže
Vodni režim / Water Regimew
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Pejić B i sar.196
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
prinose, jer biljke troše teže pristupačne kategorije i oblike vode iz zemljišta.
Za normalan rast i razviće biljaka kukuruza, te postizanje visokih i stabilnih prinosa dobrog kvaliteta, potrebna je optimalna vlažnost zemlji-šta tokom celog perioda vegetacije (Bošnjak & Dobrenov 1993). Samo u takvim uslovima bilj-ke mogu da troše vodu na nivou svojih potreba, odnosno na nivou potencijalne evapotranspi-racije (ETP). ETP kukuruza u agroekološkim uslovima Vojvodine iznosi 470-540 mm (Boš-njak 1982), a takva verovatnoća obezbeđenosti padavinama je samo 4-5%, što znači da se ne može očekivati realizacija genetskog potencijala inače vrlo rodnih hibrida kukuruza, jer količina padavina uslovljava fi toklimatski nivo prinosa i neosporno je da poljoprivredi Vojvodine ne-dostaje voda kao pokretač ostalih faktora proi-zvodnje (Vučić 1976).
Od klimatskih činilaca, padavine i tempera-tura vazduha su od posebnog značaja za uspeh biljne proizvodnje. Količina vode u zemljištu za-visi od padavina koje treba da obezbede stalan priliv vode u pristupačnom obliku za normalan rast i razviće biljaka. Autori Bošnjak & Pejić (1997) ukazuju na visoko signifi kantnu korelaci-ju između količina padavina u periodu vegetaci-je i količine padavina u letnjim mesecima (junu, julu i avgustu) i prinosa kukuruza u klimatskim uslovima Vojvodine. Utvrđena je visoko signifi -kantna korelacija između temperature vazduha i utrošene vode na evapotranspiraciju biljaka (Bošnjak 1982, Pejić 2000). Brojni istraživači
ističu da se detaljnom analizom režima padavina i temperature vazduha može jasno sagledati nji-hov uticaj na proizvodnju kukuruza u datim kli-matskim uslovima (Sabau et al. 2002, Starčević et al. 2005, Kovačević i sar. 2007, Marinković et al. 2008). Dobijeni rezultati su posebno važ-ni i mogu biti od velikog praktičnog značaja u unapređenju tehnološkog procesa proizvodnje ove biljne vrste. Kovačević i sar. (2007) ističu da se obračunom indeksa aridnosti (AI) može dobiti jasna predstava o uticaju režima padavina i temperature vazduha na proizvodnju kukuru-za. Prosečna vrednost indeksa aridnosti od 0,7 za područje istočne Hrvatske ukazuje na povolj-nost klimatskih uslova za proizvodnju kukuruza u ovom području. Međutim, vrednosti indeksa aridnosti u sušnim godinama u intervalu od 7,2-7,5 pokazuju da se moraju poznavati i moguć-nosti proizvodnje kukuruza u uslovima navod-njavanja i tako osigurati planirana proizvodnja i visoki prinosi.
Cilj ovog istraživanja je bio da se na osnovu obračunatih vrednosti indeksa aridnosti detaljno sagleda uticaj režima padavina i temperature vaz-duha na prinos kukuruza u Vojvodini.
Materijal i metod rada
Eksperimentalna istraživanja proizvodnje kukuruza u uslovima navodnjavanja kišenjem obavljena su na oglednom polju Instituta za ra-tarstvo i povrtarstvo iz Novog Sada, na Rimskim Šančevima (N 45o 19’, E 19o 50’, 84 m nadmorske
Tabela 1. Srednje mesečne temperature vazduha i mesečne količine padavina u periodu vegetacije kukuruza (Rimski Šančevi)Table 1. Mean monthly air temperatures and monthly precipitation during maize growing season (Rimski Šančevi)
GodinaYear
Mesec / MonthmajMay
junJune
julJuly
avgustAugust
septembarSeptember
oC mm oC mm oC mm oC mm oC mm1997 17,8 17 20,7 62 20,0 128 20,2 125 15,8 171998 16,1 64 21,5 104 21,6 124 21,6 82 15,7 771999 16,9 96 20,3 89 21,6 192 21,6 28 19,7 662000 18,5 39 21,4 28 22,1 29 24,0 5 17,8 132001 17.8 79 18,3 219 22,3 80 22,7 30 16,1 1622002 19,1 19 21,1 28 23,6 35 21,5 50 16,3 452003 20,6 23 24,0 31 22,6 60 24,6 30 17,2 842004 15,2 89 19,8 97 21,9 63 21,7 39 16,2 422005 17,0 38 19,3 135 21,3 122 18,3 134 17,3 672006 16,6 70 19,7 104 23,5 31 19,7 125 17,9 242007 18,4 99 22,0 71 23,2 39 21,2 80 14,6 79
ProsekAverage 17,6 58 20,7 88 22,2 82,1 21,6 66 16,8 61

Indeks aridnosti i prinos kukuruza u Vojvodini 197
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
visine), na zemljištu tipa karbonatni černozem lesne terase u periodu 1997-2007.
Ogled je postavljen po metodu blok siste-ma i prilagođen uslovima navodnjavanja kiše-njem. U ogledu su bile zastupljene varijanta sa navodnjavanjem, sa predzalivnom vlažnošću zemljišta 60-65% od poljskog vodnog kapa-citeta (PVK) i kontrolna varijanta bez navod-njavanja. Vreme zalivanja je određivano pra-ćenjem dinamike vlažnosti zemljišta dekadno, a po potrebi i u kraćem vremenskom periodu po slojevima zemljišta od po 10 cm, do 60 cm dubine. Vrednosti prosečnih prinosa kukuru-za u Vojvodini preuzeti su iz Statističkog go-dišnjaka Srbije.
Podaci o padavinama i temperaturama vazduha (Tab. 1) su uzeti sa meteorološke stanice Rimski
Šančevi koja se nalazi u sklopu Oglednog polja Instituta. Vreme zalivanja i zalivne norme su pri-kazane u tabeli 2.
Indeks aridnosti (AI - aridity index, Harouna & Carlson 1994) kao pokazatelj uticaja režima padavina i temperature vazduha na prinos ku-kuruza u Vojvodini obračunat je korišćenjem standardizovanih vrednosti srednjih mesečnih
temperatura vazduha (T`) i mesečnih suma padavina (P`) u letnjim mesecima junu, julu i avgustu.
AI = T` - P`T` = (T – Tbar)/stP` = (P – Pbar)/spT = srednje mesečne temperature vazduha (oC),P = mesečne sume padavina (mm),Tbar = prosečna vrednost srednjih mesečnih
temperatura vazduha u periodu ispitivanja (oC),Pbar = prosečna vrednost mesečnih suma pada-
vina u periodu ispitivanja (mm),st = standardna devijacija srednjih mesečnih
temperatura vazduha (oC),sp = standardna devijacija mesečnih suma pa-
davina (oC).
U ogledu je bio zastupljen hibrid NS-640, FAO grupe zrenja 600, na osnovnoj parceli od 35 m2. Berba je obavljena ručno u tehnološkoj zrelosti, a prinos zrna sa 14% vlažnosti obra-čunat je u t ha-1. Primenjena je savremena teh-nologija proizvodnje, a sve agrotehničke mere su obavljene blagovremeno, u optimalnim ro-kovima.
Tabela 2. Vreme zalivanja, zalivne norme i norma navodnjavanjaTable 2. Irrigation schedule, irrigation rates and irrigation requirements
GodinaYear
Norma zalivanja-Datum zalivanjaIrrigation rate-Date of irrigation
(mm)
Norma navodnjavanja
Irrigation requirement
(mm)
Mesec/MonthJunJune
JulJuly
AvgustAugust
1997 - - - -1998 - - 60 – 07 601999 - - - -2000 60 – 23 60 – 24 60 – 08 1802001 - 60 – 11 - 602002 60 – 24 60 – 05 - 1202003 60 – 05
60 – 20 30 – 05 - 200+
2004 - 45 – 02 60 – 17 1052005 60 – 05 - - 60
2006 -60 – 06 60 – 17 60 – 28
- 180
2007 45 – 22 60 – 19 - 165

Pejić B i sar.198
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
Rezultati i diskusija
Klimatsko područje Vojvodine odlikuje se promenljivošću meteoroloških uslova u po-jedinim godinama, što se posebno odnosi na padavine koje po godinama variraju kako po količini tako i po rasporedu. Prosečna količina padavina u Vojvodini (1964-2007) iznosi 598,7 mm, a srednja godišnja temperatura 11,20C. U periodu vegetacije (IV-IX) padne 328 mm kiše ili 55% od ukupne količine, a u istom periodu prosečna temperatura vazduha iznosi 17,80C. Samo na osnovu ukupne količine padavina u toku vegetacije ne može se govoriti o dovoljnoj ili nedovoljnoj obezbeđenosti biljaka vodom. Bošnjak (1993) ukazuje da je suša redovna poja-va u klimatskim uslovima Vojvodine, da se jav-lja kraći ili duži period svake godine i da često ostavlja ozbiljne posledice na umanjenje prinosa gajenih biljaka. Najintenzivnija suša u Vojvodi-ni je tokom jula i avgusta kada je obezbeđenost padavinama na nivou opšte potencijalne evapo-transpiracije od minimum 100 mm samo 12% (Bošnjak 1999).
Realna ocena o povoljnosti režima pada-vina i temperature vazduha za proizvodnju kukuruza u ispitivanom području može se dobiti obračunom indeksa aridnosti (Sabau et al. 2002, Kovačević i sar. 2007). Period istra-živanja (1997-2007) se odlikovao relativno po-voljnim uslovima za proizvodnju kukuruza. U povoljnim godinama za proizvodnju kukuruza (1997, 1998, 1999, 2001, 2004, 2005, 2006) in-deks aridnosti se kretao u intervalu od -0,2 u 2004. godini, do -5,8 u 2005. godini kada su postignuti najveći prosečni prinosi kukuruza u Vojvodini za posmatrani period (Tab. 3). U go-dinama nepovoljnim za proizvodnju kukuruza (2000, 2002, 2003, 2007), sa visokim defi citom lakopristupačne vode u zemljištu, indeks arid-nosti je bio u intervalu od 3,3 do 6,0. Najniži prosečni prinosi kukuruza u Vojvodini su bili u izrazito sušnim godinama 2000. i 2003. sa in-deksom aridnosti 5,6 odnosno 6,0 (Tab. 3).
U ogledima sa navodnjavanjem ostvarena je statistički signifi kantna razlika u visini prinosa kukuruza između navodnjavane i varijante bez navodnjavanja. Prinos je u uslovima navodnjava-nja, u proseku, bio veći za 2,04 t ha-1, odnosno oko 20%. U povoljnim godinama efekat navod-njavanja je izostajao ili je bio vrlo skroman (3,3% u 2005), a u sušnim vrlo visok (67,4% u 2000)
(Tab. 3). Ostvareni efekti navodnjavanja na prinos kukuruza bili su niži u odnosu na prosečno pove-ćanje od 28,7% koje je ranije utvrđeno u klimat-skim uslovima Vojvodine (Bošnjak i sar. 2005). Podaci o padavinama (Tab. 1), vremenu zalivanja i zalivnim normama (Tab. 2) ukazuju na dopunski karakter navodnjavanja u Vojvodini (Pejić et al. 2009). Padavine se ne mogu unapred predvideti za duži period vremena, odnosno posle obavlje-nih zalivanja može pasti obilnija kiša (2005. god.) što može dovesti do pogrešnih zaključaka da na-vodnjavanje nije ni bilo potrebno. Konačan sud o potrebama za zalivanjem ne treba donositi samo na osnovu mesečnih suma, već je potrebna detalj-na analiza dnevnih vrednosti padavina.
Visoko signifi kantne vrednosti koefi cijenta ko-relacije (r=0,86**) između AI i prosečnih prino-sa kukuruza ukazuju na veliku zavisnost prinosa ove biljne vrste od režima padavina i temperature vazduha u letnjim mesecima u Vojvodini. Autori Sabau et al. (2002) ističu skoro identične vrednosti koefi cijenta korelacije (r=0,81**) između AI i pri-nosa kukuruza za klimatske uslove Rumunije. Ta-kođe Kovačević et al. (2007) za period 1996-2003. za klimatske uslove Hrvatske ukazuju na visok stepen korelacije (r=0,90**) između AI i prinosa kukuruza. Isti autori ističu 2000. i 2003. kao vrlo nepovoljne godine za proizvodnju kukuruza u Hrvatskoj, sa vrednostima AI od 7,5 odnosno 7,2. Relativno niske pozitivne vrednosti koefi cijenta korelacije između AI i prinosa kukuruza dobijenih u ogledima u uslovima prirodne obezbeđenosti biljaka vodom (r=0,40ns) u odnosu na vrednosti koefi cijenta korelacije između AI i prosečnih pri-nosa kukuruza u Vojvodini (r=0,86**) ukazuju na mogućnost unapređenja tehnologije proizvodnje ove biljne vrste u Vojvodini. Naime, ukoliko se uporede prinosi u ogledu bez navodnjavanja (pro-sečno 11,05 t ha-1) sa prosečnim prinosom kuku-ruza u Vojvodini (5,01 t ha-1), uz pretpostavku po-djednakih zemljišnih i klimatskih uslova, prosečno povećanje prinosa od oko 120% se može pripisati doprinosu tehnologije proizvodnje (izbor hibri-da, plodored, obrada, đubrenje, setva, nega use-va). Činjenica je da je ovakav zaključak vrlo grub i površan, te se bez daljeg upuštanja u detaljniju analizu doprinosa agrotehnike, ipak može konsta-tovati da postoji ogromna rezerva u mogućnosti povećanja prinosa kukuruza u Vojvodini. Takođe, niske negativne vrednosti koefi cijenta korelacije (r=-0,24ns) između AI i prinosa kukuruza u uslovi-ma navodnjavanja, ukazuju na dopunski karakter navodnjavanja u Vojvodini, odnosno da nepred-vidive padavine koje prate navodnjavanje mogu uticati na visinu prinosa kukuruza ostvarenih u navodnjavanju.

Indeks aridnosti i prinos kukuruza u Vojvodini 199
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
Brojevi označeni različitim slovima u koloni ukazuju na signifi kantne razlike LSD vrednosti na pragu značajnosti P≤ 0.05
Visok stepen korelacije između prinosa kuku-ruza u Vojvodini i AI (Sl. 1); kao i između AI i defi cita lakopristupačne vode u zemljištu u peri-
GodinaYear
Prinos – Yield (t ha-1) Prosečne vrednosti prinosa za Vojvodinu
Average yield of maize in Vojvodina
(t ha-1)
ETm(mm)
ETa(mm)
Defi citVodeWaterdefi cit(mm)
Indeks aridnostiAridityindex
NavodnjavanoIrrigated
NenavodnjavanoNonirrigated
1997 14,06 12,16 5,69 450 409 41 -4,41998 10,26 9,17 4,72 461 461 - -1,41999 11,44 10,50 5,35 475 442 33 -2,92000 13,46 8,04 2,94 495 174 321 5,62001 10,77 9,61 5,59 464 383 81 -3,12002 13,60 10,21 4,99 481 237 244 4,02003 13,53 9,65 3,42 520 261 259 6,02004 12,96 10,50 5,88 458 353 105 -0,22005 14,22 13,76 6,54 442 442 - -5,82006 14,82 13,92 5,92 461 399 62 -1,12007 14,78 14,00 4,06 477 361 116 3,3
ProsekAverage 13,08A 11,05B 5,01C 471 356 115
0
2
4
6
8
10
12
14
16
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007
Godine - Year
Prin
os -
Yiel
d (t
ha-1
)
-8
-6
-4
-2
0
2
4
6
8
Arid
ity In
dex
(AI)
Navodnjavano - irrigatedNenavodnjavano - NonirrigatedProsečan prinos u Vojvodini - Average yield in VojvodinaAI
Tabela 3. Prinosi kukuruza u uslovima sa navodnjavanjem i bez navodnjavanja, prosečni prinosi kukuruza u Vojvodini, utrošak vode na potencijalnu (ETm) i stvarnu (ETa) evapotranspiraciju, defi cit lakopristupačne vode u zemljištu u periodu vegetacije i vrednosti indeksa aridnosti (AI)Table 3. Maize yields under irrigated and nonirrigated conditions, average yields of maize in Vojvodina, water used on potential (ETm) and actual (ETa) evapotranspiration, defi cit of readily available soil water in the growing season, values of aridity indeks (AI)
Slika 1: Zavisnost prinosa kukuruza od vrednosti indeksa aridnostiFig. 1: Dependence of maize yield on aridity index

Pejić B i sar.200
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
odu vegetacije kukuruza (Sl. 2) ukazuju na veliku mogućnost upotrebe AI u analizi uticaja režima padavina i temperatura vazduha na prinos kuku-ruza u Vojvodini.
Zaključak
Na osnovu rezultata istraživanja utvrđena je visoko signifi kantna korelacija (r=0,86**) izme-đu režima padavina i temperatura vazduha izra-ženog preko indeksa aridnosti (AI) i prosečnih prinosa kukuruza u Vojvodini. Korelacije između AI i prinosa kukuruza dobijenih u ogledima sa (r=-0,24) i bez (r=0,40) navodnjavanja nisu bile statistički značajne. Velike razlike u vrednostima koefi cijenta korelacije između AI i prinosa kuku-
ruza dobijenih u ogledima u uslovima prirodne obezbeđenosti biljaka vodom i prosečnih prinosa kukuruza u Vojvodini ukazuju na velike moguć-nosti unapređenja tehnologije proizvodnje ove biljne vrste.
Visok stepen korelacije između prinosa ku-kuruza u Vojvodini i AI, kao i između AI i defi cita lakopristupačne vode u periodu vege-tacije kukuruza ukazuju na veliku mogućnost upotrebe AI u analizi uticaja režima padavina i temperatura vazduha na prinos kukuruza u Vojvodini.
Ostvareni efekti navodnjavanja na prinos kuku-ruza od oko 20%, odnosno 2,04 t ha-1, ukazuju na opravdanost proizvodnje ove biljne vrste u navod-njavanju u klimatskim uslovima Vojvodine.
Slika 2: Odnos indeksa aridnosti i defi cita lakopristupačne vode u zemljištu u periodu vegetacije kuku-ruzaFig. 2: Relation between aridity index and defi cit of readily available water in the soil during the maize growing season
0
50
100
150
200
250
300
350
400
1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007
Godina - Year
Def
icit
lako
pris
tupačn
e vo
de
Def
icit
of re
adily
ava
ilabl
e w
ater
(mm
)
-8
-6
-4
-2
0
2
4
6
8
Arid
ity In
dex
WDAI

Indeks aridnosti i prinos kukuruza u Vojvodini 201
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
Literatura
Bošnjak Đ (1982): Evaporacija sa slobodne vodene površine kao osnova zalivnog režima i njen odnos prema ETP ku-kuruza i soje. Doktorska disertacija. Poljoprivredni fakultet, Novi Sad
Bošnjak Đ, Dobrenov V (1993): Efekat predzalivne vlažnosti na prinos i evapotranspiraciju kukuruza. »Korišćenje i održa-vanje melioracionih sistema«, Beograd, 155-158
Bošnjak Đ, Pejić B (1994): Realizacija racionalnog zalivnog re-žima kukuruza. Zbornik radova Instituta za ratarstvo i povr-tarstvo 22: 167-179
Bošnjak Đ, Pejić B (1997): Odnos navodnjavanja i zemljišne suše prema prinosu kukuruza u Vojvodini. Zbornik radova IX Kongresa Jugoslovenskog društva za proučavanje zemlji-šta, 624-631
Bošnjak Đ (1999): Navodnjavanje poljoprivrednih useva. Poljo-privredni fakultet, Novi Sad
Bošnjak Đ, Pejić B, Maksimović L (2005): Irrigation - a condi-tion for high and stable corn production in the Vojvodina Province. International conference on sustainable agricultu-re and European integration processes, Novi Sad. Savrem. Poljopr. 3-4: 82-87
FAOSTAT (2007): On line statistical data base of the Food and Agricultural Organization of the United Nations [Elektron-ski izvor] dostupno na http://www.fao.org
Harouna S, Carlson E (1994): Analysis of an Iowa Aridity In-dex in relationship to climate and crop yield. J. Iowa Acad. Sci. 101: 14-18
Marinković B, Crnobarac J, Marinković D, Jaćimović G, Mir-cov D V (2008): Weather conditions in the function of optimal corn yield in Serbia and the Vojvodina province. International scientifi c conference „Ist Scientifi c Agrono-mic Days“, 13-14. november 2008., Slovak University of
Agriculture Nitra, Slovak Republic, Department of Crop Production, Faculty of Agrobiology and Food Resources. Proceeding of reviewed scientifi c papers, collection of cri-tiqued scientifi c works on CD (ISBN 978-80-552-0125), 15-19
Kovačević V, Šimić D, Šoštarić J, Josipović M (2007): Precipi-tation and temperature regime impacts on maize yields in eastern Croatia. Maydica 52: 301-305
Pejić B (1996): Vodni bilans kao osnova analize prirodnog defi -cita vode i njegov odnos prema prinosu kukuruza. Zbornik radova Instituta za ratarstvo i povrtarstvo 25: 513-520
Pejić B (2000): Evapotranspiracija i morfološke karakteristike kukuruza u zavisnosti od dubine navlaženog zemljišta i nji-hov odnos prema prinosu. Doktorska disertacija, Poljopri-vredni fakultet Novi Sad.
Pejić B, Maksimović L, Škorić D, Milić S, Stričević R, Ćupina B (2009): Effect of water stress on yield and evapotranspirati-on of sunfl ower. Helia 32: 19-33
Sabau N C, Domuta C, Man T E, Sandor M, Brejea R (2002): Drought analysis by the climate indexes in link with the yield of the main crops from the Crisurilor plain, Romania. pp. 1-4. In: Proc. Intl. Conf. Drought Mitigation and Prevention of Land Desertifi cation. Bled, Slovenia, 21 - 25 April 2002, Intl. Commission on Irrigation and Drainage
Starčević Lj, Latković D (2005): Prinos kukuruza u Vojvodini, 2004. godine bio je najviši u poslednjih 10 godina (5,88 t ha-1). Da li je moglo biti više? Da! Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo 41: 385-394
Starčević Lj, Latković D, Malešević M (2005): Dependence of corn yield on weather conditions and nitrogen fertilization in IOSDV Novi Sad. Arch. Acker-Pfl . Boden 51: 513-522
Republički zavod za statistiku (2007): Statistički godišnjak Srbije [Elektronski izvor] dostupno na http://webrzs.stat.gov.rs
Vučić N (1976): Navodnjavanje poljoprivrednih kultura. Poljo-privredni fakultet, Novi Sad

Pejić B i sar.202
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 195-202
Aridity Index as a Useful Tool for Describing Impacts of Precipitation and Temperature Regime on Maize Yield in Vojvodina
Borivoj Pejić · Goran Jaćimović · Dragana Latković · Đuro Bošnjak · Branko Marinković · Ksenija Mačkić
Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8,21000 Novi Sad, Serbia
Summary: Calculated values of aridity index (AI) were used as a base for the analysis of precipitation and tempe-rature impact on maize yield in Vojvodina. An investigation of maize production with irrigation was carried out at the experiment fi eld of Institute of Field and Vegetable Crops in Novi Sad on calcareous chernozem soil on the loess terrace, in the period 1997-2007. Standardized values of temperatures (T`) and precipitation (P`) were used for AI calculation. Highly signifi cant correlation (r = 0.86**) was found between AI and average yields of maize in Vojvodina. There were no signifi cant correlations between AI and maize yield obtained in fi eld experiments without irrigation (r = 0.40) and with irrigation (r = -0.24). Low values of correlation coeffi cient between AI and maize yiel-ds obtained in fi eld experiments without irrigation and correlation coeffi cient values between AI and average yields of maize in Vojvodina indicate that production technology of maize in Vojvodina could be improved. Higher values of maize yield in irrigation conditions than in conditions without irrigation (20% or 2.04 t ha-1) confi rm justifi cation of maize production with irrigation under climatic conditions of Vojvodina.Key words: air temperature, aridity index (AI), maize, precipitation

Smanjenje rizika u proizvodnji pšenice 203
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 203-206
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 203-206originalni naučni rad / original research article
Smanjenje rizika u proizvodnji pšenice primenom vremenske prodajne opcije
Todor Marković · Milenko Jovanović
primljeno / received: 09.06.2010. prihvaćeno / accepted: 02.07.2010.© 2011 IFVC
Izvod: U radu je analiziran efekat eliminacije rizika (hedžing efekat) u proizvodnji pšenice u Sremu, primenom vremenske prodajne opcije na bazi količine padavina. Rezultati pokazuju da je upotreba vremenske prodajne opcije efi kasna samo u slučaju ako je mesto proizvodnje u neposrednoj blizini me-teorološke stanice i ako je pretpostavljena visoka korelacija između količine padavina i prinosa pšenice. Ključne reči: efi kasnost eliminacije rizika, količina padavina, proizvodnja pšenice, vremenska prodajna opcija
T. Marković · M. JovanovićPoljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Proizvodnja žitarica u Sremu, a posebno u njegovom centralnom delu zavisna je od količi-ne padavina. U mesecima kada se formira prinos (april-maj) kiša je neophodna. U poslednjih 30 godina palo je između 31,1 mm i 256,5 mm kiše (sa prosečnom količinom od približno 100 mm). Prinosi žitarica jako variraju, budući da sistemi za navodnjavanje ne stoje na raspolaganju ili nisu u dovoljnoj meri iskorišćeni. U sušnim godinama (2003) prinosi su jako podbacili, a budući da u Srbiji ne postoji osiguranje od suše, poljoprivred-nici su pretrpeli velike gubitke, koji se iz godine u godinu ponavljaju. Zbog toga bi bilo interesantno sa novim sistemima osiguranja pokušati donekle nadoknaditi nastale štete.
U prošlosti su poljoprivrednici kupovinom osiguranja pokušali da se zaštite od kolebanja prinosa useva i plodova uslovljenih vremenskim rizicima. Relativno nov instrument za upravljanje rizikom u biljnoj proizvodnji predstavljaju vre-menski derivati. Vremenski derivati defi nišu se kao fi nansijski instrumenti čija isplata zavisi od nekog vremenskog parametra, a na tržištu vre-menskih derivata dominira trgovina vremenskim opcijama (Berg 2005). Za analizu vremenskih de-rivata najčešće je uzimana temperatura, kao bazna
vrednost. Opet sa druge strane količine padavina takođe imaju veliku važnost u biljnoj proizvodnji, ali se u ovoj oblasti javlja relativno manji broj pu-blikacija (Turvey 2001, Berg et al. 2005, Mußhoff et al. 2007).
Cilj ovog rada je da se ukaže na efekat elimina-cije rizika (hedžing-efekat) u proizvodnji pšenice sa primenom vremenske prodajne opcije i bez nje. Posebno se apostrofi ra činjenica da sa uda-ljavanjem mesta proizvodnje od meteorološke stanice nastaje geografski bazni rizik (neujedna-čenost padavina na ove dve lokacije) i bazni rizik proizvodnje (slabija korelacija između količine padavina i visine prinosa pšenice).
Materijal i metod
Osnovne izvore podataka predstavlja dokumen-tacija o prosečnim prinosima i prodajnim cenama pšenice sa izabranog individualnog gazdinstva u opštini Ruma, kao i podaci o mesečnim količinama padavina sa meteorološke stanice Rimski Šančevi u Novom Sadu u periodu 1999-2008.
Uspešna primena vremenskih derivata uslov-ljena je jačinom uticaja količine padavina na pri-nos useva i plodova (Vedenov & Barnett 2004). Prilikom konstrukcije vremenske prodajne opcije ključno pitanje je izračunavanje isplate iz opcije i određivanje cene rizika (fer premije), koju je kupac spreman da plati za transfer rizika. Iz ugla kupca, isplata iz vremenske prodajne opcije (Ip) proizilazi iz razlike između graničnog nivoa (R) i realizovane
Ekonomska analiza / Economic Analysis w
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Marković T, Jovanović M204
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 203-206
količine padavina (x), multiplikovane sa novčanom vrednošću indeksa (O) (Berg et al. 2005):
[ ]0),( xRMaxOI p −⋅= (1)
Za određivanje premije koristi se metoda istorij-ske simulacije, gde fer premija predstavlja diskon-tovanu očekivanu vrednost isplate iz vremenskog derivata (Berg et al. 2005, Mußhoff et al. 2005).
)exp()( trIEP pf Δ⋅−⋅= (2)
U datoj formuli ( fP ) predstavlja fer premiju, )( pIE očekivanu vrednost isplate, )exp( tr Δ⋅−
predstavlja diskontni faktor, gde je ( r ) kamatna stopa, a ( tΔ ) vremenski period.
Po istoj metodologiji određuju se i ostali para-metri neophodni za konstrukciju vremenskih de-rivata (vremenski indeks, funkcija isplate i ogra-ničenje isplate).
Primenom kvantitativnih metoda za ocenu rizi-ka utvrđuje se da li je eliminacija rizika (hedžing) uspešnija sa primenom vremenskih derivata ili bez nje. U radu se primenjuju metod stohastičke dominacije i metod rizične vrednosti. Koncep-tom stohastičke dominacije upoređuju se funkci-je raspodele (kumulativne verovatnoće) vrednosti proizvodnje pšenice sa primenom vremenske prodajne opcije i bez nje. Sa druge strane posma-tra se varijansa, kao mera disperzije u statistici, i kvantili u raspodeli vrednosti proizvodnje, te se na osnovu njih utvrđuje mogućnost smanjivanja rizika od gubitka sa i bez vremenskog derivata (Berg et al. 2005). Sva neophodna izračunavanja vrše se uz pomoć računarskog softvera @Risk,
koji je posebno razvijen za problematiku uprav-ljanja rizikom.
Rezultati i diskusija
Uzima se u obzir vremenska prodajna opcija na bazi količine padavina, koja se odnosi na kumula-tivnu mesečnu količinu padavina koja je izmerena na meteorološkoj stanici Rimski Šančevi u periodu april-maj 2009. Poljoprivrednik ima mogućnost da kupi prodajnu opciju 01. septembra 2008. (vre-menski period iznosi 9 meseci). Ta opcija donosi poljoprivrednom proizvođaču isplatu na dan 01. juna 2009. prema formuli (1), ukoliko je kumula-tivna mesečna količina padavina ispod 95 mm (što predstavlja granični nivo, odnosno prosečnu koli-činu padavina poslednjih 10 godina).
( ) 80,95max ⋅−= xI p (3)
Ako se prekorači taj nivo, isplata je jednaka nuli dok za svaki milimetar, kada je količina padavina ispod graničnog nivoa, poljoprivrednik dobija 8 € mm-1. Međutim, ni isplata ne ide do minimalne količine padavina, već je ona limitirana na 55 mm (Graf. 1).
Ukoliko poljoprivrednik želi da osigura proi-zvodnju pšenice uz pomoć vremenske prodajne opcije neophodno je utvrditi cenu opcije. Kako bi se izbegle teškoće prilikom procenjivanja vre-menskih derivata polazi se od fer premije, kao cene opcije, kod koje je očekivana dobit za obe strane (kupca i prodavca) jednaka nuli. U tom slučaju is-ključuju se transakcioni troškovi i premija rizika. Određivanje fer premije vrši se metodom istorijske simulacije, gde se kao vremensko razdoblje uzima
(95 - x,0)
Grafi kon 1. Fer premija i isplate iz prodajne opcije u proizvodnji pšeniceGraph 1. Fair premium and payoff of put option in wheat production
-50
0
50
100
150
200
250
300
350
0 15 35 55 75 95 115 135 155
Кoličina padavina / Rainfall (mm)
Ispl
ata
iz o
pcije
/ Pay
off o
f opt
ion
(€/h
a)
Ograničenje isplate / Payoff limit = 55 mm
Izvršni nivo / Strike level= 95 mm
Fer premija / Fair price = 15,63 €

Smanjenje rizika u proizvodnji pšenice 205
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 203-206
unapred izabrani period i kamatna stopa od 5%. Uz pomoć odgovarajućeg softvera izračunata je fer premija koja iznosi 15,63 € (Graf. 1).
Za analizu korisnog dejstva vremenskih derivata u jednoj proizvodnji na ispitivanom gazdinstvu najbolje je poći od pretpostavke da prinos pšenice direktno zavisi od količine padavina u periodu april-maj, tako da svaki milimetar kiše uslovljava visinu prinosa. Me-toda stohastičke dominacije se koristi za odre-đivanje vrednosti proizvodnje sa primenom vremenskih derivata i bez nje. U slučaju bez vremenske prodajne opcije ukupna vrednost proizvodnje jednaka je tržišnoj vrednosti proi-zvodnje koja predstavlja proizvod prinosa pše-nice ( y~ ) i njene cene ( yc ), prema formuli:
yP cyV ⋅= ~~ (4)
Sa druge strane, u slučaju sa opcijom poljopri-vrednik prisvaja i dobit iz opcije, odnosno ostva-rena tržišna vrednost proizvodnje uvećava se za isplatu iz vremenske prodajne opcije ( pI~ ) i uma-njuje za fer premiju ( fP ), polazeći od formule (1):
[ ] fyP PxRMaxOcyV −−⋅+⋅= )~(,0~~
(5)
Kako je dobit nakon odbitka fer premije jed-naka nuli, tako su približno iste očekivane vred-nosti proizvodnje pšenice sa primenom vremen-ske prodajne opcije i bez nje, dok je mnogo veće smanjenje rizika u slučaju sa opcijom (Tab. 1).
Vremenska varijabla na kojoj se bazira vremen-ski derivat izmerena je na najbližoj referentnoj meteorološkoj stanici. Činjenica je da se mesto proizvodnje pšenice nalazi na većoj ili manjoj udaljenosti od meteorološke stanice. Što je veća udaljenost dolazi do pojave geografskog baznog rizika, koji najčešće uslovljava i bazni rizik pro-izvodnje, tj. slabija je korelacija između količine padavine i prinosa.
Posmatrana vremenska prodajna opcija pro-cenjena je na 15,63 €, što znači da toliko košta nadoknada niže količine padavina od graničnog nivoa (95 mm) u periodu april-maj, a koja je iz-merena na meteorološkoj stanici Rimski Šančevi u Novom Sadu. Kao što je prikazano u tabeli 1, poljoprivrednik u neposrednoj blizini referentne meteorološke stanice može prodajom opcije u potpunosti isključiti rizik od gubitka (down side risk). Takođe varijansa kod vrednosti proizvodnje može biti smanjena za 32,97%. Ako se kao mera za smanjenje rizika posmatraju kvantili, uglav-nom se u prvi plan uzima donji deo raspodele,
Tabela 1. Smanjenje rizika u proizvodnji pšenice pri različitim korelacijama između količine padavina i prinosaTable 1. Risk reduction in wheat production with different correlation coeffi cients between rainfall and yield
Vrednost proizvodnje bez opcije / Revenue without option
(€/ha)
Vrednost proizvodnje sa opcijom / Revenue with option (€/ha)
Koefi cijent korelacije / Correlation
coeffi cient1,0 1,0 0,9 0,8 0,7 0,6 0,5
Očekivana vrednost / Expected value
(€/ha)960,29 960,44 960,63 960,73 960,76 960,48 960,67
Varijansa / Variance(€/ha) 14.652 9.821 10.494 11.084 11.869 12.212 13.381
Smanjenje rizika / Risk reduction (%) - 32,97 28,38 24,34 18,99 16,65 8,67
Kva
ntili
/
Qua
ntile
s
5 % 761,13 838,64 817,21 800,49 788,44 783,64 777,12
10 % 805,88 852,99 840,95 829,26 824,17 824,60 817,50
50 % 959,99 944,37 950,09 951,25 957,11 951,25 957,58
95 % 1.156,41 1.140,79 1.140,11 1.140,92 1.141,44 1.143,66 1.146,28

Marković T, Jovanović M206
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 203-206
koji izražava najnižu vrednost proizvodnje, od-nosno gde se javljaju najlošiji rezultati. Tako npr. vrednost kvantila od 5% bez opcije iznosi 761,13 € ha-1, dok u slučaju sa opcijom raste na 838,64 € ha-1 (Tab. 1). Izvesno je da je efekat prodajne opcije utoliko niži što se više povećava udaljenost između mesta proizvodnje i referentne meteoro-loške stanice. Ako postoji još i nizak koefi cijent korelacije između količine padavina i prinosa use-va, značajno se smanjuje efekat zaštite.
Zaključak
Predstavljeni model primene vremenskih de-rivata pokazuje da geografski bazni rizik i bazni rizik proizvodnje u velikoj meri utiče na hed-žing-efekat vremenske prodajne opcije, odno-sno da su vremenski derivati na bazi količine pa-davina u velikoj meri regionalno ograničeni. U datom primeru koefi cijent korelacije između ko-ličine padavina i prinosa pšenice u velikoj meri određuje efekat smanjenja rizika. Na osnovu prethodnog može se dovesti u pitanje primena vremenskih derivata u biljnoj proizvodnji. Zbog toga je potrebno da budući ponuđači vremen-skih derivata treba da omoguće gušću mrežu referentnih meteoroloških stanica, da ponude mešoviti vremenski indeks (npr. kombinacijom količine padavina i prosečne temperature) ili je-dan vremenski indeks sa podacima iz više me-teoroloških stanica, kao i širi spektar različitih tipova vremenskih derivata (npr. kombinacija kupovne i prodajne opcije pri nedovoljnoj ili preobilnoj količini padavina).
Literatura
Berg E (2005): Integriertes Risikomanagement: Notwen-digkeit und Konzepte für die Landwritschaft. Ta-gungsband zum Fachkolloquium anlässlich des 80. Geburtstages von Prof. Em. Dr. Dr. h.c. Günter Steffen, 24.09.2004, 53-67
Berg E, Schmitz B, Starp M, Trenkel H (2005): Wetter-derivate: Ein Instrument im Risikomanagement für Landwirtschaft? Berichte über Landwirtschaft 80: 94-133
Vedenov D V, Barnett B J (2004): Effi ciency of Weather Derivatives as Primary Crop Insurance Instru-ments. J. Agric. Resour. Econ. 29: 387-403
Mußhoff O, Odening M, Xu W (2005): Zur Bewer-tung von Wetterderivaten als innovative Risiko-managementinstrumente in der Landwirtschaft. Agrarwirtschaft 54: 197-209
Mußhoff O, Odening M, Xu W (2007): Zur Quantifi -zierung des Basis-risikos von Wetterderivaten. 46. Jahrestagung. Schriften der Gesellschaft für Wirts-chafts- und Sozialwissenschaften des Landbaues (GeWiSoLa), Landwirtschaftsverlag, Münster-Hil-trup, 381-393
Turvey C G (2001): Weather Derivatives for Specifi c Event Risks in Agriculture. Rev. Agric. Econ. 23: 333-351
Risk Reduction in Wheat Production with Weather Put Option
Todor Marković · Milenko Jovanović
Faculty of Agriculture, University of Novi Sad,Trg Dositeja Obradovića 8, 21000 Novi Sad
Summary: In this paper is analysed hedging effectiveness in wheat production in Srem, Serbia with rainfall put option. It follows that effect of risk reduction is signifi cant if the fi eld of production is close to the meteorological station and if a high correlation between weather indices and yield is assumed. Key words: hedging effectiveness, rainfall, weather put option, wheat production

Proizvodni bazni rizik kod pšenice i kukuruza 207
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212originalni naučni rad / original research article
Uticaj količine padavina na prinospšenice i kukuruza kao proizvodni bazni rizik
Todor Marković · Milenko Jovanović
primljeno / received: 14.07.2010. prihvaćeno / accepted: 08.09.2010.© 2011 IFVC
Izvod: U radu je primenom metoda regresione i korelacione analize istraživana uzajamna međuzavi-snost količine padavina i prinosa pšenice i kukuruza u Vojvodini u periodu 1975-2005. Rezultati poka-zuju da količina padavina u aprilu i maju ima veoma važan uticaj na visinu prinosa pšenice, a u periodu april-avgust na visinu prinosa kukuruza. Ukoliko se u budućnosti u proizvodnji pšenice i kukuruza žele koristiti vremenski derivati, kao novi fi nansijski instrumenti u osiguranju useva, neophodno je uvažavati ove činjenice, budući da se uspešna primena vremenskih derivata zasniva na smanjenju proizvodnog baznog rizika koji nastaje pri slabijoj korelaciji između nekog vremenskog parametra i prinosa pojedinih useva. Ključne reči: količina padavina, kukuruz, prinos, proizvodni bazni rizik, pšenica, regresiona i korelaci-ona analiza, vremenski derivati
T. Marković · M. JovanovićPoljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Biljna proizvodnja, kao primarna proizvod-nja u poljoprivredi, posebno je izložena uticaju vremenskih rizika. Vremenski činioci predstav-ljaju grupu proizvodnih rizika koji umnogome određuju visinu prinosa useva, kao naturalnog pokazatelja uspeha. Između različitih vremenskih faktora poseban značaj za biljnu proizvodnju ima količina padavina. Kada nastupi sušna godi-na dolazi do smanjenja prinosa, a pošto u Srbiji nije zastupljeno osiguranje od suše, poljoprivred-ni proizvođači trpe ozbiljne štete u proizvodnji. Činjenica je da se uvek nakon poplave, suše ili snažne oluje pojača diskusija o osiguranju useva i plodova, kojim se može kompenzovati gubitak u proizvodnji (Breustedt 2003).
Pored klasičnog osiguranja od gubitka prinosa, pojedini autori ukazuju na potrebu širenja tržišta osiguranja od većeg broja rizika (sa uključenim osiguranjem od suše), odnosno vremenskih deri-vata, koje je uglavnom prisutno u razvijenim ze-mljama Evrope i Severne Amerike (Berg 2002). Vremenski derivati predstavljaju novije fi nansij-ske instrumente za transfer rizika, koji su nastali polovinom 90-ih godina prošlog veka, a njihova
isplata zavisi od nekog vremenskog parametra. Vremenskim derivatima može se trgovati na or-ganizovanom fi nansijskom tržištu (berzi) ili va-nberzanski (OTC - over the counter). Iako vremen-ski derivati pokazuju brojne prednosti u odnosu na klasično osiguranje, tržište ovih proizvoda još uvek je relativno malo. Ukoliko je težnja da se u budućnosti razvije likvidno tržište vremenskih derivata, onda će biti neophodno da se pre sve-ga osiguravaju usevi koji imaju značajno učešće u setvenoj strukturi ili viši nivo prinosa po hektaru. Posebnu pažnju treba posvetiti osnovnim žitari-cama (pšenica i kukuruz).
Cilj ovog rada je da se u dužem vremenskom periodu primenom metoda regresione i korela-cione analize ispita uticaj količina padavina na prinos osnovnih žitarica. Što je jača uzajamna međuzavisnost ova dva faktora, izražena koefi ci-jentima korelacije, smanjuje se proizvodni bazni rizik, a samim tim povećava efekat eliminacije ri-zika primenom vremenskih derivata.
Materijal i metod
Osnovne izvore podataka predstavlja doku-mentacija Statističkog zavoda Vojvodine o pro-sečnim prinosima pšenice i kukuruza u periodu 1975-2005. u pojedinim opštinama Vojvodine (Bačka Palanka, Bački Petrovac, Inđija, Irig, Ruma, Temerin, Titel i Žabalj), zatim podaci o požnjevenim površinama, ostvarenim prinosi-
Ekonomska analiza / Economic Analysisw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Marković T, Jovanović M208
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
ma i ukupnoj proizvodnji pšenice i kukuruza u Vojvodini u periodu 1975-2005. kao i podaci o dnevnim količinama padavina sa meteorološke stanice Rimski Šančevi u Novom Sadu, koji su takođe uzeti za tridesetogodišnji period (1975-2005), kako bi se dobila realnija slika o kretanju padavina.
Efi kasna primena vremenskih derivata pretpo-stavlja visok stepen korelacije između količine pa-davina i nivoa prinosa useva i plodova (Vedenov & Barnett 2004, Marković & Jovanović 2011). U radu se polazi od ukupnih mesečnih količina padavina koje se dobijaju sabiranjem dnevnih ko-ličina padavina u datom mesecu. Sa druge strane koriste se podaci o prosečnim prinosima pšenice i kukuruza u izabranim opštinama. Odnosi izme-đu uslova i rezultata proizvodnje utvrđuju se pri-menom metoda regresione i korelacione analize.
U radu se polazi od pretpostavke da prinos pšenice i kukuruza direktno zavisi od količine padavina u vegetacionom periodu, tako da svaki milimetar kiše uslovljava visinu prinosa. Stoga je neophodno prethodno eliminisati ostale vremen-ske faktore, kao i druge činioce (tehničko-tehno-loški, ekološki, ekonomski). Na osnovu toga se podaci o prinosima prethodno „očiste“ pomoću trenda (linearnog, kvadratnog i kubnog), tako da su preostala kolebanja prinosa zavisna samo od oscilacija visine vodenog taloga (Schmitz 2007).
Ispitivanje zavisnosti ostvarenog prosečnog prinosa (ŷ) od mesečne količine padavina (x) vrši se uz pomoć linearne (ŷ = a + bx), kvadratne (ŷ = a + bx + cx2) i kubne regresije (ŷ = a + bx + cx2 + dx3). Izračunavanjem koefi cijenata korelacije utvrđuje se uticaj količine padavina na prinos po-jedinih useva. Koefi cijenti korelacije izračunavaju se prema sledećem obrascu (Hartung 1998):
( ) ( )( ) ( )22 yyxx
yyxxrii
iiXY
−Σ⋅−Σ
−⋅−Σ= (1)
Na ovaj način iskazana je jačina međuzavisno-sti između nezavisne i zavisne varijable. Ovaj pokazatelj može pokazivati vrednosti između -1,0 i 1,0. U slučaju pozitivne korelacije pove-ćanje vrednosti nezavisno promenljive dovodi do povećanja zavisne varijable i obrnuto, kod negativne korelacije povećanje vrednosti jedne varijable prouzrokuje smanjenje druge promen-ljive. U slučaju korelacije koja iznosi 0 ne postoji nikakva međuzavisnost između varijabli i ovakva korelacija se neće uzimati u obzir. Svi neophodni proračuni vrše se uz pomoć računarskog softvera (Statistika 8.0), gde se podaci obrađuju primenom matematičko-statističkog instrumentarijuma.
Rezultati i diskusija
Vremenski derivati predstavljaju proizvode fi -nansijskog tržišta i javljaju se u formi vremenskih fjučersa, opcija ili forvarda. Fjučersi su standardi-zovani ugovori kojima se trguje na berzi, a forvardi su privatno dogovoreni ugovori kojima se trguje na OTC tržištu i oni su najčešće u formi svopo-va, dok se vremenskim opcijama može trgovati na berzi, ali i na OTC tržištu i one se najčešće javljaju u formi kupovne ili prodajne opcije.
Prilikom formiranja vremenskih derivata neop-hodno je odrediti sledeće parametre: tip derivata, vremenski indeks, meteorološku stanicu, vremen-ski period i funkciju isplate. Vremenski derivati najčešće se baziraju na količini padavina ili proseč-noj temperaturi. Funkcija isplate predstavlja sušti-nu vremenskih derivata i ona obuhvata utvrđiva-nje premije osiguranja, graničnog nivoa i novčane vrednosti indeksa. Granični nivo predstavlja onu vrednost indeksa od koje se vrši isplata, dok se vi-sina isplate određuje uz pomoć novčane vrednosti indeksa koja označava isplaćeni novčani iznos po jedinici indeksa, a premija predstavlja diskontova-nu očekivanu vrednost buduće isplate.
Vremenski derivati su na prvi pogled veoma slični ugovorima o klasičnom osiguranju, ali se između njih javljaju značajne razlike. Osnovne prednosti vremenskih derivata u odnosu na kla-sično osiguranje ogledaju se u sledećem (Alaton et al. 2002, Campbell & Diebold 2002):
Kod vremenskih derivata nije potreban nikakav dokaz o nastaloj šteti prouzrokovanoj nekim štet-nim događajem, a samim tim isključuje se i proce-na štete, kao i troškovi koji na taj način nastaju;
Isključuje se problematika moral-hazarda, gde pojedinci mogu svojim postupcima da utiču na visinu štete i na taj način oštete osiguravača izvla-čeći korist za sebe;
Ne postoji negativna selekcija kod koje poljo-privrednici sa manjim rizikom odustaju od plaća-nja viših premija osiguranja.
Međutim, vremenski derivati imaju i određene nedostatke u odnosu na klasično osiguranje, a oni obuhvataju (Mußhoff et al. 2005):
• Geografski bazni rizik - odnosi se na uda-ljenost mesta proizvodnje od referentne mete-orološke stanice, jer što je veće rastojanje veća je i razlika u visini vodenog taloga ili ostvarenoj dnevnoj temperaturi na te dve lokacije.
• Proizvodni bazni rizik - odnosi se na koefi cijent korelacije između prinosa nekog useva i određe-nog vremenskog parametra. Ukoliko je koefi cijent korelacije niži, povećava se bazni proizvodni rizik, a time se smanjuje korisnost vremenskih derivata. Zbog toga se u ovom radu posebna pažnja pokla-
x y

Proizvodni bazni rizik kod pšenice i kukuruza 209
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
nja odnosu količine padavina kao vremenskog in-deksa i nivoa prinosa najvažnijih žitarica.
Rezultati analize pokazuju da u poljoprivredi Vojvodine u ispitivanom periodu (1975-2005) najveću zastupljenost u zasejanim površinama pod žitima ima kukuruz (61,04%), zatim pšenica (32,99%), dok na ostale žitarice otpada 5,97%.
U posmatranom periodu (1975-2005) na po-dručju Vojvodine požnjevene površine pod pše-nicom iznose prosečno 356.695 ha ispoljavajući tendenciju pada (stopa promene -1,10%). Na ovim površinama, uz prosečan prinos od 4,34 t ha-1 ostvaruje se proizvodnja od 1.550.888 t. Vari-ranje proizvodnje jako je izraženo (koefi cijent va-rijacije 24,69%). Ovako veliko variranje ukazuje da naši prinosi, a time i proizvodnja još u velikoj meri zavise od klime, tj. u klimatski dobrim go-dinama ostvaruju se viši prinosi, a u lošim godi-nama veoma niski prinosi, što je posledica veoma loše agrotehnike (Denčić i sar. 2009). Pad proseč-nog prinosa (stopa pada -0,09%) uticao je na pad obima proizvodnje (stopa promene -1,18%) kao što je prikazano u tabeli 1.
Analiza proizvodnje kukuruza ukazuje na su-protne tendencije u pogledu dinamike razvitka zasejanih površina, prinosa i ostvarenog obima proizvodnje. Naime, površine rastu po niskim godišnjim stopama (stopa promene 0,14%) i uslovljavaju blagi porast količine proizvedenog kukuruza (stopa rasta 0,57%). U posmatranom periodu prosečna površina pod ovim usevom iznosi 659.995 ha. Prosečan prinos od 5,27 t ha-1 ima blagi porast (stopa promene 0,43%) i velike oscilacije (koefi cijent varijacije 22,10%). Proseč-
na proizvodnja kukuruza od 3.477.156 t beleži širok interval varijacije, što se refl ektuje visokim koefi cijentom varijacije od 23,69% (Tab. 1).
Analiza međuzavisnosti vremenskog činioca (količina padavina) i ostvarenog prinosa jedino je moguća ukoliko na raspolaganju stoje dugoročni meteorološki podaci, kao i ažurno praćenje kreta-nja prinosa useva u dužem vremenskom periodu. U nastavku se daje pregled ostvarenih rezultata regresione i korelacione analize kod pšenice i ku-kuruza u Vojvodini.
Ostvareni koefi cijenti korelacije kao rezultat međuzavisnosti trendom uslovljenih kolebanja prinosa pšenice i mesečnih količina padavina iskazani su po pojedinim mesecima, ali i zbirno, gde se uzima po nekoliko meseci zajedno (Tab. 2). Koefi cijenti korelacije za pojedine mesece se kod opština u Sremu (Inđija, Irig, Ruma) razlikuju od koefi cijenata korelacije kod drugih mesta. Na-ime, kod njih se uglavnom javljaju pozitivne kore-lacije između količine padavina i prinosa pšenice. Najviši koefi cijent korelacije (r = 0,51) ostvaren je u opštini Inđija u aprilu, dok su nešto niži ko-efi cijenti korelacije ostvareni u istom mesecu u opštinama Žabalj (r = 0,42) i Irig (r = 0,37).
Ako se posmatraju koefi cijenti korelacije izme-đu višemesečnih količina padavina i ostvarenog prinosa zapaža se da opštine u Sremu karakteriše pozitivna korelacija. Najviši koefi cijent korelacije (r = 0,46) zabeležen je u opštini Irig u aprilu i maju, dok je u Žablju zabeležen nešto niži koefi -cijent korelacije (r = 0,44). Zapaža se da količine padavina u aprilu i maju umnogome određuju budući prinos pšenice.
Tabela 1. Površine, prinosi i proizvodnja pšenice i kukuruza u Vojvodini (1975-2005)Table 1. Wheat and corn areas, yields and production in Vojvodina (1975-2005)
Pokazatelji IndicatorsProsečna vrednost
Average value
Koefi cijent varijacije
Variations coeffi cient
(%)
Interval varijacije Variation interval Stopa promeneChange rate
(%)Minimum Min. Maksimum Max.
Pšenica / WheatPovršina
Cultivated area (ha)356.695 13,37 268.324 437.138 -1,10
PrinosYield (t ha-1)
4,34 17,62 2,30 5,61 -0,09
Proizvodnja Production (t)
1.550.888 24,69 728.565 2.310.832 -1,18
Kukuruz / CornPovršina
Cultivated area (ha)659.995 6,80 544.421 759.296 0,14
PrinosYield (t ha-1)
5,27 22,10 2,94 6,99 0,43
Proizvodnja Production (t)
3.477.156 23,69 1.810.438 5.150.830 0,57

Marković T, Jovanović M210
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
Koefi cijenti korelacije u opštinama Temerin i Ti-tel relativno su niski, što se može tumačiti relativno malom zavisnošću prinosa pšenice od količine pa-davina. U opštinama Bačka Palanka i Bački Petro-vac koefi cijenti korelacije uglavnom su negativni.
Za razliku od pšenice čiji vegetacioni period traje od oktobra do jula, kod kukuruza je on znat-no kraći (april-oktobar). Međuzavisnost količine padavina i trendom uslovljenih kolebanja prinosa data je u tabeli 3. Posmatrajući koefi cijente ko-relacije u pojedinim mesecima primećuje se da je najviša korelacija zabeležena u julu u Bačkom Petrovcu (r = 0,64), Titelu (r = 0,63), Bačkoj Pa-lanci (r = 0,62), Inđiji (r = 0,60) i Rumi (r = 0,60). U Irigu, Temerinu i Žablju koefi cijenti korelacije
zaostaju za prethodnim. Korelacija je evidentno znatno slabija do juna i od septembra.
Ako se posmatraju višemesečne količine pa-davina, može se primetiti da uglavnom postoji pozitivna korelacija i da su koefi cijenti korelacije relativno visoki. Najviši koefi cijent korelacije je u Rumi u periodu april-avgust (r = 0,70), kao i u Temerinu (r = 0,66) i Inđiji (r = 0,64). Koefi ci-jenti korelacije u opštinama Bački Petrovac, Irig i Žabalj relativno su niži, što se može tumačiti manjom zavisnošću prinosa kukuruza od količina padavina. U veoma malom broju slučajeva koefi -cijenti korelacije su negativni, što ukazuje na to da povećanje količina padavina prouzrokuje sma-njenje prinosa kukuruza.
Tabela 2. Koefi cijenti korelacije između prosečnog prinosa pšenice i količine padavina u pojedinim mesecima u nekim opštinama Vojvodine (1975-2005) Table 2. Correlation coeffi cients between average wheat yield and rainfall in certain months in some municipalities in Vojvodina (1975-2005)
Bačka Palanka
Bački Petrovac
Inđija Irig Ruma Temerin Titel Žabalj
januar -0,15 -0,24 0,01 -0,05 -0,15 0,00 0,29 0,03februar 0,05 0,18 0,00 -0,05 -0,07 0,11 0,04 0,05mart 0,05 -0,12 0,14 0,36 0,01 0,14 0,16 0,27april 0,14 -0,02 0,51 0,37 0,28 0,29 0,20 0,42maj 0,04 0,12 0,02 0,06 -0,06 0,07 0,12 0,15jun -0,04 -0,06 0,03 0,08 -0,10 0,14 -0,10 0,05jul 0,01 0,02 0,30 0,11 0,21 0,10 0,07 -0,03avgust -0,15 -0,21 0,18 -0,05 -0,05 -0,10 0,14 -0,19septembar -0,13 -0,14 -0,12 -0,21 -0,25 -0,11 -0,19 -0,14oktobar -0,11 -0,19 -0,04 -0,21 0,06 -0,11 0,01 -0,10mar-apr 0,12 -0,08 0,35 0,25 0,20 0,28 0,23 0,34mar-maj 0,11 0,01 0,26 0,36 0,10 0,24 0,23 0,40mar-jun 0,05 -0,02 0,20 0,29 0,02 0,23 0,11 0,29mar-jul 0,05 -0,01 0,32 0,31 0,12 0,26 0,13 0,25mar-avg 0,00 -0,07 0,34 0,26 0,09 0,20 0,16 0,16mar-sep -0,04 -0,10 0,26 0,17 0,01 0,14 0,09 0,10mar-okt -0,07 -0,16 0,26 0,11 0,03 0,11 0,09 0,08apr-maj 0,11 0,07 0,24 0,46 0,11 0,21 0,19 0,44apr-jun 0,05 0,01 0,18 0,21 0,02 0,23 0,07 0,25apr-jul 0,04 0,02 0,31 0,24 0,13 0,24 0,10 0,20apr-avg -0,01 -0,05 0,33 0,19 0,09 0,18 0,13 0,11apr-sep -0,05 -0,08 0,25 0,10 0,01 0,12 0,06 0,05apr-okt -0,08 -0,14 0,24 0,04 0,03 0,09 0,06 0,02maj-jun -0,01 0,03 0,04 0,10 -0,12 0,15 -0,01 0,13maj-jul 0,00 0,04 0,22 0,15 0,05 0,18 0,04 0,08maj-avg -0,06 -0,05 0,25 0,10 0,02 0,11 0,09 -0,01maj-sep -0,09 -0,08 0,17 0,02 -0,06 0,06 0,02 -0,05maj-okt -0,13 -0,15 0,16 -0,05 -0,04 0,02 0,02 -0,09jun-jul -0,02 -0,02 0,25 0,14 0,09 0,17 -0,02 0,01jun-avg -0,08 -0,11 0,27 0,09 0,05 0,09 0,05 -0,07jun-sep -0,11 -0,13 0,18 0,00 -0,04 0,04 -0,02 -0,10jun-okt -0,14 -0,19 0,15 -0,08 -0,02 0,00 -0,02 -0,14jul-avg -0,07 -0,09 0,31 0,05 0,13 0,02 0,13 -0,12jul-sep -0,12 -0,14 0,21 -0,05 0,00 -0,03 0,03 -0,16jul-okt -0,15 -0,20 0,17 -0,13 0,02 -0,07 0,03 -0,19avg-sep -0,20 -0,24 0,04 -0,18 -0,21 -0,15 -0,03 -0,23avg-okt -0,24 -0,33 0,01 -0,29 -0,14 -0,20 -0,02 -0,26sep-okt -0,17 -0,24 -0,11 -0,30 -0,13 -0,16 -0,12 -0,17

Proizvodni bazni rizik kod pšenice i kukuruza 211
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
Sa druge strane, eksperimentalna istraživanja obavljena na oglednom polju Instituta za ratar-stvo i povrtarstvo na Rimskim Šančevima u No-vom Sadu, ukazuju na još veću međuzavisnost količine padavina i prinosa kukuruza. Ako se u obzir uzme jedan mesec, korelacionom analizom je utvrđena visokosignifi kantna zavisnost prino-sa kukuruza i količine padavina u avgustu (r = 0,93), dok je višemesečno utvrđena signifi kantna zavisnost prinosa kukuruza i količine padavina u julu i avgustu (r = 0,77) (Pejić et al. 2009). Na si-gnifi kantnu korelaciju prinosa sa padavinama kod kukuruza ukazuju i drugi autori (Bošnjak 2004, Marinković i sar. 2008).
Prilikom razmatranja međusobne uslovljenosti vremenskog parametra i visine prinosa, zapažaju
se nešto niži koefi cijenti korelacije kod pšenice. Najviša korelacija, vezana za količinu padavina, evidentirana je u mesecima april (jedan mesec), odnosno april i maj (višemesečno) u pojedinim opštinama u Sremu (Inđija, Irig i Ruma). Kod kukuruza su koefi cijenti korelacije relativno viši, pri čemu veliki uticaj na prinos ima količina pa-davina u julu (jedan mesec), odnosno u periodu april-avgust (više meseci zajedno). U Sremu je izražena jača korelacija između količine padavi-na i visine prinosa, budući da je na području op-ština u Sremu (Inđija, Irig i Ruma) konstatovan viši nivo padavina kako godišnji, tako i u vegeta-cionom periodu, u odnosu na opštine u Bačkoj (Bačka Palanka, Bački Petrovac, Temerin, Titel i Žabalj).
Tabela 3. Koefi cijenti korelacije između prosečnog prinosa kukuruza i količine padavina u pojedinim mesecima u nekim opštinama Vojvodine (1975-2005) Table 3. Correlation coeffi cients between average corn yield and rainfall in certain months in some mu-nicipalities in Vojvodina (1975-2005)
Bačka Palanka
Bački Petrovac Inđija Irig Ruma Temerin Titel Žabalj
mart -0,01 0,02 0,20 0,14 0,11 0,09 0,07 0,09april 0,16 0,25 0,43 0,19 0,41 0,34 0,07 0,26maj 0,33 0,41 0,32 0,29 0,35 0,30 0,22 0,19jun -0,07 -0,05 0,16 0,15 0,12 0,18 0,12 0,11jul 0,62 0,64 0,60 0,47 0,60 0,50 0,63 0,40avgust 0,40 0,14 0,20 0,29 0,42 0,48 0,41 0,36septembar 0,05 -0,01 0,15 -0,01 0,13 0,12 0,20 -0,06oktobar -0,04 0,00 -0,04 -0,05 -0,14 0,04 -0,05 -0,01mar-apr 0,10 0,18 0,40 0,21 0,34 0,28 0,09 0,22mar-maj 0,27 0,37 0,47 0,32 0,44 0,37 0,19 0,27mar-jun 0,15 0,23 0,40 0,29 0,36 0,34 0,19 0,24mar-jul 0,44 0,52 0,65 0,49 0,62 0,55 0,48 0,41mar-avg 0,51 0,50 0,65 0,53 0,69 0,64 0,55 0,48mar-sep 0,46 0,43 0,60 0,45 0,62 0,59 0,53 0,40mar-okt 0,46 0,44 0,60 0,45 0,60 0,61 0,53 0,40apr-maj 0,32 0,42 0,46 0,31 0,47 0,40 0,19 0,27apr-jun 0,17 0,26 0,40 0,29 0,39 0,37 0,19 0,25apr-jul 0,47 0,55 0,65 0,49 0,64 0,57 0,49 0,42apr-avg 0,55 0,53 0,64 0,53 0,70 0,66 0,57 0,49apr-sep 0,48 0,45 0,59 0,45 0,63 0,60 0,54 0,40apr-okt 0,47 0,45 0,58 0,43 0,59 0,61 0,53 0,40maj-jun 0,15 0,21 0,32 0,29 0,31 0,32 0,22 0,20maj-jul 0,52 0,58 0,63 0,52 0,63 0,57 0,58 0,41maj-avg 0,58 0,53 0,60 0,55 0,68 0,65 0,63 0,48maj-sep 0,50 0,44 0,55 0,45 0,60 0,58 0,59 0,38maj-okt 0,48 0,43 0,53 0,43 0,55 0,59 0,57 0,37jun-jul 0,41 0,44 0,56 0,45 0,54 0,49 0,55 0,37jun-avg 0,49 0,41 0,52 0,48 0,59 0,59 0,60 0,44jun-sep 0,41 0,32 0,47 0,38 0,52 0,51 0,55 0,34jun-okt 0,38 0,31 0,44 0,34 0,45 0,51 0,51 0,32jul-avg 0,65 0,53 0,54 0,49 0,65 0,60 0,66 0,47jul-sep 0,57 0,44 0,52 0,41 0,60 0,56 0,65 0,37jul-okt 0,49 0,39 0,44 0,34 0,47 0,51 0,55 0,33avg-sep 0,31 0,09 0,24 0,19 0,37 0,41 0,42 0,21avg-okt 0,24 0,08 0,19 0,13 0,23 0,38 0,33 0,17sep-okt 0,00 -0,01 0,07 -0,05 -0,02 0,11 0,10 -0,05

Marković T, Jovanović M212
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 207-212
Zaključak
Istraživanje međusobne uslovljenosti zavisnog i nezavisnog faktora olakšava konstruisanje vre-menskih derivata koji se baziraju na različitim vremenskim parametrima. Primenom vremenskih derivata najčešće se osiguravaju oni usevi i plodo-vi koji dominiraju u strukturi setve ili se ističu po visini ostvarenog prinosa. Da bi se uspešno mogli upotrebiti vremenski derivati, neophodno je uvažiti ove specifi čnosti vezane za uticaj količine padavina na visinu prinosa. Posmatrajući proizvodnju pšeni-ce evidentno je da količina padavina u aprilu i maju ima relativno visok uticaj na formiranje prinosa, dok je kod kukuruza to višemesečni period od apri-la do avgusta, pa se zato ove činjenice trebaju uzeti u obzir prilikom istraživanja mogućnosti primene vremenskih derivata u našoj poljoprivredi.
Literatura
Alaton P, Djehiche B, Stillberger D (2002): On Modelling and Pricing Weather Derivatives. Appl. Math. Finance 9: 1-20
Berg E (2002): Das System der Ernte- und Einkommensversi-cherungen in den USA - Ein Modell für Europa? Ber. Lan-dwirtsch. 80: 94-133
Bošnjak Đ (2004): Suša i njen odnos prema ratarskoj proizi-vodnji u Vojvodini. Zbornik radova Instituta za ratarstvo i povrtarstvo 40: 45-55
Breustedt G (2003): Subventionen für landwirtschaftliche Ein-kommensversicherungen - Nützlich und notwendig? [Elek-tronski izvor]. [8 str] dostupno na adresi http://www.uni-ho-henheim.de/i410b/download/gewisola/papers/breustedt.
pdf (citirano 10.03.2003, verifi kovano 06.07.2010). Proc 43. Jahrestagung der Gesellschaft für Wirtschafts- und Sozial-wissenschaften des Landbaues, Stuttgart, Deutschland
Campbell S, Diebold F (2002): Weather Forecasting for We-ather Derivatives. [Elektronski izvor]. [21 str] dostupno na adresi http://www.ifk-cfs.de (citirano 02.01.2004, verifi ko-vano 08.07.2010). Wharton Financial Institutions Center, University of Pennsylvania, Philadelphia, USA
Denčić S, Kobiljski B, Mladenov N, Pržulj N (2009): Proizvod-nja, prinosi i potreba za pšenicom u svetu i kod nas. Zbornik radova Instituta za ratarstvo i povrtarstvo 46: 367-377
Hartung J (1998): Statistik: Lehr- und Handbuch der angewand-ten Statistik, Oldenbourg Verlag, München, 73-74
Marinković B, Crnobarac J, Jaćimović G, Marinković D (2008): Tehnologija gajenja u funkciji optimalnog prinosa, prilagođena godini, njivi i hibridu/sorti. Zbornik radova In-stituta za ratarstvo i povrtarstvo 45: 159-178
Marković T, Jovanović M (2011): Smanjenje rizika u proizvodnji pšenice primenom vremenske prodajne opcije. Ratar. Povrt. / Field Veg. Crop Res. 48 (rad u štampi)
Mußhoff O, Odening M, Xu, W (2005): Zur Reduzierung nie-derschlagsbedingter Produktionsrisiken mit Wetterderivaten. In: Wirtschafts- und Sozial-wissenschaftliche Fachgebiete der Landwirtschaftlich-Gärtnerischen Fakultät der Hum-boldt-Universität 73, Working paper, 1-17
Pejić B, Maksimović L, Milić S, Simić D, Miletaški B (2010): Effect of readily available water defi cit in soil on maize yield and evapotranspiration. Ratar. Povrt. / Field Veg. Crop Res. 47: 115-121
Schmitz B (2007): Wetterderivate als Instrument im Risiko-management landwirtschaftlicher Betriebe. Doktorarbeit im wissenschaftlichen Studiengang Agrarwissenschaften an der Rheinischen Friedrich-Wilhelms-Universität zu Bonn, Institut für Lebensmittel- und Ressourcenökonomik, Bonn, 67-68
Vedenov D V, Barnett B J (2004): Effi ciency of Weather De-rivatives as Primary Crop Insurance Instruments. J. Agric. Resour. Econom. 29: 387-403
Infl uence of Rainfall on Wheat and Corn Yield as a Production-Related Basis Risk
Todor Marković · Milenko Jovanović
Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
Summary: In this paper the mutual interdependence of rainfall and yield of wheat and corn in Vojvodina in the period from 1975 to 2005 is analysed by applying the regression and correlation analysis. The results show that the amount of rainfall in April and May (wheat) and in the period from April to August (corn) has an important impact on yields height of these crops. If we want to use the weather derivatives, as new fi nancial instruments in wheat and corn production, it is necessary to take this fact into account, since successful application of weather derivatives is based on reduction of the production-related basis risk that arises when there is a weaker correlation between weather parameters and yield of certain crops.Key words: corn, production-related basis risk, rainfall, regression and correlation analysis, weather derivatives, wheat, yield

Ekonomski aspekti proizvodnje suncokreta u Srbiji 213
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218originalni naučni rad / original research article
Ekonomski aspektiproizvodnje suncokreta u Srbiji
Marija Knežević · Rade Popović
primljeno / received: 08.11.2010. prihvaćeno / accepted: 29.11.2010.© 2011 IFVC
Izvod: Suncokret se u najvećem obimu koristi za proizvodnju ulja i suncokretove sačme. S obzirom da više od 80% vrednosti u preradi suncokreta dolazi od ulja, nivo proizvodnje i cene suncokreta determi-nisane su tražnjom i cenom suncokretovog ulja (FAO 2010a). Uprkos višoj svetskoj ceni suncokretovog ulja u odnosu na druga ulja, tražnja za ovom kulturom raste. Proizvodnja suncokreta u Srbiji značajno je oscilirala u prethodnoj deceniji. Tome su doprineli nepovoljni klimatski uslovi u nekoliko godina, kao i oscilacije u površinama zasejanim ovom kulturom. Prema prosečno ostvarenim prinosima u proi-zvodnji suncokreta, Srbija zauzima sedmo mesto u svetu. U ovom radu su istraživani ekonomski aspekti proizvodnje suncokreta primenom ekonomskih kalkulacija na tri anketirana poljoprivredna gazdinstva različitih kapaciteta proizvodnje. Imajući u vidu trend rasta proizvodnje biogoriva, rast prehrambenih potreba i sve veću upotrebu biljnih u odnosu na masti životinjskog porekla, proizvodnja suncokreta predstavlja značajnu šansu za poljoprivredne proizvođače u Srbiji. Ključne reči: ekonomska kalkulacija, profi tabilnost, suncokret, tržište
M. Knežević · R. Popović Ekonomski fakultet u Subotici, Univerzitet u Novom Sadu, Segedinski put 9-11, 24000 Subotica, Srbijae-mail: [email protected]
Uvod
Proizvodnja suncokreta i suncokretovog ulja predstavlja veoma važan segment svetskog tržišta uljarica. Uprkos višim troškovima proizvodnje suncokretovog ulja u odnosu na troškove proi-zvodnje ostalih ulja, tražnja za ovom kulturom raste (APK Inform & AGRO–T.E.C.E.I. Consul-ting 2009). Suncokret predstavlja značajan izvor biljnog ulja, a uzgaja se u većini delova sveta zbog veoma kratkog perioda vegetacije (FAO 2010a). U svetu se uzgajaju dva osnovna tipa suncokreta: (i) uljani tip namenjen proizvodnji biljnog ulja i (ii) konzumni tip suncokreta namenjen ishra-ni ljudi i ptica (NDSU 2007). Cilj istraživanja u ovom radu predstavlja identifi kovanje osnovnih ekonomskih aspekata proizvodnje suncokreta na nacionalnom tržištu.
Materijal i metod rada
Istraživanje ekonomskih aspekata proizvodnje suncokreta u Srbiji izvršeno je u dva segmenta. U prvom delu analizirani su trendovi uljanih kultura
na svetskom tržištu kao i trendovi proizvodnje suncokreta na tržištu Srbije, dok je u drugom delu analizirana profi tabilnost proizvodnje suncokreta na tri anketirana poljoprivredna gazdinstva veliči-ne od 4,5 ha, 20 ha i 400 ha sa površinama pod suncokretom od 0,87 ha, 10 ha i 97 h u 2010. godini. Statistički podaci prikupljeni su iz slede-ćih izvora: Republički zavod za statistiku Srbije (RZS), Bilten poljoprivrede (2009) i Svetska or-ganizacija za hranu i poljoprivredu (FAO). U radu su korišćeni metodi ankete i polu-strukturiranog intervjua s poljoprivrednicima koji se bave proi-zvodnjom ove kulture. Prilikom izrade kalkulacija korišćen je ekonomski koncept troškova, koji za razliku od računovodstvenog pored fi ksnih i vari-jabilnih troškova, uključuje i pet kategorija opor-tunitetnih troškova.
Rezultati i diskusija
Suncokret predstavlja jednu od deset najzna-čajnijih uljarica na svetu. Sa proizvodnjom od 32 miliona t (FAO) u 2009. suncokret se nalazi na 9. mestu. Suncokretovo ulje učestvuje sa 8% u svet-skoj potrošnji biljnih ulja i sa 10% u prehrambe-noj potrošnji (APK Inform & AGRO–T.E.C.E.I. Consulting 2009). Proizvodnja suncokreta u sve-tu ima trend rasta po prosečnoj godišnjoj stopi rasta od 2,8% (Sl. 1). U periodu 1999-2008. proi-
Ekonomska analiza / Economic Analysisw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Knežević M, Popović R214
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
zvodnja suncokreta je porasla za 20%, dok je pro-izvodnja i potrošnja suncokretovog ulja porasla za 25% (FAO 2010b). Nakon toga u 2009. zbog loših klimatskih uslova i pada proizvodnje u vo-dećim proizvođačima suncokreta (Rusija za 13%, Ukrajina za 7%), usledio je blagi pad proizvod-nje suncokreta za 2% (USDA 2010). Prema FAO
prognozama za 2010-2011. očekuje se oporavak proizvodnje suncokreta (FAO 2010b).
Pored regiona EU koji ima značajan udeo kako u proizvodnji (21,73%) tako i u površinama pod ovom kulturom (16,29%), među deset najvećih proizvođača suncokreta na svetu nalaze se: Ru-sija, Ukrajina, Argentina, Francuska, Kina, SAD, Bugarska, Mađarska, Rumunija i Turska (Sl. 2. i Sl. 3). Njihov ukupan udeo u proizvodnji sunco-kreta u 2009. iznosio je 77,37%. Rusija, Ukrajina
i Argentina su tri najveća svetska proizvođača suncokreta (sunfl ower triangle). Svaka promena u obimu proizvodnje neke od ove tri zemlje ima veliki uticaj na svetsko tržište suncokreta (FAO 2010b).
U posmatranom periodu prosečni prinosi u svetu kretali su se u rasponu od 1.148 kg ha-1 do
maksimalnih 1.436 kg ha-1 ostvarenih u 2008. Prema FAO podacima u 2009. tri zemlje sa naj-većim prosečnim prinosima su Švajcarska (3.202 kg ha-1), Izrael (3.033 kg ha-1) i Hrvatska (3.000 kg ha-1). Srbija se u 2009. nalazila na sedmom mestu u svetu sa ostvarenih 2.400 kg ha-1.
Poput tržišta mnogih drugih poljoprivrednih proizvoda, globalna fi nansijska kriza ozbiljno je uticala na poljoprivrednike, trgovce i prerađiva-če uljarica kroz nižu likvidnost i porast troškova
Sl. 1. Proizvodnja, površine, prosečni prinosi i trend proizvodnje suncokreta u svetu Fig. 1. Production, harvested areas, average yield and trend of world’s sunfl ower production
y = 774.588,38x + 23.524.701,89R2 = 0,38
0
5.000.000
10.000.000
15.000.000
20.000.000
25.000.000
30.000.000
35.000.000
40.000.000
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009
Pov
ršin
a&pr
oizv
odnj
a/A
rea&
prod
uctio
n(h
a) (
t)
01002003004005006007008009001.0001.1001.2001.3001.4001.500
Prin
os /
Yie
ld (k
g/ha
)
Proizvodnja/Production (t) Površina/Area (ha)Prinos/Yield (kg/ha) Linear (Proizvodnja/Production (t))
Graf. 2Udeo najvećih proizvođača za period 1999-2003. god.
Fig. 2 Share of the largest producers in 1999-2003.
EU; 22%
Rusija; 15%
Ukrajina; 13%
Argentina; 19%
Francuska; 7%
Kina; 7%
SAD; 6%
Bugarska; 2%
Rumunija; 4% Turska; 3%Mađarska;
3%
Graf. 3 Udeo najvećih proizvođača u 2009. godini
Fig. 3 Share of the largest producers in 2009.
EU; 22%
Rusija; 20%
Ukrajina; 20%
Argentina; 8%
Francuska; 5%
Kina; 5%
SAD; 4%
Bugarska; 4%
Rumunija; 3%
Turska; 3%Mađarska;
4%
Sl. 2. Udeo najvećih proizvođača za period 1999-2003.Fig. 2. Share of the largest producers in 1999-2003.
Sl. 3. Udeo najvećih proizvođača u 2009.Fig. 3. Share of the largest producers in 2009.

Ekonomski aspekti proizvodnje suncokreta u Srbiji 215
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
fi nansiranja. Zbog lošeg roda suncokreta u 2007. svetska cena suncokreta je postigla ekstremno visok nivo u proleće 2008. Generalno, u ovom periodu se desio globalni rast cena gotovo svih poljoprivrednih proizvoda. Cena suncokreta na međunarodnom tržištu je u drugoj polovini 2008. počela da opada, nakon oporavka biljne proi-zvodnje (FAO 2010b). Rast cena uljarica otpočeo je krajem 2006. i nastavio se tokom cele 2007. i u prvoj polovini 2008. Najviši nivoi cena zabeleženi su u periodu jun-jul 2008. kada su dostigli goto-vo 2,5 puta višu vrednost u odnosu na prosek iz perioda 2002-2004. U decembru 2008. cene ulja-rica su značajno opale, da bi nakon toga došlo do blagog oporavka cena. Međutim, najavljeni rast cena uljarica u drugoj polovini 2009. je izostao, što bi se moglo objasniti oporavkom svetske pro-izvodnje u 2009. kao i daljim smanjivanjem zaliha (Popović i sar. 2009). Nivo cena uljarica iz 2009. se uz manja kolebanja održao i tokom 2010.
Zahvaljujući povoljnijim reljefnim i klimatskim uslovima, proizvodnja suncokreta u Srbiji zastu-pljenija je u ravničarskom području. Tako je u 2009. udeo AP Vojvodine u ukupnoj proizvodnji suncokreta iznosio 92,77%. Proizvodnja sunco-kreta u Srbiji se uglavnom obavlja na porodičnim
poljoprivrednim gazdinstvima. Prema podacima iz Poljoprivrednog biltena za 2009. ukupna proi-zvodnja suncokreta u privrednim društvima i ze-
GodinaYear
Proizvodnja Production
(t)
Požnjevena površina
Harvested area(ha)
PrinosYield
(kg ha-1)
1999. 272.550 183.768 1.4832000. 217.608 146.415 1.4862001. 317.878 163.155 1.9482002. 279.831 150.183 1.8632003. 353.784 199.361 1.7742004. 437.602 188.696 2.3192005. 350.762 197.843 1.7732006. 384.945 186.431 2.0652007. 294.502 154.793 1.9032008. 454.282 187.822 2.4192009. 377.549 157.337 2.400
Tabela 1. Proizvodnja, požnjevena površina i prosečan prinos suncokreta u Srbiji Table1. Sunfl ower production, harvested area and average yield in Serbia.
Tabela 2. Osnovni ekonomski indikatori proizvodnje suncokreta na tri poljoprivredna gazdinstva, sa područja Zapadnobačkog okruga u 2009/2010. proizvodnoj godini (u dinarima)Table 2. Basic economic indicators on three farms in Zapadnobački District in production season 2010 (in RSD)
Pokazatelj /Element PG 1 PG 2 PG 3Površina pod suncokretom/ Area under sunfl ower (ha) 0,87 10 97Prosečan prinos / Average yield (kg ha-1) 3.130 3.600 3.300Utrošak mineralnog đubriva / Used fertilizer (kg ha-1) 150 350 260Prodajna cena / Market price (RSD kg-1) 38 39 38Prosečan trošak / Average cost (RSD kg-1) 20,53 19,87 23,79Prosečan prihod / Average revenue (RSD kg-1) 38,00 42,89 39,06A. Prihod /Revenue
Zrno suncokreta / Sunfl ower seed1. 118.940 140.400 125.400Subvencije direktne / Subsidies per ha2. 14.000 3.500
Ukupan prihod / Total Revenue 118.940 154.400 128.900B. Trošak /Cost
Trošak materijala / Supplies cost1. 17.658 28.066 26.525Seme / Seed 4.208 4.860 4.837Mineralno đubrivo / Mineral fertilizers 4.500 11.050 8.651Pesticidi / Pesticides 1.600 4.281 5.037Dizel gorivo / Diesel 7.350 7.875 8.000
2. Trošak rada / Labour cost 3.000 3.600 6.0003. Trošak angažovanja mehanizacije / Machinery cost 21.373 17.518 15.118
4. Kamata na sopstveni operativni / Interest on own capital 839 1.333 1.2605. Zakup zemljišta / Rent for land 19.100 17.000 19.6006. Opšti troškovi (porez, itd.) / Indirect costs (taxes, etc.) 2.300 4.000 10.000
Ukupan trošak / Total Cost 64.270 71.517 78.503C. Preduzetnički profi t / Entrepreneur’s profi t 54.670 82.883 50.397Prag profi tabilnosti / Break-even price (kg ha-1) 1.691 1.834 2.066

Knežević M, Popović R216
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
mljoradničkim zadrugama iznosila je 20,5% dok je proizvodnja suncokreta na porodičnim gazdin-stvima iznosila 79,5%.
Na osnovu podataka prikazanih u tabeli 1. može se zaključiti da je maksimalan prinos od 2.419 kg ha-1 ostvaren u 2008. nakon čega je usle-dio blagi pad u 2009. kada je Srbija zauzimala sedmu poziciju u svetu.
Kako bi se sagledali ekonomski aspekti pro-izvodnje suncokreta izrađene su kalkulacije na poljoprivrednim gazdinstvima različite veličine (Tab. 2). Ekonomske kalkulacije linija biljne proizvodnje predstavljaju obračunski postupak kojim se utvrđuju tri elementa: prihod, troško-vi i profi t po 1 ha. Vremenski period za koji se izrađuje kalkulacija određena je dužinom proi-zvodnog procesa biljne proizvodnje. S obzirom da se u ovom slučaju radi o ekonomskim kal-kulacijama, neophodno je ispravno protumačiti preduzetnički profi t. Za razliku od računovod-stvenog, ekonomski koncept troškova u struk-turu ukupnih troškova uključuje i oportunitetne troškove. U oportunitetne troškove najčešće se ubrajaju: trošak rada vlasnika, kamata na sop-stveni operativni kapital, kamata na sopstveni dugoročno investiran kapital, renta za sopstve-no zemljište kao i nadoknada za menadžmet. Međutim, kako je nadoknadu za menadžment teško proceniti, ista se u kalkulaciju ne uklju-čuje, a razlika prihoda i troškova u tom slučaju predstavlja preduzetnički profi t. Preduzetnički profi t je zbir nadoknade za menadžment i eko-nomskog profi ta. Ekonomski profi t opravdava investirani kapital samo u slučaju ako je jednak ili veći od nule (Kay et al. 2008).
Pored prihoda od prodaje semena suncokreta, strukturu prihoda čine i subvencije u vidu regre-sa na repromaterijal po 1 ha do ukupnih 100 ha proizvodnje po poljoprivrednom gazdinstvu. Ostvareni prosečni prinosi anketiranih poljopri-vrednih gazdinstava sa područja Zapadnobačkog okruga bili su u rasponu od 3.130 kg ha-1 do 3.600 kg ha-1. S obzirom na male razlike u postignu-toj tržišnoj ceni, prinosi su determinisali razlike u prihodima. Poljoprivredno gazdinstvo veličine 4,5 ha u posmatranoj godini nije ostvarilo sub-vencije, s obzirom da vlasnik nije osiguranik PIO Fonda. Poljoprivredno gazdinstvo sa 400 ha (PG 3) ostvarilo je pravo na subvencije za ukupno 100 ha. Alociranjem ovog iznosa na ukupnu raspolo-živu obradivu površinu dobija se iznos od 3.500 dinara po ha.
Troškovi u proizvodnji suncokreta obuhvata-ju ekonomske troškove svih angažovanih faktora proizvodnje. Pojedinačno, za PG 2 i PG 3 naj-značajniji su troškovi materijala koji u strukturi
ukupnih troškova učestvuju sa 39,24% (PG 2) i 33,79% (PG 3). Kod PG 1 najznačajniji su troš-kovi angažovanja mehanizacije koji u ukupnim troškovima učestvuju sa 33,26%.
Troškovi angažovanja mehanizacije obračunati su na bazi cenovnika mašinskih usluga Zadruž-nog saveza Vojvodine za 2010. U slučaju PG 2 i PG 3 cene pojedinačnih usluga mehanizacije umanjene su za troškove rada koji su posebno iskazani. Udeo troškova mehanizacije na liniji proizvodnje suncokreta u ukupnim troškovima variraju od 19,26% (PG3) do 33,26% (PG1). U kalkulacijama su obuhvaćeni svi oportunitetni troškovi, s tim da su posebno iskazani jedino kamata na sopstveni operativni kapital i renta na obradivo zemljište u vlasništvu. Iskazani predu-zetnički profi t je zbir ekonomskog profi ta i na-doknade za menadžment. Nadoknada za menad-žment kao vrsta oportunitetnog troška najčešće se ne iskazuje posebno zbog poteškoća u utvrđi-vanju visine troška.
Kod sva tri poljoprivredna gazdinstva je utvr-đeno je da su ostvarila preduzetnički profi t. Naj-veći profi t na liniji proizvodnje suncokreta ostva-rilo je PG 2. Takođe, sva tri gazdinstva ostvarila su prosečne prinose iznad kritičnog nivoa (PG1 1.691 kg ha-1, PG2 1.834 kg ha-1, PG3 2.066 kg ha-1), što znači da bi uz datu prodajnu cenu sva tri poljoprivredna gazdinstva ostvarila preduzetnički profi t i uz nešto niže prinose.
Zaključak
Rast proizvodnje bio-goriva, rast prehrambenih potreba i povećana upotreba biljnih u odnosu na masti životinjskog porekla uslovili su rast svetske tražnje za uljaricama. Uprkos padu svetske pro-izvodnje suncokreta od 2% u 2009. zbog loših klimatskih uslova, u 2010-2011. očekuje se opo-ravak proizvodnje. Rast proizvodnje suncokreta u najvećoj meri određen je cenom i tražnjom za uljem. Prema dugoročnim procenama FAO i OECD, proizvodnja biljnog ulja će se povećati sa 112 miliona t iz 2008. na 155 miliona t u 2018. Procenjeno je da će do 2018. Rusija povećati pro-izvodnju biljnog ulja (uglavnom suncokretovog) za 28% a Ukrajina za 45% što će usloviti dalji rast proizvodnje suncokreta.
Ekonomski rezultati u proizvodnji suncokre-ta praćeni su na tri poljoprivredna gazdinstva sa područja Zapadnobačkog okruga u 2010. proi-zvodnoj godini. Preduzetnički profi t je ostvaren kod svih gazdinstava u značajnom obimu. Izvori visoke profi tabilnosti u proizvodnji suncokreta proizilaze prvenstveno sa strane prihoda. U na-rednim periodima, odluka da li će se poljopri-

Ekonomski aspekti proizvodnje suncokreta u Srbiji 217
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
vrednici odlučivati za proizvodnju suncokreta zavisiće od očekivanog nivoa profi tabilnosti i od budućih kretanja cena suncokretovog ulja i suncokreta. Kako se u daljem periodu očekuje rast tražnje za suncokretovim uljem, a imajući u vidu povoljne klimatske uslove, dugu tradiciju u proizvodnji i značajne prosečne prinose, u Srbiji postoje preduslovi da poljoprivredni proizvođači u ravničarskom proizvodnom području Srbije u većem obimu uključe suncokret u strukturu ra-tarske proizvodnje.
Literatura
APK Inform & AGRO–T.E.C.E.I. Consulting (2009): Sun-fl ower triangle: Argentina, Russia, Ukraine, Competitive Environment, Investing Opportunities and Prospect for 2009-2011. [Elektronski izvor]. Dostupno na: www.agrimar-ket.info/fi les/Sunfl owerTriangle.pdf (citirano: 28.10.2010.), APK Inform & AGRO–T.E.C.E.I. Consulting
FAO (2010a): Agribusiness Handbook, Sunfl ower Crude and Refi ned Oils [Elektronski izvor]. Dostupno na adre-si www.fao.org/docrep/012/al375e/al375e.pdf (citirano 24.10.2010.), FAO, Rome, Italy
FAO (2010b): Food Outlook Global Market Analysis [Elektron-ski izvor]. Dostupno na adresi www.fao.org/docrep/012/
ak349e/ak349e00.pdf (citrano 24.10.2010.), FAO, Rome, Italy
FAO (2010c): Baza podataka poljoprivrede [Elektronski izvor]. Dostupno na adresi: http//faostat.fao.org/site/567/default.aspx#ancor (citirano 28.10.2010.), FAO, Rome, Italy.
Kay R, Edwards W, Duffy P (2008): Farm management, 6st edi-tion, Mc Graw – Hill, New York
NDSU (2007): Sunfl ower production [Elektronski izvor]. Dostupno na adresi : www.ag.ndsu.edu/pubs/plantsci/rowcrops/a1331intro.pdf (citirano 13.10.2010.), North Da-kota State University
Popović R, Stojšin V, Stantić M, Knežević M, Štavljanin B (2010): Ekonomski aspekti proizvodnje uljane repice u Srbiji. Ratar. Povrt. 47: 179-185
Radić P (2010): Cenovnik mašinskih usluga, Zadružni savez Vojvodine, Novi Sad
Republički zavod za statistiku (2010a): Baza podataka poljo-privrede [Elektronski izvor]. Dostupno na adresi: http://webrzs.stat.gov.rs/axd/poljoprivreda/index1.php?ind=1 (citirano 28.10.2010.), RZS, Beograd
Republički zavod za statistiku (2010b): Poljoprivreda 2009. [Elektronski izvor]. Dostupno na adresi: http://pod2.stat.gov.rs/ObjavljenePublikacije/G2010/PdfE/G20105523.pdf (citirano 28.10.2010.), RZS, Beograd
USDA (2009): Global crop production rewiev, Prepared by USDA’s Joint Agricultural Weather Facility. [Elektronski izvor]. Dostupno na adresi: www.usda.gov/oce/weather/pubs/annual/cropproduction.pdf (citirano 29.10.2010.), USDA

Knežević M, Popović R218
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 213-218
Economics of Sunfl ower Production in Serbia
Marija Knežević · Rade Popović
Faculty of Economy in Subotica, University of Novi Sad, Segedinski put 9-11, 24000 Subotica, Serbia
Summary: Sunfl ower seed is mostly processed into oil and meal. However, more than 80% of sunfl ower’s value comes from oil, as the level of sunfl ower price and production are determined by the level of demand and the price of sunfl ower oil. Even despite the high cost of sunfl ower oil, demand for this crop is still increasing worldwide. In the last few years, sunfl ower production was unstable in Serbia. It was caused by unfavourable climate condition for few years and varying areas planted with this crop. According to average yields in sunfl ower production, Serbia is the seventh in the world. This paper considers the economic aspects of sunfl ower production, using the method of enterprise budget on three sampled farms of different sizes. Taking into account increasing bio-diesel production, increasing food demands and consumers’ nutritional preferences, the production of sunfl ower could be an impor-tant opportunity for agricultural producers in Serbia. Key words: enterprise budget, market, profi tability, sunfl ower

Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar, Serbia 219
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226original research article / originalni naučni rad
Pisum & Ervilia Tetovac - Made in Early IronAge Leskovac. Part One.
Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar in Leskovac, South Serbia
Aleksandar Medović ∙ Aleksandar Mikić ∙ Branko Ćupina · Živko Jovanović ∙ Svetlana Radović ∙ Aleksandra Nikolić · Nemanja Stanisavljević
received / primljeno: 13.10.2010. revised / prerađeno: 29.12.2010. accepted / prihvaćeno: 03.01.2011.© 2011 IFVC
Summary: A lucky fi nd of 2,572 charred pea (Pisum sativum L.) seeds in a single archaeobotanical sam-ple from the hill fort settlement Hissar near Leskovac represents a unique example in Bronze / Iron Age research in South East Europe. Another mass storage of bitter vetch (Vicia ervilia (L.) Willd.) from the same site and period (Brnjica cultural group, beginning of the Iron Age in the Morava valley) confi rms a long tradition of the city of Leskovac region as a pulse crop production centre in modern Serbia. Both pulse storages were almost pure with small amount of admixture from other crops, mainly cereals, ot-her pulses, such as lentil (Lens culinaris Medik.) and broad bean (Vicia faba L.), and oil/fi bre plants. Both pulses were cultivated as main crops and were stored separately. Key words: 12th cent. B.C., archaeobotany, Brnjica cultural group, charred bitter vetch storage, charred pea storage, Hissar, Leskovac, South Serbia
A. Medović Museum of Vojvodina, Dunavska 35, 21000 Novi Sad, Serbiae-mail: [email protected]. MikićInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, SerbiaB. ĆupinaFaculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, SerbiaŽ. Jovanović · A. Nikolić · N. StanisavljevićInstitute of Molecular Genetics and Genetic Engineering, University of Belgrade, Vojvode Stepe 444a, p.fah 23, 11010 Belgrade, Serbia S. RadovićFaculty of Biology, University of Belgrade, Studentski trg 16, 11000 Belgrade, Serbia
Introduction
The fortifi ed hill fort settlement Hissar in Leskovac, with the name derived from Turkish hissar, denoting fortress, is a multilevel settlement of the Brnjica cultural group, 1,350-1,000 B.C., Iron Age I in the Morava valley (Stojić et al. 2007).
The Hissar hill (341 m alt.) is in a strategic position over the confl uence of Jablanica and Veternica rivers in river South Morava and over the greatest part of the valley of Leskovac. The valley is 50 km long and 45 km wide. The archaeological excavations at Hissar began in 1999. In the campaign of the year 2005, two
rich pulse-crop samples were gathered from the deposits of the Brnjica II a-level from 12th cent. B.C. Out of 7 litres of earth substrate, a total number of 2,572 charred seeds of pea (Pisum sativum L.) and 3,031 charred seeds of bitter vetch (Vicia ervilia (L.) Willd.) were yielded through fl otation, respectively. In the previous archaeobotanical report (Medović 2005), only one uncertain record of pea from the deposits of Brnjica cultural group was made, whereas 37 charred items of bitter vetch in every fourth sample were found. Beside these two species, the inventory of the pulse crops from Hissar supplements fi ndings of broad bean (Vicia faba L.) and lentil (Lens culinaris Medik.).
Materials and Methods
The round seeds of many pulse crops cause considerable diffi culties for a precise archaeobotanical determination. This is the reason why normally the so-called unidentifi ed
Archaeobotany / Arheobotanikaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Acknowledgements: The authors appreciate sincere encouragement and immense enthusiasm by Dr. Gérard Duc, a widely recognised expert in the modern grain legume genetic diversity, in its ancient co-unterpart.

Medović A et al.220
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
pulses-group is proportionally more numerous than Cerealia identerminata-group. Chances of carbonization of pulses and cereals in archaeological context are not equal. There are differences in chemical composition, such as high protein content in pulses vs. high starch content in cereals, as well as in processing and usage of this two crop groups. Generally, pulses have less chances of getting carbonized than cereals and are therefore underrepresented in archaeobotanical records. Mass storages of charred pulse crops, like these two lucky fi nds from Hissar, offer a great opportunity of getting a closer look at their cultivation, crop processing and storage methods in prehistory.
Results and Discussion
Pea Pea is one of the fi rst crops cultivated by
humans. It is nearly impossible to determine a charred pea seed if a characteristic coffee bean shaped hilum is missing (Fig. 1). The seed-shape (spherical with some fl attenings, depending on their position and space within a pod) as well as its size are not enough to say whether we are dealing with pea or not. Pea seeds can be as small as some Vicia seeds and without preserved hilum they cannot be distinguished from them. In the Hissar sample,
only few pea seeds maintained almost intact seed coat (testa) with hilum. Luckily, among all the pea seeds, only 32 seeds of lentil, bitter vetch and broad bean were mixed - species whose determination is undoubtful (Table 1). Lentil and bitter vetch seeds differ from pea in shape and from broad bean in size. According to the exclusion principle we can almost be sure that the rest belongs to pea.
Beside pulses, the sample consisted of few charred residues of four cereals: hulled lax eared six-row barley (Hordeum vulgare L. subsp. vulgare), bread wheat (Triticum aestivum L.), emmer (T. dicoccon Schrank) and einkorn wheat (T. monococcum L.). 320 small seeds of common millet (Panicum miliaceum L.) are not enough to change weight percentages in the sample considerably. 20 seeds of fl ax / linseed (Linum usitatissimum L.) and a single one-seeded nutlet of dragons head (cf. Lallemantia iberica Fisch. et C.A. Mey.) represent the taxonomic group of oil and fi bre plants. Recently, archaeobotanical fi nds of dragons head are accumulating: in Serbia, it was recorded at the Bronze and Iron Age settlement of Feudvar (Becker & Kroll 2008) and in the middle Bronze Age level of Židovar (Kišgeci & Medović 2006). Both sites are located in the fertile plain of the Vojvodina region, while this is the fi rst record of this oil plant in hilly and woody central Serbia. However, most data of charred fruits of Lallemantia come from diverse Bronze Age sites in
Figures 1 and 2. A charred seed of pea (Pisum sativum L.) from Hissar, Leskovac, 12th cent. B.C. A drawing by A. Medović and a photographSlike 1 i 2. Ugljenisano seme graška (Pisum sativum L.) sa Hisara kod Leskovca, 12. vek p.n.e. Crtež A. Medovića i fotografi ja

Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar, Serbia 221
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
Table 1. Hissar, Leskovac, South Serbia, 2005, Sample 35. Macrofossil plant remains (charred seeds and one seeded fruits). Brnjica II a, 12th cent. B.C. - r < 0.5%;0.5% < + < 1.0%Tabela 1. Hisar, Leskovac, južna Srbija, 2005, uzorak 35. Makrofoslini ostaci biljaka (ugljenisana semena i jednosemeni plodovi). Brnjica II a, 12. vek p.n.e.
SpeciesVrste
QuantitityKoličina
(No.)
QuantitityKoličina
(%)
MassMasa(mg)
MassMasa(%)
Cereals / Strna žitaHordeum vulgare vulgare 10 r 126 rcf. Triticum aestivum s.l. 1 r 6 rT. aestivum s.l., rachis internodes 1 r 3 rTriticum dicoccon 1 r 11 rTriticum monococcum 1 r 2 rCerealia indeterminata 1 r 8 rMillets / Prosolika žitaPanicum miliaceum 320 10.66 553 +Pulses / VarivaPisum sativum 2572 85.68 62768 97.82Lens culinaris 21 + 113 rVicia faba 8 r 472 +Vicia ervilia 3 r 21 rOil and fi bre plants / Uljane i predivne biljkeLinum usitatissimum 21 + 20 rcf. Lallemantia iberica 1 r 2 rDye plants / Bojene biljkecf. Carthamus tinctorius 1 r 2 rFruits / VoćeSambucus 1 r 0 rWeeds and ruderals / Korovske i ruderalne biljkeConvolvulus arvensis 16 + 40 rEchinochloa crus-galli 8 r 2 rFallopia convolvulus 4 r 5 rChenopodium album 3 r 0 rSetaria viridis 2 r 0 rDigitaria sanguinalis 1 r 0 rGalium aparine 1 r 2 rLolium temulentum 1 r 3 rMedicago 1 r 2 rPlant families / Biljne familijeFabaceae 1 r 3 rPoaceae 1 r 0 rTotal / Ukupno 3002 100 64164 100Other fi nds / Ostali nalazi Insecta, larva 1 0Thousand grain weight (g) / Masa hiljadu semenaPisum sativum 24.40Panicum miliaceum 1.73

Medović A et al.222
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
northern Greece (Jones & Valamoti 2005, Becker & Kroll 2008). This plant, “exotic” for the Balkans, was probably introduced from Anatolia to Greece in the early Bronze Age and then cultivated locally (Jones & Valamoti 2005). A fi nd of saffl ower (cf. Carthamus tinctorius L.) indicates the possible use of its fl owers to dye food yellow and textile red to brown or even black. Other fi nds in the sample represent common weeds in millets and wheat. Although 24 taxa were found in this sample including three other pulse crops, pea is the dominant one, with a weight share of almost 98%. This indicates that pea was cultivated as one of the main crops and was stored separately from the other pulse crops. The thousand grain weight of these pea seeds is 24.4 g. Two thousand archaeobotanical samples from Feudvar yielded 3,695 Pisum records (Kroll 1998), on average 1.85 seeds per sample. The thousand grain weight of pea seeds in Feudvar was calculated in only one sample, 19.25 g (H. Kroll, pers. comm.) and it was 5 g lighter than the one from Hissar! At the archaeological sites of Židovar (Serbia) or Kastanas and Agios Mamas (north
Greece), there were not enough peas in material to calculate thousand grain weight. This problem indicates how rare and therefore valuable the fi nd of this pea storage at Hissar truly is.
The moisture requirement for fi eld pea is similar to that for cereal grains (Oelke et al. 1991). Plentiful rains and/or early irrigation and no rain during pod fi lling and ripening is ideal. Pea is intolerant of drought which has a particularly adverse effect on production if it occurs at fl owering. It requires free-draining soils since it is intolerant of water-logging. It can be grown on a wide range of soil types, ranging from light sandy loams to heavy clays, but in any of these soil types there must be good drainage as fi eld pea does not tolerate soggy or water-soaked conditions. The soil pH optimum is 5.5 to 6.5.
Bitter Vetch
A bitter vetch seed can be described as well-rounded tetrahedron (Fig. 2). This feature alone makes them unique among other pulses. The hilum of bitter vetch is typical, but normally it’s
Figures 3 and 4. A charred seed of bitter vetch (Vicia ervilia (L.) Willd.) from Hissar, Leskovac, 12th cent. B.C. A drawing by A. Medović and a photographSlikea 3 i 4. Ugljenisano seme urova (Vicia ervilia (L.) Willd.) sa Hisara kod Leskovca, 12. vek p.n.e. Crtež A. Medovića i fotografi ja

Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar, Serbia 223
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
missing in charred archaeobotanical samples: its hilum is shorter than of hilum from common vetch (Vicia sativa L.) and smaller and narrower than that of pea.
Bitter vetch has been one of the essential pulse crops of the southern Balkans and adjacent areas since Neolithic (Kroll 1991). It is cultivated for its edible seeds, which are fed to livestock or eaten in soups. For human consumption the bitterness of the seeds needs to be removed through leaching by several changes of boiling water or by roasting. The species of the genus Vicia are traditionally used for feeding ruminants, particularly sheep, but they are practically unused for monogastric animals due to the toxicity of their seeds and its negative effect on growth. Bitter vetch must not exceed 25% in the rations for sheep and cattle. It is very resistant to cold, because of its sparse habit and branching and slow growth (López Bellido 1994), as well as to drought, even in spring. A harvest is obtained even during excessively dry years. High grain yields (up to 3 t ha-1) may be obtained in favourable conditions. It is adapted to the regions with wide annual precipitation sums, ranging from 360 mm to 1160 mm, prefers an annual mean temperature of 14°C and is suited to neutral or lightly acid soils, tolerating limy types of soil that are not too clayey.
A rich bitter vetch sample form Hissar differs from the rich Pisum sample only in the occurrence of charred items of spelt wheat (Triticum spelta L.), which is the fi rst record of this wheat at Hissar, and in lack of pea seeds (Table 2). Reaching only 91% bitter vetch weight share is not as high as in the case of pea, the main reason for this being the increased weight share of barley. Nevertheless, the share of other legumes (such as lentil and broad bean) is insignifi cant in the sample. Just like pea from the same site, the bitter vetch from Hissar was cultivated as one of the main crops and stored separately from the other pulse crops. The thousand grain weight of its seeds is 11.74 g. In contrast to pea, it is possible to compare these values with other sites in the region. At Agios Mamas, the 1000 ervilia seeds weigh normally under 10 g (Becker & Kroll 2008), while at Feudvar they weigh as lens seeds, only 6.02 g (H. Kroll, pers. comm.). The weight of bitter vetch from Hissar fi ts into the range values from Kastanas, with 10-13 g (Kroll 1983).
Storage Pests
In the rich ervilia sample from Hissar, at least fi ve seeds had large boreholes in the place of radicle. This fi ts the description made by
H. Kroll (1983) on the charred bitter vetches seeds found at the tell of Kastanas in northern Greece. Rich bitter vetch fi nds from Kastanas were defi led by seed beetles (Brachinae). The infestation was not considerable, only 1-2%. Seed beetles infestation of pulses was also recorded on several archaeological sites in Europe, Middle East and Egypt (Panagiotakopulu 2001). In the rich Pisum sample from Hissar one larva of a holometabolous insect was found. Many seeds were defi led (G. Duc, pers. comm.), possibly by pea beetle (Bruchus pisorum L.). Both bitter vetch and pea storages from Hissar were infested by pests. The history of crops is closely related to history of pests of stored products. With a fair record of pests on archaeological sites, one could be able to reconstruct original routes of their expansion through crop trade in the past.
Soil, Climate and Biodiversity
Geographically speaking, almost two thirds of the area surrounding Hissar are hills and valleys. The rest is fl at, which, in a broader sense, constitutes the so-called Leskovac valley. The majority of soils, represented by various types of brown soils, are of medium-heavy mechanic content and medium porous. As per chemical content, the land is typically acid, while the content of humus is low, with a moderate share of physiologically active phosphorus and potassium. The climate is complex, where the main, typically continental conditions are mingling with the Mediterranean infl uences coming from the south, up the valley of the river Vardar and deeper into the hills and mountains of the south-central Balkans, where Leskovac is situated. A relatively low level of the total annual precipitation sum of between 600 mm and 800 mm represents the most important condition for agriculture.
This region is also extremely rich in plant biodiversity, making a continuum stretching from Asia Minor, over southern Bulgaria to the northern Albania and southern Montenegrin coast, with numerous crop wild relatives such as wild cereals and legumes. Considering the latter, it is only in the south, ranging from Niš, over Leskovac to Vranje, where wild pea (Pisum sativum subsp. elatius (Steven ex M. Bieb.) Asch. & Graebn.) can be found in abundance in the whole Serbia, as well as wild lentil (Lens nigricans (M. Bieb.) Godr.) and numerous wild vetch species (Bojan Zlatković, pers. comm.).
It may be assumed that this specifi c constellation of the favourable soil and climatic conditions on one side and the extreme wealth of

Medović A et al.224
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
wild and agricultural legume fl ora was the main reason why pulses have always been an important component in human diets and animal feeding as both voluminous and concentrated feed. Once, it was pea, lentil, bitter vetch, broad bean, grass pea (Lathyrus sativus L.) and chickpea (Cicer arietinum L.), while today it is common bean (Phaseolus vulgaris L.), gradually replacing the former ones during the last few centuries. At any rate, pulses were and still are there, in Leskovac and its heartland of the Serbian south, equally important as cereals, vegetables, meat and other basic nutrients.
Leskovac and its region is one of the last ancient European grain legume refugia in Serbia, where crops such as bitter vetch have been cultivated for forage and grain until relatively recently (Đorđević 1942) and where others such as broad bean still persist (Mihailović et al. 2010) and vetches are the most popular annual forage crop. Can an echo of the excavated and ringing countless bitter vetch and pea seeds from the past break the walls of oblivion and reintroduce these crops in the Serbian and Balkan agriculture, to the benefi t of Europe and its diverse agro-ecosystems?
Table 2. Hissar, Leskovac, South Serbia, 2005, Sample 36. Macrofossil plant remains (charred seeds and one seeded fruits). Brnjica II a, 12th cent. B.C. - r < 0.5%;0.5% < + < 1.0%Tabela 2. Hisar, Leskovac, južna Srbija, 2005, uzorak 36. Makrofoslini ostaci biljaka (ugljenisana semena i jednosemeni plodovi). Brnjica II a, 12. vek p.n.e.
SpeciesVrste
QuantitityKoličina
(No.)
QuantitityKoličina
(%)
MassMasa(mg)
MassMasa(%)
Cereals / Strna žitaHordeum vulgare vulgare 190 5.66 2683 6.83Triticum monococcum 14 r 116 rT. monococcum, spikelet forks / glume bases 1 r 1 rTriticum dicoccon 6 r 68 rTriticum spelta 5 r 55 rT. spelta, spikelet forks / glume bases 1 r 4 rcf. Triticum aestivum s.l. 4 r 45 rCerealia indeterminata 47 1.40 466 1.19Millets / Prosolika žitaPanicum miliaceum 27 + 42 rPulses / VarivaVicia ervilia 3031 90.24 35589 90.64Lens culinaris 11 r 90 rVicia faba 2 r 75 rWeeds and ruderals / Korovske i ruderalne biljkePolygonum convolvulus 15 r 26 rAsperula arvensis 2 r 2 rChenopodium album 2 r 0 rLactuca 1 r 2 rTotal / Ukupno 3359 100 39264 100Wood charcoal / Ugljenisano drvećeQuercus 6 361Acer 1 53Total / Ukupno 7 414Thousand grain weight (g) / Masa hiljadu semenaVicia ervilia 11.74Mineralised seeds and onseeded fruits / Mineralizovana semena i semeno voćeSambucus ebulus 2 2Total / Ukupno 2 2

Two Charred Pulse Crop Storages of the Fortifi ed Hill Fort Settlement Hissar, Serbia 225
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
Conclusions
Although pea is one of the fi rst crops cultivated by man, a lucky fi nd of 2,572 pea seeds in only one sample from Hissar represents a unique example in the archaeobotany of South East Europe. This amount of pea seeds could only be reached if hundreds of samples at one site are analysed. The reason for this should be seen not only in the fact that pulses generally have less chances of getting carbonized than cereals, and therefore are underrepresented in archaeobotanical record, but also in the diffi cult determination of Pisum seeds. Pea from Hissar was a distinct crop, stored separately from other crops. The status of pea in Hissar’s Early Iron Age economy is disputable. According to its record’s frequency, it seems that pea was cultivated on small patch-like fi elds. High 1000-grain weight of pea, with some restraints, seems to confi rm the intense crop-care on small scaled areas.
On the other hand, the records of bitter vetch at archaeological sites in South East Europe are abundant. One of the reasons may be the fact that its well-rounded tetrahedron seeds are unique and therefore can be recognized easily. Ervilia, lentil and broad bean are most frequent pulse crops at Hissar. As well as pea, bitter vetch was cultivated and stored separately.
The constellation of agro-ecological factors, such as slightly acid soil and steppe-like climate with a low level of the precipitation with drought periods, favours bitter vetch production in the Leskovac valley over pea. Nevertheless, it is suitable for a large-scale cultivation of all pulse crops. Thus this region has become an epitome for the pulse production in modern Serbia, such as in a famous proverb used to describe an extremely tasty, truly delicious and masterfully prepared pulse dish of common bean: Oh, yeah! This one’s been a genuine ‘Bean Tetovac – made in Leskovac!’
References
Becker C, Kroll H (2008): Das Prähistorische Olynth. Ausgra-bungen in der Toumba Agios Mamas 1994-1996. Ernährung und Rohstoffhandel im Wandel. Verlag Marie Leidorf GmbH, Rahden/Westf.
Đorđević V (1942): Gajenje biljaka za stočnu hranu. Srpsko po-ljoprivredno društvo, Belgrade
Jones G, Valamoti S (2005): Lallemantia, an imported or intro-duced oil plant in Bronze Age northern Greece. Veg. Hist. Archaeobot. 14: 571-577
Kišgeci J, Medović A (2006): Prehistoric use of medicinal and aromatic plants in the southeast part of the Pannonian Plain. Proceedings, Third Conference on Medicinal and Aromatic Plants of Southeast European Countries, Belgrade, Serbia, 29-32
Kroll H (1983): Kastanas. Ausgrabungen in einem Siedlun-gshügel der Bronze- und Eisenzeit Makedoniens 1975-1979. Die Pfl anzenfunde. Verlag Volker Spiess, Berlin
Kroll H (1991): Südosteuropa. In: Van Zeist W. et al. (eds.), Pro-gress in Old World Palaeoethnobotany, Balkema, Rotterdam, the Netherlands, pp. 161-177
Kroll H (1998): Die Kultur- und Naturlandschaften des Titeler Plateaus im Spiegel der metallzeitlichen Pfl anzenreste von Feudvar. In: Hänsel B, Medović P (eds.), Feudvar I. Das Pla-teau von Titel und die Šajkaška. Verlag Oetker-Voges, Kiel, Germany, pp. 305-317
López Bellido L (1994): Grain legumes for animal feed. In: Hernándo Bermejo J E, León J (eds.), Neglected Crops: 1492 from a Different Perspective. FAO, Rome, Italy, pp. 273-288
Medović A (2005): O biljnoj ekonomiji gradinskog naselja Hisar kod Leskovca, južno Pomoravlje, sa kraja bronzanog doba (oko 1350-1000 g. pre n.e). Leskov. zb. 45: 201-209
Mihailović V, Mikić A, Vasić M, Ćupina B, Đurić B, Duc G, Stoddard F L, Hauptvogel P (2010): Neglected legume crops of Serbia – Faba bean (Vicia faba). Ratar. Povrt. / Field Veg. Crop Res. 47: 27-32
Oelke E A, Oplinger E S, Hanson C V, Davis D W, Putnam D H, Fuller E I, Rosen C J (1991): Dry Field Pea. Alternative Field Crops Manual. University of Wisconsin Cooperative Extension Service, the University of Minnesota Extension Service and the Center for Alternative Plant and Animal Products
Panagiotakopulu E (2001): New Records for Ancient Pe-sts: Archaeoentomology in Egypt. J. Archaeol. Sci. 28: 1235-1246
Stojić M, Pešić J, Jović S (2007): Kulturna stratigrafi ja arheološ-kog lokaliteta Hisar u Leskovcu. Leskov. zb. 47: 29-40

Medović A et al.226
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 219-226
Grašak i urov Tetovac - made in ranogvozdenodobni Leskovac. Deo prvi.Dve zalihe ugljenisanih zrnastih mahunarki sa gradinskog
naselja Hisar kod Leskovca, južno Pomoravlje
Aleksandar Medović1 ∙ Aleksandar Mikić2 ∙ Branko Ćupina3 • Živko Jovanović4
∙ Svetlana Radović5 ∙ Aleksandra Nikolić4 • Nemanja Stanisavljević4
1Muzej Vojvodine, Dunavska 35, 21000 Novi Sad, Srbija2Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbija
3Poljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbija4Institut za molekularnu genetiku i genetičko inženjerstvo Univerziteta u Beogradu, Vojvode Stepe 444a, p.fah 23,
11010 Beograd, Srbija5Biološki fakultet Univerziteta u Beogradu, Studentski trg 16, 11000 Beograd, Srbija
Izvod: Arheološki lokalitet Hisar je gradinsko naselje Brnjičke kulturne grupe (gvozdeno doba I u Pomoravlju), podignuto na vrhu, odnosno na platou brda Hisar koje kontroliše zonu ušća Jablanice i Veternice u Južnu Moravu i najveći deo Leskovačke kotline. Prilikom arheoloških istraživanja 2005. godine u objektima koji se vezuju za Brnjica II a fazu (12. vek pre nove ere) pronađene su dve velike količine ugljenisanih semena graška (Pisum sativum L.) i urova (Vicia ervilia (L.) Willd.). Zalihe ove dve zrnaste mahunarke bile su skoro čiste, sa malim procentom nečistoća koje su pre svega činili plodovi, pre svega žitarica, drugih mahunarki (sočivo - Lens culinaris Medik. i bob - Vicia faba L.), ali i biljaka uljarica, što ukazuje da su grašak i urov bili odvojeno uzgajani, a potom i odvojeno skladišteni. Ovi nalazi, a pre svega nesvakidašnji nalaz zalihe graška, potvrđuju dugu tradiciju uzgajanja mahunarki u leskovačkom kraju.Ključne reči: 12. vek p.n.e, arheobotanika, Brnjička kulturna grupa, Hisar, južna Srbija, Leskovac, zaliha ugljenisa-nog semena graška, zaliha ugljenisanog semena urova

Extraction of the Ancient DNA from Charred Seeds from Hissar, Serbia 227
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232originalni naučni rad / original research article
Pisum & Ervilia Tetovac - Made in Early IronAge Leskovac. Part Two.
Extraction of the Ancient DNA from Charred Seeds from the site of Hissar in South Serbia
Živko Jovanović ∙ Nemanja Stanisavljević ∙ Aleksandra Nikolić ∙ Aleksandar Medović ∙ Aleksandar Mikić ∙ Svetlana Radović ∙ Vuk Đorđević
received / primljeno: 31.10.2010. revised / prerađeno: 29.12.2010. accepted / prihvaćeno: 04.01.2011.© 2011 IFVC
Summary: The extracts were prepared from the samples of 3,200-year-old charred pea and bitter vetch seeds from the site of Hissar near Leskovac, South Serbia, using two different DNA extraction pro-cedures. We used CTAB method with some modifi cation and obtained low quantity of ancient DNA in comparison with the second method used - commercial available kit. After the extraction, a whole genome amplifi cation using Phi29 DNA polymerase was performed. The amplifi ed DNAs were used for PCR reaction using primers for 26S rDNA gene, which is located on the nuclear genome. The single band corresponding to 26S rDNA fragment from modern relatives was obtained. We conclude that DNA from charred pea and vetch seed can be extracted and used for further archaeobotanical analysis at the molecular level.Key words: ancient DNA, charred pea seed, charred bitter vetch seed, PCR
Ž. Jovanović · N. Stanisavljević ∙ A. NikolićUniversity of Belgrade, Institute of Molecular Genetics and Genetic Engineering, Vojvode Stepe 444a, p. fah 23, 11010 Belgrade, Serbiae-mail: [email protected]. MedovićMuseum of Vojvodina, Dunavska 35, 21000 Novi Sad, SerbiaA. Mikić ∙ V. ĐorđevićInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, SerbiaS. RadovićFaculty of Biology, University of Belgrade, Studentski trg 16, 11000 Belgrade, Serbia
Introduction
After death DNA is degraded by various biotic and abiotic processes, resulting in fragmentation and modifi cation of original sequence information (Schlumbaum et al. 2008). It has emerged that cold, dry and low oxygen environments are benefi cial for DNA survival; for this reason, freshly excavated material is best stored cold or frozen (Burger et al. 1999, Smith et al. 2001, Pruvost et al. 2007). However, these DNA fragments still contain information which may help us to understand agricultural or vegetation history.
Ancient DNA (aDNA) has received much attention since the mid-1980s, where the fi rst sequence of an extinct animal species was
recovered from a museum specimen. Since then, the majority of ancient DNA studies have focused predominantly on animal species, while studies in plant paleogenetics have been rather limited, with the notable exception of cultivated species found in archaeological sites (Gugerli et al. 2005).
The possibility that ancient DNA is preserved in some charred plant remains was fi rst raised by Goloubinoff et al. (1993), who obtained polymerase chain reaction (PCR) products from extracts of 600-year-old maize cobs. There are many data considering general problems and doubts surrounding ancient DNA research (Cooper & Poinar 2000). Some of them are the proof of authenticity, contamination with other ancient and/or modern DNA and reproducibility of obtained results (Schlumbaum et al. 2008).
The procedure for aDNA analysis in plants involves excavation, morphological description, external cleaning if possible, powdering, DNA extraction, amplifi cation of chosen target region within the plant’s genome(s) by PCR, gel electrophoresis to establish presence of correct product size, cloning and sequencing PCR product and verifi cation.
Nuclear ribosomal DNA (rDNA) genes are
Archaeobotany / Arheobotanikaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Jovanović Ž et al.228
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
of interest for aDNA research, as they contain hundreds of unit of two variable spacer regions between the 18S, the 5.8S and the 26S genes, the so-called internal transcribed spacer regions 1 and 2, thus increasing the chance of DNA survival. These loci have been employed for phylogenetic analysis at the genus level in modern plants (Knaak et al. 1990, Soltis et al. 1992, Alvarez & Wendel 2003).
So far, there has been no preferred extraction method for ancient plant material, nor any comprehensive comparison protocols, such as exist for animals (Rohland & Hofreiter 2007). Generally, CTAB/DTAB methods, silica-based methods or DNA extraction kits are employed. All these methods were developed taking into account that plant parts are rich in secondary metabolites and other potential inhibitors of PCR.
Here we described the methods for aDNA isolation from pea (Pisum sativum), one of the fi rst crops cultivated by man, and from bitter vetch (Vicia ervilia), two charred pulse crop storaged of the fortifi ed hill settlement Hissar.
Our preliminary results show that even commercial kits for DNA extraction could be suitable for aDNA extraction from these species, but using some modifi cation.
Materials and Methods
Charred pea (Pisum sativum L.) seeds were found in a single archaeobotanical sample from the hill fort settlement Hissar near Leskovac and represent a unique sample in Bronze/Iron Age research in the South East Europe. Bitter vetch (Vicia ervilia (L.) Willd.) was from the same site and period (Brnjica cultural group, beginning of the Iron Age in the Morava valley). Both pulse storages were almost pure with small amount of admixture from other crops, mainly cereals, other pulses, such as lentil (Lens culinaris Medik.) and faba bean (Vicia faba L.).
The DNA extraction procedure was performed using two different methods - commercial kit for DNA extraction (QIAGEN DNAesy kit) and modifi ed version of the cetyltrimethylammonium bromide (CTAB) methods of Rogers & Bendich (1985). To eliminate possible contamination with DNA from other plants, we used UV treatment of laboratory over night, prior to DNA extraction. Also, all tubes and other equipment used for DNA extraction and PCR reaction were UV treated. Seeds of pea and vetch (500 mg for CTAB methods and 100 mg for DNAeasy) were
ground in liquid nitrogen to a powder form with mortar and pestle, with adding of insoluble PVPP (polyvinylpolypirrolidone-SIGMA-ALDRICH, MO, USA, #856487), 2:1 (m/m). For CTAB methods the resulting powder was added to 1 ml CTAB extraction buffer (3% CTAB, 100 mM Tris-HCl pH8.0, 20 mM EDTA, 1.4 M NaCl, preincubated at 600C), mixed thoroughly and incubated at 600C for 4 h and at 370C over night. The mixture was centrifuged in a microfuge for 10 min and the supernatant extracted two times with 24 : 1 (v/v) chloroform : isoamyl alcohol. Two volumes of CTAB precipitation solution (1% CTAB, 50 mM Tris-HCl pH 8.0, 10 mM EDTA) was added to aqueous layer and mixed well. The mixture was incubated at room temperature for 1 h, and then centrifuged in a microfuge for 10 min. The supernatant was discarded and the pellet resuspended in 350 μl 1.4 M NaCl and 350 μl of chloroform. The mixture was centrifuged in a microfuge for 10 min, the aqueous layer was mixed with 0.6 vol of cold isopropanol, centrifuged for 10 min, and the pellet was washed 3 times with 70% ethanol. The pellet was dried in speed-vac and resuspended in 50 μl of water. The second method for DNA isolation (DNAesy kit) was performed according manufacturer’s instruction.
The concentration of DNA was measured using NanoVue spectrophotometer (GE Heltcare).
For genome amplifi cation, we used Illustra GenomiPhi HY DNA Amplifi cation Kit (Amersham, GE Healthcare, UK) according to manufacturer’s instruction. Whole genome amplifi cation (WGA) is based on multiple displacement amplifi cation. This is an isothermal (30oC), linear amplifi cation using random hexamer primers and phi29 DNA polymerase. Primers attach randomly to the template adding complementary base pairs to one strand, while displacing the other DNA strand. Primary products initiate secondary priming on the displaced DNA strand and continue displacing to create multiple branches. If the initial sample of genomic DNA is high quality, it can be amplifi ed one million-fold while maintaining accurate loci and allele representation. Phi29 DNA polymerase was inactivated at 60 oC. The product of reaction was diluted 50 times, and used for PCR reaction.
PCR reaction was performed using primers for 26S rRNA:
26Sf: 5’-ttcccaaacaacccgactc-3’; 26Sr: 5’-gccgtccgaattgtagtctg-3’.
Primers used for PCR reaction were developed

Extraction of the Ancient DNA from Charred Seeds from Hissar, Serbia 229
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
in Laboratory for Plant Molecular Biology at the Institute of Molecular Genetics and Genetic Engineering, Belgrade. They are not specifi c for legumes, but primers were designed according to conserved region of 26S rDNA gene present in all plants, and these primers were tested on different plant species (Arabidopsis, tobacco, buckwheat, Ramonda nathaliae and R. serbica, pea, vetch) and after PCR reaction one single band was obtained.
The reaction product was used as a template for PCR reaction. The reaction mixture (25 μl): 2 μl DNA from genome amplifi cation reaction, 2.5 mM MgCl2, 0.2 mM dNTP mix, 0.4 μM primers, 1X Gold buffer (Applied Biosystems) and 1U Gold Taq polymerase (Applied Biosystems). PCR reaction was performed in a Biometra Thermocycler, using the following profi le:
cycle: 95°C for 5 min (initial denaturation)30 cycles: 95°C for 30 sec (denaturating),
60°C 30 sec (annealing), 72°C 1 min (extension), with fi nal extension for 10 min at 72°C.
For positive and negative control reaction genomic DNA from Pisum sativum and Vicia ervilia, extracted using DNAeasy kit, and water were used, respectively.
The PCR products were separated on 2% agarose gel and visualised by UV/EtBr staining.
Results and Discussion
To perform isolation of DNA from ancient 3,200-year-old pea and vetch seeds, we tested two different methods for conventional DNA isolation, with some modifi cations. In the fi rst method, we used conventional CTAB method, as described in the previous section. In this procedure we changed CTAB concentration in buffer, from 2% to 3%, in order to increase osmolarity of extraction buffer and obtain better disruption of the seeds. Apart of this modifi cation, we used insoluble polyvinylpolipyrrolidone (PVPP). It is exceptionally good at absorbing polyphenols during DNA purifi cation. Polyphenols are common in many plant tissues and can deactivate proteins if not removed and therefore inhibit many downstream reactions like PCR. Without the addition of PVPP, the precipitated DNA was brown, indicating the presence of oxidized phenols. This brown colour was lost on supplementation with PVPP in the extraction process (Angels et al. 2005). Regarding the fact that dry seeds possess large amounts of polyphenols and other components that may
inhibit PCR reaction, adding of PVPP is fi rst step in improving chance for isolation of ancient DNA. Using described methods, we observed that ancient pea seeds contain more fats than vetch seeds, and more extensive removing of these compounds was necessary. In addition, after removing of non-polar compounds, it was evident that vetch seed contained different pigments than pea seeds, according to colour of solution during DNA isolation. However, CTAB method yielded low-quality and low quantities of DNA, even though a detectable (about 2 ng μl-1) amount of DNA were present.
Using different methods for DNA isolation-commercially available DNAeasy kit (QIAGEN), we obtained 7.2 ng μl-1 of DNA from vetch seeds, and 0.8 ng μl-1 from pea seeds. However, using these DNA isolates, as well as CTAB obtained isolates for PCR reaction, we did not observe any amplifi cation products, nor after re-amplifi cation of PCR products. Lower DNA amount in pea seeds could be explained by the fact that distinctive types of combustion are likely to have occurred with different archaeological remains, varying from the largely anoxic or low-oxygen events occurring in an archaeological place (Threadgold & Brown 2002). Another explanation is that not all seeds contain ancient DNA, suggesting that the critical factors include some morphological feature of the seed itself (Brown et al. 1998). Regarding this fact, we decided to perform whole genome amplifi cation (WGA), using Illustra GenomiPhi HY DNA Amplifi cation Kit. The concept of whole genome amplifi cation is something that has arisen in the past few years as the polymerase chain reaction (PCR) has been adapted to replicate regions of genomes that are of biological interest. The applications are many - forensic science, embryonic disease diagnosis, bioterrorism genome detection, “immortalization” of clinical samples, microbial diversity, and genotyping. Whole-genome amplifi cation (WGA) can replenish dwindling stocks of genomic DNA or bolster low yields, which limit the number of downstream assays that can be completed. PCR-based WGA methods can introduce amplifi cation bias and generate products of insuffi cient length for some applications, such as RFLP analysis. The kit features Phi29 DNA polymerase, a bacteriophage enzyme normally involved in rolling-circle replication, which has the unique property of strand displacement. This is non-PCR method of WGA.
Using reaction product as a template for PCR

Jovanović Ž et al.230
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
with primers for 26S rDNA, we observed single band, approximately 120 bp, corresponding to PCR product obtained on modern pea and vetch DNA (Figure 1).
Our goal is further optimization of methods
in order to obtain PCR product which can be used for sequencing and archeaobotanical investigation of ancient plants and their modern relatives. The sequencing of PCR products should be imperative, in order to eliminate possible contamination with modern DNA.
Conclusions
Figure 1. PCR products obtained after amplifi cation of ancient and modern DNA from pea and bitter vetch with 26S primers. Lanes: 1 - ancient pea DNA extracted using kit, 2 - ancient pea DNA extracted using CTAB method, 3 - modern pea DNA, 4 - ancient bitter vetch DNA extracted using kit, 5 - ancient bitter vetch DNA extracted using CTAB method, 6 - modern bitter vetch DNA, 7 - negative control, 8 - DNA size marker (50bp Fermentas).Slika 1. Produkti PCR reakcije dobijeni nakon amplifi kacije drevne i moderne DNK graška i urova upotrebom 26S prajmera. Kolone: 1 - drevna DNK graška ekstrahovana upotrebom kita, 2 - drevna DNK graška ekstahovana primenom CTAB metode, 3 - DNK savremenog graška, 4 - drevna DNK urova ekstahovana upotrebom kita, 5 - drevna DNK urova ekstrahovana primenom CTAB metode, 6 - DNK savremenog urova, 7 - negativna kontrola, 8 - marker veličine DNK.

Extraction of the Ancient DNA from Charred Seeds from Hissar, Serbia 231
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
Although there are data about aDNA isolation from ancient plants, this is the fi rst time that aDNA from ancient pea and bitter vetch seed was isolated. In addition, more interesting fact is that these seeds were from a local Serbian archaeobotanical site which could be very important for further investigation. Moreover, we showed that aDNA from charred pea and bitter vetch seeds could relatively easily be isolated, using commercial kit for DNA extraction. Although some limitations for use of aDNA for PCR still exist, we showed that fragment of nuclear ribosomal DNA gene-26S rDNA can be amplifi ed by PCR and further applied for investigating and answering archaeobotanically and archaeologically relevant questions. These include questions such as species identifi cation, origin and spread of cultivated plants or monitoring the state of domestication; these are the questions that cannot be answered by morphological archaeobotanical studies alone.
References
Alvarez I, Wendel J F (2003): Ribosomal ITS sequences and plant phylogeny inference. Mol. Phylogenet. Evol. 29: 417-434
Angeles J, Laurena A, Mendoza-Tecson E (2005): Extracti-on of genomic DNA from the lipid-, polysaccharide-, and polyphenol-rich coconut (Cocos nucifera L.). Plant Mol. Biol. Report. 23: 297a-297i
Brown T A (2001): DNA sequencing. In: Brown T A (ed.), Essential Molecular Biology: A Practical Approach, Volume 2, Oxford University Press, Oxford, UK, pp.157-186
Burger J, Hummel S, Herrmann B, Henke W (1999): DNA pre-servation: a microsatellite –DNA study on ancient skeletal remains. Electrophor. 20: 1722-1728
Cooper A, Poinar N H (2000): Ancient DNA: do it right or not at all. Science 289: 1139
Goloubinoff P, Paabo S, Wilson A (1993): Evolution of maize inferred from sequence diversity of an adh2 gene segment from archaeological specimens. Proc. Nat. Acad. USA 90: 1997-2001
Gugerli F, Parducci L, Petit R J (2005): Ancient plant DNA: review and prospects. New Phytol. 166: 409-418
Knaak C, Hamby R K, Arnold M L, Leblanc M D, Chapman R L, Zimmer E A (1990): Ribosomal DNA variation and its use in plant biosystematics. In: Kawano S (ed.), Biological approaches and evolutionary trends in plants, Academic Pre-ss, London, UK, pp 135-158
Pruvost M, Schwarz R, Bessa Correira V, Champlot S, Braguier S, Morel N, Fernandez-Jalvo Y, Grange T, Geigl E M (2007): Freshly excavated fossil bones are best for amplifi cation of ancient DNA. Proc. Nat. Acad. Sci. USA 104: 739-744
Rogers S, Bendich A (1985): Extraction of DNA from milli-gram amounts of fresh, herbarium and mummifi ed plant tissues. Plant Mol. Biol. 5: 69-76
Rohland N, Hofreiter M (2007): Comparison and optimization of ancient DNA extraction. BioTech. 42: 343-352
Schlumbaum A, Tensen M, Despres-Jaenicke V (2008): Ancient plant DNA in archaeobotany. Veget. Hist. Archaeobot. 17: 223-244
Smith C I, Chamberlain A T, Riley M S, Cooper A, Stringer C B, Colins M J, Linder C R, Moore L A, Jackson R B (2001): Neanderthal DNA. Not just old but old and cold? Nature 410: 771-772.
Soltis P S, Soltis D E, Doyle J J (1992): Molecular Systematics of Plants. Chapman and Hall, New York
Threadgold J, Brown T (2003): Degradation of DNA in artifi -cially charred wheat seeds. J. Archaeol. Sci. 30: 1067-1076

Jovanović Ž et al.232
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 227-232
Grašak i urov Tetovac - made in ranogvozdenodobni Leskovac. Deo drugi. Ekstrakcija drevne DNK iz ugljenisanih semena
sa nalazišta Hisar u južnoj Srbiji
Živko Jovanović1 ∙ Nemanja Stanisavljević1 ∙ Aleksandra Nikolić1 ∙ Aleksandar Medović2 ∙ Aleksandar Mikić3 ∙ Svetlana Radović4 ∙ Vuk Đorđević3
1 Institut za molekularnu genetiku i genetičko inženjerstvo Univerziteta u Beogradu, Vojvode Stepe 444a, p.fah 23, 11010 Beograd, Srbija
2 Muzej Vojvodine, Dunavska 35, 21 000 Novi Sad3 Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbija
4 Biološki fakultet Univerziteta u Beogradu, Studentski trg 16, 11000 Beograd, Srbija
Izvod: Primenom dve različite procedure ekstrahovana je drevna DNK iz ugljenisanih semena graška i urova starih 3200 godina sa lokaliteta Hisar kod Leskovca. Korišćena je modifi kovana CTAB metoda i dobijena je relativno mala količina drevne DNK u poređenju sa drugom primenjenom metodom – komercijalno dostupnim kitom za izolaciju DNK. Nakon ekstrakcije primenjeno je umnožavanje celog genoma pomoću Phi29 DNK polimeraze. Umnožena DNK je korišćena za PCR reakciju primenom prajmera za 26S rDNK gen, koji je lociran u jedarnom genomu. Do-bijen je fragment iste veličine kao i PCR fragment 26S rDNK savremenih srodnika graška i urova. Može se zaključiti da je moguće ekstrahovati drevnu DNK iz ugljenisanih semena graška i urova i koristiti je za dalje arheobotaničke analize na molekularnom nivou. Ključne reči: drevna DNK, PCR, ugljenisano seme graška, ugljenisano seme urova

Words Denoting Faba Bean in European Languages 233
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238original research article / originalni naučni rad
Words Denoting Faba Bean(Vicia faba) in European Languages
Aleksandar Mikić
received / primljeno: 18.11.2010. accepted / prihvaćeno: 23.12.2010.© 2011 IFVC
Summary: Faba bean (Vicia faba L.) took part in the ‘agricultural revolution’ of post-glacial Europe along with other grain legumes and cereals. In order to assess the diversity and the origin of the words denoting faba bean in the languages of Europe, a lexicological study was carried out with emphasis upon etymological dictionaries. The words in almost all modern Indo-European languages in Europe owe their origin to the Proto-Indo-European root *bhabh-, bhabhā, also denoting faba bean. The Proto-Altaic root *bŭkrV, denoting pea, nut and cone, through the Proto-Turkic *burčak, denoting both pea and bean, is responsible for the words in several modern Altaic languages of Europe, while the others are borrowings from Arabic. The origin of the words in modern Caucasian languages is the Proto-Cau-casian root *howł[ā], meaning both bean and lentil. The words in Uralic languages are either borrowings, mostly from Slavic, or derived from their own words denoting pea. Key words: crop history, etymology, Europe, faba bean, lexicology, Vicia faba
A. Mikić Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbiae-mail: [email protected]
Introduction
Faba bean (Vicia faba L.) originated in the Central Asian centre of diversity (Zeven & Zhukovsky 1975), unlike many other botanically related grain legume species such as pea (Pisum sativum L.), vetches (Vicia spp.) and vetchlings (Lathyrus spp.) which came mostly from the Near East and Mediterranean. Another peculiarity of faba bean in comparison to its close legume relatives is a lack of evidence about its non-cultivated forms and a potential progenitor, being usually considered completely extinct (Tanno & Willcox 2006). It is, however, one of the fi rst domesticated crops, together with several other legumes, namely pea, bitter vetch (V. ervilia (L.) Willd.), lentil (Lens culinaris Medik.) and chickpea (Cicer arietinum L.), and cereals, with the earliest well preserved archaeobotanical fi nds 10 millennia old and located in Syria (Zohary & Hopf 2000). Together with these crops, faba bean took part in the ‘agricultural revolution’ of post-glacial Europe and quickly spread from Asia Minor, over the Balkans climbing up the fl ow of Danube, and reached all over the central, western
and northern parts of the continent already in 6th millennium BC (Rottoli & Castiglioni 2009). It was simultaneously widespread towards and across the Caucasus to the easternmost borders of Europe and also to other regions of Asia and North Africa. From this time until today, faba bean has become one of the economically most important grain legumes in Europe and the world, used for human consumption, animal feeding and various non-food purposes (Mihailović et al. 2010).
Those bringing what we call today agriculture to Europe met with those who had already lived there and who came there from other directions: some peoples were assimilated by the others, some have survived until the modern day and many merged together and brought forward something new. All these processes resulted in a great ethnic and linguistic diversity but strongly correlated (Cavalli-Sforza & Seielstad 2001), making Europe a common home to at least three hundred extinct and living languages of six great families (Price 1998). The richest family is Indo-European, regarded as stemming from a single ancestral language, Proto-Indo-European, fully developed during the 5th and 4th millennia in the Pontic-Caspian steppe (Anthony 2007) and later divided into present Albanian, Armenian, Baltic, Celtic, Germanic, Hellenic, Indo-Iranian, Italic and Slavic branches. The great and still
Ethnobotany / Etnobotanikaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Mikić A234
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
disputed Altaic family is represented in Europe mostly by its Turkic languages. The Basque language, usually considered a language isolate, might be related to the Caucasian languages, making their speakers the descendants of the Paleolithic inhabitants of Europe who retreated before the newcomers to the Pyrenees and the North Caucasus and including both groups into a supposed Dené-Caucasian language superfamily (Ruhlen 2001). The south-east regions of the Caucasus are home to Georgian and three more Kartvelian languages. The Uralic language family in Europe is represented by their rich Finno-Ugric branch, of which Finnish and Hungarian are most known and spoken, while the Maltese language is the only Afro-Asiatic language spoken in Europe.
The (r)evolution of agriculture in Europe, as we fi nd, went in parallel with the development of its languages. In some cases, peoples welcomed faba bean and other primeval crops with the newcomers who had brought them from their original homeland, and they imported the crops together with their names. In other cases, those who had migrated found the familiar crops already present in their new homes and started to use the words the aboriginal population had been using. In some cases, the original words happened to be forgotten and were replaced with those from neighbouring languages, especially if new cropping systems were introduced. Although all this makes a rather complex background, this research was aimed at assessing the diversity and the origin of the words denoting faba bean in the languages of Europe.
Materials and Methods
In order to complete the goals of this preliminary lexicological essay, the analysis of the origin and diversity of the words denoting faba bean in European languages has been carried out in two major steps.
The fi rst action targeted the past and the present diversity and comprised searching available printed and online dictionaries of the most important living and extinct European languages and collecting all the words, together with their modifi cations, that denote faba bean.
The second task was to examine the existing etymological dictionaries and other relevant historical linguistics resources, related to a single language and a whole language family, in order to collect all the root words related to faba bean in the ancestors of modern European languages and thus shed light on the question of the origin.
Finally, I have tried to bring these two steps together in an integrated analysis that could provide both linguists and crop historians with attested and assumed forms of the words denoting faba bean in living and extinct European languages, in order to try to explain how they developed, diversifi ed, disappeared and survived. This should also initiate further interdisciplinary approaches to this complex and interesting issue, proving that natural and social sciences may assist each other to better understand the past of mankind.
Results and Discussion
Indo-European languages
The collected words related to faba bean in the modern Indo-European languages and their major dialects show a high degree of mutual similarity, especially between and within the most abundant branches such as Germanic, Italic and Slavic (Tab. 1).
It is considered that the Proto-Indo-European root denoting faba bean was *bhabh-, bhabhā, meaning literally something swelling. Its evolution followed the migrations of the Indo-European tribes and brought forth numerous derivations (Pokorny 1959, Nikolaev 2007):
- The supposed Proto-Albanian root *bhakā gave the form in Modern Albanian;
- The extinct Old Prussian language preserved the primeval Proto-Indo-European root to a greater extent, with baba and babo, than its surviving relatives such as Latvian and Lithuanian;
- The Proto-Germanic language root *bau-nō(n-), with a presumed more ancient form of *bab-n-ō, is responsible for the words denoting faba bean in all modern Germanic languages, as witnessed by the extinct and mediating bōne in Middle Dutch, bēan in Old English, bāne in Old Frisian, bōna in both Old High German and Old Low German and baun in Old Norse, with the last one passing it along with its meaning to all modern Goidelic Celtic languages, namely Irish, Manx and Scottish Gaelic (Macbain 1911);
- In the Hellenic languages, there was a shift of meaning from faba bean to lentil, making φακóς denoting the latter in both Old and Modern Greek, while some connect it to the Greek verb φάγέώ, to eat, highlighting its use for food and feed by the ancient Greeks and Romans (Duc et al. 2010);
- In the Italic languages, there are haba in extinct Faliscan and faba in Latin, with the latter

Words Denoting Faba Bean in European Languages 235
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
being ultimate source of the words denoting faba bean in all modern Italic languages derived from Vulgar Latin, and commonly known as Romance languages. From this Latin it was borrowed by Brythonic, the language which gave the Brythonic Celtic languages, namely Breton, Cornish and Welsh;
- The Proto-Slavic root denoting faba bean, *bobŭ, proved to be also conservative, retaining almost the unchanged form in all its living descendants (Vasmer 1953, Skok 1971). It was imported by Romanian, as bob, and Romani, as boba.
The Armenian word denoting faba bean might be a borrowing of lobó-s, the Old Greek word
Fig. 1. A simplifi ed geographic and historical evolution of the Proto-Indo-European word denoting faba bean into its direct descending languagesSlika 1. Pojednostavljena geografska i istorijska evolucija praindoevropske reči koja označava bob u njenim neposrednim jezicima naslednicima
Table 1. The words denoting faba bean in modern Indo-European languages and dialects of EuropeTabela 1. Reči koje označavaju bob u savremenim indoevropskim jezicima i dijalektima Evrope
BranchGrana
LanguageJezik
WordReč
BranchGrana
LanguageJezik
WordReč
Albanian bathë
Italic
Friulian faveArmenian lobi Galician fabaBaltic Latvian pupas Italian fava
Lithuanian pupa Judaeo-Spanish ava
Celtic
Breton fav Ligurian bazann-aCornish fav Occitan favaIrish pónaire Picard fèfeManx poanrey Portuguese favaScottish Gaelic pònair Romanian bobWelsh ffa Sardinian fa
Germanic
Danish bønne Spanish habaDutch boon Walloon féveEnglish bean
Slavic
Belarusian bobFaroese bøna Bulgarian bobFrisian beanne Croatian bobGerman Bohne Czech bobIcelandic baun Kashubian bóbNorwegian bønne Lower Sorbian bobSwedish böna Macedonian bobYiddish bob Polish bób
Hellenic Greek koukiá Russian bobIndo-Iranian Ossetic qædur Rusyn bob
Romany boba Serbian bob
ItalicAragonese faba Slovak bobCatalan fava Slovenian bobCorsican fava Ukrainian bibFrench fève; fèverole Upper Sorbian bob

Mikić A236
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
denoting a pod of leguminous plants that, in its turn, was derived from the Proto-Indo-European *leb-, denoting blade (Nikolaev 2007).
Altaic languages
The Proto-Altaic root *bŭkrV, denoting pea, nut and cone (Starostin et al. 2003), through the Proto-Turkic *burčak, denoting both pea and bean, is responsible for the words denoting faba bean in several modern Altaic languages of Europe, such as Bashkir, Chuvash, Crimean Tatar, Karaim, Kazakh, Kumyk and Tatar (Tab. 2), where in most cases and with an adjective, it literally means black pea, probably because of the black colour of mature faba bean pods. The fact that the Proto-Turkic word denoted both pea and bean may mean that it simply denoted a pulse, that is, something that was cooked and eaten in such way, while they differed from each other only in seed size and shape, with pea smaller and more round and bean larger and more irregularly shaped.
The Arabic word denoting faba bean, bakl, was imported in some Altaic languages, most notably Turkish (Nişanyan 2002) and Karaim.
Due to the geographic proximity, the Azeri word denoting faba bean could have the same origin as the one in Armenian language, while the Kalmyk word denoting faba bean is obviously a borrowing from Russian.
Caucasian languages
The Proto-Caucasian root *howł[ā], meaning both bean and lentil (Nikolayev & Starostin 1994), is responsible for the words denoting faba bean in some modern Caucasian languages such as Aknadian, Avar and Tindi (Tab. 3). This root is considered to be related to the Proto-Basque *iłhař, denoting pea, faba bean, vetch and heather and giving the Modern Basque ilar, denoting only pea (Bengtson 2007), and thus another proof of the theory on the Dené-
Caucasian language superfamily.Another Proto-Caucasian root, *qŏr’ā,
denoting solely pea (Nikolaev & Starostin, 1994), gave rise to the words denoting faba bean in some other modern Caucasian languages such as Archi, Chechen, Ingush, Lak, Lezgian, Tabassaran and Adyghe, with very little difference in comparison to the words denoting pea (Mikić 2009).
The alternative Lezgian word denoting faba bean, paxla, is a borrowing from the Turkish language.
Uralic languages
The majority of the words denoting faba bean in the modern Uralic languages of Europe are borrowings. The speakers of the westernmost ones such as Estonian, Finnish, Ingrian, Livonian, Sami and Võro (Tab. 4) imported both the faba bean crop and the word denoting it from their Slavic neighbours (Stoddard 2009). The same is true for Hungarian, which borrowed its word denoting faba bean from the South Slavs who welcomed Hungarian tribes in the Carpathian Basin in 9th century AD (Skok 1971). Some, like Mari, obviously borrowed their words denoting faba bean from their Turkic neighbours.
There are some indications that the Proto-Uralic root kača, denoting hole and cavity (Starostin 2005), could have given origin to the words denoting pea and faba bean in some of the contemporary Uralic languages, closest to their original homeland, such as Erzya, Moksha and Udmurt (Lytkin & Gulyaev 1970).
Other language families of Europe
Similarly to Armenian and Azeri, the words denoting faba bean in modern Kartvelian languages, such as the Georgian lobio, may be borrowings of the Old Greek lobó-s.
Table 2. The words denoting faba bean in modern Altaic languages of EuropeTabela 2. Reči koje označavaju bob u savremenim altajskim jezicima EvropeBranchGrana
LanguageJezik
WordReč
Mongolian Kalmyk bob
Turkic
Azeri lobyaBashkir baqsa borsağıChuvash nímëş parşíCrimean Tatar pasleKarachay-Balkar hans qudoruKaraim baqla; burcacyk; bobKazakh iri burşaqKumyk burçaqTatar bakça borçagıTurkish bakla

Words Denoting Faba Bean in European Languages 237
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
The Maltese word denoting faba bean, fula, is a modifi cation of the Arabic fūl-, originating directly from the Proto-Semitic *pūl-, denoting beans, and ultimately from the Proto-Afroasiatic *pal-, denoting both bean and corn in general (Militarev 2006).
The word denoting faba bean in modern Basque, baba, is a borrowing from the Latin language (Trask 2008).
Conclusions
Although preliminary and highly incomplete, the gathered and presented lexicological evidence may provide both the linguistic and the life sciences’ audience with a valuable testimony to the role faba bean and other traditional grain legumes had in everyday life of the Europe nations. Some of the collected words, especially those of the Indo-European language family, prove to be highly conservative and thus preserve their most ancestral forms in almost unchanged forms. Others witness a rich and continuous exchange of both crops and words denoting them among the European peoples that brought forth modern nations. This research may hopefully contribute to the interdisciplinary approach by agricultural sciences, archaeobotany
and linguistics to the ever puzzling issue of the very origins of European civilization.Future research on the legume lexicology should surely aim not only to determining the origins and the evolution of the words denoting individual species, but also to give clues of their mutual horizontal transfer where faba bean played a very important role. In many languages, such as Germanic, the words such as the English bean became much more associated to Phaseolus beans than to V. faba. At the same time, bean became a kind of root word for numerous bean-like seeds or products, such as coffee bean, adzuki bean or soybean. Among the next steps on the faba bean lexicology will surely be its agronomical aspects which deserve a study of their own that would deal with a rather rare multitude of its names denoting various types, depending on seed size or growing conditions, such as broad bean or horse bean in English, fève des marais in French or Ackerbohne in German, as recognised already by Pliny.Regardless of the extent it is used at this very moment or its form, what the scientists recently labelled as faba bean and what is globally known as Vicia faba remains an important part of our everyday life, our diets, our speech and our culture.
Table 3. The words denoting faba bean in modern Caucasian languagesTabela 3. Reči koje označavaju bob u savremenim kavkaskim jezicima
BranchGrana
LanguageJezik
WordReč
Northeast
Aknadian haleArchi bex:`e čaqAvar holoChechen qoIngush qeşLak luћi qjuruLezgian xaru; paxlaTabasaran xaruTindi beč’at’ub hali
Northwest Adyghe ceshä
Table 4. The words denoting faba bean in modern Uralic languages of EuropeTabela 4. Reči koje označavaju bob u savremenim uralskim jezicima Evrope
BranchGrana
LanguageJezik
WordReč
Finno-Permic
Erzya kuvtjolEstonian ubaFinnish papuIngrian papuKarelian papuLivonian pubādMari neməčpursaMoksha babanjsnavSami báhpuUdmurt s’öd köžyVõro uba
Ugric Hungarian bab

Mikić A238
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 233-238
Inwitári! Mistress of Bean!No mortal eye has ever seenThy rosy cheeks as bean in bloomTo love thee’s my eternal doom.
References
Anthony D W (2007): The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton University Press, New Jersey
Bengtson J (2007): Basque etymology [Online]. Available at http://starling.rinet.ru (Cited 18 November 2010, verifi ed 18 November 2010). The Tower of Babel, an International Etymological Database Project
Cavalli-Sforza L L, Seielstad M (2001): Genes, Peoples and Lan-guages. Penguin Press, London
Duc G, Bao S, Baum M, Redden B, Sadiki M, Suso M J, Vish-niakova M, Zong X (2010): Diversity maintenance and use of Vicia faba L. genetic resources. Field Crop. Res. 115: 270-278
Lytkin V I, Gulyaev E S (1970) Kratkii etimologicheskii slovar’ komi yazyka. Nauka, Moscow
Macbain A (1911): Etymological Dictionary of the Gaelic Lan-guage. Eneas Mackay, Inverness
Mihailović V, Mikić A, Vasić M, Ćupina B, Đurić B, Duc G, Stoddard F L, Hauptvogel P (2010): Neglected legume crops of Serbia – Faba bean (Vicia faba). Ratar. Povrt. / Field Veg. Crop. Res. 47: 27-32
Mikić A (2009): Words denoting pea (Pisum sativum) in Europe-an languages. Pisum Genet. 41: 29-33
Militarev A. (2006): Semitic etymology [Online]. Available at http://starling.rinet.ru (Cited 18 November 2010, verifi ed 18 November 2010). The Tower of Babel, an International Etymological Database Project
Nikolayev S L (2007): Indo-European etymology [Online]. Available at http://starling.rinet.ru (Cited 18 November 2010, verifi ed 18 November 2010). The Tower of Babel, an International Etymological Database Project
Nikolaev S L, Starostin S A (1994): A North Caucasian Etymo-
logical Dictionary. Asterisk, MoscowNişanyan S (2002): Sözlerin Soyağaci: Çağdaş Türkçenin Eti-
molojik Sözlüğü. Adam yay., IstanbulPokorny J (1959): Indogermanisches Etymologisches Wörter-
buch, 1. Francke, BernPrice G (1998): An Encyclopedia of the Languages of Europe.
Blackwell, OxfordRottoli M, Castiglioni E (2009): Prehistory of plant growing
and collecting in northern Italy, based on seed remains from the early Neolithic to the Chalcolithic (c. 5600–2100 cal B.C.). Veget. Hist. Archaeobot. 18: 91-103
Ruhlen M (2001): Il Dene-caucasico: una nuova famiglia lingui-stica. Pluriverso 2: 76-85
Skok P (1971): Etimologijski rječnik hrvatskoga ili srpskoga je-zika, 1 (A - J). Jugoslavenska akademija znanosti i umjetnosti, Zagreb
Starostin S, Dybo A, Mudrak O (2003): Etymological Dictiona-ry of the Altaic Languages. Brill, Leiden
Starostin S (2005): Uralic etymology [Online]. Available at http://starling.rinet.ru (Cited 18 November 2010, verifi ed 18 November 2010). The Tower of Babel, an International Etymological Database Project
Stoddard F L, Hovinen S, Kontturi M, Lindström K, Nykänen A (2009): Legumes in Finnish agriculture: history, present status and future prospects. Agric. Food Sci. 18: 191-205
Tanno K, Willcox G (2006): The origins of cultivation of Cicer arietinum L. and Vicia faba L.: early fi nds from Tell el-Kerkh, north-west Syria, late 10th millennium B.P. Veget. Hist. Ar-chaeobot. 15: 197-204
Trask R L (2008): Etymological Dictionary of Basque. Univer-sity of Sussex, Brighton
Vasmer M (1953): Russisches etymologisches Wörterbuch, 1 (A–K). Carl Winters Universitätsverlag, Heidelberg
Zeven A C, Zhukovsky P M (1975): Dictionary of Cultivated Plants and Their Centres of Diversity. Centre for Agricul-tural Publishing and Documentation, Wageningen
Zohary D, Hopf M (2000): Domestication of Plants in the Old World: the Origin and Spread of Cultivated Plants in West Asia, Europe and the Nile Valley. Oxford University Press, Oxford
Reči koje označavaju bob (Vicia faba) u evropskim jezicima
Aleksandar Mikić
Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbija
Izvod: Bob (Vicia faba L.) je uzeo učešća u tzv. poljoprivrednoj revoluciji Evrope nakon poslednjeg ledenog doba zajedno sa ostalim zrnenim mahunarkama i strninama. U cilju utvrđivanja raznolikosti i porekla reči koje označava-ju bob u evropskim jezicima, izvršeno je leksikološko istraživanje sa naglaskom na etimološkim rečnicima. Reči u skoro svim savremenim indoevropskim jezicima Evrope vode poreklo od praindoevropskog korena *bhabh-, bhabhā, koji takođe označava bob. Praaltajski koren *bŭkrV, koji označava grašak, orah i šišarku, kroz pratursko *burčak, koje označava grašak i bob, dao je reči u nekim savremenim altajskim jezicima Evrope, dok se u ostalim radi o po-zajmicama iz arapskog. Poreklo reči u savremenim kavkaskim jezicima nalazi se u prakavkaskom korenu *howł[ā], značenja bob i sočivo. Reči u uralskim jezicima su pozajmice, uglavnom iz slovenskih jezika, ili izvedene iz izvornih uralskih reči koje označavaju grašak. Ključne reči: bob, etimologija, Evropa, istorija gajenih biljaka, leksikologija, Vicia faba

Varijabilnost nekih morfoloških svojstava suncokreta 239
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244prethodno saopštenje / research note
Varijabilnost visine biljke i prečnika glavekod novih hibridnih kombinacija suncokreta
(Helianthus annuus L.)
Radovan Marinković · Milan Jocković · Siniša Jocić · Mihajlo Ćirić
primljeno / received: 18.10.2010. prerađeno / revised: 11.11.2010. prihvaćeno / accepted: 30.11.2010.© 2011 IFVC
Izvod: Proučavana je visina biljke i prečnik glave kod 30 novih hibridnih kombinacija suncokreta. Is-pitivanja su obavljena u poljskim uslovima na oglednom polju Instituta za ratarstvo i povrtarstvo na Rimskim Šančevima. Utvrđene su značajne razlike između genotipova u pogledu ispitivanih svojstava. Visina biljaka se kretala od 192,83 cm (OD 3369 x RHA-B-1) do 250,17 cm (L-48 x Snrf-583), a prečnik glave između 20,47 cm (UK-PA 45 x Snrf 583) i 24,53 cm (UK-PA 243 x RHA-B-1). Grupisanje geno-tipova prema sličnosti urađeno je klaster analizom. Kod visine biljaka konstatovane su četiri grupe i dva zasebna genotipa, a kod prečnika glave tri grupe i osam nezavisnih genotipova.Ključne reči: analiza varijanse, klaster analiza, suncokret
R. Marinković · M. Jocković · S. Jocić · M. Ćirić Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbijae-mail: [email protected]
Uvod
Suncokret je osnovna biljka za proizvodnju jestivog ulja u Srbiji kao i u mnogim zemljama sveta. Imajući u vidu specifi čnosti agroekoloških uslova određenih proizvodnih regiona potrebno je posedovati takvu genetsku varijabilnost koja će nam omogućiti kreiranje hibrida za odgovara-juće proizvodne regione kako bi se maksimalno iskoristile osobine hibrida i faktori spoljne sredi-ne. Visina biljke i prečnik glave su važne osobine koje utiču na prinos kod suncokreta (Gvozdeno-vić et al. 2005).
Visini biljke i prečniku glave kod suncokreta uvek je poklanjana određena pažnja. U ranijim periodima gajenja ruski seljaci su uzimali seme za setvu sa biljaka koje su imale visoko i snažno stablo i veliku glavu. Kasnije su se pojavile sorte i hibridi koji su se značajno razlikovali u pogledu ovih svojstava (Marinković i sar. 2002).
Shabana (1974) navodi da je optimalna visina biljke suncokreta između 160 cm i 180 cm. Da bi se povećala otpornost na poleganje i olakšala primena mehanizacije potrebno je stvoriti hibri-de visine od 120 cm do 150 cm (Schneiter et al. 1988). Genotipovi niže stabljike su otporniji na poleganje u uslovima velikih kiša i jakih vetrova,
pa im treba dati prednost u proizvodnji u takvim uslovima zbog jednostavnije žetve (Fick et al. 1985).
Posebnu pažnju u oplemenjivanju suncokre-ta treba posvetiti morfološkim osobinama glave (Mijić i sar. 2005). Prečnik glave utiče na broj cve-tova i broj semena po glavi, što direktno utiče na prinos semena po biljci (Tahir et al. 2002, Hladni 2010).
Cilj ovog rada je bio da se utvrdi genetska varijabilnost visine biljke i prečnika glave novih hibridnih kombinacija suncokreta.
Materijal i metod
Genetski materijal za ovo istraživanje sasto-jao se od 30 novih hibridnih kombinacija koje su stvorene u Institutu za ratarstvo i povrtarstvo u Novom Sadu. Ogled je postavljen u toku 2009. na oglednom polju Instituta za ratarstvo i povr-tarstvo na Rimskim Šančevima u tri ponavljanja po slučajnom blok sistemu. Materijal je bio zase-jan mašinski u četiri reda sa međurednim rastoja-njem od 70 cm i razmakom između biljaka u redu od 25 cm, u svakom ponavljanju.
Merenja visine biljke i prečnika glave urađena su u poljskim uslovima u fazi fi ziološke zrelosti.
Genetika i oplemenjivanje / Genetics and Breedingw
ww.
nsse
me.
com
/jour
nal.h
tml
( )Ovo istraživanje je deo projekta broj TR-31025 „Razvoj novih sorti i poboljšanje tehnologija proizvodnje uljanih biljnih vrsta za različite namene“ Ministarstva za nauku i tehnološki razvoj Republike Srbije

Marinković R i sar.240
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Veličina uzorka za merenje bila je 20 biljaka po ponavljanju, odnosno 60 biljaka na nivou ogle-da. Za ispitivanje varijabilnosti hibridnih kom-binacija primenjena je analiza varijanse prema Hadživukoviću (1973). Izvršeno je grupisanje po sličnosti klaster analizom da bi dobili jasniju sliku o razlikama između genotipova u posmatranim osobinama.
Rezultati i diskusija
Visina biljke
Analizirane hibridne kombinacije suncokreta pokazale su veliku varijabilnost visine biljke, što se može zaključiti iz visoko značajne vrednosti F testa za genotipove (Tab. 1).
Prosečna vrednost visine biljke kod hibrid-nih kombinacija kretala se od 192,83 cm (OD 3369 x RHA-B-1) do 250,17 cm (L-48 x Sn-rf-583). Kod tri hibridne kombinacije visina je bila ispod 200 cm, kod četrnaest kombinacija između 200 cm i 220 cm, kod dvanaest kom-
binacija između 220 cm i 240 cm i kod jedne kombinacije preko 240 cm. Primenom LSD vrednosti utvrđeno je da genotip L-48 x Sn-rf-583 ima visoko značajno veću (prag rizika od 1%) visinu biljke od svih ostalih hibridnih kombinacija. Najveće razlike su bile između genotipa L-48 x Snrf 583 i kombinacija OD 3369 x RHA-B-1, L-1 x RHA-B-1 i UK-PA 63 x RHA-B-1 (Sl. 1).
U regionima sa jakim vetrovima i obilnim pa-davinama izraženije je poleganje kod viših geno-tipova što dovodi do smanjenja prinosa i kvaliteta semena (Marinković i sar. 2002). Genotipovi niže stabljike imaju sličan potencijal prinosa kao i ge-notipovi više stabljike (Schneiter 1992, Velasco et al. 2003). Smanjivanjem visine biljke moguće je povećati broj biljaka po jedinici površine (Suzer & Atakisi 1993). Uzimajući u obzir navedene či-njenice, a na osnovu izračunatih rezultata visine biljke vidimo da su najpoželjniji genotipovi OD 3369 x RHA-B-1, L1 x RHA-B-1 i UK-PA 63 x RHA-B-1, koji se odlikuju najmanjom visinom biljke (Sl. 1).
Izvor varijacijeSource of variation df MS F test
Ponavljanje / Replication 2 19,62 0,38Genotip / Genotype 29 534,41 10,26**Greška / Error 58 52,07Ukupno / Total 89
Tabela 1. Analiza varijanse visine biljke 30 hibridnih kombinacija suncokreta Table 1. Analysis of variance of plant height of 30 hybrid combinations of sunfl ower
Slika 1. Prosečne vrednosti visine biljke 30 hibridnih kombinacija suncokreta sa LSD (1%)Fig. 1. Average values of plant height of 30 hybrid combinations of sunfl ower

Varijabilnost nekih morfoloških svojstava suncokreta 241
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Primenom klaster analize za visinu biljke izvrše-no je grupisanje hibridnih kombinacija prema slič-nosti u četiri grupe i dva zasebna genotipa. Hibridna kombinacija L-48 x Snrf-583 izdvojila se najvećom srednjom vrednošću visine biljke. Zajedno sa kom-binacijom UK-PA 243 x Snrf-583 spojena je na naj-višem hijerarhijskom nivou sa ostalim kombinacija-ma. Grupe 1 i 3 su na nižim hijerarhijskim nivoima podeljene u više podgrupa. U grupi 3 su se izdvo-jili genotipovi sa najmanjom prosečnom vrednošću visine biljke i oni su bolji adaptirani proizvodnji u plodnijim regionima nego genotipovi više stabljike (Miller & Hammond 1991) (Sl. 2).
Prečnik glave
Analizom varijanse utvrđena je visoko zna-čajna vrednost F testa za genotipove koja nam govori da između novih hibridnih kombinaci-ja suncokreta postoje velike razlike u prečniku glave (Tab. 2). Najmanji prečnik glave konsta-tovan je kod hibridne kombinacije UK-PA 45
x Snrf 583 (20,47 cm), a najveći kod hibridne kombinacije UK-PA 243 x RHA-B-1 (24,53 cm). Ova kombinacija je imala značajno veće vrednosti od svih ostalih kombinacija, osim od UK-PA 229 x Snrf 583, Nil-rm 23 x RHA-B-1, UK-PA 63 x RHA-B-1, CMS-30 x RHA-B-1, L-48 x Snrf 583, L-26 x RHA-B-1, L-98 x Snrf 583 i Nil-rm 23 x Snrf 583. Najveće razlike su utvrđene između hibridne kombi-nacije UK-PA 243 x RHA-B-1 i kombinaci-ja UK-PA 45 x Snrf 583, UK-PA 101 x Snrf 583, UK-PA 63 x Snrf 583 i UK-PA 73 x Snrf 583 (Sl. 3).
Brojnim istraživanjima utvrđena je značajna pozitivna korelacija između prečnika glave i visine prinosa semena (Marinković 1992, Petakov 1994, Punia & Gill 1995, Arshad et al. 2007, Behradfar et al. 2009), dok su značajan direktan efekat preč-nika glave na prinos semena utvrdili Alba et al. (1979), Giriraj et al. (1979) i Marinković (1987).
Slika 2. Klaster analiza za visinu biljke 30 hibridnih kombinacija suncokretaFig. 2. Cluster analysis of plant height of 30 hybrid combinations of sunfl ower
Izvor varijacijeSource of variation df MS F test
Ponavljanje / Replication 2 3,01 2,49Genotip / Genotype 29 2,76 2,29**Greška / Error 58 1,21Ukupno / Total 89
Tabela 2. Analiza varijanse prečnika glave 30 hibridnih kombinacija suncokretaTable 2. Analysis of variance of head diameter of 30 hybrid combinations of sunfl ower

Marinković R i sar.242
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Klaster analiza prečnika glave podelila je ispitivane hibridne kombinacije na tri grupe i osam pojedinačnih genotipova. Grupa 1 se sastoji iz 14 genotipova kod kojih je uočljiva podela i na nižim hijerarhijskim nivoima. U grupi 2 nalazi se 6 genotipova koji su svrstani u dve podgrupe i dva zasebna genotipa. Gru-pu 3 čine 3 genotipa. Na grafi konu se poseb-no ističu genotipovi UK-PA 243 x RHA-B-1
i Nil-rm 23 x RHA-B-1 koji su se odlikovali najvećim prosečnim vrednostima ispitivanog svojstva (Sl. 4).
Uzimajući u obzir mišljenje Škorića i sar. (1989) može se zaključiti da se sve hibridne kom-binacije u klasteru odlikuju optimalnim prečni-kom glave (20-25 cm). Buduće analize međuzavi-snosti prečnika glave i prinosa semena pokazaće koji prečnik je najpoželjniji.
Slika 3. Prosečne vrednosti prečnika glave 30 hibridnih kombinacija suncokreta sa LSD (1%)Fig. 3. Average values of head diameter of 30 hybrid combinations of sunfl ower
Slika 4. Klaster analiza prečnika glave 30 hibridnih kombinacija suncokretaFig. 4. Cluster analysis of head diameter of 30 hybrid combinations of sunfl ower

Varijabilnost nekih morfoloških svojstava suncokreta 243
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Zaključci
Srednja vrednost visine biljke ispitivanih hi-bridnih kombinacija kretala se od 192,83 cm (OD 3369 x RHA-B-1) do 250,17 cm (L-48 x Snrf-583). Utvrđena je velika varijabilnost u visini biljke iz-među genotipova. Klaster analiza visine biljke gru-pisala je hibridne kombinacije u četiri grupe i dva nezavisna genotipa, a najpoželjniji genotipovi za gajenje u različitim uslovima svrstani su u 3. grupu s obzirom da se odlikuju najmanjom prosečnom visinom biljke, a među njima su najinteresantniji za dalja istraživanja OD 3369 x RHA-B-1, L1 x RHA-B-1 i UK-PA 63 x RHA-B-1.
Prečnik glave kretao se između 20,47 cm kod hibridne kombinacije UK-PA 45 x Snrf 583 i 24,53 cm kod hibridne kombinacije UK-PA 243 x RHA-B-1. Analizom varijanse prečnika gla-ve utvrđeno je postojanje velike varijabilnosti u prečniku glave između proučavanih genotipova. Klaster analiza je prema sličnosti razvrstala hi-bridne kombinacije u tri grupe i osam pojedinač-nih genotipova od kojih se posebno ističu geno-tipovi sa najvećim prečnikom glave UK-PA 243 x RHA-B-1 i Nil-rm 23 x RHA-B-1. Svi genotipovi u ogledu su se odlikovali optimalnim prečnikom glave.
Literatura
Arshad M, Ilyas M K and Khan M A (2007): Genetic divergence and path coeffi cient analysis for seed yield traits in sunfl ower (Helianthus annuus L.) hybrids. Pak. J. Bot. 39: 2009-2015
Alba E, Benvenuti A, Tuberosa R, Vanozzi G O (1979): A path coeffi cient analysis of some yield components in sunfl ower. Helia 2: 25-29
Behradfar A, Gorttapeh A H, Zardashty M R and Talat F (2009): Evaluation correlated traits for seed and oil yield in sunfl ower (Helianthus annuus L.). Res. J. Biol. Sci. 4: 82-85
Fick G N, Caroline J J, Auwater G E, Duhigg P M (1985): Agronomic characteristics and performance of dwarf sun-fl ower hybrids. Proc. 11th Inter. Sunfl . Conf., Mar Del Plata, Argentina, 739-742
Giriraj K, Vidyashankart S, Venkataram M N, Seetharam S (1979): Path coeffi cient analysis of seed yield in sunfl ower. The Sunfl ower Newsletter 3: 10-12
Gvozdenović Sandra, Joksimović J, Škorić D (2005): Gene ef-fect and combining abilities for plant height and head diame-ter in sunfl ower. Genetika 37: 57-64
Hadživuković S (1973): Statistički metodi. Radivoj Ćirpanov, Novi Sad
Hladni Nada (2010): Geni i prinos suncokreta (monografi ja). Zadužbina Andrejević, posebna izdanja
Marinković R (1987): Analiza komponenti prinosa semena suncokreta (Helianthus annuus L.) koefi cijentom putanje I. Abstrakt, III Kongres genetičara Jugoslavije sa međunarod-nim učešćem, Ljubljana, Jugoslavija
Marinković R (1992): Path-coeffi cient analysis of some yield components of sunfl ower(Helianthus annuus L.). Euphytica 60: 201-205
Marinković R, Škorić D, Jovanović D (2002): Efekat heterozisa za visinu biljke i prečnik glave kod suncokreta (Helianthus annuus L.). Zbornik radova instituta za
ratarstvo i povrtarstvo, Novi Sad. Sveska 36, 169-177.Marinković R, Dozet B, Vasić D (2003): Oplemenjivanje sunco-
kreta (monografi ja). DOO ’’Školska knjiga’’, Novi SadMijić A, Krizmanić M, Liović I, Bilandžić M, Zdunić Z, Ko-
zumplik V (2005): Procjena kombinacijskih sposobnosti i genetskih učinaka za visinu biljke i promjer glave suncokreta. Poljoprivreda 11: 18-23
Miller J F and Hammond J J (1991): Inheritance of reduced height in sunfl ower. Euphytica 53: 131-136
Petakov D (1994): Correlation and heritability of some quan-titative characters in sunfl ower diallel crosses. Symposium on breeding of oil and protein crops, Albena, Bulgaria, 162-164
Punia M S and Gill H S (1995): Correlation and path coeffi cient analysis for seed yield traits in sunfl ower (Helianthus annuus L.). Plant Breed. Abs. 65: 915
Schneiter A, Curadar B, Zaffdroni G, Maid H (1988): Agro-nomic evaluation of semidwrf sunfl ower. Proc of the 12th Inter. Sunfl . Conf.1, 406-408, Novi Sad, Yugoslavia
Schneiter A A (1992): Production of semidwarf and dwarf in the northern Great Plains of the United States. Field Crop Res. 30: 391-401
Shabana M R (1974): Genetic variability of the yield compo-nents of oil in differentsunfl ower variaties and inbred lines. PhD. thesis, University of Novi Sad, Faculty of Agriculture
Suzer S and Atakisi I (1993): Yield components of sunfl ower hybrids of different height. Helia 16: 35-40
Škorić D, Vrebalov T, Ćupina T, Turkulov J, Marinković R, Ma-širević S, Atlagić J, Tadić L, Sekulić R, Stanojević D, Kovače-vić M, Jančić V i Sakač Z (1989): Suncokret (monografi ja), Nolit, 613
Tahir M H N, Sadaqat H A, Bashir S (2002): Correlation and path coeffi cient analysis of morphological traits in sunfl ower (Helianthus annuus L.) Populations. Int. J. Agri. Biol. 4: 341-343
Velasco L, Dominguez J, Munoz-Ruz J, Perez-Vich B, Fer-nandez-Martinez J M (2003): Registration of ’Dw 89’ and ’Dw 271’ parental lines of sunfl ower. Crop Sci. 43: 1140-1141
Yagoub S O, Osman A A M, Abdesalam K A (2010): Effect of watering intervals and weeding on grouth and yield of sun-fl ower (Helianthus annus L.). J. Sci. Technol. 11: 137-148

Marinković R i sar.244
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 239-244
Variability of Plant Height and Head Diameter in New Hybrid Combinations of Sunfl ower
Radovan Marinković · Milan Jocković · Siniša Jocić · Mihajlo Ćirić
Institute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbia
Summary: Plant height and head diameter of new hybrid combinations of sunfl ower were investigated. Tests were done under fi eld conditions at experimental fi eld of Institute of Field and Vegetable Crops in Rimski Šančevi. Signifi cant differences were determined between genotypes observing the studied characters. Plant height ranged from 192.83 cm (OD 3369 x RHA-B-1) to 250.17 cm (L-48 x Snrf-583), head diameter ranged between 20.47 cm (UK-PA 45 x Snrf 583) and 24.53 cm (UK-PA 243 x RHA-B-1). Genotypes were grouped by cluster analysis. Four groups and two separate genotypes were found regarding plant height, and three genotypes and eight independent genotypes regarding head diameter. Key words: analysis of variance, cluster analysis, sunfl ower

Preliminary Results on Agronomic Performance of Barrel Medic in Serbia 245
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252research note / prethodno saopštenje
Preliminary Results on Agronomic Performanceof Barrel Medic (Medicago truncatula) in Serbia
Vojislav Mihailović · Aleksandar Mikić · Branko Ćupina · Dragan Milić · Đorđe Krstić · Slobodan Katić · Sanja Vasiljević · Đura Karagić
received / primljeno: 29.11.2010. revised / prerađeno: 27.12.2010. accepted / prihvaćeno: 30.12.2010.© 2011 IFVC
Summary: A small-plot trial with eight Australian barrel medic (Medicago truncatula Gaertn.) cultivars was carried out in 2010 at Rimski Šančevi. The average green forage and forage dry matter yields were highest in Jemalong (30.7 t ha-1 and 7.3 t ha-1) and Parabinga (30.7 t ha-1 and 8.0 t ha-1). Forage dry matter crude protein content ranged between 150.8 g kg-1 in Parabinga and 179.4 g kg-1 in Jester. Forage neutral detergent fi bre content varied from 305.2 g kg-1 in Sephi to 458.8 g kg-1 in Caliph, while the average fo-rage acid detergent fi bre content was 312.8 g kg-1. The average seed yield for all cultivars was 281 kg ha-1 and may be considered satisfying, as it was obtained in a very rainy and warm growing season. Jemalong and Parabinga had the highest aboveground nitrogen yield (190 kg ha-1 and 193 kg ha-1) and thus the greatest potential for green manure.Key words: acid detergent fi bre, barrel medic, forage crude protein yield, forage quality, forage yield, green manure, Medicago truncatula, model legume, neutral detergent fi bre, seed yield
V. Mihailović · A. Mikić · D. Milić · S. Katić · S. Vasiljević · Đ. KaragićInstitute of Field and Vegetable Crops, Maksima Gorkog 30, 21000 Novi Sad, Serbiae-mail: [email protected]. Ćupina · Đ. KrstićFaculty of Agriculture, University of Novi Sad, Trg Dositeja Obrado-vića 8, 21000 Novi Sad, Serbia
Introduction
The most important forage fi eld legumes in Serbia are lucerne (Medicago sativa L.), red clover (Trifolium pratense L.), pea (Pisum sativum L.) and vetches (Vicia spp.). It is estimated that the cultivation area of lucerne varies between 180,000 ha and 200,000 ha, while red clover takes from 80,000 ha to 120,000 ha (Karagić et al. 2010). Lucerne is dominant in the northern parts of the country with rich chernozem soils, while red clover is more widely distributed in central Serbia, being more tolerant to prevailing acid soils. Pea and vetches are the most signifi cant annual forage legumes, appreciated for the diversity of their use, ranging from green forage, forage dry matter and forage meal, over silage and haylage to dry grain and straw (Mikić et al. 2010a). Field pea (comprising forage and dry pea) is grown on about 25,000 ha in Serbia today, while vetches are grown on 7,500 ha (Mihailović et al. 2005).
Barrel medic (Medicago truncatula Gaertn.) originated in the Mediterranean basin (Rubiales et al. 2009) and became widely naturalised in other regions of the world such as Australia, where annual medics are used as forages and soil improvers on about 50 million ha (Crawford et al. 1989). During the last decade of the last century, barrel medic was recognised as appropriate to be a model species for legumes, especially due to its diploid and small (about 5 108 base pairs) genome, its self-fertile nature, its prolifi c seed production and its rapid generation time (Cook 1999, Young et al. 2003, Tivoli et al. 2008, Thompson et al. 2009). This was confi rmed by various researches revealing a high synteny, i.e. an extensive conservation of gene order, not only between barrel medic and lucerne (Choi et al. 2004), but also between barrel medic and pea (Kaló et al. 2004). In this way, efforts in genomic research of barrel medic give new perspectives for lucerne breeding (Annicchiarico & Scotti 2009, Julier & Meusnier 2010). The oldest and largest collection of barrel medic is housed at the South Australian
Soil and Crop Management / Gajenje i agrotehnikaw
ww.
nsse
me.
com
/jour
nal.h
tml
( )
Acknowledgements: This research was supported by the Ministry of Science and Technological Development of the Republic of Ser-bia. The authors are grateful to Rade Matić and his colleagues from SARDI for kindly donating the barrel medic seed material for this research.

Mihailović V et al.246
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
Research and Development Institute (SARDI), with more than 5,000 accessions (Ellwood 2006).
The main goal of this very preliminary research was to assess the possibility of growing barrel medic for forage production in the prevailing agro-ecological conditions of northern Serbia. It was especially aimed at assessing the length of its growing season when cultivated for forage and thus the possibility to include it as a short-season crop in prevailing cropping systems in Serbia, with potentially preceding autumn-sown brassicas and succeeding sorghum, Sudan grass or short-season maize. At the same time, this research was aimed at determining the potential of barrel medic for seed production in the conditions of Serbia, as well as its hypothetical use as green manure crop.
Materials and Methods
A small-plot trial was carried out in 2010 at the experimental fi eld of the Institute of Field and Vegetable Crops at Rimski Šančevi, near Novi Sad, 45o20’ N, 19o51’ E and 84 m asl. It included eight barrel medic cultivars of Australian origin, namely Borung, Caliph, Jemalong, Jester, Mogul, Parabinga, Paraggio and Sephi, kindly provided by SARDI.
The trial was established on April 23. All eight cultivars were sown at a density of about 250
viable seeds m-2 (Walsh et al. 2001), a plot size of 1 m2, three replicates for forage production and one plot for seed production, a row spacing of 20 cm both within and between plots in order to reduce excessive growth of border rows of a plot. In fact, each cultivar was sown in seven rows, but only fi ve inside the plot were taken into account, while two border rows were discarded. The dominant weather and soil conditions during the trial period at Rimski Šančevi in 2010 are given in Tables 1 and 2.
All eight cultivars were cut in full bloom and while forming fi rst pods, being a desirable balance between yield and quality in majority of annual forage legumes (Mihailović et al. 2008). Quantitative morphological characteristics, such as average stem length (cm), number of stems (plant-1), number of leaves (plant-1), number of fl owers (plant-1) and number of leaves (plant-1) were measured on the plant samples taken before the cutting. The plant samples of 30 plants per replication were taken from the middle rows of each plot. The same samples were used to measure the proportion of each plant part, namely stems, leaves, fl owers and pods, in the total green forage yield and, after they were dried at the room temperature to a constant mass, in the total dry forage matter yield. Green forage yield (t ha-1) was calculated on the basis of the green forage yield (kg m-2), measured immediately after
Table 1. Weather conditions during the trial with barrel medic at Rimski Šančevi in 2010Tabela 1. Vremenski uslovi tokom ogleda sa buretastom lucerkom na Rimskim Šančevima 2010. godine
Month / MesecYear / Godina
April /April
May /Maj
June /Jun
July /Jul
August /Avgust
Average / SumProsek / Suma
Average monthly temperature (ºC) / Srednja mesečna temperatura2010 13 17 20 23 22 19.0
Long-term averageVišegodišnji prosek 11 17 20 21 21 18.0
Monthly precipitation sum (mm) / Mesečna količina padavina2010 71 95 174 98 169 607
Long-term averageVišegodišnji prosek 47 59 85 70 59 320
Table 2. Chemical properties of the surface chernozem soil layers during the trial with barrel medic at Rimski Šančevi in 2010Tabela 2. Hemijske osobine površinskog sloja zemljišta tipa černozem tokom ogleda sa buretastom lucerkom na Rimskim Šančevima 2010. godine
Depth(cm)Dubina
pHKCl
pHH2O
CaCO3(%)
Humus(%)
N total(%)
Ukupni N
Al-P2O5(mg 100g-1)
Al-K2O(mg 100g-1)
0 - 30 7.41 7.90 5.61 2.97 0.196 17.99 21.00
30 - 60 7.50 7.95 7.52 2.05 0.185 13.45 18.23

Preliminary Results on Agronomic Performance of Barrel Medic in Serbia 247
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
the cutting, while forage dry matter yield (t ha-1) was calculated from plot fresh yield and forage moisture of the forage samples.
The chemical analysis of the forage dry matter in all eight cultivars included crude protein content (g kg-1), by multiplying the forage dry matter nitrogen content (g kg-1) by 6.25, and the content (g kg-1) of neutral detergent fi bre (NDF), acid detergent fi bre (ADF) and lignin. Forage crude protein yield (kg ha-1) was calculated on the basis of the forage dry matter crude protein content and forage dry matter yield.
In the stage of full maturity of the seeds in the fi rst pods, all eight cultivars were harvested by hand. Seed yield (kg ha-1) was calculated on the basis of the seed yield (g m-2), at a moisture content of 14%. Thousand seed weight (g) and seed viability (%) were determined by the samples of 100 seeds one month after the harvest of each cultivar. In order to enable a basic statistical analysis with one plot only, the separate samples were taken from three middle rows, with the seed yield of each recalculated for the statistical purposes.
On the basis of forage dry matter nitrogen content and forage dry matter yield, aboveground nitrogen yield (kg ha-1) was calculated in order to assess the potential of the tested barrel medic cultivars for green manure.
Number of days from sowing to cutting and number of days from sowing to harvest for each of eight cultivars were also recorded.
The results were processed by the method of
analysis of variance (ANOVA) with the LSD test, using the MSTAT-C software.
Results and Discussion
Weather and soil conditions
The growing season of barrel medic at Rimski Šančevi was by 1°C warmer in comparison to long-term average (Tab. 1). At the same time, fi ve months of the barrel medic growing season at Rimski Šančevi were characterised by almost twice precipitation relative to long-term average.
The chernozem soil at Rimski Šančevi was well provided with basic mineral nutrients and humus, with a slight alkaline reaction (Tab. 2), and thus might be considered favourable for the barrel medic cultivation.
Forage yield
There were signifi cant differences among the average values of number of days from sowing to cutting and quantitative morphological characteristics related to forage yield in eight tested barrel medic cultivars (Tab. 3). Number of days from sowing to cutting varied between 55 in Parabinga and 71 in Mogul, pointing that barrel medic has much shorter growing season for forage production in comparison to traditional annual forage legumes such as common vetch, where it varies between 88 days and 102 days (Mikić 2008). On average, stem length in barrel
Table 3. Number of days from sowing to cutting and quantitative morphological characteristics related to forage yield in barrel medic cultivars at Rimski Šančevi in 2010Tabela 3. Broj dana od setve do kosidbe i kvantitativne morfološke osobine vezane za prinos krme sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar /Sorta
Number of days from sowing to cutting /Broj dana
od setve do kosidbe
Average stem length
(cm plant-1) /Prosečna dužina
stabala po biljci
Number of stems
(plant-1) /Broj stabala
po biljci
Number of leaves
(plant-1) /Broj listova
po biljci
Number of fl owers
(plant-1) /Broj cvetova
po biljci
Number of pods
(plant-1) /Broj mahuna
po biljci
Borung 60 50 3.1 122 23 9Caliph 58 45 4.9 90 16 32Jemalong 60 41 6.9 122 35 12Jester 62 38 4.0 120 14 6Mogul 71 45 7.1 110 15 5Parabinga 55 45 3.2 136 22 38Paraggio 70 30 2.2 56 10 7Sephi 62 29 3.6 50 9 5Average / Prosek 62 40 4.3 101 18 14LSD0.05 2 4 2.8 29 8 10

Mihailović V et al.248
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
medic was 40 cm, similar to plant height in the fi rst cut of lucerne, especially in the year of establishment (Mikić et al. 2007). With the number of stems ranging from 2.2 plant-1 in Paraggio to 7.1 plant-1 in Mogul, the barrel medic cultivars, in average, were moderately prone to basal branching in comparison to other annual forage legumes such as large-fl owered (Vicia grandifl ora Scop.) and other vetches (Mikić et al. 2009). The highest number of leaves was in Parabinga (136 plant-1). The cultivars Caliph and Parabinga had greater number of pods (32 plant-1 and 38 plant-1) than fl owers in cutting and as compared to other six cultivars, witnessing their earliness and a faster transition from fl owering to pod forming.
The average green forage and forage dry matter yields were highest in Jemalong (30.7 t
ha-1 and 7.3 t ha-1 respectively), and Parabinga (30.7 t ha-1 and 8.0 t ha-1 respectively) (Tab. 4). The forage dry matter yield in most of the tested cultivars at Rimski Šančevi was within the limits barrel medic achieved in the upper Midwest of USA on Hubbard loamy sand, ranging from about 4.7 t ha-1 to 9.1 t ha-1 (Zhu et al. 1998). This confi rms that barrel medic has the potential for producing high forage yields in temperate regions such as the north-central USA and the northern Balkans, as well as that it might be used when traditional forage supplies are inadequate (Zhu et al. 1996).
It is considered that a higher ratio between leaves and stems in lucerne is mainly associated with shorter and thinner main stems, as well as that it is modestly and inversely correlated with forage yield (Annicchiarico 2007). In all eight
Table 4. Parameters related to forage yield in barrel medic cultivars at Rimski Šančevi in 2010Tabela 4. Veličine vezane za prinos krme sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar /Sorta
Green forage yield (t ha-1) /
Prinos zelene krme
Forage dry matter yield(t ha-1) /
Prinos suve materije krme
Forage dry matter proportion /
Udeo suve materije krme
Borung 28.1 5.9 0.21Caliph 26.2 6.6 0.25Jemalong 30.7 7.3 0.24Jester 22.5 4.9 0.22Mogul 29.3 5.9 0.20Parabinga 30.7 8.0 0.26Paraggio 11.8 3.1 0.26Sephi 11.4 2.7 0.24Average / Prosek 23.8 5.6 0.24LSD0.05 8.2 1.9 0.03
Table 5. Proportion of stems, leaves, fl owers and pods in forage yield in barrel medic cultivars at Rimski Šančevi in 2010Tabela 5. Udeo stabala, listova, cvetova i mahuna u prinosu krme sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar /Sorta
Green forage yield / Prinos zelene krme
Forage dry matter yield / Prinos suve materije krme
Proportion of stems /
Udeo stabala
Proportion of leaves / Udeo listova
Proportion of fl owers
/ Udeo cvetova
Proportion of pods /
Udeo mahuna
Proportion of stems /
Udeo stabala
Proportion of leaves /
Udeo listova
Proportion of fl owers
/ Udeo cvetova
Proportion of pods /
Udeo mahuna
Borung 0.63 0.32 0.03 0.02 0.58 0.38 0.03 0.01Caliph 0.56 0.34 0.01 0.09 0.54 0.35 0.02 0.10Jemalong 0.49 0.43 0.04 0.04 0.49 0.42 0.04 0.04Jester 0.60 0.36 0.01 0.02 0.59 0.38 0.01 0.02Mogul 0.60 0.37 0.01 0.01 0.57 0.41 0.02 0.01Parabinga 0.52 0.34 0.04 0.10 0.51 0.35 0.01 0.13Paraggio 0.58 0.38 0.02 0.02 0.55 0.41 0.01 0.03Sephi 0.51 0.41 0.03 0.04 0.49 0.42 0.05 0.05Average / Prosek 0.56 0.37 0.02 0.05 0.54 0.39 0.02 0.05

Preliminary Results on Agronomic Performance of Barrel Medic in Serbia 249
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
tested barrel medic cultivars, stems had higher proportion than leaves (Table 5). The smallest difference in the stems to leaves ratio in forage dry matter yield was in both Jemalong and Sephi (49 : 42). Caliph and Parabinga were the only cultivars where the joint proportion of fl owers and pods was higher than 10%.
Forage quality
Barrel medic is considered a valuable source of forage protein, especially in mixtures with grasses (Stout et al. 1997). The average forage dry matter crude protein content in eight tested barrel medic cultivars ranged between 150.8 g kg-1 in Parabinga and 179.4 g kg-1 in Jester (Tab. 6). This was lower in comparison to the values
of forage dry matter crude protein content in lucerne cultivars, with a variation from 177 g kg-1 to 194 g kg-1 (Katić et al. 2009), and could partially be explained by an excessively rainy barrel medic growing season in 2010. Paraggio and Sephi had signifi cantly lower forage crude protein yield (511 kg ha-1 and 473 kg ha-1) than the rest of the tested barrel medic cultivars, led by Parabinga (1,206 kg ha-1). Due to the prevailing conditions during its growing period in 2010, barrel medic had a lower average forage crude protein yields in comparison to other annual forage legumes such as pea, with more than 1,500 kg ha-1 (Mihailović et al. 2010).
The content of forage NDF in eight barrel medic cultivars varied between 305.2 g kg-1 in Sephi and 458.8 g kg-1 in Caliph and with an average value of 363.5 g kg-1. In this way, barrel medic had lower forage NDF content than lucerne, with an average value of 391 g kg-1 (Julier et al. 2000) and almost equal to an average seasonal value of 369.3 g kg-1 in red clover (Vasiljević et al. 2008). In a similar way, the average forage ADF content in eight barrel medic cultivars (312.8 g kg-1) was lower in comparison to that in lucerne cultivars in the same agro-ecological conditions, with a range from 374.4 g kg-1 to 395.2 g kg-1 (Katić et al. 2008), but also higher than an average seasonal value of 289.1 g kg-1 in red clover (Vasiljević et al. 2009). Sephi had the lowest forage lignin content (62.5 g kg-1), while had the highest forage lignin content (103.8 g kg-1).
Seed yield
The number of days from sowing to harvest in eight barrel medic cultivars varied between 90 in Jester and Parabinga and 106 in Mogul (Tab. 7), with a similar general trend as the number of days from sowing to cutting and confi rming once again that barrel medic also has shorter growing season for seed production in comparison to the traditional Serbian annual forage legumes such as common vetch, where it varies between 116 days and 136 days (Mikić 2008). The average seed yield in the tested barrel medic cultivars was 281 kg ha-1 and may be considered satisfying bearing in
Table 6. Parameters related to forage dry matter chemical composition (g kg-1) and forage crude protein yield (kg ha-1) in barrel medic cultivars at Rimski Šančevi in 2010Tabela 6. Veličine vezane za hemijski sastav suve materije krme (g kg-1) i prinos sirovih proteina krme (kg ha-1) sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar / Sorta
Forage dry matter crude
protein content /
Sadržaj sirovih proteina suve materije krme
Forage crude protein yield /Prinos sirovih proteina krme
Forage neutral detergent fi bre
content /Sadržaj
neutralno-deterdžentskih vlakana krme
Forage acid detergent fi bre
content /Sadržaj kiselo-deterdženstkih vlakana krme
Forage lignin content /
Sadržaj lignina krme
Borung 165.7 985 370.5 327.7 78.9Caliph 162.5 1079 458.8 359.6 78.1Jemalong 162.6 1187 335.1 290.3 75.7Jester 179.4 879 377.8 311.8 103.8Mogul 159.8 950 340.6 309.1 87.3Parabinga 150.8 1206 386.7 327.5 101.6Paraggio 167.3 511 333.3 280.2 65.3Sephi 174.7 473 305.2 296.5 62.5Average / Prosek 165.4 909 363.5 312.8 81.6LSD0.05 13.9 376 68.4 40.1 19.8

Mihailović V et al.250
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
mind extremely unfavourable conditions for seed production during its growing season in 2010 and the average lucerne seed yields in Serbia between 255 kg ha-1 and 468 kg ha-1 (Karagić et al. 2010). The average thousand seed weight in eight barrel medic cultivars ranged from 3.6 g in Jester to 4.1 g in Paraggio, remaining within the limits of the species (Frame 2010) and confi rming that the seeds in all barrel medic cultivars reached their ordinary size. A large variation in the values of seed viability among the tested cultivars, from 49% in Parabinga to 95% in Mogul, witness the existence of hard seeds in certain cultivars, while it remains unexplained if it is a specifi c interaction of individual genotype with environment or a consequence of different proteomic processes during seed development in diverse cultivars (Gallardo et al. 2003).
Green manure
The cultivar Jester had the largest forage dry matter nitrogen content (28.7 g kg-1) while the cultivar Parabinga had the lowest forage dry matter nitrogen content (24.1 g kg-1) (Tab. 8). Paraggio and Sephi produced signifi cantly lower aboveground nitrogen yield (82 kg ha-1 and 76 kg ha-1) than the remaining barrel medic cultivars, especially Jemalong (190 kg ha-1) and Parabinga (193 kg ha-1). Other annual legumes, such as grass pea (Lathyrus sativus L.), had the aboveground nitrogen yield of about 340 kg ha-1 in the same agro-ecological conditions (Mikić et al. 2010).
Table 7. Parameters related to seed yield in barrel medic cultivars at Rimski Šančevi in 2010Tabela 7. Veličine vezane za prinos semena sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar / Sorta
Number of days from sowing to
harvest /Broj dana od setve
do žetve
Seed yield (kg ha-1) / Prinos semena
Thousand seed weight (g) /Masa hiljadu
semena
Seed viability (%) / Klijavost
semena
Borung 92 159 3.7 55Caliph 92 217 3.9 79Jemalong 90 228 3.8 57Jester 92 338 3.6 81Mogul 106 199 3.9 95Parabinga 90 276 4.0 49Paraggio 104 395 4.1 94Sephi 94 432 3.9 52Average / Prosek 95 281 3.9 70LSD0.05 3 121 1.5 23
Table 8. Forage dry matter nitrogen content (g kg-1) and aboveground nitrogen yield (kg ha-1) in barrel medic cultivars at Rimski Šančevi in 2010Tabela 8. Sadržaj azota suve materije krme (g kg-1) i prinos nadzemnog azota (kg ha-1) sorti buretaste lucerke na Rimskim Šančevima 2010. godine
Cultivar / Sorta
Forage dry matter nitrogen content (g kg-1) /
Sadržaj azota suve materije krme
Aboveground nitrogen yield (kg ha-1) /
Prinos nadzemnog azota
Borung 26.5 158Caliph 26.0 173Jemalong 26.0 190Jester 28.7 141Mogul 25.6 152Parabinga 24.1 193Paraggio 26.8 82Sephi 28.0 76Average / Prosek 26.5 145LSD0.05 2.1 48

Preliminary Results on Agronomic Performance of Barrel Medic in Serbia 251
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
Conclusions
Although preliminary, the results of this agronomic performance evaluation of barrel medic in Serbia show that it has a certain potential for forage production. It is noteworthy that, even in a rainy and a warm spring and summer of 2010, it achieved satisfying forage yields of good quality and, in several cases, reliable seed yields, both during a relatively brief growing season. In that way, barrel medic could fi nd its place as a short-lived forage or green manure crop in diverse cropping systems with benefi ts for both animal husbandry and soil fertility. At the same time, its promising preliminary agronomic performance opens a possibility for the evaluation of the results of various genomic research in fi eld trials on chernozem and other prevailing soil and weather conditions in Serbia. Another fi eld trial season with barrel medic has already begun with establishing the evaluation of cold tolerance in the autumn 2010.
References
Annicchiarico P (2007): Inter- and intra-population genetic variation for leaf : stem ratio in landraces and varieties of lucerne. Grass Forage Sci. 62: 100-103
Annicchiarico P, Scotti C (2009): Some burning questions for lucerne breeders. Grain Legum. 51: 18-19
Choi H-K, Kim D, Uhm T, Limpens E, Lim H, Mun J-H, Kalo P, Varma Penmetsa R, Seres A, Kulikova O, Roe B A, Bi-sseling T, Kiss G B, Cook D R (2004): A sequence-based genetic map of Medicago truncatula and comparison of marker colinearity with M. sativa. Genet. 166: 1463-1502
Cook D R (1999): Medicago truncatula—a model in the ma-king! Curr. Opin. Plant Biol. 2: 301-304
Crawford E J, Lake A W H, Boyce K G (1989): Breeding annual Medicago species for semi-arid conditions in southern Austra-lia. Adv. Agron. 42: 399-437
Ellwood S R, D’ Souza N K, Kamphuis L G, Burgess T I, Nair R M, Oliver R P (2006): SSR analysis of the Medicago trunca-tula SARDI core collection reveals substantial diversity and unusual genotype dispersal throughout the Mediterranean basin. Theor. Appl. Genet. 112: 977-983
Frame J (2010): Grassland Species Profi les [Online]. [1 p] Avai-lable at http://www.fao.org/ag/AGP/AGPC/doc/Gbase/data/meditrunc.htm (cited 29 November 2010, verifi ed 29 November 2010). FAO, Rome
Gallardo K, Le Signor C, Vandekerckhove J, Thompson R D, Bur-stin J (2003): Proteomics of Medicago truncatula seed development establishes the time frame of diverse metabolic processes rela-ted to reserve accumulation. Plant Physiol. 133: 664-682
Julier B, Meusnier I (2010): Alfalfa breeding benefi ts from ge-nomics of Medicago truncatula. Ratar. Povrt. / Field Veg. Crop Res. 47: 395-402
Julier B, Huyghe C, Ecalle C (2000): Within- and among-culti-var genetic variation in alfalfa: Forage quality, morphology, and yield. Crop Sci. 40: 365-369
Kaló P, Seres A, Taylor S A, Jakab J, Kevei Z, Kereszt A, En-dre G, Ellis T H N, Kiss G B (2004): Comparative mapping between Medicago sativa and Pisum sativum. Mol. Genet. Ge-nomics 272: 235-246
Karagić Đ, Jevtić G, Terzić D (2010): Forage legumes seed pro-duction in Serbia. Biotechnol. Anim. Husb. 26 (special issue): 1: 133-148
Katić S, Mihailović V, Milić D, Karagić Đ, Glamočić D, Jajić I (2008): Genetic and seasonal variations of fi bre content in lucerne. Proceedings, XXVIIth EUCARPIA Symposium on Improvement of Fodder Crops and Amenity Grasses, Copenhagen, Denmark, 19-23 August 2007, 130-135
Katić S, Milić D, Karagić Đ, Vasiljević S, Glamočić D, Jajić I (2009): Variation of protein, cellulose and mineral contents of lucerne as infl uenced by cultivar and cut. Biotechnol. Anim. Husb. 25: 1189-1195
Mihailović V, Mikić A, Ćupina B, Erić P (2005): Field pea and vetches in Serbia and Montenegro. Grain Legum. 44: 25-26
Mihailović V, Mikić A, Vasiljević S, Katić S, Karagić Đ, Ćupina B (2008): Forage yields in urban populations of hairy vetch (Vi-cia villosa Roth) from Serbia. Grassl. Sci. Eur. 13: 281-283
Mihailović V, Mikić A, Katić S, Karagić Đ, Milošević B (2010): Potential of fi eld pea for forage and grain protein yields. Ra-tar. Povrt. / Field Veg. Crop Res. 47: 43-48
Mikić A (2008): Genetic variability of agronomic characteristics of common vetch (Vicia sativa L.). MSc thesis. Faculty of Agriculture, University of Novi Sad, Novi Sad, Serbia
Mikić A, Mihailović V, Ćupina B, Krstić Đ, Hauptvogel R, Drobná J, Antalíková G (2009): Forage yields in urban popu-lations of large-fl owered vetch (Vicia grandifl ora Scop.) from Serbia. Grassl. Sci. Eur. 14: 421-424
Mikić A, Mihailović V, Ćupina B, Đorđević V, Stoddard F L (2010a): Introduction of novel legume crops in Serbia – Whi-te lupin (Lupinus albus). Field Veget. Crop. Res. 47: 21-26
Mikić A., Mihailović V, Ćupina B, Ðurić B, Krstić Đ, Vasić M, Va-siljević S, Karagić Đ, Ðorđević V (2010b): Towards the re-intro-duction of grass pea (Lathyrus sativus) in the West Balkan Coun-tries: The case of Serbia and Srpska (Bosnia and Herzegovina). Food Chem. Toxicol. (in press) doi:10.1016/j.fct.2010.07.052
Mikić V, Radović J, Vasiljević S, Milić D (2007): Correlations between agronomic characteristics of lucerne (Medicago sati-va L.) genotypes as affected by cutting. Proceedings, XXVI EUCARPIA Fodder Crops and Amenity Grasses Section and XVI Medicago spp. Group Joint Meeting, Perugia, Italy, 3-7 September 2006, 172-175
Rubiales D, Fernández-Aparicio M, Moral A, Pujadas A (2009): Collection and conservation of Iberian Medicago spp. germ-plasm. Grain Legum. 53: 29
Stout D G, Brooke B, Hall J W, Thompson D J (1997): Forage yield and quality from intercropped barley, annual ryegrass and different annual legumes. Grass Forage Sci. 52: 298-308
Thompson R, Aubert G, Duc G, Gallardo K, Lesignor C (2009): Medicago truncatula as a model legume. Grain Legum. 53: 5
Tivoli B, Baranger A, Sivasithamparam K, Barbetti M J (2008): Annual Medicago: From a model crop challenged by a spec-trum of necrotrophic pathogens to a model plant to explore the nature of disease resistance. Ann. Bot. 98: 1117-1128
Vasiljević S, Glamočić D, Jajić I, Ćupina B, Katić S, Milić D, Mikić A (2008): Fibre fraction of red clover (Trifolium pratense L.) at dif-ferent harvest over two seasons. Grassl. Sci. Eur. 13: 510-512
Vasiljević S, Milić D, Mikić A (2009): Chemical attributes and quality improvement of forage legumes. Biotechnol. Anim. Husb. 25: 493-504
Walsh M J, Delaney R H, Groose R W, Krall J M (2001): Per-formance of annual medic species (Medicago spp.) in south-eastern Wyoming. Agron. J. 93: 1249-1256
Young N, Mudge J, Ellis T (2003): Legume genomes: more than peas in a pod. Curr. Opin. Plant Biol. 6: 199-204
Zhu Y, Sheaffer C C, Barnes D K (1996): Forage yield and qual-ity of six annual Medicago species in the north-central USA. Agron. J. 88: 955-960
Zhu Y, Sheaffer C C, Russelle M P, Vance C P (2008): Dry mat-ter accumulation and dinitrogen fi xation of annual Medicago species Agron. J. 90: 103-108

Mihailović V et al.252
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 245-252
Prvi rezultati gajenja buretaste lucerke (Medicago truncatula) u Srbiji
Vojislav Mihailović1 · Aleksandar Mikić1 · Branko Ćupina2 · Dragan Milić1 · Đorđe Krstić2 · Slobodan Katić1 · Sanja Vasiljević1 · Đura Karagić1
1Institut za ratarstvo i povrtarstvo, Maksima Gorkog 30, 21000 Novi Sad, Srbija2Poljoprivredni fakultet Univerziteta u Novom Sadu, Trg Dositeja Obradovića 8, 21000 Novi Sad, Srbija
Izvod: Mikro-ogled sa osam australijskih sorti buretaste lucerke (Medicago truncatula Gaertn.) izveden je na Rim-skim Šančevima 2010. Najveći prosečni prinos zelene krme i suve materije krme bio je kod sorti Jemalong (30,7 t ha-1 i 7,3 t ha-1) i Parabinga (30,7 t ha-1 i 8,0 t ha-1). Sadržaj sirovih proteina suve materije krme kretao se između 150,8 g kg-1 kod sorte Parabinga i 179,4 g kg-1 kod sorte Jester. Sadržaj neutralno-deterdžentskih vlakana (NDF) krme kretao se od 305,2 g kg-1 kod sorte Sephi do 458,8 g kg-1 kod sorte Caliph, dok je prosečni sadržaj kiselo-deterdžentskih vlakana (ADF) bio 312,8 g kg-1. Prosečan prinos semena svih sorti iznosio je 281 kg ha-1 i može se smatrati zaodovoljavajućim s obzirom na veoma kišoviti i topli vegetacioni period vrste. Sorte Jemalong i Parab-inga ostvarile su najveći prinos nadzemnog azota (190 kg ha-1 i 193 kg ha-1) i time najveći potencijal za zelenišno đubrenje.Ključne reči: buretasta lucerka, kiselo-deterdžentska vlakna, kvalitet krme, Medicago truncatula, model mahunarke, neutralno-deterdžentska vlakna, prinos krme, prinos semena, prinos sirovih proteina krme, zelenišno đubrenje

The Legume Manifesto 253
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
The Legume Manifesto: (Net)workers on Fabaceae, Unite!
Aleksandar Mikić · Diego Rubiales · Petr Smýkal · Frederick L. Stoddard
received / primljeno: 25.11.2010. accepted / prihvaćeno: 26.11.2010.© 2011 IFVC
Summary: Legumes have been an important part of cropping systems since the dawn of agriculture. The shift in Europe from draught animals to meat animals coincided with the increasing availability of soybean meal from North and South America, and the Common Agricultural Policy of the European Union promoted the growing of cereals and oilseeds at the expense of other crops, so legumes fell out of favour with farmers and decision-makers. Continental concerns about food and feed security, high prices of oil and soybean meal, and advances in the application of fundamental molecular genetics to crop species, all mean that now is a good opportunity to promote the return of legumes to European cropping systems by enhancing the effi ciency of research and development on this family. Hence we propose the establishment of a Legume Society that will promote information exchange and scientifi c productivity by uniting the various legume research communities.Key words: food and feed security, Legume Society, modelling, networking, sustainability
A. Mikić Institute of Field and Vegetable Crops, Novi Sad, SerbiaD. RubialesCSIC, Institute for Sustainable Agriculture, Córdoba, SpainP. SmýkalAgritec Plant Research Ltd., Šumperk, Czech RepublicF. L. StoddardUniversity of Helsinki, Department of Agricultural Sciences, PO Box 27 (Latokartanonkaari 5), FIN-00014 Helsinki, Finlande-mail: [email protected]
Legumes, Past Tense: Once Upon a Time Th ere Were Legumes
The Fabaceae is the third largest family of fl owering plants, after Orchidaceae and Asteraceae, with over 650 genera and 20,000 species (Lewis et al. 2005). It is an extremely diverse family of worldwide distribution, encompassing everything from arctic alpine herbs, to annual xerophytes and equatorial forest trees. Members of the family are characterized by the distinct fruit, termed a legume or pod, which gives the family its name. Another peculiarity of legumes is their symbiosis with nitrogen-fi xing bacteria that provides not only added value in agriculture, but also plays important role in natural ecosystems.
Legumes have always been a part of the everyday life of humans. Grain legumes such as pea (Pisum sativum L.), bitter vetch (Vicia ervilia (L.) Willd.), lentil (Lens culinaris Medik.), chickpea
(Cicer arietinum L.), faba bean (Vicia faba L.) and grass pea (Lathyrus sativus L.) are commonly recognized as among the fi rst domesticated plant species and the most ancient crops, contributing to the ‘agricultural revolution’ in the Fertile Crescent at the end of the last Ice Age (Bellwood 2005). Subsequently and independently, soybean (Glycine max (L.) Merr.) was domesticated in China, cowpea (Vigna unguiculata (L.) Walp.) in Africa, pigeon pea (Cajanus cajan (L.) Millsp.) in India and common bean (Phaseolus vulgaris L.) twice, in central and South America. Apart from these food legumes, forage legumes have been adapted from wild fl ora and used in managed grazing lands, and include lucerne (Medicago sativa L.), clovers (Trifolium spp.) and birdsfoot trefoil (Lotus corniculatus L.), providing animal husbandry with quality forage, although the history of domestication is much more obscure.
The most important and widely cultivated legume crops began to be studied fi rst, such as pea, the key experimental organism for Mendel’s pioneering genetic research (Ellis 2007), soybean, lucerne and white clover. They are followed progressively by the less widespread ones, now reaching vetchlings (Lathyrus spp.) and vetches (Vicia spp.), legumes with pharmaceutical properties including fenugreek (Trigonella foenum-graecum L.) and liquorice (Glycyrrhiza glabra L.), economically important tree legumes such as
Views / Stavoviw
ww.
nsse
me.
com
/jour
nal.h
tml
( )

Mikić A et al.254
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
Robinia and Pongamia, and garden ornamentals such as sweet pea (Lathyrus odoratus L.). Certain legume crops have well-studied genetic systems. Yet, in many of these, comprehensive genetic analysis is limited due to the large size of their genomes. Thus in order to expedite and simplify genome analysis of crop legumes, model species with smaller genomes were selected, one being Medicago truncatula and Lotus japonicus another. Genomes of both species have been sequenced extensively by end of 2008, as has the genome of soybean in 2010, economically the most important legume crop.
Each segment of legume research began to develop its own network, maintained by diverse means, ranging from pure enthusiasm to specifi c fi nancial support by projects, and improved through regular meetings. Some of the largest networks dealing with legumes such as European Grassland Federation (EGF) organize annual meetings, while by contrast the European Association for Plant Breeding Research (EUCARPIA), with its sections on fodder crops and amenity grasses, oil and protein crops, ornamental plants, and genetic resources, has irregular meetings where different aspects of legumes are covered by different groups. The European Association for Grain Legume Research (AEP) was established to support specifi cally grain legumes and, apart from its triennial conferences, actively participated in establishing various Framework Programme (FP) projects (Ellis et al. 2005, Schneider 2007) such as FP4 LINK, FP5 EU-Faba, FP5 GL-Pro, FP6 EUROCROP and FP6 Grain Legumes Integrated Project (GLIP) with its Grain Legumes Technology Transfer Platform (GL-TTP). Some global legume communities have existed mainly in the form of regular conferences such as the International Food Legume Research Conference (IFLRC), International Conference on Legume Genomics and Genetics (ICLGG) and European Nitrogen Fixation Conference. There are also long-term programmes such as the European Cooperative Programme for Plant Genetic Resources (ECPGR) with its working groups and narrowly specialized groups such as the Pisum Genetics Association (PGA) and EUCARPIA Medicago species group.
Legumes, Present Tense: How Can We Res-cue the Disappearing Legume Community?
Legumes are established as one of the most important crop groups with extraordinarily diverse uses. They are recognized as a quality source of
protein for human consumption and in animal feed, their starch has the nutritional advantage of being slowly digestible, while others such as soybean are rich in oil (Đorđević 2009). Their impact on soil and its properties is positive for all succeeding crops, since they live in symbiotic community with nitrogen-fi xing bacteria and both renew and enrich the ecosystem fertility (Stoddard et al. 2009). They are also a source of various bioactive compounds that are used in pharmaceutical industries. Perennial and tree legumes may represent a valuable biofuel and ornamental legumes contribute to the beauty of living.
Despite all these and many other benefi ts for all of society, legume cropping has been in decline throughout Europe. Legumes are often considered not suffi ciently profi table in comparison to other major crops such as cereals or oilseed brassicas, in a context where crop profi tability is largely determined by subsidies set by decision-makers. The decrease in the legume cultivation area causes a decrease in the support for legume research. In the end, this badly affects the legume research communities and their networks, until the European legume research community may be likened to galaxies following the cosmological Big Bang (Fig. 1).
Fig. 1. Current situation in the European legume research community as compared to the Big Bang model in cosmology: with the fl ow of time and in ever-expanding space, each community behaves like a galaxy and becomes more and more remote from the others and with continuously weakened mutual bonds
Slika 1. Trenutne okolnosti u evropskoj istraživačkoj zajednici mahunarki u poređenju sa modelom Velikog praska u kosmologiji: tokom vremena i u svemiru koji se večno širi, svaka zajednica se ponaša poput galaksije i sve se više udaljava od ostalih uz postojano slabljenje međusobnih veza

The Legume Manifesto 255
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
Legume research in the USA, Australia and Canada, unlike that in Europe, demonstrates how a feasible and sustainable link between basic science, applied research and end-users may be established and maintained (Warkentin et al. 2008). Organizations such as Pulse Australia and Pulse Canada thrive much better than their counterparts in Europe, in spite of the latter being the home to many of the fi nest discoveries in legume research, especially its basic science. This is the reason why legumes occupied less than 2% of European arable area in 2007-2009, in contrast to 6% in Australia and 10% in Canada, and why the EU imports over 70% of its plant-derived protein, including 21-32 Mt soy meal annually from 2000 to 2007.
Perhaps the EU FP6 project GLIP was the fi nal mover in the European legume research community. Its support to the networks such as AEP and GL-TTP was essential and since it ended, both are in serious fi nancial trouble and unable to carry out their mission of enhancing legume research and even to maintain their membership. With the exception of FP7 project Legume Futures and some multilateral projects, there is currently little EU investment in legumes. Opportunities for synergy and interaction are therefore progressively harder to fi nd and develop.
Many legume scientists therefore became concerned during 2009 and 2010: How could the disintegration of the existing legume research communities and their networks in Europe be reversed?
Legumes, Future Tense: And Th ey Grew Legumes Happily Ever After
The critical time for action has come and the next fi ve years must be a turning point for legume research networking (Mikić & Smýkal 2010). A society to assemble all legume researchers in Europe will be established, provisionally named the Legume Society (LS), and its network will assist in healing the wounds in the European legume research community.
The purposes of such pan-European, all-legume, interactive, multifunctional and sustainable society and its network are three-fold: scientifi c, economic and social.
1. Scientifi c benefi ts.- The national, bilateral and multilateral
projects, funded by national and international bodies, will be linked together in an effi cient, useful and productive way;
- The results and the databases will be more accessible, available and applicable and the general fl ow of information will be eased;
- Improved coordination will lead to a more sustainable actions and better use of existing fi nancial resources on national and EU levels;
- The network will interact with non-European legume science networks, exchanging ideas and best practices.
2. Economic benefi ts.- The network will include food, feed and non-
food industry, extension services, farmers, stake-holders and all other end-users (Smýkal et al. 2010);
- The network will lobby European decision-makers for more favourable solutions for legumes, such as an end to the anti-legume weighting of the Common Agricultural Policy, which would allow replacement of at least 50% of imported soybean meal by a 4-fold increase in European grain legume areas;
- The network will promote legumes as the crops that:
symbiotically fi x nitrogen and do not require synthetic N fertilizers, resulting in cost savings and reductions in energy inputs, greenhouse gas emission and water pollution;
when handled correctly, are competitive with weeds and are highly effective in mixed cropping, particularly of annual legumes either with annual cereals or, at the establishment phase, with perennial forage legumes, resulting in reduced costs and pollution together with higher forage and feed yields;
often have allelopathic properties that destroy or suppress parasites such as broomrape, some nematodes and certain pathogenic fungi, allowing a reduction in the use of agrochemicals in integrated systems;
and are multi-functional crops providing food, feed, fi bre and fuel, with benefi ts to the following crop, thus enhancing the diversity of farm income streams, farm economic viability, and sustainability, and providing a broader range of raw materials to processing industries.
Social benefi ts.- The network will promote the benefi t to
the whole system, encompassing ecological and social aspects of legumes as crops that:
improve mechanical and chemical soil properties;
have benefi cial effects by their root exudates upon soil microorganisms;
are the best choice to remediate soils contaminated by mines or power plants;
reduce cereal crop diseases;renew soil fertility;

Mikić A et al.256
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
reduce the negative impacts of agriculture on climate and help mitigate negative climatic changes;
enrich agroecosystems and landscapes, contribute to regional biodiversity and support pollinating bee and other useful insect populations;
assist in the conservation of the traditional ways of living, improve the viability of rural communities, and renew the interest of younger generations in agricultural production and its benefi ts;
provide the food and feed sector with healthy, safe and rich source of plant protein, the pharmaceutical sector with raw materials and the energy sector with sustainable biofuels;
and contribute to the overall health and thus to the good of society.
The structure of LS should be light and effi cient, comprising both managing roles, in the form of Executive Committee headed by President, and scientifi c counselling, represented by Scientifi c Committee with several sections following the legume research topics as the most natural links between the individual legume crop groups.
Among the innovative aspects of the idea about ILS and its network there are:
- The fact that all legumes are to be covered by one society and managed by one network;
- The transfer of information from the model legumes to the cultivated species will be made easier;
- Novel uses will no longer risk falling into the gaps between specialties, for example when a grain legume is assessed for use as a forage or green manure;
- Facilitation of re-introduction of neglected and underutilized crops such as grass pea and cowpea in southern Europe, or faba bean and lentil in the Balkans;
- Facilitation of introduction of novel crops, such as alkaline soil-tolerant white lupin or photoperiod-neutral pigeon pea in temperate Europe for forage and grain production;
- Use of legumes as bioherbicides, bioinsecticides, biofumigants and other biopesticides, such as fenugreek against broomrape in faba bean, pea against weeds in perennial forage legume establishment (Ćupina et al. 2010);
- Use of legumes as cover crops, catch crops, mulch and biofuel;
- Use of legumes in soil remediation and erosion prevention;
- Innovations in genomics and other omic basic sciences;
- Innovations in legume quality, chemical composition and biofortifi cation;
- Innovations in place, role and potential in various cropping systems.
The rough agenda of the LS development during the years to come comprises three major steps (Fig. 2).
- Firstly, enhanced contact among legume scientists belonging to different crop communities should result in enriched exchanges of ideas and scientifi c outcomes;
- Gradually, the expected synergy between soybean, grain, forage and tree legume research will develop (Rubiales 2009);
- Legume scientists from the EU and its converging regions will be brought together so the network covers the whole Enlarged European Research Area, fi lling the geographical gaps that have existed for decades and caused delays in progress;
Fig. 2. A provisional LS agenda: existing contacts between various legume communities, as initiated especially by AEP and GL-TTP, serve as a condensation centre that delivers the LS, during the next fi ve years and at least on a European level, as the main gear that would give power to diverse legume research communities and that later would bind together the legume research on a global level, integrating legume research in Europe as a whole (EU+) with that in USA, Canada (CAN), Australia (AUS), Brazil, Russia, India and China (BRIC) and third countries (3rd C)
Slika 2. Okvirni rokovnik Društva za mahunarke (LS): postojeće veze između različitih zajednica mahunarki, začete posebno od strane AEP i GL-TTP, služe kao središte zgušnjavanja koje rađa LS, tokom narednih pet godina i najmanje u evropskoj ravni, kao glavni zupčanik koji pokreće razne istraživačke zajednice mahunarki i kasnije objedinjava istraživanje mahunarki u svetskoj ravni, integrišući istraživanje Evrope u celini (EU+) sa onim u SAD (USA), Kanadi (CAN), Australiji (AUS), Brazilu, Rusiji, Indiji i Kini (BRIC) i zemljama Trećeg sveta (3rd C)

The Legume Manifesto 257
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
- The LS will welcome participants from non-European legume communities, from: (1) large producers, such as USA, Canada, Australia, Brazil, Russia, India and China, with the main goal of exchange of knowledge and joint enhancement of the global legume production and use; (2) third countries, such as those in Asia, Africa and South America, for technology transfer from the core countries, know-how improvement in target regions, and unbiased information exchange, with essential help from the CGIAR centres such as ICARDA, ICRISAT and IITA and with the aim of increasing food, feed, fi bre and fuel supplies as well as the quality of life in all countries.
***
Establishment of the LS requires little more than enthusiasm, good will, and members’ time. Its maintenance will require the continuing efforts of the current legume research network in Europe to provide it with the necessary token fi nancial support and, at times, a little volunteer labour. We expect that this support will be returned many times over in the improved success rate of research grant applications and research outcomes. Perhaps the greatest opportunity to fulfi l this task lies in the form of a COST action, where the proposal on a European legume research network has been submitted and is being processed at the time of writing. In fact, LS and its network already exist in an embryonic form, demonstrating that the goals of this manifesto are feasible and already half-achieved.
References
Bellwood P (2005) First Farmers, the Origins of Agricultural Societies. Blackwell Publishing, Oxford
Ćupina B, Krstić Đ, Mikić A, Erić P, Vučković S, Pejić B (2010): The effect of fi eld pea (Pisum sativum L.) companion crop ma-nagement on red clover (Trifolium pratense L.) establishment and productivity. Turk. J. Agric. For. 34: 275-283
Đorđević V (2009): Perspective of soybean in Europe. Grain Legum. 51: 16-17
Ellis T H N (2007): Approaches to the isolation of genes of agronomic importance in pea. Zbornik radova Instituta za ratarstvo i povrtarstvo / A Periodical of Scientifi c Research on Field and Vegetable Crops 44: 45-47
Ellis N, Schneider A, Golstein C (2005): GLIP grain legumes integrated project, new strategies to improve grain legumes for food and feed. Abstracts, 4th International Food Legu-mes Research Conference Food Legumes for Nutritional Security and Sustainable Agriculture, New Delhi, India, 18-22 October 2005, 42.
Lewis G, Schrire B, MacKinder B, Lock M (2005): Legumes of the World. Royal Botanical Gardens, Kew
Mikić A, Smýkal P (2010): International Legume Society - the future of all-legume networking. Book of Abstracts, 5th In-ternational Food Legumes Research Conference & 7th Eu-ropean Conference on Grain Legumes Legumes for Global He-alth – Legume Crops and Products for Food, Feed and Environmental Benefi ts, Antalya, Turkey, 26-30 April 2010, 269
Rubiales D. (2009): This is the time for grain and forage legu-mes to come back! Grain Legum. 51: 4
Schneider A (2007): Opportunities for complementary sources of funds for GL-TTP projects. Abstracts, 1st Grain Legu-mes Technology Transfer Platform (GL-TTP) Workshop Targeting Science to Real Needs, Paris, France, 23-25 April 2007, 17
Smýkal P, Ellis T H N, Bochard A M, Warkentin T, Mikić A, Schneider A (2010): Grain Legumes Technology Transfer Platform (GL-TTP): a step towards the integration of grain, forage and other legume communities. Biotechnol. Anim. Husb. 26 (special issue) 2: 111-116
Stoddard F L, Hovinen S, Kontturi M, Lindström K, Nykänen A (2009): Legumes in Finnish agriculture: history, present status and future prospects. Agric. Food Sci. 18: 191-205
Warkentin T, Vandenberg A, Tar’an B, Banniza S, Bett K (2008): Field pea genetic improvement at the University of Saskat-chewan. Proceedings, International Conference Conventional and Molecular Breeding of Field and Vegetable Crops, Novi Sad, Serbia, 21-24 November 2008, 372

Mikić A et al.258
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 253-258
Legumistički manifest: (Net)proleteri svih Fabaceae, ujedinite se!
Aleksandar Mikić1 · Dijego Rubijales2 · Petr Smikal3 · Frederik L. Stodard4
1Institut za ratarstvo i povrtarstvo, Novi Sad, Srbija2CSIC, Institut za održivu poljoprivredu, Kordova, Španija
3Istraživanja na biljkama Agritek, s.o.o, Šumperk, Češka4Univerzitet u Helsinkiju, Departman poljoprivrednih nauka, Helsinki, Finska
Izvod: Mahunarke su bile važan deo sistema ratarenja od osvita poljoprivrede. Pomak sa ispaše na stajaću stoku u Evropi odigrao se istovremeno sa povećanjem raspoloživih količina sojine sačme iz Severne i Južne Amerike, dok je Zajednička poljoprivredna politika Evropske Unije promovisala gajenje žitarica i kupusnjača na račun ostalih useva, te su mahunarke izgubile ugled kod proizvođača i struktura vlasti. Pažnja koju Evropa ukazuje bezbednosti ljudske i stočne hrane, visoke cene uljane i sojine sačme i dostignuća u primeni osnovne molekularne genetike kod gajenih vrsta ukazuju da je u ovom trenutku dobra prilika za promociju povratka mahunarki u evropske sisteme ratarenja putem povećanja efi kasnosti istraživanja i razvoja na ovoj familiji. Stoga predlažemo osnivanje Društva za mahunarke koje će promovisati razmenu informacija i naučnog stvaralaštva ujedinjenjem različitih istraživačkih zajednica mahunarki.Ključne reči: bezbedna ljudska i stočna hrana, Društvo za mahunarke, modeliranje, netvorking, održivost

Crvenilo kukuruza: pola veka kasnije 259
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 259
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 259
Živko Ćurčić: Oplemenjivanje šećerne repe na rizomaniju / Sugar Beet
Breeding on RhizomaniaZadužbina Andrejević, Beograd i Institut za ratarstvo i
povrtarstvo, Novi Sad, 2010, 80 str.
Monografi ja „Oplemenjivanje šećerne repe“ predstavlja pregledno i jasno izloženo štivo u 7 poglavlja. Poglavlja su napisana logičnim redo-sledom sa jasnim naslovima, tako da se prate i razumeju bez poteškoća. Proučena brojna litera-tura (165 navoda) koja se odnosi na problematiku istraživanja doprinela je da se dobijeni rezultati istraživanja prodiskutuju i uporede sa rezultatima drugih autora, koji su radili na istoj ili sličnoj pro-blematici.
Autor nas u prvim poglavljima upoznaje sa rizomanijom, jednim od najznačajnijih oboljenja šećerne repe i mogućnostima njenog suzbijanja. U četvrtom poglavlju su prikazane metode ople-menjivanja šećerne repe na otpornost prema ri-zomaniji, od klasičnih metoda oplemenjivanja do savremenih, pomoću molekularnih markera i po-stizanja transgene otpornosti prema ovoj bolesti. Takođe je prikazan značaj poznavanja kombina-
cionih sposobnosti linija različitih tipova otpor-nosti prema rizomaniji, zbog stvaranja hibrida visokih prinosa odgovarajućeg kvaliteta korena.
Naučni doprinos ovog rada ogleda se pre sve-ga u njegovoj originalnosti, jer je ispitivana veza između kombinacionih sposobnosti linija šećerne repe i tipova otpornosti prema rizomaniji o koji-ma ima malo literaturnih podataka. Zaključeno je da bi uticaj izvora otpornosti prema rizomaniji na kombinacione sposobnosti trebalo još ispitivati, kako bi se došlo do pravilnih zaključaka, jer na osnovu njegovog istraživanja nisu uočene značaj-ne razlike između Holly tipa otpornosti i WB42 tipa otpornosti. Istraživanja bi trebalo izvesti na zemljištima sa visokim stepenom infekcije viru-som rizomanije, kako bi se mogle bolje ispitati razlike između ova dva tipa otpornosti.
dr Nevena Nagl
Prikaz knjige / Book Revieww
ww.
nsse
me.
com
/jour
nal.h
tml
Živko Ćurčić: Oplemenjivanje šećerne repe na rizomaniju / Sugar Beet Breeding on Rhizomania
Zadužbina Andrejević, Beograd and Institute of Field and Vegetable Crops, Novi Sad, 2010, 80 pages
Published in Serbian and English, this monograph is a clear review written in 7 chapters logically ordered with clear titles, so that it is easily followed and understood. A large number of references (165) regarding the research issue enabled extensive discussion of the results and comparison to other relevant articles. The fi rst chapters introduce rhizomania as one of the most important sugar beet diseases, while the fourth chapter displays sugar beet breeding methods aiming at resistance to rhizomania, from classic breeding methods to contemporary ones, including molecular markers and transgenic resistance to this disease. Scientifi c contribution of this monograph lies primarily in its original research on the connection between combining abilities of sugar beet lines on the one hand, and types of resistance to rhizomania on the other hand, which is generally seldom dealt with.

Bekavac G i sar.260
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 260
Ratar. Povrt. / Field Veg. Crop Res. 48 (2011) 260
Đuro Gvozdenović: PaprikaInstitut za ratarstvo i povrtarstvo,
Novi Sad, 2010, 286 str.
Monografi ja „Paprika“ obrađuje istorijat i poreklo paprike, površine i prinose, privredni značaj, hemijski sastav i hranljivu vrednost, bo-taničku klasifi kaciju, morfološke osobine, uslove uspevanja, tehnologiju gajenja, obradu u toku vegetacije, proizvodnju paprike na polietilenskoj foliji, proizvodnju paprike direktnom setvom semena, proizvodnju paprike u zaštićenom pro-storu, zaštitu paprike, hidroponski način gajenja paprike, ekološku (organsku) proizvodnju, ople-menjivanje i proizvodnju semena paprike.
Gajenje i oplemenjivanje paprike zauzimaju centralni i najobimniji deo ove monografi je, gde je autor koristio svoje bogato iskustvo i rezulta-te vlastitih istraživanja, kao i istraživanja velikog broja istraživača kako naših tako i inostranih, oblikujući ih u jednu homogenu celinu, dajući na taj način veću vrednost pojedinačnim rezultatima i svim rezultatima zajedno.
Autor je posebno obradio poglavlje „Proi-zvodnja semena paprike“, veoma značajnu oblast u gajenju paprike, koja nije do sada bila lako dostu-pna čitaocima, a posebno proizvođačima paprike. Naime, proizvodnja semena paprike predstavlja značajnu privrednu aktivnost u našoj zemlji, pre svega jer je ekonomski veoma rentabilna.
Ova monografi ja predstavlja značajnu publi-kaciju iz oblasti gajenja, oplemenjivanja i proi-zvodnje semena paprike. Napisana je lakim sti-lom, za sve čitaoce razumljivim tečnim jezikom, tako da je pristupačna širem krugu čitalaca, kako stručnjacima iz ove oblasti tako i drugim kori-snicima u produbljivanju njihovih znanja iz ove tematike, a pre svega proizvođačima paprike, stu-dentima master i doktorskih studija poljoprivred-ne i biološke struke.
prof. dr Žarko Ilin
Prikaz knjige / Book Revieww
ww.
nsse
me.
com
/jour
nal.h
tml
Đuro Gvozdenović: Pepper (in Serbian)Institute of Field and Vegetable Crops, Novi Sad, 2010, 286 pages
This monograph deals with history and origins of pepper, its acreage and yield, economic importance, chemical compositions and nutritional values, as well as morphologic features, growing conditions, cultivation practices, various production modes (including hydroponic and organic production), breeding and seed production. Central and most extensive part of this book regards pepper growing and breeding, supported by author’s own rich experience and research, together with numerous results of other relevant authors. The issue of seed production is separately tackled as a very important part of growing peppers, which has not been easily available to readership so far, especially to pepper producers. This monograph is a signifi cant publication from the fi elds of pepper growing, breeding and seed production. It is written in clear style, easily readable and accessible to a wide range of readers, including experts, producers, and post-graduate students.

Rat
arst
vo i
pov
rtar
stvo
/ F
ield
an
d V
eget
able
Cro
ps
Res
earc
h 2
011:
48
(1)
Institut za ratarstvo i povrtarstvo / Institute of Field and Vegetable Crops, Novi Sad
UDK: 631/635(051) ISSN 1821-3944
INSTITUT ZA RATARSTVO I POVRTARSTVO
Maksima Gorkog 30,21000 Novi Sad,
Tel.: 021/4898-100, Fax: 021/6621-212www.nsseme.com Ratar. Povrt. / Field Veg. Crop Res. 2011: 48 (1)




















