
Molecular Epidemiology of Human Cancer Viruses
94
MOLECULAR EPIDEMIOLOGY OF HUMAN CANCER VIRUSES BY IBEMGBO, SYLVESTER AGHA B.Sc. Hons. (Ife) MAY,2013 1
Transcript of Molecular Epidemiology of Human Cancer Viruses

MOLECULAR EPIDEMIOLOGY OF HUMAN
CANCER VIRUSESBY
IBEMGBO, SYLVESTER AGHAB.Sc. Hons. (Ife)
MAY,2013
1

ABSTRACT
Both DNA and RNA viruses have been shown to be capable of causing
cancer in humans. This review deals with the molecular
epidemiology of viruses that are strongly associated with human
cancers, and these viruses include; Epstein-Barr virus(EBV),
human papilloma virus(HPV), hepatitis B virus(HBV), human herpes
virus-8(HHV-8), Human T lymphotrophic virus type 1(HTLV-1) and
hepatitis C viruses(HCV).
2

1.0 INTRODUCTION
Cancer results from alterations in critical regulatory genes that
control cell proliferation, differentiation, and survival.
Studies of tumor viruses revealed that specific genes (called
oncogenes) are capable of inducing cell transformation, thereby
providing the first insights into the molecular basis of cancer
(Rao, 2012).Cancer is a major public health challenge all over
the world and lots of current medical research works are aimed at
finding a solution to this problem. With 10.9 million new cases
and 6.7 million deaths per year, cancer is a distressing disease,
presenting amassive disease burden to those affected and their
families as well as health care systems (Parkin et al.,2005). It
3

is estimated that 15-20% of all human cancers are etiologically
linked to viruses (Hausen,2001 and Parkin,2002), these viruses
are called oncoviruses. As obligate intracellular parasites,
oncoviruses encode proteins that reprogram host cellular
signaling pathways that control proliferation, differentiation,
cell death, genomic integrity and recognition by the immune
system (Margaret and Karl,2008).
Both DNA and RNA viruses have been shown to be capable of causing
cancer in humans. Epstein-Barr virus, human papilloma virus,
hepatitis B virus, and human herpes virus-8 are four DNA viruses
that are strongly associated with the development of human
cancers. Human T lymphotrophic virus type 1 and hepatitis C
viruses are two RNA viruses that contribute to human cancers
(Liao, 2006)
Viral carcinogenesis provided the breakthroughs that crystallized
current concepts of cancer development and revealed mechanisms
responsible for the orchestration of normal cell growth control.
Both the RNA tumor viruses and the DNA tumor viruses played
pivotal roles in the establishment of paradigms that extend far
4

beyond virology to form the foundation of contemporary cancer
biology (Butel, 2000).
2.0 OVERVIEW OFHUMAN CANCER VIRUSES
Human tumor viruses belong to a number of virus families,
including the RNA virus families Retroviridae and Flaviviridae
and the DNA virus families Hepadnaviridae, Herpesviridae, and
Papillomaviridae. Presently, the viruses that are strongly
associated with human malignancies include; HTLV-1, etiologically
linked to adult T-cell leukemia/lymphoma (ATLL) (Poiesz et al.,
1980; Boshart et al., 1984); HPV, etiologically linked to cervical
cancer, skin cancer in patients with epidermodysplasia
verruciformis (EV), head and neck cancers, and other anogenital
cancers(Durst et at., 1983,); HHV-8 etiologically linked to
Kaposi's sarcoma (KS), primary effusion lymphoma, and Castleman's
disease (Chang et al., 1994); EBV, etiologically linked to
Burkitt's Lymphoma (BL), nasopharyngeal carcinoma (NPC), post-
transplant lymphomas, and Hodgkin's disease (Epstein et al.,
1964); and, HBV andHCV which are both linked to hepatocellular
carcinoma (HCC) (Dane et al., 1970 and Choo et al., 1979).
5

Other viruses which may play a role in human cancers
include;simian vacuolating virus 40 (SV40) (brain cancer, bone
cancer, and mesothelioma) (Sweet and Hilleman, 1960); BK virus
(BKV) (prostate cancer) (Gardner et al., 1971); JC virus (JCV)
(brain cancer) (Padgett et al., 1971); human endogenous
retroviruses (HERVs) (germ cell tumors, breast cancer, ovarian
cancer, and melanoma) (Bannert and Kurth., 2004; Gifford and
Tristem., 2003); human mammary tumor virus (HMTV) (breast cancer)
(Holland and Pogo, 2004); and Torque teno virus (TTV)
(gastrointestinal cancer, lung cancer, breast cancer, and
myeloma) (Nishizawa et al., 1997). Studies of the RNA and DNA
tumor viruses have led to the discovery of oncogenes and tumor
suppressors and have greatly added to our understanding of the
etiology of carcinogenesis, both virally and non-virally induced
(McLaughlin-Drubin and Munger, 2007)
Table 1 shows the route of transmission and the kind of human
malignancy caused by some human cancer viruses.
Table 1
Virus Family Cells Human Transmission
6
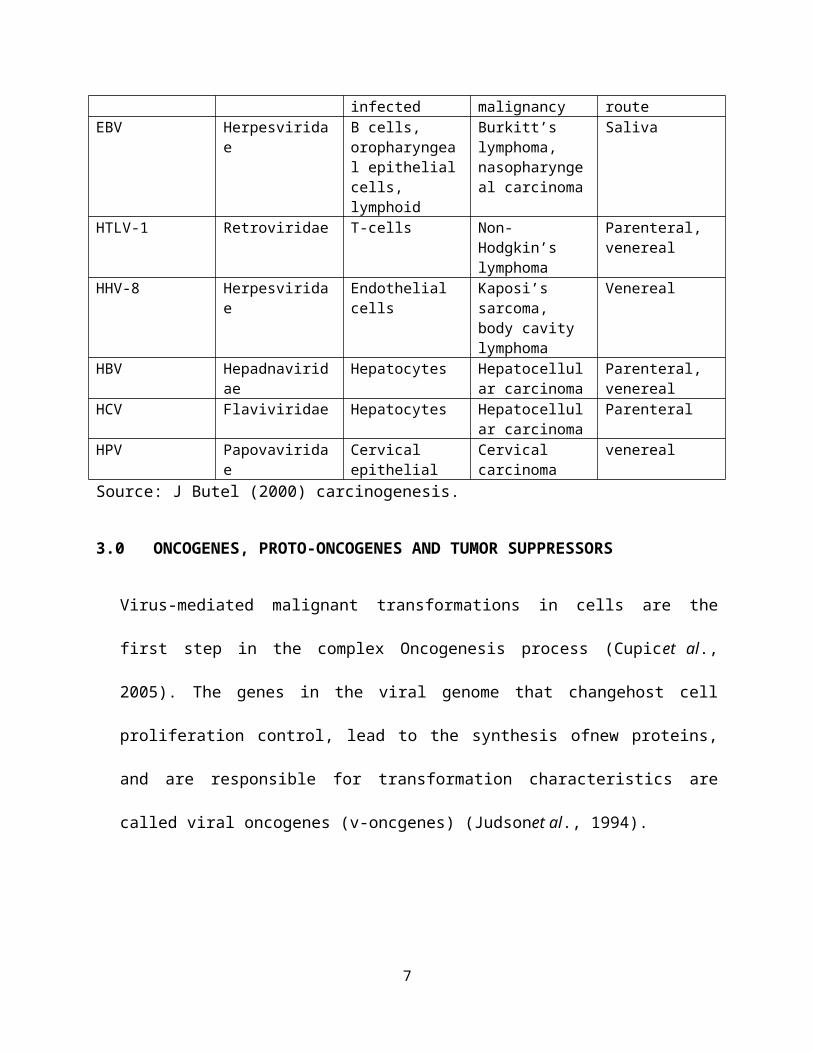
infected malignancy routeEBV Herpesvirida
eB cells, oropharyngeal epithelialcells, lymphoid
Burkitt’s lymphoma, nasopharyngeal carcinoma
Saliva
HTLV-1 Retroviridae T-cells Non-Hodgkin’s lymphoma
Parenteral, venereal
HHV-8 Herpesviridae
Endothelial cells
Kaposi’s sarcoma, body cavity lymphoma
Venereal
HBV Hepadnaviridae
Hepatocytes Hepatocellular carcinoma
Parenteral, venereal
HCV Flaviviridae Hepatocytes Hepatocellular carcinoma
Parenteral
HPV Papovaviridae
Cervical epithelial
Cervical carcinoma
venereal
Source: J Butel (2000) carcinogenesis.
3.0 ONCOGENES, PROTO-ONCOGENES AND TUMOR SUPPRESSORS
Virus-mediated malignant transformations in cells are the
first step in the complex Oncogenesis process (Cupicet al.,
2005). The genes in the viral genome that changehost cell
proliferation control, lead to the synthesis ofnew proteins,
and are responsible for transformation characteristics are
called viral oncogenes (v-oncgenes) (Judsonet al., 1994).
7

Protooncogenes (c-onc genes) are the cellular counterparts of
v-onc genes. Their functions arecellular growth and
development. The activationof c-onc genes with mutation leads
to uncontrolledcell growth (Vats and Emami, 1993). C-onc genes
are transformed intooncogenic form by amplification, point
mutation,deletion, or chromosomal translocation (Bell, 1988).
C-onc genes can beclassified into different groups in termsof
their protein products, such as protein kinases,growth
factors, growth factor receptors, and DNAbinding proteins.
Tumor suppressor genes are genes that prevent malignant
transformation. They are called anti-oncogenes(tumor
suppressor genes). When these genes losetheir suppressive
effects, unpreventable growthoccurs (Vats and Emami,
1993).Their normal functions are to prevent and regulate cell
growth. Among tumor suppressorgenes, the retinoblastoma gene
(Rb) and p53 are themost studied (Stass andMixson, 1997)
4.0 MECHANISM OF ONCOGENESIS IN VIRUSES
Oncogenesis is a cytological, genetic, and cellular
transformation process that results in malignant tumors.The
8

activation of oncogenes requires genetic changes in cellular
proto-oncogenes. Oncogenes are activated by 3 genetic mechanisms:
Mutation, Gene amplification, and Chromosome rearrangements.These
mechanisms result in either an increase inprotooncogene
expression or a change in protooncogene structure. Neoplasia is a
multistep process; therefore, more than one of these mechanisms
contribute to the formation of tumors. Expression of the
neoplastic phenotype includes the capacity for metastasis and
usually requires a combination of protooncogene activation and
tumor suppressor gene loss or inactivation (Pierottiet al., 2010).
4.1 MECHANISM OF ONCOGENESIS IN DNA TUMOR VIRUSES
DNA tumor viruses have 2 life forms. In permissive cells, viral
replication causes cell lysis and cell death. In non-permissive
cells, viral DNA is mostly integrated into the different sites of
cell chromosomes. It encodes binding proteins and inactivates
cell growth, regulating proteins like p53 and retinoblastoma. The
9

cell is transformed as a result of the expression of proteins
that control viral and cellular DNA synthesis (Cupicet al., 2005).
4.2 MECHANISM OF ONCOGENESIS INRNA TUMOR VIRUSES
All oncogenic RNA viruses are retroviruses (Klein,
2002).Retroviruses have 3 basic genes (gag, pol, andenv), which
are used for the synthesis of structuralproteins, virion-
associated enzymes, and envelope glycoproteins (Cullen, 1992).
Although retroviruses have been associated with many animal
tumors, to date, only one human retrovirus, HTLV-1, has been
associated with human cancers. Complex retroviruses such as
lentiviruses have an extra non-structural gene (v-onc) that
allows them to transform the cell (Weiss, 1996).
10
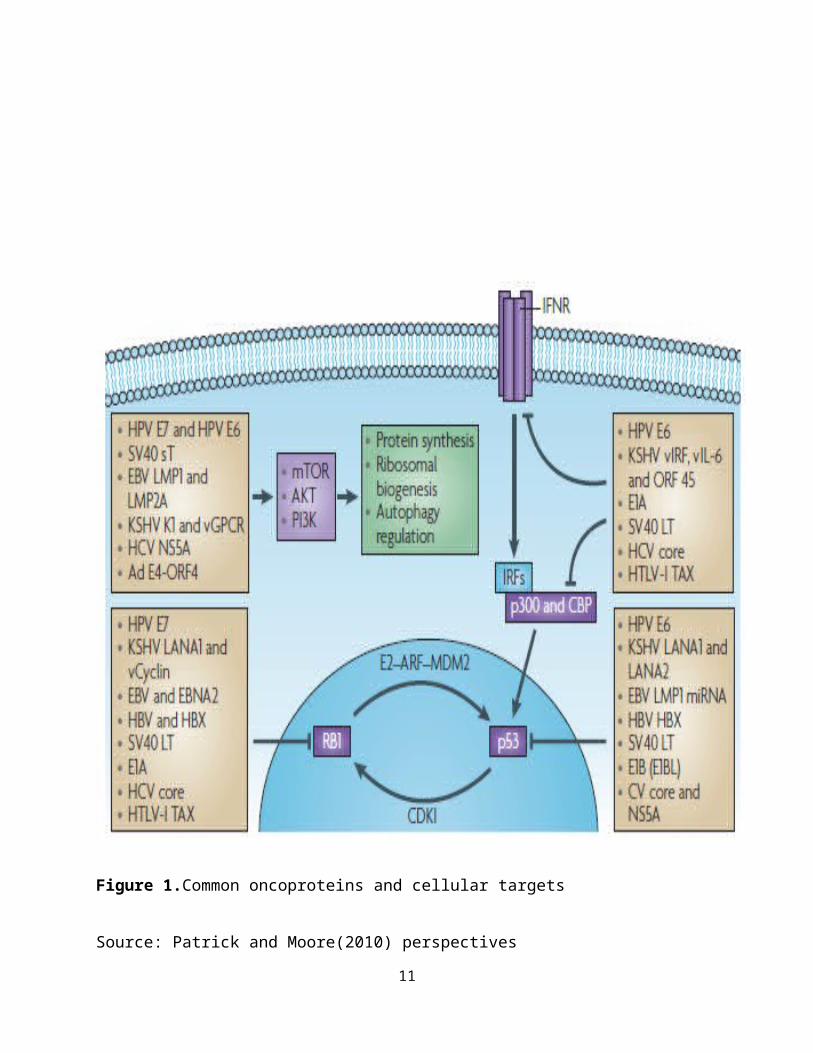
Figure 1.Common oncoproteins and cellular targets
Source: Patrick and Moore(2010) perspectives
11
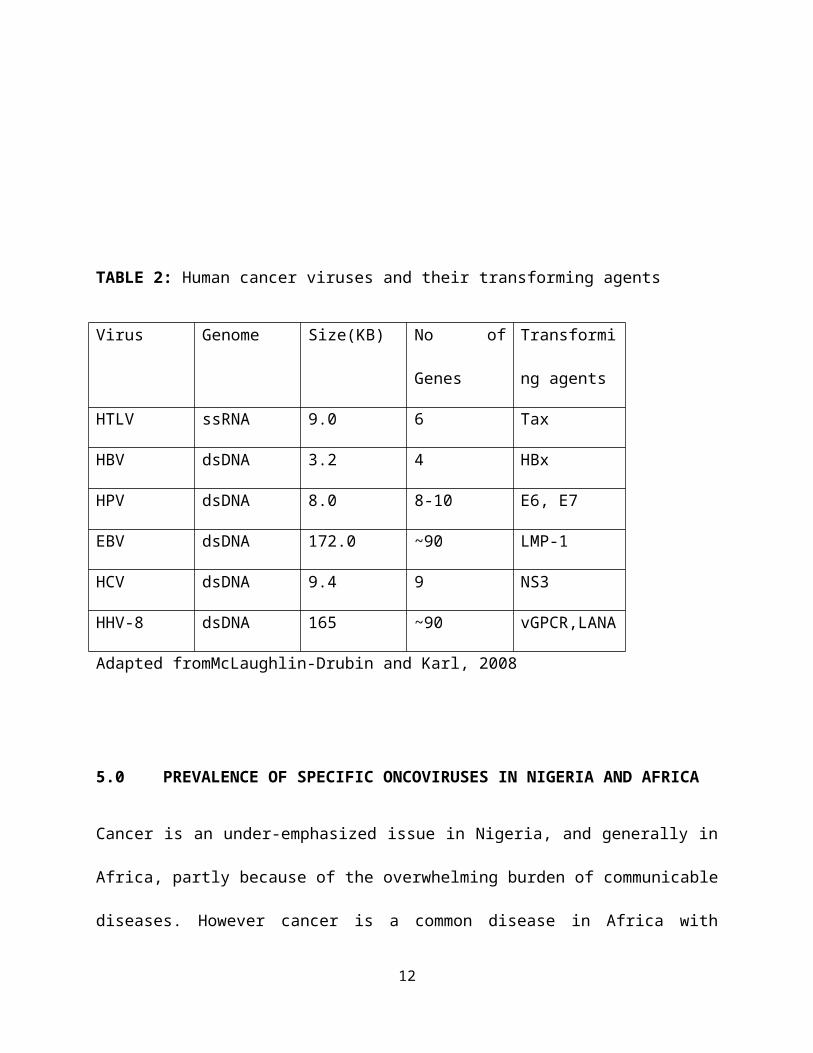
TABLE 2: Human cancer viruses and their transforming agents
Virus Genome Size(KB) No of
Genes
Transformi
ng agents
HTLV ssRNA 9.0 6 Tax
HBV dsDNA 3.2 4 HBx
HPV dsDNA 8.0 8-10 E6, E7
EBV dsDNA 172.0 ~90 LMP-1
HCV dsDNA 9.4 9 NS3
HHV-8 dsDNA 165 ~90 vGPCR,LANA
Adapted fromMcLaughlin-Drubin and Karl, 2008
5.0 PREVALENCE OF SPECIFIC ONCOVIRUSES IN NIGERIA AND AFRICA
Cancer is an under-emphasized issue in Nigeria, and generally in
Africa, partly because of the overwhelming burden of communicable
diseases. However cancer is a common disease in Africa with
12

650,000 people, of a population of 965 million, diagnosed
annually (Parkin et al., 2008). In females, the lifetime risk of
dying from cancer in Africa is almost double the risk in
developed countries. The cancers with the highest incidence are
cervical, breast, and now HIV-associated Kaposi's sarcoma. Some
of the most common cancers in males; Kaposi's sarcoma
(constituting 12.9% of all cancers in males) and cancer of the
liver (14.8%), and in females;cancer of the cervix (constituting
23.3% of all cancers in females) and Kaposi's sarcoma (5.1%),
cancer of the liver (5.0%) (Parkin et al.,2008), are strongly
associated with viruses. Some of the important oncoviruses and
their activities in Nigeria and Africa are discussed below.
5.1 HUMAN PAPILLOMAVIRUS IN NIGERIA AND AFRICA
Papillomaviruses (PVs) are double-stranded DNA viruses that
constitute the Papillomaviridae family.Approximately 200 human
papillomavirus (HPV) types have been described (de Villiers et
al., 2004). HPVs cause a range of epithelial hyperplastic lesions
and can be classified into two groups: mucosal and cutaneous.
13

These groups can be further divided into low- and high-risk,
depending on the associated lesion's propensity for malignant
progression. The concept of low-risk and high-risk HPVs has been
most clearly established with the mucosal HPVs (McLaughlin-Drubin
and Munger, 2008).
HPV is also associated with oral and other anogenital
malignancies, however, it is most commonly associated with
cervical cancer; over 99% of all cervical cancers are associated
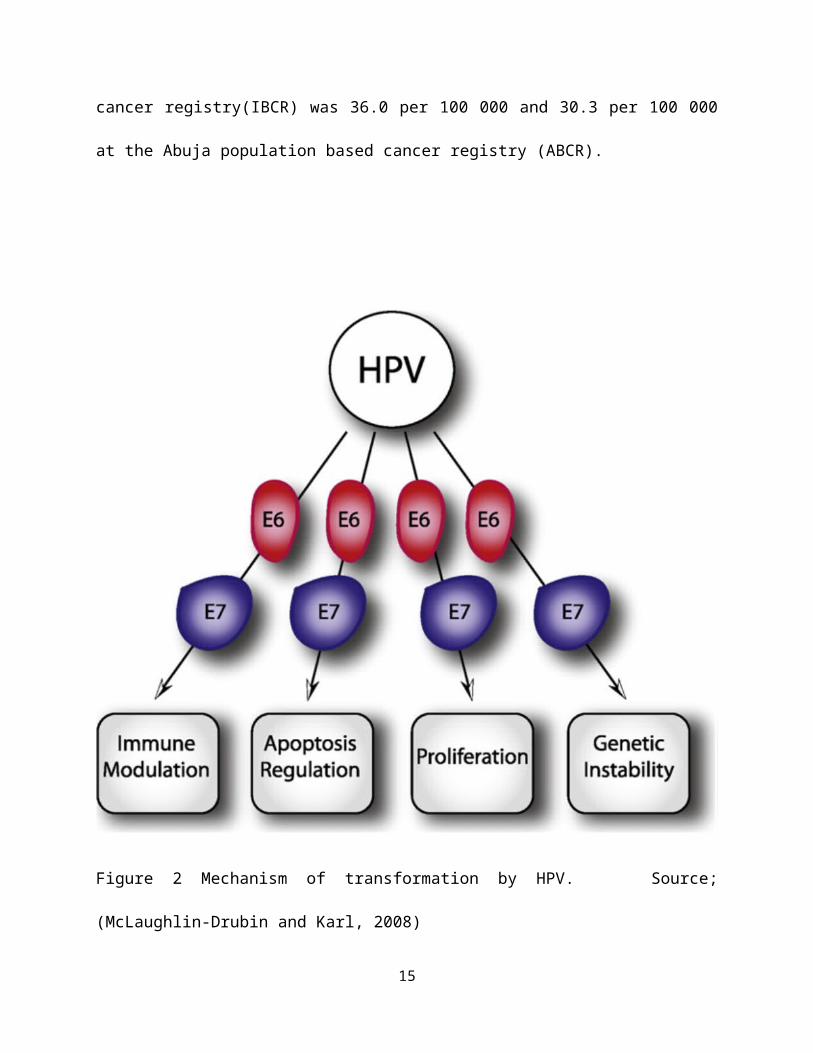
with high-risk HPV infections (Lowy and Howley, 2001). Figure 2
shows the mechanism of transformation by HPV
The incidence of cervical cancer in sub-Saharan Africa is among
the highest worldwide, with age-standardized rates of 35.7 per
100,000 in Bamako, Mali, and 41.7 per 100,000 in Kyadondo, Uganda
(Parkin et al, 2002). In a population based study, Thomas et al.,
(2004) stated that the prevalence of HPV of any type was 26.3,
24.8 and 40.9%, respectively, among women with normal and
abnormal cervical findings in Ibadan, Nigeria. According to
Elimaet al., (2012), the age standardized incidence rate (ASR) of
cervical cancer in Nigeria from the Ibadan population based
14
cancer registry(IBCR) was 36.0 per 100 000 and 30.3 per 100 000
at the Abuja population based cancer registry (ABCR).
Figure 2 Mechanism of transformation by HPV. Source;
(McLaughlin-Drubin and Karl, 2008)
15

5.2 EBV AND BURKITT’S LYMPHOMA IN NIGERIA
EBV is a ubiquitous double-stranded DNA virus of the γ-
herpesviruses subfamily of the Lymphocryptovirus (LCV) genus
(McLaughlin-Drubin and Munger, 2008). The EBV genome is a linear,
double-stranded, 184 kbp DNA composed of 60 mole percent guanine
and cytosine (Baer et al., 1984; Kieff and Levine, 1974 ;
Pritchett et al., 1975). As with other herpesviruses, EBV has a
toroid shaped DNA core in a nucleocapsid with 162 capsomeres, an
outer envelope with external glycoprotein spikes, and a protein
tegument between the nucleocapsid and envelope (Dolyniuk et al.,
1976; Dolyniuk et al.,1976; Epstein et al., 1965; Johannsen et
al., 2004)
Burkitt's lymphoma (BL) is an unusual type of non-Hodgkin's
lymphoma occurring endemically in the narrow zone across central
Africa, where malaria is endemic. (Ferry, 2006; Cheneet al.,
2009).). In a study that analyzed the profile of cancers recorded
in the first decade (1995-2004) of establishment of the Kano
16

cancer registry (KCR), a histology/cytology-based registry in
Kano, Nigeria, BL (31.4%), other lymphoreticular cancers (23.8%)
and retinoblastoma (20%) predominated in children of a total of
1990 cancer cases. (Mohammed et al., 2008).Another study published
in 2009, comprising 5-year retrospective review of pediatric
solid tumors as seen at the Jos University Teaching Hospital,
Nigeria, non-hodgkin's lymphoma and BL accounted for 17 (19.5%)
and 12 (13.8%), respectively, of a total of 181 solid tumors of
children. (Tankoet al., 2009)
5.3 KAPOSI'S SARCOMA-ASSOCIATED HERPESVIRUS (KSHV)/HUMAN HERPES
VIRUS 8 (HHV-8) IN NIGERIA
HHV-8, also known as KSHV is a double-stranded DNA virus,
initially discovered on the basis of its association with
Kaposi's sarcoma, an endothelial neoplasm, it is now known to be
a member of the lymphotropic γ-herpesvirus subfamily (Knipe and
Howley, 2006). The KSHV genome is a double-stranded linear DNA of
165 to 170 kb (Renneetal., 1996). The genome contains a central
17

unique region of 145 kb, which has all of the viral open reading
frames (ORF) (Russo et al.,1996), flanked by a series of highly
GC-rich direct terminal repeats (TR). Each TR unit is 801 base
pair (bp) long, with a base composition of 85% G+C, and is devoid
of substantial ORFs. In general, isolates have 20 to 25 kb of
total TR DNA per genome, but the number of repeats present at
each terminus can vary substantially (Lagunoff and Ganem, 1997).
The neoplastic potential of KSHV, especially in immunocompromised
individuals, is well-established: epidemiological studies link
KSHV to human malignancies (Parkin, 2006) KSHV transforms
endothelial cells (Flore et al., 1998)
A South African study showed rates of KSHV infection to vary from
35% to 49% across different municipalities within one province
(Maskew et al., 2011)
18

5.4 HUMAN T-CELL LEUKEMIA VIRUS TYPE 1 (HTLV-1)
HTLV-1 is a deltaretrovirus first isolated from the blood of an
African-American patient with cutaneous T-cell lymphoma in the
late 1970s (Poiesz et al., 1980); Human T-cell leukemia virus type
1(HTLV-1) is a slow transforming, single stranded RNA retrovirus
and is associated with adult T-cell leukemia (Gallo et al.,1983).
It possesses a diploid genome. The genome contains gag, pro, pol,
and env genes; the gag, pro, and pol genes are present in three
different reading frames, and expression of the Gag-Pro-Pol
protein requires two successive frameshifts (Lairmoreand
Franchini, 2007).
While the exact cellular events remain unclear, a variety of
steps, including virus, host cell, and immune factors, are
implicated in the leukemogenesis of ATLL (Matsuoka, 2003). A
number of studies indicate that the multifunctional viral
accessory protein Tax is the major transforming protein of HTLV-1
(Nerenberg et al., 1987; Grassmann et al., 1992; Pozzatti et al.,
19

1990; Tanaka et al., 1990; Smith and Greene, 1991). Figure 3 shows
the mechanism of transformation by HTLV.
It has been shown that both HTLV-I and HTLV-II are endemic in
Nigeria (Olaleyeet al., 1998).Several studies haveshown that the
prevalence of HTLV-I is high in Africa. Seropositivity for HTLV-I
in the region range from 0.6% in Morocco, to 3.5% in South Africa
and 16.9% in Tanzania (Olaleye et al., 1998)
Figure shows the mechanism of cell transformation y HTLV-1
20
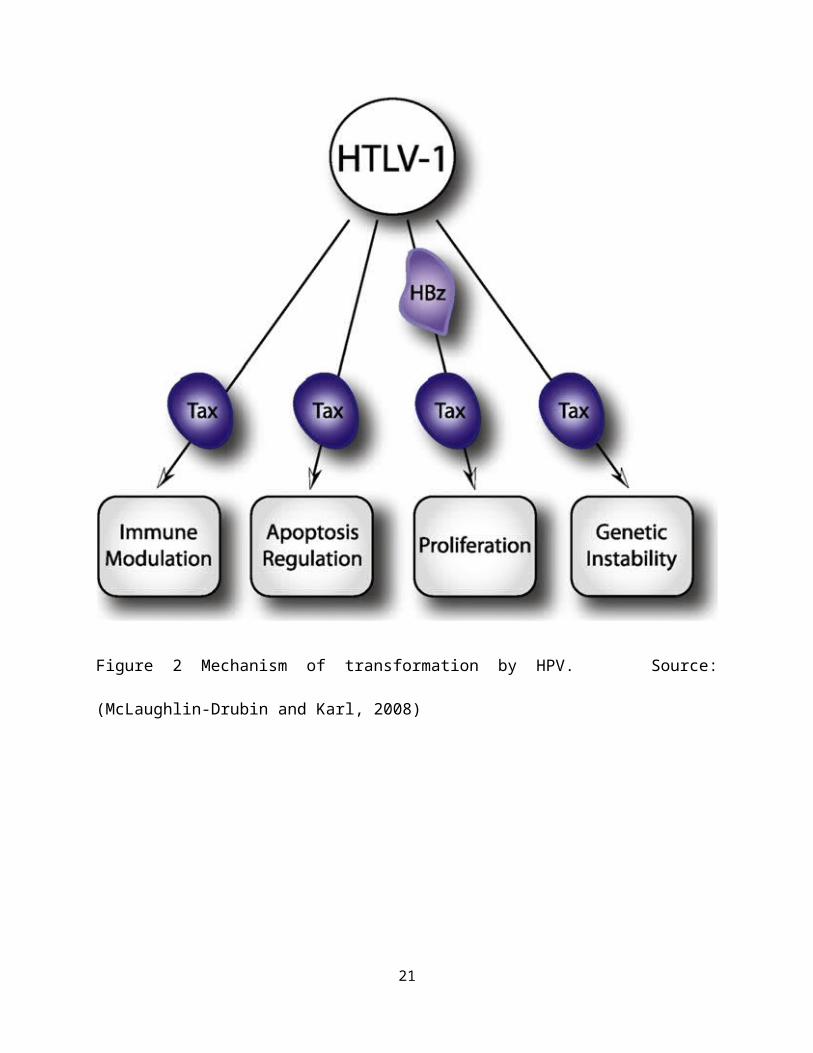
Figure 2 Mechanism of transformation by HPV. Source:
(McLaughlin-Drubin and Karl, 2008)
21

5.5 HEPATITIS B VIRUS (HBV) IN NIGERIA
HBV is a member of the Hepadnaviridae family. It is the only DNA
virus among the agents which commonly cause viral hepatitis. The
viral particle (called the Dane particle) is 42 nm in diameter
(Haaheimet al., 2002). HBV has a circular, partially double-
stranded, DNA genome with four overlapping open reading frames
(ORFs) that encode for the envelope (preS/S), core (preC/C),
polymerase, and X proteins (Seeger and Mason, 2000). Like
retroviruses, the replication of HBV is dependent on reverse
transcription; unlike retroviruses, integration of the viral
genome into the host chromosome is not necessary for viral
replication but does allow for persistence of the viral genome
(Feitelson and Lee, 2007).This virus is responsible for 80% of
all cases of primary liver cancer, which is one of leading causes
of death in Asia and Africa (Clement et al., 1990).
Hepatitis B Virus, a major public health problem worldwide is
more prevalent in the developing countries (Johnson et al.,
1986).In Nigeria, 11.6% prevalence rate has been reported from
Maiduguri (Harry et al., 1994), 13.8% from Lagos (Nasidiet al.,
22

1993), 4.3% from Port Harcourt (Akaniet al., 2005), 5.7% from
Ilorin (Agbedeet al., 2007), 8.3% from Zaria (Luka et al., 2008),
17.1% from female sex workers, 14.9% (Forbi et al., 2008) from
healthy blood donors (Ejeleet al., 2004) and 25.7% among surgeons
(Belo, 2000). Although HBV prevalence varies widely across the
African continent, hepatitis B surface antigen (HBsAg) positivity
is estimated at 8-20% (Apurva and Jordan, 2007).
5.6 HEPATITIS C VIRUS (HCV) IN NIGERIA
HCV is a single-stranded RNA virus of the Hepacivirus genus in
the Flaviviridae family and is the only positive-stranded RNA
virus among the human oncogenic viruses. Its approximately 9.6 kb
genome contains an open reading frame (ORF) that codes for a 3000
amino acid residue polyprotein precursor (Choo et al., 1989) that
is cleaved by cellular and viral proteases into three structural
proteins (core, E1, E2) and seven nonstructural proteins (p7,
23

NS2, NS3, NS4a, NS4B, NS5A, and NS5B) (Lindenbach and Rice,
2005).
HCV proteins have been reported to activate cellular oncoproteins
and inactivate tumor suppressors, such as p53, CREB2/LZIP, and
the retinoblastoma protein (pRB). HCV causes genome instability,
signifying that certain HCV proteins may have a mutator function
(McLaughlin-Drubin and Munger, 2008).
In 2005, Inyamaet al., (2005) put the prevalence of HCV among
Nigerian patients with HIV in Jos infection at 5.7%.So far the
prevalence of HCV infection is increasing in Nigeria, ranging
from 4.7-5% in Ilorin, to 5.3-6.6% in Enugu, to 11% in Ibadan and
20% in Benin (Ejioforet al., 2010)
24

2.1.1 MOLECULAR EPIDEMIOLOGY OF HTLV-1
HTLV-1 has a worldwide distribution, with an estimated 12 to 25
million people infected. However, disease is only observed in
less than 5 percent of infected individuals. It is endemic to
Japan, South America, Africa, and the Caribbean (Gessian et al.,
1991; Wong-Staal and Gallo, 1985). The seroprevalence of HTLV-1
varies between 0.1% and 30% within endemic populations (Gessain
and Mahieux, 2000). Sporadic HTLV-1 infection occurs among at-
risk groups from non-endemic regions, including metropolitan
areas in the United States and western European countries
(Matsuoka and Jeang, 2005).The incidence rate is 2 to 4 per
100,000 person-years and males have higher risk than females
(Cleghornet al.,1995; Kondo et al., 1989).
2.1.2 TRANSMISSION AND PATHOGENESIS OF HTLV-1
HTLV-1 is transmitted through blood transfusions, sexual
contact, and during parturition. Typically, 1% to 5% of
individuals infected with HTLV-1 develop ATLL after a latent
period of 20 to 30 years (Cleghornet al.,1995; Yamaguchi and
Takatsuki, 1993).
25

ATLL is an aggressive malignancy of T lymphocytes,
immunophenotypically characterized by multiple distinct cell
surface markers, including CD3+/CD4+/CD8-/CD25+/HLA-DR+ T-cells
(Uchiyama et al., 1977). The typical clinical symptoms and
presentations of patients with ATLL include malaise, fever,
lymphoadenopathy, hepatosplenomegaly, jaundice, weight loss, and
various opportunistic infections, such as Pneumocystis carinii (White
et al., 1995). Based on the clinical course of the disease, four
classifications are applied for ATLL, including (a) asymptomatic,
(b) preleukemic, (c) chronic smoldering, and (d) acute stages
(Kawano et al., 1985,Shimoyama, 1991, Yamaguchi et al., 1983). The
latter two forms represent the typical clinical presentation
mentioned above. Clinicopathologically, the chronic stage ATLL is
characterized by leukocytosis, whereas the acute stage is
characterized by lymphocytes with atypical cell morphology,
including multilobulated nucleus, lymphoadenopathy, and
hepatosplenomegaly (Yamaguchi and Takatsuki, 1993, Yamaguchi,
1994). Other clinicopathologic abnormalities seen in patients
with ATLL include hypercalcemia of malignancy, in association
with lytic bone lesions, elevated lactate dehydrogenase (LDH),
26

and the presence of soluble form of interleukin-2 (IL-2) receptor
in serum samples. Infiltration of the skin by neoplastic
lymphocytes frequently occurs in patients with ATLL. Diagnostic
criteria of ATLL include HTLV-1 seropositivity, leukocytosis,
increased serum levels of IL-2 receptor and LDH, demonstration of
neoplastic T cells with polylobulated nuclear morphology (flower
cells), and the presence of clonally integrated HTLV-1 genomes
within the chromosomes of neoplastic lymphocytes (Shimoyama,
1991).The long clinical latency, together with the relatively low
cumulative lifetime risk of a carrier developing Adult T-cell
Leukemia (ATL), indicates that HTLV-1 infection alone is not
sufficient to elicit T-cell transformation.
2.1.3 T-CELL IMMORTALIZATION AND TRANSFORMATION BY HTLV-1
The terms immortalization and transformation of T cells by HTLV-1
refer to ligand (IL-2)-dependent or -independent indefinite
growth of infected T cells, respectively. Biochemically, this
distinction has been associated in most, but not all, HTLV-
1infected T cells with constitutive activation of the Jak/STAT
27

signaling pathways (Migoneet al.,1995; Mulloyet al., 1998; Xuet al.,
1997), as well as with a reduction of the level of Src homology 2
(SH2)-containing tyrosine phosphatase-1 (SHP-1) (which could
result in decreased phosphorylation of STAT) (Migoneet al.,1998).
However, an inhibitor of Jak3, AG-490, does not affect the growth
of the HTLV-1infected T-cell line MT-2 (Kirkenet al., 2000),
suggesting that other pathways may be involved in the
proliferation of these cells.
Immortalization of primary human T cells in the presence of IL-2
can be induced by infection with various HTLV-1 isolates or by
Tax-1 expression alone (Grassmann et al., 1989; Iwanagaet al., 1999;
Tanaka et al., 1991). In some cases, however, continuous
stimulation of the TCR and Tax-1 expression is required (Tanaka
et al., 1991). The oncogenic properties of Tax-1 may be related to
its effect on cell cycle checkpoint inhibitors, cyclins,
telomerase activity, and transcriptional modulation of a wide
range of cellular genes involved in cellular growth and survival
(Kelly et al., 1992). Exploiting the NF-kB pathway, Tax activates a
large number of cellular genes that promote lymphocyte growth,
activation, and survival. These genes fall into several28

categories, including cytokines and growth factors such as IL-1,
IL-2, IL-6, IL-8, IL-15, and TGFb, growth factor receptors such
as IL-2Ra and OX40, and antiapoptotic factors such as BCL-xl
(Lemoine et al., 2001). Special importance is attributed to Tax
activation of the cellular proto-oncogene, c-MYC, which can
independently mediate cellular transformation via NF-kB
regulation (Keath et al., 1984; Adams et al., 1985).
Schematic depiction of the major biological activities that
contribute to the transforming activities of HTLV-1 (McLaughlin-
Drubin and Karl, 2008)
The ability of Tax to disable an important protective mechanism
that protects against chromosomal missegregation during mitosis
provides a potential mechanism for the high incidence of
aneuploidy in ATL cells (McLaughlin-Drubin and Karl, 2008)
2.1.4 TREATMENT, PREVENTION AND CONTROL OF HTLV-1
29

Depending on the stage of disease progression, ATLL is highly
refractory to most forms of conventional therapy and, thus, has a
poor prognosis, especially for the acute form of the malignancy
(Taylor and Matsuoka, 2005). The median time of survival of
patients with ATLL is typically less than 1 year despite
aggressive chemotherapy. Complicating the treatment of ATLL are
paraneoplastic syndromes (e.g., hypercalcemia), which contribute
significantly to overall mortality of these patients (Blayneyet
al., 1983; Jaffe et al., 1984; Prageret al., 1997). Standard
cytotoxic or cytostatic chemotherapeutic agents (e.g.,
cyclophosphamide, adriamycin, vincristine, and prednisolone so
called CHOP therapy) remain an initial approach to treating
patients with ATLL despite variable success in median survival
times in most cohorts (Mufti et al., 1998; Ohshima et al.,1999;
Pawsonet al., 1998; Yamaguchi et al., 1983). Improved outcomes have
been reported for standard chemotherapy supplemented with the
cytokine, G-CSF (Bohliuset al., 2004). The relative insensitivity
of ATLL to chemotherapy has been speculated to be caused by the
enhanced expression of the multidrug resistance gene (Chuang
etal., 1997; Lau et al., 1998).
30

Targeting NF-kB in ATLL is based on experimental evidence of its
importance in Tax-induced transformation. Bortezomib (PS-341) has
been used in an effort to block proteasome processing and thereby
interrupt NF-kB signaling in ATLL cells (Sato et al., 1991).
Sodium valproate, an inhibitor of deacetylases, has been reported
to reduce lymphoma growth in a sheep model of BLV, a member of
the deltaretroviruses, offering a new direction for ATLL therapy
(Achachiet al., 2005).
Monoclonal antibodies directed at cell surface markers have been
employed to target and kill ATLL cells. This therapy has the
potential to allow more selective targeting of molecules on the
surface of ATLL cells, such as the IL-2 receptor (CD25) (Green et
al.,1995;Waldmannet al., 1988).
The prevention of transmission of HTLV-1 and HTLV-2 has been
accomplished based on elimination of breastfeeding from HTLV-
1infected mothers in endemic regions (Hino et al., 1994), but
this intervention is problematic on a worldwide scale.
The viral envelope glycoproteins appear to be good candidates for
a subunit vaccine. The surface glycoprotein (SU) component of
31

envelope mediates viral infection of cells by binding to the
cellular entry factors heparan sulfate proteoglycans (Pinonet
al.,2003; Jones et al., 2005). Subsequently, through a cascade of
conformational changes in envelope that includes isomerization of
a SU-transmembrane glycoprotein (TM) intersubunit disulfide
(Wallinet al., 2004; Li et al., 2008),envelope catalyzes the fusion
of the viral and cell membranes allowing entry of the virus into
the cell. Neutralizing Abs and synthetic inhibitory peptides
block envelope-mediated entry (Sagaraet al., 1996; Piñónet al.,
2003; Brighty and Jassal, 2001; Lamb et al.,2009), suggesting that
interfering with envelope function can prevent dissemination of
viral infection
3.0 HUMAN PAPILLOMAVIRUS (HPV)
The papillomaviruses (PVs) comprise a group of nonenveloped,
epitheliotropic DNA viruses that induce benign lesions of the
skin (warts) and mucous membranes (condylomas) (Knipe and Howley,
2006). Papillomaviruses (PVs) are adouble-stranded DNA viruses
that constitute the Papillomaviridae family.Approximately 200
human papillomavirus (HPV) types have been described (de Villiers
32

et al., 2004). HPVs cause a range of epithelial hyperplastic
lesions and can be classified into two groups: mucosal and
cutaneous. These groups can be further divided into low- and
high-risk, depending on the associated lesion's propensity for
malignant progression. The concept of low-risk and high-risk HPVs
has been most clearly established with the mucosal HPVs
(McLaughlin-Drubin and Munger, 2008).
3.1.0 MOLECULAR EPIDEMIOLOGY OF HPV
Both epidemiological and molecular evidence strongly supports the
link between infection with high-risk HPVs and the development of
cervical cancer. Nonetheless, the incidence of malignant
progression of high-risk HPV associated lesions is relatively
low; malignant progression usually occurs with other risk
factors, such as, decreased immune function, and/or a long
latency period after other genomic alterations in the host cell
DNAhave occurred. Smoking and prolonged use of birth control
pills have also been implicated as risk factors for progression.
The low-risk mucosal HPVs, such as HPV6 and 11, cause genital
warts, whereas the high-risk mucosal HPVs, such as HPV16 and 18,
33

cause squamous intraepithelial lesions that can progress to
invasive squamous cell carcinoma. HPV is also associated with
oral and other anogenital malignancies, however, it is most
commonly associated with cervical cancer; over 99% of all
cervical cancers are associated with high-risk HPV infections
(Lowy and Howley, 2001).
3.1.1 PREVALENCE OF HPV
Over a ten-year period (1991-2000), 815 women had Pap smears at
the Medical Women Centre in Enugu of whom only 4 (0.5%) ever had
a previous smear. The prevalence of abnormal smears was 12.2%,
the proportion rising with increasing age and doubling after the
age of 54 years. All the women with positive smears were or had
been coitally active; 97% were parous and 81% grand multiparous
(Chukwuali et al., 2003)
In a demonstration project, ‘prevention of cervical cancer
through screening using visual inspection with acetic acid(VIA)
and treatment with cryotherapy’, WHO reported that 5.7percent of
the Nigerians screened were positive.
34

According to CDC in 2012, Overall, about 26,000 (18,000 among
females and 8,000 among males) of 33,369 cancers at HPV-
associated sites were attributed to HPV infection. For the
individual cancers, the number and percentages attributable to
HPV were:
Cervix, 11,500 (96%)
Vulva, 1,600 (51%)
Vagina, 500 (64%)
Penis, 400 (36%)
Anus(women), 2,900 (93%)
Anus(men), 1,600 (93%)
Oropharynx(women) 1,500 (63%)
Oropharynx (men), 5,900(63%).
Infections with cutaneous HPVs appear extremely frequently in the
general population and some of these viruses, even the presumed
high-risk HPV, may be part of the normal “flora” of the skin as
they can be detected in follicles of plucked hair (Boxmanet al.,
1997; Boxmanet al., 1999).
35

A 2007 study in United States found approximately 26.8% of all
women examined (aged 14-59) were infected with the human
papillomavirus. It is important to note that most HPV infections
are cleared within two years (i.e. most infections are not
lifelong) (Dunne et al., 2007)
• 14-19 ~ 24.5%
• 20-24 ~ 44.8%
• 25-29 ~ 27.4%
• 30-39 ~ 27.5%
• 40-49 ~ 25.2%
• 50-59 ~ 19.6%
The true incidence of cervical cancer in most African countries
is unknown and there is gross under reporting. Only a few
countries have functional cancer registers and record keeping is
minimal or non-existent. Some of the figures quoted in the
literature are hospital-based which represents a small fraction
of women dying of cancer of the cervix as most women cannot
assess hospital care and die at home. (Anorlu, 2008)
36

3.1.2 TRANSMISSION OF HPV
The human papillomavirus is transmitted via skin-skin contact.
Sexual intercourse is not necessary for transmission, but is the
most common route. The virus can infect the genital, anal, and
oral regions of the body. Infection occurs when viruses enter
into small breaks in the skin or mucous membranes. The
probability of acquiring HPV from a single sexual encounter is
not known, but is probably high (Burchellet al., 2006)
Other than the appearance of genital warts, infection with the
human papillomavirus is asymptomatic. In most cases HPV doesn't
cause any problems and is cleared by the immune system (Hausen,
2001)
3.1.3 HPV IMMORTALIZATION AND TRANSFORMATION
The molecular mechanisms by which high-risk HPV causes cervical
cancer have been studied extensively, and numerous viral and host
37

interactions that may contribute to transformation and malignancy
have been described (Munger et al., 2004).HPV5 and 8 as well as
related cutaneous HPVs may contribute to the development of
nonmelanoma skin cancers (NMSC), particularly in
immunocompromised patients (Shamaninet al., 1994; Shamaninet al.,
1996).
It has been shown that E6 proteins of cutaneous HPVs can target
the proapoptotic Bcl-2 family member, Bak, for degradation. Bak
plays an important role in signaling apoptosis in response the UV
irradiation and hence it has been postulated that cutaneous
HPVexpressing cells may be less prone to undergo apoptosis after
UV induced DNA damage (Jackson et al., 2000).
Cellular targets of the papillomavirus E6 oncoproteins
Associated Cellular ProteinsFunctional Consequences
E6-associated protein (E6AP)Ubiquitylation and
proteolysis of associated
proteins
Ubiquitylation of E6
p53 E6AP-dependent ubiquitylation
38

and proteolysis
E6BP (Erc55) Unknown
Paxillin Disruption of the actin
cytoskeleton
HDlg MUPP1 and hScrib E6AP dependent ubiquitination
and proteolysis
IRF-3 Inhibition of β-interferon
induction
Clatherin adaptor complex
AP-1
Unknown
Bak Inhibition of Bak-induced
apoptosis
CBP/p300 Inhibition of p53
transcriptional activity
Myc Activation of cellular
telomerase
NFX1 Activation of cellular
telomerase
E6TP1 E6-induced degradation
hADA3 Transcriptional regulation
39

Source: Fields Virology, 5th Edition Copyright ©2007 Lippincott
Williams & Wilkins
During carcinogenic progression the HPV genome frequently
integrates into a host cell chromosome and, as a result, the
viral oncoproteins, E6 and E7, are the only viral proteins that
are consistently expressed inHPV positive cervical carcinomas.HPV
genome integration is often associated with higher dysregulated
E6/E7 expression (Jeonet al., 1995). Persistent expression of E6
and E7 is necessary for maintenance of the transformed phenotype
of cervical carcinoma cells (Goodwin and DiMaio, 2000; Wells et
al., 2000).
The transforming activities of the high-risk E6 and E7
oncoproteins is related to their ability to associate with and
dysregulate cellular regulatory protein complexes, most notably
p53 and pRB (Munger and Howley, 2002). As p53 and pRB normally
control cellular proliferation, differentiation, and apoptosis,
the abrogation of their normal biological activities places such
a cell at a risk of malignant progression. In addition, high-risk
HPV E6 and E7 expressing cells have a decreased ability to
40

maintain genomic integrity (White et al., 1994). The high-risk HPV
E7 oncoprotein acts as a mitotic mutator and induces multiple
forms of mitotic abnormalities, including anaphase bridges,
unaligned or lagging chromosomes, and most notably multipolar
mitoses (Duensing and Munger, 2002). Multipolar mitoses are
histopathological hallmarks of high-risk HPV associated cervical
lesions and cancer (Crum et al., 1984) and are caused by the
ability of high-risk HPV to uncouple centrosome duplication from
the cell division cycle (Duensing et al., 2000; Duensing et al.,
2001).
41
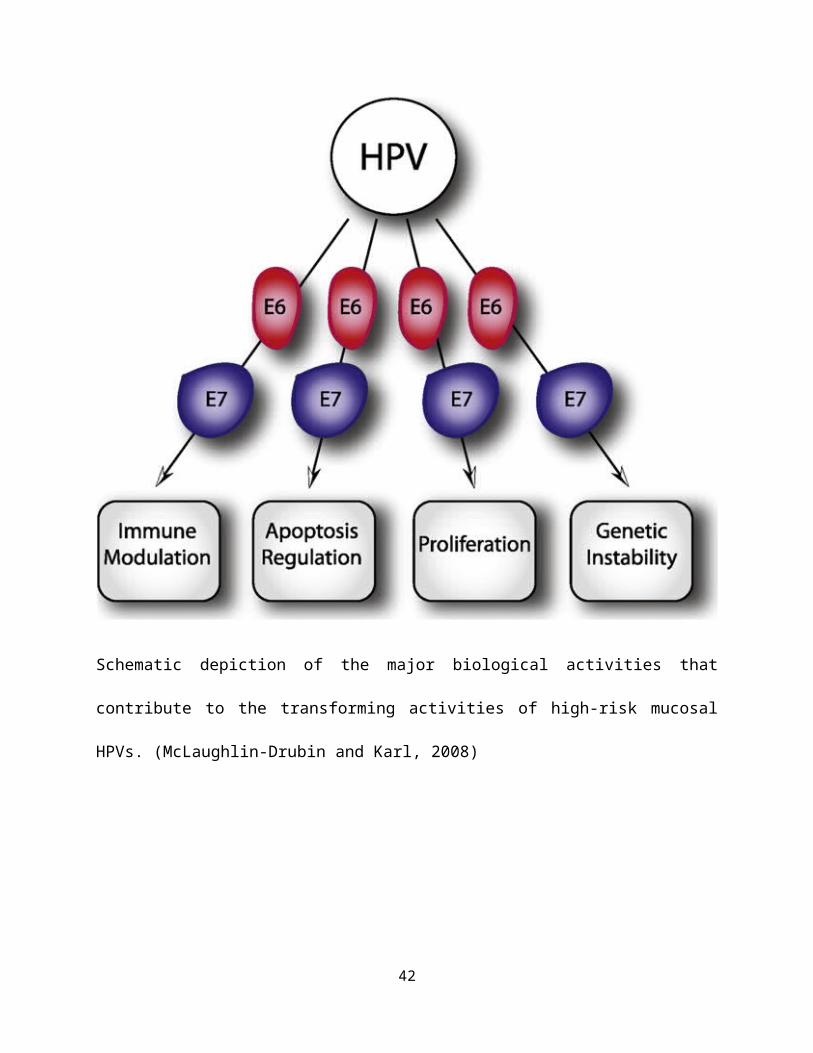
Schematic depiction of the major biological activities that
contribute to the transforming activities of high-risk mucosal
HPVs. (McLaughlin-Drubin and Karl, 2008)
42

3.1.3 TREATMENT, PREVENTION AND CONTROL OF HPV/ CERVICAL
CANCER
High-grade dysplasias represent precancerous lesions that are
unlikely to resolve spontaneously, and their treatment is
recommended to prevent cervical cancer. HPV testing can be done
in case, because most successfully treated cases become negative
for HPV DNA, whereas incompletely treated cases may remain
positive (Zielinski et al., 2004). Cervical cancer is treated by
surgery, radiotherapy, and chemotherapy, with early stage tumors
having a better prognosis than more advanced tumors (McCreathet
al., 2005; Serkies and Jassem, 2005).
Public health programs aimed at controlling other sexually
transmitted infections should also be effective in preventing
genital HPV infection. Control of sexual promiscuity on the part
of both sexual partners should reduce the likelihood of exposure
to genital HPV.
43

Given the high rate of HPV infection that follows initiation of
sexual activity, the VLP vaccines would be most cost-effective
when given to young adolescents who are not yet sexually active
(Knipe and Howley, 2007). Gardasil® is FDA approved for the
prevention of HPV types 6, 11, 16 and 18 infection in young women
aged 9-26. On October 16, 2009, the FDA also approved Gardasil®
to prevent genital warts caused by HPV types 6 and 11 in young
men aged 9-26 (Markowitz et al., 2007; FDA, 2009).Cervarix® also
prevents infection by HPV 16 and 18. Cervarix® was approved for
use in the United States in October 2009 (FDA, 2009). Both the
quadrivalent HPV 16/18/6/11 vaccine (Gardasil®, Merck & Co.,
Inc.) and the bivalent HPV 16/18 vaccine (Cervarix™,
GlaxoSmithKline Biologicals) have shown high safety, efficacy,
and immunogenicity. HPV types 16 and 18 are responsible for 70%
of the cases of cervical cancer worldwide. The three dose
regimen required for vaccination with these vaccines costs $360,
the most expensive vaccine in history. When 80% of the cases of
cervical cancer occur in developing countries, the expense of the
vaccines becomes a difficult barrier (Goldie,2008)
4.0 MOLECULAR EPIDEMIOLOGY OF KSHV44

Several genomic regions display remarkable sequence variability,
making them useful as markers of strain diversity and potential
epidemiologic markers of spread. These include regions
surrounding internal sequence repeats and, especially, the coding
regions directly adjacent to the left- and right-hand terminal
repeats: ORFs K1 and K15, respectively (Knipe and Howley, 2006).
When many isolates are examined from across the globe, it is
clear that the K1 sequence allows separation into four major
subtypes: A, B, C, and D. These subgroup identifications are also
supported by sequence variations in other genomic loci (Zong et
al., 1999; Zong et al., 2002). The A and C subgroups have been
isolated from KSHV-infected subjects in Europe, the United
States, Asia, and the Middle East, whereas subgroup B strains are
found predominantly in sub-Saharan Africa; type D strains are
found primarily in south Asia, Australia, and the Pacific
islands. These findings suggest that KSHV is an ancient human
infection, having entered the human population at or around the
time that modern humans arose in Africa in the early Pleistocene
Age (Hayward, 1999).
45

and KSHV-encoded transforming genes have been identified. The
current model of KSHV-induced malignancy involves a combination
of proliferation, survival, and transformation mediated by
latently expressed viral proteins together with a paracrine
mechanism that is exerted directly or indirectly by the lytically
expressed v-cytokines and viral G-protein coupled receptor
(vGPCR). HIV-1 infected individuals are at the highest risk for
developing KS. Interestingly, KS lesions and tumors appear to
regress in patients who receive Highly Active Antiretroviral
Therapy (HAART), suggesting that KSHV gene expression may be
insufficient to initiate or maintain transformation (Lebbeet al.,
1998; Gill et al.,2002).
Patterns of KSHV Infection and non-HIV KS
KS incidence Regions
Population KSHVprevalence
Transmission Risk groups
Low North America,North Europe,
<5% Sexual Homosexual men, STDclinic attendees,
46

Asia transplantrecipients
Intermediate
Mediterranean,Middle Easterncountries,Caribbean
5-20% Sexual,nonsexual
Homosexual men, STDclinic attendees,transplantrecipients, olderadults
High Africa, parts ofAmazon basin
>50% Nonsexual,sexual
Older adults, lowersocioeconomicstatus
AIDS-KS rates are highly dependent on local HIVinfection rates and risk groups.Source: Fields Virology, 5th Edition Copyright ©2007 LippincottWilliams & Wilkins
4.1 TRANSMISSION OF KSHV
The relatively close conformity of the subtypes with ethnic and
geographic clusters also implies that (at least before the
context of the AIDS epidemic) the virus is primarily transmitted
in a familial pattern (e.g., vertically from parent to child and
horizontally among siblings within the family unit), with wider
horizontal transmission being less efficient. This inference is47

consistent with much of what is known from direct study ofKSHV
epidemiology (Knipe and Howley, 2006)
4.2 TREATMENT, PREVENTION AND CONTROL
Kaposi's sarcoma can be treated either surgically or through
local irradiation. Chemotherapy with drugs such as liposomal
anthracyclines or paclitaxel may be used, particularly for
invasive disease. Antiviral drugs, such as ganciclovir, that
target the replication of KSHV have been used to successfully
prevent development of Kaposi's sarcoma(Martin et al., 1999). For
patients with AIDS-KS, as mentioned earlier, the most effective
therapy is highly active antiretroviral therapy to reduce HIV
infection.
48

5.0 EPSTEIN-BARR VIRUS
Epstein-Barr virus (EBV) was discovered through the seminal
observations of Denis Burkitt, a British colonial surgeon in
Uganda, and Anthony Epstein, a pathologist in the United Kingdom.
In 1957, in Kampala, Burkitt encountered two young children with
multifocal jaw tumors, a disease for which he had no knowledge or
previous experience. He established that the tumors were
lymphomas and undertook a journey through sub-Saharan Africa to
study the prevalence of the lymphoma among the various tribes
living in different areas. Surprisingly, the lymphomas were
prevalent in children in climates supportive of holoendemic
malaria (Knipe and Howley, 2006).
Other diseases associated with EBV include; infectious
mononucleosis, Hodgkin’s lymphoma, and nasopharyngeal carcinoma.
In addition, EBV is linked with oral hairy leukoplakia, which may
be an earliest oral manifestation of HIV infection (Jain et al.,
2011)
49

EBV is a ubiquitous double-stranded DNA virus of the γ
herpesviruses subfamily of the Lymphocryptovirus (LCV) genus
(McLaughlin-Drubin and Munger, 2008). The EBV genome is a linear,
double-stranded, 184 kbp DNA composed of 60 mole percent guanine
and cytosine (Baer et al., 1984; Kieff and Levine, 1974 ;
Pritchett et al., 1975). As with other herpesviruses, EBV has a
toroid shaped DNA core in a nucleocapsid with 162 capsomeres, an
outer envelope with external glycoprotein spikes, and a protein
tegument between the nucleocapsid and envelope (Dolyniuk et al.,
1976; Dolyniuk et al.,1976; Epstein et al., 1965; Johannsen et
al., 2004)
5.1 MOLECULAR EPIDEMIOLOGY OF EBV
The Epstein-Barr virus is a human herpes virus (human herpes
virus 4) that infects over 90% of adults worldwide, irrespective
of race, ethnicity or geography (Rickinson and Kieff, 1996).
Worldwide, more than 95% of the population is infected with EBV
(Evans and Niederman, 1989); the majority of EBV infections occur
50

during childhood without causing overt symptoms. Post adolescent
infection with EBV frequently results in mononucleosis, a self-
limiting lymphoproliferative disease. EBV infects and replicates
in the oral epithelium, and resting B lymphocytes trafficking
through the oral pharynx become latently infected. Infected B
lymphocytes resemble antigen activated B cells, and EBV gene
expression in these cells is limited to a B cell growth program,
termed Latency III, that includes LMP1, LMP2a/b, EBNAs -1, -2, -
3a-3b, -3c, and -LP, miRNAs, BARTs, and EBERs. These cells are
eliminated by a robust immune response to EBNA3 proteins,
resulting in Latency I, a reservoir of latently infected resting
memory B cells expressing only EBNA1 and LMP2. The
differentiation of memory cells to plasma cells results in
reactivation of the replication phase of the viral life cycle
that includes expression of latency III gene products. In
addition, there is likely another amplification step by
reinfection of the epithelial cells followed by shedding virus in
the saliva to the next host.
51

5.2 TRANSMISSION OF EBV
Epstein-Barr virus is spread by the oral route. Most children in
the developing world become infected within the first 3 years of
life (IARC, 1997; Evans et al., 1989). In crowded, poor socio-
economic environments, primary infection occurs early (de Thé et
al., 1975) and over 80% of children in Uganda are thought to be
sero-positive by the age of one (Kafuko et al., 1972). Early
infection is generally clinically silent. Under more affluent,
less crowded conditions, primary infection may not occur until
adolescence or young adulthood, when a round 50% of cases present
as infectious mononucleosis or ‘kissing disease’. Those infected
remain life-long carriers (Baumforth et al., 1999). In apparent
contradiction to its ubiquity, EBV has oncogenic potential and is
able to transform B-cells in culture into a state of continuous
proliferation or ‘immortalization’(Khannaet al., 1995).
5.3 EBV AND BURKITT’S LYMPHOMA IN NIGERIA
Burkitt's lymphoma (BL) is an unusual type of non-Hodgkin's
lymphoma occurring endemically in the narrow zone across central
Africa, where malaria is endemic. (Ferry, 2006; Cheneet al., 2009)
52

As of a study published in 2009, comprising 5-year retrospective
review of pediatric solid tumors as seen at the Jos University
Teaching Hospital, Nigeria, non-hodgkin's lymphoma and BL
accounted for 17 (19.5%) and 12 (13.8%), respectively, of a total
of 181 solid tumors of children. (Tankoet al., 2009)
In an another study that analyzed the profile of cancers recorded
in the first decade (1995-2004) of establishment of the Kano
cancer registry (KCR), a histology/cytology-based registry in
Kano, Nigeria, BL (31.4%), other lymphoreticular cancers (23.8%)
and retinoblastoma (20%) predominated in children of a total of
1990 cancer cases. (Mohammed et al., 2008)
6.0 MOLECULAR EPIDEMIOLOGY OF HBV
Hepatitis B Virus, a major public health problem world wide is
more prevalent in the developing countries. (Johnson et al., 1986)
More than 2 billion people are infected with HBV world-wide while
53

some 280 million are chronic carriers, harboring the virus in
their liver (Clement et al., 1990). About 2 million of these
carriers die each year as a result of cirrhosis or primary liver
cell cancer induced by the virus. This virus is responsible for
80% of all cases of primary liver cancer, which is one of leading
causes of death in Asia and Africa (Clement et al., 1990).
In Nigeria, 11.6% prevalence rate has been reported from
Maiduguri (Harry et al., 1994), 13.8% from Lagos (Nasidi et al.,
1993), 4.3% from Port Harcourt (Akani et al., 2005), 5.7% from
Ilorin (Agbede et al., 2007), 8.3% from Zaria (Luka et al.,
2008), 17.1% from female sex workers, 14.9% (Forbi et al., 2008)
from healthy blood donors (Ejele et al., 2004) and 25.7% among
surgeons (Belo, 2000). Although HBV prevalence varies widely
across the African continent, hepatitis B surface antigen (HBsAg)
positivity is estimated at 8-20% (Apurva and Jordan, 2007).
Zonal distribution of subjects assayed for Hepatitis B viral load in Nigeria
Geographical
Zones
HBV viral load
tests
54
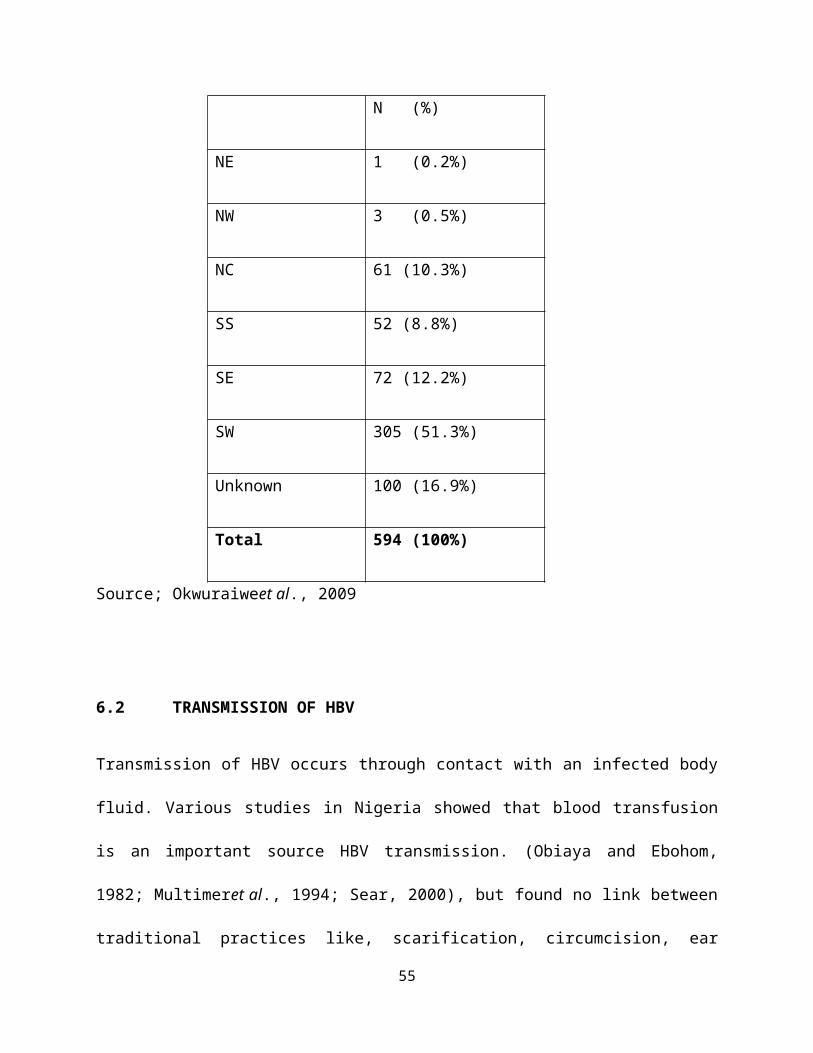
N (%)
NE 1 (0.2%)
NW 3 (0.5%)
NC 61 (10.3%)
SS 52 (8.8%)
SE 72 (12.2%)
SW 305 (51.3%)
Unknown 100 (16.9%)
Total 594 (100%)
Source; Okwuraiweet al., 2009
6.2 TRANSMISSION OF HBV
Transmission of HBV occurs through contact with an infected body
fluid. Various studies in Nigeria showed that blood transfusion
is an important source HBV transmission. (Obiaya and Ebohom,
1982; Multimeret al., 1994; Sear, 2000), but found no link between
traditional practices like, scarification, circumcision, ear
55

piercing and HBV infection (Chukwukaet al., 2003; Angyo and
Yakubu, 2001). Higher HBsAg prevalence noted among prisoners and
rural dwellers were attributed overcrowding and clustering.
(Amazigo and Chime, 1990). Studies from north-central Nigeria
indicates that unprotected sex is implicated in the transmission
of HBV (Ivlustapha and Jibrin, 2004; Sirisenaet al., 2002 )
The greatest sources of new infections worldwide have been from
infected mothers to the newborn, or among very young children.
The risk of vertical transmission varies, depending on geographic
regions. In North America, western Europe, and Africa, the risk
of vertical transmission from mothers who are chronically
infected is approximately 10% (Knipe and Howley, 2006)
6.3 HEPATOCARCINOGENESIS OF HBV
HBV-associated liver carcinogenesis is viewed a multifactorial
process. The integration of the HBV genome into the host
chromosome at early stages of clonal tumor expansion has been
demonstrated to both affect a variety of cellular genes as well
as exert insertional mutagenesis, while chronic liver
56

inflammation confers the accumulation of mutations in the host
genome.
HBV encodes HBx, a 154 amino acid multifunctional regulatory
protein that is highly conserved among all of the mammalian
hepadnaviruses(Diaoet al., 2001). HBx stimulates signal
transduction pathways such as MAPK/ERK and can also up-regulate
the expression of genes such as c-Myc, c-Jun, NF-κB, AP 1, Ap-2,
RPB5 subunit of RNA polymerase II, TATA binding protein, and CREB
(Pang etal., 2006).PreS2 activators, and HBSP may exert oncogenic
functions. However, exactly how these viral factors contribute to
higher risks of HCC development remains unclear.
6.4 CONTROL AND PREVENTION OF HBV INFECTION AND HCC
According to J Trop Paed (2003), there are broadly three
strategies for dealing with HBV infection in the developed
countries; immunization for at risk population, antiviral drugs
(lamivudine, adeforvir and dipivoxil) and immunostimulatory
therapy with alpha-interferon for those affected.
57

7.0 HEPATITIS C VIRUS (HCV)
HCV is a single-stranded RNA virus of the Hepacivirus genus in
the Flaviviridae family and is the only positive-stranded RNA
virus among the human oncogenic viruses. Its approximately 9.6 kb
genome contains an open reading frame (ORF) that codes for a 3000
amino acid residue polyprotein precursor (Choo et al., 1989) that
is cleaved by cellular and viral proteases into three structural
proteins (core, E1, E2) and seven nonstructural proteins (p7,
NS2, NS3, NS4a, NS4B, NS5A, and NS5B) (Lindenbach and Rice,
2005).
7.1 MOLECULAR EPIDEMIOLOGY OF HCV
HCV is the etiologic agent of posttransfusion and sporadic non-A,
non-B hepatitis (Choo et al., 1989) and infects approximately 2%
of the population worldwide, although the prevalence of HCV
infection varies by geographical location (Shepard et al., 2001)
HCV proteins have been reported to activate cellular oncoproteins
and inactivate tumor suppressors, such as p53, CREB2/LZIP, and
the retinoblastoma protein (pRB). HCV causes genome instability,
58

signifying that certain HCV proteins may have a mutator function
(McLaughlin-Drubin and Munger, 2008).
So far the prevalence of hepatitis C. virus infection is
increasing in Nigeria, ranging from 4.7-5% in Ilorin, to 5.3-6.6%
in Enugu, to 11% in Ibadan and 20% in Benin (Ejioforet al., 2010)
7.2 CONTROL AND PREVENTION OF HCV INFECTION
Available options for the management of HCV include; the use of
interferons, which are immune response modifiers; Ribavirin,
which is an antiviral drug; a combination of interferon and
ribavirin; or liver transplantation in the case of failed
chemotherapy (Ejioforet al., 2010)
CONCLUSION
Identification of viruses strongly associated with human cancers
provides an opportunity to advance preventive measures to inhibit
virus infections, hence reducing the risk of cancer.Development
of vaccines against cancer viruses is capable of reducing the
global incidence of cancer.
59

In sub-Saharan African countries, including Nigeria, where the
incidence of cervical cancer and other cancers of viral etiology
are on the increase, there is need to sensitize the citizenry on
the importance of early diagnosisand make vaccines available for
all those who are at risk.
60

REFERENCES
Achachi A, Florins A, Gillet N (2005). Valproate activates bovine
leukemia virus gene expression, triggers apoptosis, and induces
leukemia/lymphoma regression in vivo. ProcNatlAcad Sci U S
102(29):10309-10314.
Adams JM, Harris AW, Pinkert CA, Corcoran LM, Alexander WS, Cory
S, Palmiter RD,Brinster RL.(1985). The c-myc oncogene driven by
immunoglobulin enhancers induces lymphoid malignancy in
transgenic mice. Nature318:533–538.
Agbede, OO, Iseniyi, JO, Kolewale, MO, and Ojuowa, A. (2007).
RiskFactors and seroprevalence of hepatitis B antigenemia in
mothers and their preschool children in Ilorin, Nigeria.Therapy,
4(1): 67-72.
Akani, CI, Ojule, AC, Opurum, HC, and Ejilemele, AA (2005).
Seroprevalence of HBs Ag in pregnant women in Port Harcourt,
Nigeria.Post graduate Medical Journal12(4): 266-270.
61

Angyo I. A., Yakubu A. M. (2001) Lack of association between some
risk factors and Hepatitis B Surface Antigenaemia in children
with sickle cell anaemia.W Afr Med J 20: 214-8.
Anorlu RI, (2008)Cervical cancer: The Sub-Saharan African
Perspective. Reproduction Health Matters16(32):41-49
http://simplelink.library.utoronto.ca.myaccess.library.utoronto.c
a/url.cfm/107825
Apurva A.M. and Jordan J.F. (2007).Viral hepatitis and HIV in
Africa. AIDS Reviews9: 25-39.
Azimi N, Brown K, Bamford RN, (1998). Human T cell lymphotropic
virus type I Tax protein trans-activates interleukin 15 gene
transcription through an NF-kappa B site. ProcNatl Acad Sci U S A
95:2452-2457
B.H. Sweet, M.R. Hilleman, (1960)The vacuolating virus, S.V. 40,
Proc. Soc. Exp. Biol. Med. 105 420–427.
Baer R (1984). DNA sequence and expression of the B95-8 Epstein-
Barr virus genome. Nature 310(5974):207-211.
62

BannertN, KurthR. (2004) Retroelements and the human genome: new
perspectives on an old relation, Proc. Natl. Acad. Sci. U. S. A. 101
(Suppl 2) 14572–14579.
Baumforth KRN, Young LS, Flavell KJ, Constandinou C, Murray PG.
(1999) Demystified ... The Epstein-Barr virus and its
association with human cancers. J Clin Pathol: Mol Pathol; 52:307-322.
Belo, A.C. (2000). Prevalence of hepatitis B virus markers in
Surgeons in Lagos, Nigeria.East African Medical JournalVol. 77 No.
5 May 2000; 283-285.
Beraud C, Sun S-C, Ganchi P, Ballard DW, Greene WC. (1994). Human
T-cell leukemia virus type I Tax associates with and is
negatively regulated by the NF-kB2 p100 gene product:
implications for viral latency. Mol Cell Biol14:1374–1382
Blayney DW, Jaffe ES, Fisher RI (1983) . The human T-cell
leukemia/lymphoma virus, lymphoma, lytic bone lesions, and
hypercalcemia. Ann Intern Med;98(2):144-151.
Boshart M, Gissmann L, Ikenberg H, Kleinheinz A, Scheurlen W, zur
Hausen H.(1984)A new type of papillomavirus DNA, its presence
63

in genital cancer biopsies and in cell lines derived from
cervical cancer. Embo J.;3:1151–7. [PMC free article] [PubMed]
BoxmanIL,BerkhoutRJ, MulderLH,WolkersMC,BouwesBavinckJN,
VermeerBJ, ter ScheggetJ. (1997) Detection of human
papillomavirus DNA in plucked hairs from renal transplant
recipients and healthy volunteers, J. Invest. Dermatol.108 712–715.
BoxmanIL. MulderLH, RussellA, BouwesBavinckJN, Green A,
TerScheggetJ.(1999) Human papillomavirus type 5 is
commonly present in immunosuppressed and immunocompetent
individuals, Br. J. Dermatol. 141 246–249.
Brighty, DW, JassalSR.(2001). The synthetic peptide P-197
inhibits human T-cell leukemia virus type 1 envelope-mediated
syncytium formation by a mechanism that is independent of
Hsc70. J. Virol. 75: 10472–10478.Abstract/FREE Full Text.
Burchell AN, Winer RL, de Sanjosé S, Franco EL (2006). Chapter 6:
Epidemiology and transmission dynamics of genital HPV
infection. Vaccine. Aug 31;24Suppl 3:S3/52-61. Epub 2006 Jun
2. [PUBMED
64

Centre for Disease Control and Prevention (CDC) 2012.
Chang Y,Cesarman E,Pessin MS, Lee F, Culpepper J, Knowles DM,
Moore PS.(1994) Identification of herpesvirus-like DNA
sequences in AIDS-associated Kaposi's sarcoma, Science2661865–
1869.
Chene A, Donati D, Orem J, MbiddeER, Kironde F, Wahlgren M.(2009)
Endemic Burkitt's lymphoma as a polymicrobial disease: New
insights on the interaction between Plasmodium falciparum and
Epstein-Barr virus. Semin Cancer Biol;19:411-20.
Choo QL,Kuo G, Weiner AJ, Overby LR, BradleyDW, Houghton M.
(1989) Isolation of a cDNA clone derived from a blood-borne
non-A, non-B viral hepatitis genome, Science 244 359–362.
Chuang SE, Doong SL, Lin MT. (1997) Tax of the human T-
lymphotropic virus type transactivates promoter of the MDR-1
gene. BiochemBiophys Res Commun238:482- 486.
Chukwuka JO, Ezechukwu CC, Egbuonu I. (2003) Cultural Influences
on Hepatitis B Surface Antigen Seropositivity in Primary School
in Nnewi. Nig J Paed30 :140-2.
65

Cleghorn FR, Manns A, Falk R(1995). Effect of human T-
lymphotropic virus type I infection on non-Hodgkin's lymphoma
incidence.J Nat Cancer Inst;87:1009-1014.
Clement CJ, Kane M, Hu DJ, Kim-farleyR.l. (1990) Hepatitis B
vaccine joins fight against Pandemic disease. World Health
Forum11:165 - 8.
CrumCP, IkenbergH, RichartRM, GissmanL.(1984) Human
papillomavirus type 16 and early cervical neoplasia, N. Engl. J.
Med. 310 880–883.
DaneDS, CameronCH, BriggsM. (1970) Virus-like particles in serum
of patients with Australia-antigen-associated hepatitis,
Lancet1 695–698.
de Thé G, Day NE, Geser A, Lavoué MF, Ho JHC, Simons (1975).
Sero-epidemiology of the Epstein-Barr virus : preliminary
analysis of an international study - a review, In: de-Thé G,
Epstein MA, zur Hausen H, editors. Oncogenesis and Herpes Viruses
II. Lyon: International Agency for Research on Cancer; p. 3-16.
66

de VilliersEM, FauquetC, BrokerTR, BernardHU, zur HausenH. (2004)
Classification of papillomaviruses, Virology324 17–27.
DiaoJ, GarcesR, RichardsonCD.(2001) X protein of hepatitis B
virus modulates cytokine and growth factor related signal
transduction pathways during the course of viral infections
and hepatocarcinogenesis, Cytokine Growth Factor Rev. 12 189–205
Dolyniuk M, Pritchett R, Kieff E.(1976)Proteins of Epstein-Barr
virus. I. Analysis of the polypeptides of purified enveloped
Epstein-Barr virus. J Virol;17(3):935-949.
Dolyniuk M, Wolff E, Kieff E. (1976)Proteins of Epstein-Barr
virus. II. Electrophoretic analysis of the polypeptides of
the nucleocapsid and the glucosamine- and polysaccharide-
containing components of enveloped virus. J Virol;18(1):289-
297.
Duensing S, Duensing A, Crum CP, Munger K. (2001) Human
papillomavirus type 16 E7 oncoprotein-induced abnormal
centrosome synthesis is an early event in the evolving
malignant phenotype, Cancer Res. 61 2356–2360.
67

Duensing S, Lee LY, Duensing A, Basile J,Piboonniyom S, Gonzalez
S, Crum CP, Munger K. (2000) The human papillomavirus type 16
E6 and E7 oncoproteins cooperate to induce mitotic defects and
genomic instability by uncoupling centrosome duplication from the
cell division cycle, Proc. Natl. Acad. Sci. U. S. A. 97 10002–
10007.
Duensing S, Munger K.(2002) The human papillomavirus type 16 E6
and E7 oncoproteins independently induce numerical and
structural chromosome instability, Cancer Res. 62 7075–7082.
Dunne EF, Unger ER, Sternberg M, McQuillan G, Swan DC, Patel SS,
Markowitz LE.(2007) Prevalence of HPV infection among females in
the United States.JAMA.297(8):813-9. [PUBMED]
Durst M, Gissmann L, IkenbergH, zur Hausen HA.(1983)
papillomavirus DNA from a cervical carcinoma and its
prevalence in cancer biopsy samples from different geographic
regions. ProcNatlAcad Sci U S A.;80:3812–5. [PMC free article]
[PubMed]
68

Ejele O, Nwauche C,Erhabor O. (2004). The prevalence of hepatitis
B surface antigen in HIV- positive patients in the Niger
Delta Nigeria.Niger J Med 2004, 13: 175-9.
Ejiofor OS, Emechebe GO, Igwe WC, Ifeadike CO, Ubajaka CF. (2010)
Hepatitis C virus infection in Nigerians. Niger Med J [serial
online] [cited 2013 May 8];51:173-6. Available from:
http://www.nigeriamedj.com/text.asp?2010/51/4/173/73290
Epstein M, Achong B, Barr Y. (1965)Morphological and biological
studies on a virus in cultured lymphoblasts from Burkitt's
lymphoma. J Exp Med;121:761-770
EpsteinMA,AchongBG, BarrYM(1964).Virus particles in cultured
lymphoblasts from Burkitt's lymphoma, Lancet1 702–703.
Ezegbudo CN, Agba MI, Agbonlahor DE, Nwobu GO, Igwe CU, Agba MI.
(2004) The seroprevalence of hepatitis B Surface antigen and
human immuno deficiency virus among pregnant women in
Anambra state Nigeria. Shir E-Med J; 5 : 20 - 2.
69

Fakunle YM, Abdulrahman MB, Whittle AC. Hepatitis B virus
infection in children and adults in Northern Nigeria, a
Preliminary Survey. Trans R Soc Trop Med Hyg,;75 : 626-9.
FeitelsonMA, LeeJ. (2007) Hepatitis B virus integration, fragile
sites, and hepatocarcinogenesis, Cancer Lett. 252 157–170.
Ferry JA. (2006) Burkitt's lymphoma: Clinicopathologic features
and differential diagnosis. Oncologist;11:375-83.
FloreO, RafiiS, ElyS, O'LearyJJ, HyjekEM, CesarmanE. (1998)
Transformation of primary human endothelial cells by Kaposi's
sarcomaassociated herpesvirus, Nature394 588–592.
Food and Drug and Administration Vaccines, Blood, & Biologics
Accessed October 21st, 2009
[http://www.fda.gov/BiologicsBloodVaccines/Vaccines/ApprovedProdu
cts/ucm186991.h tm]
Forbi JC, Onyemauwa N, Gyar SD, Oyeleye AO, Entonu P, Agwale SM.
(2008) High Prevalence of Hepatitis B Virus among Female Sex
Workers in Nigeria. Rev. Inst. Med. trop. S. Paulo50(4):219-221.
70

Franchini G, Cereseto A, Tryniszewska E. (1999) Telomerase
activation, rearrangement, and inhibition of cell-cycle
inhibitors in human T-cells immortalized or transformed by HTLV-
I. In: Semmes OJ, Hammarskjold M-L, eds. Molecular Pathogenesis
of HTLV-I: A Current Perspective. Arlington: ABI Professional
Publications; 59-70.
Franzese O, Balestrieri E, Comandini A.(2002) Telomerase activity
of human peripheral blood mononuclear cells in the course of
HTLV type 1 infection in vitro. AIDS Res Hum
Retroviruses;18(4):249-251.
Gallo RC. (1983) Association of the human type C retrovirus with
a subset of adult T-cell cancers.Cancer Res.;43(8):3892–3899
GardnerSD, FieldAM, ColemanDV, HulmeB.(1971) New human
papovavirus (B.K.) isolated from urine after renal
transplantation, Lancet1 1253–1257
Gessain A, Mahieux R. (2000) A virus called HTLV-1.
Epidemiological aspects.Presse Med;29(40):2233-2239
71

Gessian A, Yanagihara R, Franchini G, Garruto RM, Jenkins CL,
Ajdukiewicz AB, Gallo RC, Gajdusek DC (1991). Highly
divergent molecular variants of human T- lymphotropic virus
type I from isolated populations in Papua New Guinea and the
Solomon Islands, Proc. Natl. Acad. Sci. U. S.A. 88: 7694–7698.
Gifford R, Tristem M. (2003) The evolution, distribution and
diversity of endogenous retroviruses, Virus Genes 26 291–315.
GillJ, BourbouliaD, WilkinsonJ, HayesP, CopeA, MarcelinAG,
CalvezV, Gotch F,Boshoff C, Gazzard B. (2002) Prospective
study of the effects of antiretroviral therapy on Kaposi
sarcoma-associated herpesvirus infection in patients with and
without Kaposi sarcoma, J. Acquir. Immune.Defic.Syndr. 31 384–390.
Goldie SJ.(2008) Health and economic outcomes of HPV 16,18
vaccination in 72 GAVI-eligible countries, Vaccine,26:4080-
4093
http://simplelink.library.utoronto.ca.myaccess.library.utoronto.c
a/url.cfm/107855
72

GoodwinEC, DiMaioD. (2000) Repression of human papillomavirus
oncogenes in HeLa cervical carcinoma cells causes the orderly
reactivation of dormant tumor suppressor pathways, Proc. Natl.
Acad. Sci. U. S. A. 97 12513–12518.
Grassmann R, Berchtold S, Radant I, Alt M, Fleckenstein B,
Sodroski JG, Haseltine WA, Ramstedt U.(1992) Role of human T-
cell leukemia virus type 1 X region proteins in immortalization
of primary human lymphocytes in culture. J Virol.;66:4570–5. [PMC
free article] [PubMed]
Grassmann R, Dengler C, Muller-Fleckenstein I.(1989)
Transformation to continuous growth of primary human T
lymphocytes by human T-cell leukemia virus type I X-region genes
transduced by a herpesvirus saimiri vector. ProcNatlAcad Sci U
S A;86(9):3351-3355.
Greene WC, Leonard WJ, Depper JM. (1986) The human interleukin-2
receptor: normal and abnormal expression in T cells and in
leukemias induced by the human T-lymphotropic retroviruses.
Ann Intern Med;105(4):560-572.
73

H. zur Hausen, Viruses in human cancers.(2001) Curr. Sci.81 523–
527.
HaaheimLR, PattisonJR, WhitleyRJ.(2002) A Practical Guide to Clinical
Virology. John Wiley & Sons, Ltd. ISBNs: 0-470-84429-9 (HB); 0-
471-95097-1 (PB)
Harry TO, Bajani MD, Moses AE.(1994). Hepatitis B virus infection
among blood donors and pregnant women in Maiduguri,
Nigeria.East Africa Medical Journal70: 596-597.
Hayward GS.(1999) KSHV strains: the origins and global spread of
the virus. Semin Cancer Biol;9(3):187-199
Hepatitis B. Immunization.(2003) [Editorial] J Trop Paed.; 48: 256-
7.
Hino S, Katamine S, Kawase K.(1994) Intervention of maternal
transmission of HTLV-1 in Nagasaki, Japan. Leukemia;8[Suppl
1]:S68-70.
Hirai H, Fujisawa J, Suzuki T, Ueda K, Muramatsu M, Tsuboi A,
Arai N, Yoshida M. 1992. Transcriptional activator Tax of
74

HTLV-1 binds to the NF-kappa B precursor p105. Oncogene7:1737–
1742.
Holland JF, Pogo BGT, (2004) Mouse mammary tumor virus-like
infection and human breast cancer, Clin. Cancer Res. 10 5647–5649.
Iwanaga Y, Tsukahara T, Ohashi T.(1999) Human T-cell leukemia
virus type 1 tax protein abrogates interleukin-2 dependence
in a mouse T-cell line. J Virol;73(2):1271-1277
Jackson S, Harwood C, Thomas M, Banks L, Storey A. (2000) Role of
Bak in UV-induced apoptosis in skin cancer and abrogation by
HPV E6 proteins, Genes Dev. 14 3065–3073.
Jaffe ES, Blattner WA, Blayney DW.(1984) The pathologic spectrum
of adult T-cell leukemia/lymphoma in the United States. Human
T-cell leukemia/lymphoma virus- associated lymphoid
malignancies. Am J Surg Pathol;8(4):263-275.
Jain N, Bhatia V, Lattoo S. (2011) Epstein-Barr virus and
associated head and neck manifestations. Ann Nigerian Med
[serial online] [cited 2013 May 6];5:38-41. Available from:
http://www.anmjournal.com/text.asp?2011/5/2/38/92947
75

Janet S. Butel,(2000) Carcinogenesis21 (3): 405-426. doi:
10.1093/carcin/21.3.405
Jeon S, Allen-Hoffmann BL,Lambert PF. (1995) Integration of
human papillomavirus type 16 into the human genome correlates
with a selective growth advantage of cells, J. Virol. 69 2989–2997.
Jin DY, Spencer F, Jeang KT. (1998) Human T cell leukemia virus
type 1 oncoprotein tax targets the human mitotic checkpoint
protein MAD1. Cell 93:81–91.
Johannsen E. Proteins of purified Epstein-Barr virus.(2004)
ProcNatlAcad Sci USA;101(46):16286-16291.
John B Liao. (2006)Yale J Biol Med. 2006 December; 79(3-4): 115–122.
Published online 2007 October
Johnson AOK., Sodeinde O, Odeola HA, Ayoola EA. (1986) Survey of
Hepatitis A and B infections in childhood in Ibadan -
Preliminary Study: Nig J Paed; 13: 83 -6.
Jones KS.Petrow-Sadowski C, BertoletteDC, Huang Y,Ruscetti FW.
(2005) Heparan sulfate proteoglycans mediate attachment and
76

entry of human T-cell leukemia virus type 1 virions into CD4+ T
cells. J. Virol. 79: 12692–12702.Abstract/FREE Full Text.
Kafuko GW, Henderson BE, Kirya BG, Munube GM, Tukei PM, Day NE.
(1972) Epstein-Barr virus antibody levels in children from
the West Nile District of Uganda. Report of a field
study.Lancet;1:706-709.
Kanno T, Brown K, Franzoso G, Siebenlist U. (1994) Kinetic
analysis of human T-cell leukemia virus type I tax-mediated
activation of NF-kappaB. Mol Cell Biol14:6443–6451.
Kasai T, Iwanaga Y, Iha H, Jeang KT. (2002) Prevalent loss of
mitotic spindle checkpoint in adult T-cell leukemia confers
resistance to microtubule inhibitors. J BiolChem 277:5187–5193
Kawano F, Yamaguchi K, Nishimura H. (1985) Variation in the
clinical courses of adult T-cell leukemia. Cancer 55(4):851-856
Keath EJ, Caimi PG, Cole MD. (1984) Fibroblast lines expressing
activated c-myc oncogenes are tumorigenic in nude mice
and syngeneic animals. Cell39:339–348.
77

Khanna R, Burrows SR, Moss DJ. (1995) Immune regulation in
Epstein-Barr virus-associated diseases.Microbiol Rev59:387-405.
Kieff E, Levine J.(1974) Homology between Burkitt herpes viral
DNA and DNA in continuous lymphoblastoid cells from patients
with infectious mononucleosis. ProcNatlAcad Sci USA;71(2):355-
358.
Kire CF. (1993)The epidemiology and control of hepatitis B in
Sub Saharan Africa, Prog Med Virology; 40 : 143-56.
Kirken RA, Erwin RA, Wang L. (2000) Functional uncoupling of the
janus kinase 3-stat5 pathway in malignant growth of human T
cell leukemia virus type 1-transformed human T cells. J
Immunol;165(9):5097-5104
Knipe DM, Howley PM.(2007) Title: Fields Virology, 5th Edition
Copyright Lippincott Williams & Wilkins, p.2300
Kondo T, Kono H, Miyamoto N. (1989) Age- and sex-specific
cumulative rate and risk of ATLL for HTLV-I carriers. Int J
Cancer;43(6):1061-1064.
78

Lagunoff M, Ganem D. (1997) Organization of the termini of the
genome of the Kaposi's sarcoma-associated herpesvirus (human
herpesvirus 8). Virology;236:147-154
Lamb DA, Mirsaliotis SM, Kelly DW.(2009). Basic residues are
critical to the activity of peptide inhibitors of human T cell
leukemia virus type 1 entry. J. Biol. Chem. 284: 6575–
6584.Abstract/FREE Full Text
Lau A, Nightingale S, Taylor GP.(1998) Enhanced MDR1 gene
expression in human T-cell leukemia virus-I-infected patients
offers new prospects for therapy. Blood;91:2467-2474.
LebbeC, Blum L, PelletC, BlanchardG, VerolaO, MorelP, DanneO,
CalvoF. (1998) Clinical and biological impact of antiretroviral
therapy with protease inhibitors on HIV-related Kaposi's
sarcoma, Aids12F45–F49.
Lemoine FJ, Wycuff DR, Marriott SJ.(2001). Transcriptional
activity of HTLV-1 Tax influences the expression of marker genes
associated with cellular transformation. Dis Markers 17:1–9.
79

Leonard IC, Wilson IBO, Nnenna CM. (2003) Trop J
ObstetGynaecol, ,20: 109-112
Li K, Zhang S, Kronqvist M, Wallin M, Ekström M,Derse D, Garoff
H. (2008)Intersubunit disulfide isomerization controls
membrane fusion of human T-cell leukemia virus Env. J. Virol. 82:
7135–7143.Abstract/FREE Full Text25.
LowyDR, HowleyPM,(2001) in: D.M. Knipe, P.M. Howley (Eds.), Fields'
Virology, Lippincott Williams and Wilkins, New York, pp. 2231–
2264.
Luka SA, Ibrahim MB, Iliya, SN. (2008). Seroprevalence of
Hepatitis B surface antigen among pregnant women attending
Ahmadu Bello University Teaching Hospital Zaria, Nigerian
Journal of Parasitology 29 (1):38-41.
Margaret E. McLaughlin-Drubin and Karl Munger.
(2008)BiochimBiophysActa 2008 March; 1782(3): 127–150.
Published online 2007 December 23. doi:
10.1016/j.bbadis.2007.12.005
80

Mariner JM, Lantz V, Waldmann TA.(2001) Human T cell lymphotropic
virus Type I Tax activates IL-15Ralpha gene expression through
an NF-kappaB site. J Immunol;166(4):2602-2609
Markowitz LE, Dunne EF, Saraiya M, Lawson HW, Chesson H, Unger ER
(2007) ; Centers for Disease Control and Prevention (CDC);
Advisory Committee on Immunization Practices
(ACIP).Quadrivalent Human Papillomavirus Vaccine:
Recommendations of the Advisory Committee on Immunization Practices (ACIP).
MMWR Recomm Rep. 2007 Mar 23;56(RR-2):1-24. [PUBMED]
Matsuoka M, Jeang KT. (2005) Human T-cell leukemia virus type I
at age 25: a progress report. Cancer Res;65(11):4467-4470.
Matsuoka M.(2003) Human T-cell leukemia virus type I and adult T-
cell leukemia. Oncogene.;22:5131–40.
McCreath WA, Salom E, Chi DS.(2005)Cervical cancer: current
management of early/late disease. SurgOncol Clin N Am;14:249-
266
MigoneTS, Cacalano NA, Taylor N. (1998) Recruitment of SH2-
containing protein tyrosine phosphatase SHP-1 to the
81

interleukin 2 receptor; loss of SHP-1 expression in human T-
lymphotropic virus type I-transformed T cells. ProcNatlAcad Sci
U S A;95:3845-3850.
Migone TS, Lin JX, Cereseto A.(1995) Constitutively activated
Jak-STAT pathway in T cells transformed with HTLV-I.
Science;269:79-81
Mohammed AZ, Edino ST, Ochicha O, Gwarzo AK, Samaila AA. (2008)
Cancer in Nigeria: A 10-year analysis of the Kano cancer
registry. Niger J Med;17:280-4.
Mufti GJ, Pawson R, Pagliuca A. (1998) Management of adult T-cell
leukaemia/lymphoma.Br J Haematol;100:453-458.
Mulloy JC, Migone TS, Ross TM.(1998) Human and simian T-cell
leukemia viruses type 2 (HTLV-2 and STLV-2(pan-p)) transform T
cells independently of Jak/STAT activation. J Virol;72(5):4408-
4412.
Multimer DJ, Olomi A, Skidmore S, Olomu N, Ratcliffe D, Rodger S.
(1994) Viral hepatitis in Nigeria-sickle cell disease and
commercial blood donors. QJM; 87: 407-11
82

Munger K, Baldwin A, Edwards KM, Hayakawa H, Nguyen CL, Owens M,
Grace M, Huh K. (2004) Mechanisms of human papillomavirus-
induced oncogenesis, J. Virol.78 11451– 11460.
Munger K, Howley PM.(2002) Human papillomavirus immortalization
and transformation functions, Virus Res. 89 213–228.
Nasidi ATO, Vyazor SO, Numumbe GMR, Azzan BB, Ancinlev VA.(1983)
Prevalence of Hepatitis B Infection Marker in two different
geographical areas of Nigeria.Preceedings of the first international
conference, 12-15 December, 1983, Lagos, Nigeria.
Nerenberg M, Hinrichs SH, Reynolds RK, Khoury G, Jay G. (1987)
The tat gene of human T- lymphotropic virus type 1 induces
mesenchymal tumors in transgenic mice. Science.;237:1324–9.
[PubMed]
NishizawaT, OkamotoH, KonishiK, YoshizawaH, MiyakawaY, MayumiM.
(1997) A novel DNA virus (TTV) associated with elevated
transaminase levels in post transfusion hepatitis of unknown
etiology, Biochem.Biophys. Res. Commun. 241 92–97.
83

Obiaya MO, Ebohom PA. (1982) Blood Transfusion Hazards in Benin
City, Nigeria.Nig Med J; 12: 251-4.
Ohshima K, Suzumiya J, Sato K. (1999) Survival of patients with
HTLV-I-associated lymph node lesions. J Pathol;189:539-545.
PadgettBL, WalkerDL, ZuRheinGM, EckroadeRJ,DesselBH, (1971)
Cultivation of papova- like virus from human brain with
progressive multifocal leucoencephalopathy, Lancet 1 1257–1260.
Parkin DM, Bray F, Ferlay J, Pisani P.(2005) Global cancer
statistics, 2002. CA Cancer J Clin.;55:74–108. [PubMed
ParkinDM.(2006) The global health burden of infection-associated
cancers in the year 2002, Int. J. Cancer 118 3030–3044.
Pawson R, Richardson DS, Pagliuca A. (1998) Adult T-Cell
leukemia/lymphoma in London: clinical experience of 21 cases.
Leuk Lymphoma;31:177-185.
Piñón JD, KellySM, PriceNC, FlanaganJU,BrightyDW. (2003). An
antiviral peptide targets a coiled-coil domain of the
human T-cell leukemia virus envelope glycoprotein. J. Virol. 77:
3281–3290.Abstract/FREE Full Text.
84

Piñon, JD,KlassePJ,JassalSR, WelsonS,
WeberJ,BrightyDW,SattentauQJ. (2003). Human T-cell leukemia
virus type 1 envelope glycoprotein gp46 interacts with cell
surface heparan sulfate proteoglycans. J. Virol. 77: 9922–
9930.Abstract/FREE Full Text.
Poiesz BJ, Ruscetti FW, Gazdar AF, Bunn PA, Minna JD, Gallo RC.
(1980) Detection and isolation of type C retrovirus particles
from fresh and cultured lymphocytes of a patient with cutaneous
T-cell lymphoma.ProcNatlAcad Sci U S A.;77:7415–9. [PMC free
article] [PubMed]
Pozzatti R, Vogel J, Jay G.(1990)The human T-lymphotropic virus
type I tax gene can cooperate with the ras oncogene to
induce neoplastic transformation of cells. Mol Cell Biol.;10:413–7.
[PMC free article] [PubMed]
Prager D, Rosenblatt JD, Ejima E. (1997) Hypercalcemia,
parathyroid hormone-related protein expression and human T-
cell leukemia virus infection. In: Polliack A, ed. Leukemia & Lymphoma
Reviews 6. Harwood Academic Pub. Newark, NJ:77-82.
85

Proietti FA, Carneiro-Proietti ABF, Catalan-Soares BC, Murphy EL.
(2005) Global epidemiology of HTLV-I infection and associated
diseases.Oncogene.;24:6058–68. DOI: 10.1038/sj.onc.1208968
Renne R, Lagunoff M, Zhong W. (1996)The size and conformation of
Kaposi's sarcoma- associated herpesvirus (human herpesvirus 8)
DNA in infected cells and virions. J Virol;70:8151-8154
Rickinson AB, Kieff E.(1996) Epstein-Barr virus, In: Field's Virology
Fields BN, Knipe DM, Howley PM, Chanock RM,
Melnick JL, Monath T P, editors.. Philadelphia, P. A . :
Lippincott-Raven;. p. 2397-2446.
Russo JJ, Bohenzky RA, Chien MC.(1996) Nucleotide sequence of the
Kaposi sarcoma- associated herpesvirus (HHV8). ProcNatlAcad Sci U
S A;93:14862-14867.
Sagara Y, InoueY,ShirakiH,JinnoA, HoshinoH, MaedaY. (1996).
Identification and mapping of functional domains on human T-
cell lymphotropic virus type 1 envelope proteins by using
synthetic peptides. J. Virol. 70: 1564–1569.Abstract/FREE Full Text.
86

Sato K, Maruyama I, Maruyama Y.(1991) Arthritis in patients
infected with human T lymphotropic virus type I. Clinical and
immunopathologic features. Arthritis Rheum;34(6):714-721
Sear M. (2000) The Jaundiced Child. In: a Manual of Tropical Paediatrics.
Cambridge University press, 185-200.
SeegerC, MasonWS,(2000) Hepatitis B virus biology, Microbiol MolBiol
Rev 64 51–68.
Serkies K, Jassem J. (2005) Chemotherapy in the primary treatment
of cervical carcinoma. Crit Rev Oncol Hematol;54:197-208
Shamanin V, Glover M, Rausch C,Proby C, Leigh IM, zur Hausen H,
de Villiers EM. (1994) Specific types of human papillomavirus
found in benign proliferations and carcinomas of the skin in
immunosuppressed patients, Cancer Res. 54 4610–4613.
ShamaninV, zur HausenH, LavergneD, ProbyCM, LeighIM, NeumannC,
HammH, GoosM, HausteinUF, JungEG, PlewigG, WolffH, de
VilliersEM. (1996) Human papillomavirus infections in non-
melanoma skin cancers from renal transplant recipients and
no-nimmunosuppressed patients, J. Natl. Cancer Inst. 88 802–811.
87

ShepardCW, Finelli.L, AlterMJ,(2005) Global epidemiology of
hepatitis C virus infection, Lancet, Infect. Dis. 5 558–567.
Shimoyama M. (1991) Diagnostic criteria and classification of
clinical subtypes of adult T-cell leukaemia-lymphoma.A
report from the Lymphoma Study Group (1984-1987).Br J
Haematol;79(3):428-437.
Sinha-Datta U,Horikawa I, Michishita E. (2004) Transcriptional
activation of hTERT through the NF-kappaB pathway in
HTLV-I-transformed cells. Blood;104(8):2523-2531.
Smith MR, Greene WC.(1991) Functional analyses of the type I
human T-cell leukemia virus tax gene product. Trans Assoc Am
Physicians.;104:78–91. [PubMed]
Suzuki T, Hirai H, Murakami T, Yoshida M. (1995) Tax protein of
HTLV-1 destabilizes the complexes of NF-kappaB and IkappaBa and
induces nuclear translocation of NF-kappaB for transcriptional
activation. Oncogene10:1199–1207.
Tanaka A, Takahashi C, Yamaoka S, Nosaka T, Maki M, Hatanaka M.
(1990) Oncogenic transformation by the tax gene of human T-
88

cell leukemia virus type I in vitro. ProcNatl Acad Sci U S
A;87:1071–5. [PMC free article] [PubMed]
Tanaka Y, Yoshida A, Tozawa H. (1991) Production of a recombinant
human T-cell leukemia virus type-I trans-activator (tax1)
antigen and its utilization for generation of monoclonal
antibodies against various epitopes on the tax1 antigen.Int J
Cancer;48(4):623-630.
Tanko NM, Echejoh GO, Manasseh NA, Mandong MB, Uba AF.
(2009)Paediatric solid tumours in Nigerian children:
A changing pattern? Afr J Paediatr Surg;6:7- 10.[PUBMED]
Taylor GP, Matsuoka M.(2005) Natural history of adult T-cell
leukemia/lymphoma and approaches to
therapy.Oncogene;24(39):6047-6057
U.S. Food and Drug Administration.(2009) News Release: FDA
Approves New Indication for Gardasil to Prevent Genital Warts
in Men and Boys. October 16, 2009.
[http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm187
003.htm]
89

Uchiyama T, Yodoi J, Sagawa K. (1977)Adult T-cell leukemia:
clinical and hematologic features of 16 cases 5209.
Blood;50(3):481-492.
Ukaeje CJ, Ogbu O, Inyama PU, Anyanwu GI, Njoku IO.(2005)
Prevalence of hepatitis B surface antigen among blood donors
and human immunodeficiency virus infected patients in Jos,
Nigeria. Mom do instOswaldo Cruz; 100: 13-6.
Verdonck K, Gonzalez E, Van Dooren S, Vandamme A-M, Vanham G,
Gotuzzo E.(2007) Human T-lymphotropic virus 1: recent
knowledge about an ancient infection. LancetInfectDis.;7:266–81.
DOI: 10.1016/S1473-3099(07)70081-6
Waldmann TA, Goldman CK, Bongiovanni KF. (1988) Therapy of
patients with human T-cell lymphotrophic virus I-induced adult
T-cell leukemia with anti-Tac, a monoclonal antibody to the
receptor for interleukin-2. Blood;72(5):1805-1816.
Wallin, M., M. Ekström, H. Garoff.(2004). Isomerization of the
intersubunitdisulphide-bond in Env controls retrovirus
fusion. EMBO J.23: 54–65.CrossRefMedline
90

WellsSI, FrancisDA,KarpovaAY,DowhanickJJ, BensonJD, HowleyPM.
(2000) Papillomavirus E2 induces senescence in HPV-
positive cells via pRB- and p21(CIP)- dependent pathways, Embo.
J. 19 5762–5771.
WhiteAE, LivanosEM, TlstyTD (1994), Differential disruption of
genomic integrity and cell cycle regulation in normal human
fibroblasts by the HPV oncoproteins, Genes Dev.8 666–677.
White JD, Zaknoen SL, Kastensportes C.(1995) Infectious
complications and immunodeficiency in patients with human
T-cell lymphotropic virus I-associated adult T-cell
leukemia/lymphoma. Cancer;75:1598-1607
Wong-StaalF, GalloRC.(1985) Human T-lymphotropic retroviruses,
Nature317 395–403.
XuXN, Screaton GR, Gotch FM.(1997) Evasion of cytotoxic T
lymphocyte (CTL) responses by nef-dependent induction of Fas
ligand (CD95L) expression on simian immunodeficiency virus-
infected cells. J Exp Med;186(1):7-16
91

Yamaguchi K, Nishimura H, Kohrogi H.1983 A proposal for
smoldering adult T-cell leukemia: a clinicopathologic study of
five cases. Blood;62(4):758-766.
Yamaguchi K, Takatsuki K. (1993) Adult T cell leukaemia-lymphoma.
Clin Haematol; 6(4):899-915.
ZongJC, Ciufo DM, Alcendor DJ.(1999) High-level variability in
the ORF-K1 membrane protein gene at the left end of the
Kaposi's sarcoma-associated herpesvirus genome defines four
major virus subtypes and multiple variants or clades in different
human populations. J Virol;73(5):4156-4170.
Zong JC, Metroka C, Reitz MS. (1997) Strain variability among
Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8)
genomes: evidence that a large cohort of United States AIDS
patients may have been infected by a single common isolate.
Journal of Virology;71(3):2505-2511.
Cupic, M, Lazarevic, I, Kuljić-Kapulica, N. (2005)Oncogenic
viruses and their role in tumor formation. Srp.
Arh.Celok.Lek.,133: 384-387 (article in Serbian with an abstract in
English).
92

Moscow, JA, Cowan, KH.(2007) Biology of cancer. In: Goldman, L.,
Ausiello, D., Eds. Cecil Medicine. 23rd edn., Saunders Elsevier,
Philadelphia.; 1340-1348.
Vats, TS.,Emami, A.(1993) Oncogenes: present status. Indian
Journal ofPediatrics.,;60: 193- 201.
Bell, JC.(1988) Oncogenes:Cancer Letters; 40: 1-5.
Pierotti, MA, Frattini, M, Sozzi, G, Croce, C.M.(2010):
Oncogenes. In: Hong, W.K, Bast, R.C., Hait, W.N, Kufe, D.W,
Pollock, R.E, Weichselbaum, R.R, Holland, J.F, Frei, E, Eds.
Holland- Frei Cancer Medicine. 8th edn., People’s Medical
Publishing House, Shelton, Connecticut. pp68-85.
Cullen, BR. (1992) Mechanism of action of regulatory proteins
encoded by complex retroviruses. Microbiology.Molcular
BiologyReview.,;56: 375-394.
Weiss, RA. (1996) Retrovirus classification and cell
interactions. J. Antimicrob. Chemother.,; 37: 1-11.
Elima, J, Maria, PC,Ogunbiyi, O, Oga, E, Fabowale T,Igbinoba
F,Osubor G,Otu T, Kumai H,Koechlin A,Osinubi P,Dakum P,Blattner
93

W,Adebamowo CA. Cancer incidence in Nigeria: A report from
population-based cancer registries. Cancer Epidemiology 36
pp271- 278, October 2012
94




















