
Dynamic supply chain coordination games with repeated bargaining

Microbes and Infection 10 (2008) 716e725www.elsevier.com/locate/micinf
Original article
Molecular characterization of ribonucleoproteic antigens containingrepeated amino acid sequences from Trypanosoma cruzi
Fabiano S. Pais a, Wanderson D. DaRocha a, Ramon M. Almeida a, Sophie Y. Leclercq a,Marcus L. Penido a, Stenio P. Fragoso b, Daniella C. Bartholomeu c,
Ricardo T. Gazzinelli a, Santuza M.R. Teixeira a,*
a Departamento de Bioquımica e Imunologia, ICB, Universidade Federal de Minas Gerais, UFMG,
Avenida Antonio Carlos, 6627, Pampulha, Belo Horizonte, MG, 31270-901, Brazilb Instituto de Biologia Molecular do Parana. Rua, Professor Algacyr Munhoz Mader 3775, Cidade Industrial 81350-010 e Curitiba, PR, Brazil
c Departamento de Parasitologia, ICB, Universidade Federal de Minas Gerais, UFMG, Avenida Antonio Carlos,
6627, Pampulha, Belo Horizonte, MG, 31270-901, Brazil
Received 4 January 2008; accepted 14 March 2008
Available online 29 March 2008
Abstract
Trypanosoma cruzi expresses several proteins containing antigenic amino acid repeats. Here we characterized TcRpL7a and TcRBP28, whichcarry similar repeat motifs and share homology to the eukaryotic L7a ribosomal protein and to a Trypanosoma brucei RNA binding protein,respectively. Analyses of the full length and truncated recombinant TcRpL7a showed that the humoral response of patients with Chagas diseaseis directed towards its repetitive domain. Sequence analyses of distinct copies of TcRpL7a genes present in the genome of six T. cruzi strainsindicate that the number of repeats is higher in proteins from T. cruzi II than T. cruzi I strains. A serum panel of 59 T. cruzi infected patientsshowed that 73% reacted with TcRpL7a, 71% reacted with TcRBP28 and 80% reacted with 1:1 mixture of both antigens. Synthetic peptidesharboring the TcRpL7a repeat motif reacted with 46% of the serum samples. Antibodies raised against both antigens identified equivalentamounts of the native proteins in all three stages of the parasite life cycle. Analyses of subcellular fractions indicated that TcRBP28 is presentin the cytoplasm whereas TcRpL7a co-fractionates with polysomes. Confirming their predicted cellular localization, GFP fusions showed that,whereas GFP::TcRBP28 localizes in the cytoplasm, GFP::TcRpL7a accumulates in the nucleus, where ribosome biogenesis occurs.� 2008 Elsevier Masson SAS. All rights reserved.
Keywords: Trypanosoma cruzi; Antigens; Amino acid repeats
1. Introduction
Infection by the protozoan Trypanosoma cruzi, the etiolog-ical agent of American trypanosomiasis or Chagas disease, isa major health problem in Latin America. WHO estimates aninfection prevalence of 12 million, with 3.0 million symptom-atic cases in 15 countries and a large contingent of nonsymp-tomatic infected individuals (http://www.who.int/tdr/dw/chagas2003.htm). Therefore, there is a need to continue
* Corresponding author. Tel.: þ55 31 3409 2665; fax þ55 31 3409 2614.
E-mail address: [email protected] (S.M.R. Teixeira).
1286-4579/$ - see front matter � 2008 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.micinf.2008.03.005
screening blood donors in endemic and non-endemic areassince migrations potentially compromise blood supplies. Rou-tinely, Chagas disease is diagnosed through serological testssuch as enzyme-linked immunosorbent assay (ELISA), indi-rect immunofluorescence (IIF) and indirect hemagglutination(IHA) with protein extracts prepared from epimastigotes, theinsect stage of the parasite life cycle [1]. However, the useof total parasite protein extracts results in a significant numberof false positives due to cross reactivity with antigens fromother related parasites such as Leishmania. Thus, the searchfor T. cruzi antigens that can be employed as specific targetsfor immune diagnosis of Chagas disease has been a majorgoal pursued by several laboratories. In the past decade several

717F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
recombinant antigens have been tested and a few of them havebeen incorporated in commercially available diagnostic kits.Although the use of recombinant antigens increases specific-ity, in order to achieve appropriated sensitivity, a panel withdifferent recombinant proteins is necessary. The combinationof two or more recombinant antigens has been employed todevelop novel immunodiagnostic assays that resulted in highersensitivity and specificity [2,3]. Chimeric recombinant pro-teins generated by the fusion of sequences from two antigensor four distinct peptides also proved to be highly efficient forthe diagnostic of Chagas disease [4,5].
In the majority of the work described above, the parasite re-combinant antigens were identified by immunoscreening ofcDNA libraries derived from epimastigote and trypomastigotestages of the parasite [6]. However, because of its intracellularlocation, amastigote antigens may be more relevant to thestudy of chronic chagasic pathology and more likely to repre-sent useful targets for serodiagnosis and vaccine development[7]. Using the immunoscreening approach, we isolated severalantigens derived from an amastigote cDNA library thatstrongly react with sera from chronic Chagas disease patients.Sequence analyses of positive clones indicate that ribonucleo-proteins and proteins containing tandem amino acids repeatsare among the most prominent antigens expressed in T. cruziamastigotes [8]. Several proteins containing repeats havebeen described as targets of the immune response duringT. cruzi infection, among them, members of the trans-sialidasefamily, the cytoplasmic repetitive antigen (CRA), the flagellarrepetitive antigen (FRA) and the trypomastigote surface pro-tein B13 [2,9,10]. Antigens containing repetitive motifs havealso been identified in other protozoan parasites such as thePlasmodium vivax circumsporozoite protein (CSP) and mero-zoite surface protein-2 (MSP-2) [11] and the Leishmaniaamastigote surface protein A2 [12].
Here we characterized two T. cruzi antigens containingsimilar Ala-Lys-Pro-rich repetitive motifs at their N-terminusand showing homology to the eukaryotic L7a ribosomal pro-tein [8] and to Trypanosoma brucei RNA binding proteins[13]. Our goal was to evaluate the humoral response of Chagasdisease patients against both proteins and to provide evidencesindicative of their function in the parasite. Because the aminoacid repeats were found to be the target of the antibodies pres-ent in patients’ sera, we also evaluated the presence of repeti-tive motifs in other T. cruzi ribosomal proteins and analyzedthe repetitive domain in TcRpL7a genes present in the genomeof T. cruzi strains that belong to the two major parasitelineages.
2. Materials and methods
2.1. Human sera
Serum samples were obtained from 59 patients with Chagasdisease from the Hospital das Clınicas of the UniversidadeFederal de Minas Gerais in Belo Horizonte, Brazil, whichhave been followed up by Mauro M. Teixeira. All Chagas dis-ease patients were chronically infected with T. cruzi and came
from different areas of the state of Minas Gerais, Brazil. Thediagnosis of Chagas disease patients was based on ELISAand IIF. Nine sera from individuals infected with Leishmaniachagasi and Toxoplasma gondii were also obtained. Age-and sex-matched seronegative individuals from the same com-munity were used as negative controls.
2.2. Parasite cultures
Epimastigote forms of all strains or clones of T. cruzi weremaintained in logarithmic growth phase at 28 �C in LIT me-dium as described [14]. Tissue culture trypomastigotes andamastigotes were obtained after infecting Vero cell cultureswith stationary phase epimastigotes. Purification of amasti-gotes and trypomastigotes from infected cultures were per-formed using discontinuous metrizamide gradients asdescribed [8].
2.3. DNA extraction, amplification andsequencing of TcRpL7a genes
Total DNA was isolated from T. cruzi epimastigotes accord-ing to previously described protocols. For polymerase chainreaction (PCR) amplification of TcRpL7a, 50 ng of purifiedDNA were used as described [14] using primers based onthe TcRpL7a sequence deposited in GenBank (accession num-ber AF316150). Amplicons derived from genomic DNA ofeach strain were cloned in pCR4-TOPO (Invitrogen) and ap-proximately 10e12 clones from each strain were sequencedusing automated sequencing protocols [14]. Sequences ob-tained from the T. cruzi genome consortium containing readswith homology to L7a from the clone CL Brener were also in-cluded in our analysis (http://tigrblast.tigr.org).
2.4. Production of recombinant proteins andpolyclonal antisera
cDNA fragments corresponding to the truncated C-terminalnon-repetitive region (amino acid 88e300) of the TcRpL7aprotein were obtained from digestion of the plasmidpGEX::TcRpL7a [8] and cloned at available sites into thepGEX vector (GE HealthCare) to generate recombinant pro-tein TcRpL7aDRep with glutathione S-transferase (GST)fused at the N-terminus. Digested plasmid containing the N-terminal repetitive region (amino acid 1e87) was re-ligatedto generate recombinant protein GST::TcRpL7aRep. The com-plete TcRpL7a open reading frame was also cloned into avail-able sites in pET21a vector (Novagen), generating His-taggedproteins. All GST fusion and TcRpL7a-His proteins wereobtained from isopropylthiogalactoside induced bacterial cul-tures and purified by chromatography using glutathioneeSepharose 4B or HisTrap column (GE HealthCare). Purifiedrecombinant proteins GST::TcRBP28 and GST::TcRpL7awere used to raise polyclonal antibodies in BALB/c mice bysubcutaneous inoculation of 10 mg of the antigen with Alumin-ium hydroxide as an adjuvant. Animals were boosted twice at2 weeks intervals and bled after 6 weeks.

718 F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
2.5. Peptide synthesis
The peptide TcRep AAKTAAKPAAKSAAKPAAKP(amino acid 17e37 of TcRpL7a) was prepared on a PSSM8multispecific peptide synthesizer (Shimadzu) by solid-phasesynthesis with a scale of 30 mmol as described [15]. Peptideswere purified by high-pressure liquid chromatography in a Shi-madzu system and analyzed in a C18 Vydac column. Theiridentities were confirmed by Q-TOF Micro� equipped withan electrospray ionization source (Micromass).
2.6. Western blot assays and ELISA
Bacteria and parasite total cell lysates were prepared by ho-mogenizing washed cell pellets in PBS. Polysomal extractswere obtained as described [16]. Proteins were separated in12.5% SDSePAGE, transferred to nitrocellulose membranes,blocked with PBS containing 5% of non-fat dried milk andallowed to react with human, rabbit, mice or anti-GST mono-clonal antibodies. Immunocomplexes were detected with sec-ondary horseradish peroxidase conjugated anti-IgG antibodiesand ECL Plus (GE HealthCare). Purified recombinant proteinsor peptides were coated onto 96-well plates, blocked with 5%non-fat milk and, after antibody incubations, revealed with hy-drogen peroxide and O-phenylenediamine dihydrochloridefollowed by addition of H2SO4. A450 nm was measured ina plate ELISA reader. Cutoff values were calculated for eachplate as follows: CO¼mþ 3$SD, in which m is the absor-bance average of the negative controls and SD is the standarddeviation. For competition assay, diluted serum samples werepre-incubated overnight at 4 �C with either the TcRep or witha non-related peptide (100 mg/ml) and used in ELISA as de-scribed above.
2.7. Parasite transfections and confocalmicroscopy analyses
Table 1
Amino acid repeats in T. cruzi ribosomal proteins
Protein Repeat consensus Repeat Position
Full-length cDNAs TcAG48 and TcAG29 [8] were trans-ferred after digestion of pBluescript vectors to thepTREXnGFP T. cruzi expression vector [17], which is a deriv-ative of the pTREX vector [18] containing a GFP without thestop codon. The resulting constructs were denominatedpTREXGFP::TcRBP28 and pTREXGFP::TcRpL7a. Epimasti-gotes were transfected as described [17] using 50 mgpTREXGFP DNA and 100 mg of pTREXGFP::TcRBP28 orpTREXGFP::TcRpL7a DNA. Twenty-four hours after trans-fection, parasites were analyzed by confocal microscopy.
3. Results
description number60S ribosomal
protein
L7a AAK(P/T/S) 12 N-ter
L14 IY 7 N-ter
3.1. Repetitive domains present in T. cruziribosomal proteins
L19 AKAA(A/T)AP 21 C-ter
L23a (A/T)AAP(K/Q)K 6 N-ter
40S ribosomal
protein
S21 QAR(K/R)QQGR(N/D)(A/E)P 6 C-ter
S10 (R/E)G 15 C-ter
S2 RGG 5 N-ter
In previous studies we isolated several cDNAs encoding ri-bosomal proteins after immunoscreening an expression cDNAlibrary derived from T. cruzi amastigotes with sera from pa-tients with Chagas disease [8]. The majority of positive clones
(54%) encodes four T. cruzi ribosomal proteins (TcP0, TcP2b,TcRpL19 and TcRpL7a), two of them, TcRpL7a andTcRpL19 (originally named TcAG29 and TcAG26), contain-ing similar tandem amino acid repeats. After searching theT. cruzi genome database (www.genedb.org), we identifieda total of seven orthologs of eukaryotic ribosomal proteinsthat contain amino acid repeats (Table 1). An Ala-Lys-Prorich repeated motif was found in T. cruzi orthologs of L7a,L19 and L23a, whereas Gly-Arg rich repeats are present inS2 and S10 proteins and Ile-Tyr repeats are present in L14.T. cruzi S21 contains a fourth type of amino acid repeat. Theirpositions in the sequence are both in the N-terminal (L7a, L14,L23a and S2) and the C-terminal (L19, S21 and S10) regionsof the proteins (see Supplementary Fig. 1). Remarkably, se-quence alignments with homologous ribosomal proteinsfrom other eukaryotes including the two related trypanosoma-tids T. brucei and Leishmania major, indicate that the presenceof long amino acid repeats is much more frequent in T. cruzi(not shown).
3.2. Sequence variation in the repeat domain of TcRpL7a
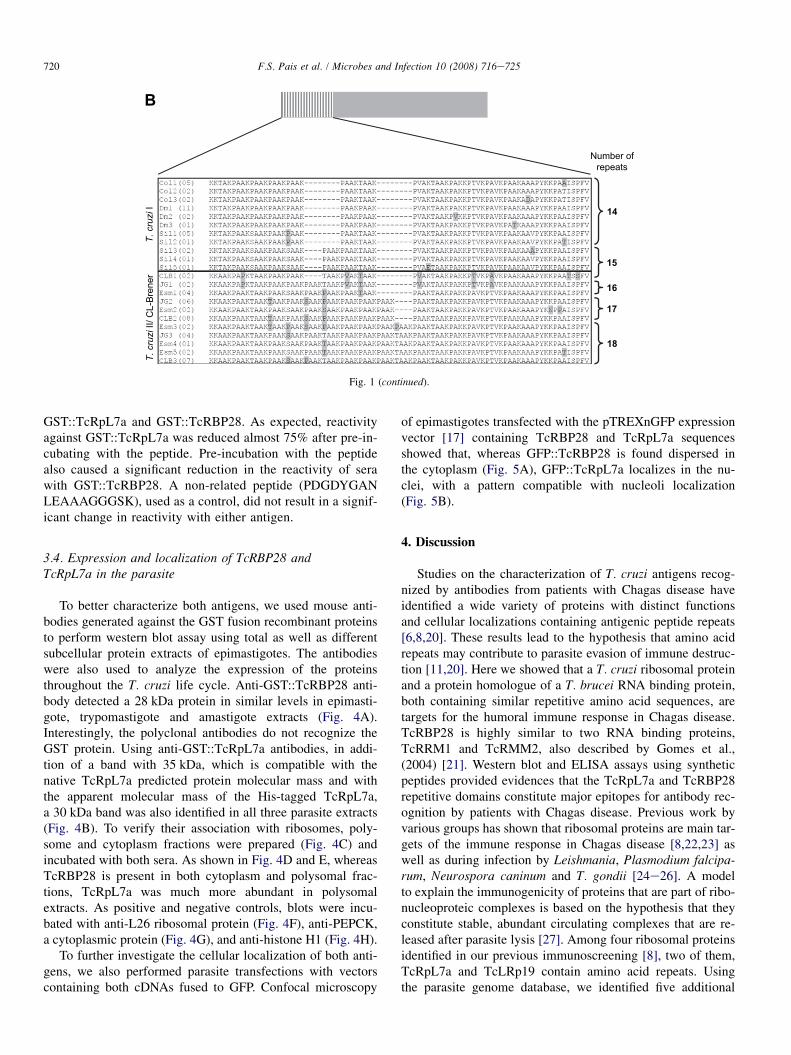
To evaluate the sequence variability and the number ofamino acid repeats present in TcRpL7a from different parasitestrains, we analyzed PCR amplified genomic sequences en-coding TcRpL7a obtained from six T. cruzi strains: Colombi-ana, Silvio X10 and Dm28c, which belong to the T. cruzi Ilineage, Esmeraldo and JG, belonging to the T. cruzi II lineageand the CL Brener clone, the genome project reference strain,known to be a hybrid strain [19]. As shown in Fig. 1A, a neigh-bor-joining tree generated with all TcRpL7a nucleotide se-quences clearly supports the division in T. cruzi I and IIlineages. Furthermore, as indicated by the presence of threemain branches containing all amplicons derived from eachT. cruzi I strain, TcRpL7a genes from T. cruzi I genomes sharemore similarity among paralogs than to their orthologs fromother strains. In contrast, almost all TcRpL7a sequencesfrom T. cruzi II strain are grouped together in one main cluster,with the exception of few divergent sequences derived from JGand CL Brener strains. The alignment showed in Fig. 1B indi-cated that the number of the AKP-rich repeat motifs is lowerin sequences derived from T. cruzi I strains, which contains14e15 repeats, than in T. cruzi II and hybrid strains, in which15e18 degenerated repeats were found.

719F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
3.3. Reactivity of TcRBP28 and TcRpL7a with sera fromT. cruzi infected patients
The reactivity of recombinant TcRBP28 (originally namedTcAG48) and TcRpL7a proteins has been previously tested inwestern blot assays using pools of sera derived from Chagasdisease patients and normal individuals [8]. To better evaluatethe immune response against the repetitive amino acid do-main, we generated truncated versions of the recombinant an-tigen containing only the repeat domain of TcRp7a and thenon-repetitive C-terminal region fused to GST. Blots contain-ing the full length recombinant protein GST::TcRpL7a(61 kDa) as well as truncated versions containing the repetitiveregion, GST::TcRpL7aRep (34 kDa), and the non-repetitivedomain, GST::TcRpL7aDRep (53 kDa) were incubated withpool of sera from chronic Chagas disease patients and healthydonors. As shown in Fig. 2A, antibodies from T. cruzi infectedpatients react exclusively with the repetitive region ofTcRpL7a. As positive and negative controls, blots were incu-bated with anti-GST antibodies (Fig. 2B) and with sera fromnon-infected individuals (Fig. 2C), respectively.
The individual immune response against GST::TcRBP28and GST::TcRpL7aRep was also evaluated by ELISA withsera from several individuals with Chagas disease, leishman-iasis and toxoplasmosis (Fig. 3A). About 71% of the T. cruziinfected patients presented reactivity above the cutoff valueagainst GST::TcRBP28 and 73% against GST::TcRpL7aRep.A small increase in reactivity (80%) was observed when bothantigens were coated together. To evaluate the efficacy of therepetitive sequence for diagnostic proposes, a peptide cover-ing part of the repetitive domain of TcRpL7a containingfive times the AAKX motif was generated. Using 100 ng ofthe synthetic TcRep peptide (AAKTAAKPAAKSAAK-PAAKP), 46% of the patients with Chagas disease presentedreactivity above the cutoff value (Fig. 3A). Since the repeti-tive domains in TcRpL7a and TcRBP28 are very similar(Fig. 3B), we decided to evaluate if the repetitive region ofTcRBP28 may also constitute a major epitope in this proteinby performing competition assays in the presence of the syn-thetic peptide (Fig. 3C). Sera from 11 T. cruzi infected pa-tients were previously incubated with 100 ng of the TcReppeptide and then incubated with ELISA plates coated with
Fig. 1. Sequences encoding TcRpL7a in different strains. After amplification
and cloning fragments containing part of the TcRpL7a genes from the genome
of six T. cruzi strains, DNA sequences were obtained from approximately 10
clones derived from each strain. (A) Nucleotide sequences were used to build
an unrooted neighbor-joining tree, in which the shape and the shade of each
node indicate the strains according to the legend. The length of the pathway
connecting each pair of nodes roughly indicates the level of dissimilarity be-
tween sequences. Bootstrap values for major branches are shown near their re-
spective fork. (B) Amino acid sequences from each strain indicated on the left
were aligned with the number in parenthesis indicating the number of identical
sequences in each case (Col, Colombiana; Dm, Dm28c; Sil, Silvio X10; Esm,
Esmeraldo; CLB, CL Brener and JG) and the number of degenerated repeat
motifs within each sequence shown on the right. The rectangle above the se-
quences indicates the localization of the repetitive domain in the protein
sequence.
B
Number ofrepeats
T. cru
zi I
T. cru
zi II/
CL-
Bren
er
14
15
16
17
18
Fig. 1 (continued).
720 F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
GST::TcRpL7a and GST::TcRBP28. As expected, reactivityagainst GST::TcRpL7a was reduced almost 75% after pre-in-cubating with the peptide. Pre-incubation with the peptidealso caused a significant reduction in the reactivity of serawith GST::TcRBP28. A non-related peptide (PDGDYGANLEAAAGGGSK), used as a control, did not result in a signif-icant change in reactivity with either antigen.
3.4. Expression and localization of TcRBP28 andTcRpL7a in the parasite
To better characterize both antigens, we used mouse anti-bodies generated against the GST fusion recombinant proteinsto perform western blot assay using total as well as differentsubcellular protein extracts of epimastigotes. The antibodieswere also used to analyze the expression of the proteinsthroughout the T. cruzi life cycle. Anti-GST::TcRBP28 anti-body detected a 28 kDa protein in similar levels in epimasti-gote, trypomastigote and amastigote extracts (Fig. 4A).Interestingly, the polyclonal antibodies do not recognize theGST protein. Using anti-GST::TcRpL7a antibodies, in addi-tion of a band with 35 kDa, which is compatible with thenative TcRpL7a predicted protein molecular mass and withthe apparent molecular mass of the His-tagged TcRpL7a,a 30 kDa band was also identified in all three parasite extracts(Fig. 4B). To verify their association with ribosomes, poly-some and cytoplasm fractions were prepared (Fig. 4C) andincubated with both sera. As shown in Fig. 4D and E, whereasTcRBP28 is present in both cytoplasm and polysomal frac-tions, TcRpL7a was much more abundant in polysomalextracts. As positive and negative controls, blots were incu-bated with anti-L26 ribosomal protein (Fig. 4F), anti-PEPCK,a cytoplasmic protein (Fig. 4G), and anti-histone H1 (Fig. 4H).
To further investigate the cellular localization of both anti-gens, we also performed parasite transfections with vectorscontaining both cDNAs fused to GFP. Confocal microscopy
of epimastigotes transfected with the pTREXnGFP expressionvector [17] containing TcRBP28 and TcRpL7a sequencesshowed that, whereas GFP::TcRBP28 is found dispersed inthe cytoplasm (Fig. 5A), GFP::TcRpL7a localizes in the nu-clei, with a pattern compatible with nucleoli localization(Fig. 5B).
4. Discussion
Studies on the characterization of T. cruzi antigens recog-nized by antibodies from patients with Chagas disease haveidentified a wide variety of proteins with distinct functionsand cellular localizations containing antigenic peptide repeats[6,8,20]. These results lead to the hypothesis that amino acidrepeats may contribute to parasite evasion of immune destruc-tion [11,20]. Here we showed that a T. cruzi ribosomal proteinand a protein homologue of a T. brucei RNA binding protein,both containing similar repetitive amino acid sequences, aretargets for the humoral immune response in Chagas disease.TcRBP28 is highly similar to two RNA binding proteins,TcRRM1 and TcRMM2, also described by Gomes et al.,(2004) [21]. Western blot and ELISA assays using syntheticpeptides provided evidences that the TcRpL7a and TcRBP28repetitive domains constitute major epitopes for antibody rec-ognition by patients with Chagas disease. Previous work byvarious groups has shown that ribosomal proteins are main tar-gets of the immune response in Chagas disease [8,22,23] aswell as during infection by Leishmania, Plasmodium falcipa-rum, Neurospora caninum and T. gondii [24e26]. A modelto explain the immunogenicity of proteins that are part of ribo-nucleoproteic complexes is based on the hypothesis that theyconstitute stable, abundant circulating complexes that are re-leased after parasite lysis [27]. Among four ribosomal proteinsidentified in our previous immunoscreening [8], two of them,TcRpL7a and TcLRp19 contain amino acid repeats. Usingthe parasite genome database, we identified five additional

30 -
43 -
67 -
20 -
1 2 3 4 1 2 3 4 1 2 3 4kDaA B C
Fig. 2. Reactivity of pools of sera from patients with Chagas disease against the recombinant T. cruzi antigen TcRpL7a. (A) Immunoblot containing purified re-
combinant proteins GST (lane 1), GST::TcRpL7aRep (lane 2), GST::TcRpL7aDRep (lane 3) and GST::TcRpL7a (lane 4) were incubated with pool of sera from 16
T. cruzi infected patients diluted 1:1.000. (B) A similar immunoblot was incubated with anti-GST antibody diluted 1:4.000 and, (C), with pool of sera from non-
infected individuals diluted 1:1.000.
721F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
T. cruzi ribosomal proteins containing repetitive sequences,which are currently being tested to verify whether they arealso targets of the immune response in Chagas disease. In spiteof the presence of its large repetitive N-terminal extension,which is exclusive for the T. cruzi protein, western blot assaysindicated that TcRpL7a is likely part of the ribosome.However, parasites transfected with GFP::TcRpL7a showeda nucleolar localization of the fusion protein, a result that isconsistent with the localization of the human homologue L7aprotein in transfected HeLa cells [28]. It is also consistentwith the presence of three nuclear localization signals presentin the T. cruzi protein (amino acids 118e121, 160e176 and304e307). Hence, it is possible that over expression ofTcRpL7a causes its accumulation in the nucleus where thebiogenesis of the ribosome takes place. Still, additional rolesof the T. cruzi ribosomal protein cannot be ruled out sincethe L12 ribosomal protein from Gonococcus spp. was alsofound associated with the cell membrane and participating incell invasion [29] and the Drosophila L22 ribosomal proteinwas found associated with histone H1 and controlling genetranscription [30]. Regarding the cellular localization ofTcRBP28, further studies will be needed to clarify its function,since the cytoplasmic localization of the GFP fusion protein isin sharp contrast with nuclear localization found for theT. brucei homologues p34 and p37. It is noteworthy that, differ-ently from the T. cruzi homologue, the T. brucei proteins containseveral characteristic bipartite nuclear localization signals [13].
Interestingly, within the T. cruzi population, the number ofrepeats in TcRpL7a shows a tendency to be higher in parasitestrains belonging to T. cruzi II lineage when compared toT. cruzi I strains. Also, their sequences are more heteroge-neous in T. cruzi II than in T. cruzi I strains. These findingsare in agreement with previous work from our group showingan increased number and variability in the repeat motifs ofTcRBP28 in T. cruzi II strains and in the hybrid CL Brenerclone [14]. The fact that few sequences from CL Brener are
grouped more closely with T. cruzi I strains is in agreementwith previous analyses showing evidences of its hybrid nature[14,19]. An expansion of amino acid repeats in antigens fromT. cruzi II strains, if it is confirmed after the analysis of greaternumber of strains, is particularly striking since various studieshave indicated a preferential association of T. cruzi II with thehuman infection in endemic areas in South America [14,31].Thus, we may ask why amino acid repeats are so widespreadin T. cruzi antigens, and, more importantly, why are they beenexpanded in T. cruzi II strains?
Studies with trans-sialidase, a unique T. cruzi enzyme thattransfers sialyl residues from host glycoconjugates to theparasite acceptor surface molecules and it is a key playerin parasite cell invasion, have provided some evidences forthe role of antigenic amino acid repeats during the T. cruziinfection. This surface enzyme has two clearly defined re-gions: the amino terminus containing the catalytic activityand a C-terminal domain known as SAPA that contains vari-able number of a 12 amino acid repeat [9]. SAPA repeatselicit a strong humoral response early in the infection, whichis followed by the induction of antibodies against the cata-lytic domain only during later phases of the infection [32].Since anti-SAPA antibodies are unable to neutralize trans-sialidase activity, it has been suggested that this immunodo-minant epitope plays a distractive role that is able to delaythe induction of a neutralizing humoral response. A secondrole for the repetitive domain in trans-sialidases may be in-volved with extending the protein half-life in the blood [33].It can be thus speculated that the presence of amino acid re-peats in T. cruzi antigens that are part of ribonucleoproteincomplexes may fulfill both roles, i.e., to generate a strong,but not necessarily effective humoral response against lysedparasites, and also to contribute to increasing the persistencein the blood of antigens acting as immunomodulators, whichmay be essential for long-term survival of the parasite in themammalian host.
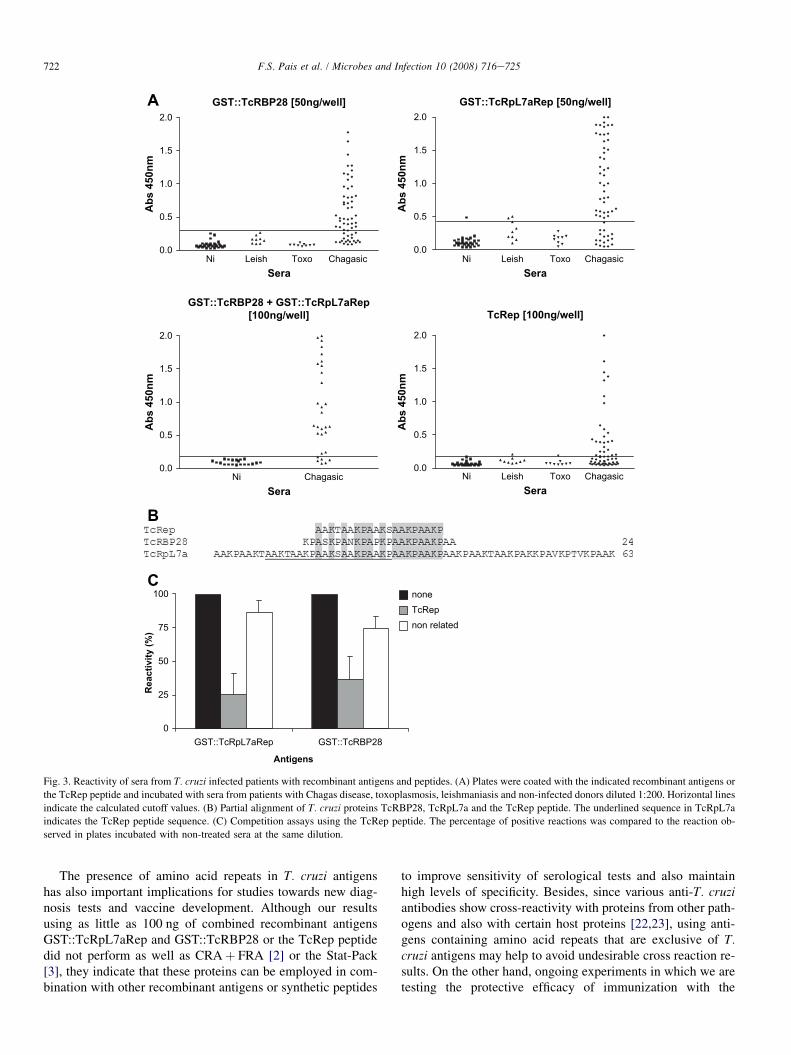
GST::TcRpL7aRep [50ng/well]
0.0
0.5
1.0
1.5
2.0
Ab
s 450n
m
GST::TcRBP28 [50ng/well]
0.0
0.5
1.0
1.5
2.0A
B
C
Ni Leish Toxo ChagasicSera
Ni Leish Toxo ChagasicSera
Ab
s 450n
m
0.0
0.5
1.0
1.5
2.0
GST::TcRBP28 + GST::TcRpL7aRep
[100ng/well] TcRep [100ng/well]
0.0
0.5
1.0
1.5
2.0
Ab
s 450n
m
Ni Chagasic Ni Leish Toxo ChagasicSeraSera
Ab
s 450n
m
0
25
50
75
100
Reactivity (%
)
GST::TcRpL7aRep GST::TcRBP28
Antigens
noneTcRepnon related
Fig. 3. Reactivity of sera from T. cruzi infected patients with recombinant antigens and peptides. (A) Plates were coated with the indicated recombinant antigens or
the TcRep peptide and incubated with sera from patients with Chagas disease, toxoplasmosis, leishmaniasis and non-infected donors diluted 1:200. Horizontal lines
indicate the calculated cutoff values. (B) Partial alignment of T. cruzi proteins TcRBP28, TcRpL7a and the TcRep peptide. The underlined sequence in TcRpL7a
indicates the TcRep peptide sequence. (C) Competition assays using the TcRep peptide. The percentage of positive reactions was compared to the reaction ob-
served in plates incubated with non-treated sera at the same dilution.
722 F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
The presence of amino acid repeats in T. cruzi antigenshas also important implications for studies towards new diag-nosis tests and vaccine development. Although our resultsusing as little as 100 ng of combined recombinant antigensGST::TcRpL7aRep and GST::TcRBP28 or the TcRep peptidedid not perform as well as CRAþ FRA [2] or the Stat-Pack[3], they indicate that these proteins can be employed in com-bination with other recombinant antigens or synthetic peptides
to improve sensitivity of serological tests and also maintainhigh levels of specificity. Besides, since various anti-T. cruziantibodies show cross-reactivity with proteins from other path-ogens and also with certain host proteins [22,23], using anti-gens containing amino acid repeats that are exclusive of T.cruzi antigens may help to avoid undesirable cross reaction re-sults. On the other hand, ongoing experiments in which we aretesting the protective efficacy of immunization with the
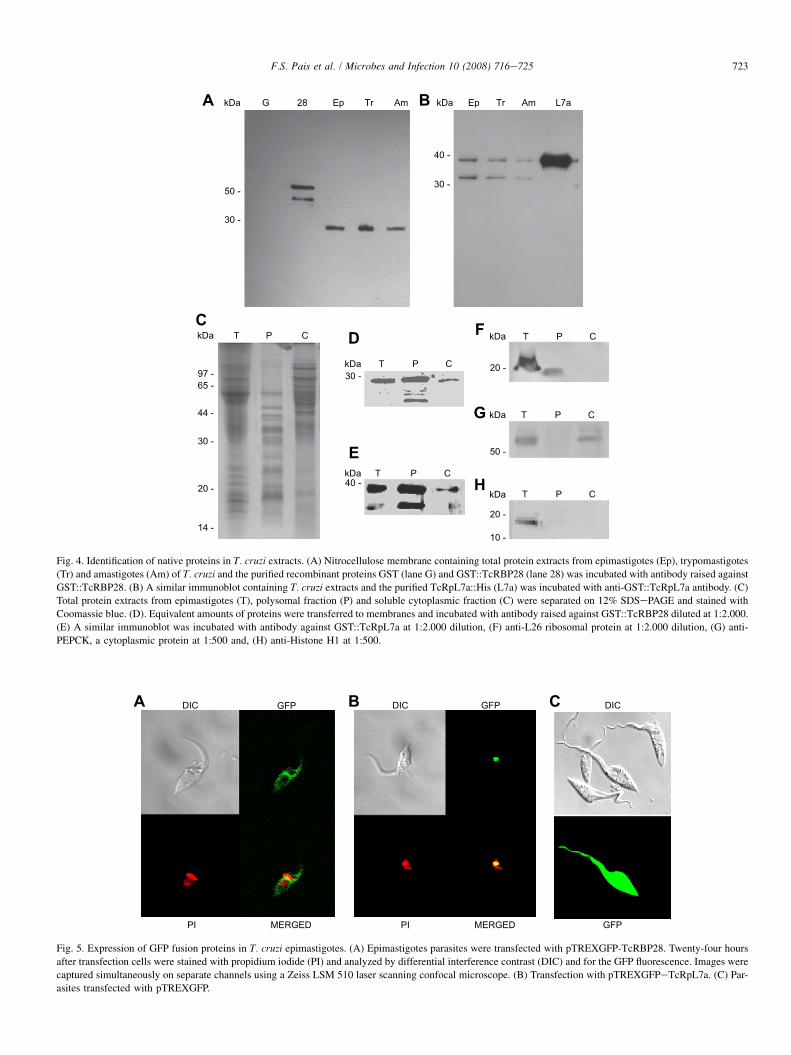
50 -
30 -
40 -
30 -
kDakDaA
C
D
E
F
G
H
BAmTrEp28G Ep Tr L7aAm
kDa
30 -
20 -
14 -
65 -
44 -
97 -
50 -
kDa
20 -
10 -
kDa40 -kDa
kDa
20 -30 -kDa
T P C
T P C
T P C
T P C
T P C
T P C
Fig. 4. Identification of native proteins in T. cruzi extracts. (A) Nitrocellulose membrane containing total protein extracts from epimastigotes (Ep), trypomastigotes
(Tr) and amastigotes (Am) of T. cruzi and the purified recombinant proteins GST (lane G) and GST::TcRBP28 (lane 28) was incubated with antibody raised against
GST::TcRBP28. (B) A similar immunoblot containing T. cruzi extracts and the purified TcRpL7a::His (L7a) was incubated with anti-GST::TcRpL7a antibody. (C)
Total protein extracts from epimastigotes (T), polysomal fraction (P) and soluble cytoplasmic fraction (C) were separated on 12% SDSePAGE and stained with
Coomassie blue. (D). Equivalent amounts of proteins were transferred to membranes and incubated with antibody raised against GST::TcRBP28 diluted at 1:2.000.
(E) A similar immunoblot was incubated with antibody against GST::TcRpL7a at 1:2.000 dilution, (F) anti-L26 ribosomal protein at 1:2.000 dilution, (G) anti-
PEPCK, a cytoplasmic protein at 1:500 and, (H) anti-Histone H1 at 1:500.
DICA B CGFP DIC
GFP
GFP
PI MERGEDPI MERGED
DIC
Fig. 5. Expression of GFP fusion proteins in T. cruzi epimastigotes. (A) Epimastigotes parasites were transfected with pTREXGFP-TcRBP28. Twenty-four hours
after transfection cells were stained with propidium iodide (PI) and analyzed by differential interference contrast (DIC) and for the GFP fluorescence. Images were
captured simultaneously on separate channels using a Zeiss LSM 510 laser scanning confocal microscope. (B) Transfection with pTREXGFPeTcRpL7a. (C) Par-
asites transfected with pTREXGFP.
723F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725

724 F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
repetitive and the non-repetitive domains of TcRpL7a mayalso help determining whether or not the repeated sequencesshould be included or rather avoided in the design of new an-tigenic targets for vaccine development.
Acknowledgements
The authors are grateful to Mauro M. Teixeira from UFMGfor providing the serum samples, Fernando Souza for technicalsupport and Sergio Schenkman from UNIFESP for providingantibodies anti-histone H1. This work was supported by grantsfrom CAPES, CNPq and FAPEMIG, Brazil and by Interna-tional Research Scholars Grant from the Howard HughesMedical Institute.
Appendix A. Supplementary information
Supplementary material can be found, in the online version,at doi: 10.1016/j.micinf.2008.03.005
References
[1] Z. Brener, A. Andrade, M. Barral-Netto, Trypanosoma cruzi e a doenca
de Chagas, 2a ed. Guanabara Koogan, Rio de Janeiro, 2000.
[2] M.A. Krieger, E. Almeida, W. Oelemann, J.J. Lafaille, J.B. Pereira,
H. Krieger, M.R. Carvalho, S. Goldenberg, Use of recombinant antigens
for the accurate immunodiagnosis of Chagas’ disease, Am. J. Trop. Med.
Hyg. 46 (1992) 427e434.
[3] C. Ponce, E. Ponce, E. Vinelli, A. Montoya, V. de Aguilar, A. Gonzalez,
B. Zingales, R. Rangel-Aldao, M.J. Levin, J. Esfandiari, E.S. Umezawa,
A.O. Luquetti, J.F. da Silveira, Validation of a rapid and reliable test for
diagnosis of Chagas’ disease by detection of Trypanosoma cruzi-specific
antibodies in blood of donors and patients in Central America, J. Clin.
Microbiol. 43 (2005) 5065e5068.
[4] S. Aguirre, A.M. Silber, M.F. Brito, M.E. Ribone, C.M. Lagier,
I.S. Marcipar, Design, construction, and evaluation of a specific chimeric
antigen to diagnose chagasic infection, J. Clin. Microbiol. 10 (2006)
3768e3774.
[5] A.W. Ferreira, Z.R. Belem, E.A. Lemos, S.G. Reed, A. Campos-Neto,
Enzyme-linked immunosorbent assay for serological diagnosis of
Chagas’ disease employing a Trypanosoma cruzi recombinant antigen
that consists of four different peptides, J. Clin. Microbiol. 39 (2001)
4390e4395.
[6] J.F. da Silveira, E.S. Umezawa, A.O. Luquetti, Chagas disease: recombi-
nant Trypanosoma cruzi antigens for serological diagnosis, Trends
Parasitol. 17 (2001) 286e291.
[7] M.M. Rodrigues, S.B. Boscardin, J.R. Vasconcelos, M.I. Hiyane,
G. Salay, I.S. Soares, Importance of CD8 T cell-mediated immune re-
sponse during intracellular parasitic infections and its implications for
the development of effective vaccines, An. Acad. Bras. Cienc. 75
(2003) 443e468.
[8] W.D. DaRocha, D.C. Bartholomeu, C.D. Macedo, M.F. Horta, E. Cunha-
Neto, J.E. Donelson, S.M.R. Teixeira, Characterization of cDNA clones
encoding ribonucleoprotein antigens expressed in Trypanosoma cruzi
amastigotes, Parasitol. Res. 88 (2002) 292e300.
[9] A.J. Parodi, G.D. Pollevick, M. Mautner, A. Buschiazzo, D.O. Sanchez,
A.C. Frasch, Identification of the gene(s) coding for the trans-sialidase of
Trypanosoma cruzi, EMBO J. 11 (1992) 1705e1710.
[10] A. Gruber, B. Zingales, Trypanosoma cruzi: characterization of two re-
combinant antigens with potential application in the diagnosis of Chagas’
disease, Exp. Parasitol. 76 (1993) 1e12.
[11] D.J. Kemp, R.L. Coppel, R.F. Anders, Repetitive proteins and genes of
malaria, Ann. Rev. Microbiol. 41 (1987) 181e208.
[12] H. Charest, G. Matlashewski, Developmental gene expression in
Leishmania donovani: differential cloning and analysis of an amasti-
gote-stage-specific gene, Mol. Cell. Biol. 14 (1994) 2975e2984.
[13] J. Zhang, W. Ruyechan, N. Williams, Developmental regulation of two
nuclear RNA binding proteins, p34 and p37, from Trypanosoma brucei,Mol. Biochem. Parasitol. 92 (1998) 79e88.
[14] G.C. Cerqueira, D.C. Bartholomeu, W.D. DaRocha, L. Hou,
D.M. Freitas-Silva, C.R. Machado, N.M. El-Sayed, S.M.R. Teixeira, Se-
quence diversity and evolution of multigene families in Trypanosoma
cruzi, Mol. Biochem. Parasitol 157 (2008) 65e72.
[15] B.C. de Alencar, A.F. Araujo, M.L. Penido, R.T. Gazzinelli,
M.M. Rodrigues, Cross-priming of long lived protective CD8þ T cells
against Trypanosoma cruzi infection: importance of a TLR9 agonist
and CD4þ T cells, Vaccine 25 (2007) 6018e6027.
[16] A.R. Avila, S.F. Yamada-Ogatta, V. da Silva Monteiro, M.A. Krieger,
C.V. Nakamura, W. de Souza, S. Goldenberg, Cloning and characteriza-
tion of the metacyclogenin gene, which is specifically expressed during
Trypanosoma cruzi metacyclogenesis, Mol. Biochem. Parasitol. 117
(2001) 169e177.
[17] W.D. DaRocha, R.A. Silva, D.C. Bartholomeu, S.F. Pires, J.M. Freitas,
A.M. Macedo, M.P. Vazquez, M.J. Levin, S.M.R. Teixeira, Expression
of exogenous genes in Trypanosoma cruzi: improving vectors and
electroporation protocols, Parasitol. Res. 92 (2004) 113e120.
[18] M. Vazquez, M.J. Levin, Functional analysis of the intergenic regions
of TcP2beta gene loci allowed the formulation of an improved Trypano-
soma cruzi expression vector, Gene 239 (1999) 217e225.
[19] N.M. El-Sayed, P.J. Myler, D.C. Bartholomeu, D. Nilsson, G. Aggarwal,
A.N. Tran, E. Ghedin, E.A. Worthey, A.L. Delcher, G. Blandin,
S.J. Westenberger, E. Caler, G.C. Cerqueira, C. Branche, B. Haas,
A. Anupama, E. Arner, L. Aslund, P. Attipoe, E. Bontempi,
F. Bringaud, P. Burton, E. Cadag, D.A. Campbell, M. Carrington,
J. Crabtree, H. Darban, J.F. da Silveira, P. de Jong, K. Edwards,
P.T. Englund, G. Fazelina, T. Feldblyum, M. Ferella, A.C. Frasch,
K. Gull, D. Horn, L. Hou, Y. Huang, E. Kindlund, M. Klingbeil,
S. Kluge, H. Koo, D. Lacerda, M.J. Levin, H. Lorenzi, T. Louie,
C.R. Machado, R. McCulloch, A. McKenna, Y. Mizuno, J.C. Mottram,
S. Nelson, S. Ochaya, K. Osoegawa, G. Pai, M. Parsons, M. Pentony,
U. Pettersson, M. Pop, J.L. Ramirez, J. Rinta, L. Robertson,
S.L. Salzberg, D.O. Sanchez, A. Seyler, R. Sharma, J. Shetty,
A.J. Simpson, E. Sisk, M.T. Tammi, R. Tarleton, S. Teixeira, S. Van
Aken, C. Vogt, P.N. Ward, B. Wickstead, J. Wortman, O. White,
C.M. Fraser, K.D. Stuart, B. Andersson, The genome sequence of Trypa-nosoma cruzi, etiologic agent of Chagas disease, Science 309 (2005)
409e415.
[20] D.E. Hoft, K.S. Kim, K. Otsu, D. Moser, W.J. Yost, J.H. Blumin,
J.E. Donelson, L.V. Kirchhoff, Trypanosoma cruzi expresses diverse
repetitive protein antigens, Infec. Immun. 57 (1989) 1959e1967.
[21] G. Gomes, T. Peter Urmenyi, E. Rondinelli, N. Williams, R. Silva,
TcRRMs and Tcp28 genes are intercalated and differentially expressed
in Trypanosoma cruzi life cycle, Biochem. Biophys. Res. Commun. 24
(2004) 985e992.
[22] M.J. Levin, E. Mesri, R. Benarous, G. Levitus, A. Schijman, P. Levy-
Yeyati, P. Chiale, A. Ruiz, A. Kahn, M. Rosembaum, H.N. Torres,
E.L. Segura, Identification of major Trypanosoma cruzi antigenic
determinants in chronic Chagas’ heart disease, Am. J. Trop. Med. Hyg.
41 (1989) 530e539.
[23] Y.A. Skeiky, D.R. Benson, J.A. Guderian, P.R. Sleath, M. Parsons,
S.G. Reed, Trypanosoma cruzi acidic ribosomal P protein gene family.
Novel P proteins encoding unusual cross-reactive epitopes, J. Immunol.
151 (1993) 5504e5515.
[24] S. Iborra, D.R. Abanades, N. Parody, J. Carrion, R.M. Risue~no,
M.A. Pineda, P. Bonay, C. Alonso, M. Soto, The immunodominant T
helper 2 (Th2) response elicited in BALB/c mice by the Leishmania
LiP2a and LiP2b acidic ribosomal proteins cannot be reverted by strong
Th1 inducers, Clin. Exp. Immunol. 50 (2007) 375e385.
[25] S. Singh, S. Chatterjee, R. Sohoni, S. Badakere, S. Sharma, Sera from
lupus patients inhibit growth of Plasmodium falciparum in culture,
Autoimmunity 33 (2001) 253e263.

725F.S. Pais et al. / Microbes and Infection 10 (2008) 716e725
[26] H. Zhang, E.G. Lee, M. Liao, M.K. Compaore, G. Zhang, O. Kawase,
K. Fujisaki, C. Sugimoto, Y. Nishikawa, X. Xuan, Identification of ribo-
somal phosphoprotein P0 of Neospora caninum as a potential common
vaccine candidate for the control of both neosporosis and toxoplasmosis,
Mol. Biochem. Parasitol. 153 (2007) 141e148.
[27] J.M. Requena, C. Alonso, M. Souto, Evolutionarily conserved proteins as
proeminent immongens during Leishmania infections, Parasitol. Today
16 (2000) 246e250.
[28] G. Russo, G. Ricciardelli, C. Pietropaolo, Different domains cooperate to
target the human ribosomal L7a protein to the nucleus and to the nucle-
oli, J. Biol. Chem. 272 (1997) 5229e5235.
[29] J.M. Spence, V.L. Clark, Role of ribosomal protein L12 in gonococcal
invasion of Hec1B cells, Infect. Immun. 68 (2000) 5002e
5010.
[30] J.Q. Ni, L.P. Liu, D. Hess, J. Rietdorf, F.L. Sun, Drosophila ribosomal
proteins are associated with linker histone H1 and suppress gene tran-
scription, Genes Dev. 20 (2006) 1959e1973.
[31] B. Zingales, R.P. Souto, R.H. Mangia, C.V. Lisboa, D.A. Campbell,
J.R. Coura, A. Jansen, O. Fernandes, Molecular epidemiology of Amer-
ican trypanosomiasis in Brazil based on dimorphisms of rRNA and mini-
exon gene sequences, Int. J. Parasitol. 28 (1998) 105e112.
[32] M.S. Leguizamon, O.E. Campetella, S.M. Gonzalez Cappa, A.C. Frasch,
Mice infected with Trypanosoma cruzi produce antibodies against the en-
zymatic domain of trans-sialidase that inhibit its activity, Infect. Immun.
62 (1994) 3441e3446.
[33] C.A. Buscaglia, J. Alfonso, O. Campetella, A.C. Frasch, Tandem amino
acids repeats from Trypanosoma cruzi shed antigens increase the half-life
of proteins in blood, Blood 93 (1999) 2025e2032.
Copyright © 2022 FDOKUMEN




















