
Analogue encoding of physicochemical properties of proteins in their cognate messenger RNAs

Molecular characterization of a putative heat shock proteincognate gene in Rhynchosciara americana
Alexandre de Andrade & Fabio Siviero & Paula Rezende-Teixeira &
Roberto Vicente Santelli & Glaucia Maria Machado-Santelli
Received: 18 July 2009 /Accepted: 2 September 2009 /Published online: 19 September 2009# Springer Science + Business Media B.V. 2009
Abstract An hsc70 homologue gene (Rahsc70) ofthe diptera Rhynchosciara americana was isolatedand characterized. We were able to determine themRNA sequence from an EST of salivary glandcDNA library, and a Rahsc70 cDNA cassette wasused as a probe to isolate the genomic region from agenomic library. The mRNA expression of this geneparallels the 2B puff expansion, suggesting its
involvement in protein processing, since this larvalperiod corresponds to a high synthetic activity period.During heat shock stress conditions, hsc70 expressiondecreased. In situ hybridization of polytene chromo-somes showed that the Rahsc70 gene is located nearthe C3 DNA puff. The cellular localization of Hsc70protein showed this protein in the cytoplasm and inthe nucleus.
Keywords Rhynchosciara americana . polytenechromosome . DNA puff . heat shock protein .
development
AbbreviationsHsp heat shock proteinHsc heat shock cognateRahsc Rhynchosciara americana heat shock
cognateB2 DNA puff at region 2 of chromosome BC3 DNA puff at region 3 of chromosome CC8 DNA puff at region 8 of chromosome CIR beginning of cocoon spinningEcR ecdysone receptorUSP ultraspiracle protein
Introduction
Some chaperones involved in the folding process areknown as heat shock proteins (Hsp) (Craig et al.
Chromosome Research (2009) 17:935–945DOI 10.1007/s10577-009-9081-7
Responsible Editor: Hans-Joachim Lipps.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10577-009-9081-7) containssupplementary material, which is available to authorized users.
A. de Andrade : F. Siviero : P. Rezende-Teixeira :R. V. SantelliInstituto de Química, Departamento de Bioquímica,Universidade de São Paulo,Av. Prof. Lineu Prestes 748,São Paulo, SP, 05508-900, Brazil
A. de Andrade : F. Siviero : P. Rezende-Teixeira :G. M. Machado-SantelliInstituto de Ciências Biomédicas,Departamento de Biologia Celular e Desenvolvimento,Universidade de São Paulo,Av. Prof. Lineu Prestes 1524,São Paulo, SP, 05508-900, Brazil
G. M. Machado-Santelli (*)Departamento de Biologia Celular e do Desenvolvimento,Universidade de São Paulo,Av. Prof. Lineu Prestes 1524,São Paulo, SP, 05508-900, Brazile-mail: [email protected]

1983; Rybczynski and Gilbert 2000). These proteinsare among the most conserved proteins in differentbiological kingdoms; they utilize ATP-dependentcycles to lead conformational changes in their targetproteins (Beinssinger and Buchner 1998). There aremany chaperone systems, and they are also known asHsp families; among them, Hsp70, Hsp90, and Hsp40illustrate the major conserved groups. Members of theHsp70 family participate in a variety of cellularprotein folding processes, including folding of newlysynthesized proteins in the cytosol, translocation ofproteins into organelles, assembly and disassembly ofprotein complexes, and control of the biologicalactivity of folded regulatory proteins (Rudiger et al.2001; Soti and Csermely 2002). The cellularresponse to high-temperature stress appears to besimilar in different organisms. Shortly after a heatshock stress, the synthesis of the Hsp increase andtranslation of normal proteins is down regulated. InDrosophila, for example, the time required to detectthis new scenario is around 20 min (DiDomenico etal. 1982).
Besides the stress-induced proteins, the foldingprotein machinery also includes chaperones that areconstitutively expressed. These proteins are calledheat shock cognates proteins (Hsc). In Drosophilamelanogaster, seven different constitutive genesbelonging to the hsp70 family have been described.The seven hsc70 genes are denominated hsc70-1through hsc70-7, and are expressed during normalgrowth. The most highly expressed is the hsc70-4gene that has an ORF of 1,953 bp and a single intronlocated in the 5′UTR region (Perkins et al. 1990).Also, hsc70-4 homologues were found in otherinsects, including Manduca sexta (Rybczynski andGilbert 2000), Chironomus tentans, and Chironomusyoshimatsui (Karouna-Renier et al. 2003).
For over five decades, Rhynchosciara americanahas been utilized as a model for the study of thepolytenization/amplification phenomenon that occursin its chromosomes from several tissues during larvaldevelopment (Breuer and Pavan 1955; Machado-Santelli and Basile 1973; Glover et al. 1982). Thesalivary glands show the largest polytene chromo-somes, and during larval development, some structuralchanges can be easily noticed. The chemical changesoccurring during the swelling of specific regions ofthe polytene chromosomes are related to the synthe-sis of RNA and/or synthesis and accumulation of
DNA. This process is rigorously regulated duringdevelopment (Pavan 1965). These structural special-izations are called puffs, and among them, thosecalled DNA puffs were targets of intensive study. InR. americana, three important DNA puffs werecharacterized: B2 (Santelli et al. 2004), C3 (Santelliet al. 1991; Penalva et al. 1997), and C8 (Frydman etal. 1993). The proteins coded by the amplifiedsequences in these DNA puffs are usually related tothe construction of the communal cocoon, where themetamorphosis from the larval stage to adult animalstakes place (Winter et al. 1977a, b; Orkretic et al.1977). Although this organism has been studied fora long time, a limited amount of data regarding itsdevelopment, molecular mechanisms, and DNAsequences is available. The most recent findingsabout R. americana development and molecularmechanisms were published by Rezende-Teixeira etal. (2008a, b, 2009), Gouveia et al. (2008), Madalenaet al. (2007), and Rossato et al. (2007).
Recently, our group sequenced a cDNA library fromsalivary glands of R. americana fourth-instar larvae.With this sequencing, some transcripts involved inheat shock response itself or with its pathway wereidentified: hsc70, hsp83, hip, hop, dnaj, trap1 andprolyl isomerase (Siviero et al. 2006). The period ofconstruction of this library is characterized by anintense protein synthesis and secretion. The proteinfolding and transport among the cellular compart-ments are processes usually involving chaperone andco-chaperone activity. The hsc70, hip, and hsp83messages may be required, especially in this larvalage, due to their commitment with the cocoonconstruction.
The folding process is an essential step for proteinsto reach their functional conformation. Interestingly,as was reported in D. melanogaster, the proteinsHsc70 and Hsp90, together with its co-chaperonesHip, Hop, and p23, form the heterodimer ecdysonereceptor EcR/USP (EcR, Ecdysone Receptor; USP,Ultraspiracle) (Arbeitman and Hogness 2000). Thehormone ecdysone plays an important role in theregulation of insect development. Amabis and Cabral(1970) and Stocker and Pavan (1974) described theactivity of this hormone in the genus Rhynchosciara.
In the present study, we report the characterizationof a heat shock cognate gene present in the R.americana genome, hereafter called Rahsc70. Thestudy of this gene may unveil new molecular aspects
936 A. de Andrade et al.

yet to be shown in R. americana. To address somequestions, we report a Rahsc70 molecular depiction,its normal expression profile, and its expressionduring heat shock stress situations.
Methods
Animals and salivary glands
Larvae were collected in Ubatuba, Sao Paulo, Brazil.Salivary glands were stored using the conditionsestablished by Lara et al. (1965). Larvae at theappropriate age were washed and dissected inphosphate-buffered saline (PBSA, 137 mM NaCl,2.7 mM KCl, 10 mM Na2HPO4, and 2 mM KH2PO4,for 1-L solution after pH 7.4 adjusted with HCl), andsalivary glands were fixed in ethanol 70% for 10 minand stored in ethanol/glycerol 1:1 at −20°C.
Rhynchosciara americana genomic phage libraryscreening
Utilizing as a probe the clone revealed by the R.americana cDNA sequencing, a screening of a R.americana genomic library constructed in phagelambda DASH II was made. The protocol followedwas a modification of CsCl lambda DNA purificationdescribed in Santelli and Navarro-Cattapan (2000)and in Andrade et al. (2005).
Sequencing and assembly
The R. americana cDNA sequencing reactions wereperformed in a polymerase chain reaction (PCR)GeneAmp 9700 (Perkin Elmer), using a BigDye(PerkinElmer) terminator (96°C, 1 min; 35 cycles of96°C, 20 s; 50°C, 1 min; 60°C, 4 min). The sampleswere read in an ABI-Prism 377 (PerkinElmer)automatic DNA sequencer, and the sequences wereanalyzed using the Sequence Analysis software(Perkin Elmer). The electropherograms were analyzedwith a Linux (Debian) workstation, where the set ofsoftware Phred, Phrap Crossmatch, and Consed 11was used to assemble the sequences (Ewing andGreen 1998; Ewing et al. 1998). Phylogeneticanalyses were made utilizing the CLUSTALX 1.83program (Thompson et al. 1997). The accessionnumbers of the Hsc70 proteins sequences used to
construct the phylogenetic tree and the alignment are:R. americana DQ402054 , D. melanogasterAAA28627, Ceratitis capitata AAC23392, Anophelesgambiae XP320971, M. sexta AAF09496, Bombyxmori BAB92074, Trichoplusia ni ABH09732, Cras-sostrea gigas AA041703, Urechis caupo AAA74394,Rivulus marmoratus AAF71255, Xiphophorus mac-ulatus BAB72169, Oncorhynchus mykiss P08108,Danio rerio AAH63228, Ambystoma mexicanumAAK31583, Gallus gallus NP990334, Homo sapiensAAK17898, Bos taurus NP776770, Rattus novergicusNP077327, Mus musculus AAA37869, Cricetulusgriseus P19378, and Artemia franciscana AAL27404.
RNA preparation
Rhynchosciara americana salivary glands were trans-ferred to a digestion solution [Tris (pH 7.8), NaCl20 mM, EDTA 20 mM, SDS 0.5%, and 300 μg/mL ofproteinase K (Invitrogen)], maintained 1–2 h at 60°C,extracted with phenol (1:1), and phenol/chloroform (1:1).Following precipitation with 0.3MNaAc and 2× volumeof ethanol, the RNA was treated with DNAseI (Sigma)and quantified in a NanoDrop 1000 Spectrophotometer.
Larvae heat shock
Larvae were transferred to preheated glass plates(containing moistened paper) and placed in a 30°Cincubator. Animals were kept in those conditions for30 min or for 1 h. Afterwards, animals were dissectedand the salivary glands were removed. RNA wasextracted from the salivary glands.
Real-time reverse transcriptase-PCR
Real time reverse transcriptase (RT)-PCR was per-formed using SuperScript III Platinum SYBR GreenOne-Step RT-PCR kit in triplicate. The RNA wasprepared as described above and the primers usedwere RaHSC70-QL (AACTAATTAAAGATGGCCGCAC) and RaHSC70-QR (CATCGCCAATTAATCGTTCTG). The normalization was done againsttotal RNA (Bustin 2000, 2002). This was donebecause the expression of a reference gene (GAPDH,actin and tubulin) present in the cDNA library of R.americana salivary glands varied greatly during thelarval development, as described in Xenopus (Sindelkaet al. 2006).
Characterization of a Hsp cognate gene in Rhynchosciara americana 937

Immunofluorescence
Salivary gland chromosomes were treated for fixationwith ethanol/acetic acid (3:1) for 30 s, formaldehydesolution (3.7%)/acetic acid (45%) for 3 min washed inPBSA and stored in glycerol/PBSA (2:1) at −20°C.The glands were squashed and fixed in a microscopeslide treated with polylysine and coverslips wereremoved after freezing the slides on dry ice, and theslides were washed 2× for 5 min in PBSA, incubatedfor 10 min in PBSA/0.5%Triton X100 solution, andwashed in PBSA 1× for 5 min. To block nonspecificbinding on glands, the slides were incubated inPBSA/1%BSA solution for 30 min, and after that,they were washed in PBSA 1× for 5 min, then 15 μLof the antibody Hsc/Hsp70 (Sigma, BRM-22) wasadded in a 1/100 dilution, covered with ParafilmR andstored in a moist chamber overnight at 4°C. Theslides were washed 2× for 5 min with PBSA, 15 μLof anti-mouse FITC antibody was added in a 1/50dilution, and they were covered with ParafilmR andincubated for 1.5 h at room temperature in a moistchamber protected from light. The slides were againwashed 3× for 5 min with PBSA and 10 μL RNAse(10 mg/mL) was added onto the slides. The slideswere then covered with ParafilmR for 30 min, washed3× for 5 min with PBSA, and after that, 10 μL ofpropidium iodide was added. Slides were thencovered with ParafilmR and incubated for 20 min.After washing the slides 1× for 5 min with PBSA,10 μL VectaShieldR was added and the slides werecovered with coverslips. Cytological preparationswere analyzed in a laser scanning confocal micro-scope (LSM-510—Zeiss).
In situ hybridization
Salivary gland chromosomes were fixed with ethanol/acetic acid solution (3:1) and then were squashed in45% acetic acid. Coverslips were removed afterfreezing the slides on dry ice. The preparations werewashed in PBSA, denatured for 3–5 min in NaOH0.07 M, washed 3× with alcohol, and dried. Theprobe was labeled with dUTP-digoxigenin by randomprimers (Boehringer-Mannhein). The hybridizationwas performed overnight in a moist chamber at 58°C.The labeling was revealed by antibody anti-digoxigenin-fluorescein and the chromosomes werecounterstained with propidium iodide. Cytological
preparations were analyzed in a laser scanning confo-cal microscope (LSM-510—Zeiss).
Results
Characterization of Rahsc70 gene
Screening of a R. americana genomic library usingRahsc70 cDNA as a probe identified a Rahsc70genomic recombinant containing the complete genesequence (accession number: DQ402054). It spans4,450 bp and presents one single open reading framecoding a protein of 656 amino acids (Fig. 1 andFig. 1SM—Supplemental Material). A canonicalTATA box signal was found at positions 752–759, inthe 5′UTR. An intron 796-bp long is present fromnucleotides 902 to 1698, immediately preceding thestart codon. It is in a similar arrangement as thatobserved in the Drosophila hsc70-4 gene (Perkins etal. 1990). By comparing the genomic DNA withcDNA sequences, it was found that the Rahsc70intron presents conventional splicing sites (GT-AG).The Rahsc70 intron is shorter than that found in theDrosophila hsc70-4 gene, which is 1.9-kb long.However, the Rahsc70 intron is about the same sizeas those present in the 5′UTR region of C. tentans andC. yoshimatsui (Karouna-Renier et al. 2003). Intronsare usually found in the constitutive heat shock genes,in contrast to what happens with heat shock induciblegenes, which, in the majority of cases, have no intronsin their structure. The usual absence of introns inheat-inducible genes is related to its rapid expressionduring stress situations. In eukaryotic DNA, thetranscription start site is typically found 23–30
Fig. 1 Schematic structure of RaHsc70 gene in R. americana.Above are showed the two exons, the RaHsc70 ORF coded656aa is indicated by dark gray box. The canonical TATA box(TATAA) and the possible polyadenylation signal are indicatedin the scheme
938 A. de Andrade et al.
nucleotides downstream of the consensus TATAAAA(Verbeet et al. 1984; Thomm and Wich 1988). In theRahsc70 sequence, there is a putative transcriptionstart site (CAATTCA) in the sequence from nucleo-tides 789 to 795, 29 nucleotides downstream of thecanonical TATA box. This transcription start site isidentical to the counterpart present in the sequence ofhsp70 gene in D. melanogaster (Ingolia et al. 1980).A canonical polyadenylation signal is located at thenucleotide 3868, and a polyadenylation site is locatedat nucleotide 3888. The 3′UTR sequence of hsp genesusually has regulatory AU-rich elements (AUUUA)that may be responsible for stabilizing the transcriptsduring heat shock (Lindquist and Petersen 1990). TheRahsc70 gene, similar to Chironomus and Drosophilaorthologues, has these AU-rich elements in its 3′UTRthat can be related to stabilization of its transcript atnormal temperatures (Rubenstein and Lyons 2001).
The RaHsc70 amino acid consensus sequenceshowed putative sites for phosphorylation by caseinkinase II, tyrosine kinase, protein kinase C, andcAMP/cGMP-protein kinase dependent. The threeHsp70 family signatures were found in RaHsc70:Hsp70_1 at positions 12–19 (IDCGTTYS), Hsp70_2at positions 200–213 (IFDLGGGTFDVSIL), andHsp70_3 at positions 337–351 (IVLVGGSTRIPKVQK) PROSITE (http://www.expasy.ch/prosite).These signatures have been found in other membersof the Hsc70 family (Karouna-Renier et al. 2003). Abipartite nuclear localization signal was identified atpositions 249–265 and 269–276 (KRKHKRDLTTNKRSLRR // KRSLRRLRTACERAKRT), suggestingthe RaHsc70 protein’s participation in nuclear process-
es. This nuclear localization signal is also present in thehuman Hsc70 protein homologue (Demand et al. 1998)in a similar position as the signal found in R.americana. The sequence tag present in the carboxi-terminal region of the RaHsc70 protein (EEVD) is amotif widely conserved in all eukaryotic cytosol/nuclear Hsc70s and Hsp70s, which was previouslyidentified in two species of Chironomus (Karouna-Renier et al. 2003). This region plays an importantrole in the association of co-factors that regulate thechaperone activity of Hsp70 (Tsukahara and Maru2004).
The conceptual translation of the Rahsc70 cDNAshowed strong similarity with some domains from theProDom data bank (http://proteintoulouse.inra.fr/prodom/current/html/home.php): heat-shock ATPbinding, heat shockATP-binding endoplasmic reticulumcognate, and kinesin-like ATP-binding motor domains.The ATP-binding domain found in the RaHsc70 proteinis localized at the N-terminal region and is believed toact in the control of chaperone-substrate affinity.Another domain found was an endoplasmic reticulumATP-binding domain, which could be related to theHsc70 chaperones role in the folding and translocationinto the endoplasmic reticulum.
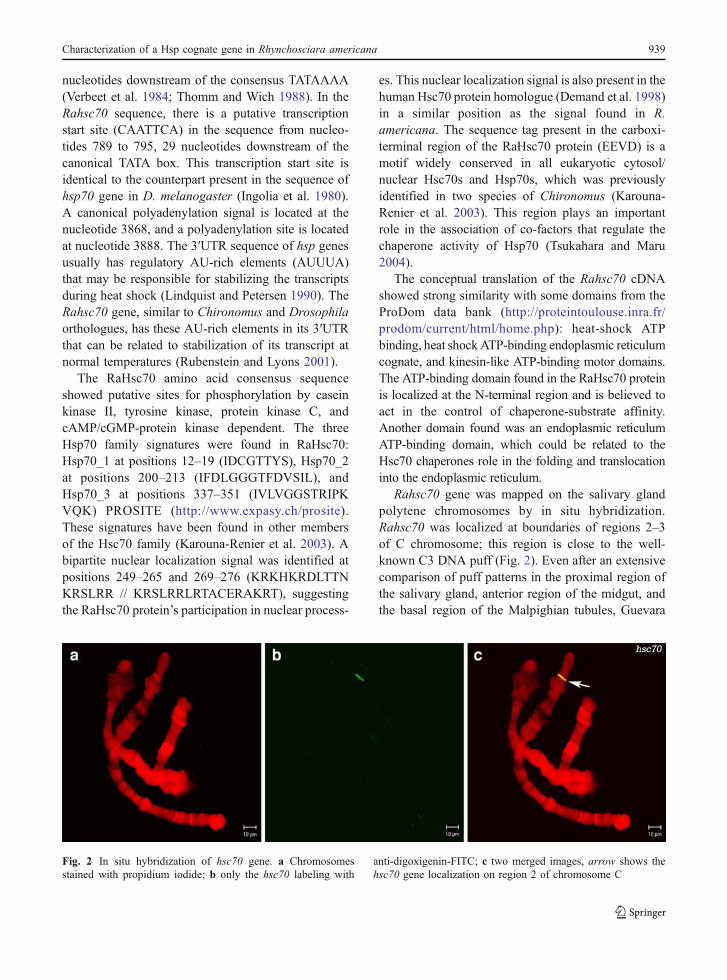
Rahsc70 gene was mapped on the salivary glandpolytene chromosomes by in situ hybridization.Rahsc70 was localized at boundaries of regions 2–3of C chromosome; this region is close to the well-known C3 DNA puff (Fig. 2). Even after an extensivecomparison of puff patterns in the proximal region ofthe salivary gland, anterior region of the midgut, andthe basal region of the Malpighian tubules, Guevara
Fig. 2 In situ hybridization of hsc70 gene. a Chromosomesstained with propidium iodide; b only the hsc70 labeling with
anti-digoxigenin-FITC; c two merged images, arrow shows thehsc70 gene localization on region 2 of chromosome C
Characterization of a Hsp cognate gene in Rhynchosciara americana 939

and Basile (1973) could not find any transcription/amplification puff at this locus.
RaHsc70 phylogenetic analysis
To investigate the similarity of RaHsc70 with otherspecies, a phylogenetic tree was generated. To do
this, the conceptual translation of the Rahsc70cDNA was utilized. Figure 3 shows a phylogenetictree comparing the protein sequence of RaHsc70with the Hsc70 proteins from other species. TheHsc70 protein of R. americana, as expected, ishighly similar to the Hsc70 proteins from otherinsects.
Fig. 3 Phylogenetic treeconstructed utilizingRaHsc70 protein sequenceand its homologues
940 A. de Andrade et al.
An alignment of the RaHsc70 amino acid sequencewith other species showed a high degree of sequenceidentity with orthologs in A. gambiae, C. capitata, M.sexta, and B. mori, ranging between 88.2–90.8%amino acid identity and 95.7–96.5% conservativereplacements. The D. melanogaster Hsc70-4 aminoacid sequence appears to be the closest heat shockcognate protein to Rahsc70, with 90.9% and 96.6%
amino acid identity and conservative replacements,respectively (Figure 2SM—Supplemental Material).
Rahsc70 gene expression profile in normal and stressconditions
The analysis of the Rahsc70 expression pattern duringR. americana development could reveal whether this
a
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
2P IR 2B 2B - 7h 2B - 24h 3C
Periods of fourth instar of larval development
Rel
ativ
e E
xpre
ssio
n R
aHS
C70
b
c(a) (b) (c)
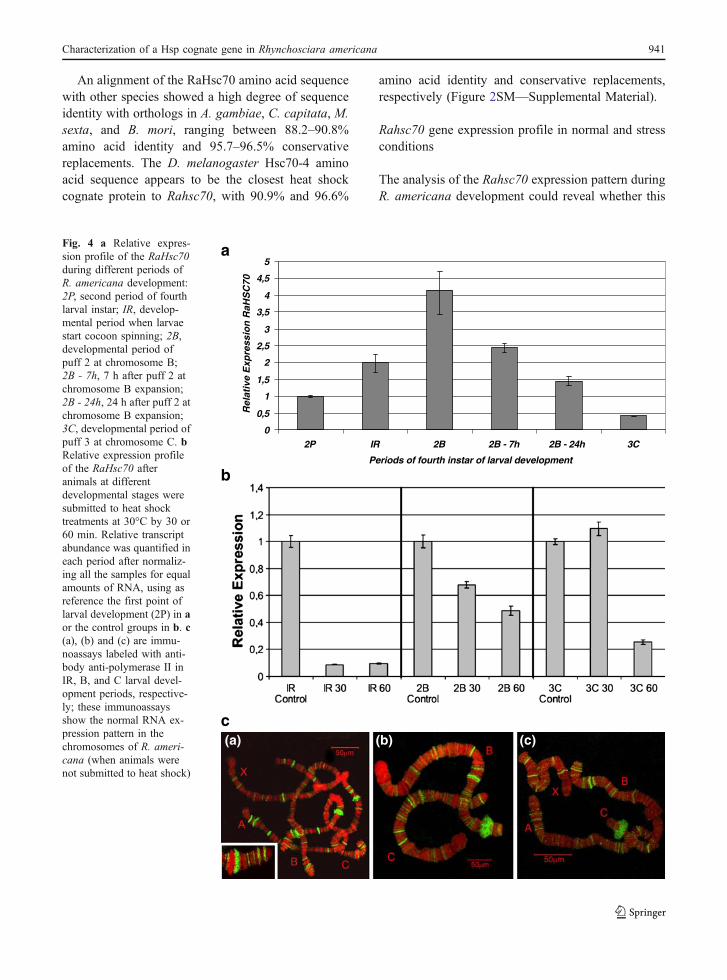
Fig. 4 a Relative expres-sion profile of the RaHsc70during different periods ofR. americana development:2P, second period of fourthlarval instar; IR, develop-mental period when larvaestart cocoon spinning; 2B,developmental period ofpuff 2 at chromosome B;2B - 7h, 7 h after puff 2 atchromosome B expansion;2B - 24h, 24 h after puff 2 atchromosome B expansion;3C, developmental period ofpuff 3 at chromosome C. bRelative expression profileof the RaHsc70 afteranimals at differentdevelopmental stages weresubmitted to heat shocktreatments at 30°C by 30 or60 min. Relative transcriptabundance was quantified ineach period after normaliz-ing all the samples for equalamounts of RNA, using asreference the first point oflarval development (2P) in aor the control groups in b. c(a), (b) and (c) are immu-noassays labeled with anti-body anti-polymerase II inIR, B, and C larval devel-opment periods, respective-ly; these immunoassaysshow the normal RNA ex-pression pattern in thechromosomes of R. ameri-cana (when animals werenot submitted to heat shock)
Characterization of a Hsp cognate gene in Rhynchosciara americana 941

gene is constitutively or variably expressed. Theexpression profile of Rahsc70 was determined byreal-time RT-PCR in different developmental periodsof larval fourth-instar (Fig. 4a). Notably, Rahsc70expression rose progressively from the second periodlarval (2P) until B2 puff expansion (2B), beingreduced at the moment of C3 puff expansion (3C).High levels of transcription and protein synthesischaracterize some periods of larval fourth-instar, likewhen the cocoon is been elaborated, that correspondto the third (IR) to fourth periods (2B puff). Therelative expression observed in Rahsc70 suggests apossible participation in the folding process ofpeptides at the period that correspond to cocoonformation. The northern blot patterns (data notshown) and the quantitative PCR profile in R.americana led us to speculate that the Rahsc70transcript, similar to what happens with Sesamianonagrioides, would be under strict developmentalregulation (Gkouvitsas et al. 2009).
After determining the pattern of expression ofRahsc70 at normal conditions, it would be relevant tostudy how its expression is affected during a stresssituation. To establish the behavior of Rahsc70 duringheat shock and confirm that it is a cognate homo-logue, its expression profile was determined throughreal-time RT-PCR during heat shocks at differentdevelopmental stages (Fig. 4b). In the initial steps ofthe cocoon spinning, the Rahsc70 expression leveldecreased due to heat shock. During the fourth andfifth periods (2B and 3C cytological markers, respec-tively), the heat shock effects in the gene expressionare less intense. The differential expression ofRahsc70 during shock conditions in the study periodsshowed that the regulation of the response to hightemperature can have a fine control throughout larvaldevelopment in the salivary gland. This control couldbe related to physiological or metabolic status of thelarvae.
The expression level of Rahsc70 during a heatshock stress indicates that it is not a heat shock-inducible gene, as hsp70. In fact, a closer look at theRahsc70 profile reveals that the decrease of itsexpression levels is more severe in the period justbefore the opening of the puffs (period IR). Conse-quently, as Rahsc70 function is related to proteinfolding, it may be important for the organism tomaintain relatively high Rahsc70 levels in order tosustain the folding processes at work in the cell. It is
also possible that this is part of the heat stressresponse in R. americana, a mechanism mediated bythe Hsp.
The normal RNA transcription patterns in salivarychromosomes are also shown in Fig. 4c. Immunoflu-orescence of chromosomes from larvae in initial stepsof cocoon spinning, puffs 2B and 3C developmentalages, using antibody anti-polymerase II, showedpositive bands all over the chromosomes in youngerlarvae. Labeling is less intense in older larvaechromosomes.
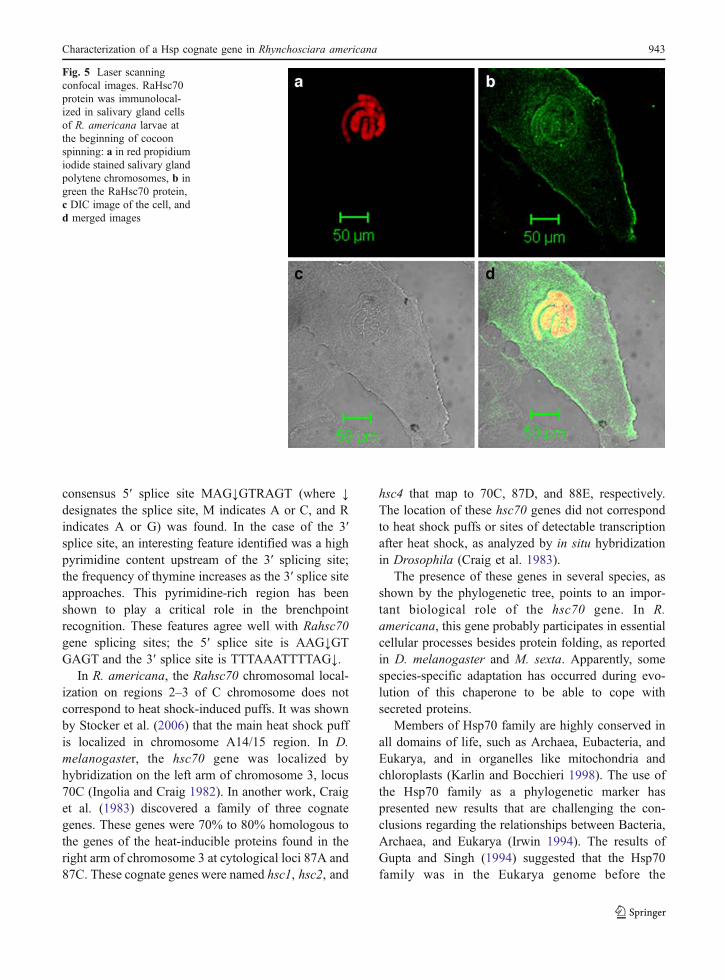
RaHsc70 protein is localized in the cytoplasmand nucleus
Immunofluorescence with anti-hsp70 antibodyshowed positive labeling over the cytoplasm andnucleus of salivary gland cells of R. americana at thebeginning of cocoon spinning (Fig. 5). The antibodyutilized recognizes both the Hsc70 and Hsp70proteins. As in the non-stressed situation, Hsp70 isvirtually absent and the signal can be interpretedmostly as the RaHsc70 protein. In R. americana, thepattern of Hsc70 distribution was granular.
Discussion
The cognate gene member of the Hsp70 familyidentified in R. americana is an orthologue of themajor constitutive gene previously identified in D.melanogaster (hsc70-4) by Craig et al. (1983).Rahsc70 gene expression could be under R. americanadevelopmental regulation. Its expression was detectedduring the beginning of cocoon spinning until theexpansion of puff 3C, the larval age characterized by ahuge level of protein synthesis. It is possible tohypothesize that, at this time, Rahsc70 gene expres-sion rises due to the need for the protein coded by thisgene in folding processes or that the cellular pathwayin which hsc70 participates has been turned on.
As described by Yost et al. (1990), stress caninterfere with mRNA processing. In the report ofMount et al. (1992), a database of 209 Drosophilaintrons was extracted from Genbank and examined tocharacterize features that might serve as signals formessenger RNA splicing. A comparison of thenucleotide frequency distribution of the Drosophila5′ splice sites showed considerable similarity. A
942 A. de Andrade et al.
consensus 5′ splice site MAG↓GTRAGT (where ↓designates the splice site, M indicates A or C, and Rindicates A or G) was found. In the case of the 3′splice site, an interesting feature identified was a highpyrimidine content upstream of the 3′ splicing site;the frequency of thymine increases as the 3′ splice siteapproaches. This pyrimidine-rich region has beenshown to play a critical role in the brenchpointrecognition. These features agree well with Rahsc70gene splicing sites; the 5′ splice site is AAG↓GTGAGT and the 3′ splice site is TTTAAATTTTAG↓.
In R. americana, the Rahsc70 chromosomal local-ization on regions 2–3 of C chromosome does notcorrespond to heat shock-induced puffs. It was shownby Stocker et al. (2006) that the main heat shock puffis localized in chromosome A14/15 region. In D.melanogaster, the hsc70 gene was localized byhybridization on the left arm of chromosome 3, locus70C (Ingolia and Craig 1982). In another work, Craiget al. (1983) discovered a family of three cognategenes. These genes were 70% to 80% homologous tothe genes of the heat-inducible proteins found in theright arm of chromosome 3 at cytological loci 87A and87C. These cognate genes were named hsc1, hsc2, and
hsc4 that map to 70C, 87D, and 88E, respectively.The location of these hsc70 genes did not correspondto heat shock puffs or sites of detectable transcriptionafter heat shock, as analyzed by in situ hybridizationin Drosophila (Craig et al. 1983).
The presence of these genes in several species, asshown by the phylogenetic tree, points to an impor-tant biological role of the hsc70 gene. In R.americana, this gene probably participates in essentialcellular processes besides protein folding, as reportedin D. melanogaster and M. sexta. Apparently, somespecies-specific adaptation has occurred during evo-lution of this chaperone to be able to cope withsecreted proteins.
Members of Hsp70 family are highly conserved inall domains of life, such as Archaea, Eubacteria, andEukarya, and in organelles like mitochondria andchloroplasts (Karlin and Bocchieri 1998). The use ofthe Hsp70 family as a phylogenetic marker haspresented new results that are challenging the con-clusions regarding the relationships between Bacteria,Archaea, and Eukarya (Irwin 1994). The results ofGupta and Singh (1994) suggested that the Hsp70family was in the Eukarya genome before the
Fig. 5 Laser scanningconfocal images. RaHsc70protein was immunolocal-ized in salivary gland cellsof R. americana larvae atthe beginning of cocoonspinning: a in red propidiumiodide stained salivary glandpolytene chromosomes, b ingreen the RaHsc70 protein,c DIC image of the cell, andd merged images
Characterization of a Hsp cognate gene in Rhynchosciara americana 943

endosymbiotic event that led to the acquisition ofmitochondria and chloroplasts. It is possible that theHsp70 family arose from a common ancestor andspread later on. An analysis of 29 Hsp70 genes fromeukaryotic and prokaryotic species demonstrated anextreme conservation of it (Boorstein et al. 1994).
The localization of RaHsc70 indicates its associa-tion with cell physiology. The presence of RaHsc70 inboth compartments, cytoplasm and nucleus, indicatesparticipation of RaHsc70 not only in protein folding,assembly, and disassembly of protein complexes, butalso in translocation across membranes. It alsosuggests the participation of RaHsc70 in nuclearprocesses. The Hsc70 pattern of distribution did notcorrespond to the fibrous meshwork found in Dro-sophila by Palter et al. 1986. The localization of thisprotein in the nucleus is consistent with the nuclearsignal localization found in the RaHsc70 proteinsequence, which may be necessary for RaHsc70-selective translocation into the nucleus. The presenceof Hsp/Hsc70 in the nucleus may be required tomaintain the correct folding of the protein machinerypresent there. It was demonstrated by Mandell andFeldherr (1990) that these proteins shuttle between thenucleus and the cytoplasm. The kind of cellularprocesses in which RaHsc70 is participating is stillunclear, and it deserves further investigation.
Acknowledgments We thank Dr. Hamza El Dorry forsequencing support and Roberto Cabado Modia Jr. for helpingwith image acquisition. Also, we would like to thank thefinancial support and fellowship from CAPES, CNPq, andFAPESP.
References
Amabis JM, Cabral D (1970) RNA and DNA puffs in polytenechromosomes of Rhynchosciara: inhibition by extirpationof prothorax. Science 169:692–694
Andrade A, Rezende-Teixeira P, Siviero F, Santelli RV (2005)A shortcut in phage screening technique. Gen Mol Biol1:150–151
Arbeitman MN, Hogness DS (2000) Molecular chaperonesactive the Drosophila ecdysone receptor, an RXR hetero-dimer. Cell 101:67–77
Beinssinger M, Buchner J (1998) How chaperones foldproteins. Biol Chem 379:245–259
Boorstein WR, Ziegelhoffer T, Craig EA (1994) Molecularevolution of the HSP70 multigene family. J Mol Evol 38:1–17
Breuer ME, Pavan C (1955) Behavior of polytene chromo-somes of Rhynchosciara angelae at different stages oflarval development. Chromosoma 7:371–386
Bustin SA (2000) Absolute quantification of mRNA using real-time reverse transcription polymerase chain reactionassays. J Mol Endocrinol 25:169–193
Bustin SA (2002) Quantification of mRNA using real-timereverse transcription PCR (RT-PCR): trends and problems.J Mol Endocrinol 29:23–39
Craig EA, Ingolia TD, Manseau LJ (1983) Expression ofDrosophila heat shock cognate genes during heat shockand development. Dev Biol 99:418–426
Demand J, Luders J, Hohfeld J (1998) The carboxy-terminaldomain of Hsc70 provides binding sites for a distinct setof chaperone cofactors. Mol Cell Biol 18:2023–2028
DiDomenico BJ, Bugaisky GE, Lindquist S (1982) Heat shockand recovery are mediated by different translationalmechanisms. PNAS USA 79:6181–6185
Ewing B, Green P (1998) Base-calling of automated sequencertraces using Phred, 2. Error probabilities. Genome Res8:186–194
Ewing B, Hillier LD, Wendl MC, Green P (1998) Base-callingof automated sequencer traces using Phred, 1. Accuracyassessment. Genome Res 8:175–185
Frydman HM, Cadavid EO, Yokosawa J, Silva FH, Navarro-Cattapan LD, Santelli RV, Jacobs-Lorena M, GraessmannM, Graessmann A, Stocker AJ, Lara FJS (1993) Molecularcharacterization of the DNA puff C-8 gene of Rhyncho-sciara americana. J Mol Biol 233:799–803
Gkouvitsas T, Kontogiannatos D, Kourti A (2009) Cognate Hsp70gene is induced during deep larval diapause in the mothSesamia nonagrioides. Insect Mol Biol 18(2):253–264
Glover DM, Zaha A, Stocker AJ, Santelli RV, Pueyo MT,Toledo SM, Lara FJS (1982) Gene amplification inRhynchosciara salivary gland chromosomes. PNAS USA79:2947–2951
Gouveia Fde S, Gimenes F, Fiorini A, Fernandez MA (2008)Intrinsic bent DNA sites in the developmentally amplified C3–22 gene promoter of Rhynchosciara americana (Diptera:Sciaridae). Biosci Biotechnol Biochem 72(5):1190–1198
Guevara M, Basile R (1973) DNA and RNA puffs inRhynchosciara. Caryologia 26:275–295
Gupta RS, Singh B (1994) Phylogenetic analysis of 70 kDaheat shock protein sequences suggests a chimeric originfor the eukaryotic cell nucleus. Curr Biol 12:1104–1114
Ingolia TD, Craig EA (1982) Drosophila gene related to themajor heat shock-induced gene is transcribed at normaltemperatures and not induced by heat shock. PNAS USA79:525–529
Ingolia TD, Craig EA, McCarthy BJ (1980) Sequence of threecopies of the gene for the major Drosophila heat shockinduced protein and their flanking regions. Cell 21:669–679
Irwin DM (1994) Molecular evolution. Who are the parents ofeukaryotes? Curr Biol 12:1115–1117
Karlin S, Bocchieri L (1998) Heat shock protein 70 family:multiple sequence comparisons, function and evolution. JMol Evol 47:565–577
Karouna-Renier NK, Yang WJ, Rao KR (2003) Cloning andcharacterization of a 70 kDa heat shock cognate gene(HSC70) from two species of Chironomus. Insect MolBiol 12:19–26
Lara FJS, Tamaki H, Pavan C (1965) Laboratory culture ofRhynchosciara angelae. Am Nature 99:189–191
944 A. de Andrade et al.

Lindquist S, Petersen S (1990) Selective translation anddegradation of heat shock messenger RNA’s in Drosoph-ila. Enzyme 44:147–166
Machado-Santelli GM, Basile R (1973) Studies of DNA puff inRhynchosciara. Genetics 74:168
Madalena CR, Amabis JM, Stocker AJ, Gorab E (2007) Thelocalization of ribosomal DNA in Sciaridae (Diptera:Nematocera) reassessed. Chromosome Res 15(4):409–416
Mandell RB, Feldherr CM (1990) Identification of two HSP70-related Xenopus oocyte proteins that are capable ofrecycling across the nuclear envelope. J Cell Biol111:1775–1783
Mount SM, Burks C, Hertz G, Stormo GD, White O, Fields C(1992) Splicing signals in Drosophila: intron size, infor-mation content, and consensus sequences. Nucleic AcidsRes 20:4255–4262
Orkretic MC, Penoni JS, Lara FJS (1977) Messenger-like RNAsynthesis and DNA chromosomal puffs in the salivaryglands of Rhynchosciara americana. Arch BiochemBiophys 178:158–165
Palter KB, Watanabe M, Stinson L, Mahowald AP, Craig EA(1986) Expression and localization of Drosophila mela-nogaster hsp70 cognate proteins. Mol Cell Biol 6:1187–1203
Pavan C (1965) Nucleic acid metabolism in polytene chromo-somes and the problem of differentiation. BrookhavenSysp Biol 18:222–241
Penalva LO, Yokosawa J, Stocker AJ, Soares MA, GraessmannM, Orlando TC, Winter CE, Botella LM, Lara FJS (1997)Molecular characterization of the C-3 DNA puff gene ofRhynchosciara americana. Gene 2:163–172
Perkins LA, Doctor JS, Zhang K, Stinson L, Perrimon N, CraigEA (1990) Molecular and developmental characterizationof the heat shock cognate 4 gene of Drosophila mela-nogaster. Mol Cell Biol 10:3232–3238
PRODOM (2008). http://proteintoulouse.inra.fr/prodom/current/html/home.php
PROSITE (2009). PROSITE homepage. http://www.expasy.ch/prosite
Rezende-Teixeira P, Siviero F, Andrade A, Santelli RV,Machado-Santelli GM (2008a) Mariner-like elements inRhynchosciara americana (Sciaridae) genome: molecularand cytological aspects. Genetica 2:137–145
Rezende-Teixeira P, Siviero F, Brandão AS, Santelli RV,Machado-Santelli GM (2008b) Molecular characterizationof a retrotransposon in the Rhynchosciara americanagenome and its association with telomere. ChromosomeRes 5:729–742
Rezende-Teixeira P, Siviero F, da Costa Rosa M, Machado-Santelli GM (2009) The R2 mobile element of Rhyncho-sciara americana: molecular, cytological and dynamicaspects. Chromosome Res 17:455–467
Rossato RM, Madalena CR, Gorab E (2007) Unusually shorttandem repeats in the chromosome end structure of Rhyn-chosciara (Diptera: Sciaridae). Genetica 131(2):109–116
Rubenstein RC, Lyons BM (2001) Sodium 4-phenylbutyratedownregulates HSC70 expression by facilitating mRNAdegradation. Am J Physiol, Lung Cell Mol Physiol 281:L43–L51
Rüdiger S, Schneider-Mergener J, Bukau B (2001) Its substratespecificity characterizes the DnaJ co-chaperone as a
scanning factor for the DnaK chaperone. EMBO J20:1042–1050
Rybczynski R, Gilbert LI (2000) cDNA Cloning and expressionof a hormone-regulated heat shock protein (hsc70) fromthe prothoracic gland of Manduca sexta. Insect BiochemMol Biol 30:579–589
Santelli RV, Navarro-Cattapan LD (2000) A simplified CsClprotocol for lambda DNA purification: no enzymatictreatment/one phenol extraction. Gen Mol Biol 23:65–66
Santelli RV, Siviero F, Machado-Santelli GM, Lara FJS,Stocker AJ (2004) Molecular Characterization of the B2DNA Puff Gene of Rhynchosciara americana. Chromo-soma 113:167–176
Santelli RV, Machado-Santelli GM, Pueyo MT, Navarro-CattapanLD, Lara FJS (1991) Replication transcription in the course ofDNA amplification of the C3 and C8 DNA puffs ofRhynchosciara americana. Mech Dev 36:59–66
Sindelka R, Ferjentsik Z, Jonak J (2006) Developmentalexpression profiles of Xenopus laevis reference genes.Dev Dyn 235:754–758
Siviero F, Rezende-Teixeira P, Andrade A, Machado-SantelliGM, Santelli RV (2006) Analysis of expressed sequencetags from Rhynchosciara americana salivary glands.Insect Mol Biol 15:109–118
Söti C, Csermely P (2002) Chaperones and aging: role inneurodegeneration and in other civilization diseases.Neurochem Int 41:383–389
Stocker AJ, Madalena CR, Gorab E (2006) The effects oftemperature shock on transcription and replication inRhynchosciara americana (Diptera: Sciaridae). Genetica126:277–290
Stocker AJ, Pavan C (1974) The influence of the ecdysteroneon gene amplification, DNA synthesis and puff formationin the salivary glands of Rhynchosciara hollanderi.Chromosoma 45:295–319
Thomm M, Wich G (1988) An archaebacterial promoter elementfor stable RNA genes with homology to the TATA box ofhigher eukaryotes. Nucleic Acids Res 1:151–163
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, HigginsDG (1997) The CluxtalX windows interface: flexiblestrategies for multiple sequence alignment aided by qualityanalysis tolls. Nucleic Acids Res 25:4876–4882
Tsukahara F, Maru Y (2004) Identification of novel nuclearexport a nuclear localization related signals in human heatshock cognate protein 70. J Biol Chem 279:8867–8872
Verbeet MP, Klootwijk J, van Heerikhuizen H, Fontijn RD,Vreugdenhil E, Planta RJ (1984) A conserved sequenceelement is present around the transcription initiation sitefor polymerase A in Saccharomycetoideae. Nucleic AcidsRes 2:1137–1148
Winter CE, De Bianchi AG, Terra WR, Lara FJS (1977a)Relationships between newly synthesized proteins andpuff patterns in salivary glands of Rhynchosciara ameri-cana. Chromosoma 61:193–206
Winter CE, De Bianchi AG, Terra WR, Lara FJS (1977b) Thegiant DNA puffs of Rhynchosciara americana code forpolypeptides of the salivary gland secretion. J InsectPhysiol 23:453–1459
Yost HJ, Petersen RB, Lindquist S (1990) RNA Metabolism:strategies for regulation in the heat shock response. TrendsGenet 6:223–227
Characterization of a Hsp cognate gene in Rhynchosciara americana 945
Copyright © 2022 FDOKUMEN




















