
Molecular basis of the optochin-sensitive phenotype of pneumococcus: characterization of the genes...
13
Molecular Microbiology (1994) 12{4), 587-598 Molecular basis of the optochin-sensitive phenotype of pneumococcus: characterization of the genes encoding the FQ complex of the Streptococcus pneumoniae and Streptococcus oralis H"^-ATPases Asuncion Fenoll,^ Rosario Munoz,' Ernesto Garcia^* and Adela G. de la Campa^ ^Centra Nacional de Microbiologia, Instituto de Salud Carlos III. 28220 Majadahonda, Spain. ^Centro de Investigaclones Biologicas, CSIC, Velazquez 144, 28006. Madrid, Spain. Summary The gene responsible for the optochin-sensitive (Opt^) phenotype of Streptococcus pneumoniae has been characterized. Sequence comparisons indicated that the genes involved encoded the subunits of the FQ complex of an H^-ATPase. Sequence analysis and transformation experiments showed that the atpC gene Is resfionsible for the optochin-sensitive resistant (Opt^/Opt'^) phenotype. Our results aiso show that naturai as well as laboratory Opt" isolates have arisen by point mutations that produce different amino acid changes at positions 48, 49 or 50 of the ATPase c subunit. The nucleotide sequence of the Fo compiex of the Streptococcus oralis ATPase has also been determined. In addition, comparison of the sequence of the atpCAB genes of S. prieumoniae R6 (Opt^) and M222 (an Opt" strain produced by inter- species recombination between pneumococcus and S. oralis), and S. oralis reveaied that, in M222, an inter- change of atpC and atpA had occurred. We also demonstrate that optochin specificaily inhibited the membrane-bound ATPase activity of the S. pneumo- niae wild-type (Opt^) strains, and found a 100-fold dif- ference between Opt^ and Opt" strains, both in growth Inhibition and in membrane ATPase resistance. colony morphology on blood agar plates and immunoiogi- cal reaction with type-specific antisera ('Quellung' test) ailow the identification of pneumococcus (Lund and Henrichsen, 1978), However, pneumococcal isolates showing atypical responses in one or more of these tests, and viridans streptococci producing positive results in some of them, have been reported (Lund, 1950; Yurchah and Austrian, 1966; Wasilauskas and Hampton, 1984; Hoimerg etal, 1985; Pease etai, 1986; Kontiainen and Sivonen, 1987; Phiiiips et ai. 1988). Among a-haemolytic streptococci, S. pneumoniae and Streptococ- cus oralis are genetically related with a 56% overall simi- larity between their DNAs, and both possess choline as a cell wall component (Kilpper-Balz et ai, 1985), DNA probes derived either from S. oralis (Schmidhuber ef al., 1988) or S. pneumoniae (Pozzi ef ai. 1989; Fenoll ef al, 1990) have been used for species identification. Although the use of optochin was introduced early in this century for the treatment of experimentai pneumococcal infections, and optochin resistance was probably the first observation of the development of resistance in vivo to an antibacterial drug, both in mice (Morgenroth and Kaufmann, 1912) and man (Moore and Chesney, 1917), very little is known about the genetic and biochemical bases of the optochin-sensitive/resistant phenotype. Kreger and Olsen (1968) claimed that Opt inhibited an S. pneumoniae diaphorase (dehydrogenase) but a direct correlation between growth inhibition and the ievei of resistance of the purified diaphorase couid not be demonstrated. In the present study, we have cloned, sequenced, and characterized the gene responsible for the optochin-sensitivity/resistance trait of S. pneumoniae and S. oralis. Introduction Optochin (ethylhydrocupreine hydrochioride) (Opt) sen- sitivity and biie soiubility are two characteristics that differentiate Streptococcus pneumoniae from other a-haemolytic streptococci. These features, together with Received 26 November, 1993; revised 18 February, 1994; accepted 21 February, 1994. 'For correspondence. Tel. (1) 5611800; Fax (1) 5627518; e-mail; cibeg47@cc,csic,es. Results Constnjction ot Opt" S. pneumoniae strains To obtain an Opt" strain of pneumococcus, we first trans- formed the pneumocoocai M22 strain (Opt^) using DNAs from different Opt" streptococci (S. sanguis, S. mitis, S. gordonii and S. oralis) taxonomicaily related to S. pneumoniae. Only S. oralis DNA showed significant
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Molecular basis of the optochin-sensitive phenotype of pneumococcus: characterization of the genes...

Molecular Microbiology (1994) 12{4), 587-598
Molecular basis of the optochin-sensitive phenotype ofpneumococcus: characterization of the genes encodingthe FQ complex of the Streptococcus pneumoniae andStreptococcus oralis H"̂ -ATPases
Asuncion Fenoll,^ Rosario Munoz,' Ernesto Garcia^*and Adela G. de la Campa^^Centra Nacional de Microbiologia, Instituto de SaludCarlos III. 28220 Majadahonda, Spain.^Centro de Investigaclones Biologicas, CSIC, Velazquez144, 28006. Madrid, Spain.
Summary
The gene responsible for the optochin-sensitive (Opt^)phenotype of Streptococcus pneumoniae has beencharacterized. Sequence comparisons indicated thatthe genes involved encoded the subunits of the FQcomplex of an H^-ATPase. Sequence analysis andtransformation experiments showed that the atpCgene Is resfionsible for the optochin-sensitiveresistant (Opt^/Opt'̂ ) phenotype. Our results aisoshow that naturai as well as laboratory Opt" isolateshave arisen by point mutations that produce differentamino acid changes at positions 48, 49 or 50 of theATPase c subunit. The nucleotide sequence of the Focompiex of the Streptococcus oralis ATPase hasalso been determined. In addition, comparison of thesequence of the atpCAB genes of S. prieumoniae R6(Opt^) and M222 (an Opt" strain produced by inter-species recombination between pneumococcus andS. oralis), and S. oralis reveaied that, in M222, an inter-change of atpC and atpA had occurred. We alsodemonstrate that optochin specificaily inhibited themembrane-bound ATPase activity of the S. pneumo-niae wild-type (Opt^) strains, and found a 100-fold dif-ference between Opt^ and Opt" strains, both in growthInhibition and in membrane ATPase resistance.
colony morphology on blood agar plates and immunoiogi-cal reaction with type-specific antisera ('Quellung' test)ailow the identification of pneumococcus (Lund andHenrichsen, 1978), However, pneumococcal isolatesshowing atypical responses in one or more of thesetests, and viridans streptococci producing positive resultsin some of them, have been reported (Lund, 1950;Yurchah and Austrian, 1966; Wasilauskas and Hampton,1984; Hoimerg etal, 1985; Pease etai, 1986; Kontiainenand Sivonen, 1987; Phiiiips et ai. 1988). Amonga-haemolytic streptococci, S. pneumoniae and Streptococ-cus oralis are genetically related with a 56% overall simi-larity between their DNAs, and both possess choline as acell wall component (Kilpper-Balz et ai, 1985), DNAprobes derived either from S. oralis (Schmidhuber ef al.,1988) or S. pneumoniae (Pozzi ef ai. 1989; Fenoll ef al,1990) have been used for species identification.
Although the use of optochin was introduced early in thiscentury for the treatment of experimentai pneumococcalinfections, and optochin resistance was probably the firstobservation of the development of resistance in vivo toan antibacterial drug, both in mice (Morgenroth andKaufmann, 1912) and man (Moore and Chesney, 1917),very little is known about the genetic and biochemicalbases of the optochin-sensitive/resistant phenotype.Kreger and Olsen (1968) claimed that Opt inhibited anS. pneumoniae diaphorase (dehydrogenase) but a directcorrelation between growth inhibition and the ievei ofresistance of the purified diaphorase couid not bedemonstrated. In the present study, we have cloned,sequenced, and characterized the gene responsible forthe optochin-sensitivity/resistance trait of S. pneumoniaeand S. oralis.
Introduction
Optochin (ethylhydrocupreine hydrochioride) (Opt) sen-sitivity and biie soiubility are two characteristics thatdifferentiate Streptococcus pneumoniae from othera-haemolytic streptococci. These features, together with
Received 26 November, 1993; revised 18 February, 1994; accepted21 February, 1994. 'For correspondence. Tel. (1) 5611800; Fax (1)5627518; e-mail; cibeg47@cc,csic,es.
Results
Constnjction ot Opt" S. pneumoniae strains
To obtain an Opt" strain of pneumococcus, we first trans-formed the pneumocoocai M22 strain (Opt^) usingDNAs from different Opt" streptococci (S. sanguis,S. mitis, S. gordonii and S. oralis) taxonomicaily related toS. pneumoniae. Only S. oralis DNA showed significant

588 A. Fenoll, R. Munoz, E. Garcia and A. G. de la Campa
transformation efficiency and one Opt" M22 colony wasselected and designated as strain M222. Aftenwards, weisotated a spontaneous Opt" mutant (MJ1) by growingS. pneumoniae R6 at inhibitory concentrations of optochin(the frequency of spontaneous mutation to Opt" beingabout 10"^). Then, DNA from MJ1 was used to transformcompetent cells of strain R6, and one Opt" colony, namedMJ11, was seiected for further study. Other Opt" pneumo-coccal strains, either clinical or laboratory isolates, aredescribed below.
Cloning and sequencing of the genes conferring Optresistance on M222
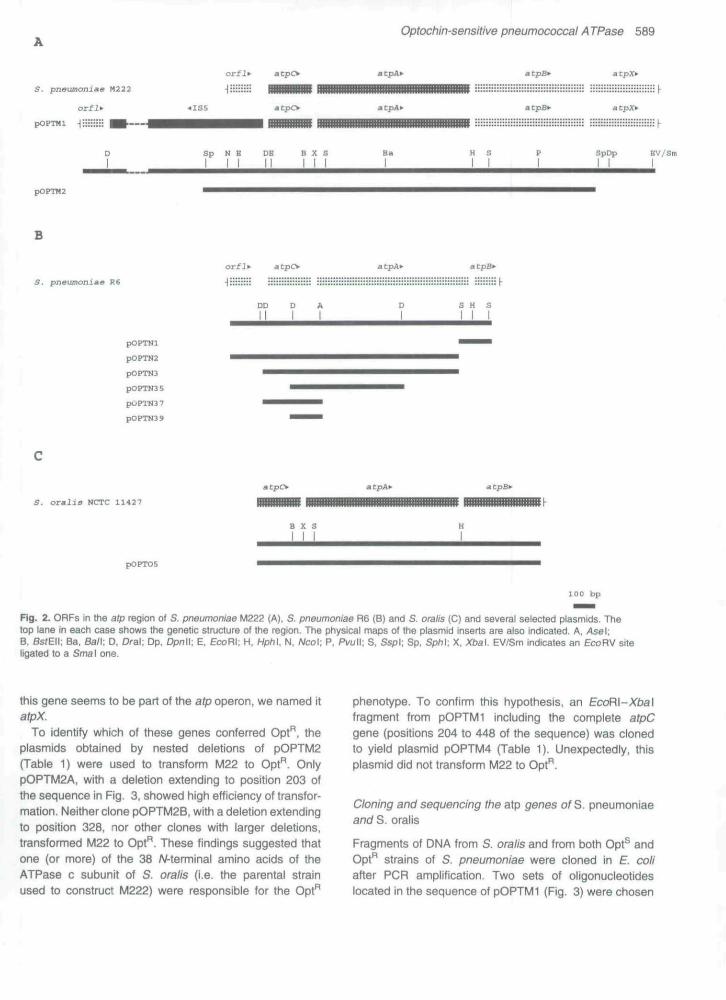
Chromosomal M222 DNA was digested with severalrestriction endonucleases and the residual Opt'̂ transform-ing activity was assayed using strain M22 (Opt®) as recipi-ent. EcoRV fragments of 5-7 kb, isolated from agarosegels and showing the highest transformation activity,were cloned into Escherichia coli Jfvl83 using Smal-cutpUC13. Alkaline lysates of E. coli transformants weretested for the ability to transform the M22 strain to Opt".One clone harbouring a recombinant plasmid (pOPTMl)with a 6.7 kb DNA insert was isolated from about 1500recombinant colonies, and used for further studies. Tolocalize the DNA region responsible for the Opt" pheno-type, different restriction fragments of pOPTfuli (Fig. 1)were isolated and used to transform the M22 strain toOpt". The gene responsible for the Opt" phenotype waslocalized in a 1.8 kb Sph\ fragment. This fragment wascloned into the Sph\ site of pBGS19+, yielding plasmidpOPTM2 (Table 1), and sequenced using synthetic oligo-nucleotides. Figure 2 A shows the restriction map andgene order. We found four complete open reading frames
(ORFs) of substantial length. The first ORF having an ATGcodon near its origin had an orientation opposite to that ofthe other three. The deduced amino acid sequence of thisORF proved to be identical to the lns5A protein encoded bythe IS5 eiement of E. coli (Kroger and Hobom, 1982).Sequencing, restriction mapping and polymerase chainreaction (PCR) amplification analyses (data not shown)indicated that IS5was not present in either of the strepto-coccal genomes and, consequently, is presumed to be acloning artefact.
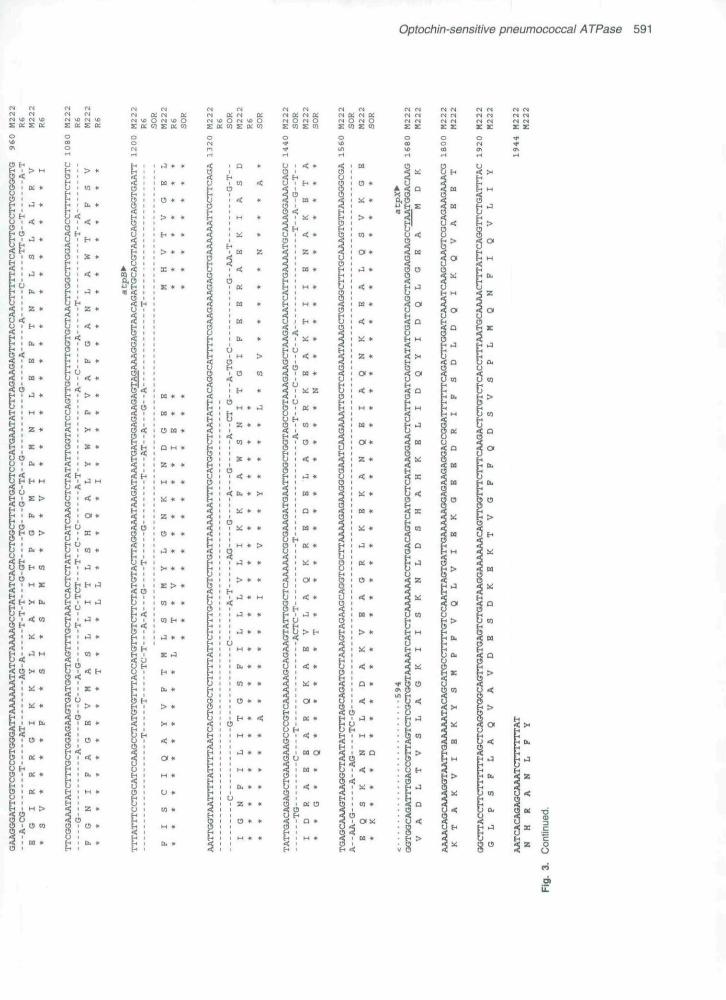
Figure 3 shows the nucieotide sequence of the DNAinsert of plasmid pOPTMl (M222 strain) with exclusion ofthe 1S5 element. The site of integration of the IS5 element(CTAA) is located at nucleotides 176-179. Three completeORFs beginning after this CTAA box. and oriented In theopposite direction for transcription to IS5, were found.The first one (positions 213-413) could encode a proteinof about 7 kDa, and the second (positions 448-1164)and the third ones (positions 1178-1672) would encodeproteins of 27 kDa and 18 kDa, respectively. Amino acidsequence comparison with the EMBL data bank, showedthat the three ORFs were homologous to the genescoding for the c, a, and b subunits, respectively, of the Foportion of the H*-ATPase (data not shown). We thereforenamed these genes atpC. A and B. A good consensuspromoter sequence was located upstream from theoperon and all three genes were preceded by putativeribosome-binding sites (RBS) (Shine and Dalgamo,1975). An incomplete ORF beginning in an ATG codonthat overlapped the termination codon of atpB was locatedat the 3' end of the atpCAB operon. A putative RBS alsopreceded this gene. We were not able to find any signifi-cant similarity between the amino acid sequence of thisgene and those available at the EMBL data bank, but as
( • )
( • )
lkh
Fig, 1. Localization oi the Opt*^-coding region in plasmid pOPTMl. The restriction map of pOPTMl is shown linearized at the vector EcoRIsite. Restriction fragments used to test transforming activity are indicated. {+). and ( - ) indicates that Ihe efficiency of transformation was higherthan lO"" or lower than 10 transformants ml \ respectively.
Optochin-sensitive pneumococcal ATPase 589
S. pneumoniae N222
pOPTHi -I:::;::::
orfl* a cpcv
mtpO a tpB^
S p N B DB B X S SpDp BV/Sm
pOPTM2
B
8. pnauncinia« R6
atpA*-
DD D A
pOPTNl
pOPTN2
pOPTN3
pOPTN3 5
POPTN39
S. ovalia NCTC 11427
pOPTOS
100 bp
Fig. 2. ORFs in the afp region of S. pneumoniae M222 (A), S, pneumoniae R6 (B) and S. ora//s (C) and several selected plasmids. Thetop lane in each case shows the genetic structure of the region. The physical maps of the plasmid inserts are aiso indicated. A, Asel.B, Ss/Eii; Ba, Ban. D. Oral; Dp. Dpnll; E, EcoHl. H. Hph\, N, Ncol; P, Pvull; S. Ssp\; Sp. SpM; X. Xbal EV/Sm indicates an £coRV siteiigated to a Sma\ one.
this gene seems to be part of the atp operon, we named itatpX.
To identify which of these genes conferred Opt" ,̂ theplasmids obtained by nested deletions of pOPTM2(Tabie 1) were used to transform M22 to Opt". OnlypOPTM2A, with a deletion extending to position 203 ofthe sequence in Fig. 3. showed high efficiency of transfor-mation. Neither clone pOPTM2B, with a deletion extendingto position 328, nor other clones with larger deletions,transformed M22 to Opt". These findings suggested thatone (or more) of the 38 /V-terminal amino acids of theATPase c subunit of S. oralis (i.e. the parental strainused to construct M222) were responsible for the Opt"
phenotype. To confirm this hypothesis, an EcdR\~Xba\fragment from pOPTMl including the complete atpCgene (positions 204 to 448 of the sequence) was clonedto yield plasmid pOPTM4 (Table 1). Unexpectedly, thisplasmid did not transform M22 to Opt".
Cloning and sequencing the atp genes of S. pneumoniaeand S. oralis
Fragments of DNA from S. oralis and from both Opt^ andOpt" strains of S. pneumoniae were cloned in E. coliafter PCR amplification. Two sets of oligonucleotideslocated in the sequence of pOPTMl (Fig. 3) were chosen

590 A. Fenoll, R. Mufioz, E. Garcia and A. G. de la Campa
IV a IN
Eh k k P
I- • •
> • k
w S C O< -^ £9S: D ) ^ C^ c -a «E nj c CD
-a ol .y s
CO C m
^ ,_ Q) gTJ » O «
^ To a.%
« — .> —
2 3 ™ inc sr Q. „ CO
1 g
I 111 I^ £ iS•D c S
lC*" i : O Oi n n > i =
Optochin-sensitive pneumococcal ATPase 591
B k
•1 kI
•J k
M k
2 «
E kI
Oi •
cs(SE
KO
m
cscsE
CKO
mINE
taom
fiINE
KOCO
<NCNE
c-tt sE
O o "< z q3 •? O
ri
i
592 A. Fenoll. R. hAunoz, E. Garcia and A. G. de la Campa
Table 1. Recombinant plasmids used in this study.
Name
pOPTMlpOPTM2pOPTM2ApOPTM2BpOPTM2CP0PTM2EpOPTM2FpOPTM2GP0PTM2HpOPTM2lpOPTM3pOPTM4pOPTNlpOPTN2pOPTN3POPTN35POPTN37POPTN39pOPTO5
Mariners
amp, atpCAB, 1S5kan. atpCABkan. atpCABkan. atpABkan, atpABkan. atpBkan. atpBkan. alpskan. atpBkanamp. atpCABamp. atpCampamp, atpCAamp, atpCamp, AatpC, AatpAamp, atpCamp. XatpCamp, atpCA
Insert^
1-19441-1715
203-1715328-1715440-1715642-1715832-1715975^1715
1312-17151498-1715203-1944203-447
1152-12791-1152
218-1148292-864218-462294-462204-1560
Transformation"
-1-
++-—-----+-—_+++++
Constnjction
pUC13/Smal Iigated to M222/EcoRVpBGS19+/Sp/il Iigated to pOPTMI/SphIpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM2 5' deletionpOPTM1/£coHI self-ligatedpOPTM3/Xtjai self-ligatedSee text for detailsSee text for detailsSee text for detailspOPTN3/Dral iigated to pUC18/SmalpOPTN3/ecoRI + Asel Iigated to pUC18/SmalpOPTN3/Dral + Ase\ Iigated to pUC18/SmalSee text for details
a. Nucleotides included in the Insert refer to the positions indicated in Fig. 3.b, Transformation of M22 to Opt" with an efficiency higher than lO'' transfonnants ml"
to amplify atp genes: oligonucleotides 660 and 512 arelocated respectively before and after the insertion site ofIS5 in pOPTMl, whereas oligonucleotide 594 is locatednear the 3' end of atpB. When the set 660/594 was used,all S. pneumoniae DNAs tested (including M222 DNA)showed a major band of about 1.4 kb. No amplificationwas observed with S. oralis DNA (data not shown). Withthis set of primers, pOPTMl showed a band of about2.5 kb, as should be expected from the insertion of anIS5 element (1165 bp) in this plasmid. When oligonucleo-tides 512/594 were used, M222, pOPTMl and S. oralisshowed a band of the expected size (1.4 kb), but theband observed with other S. pneumoniae DNAs was ofabout 1.7 kb (data not shown). To check for the accuracyof the amplified products, the bands of 1.4 and 1.7 kbcorresponding to the amplification of the MJ11 (Opt*̂ )strain were assayed for transformation activity. Both DNAfragments transformed M22 to Opt" with high efficiency.For cloning afp genes of S. pneumoniae, the 1.7 kb PCRfragments of R6 and MJ11 DNAs obtained by amplifi-cation with oligonucleotides 512/594 were cut with EcoRI(a target included in the oligonucleotide 512) and Ssp\and Iigated to EcoRI/Smal-cut pUC18- Clones derivedfrom R6 were checked by in situ hybridization with^P-labelled pOPTM2 A and those derived from MJ11 werechecked for transformation activity to the Opt" phenotype.Three independent clones were isolated and their plasmidinserts were sequenced: pOPTNl and pOPTN2 derivedfrom R6, and pOPTN3 from MJ11 (Table 1, Fig. 2B).
A comparison between the DNA sequences of M222(plasmid pOPTMl) and R6 (ptasmids pOPTNl andpOPTN2) showed an identical region extending from the5' end of the sequence in Fig. 3 to near the CTAA box.
Variations in the sequence were observed in atpC andatpA genes but identity was restored in the atpB gene(Fig. 3). Three nucleotide changes were observed in thecomparison of the nucleotide sequences of R6(pOPTN2) and MJ11 (pOPTN3): two of them were inatpC and one in atpA. Among the changes in atpC, thefirst one was In the cloning site (i.e. EcoRI target inpOPTN3 but not in pOPTN2) at position 220 (Fig. 3), thesecond one (position 357) was a G to A transition thatwould change Ala to Thr in the amino acid chain of theresistant allele, and the third change (position 897)affected atpA, Transformation experiments using differentrestriction fragments obtained from pOPTN3 (Table 1,Fig. 2B) indicated that the mutation responsible for theOpt"̂ phenotype was the G to A transition at position357. in addition, 1.4 kb DNA fragments obtained by ampli-fication of R6 and MJ11 DNA with oligonucleotides 660and 594 (see above) were used as templates employingthe oligonucleotide 660 as a primer to detemiine the com-plete sequence of the atpC gene. This experimentalapproach confirmed the mutation in MJ11 at position357- Moreover, the nucleotide sequence of the atpCgene from four additional Opt" R6 derivatives and severalclinical isolates of S. pneumoniae. either Opt" (four strains)or Opt^ (three strains), was determined in the same way.Furthermore, we employed the PCR products to transfomithe Opt" trait and observed that only those obtained fromOpt" isolates did transfonn the sensitive strain R6. Onthe other hand, the nucleotide sequence of the atpCgene from the Opt^ clinical isolates was identical to thatof R6 (data not shown) whereas all the Opt" isolateshave point mutations located at nucleotides 354 (strainsMJ2. MJ4. MJ5, MJ6, MJ455/89, and MJ4133/89), 357
Optochin-sensitlvB pneumococcai A TPase 593
Table 2. Mutational substitutions on severalnatural and laboratory Opt" S. pneumoniaeisolates.
strain
R6 derivatives
MJ11MJ2MJ4MJ6MJ6
Clinical isolates
MJ959/89MJ455/89MJ1996/89MJ4133/89
Source
SM^SM"SM^SM^SM^
BloodCSF"BloodSputum
Capsulargroup
Rough"Rough"Rough"Rough"Rough"
196
14NT"
Mutation
nucleotide amino acidsubstitution substitution
G ^ " - A Ala-49 - ThrG *̂** — C Val-48 -^ LeuG^^ -^ C Val-48 - LeuG^** -> C Val-48 -- LeuG ^ - C Val-48 -• Leu
G ^ ' — A Ala-49 -- ThrQ^ -, T Val-48 - PheT * ^ — G Phe-50 - . LeuG ^ -^ T Val-48 - . Phe
a. SM, spontaneous mutant isolated in the laboratory.b. The progenitor of these strains was of serogroup 2.c. CSF, cerebrospinal fluid.d. NT, non-typable.
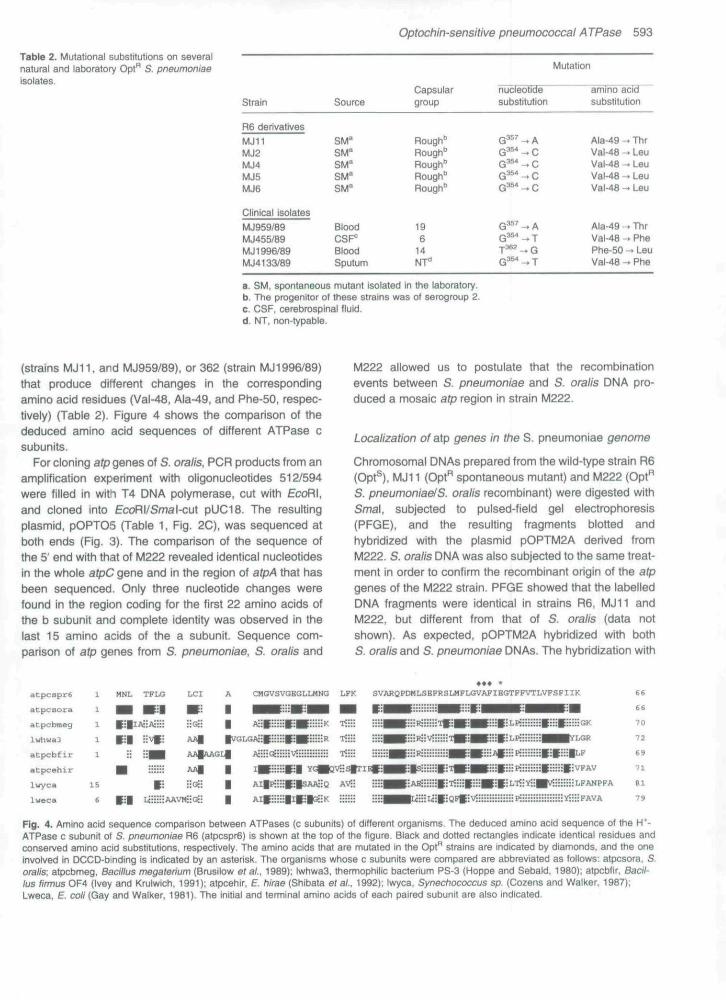
(strains MJ11. and MJ959/89), or 362 (strain MJ1996/89)that produce different changes in the correspondingamino acid residues (Val-48, Ala-49, and Phe-50, respec-tively) (Table 2). Figure 4 shows the comparison of thededuced amino acid sequences of different ATPase csubunits.
For ctoning afp genes of S. oraiis, PCR products from anamplification experiment with oligonucleotides 512/594were filled in with T4 DNA polymerase, cut with EcoRI,and cloned into EcoRI/Smal-cut pUC18. The resultingplasmid, pOPTO5 (Table 1, Fig. 2C), was sequenced atboth ends (Fig. 3). The comparison of the sequence ofthe 5' end with that of fvl222 revealed identical nucleotidesin the whole afpC gene and in the region of atpA that hasbeen sequenced. Only three nucleotide changes werefound in the region coding for the first 22 amino acids ofthe b subunit and complete identity was observed in thelast 15 amino acids of the a subunit. Sequence com-parison of afp genes from S. pneumoniae, S. oraiis and
M222 allowed us to postulate that the recombinationevents between S, pneumoniae and S. oraiis DNA pro-duced a mosaic atp region in strain M222.
Locaiization of atp genes in the S. pneumoniae genome
Chromosomal DNAs prepared from the wild-type strain R6(Opt^), MJ11 (Opt" spontaneous mutant) and M222 (Opt"S, pneumoniaelS. oraiis recombinant) were digested withSmal, subjected to pulsed-field gel electrophoresis(PFGE), and the resulting fragments blotted andhybridized with the plasmid pOPTM2A derived fromM222. S. oratis DNA was also subjected to the same treat-ment in order to confirm the recombinant origin of the atpgenes of the M222 strain. PFGE showed that the labelledDNA fragments were identical in strains R6, fvlJII andM222, but different from that of S. oraiis (data notshown). As expected, pOPTM2A hybridized with bothS. oraiis and S. pneumoniae DNAs. The hybridization with
AtpCBprSacpcHora
atpcbmeg
Iwhwa3
atpcbfir
lwyca
lweca
1
1
1
1
1
IS
6
MNL TPLG
PI
AA|
CMGVSVGBGLUING LFK
AV-
SVARQPDKLSBPRSLHPLGVAFIEGTFFVTLVFSFIIK 66
66
70
72
69
71
Bl
79
Fig. 4. Amino acid sequence comparison between ATPases (c subunits) of different organisms. The deduced amino acid sequence of the H*-ATPase c subunit of S. pneumoniae R6 {atpcspr6) is shown at the top of the figure. Black and dotted rectangles indicate identical residues andconserved amino acid substitutions, respectiveiy. The amino acids (hat are mutated in the Opt'^ strains are indicated by diamonds, and Ihe oneinvolved in DCCD-bmding is indicated by an asterisk. The organisms whose c subunits were compared are abbreviated as follows: atpcsora, S,orate; atpcbmeg, Badtlus megaterium (Brusilow ef ai, 1989); Iwhwa3, thennophiilc bacterium PS-3 (Hoppe and Sebaid, 1980); atpcbfir, Bacit-tus firmus 0F4 (Ivey and Krulwich. 1991); atpcehir, E, hirae (Shibata etal.. 1992); iwyca. Synechococcus sp. (Cozens and Walker. 1987);Lweca, £". cott (Gay and Walker, 1981). The initial and terminal amino acids of each paired subunit are aiso indicated.
594 A. Fenoll, R. Mufioz, E. Garcia and A. G. de la Campa
the S. pneumoniae DNAs localized the atp genes in aSmal fragment of 260kb (fragment number 4). Theseresults agree with the previous localization of the opt-r2allele by chromosomai transformation (Gasc etai, 1991).
In vitro measurements of ATPase activity
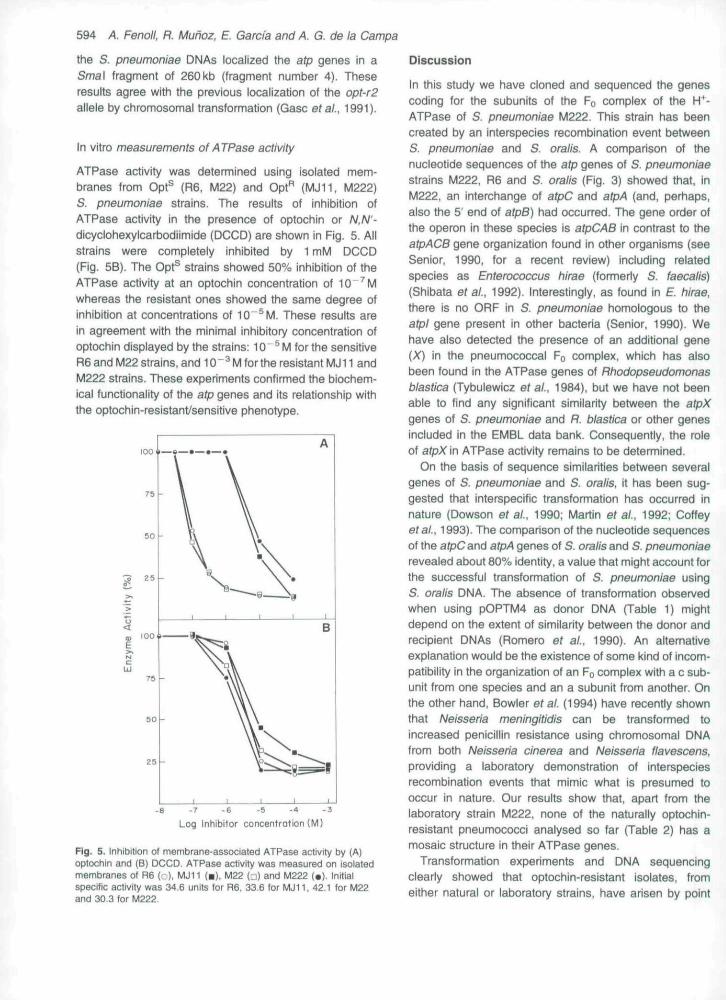
ATPase activity was determined using isolated mem-branes from Opt^ (R6, M22) and Opt"̂ (MJ11, M222)S. pneumoniae strains. The resuits of Inhibition ofATPase activity in the presence of optochin or N.N'~dicyclohexylcarbodiimide (DCCD) are shown in Fig. 5. Allstrains were completely inhibited by 1 mM DCCD(Fig. 5B). The Opt^ strains showed 50% inhibition of theATPase activity at an optochin concentration of 10"^Mwhereas the resistant ones showed the same degree ofinhibition at concentrations of 10"^M. These results arein agreement with the minimal inhibitory concentration ofoptochin displayed by the strains: 10"^ M for the sensitiveR6 and M22 strains, and 10 " ^ M for the resistant MJ11 andM222 strains. These experiments confirmed the biochem-ical functionality of the atp genes and its relationship withthe optochin-resistanfsensitive phenotype.
100 11 9—
-B -7 -6 -S -4 -3Log Inhibitor concentration (M)
Fig. 5. Inhibition of membrane-associated ATPase activity by (A)optochin and (B) DCCD. ATPase activity was measured on isolatedmembranes of R6 (•..), MJ11 (•), U22 (j) and M222 (•). Initialspecific activity was 34.6 units for R6. 33.6 for MJII, 42.1 for M22and 30.3 for M222.
Discussion
In this study we have cloned and sequenced the genescoding for the subunits of the FQ complex of the H"*-ATPase of S. pneumoniae M222. This strain has beencreated by an interspecies recombination event betweenS. pneumoniae and S. oralis. A comparison of thenucieotide sequences of the atp genes of S. pneumoniaestrains M222, R6 and S. oralis (Fig. 3) showed that, inM222, an interchange of atpC and atpA (and, perhaps,also the 5' end of atpB) had occurred. The gene order ofthe operon in these species is atpCAB in contrast to theatpACB gene organization found in other organisms (seeSenior, 1990, for a recent review) including relatedspecies as Enterococcus hirae (formerly S. faecalis)(Shibata etai, 1992). Interestingly, as found in E. hirae,there is no ORF in S. pneumoniae homologous to theatpl gene present in other bacteria (Senior, 1990). Wehave also detected the presence of an additional gene(X) in the pneumococcal Fo complex, which has alsobeen found in the ATPase genes of Rhodopseudomonasblastica (Tybutewicz etal., 1984). but we have not beenable to find any significant similarity between the atpXgenes of S. pneumoniae and R. blastica or other genesincluded in the EMBL data bank. Consequently, the roleof atpX in ATPase activity remains to be determined.
On the basis of sequence similarities between severalgenes of S. pneumoniae and S. oralis, it has been sug-gested that interspecific transformation has occurred innature (Dowson et ai, 1990; Martin et ai, 1992; Coffeyef ai, 1993). The comparison of the nucieotide sequencesof the a^Cand atpA genes of S. oralis and S. pneumoniaerevealed about 80% identity, a value that might account forthe successful transfonnation of S. pneumoniae usingS. oralis DNA. The absence of transfonnation observedwhen using pOPTM4 as donor DNA (Table 1) mightdepend on the extent of similarity between the donor andrecipient DNAs (Romero et ai. 1990). An alternativeexplanation would be the existence of some kind of incom-patibility in the organization of an FQ complex with a c sub-unit from one species and an a subunit from another. Onthe other hand. Bowler et ai (1994) have recently shownthat Neisseria meningitidis can be transformed toincreased penicillin resistance using chromosomal DNAfrom both Neisseria cinerea and Neisseria flavescens.providing a laboratory demonstration of interspeciesrecombination events that mimic what is presumed tooccur in nature. Our results show that, apart from thelaboratory strain M222, none of the naturally optochin-resistant pneumococci analysed so far (Table 2) has amosaic structure in their ATPase genes.
Transformation experiments and DNA sequencingclearly showed that optochin-resistant isolates, fromeither natural or laboratory strains, have arisen by point

Optochin-sensitive pneumococcai A TPase 595
mutations in the atpC gene altering Ala-49 or its neigh-bours. These amino acid residues are near to Glu-52which, in all the species studied so far. is directly involvedin proton translocation (Senior, 1990). fviutationa! analysescarried out in E. coli have shown that Gly-58 (equivalent toAla-49 in S, pneumoniae) is also an important residue forproton conduction (Mosher et ai, 1983). Moreover, inSaccharomyces cerevisiae, oligomycin and venturicidinresistance can be induced by an Ala to Thr change in themitochondrial H*-ATPase c subunit in a positionequivalent to the Ala-49 residue of S. pneumoniae (Nag-ley et ai, 1986), However, S. oraiis. a naturally Opt"species, retains unchanged the Ala-49 and the Phe-50residues and a conserved substitution (Val to He) is pre-sent at position 48 of the c subunit, a finding whichindicates that alterations other than those found in themutants reported here might also confer an Opt"̂ pheno-type. Transfonnation experiments using plasmids with dif-ferent deletions allowed us to conclude that the nucleotidechange(s) responsible for this phenotype are some ofthose encoding the 38 W-tenninal amino acids of the c sub-unit of S. oraiis (Table 1). On the other hand, it is well docu-mented that the DCCD-binding site in E. coli is Asp-61 ofthe c subunit (equivalent to Glu-52 of S, pneumoniae)and mutations that eliminate the carboxyl side chainshow a DCCD-resistant phenotype (Senior, 1990), butother DCCD-resistant mutations reside in lle-28. which isclosely juxtaposed to Asp-61 within the membranebilayer. Consequently, it is conceivable that mutationsother than those affecting amino acids 48, 49 or 50 in theS. pneumoniae ATPase c subunit would also produce anOpt"̂ phenotype. Nevertheless, further experimentswould be needed to prove this hypothesis.
Optochin inhibition to a S. pneumoniae diaphorase hadbeen reported (Kreger and Olsen, 1968), but a directrelationship between growth inhibition and the level ofresistance of the purified diaphorases was not seen.Thus, the Opt" pneumococci are 140- to 240-fold moreresistant to growth inhibition than the wild-type pneumo-cocci but the mutant diaphorases were only four- to eight-fold more resistant to optochin inhibition than those of theOpt^ strains. In contrast, the results reported in thepresent study demonstrate a direct relationship betweenOpt sensitivity and Opt-induced ATPase inhibition asa 100-fold difference (Fig. 5) was found between Opt-sensitive and -resistant strains both in growth inhibition andin membrane ATPase resistance.
(Ronda et ai, 1988), and three clinical isolates (MJ425,MJ647, and MJ803), MJ1, MJ2, MJ4, MJ5, and MJ6 arespontaneous Opt"̂ pneumococcai mutants obtained byplating about 10'' cells of strain R6 on blood agar platescontaining optochin (Sigma) at 12.5|,igml '. The clinicalOpt*̂ isolates of S. pneumoniae MJ959/89, MJ455/89,MJ1996/89, and MJ4133/89 were isolated as previouslydescribed (Kontiainen and Sivonen, 1987), Strain MJ11 wasobtained by transformation ot R6 with MJ1 DNA and selectionfor Opt" colonies, M222 is an Opt" derivative ot M22 obtainedby transformation with S. oratis DNA, Streptococcal strains,other than S. pneumoniae, were: S. oratis NCTC 11427,S. sanguisATCC 10556. S. m/f/sATCC 9811 and S. gordoniiATCC 33399. The plasmids used for cloning were pUCi3(Messing, 1983), pUC18 (Yanisch-Perron etai. 1985) andpBGS19+ (Spratt ef at.. 1986) and the recombinant plasmidsobtained in this study are listed in Table 1,
Growth and transformation of bacteria
E. coii was grown in LB medium (Sambrook et ai, 1989).Streptococci were grown in Todd-Hewitt broth (Difco),supplemented with 0.5% yeast extract (Difco), Mueller-Hinton agar plates (Difco) supplemented with 5% defibri-nated sheep blood were used for optochin-sensitivity tests,S. pneumoniae was also grown in liquid C medium (Tomasz,1970), containing 0.08% yeast extract (C+Y). Transfonnationof S. pneumoniae was performed according to a procedurepreviously described (Tomasz, 1970). Transformants ofS. pneumoniae ware selected in C+Y plates containing Opt at5|.Lgml" ^ and incubated at 37 C in a 5% CO? atmosphere.E. co//transformants were selected in LB medium containingampicillin at 100 HQ ml ^ or kanamycin at 25 ng ml \
Chromosomal DNA and plasmid preparation
Streptococcal strains other than S. pneumoniae were lysedwith achromopeptidase (Sigma) as described by Ezaki et ai(1988). S. pneumomae chromosomal DNA was obtained asfollows: cells from an exponentially growing culture in C+Y orTodd-Hewitt medium were washed twice with lOmM Tris-HCI (pH8,0), 10mM EDTA, 150mM NaCI, and resuspendedin 1/50 of the initial volume ol 50 mM Tris-HCI (pH 8,0), 30 mMEDTA. 0.4% sarkosyl and 0.1% Triton X-100 (Morrison,1978), Cells were incubated at 37 C until lysis and thentreated with RNase (100|.igml"') and proteinase K(100|,igml ^). Then, CsCI was added to 1,7 g cm ^densityand ultracentrifuged at 80 000 x g for 60 h (Sambrook et ai,1989), Plasmids were prepared from E. co//by the method ofBimboim and Doly (1979). Purified plasmids were preparedby equilibrium centrifugation in CsCI-ethidium bromidegradients (Sambrook etai, 1989).
Experimental procedures
Bacterial strains and plasmids
The E coli strains used were DH5IL (Hanahan, 1983) andJM83 (Yanisch-Perron et ai. 1985), The Opt® strains ofS. pneumoniae used were: the wild-type strain R6 and M22
DNA manipulations
Restriction andonucleases, T4 DNA ligase, T4 and Kienowfragment DNA polymerases were obtained commercially andused as specified by the suppliers. Unidirectional deletions ofthe DNA were generated by Exolll and SI nuclease using theDouble-Stranded Nested Deletion Kit (Pharmacia), Gel

596 A. Fenoli R. Muhoz, E. Garcia and A. G. de la Campa
eiectrophoresis of plasmids, restriction fragments, and PCRproducts were carried out in agarose gels as described(Sambrook etal.. 1989). DNA was recovered from gel sliceswith the Gene Clean II Kit (Bio 101).
Polymerase chain reaction amplification
PCR amplifications were performed using 2 units of Taqpolymerase (Parkin Elmer Cetus), 1 |.ig of chromosomalDNA, 1 |.iM of each synthetic oiigonucleotide primer, 200 nMof each dNTP, and 5mM MgCl2 in the buffer recommendedby the manufacturers. Amplification was achieved with acycle of 1 min denaturation at 95 C, 2 min annealing at55 C and 2.5 min polymerase extension at 72 C repeated 20times with a final 7.5 min 72 C extension step and slowcooling to 4 X using a Linus Dualcycler. The followingsynthetic oiigonucleotide primers were used: 512, 5 -AGTAAATTTTAAATAAAAGGAGAATTCATC-3; 594, 5 -CCAGCGAGACTAACGGTCAAATCTGCCACC-3'; 660, 5-CGCTTCCAATAGCGGTTAAAAGTTGAC AAA-3'.
In situ colony hybhdization
In situ colony hybridization with radioactive ^^P-iabelledplasmid pOPTM2A was performed as described by Sam-brook et ai (1989). DNA was radiolabelled with 50nCi of[a-^^P]-dCTP (3000Ci mmol" ' ) using the Multiprime DNALabelling System (Amersham).
DNA sequence determination and analysis
DNA sequencing was carried out by the dideoxy-chaintermination method (Sanger ef ai. 1977), using theSequenase Kit from US Biochemicals. Oiigonucleotideprimers were synthesized in a Pharmacia UKB GeneAssembler Plus DNA synthesizer. Atl sequences showed inthis study were determined in both DNA strands. Direct PCRsequencing was carried out with the fmof^ DNA SequencingSystem from Promega. DNA and protein sequence com-parisons were done using software from the WisconsinGenetics Computer Group.
PFGE of chromosomal DNA
DNA embedded in agarose plugs was prepared from S. oralisand S. pneumoniae strains R6 and MJ11 as previouslydescribed (Gasc ef ai, 1991). Gels were cast and run in75mfyl Tris, 25 mM boric acid, 0.1 mM EDTA at 7 C in acontour-clamped homogeneous electric field-DRII apparatus(Bio-Rad) at 200 V with the switch-time ramped from 1 to 50 sover the 30 h running time. DNA probes were labelled withthe PolarPlex^" Chemiluminescent Blotting Kit (Millipore).Southern blots and hybridization followed the manufacturer'sinstructions.
ATPase activity measurements
Cultures were grown in Todd-Hewitt broth supplementedwith 0.5% yeast extract until late-exponential phase.
S. pneumoniae membranes were prepared by sonication(6 min) in 20 mM Tris-HCI (pH7.5), 1 mM MgClg (Trombeef al., 1984). Protein content was determined by the ProteinAssay Kit from Bio-Rad. The assay buffer used was lOOmMTris-HCI (pH7.5), 5mM MgCIs with 5mM ATP as substrate.To determine the susceptibility of the membrane ATPase toDCCD and optochin, extracts were incubated with theinhibitor for 20 min at room temperature before the additionof ATP. ATPase activity was assessed in terms of the releaseof phosphate from ATP and was expressed per mg ofmembrane protein. Inorganic phosphate (Pi) was assayedas previousiy described (Chen et ai, 1956), and an enzymeunit was taken as the amount of enzyme that resulted in therelease of 1 nmol of Pi per ml per 30 min.
Acknowledgements
We thank Rubens Lopez, Pedro Garcia, and Jose L. Garciafor stimulating suggestions and for the critical reading of themanuscript. We also thank William Sanders for correcting theEnglish version. The help of Nuria de Pedro on the firstoloning experiments is greatly appreciated. R.M. ts thebeneficiary of a postdoctoral fellowship from the Instituto deSalud Carlos III. This research was supported by GrantsSAL91-0898-C02-01/02 from the Comision Interministerial deCiencia y Tecnologia (CICYT).
References
Bimboim, H.C, and Doly. J. (1979) A rapid alkaline extractionprocedure for screening recombinant DNA. NucI Acids Res7: 1513-1523.
Bowler, L.C. Zhang, Q-Y., Riou. J-Y.. and Spratt. B. G.(1994) Interspecies recombination between the penAgenes of Neisseria meningitidis and commensal Neisseriaspecies during the emergence of penicillin resistance in N.meningitidis. natural events and laboratory simulation. JBacteriol 176: 333-337.
Brusilow, W.S., Scarpetta, M.A., Hawthorne, C.A., and Clark,W.P. (1989) Organization and sequence of the genescoding for the proton-translocating ATPase of Bacillusmegatehum. J Biol Chem 284: 1528-1533.
Chen, Jr, P.S., Toribara, T.Y., and Warner, H. (1956)Microdetermination of phosphorus. Anal Chem 28:1756-1758.
Coffey, T.J., Dowson, C.G., Danielts, M., and Spratt, B.G.(1993) Horizontal spread of an altered penicillin-bindingprotein 2B gene between Streptococcus pneumoniae andStreptococcus oralis. FEMS Microbioi Lett 110: 335-340.
Cozens, A.L., and Walker. J.E. (1987) The organization andsequence of the genes for the ATP synthase in thecyanobacterium Synechococcus 6301. Support for anendosymblotic origin of chloroplasts. J Mol Biol 194:359-383.
Dowson, C.G., Hutchison, A., Woodford, N., Johnson, A.P.,George, R.C, and Spratt, B.G. (1990) Penicillin-resistantviridans streptococci have obtained altered penicillin-binding protein genes from penicillin-resistant strains ofStreptococcus pneumoniae. Proc NatI Acad Sci USA 87:5858-5862.

Optochin-sensitive pneumococcai ATPase 597
Ezaki, T., Hashimoto, Y., Takeuchi. N., Yamamoto, H., Liu,S.-L, Miura, H., Matsui, K,, and Yabuuchi, E, (1988)Simple genetic method to identify viridans group strepto-cocci by colorimetric dot hybridization and fluorometrichybridization in microdilution wells, J Clin Microbiot 26:1708-1713.
Fenoll, A.. Martinez-Suarez, J.V., Munoz, R., Casal, J., andGarcia, J.L. (1990) Identification of atypical strains ofStreptococcus pneumoniae by a specific DNA probe. EurJCtin Microbiot Inf Dis 9: 396-401.
Gasc, A.-M., Kauc, L,, Barraille, P., Sicard, M., and Goodgal,S, (1991) Gene localization, size, and physical map of thechromosome of Streptococcus pneumoniae. J Bacteriol173: 7361-7367.
Gay, N.J., and Walker, J,E, (1981) The a/poperon: nucleotidesequence of the promoter and the genes for the membraneproteins, and the 6 subunit of Escherichia coli ATP-synthase, NucI Acids Res 9: 3919-3926,
Hanahan, D. (1983) Studies on transformation of Escherichiacoli with plasmids. J Mot Biot 166: 557-580,
Holmberg, H., Danielsson. D,. Hardie, J,. Krook, A., andWhiley, R. (1985) Cross-reactions between alpha-streptococci and omniserum. a polyvalent pneumococcaiserum, demonstrated by direct immunofluorescence,immunoeiectroosmophoresis and latex agglutination. JCtin MiCfVbiot 21: 745-748.
Hoppe, J., and Sebaid, W, (1980) Amino acid sequence of theproteolipid subunit of the proton-translocating ATPasecomplex from the thermophilic bacterium PS-3. Eur JBiochem 107: 57-65,
Ivey, D,M,, and Krulwich, T,A, (1991) Organization andnucleotide sequence of the atp genes encoding the ATPsynthase from alkaliphilic Bacillus firmus 0F4. Mol GenGenet 229: 292-300.
Kilpper-Balz, R., Wenzig, P,, and Schleifer, K.H. (1985)Molecular relationships and classification of some viridansstreptococci as Streptococcus oraiis and amended descrip-tion of Streptococcus oratis (Bridge and Sneath 1982), IntJSyst Bacteriol 35: 482-488.
Kontiainen, S,, and Sivonen. A, (1987) Optochin resistancein Streptococcus pneumoniae strains isolated fromblood and middle ear fluid. Eur J Ctin Microbiol 6:422-424.
Kreger, AS., and Olsen. R,H, (1968) Purification andproperties of mutants and wild-type diaphorases fromDiplococcus pneumoniae. J Bacteriol 96: 1029-1036.
Kroger, M., and Hobom, G, (1982) Structural analysis ofinsertion sequence IS5, Nature97: 159-162.
Lund, E. (1950) Antigenic relationship between pneumococciand non-hemolytic streptococci. Acta Pathol MicrobiotScand 27: 110-118.
Lund, E,. and Henrichsen, J. (1978) Laboratory diagnosis,serology and epidemiology of Streptococcus pneumoniae.Meth Microbiot 12: 241-262.
Martin, C, Briese, T,, and Hakenbeck, R. (1992) Nucleotidesequences of genes encoding penicillin-binding proteinsfrom Streptococcus pneumoniae and Streptococcus oratiswith high homology to Escherichia coti penicillin-bindingproteins 1Aand IB, J Bacteriol 17^: 4517-4523.
Messing, J, (1983) New M13 vectors for cloning. MethEnzymot 101: 20-78,
Moore, H.F., and Chesney, A.M. (1917) A study ofethylhydrocupreine (optochin) in the treatment of acutelobar pneumonia. Arch Intem Med 19: 611-682,
Morgenroth, J,, and Kaufmann, M, (1912) Arzneifestigkeit beibakterien (pneumokokken), Z Immunltatsforsch Exp Thar15:610-624,
Morrison, D,A. (1978) Transformation in pneumococcus:protein content of eclipse complex. J Bacteriol 136:548-557,
Mosher, M.E., Peters, LK., and Fillinganne, R,H, (1983) Useof lambda unc transducing bacteriophages in genetic andbiochemical characterization of H*-ATPase mutants ofEscherichia coli. J Bacteriol 156: 1078-1092,
Nagley, P,, Hail, R.M., and Ooi, B.G, (1986) Amino acidsubstitutions in mitochondrial ATPase subunit 9 ofSaccbaromyces cerevisiae leading to oligomycin orventuricidin resistance, FEBSLett1B5: 159-163.
Pease, AA,, Douglas, C.WI,, and Spencer, R,C, (1986)Identifying non-capsulated strains of Streptococcus pneu-moniae isolated from eyes. J Clin Pattiol 39: 871-875,
Phillips, G,, Barker, R., and Brogan, O, (1988) Optochin-resistant Streptococcus pneumoniae. Lancet i\: 281.
Pozzi, G., Oggioni, M,R,, and Tomasz, A. (1989) DNA probefor Identification of Streptococcus pneumoniae. J ClinMicrobiol 27: 370-372,
Romero, A., Lopez, R., and Garcia. P, (1990) Sequence ofthe Streptococcus pneumoniae bacteriophage HB-3 ami-dase reveals high homology with the major host autolysin. JBacteriol 172: 5064-5070,
Ronda, C, Garcia, J,L., and Lopez, R. (1988) Characteriza-tion of genetic transformation in Streptococcus oratis NCTC11427: expression of the pneumococcai amidase inS, oratts using a new shuttle vector. Mol Gen Genef 215:53-57,
Sambrook, J., Fritsch, E.F., and Maniatis, T, (1989) Mol-ecular Cloning: A Laboratory Manuai 2nd edn. CoidSpring Harbor, New York: Coid Spring Harbor LaboratoryPress,
Sanger, F,, Nicklen, S,, and Coulson, AR, (1977) DNAsequencing with chain-terminating inhibitors. Proc NattAcad Sci USA 74: 5463-5467.
Schmidhuber, S,, Ludwing, W,, and Schleiffer, K.H,(1988) Construction cf a DNA probe for the specificidentification of Streptococcus oraiis. J Clin Microbiol 26:1042-1044,
Senior, A,E. (1990) The proton-translocating ATPase ofEscherichia coti Annu Rev Btophys Biophys Chem 19:7-41.
Shibata, C, Ehara, T,, Tomura, K., Igarashi, K., andKobayashi, H. (1992) Gene structure of Enterococcushirae (Streptococcus faecatis) FiFo-ATPase, which func-tions as a regulator of cytoplasmic pH, J Bacteriot 174:6117-6124.
Shine, J., and Dalgarno, L, (1975) Determinants of cistronspecificity in bacterial ribosomes. Nature 254: 34-38,
Spratt, B,G., Hedge, P.J., te Heesen, S,, Edeiman, A,, andBroome-Smith, J,K, (1986) Kanamycin-resistant vectorsthat are analogues of plasmids pUC8, pUC9, pEMBL8 andpEMBL9. Gene 41: 337-342.
Tomasz, A, (1970) Cellular metabolism in genetic trans-formation of pneumococci: requirement for protein

598 A. Fenoli R. Mufioz, E. Garcia and A. G. de la Campa
synthesis during induction of competence. J Bacteriol 101:860-871.
Trombe, M-C, Laneelle, G., and Sicard, A.M. (1984)Characterization of a Streptococcus pneumoniae mutantwith altered electric transmembrane potential. J Bacteriol158: 1109-1114.
Tybulewicz, V.LJ., Falk, G., and Walker, J.E. (1984)Rhodopseudomonas blastica atp operon. Nucieotidesequence and transcription. J Mol Biol 179: 185-214.
Wasilauskas, B.I., and Hampton, K.D. (1984) An analysisof Streptococcus pneumoniae, identification using
biochemical and serological procedures. Diagn Microbioi/n/D/s 2: 301-307.
Yanisch-Perron, C, Vieira, J., and Messing, J. (1985)Improved Ml 3 phage cloning vectors and host strains:nucieotide sequences of the M13mpl8and pUCi9 vectora.Gene 33: 103-119.
Yurchah, A., and Austrian, R. (1966) Serologic andgenetic relationship between pneumococci and otherrespiratory streptococci. Trans Assoc Amer Phys 79:368-375.





















