
Porin expression in clinical isolates of Klebsiella pneumoniae
Upload
independentCategory
view
4download
0
MICROBIAL DRUG RESISTANCEVolume 3, Number 1, 1997Mary Ann Liebert, Inc.
Ubiquitous Distribution of the Competence Related GenescomA and comC Among Isolates of Streptococcus pneumoniae
MARIO RAMIREZ,1 DONALD A. MORRISON,2 and ALEXANDER TOMASZ1
ABSTRACT
DNA probes specific for the pneumococcal competence regulatory genes comA and comC were used to testthe presence of these genes in 214 pneumococcal isolates selected to represent a wide variety of clonal typesand genetic backgrounds (as defined by chromosomal macrorestriction patterns and multilocus enzyme analy-sis), a variety of serotypes, isolation dates (between 1916 and 1996), geographic origins (on four continents),as well as different clinical origins (including both infection sites and colonization sites). Each isolate gave pos-itive signal with both DNA probes. The comA and comC genes were never on the same Sma\ restriction frag-ment and the comA gene showed a considerable degree of polymorphism from one strain to another. WhileDNA sequencing of 50 of the isolates have identified three distinct alíeles of the comC gene, the number ofmutations within the leader peptide were minimal. The results suggest that the mechanism to undergo genetictransformation is widespread within the species of Streptococcus pneumoniae.
INTRODUCTION
Competence, the ability of Streptococcus pneumoniae to
capture DNA molecules from the environment and un-
dergo genetic transformation, has been known to be inducibleby a pheromone—called "activator"25 or "competence fac-tor"24—that regulates the property of competence on the levelof pneumococcal populations in a manner reminiscent of a hor-monal control.23 Recent evidence accumulating in the literaturehas documented the frequent acquisition by pneumococci ofboth homologous as well as heterologous genetic elements suchas determinants of antibiotic resistance and capsular type,2-4 andit has been suggested that the mechanism involving these re-
combinational events may be genetic transformation. However,virtually all experiments on genetic transformation of pneumo-cocci have been done using derivatives of a single pneumo-coccal strain originally isolated by Oswald Avery at The Rock-efeller University.1 Therefore, the presence or absence of thisrecombinational system among natural isolates of pneumococcihas remained an open question. In order to test whether or not
competence could have the postulated role in the epidemiologyof pneumococcus we used two DNA probes: one for the re-
cently-cloned comC gene (determinant of a competence-stim-ulating peptide [CSP] required for competence induction)6 andanother for comA (involved with the release of CSP from theproducer cells)9'10 to test for the distribution of these key com-
ponents of the genetic transformation system among a largenumber of pneumococcal isolates.
MATERIALS AND METHODS
Strains and growth conditions
The pneumoccocal strains used in this study were from TheRockefeller University collection and their relevant propertiesare listed in Tables 1A through 6A. Strain RC052 used as con-
trol for competence induction was constructed by transformingstrain R36A with DNA from the mutant strain CP1415.14 StrainCP1500 carries Streptomycin and Novobiocin resistance mark-ers.13 (Strains B, L, V, N, M, and S are recent clinical isolatesfrom hospitals in the US.) Bacterial cultures were grown in a
semi-synthetic medium (C + Y) described previously." To in-duce competence, synthetic CSPa was added to the medium at
'Laboratory of Microbiology, The Rockefeller University, 1230 York Avenue, New York, New York 10021.laboratory for Molecular Biology, The University of Illinois at Chicago, 900 South Ashland Avenue, Chicago, Illinois 60607
39

40 RAMIREZ ET AL.
250 ng/ml. The antibiotics used for selecting genetic transfor-mants were streptomycin (100 /xg/ml) and novobiocin (5 /ng/ml).
ProbesA comA fragment internal to the coding region of the gene
(288 bp—1951 bp), was obtained through polymerase chain re-action (PCR) using DNA from strain R36A as template andprimers: CTACTATGTGGTGACTGGGCAGG (COMAd) andCCAAACTGCTAGTCGCCTCATC (COMAr). The primerswere designed based on the submitted DNA sequence (acces-sion M36180, L15190) of the comA gene.9
The resulting 1663 bp fragment was cloned into plasmidpCRII and transformed into Escherichia coli strain INVaF' us-
ing the TA cloning kit according to the manufacturer's recom-
mendations (Invitrogen, CA). The identity of the cloned frag-ment was verified by restriction analysis.
An EcoRI band of 1.6 kb was purified after agarose gel elec-trophoresis using the Wizard PCR Preps Purification System(Promega, WI) and labeled with the ECL Random Prime La-beling kit (Amersham, UK) according to the manufacturer'srecommendations.
A comC probe was prepared by labeling a 315 bp fragmentresulting from a PCR reaction using DNA from strain R36A as
template and primers: CAAATAAAATGGTAACTGTG(CSPr) and GACAGTTGAGAGAATTTTATC (CSPd). Theprimers design was based on a sequence of the comC611 geneand neighboring area (accession U33315). The fragment en-
compasses the entire coding region of comC as well as 83 bpupstream and 93 bp downstream. The fragment was purifiedfrom an agarose gel using the Wizard PCR Preps PurificationSystem (Promega, WI) and labeled with the ECL RandomPrime Labeling kit (Amersham, UK) according to the manu-facturer's recommendations.
Table 1A. Streptococcus pneumoniae Isolates with Different Serotypes, Isolation Dates and Geographic Sources
Strain code SerotypeIsolation
year Geographic criginClonal type
(PFGE)D39SMD10789K3K634034A89SA23A112MA 1347A661K15K40N7SP96K41Noc211MD522910545SP108K29SP114117458249LI3186OP5148K10K30K48BSH36A101K8K17K18SP140SP144SP1481138-801139-80Pen6
22334466A6A6B6B9V9N
10A11A141415B15B15B18A19A19A19F19F19F19F19F23F23F23F23F24F33A33F36
nontypablenontypable
rough
1916a1985199019901984198519851982
1985199019901995193819911990-911985
1991
1977
1990199019901996198419901990199019421942194219801980
USASpainSeoul, South KoreaSeoul, South KoreaRepublic of South AfricaUSARepublic of South AfricaUSA
USASeoul, South KoreaSeoul, South KoreaNew York, USAUSASeoul, South KoreaUSASpainUSASeoul, South Korea
Czech RepublicSeoul, South KoreaSeoul, South KoreaSeoul, South KoreaNew York, USAUSASeoul, South KoreaSeoul, South KoreaSeoul, South Korea
USAUSAUSA
ndbndGHndndndnd
ndLN8ndOndndndndIndndndndndAAAndndAAMndndndndndnd
aShaded dates highlight early isolates.bNot done.

GENE COMPETENCE IN S. PNEUMONIAE 41
Table IB. Hybridization of Streptococcus pneumoniae with the DNA Probes comA, comC and Response to CSPa
Strain code Serotype
Chromosomal location(PFGE Smal band) Spontaneous Response Sequence
comA Kb comC Kb competence to CSPa comC
D39SMD10789K3K634034A89SA23A112MAI 347A661K15K40N7SP96K41Noc211MD522910545SP108K29SP114117458249LI3186OP5148K10K30K48BSH36A101K8K17K18SP140SP144SP1481138-801139-80Penó
22334466A6A6B6B9V9N10A11A141415B15B15B18A19A19A19F19F19F19F19F23F23F23F23F24F33A33F36
nontypablenontypable
rough
110969691
107103177 G 153104
99160140
I 116H 104
10397
175101113102
D 214.153
FGFGFGE
EFF
EGFFGH
G 122L93
H 92
98103
9110594
132173
G 177G 145H 149H 88F 150
F 100I 101J 180
132F 102
107F 110H 151 L91
112
FG
129111
C202E 187D 198E 189D 199D202D203
ndD225D202D222E209H 139C206D 191D202E209E202E218E201
nd 110E 115D200H 111D201F 132D205D207D210E 127D219D216
nd 132E
ndEFEE
129
127195196197
LNoneNoneNoneHNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneHNoneHNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneHHIHNoneH
IHLLHNonebLINoneIHIHNoneINoneNoneIHHHIINoneNoneHLLNoneHLNoneHHHIHNonend
ndandndndnd02ya
aa
a
ndnd02nd01a
aandnda
nd02ndndndnd01a
a
ndndnda
ndnd02nd
aNot done.bShading indicates lack of response to CSPa.
Assay of endogenous and CSPa inducible competenceDNA from strain CP1500 was prepared according to a pub-
lished procedure,12 and was used in transformation experimentsat a concentration of 1 pglml. Cultures were grown to station-ary phase and diluted in fresh medium containing the trans-
forming DNA to OD620 < 0.005, corresponding to a viable titerof about 2-5 X 106cfu/ml. CSPa was added to one sample whilethe other received no treatment. Both samples were incubatedat 37°C until the optical density of the culture reached OD62o ""
0.6 (about 6 hours). Transformants were scored by plating on
streptomycin and/or novobiocin containing blood agar plates.
Localization of comA and comC
DNA was prepared, digested by Smal and processed bypulsed-field gel electrophoresis (PFGE) according to a previ-
ously published procedure.21 Transfer of the gels was done withthe VacumGene System (Pharmacia Biotec, NJ) according tothe manufacturer's recommendations. The resulting membraneswere hybridized to comA and comC probes using the ECL sys-tem (Amersham, UK) according to the manufacturer's recom-
mendations. The molecular weight of the hybridization signal(s)and the corresponding Smal fragments were determined.
DNA sequencingDNA for sequencing was prepared according to the same
procedure used for probe preparation. Sequencing was per-formed in a Perkin Elmer/Applied Biosystems model 373 DNAStretch Sequencer by the fluorescence based Taq FS dye ter-minator cycle sequencing method. The sequence of both strandswas determined in most cases using either the CSPr or CSPdprimer. In the case of the single isolate representing the comC-y

Table 2A. Multidrug Resistant Clones of Streptococcus pneumoniae
Strain code Geographic originIsolation
year SerotypePEN MIC
p.g/mlClonal type
(PFGE)
Sp2150Sp2186Sp2235ClevlClev2Clev3Clev4Clev5ZlZ3Z4Z12K2K4K5K7K9K13K14K16K21K23K38K42K49K50NoelNoel 87Noc230
Madrid, Spain 1990Madrid, Spain 1990Madrid, Spain 1990Cleveland, OH, USA 1989Cleveland, OH, USA 1989Cleveland, OH, USA 1989Cleveland, OH, USA 1989Cleveland, OH, USA 1989Zagreb, Croatia 1990-93Zagreb, Croatia 1990-93Zagreb, Croatia 1990-93Zagreb, Croatia 1990-93Seoul, South Korea 1989Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1990Seoul, South Korea 1991Seoul, South Korea 1990Seoul, South Korea 1992Seoul, South Korea 1992Seoul, South Korea 1992Durham, NC, USA 1990-91Durham, NC, USA 1990-91Durham, NC, USA 1990-91
23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F23F
1I1121-221-21-21-21-21-212222221222222222
AAAAAAAAAAAAAAAAAAAAAAAAAAAAA

Table 3A. Multidrug Resistant Clones of Streptococcus pneumoniae
Strain code Geographic originIsolation
year SerotypePEN MIC
p,g/mlClonal type
(PFGE)
K27KllK22K25K28K33K34K39K44K45K46Ic2Ic7Ic8Icl4Ic22Ic24Icl41AlaiAla 16Ala39Ala46Ala55Hunl4315Hun524Hun633CzlOCzl9
Seoul, South Korea 1990Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1991Seoul, South Korea 1992Seoul, South Korea 1992Iceland 1989-91Iceland 1989-91Iceland 1989-91Iceland 1989-91Iceland 1989-91Iceland 1989-91Iceland 1992Anchorage, AK, USA 1982Fairbanks, AK, USA 1986Bethel, AK, USA 1989Bethel, AK, USA 1989Anchorage, AK, USA 1989Hungary 1989Hungary 1989Hungary 1989Czech Republic 1990Czech Republic 1990
19F19F19F19F19F19F19F19F19F19F19F6B6B6B6B6B6B6B6B6B6B6B6B
19A19A19A1419A
212212222221-21-21-21-21-21-210.120.120.250.250.25224100.8
AAAAAAAAAA
BBBBBBBCCCCCDDDEF
Table 3B. Hybridization of Multidrug Resistant Streptococcus pneumoniaewith the DNA Probes comA, comC and Response to CSPa
Chromosomal locationClonal type (PFGE Smal band) Spontaneous Response Sequence
(PFGE) comA Kb comC Kb competence to CSPa comCStraincode Serotype
K27KllK22K25K28K33K34K39K44K45K46Ic2Ic7Ic8Icl4Ic22Ic24Icl41AlaiAla 16Ala39Ala46Ala55Hunl4315Hun524Hun663CzlOCzl9
19F19F19F19F19F19F19F19F19F19F19F6B6B6B6B6B6B6B6B6B6B6B6B
19A19A19A1419A
AAAAAAAAAAABBBBBBBCCCCCDDDEF
FGFFFF
GFGFG
178167164170178170177178177176177
FGFFFCCC
G 154H 140
G 142G 154G 142H 149G 148H 149G 148H 149
G 99nd 101G 99
nd99G 104G 98G 101
104104108106125264264264
I 104G 102
D 216D 215E 207G 202D 213D 206D 204D 216D 207D 209D 207C 213
nd 213C 216
nd ndC 213E 213C 211D 211E 208D 217D 217D 204E 76E 76E 78E 213E 203
NoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNonendLLNoneNoneNoneINoneNoneNoneNone
LLLLNonebLIINoneLNoneLILILNonendHLHINoneHIHNoneNone
01ndandndndndndndndndndnda
ndndndndndndndndnda
nda
nda
nd
'Not done; bShading indicates lack of response to CSPa.
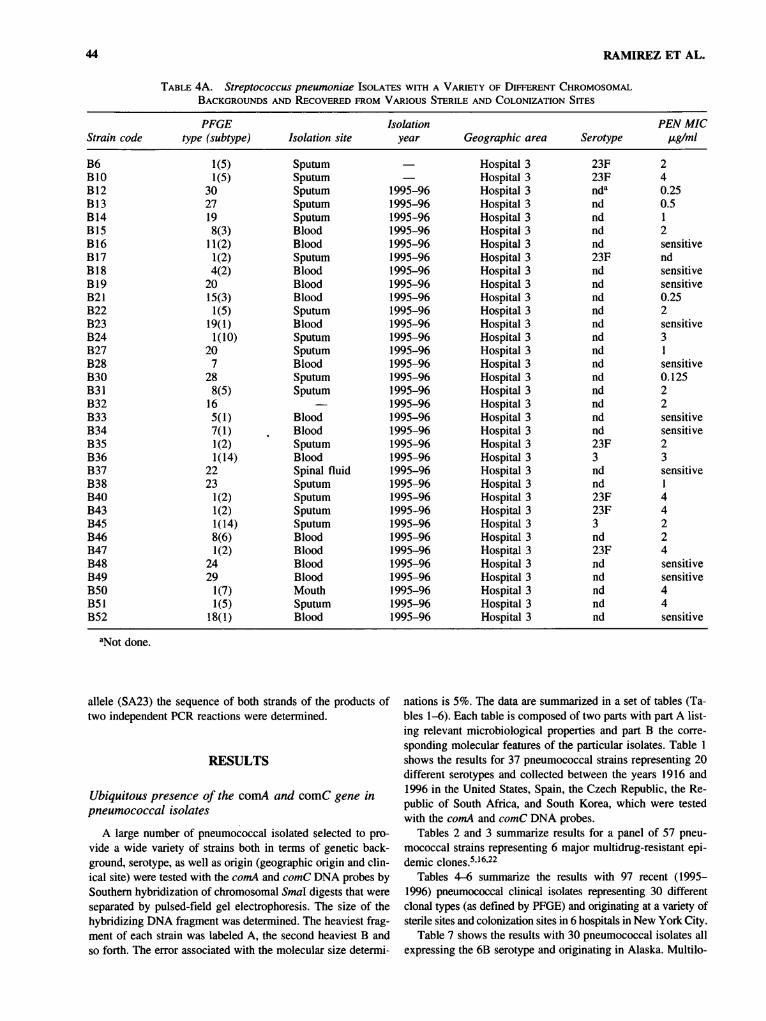
44 RAMIREZ ET AL.
Table 4A. Streptococcus pneumoniae Isolates with a Variety of Different ChromosomalBackgrounds and Recovered from Various Sterile and Colonization Sites
Strain codePFGE
type (subtype) Isolation siteIsolation
year Geographic area SerotypePEN MIC
p.g/mlB6BIOB12B13B14B15B16B17B18B19B21B22B23B24B27B28B30B31B32B33B34B35B36B37B38B40B43B45B46B47B48B49B50B51B52
1(5)1(5)
302719
8(3)11(2)
1(2)4(2)
2015(3)
1(5)19(1)
1(10)20
728
8(5)16
5(1)7(1)1(2)1(14)
2223
1(2)1(2)1(14)8(6)1(2)
2429
1(7)1(5)
18(1)
SputumSputumSputumSputumSputumBloodBloodSputumBloodBloodBloodSputumBloodSputumSputumBloodSputumSputumBloodBloodSputumBloodSpinal fluidSputumSputumSputumSputumBloodBloodBloodBloodMouthSputumBlood
1995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-961995-96
HospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospital 3Hospital 3Hospital 3Hospital 3Hospital 3HospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospital
23F23Fndandndndnd23Fndndndndndndndndndndndndnd23F3ndnd23F23F3nd23Fndndndndnd
240.250.5
12sensitivendsensitivesensitive0.252sensitive31sensitive0.12522sensitivesensitive23sensitive144224sensitivesensitive44sensitive
aNot done.
alíele (SA23) the sequence of both strands of the products oftwo independent PCR reactions were determined.
RESULTS
Ubiquitous presence of the comA and comC gene inpneumococcal isolates
A large number of pneumococcal isolated selected to pro-vide a wide variety of strains both in terms of genetic back-ground, serotype, as well as origin (geographic origin and clin-ical site) were tested with the comA and comC DNA probes bySouthern hybridization of chromosomal Smal digests that were
separated by pulsed-field gel electrophoresis. The size of thehybridizing DNA fragment was determined. The heaviest frag-ment of each strain was labeled A, the second heaviest B andso forth. The error associated with the molecular size determi-
nations is 5%. The data are summarized in a set of tables (Ta-bles 1-6). Each table is composed of two parts with part A list-ing relevant microbiological properties and part B the corre-
sponding molecular features of the particular isolates. Table 1shows the results for 37 pneumococcal strains representing 20different serotypes and collected between the years 1916 and1996 in the United States, Spain, the Czech Republic, the Re-public of South Africa, and South Korea, which were testedwith the comA and comC DNA probes.
Tables 2 and 3 summarize results for a panel of 57 pneu-mococcal strains representing 6 major multidrug-resistant epi-demic clones.5-16-22
Tables 4-6 summarize the results with 97 recent (1995-1996) pneumococcal clinical isolates representing 30 differentclonal types (as defined by PFGE) and originating at a variety ofsterile sites and colonization sites in 6 hospitals in New York City.
Table 7 shows the results with 30 pneumococcal isolates allexpressing the 6B serotype and originating in Alaska. Multilo-

GENE COMPETENCE IN S. PNEUMONIAE 45
Table 4B. Hybridization of Streptococcus pneumoniae with the DNA Probes comA, comC and Response to CSPa
Chromosomal locationStrain Isolation PFGE (PFGE Smal band) Spontaneous Response Sequencecode site type (subtype) comA Kb comC Kb competence to CSPa comC
B6BIOB12B13B14B15B16B17B18B19B21B22B23B24B27B28B30B31B32B33B34B35B36B37B38B40B43B45B46B47B48B49B50B51B52
SputumSputumSputumSputumSputumBloodBloodSputumBloodBloodBloodSputumBloodSputumSputumBloodSputumSputumBloodBloodSputumBloodSpinal fluidSputumSputumSputumSputumBloodBloodBloodBloodMouthSputumBlood
1(5)1(5)
302719
8(3)11(2)
1(2)4(2)
2015(3)
1(5)19(1)
1(10)20
728
8(5)165(1)7(1)1(2)1(14)
2223
1(2)1(2)1(14)8(6)1(2)
2429
1(7)1(5)
18(1)
F 177F 177F 162F 129F 112G 108F 161F 180G 164F 94F 116G 154H 111G 153F 102G 92F 127F 103F 174G 97F 101F 150F 150E97E 152F 150F 153F 153F 99F 153F 101G 90G 177G 153F 101
G 148G 148
93
G 151
H 86H 88G 78
H 86H 90H 90
H 90G 92
I 90
D205D207G138D219D219F 143D202D206E207D204E 193D210F 209E211E 131D 193D207E 140C 222D202D204D210D210C 204G 109D 207D212D 209E 142D 209D206E 129E209D212E 121
NoneNoneNoneNoneNoneNoneNoneNoneNoneNoneLNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNone
NoneaNoneIndHHHLHNoneHIHNoneHHLHHINoneNoneNoneHNoneNoneNoneLNoneNoneNoneLLNoneNone
0101ndbndndndndndnd02ndnda
01ndndndndndndndnd01ndndndndndndndndndndndnd
"Shading indicates lack of response to CSPa.bNot done.
cus enzyme analysis indicated that these strains represent a con-
siderable variation of the genetic backgrounds.Each of the 214 strains listed through Tables 1-7 gave pos-
itive hybridization signal with both the comA and comCprobes.
Strain to strain variation in the molecular size of DNAfragments hybridizing with comA and comC
Tables 1-7 illustrate the considerable variation in the mole-cular size of the comA hybridizing DNA fragments. More thanone comA hybridizing fragment was identified in a number ofstrains. Considerably less variation was observed in the molec-ular size of the comC reacting DNA fragments. The spread ofthese fragment sizes is more graphically illustrated by Figures1-3.
Multiple alíeles of the comC geneFor fifty of the pneumococcal strains examined the sequence
of the comC gene was determined. Three alíeles: comCa, comCßand a new alíele, comCy were identified. The comparison of se-
quences is shown in Figure 4. The genes comCa and comCßare identical to comCl and comC2 described by Pozzi et a/.18
Response of pneumococcal isolates to the comCapeptide
Each of the 214 pneumococcal isolates was also tested forresponsiveness to the competence-stimulating activity of syn-thetic CSPa, used at a level that saturates the response in Rxand R36A.6 In parallel, the capacity of the same strains for de-velopment of spontaneous competence during a cycle of

46 RAMIREZ ET AL.
Table 5A. Streptococcus pneumoniae Isolates with a Variety of Different ChromosomalBackgrounds and Recovered from Various Sterile and Colonization Sites
Strain codePFGE
type (subtype) Isolation siteIsolation
year Geographic area SerotypePEN MIC
p,g/mlB53B54B56B57B58LILIOLUL12L13L14L15L16L17L18L19L2L20L21L24L25L26L27L3L4L5L6L7L8L9MlM2M3M4M5
1(5)25
8(4)19(2)
261
101
1111
1(8)12
11(1)1(12)1(9)
1(10)1
1(11)1(10)
16171711
11
1(3)1(7)1(2)
11983
1(4)1(4)
SputumBloodBloodBloodBlood
NasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingeal
BloodSputum
EarSputumSputum
19951995199519951995199519951995199519951995199519951995199519951995199519951995199519951995199519951995199519951995199519941994199419941994
HospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospitaHospita'Hospita!HospitaHospita!HospitaHospitaHospitaHospita!HospitaHospitaHospita!HospitaHospitaHospita
ndandndndnd232323
66
non-typable106
non-typable2323232323
91010
623232323232323ndndndndnd
0.250.120.5
sensitivesensitive
40.1251.50.0940.09410.0230.0640.1942231.50.0940.0230.0230.0471.511.51.51.523
ndndndndnd
aNot done.
growth, selected as optimal for the expression of competencein strain R6,24 was also determined. As shown in the tables andin Figure 5, individual strains showed considerable variationboth with respect to their spontaneous competence and also withrespect to their degree of responsiveness to CSPa. The rangeof response included no detectable response and a spectrum ofvariation from low (transformation rate 10"8) through inter-mediate to high (transformation rate 10~3) response.
DISCUSSION
The great majority of penicillin resistant and multidrug re-
sistant 5. pneumoniae strains express only a few (about 5 to 6)
of the 90 pneumococcal polysaccharides and only a relativelysmall group of the very large number of the multidrug-resistantclones appear to be epidemic, i.e., capable of expansion over
large geographic areas.15'21 It was a priori conceivable thatthese phenomena were related to the uneven distribution ofcompetence related genes among natural isolates of pneumo-cocci, since acquisition of drug resistance traits as well as vir-ulence determinants may occur by recombinational events.4
However the observations described in this paper stronglysuggest that the capacity to undergo genetic transformation isa ubiquitous property within the species of S. pneumoniae. Twoimportant genetic elements that are indispensable for the ex-
pression of this property were found to be uniformly present inpneumococcal isolates irrespective of their isolation dates,genetic backgrounds, serotypes, and other relevant microbio-

GENE COMPETENCE IN S. PNEUMONIAE 47
Table 5B. Hybridization of Streptococcus pneumoniae with the DNA Probes comA, comC and Response to CSPa
Strain PFGEcode Isolation site type (subtype)
Chromosomal location(PFGE Smal band) Spontaneous Response Sequence
comA Kb comC Kb competence to CSPa comC
B53B54B56B57B58LILIOLUL12L13L14L15L16L17L18L19L2L20L21L24L25L26L27L3L4L5L6L7L8L9MlM2M3M4M5
SputumBloodBloodBloodBloodNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealNasopharingealBloodBputumEarSputumSputum
1(5)25
8(4)19(2)26
110
11111
1(8)1211(1)
1(12)1(9)1(10)11(11)1(10)
16171711
111(3)1(7)1(2)119831(4)1(4)
GGF
163103110
I 90
nd 132 nd 106
H 152
G 149
FGGFFFF
FFGGF
GFGF
103181102175156156192
G 96F 159H 100
180151178139148
H 107G 97
98159179178
H 101G 151
G 163
G 152
H 150
FFFG
152178181109
H 107G 101G 179G 182
H 152G 150
G 150G 154
H 149H 154
D206C 194E 143
nd201D 189E211E208E209D204D204D208E213D206D208E209E207E210E205E208D218E213E213D202E210E210E211E210D211E210E211E217F 142E208D211D211
NoneNoneNoneNoneNoneNoneNoneLNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneNoneINoneNoneNoneNone
None"HHNoneILNoneNoneINoneNoneNoneNoneNoneLNoneNoneNoneNoneHNoneNoneHNoneNoneNoneNoneNoneNoneNoneHIHLNone
ndbndndndnd01ndndndndnd02ndndndndndnd01ndndndnd02ndndndndndndndnda
nd01
"Shading indicates lack of response to CSPa.bNot done.
logical differences among the strains. Mutations in the leaderpeptide of ComC were minimal which is consistent with theidea that this part of the molecule is essential for the properrecognition and processing of ComC by its cognate transporter(ComA).6-7 This finding is compatible with the notion that thecomC dependent pathway of genetic exchange is not only ubiq-uitous but is actually widely "used" in pneumococci for cat-
alyzing genetic exchange.The 0 alíele of comC was identified with two different leader
peptides (lysine to aspargine substitution at aminoacid 20 ofthe N terminus). This substitution was shared by the y alíelethat had another nonconserved substitution in the leader pep-
tide at residue 22 (lysine to glutamine). All alíeles share thefour residues conserved among more than 90% of the knownmembers of the double glycine family of processed secretedpeptides,7 but it is not clear at present whether the observedsubstitutions interfere with the selective recognition and pro-cessing of ComC or if they have corresponding allelic varia-tions in the comA locus. The variation in the signaling peptideis much more pronounced. Although there are only three non-
conserved substitutions in the a and 0 alíeles the differencesin the y alíele are much more extensive. The functionality ofthe ComC y signaling peptide remains an open question butit is possible that matching receptors may exist for the differ-

Table 6A. Streptococcus pneumoniae Isolates with a Variety of Different ChromosomalBackgrounds and Recovered from Various Sterile and Colonization Sites
Strain codePFGE
type (subtype) Isolation siteIsolation
year Geographic area SerotypePEN MIC
p.g/mlNlN10NilN12N2N3N4N5N6N7N8N9RlR2R4R7R9S2S4S5S6S7VIV2V3V5AV5B
141(2)
1718
8(1)1(13)4(1)
1515(1)
8(2)16
1(6)2(1)2(2)2(1)2(3)2(4)76543
131313(1)
1(4)1(4)
Eye
BloodBloodSputumSputumBloodCsfSputumTrachéal asp.
EarEarEarThroatThroatSputumBloodBloodBloodBloodCsfCsfCsfBloodCsf
1994
1994199419941994199519951995
199419941994199419941994199419941994199419931993199319931993
Hospital 2Hospital 2HospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalPrivate ClinicPrivate ClinicPrivate ClinicPrivate ClinicPrivate ClinicHospital 6HospitalHospitalHospitalHospitalHospitalHospitalHospitalHospitalHospital
15ndandnd
91919
9149N19nd
non-typablenon-typable
6Bnon-typable
ndndndndndndndndndndnd
2ndndnd220.120.120.1222ndndndndndndndndndndnd20.250.06ndnd
"Not done.
Table 6B. Hybridization of Streptococcus pneumoniae with the DNA Probes comA, comC and Response to CSPa
Strain PFGEcode Isolation site type (subtype)
Chromosomal location(PFGE Smal band) Spontaneous Response Sequence
comA Kb comC Kb competence to CSPa comC
NlN10NilN12N2N3N4N5N6N7N8N9RlR2R4R7R9S2S4S5S6S7VIV2V3V5AV5B
eye
bloodbloodsputumsputumbloodcsfsputumtrachea! asp.
earearthroatthroatsputumbloodbloodbloodbloodcsfcsfcsfbloodcsf
141(2)
17188(1)1(13)4(1)
1515(1)8(2)
161(6)2(1)2(2)2(1)2(3)2(4)76543
131313(1)
1(4)1(4)
EGHF
GGFFFHFFF
GF
97159104100101134148101103104163132150101150 G 99
G 99
nd 96H 99FGFFGFF
991031551506410199
G 99G 144G 144
D215D 203G 115E 117F 139D215D 197F 139G 139H 139C 207D 204D 198D 244D 200F 200E 202D 205F 140D 204D 204E 204E 128E 128F 128D 207D 207
NoneNoneNoneNoneNoneNoneHNoneNoneNoneNoneNoneNoneLNoneNoneNoneNoneNoneNoneNoneNoneLNoneNoneNoneNone
None"NoneHNoneHLHNoneHHHNoneIHLNoneNoneHHLNoneHHHHLI
andbndndndndndndndndndndnda
ndndndndndnda
ndndnd
02nd
'Shading indicates lack of response to CSPa; bNot done.

GENE COMPETENCE IN S. PNEUMONIAE 49
Table 7. comA and comC Genes of Streptococcus pneumoniae Isolates with Different Genetic Backgrounds
Clonal Alíeles Chromosomal location ResponseStrain type PspA different PFGE Smal band Spontaneous to Sequencecode (ET) type (type A) comA Kb comC Kb competence CSPa comC
Ala26Ala37Ala44Ala6Ala 13Ala43Ala24Ala38Ala5Alai 1Ala3Ala7Ala29Ala41Ala40Ala54Ala36Ala35Ala20Alal7Ala 10Ala55
BBCCDDEEEFGHIJNOPQRTUWXYS
19191916
019
91916
000
191634
012
031351616
F 106G 110F 106F 99G 104
106106 G 1081089998
F 99F 99
114127125123126108135161160125
D 205E210D 207D 200F 202C 217D215E210E211D 207D216D200D208D 204D 207D 204D 202D221D205D202D 203D 204
NoneILNoneLNoneHLLLNoneINoneNoneNoneNoneNoneNoneNoneNoneNoneNone
HHLNonebHHHIHHHHHHHIIHNoneHLNone
nd"ndndndndndndandndndndndndndndndndndndnd
"Not done.bShading indicates lack of response to CSPa.
ent comC alíeles. Evidence for such complementary inpheromone/receptor interactions have recently been describedin S. sanguis?
Sequencing of the comC from additional isolates will be nec-
essary to find a possible epidemiological correlate of the dif-ferent comC alíeles. It seems that the variation in this locus isindependent from the general chromosomal background sincestrains sharing common PFGE background were found to oc-
casionally carry different alíeles of comC.The presence of an intact comA gene has been shown to be
necessary for the release of CSP from the bacteria, which inturn is known to be essential for the triggering of the autocat-
alytic process leading to the rapid spread of the competent phys-iology to a bacterial population.3-23-24 An examination of themolecular sizes of comA and comC hybridizing DNA fragmentsin the various pneumococcal isolates shows that these two geneswere never on the same Smal fragment; moreover in strain R6comC localizes to fragment 5 which puts at least 25 Kb be-tween the two loci. This suggests a complex control for the ex-
pression of the competent physiology. In addition, two bandshave been identified by high stringency hybridization with thecomA probe in several of the strains. It is not clear at presentwhether this results from the presence of a Smal site internalto the probe or to the presence of homologous or duplicatecopies of comA. It is conceivable that comA also participates inthe control of export phenomena other than the release of thecomC gene product.
Different pneumococcal strains showed a wide variety of re-
MW 12 3 4 5 6 7 8 MW291Kb
2425 Kb
194 Kb
145.5 Kb
97 Kb
4825 Kb
FIG. 1. Hybridization patterns of comC reacting DNA frag-ments in Smal digests of pneumococci. Figure 1 illustrates thevariation in the molecular sizes oicomC hybridizing DNA frag-ments as observed by the analysis of a select group of strains.

50
MW 12 3 4 5 6 7 9 10 11 MW
291Kb
242.5 Kb
194 Kb
145.5 Kb
97 Kb
485 Kb
FIG. 2. Hybridization patterns of comA reacting DNA frag-ments in Smal digests of pneumococci. Variation in the mole-cular sizes of comA hybridizing DNA fragments as observedby the analysis of a select group of strains that showed two hy-bridizing bands.
MW 12 3 4 5 6 7
RAMIREZ ET AL.
9 10 11 MW291Kb
242.5 Kb
194 Kb
145.5 Kb
97 Kb
485 Kb
FIG. 3. Hybridization patterns of comA reacting DNA frag-ments in Smal digests of pneumococci. Variation in the mole-cular sizes of comA hybridizing DNA fragments as observedby the analysis of strains that showed single comA hybridizingband.
sponses when tested with the CSPa synthetic peptide. The rea-
sons for the lack of activation of cells to competence underthese conditions are not clear. At least in some cases it appearsthat bacterial strains that did not respond to CSPa carried a dif-ferent alíele of comC suggesting that for successful interactionwith the peptide pheromone the bacteria also have to carry a
matching receptor as discussed above.One should also remember, however, that the competent
physiology of pneumococci is a complex phenotype and lackof appearance of competence assayed by the success of the cellsin undergoing genetic transformation with a genetically-markedDNA may be caused by a block in any one of a number of steps
in the pathway of DNA binding, transport and recombinationthat have already been identified in this bacterium.13-19-20 A case
observed in this study may be the "genetically incompetent"strain RA724 which although carrying the comA and comCagenes remained insensitive to the addition of CSPa to themedium.
The ubiquitous presence of the quorum-sensing gene comCin pneumococci indicates that genetic transformation as a mech-anism for exchange and acquisition of genetic elements may bewidely available within the species of S. pneumoniae, and thatthis mechanism may indeed be involved with the global spreadof antibiotic-resistant clones of pneumococci.
a
ßl
ß2
Y
Leader Peptide
MKNTVKLEQFVALKEKDLQMKNTVKLEQFVALKEKDLQMKNTVKLEQFVALKEKDLQMKNTVKLEQFVALKEKDLQ
KŒKGGEMRLSKFFRbFŒLQRKKI Signaling Peptide
EKGGEMRLSKGGEMRLS
RIIL
RIILDFDFL
ni|q|ggemr[kmneksfniLFL
RKK
RKK
FNFFNFFNFFRRR
FIG. 4. DNA sequence of comC alíeles. DNA sequences were determined as described in Materials and Methods. Identicalresidues are matched by boxes. Underlined residues are conserved in more than 90% of the members of the double glycine fam-ily.

GENE COMPETENCE IN S. PNEUMONIAE 51
10H
& 10
XI2'>
i 10essE
.o
10
DNoCSPa
BCSPa(250ng/ml)
BSH19None (ß)
SP2235
Low (a)
IC7
Intermediate (a)
BSH 12
High (a?)
FIG. 5. Response of clinical isolate to treatment with CSPa—in vitro. The numbers of spontaneous and CSPa induced trans-formants were determined as described in Materials and Methods.
ACKNOWLEDGMENTS
The authors wish to thank Elena Severina for technical as-
sistance in obtaining the PFGE results of part of the strains.Partial support for these studies was received from grant no.
ROI AI 16794 of the National Institutes of Health. M.R. is on a
fellowship from the Gulbenkian Foundation (PGDBM) andFundaçâo Luso Americana para o Desenvolvimento (FLAD).
REFERENCES
1. Avery, O.T., CM. MacLeod, and M. McCarty. 1944. Studieson the chemical nature of the substance inducing transformation ofpneumococcal types. J. Exp. Med. 79:137-158.
2. Barnes, D.M., S. Whittier, P.H. Gilligan, S. Soares, A. Tomasz,and F.W. Henderson. 1995. Transmission of multidrug-resistantserotype 23F Streptococcus pneumoniae in group day care: Evi-dence suggesting capsular transformation of the resistant strain invivo. J. Infect. Dis. 171:890-896.
3. Chandler, M.S., and D.A. Morrison. 1987. Competence for ge-netic transformation in Streptococcus pneumoniae: Molecularcloning of com, a competence control locus. J. Bacteriol. 169:2005-2011.
4. Dowson, C.G., A. Hutchison, J.A. Brannigan, R.C. George, D.Hansman. J. Linares, A. Tomasz, J.M. Smith, and B.G. Spratt.1989. Horizontal transfer of penicillin-binding protein genes inpenicillin-resistant clinical isolates of Streptococcus pneumoniae.Proc. Nati. Acad. Sei. USA 86:8842-8846.
5. Figueiredo, A.M.S., R. Austrian, P. Urbaskova, L.A. Teixeira,and A. Tomasz. 1995. Novel penicillin-resistant clones of Strep-tococcus pneumoniae in the Czech Republic and in Slovakia. Mi-crobial Drug Resistance. 1:71-78.
6. Havarstein, L.S., G. Coomaraswamy, and D.A. Morrison. 1995.An unmodified heptadecapeptide pheromone induces competencefor genetic transformation in Streptococcus pneumoniae. Proc.Nati. Acad. Sei. USA 92:11140-11144.
7. Havarstein, L.S., D.B. Diep, and I.F. Nes. 1995. A family of bac-teriocin ABC transporters carry out proteolytic processing of theirsubstrates concomitant with export. Molec. Microbiol. 16:229-240.
8. Havarstein, L.S., P. Gaustad, I.F. Nes, and D.A. Morrison.1996. Identification of the streptococcal competence-pheromonereceptor. Molec. Microbiol. 2:863-869.
9. Hui, F.M., and D.A. Morrison. 1991. Genetic transformation inStreptococcus pneumoniae: nucleotide sequence analysis showscomA, a gene required for competence induction, to be a memberof the bacterial ATP-dependent transport protein family. J. Bacte-riol. 173:372-381.Hui, F.M., L. Zhou, and D.A. Morrison. 1995. Competence forgenetic transformation in Streptococcus pneumoniae: Organizationof a regulatory locus with homology to two lactococcin A secre-
tion genes. Gene 153:25-31.Lacks, S., and R.D. Hotchkiss. 1960. A study of the genetic ma-
terial determining an enzyme activity in pneumococcus. Biochim.Biophys. Acta. 39:508-517.
12. Marmur, J. 1961. A procedure for isolation of deoxiribonucleicacid from microorganisms. J. Molec. Biol. 3:208-218.
13. Morrison, D.A., S.A. Lacks, W.R. Guild, and J.M. Hageman.1983. Isolation and characterization of three new classes of trans-
id.
11

52 RAMIREZ ET AL.
formation-deficient mutants of Streptococcus pneumoniae that are
defective in DNA transport and genetic recombination. J. Bacte-riol. 156:281-290.
14. Morrison, D.A., M.C. Trombe, M.K. Hayden, G.A. Waszak,and J.D. Chen. 1984. Isolation of transformation-deficient Strep-tococcus pneumoniae mutants defective in control of competence,using insertion-duplication mutagenesis with the erythromycin re-
sistance determinant of pAM beta 1. J. Bacteriol. 159:870-876.15. Munoz, R., T.J. Coffey, M. Daniels, CG. Dowson, G. Laible,
J. Casal, R. Hakenbeck, M. Jacobs, J.M. Musser, B.G. Spratt,and A. Tomasz. 1991. Intercontinental spread of a multiresistantclone of serotype 23F Streptococcus pneumoniae. J. Infect. Dis.164:302-306.
16. Munoz, R., J.M. Musser, M. Crain, D.E. Briles, A. Marion, AJ.Parkinson, U. Sorensen, and A. Tomasz. 1992. Geographic dis-tribution of penicillin-resistant clones of Streptococcus pneumo-niae: Characterization by penicillin-binding protein profile, surfaceprotein A typing, and multilocus enzyme analysis. Clin. Infect. Dis.15:112-118.
17. Pestova, E.V., L.S. Havarstein, and D.A. Morrison. 1996. Reg-ulation of competence for genetic transformation in Streptococcuspneumoniae by an auto-induced peptide pheromone and a two-
component regulatory system. Molec. Microbiol. 21:853-862.18. Pozzi, G., L. Másala, F. Iannelli, R. Manganelli, L.S.
Havarstein, L. Piccoli, D. Simon, and D.A. Morrison. 1996.Competence for genetic transformation in encapsulated strains ofStreptococcus pneumoniae: Two allelic variants of the peptidepheromone. J. Bacteriol. 178:6087-6090.
19. Priebe, S.D., S.M. Hadi, B. Greenberg, and S.A. Lacks. 1988.Nucleotide sequence of the hexA gene for DNA mismatch repairin Streptococcus pneumoniae and homology of hexA to mutS of
Escherichia coli and Salmonella typhimurium. J. Bacteriol. 170:190-196.
20. Puyet, A., B. Greenberg, and S.A. Lacks. 1990. Genetic andstructural characterization of endA. A membrane-bound nucleaserequired for transformation of Streptococcus pneumoniae. J. Molec.Biol. 213:727-738.
21. Soares, S., K.G. Kristinsson, J.M. Musser, and A. Tomasz.1993. Evidence for the introduction of a multiresistant clone ofserotype 6B Streptococcus pneumoniae from Spain to Iceland inthe late 1980s. J. Infect. Dis. 168:158-163.
22. Tarasi, A., N. Sterk-Kousmanovic, K. Sieradzki, S. Schoen-wald, R. Austrian, and A. Tomasz. 1995. Penicillin-resistant nadmultidrug resistant Streptococcus pneumoniae in a pédiatrie hos-pital in Zagreb, Croatia. Microbial Drug Resistance. 1:169-176.
23. Tomasz, A. 1965. Control of competent state in Pneumococcus bya hormone-like cell product: An example for a new type of regu-latory mechanism in bacteria. Nature 208:155-159.
24. Tomasz, A. 1966. Model for the mechanism controlling the ex-
pression of competent state in Pneumococcus cultures. J. Bacte-riol. 91:1050-1061.
25. Tomasz, A., and J.L. Mosser. 1966. On the nature of the pneu-mococcal activator substance. Proc. Nati. Acad. Sei. USA 55:58-66.
Address reprint requests to:Alexander Tomasz
Laboratory of MicrobiologyThe Rockefeller University
1230 York Ave.New York, NY 10021

This article has been cited by:
1. Mario RamirezStreptococcus pneumoniae 1529-1546. [CrossRef]2. Carina Valente, Hermínia De Lencastre, Raquel Sá-Leão. 2012. Pherotypes of Co-Colonizing Pneumococci Among
Portuguese Children. Microbial Drug Resistance 18:6, 550-554. [Abstract] [Full Text HTML] [Full Text PDF] [FullText PDF with Links]
3. Heather Maughan, Rosemary J. Redfield. 2009. EXTENSIVE VARIATION IN NATURAL COMPETENCE INHAEMOPHILUS INFLUENZAE. Evolution 63:10.1111/evo.2009.63.issue-7, 1852-1866. [CrossRef]
4. Jean-Pierre Claverys, Leiv S. Håvarstein. 2007. Cannibalism and fratricide: mechanisms and raisons d'être. NatureReviews Microbiology 5, 219-229. [CrossRef]
5. Jean-Pierre Claverys, Marc Prudhomme, Bernard Martin. 2006. Induction of Competence Regulons as a GeneralResponse to Stress in Gram-Positive Bacteria. Annual Review of Microbiology 60, 451-475. [CrossRef]
6. Jean-Pierre Claverys, Bernard Martin. 2003. Bacterial ‘competence’ genes: signatures of active transformation, or onlyremnants?. Trends in Microbiology 11, 161-165. [CrossRef]
7. Lam-Son Phan Tran, Toshiro Nagai, Yoshifumi Itoh. 2000. Divergent structure of the ComQXPA quorum-sensingcomponents: molecular basis of strain-specific communication mechanism in Bacillus subtilis. Molecular Microbiology37, 1159-1171. [CrossRef]
8. Regine Hakenbeck, Thorsten Grebe, Dorothea Zahner, Jeffry B. Stock. 1999. beta-Lactam resistance in Streptococcuspneumoniae: penicillin-binding proteins and non-penicillin-binding proteins. Molecular Microbiology 33, 673-678.[CrossRef]
9. A TOMASZ. 1999. New faces of an old pathogen: emergence and spread of multidrug-resistant Streptococcuspneumoniae*1. The American Journal of Medicine 107, 55-62. [CrossRef]
10. 1998. Molecular Characterization of Penicillin-Resistant Streptococcus pneumoniae Isolates Causing Respiratory Diseasein the United States. Microbial Drug Resistance 4:4, 325-337. [Abstract] [Full Text PDF] [Full Text PDF with Links]
11. R.Dwayne Lunsford. 1998. Streptococcal Transformation: Essential Features and Applications of a Natural GeneExchange System. Plasmid 39, 10-20. [CrossRef]
12. Mirjana Nesin, Anatoly Severin, Alexander Tomasz. 1997. Stability of clonally related DNA fingerprints and cell-wallpeptide patterns in geographic isolates of multiresistant epidemic clones of Streptococcus pneumoniae. InternationalJournal of Infectious Diseases 2, 91-98. [CrossRef]
Copyright © 2022 FDOKUMEN




















