
Molecular Analysis of Deletions of the Short Arm of Chromosome9 in Human Gliomas
8
1992;52:2523-2529. Cancer Res Olufunmilayo I. Olopade, Robert B. Jenkins, David T. Ransom, et al. Chromosome 9 in Human Gliomas Molecular Analysis of Deletions of the Short Arm of Updated version http://cancerres.aacrjournals.org/content/52/9/2523 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications on March 11, 2014. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on March 11, 2014. © 1992 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Transcript of Molecular Analysis of Deletions of the Short Arm of Chromosome9 in Human Gliomas

1992;52:2523-2529. Cancer Res Olufunmilayo I. Olopade, Robert B. Jenkins, David T. Ransom, et al. Chromosome 9 in Human GliomasMolecular Analysis of Deletions of the Short Arm of
Updated version
http://cancerres.aacrjournals.org/content/52/9/2523
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

(CANCER RESEARCH 52. 2523-2529. May I. 1992)
Molecular Analysis of Deletions of the Short Arm of Chromosome 9 in HumanGliomas'
Olufunmilayo I. Olopade,2 Robert B. Jenkins, David T. Ransom, Kathleen Malik, Helen Pomykala, Tsutomu Nobori,
Janet M. Cowan, Janet D. Rowley, and Manuel O. DiazSection of Hematology/Oncology, Department of Medicine, University of Chicago, Chicago, Illinois 60637 [O. I. O., K. M., H. P., J, D. R„M. O. D.]; Section ofLaboratory Genetics, Department of Laboratory Medicine and Pathology, Mayo Clinic and Mayo Foundation, Rochester, Minnesota 55905 [R. B. J., D. T. R.J;Department of Paediatrics/Genetics, Virginia Medical School. Norfolk, Virginia 23507 [J. M. C.]; and Department of Medicine, University of California, San Diego,California 92093 ¡T.N.]
ABSTRACT
Previous studies have suggested that structural abnormalities involvingthe short arm of chromosome 9 are frequently associated with gliomas.The a-, 0-, and u-interferon (IFNA, IFNB1, and IFNW, respectively)and the methylthioadenosine phosphorylase (Ml.II') genes have been
mapped to the short arm of chromosome 9, band p22. Homozygousdeletions of these genes have been reported in many leukemia- andglioma-derived cell lines. In this report, we present a detailed analysis ofpartial and complete homozygous or hemizygous deletions of DNAsequences on 9p in human cell lines and primary tumor samples of gliomapatients. Ten of 15 (67%) glioma-derived cell lines had hemizygous orhomozygous deletion of / / \ genes or rearrangement of sequences aroundthese genes, while 13 of 35 (37%) primary glioma tumor samples hadhemizygous (8 tumors) or homozygous (5 tumors) deletion of the lf'\
genes. The shortest region of overlap of these deletions maps in theinterval between the centramene end of the II \ gene cluster and theMI AT gene. In the cell lines and primary tumors examined, these grossgenomic alterations were seen only in association with high grade orrecurrent gliomas. Our observations confirm that loss of DNA sequenceson 9p, particularly the II \ genes, occurs at a significant frequency ingliomas, and may represent an important step in the progression of thesetumors. These results are consistent with a model of tumorigenesis inwhich the development or progression of cancer involves the loss orinactivation of a gene or several genes that normally act to suppresstumorigenesis. One such gene may be located on 9p; this gene may beclosely linked to the IFN genes. Nevertheless, loss of the /F7V genes,when it occurs, may play an additional role in the progression of thesetumors.
INTRODUCTION
Tumor suppressor genes were first identified and mapped bythe observation of frequent deletion of specific chromosomalregions in tumor cells, which was later confirmed by molecularanalysis. The deletions of band 13ql4, which were observed innormal somatic and tumor cells of some patients with retino-blastoma, prompted investigators to perform detailed molecularanalysis of this region of the chromosome (1,2). This eventuallyled to the cloning of the RBI gene (3, 4). The retinoblastomagene, as well as other putative tumor suppressor genes, havenow been implicated in other types of neoplasia in adults,including renal cell carcinoma (5), small cell lung cancer (6),colorectal cancers (7), and osteosarcomas and other sarcomas(8). Recent cytogenetic and molecular studies have indicatedthat deletions of part or all of chromosomes 8, 9, 10, 17, and
Received 10/23/91; accepted 2/19/92.The costs of publication of this article were defrayed in part by (he payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
' The work was supported in part by the Department of Health and Human
Services. Grants CA49I33, CA42557, and CA50905; Association for BrainTumor Research/Ellen Ruth Lebow Fellowship; Rochester Eagle's Cancer Re
search Foundation; and the University of Chicago Cancer Research Center.2To whom requests for reprints should be addressed, at The Department of
Medicine, Section of Hematology/Oncology, University of Chicago. 5841 SouthMaryland Avenue, Box 420, Chicago, IL 60637.
22 occur frequently in gliomas (9-14). Mutations in the 77*55gene have also been observed in a few glioma-derived cell lines
(15). However, structural abnormalities involving the short armof chromosome 9 (9p) appear to be the most common structuralabnormalities observed in gliomas (9). The IFNA/IFNW andIFNB1 as well as the gene that codes for the enzyme methylthioadenosine phosphorylase (MTAP) have been localized to9p (16-18). We previously reported the homozygous or hemizygous deletion of the IFN gene cluster in 7 of 15 (47%)malignant hematopoietic cell lines and in 18 primary leukemiacells of 62 (28%) patients with ALL1 examined (19, 20). Some
of these cell lines with IFN gene deletions also lack MTAPenzyme activity, suggesting that the MTAP gene is included inthese deletions. We have also previously reported IFN genedeletions in a few glioma-derived cell lines and primary tumor
tissue (21, 22). Miyakoshi et al. (23) as well as James et al. (24)have reported similar observations in cell lines and primarytumor samples from glioma patients. However, neither theextent of these genomic alterations on 9p nor the involvementof the different IFN genes has been studied in great detail.
To extend our earlier results and to confirm that the loss ofDNA sequences, which include the IFN gene cluster and theMTAP gene, is associated with the malignant process ingliomas, we have performed detailed analyses of several morecell lines and primary tumor tissue derived from these patients.We sought to determine the full extent of these deletions andtheir shortest region of overlap, in order to clone and identifycandidate tumor suppressor genes from this region on the shortarm of chromosome 9.
MATERIALS AND METHODS
Tumor and Cell Lines. We studied cell lines established from patientswith glioma, which were either provided by the investigators who hadestablished them or were obtained from the American Type CultureCollection. The cell lines used and references to their origin are listedin Table 1. We also examined primary tumors from patients withglioma (see Table 2). Tumor and matched blood samples were obtainedfrom patients undergoing biopsy or surgical resection of their tumorsat the Mayo Clinic between May 1988 and June 1990. For the purposesof this study, only primary tumor samples with sufficient tumor material for cytogenetic and molecular studies were analyzed.
Cytogenetic Analysis. Cytogenetic analysis was undertaken on cellsin logarithmic growth phase, using standard techniques as describedpreviously (11). Metaphase cells were analyzed following banding witheither quinacrine mustard or trypsin-Giemsa. Chromosomal abnormalities were described in accordance with the International System forHuman Cytogenetic Nomenclature (25). The complete karyotypes ofthese tumors will be included in a separate publication; only partial
3The abbreviations used are: ALL, acute lymphocytic leukemia; MTAP,
methylthioadenosine phosphorylase.
2523
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF 9p DELETIONS IN HUMAN GL1OMAS
karyotypes are included in this report.4
Preparation of DNA, Gel Electrophoresis, and Southern Transfers.The primary tumors were classified morphologically in accordance withthe WHO schema and were graded by the method of Daumas-Duportet al. (26) and Zulch (27). The first, middle, and last sections were air-dried, fixed in acetone, and stained with hematoxylin and eosin. Thesections were examined under the microscope for an estimate of theproportion of tumor cells. High molecular weight DNA was isolatedfrom the pooled primary tumor slices and matched peripheral bloodcells as well as from the different cell lines using an automated DNAextraction instrument (model 340; Applied Biosystems, Inc.). PurifiedDNA (5-8 /jg) was digested with different restriction enzymes, whichinclude Hindlll, Msp\, Taql, Bgl\\, and EcoRl, and separated by elec-trophoresis in 0.8% agarose gel slabs. DNA was transferred from thegels to nylon membranes (Gene Screen Plus, New England Nuclear) asdescribed previously (28).
Probes and Hybridization. Since we have previously determined thecritical region on chromosome 9 to be closely linked to the /FY genecluster, and due to the fact that limited amount of tumor DNA wasavailable, only the following cloned DNA were used as probes in theSouthern hybridization: (a) pL-FA, an IFNA2 cDNA clone that cross-hybridizes to all the genes of the IFNA gene family (29); (b) L130, agenomic clone that recognizes the a-like or u-interferon (IFNW} genes(previously named «„)(30); (c) pHFb, a genomic clone of the IFNBIgene (31); (d) CRLI263 (D9S19) and CRL1022 (D9S25), 2 polymorphic human DNA segments that map to 9p (Collaborative ResearchInstitute); and (e) pTR 48, a partial cDNA clone of the transferrinreceptor (TFR) gene (localized on 3q) that was used as control for theamount of DNA loaded in the Southern blots (32). The TFR probe wasused as a control because abnormalities of 3q are uncommon in gliomas.Probes were prepared by isolation of appropriate restriction fragmentsfrom 0.8% low melting agarose. The inserts were used as templates toprepare labeled probes with Klenow fragment of DNA polymerase Iand [a-';P]dCTP, after priming with random hexanucleotides (33).
Hybridization and washing of the blots were as described (28). Toquantitate the changes affecting the IFN genes, autoradiographs of theblots were examined by scanning densitometry, and peak areas corresponding to each hybridization signal were calculated by electronicintegration. These ratios were corrected for differences in the amountof DNA loaded in the gels, using the control probe for a locus outsideof chromosome 9 (TFR).
IFNA and IFNW refer to about 26 IFN genes and pseudogenes thatare interspersed and clustered within a short segment of 9p, while theIFNB1 gene maps distal to this cluster5. We have used the length of
restriction fragments from published partial restriction maps of theIFN gene cluster to identify the different IFN genes that hybridize tothe probes used in this study (34, 35). We have assigned each restrictionfragment identified on a Hindlll digest to individual /FA'genes. How
ever, several of the IFN genes overlap on the same fragment, thereforeonly a few are identified in Fig. 1. Two restriction fragments thathybridize to an IFNA probe but do not correspond to any of thepreviously mapped IFNA genes are called IFNA21 and IFNA22 in thisreport. Many of the cell lines and primary tumors with complete orpartial homozygous deletions of these genes have at least one chromosome 9 homologue that appeared normal at the light microscope level(19, 20). We have used these partial deletions to determine the orderof some of these genes or group of genes on 9p/ This allowed us todetermine the extent of some of the deletions and the genes involved.
To determine whether there were submicroscopic rearrangementsclosely linked to, but excluding, the IFN genes, pulsed-field gel electro-phoresis was performed on several of the cell lines. DNA was obtainedby lysing cells prepared in solid agarose plugs, and was digested in theplugs with the restriction enzyme Noti, which recognizes the 8-base
4 D. T. Ransom, S. R. Ritland, C. P. Moertel. R. J. Dahl. J. R. O'Fallon. B.
W. Scheithaur, D. W. Kimmel, P. J. Kelly, O. I. Olopade. M. O. Diaz, and R. B.Jenkins. Correlation of cytogenetic and diffuse loss of heterozygosity studies inhuman astrocytomas and mixed oligo-astrocytomas, submitted for publication.
'O. I. Olopade, S. Bohlander, H. Pomykala, M. Le Beau, and M. Diaz.Deletion mapping of the interferon gene cluster on chromosome region 9p22,submitted for publication.
pair sequence GCGGCCGC. The plugs were then placed into slots ina 0.8% agarose gel (5 mm depth) and electrophoresed for 18 h at 14°C
using a transverse alternating field electrophoresis (Beckman Geneline)system. The electrophoresis was performed under constant current 160mA (310 V) in a running buffer of 10 rnvTris base/0.5 mM EDTA/glacial acetic acid. Gels were prepared for Southern blotting by 20-mindepurination in 0.25 N HCI, followed by denaturation in 0.4 N NaOHfor 90 min and neutralization in 0.5 M Tris for 60 min (36). DNA wastransferred to nylon membranes and hybridized to IFN gene probes asdescribed previously (28).
Expression of the MTAP enzyme was determined both by Westernblot analysis with an antibody specific for the MTAP protein (37) andby a radiochemical assay for enzyme activity (38, 39). In every case, theresults of the Western blot analysis were concordant with enzymeactivity assay; the results of enzyme activity only are listed in Table 1.The MTAP assay was performed only on the established cell lines,because there was often not enough tumor tissue to perform the assayon the primary tumors.
RESULTS
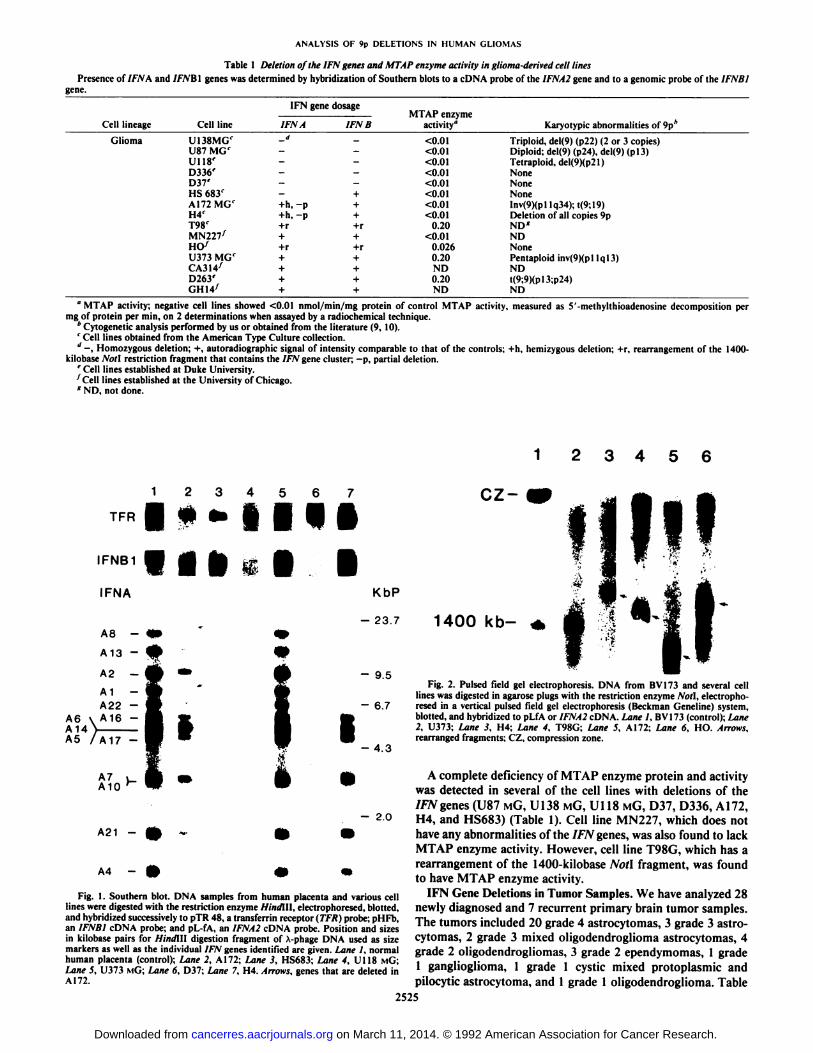
IFN Gene Deletions in Cell Lines. Of the 15 glioma-derivedcell lines examined, we found 5 cell lines (U87 MG, U138 MG,U118 MG, D37, and D336) that have homozygous deletion ofthe entire 1FNA/IFNW gene cluster and of the IFNB1 gene(Table 1; Fig. 1). Three other glioma-derived cell lines hadhomozygous deletions of various IFN genes in the cluster. A172showed the loss of 2 bands in Southern blot after Hindilldigestion and hybridization to the IFNA2 probe (Fig. 1) andloss of 3 bands after hybridization to the L-130 probe (data notshown). These bands correspond to DNA fragments that wehave assigned to IFNAI, 1FNA8, 1FNW2, and ¡FNW19[usingthe restriction maps previously published by Heneo et al. (35)and unpublished maps by Maltepe and Diaz]. The deletionswere seen with other restriction enzyme digests, and the sizesof the missing fragments corresponded to the expected sizes forIFNAI, IFNA8, IFNW2, and 1FNW19 genes, thus confirmingthat the band losses observed were not a result of polymorphism(data not shown). In the cell line H4, homozygous deletion ofpart of the IFNA/IFNW gene cluster including IFNAI, IFNA8,IFNA22, IFNAI3, IFNA2, and IFNW19 was observed. In thecell line HS683, the deletion included all of the genes in thecluster except for IFNW1 (data not shown) and IFNBI (Fig. 1).The other genes in the IFNA/IFNW gene cluster were deletedfrom only one chromosome 9 homologue (hemizygous deletion)in the 3 cell lines. The IFNBI gene was not deleted in H4 (Fig.1). Cell lines U373, D263, MN227, CA314, GH14, HO, andT98G have a complete diploid complement of the IFN genes(Table 1). The probes CRL1263 and CRL1022 did not detectany deletions or rearrangements in any of the cell lines describedabove (data not shown).
By long range restriction mapping, we have determined thatthe IFNA/IFNW and IFNBI genes are contained within one1400-kilobase Noil fragment. Therefore, any loss or rearrangement of sequences within this region can be seen as ashift in the size of the Non fragment. Rearrangement of thisNoti fragment was observed in all cell lines with partial deletions (A 172, HS683, and H4) and 2 cell lines that have nodeletion of the IFN gene cluster (T98G and HO) (Fig. 2). Wehave also observed rearrangements in 2 melanoma-derived celllines, a lung cancer-derived cell line and a leukemia-derived cellline.6 We did not observe any rearrangements in MN227, U373
MG, CA314, D263, or GH14.
6O. I. Olopade, D. Buchhagen. J. Minna, J. D. Rowley, and M. O. Diaz,
unpublished data.
2524
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
ANALYSIS OF 9p DELETIONS IN HUMAN GLIOMAS
Table 1 Deletion of the IFN genes and MTAP enzyme activity in glioma-derired cell lines
Presence of 7ÕWAand IFNBÌgenes was determined by hybridization of Southern blots to a cDNA probe of the ¡FNA2gene and to a genomic probe of the IFNBlgene.
Cell lineage CelllineGliomaU138MG'U87MG'U118'D336-'D37'HS
683'A172MG'H4'T98'MN227'HO'U373
MG'CA314'D263'GH14yIFN
gene dosageMTAPenzymeIFN
A IFN Bactivity"-d
-<0.01<0.01<0.01<0.01<0.01+
<0.01+h,-p +<0.01+h.-p +<0.01+r
+r0.20+-1-<0.01+r+r0.026++0.20++ND+-1-0.20++ NDKaryotypic
abnormalities of9p*Triploid,
del(9) (p22) (2 or 3copies)Diploid;del(9) (p24), del(9)(pl3)Tetraploid,
del(9)(p21)NoneNoneNoneInv(9)(pllq34);t(9;19)Deletion
of all copies9pND*NDNonePentaploid
inv(9)(plIql3)NDt(9;9)(pl3;p24)ND
"MTAP activity; negative cell lines showed <0.01 nmol/min/mg protein of control MTAP activity, measured as 5'-methylthioadenosine decomposition per
mg of protein per min. on 2 determinations when assayed by a radiochemical technique.* Cytogenetic analysis performed by us or obtained from the literature (9, 10).' Cell lines obtained from the American Type Culture collection.J —¿�,Homozygous deletion; +, autoradiographic signal of intensity comparable to that of the controls; +h, hemizygous deletion; +r, rearrangement of the 1400-
kilobase Noll restriction fragment that contains the IFN gene cluster; -p, partial deletion.' Cell lines established at Duke University.1Cell lines established at the University of Chicago.*ND, not done.
3 4
cz-TFR
IFNB1
IFNA
•¿�••••ilKbP
- 23.7
- 9.5
- 6.7
- 4.3
- 2.0
A21 -
A4 -
Fig. 1. Southern blot. DNA samples from human placenta and various celllines were digested with the restriction enzyme Hindlll, electrophoresed. blotted,and hybridized successively to pTR 48, a transferrin receptor (TFR) probe; pHFb,an IFNBl cDNA probe; and pL-fA, an IFNA2 cDNA probe. Position and sizesin kilobase pairs for Hindltl digestion fragment of A-phage DNA used as sizemarkers as well as the individual IFN genes identified are given. Lane I, normalhuman placenta (control); Lane 2, A172; Lane 3, HS683; Lane 4, U118 MG;Lane S, U373 MÃ’;Lane 6, D37; Lane 7, H4. Arrows, genes that are deleted inA172.
1400 kb-
Fig. 2. Pulsed field gel electrophoresis. DNA from BV173 and several celllines was digested in agarose plugs with the restriction enzyme Noll, electrophoresed in a vertical pulsed field gel electrophoresis (Beckman Geneline) system,blotted, and hybridized to pLfA or IFNA2 cDNA. Lane I, BV173 (control); Lane2, U373; Lane 3. H4; Lane 4, T98G; Lane 5. A172; Lane 6, HO. Arrows,rearranged fragments; CZ, compression zone.
A complete deficiency of MTAP enzyme protein and activitywas detected in several of the cell lines with deletions of theIFN genes (U87 MG, U138 MG, Ul 18 MG, D37, D336, A172,H4, and HS683) (Table 1). Cell line MN227, which does nothave any abnormalities of the IFN genes, was also found to lackMTAP enzyme activity. However, cell line T98G, which has arearrangement of the 1400-kilobase Noil fragment, was foundto have MTAP enzyme activity.
IFN Gene Deletions in Tumor Samples. We have analyzed 28newly diagnosed and 7 recurrent primary brain tumor samples.The tumors included 20 grade 4 astrocytomas, 3 grade 3 astro-cytomas, 2 grade 3 mixed oligodendroglioma astrocytomas, 4grade 2 oligodendrogliomas, 3 grade 2 ependymomas, 1 grade1 ganglioglioma, 1 grade 1 cystic mixed protoplasmic andpilocytic astrocytoma, and 1 grade 1 oligodendroglioma. Table
2525
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSISOF 9p DELETIONS IN HUMAN GL1OMAS
Table 2 Summary of the molecular analysis of 9p deletions in primary gliomas
UPN626667707172737475798283848586103113122125134136140150154157158159160162196197215219231235Grade24244114441242444423443334444443422MorphologyOligodendrogliomaAstrocytomaOligodendroglioma
with minorastrocy-ticcomponentAstrocytomaAstrocytomaCystic
mixed protoplasmic andpilocyticastrocytoma(recurrent)GangliogliomaAstrocytoma
(recurrent)AstrocytomaAstrocytomaOligodendrogliomaEpendymomaAstrocytomaCellular
ependymoma(recurrent)Astrocytoma(recurrent)AstrocytomaAstrocytomaAstrocytomaOligodendroglioma
(recurrent)Mixedoligodendroglioma-astrocytomaAstrocytomaAstrocytomaAstrocytomaAstrocytoma
(recurrent)Mixedoligodendroglioma-astrocytoma(recurrent)AstrocytomaAstrocytomaAstrocytomaAstrocytomaAstrocytomaAstrocytomaAstrocytomaAstrocytomaOligodendrogliomaEpendymomaIFNA
IFNB1+"++
++++h*
++h
++++
++++h+h
++++++++h—
—¿�——¿�+h+
++h-t-h++-t-h+h+++++h+h+++
++++h+h++++++++-t-h+h+++
+Cytogenetic
abnormality of9pNoneNo
growth incultureNonedel(9)(pl3)(2or9cells)NoneNoneNoneNoneNoneNoneNone-9
(in triploid cell) (1 of 5cells)der(9)t(9;?)(p?22?;?)(1 of 2cells)NoneNoneNoneder(9)t(9;?)(pl3;?)
(5 of 25cells)-9,-9(in tetraploid cell) (1 of 20cells)der(9)t(9;10)(pl3;ql
1.2) (1 of 7cells)Nonedel(9)(pl3p22)(23of25cells)del(9)(p21.2p22)(21
of 21cells)Nonedel(9)(pl3)(l
of 12cells)Nogrowth incultureNoneNonedel(9)(pll)(5of
21cells)NoneNone+9
(23 of 30cells)NoneNoneNone+9,
+ (16 of 30 cells)°+, Autoradiographic signal of intensity comparable to that of controls.* +h, Hemizygous deletions. Gene dosage was calculated by normalizing to a control probe (TFR) and using the following calculation: (IFNj/TFRi)/(/FNN/TFRN)
x 2, where ¡FNA2represents the signal response for IFNA2 in corresponding normal (N) and tumor (T) DNA samples, and TFR represents the signal response atthe TFRC locus on chromosome 3. Using this formula, a value of 2.00 would represent a normalized tumor DNA IFNA2 signal response equivalent to that producedby the control DNA. The dosage values for IFNA2 in the tumors with deletions are: tumors 70, 0.8; 71, 0.9; 75, 0.4; 79, 0.9; 85, 0; 86, 0.37; 103, 0.26; 113, 0.37;125, 0.83; 136, 0.90; 160, 0.92; 154, 0.89; and 219. 1.1. Dosage values higher than 0 in the tumors with homozygous deletion are due to admixed normal cells.
' —¿�,Homozygous deletion.
PatientNumber
62 66 67 70 71 72 73 74 75 79 82 83 84 85 86
TFR •¿�
IFNB1
IFNA2 •¿�
Fig. 3. Southern blot. DNA from primary glioma tissues was digested with the restriction enzyme HindlU. electrophoresed, blotted, and hybridized, successivelyto TFR, ¡FNB1,and ¡FNA2probes. The band displayed corresponds to IFNA2, which gives the strongest and readily interpretable signal when this probe is hybridizedto //i/i<flll-digested DNA.
2 contains a summary of the results and the partial karyotypesof these tumors. Using Taq\ and Mspl restriction enzymes, only2 of the patients showed restriction fragment length polymorphism with IFNA2 gene probe; there was loss of one IFNA2alÃelein tumor cells from one of these 2 patients (see Fig. 4,patient 136). This patient was also informative for theCRL1263 probe and demonstrated loss of one alÃelein thislocus. Thus, for the majority of the tumors, we have relied onthe densitometry analysis for a determination of gene dosage.Homozygous deletion of the entire IFNA/IFNW and IFNB1
gene cluster was observed in tumors 86 and 103, while homozygous deletion of only the IFNA/IFNW genes was observed intumors 75, 85, and 113 (Figs. 3 and 4; Table 2) In the latertumors, hemizygous deletion of IFNB1 gene was also observed.Tumors 70, 71, 79,125, 136,154,160, and 219 had hemizygousdeletion of the IFNA/IFNW gene cluster; in addition, thesedeletions were associated with hemizygous loss of IFNB1 genein tumors 125, 160, 154, and 219. Thus, 13 of 35 (37%) primarytumors had hemizygous or homozygous deletions of the IFNgenes. We were unable to ascertain any additional partial dele-
2526
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF 9p DELETIONS IN HUMAN GL1OMAS
Patient No.:
TFRControl Probe
IFNA2
kbp9.5-
6.7-
-4.3-
2.3- im
Fig. 4. Taql digest of primary tumor tissues hybridized to an IFNA2 probe. B-normal blood, T-tumor tissue. Arrow, loss of a restriction fragment from apolymorphic allelic pair. Residual hybridization signal in patient 103 tumor laneis due to admixed normal tissue.
lions among these tumors. Ten of the 13 tumors with 1FN genedeletions were grade 4, while the remaining 3 were grades 2and 3. The 3 lower grade tumors were studied after recurrence.
Correlation of Cytogenetics with Gene Loss. Cytogeneticanalysis showed the involvement of 9p in deletion or rearrangement in at least one cell in 10 of the 35 (29%) primarytumors studied, and 7 of the 11 (64%) cell lines for whichcytogenetic data are available (Tables 1 and 2). However, for arearrangement to be regarded as clonal, it should be observedin at least 2 metaphase cells in the case of structural rearrangements, and 3 cells for numerical changes. Thus, thevisible involvement of 9p is questionable in tumors 83, 84, 122,and 154 since in each case the change was seen in only 1 or 2metaphase cells. In tumors 71, 75, 79, 85, 86, 103, and 219,homozygous or hemizygous deletion of the IFN genes wasobserved without any visible cytogenetic abnormality of 9p. Intumor 140, a del(9)(p21.2 p22) was observed in all the tumorcells with no apparent deletion of IFN genes.
DISCUSSION
The observation of a high frequency of homozygous or hemizygous loss of sequences on 9p, which include the IFN genesin the primary tumor of patients with gliomas, suggests thatthese gene deletions are associated with the malignant processin gliomas. In this report, we have demonstrated that thesedeletions are sometimes submicroscopic, since deletion of DNAsequences from the region is found in cases without cytogeneticabnormalities of 9p. While we cannot rule out genomic instability as being responsible for the observed hemizygous deletions, the 5 primary tumors with homozygous deletion provideus with the strongest evidence that the deletion of sequences inthis region on 9p is nonrandom and may play an importantrole in glioma tumorigenesis.
The high frequency of IFN gene deletions or rearrangementsin glioma-derived cell lines (67%) may be due to the fact thatmost of these cell lines were derived from patients with glio-blastoma multiforme (grade 4 astrocytomas), since the incidence of structural rearrangements of 9p is higher in high grade
tumors (9). It is unlikely that the IFN gene deletion is a mutationacquired during growth of the cells in vitro because we haveobserved similar deletions at a significant frequency (37%) inprimary glioma tumor tissue. The increased frequency of thedeletions in glioma-derived cell lines may be due in part to aselective advantage conferred by the deletion for establishmentor growth of the cells in vitro. A similar hypothesis has beenproposed to explain the predominance of RBI deletions inbreast cancer cell lines established in vitro. In addition, ourobservation of IFN gene deletions only in association with newlydiagnosed primary grade 4 tumors or recurrent tumors suggeststhat this genetic mutation is important for tumor progression.These results are consistent with the report by James et al. (24),who found IFN gene deletions in 10 of 20 high grade tumors,while none of the 10 low grade tumors had deletions; thedeletions were seen frequently in association with alterations ofchromosome 10, chromosome region 17p, or amplification ofEGFR. In this study, each of the tumors with 9p abnormalitiesalso has additional alterations, often involving chromosomesor chromosome regions 7q, 10, 13q, 17p, and 19.4
In our previous study of ALL, we had observed 2 cases ofpartial deletion of the IFN gene cluster (20). In this study,several cell lines have partial deletion with breakpoints that arescattered within the IFN gene cluster. We found partial deletionor rearrangement in 3 cell lines (A 172, T98G, and H4) thatwere previously studied by Miyakoshi et al. (23). These werenot reported as partial deletions by the previous authors becausethey only considered the restriction fragment that containsIFNA2 for their evaluation oflFNA gene losses. The observationof several deletion breakpoints within the IFN gene clustersuggests that this locus may contain nucleotide sequences thatparticipate in unequal crossing over, abnormal recombination,or chromosome breakage. Detailed studies of these deletionbreakpoints should provide us with clues about the possiblemechanisms involved in these deletions.
The rearrangements of the Noil fragment that contains theIFN gene cluster observed in 5 different glioma-derived celllines, as well as in ALL-, melanoma-, and lung cancer-derivedcell lines, suggests that the mutational event in these tumorsmay have eliminated one of the Noti sites flanking the IFNgene cluster. The observed rearrangement does not appear tobe due to methylation, because we have examined 25 differentcell lines that show no evidence of deletion of either 9p or theIFN genes, and have found no variation in the 1400-kilobaseNoti fragment. Other enzymes such as Sacll, 5/71, and BssHllcould not be reliably used in this analysis. We believe that the2 Noti sites encompassing the IFN genes are always unmethy-lated, and that no other Noti site is located between them.Moreover, the size of the rearranged Noil fragment varies inthe different cell lines with abnormalities, including those withpartial deletions, which is inconsistent with products of methylation. It is therefore likely that the rearrangements seen incell lines without deletion of the IFN genes are due to deletionswith breakpoints inside the 1400-kilobase Noil fragment, butoutside the IFN gene cluster. No germline band was observedin T98G and HO, perhaps because the rearranged alÃelewasduplicated, while the homologous alÃelewas deleted from themalignant cells following somatic recombination or chromosome nondisjunction. As we obtain additional probes fromwithin this region of 9p, the extent and mechanism of thedifferent deletions will be studied more accurately.
We have observed a deficiency of MTAP enzyme activity inseveral of these cell lines, suggesting that the MTAP gene is
2527
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF 9p DELETIONS IN HUMAN GLIOMAS
p-telomere
Not I restriction siteIFNB1
IFNW1
IFNA21, IFNWP1S, IFNA4
U87U118U138D37D336
Band9p22
IFNWP9, IFNA7, IFNA10
IFNWP18, IFNA16, IFNA17
IFNA5, IFNAÌ4,IFNA22
IFNA2, IFNA6, IFNA13, IFNAP20
IFNA8, IFNW2, IFNWP19,IFNA1
Not I restriction site
I shortest region of overlap
MTAP
centromere
Fig. 5. Schematic diagram of//W genes lost in several glioma-derivedcell lineswith deletions, showing the shortest region of overlap.
involved in the same event that resulted in homozygous loss ofthe 1FN genes. In fact, Nobori et al. (37) recently reportedabsence of MTAP enzyme in 9 of 12 (75%) primary gliomas.Thus, the loss of MTAP protein or enzyme activity can be usedas a marker of deletion in the cases in which there is no directevidence of such an event, such as in cell line MN227, whichshows no deletion or rearrangement of the IFN genes. However,since the MTAP gene has not been cloned, we have not beenable to link all these genes on a long range map.
To determine the shortest region of overlap of these deletions,we have combined the information obtained from cytogeneticanalysis, standard Southern analysis, pulsed field gel electro-phoresis, and MTAP enzyme assay. Among the deletions detected in glioma-derived cell lines, some have breakpointswithin the IFN gene cluster and, therefore, delete only part ofthe IFN genes; other deletions occur within the 1400-kilobaseNon fragment and extend to MTAP but do not include the IFNgenes, so that a different sized Noti fragment is seen; otherdeletions occur within the 1400-kilobase Noti fragment but donot extend to MTAP; still other deletions may occur beyondthe boundaries of the Noti fragment so that no abnormalitiesare detected by our currently available probes but the cells lackMTAP enzyme activity. By comparing the different deletionsthat are visible at the light microscope level with the differentgenes lost at the molecular level, we have tentatively determinedthe order of the genes on 9p to be te\omeTe-IFNBl-IFNA//F7VH/cluster-M7/i/>-centromere.5 A schematic diagram of all
complete and partial deletions and rearrangements is shown inFig. 5. The diagram indicates that the shortest region of overlapof the deletions as determined by the materials currently available to us lies in the interval between the centromeric end ofthe IFN gene cluster and the MTAP gene locus. We had previously suggested that the IFN genes could be the tumor suppressor genes involved in ALL (19, 20). Miyakoshi et al. (23) alsopreviously proposed that the IFN genes are the tumor suppressor genes. However, the shortest region of overlap of thesedeletions using data obtained from long range mapping and
MTAP analysis excludes the IFN gene cluster. We believe thatanother gene closely linked to the IFN genes is the relevanttumor suppressor gene. However, the loss of the IFN geneswhen it occurs may play an additional role in the progressionof these tumors.
In conclusion, we have confirmed that homozygous deletionsor rearrangement of the IFN and MTAP genes occurred frequently in primary glioma samples, as previously described inpatients with ALL. There have been several reports of survivorsof childhood ALL who develop gliomas as secondary cancer.These brain tumors have frequently been thought to arise as aresult of the prophylactic cranial irradiation to which thesechildren were exposed (40-43). However, Meadows et al. (44)have suggested that the coincidence of gliomas and hematolog-ical cancers is nonrandomly distributed in the group of patientswith second cancers and may represent a new "cancer syndrome." The fact that both gliomas and ALL have occurred in
the same individual in the absence of radiation, and that thereare families in which both tumors have been documented,suggest that the same genetic mutation may predispose to thedevelopment of both cancers. Germline mutations in the putative tumor suppressor gene on 9p could be the predisposingfactor in such families. Studies are under way to define theextent of the deletions on 9p, and to delimit their shortestregion of overlap, in order to clone and characterize the putativetumor suppressor gene in this locus.
ACKNOWLEDGMENTS
We are grateful to Elizabeth Van Melle and Steve Ritland for theirtechnical help, Paula Pitha for the pLfa and pHfb probes, ClaudioSchneider for the pTR48 probe, Sandra H. Bigner and Kari Steffansonfor providing the cell lines, and Joyce Trice for her secretarialassistance.
REFERENCES
1. Cavenee, W. K., Dryja, T. P., Phillips, R. A., Benedict, W. F., Godbout, R.,Gallie, B. L., Murphree, A. L., Strong, L. C, and White, R. L. Expressionof recessive alÃelesby chromosomal mechanisms in retinoblastoma. Nature(Lond.), 305: 779-784, 1983.
2. Dryja, T. P., Cavenee. W., White, R. M, Rapaport, J. M., Petersen, R.,Albert, D. M., and Bruns, G. A. Homozygosity of chromosome 13 inretinoblastoma. N. Engl. J. Med., 310: 550-553, 1984.
3. Friend, S. H., Bernards, R., Rogelj, S., Weinberg, R. A., Rapaport, J. M.,Albert, D. M., and Dryja, T. P. A human DNA segment with properties ofthe gene that predisposes to retinoblastoma. Nature (Lond.), 323: 643-646,1986.
4. Lee, W. H., Bookstein, R., Hong, F., Young, L. J., Shew, J. Y., and Lee, E.Y-H-P human retinoblastoma susceptibility gene: cloning, identification andsequence. Science (Washington DC), 235: 1394-1399, 1987.
5. Zbar, B., Brauch, H., Talmadge, G., and Linehan, M. Loss of alÃelesof locion the short arm of chromsome 3 in renal cell carcinoma. Nature (Lond.),327:721-724,1987.
6. Naylor, S. L., Johnson, B. E., Minna, J. D., and Sakaguchi, A. Y. Loss ofheterozygosity of chromosome 3p markers in small cell lung cancer. Nature(Lond.), 329:451-454, 1987.
7. Vogelstein, B., Fearon, E. R., Hamilton, S. R., Kern, S. E., Preisinger, A.L., Leppert, M., Nakamura, Y., White, R., Smits, A. M., and Bos, J. L.Genetic alterations during colorectal tumor development. N. Engl. J. Med.,379:525-532, 1988.
8. Horowitz, J. M., Sang-Ho, P., Bogenmann, E., Cheng, J. C., Yandell, D.W., Kaye, F. J., Minna, J. D., Dryja, T. P., and Weinberg, R. A. Frequentinactivation of the retinoblastoma anti-oncogene is restricted to a subset ofhuman tumor cells. Proc. Nati. Acad. Sci. USA, S7: 2775-2779, 1990.
9. Bigner, S. H., Mark, J., Burger, R. C., Malaley, M. S., Bullard, D. E.,Muhlbaser, L. H., and Bigner, D. D. Specific chromosomal abnormalities inmalignant human gliomas. Cancer Res., 58: 405-411, 1988.
10. Bigner, S. H., Humphrey, P. A., Wong, A. J., Vogelstein, B., Mark, J.,Friedman, H. S., and Bigner, D. D. Characterization of the epidermal growthfactor in human glioma cell lines and xenografts. Cancer Res., 50: 8017-8022, 1990.
11. Jenkins, R. B., Kimmel, D. W., Moertel, C. A., Schultz, C. G., Scheithauer,
2528
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF 9p DELETIONS IN HUMAN GLIOMAS
B. W., Kelly, P. J., and Dewald, G. W. A cytogenetic study of 53 humangliomas. Cancer Genet. Cytogenet., 39: 253-279, 1989.
12. James, C. D., Carlbom, E., Dumanski, J. D.. Hansen, M., Nordenskjold, M.,Collins, V. P., and Cavenee, W. K. Clonal genomic alterations in gliomamalignancy stages. Cancer Res., 48: 5546-5551, 1988.
13. EI-Azouzi, Chung. R. Y., Farmer, G. E., Martu, R. L., Black, P. M, Rouleau,G. A., Hettlich, C., Hedley-Whyte, E. T., Zervas, N. T., Panagopoulos. K.,Nakamura, Y., Gunsella, J. F., and Seizinger, B. R. Loss of distinct regionson the short arm of chromosome 17 associated with tumorigenesis of humanastrocytogenetiomas. Proc. Nati. Acad. Sci. USA. 86: 7186-7190, 1989.
14. Fults. D.. Pedone, C. A., Thomas, G. A., and White, R. Allelotype of humanmalignant astrocytoma. Cancer Res., 50: 5784-5789, 1990.
15. Nigro, J. M., Baker, S. J., Preisinger. A. L., Jessup, J. M., Hosteller, R.,Cleary, K., Bigner, S. H., Davidson, N., Baylin, S., Devilee, P., Glover, T.,Collins, F. S., Weston, A., Modali, R., Harris. C. C., and Vogelstein, B.Mutations in the p53 gene occurs in diverse human tumor types. Nature(Lond.), 342: 705-708, 1989.
16. Trent, J. M., Olson, S., and Lawn, R. M. Chromosomal localization ofhuman leukocyte, fibroblast, and immune Interferon genes by means of insitu hybridization. Proc. Nati. Acad. Sci. USA, 79: 7809-7813, 1982.
17. Pitha. P. M., Slate, N. B., Raj, N. B. K., and Ruddle, F. H. Human interferongene localization and expression in somatic cell hybrids. Mol. Cell Biol., _'.•564-570. 1982.
18. Carrera, C. J., Eddy, R. L., Shows, T. B., and Carson, D. A. Assignment ofthe gene for methylthioadenosine phosphorylase to human chromosome 9by mouse-human somatic cell hybridization. Proc. Nati. Acad. Sci. USA, 81:2665-2668, 1984.
19. Diaz. M. O., Rubin, C. M., Harden, A., Ziemin. S., Larson, R. A., Le Beau.M. M., and Rowley, J. D. Deletions of interferon genes in acute lymphoblasticleukemia. N. Engl. J. Med., 322: 77-82, 1990.
20. Diaz, M. O., Ziemin, S., LeBeau. M. M.. Pitha, P., Smith, S. D., Chilcote,R. R., and Rowley, J. D. Homozygous deletion of the alpha-a and beta-linterferon genes in human leukemia and derived cell lines. Proc. Nati. Acad.Sci. USA, 85: 5259-5263. 1988.
21. Olopade, O. I., Jenkins, R., Linnenbach. A. J., Cowan, J. M., Pomykala, H.,Rowley, J. D.. and Diaz, M. O. Molecular analysis of chromosome 9pdeletion in human solid tumors. Proc. Am. Assoc. Cancer Res., 31: 318,1990.
22. Ransom. D., Jenkins, R., Olopade. F., Diaz, M., Ritland, S., and Bren, C.T. Loss of chromosome 10 alÃelesand loss of alpha-inlerieron genes inhuman gliomas. Proc. Am. Assoc. Cancer Res., 31: 38, 1990.
23. Miyakoshi, J., Dobler, K. D., Allalunis-Turner, J., McKean, J. D. S., Petruk,K., Allen, P. B. R., Aronyk, K. N., Weir, B., Huyser-Wierenga, D., Fulton,D.. Urtasun, R. C., and Day, R. S. III. Absence of IFNA and 1FNB1 genesfrom human malignant glioma cell lines and lack of correlation with cellularsensitivitiy to interferons. Cancer Res., 50: 278-283. 1990.
24. James, C. D., He, J., Carlbom, E., Nordenskjold, M., Cavenee, W. K., andCollins, V. P. Chromosome 9 deletion mapping reveals interferon A andinterferon Bl gene deletions in human glial tumors. Cancer Res., 51: 1684-1688, 1991.
25. Harnden, D. G., and Klinger, H. P. (eds.) An International System forHuman Cytogenetic Nomenclature. Basel: Karger (published in collaborationwith Cytogenet. Cell Genet.), 1985.
26. Daumas-Duport. C., Scheithauer, B., Ofallen, J., and Kelly, P. J. Grading ofastrocytomas: a simple and reproducible method. Cancer (Phila.), 62: 2152-2165, 1988.
27. Zulch, K. J. Histological Typing of Tumors of the Central Nervous System.International Histological Classificatoin of Tumor No. 21. Geneva: WHO,1979.
28. Shima, E., Le Beau, M. M., McKeithan, T. W., Minowada, J., Showe, L. C.,Mack, T. W., Minden, M. D., Rowley, J. D., and Diaz, M. O. T-cell receptoralpha chain gene moves immediately downstream of c-myc in a chromosomal8;14 translocation in a cell line from a human T-cell leukemia. Proc. Nati.Acad. Sci. USA. 83: 3439-3443. 1986.
29. Mizoguchi, J. Pitha, P. M., and Raj, N. B. Efficient expression in Escherichiacoli of two species of human interferon-a/pAa and their hybrid molecules.DNA, 4:221-232, 1985.
30. Feinstein, S. I., Mory, Y., Chernajovsky, Y., Maroteaux, L., Nir, V., Lavie,V., and Revel, M. Family of human interferon-like sequences. Mol. Cell.Biol.. 5:510-517, 1985.
31. Raj, B. K., and Pitha, P. Two levels of regulation of beta interferon geneexpression in human cells. Proc. Nati. Acad. Sci. USA, 80:3923-3927,1983.
32. Schneider, C., Owen, M. J., Banville, D., and Williams, J. G. Primarystructure of the human transferrin receptors deduced from the mRNAsequence. Nature (Lond.), 311: 675-678. 1984.
33. Feinberg, A. P., and Vogelstein, B. A technique for radiolabelling DNArestriction endonuclease fragments to high specific activity. Ann. Biochem.,132:6-\3, 1983.
34. Brack, C., Nagata, S., Martei, W., and Weissman, C. Molecular analysis ofthe human interferon-a//)/ia gene family. Gene, 15: 379-394, 1981.
35. Heneo, K., Brosius, J., Fujisawa, A., Fujisawa, J. 1., Haynes, J. R., Hochstadt,J., Kovacic, T., Pasek, M., Schambock, A., Schmid, S., Todokoro, K.,Walehli, M., Nagata, S., and Weissman, C. Structural relationship of humaninterferon alpha genes and pseudogenes. J. Mol. Biol., 185: 227-260. 1988.
36. Gardiner, K., Laas, W., and Patterson, D. Fractionation of large mammnalianDNA restriction fragments using vertical pulsed-field gradient gel electro-phoresis. Somatic. Cell Mol. Genet., 12: 185-195, 1986.
37. Nobori, T., Karras J. G., Ragione, F. D., Walttz, T. A., Chen, P. P., andCarson, D. A. Absence of methylthioadenosine phosphorylase in humangliomas. Cancer Res.. 51: 3193-3197. 1991.
38. Kamatani, N., and Carson, D. A. Dependence of adenine production uponpolyamine synthesis in cultured human lymphoblasts. Biochim. Biophys.Acta., 675: 344-350, 1981.
39. Bradford, M. M. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding.Anal. Biochem., 72: 248-254, 1976.
40. Li, F. P., Cassady, J. R., and Jaffer W. Risk of second tumors in survivorsof childhood cancer. Cancer (Phila.). 35: 1230-1235, 1975.
41. Farwell, J., and Flannery, J. T. Second primaries in children with centralnervous system tumors. J. Neuro-Oncol., 2: 371-375, 1984.
42. Farwell, J., and Planner). J. T. Cancer in relatives of children with centralnervous system neoplasms. N. Engl. J. Med., 311: 749-753. 1984.
43. Meadows, A. T., O'Angno C., and Mike, U. M. Patterns of second malignantneoplasms in children. Cancer (Phila.), 40: 1903-1911, 1977.
44. Meadows, A. T., Barron, E., Fossati-Behain. F., et al. Second malignantneoplasms in children; an update from the Late Effects Study Group. J. Clin.Oncol., 3: 532-538, 1985.
2529
on March 11, 2014. © 1992 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from




















