
Hsp60C is required in follicle as well as germline cells during oogenesis in Drosophila melanogaster

Insect Biochemistry and Molecular Biology 28 (1998) 309–319
Molecular adaptation ofDrosophila melanogasterlysozymes to adigestive function
Rosana Regela, Sergio R. Matiolib, Walter R. Terraa,*
a Departamento de Bioquı´mica, Instituto de Quı´mica, Universidade de Sa˜o Paulo, C.P. 26077, 05599-970, Sa˜o Paulo, Brasilb Departamento de Biologia, Instituto de Biocieˆncias, Universidade de Sa˜o Paulo, C.P. 11461, 05422-970, Sa˜o Paulo, Brasil
Received 9 July 1997; accepted 7 November 1997
Abstract
A lysozyme (pI 5.5) was purified to homogeneity from heated acid extracts ofDrosophila melanogasterlarvae, using gel filtrationin a Superose column and ion-exchange chromatography in a Mono Q column. The final yield was 67%. The purified lysozymewith Mr 13 700 (determined by SDS-polyacrylamide gel electrophoresis) decreases in activity and has its pH optimum displacedtowards acidic values andKm increases as the ionic strength of the medium becomes higher. The lysozyme is resistant to a cathepsinD-like proteinase present in cyclorrhaphous Diptera and displays a chitinase activity which is 11-fold higher than that of chickenlysozyme. Microsequencing of an internal peptide of the purified lysozyme showed that this enzyme is the product of the previouslysequenced Lys D gene. The results suggest that the product of the Lys P gene has pI 7.2, a pH optimum around 5 and is not atrue digestive enzyme. The most remarkable sequence convergence ofD. melanogasterlysozyme D and lysozymes from vertebrateforegut fermenters are serine 104 and a decrease in the number of basic amino acids, suggesting that these features are necessaryfor digestive function in an acid environment. Adaptive residues putatively conferring stability in an acid proteolytic environmentdiffer between insects and vertebrates, probably because they depend on the overall three-dimensional structure of the lysozymes.A maximum likelihood phylogeny and inferences from insect lysozyme sequences showed that the recruitment of lysozymes asdigestive enzymes is an ancestral condition of the flies (Diptera: Cyclorrhapha). 1998 Elsevier Science Ltd. All rights reserved
Keywords:Lysozyme properties; Lysozyme purification; Digestive lysozyme; Lysozyme sequences; Ruminant lysozyme
1. Introduction
Lysozyme (EC 3.2.1.17) catalyzes the hydrolysis ofthe 1,4-b-glycosidic linkage between N-acetylmuramicacid and N-acetylglucosamine of the peptidoglycanpresent in the cell wall of many bacteria, causing celllysis. Lysozyme is part of the defense mechanismagainst bacteria and has been described in most animals(Jolles and Jolle`s, 1984), including insects (Dunn, 1986;Hultmark, 1996).
Lysozyme has also been implied in the midgut diges-tion of bacteria by organisms which ingest large amountsof them such as marine bivalves (McHenery et al.,1979), or that harbor a bacterial culture in their guts,exemplified by vertebrate foregut fermenters (ruminants,see Dobson et al., 1984; colobine leaf-eating monkeys,
* Corresponding author. Fax:+ 55 11 818-7986; E-mail [email protected]
0965-1748/98/$19.00 1998 Elsevier Science Ltd. All rights reservedPII: S0965-1748 (97)00108-2
see Stewart et al., 1987). Bacteria are digested in foregutfermenters with the aid of a stomach lysozyme.
Among the insects, the capacity of digesting bacteriain the midgut seems to be an ancestral trait of DipteraCyclorrhapha (Lemos and Terra, 1991a), which agreeswith the fact that most Diptera Cyclorrhapha larvae aresaprophagous, feeding largely on bacteria (Terra, 1990).These insects have midgut lysozymes (Lemos et al.,1993) similar to those of vertebrate fermenters (Dobsonet al., 1984; Stewart and Wilson, 1987). Thus, the majorlysozymes purified from Musca domestica larvalmidguts have low pI values (7.9 and 8.2), are moreactive at acid pH values, when present in media withphysiological ionic strengths, and are resistant to thecathepsin D (acid proteinase) present inM. domesticamidguts (Lemos et al., 1993).
Sequence analyses showed that specific amino acidsubstitutions are associated with the adaptation of lyso-zymes to a digestive role in ruminants and colobinemonkeys (Stewart et al., 1987; Stewart and Wilson,

310 R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
1987). As the digestive lysozymes from insects and ver-tebrate foregut fermenters have similar properties,sequence analyses could reveal the molecular basis forthese shared properties. This is particularly interesting,when it is realized that vertebrates and insects belong towidely divergent phyla.
Digestive lysozymes in vertebrate foregut fermentersare encoded by multiple genes that evolved in parallel(Irwin and Wilson, 1990), the same being true forDro-sophila melanogaster. This insect has at least eight lyso-zyme genes: five closely related genes (the Lys D family:Lys A, B, C, D and E) expressed strongly in the midgutof larvae and adults, Lys P expressed in salivary glands(mainly of adults), Lys S expressed chiefly in larval gas-tric caeca and, finally, Lys X expressed primarily inpharate and early pupa (Kylsten et al., 1992; Daffre etal., 1994). All these genes, except Lys P and Lys X,encode acidic proteins. Although all these genes weresequenced and phylogenetic trees were produced(Kylsten et al., 1992; Daffre et al., 1994), a thoroughcomparison between vertebrate digestive lysozymes andD. melanogasterlysozymes was not accomplished dueto the lack of pureD. melanogasterenzymes and knowl-edge on sequences of non-digestive lysozymes in a closerelative toD. melanogaster.
One of the threeM. domesticalysozymes charac-terized by Lemos et al. (1993) was also purified by Itoet al. (1995) and sequenced. No detailed comparison oflysozyme sequences were described and the phylogen-etic tree (obtained by the neighbor-joining method) oflysozymes lead them to the conclusion thatM. domesticalysozymes had diverged fromD. melanogasterlyso-zymes before these underwent multiple gene dupli-cations. This is a unlikely hypothesis, asM. domesticaalso have several different lysozymes (Lemos et al.,1993).
In this paper, the purification and characterization ofthe major (lysozymes D)D. melanogasterlarval lyso-zymes are described and shown to be similar to the ver-tebrate stomach lysozymes. Furthermore, the finding thatD. melanogasterlysozymes D have a serine residue inposition 104 and a low number of basic amino acids, inparallel to the vertebrate stomach lysozymes, suggeststhat these features are necessary for a digestive role inan acid environment. Finally, the use of the maximumlikelihood method of phylogenetic inference indicatedthat the recruitment of lysozyme as a digestive enzymeis an ancestral condition of the flies.
2. Material and methods
2.1. Animals
Larvae of Drosophila melanogasterCanton S(Diptera: Cyclorrhapha) were reared on a corn meal diet
with the following composition: 1000 ml distilled water,175 g corn meal, 9 g agar, 30 g yeast, 40 g soybean meal,20 ml of 0.06 g/ml malt solution, 30 ml sugar canesyrups and 30 ml of 10% nipagin in alcohol. Larvae ofMusca domestica(Diptera: Cyclorrhapha) were rearedon a mixture of fermented commercial pig food and ricehull (1:2, v/v). The larvae used in this study wereactively feeding individuals at third larval instar.
2.2. Preparation ofD. melanogasterextracts
Larvae were rinsed in water, blotted in filter paper andimmobilized by placing them on ice. About 11 000 lar-vae (15.5 g of larvae) were homogenized in 100 mMsodium acetate buffer pH 3.5 with the aid of an Omni-mixer (Omni, USA) at 6000 rev/min for 3 pulses of 20 s,with 1 min intervals. The homogenate was centrifuged at20 000g for 30 min at 4°C and the resulting supernatantpassed through glass wool to be freed from fats. Thepellet was resuspended in the same buffer as before withthe aid of an Omni-mixer at first and then with a semim-icroprobe of a sonicator Branson 250, with the outputset at 3 by using three pulses of 20 s each at 1 min inter-vals. The dispersed pellet was centrifuged at 20 000gfor 30 min at 4°C and the resulting supernatant, afterpassing through glass wool, was added to the first one.The filtrate was adjusted with buffer to contain 700 mgprotein/ml and was labeled "acid extract".
The acid extract after being frozen for 24 h wasthawed and centrifuged at 20 000g for 30 min at 4°C.The resulting supernatant was then maintained in a boil-ing water bath for 5 min and centrifuged at 10 000g for10 min at 4°C. Finally, the heated acid extract was dia-lysed overnight against 150 vol of distilled water witha single water change.
2.3. Preparation ofM. domesticamidgut homogenates
Larvae were rinsed in water, blotted in filter paper,immobilized by placing them on ice and dissected incold 150 mM NaCl. Midguts were homogenized in dou-ble distilled water (10 midguts/ml) with the aid of a Pot-ter-Elvehjem homogenizer. The homogenates after beingfiltered through a piece of nylon mesh of 45 mm poresize were centrifuged at 100 000g for 60 min at 4°C.The supernatants were passed through glass wool anddialysed overnight against 1000 vol of water with twochanges. The resulting dialysate is a good source ofcathepsin D (Lemos and Terra, 1991b).
2.4. Purification ofD. melanogasterlysozymes
Dialysed acid extract fromD. melanogasterlarvae(500ml) was applied to a column of Superose 12 HR10/30, equilibrated and eluted with 20 mM imidazolebuffer pH 6.4 in a FPLC system (Pharmacia-LKB

311R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
Biotechnology, Sweden). The flux was 0.5 ml/min andfractions of 0.4 ml were collected. The active fractionswere pooled and applied to a column of Mono Q HR5/5 equilibrated with 20 mM imidazole buffer pH 6.4,in a FPLC system. The column was eluted with 0–600 mM NaCl gradient in the same buffer. The flux was1 ml/min and fractions of 0.4 ml were collected. Theactive fractions were pooled and dialysed overnightagainst 100 vol of water, with two changes.
2.5. Density-gradient ultracentrifugation
Samples (0.2 ml) of the acid extracts containing1.5 mg of bovine hemoglobin and 50 mg of bovine livercatalase were layered on the top of 10 ml glycerol gradi-ents (10–30%, w/w) made up in 50 mM sodium acetatepH 3.5. Centrifugation and collection of fractions wereperformed as described previously (Terra and Ferreira,1983). Mr (relative molecular weight) values of enzymesassayed in the fractions were calculated by the methodof Martin and Ames (1961), by using the sedimentationrates of bovine hemoglobin (Mr 64 500) and bovine livercatalase (Mr 232 000) as reference standards.
2.6. Isoelectric focusing in polyacrylamide gels
Isoelectric focusing was performed as described byTerra et al. (1978), in columns of 7% polyacrylamidegels containing 1% ampholytes pH 3–10. Samples wereapplied after polymerization and pre-focusing (30 min at31 V/cm) on the top of the alkaline side of the gels.Before electrofocusing, the samples were concentratedby centrifugation through Microcon 10 (Amicon, USA).
2.7. SDS-polyacrylamide gel electrophoresis
Samples containing approximately 4 mg protein werecombined with sample buffer containing 60 mM Tris-HCl buffer pH 6.8, 2.5% (w/v) SDS, 0.36 mMb-mer-captoethanol, 0.5 mM EDTA, 10% (v/v) glycerol and0.005% (w/v) bromophenol blue. The samples wereheated for 5 min at 95°C in a water bath before beingloaded onto a 15% (w/v) polyacrylamide gel slab con-taining 0.1% (w/v) SDS (Laemmli, 1970). The gels wererun at constant voltage of 200 V and stained for proteinusing a silver stain (Blum et al., 1987). Lysozyme Mrvalues were calculated according to Shapiro et al. (1967)using the following Mr standards: chicken lysozyme (Mr14 400); soybean trypsin inhibitor (Mr 21 500), carbonicanhydrase (Mr 31 000); ovalbumin(Mr 45 000), bovineserum albumin(Mr 66 200) and phosphorylase b (Mr97 400).
2.8. Microsequencing of purified lysozyme
D. melanogasterpurified lysozyme was concentratedby centrifuging in Microcon 10 (Amicon, USA) and then
combined with a volume of sample buffer containing62.5 mM Tris-HCl pH 6.75, 2% (w/w) SDS, 5% (v/v)b-mercaptoethanol, 10% (v/v) glycerol and 0.001% (v/v)bromophenol blue. The samples stood for 30 min at37°C before being loaded onto gel slabs. The 15% poly-acrylamide gel slabs containing 0.1% SDS were pre-pared 24 h before the run and were stored refrigeratedin a humid chamber. After a pre-run (10 mA, 10 min)with 0.1 M sodium thioglycolate in the run buffer, thesamples, pre-stained standards, and chicken lysozyme(2 mg) were loaded and electrophoretically resolved asdescribed above. The resolved peptides in the gel wereelectroblotted onto PVDF (polyvinylidene difluoride)membranes according to Matsudaira (1987). The PVDFmembranes were stained for proteins using 0.1% Coom-assie Blue R-250 in a 50% (v/v) methanol solution, andwere destained with a 50% methanol solution. DriedPVDF membranes were the source of peptides formicrosequencing.
After alkylation, the PVDF membrane was digestedby endoproteinase Lys-C, peptides resolved by reverse-phase HPLC and the chosen peptide (retention time81.8 min) was sequenced. Protein alkylation, enzymaticcleavage, peptide purification and sequence analysiswere performed at the Macromolecular Structure Analy-sis Facility at the University of Kentucky, Lexington,KY, USA.
2.9. Protein determination and hydrolase assays
Protein was determined according to Bradford (1976),using ovalbumin as a standard, or following the methodof Smith et al. (1985), as modified by Morton and Evans(1992), employing bovine serum albumin as a standard.
Lysozyme activity was determined by measuring thedecrease of turbidity at 650 nm of a suspension(1 mg/ml) ofMicrococcus lysodeikticusin (except other-wise specified) 50 mM citrate-sodium phosphate pH 3.5containing 150 mM NaCl, in a reaction volume of0.5 ml. The reaction was stopped by the addition of0.25 ml of 0.5 M sodium carbonate and the absorbancewas measured. One lysozyme unit is defined as theamount of enzyme that causes a change in theabsorbance of the reaction medium of 0.01/min.
Chitinase activity was measured using the viscometricassay of Winicur and Mitchell (1974), as modified byLemos and Terra (1991a), with chitosan as the substrate.The slopes of the plots of the inverse of specific activityagainst time in min were calculated. One chitinase unitis defined as the amount of enzyme that results in a slopeof 1000/min.
In each determination of the enzyme activities, incu-bations were carried out at 30°C for at least four differentperiods of time and the initial rates of hydrolysis werecalculated. All assays were performed under conditionssuch that activity was proportional to protein concen-

312 R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
tration and to time. Controls without enzyme or withoutsubstrate were included.
2.10. Kinetic studies
The effect of pH and ionic strength of reaction mediaon the activity of the purified lysozyme was determinedusing the following buffers (Miller and Golder, 1950):glycine-HCl (pH 2.0–3.5), sodium acetate (pH 4.0–5.5),sodium phosphate (pH 6.0–7.5) and Tris-HCl (pH 8.0)prepared with ionic strengths from 0.02 up to 0.2.
The effect ofM. lysodeikticusconcentration on thepurified lysozyme activity was determined at pH 4.3 atdifferent ionic strengths.Km values (means± SEM)were determined by a weighted linear regression usingthe program Enzfitter.
2.11. Amino acid and nucleotide sequence analyses
The sequences were visually aligned by using theSeqPup program (Gilbert, 1996). This program assignsdifferent colors to the residue letters, reflecting theamino acid chemical group in case of protein alignmentand the nucleotide in case of DNA alignment. The DNAalignment was constrained to the protein alignment.
The strategy to obtain the phylogenetic relationshipsamong the sequences and the ancestor sequences was asfollows: In the first step, an exhaustive search for themost parsimonious tree (the tree that requires the mini-mum number of amino acid substitutions) was perfor-med using the PAUP 3.1 program (Swofford, 1993) ina PowerMacintosh computer. This first step was perfor-med because a complete search with the likelihoodmethod would be prohibitive in terms of computationaltime. The most parsimonious trees with identical numberof steps were grouped in a single tree through the strictconsensus method. This tree was then used as a startingpoint to search for the maximum likelihood tree. To per-form this search, and also to obtain the estimated ances-tor amino acid sequences, we used the PAML package(Yang, 1997). This package was installed in a Cray EL98 supercomputer from the Laboratory of AdvancedScientific Computation of the University of Sa˜o Paulo(LCCA-USP). The maximum likelihood phylogeneticinferences of amino acid sequences were performedusing the empirical substitution matrix of Jones et al.(1992). The estimated nucleotide ancestor sequenceswere obtained by using the model of nucleotide substi-tution proposed by Felsenstein (1984), in his programDNAML and described by Hasegawa and Kishino(1989), that accounts for the nucleotide frequencies atthe extant sequences, and different transition and trans-version substitution rates.
3. Results
3.1. Purification of theD. melanogastermajor larvallysozyme
Based on the fact that the lysozymes from the relatedinsectMusca domesticaare acid and heat stable (Lemoset al., 1993), D. melanogasterlarval midguts werehomogenized in 100 mM acetate buffer pH 3.5, centri-fuged, and the resulting supernatants heated for 5 min ina boiling water bath before being centrifuged again and,finally, dialysed against water. The dialysates were usedas a source of lysozyme.
Three lysozymes fromD. melanogasterlarval dialys-ate acid extract were resolved by isoelectric focusing(Fig. 1A) with pI values (four determinations, mean±SEM): 4.7± 0.1, 5.5± 0.1, and 7.2± 0.2. Only pI 5.5 —lysozyme is detected if fractions obtained by isoelectricfocusing are assayed at pH 3.5 (Fig. 1B). Three lyso-zymes were also resolved by polyacrylamide gel electro-
Fig. 1. Isoelectric focusing of lysozymes in the dialysed acid extractof D. melanogasterlarvae. (A) Assays in 50 mM citrate-sodium phos-phate pH 5.0 containing 150 mM NaCl. (B) Assays in 50 mM citrate-sodium phosphate pH 3.5 containing 150 mM NaCl.
313R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
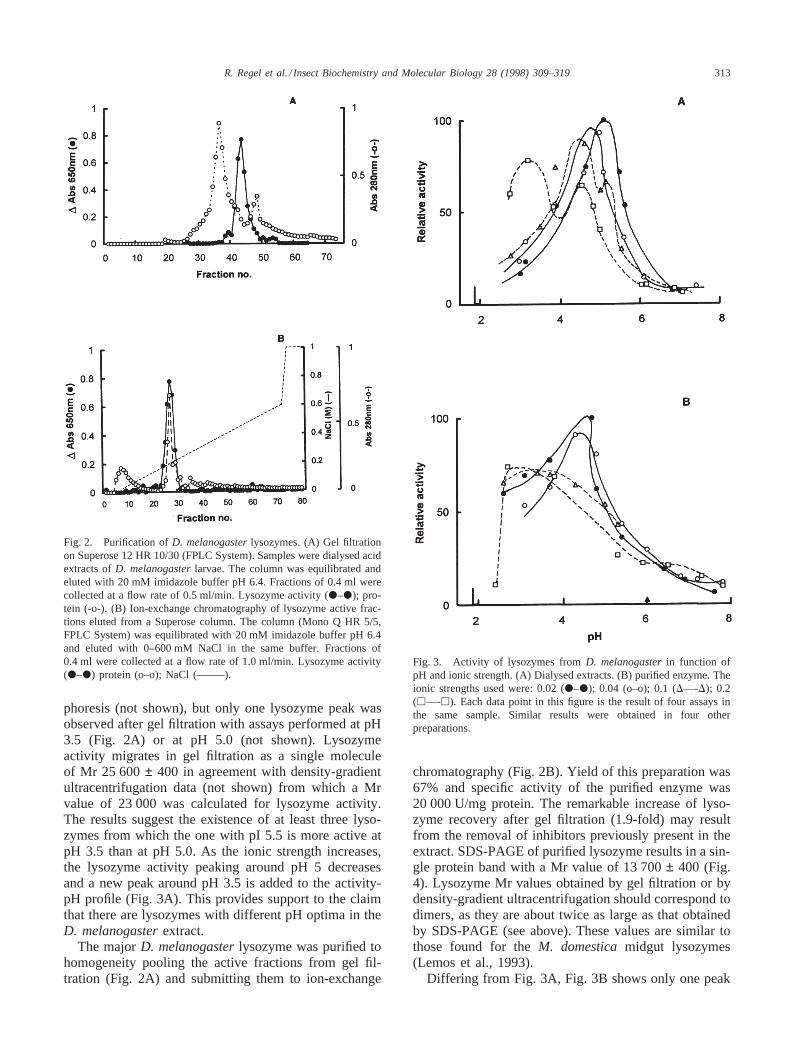
Fig. 2. Purification ofD. melanogasterlysozymes. (A) Gel filtrationon Superose 12 HR 10/30 (FPLC System). Samples were dialysed acidextracts ofD. melanogasterlarvae. The column was equilibrated andeluted with 20 mM imidazole buffer pH 6.4. Fractions of 0.4 ml werecollected at a flow rate of 0.5 ml/min. Lysozyme activity (P–P); pro-tein (-o-). (B) Ion-exchange chromatography of lysozyme active frac-tions eluted from a Superose column. The column (Mono Q HR 5/5,FPLC System) was equilibrated with 20 mM imidazole buffer pH 6.4and eluted with 0–600 mM NaCl in the same buffer. Fractions of0.4 ml were collected at a flow rate of 1.0 ml/min. Lysozyme activity(P–P) protein (o–o); NaCl (——–).
phoresis (not shown), but only one lysozyme peak wasobserved after gel filtration with assays performed at pH3.5 (Fig. 2A) or at pH 5.0 (not shown). Lysozymeactivity migrates in gel filtration as a single moleculeof Mr 25 600± 400 in agreement with density-gradientultracentrifugation data (not shown) from which a Mrvalue of 23 000 was calculated for lysozyme activity.The results suggest the existence of at least three lyso-zymes from which the one with pI 5.5 is more active atpH 3.5 than at pH 5.0. As the ionic strength increases,the lysozyme activity peaking around pH 5 decreasesand a new peak around pH 3.5 is added to the activity-pH profile (Fig. 3A). This provides support to the claimthat there are lysozymes with different pH optima in theD. melanogasterextract.
The majorD. melanogasterlysozyme was purified tohomogeneity pooling the active fractions from gel fil-tration (Fig. 2A) and submitting them to ion-exchange
Fig. 3. Activity of lysozymes fromD. melanogasterin function ofpH and ionic strength. (A) Dialysed extracts. (B) purified enzyme. Theionic strengths used were: 0.02 (P–P); 0.04 (o–o); 0.1 (D—-D); 0.2(h—-h). Each data point in this figure is the result of four assays inthe same sample. Similar results were obtained in four otherpreparations.
chromatography (Fig. 2B). Yield of this preparation was67% and specific activity of the purified enzyme was20 000 U/mg protein. The remarkable increase of lyso-zyme recovery after gel filtration (1.9-fold) may resultfrom the removal of inhibitors previously present in theextract. SDS-PAGE of purified lysozyme results in a sin-gle protein band with a Mr value of 13 700± 400 (Fig.4). Lysozyme Mr values obtained by gel filtration or bydensity-gradient ultracentrifugation should correspond todimers, as they are about twice as large as that obtainedby SDS-PAGE (see above). These values are similar tothose found for theM. domesticamidgut lysozymes(Lemos et al., 1993).
Differing from Fig. 3A, Fig. 3B shows only one peak

314 R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
Fig. 4. Electrophoresis in SDS 15% polyacrylamide gel slab of larvalextracts fromD. melanogaster. Lane 1, acid extract; 2, heated acidextract; 3, dialysed acid extract; 4, Superose eluate; 5, Mono Q eluate;6, molecular weight markers. About 3 mg of proteins were applied toeach lane. Lane 2 apparently has less protein than the other lanesbecause heated acid extracts contain compounds that interfere withprotein determination.
of activity, whichever ionic strength has been used inthe assay. This confirms that the purified enzyme corre-sponds to a single molecular species. The same con-clusion can be drawn from the finding of a single peakof lysozyme activity with pI 5.5 after isoelectric focusingfollowed by assays at pH 3.5 or pH 5.0 (not shown).
3.2. Origin ofD. melanogasterlarval lysozymes
The purified lysozyme was cleaved with an endoprote-inase, the peptides resolved by reverse phase chromatog-raphy and one of them (retention time 81.8 min) wassequenced. The sequence found was: DDITHSVxCAQxVLSQQG, where x corresponds to non-identifiedamino acids. A search on the GenBank showed that thepeptide corresponds to a sequence of the D-family lyso-zymes fromD. melanogaster. This family contains 5lysozymes, from which lysozyme D differs from lyso-zymes B and C only in the signal peptide and from lyso-zymes E and A in one and two amino acids, respectively.Further support for the identification of the purifiedD.melanogasterlysozyme with the lysozymes D comesfrom the finding that the pI determined for the purifiedlysozyme (5.5, see above) is close to that one calculated(5.1) by Daffre et al. (1994) from the sequences of thelysozyme D. These lysozymes are the majorD. mel-anogasterlysozymes and are expressed only in midguts(Kylsten et al., 1992). Thus the purified enzyme corre-sponds to the major midgut lysozyme fromD. mel-anogaster. The minor lysozymes with pI values 4.7 and7.2 should correspond to lysozyme S and lysozyme Pfor which Daffre et al. (1994) calculated pI values of 4.4and 8.6, respectively.
3.3. Kinetic properties of the purifiedD. melanogasterlysozyme
D. melanogasterpurified lysozyme activity decreases(Fig. 3B), its pH optimum is displaced toward acidic pHvalues (Fig. 3B) and itsKm value increases (Fig. 5) asthe ionic strength of its assay medium becomes higher.Similar results were also found for other enzymes(Maurel and Douzou, 1976; Dobson et al., 1984), includ-ing M. domestica(Lemos et al., 1993). In contrast towhat is observed with the enzyme fromM. domestica(Lemos et al., 1993), excess substrate does not causeinhibition of the purified D. melanogasterlysozyme(not shown).
Ten units of the purified lysozyme display 2.1±0.1 mU of chitinase (viscometric assay) at pH 3.5,whereas chicken lysozyme presents 0.20± 0.02 mU inthe same conditions. Thus,D. melanogasterlysozymeshows a chitinase activity about 11-fold higher than thatof chicken lysozyme.
D. melanogasterpurified lysozyme maintained in thepresence ofMusca domesticamidgut homogenates con-taining 2.0 units of cathepsin D for 180 min in 100 mMacetate buffer pH 3.3 at 30°C retains full activity.Cathepsin D is the acid proteinase present in the middleregion of midgut from Diptera Cyclorrhapha (Terra andFerreira, 1994). Nevertheless, the purified lysozymeincubated with 2.4 units of bovine pepsin loses 42% ofits activity after 180 min in 100 mM acetate buffer pH3.5 at 30°C.
3.4. Reconstructing lysozyme amino acid sequences
A maximum likelihood method was employed toreconstruct the ancestral amino acid sequences of insectlysozymes. With this aim, enzymes from the followinginsects were used: The DipteraD. melanogaster
Fig. 5. Effect of the ionic strength onKm values of the purifiedlysozyme fromD. melanogasterat pH 4.3.

315R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
(Lysozymes D, P and S) (Kylsten et al., 1992; Daffre etal., 1994),M. domestica(Ito et al., 1995),Anophelesdarlingi (Moreira et al., GenBank — accession numberAF 003945) andAnopheles gambiae(Kang et al., Gen-Bank — accession number U 28809), and the Lepidop-tera Hyalophora cecropia(Engstrom et al., 1985; Sunet al., 1991),Manduca sexta(Mulnix and Dunn, 1994),Bombyx mori(Lee and Brey, 1995),Hyphantria cunea(Park et al., GenBank — accession number U 23788)andTrichoplusia ni(Kang et al., GenBank — accessionnumber U 38782).
A phylogenetic tree was produced (Fig. 6) definingseveral ancestors. The common ancestor for Lepidopteraand Diptera was named Panorpoid ancestor, becauseLepidoptera and Diptera are panorpoid orders. The otherancestors were Diptera, for Nematocera andCyclorrhapha; Nematocera, forA. gambiaand A. dar-lingi; Cyclorrhapha, forD. melanogasterand M. dom-estica; digestive lysozyme ancestor, forM. domesticaandD. melanogaster(except for LysP) and Lepidoptera,for the other insects. The ancestor sequences and phylo-genetic trees were also estimated by using the nucleotidedata (not shown). The results were slightly different butthey lead to the same conclusions.
The tree in Fig. 6 differs from the one from Ito et al.(1995) in placing theM. domesticalysozyme sequencewithin theD. melanogasterlysozyme family. The differ-ence found probably resulted from the fact that, in con-trast to Ito et al. (1995), we included two Nematocerasequences and employed the maximum likelihoodmethod.
Relevant reconstructed ancestor sequences, the actualinsect sequences, together with the sequences of some
Fig. 6. Phylogenetic tree of insect lysozymes obtained by the maximum likelihood method for reconstruction of ancestral sequences. 1, Panorpoidancestor; 2, Lepidoptera ancestor; 3, Diptera ancestor; 4, Nematocera ancestor; 5, Cyclorrhapha ancestor; 6, digestive lysozyme ancestor.
vertebrate lysozymes are shown in Fig. 7. The vertebratesequences come from the following species: human(Homo sapiens, Chung et al., 1988), the ruminant cow(Bos taurus, Jolles et al., 1984, 1989; Irwin and Wilson,1989) and the leaf-fermenter monkeyPresbytis entellus(Stewart et al., 1987). An inspection of Fig. 7 showsthat some positions are occupied by the same amino acidresidues, such as those of the 8 cysteines, and position36, occupied by glutamate, known to be involved incatalysis in chicken lysozyme (Imoto et al., 1972). Pos-itions 54–61 are also occupied by the same amino acidsin all lysozymes and include aspartate 54, which isinvolved in catalysis, and other residues shown to bindthe substrate in chicken lysozyme (Imoto et al., 1972).In some lysozymes, leucine was replaced by isoleucine57, although this change is not expected to affect sig-nificantly the binding site. Some amino acid sequencesoccur only in insect lysozymes, such as those of pos-itions 50–52 and 90–93. The replacement of serine forthreonine 93 inH. cecropiaand M. domesticais not amajor change. The same is true for the replacement ofasparagine for aspartate 91 inM. domestica. Perhaps themost interesting observation is the occurrence of serineat the position 104 only in Cyclorrhapha and vertebratedigestive lysozymes.
A comparison among the reconstructed ancestralsequences of insect lysozymes shows that the digestivelysozyme ancestor sequence has a decrease in the num-ber of basic amino acids, caused by a diminution inlysine residues.

316R
.R
eg
el
et
al./In
sect
Bio
che
mistry
an
dM
ole
cula
rB
iolo
gy
28
(19
98
)3
09
–3
19
Fig. 7. Amino acid sequences of insect lysozymes, some vertebrate lysozymes, and reconstructed ancestral insect lysozymes. Ancestral insect lysozyme sequences were reconstructed with themaximum likelihood method. The residues are shown only where different from the preceding baseline sequence, with identity indicated by a dot. Dashes denote deletions relative to othersequences and question marks, undetermined residues. The sequences were aligned with the aid of the program SeqPup-version 0.6e which colors differentially the different amino acids makingeasy visual alignment.

317R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
4. Discussion
4.1. D. melanogasterlarval lysozymes
D. melanogasterlarval homogenates have a major (pI5.5) and two minor (pI 4.7 and 7.2) lysozyme activities.The major lysozyme activity was purified to homogen-eity and was partially sequenced, revealing that it corre-sponds to the D-lysozyme family. These lysozymes areexpressed only in the midgut ofD. melanogaster(Kylsten et al., 1992; Daffre et al., 1994).
The purified lysozyme has a Mr value of 13 700, whenSDS-polyacrylamide gel electrophoresis was employed,and may be found as dimers (Mr 23 000) or tetramers(Mr 42 000) during ultracentrifugation in glycerol gradi-ents. Thus,D. melanogastermidgut lysozyme has a Mrvalue which is similar to that of most lysozymes, includ-ing those fromM domestica(Lemos et al., 1993).
Like M. domesticalysozymes (Lemos et al., 1993),D. melanogasterpurified lysozyme shows a chitinaseactivity much higher than that of chicken lysozyme, apI value lower than those of most animals (Imoto et al.,1972), although it is similar to that of ruminant lyso-zymes (Pahud and Widmer, 1982), and the samebehavior in relation to pH. ThusD. melanogasterpur-ified lysozyme becomes less active, its pH-activity pro-file is displaced toward the acid side, andKm valuesenlarge as the ionic strength of the medium becomeshigher. Also in agreement withM. domesticadata, themagnitude of the observed changes are only comparablewith ruminant lysozymes (Dobson et al., 1984). Theseproperties, as discussed below, are related to a diges-tive function.
The pH-lysozyme activity profiles ofD. melanogasterlarval homogenates show that there is an activity peakaround pH 5, in addition to peak at pH 3.5 correspondingto the major midgut lysozyme.M. domesticahas a minorlysozyme activity in the salivary glands which is moreactive at pH 6, than at pH 3.8 (Lemos and Terra, 1991a).Thus, it is probable that the pH 5 lysozyme activity peakobserved inD. melanogasterhomogenates (Fig. 3A) cor-responds to lysozyme P, which is only expressed in sali-vary glands (Kylsten et al., 1992). If true, this meansthat lysozyme P is not adapted to function in the veryacid middle midgut contents, but only in the buccal cav-ity and anterior midgut contents. Although lysozyme Pmay initiate bacterial lysis in anterior midgut, its physio-logical meaning there should not be important, becauseit occurs in low amounts andM. domesticadata showedthat bacteria is significantly killed only in the acidmiddle midgut (Greenberg, 1959). According to thisview, lysozyme P is not a true digestive enzyme, as lyso-zyme D, and it also differs from the more common lyso-zymes (those involved in defense) in having a low pI(7.2). It seems that this enzyme is an intermediate inthe evolution of the digestive lysozymes. The fact that
lysozyme P sequence is not close to the otherD. mel-anogasterlysozymes favors this view (see also below).
The pI 4.7-lysozyme activity observed inD. mel-anogaster larval homogenates should correspond tolysozyme S, which is expressed only in midgut caeca(Kylsten et al., 1992; Daffre et al., 1994). The data isnot sufficient to advance any hypothesis on the meaningof this enzyme, which is present in small amounts.
4.2. Molecular adaptation of lysozymes to a digestivefunction
The features invariably shared by the vertebrate diges-tive lysozymes are a high level in the stomach, a low pHoptimum at physiological ionic strength and resistance topepsin (Prager, 1996). A low pH optimum seems toresult from the presence of serine 104 (numbered as inFig. 7), responsible for the activity at low pH values andfrom a low pI value, which causes a decrease of theactivity at high pH values (Stewart and Wilson, 1987).It is not clear how serine 104 acts, but since it islocalized near the substrate binding cleft (Stewart andWilson, 1987), it should affect thepKa values of thecatalytic active groups changing the enzyme activity.The decrease in activity observed at higher pH valuesresults from a diminished interaction of the negative sub-strate (polyanionic carbohydrate chains in the bacterialcell walls) with an increasingly less positive enzyme.Lower pI values are a consequence of a smaller contentin basic amino acids and a greater amount of acid aminoacids (Prager, 1996). Resistance to pepsin seems todepend on the elimination of susceptible peptide bondsand/or on conformational changes that hide them. Thus,the nature of the adaptation to an acid proteolyticenvironment is largely dependent on the ancestor of thelysozyme present sequence and is, as a consequence,highly variable (Prager, 1996).
D. melanogastermajor midgut lysozyme (lysozymesD) andM domesticalysozyme have, in their sequences,positions occupied by the same amino acids as in allvertebrate lysozymes C. This is true for 8 cysteines, pos-itions 55–61 (numbered as in Fig. 7) (substrate bindingcleft), glutamate 36 and aspartate 54 (Fig. 7 and Imotoet al., 1972). Glutamate 36 is believed to act as a generalacid in catalysis (Imoto et al., 1972), whereas aspartate54 is postulated to stabilize the resulting metastableoxocarbonium intermediate (Imoto et al., 1972),although recently aspartate 54 has been implicated morestrongly in catalysis of the hydrolysis of chitin-derivedsubstrates (Matsumura and Kirsch, 1996). The ability ofD. melanogasterand M. domesticapurified lysozymesin hydrolyzing chitosan favors this view.
The most remarkable sequence convergence ofD.melanogasterlysozymes D andM. domesticalysozymesfrom vertebrate foregut fermenters are serine 104 and adecrease in the number of basic amino acids, suggesting

318 R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
that these features are necessary for a digestive role in anacid environment. In vertebrate adaptation to digestivefunction, the lysozymes presented an increase in acidamino acids accompanied by a reduction in basic aminoacid numbers. InD. melanogasterandM. domestica, wecould only observe a reduction in the basic amino acidnumber. However, this reduction was enough to reducethe observed pI values. The lack of putative adaptiveresidues in lysozyme D andM. domesticalysozyme(those conferring stability in an acid proteolyticenvironment) described for digestive vertebrate lyso-zymes (Prager, 1996) is not surprising. Both insect lyso-zymes sequences are shorter, with gaps assigned relativeto the vertebrate enzymes in a number of places (Prager,1996), and have specific sequences and more prolines.Although some convergences between theD. mel-anogasterand M. domesticaand vertebrate lysozymescan be postulated, the overall pattern is different. Thismay be attributed by structural constraints that may haveevolved in a different fashion during the evolution of thelysozymes in vertebrate and insect lineages. Changes indifferent sites may be also adaptive to the digestiveactivity.
D. melanogasterlysozyme P lacks serine 104, whichagrees with the hypothesis presented above that it is nota true digestive enzyme and that it is not active at lowpH values. Since it has a low pI value, it seems to bestanding midway between a typical C-type lysozyme anda digestive one.
The placement of theD. domestica lysozymesequence within theD. melanogasterlysozyme family(Fig. 6) may be regarded as a result of adaptive evolutionof both M. domesticaand M. melanogasterdigestivelysozymes from an ancestral molecule that was nondigestive. In other words, we may hypothesize that inCyclorrhapha, ancestor lysozyme gene duplications haveoccurred and some lysozyme gene products wererecruited to a digestive function and others not. The twosets of genes evolved independently. The fact thatD.melanogasterlysozyme P (which probably is not a truedigestive enzyme) sequence stands outside the cluster ofM. domesticaandD. melanogasterdigestive lysozymesagrees with this hypothesis. Knowledge on the otherM.domestica lysozyme sequences would certainly shedmore light on this subject.
Acknowledgements
We are much indebted to Dr. C. R. Vilella for his helpin D. melanogasterraising, to Drs. C. Ferreira and S.Daffre for helpful discussion, to the Laboratory ofAdvanced Scientific Computation (University of Sa˜oPaulo) for computational facilities, and to Miss L. Y.Nakabayashi for technical assistance This work was sup-ported by the Brazilian Research Agencies FAPESP and
CNPq. R. Regel is a graduate fellow of FAPESP. S. R.Matioli and W. R. Terra are staff members of theirrespective departments and research fellows of CNPq.
References
Blum, H., Beir, H., Gross, H., 1987. Improved silver staining of plantproteins, RNA and DNA in polyacrylamide gels. Electrophoresis8, 93–99.
Bradford, M.M., 1976. A rapid and sensitive method for the quantit-ation of microgram quantities of protein utilizing the principle ofprotein-dye binding. Analyt. Biochem. 72, 248–254.
Chung, L.P., Keshav, S., Gordon, S., 1988. Cloning the human lyso-zyme cDNA: invertedAlu repeat in the mRNA andin situhybridiz-ation for macrophages and Paneth cells. Proc. Natl. Acad. Sci. USA85, 6227–6231.
Daffre, S., Kylsten, P., Samakovlis, C., Hultmark, D., 1994. The lyso-zyme locus inDrosophila melanogaster: an expanded gene familyadapted for expression in the digestive tract. Mol. Gen. Genet. 242,152–162.
Dobson, D.E., Prager, E.M., Wilson, A.C., 1984. Stomach lysozymesof ruminants. I. Distribution and catalytic properties. J. Biol. Chem.259, 11607–11616.
Dunn, P.E., 1986. Biochemical aspects of insect immunology. A. Rev.Ent. 31, 321–339.
Engstrom, A., Xanthopoulos, K.G., Boman, H.G., Bennich, H., 1985.Amino acid and cDNA sequences of lysozyme fromHyalophoracecropia. EMBO J. 4, 2119–2122.
Gilbert, D. 1996. SeqPup: a biosequence editor and analysis appli-cation, version 0.6d Biology Department, Indiana University.
Greenberg, B., 1959. Persistence of bacteria in the developmentalstages of the housefly. II. Quantitative study of the host-contami-nant relationship in flies breeding under natural conditions. Am. J.Trop. Med. Hyg. 8, 412–416.
Hasegawa, M., Kishino, H., 1989. Confidence limits on the maximumlikelihood estimation of the hominoid tree from mitochondrialDNA sequences. Evolution 43, 672–677.
Hultmark, D. 1996. Insect lysozymes. In: Jolle`s, P. (Ed.), Lysozymes:Model Enzymes in Biochemistry and Biology, Birkha¨user Verlag,Basel, pp. 87–102.
Imoto, T., Johnson, L. N., North, A. C. T., Phillips, D. C. and Rupley,J. A. 1972. Vertebrate Lysozymes. In: Boyer, P. D. (Ed.), TheEnzymes, Vol. 7, Academic Press, New York, pp. 665–868.
Irwin, D.M., Wilson, A.C., 1989. Multiple cDNA sequences and theevolution of bovine stomach lysozyme. J. Biol. Chem. 264,11387–11393.
Irwin, D.M., Wilson, A.C., 1990. Concerted evolution of ruminant sto-mach lysozymes: characterization of lysozyme cDNA clones fromsheep and deer. J. Biol. Chem. 265, 4944–4952.
Ito, Y., Nakamura, M., Hotani, T., Imoto, T., 1995. Insect lysozymefrom house fly (Musca domestica) larvae: possible digestive func-tion based on sequence and enzymatic properties. J. Biochem.Tokyo 118, 546–551.
Jolles, P., Jolle`s, J., 1984. What’s new in lysozyme research? Molec.Cell Biochem. 63, 165–189.
Jolles, P., Schoentgen, F., Jolle`s, J., Dobson, D.E., Prager, E.M., Wil-son, A.C., 1984. Stomach lysozymes of ruminants. II. Amino acidsequence of cow lysozyme 2 and immunological comparisons withother lysozymes. J. Biol. Chem. 259, 11617–11625.
Jolles, J., Jolle`s, P., Bowman, B.H., Prager, E.M., Stewart, C.B., Wil-son, A.C., 1989. Episodic evolution in the stomach lysozymes ofruminants. J. Mol. Evol. 28, 528–535.
Jones, D.T., Taylor, W.R., Thornton, J.M., 1992. The rapid generationof mutation data matrices from protein sequences. Computer Appli-cation in Biosciences 8, 275–282.

319R. Regel et al. / Insect Biochemistry and Molecular Biology 28 (1998) 309–319
Kylsten, P., Kimbrell, D.A., Daffre, S., Samakovlis, C., Hultmark, D.,1992. The lysozyme locus inDrosophila melanogaster: differentgenes are expressed in midgut and salivary glands. Molec. Gen.Genet. 232, 335–343.
Laemmli, U.K., 1970. Cleavage of structural proteins during theassembly of the head of the bacteriophage T4. Nature 227, 680–685.
Lee, W.J., Brey, P.T., 1995. Isolation and characterization of the lyso-zyme-encoding gene from the silkwormBombyx mori. Gene 161,199–203.
Lemos, F.J.A., Terra, W.R., 1991a. Digestion of bacteria and the roleof midgut lysozyme in some insect larvae. Comp. Biochem. Phy-siol. 100B, 265–268.
Lemos, F.J.A., Terra, W.R., 1991b. Properties and intracellular distri-bution of a cathepsin D-like proteinase active at the acid region ofMusca domesticamidgut. Insect Biochem. 21, 457–465.
Lemos, F.J.A., Ribeiro, A.F., Terra, W.R., 1993. A bacteria-digestingmidgut-lysozyme fromMusca domestica(Diptera) larvae. Purifi-cation, properties and secretory mechanism. Insect Biochem.Molec. Biol. 23, 533–541.
Martin, R.G., Ames, B.N., 1961. A method for determining the sedi-mentation behavior of enzymes: application to protein mixtures. J.Biol. Chem. 236, 1372–1379.
Matsudaira, P., 1987. Sequence from picomole quantities of proteinselectroblotted onto polyvinylidine difluoride membranes. J. Biol.Chem. 262, 10035–10038.
Matsumura, I., Kirsch, J.F., 1996. Is Aspartate 52 essential for catalysisby chicken egg white lysozyme? The role of natural substrate-assisted hydrolysis. Biochemistry 35, 1881–1889.
Maurel, P., Douzou, P., 1976. Catalytic implications of electrostaticpotentials: the lytic activity of lysozyme as a model. J. Molec. Biol.102, 253–264.
McHenery, J.G., Birkbeck, T.H., Allen, S.A., 1979. The occurrenceof lysozyme in marine bivalves. Comp. Biochem. Physiol. 63B,25–28.
Miller, G.L., Golder, R.H., 1950. Buffers of pH 2 to 12 for use inelectrophoresis. Arch. Biochem. 29, 420–423.
Morton, R.E., Evans, T.A., 1992. Modifications of the bicinchoninicacid protein assay to eliminate lipid interference in determininglipoprotein protein content. Anal. Biochem. 204, 332–334.
Mulnix, A.M., Dunn, P.E., 1994. Structure and induction of a lyso-zyme gene from the tobacco hornwormManduca sexta. InsectBiochem. Molec. Biol. 24, 271–281.
Pahud, J.J., Widmer, F., 1982. Calf rennet lysozyme. Biochem. J. 201,661–664.
Prager, E. M. 1996. Adaptive evolution of lysozyme: changes in aminoacid sequence, regulation of expression and gene number. In Jolle`s,P. (Ed.), Lysozymes: Model Enzymes in Biochemistry andBiology, Birkhauser Verlag, Basel, pp. 323–345.
Shapiro, A.L., Vinuela, E., Maizel, J.V., 1967. Molecular weight esti-mation of polypeptide chains by electrophoresis in SDS-polyacryla-mide gels. Biochem. Biophys. Res. Commun. 28, 815–820.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gartner,F.H., Provenzano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J.,Klerk, D.C., 1985. Measurements of protein using bicinchoninicacid. Anal. Biochem. 150, 76–85.
Stewart, C.B., Wilson, A.C., 1987. Sequence convergence and func-tional adaptation of stomach lysozymes from foregut fermenters.Cold Spring Harb. Symp. Quant. Biol. 52, 891–899.
Stewart, C.B., Schilling, J.W., Wilson, A.C., 1987. Adaptive evolutionin the stomach lysozymes of foregut fermenters. Nature 330,401–404.
Sun, S.C., Asling, B., Faye, I., 1991. Organization and expression ofthe immunoresponsive lysozyme gene in the giant silk mothHyalo-phora cecropia. J. Biol. Chem. 266, 6644–6649.
Swofford, D. L. 1993. Phylogenetic Analysis using Parsimony(PAUP), version 3.2. University of Illinois, Champaign.
Terra, W.R., 1990. Evolution of digestive systems of insects. A. Rev.Ent. 35, 181–200.
Terra, W.R., Ferreira, C., De Bianchi, A.G., 1978. Physical propertiesand Tris inhibition of an insect trehalase and a thermodynamicapproach to the nature of its active site. Biochim. Biophys. Acta524, 131–141.
Terra, W.R., Ferreira, C., 1983. Further evidence that enzymesinvolved in the final stages of digestion ofRhynchosciara amer-icana do not enter the endoperitrophic space. Insect Biochem. 13,143–150.
Terra, W.R., Ferreira, C., 1994. Insect digestive enzymes: properties,compartmentalization and function. Comp. Biochem. Physiol.109B, 1–62.
Winicur, S., Mitchell, H.K., 1974. Chitinase activity duringDrosophiladevelopment. J. Insect Physiol. 20, 1775–1805.
Yang, Z. 1997. Phylogenetic analysis by Maximum Likelihood(PAML), version 1.3. Department of Integrative Biology, Univer-sity of California at Berkeley.
Copyright © 2022 FDOKUMEN




















