
Modulation of medial temporal lobe activity in epilepsy patients with hippocampal sclerosis during...
11
Journal of the International Neuropsychological Society (2009), 15, 1–10. Copyright © 2009 INS. Published by Cambridge University Press. Printed in the USA. doi:10.1017/S135561770909078X 1 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 INTRODUCTION Although traditional models have long considered that me- dial temporal lobe (MTL) structures are crucial for the for- mation of long-term memories (Eichenbaum, 1997), but not for working memory (WM) (Cave & Squire, 1992; Gabrieli, 1998; Squire et al., 2004), there is an emerging view support- ing the essential contribution of MTL in WM processes (for a review, see Gazzaley et al., 2004; Ranganath & Blumenfeld, 2005). Data from animal lesion and neurophysiological ex- periments (Davachi & Goldman-Rakic, 2001; Holscher & Rolls, 2002; Suzuki et al., 1997; Young et al., 1997), as well as findings from human neuropsychological studies (Haettig et al., 2004; Lancelot et al., 2003; Nichols et al., 2006; Olson et al., 2006; Owen et al., 1996; Wagner et al., 2008), provide evidence for the critical involvement of MTL in WM. Recent human neuroimaging studies provide further support for the relevant contribution of MTL activation during WM (Axmacher et al., 2007, 2008; Cabeza et al., 2002; Glabus et al., 2003; Karlsgodt et al., 2005; Lee et al., 2006; Manoach et al., 2003; Monk et al., 2002; Petersson et al., 2006; Ranganath & D’Esposito, 2001; Schon et al., 2004; Stern et al., 2001; Tesche & Karhu, 2000). While most of the neuroscientific research has focused on the study of brain activity during WM maintenance, “the neu- ral mechanisms for WM encoding have not been extensively investigated” (Ranganath et al., 2004). Several authors AU2 Modulation of medial temporal lobe activity in epilepsy patients with hippocampal sclerosis during verbal working memory PABLO CAMPO, 1 FERNANDO MAESTÚ, 1,2 IRENE GARCÍA-MORALES, 3,4 ANTONIO GIL-NAGEL, 4 BRYAN STRANGE, 5 MANUEL MORALES, 6 and TOMÁS ORTIZ 1 1 Center of Magnetoencephalography Dr. Pérez-Modrego, Complutense University of Madrid, Madrid, Spain 2 Department of Basic Psychology II (Cognitive Processes), Faculty of Psychology, Complutense University of Madrid, Madrid, Spain 3 Epilepsy Unit, Neurology Department, University Hospital of San Carlos, Madrid, Spain 4 Epilepsy Unit, Neurology Department, Hospital Rúber Internacional, Madrid, Spain 5 Wellcome Department of Imaging Neuroscience, Institute of Neurology, London, United Kingdom 6 Department of Experimental Psychology, University of Seville, Seville, Spain (Received Month 00, 2008; Final Revision Month 00, 2008; Accepted Month 00, 2008) Abstract It has been traditionally assumed that medial temporal lobe (MTL) is not required for working memory (WM). However, animal lesion and electrophysiological studies and human neuropsychological and neuroimaging studies have provided increasing evidences of a critical involvement of MTL in WM. Based on previous findings, the central aim of this study was to investigate the contribution of the MTL to verbal WM encoding. Here, we used magnetoencephalogra- phy (MEG) to compare the patterns of MTL activation of 9 epilepsy patients suffering from left hippocampal sclerosis with those of 10 healthy matched controls while they performed a verbal WM task. MEG recordings allow detailed tracking of the time course of MTL activation. We observed impaired WM performance associated with changes in the dynamics of MTL activity in epilepsy patients. Specifically, whereas patients showed decreased activity in damaged MTL, activity in the contralateral MTL was enhanced, an effect that became significant in the 600- to 700-ms interval after stimulus presentation. These findings strongly support the crucial contribution of MTL to verbal WM encoding and provide compelling evidence for the proposal that MTL contributes to both episodic memory and WM. Whether this pattern is signaling reorganization or a normal use of a damaged structure is discussed. ( JINS, 2009, 15, 1–10.) Keywords: Magnetoencephalography, Hippocampus, Memory, Memory disorders, Epilepsy, Epilepsy surgery AU1 P.C. and F.M. contributed equally to the study. Correspondence and reprint requests to: Pablo Campo, Centro de Mag- netoencefalografía Dr. Pérez Modrego, Pabellón nº 8 Facultad de Medicina, Universidad Complutense de Madrid, 28040 Madrid, Spain. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Modulation of medial temporal lobe activity in epilepsy patients with hippocampal sclerosis during...

Journal of the International Neuropsychological Society (2009), 15 , 1 – 10 .Copyright © 2009 INS. Published by Cambridge University Press. Printed in the USA.doi:10.1017/S135561770909078X
1
12345678910111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455
INTRODUCTION
Although traditional models have long considered that me-dial temporal lobe (MTL) structures are crucial for the for-mation of long-term memories (Eichenbaum, 1997 ), but not for working memory (WM) (Cave & Squire, 1992 ; Gabrieli, 1998 ; Squire et al., 2004 ), there is an emerging view support-ing the essential contribution of MTL in WM processes (for a review, see Gazzaley et al., 2004 ; Ranganath & Blumenfeld, 2005 ). Data from animal lesion and neurophysiological ex-periments (Davachi & Goldman-Rakic, 2001 ; Holscher &
Rolls, 2002 ; Suzuki et al., 1997 ; Young et al., 1997 ), as well as fi ndings from human neuropsychological studies (Haettig et al., 2004 ; Lancelot et al., 2003 ; Nichols et al., 2006 ; Olson et al., 2006 ; Owen et al., 1996 ; Wagner et al., 2008 ), provide evidence for the critical involvement of MTL in WM. Recent human neuroimaging studies provide further support for the relevant contribution of MTL activation during WM (Axmacher et al., 2007 , 2008 ; Cabeza et al., 2002 ; Glabus et al., 2003 ; Karlsgodt et al., 2005 ; Lee et al., 2006 ; Manoach et al., 2003 ; Monk et al., 2002 ; Petersson et al., 2006 ; Ranganath & D’Esposito, 2001 ; Schon et al., 2004 ; Stern et al., 2001 ; Tesche & Karhu, 2000 ).
While most of the neuroscientifi c research has focused on the study of brain activity during WM maintenance, “the neu-ral mechanisms for WM encoding have not been extensively investigated” (Ranganath et al., 2004 ). Several authors
AU2
Modulation of medial temporal lobe activity in epilepsy patients with hippocampal sclerosis during verbal working memory
PABLO CAMPO , 1 FERNANDO MAESTÚ , 1 , 2 IRENE GARCÍA-MORALES , 3 , 4 ANTONIO GIL-NAGEL , 4 BRYAN STRANGE , 5 MANUEL MORALES , 6 and TOMÁS ORTIZ 1 1 Center of Magnetoencephalography Dr. Pérez-Modrego , Complutense University of Madrid , Madrid , Spain 2 Department of Basic Psychology II (Cognitive Processes) , Faculty of Psychology , Complutense University of Madrid , Madrid , Spain 3 Epilepsy Unit , Neurology Department , University Hospital of San Carlos , Madrid , Spain 4 Epilepsy Unit , Neurology Department , Hospital Rúber Internacional , Madrid , Spain 5 Wellcome Department of Imaging Neuroscience , Institute of Neurology , London , United Kingdom 6 Department of Experimental Psychology , University of Seville , Seville , Spain
(Received Month 00 , 2008 ; Final Revision Month 00 , 2008 ; Accepted Month 00 , 2008 )
Abstract
It has been traditionally assumed that medial temporal lobe (MTL) is not required for working memory (WM). However, animal lesion and electrophysiological studies and human neuropsychological and neuroimaging studies have provided increasing evidences of a critical involvement of MTL in WM. Based on previous fi ndings, the central aim of this study was to investigate the contribution of the MTL to verbal WM encoding. Here, we used magnetoencephalogra-phy (MEG) to compare the patterns of MTL activation of 9 epilepsy patients suffering from left hippocampal sclerosis with those of 10 healthy matched controls while they performed a verbal WM task. MEG recordings allow detailed tracking of the time course of MTL activation. We observed impaired WM performance associated with changes in the dynamics of MTL activity in epilepsy patients. Specifi cally, whereas patients showed decreased activity in damaged MTL, activity in the contralateral MTL was enhanced, an effect that became signifi cant in the 600- to 700-ms interval after stimulus presentation. These fi ndings strongly support the crucial contribution of MTL to verbal WM encoding and provide compelling evidence for the proposal that MTL contributes to both episodic memory and WM. Whether this pattern is signaling reorganization or a normal use of a damaged structure is discussed. ( JINS , 2009, 15 , 1– 10 .)
Keywords : Magnetoencephalography , Hippocampus , Memory , Memory disorders , Epilepsy , Epilepsy surgery
AU1
P.C. and F.M. contributed equally to the study. Correspondence and reprint requests to: Pablo Campo, Centro de Mag-
netoencefalografía Dr. Pérez Modrego, Pabellón nº 8 Facultad de Medicina, Universidad Complutense de Madrid, 28040 Madrid, Spain. E-mail: [email protected]

P. Campo et al.2
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
(Cowan, 1995 , 1999 ; Miyake & Shah, 1999 ) have considered an issue of central importance to understanding WM, the pre-cise characterization of how the encoding of task-relevant information takes place in the context of WM tasks. In a pre-vious study (Campo et al., 2005), we used magnetoencepha-lography (MEG), a noninvasive technique with high temporal resolution and good spatial resolution, to investigate the en-coding phase of a verbal task and a spatial WM task. The temporal pattern of neural activity provides valuable infor-mation to understand the cognitive functions supported by different cortical areas (Axmacher et al., 2006 ; Helenius et al., 1998 ). Thus, using MEG, we could specify not only which brain structures were involved but also a detailed time course of their activation. We observed a material-specifi c modulation of the activity of MTL, consisting of a greater activation in left MTL between 500 and 700 ms after stimu-lus presentation during the verbal task and a preponderance of right MTL activation between 400 and 800 ms during the spatial task. These fi ndings, along with others (Karlsgodt et al., 2005 ; Ranganath et al., 2004 ), provide support for the contribution of MTL activation in WM encoding operations (Martin, 1999 ).
The aim of the current study was to further investigate our previous fi ndings suggesting the involvement of left MTL during the encoding phase of verbal WM. As stated by some authors (Grippo et al., 1996 ; Sawrie et al., 2001 ), epilepsy patients suffering from hippocampal sclerosis (HS), consist-ing of neural loss and gliosis restricted to hippocampus and surrounding tissue (Thom et al., 2005 ; Trenerry et al., 1993 ), provide a useful lesional model for studying memory pro-cesses. While patients with HS typically exhibit episodic memory impairment (Jones-Gotman et al., 1997 ; Rausch & Babb, 1993 ; Sass et al., 1995 ), some studies have shown that performance in WM tasks is also reduced (Abrahams et al., 1999 ; Axmacher et al., 2007 ; Grippo et al., 1996 ; Krauss et al., 1997 ; Wagner et al., 2008 ). The nature of this localized lesion allows furthering exploration of the essential contri-bution of MTL activation in WM. Of these mentioned stud-ies, only two reported neurophysiological data comparing verbal WM functioning in epilepsy patients and matched healthy controls. Grippo et al. ( 1996) used event-related po-tentials, a methodology inferior to MEG in terms of precise source localization. A study by Axmacher et al. ( 2007) used intracranial electroencephalogram (iEEG), a methodology with excellent temporal and spatial resolution but a highly invasive neurosurgical procedure.
In this study, we used MEG, a noninvasive procedure, to compare the neuromagnetic patterns of MTL activation evoked by a verbal–semantic WM task in a group of epilepsy patients with left HS, who were evaluated for surgical treat-ment, versus a group of matched healthy controls. The ver-bal WM task was designed to ensure that participants encoded words semantically (i.e., deep encoding) as prior neuroimaging investigations have demonstrated that the depth to which verbal stimuli are processed modulates the activity in the MTL (Kapur et al., 1994 ; Lepage et al., 2000 ). The focus of the study was to determine whether MTL is es-
sentially contributing to verbal WM encoding. If so, re-stricted left MTL damage should modify the dynamics of the activity of its constituent structures during the process of en-coding. Based on previous neuroimaging studies investigating verbal episodic memory in patients with HS (Golby et al., 2002 ; Henke et al., 2003 ; Rabin et al., 2004 ; Richardson et al., 2003 , 2006 ), we hypothesized that left MTL activation would be greater in the control group and that these differ-ences would be time modulated. A reorganization of verbal WM to the undamaged MTL in patients with HS was also expected.
METHODS
Participants
Nine patients (fi ve males) with medically refractory MTL epilepsy were consecutively recruited following evaluation at the “Hospital Ruber Internacional” and participated in the study. They ranged in age from 24 to 43 years ( M = 33.00, SD = 7.08). Diagnosis was established according to clinical, electroencephalogram (EEG), and magnetic resonance imaging (MRI) data. All of them underwent neurological examination, continuous video-EEG monitoring, high-resolution 1.5-T brain MRI, and neuropsychological test-ing ( Table 1 ). Patients were included in the study when clinical data and MRI and EEG fi ndings were suggestive of unilateral mesial temporal lobe epilepsy related to left HS. All patients had seizures with typical temporal lobe semiology that were not controlled with antiepileptic drugs and moderate-to-severe decreased volume, and abnormally increased T2 and FLAIR signal, of the left hippocampus on brain MRI. No lesions were observed in other structures beyond left MTL. Bedside video-EEG monitoring showed interictal epileptiform activity ipsilateral to the side of HS and in fi ve cases complex partial seizures with an ictal onset
AU3
AU4
Table 1. Neuropsychological memory performance of epilepsy patients and controls
Neuropsychological memory performance
Patients Controls
LM 70% LM 75% HVLT 87% LM 87% HVLT 33% LM 90% LM 60% LM 69% LM 61% LM 86% LM 60% LM 73% LM 59% LM 92% LM 58% LM 89% LM 57% LM 84%
LM 82%
Note. Verbal memory was assessed by two free recall tests: the HVLT (max-imum score of 12) and the LM subtest from the Wechsler Memory Scale-Revised (maximum score of 50). The measures represent the percentage of words on immediate recall that are remembered on the delayed free recall trial. HVLT, Hopkins Verbal Learning Test; LM, Logical Memory.

Hippocampal sclerosis and working memory 3
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
in left anterior temporal electrodes. No seizure occurred within 24 hr before the experiment.
The control group comprised 10 healthy volunteers (5 males) ranging in age from 24 to 37 years ( M = 30.9, SD = 4.82). Participants were interviewed and entered in the study if they met the following inclusion criteria: (a) absence of a history of neuropathological conditions or psychopatho-logical diseases and (b) no antecedent of drug or alcohol abuse. There was no signifi cant difference between groups in terms of age ( t = 0.76; p < .05).
Participants were right-handed according to the Edinburgh Handedness Inventory (Oldfi eld et al., 1971), and Spanish was their primary language. We did not make any formal studies for hemispheric language dominance in the patient group. Only handedness was evaluated. However, a recent study (Fontoura et al., 2008 ) found that 92.3% of right-handed epilepsy patients with HS showed left-hemisphere language dominance (see also Springer et al., 1999 ). Loring et al. ( 1999) observed that language transfer to the right hemisphere is more related to extensive lesions in areas be-yond the temporal lobe. Chugani et al. ( 1996) reported that small neocortical lesions are associated with compensation of ipsilateral brain regions, whereas more extensive lesions induce compensatory modifi cations on the contralateral hemisphere. Based on these results, it can be considered that our patients have a left dominance for language, although it has not been systematically explored.
All participants signed a consent form detailing the proce-dures of the study in accordance with the Declaration of Helsinki (1991).
Stimuli and Tasks
We employed the same verbal WM task as in our previous studies (Campo et al., 2005, 2008 ). In each trial, participants fi rst saw a stimulus array comprising four words, located centrally in the display, which was projected for 3000 ms (encoding phase). Participants were informed that, later on, three consecutive probe displays comprising a semantic cat-egory name would be presented and that they would be asked to decide if one of the words in the stimulus display belonged to the semantic category represented by one of the words in the probe displays. Participants had 500 ms to respond to each probe (recognition phase). There was an interval be-tween probe displays of 500–700 ms. A delay interval of 2500 ms during which participants viewed a black screen (maintenance phase) was presented between the offset of the stimulus display and the onset of the probe displays. Con-crete words were used, four to seven letters in length (5.62 ± 1.57) and of moderate frequency (Algarabel, 1996 ). A total of 120 trials were presented.
The stimuli were projected through an LCD video pro-jector (SONY VPL-X600E), situated outside the shielded room, onto a series of in-room mirrors, the last of which was suspended approximately 50 cm above the subject’s face and subtended a visual angle of 1–3° horizontally and 0.5° vertically.
AU5
AU6
Data Collection and Analysis
All MEG recordings were done using a whole-head neuro-magnetometer comprising an array of 148 magnetometers (4-D 2500 ® ; San Diego, CA) housed in a magnetically shielded room. The magnetic fl ux measurements were band-pass fi ltered between 0.1 and 50 Hz and digitized at 678 Hz. MEG data were submitted to an interactive noise reduction procedure to reduce environmental noise (4-D 2500 ® ). Single-trial event-related fi elds (ERFs) in response to 120 stimulus presentations were averaged separately for each sensor after excluding those containing eye movement (as indicated by peak-to-peak amplitudes in the electrooculogram channel in excess of 50 µV) or other myogenic or mechanical artifacts. Finally, the averaged epochs (3000-ms duration) were digi-tally fi ltered with a low-pass 20-Hz fi lter to improve signal quality and adjusted relative to the mean amplitude in the 150-ms prestimulus baseline. A minimum of 80 ERF epochs were collected to calculate each averaged waveform.
The sources of magnetic fi elds were modeled as equiva-lent current dipoles (ECDs). The location, orientation, and amplitude of the best-fi tting single ECD were estimated us-ing a spherical volume conductor model (Sarvas, 1987 ). For a given point in time, the ECD-fi tting algorithm (Levenberg–Marquardt; Moré, 1977 ) was applied to the magnetic fl ux measurements obtained from a group of 34–38 magnetom-eters, always including both magnetic fl ux extremes. The ECD solutions were considered satisfactory after meeting the following criteria: (a) correlation coeffi cient of at least .9 between the observed and the “best” predicted magnetic fi eld distribution, (b) a goodness of fi t of at least 0.9 or higher, and (c) a confi dence volume <1 cm 3 .
Brain response sources, represented as ECDs, were super-imposed on the individual T1-weighted magnetic resonance images (1.5 T, TR 13.6 ms, TE 4.8 ms, recording matrix 256 × 256 pixels, one excitation, 240 mm fi eld of view, and 1.4 mm slice thickness) to delineate the anatomical location. Ana-tomical landmarks (nasion, cz, inion, and the two auricular points corresponding to the external meati) were localized in each individual and used for the coregistration of the MRI and MEG scans. The degree of activation of a particular area was estimated by the total number of successive dipoles that accounted for the ERF components. The reason to elect this procedure for constructing brain activation is twofold: First, because it has demonstrated suffi cient accuracy for lateral-izing neurophysiological activity associated with language function (Breier et al., 2000 ; Maestu et al., 2002 ; Papanico-laou et al., 2004 ) and for distinguishing healthy controls from patients with moderate-to-severe memory impairments (Maestu et al., 2003 , 2004 , 2005 ). Second, its concurrent va-lidity has been successfully tested in comparison with the results of invasive techniques considered as the “gold stan-dards” in the fi eld (Wada Test and electrocortical stimula-tion) in a large patient series (Papanicolaou et al., 2004 ). The rationale for using the number of sequential activity sources as a dependent measure is based on the premise that process-ing of an incoming stimulus requires a transient increase in
AU7
AU8
AU9

P. Campo et al.4
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
neural signaling in one or more neural populations. This in turn produces a time-limited increase in intracellular currents that, once integrated, can be represented as an elec-trical dipole.
While detection of activity generated in deep sources has been a major concern in the MEG literature, several studies have reported activation of MTL during encoding or retrieval phase of memory tasks, showing similar results to those commonly reported by studies using hemodynamic tech-niques, which also constitute a convergent validity (Castillo et al., 2001 ; Hanlon et al., 2003 ; Maestu et al., 2006 ; Papanicolaou et al., 2002 ; Riggs et al., 2009 ; Tendolkar et al., 2000 ; Tesche & Karhu, 2000 ; Ver Hoef et al., 2008 ).
As the accuracy of MEG localization in deep structures is about 1 cm (Tarkiainen et al., 2003 ; see also Gonsalves et al., 2005), we decided to collapse all the sources identifi ed on different MTL structures into one anatomical variable re-ferred as “MTL.” The identifi cation of the cerebral areas where activity sources were observed was determined using a standard MRIcro software atlas of the human brain (Rorden & Brett, 2000 ) as guidance. Here , our interest focused on MTL, including hippocampal gyrus, parahippocampal gyrus, enthorhinal cortex, and subiculum as described by Amaral and Insausti ( 1990) .
For statistical purposes, activity sources observed were grouped into nonoverlapping successive latency windows of 100-ms duration from 200 to 800 ms. We only considered physiological effects occurring in a 200- to 800-ms interval since most previous studies have found effects in this time period (Aine et al., 2005 ; Campo et al., 2005a , 2005b ; Fernandez et al., 1999 , 2002 ; Paller & Wagner, 2002 ; Stare-sina et al., 2005 ).
RESULTS
Behavioral Performance
Performance in the verbal WM task was assessed in terms of corrected hits for each set of stimuli. We observed a mean accuracy level of 76.44% ( SD = 8.29) in the control group and mean accuracy level of 56.23% ( SD = 16.24) in the patient group. Control subjects performed signifi cantly better than patients in terms of accuracy ( t [17] = 3.47; p < .05).
Brain Imaging Data
The number of sequential activity sources in MTL regions was used as a dependent measure. Data were analyzed using a repeated measures analysis of variance (ANOVA) with one between-subject factor, group (patient and control), and two within-subject factors, hemisphere (left and right) and la-tency (200–300, 300–400, 400–500, 500–600, 600–700, and 700–800 ms). Due to the large number of comparisons, re-sults were Bonferroni corrected. Effects were regarded as statistically signifi cant when p remained <.05 after correc-tion. Post hoc analyses were conducted using unpaired t tests.
Group Differences in Spatiotemporal Activation Profi les in MTL Activity
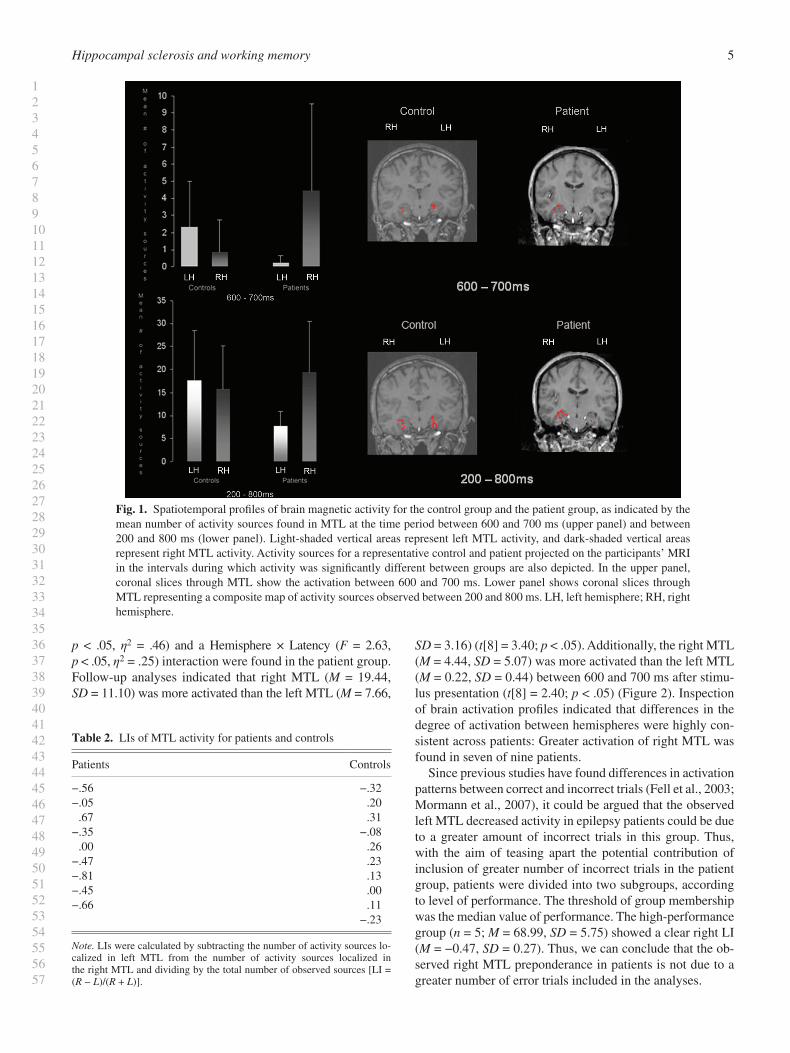
A signifi cant Group × Hemisphere × Latency interaction was explained by a cubic effect ( F = 12.55, p < .05, η 2 = .43). Post hoc comparisons indicated that left MTL showed a greater number of activity sources in the control group ( M = 2.33, SD = 2.66) as compared to the patient group ( M = 0.22, SD = 0.44) between 600 and 700 ms ( t [17] = 2.31; p < .05) ( Figure 1 ). In the same interval, patients showed a higher activation in right MTL ( M = 4.44, SD = 5.07) than controls ( M = 0.80, SD = 1.96), although the results of the post hoc comparisons yielded a marginal effect ( t [17] = 2.11; p = .050). A Group × Hemisphere effect was also found ( F = 4.56, p < .05, η 2 = .21). Further analyses revealed that left MTL showed more activity sources in the control group ( M = 17.70, SD = 10.77) as compared to the patient group ( M = 7.67, SD = 3.16) in the analyzed interval (between 200 and 800 ms) ( t [17] = 2.68; p < .05) ( Figure 1 ).
Laterality Indexes of MTL Activity
Previous language lateralization studies with English- and Spanish-speaking epilepsy patients adopted a cutoff lateral-ity index (LI) value of .25, which was associated with the highest concordance with judgments of hemispheric domi-nance derived from the Wada Test (Maestu et al., 2002 ; Papanicolaou et al., 1999, 2004 ). Additionally, as pointed out by Binder et al. ( 2005) , “the capacity to detect a unilateral abnormality of hippocampal activation is greatly enhanced by computing an asymmetry index, which is feasible only when activation is bilateral.” Here, we apply this criterion, focusing on MTL activity and increasing the interval consid-ered, in our case 200–800 ms. This modifi cation was based on previous iEEG studies of verbal memory where relevant activity in MTL structures in very early latencies was ob-served (between 150 and 300 ms) (Fell et al., 2002 , 2003 ; Fernandez & Tendolkar, 2001 ; Fernandez et al., 1998 ). In the current study, the LI intends to refl ect the relatively greater activation of one MTL as compared to the other. Accord-ingly, we calculated the LI by subtracting the number of ac-tivity sources localized in left MTL from the number of activity sources localized in the right MTL and dividing by the total number of observed sources [LI = ( R − L )/( R + L )] (Ver Hoef et al., 2008 ). According to this criterion, the pro-portions of patients who would be classifi ed as right MTL dominant for memory were six ( M = −0.55, SD = 0.16), two were bilateral ( M = −0.02, SD = 0.03), and one was left MTL dominant (LI = .67). Considering controls, two were classi-fi ed as left MTL dominant ( M = 0.29, SD = 0.04), seven were bilateral ( M = 0.05, SD = 0.16), and one was considered as right MTL dominant (LI = −.32). LI data for each participant are presented in Table 2 .
In order to further explore the asymmetry of activation of MTL, we conducted a two-way ANOVA (Hemisphere × Latency) for each group. No signifi cant effects were found in the control group. A main effect of hemisphere ( F = 6.82,
AU10
AU11
AU12
Hippocampal sclerosis and working memory 5
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
p < .05, η 2 = .46) and a Hemisphere × Latency ( F = 2.63, p < .05, η 2 = .25) interaction were found in the patient group. Follow-up analyses indicated that right MTL ( M = 19.44, SD = 11.10) was more activated than the left MTL ( M = 7.66,
SD = 3.16) ( t [8] = 3.40; p < .05). Additionally, the right MTL ( M = 4.44, SD = 5.07) was more activated than the left MTL ( M = 0.22, SD = 0.44) between 600 and 700 ms after stimu-lus presentation ( t [8] = 2.40; p < .05) ( Figure 2 ). Inspection of brain activation profi les indicated that differences in the degree of activation between hemispheres were highly con-sistent across patients: Greater activation of right MTL was found in seven of nine patients.
Since previous studies have found differences in activation patterns between correct and incorrect trials (Fell et al., 2003 ; Mormann et al., 2007 ), it could be argued that the observed left MTL decreased activity in epilepsy patients could be due to a greater amount of incorrect trials in this group. Thus, with the aim of teasing apart the potential contribution of inclusion of greater number of incorrect trials in the patient group, patients were divided into two subgroups, according to level of performance. The threshold of group membership was the median value of performance. The high-performance group ( n = 5; M = 68.99, SD = 5.75) showed a clear right LI ( M = −0.47, SD = 0.27). Thus, we can conclude that the ob-served right MTL preponderance in patients is not due to a greater number of error trials included in the analyses.
Fig. 1. Spatiotemporal profi les of brain magnetic activity for the control group and the patient group, as indicated by the mean number of activity sources found in MTL at the time period between 600 and 700 ms (upper panel) and between 200 and 800 ms (lower panel). Light-shaded vertical areas represent left MTL activity, and dark-shaded vertical areas represent right MTL activity. Activity sources for a representative control and patient projected on the participants’ MRI in the intervals during which activity was signifi cantly different between groups are also depicted. In the upper panel, coronal slices through MTL show the activation between 600 and 700 ms. Lower panel shows coronal slices through MTL representing a composite map of activity sources observed between 200 and 800 ms. LH, left hemisphere; RH, right hemisphere.
Table 2. LIs of MTL activity for patients and controls
Patients Controls
−.56 −.32 −.05 .20
.67 .31 −.35 −.08
.00 .26 −.47 .23 −.81 .13 −.45 .00 −.66 .11
−.23
Note. LIs were calculated by subtracting the number of activity sources lo-calized in left MTL from the number of activity sources localized in the right MTL and dividing by the total number of observed sources [LI = ( R − L )/( R + L )].
P. Campo et al.6
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
DISCUSSION
In the current study, we investigated the contribution of MTL to encoding processes of verbal WM by comparing MEG profi les of a group of epilepsy patients with temporal lobe epilepsy related to HS with those of a group of healthy con-trols. Our fi ndings corroborate our previous results with healthy controls (Campo et al., 2005) and demonstrate be-havioral verbal WM impairment and changes in the dynam-ics of MTL activity in epilepsy patients. Consistent with previous neuropsychological studies, we observed that pa-tients with left MTL sclerosis demonstrated a reduced per-formance in verbal WM in comparison to healthy controls (Grippo et al., 1996 ; Owen et al., 1996 ; Petrides & Milner, 1982 ; Wagner et al., 2008 ).
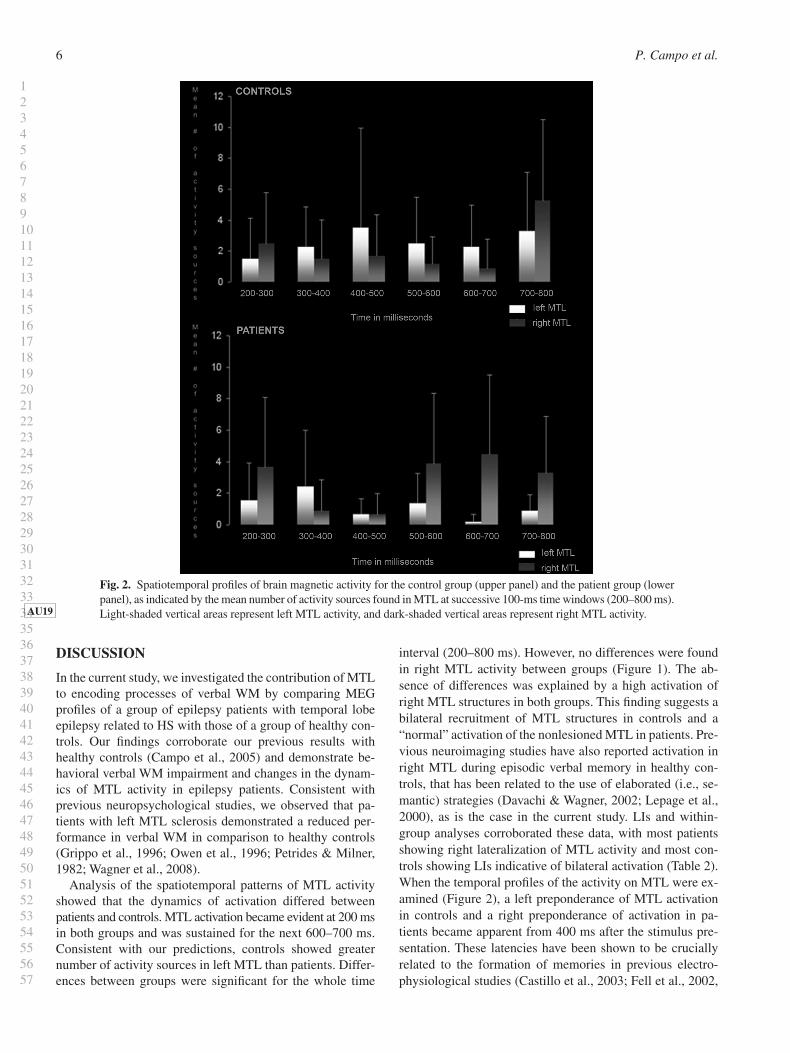
Analysis of the spatiotemporal patterns of MTL activity showed that the dynamics of activation differed between patients and controls. MTL activation became evident at 200 ms in both groups and was sustained for the next 600–700 ms. Consistent with our predictions, controls showed greater number of activity sources in left MTL than patients. Differ-ences between groups were signifi cant for the whole time
interval (200–800 ms). However, no differences were found in right MTL activity between groups ( Figure 1 ). The ab-sence of differences was explained by a high activation of right MTL structures in both groups. This fi nding suggests a bilateral recruitment of MTL structures in controls and a “normal” activation of the nonlesioned MTL in patients. Pre-vious neuroimaging studies have also reported activation in right MTL during episodic verbal memory in healthy con-trols, that has been related to the use of elaborated (i.e., se-mantic) strategies (Davachi & Wagner, 2002 ; Lepage et al., 2000 ), as is the case in the current study. LIs and within-group analyses corroborated these data, with most patients showing right lateralization of MTL activity and most con-trols showing LIs indicative of bilateral activation ( Table 2 ). When the temporal profi les of the activity on MTL were ex-amined ( Figure 2 ), a left preponderance of MTL activation in controls and a right preponderance of activation in pa-tients became apparent from 400 ms after the stimulus pre-sentation. These latencies have been shown to be crucially related to the formation of memories in previous electro-physiological studies (Castillo et al., 2003 ; Fell et al., 2002 ,
Fig. 2. Spatiotemporal profi les of brain magnetic activity for the control group (upper panel) and the patient group (lower panel), as indicated by the mean number of activity sources found in MTL at successive 100-ms time windows (200–800 ms). Light -shaded vertical areas represent left MTL activity, and dark-shaded vertical areas represent right MTL activity. AU19

Hippocampal sclerosis and working memory 7
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
2003 ; Fernandez et al., 2002 ). However, we only found time-modulated signifi cant differences in MTL activation between groups in the 600- to 700-ms interval ( Figure 1 ).
Specifi cally, controls exhibited greater activation than patients in left MTL, while patients showed an opposite pat-tern, greater right MTL activation than controls. Decreases in MTL activity ipsilateral to lesion and greater activation of the contralateral MTL have been previously reported in epilepsy patients with HS for episodic memory, either for verbal or for nonverbal material, with functional magnetic resonance imaging (Dupont et al., 2000 ; Golby et al., 2002 ; Powell et al., 2007 ; Richardson et al., 2003 ) and iEEG/MEG (Mormann et al., 2007 ; Ver Hoef et al., 2008 ). This fi nding has been interpreted as a reorganization of memory function, although not suffi cient to enable normal memory perfor-mance (Dupont et al., 2000 ; Powell et al., 2007 ). The double dissociation we observe between 600 and 700 ms after stim-ulus presentation is consistent with this explanation. As stated by Ver Hoef et al. ( 2008) , the identical temporal char-acter of left and right MTL activity supports its similar func-tion. However, the fact that patients exhibited a decreased activity in the lesioned MTL and a similar activation in the nonlesioned MTL to that of controls in the interval between 200 and 800 ms, along with a worse performance, could be indicating that patients with left HS, rather than showing a reallocation, are using a residual component of a damaged memory structure subserving verbal WM. Further studies should be conducted to explore this alternative explanation. Nonetheless, current results suggest that MTL activation is contributing to verbal WM and that this activation is not sig-naling a parallel process not specifi cally related to this cog-nitive function (Wagner et al., 2008 ).
Three concerns should be considered related to current results. First, as previous studies have found differences in activation patterns between correct and incorrect trials (Fell et al., 2003 ; Mormann et al., 2007 ), it could be argued that the decrease of activation on the pathological MTL could be due to a greater amount of incorrect trials made by epilepsy patients. To address this possible caveat, we demonstrate that a subgroup of patients with better performance (i.e., greater amount of correct trials) showed a clear right MTL preponderance of activation (Mormann et al., 2007 ). Thus, the signifi cant decrease in left MTL activity is due to under-lying pathology. Second, MTL fi ndings could be refl ecting retrieval from semantic memory rather than specifi c WM ac-tivity. However, several lines of evidence suggest that while patients with damage to hippocampal structures sometimes exhibit impairments in semantic abilities, it appears that this defi cit is less apparent in epilepsy patients than in patients with MTL damage resulting from other etiologies (Giovagnoli et al., 2005 ; Ribbler & Rausch, 1990 ). Additionally, neu-roimaging and neuropsychological studies suggest that semantic processes rely more on lateral aspects of the tem-poral lobe (Levy et al., 2004 ). Therefore, we suggest that MTL activation observed in this study is not signaling a se-mantic retrieval process but an encoding mechanism. Finally, some authors have claimed that MTL is recruited in WM
only during processing of novel or complex stimuli (Ranganath & Blumenfeld, 2005 ). This notion has led to an overwhelm-ing number of studies investigating the role of MTL in WM using novel/complex visual stimuli (Della-Maggiore et al., 2000; Haxby et al., 1995 ; Piekema et al., 2006 ; Ranganath & D’Esposito, 2001 ). However, neuropsychological studies (Owen et al., 1996 ; Petrides & Milner, 1982 ; Wagner et al., 2008 ) and neuroimaging experiments of WM in young controls (Cabeza et al., 2002 ; Campo et al., 2005; Karlsgodt et al., 2005 ; Petersson et al., 2006 ), older controls (Cabeza et al., 2004 ), and patient groups (Grippo et al., 1996 ) demon-strate an additional involvement of MTL in WM for familiar verbal stimuli. The present study provides neurophysiologi-cal support for these fi ndings, and highlights the potential application of MEG in exploring material-specifi c memory effects for WM in epilepsy patients (Wagner et al., 2008 ), similar to that reported for episodic memory (Golby et al., 2002 ; Jones-Gotman et al., 1997 ; Powell et al., 2007 ).
Although it was not the main focus of our study, our results suggest the potential clinical utility of the current paradigm for the presurgical evaluation protocol assessing the suitabil-ity of epilepsy patients for surgery. However, larger samples should be used before this can be considered, and taking into account the material-specifi c lateralization of memory pro-cessing (Golby et al., 2002 ; Powell et al., 2004 ; Wagner et al., 2008 ), nonverbal tasks should also be included.
In summary, the present study provides, for the fi rst time, direct evidence for the alteration of the dynamics of MTL activity in epilepsy patients suffering from left HS in a verbal WM task using MEG. The observed changes in the timing and laterality of MTL activation, as well as a reduced behav-ioral performance, in patients with a restricted left MTL damage strongly support the crucial contribution of these structures to verbal WM encoding. Along with previous neu-ropsychological and neuroimaging studies, current results provide compelling evidence for the proposal that MTL con-tributes to both episodic memory and WM encoding.
ACKNOWLEDGMENTS
The manuscript is new and original, is not currently under review by any other publication, and has not been previously published either electronically or in print. The project did not receive any fi nancial sup-port, and there are no confl icts of interest affecting this manuscript.
REFERENCES
Abrahams , S. , Morris , R.G. , Polkey , C.E. , Jarosz , J.M. , Cox , T.C. , Graves , M. , & Pickering , A. ( 1999 ). Hippocampal involvement in spatial and working memory: A structural MRI analysis of patients with unilateral mesial temporal lobe sclerosis . Brain and Cognition , 41 ( 1 ), 39 – 65 .
Aine , C.J. , Adair , J.C. , Knoefel , J.E. , Hudson , D. , Qualls , C. , Kova-cevic , S. , Woodruff , C.C. , Cobb , W. , Padilla , D. , Lee , R.R. , & Stephen , J.M. ( 2005 ). Temporal dynamics of age-related differences in auditory incidental verbal learning . Brain Research. Cognitive Brain Research , 24 ( 1 ), 1 – 18 .
Algarabel , S. ( 1996 ). Indices de interés psicolinguístico de 1917 palabras castellanas . Cognitiva , 8 , 43 – 88 .
AU13

P. Campo et al.8
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Amaral , D. & Insausti , R. ( 1990 ). Hippocampal formation . In G. Paxinos (Ed.), The human nervous system (pp. 711 – 755 ). San Diego, CA : Academic Press .
Axmacher , N. , Mormann , F. , Fernandez , G. , Cohen , M.X. , Elger , C.E. , & Fell , J. ( 2007 ). Sustained neural activity patterns during working memory in the human medial temporal lobe . The Jour-nal of Neuroscience , 27 ( 29 ), 7807 – 7816 .
Axmacher , N. , Mormann , F. , Fernandez , G. , Elger , C.E. , & Fell , J. ( 2006 ). Memory formation by neuronal synchronization . Brain Research Review , 52 ( 1 ), 170 – 182 .
Axmacher , N. , Schmitz , D.P. , Wagner , T. , Elger , C.E. , & Fell , J. ( 2008 ). Interactions between medial temporal lobe, prefrontal cor-tex, and inferior temporal regions during visual working memory: A combined intracranial EEG and functional magnetic resonance imaging study . The Journal of Neuroscience , 28 ( 29 ), 7304 – 7312 .
Binder , J.R. , Bellgowan , P.S. , Hammeke , T.A. , Possing , E.T. , & Frost , J.A. ( 2005 ). A comparison of two FMRI protocols for elic-iting hippocampal activation . Epilepsia , 46 ( 7 ), 1061 – 1070 .
Breier , J.I. , Simos , P.G. , Zouridakis , G. , & Papanicolaou , A.C. ( 2000 ). Lateralization of activity associated with language func-tion using magnetoencephalography: A reliability study . Journal of Clinical Neurophysiology , 17 ( 5 ), 503 – 510 .
Cabeza , R. , Daselaar , S.M. , Dolcos , F. , Prince , S.E. , Budde , M. , & Nyberg , L. ( 2004 ). Task-independent and task-specifi c age ef-fects on brain activity during working memory, visual attention and episodic retrieval . Cerebral Cortex , 14 ( 4 ), 364 – 375 .
Cabeza , R. , Dolcos , F. , Graham , R. , & Nyberg , L. ( 2002 ). Similari-ties and differences in the neural correlates of episodic memory retrieval and working memory . NeuroImage , 16 ( 2 ), 317 – 330 .
Campo , P. , Maestu , F. , Capilla , A. , Fernandez , S. , Fernandez , A. , & Ortiz , T. ( 2005 a). Activity in human medial temporal lobe asso-ciated with encoding process in spatial working memory revealed by magnetoencephalography . The European Journal of Neuro-science , 21 ( 6 ), 1741 – 1748 .
Campo , P. , Maestu , F. , Capilla , A. , Morales , M. , Fernandez , S. , del Rio , D. , & Ortiz , T. ( 2008 ). Temporal dynamics of parietal activity during word-location binding . Neuropsychology , 22 ( 1 ), 85 – 99 .
Campo , P. , Maestu , F. , Ortiz , T. , Capilla , A. , Fernandez , S. , & Fernandez , A. ( 2005 b). Is medial temporal lobe activation specifi c for encoding long-term memories? NeuroImage , 25 ( 1 ), 34 – 42 .
Castillo , E.M. , Simos , P.G. , Davis , R.N. , Breier , J. , Fitzgerald , M.E. , & Papanicolaou , A. ( 2003 ). Temporal dynamics of medial temporal lobe activation during encoding and recognition. A magnetoencephalography study . Revista Española de Neurop-sicología , 5 ( 2 ), 99 – 120 .
Castillo , E.M. , Simos , P.G. , Davis , R.N. , Breier , J. , Fitzgerald , M.E. , & Papanicolaou , A.C. ( 2001 ). Levels of word processing and incidental memory: Dissociable mechanisms in the temporal lobe . Neuroreport , 12 ( 16 ), 3561 – 3566 .
Cave , C.B. & Squire , L.R. ( 1992 ). Intact verbal and nonverbal short-term memory following damage to the human hippocam-pus . Hippocampus , 2 ( 2 ), 151 – 163 .
Chugani , H.T. , Muller , R.A. , & Chugani , D.C. ( 1996 ). Functional brain reorganization in children . Brain & Development , 18 ( 5 ), 347 – 356 .
Cowan , N. ( 1995 ). Attention and memory: An integrated frame-work . Oxford, UK : Oxford University Press .
Cowan , N. ( 1999 ). An embedded-process model of working mem-ory . In P. Shah (Ed.), Models of working memory . Cambridge, UK : Cambridge University Press .
Davachi , L. & Goldman-Rakic , P.S. ( 2001 ). Primate rhinal cortex participates in both visual recognition and working memory
tasks: Functional mapping with 2-DG . Journal of Neurophysiol-ogy , 85 ( 6 ), 2590 – 2601 .
Davachi , L. & Wagner , A.D. ( 2002 ). Hippocampal contributions to episodic encoding: Insights from relational and item-based learn-ing . Journal of Neurophysiology , 88 ( 2 ), 982 – 990 .
Dupont , S. , Van de Moortele , P.F. , Samson , S. , Hasboun , D. , Poline , J.B. , Adam , C. , Lehericy , S. , Le Bihan , D. , Samson , Y. , & Baulac , M. ( 2000 ). Episodic memory in left temporal lobe epilepsy: A functional MRI study . Brain , 123 ( Pt 8 ), 1722 – 1732 .
Eichenbaum , H. ( 1997 ). Declarative memory: Insights from cogni-tive neurobiology . Annual Review of Psychology , 48 , 547 – 572 .
Fell , J. , Klaver , P. , Elfadil , H. , Schaller , C. , Elger , C.E. , & Fernandez , G. ( 2003 ). Rhinal-hippocampal theta coherence during declarative memory formation: Interaction with gamma synchronization? The European Journal of Neuroscience , 17 ( 5 ), 1082 – 1088 .
Fell , J. , Klaver , P. , Elger , C.E. , & Fernandez , G. ( 2002 ). The interac-tion of rhinal cortex and hippocampus in human declarative mem-ory formation . Review in the Neurosciences , 13 ( 4 ), 299 – 312 .
Fernandez , G. , Effern , A. , Grunwald , T. , Pezer , N. , Lehnertz , K. , Dumpelmann , M. , Van Roost , D. , & Elger , C.E. ( 1999 ). Real-time tracking of memory formation in the human rhinal cortex and hippocampus . Science , 285 ( 5433 ), 1582 – 1585 .
Fernandez , G. , Klaver , P. , Fell , J. , Grunwald , T. , & Elger , C.E. ( 2002 ). Human declarative memory formation: Segregating rhinal and hippocampal contributions . Hippocampus , 12 ( 4 ), 514 – 519 .
Fernandez , G. & Tendolkar , I. ( 2001 ). Integrated brain activity in medial temporal and prefrontal areas predicts subsequent mem-ory performance: Human declarative memory formation at the system level . Brain Research Bulletin , 55 ( 1 ), 1 – 9 .
Fernandez , G. , Weyerts , H. , Schrader-Bolsche , M. , Tendolkar , I. , Smid , H.G. , Tempelmann , C. , Hinrichs , H. , Scheich , H. , Elger , C.E. , Mangun , G.R. , & Heinze , H.J. ( 1998 ). Successful verbal encoding into episodic memory engages the posterior hippocam-pus: A parametrically analyzed functional magnetic resonance imaging study . The Journal of Neuroscience , 18 ( 5 ), 1841 – 1847 .
Fontoura , D.R. , Branco Dde , M. , Anes , M. , Costa , J.C. , & Portuguez , M.W. ( 2008 ). Language brain dominance in patients with refractory temporal lobe epilepsy: A comparative study between functional magnetic resonance imaging and dichotic listening test . Arquivos de Neuro-psiquiatria , 66 ( 1 ), 34 – 39 .
Gabrieli , J.D. ( 1998 ). Cognitive neuroscience of human memory . Annual Review of Psychology , 49 , 87 – 115 .
Gazzaley , A. , Rissman , J. , & Desposito , M. ( 2004 ). Functional con-nectivity during working memory maintenance . Cognitive, Af-fective & Behavioral Neuroscience , 4 ( 4 ), 580 – 599 .
Giovagnoli , A.R. , Erbetta , A. , Villani , F. , & Avanzini , G. ( 2005 ). Semantic memory in partial epilepsy: Verbal and non-verbal defi cits and neuroanatomical relationships . Neuropsychologia , 43 ( 10 ), 1482 – 1492 .
Glabus , M.F. , Horwitz , B. , Holt , J.L. , Kohn , P.D. , Gerton , B.K. , Callicott , J.H. , Meyer-Lindenberg , A. , & Berman , K.F. ( 2003 ). Interindividual differences in functional interactions among pre-frontal, parietal and parahippocampal regions during working memory . Cerebral Cortex , 13 ( 12 ), 1352 – 1361 .
Golby , A.J. , Poldrack , R.A. , Illes , J. , Chen , D. , Desmond , J.E. , & Gabrieli , J.D. ( 2002 ). Memory lateralization in medial temporal lobe epilepsy assessed by functional MRI . Epilepsia , 43 ( 8 ), 855 – 863 .
Grippo , A. , Pelosi , L. , Mehta , V. , & Blumhardt , L.D. ( 1996 ). Working memory in temporal lobe epilepsy: An event-related potential study . Electroencephalography and Clinical Neuro-physiology , 99 ( 3 ), 200 – 213 .
AU14
AU15

Hippocampal sclerosis and working memory 9
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
Haettig , H. , Schepler , P. , Lehmann , T.N. , & Meencke , H.J. ( 2004 ). Short-term memory outcome after temporal lobe surgery for epi-lepsy: A 12 month follow up . Paper presented at the Human Brain Mapping, Budapest .
Hanlon , F.M. , Weisend , M.P. , Huang , M. , Lee , R.R. , Moses , S.N. , Paulson , K.M. , Thoma , R.J. , Miller , G.A. , & Canive , J.M. ( 2003 ). A non-invasive method for observing hippocampal function . Neuroreport , 14 ( 15 ), 1957 – 1960 .
Haxby , J.V. , Ungerleider , L.G. , Horwitz , B. , Rapoport , S.I. , & Grady , C.L. ( 1995 ). Hemispheric differences in neural systems for face working memory: A PET-rCBF study . Human Brain Mapping , 3 , 68 – 82 .
Helenius , P. , Salmelin , R. , Service , E. , & Connolly , J.F. ( 1998 ). Distinct time courses of word and context comprehension in the left temporal cortex . Brain , 121 ( Pt 6 ), 1133 – 1142 .
Henke , K. , Treyer , V. , Weber , B. , Nitsch , R.M. , Hock , C. , Wieser , H.G. , & Buck , A. ( 2003 ). Functional neuroimaging predicts indi-vidual memory outcome after amygdalohippocampectomy . Neu-roreport , 14 ( 9 ), 1197 – 1202 .
Holscher , C. & Rolls , E.T. ( 2002 ). Perirhinal cortex neuronal activ-ity is actively related to working memory in the macaque . Neural Plasticity , 9 ( 1 ), 41 – 51 .
Jones-Gotman , M. , Zatorre , R.J. , Olivier , A. , Andermann , F. , Cendes , F. , Staunton , H. , McMackin , D. , Siegel , A.M. , & Wieser , H.G. ( 1997 ). Learning and retention of words and designs fol-lowing excision from medial or lateral temporal-lobe structures . Neuropsychologia , 35 ( 7 ), 963 – 973 .
Kapur , S. , Craik , F.I. , Tulving , E. , Wilson , A.A. , Houle , S. , & Brown , G.M. ( 1994 ). Neuroanatomical correlates of encoding in episodic memory: Levels of processing effect . Proceedings of the National Academy of Sciences of the United States of Amer-ica , 91 ( 6 ), 2008 – 2011 .
Karlsgodt , K.H. , Shirinyan , D. , van Erp , T.G. , Cohen , M.S. , & Cannon , T.D. ( 2005 ). Hippocampal activations during encoding and retrieval in a verbal working memory paradigm . NeuroImage , 25 ( 4 ), 1224 – 1231 .
Krauss , G.L. , Summerfi eld , M. , Brandt , J. , Breiter , S. , & Ruchkin , D. ( 1997 ). Mesial temporal spikes interfere with working memory . Neurology , 49 ( 4 ), 975 – 980 .
Lancelot , C. , Samson , S. , Ahad , P. , & Baulac , M. ( 2003 ). Effect of unilateral temporal lobe resection on short-term memory for au-ditory object and sound location . Annals of the New York Acad-emy of Sciences , 999 , 377 – 380 .
Lee , A.C. , Buckley , M.J. , Gaffan , D. , Emery , T. , Hodges , J.R. , & Graham , K.S. ( 2006 ). Differentiating the roles of the hippocam-pus and perirhinal cortex in processes beyond long-term declara-tive memory: A double dissociation in dementia . The Journal of Neuroscience , 26 ( 19 ), 5198 – 5203 .
Lepage , M. , Habib , R. , Cormier , H. , Houle , S. , & McIntosh , A.R. ( 2000 ). Neural correlates of semantic associative encoding in episodic memory . Brain Research. Cognitive Brain Research , 9 ( 3 ), 271 – 280 .
Levy , D.A. , Bayley , P.J. , & Squire , L.R. ( 2004 ). The anatomy of semantic knowledge: Medial vs. lateral temporal lobe . Proceed-ings of the National Academy of Sciences of the United States of America , 101 ( 17 ), 6710 – 6715 .
Loring , D.W. , Strauss , E. , Hermann , B.P. , Perrine , K. , Trenerry , M.R. , Barr , W.B. , Westerveld , M. , Chelune , G.J. , Lee , G.P. , & Meador , K.J. ( 1999 ). Effects of anomalous language representa-tion on neuropsychological performance in temporal lobe epi-lepsy . Neurology , 53 ( 2 ), 260 – 264 .
Maestu , F. , Arrazola , J. , Fernandez , A. , Simos , P.G. , Amo , C. , Gil-Gregorio , P. , Fernandez , S. , Papanicolaou , A. , & Ortiz , T. ( 2003 ). Do cognitive patterns of brain magnetic activity correlate with hippocampal atrophy in Alzheimer’s disease? Journal of Neurol-ogy, Neurosurgery, and Psychiatry , 74 ( 2 ), 208 – 212 .
Maestu , F. , Campo , P. , Gil-Gregorio , P. , Fernandez , S. , Fernandez , A. , & Ortiz , T. ( 2006 ). Medial temporal lobe neuromagnetic hypo-activation and risk for developing cognitive decline in elderly population: A 2-year follow-up study . Neurobiology of Aging , 27 ( 1 ), 32 – 37 .
Maestu , F. , Fernandez , A. , Simos , P.G. , Lopez-Ibor , M.I. , Campo , P. , Criado , J. , Rodriguez-Palancas , A. , Ferre , F. , Amo , C. , & Ortiz , T. ( 2004 ). Profi les of brain magnetic activity during a memory task in patients with Alzheimer’s disease and in non-demented elderly subjects, with or without depression . Journal of Neurology, Neu-rosurgery, and Psychiatry , 75 ( 8 ), 1160 – 1162 .
Maestu , F. , Garcia-Segura , J. , Ortiz , T. , Montoya , J. , Fernandez , A. , Gil-Gregorio , P. , Campo , P. , Fernandez , S. , Viano , J. , & Portera , A. ( 2005 ). Evidence of biochemical and biomagnetic interactions in Alzheimer’s disease: An MEG and MR spectroscopy study . Dementia and Geriatric Cognitive Disorders , 20 ( 2–3 ), 145 – 152 .
Maestu , F. , Ortiz , T. , Fernandez , A. , Amo , C. , Martin , P. , Fernan-dez , S. , & Sola , R.G. ( 2002 ). Spanish language mapping using MEG: A validation study . NeuroImage , 17 ( 3 ), 1579 – 1586 .
Manoach , D.S. , Greve , D.N. , Lindgren , K.A. , & Dale , A.M. ( 2003 ). Identifying regional activity associated with temporally sepa-rated components of working memory using event-related func-tional MRI . NeuroImage , 20 ( 3 ), 1670 – 1684 .
Martin , A. ( 1999 ). Automatic activation of the medial temporal lobe during encoding: Lateralized infl uences of meaning and novelty . Hippocampus , 9 ( 1 ), 62 – 70 .
Miyake , A. & Shah , P. ( 1999 ). Models of working memory . Cam-bridge, UK : Cambridge University Press .
Monk , C.S. , Zhuang , J. , Curtis , W.J. , Ofenloch , I.T. , Tottenham , N. , Nelson , C.A. , & Hu , X. ( 2002 ). Human hippocampal activation in the delayed matching- and nonmatching-to-sample memory tasks: An event-related functional MRI approach . Behavioral Neuroscience , 116 ( 4 ), 716 – 721 .
Moré , J. ( 1977 ). The Levenberg-Marquardt algorithm: Implemen-tation and theory . In G. Watson (Ed.), Numerical analysis (pp. 105 – 116 ). Springer Verlag .
Mormann , F. , Fernandez , G. , Klaver , P. , Weber , B. , Elger , C.E. , & Fell , J. ( 2007 ). Declarative memory formation in hippocampal sclerosis: An intracranial event-related potentials study . Neurore-port , 18 ( 4 ), 317 – 321 .
Nichols , E.A. , Kao , Y.C. , Verfaellie , M. , & Gabrieli , J.D. ( 2006 ). Working memory and long-term memory for faces: Evidence from fMRI and global amnesia for involvement of the medial temporal lobes . Hippocampus , 16 ( 7 ), 604 – 616 .
Olson , I.R. , Moore , K.S. , Stark , M. , & Chatterjee , A. ( 2006 ). Visual working memory is impaired when the medial temporal lobe is damaged . Journal of Cognitive Neuroscience , 18 ( 7 ), 1087 – 1097 .
Owen , A.M. , Morris , R.G. , Sahakian , B.J. , Polkey , C.E. , & Robbins , T.W. ( 1996 ). Double dissociations of memory and executive functions in working memory tasks following frontal lobe exci-sions, temporal lobe excisions or amygdalo-hippocampectomy in man . Brain , 119 ( Pt 5 ), 1597 – 1615 .
Paller , K.A. & Wagner , A.D. ( 2002 ). Observing the transformation of experience into memory . Trends in Neurosciences , 6 , 93 – 102 .
Papanicolaou , A.C. , Simos , P.G. , Castillo , E.M. , Breier , J.I. , Katz , J.S. , & Wright , A.A. ( 2002 ). The hippocampus and memory of
AU16
AU17

P. Campo et al.10
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657
verbal and pictorial material . Learn & Memory (Cold Spring Harbor, N.Y.) , 9 ( 3 ), 99 – 104 .
Papanicolaou , A.C. , Simos , P.G. , Castillo , E.M. , Breier , J.I. , Sarkari , S. , Pataraia , E. , Billingsley , R.L. , Buchanan , S. , Wheless , J. , Maggio , V. , & Maggio , W.W. ( 2004 ). Magneto-cephalography: A noninvasive alternative to the Wada procedure . Journal of Neurosurgery , 100 ( 5 ), 867 – 876 .
Petersson , K.M. , Gisselgard , J. , Gretzer , M. , & Ingvar , M. ( 2006 ). Interaction between a verbal working memory network and the medial temporal lobe . NeuroImage , 33 ( 4 ), 1207 – 1217 .
Petrides , M. & Milner , B. ( 1982 ). Defi cits on subject-ordered tasks after frontal- and temporal-lobe lesions in man . Neuropsycholo-gia , 20 ( 3 ), 249 – 262 .
Piekema , C. , Kessels , R.P. , Mars , R.B. , Petersson , K.M. , & Fernandez , G. ( 2006 ). The right hippocampus participates in short-term memory maintenance of object-location associations . NeuroImage , 33 ( 1 ), 374 – 382 .
Powell , H.W. , Koepp , M.J. , Richardson , M.P. , Symms , M.R. , Thompson , P.J. , & Duncan , J.S. ( 2004 ). The application of func-tional MRI of memory in temporal lobe epilepsy: A clinical re-view . Epilepsia , 45 ( 7 ), 855 – 863 .
Powell , H.W. , Richardson , M.P. , Symms , M.R. , Boulby , P.A. , Thomp-son , P.J. , Duncan , J.S. , & Koepp , M.J. ( 2007 ). Reorganization of verbal and nonverbal memory in temporal lobe epilepsy due to uni-lateral hippocampal sclerosis . Epilepsia , 48 ( 8 ), 1512 – 1525 .
Rabin , M.L. , Narayan , V.M. , Kimberg , D.Y. , Casasanto , D.J. , Glosser , G. , Tracy , J.I. , French , J.A. , Sperling , M.R. , & Detre , J.A. ( 2004 ). Functional MRI predicts post-surgical memory following temporal lobectomy . Brain , 127 ( Pt 10 ), 2286 – 2298 .
Ranganath , C. & Blumenfeld , R.S. ( 2005 ). Doubts about double dissociations between short- and long-term memory . Trends in Cognitive Sciences , 9 ( 8 ), 374 – 380 .
Ranganath , C. , DeGutis , J. , & D’Esposito , M. ( 2004 ). Category-specifi c modulation of inferior temporal activity during working memory encoding and maintenance . Cognitive Brain Research , 20 , 37 – 45 .
Ranganath , C. & D’Esposito , M. ( 2001 ). Medial temporal lobe ac-tivity associated with active maintenance of novel information . Neuron , 31 ( 5 ), 865 – 873 .
Rausch , R. & Babb , T.L. ( 1993 ). Hippocampal neuron loss and memory scores before and after temporal lobe surgery for epi-lepsy . Archives of Neurology , 50 ( 8 ), 812 – 817 .
Ribbler , A. & Rausch , R. ( 1990 ). Performance of patients with uni-lateral temporal lobectomy on selective reminding procedures using either related or unrelated words . Cortex , 26 ( 4 ), 575 – 584 .
Richardson , M.P. , Strange , B.A. , Duncan , J.S. , & Dolan , R.J. ( 2003 ). Preserved verbal memory function in left medial tempo-ral pathology involves reorganisation of function to right medial temporal lobe . NeuroImage , 20 (Suppl 1), S112 – 119 .
Richardson , M.P. , Strange , B.A. , Duncan , J.S. , & Dolan , R.J . ( 2006 ). Memory fMRI in left hippocampal sclerosis: Optimizing the approach to predicting postsurgical memory . Neurology , 66 ( 5 ), 699 – 705 .
Riggs , L. , Moses , S.N. , Bardouille , T. , Herdman , A.T. , Ross , B. , & Ryan , J.D. ( 2009 ). A complementary analytic approach to exam-ining medial temporal lobe sources using magnetoencephalogra-phy . NeuroImage , 45 ( 2 ), 627 – 642 .
Rorden , C. & Brett , M. ( 2000 ). Stereotaxic display of brain lesions . Behavioral neurology , 12 , 191 – 200 .
Sarvas , J. ( 1987 ). Basic mathematical and electromagnetic con-cepts of the biomagnetic inverse problem . Physics in Medicine and Biology , 32 ( 1 ), 11 – 22 .
Sass , K.J. , Buchanan , C.P. , Kraemer , S. , Westerveld , M. , Kim , J.H. , & Spencer , D.D. ( 1995 ). Verbal memory impairment resulting from hippocampal neuron loss among epileptic patients with structural lesions . Neurology , 45 ( 12 ), 2154 – 2158 .
Sawrie , S.M. , Martin , R.C. , Knowlton , R. , Faught , E. , Gilliam , F. , & Kuzniecky , R. ( 2001 ). Relationships among hippocampal volum-etry, proton magnetic resonance spectroscopy, and verbal memory in temporal lobe epilepsy . Epilepsia , 42 ( 11 ), 1403 – 1407 .
Schon , K. , Hasselmo , M.E. , Lopresti , M.L. , Tricarico , M.D. , & Stern , C.E. ( 2004 ). Persistence of parahippocampal representa-tion in the absence of stimulus input enhances long-term encod-ing: A functional magnetic resonance imaging study of sub sequent memory after a delayed match-to-sample task . The Journal of Neuroscience , 24 ( 49 ), 11088 – 11097 .
Springer , J.A. , Binder , J.R. , Hammeke , T.A. , Swanson , S.J. , Frost , J.A. , Bellgowan , P.S. , Brewer , C.C. , Perry , H.M. , Morris , G.L. , & Mueller , W.M. ( 1999 ). Language dominance in neurologically normal and epilepsy subjects: A functional MRI study . Brain , 122 ( Pt 11 ), 2033 – 2046 .
Squire , L.R. , Stark , C.E. , & Clark , R.E. ( 2004 ). The medial tempo-ral lobe . Annual Review of Neuroscience , 27 , 279 – 306 .
Staresina , B.P. , Bauer , H. , Deecke , L. , & Walla , P. ( 2005 ). Neurocog-nitive correlates of incidental verbal memory encoding: A magne-toencephalographic (MEG) study . NeuroImage , 25 ( 2 ), 430 – 443 .
Stern , C.E. , Sherman , S.J. , Kirchhoff , B.A. , & Hasselmo , M.E. ( 2001 ). Medial temporal and prefrontal contributions to working memory tasks with novel and familiar stimuli . Hippocampus , 11 ( 4 ), 337 – 346 .
Suzuki , W.A. , Miller , E.K. , & Desimone , R. ( 1997 ). Object and place memory in the macaque entorhinal cortex . Journal of Neu-rophysiology , 78 ( 2 ), 1062 – 1081 .
Tarkiainen , A. , Liljestrom , M. , Seppa , M. , & Salmelin , R. ( 2003 ). The 3D topography of MEG source localization accuracy: Ef-fects of conductor model and noise . Clinical Neurophysiology , 114 ( 10 ), 1977 – 1992 .
Tendolkar , I. , Rugg , M. , Fell , J. , Vogt , H. , Scholz , M. , Hinrichs , H. , & Heinze , H.J. ( 2000 ). A magnetoencephalographic study of brain activity related to recognition memory in healthy young human subjects . Neuroscience Letters , 280 ( 1 ), 69 – 72 .
Tesche , C.D. & Karhu , J. ( 2000 ). Theta oscillations index human hippocampal activation during a working memory task . Proceed-ings of the National Academy of Sciences of the United States of America , 97 ( 2 ), 919 – 924 .
Thom , M. , Zhou , J. , Martinian , L. , & Sisodiya , S. ( 2005 ). Quantita-tive post-mortem study of the hippocampus in chronic epilepsy: Seizures do not inevitably cause neuronal loss . Brain , 128 ( Pt 6 ), 1344 – 1357 .
Trenerry , M.R. , Jack , C.R. , Jr ., Ivnik , R.J. ,, Sharbrough , F.W. , Cascino , G.D. , Hirschorn , K.A. , Marsh , W.R. , Kelly , P.J. , & Meyer , F.B. ( 1993 ). MRI hippocampal volumes and memory function before and after temporal lobectomy . Neurology , 43 ( 9 ), 1800 – 1805 .
Ver Hoef , L.W. , Sawrie , S. , Killen , J. , & Knowlton , R.C. ( 2008 ). Left mesial temporal sclerosis and verbal memory: A magneto-encephalography study . Journal of Clinical Neurophysiology , 25 ( 1 ), 1 – 6 .
Wagner , D.D. , Sziklas , V. , Garver , K.E. , & Jones-Gotman , M. ( 2008 ). Material-specifi c lateralization of working memory in the medial temporal lobe . Neuropsychologia .
Young , B.J. , Otto , T. , Fox , G.D. , & Eichenbaum , H. ( 1997 ). Mem-ory representation within the parahippocampal region . The Jour-nal of Neuroscience , 17 ( 13 ), 5183 – 5195 .
AU18

AUTHOR QUERIES
1 Please clarify if the representation of “Rúber” in the fourth affi liation line can be changed to “Ruber” as in the sentence “Nine patients (fi ve males) with …”
2 Please note that the section heading “Introduction” has been introduced as per style. Please check if this is correct.
3 Please clarify in all the reference citations of “Campo et al. (2005)” whether the year is “2005a or 2005b” throughout the text.
4 Please clarify whether “mesial temporal lobe” in the sentence “Patients were included in the study …” can be changed to “medial temporal lobe” and hence can be abbreviated as “MTL.”
5 Please note that “Oldfi eld et al. (1971)” is not listed. Please add to the list or delete the citation.
6 Please provide the complete supplier name and location for “SONY VPL-X600E” in the sentence “The stimuli were projected …”
7 Please provide the complete supplier name in the sentence “All MEG recordings …”
8 Please spell out “TR” and “TE” in the sentence “Brain response sources, represented …”
9 Please clarify if the spelling of “meati” in the sentence “Anatomical landmarks …” can be changed to “meatus.”
10 Please note that “Gonsalves et al. (2005)” is not listed. Please add to the list or delete the citation.
11 Please clarify if the spelling of “enthorhinal” in the sentence “Here, our interest …” can be changed to “entorhinal.”
12 Please note that “Papanicolaou et al. (1999)” is not listed. Please add to the list or delete the citation.
13 Please note that “Della-Maggiore et al. (2000)” is not listed. Please add to the list or delete the citation.
14 Please check if the edits made to the book title in the reference “Cowan (1995)”are correct.
15 Please provide the page range for the reference “Cowan (1999).”
16 Please provide the date of the symposium in the reference “Haettig et al. (2004).”
17 Please provide the publisher location for the reference “Moré(1977).”
18 Please provide the volume number and page range for the reference “Wagner et al. (2008).”
19 Please check if the edits made to the sentence “Light-shaded vertical …” are correct.




















