
Modulation of expression of IL-8 gene in bronchial epithelial cells by 5-methoxypsoralen
12
Modulation of expression of IL-8 gene in bronchial epithelial cells by 5-methoxypsoralen Elena Nicolis a,1 , Ilaria Lampronti b,1 , Maria Cristina Dechecchi a , Monica Borgatti b , Anna Tamanini a , Valentino Bezzerri a , Nicoletta Bianchi b , Martina Mazzon a , Irene Mancini b , Maria Grazia Giri c , Paolo Rizzotti a , Roberto Gambari b, ⁎ ,2 , Giulio Cabrini a, ⁎ ,2 a Laboratory of Molecular Pathology, Laboratory of Clinical Chemistry and Haematology, University Hospital of Verona, Italy b ER-GenTech, Department of Biochemistry and Molecular Biology Interdisciplinary Center for the Study of Inflammation, University of Ferrara, Italy c Department of Medical Physics, University Hospital of Verona, Italy abstract article info Article history: Received 7 April 2009 Received in revised form 29 July 2009 Accepted 19 August 2009 Keywords: Cystic fibrosis Cytokine Inflammation Pseudomonas aeruginosa Psoralen Persistent recruitment of neutrophils in the bronchi of cystic fibrosis patients contributes to airway tissue damage, suggesting the importance of intervening on the expression of the neutrophil chemokine IL-8. Extracts from plants have been investigated to select components able to reduce IL-8 expression in bronchial epithelial cells challenged with Pseudomonas aeruginosa. Extracts and purified components have been added to cells 24 h before pro-inflammatory challenge with P. aeruginosa and IL-8 transcription was quantified in the IB3-1 CF cells in vitro. P. aeruginosa-dependent IL-8 mRNA induction was increased by Argemone mexicana and Vernonia anthelmintica whereas no significant modification of transcription was observed with Aphanamixis polystachya, Lagerstroemia speciosa and Hemidesmus indicus. Finally, inhibition of IL-8 was observed with Polyalthia longifolia (IC 50 = 200 μg/ml) and Aegle marmelos (IC 50 =20 μg/ml). Compounds from A. marmelos were isolated and identified by GC–MS. No significant effect was observed with butyl-p- tolyl sulphate, whereas the inhibition obtained with 6-methyl-4-chromanone concentration was accompanied by an anti-proliferative effect. On the contrary, 5-methoxypsoralen resulted in IL-8 inhibition at 10 μM concentration, without effects on cell proliferation. In synthesis, 5-methoxypsoralen can be taken into consideration to investigate mechanisms of neutrophil chemotactic signalling and for its potential application in modulating the excessive CF lung inflammation. © 2009 Elsevier B.V. All rights reserved. 1. Introduction Cystic fibrosis (CF) is a genetic disease caused by mutations of the Cystic Fibrosis Transmembrane conductance Regulator (CFTR) gene which encodes for a chloride channel. Although CFTR is expressed in several epithelia, the consequences of mutated CFTR for morbidity and mortality in CF are most important in the respiratory tract. Data from CF patients and experimental models supported the concept that mutated CFTR is associated with a hyper-inflammatory background originating from CF respiratory epithelial cells, which results in PMN recruitment, mainly sustained by release of IL-8 [1–5]. Different mechanisms have been proposed to explain the hyper-expression of IL-8 [6,7], possibly intervening on histone acetylation [8]. Within the lumen of CF airways, nucleic acids derived from PMNs further reduce mucociliary clearance, thus favouring bacterial colonization by S. aureus, H. influenzae and ultimately Pseudomonas aeruginosa. PMNs, continuously activated by the presence of bacterial products, release Reactive Oxygen Species and proteases, and contribute to lung tissue injury, with progressive bronchial wall damage. Hence, the exagger- ated PMN-dominated inflammatory process is a major determinant of the gradual decline in CF lung function [9,10]. Breaking this circle will require not only eradication of the infection but also reduction of the inflammatory background. Strategies to improve this situation for the patients concentrate on (1) the correction of the basic defect by CFTR gene transfer [11], (2) the correction of the CFTR alterations due to stop codon mutations [12], (3) the development of novel pharmacological correctors and potentiators of mutated CFTR, (4) the development of anti-bacterial compounds and (5) the development of anti-inflamma- tory compounds. Correction of the basic defect by CFTR gene transfer has been shown to be a long way to go and obstacles primarily concern safety and efficacy of CFTR viral or liposomal constructs [11]. The International Immunopharmacology 9 (2009) 1411–1422 ⁎ Corresponding authors. Gambari is to be contacted at the Department of Biochemistry and Molecular Biology, Via L. Borsari 46, I-44100 Ferrara, Italy. Tel.: + 39 0532 974443; fax: + 39 0532 974500. Cabrini, Laboratory of Molecular Pathology, Laboratory of Clinical Chemistry and Haematology, University Hospital of Verona, Piazzale Stefani 1, I-37126 Verona, Italy. Tel.: +39 045 812 2364; fax: +39 045 812 2840. E-mail addresses: [email protected] (R. Gambari), [email protected] (G. Cabrini). 1 Contributed equally to this work. 2 Share senior authorship. 1567-5769/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.intimp.2009.08.013 Contents lists available at ScienceDirect International Immunopharmacology journal homepage: www.elsevier.com/locate/intimp
Transcript of Modulation of expression of IL-8 gene in bronchial epithelial cells by 5-methoxypsoralen

International Immunopharmacology 9 (2009) 1411–1422
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r.com/ locate / in t imp
Modulation of expression of IL-8 gene in bronchial epithelial cells by5-methoxypsoralen
Elena Nicolis a,1, Ilaria Lampronti b,1, Maria Cristina Dechecchi a, Monica Borgatti b, Anna Tamanini a,Valentino Bezzerri a, Nicoletta Bianchi b, Martina Mazzon a, Irene Mancini b, Maria Grazia Giri c,Paolo Rizzotti a, Roberto Gambari b,⁎,2, Giulio Cabrini a,⁎,2
a Laboratory of Molecular Pathology, Laboratory of Clinical Chemistry and Haematology, University Hospital of Verona, Italyb ER-GenTech, Department of Biochemistry and Molecular Biology Interdisciplinary Center for the Study of Inflammation, University of Ferrara, Italyc Department of Medical Physics, University Hospital of Verona, Italy
⁎ Corresponding authors. Gambari is to be contBiochemistry and Molecular Biology, Via L. Borsari 4+39 0532 974443; fax: +39 0532 974500. Cabrini, LaboLaboratory of Clinical Chemistry and Haematology, UPiazzale Stefani 1, I-37126 Verona, Italy. Tel.: +39 0452840.
E-mail addresses: [email protected] (R. Gambari), giulio.c1 Contributed equally to this work.2 Share senior authorship.
1567-5769/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.intimp.2009.08.013
a b s t r a c t
a r t i c l e i n f oArticle history:Received 7 April 2009Received in revised form 29 July 2009Accepted 19 August 2009
Keywords:Cystic fibrosisCytokineInflammationPseudomonas aeruginosaPsoralen
Persistent recruitment of neutrophils in the bronchi of cystic fibrosis patients contributes to airway tissuedamage, suggesting the importance of intervening on the expression of the neutrophil chemokine IL-8.Extracts from plants have been investigated to select components able to reduce IL-8 expression in bronchialepithelial cells challenged with Pseudomonas aeruginosa. Extracts and purified components have been addedto cells 24 h before pro-inflammatory challenge with P. aeruginosa and IL-8 transcription was quantified inthe IB3-1 CF cells in vitro. P. aeruginosa-dependent IL-8 mRNA induction was increased by Argemonemexicana and Vernonia anthelmintica whereas no significant modification of transcription was observed withAphanamixis polystachya, Lagerstroemia speciosa and Hemidesmus indicus. Finally, inhibition of IL-8 wasobserved with Polyalthia longifolia (IC50=200 μg/ml) and Aegle marmelos (IC50=20 μg/ml). Compoundsfrom A. marmelos were isolated and identified by GC–MS. No significant effect was observed with butyl-p-tolyl sulphate, whereas the inhibition obtained with 6-methyl-4-chromanone concentration wasaccompanied by an anti-proliferative effect. On the contrary, 5-methoxypsoralen resulted in IL-8 inhibitionat 10 μM concentration, without effects on cell proliferation. In synthesis, 5-methoxypsoralen can be takeninto consideration to investigate mechanisms of neutrophil chemotactic signalling and for its potentialapplication in modulating the excessive CF lung inflammation.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Cystic fibrosis (CF) is a genetic disease caused by mutations of theCystic Fibrosis Transmembrane conductance Regulator (CFTR) genewhich encodes for a chloride channel. Although CFTR is expressed inseveral epithelia, the consequences ofmutated CFTR formorbidity andmortality in CF are most important in the respiratory tract. Data fromCF patients and experimental models supported the concept thatmutated CFTR is associated with a hyper-inflammatory backgroundoriginating from CF respiratory epithelial cells, which results in PMNrecruitment, mainly sustained by release of IL-8 [1–5]. Different
acted at the Department of6, I-44100 Ferrara, Italy. Tel.:ratory of Molecular Pathology,niversity Hospital of Verona,812 2364; fax: +39 045 812
[email protected] (G. Cabrini).
l rights reserved.
mechanisms have been proposed to explain the hyper-expression ofIL-8 [6,7], possibly intervening on histone acetylation [8]. Withinthe lumen of CF airways, nucleic acids derived from PMNs furtherreduce mucociliary clearance, thus favouring bacterial colonization byS. aureus, H. influenzae and ultimately Pseudomonas aeruginosa. PMNs,continuously activated by the presence of bacterial products, releaseReactive Oxygen Species and proteases, and contribute to lung tissueinjury, with progressive bronchial wall damage. Hence, the exagger-ated PMN-dominated inflammatory process is a major determinant ofthe gradual decline in CF lung function [9,10]. Breaking this circle willrequire not only eradication of the infection but also reduction of theinflammatory background. Strategies to improve this situation for thepatients concentrate on (1) the correction of the basic defect by CFTRgene transfer [11], (2) the correction of the CFTRalterations due to stopcodon mutations [12], (3) the development of novel pharmacologicalcorrectors and potentiators of mutated CFTR, (4) the development ofanti-bacterial compounds and (5) the development of anti-inflamma-tory compounds. Correction of the basic defect by CFTR gene transferhas been shown to be a longway to go and obstacles primarily concernsafety and efficacy of CFTR viral or liposomal constructs [11]. The

1412 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
strategy based on the use of correctors of the stop mutations of theCFTR mRNA affecting a sub-set of CF patients has been demonstratedeffective on in vitro cell systems and experimental animals and isunder clinical trial [13,14]. Novel pharmacological correctors andpotentiators of mutated CFTR protein leave many unanswered ques-tions concerning their application in CF, including the degree ofcorrection of the defect that can be obtained, the safety in long termadministration and the efficacy in reverting the extensive lunginflammatory pathology [15]. Although anti-bacterial therapy hasbeen shown to be more promising since early eradication therapy hasbeen established in some centres, it remains questionable whether allpatientswill profit from this strategy [9,16,17]. In addition, besides thefact that anti-inflammatory molecules have demonstrated potentialbenefits in CF patients, enthusiasm has been tempered either bythe limited efficacy or the occurrence of undesired effects [9,10,18].Therefore considering that at present no fully satisfactory anti-inflammatory treatment is available for clinical use in CF patients,investigation on novel anti-inflammatory molecules for the treatmentof lung inflammation is considered a major priority in the CF commu-nity (www.cff.org). With respect to pro-inflammatory mechanisms,P. aeruginosa products interact with Toll-like and asialoGM1 receptorsexpressed on bronchial epithelial cells, which leads to downstreamactivation of nuclear transcription factors, including NF-κB [19] andtranscription of a cascade of pro-inflammatory cytokines andchemokines, first of all IL-8 [20]. Natural products have been used assources for the discovery of newdrugs on the basis of theirwidespreadapplication in traditional medicine to cure several human diseases[21]. Medicinal plants can be also employed to interfere with protein/DNA interactions, including inhibition of NF-κB activity [22–24]. Thus,screening libraries of plant extracts and theirmajor active componentsfor their anti-inflammatory activity, in suitable respiratory epithelialcells in vitro and pre-clinical animal models in vivo, could selectcandidate molecules of potential therapeutic effect. These anti-inflammatory molecules for respiratory diseases could be adminis-tered both systemically (e.g. orally) or locally (e.g. by aerosol). Herewefirstly screened the anti-inflammatory effect of a series of medicinalplant extracts from Bangladesh, such as Argemone mexicana, Vernoniaanthelmintica, Aphanamixis polystachya, Polyalthia longifolia, Hemides-mus indicus, Lagerstoemia speciosa and Aegle marmelos, and we foundthat the whole extract of A. marmelos has the strongest inhibitoryeffect on the P. aeruginosa-dependent IL-8 induction in human CF-derived bronchial IB3-1 cells without affecting cell proliferation. Byseparating the major components contained in A. marmelos by gas
Table 1Primers for quantification of gene transcripts.
Primer Sequence (5′–3′) Accessionnumber
nM Calibratorgene
ICAM-1-F TATGGCAACGACTCCTTCTCG NM_000201 15 GAPDHICAM-1-R CTCTGCGGTCACACTGACTGA 15IL 8-F GACCACACTGCGCCAACA AF_385628.2 15 GAPDHIL 8-R GCTCTCTTCCATCAGAAAGTTACATAATTT 15GROα-F GCCCAAACCGAAGTCATAGC NM_001511.1 2.5 RPIIGROα-R GAGGCAAGCTTTCCGCC 45GROγ-F CCGGACCCCACTGCG M_36821 2.5 G6PDGROγ-R TTCCCATTCTTGAGTGTGGCTA 15IL-6-F CGGTACATCCTCGACGGC NM_000600 2.5 GAPDHIL6-R CTTGTTACATGTTCTCCTTTCTCAGG 45IL1β-F CTCCACCTCCAGGGACAGGA BT_007213.1 45 EEF1γIL1β-R GGACATGGAGAACACCACTTGTT 15GAPDH-F GTGGAGTCCACTGGCGTCTT J0_4038 2.5GAPDH-R GCAAATGAGCCCAGCCTTC 15G6PD-F GCCAAGAAGAAGATCTACCCCA NM_000402.2 15G6PD-R AAGGCCATCCCGGAACAG 2.5RPII-F TCGAGCAGATCAGCAAGGTG NM_000937.2 2.5RPII-R TCTTGTTGTCTGTCTGTGGCAA 2.5eEF1γ-F AAACTGTGTGAGAAGATGGCCC NM_001404.3 15eEF1γ-R GGGTCTCTGCAAACTTTTTAGCA 2.5
chromatography and identifying their structure bymass spectrometry,we found that three major components, namely 5,6-dimethoxy-1-indanone, 2-hydroxy-cinnamic acid and 5-methoxy psoralen, repro-duce the inhibitory effect observed with the whole extract from A.marmelos.
2. Materials and methods
2.1. Materials
IB3-1 cells were purchased from LGC Promochem (Teddington,Middlesex, UK). LHC-8 basal medium and Platinum SYBR Green qPCRSuper Mix-UDG were purchased from Invitrogen (Carlsbad, CA, USA).Azithromycin (AZM) was from Pfizer (Roma, Italy). Trypticase soybroth (TSB) or agar (TSA) was purchased from Difco (Detroit, MI,USA). Fisons model GC8000 gas chromatograph was purchased fromThermo Finnigan (San Jose, CA, USA) and was interfaced to a Fisonmodel MD 800 quadrupole mass spectrometer. The software utilizedwas the Mass Spectrometry Data Handling System (version 1.12,Thermo Finnigan, San Jose, CA, USA) with NIST library to recognize allthe derivatives found in plant extracts. High Pure RNA isolation kitwas purchased from Roche (Mannheim, Germany). High CapacitycDNA Archive kit and Primer Express Software were purchased fromApplied Biosystems (Foster City, CA, USA). Sysmex XE-2100 Cyt-ometer was purchased from Dasit (Milan, Italy).
2.2. Cell cultures and bacteria
IB3-1 cells, derived from a CF patient with a ΔF508/W1282Xmutant genotype and immortalised with adeno12/SV40 [25], weregrown in LHC-8 basal medium, supplemented with 5% FBS in theabsence of gentamycin, at 37 °C/5% CO2. The laboratory strain ofP. aeruginosa PAO1 was grown in TSB or TSA.
2.3. Cell infection
This was performed as previously described [26]. Briefly, IB3-1cells were seeded at a density of 200,000 cells/cm2, 24 h beforeinfection. After adhesion, cells were starved in serum free LHC-8 for18 h. Before the experiment, PAO1 from overnight cultures in TSAplates were grown in 20 ml TSB broth at 37 °Cwith shaking until therewas an OD at 660 nm of about 1×109 CFU/ml, determined by dilutionplating. Bacteria were washed twice by resuspension in PBS and thento a final dilution in cell culture medium. The doses of PAO1 weredetermined by plating aliquots of dilutions on TSA plates and areexpressed as CFU/cell. Monolayers of cells were infected with PAO1 at37 °C/5% CO2 for 4 h with a dose ranging from 30 to 100 CFU/cell.
2.4. Medicinal plant extracts
The ethanolic extracts ofA.marmelosweredefattedwithpetroleumether and the defatted extract was successively fractionated withdifferent systems on the polarity basis as previously described [27].
Table 2
Gene RQ⁎ SEM# p§
ICAM-1 10 1 ⁎⁎
IL-8 2072 48 ⁎⁎⁎
GRO-α 2266 514 ⁎
GRO-γ 661 32 ⁎⁎
IL-6 106 17 ⁎
IL1-β 112 17 ⁎
IFN-γ 2 0.7 n.s.
RQ, Relative Quantification: average fold induction by PAO1, with respect to the basallevel of uninfected cells; #SEM, standard error of the mean; §statistical significancemade by paired Student's t test at p<0.05 (⁎), p<0.01 (⁎⁎), and p<0.001 (⁎⁎⁎).

Fig. 1. Effect of Polyalthia longifolia and Hemidesmus indicus on P. aeruginosa-dependent IL-8 transcription, on cell proliferation and on bacterial growth. IB3-1 cells were firstly pre-incubated for 24 or 48 h with 5, 50, 100, 200 or 500 μg/ml of extracts. Cells were then infected with PAO1 at doses ranging from 50 to 100 CFU/cell. Total RNA was isolated 4 h afterinfection and IL-8 transcript was quantified by real-Time qPCR. (A) Polyalthia longifolia, but not (B) Hemidesmus indicus, inhibited the transcription of IL-8. Results are expressed aspercentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data are mean±SEM from three independent experiments performed in duplicate.**p<0.01; ***p<0.001. (C) Polyalthia longifolia and (D) Hemidesmus indicus were tested in a cell proliferation assay after 24 or 48 h of incubation WITH IB3-1 cells. Results areexpressed as percentage, indicated by a dotted line, of the values obtained in untreated cells. Data are mean±SEM. Anti-bacterial activity of Polyalthia longifolia (E) and Hemidesmusindicus (F) against PAO1 strain after 24 h of incubation.
1413E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
2.5. Gas chromatography/mass spectrometry (GS/MS) analysis of thefractions of A. marmelos extracts
The GS/MS was performed as previously described [27]. Briefly, aFisons model GC8000 gas chromatograph interfaced to a Fison modelMD 800 quadrupolemass spectrometerwas used for all measurements.The fused-silica gas chromatographic capillary columnwasaMegaSE54(methyl phenyl polysiloxanes), 25 m×0.25 mm I.D. and 0.25 μm filmthickness [28]. One μl of sample dissolved into appropriate solvents wasinjected into thegas chromatograph. The software utilizedwas theMassSpectrometry Data Handling System with NIST library to recognize allthe derivatives found in plant extracts.
2.6. Cell treatment
IB3-1 cells were incubated for 24 or 2 h with different concentra-tions of the plant extracts or of the components identified in A. mar-melos. Cells were exposed to 8 µg/ml AZM.
2.7. Quantitation of transcripts of pro-inflammatory genes
This was carried out as described previously [29]. Briefly, totalRNA was isolated using High Pure RNA isolation kit following thesupplier's instructions. Reverse transcription (RT) was performedusing the High Capacity cDNA Archive kit: 1 μg of total RNA was
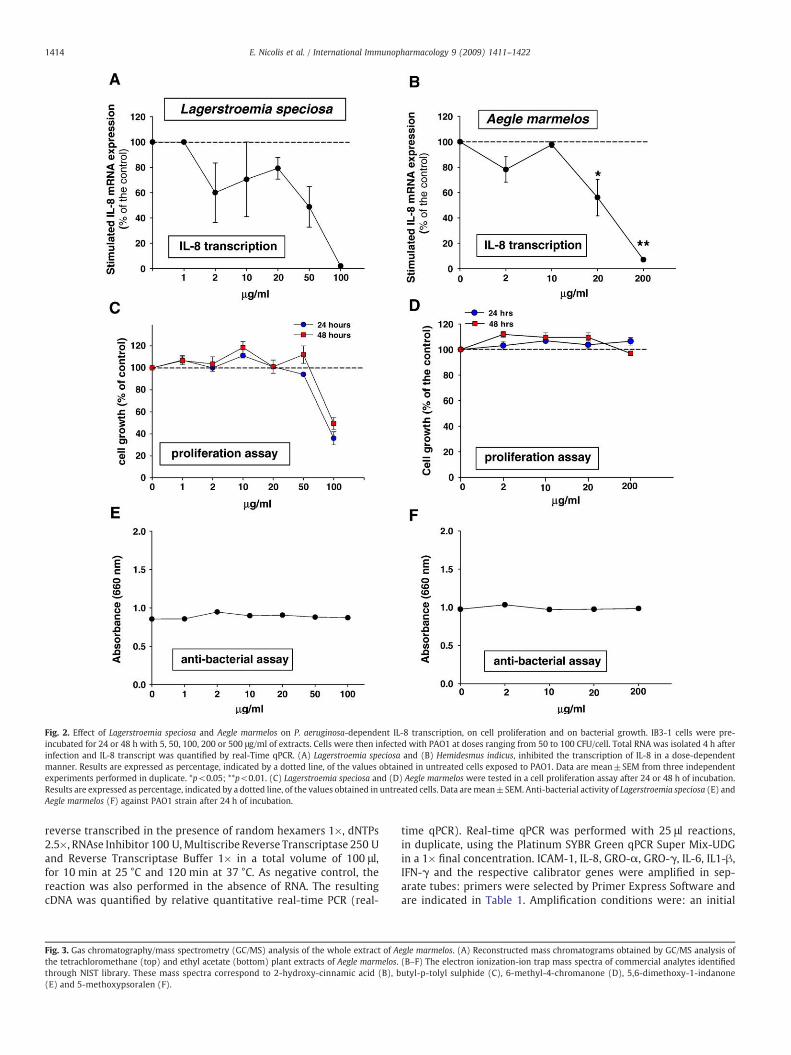
Fig. 2. Effect of Lagerstroemia speciosa and Aegle marmelos on P. aeruginosa-dependent IL-8 transcription, on cell proliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 h with 5, 50, 100, 200 or 500 μg/ml of extracts. Cells were then infected with PAO1 at doses ranging from 50 to 100 CFU/cell. Total RNA was isolated 4 h afterinfection and IL-8 transcript was quantified by real-Time qPCR. (A) Lagerstroemia speciosa and (B) Hemidesmus indicus, inhibited the transcription of IL-8 in a dose-dependentmanner. Results are expressed as percentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data are mean±SEM from three independentexperiments performed in duplicate. *p<0.05; **p<0.01. (C) Lagerstroemia speciosa and (D) Aegle marmelos were tested in a cell proliferation assay after 24 or 48 h of incubation.Results are expressed as percentage, indicated by a dotted line, of the values obtained in untreated cells. Data aremean±SEM. Anti-bacterial activity of Lagerstroemia speciosa (E) andAegle marmelos (F) against PAO1 strain after 24 h of incubation.
1414 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
reverse transcribed in the presence of random hexamers 1×, dNTPs2.5×, RNAse Inhibitor 100 U,Multiscribe Reverse Transcriptase 250 Uand Reverse Transcriptase Buffer 1× in a total volume of 100 μl,for 10 min at 25 °C and 120 min at 37 °C. As negative control, thereaction was also performed in the absence of RNA. The resultingcDNA was quantified by relative quantitative real-time PCR (real-
Fig. 3. Gas chromatography/mass spectrometry (GC/MS) analysis of the whole extract of Athe tetrachloromethane (top) and ethyl acetate (bottom) plant extracts of Aegle marmelos.through NIST library. These mass spectra correspond to 2-hydroxy-cinnamic acid (B), b(E) and 5-methoxypsoralen (F).
time qPCR). Real-time qPCR was performed with 25 μl reactions,in duplicate, using the Platinum SYBR Green qPCR Super Mix-UDGin a 1× final concentration. ICAM-1, IL-8, GRO-α, GRO-γ, IL-6, IL1-β,IFN-γ and the respective calibrator genes were amplified in sep-arate tubes: primers were selected by Primer Express Software andare indicated in Table 1. Amplification conditions were: an initial
egle marmelos. (A) Reconstructed mass chromatograms obtained by GC/MS analysis of(B–F) The electron ionization-ion trap mass spectra of commercial analytes identifiedutyl-p-tolyl sulphide (C), 6-methyl-4-chromanone (D), 5,6-dimethoxy-1-indanone

1415E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
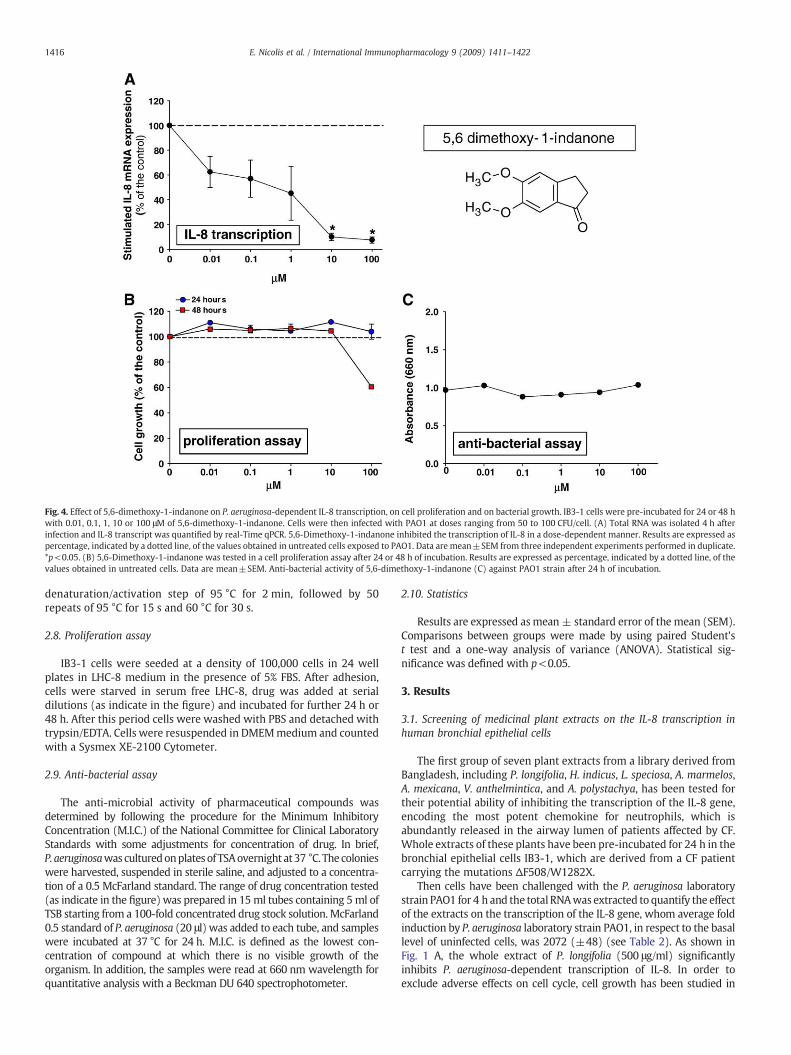
Fig. 4. Effect of 5,6-dimethoxy-1-indanone on P. aeruginosa-dependent IL-8 transcription, on cell proliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 hwith 0.01, 0.1, 1, 10 or 100 μM of 5,6-dimethoxy-1-indanone. Cells were then infected with PAO1 at doses ranging from 50 to 100 CFU/cell. (A) Total RNA was isolated 4 h afterinfection and IL-8 transcript was quantified by real-Time qPCR. 5,6-Dimethoxy-1-indanone inhibited the transcription of IL-8 in a dose-dependent manner. Results are expressed aspercentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data are mean±SEM from three independent experiments performed in duplicate.*p<0.05. (B) 5,6-Dimethoxy-1-indanone was tested in a cell proliferation assay after 24 or 48 h of incubation. Results are expressed as percentage, indicated by a dotted line, of thevalues obtained in untreated cells. Data are mean±SEM. Anti-bacterial activity of 5,6-dimethoxy-1-indanone (C) against PAO1 strain after 24 h of incubation.
1416 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
denaturation/activation step of 95 °C for 2 min, followed by 50repeats of 95 °C for 15 s and 60 °C for 30 s.
2.8. Proliferation assay
IB3-1 cells were seeded at a density of 100,000 cells in 24 wellplates in LHC-8 medium in the presence of 5% FBS. After adhesion,cells were starved in serum free LHC-8, drug was added at serialdilutions (as indicate in the figure) and incubated for further 24 h or48 h. After this period cells were washed with PBS and detached withtrypsin/EDTA. Cells were resuspended in DMEMmedium and countedwith a Sysmex XE-2100 Cytometer.
2.9. Anti-bacterial assay
The anti-microbial activity of pharmaceutical compounds wasdetermined by following the procedure for the Minimum InhibitoryConcentration (M.I.C.) of the National Committee for Clinical LaboratoryStandards with some adjustments for concentration of drug. In brief,P. aeruginosawasculturedonplates of TSAovernight at37 °C. The colonieswere harvested, suspended in sterile saline, and adjusted to a concentra-tion of a 0.5 McFarland standard. The range of drug concentration tested(as indicate in the figure) was prepared in 15 ml tubes containing 5ml ofTSB starting from a 100-fold concentrated drug stock solution. McFarland0.5 standard of P. aeruginosa (20 μl) was added to each tube, and sampleswere incubated at 37 °C for 24 h. M.I.C. is defined as the lowest con-centration of compound at which there is no visible growth of theorganism. In addition, the samples were read at 660 nm wavelength forquantitative analysis with a Beckman DU 640 spectrophotometer.
2.10. Statistics
Results are expressed asmean ± standard error of the mean (SEM).Comparisons between groups were made by using paired Student'st test and a one-way analysis of variance (ANOVA). Statistical sig-nificance was defined with p<0.05.
3. Results
3.1. Screening of medicinal plant extracts on the IL-8 transcription inhuman bronchial epithelial cells
The first group of seven plant extracts from a library derived fromBangladesh, including P. longifolia, H. indicus, L. speciosa, A. marmelos,A. mexicana, V. anthelmintica, and A. polystachya, has been tested fortheir potential ability of inhibiting the transcription of the IL-8 gene,encoding the most potent chemokine for neutrophils, which isabundantly released in the airway lumen of patients affected by CF.Whole extracts of these plants have been pre-incubated for 24 h in thebronchial epithelial cells IB3-1, which are derived from a CF patientcarrying the mutations ΔF508/W1282X.
Then cells have been challenged with the P. aeruginosa laboratorystrain PAO1 for 4 h and the total RNAwas extracted to quantify the effectof the extracts on the transcription of the IL-8 gene, whom average foldinduction by P. aeruginosa laboratory strain PAO1, in respect to the basallevel of uninfected cells, was 2072 (±48) (see Table 2). As shown inFig. 1 A, the whole extract of P. longifolia (500 μg/ml) significantlyinhibits P. aeruginosa-dependent transcription of IL-8. In order toexclude adverse effects on cell cycle, cell growth has been studied in
1417E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
the IB3-1 cells pre-incubated with the extracts for 24 and 48 h. Nochanges in cell proliferation have been observed even at the highestconcentration for twice the pre-incubation time showing an anti-inflammatory effect (48 h), as shown in Fig. 1 C. Thirdly, to exclude thatthe inhibition of the P. aeruginosa-dependent induction of IL-8 is due toan indirect anti-bacterial effect on P. aeruginosa, we excluded an anti-bacterial effect of the extract, as shown in Fig. 1 E. The analysis has beenextended to the other plant extracts available. As shown in Fig. 1 B,H. indicus does not significantly inhibit P. aeruginosa-dependent IL-8 transcription. The whole extract of L. speciosa (100 μg/ml) inhibitsP. aeruginosa-dependent IL-8 transcription (Fig. 2 A), however this isclearly associated with a sharp decrease in cell proliferation (Fig. 2 C),thus the effect on IL-8 mRNA is possibly dependent on a generalizedeffect on transcription. Whole extract of A. marmelos significantlyinhibits P. aeruginosa-dependent IL-8 transcription (Fig. 2 B), where50% decrease of P. aeruginosa-dependent IL-8 transcription has beenobtained with 20 μg/ml whole extract and almost complete inhibi-tion at 200 μg/ml concentration. Neither changes in cell proliferation(Fig. 2 D) nor effects on the viability of P. aeruginosa (Fig. 2 F) havebeen observed at the highest concentrations of thewhole extract. Thesame analysis has been carried out for the whole extracts of A. mexi-cana, V. anthelmintica, and A. polystachya. No significant changes inP. aeruginosa-dependent IL-8 transcription have been observed by pre-incubating the whole extracts of these plants with the bronchial cells(data not shown). In synthesis, within the group of 7 plant extractstested, a significant inhibitory effect on P. aeruginosa-dependent IL-8 transcription has been observed mainly with the whole extract ofA. marmelos, without undesired effects on cell proliferation, and a lessstrong effect with P. longifolia. Therefore we focused our attention on
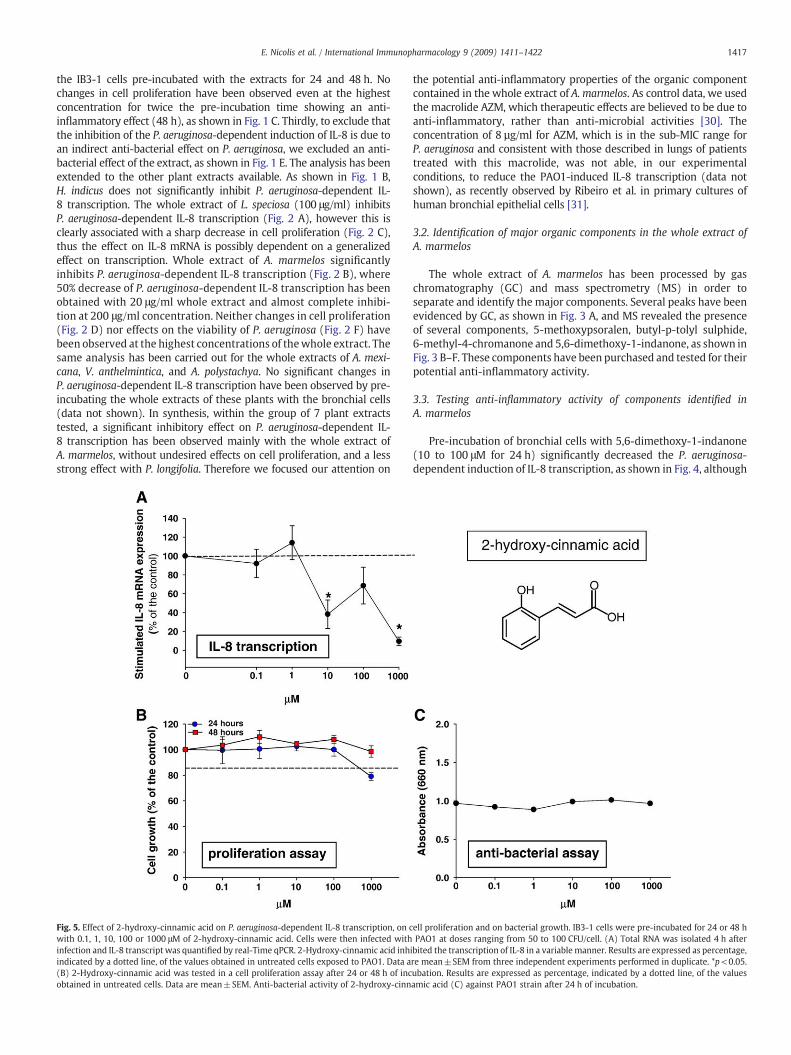
Fig. 5. Effect of 2-hydroxy-cinnamic acid on P. aeruginosa-dependent IL-8 transcription, on cwith 0.1, 1, 10, 100 or 1000 μM of 2-hydroxy-cinnamic acid. Cells were then infected withinfection and IL-8 transcript was quantified by real-Time qPCR. 2-Hydroxy-cinnamic acid inhindicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data a(B) 2-Hydroxy-cinnamic acid was tested in a cell proliferation assay after 24 or 48 h of incobtained in untreated cells. Data are mean±SEM. Anti-bacterial activity of 2-hydroxy-cinn
the potential anti-inflammatory properties of the organic componentcontained in the whole extract of A. marmelos. As control data, we usedthe macrolide AZM, which therapeutic effects are believed to be due toanti-inflammatory, rather than anti-microbial activities [30]. Theconcentration of 8 µg/ml for AZM, which is in the sub-MIC range forP. aeruginosa and consistent with those described in lungs of patientstreated with this macrolide, was not able, in our experimentalconditions, to reduce the PAO1-induced IL-8 transcription (data notshown), as recently observed by Ribeiro et al. in primary cultures ofhuman bronchial epithelial cells [31].
3.2. Identification of major organic components in the whole extract ofA. marmelos
The whole extract of A. marmelos has been processed by gaschromatography (GC) and mass spectrometry (MS) in order toseparate and identify the major components. Several peaks have beenevidenced by GC, as shown in Fig. 3 A, and MS revealed the presenceof several components, 5-methoxypsoralen, butyl-p-tolyl sulphide,6-methyl-4-chromanone and 5,6-dimethoxy-1-indanone, as shown inFig. 3 B–F. These components have been purchased and tested for theirpotential anti-inflammatory activity.
3.3. Testing anti-inflammatory activity of components identified inA. marmelos
Pre-incubation of bronchial cells with 5,6-dimethoxy-1-indanone(10 to 100 μM for 24 h) significantly decreased the P. aeruginosa-dependent induction of IL-8 transcription, as shown in Fig. 4, although
ell proliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 hPAO1 at doses ranging from 50 to 100 CFU/cell. (A) Total RNA was isolated 4 h after
ibited the transcription of IL-8 in a variable manner. Results are expressed as percentage,re mean±SEM from three independent experiments performed in duplicate. *p<0.05.ubation. Results are expressed as percentage, indicated by a dotted line, of the valuesamic acid (C) against PAO1 strain after 24 h of incubation.
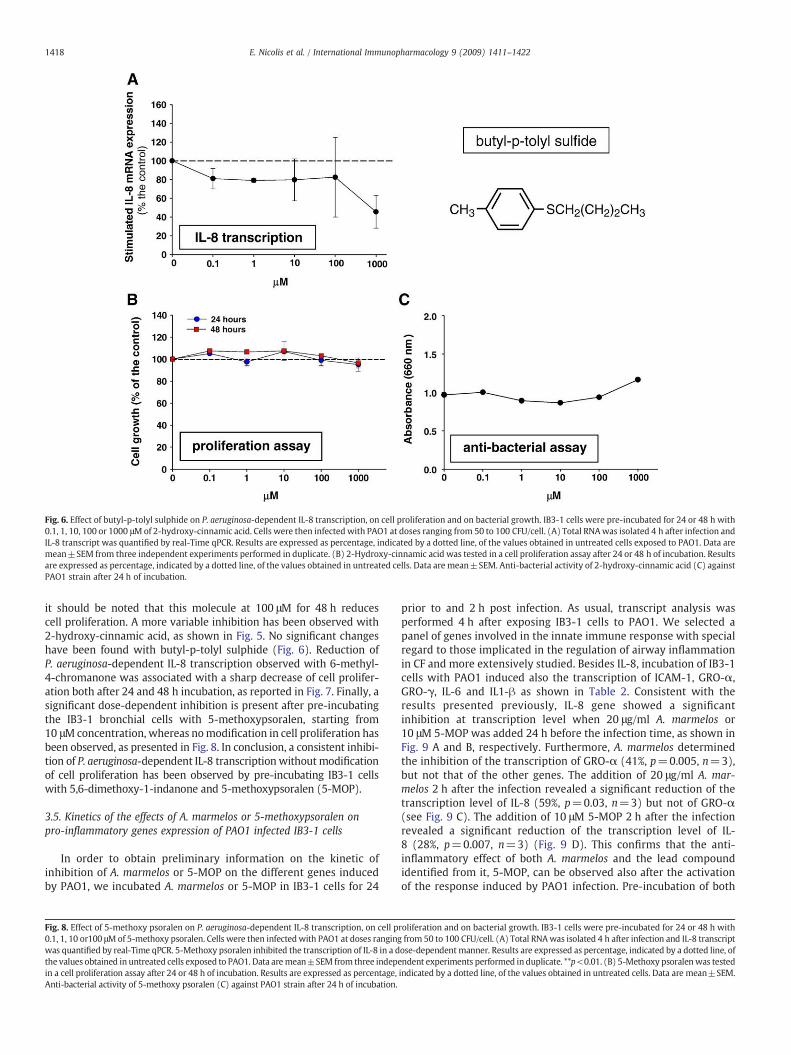
Fig. 6. Effect of butyl-p-tolyl sulphide on P. aeruginosa-dependent IL-8 transcription, on cell proliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 h with0.1, 1, 10, 100 or 1000 μMof 2-hydroxy-cinnamic acid. Cells were then infected with PAO1 at doses ranging from 50 to 100 CFU/cell. (A) Total RNAwas isolated 4 h after infection andIL-8 transcript was quantified by real-Time qPCR. Results are expressed as percentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data aremean±SEM from three independent experiments performed in duplicate. (B) 2-Hydroxy-cinnamic acid was tested in a cell proliferation assay after 24 or 48 h of incubation. Resultsare expressed as percentage, indicated by a dotted line, of the values obtained in untreated cells. Data are mean±SEM. Anti-bacterial activity of 2-hydroxy-cinnamic acid (C) againstPAO1 strain after 24 h of incubation.
1418 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
it should be noted that this molecule at 100 μM for 48 h reducescell proliferation. A more variable inhibition has been observed with2-hydroxy-cinnamic acid, as shown in Fig. 5. No significant changeshave been found with butyl-p-tolyl sulphide (Fig. 6). Reduction ofP. aeruginosa-dependent IL-8 transcription observed with 6-methyl-4-chromanone was associated with a sharp decrease of cell prolifer-ation both after 24 and 48 h incubation, as reported in Fig. 7. Finally, asignificant dose-dependent inhibition is present after pre-incubatingthe IB3-1 bronchial cells with 5-methoxypsoralen, starting from10 μMconcentration, whereas nomodification in cell proliferation hasbeen observed, as presented in Fig. 8. In conclusion, a consistent inhibi-tion of P. aeruginosa-dependent IL-8 transcription without modificationof cell proliferation has been observed by pre-incubating IB3-1 cellswith 5,6-dimethoxy-1-indanone and 5-methoxypsoralen (5-MOP).
3.5. Kinetics of the effects of A. marmelos or 5-methoxypsoralen onpro-inflammatory genes expression of PAO1 infected IB3-1 cells
In order to obtain preliminary information on the kinetic ofinhibition of A. marmelos or 5-MOP on the different genes inducedby PAO1, we incubated A. marmelos or 5-MOP in IB3-1 cells for 24
Fig. 8. Effect of 5-methoxy psoralen on P. aeruginosa-dependent IL-8 transcription, on cell p0.1, 1, 10 or100 μMof 5-methoxy psoralen. Cells were then infectedwith PAO1 at doses rangingwas quantified by real-Time qPCR. 5-Methoxy psoralen inhibited the transcription of IL-8 in a dthe values obtained in untreated cells exposed to PAO1. Data aremean±SEM from three indepein a cell proliferation assay after 24 or 48 h of incubation. Results are expressed as percentage,Anti-bacterial activity of 5-methoxy psoralen (C) against PAO1 strain after 24 h of incubation.
prior to and 2 h post infection. As usual, transcript analysis wasperformed 4 h after exposing IB3-1 cells to PAO1. We selected apanel of genes involved in the innate immune response with specialregard to those implicated in the regulation of airway inflammationin CF and more extensively studied. Besides IL-8, incubation of IB3-1cells with PAO1 induced also the transcription of ICAM-1, GRO-α,GRO-γ, IL-6 and IL1-β as shown in Table 2. Consistent with theresults presented previously, IL-8 gene showed a significantinhibition at transcription level when 20 μg/ml A. marmelos or10 μM 5-MOP was added 24 h before the infection time, as shown inFig. 9 A and B, respectively. Furthermore, A. marmelos determinedthe inhibition of the transcription of GRO-α (41%, p=0.005, n=3),but not that of the other genes. The addition of 20 μg/ml A. mar-melos 2 h after the infection revealed a significant reduction of thetranscription level of IL-8 (59%, p=0.03, n=3) but not of GRO-α(see Fig. 9 C). The addition of 10 μM 5-MOP 2 h after the infectionrevealed a significant reduction of the transcription level of IL-8 (28%, p=0.007, n=3) (Fig. 9 D). This confirms that the anti-inflammatory effect of both A. marmelos and the lead compoundidentified from it, 5-MOP, can be observed also after the activationof the response induced by PAO1 infection. Pre-incubation of both
roliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 h withfrom 50 to 100 CFU/cell. (A) Total RNAwas isolated 4 h after infection and IL-8 transcriptose-dependentmanner. Results are expressed as percentage, indicated by a dotted line, ofndent experiments performed in duplicate. **p<0.01. (B) 5-Methoxy psoralenwas testedindicated by a dotted line, of the values obtained in untreated cells. Data are mean±SEM.
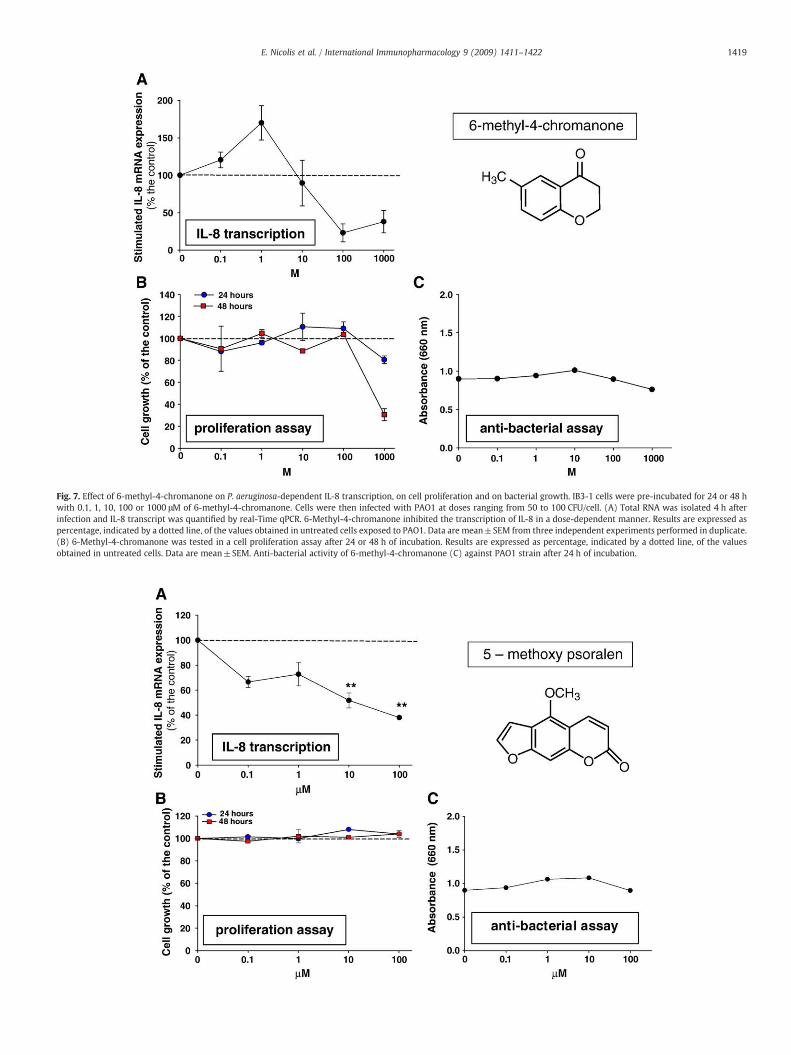
Fig. 7. Effect of 6-methyl-4-chromanone on P. aeruginosa-dependent IL-8 transcription, on cell proliferation and on bacterial growth. IB3-1 cells were pre-incubated for 24 or 48 hwith 0.1, 1, 10, 100 or 1000 μM of 6-methyl-4-chromanone. Cells were then infected with PAO1 at doses ranging from 50 to 100 CFU/cell. (A) Total RNA was isolated 4 h afterinfection and IL-8 transcript was quantified by real-Time qPCR. 6-Methyl-4-chromanone inhibited the transcription of IL-8 in a dose-dependent manner. Results are expressed aspercentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data are mean±SEM from three independent experiments performed in duplicate.(B) 6-Methyl-4-chromanone was tested in a cell proliferation assay after 24 or 48 h of incubation. Results are expressed as percentage, indicated by a dotted line, of the valuesobtained in untreated cells. Data are mean±SEM. Anti-bacterial activity of 6-methyl-4-chromanone (C) against PAO1 strain after 24 h of incubation.
1419E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
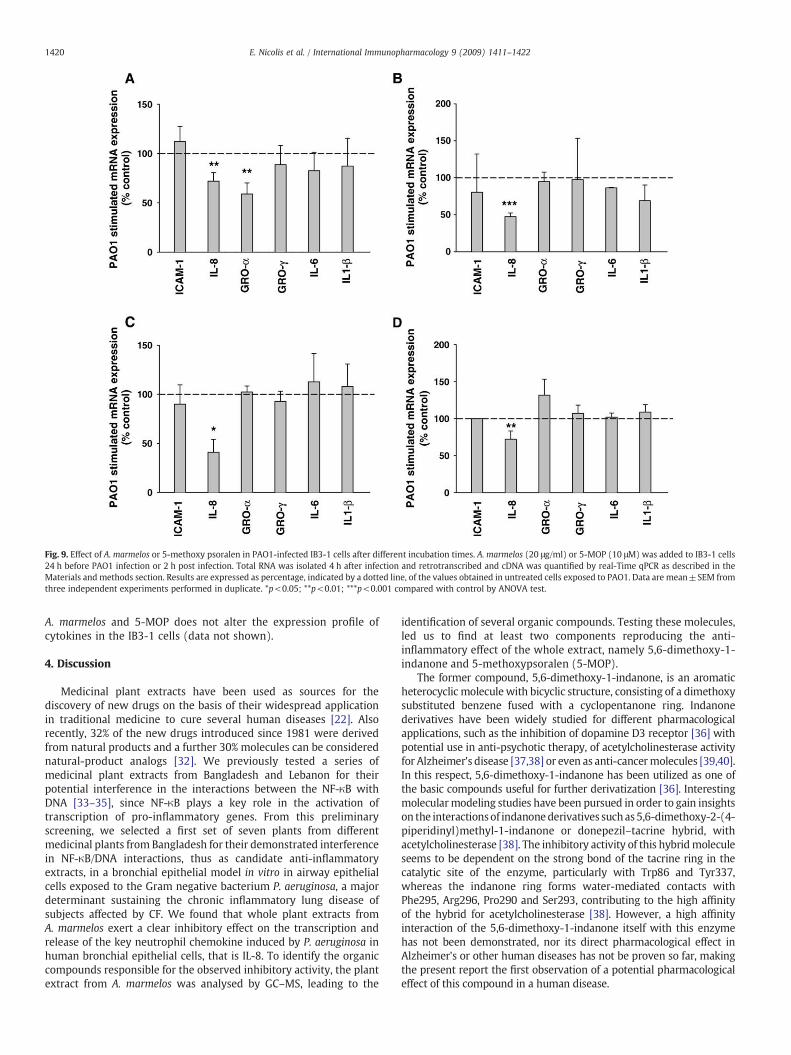
Fig. 9. Effect of A. marmelos or 5-methoxy psoralen in PAO1-infected IB3-1 cells after different incubation times. A. marmelos (20 μg/ml) or 5-MOP (10 μM) was added to IB3-1 cells24 h before PAO1 infection or 2 h post infection. Total RNA was isolated 4 h after infection and retrotranscribed and cDNA was quantified by real-Time qPCR as described in theMaterials and methods section. Results are expressed as percentage, indicated by a dotted line, of the values obtained in untreated cells exposed to PAO1. Data are mean±SEM fromthree independent experiments performed in duplicate. *p<0.05; **p<0.01; ***p<0.001 compared with control by ANOVA test.
1420 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
A. marmelos and 5-MOP does not alter the expression profile ofcytokines in the IB3-1 cells (data not shown).
4. Discussion
Medicinal plant extracts have been used as sources for thediscovery of new drugs on the basis of their widespread applicationin traditional medicine to cure several human diseases [22]. Alsorecently, 32% of the new drugs introduced since 1981 were derivedfrom natural products and a further 30% molecules can be considerednatural-product analogs [32]. We previously tested a series ofmedicinal plant extracts from Bangladesh and Lebanon for theirpotential interference in the interactions between the NF-κB withDNA [33–35], since NF-κB plays a key role in the activation oftranscription of pro-inflammatory genes. From this preliminaryscreening, we selected a first set of seven plants from differentmedicinal plants from Bangladesh for their demonstrated interferencein NF-κB/DNA interactions, thus as candidate anti-inflammatoryextracts, in a bronchial epithelial model in vitro in airway epithelialcells exposed to the Gram negative bacterium P. aeruginosa, a majordeterminant sustaining the chronic inflammatory lung disease ofsubjects affected by CF. We found that whole plant extracts fromA. marmelos exert a clear inhibitory effect on the transcription andrelease of the key neutrophil chemokine induced by P. aeruginosa inhuman bronchial epithelial cells, that is IL-8. To identify the organiccompounds responsible for the observed inhibitory activity, the plantextract from A. marmelos was analysed by GC–MS, leading to the
identification of several organic compounds. Testing these molecules,led us to find at least two components reproducing the anti-inflammatory effect of the whole extract, namely 5,6-dimethoxy-1-indanone and 5-methoxypsoralen (5-MOP).
The former compound, 5,6-dimethoxy-1-indanone, is an aromaticheterocyclic molecule with bicyclic structure, consisting of a dimethoxysubstituted benzene fused with a cyclopentanone ring. Indanonederivatives have been widely studied for different pharmacologicalapplications, such as the inhibition of dopamine D3 receptor [36] withpotential use in anti-psychotic therapy, of acetylcholinesterase activityfor Alzheimer's disease [37,38] or even as anti-cancermolecules [39,40].In this respect, 5,6-dimethoxy-1-indanone has been utilized as one ofthe basic compounds useful for further derivatization [36]. Interestingmolecularmodeling studies have been pursued in order to gain insightson the interactionsof indanonederivatives suchas5,6-dimethoxy-2-(4-piperidinyl)methyl-1-indanone or donepezil–tacrine hybrid, withacetylcholinesterase [38]. The inhibitory activity of this hybridmoleculeseems to be dependent on the strong bond of the tacrine ring in thecatalytic site of the enzyme, particularly with Trp86 and Tyr337,whereas the indanone ring forms water-mediated contacts withPhe295, Arg296, Pro290 and Ser293, contributing to the high affinityof the hybrid for acetylcholinesterase [38]. However, a high affinityinteraction of the 5,6-dimethoxy-1-indanone itself with this enzymehas not been demonstrated, nor its direct pharmacological effect inAlzheimer's or other human diseases has not be proven so far, makingthe present report the first observation of a potential pharmacologicaleffect of this compound in a human disease.

1421E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
5-MOP belongs to a class of tricyclic aromatic molecules also knownas furocoumarins found in different plant species that are utilized inthe therapy of chronic inflammatory diseases, such as severe forms ofpsoriasis, eczema, lichen planus and cutaneous T-cell lymphoma, asFood and Drug Administration-approved drugs known as bergapten(5-MOP) or methoxalen (8-methoxypsoralen) [41]. Mechanism ofanti-inflammatory action in skin diseases has been elucidated, asmethoxypsoralens penetrate psoriatic skin cells and intercalatebetween nucleic acids. Application of Ultraviolet A radiation causesformation of monofunctional and then bifunctional cross-linkagesbetween psoralen and pyrimidine bases [42,43]. This leads toapoptosis of different cells within the psoriatic plaque, includingT cells and antigen presenting cells, which results in overall reductionof release of chemokines and pro-inflammatory cytokines as a functionof reduction in the number of these immune cells. Besides the appli-cation in chronic inflammatory diseases of the skin, we recently foundthat furocoumarins associated with Ultraviolet A irradiation promote atlow concentration a preferential increase of gamma-globinmRNA tran-scription whereas at high doses they activate caspases and mitochon-drial depolarization, leading to apoptosis, in K562 erythroid cells[44,45]. However, as far as 5-MOP is concerned, although its use inassociationwithUltraviolet A irradiation for skin diseases is known froma long time [46], to the best of our knowledge, this is thefirst report of itsdirect anti-inflammatory effect in the absence of irradiation, whosemechanisms require further investigation. As far as the clinical indi-cation of the lead compounds identified here to cure the chronic lungdisease of patients affected by cystic fibrosis, consensus is growing onthe need of intervening in parallel on the three major pathophysiolog-ical issues of CF lung pathology [47], that is the basic defect of iontransport regulation and airway surface fluid hydration, the chronicbacterial colonization by P. aeruginosa and the excessive inflammatoryresponse, which contributes to the progressive damage of therespiratory tissue and lung function insufficiency. Thus potential clinicalapplication ofmolecules such as 5-MOP, an already FDA-approved drug,as an anti-inflammatory molecule for cystic fibrosis patients deservesfurther investigation in pre-clinical and clinical studies.
Acknowledgements
This work was supported by grants from Fondazione Cariverona –
Bando 2005 – Malattie rare e della povertà (to GC), the Italian CysticFibrosis Research Foundation (grant FFC # 13/2007) with the contri-bution of “Associazione Veneta per la lotta alla fibrosi cistica” (to GC andRG), the Fondazione Cariparo (to RG) andMUR COFIN-2005 (to RG). VBis fellow of the “Fondazione Cariverona”, EN is fellow of the “AziendaOspedaliera di Verona”. We are grateful to Alice Prince (ColumbiaUniversity, NY, U.S.A.) for donating the P. aeruginosa laboratory strainPAO1 and to Federica Quiri for excellent technical assistance.
References
[1] Khan TZ, Wagener JS, Bost T, Martinez J, Accurso FJ, Riches DW. Early pulmonaryinflammation in infants with cystic fibrosis. Am J Respir Crit Care Med 1995;151:1075–82.
[2] Noah TL, Black HR, Cheng PW, Wood RE, Leigh MW. Nasal and bronchoalveolarlavage fluid cytokines in early cystic fibrosis. J Infect Dis 1997;175:638–47.
[3] DiMango E, Ratner AJ, Bryan R, Tabibi S, Prince A. Activation of NF-kB by adherentPseudomonas aeruginosa in normal and cystic fibrosis respiratory epithelial cells.J Clin Invest 1998;101:2598–605.
[4] Puchelle E, De Bentzmann S, Hubeau C, Jacquot J, Gaillard D. Mechanisms involvedin cystic fibrosis airway inflammation. Pediatr Pulmonol 2001;S23:143–5.
[5] Joseph T, Look D, Ferkol T. NF-kB activation and sustained IL-8 gene expression inprimary cultures of cystic fibrosis airway epithelial cells stimulated with Pseudo-monas aeruginosa. Am J Physiol Lung Cell Mol Physiol 2005;288:L471–9.
[6] Meusser B, Hirsch C, Jarosch E, Sommer T. ERAD: the long road to destruction. NatCell Biol 2005;7:766–72.
[7] Vij N, Fang S, Zeitlin PL. Selective inhibition of endoplasmic reticulum-associateddegradation rescues DeltaF508-cystic fibrosis transmembrane regulator andsuppresses interleukin-8 levels: therapeutic implications. J Biol Chem2006;281:17369–78.
[8] Bartling TR, Drumm ML. Oxidative stress causes IL8 promoter hyperacetylation incystic fibrosis airway cell models. Am J Respir Cell Mol Biol 2009;40:58–65.
[9] DöringG,Worlitzsch D. Inflammation in cysticfibrosis and itsmanagement. PaediatrRespir Rev 2000;1:101–6.
[10] Koehler DR, Downey GP, Sweezey NB, Tanswell AK, Hu J. Lung inflammation as atherapeutic target in cystic fibrosis. Am J Respir Cell Mol Biol 2004;31:377–81.
[11] Griesenbach U, Geddes DM, Alton EW. Gene therapy progress and prospects:cystic fibrosis. Gene Ther 2006;13:1061–7.
[12] Wilschanski M, Yahav Y, Yaacov Y, Blau H, Bentur L, Rivlin J, et al. Gentamicin-induced correction of CFTR function in patients with cystic fibrosis and CFTR stopmutations. N Engl J Med 2003;349:1433–41.
[13] Du M, Liu X, Welch EM, Hirawat S, Peltz SW, Bedwell DM. PTC124 is an orallybioavailable compound that promotes suppression of the human CFTR-G542Xnonsense allele in a CF mouse model. Proc Natl Acad Sci U S A 2008;105:2064–9.
[14] Kerem E, Hirawat S, Armoni S, Yaakov Y, Shoseyov D, Cohen M, et al. Effectivenessof PTC124 treatment of cystic fibrosis caused by nonsensemutations: a prospectivephase II trial. Lancet 2008;372:719–27.
[15] MacDonald KD, McKenzie KR, Zeitlin PL. Cystic fibrosis transmembrane regulatorprotein mutations: ‘class’ opportunity for novel drug innovation. Paediatr Drugs2007;9:1–10.
[16] Ratjen F. What's new in CF airway inflammation: an update. Paediatr Respir Rev2006;7(Suppl 1):S70–2.
[17] Rubin BK. CFTR is a modulator of airway inflammation. Am J Physiol Lung Cell MolPhysiol 2007;292:L381–2.
[18] Konstan MW, Doring G, Lands LC, Hilliard KA, Koker P, Bhattacharya S, et al.Results of a phase II clinical trial of BIIL284 BS for the treatment of CF lung disease.Pediatr Pulmonol S 2005;28:125–6.
[19] Kawai T, Akira S. TLR signaling. Cell Death Differ 2006;13:816–25.[20] Hoffmann E, Dittrich-Breiholz O, Holtmann H, Kracht M. Multiple control of
interleukin-8 gene expression. J Leukoc Biol 2002;72:847–55.[21] Koehn FE, Carter GT. The evolving role of natural products in drug discovery. Nat
Rev Drug Discov 2005;4:206–20.[22] Akesson C, Lindgren H, Pero RW, Leanderson T, Ivars F. An extract of Uncaria
tomentosa inhibiting cell division and NF-kappa B activity without inducing celldeath. Int Immunopharmacol 2003;3:1889–900.
[23] Hwang BY, Lee JH, Koo TH, Kim HS, Hong YS, Ro JS, et al. Kaurane diterpenes fromIsodon japonicus inhibit nitric oxide and prostaglandin E2 production and NF-kappaB activation in LPS-stimulated macrophage RAW264.7 cells. Planta Med2001;67:406–10.
[24] Lee JH, Ko WS, Kim YH, Kang HS, Kim HD, Choi BT. Anti-inflammatory effect of theaqueous extract from Lonicera japonica flower is related to inhibition of NF-kappaB activation through reducing I-kappaBalpha degradation in rat liver. Int JMol Med 2001;7:79–83.
[25] Zeitlin PL, Lu L, Rhim J, Cutting G, Stetten G, Kieffer KA, et al. A cystic fibrosisbronchial epithelial cell line: immortalization by adeno-12-SV40 infection. Am JRespir Cell Mol Biol 1991;14:313–9.
[26] Dechecchi MC, Nicolis E, Bezzerri V, Vella A, Colombatti M, Assael BM, et al. MPB-07 reduces the inflammatory response to Pseudomonas aeruginosa in cysticfibrosis bronchial cells. Am J Respir Cell Mol Biol 2007;36:615–24.
[27] Khan MTH, Lampronti I, Martello D, Bianchi N, Jabbar S, Choudhuri MSK, et al.Identification of pyrogallol as an antiproliferative compound present in extractsfrom the medicinal plant Emblica officinalis: effects on in vitro cell growth ofhuman tumor cell lines. Int J Onc 2002;20:187–92.
[28] Tomkins BA, Sega GA. Determination of thiodyglycol in groundwater using solid-phase extraction followed by gas chromatography with mass spectrometricdetection in the selected-ion mode. J Chromatogr 2001;909:13–28.
[29] Nicolis E, Lampronti I, Dechecchi MC, Borgatti M, Tamanini A, Bianchi N, et al.Pyrogallol, an active compound from the medicinal plant Emblica officinalis,regulates expression of pro-inflammatory genes in bronchial epithelial cells. IntImmunopharmacol 2008;10:1672–80.
[30] Jaffe A, Bush A. Anti-inflammatory effects of macrolides in lung disease. PediatrPulmonol 2001;31:464–73.
[31] Ribeiro CMP, Hurd H, Wu Y, Martino MEB, Jones L, Brighton B, et al. Azithromycintreatment alters gene expression in inflammatory, lipid metabolism, and cell cyclepathways in well-differentiated human airway epithelia. PLoS One 2009;4:e5806.
[32] Schmidt BM, Ribnicky DM, Lipsky PE, Raskin I. Revisiting the ancient concept ofbotanical therapeutics. Nat Chem Biol 2007;3:360–6.
[33] Lambertini E, Piva R, Khan MT, Lampronti I, Bianchi N, Borgatti M, et al. Effects ofextracts from Bangladeshi medicinal plants on in vitro proliferation of humanbreast cancer cell lines and expression of estrogen receptor alpha gene. Int J Oncol2004;24:419–23.
[34] Lampronti I, KhanMT, Bianchi N, Ather A, Borgatti M, Vizziello L, et al. Bangladeshimedicinal plant extracts inhibiting molecular interactions between nuclear factorsand target DNA sequencesmimicking NF-kappaB binding sites. Med Chem2005;1:327–33.
[35] Lampronti I, Khan MT, Borgatti M, Bianchi N, Gambari R. Inhibitory effects ofBangladeshi medicinal plant extracts on interactions between transcriptionfactors and target DNA sequences. Evid Based Complement Alternat Med2008;5:303–12.
[36] Haadsma-Svensson SR, Cleek KA, Dinh DM, Duncan JN, Haber CL, Huff RM, et al.Dopamine D(3) receptor antagonists. 1. Synthesis and structure–activity relation-ships of 5, 6-dimethoxy-N-alkyl- and N-alkylaryl-substituted 2-aminoindans.J Med Chem 2001;44:4716–32.
[37] Sheng R, Xu Y, Hu C, Zhang J, Lin X, Li J, et al. Design, synthesis and AChE inhibitoryactivity of indanone and aurone derivatives. Eur J Med Chem 2009;44:772–84.

1422 E. Nicolis et al. / International Immunopharmacology 9 (2009) 1411–1422
[38] Camps P, Formosa X, Galdeano C, Gómez T, Muñoz-Torrero D, Scarpellini M, et al.Novel donepezil-based inhibitors of acetyl- and butyrylcholinesterase and acetyl-cholinesterase-induced beta-amyloid aggregation. J Med Chem 2008;51:3588–98.
[39] Saxena HO, Faridi U, Kumar JK, Luqman S, Darokar MP, Shanker K, et al. Synthesisof chalcone derivatives on steroidal framework and their anticancer activities.Steroids 2007;72:892–900.
[40] Saxena HO, Faridi U, Srivastava S, Kumar JK, Darokar MP, Luqman S, et al. Gallicacid-based indanone derivatives as anticancer agents. Bioorg Med Chem Lett2008;18:3914–8.
[41] MenterA,GriffithsCE. Current and futuremanagement of psoriasis. Lancet2007;370:272–84.
[42] Kitamura N, Kohtani S, Nakagaki R. Molecular aspects of furocoumarin reactions:photophysics, photochemistry, photobiology, and structural analysis. J PhotochemPhotobiol 2007;6:168–85.
[43] Stern RS. Psoralen and ultraviolet A light therapy for psoriasis. N Engl J Med2007;357:682–90.
[44] Viola G, Vedaldi D, Dall'Acqua F, Lampronti I, Bianchi N, Zuccato C, et al.Furocoumarins photolysis products induce differentiation of human erythroidcells. J Photochem Photobiol 2008;92:24–8.
[45] Viola G, Vedaldi D, Dall'Acqua F, Fortunato E, Basso G, Bianchi N, et al. Induction ofgamma-globin mRNA, erythroid differentiation and apoptosis in UVA-irradiatedhuman erythroid cells in the presence of furocumarin derivatives. BiochemPharmacol 2008;75:810–25.
[46] Tanew A, Ortel B, Rappersberger K, Hönigsmann H. 5-Methoxypsoralen (bergap-ten) for photochemotherapy. Bioavailability, phototoxicity, and clinical efficacy inpsoriasis of a new drug preparation. J Am Acad Dermatol 1988;18:333–8.
[47] Frerichs C, Smyth A. Treatment strategies for cystic fibrosis: what's in the pipeline?Expert Opin Pharmacother 2009;10:1191–202.











![bronchial-hygiene-therapy.ppt [Read-Only] - Semantic Scholar](https://static.fdokumen.com/doc/165x107/6317b9679076d1dcf80beb6a/bronchial-hygiene-therapyppt-read-only-semantic-scholar.jpg)








