
Mills mol micro 2002
11
© 2002 Blackwell Science Ltd Truncation of merozoite surface protein 3 disrupts its trafficking and that of acidic–basic repeat protein to the surface of Plasmodium falciparum merozoites nevertheless suffering significant morbidity. Drug resist- ance continues to increase and as yet there is no effec- tive vaccine against malaria. A large number of potential vaccine candidates have been identified; however, little is known of their functions (Barnwell and Galinski, 1998). Many of the putative vaccine antigens are located on the merozoite surface. As a major interface between host and pathogen, the merozoite surface is an obvious target for the development of a vaccine. Proteins comprising the merozoite surface are exposed to the host immune system before and during invasion into the erythrocyte, albeit for a short time. Thus far, several merozoite surface proteins have been identified, and included in this group are the mero- zoite surface proteins 1–6 and 8 (Smythe et al., 1988; Blackman et al., 1990; McColl et al., 1994; Oeuvray et al., 1994a; Marshall et al., 1997; Marshall et al., 1998; Black et al., 2001; Trucco et al., 2001). MSP1 is an important vaccine candidate, and there is evidence to suggest that it can bind to the surface of the eryth- rocyte (Kaslow et al., 1994), whereas MSP2–6 and 8 are less characterized. Indeed, there is very little data defining the roles of any of these antigens in parasite invasion. Merozoite surface protein 3 (MSP3) is a polymorphic antigen, which has a number of structural domains (Fig. 1A). These domains include three blocks of four heptad repeats of the type AXXAXXX, a hydrophilic region and a putative leucine zipper sequence at the C-terminus (Fig. 1A) (McColl et al., 1994). The solution structure of the heptad repeats has been solved, and each block appears to form an a-helix, which may interact to form a somewhat unstable coiled-coil (Mulhern et al., 1995). Despite some sequence diversity of the msp3 gene between P. falciparum isolates, there is significant con- servation (McColl et al., 1994; Escalante et al., 1998). Indeed, the alanine residues within the heptad repeat regions and the C-terminal half of the protein, which includes the putative leucine zipper region, are strongly conserved (McColl and Anders, 1997). Current evidence supports MSP3 as a vaccine candi- date. MSP3 was first detected using human hyperimmune serum from Papua New Guinea (McColl et al., 1994) and also with antibodies that inhibit P. falciparum in vitro Molecular Microbiology (2002) 43(6), 1401–1411 Kerry E. Mills, J. Andrew Pearce, Brendan S. Crabb and Alan F. Cowman* The Walter and Eliza Hall Institute of Medical Research, Melbourne 3050, Australia. Summary Merozoite surface protein 3 (MSP3), an important vaccine candidate, is a soluble polymorphic antigen associated with the surface of Plasmodium falci- parum merozoites. The MSP3 sequence contains three blocks of heptad repeats that are consistent with the formation of an intramolecular coiled-coil. MSP3 also contains a glutamic acid-rich region and a putative leucine zipper sequence at the C-terminus. We have disrupted the msp3 gene by homologous recombination, resulting in the expression of a trun- cated form of MSP3 that lacks the putative leucine zipper sequence but retains the glutamic acid-rich region and the heptad repeats. Here, we show that truncated MSP3, lacking the putative leucine zipper region, does not localize to the parasitophorous vacuole or interact with the merozoite surface. Fur- thermore, the acidic–basic repeat antigen (ABRA), which is present on the merozoite surface, also was not localized to the merozoite surface in parasites expressing the truncated form of MSP3. The P. falci- parum merozoites lacking MSP3 and ABRA on the surface show reduced invasion into erythrocytes. These results suggest that MSP3 is not absolutely essential for blood stage growth and that the putative leucine zipper region is required for the trafficking of both MSP3 and ABRA to the parasitophorous vacuole. Introduction The protozoan parasite Plasmodium falciparum causes the most severe form of malaria in humans. The majority of victims are young children, with adult populations Accepted 7 December, 2001. *For correspondence. E-mail [email protected]; Tel. (+61) 3 9345, 2555; Fax (+61) 3 934 70852.
Transcript of Mills mol micro 2002

© 2002 Blackwell Science Ltd
Truncation of merozoite surface protein 3 disrupts its trafficking and that of acidic–basic repeat protein to the surface of Plasmodium falciparum merozoites
nevertheless suffering significant morbidity. Drug resist-ance continues to increase and as yet there is no effec-tive vaccine against malaria. A large number of potentialvaccine candidates have been identified; however, little isknown of their functions (Barnwell and Galinski, 1998).Many of the putative vaccine antigens are located on themerozoite surface. As a major interface between host and pathogen, the merozoite surface is an obvious targetfor the development of a vaccine. Proteins comprising the merozoite surface are exposed to the host immunesystem before and during invasion into the erythrocyte,albeit for a short time.
Thus far, several merozoite surface proteins have been identified, and included in this group are the mero-zoite surface proteins 1–6 and 8 (Smythe et al., 1988;Blackman et al., 1990; McColl et al., 1994; Oeuvray et al., 1994a; Marshall et al., 1997; Marshall et al., 1998;Black et al., 2001; Trucco et al., 2001). MSP1 is an important vaccine candidate, and there is evidence to suggest that it can bind to the surface of the eryth-rocyte (Kaslow et al., 1994), whereas MSP2–6 and 8 are less characterized. Indeed, there is very little datadefining the roles of any of these antigens in parasite invasion.
Merozoite surface protein 3 (MSP3) is a polymorphicantigen, which has a number of structural domains(Fig. 1A). These domains include three blocks of fourheptad repeats of the type AXXAXXX, a hydrophilic regionand a putative leucine zipper sequence at the C-terminus(Fig. 1A) (McColl et al., 1994). The solution structure ofthe heptad repeats has been solved, and each blockappears to form an a-helix, which may interact to form a somewhat unstable coiled-coil (Mulhern et al., 1995).Despite some sequence diversity of the msp3 genebetween P. falciparum isolates, there is significant con-servation (McColl et al., 1994; Escalante et al., 1998).Indeed, the alanine residues within the heptad repeatregions and the C-terminal half of the protein, whichincludes the putative leucine zipper region, are stronglyconserved (McColl and Anders, 1997).
Current evidence supports MSP3 as a vaccine candi-date. MSP3 was first detected using human hyperimmuneserum from Papua New Guinea (McColl et al., 1994) andalso with antibodies that inhibit P. falciparum in vitro
Molecular Microbiology (2002) 43(6), 1401–1411
Kerry E. Mills, J. Andrew Pearce, Brendan S. Crabband Alan F. Cowman*The Walter and Eliza Hall Institute of Medical Research,Melbourne 3050, Australia.
Summary
Merozoite surface protein 3 (MSP3), an importantvaccine candidate, is a soluble polymorphic antigenassociated with the surface of Plasmodium falci-parum merozoites. The MSP3 sequence containsthree blocks of heptad repeats that are consistentwith the formation of an intramolecular coiled-coil.MSP3 also contains a glutamic acid-rich region and aputative leucine zipper sequence at the C-terminus.We have disrupted the msp3 gene by homologousrecombination, resulting in the expression of a trun-cated form of MSP3 that lacks the putative leucinezipper sequence but retains the glutamic acid-richregion and the heptad repeats. Here, we show thattruncated MSP3, lacking the putative leucine zipperregion, does not localize to the parasitophorousvacuole or interact with the merozoite surface. Fur-thermore, the acidic–basic repeat antigen (ABRA),which is present on the merozoite surface, also wasnot localized to the merozoite surface in parasitesexpressing the truncated form of MSP3. The P. falci-parum merozoites lacking MSP3 and ABRA on thesurface show reduced invasion into erythrocytes.These results suggest that MSP3 is not absolutelyessential for blood stage growth and that the putativeleucine zipper region is required for the trafficking of both MSP3 and ABRA to the parasitophorousvacuole.
Introduction
The protozoan parasite Plasmodium falciparum causesthe most severe form of malaria in humans. The majorityof victims are young children, with adult populations
Accepted 7 December, 2001. *For correspondence. [email protected]; Tel. (+61) 3 9345, 2555; Fax (+61) 3 93470852.

© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
1402 K. E. Mills, J. A. Pearce, B. S. Crabb and A. F. Cowman

Trafficking of MSP3 and ABRA to the parasitophorous vacuole 1403
growth by co-operation with blood monocytes in an anti-body-dependent cellular inhibition assay (ADCI) (Oeuvrayet al., 1994a). The ADCI assay selects for moleculescapable of producing a protective immune response. Thisstrengthens the case for MSP3 as a vaccine candidate(Oeuvray et al., 1994a). Recently, it has been shown inan immunocompromised mouse model infected with P. falciparum that human antibodies purified on peptidesfrom MSP3 are able to suppress P. falciparum growth inthe presence of human monocytes (Badell et al., 2000).
Several MSP3 homologues have been found in otherPlasmodium species. Plasmodium vivax express aprotein similar in structure to PfMSP3, which containsblocks of heptad repeats predicted to form a coiled-coiland is also polymorphic (Galinski et al., 1999). Also, thePlasmodium knowlesi 140 kDa protein has homology withPfMSP3 (Hudson et al., 1983). Interestingly, MSP6 showssome structural similarity and homology with MSP3.MSP6 has been shown to form part of the MSP1 complexon the surface of the merozoite of P. falciparum (Truccoet al., 2001).
Given that MSP3 is conserved over diverse Plasmod-ium species, is recognized by hyperimmune sera from amalaria-endemic region and antibodies to MSP3 appearto be capable of inhibiting in vitro growth by ADCI assays,we sought to elucidate further the role of MSP3 in the P.falciparum merozoite. In this report, we describe the firsttargeted disruption of a gene expressing a protein locatedon the merozoite surface of P. falciparum. Targeting ofmsp3 resulted in truncation of 37 amino acids from the C-terminus of the MSP3 protein and removal of the putativeleucine zipper sequence. This truncation disrupted thetrafficking of this protein to the parasitophorous vacuoleand interaction with the merozoite surface. Furthermore,the acidic–basic repeat antigen (ABRA), another mero-zoite surface protein, was also found to be absent fromthe merozoite surface. Interestingly, the loss of these proteins from the merozoite led to a reduction in invasionefficiency. These results have important implications for
the function of MSP3 in merozoite invasion and its poten-tial as a vaccine candidate.
Results
Targeted disruption of the msp3 gene
In order to study the role of MSP3 in merozoite invasioninto human erythrocytes, we constructed two P. falci-parum transgenic lines. In the first parasite line, the singlewild-type copy of msp3 was disrupted, whereas in thesecond, the msp3 gene was replaced intact. This wasachieved by introducing either the transfection vectorpHDm3 or pH-m3/3¢, respectively, into D10 parasites(Fig. 1B). Transfected parasites were selected onpyrimethamine, cultured on drug alternating with cycleson and off drug to facilitate loss of episomal plasmid andselection for parasites with integrated copies (Crabb andCowman, 1996; Wu et al., 1996; Crabb et al., 1997).
To determine whether the transfection plasmids hadintegrated into the parasite genome, chromosomes of theD10 parental line and the transfected lines were analysedby pulsed-field gel electrophoresis (PFGE) (Fig. 1C).When probed with the msp3 gene, D10 chromosomesexhibit a single band at a position corresponding to chro-mosome 10. However, after three rounds of cycling, chro-mosomes from the transfected parasites D10Dm3 shownot only the endogenous gene, but also further bandsconsistent with migration of unintegrated plasmids underthese electrophoresis conditions (Crabb and Cowman,1996). In contrast, after six rounds of cycling, chromo-somes from D10Dm3-transfected parasites show only asingle band corresponding to chromosome 10, indicatingcomplete integration of pHDm3 into chromosome 10(Fig. 1B). To prove that this single band did not arise asa result of complete loss of plasmid, the same chromo-somes were subjected to hybridization with the plasmidbackbone, pGEM (Fig. 1C). As expected, no hybridizationoccurred with D10 chromosomes; however, a pattern
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
Fig. 1. Structure of MSP3 and disruption of the msp3 gene.A. Structure of MSP3 showing the putative signal sequence (SS), the three blocks of four heptad repeats (H1, H2, H3), the acidic region andthe putative leucine zipper domain.B. Disruption of the msp3 gene in D10 parasites. The plasmids pHDm3 and pHm-3/3¢ are shown and were transfected into D10 andtransfected parasites selected with pyrimethamine. Cycling of transfected parasites on and off drug allowed selection of those with integratedcopies of the plasmid into the msp3 gene as shown. The msp3 gene is divided into three shaded regions to show the regions used in pHDm3and pHm3/3¢. The plasmid pHDm3 integrated via a single recombination event to produce the parasite line D10Dm3 that contained a truncatedmsp3 gene. The plasmid pHm-3/3¢ integrated via a single recombination event to produce the parasite line D10-3/3¢, in which the msp3 genehad been reconstituted. S represents SwaI.C. Pulsed-field gel electrophoresis analysis of chromosomes from D10 and transfected parasites to determine whether integration of thetransfection plasmids had occurred. The first panel shows chromosomes from parasites after transfection with pHDm3 and three and sixcycles on and off drug probed with the msp3 gene (first panel) and the plasmid backbone pGEM (second panel). The third blot showschromosomes from parasites after transfection with pHm-3/3¢ and one and two cycles on and off drug probed with the msp3 gene (third panel)and the plasmid backbone pGEM (fourth panel). Chromosome 10 is indicated, and a single band migrating at the position of chromosome 10suggests integration of the transfection plasmid.D. Southern hybridization of genomic DNA from D10, D10Dm3 after cycle six (first panel) as well as D10, D10m3/3¢ after one and two cycleson and off drug (second panel), indicating that the transfection plasmids had integrated into the msp3 gene in both D10Dm3 and D10m3/3¢.Sizes of the hybridizing bands are shown in kb at the side.
consistent with unintegrated plasmid occurred with chro-mosomes from D10Dm3 parasites after three rounds ofcycling. A single band corresponding to chromosome 10was seen in chromosomes from transfected parasitesafter six rounds of cycling. These results indicate that,after six rounds of cycling, complete integration of pHDm3into chromosome 10 had occurred in the D10Dm3-transfected parasites.
The plasmid pH-m3/3¢ was also transfected into D10,and parasites were cycled through two rounds of off drugselection to obtain parasites with integrated copies of theplasmid. Chromosomes from D10 and the transfected lineD10-M3/3¢ were probed with the msp3 gene. After twocycles of selection, all the pH-m3/3¢ plasmid was appar-ently integrated into chromosome 10. This was confirmedby hybridization of a pGEM probe (Fig. 1C).
In order to determine whether the pHDm3 and pH-m3/3¢plasmids had integrated into the msp3 gene on chromo-some 10 of the D10 transfectants, Southern blot analysesof SwaI restriction-digested genomic DNA were per-formed (Fig. 1D). Hybridization with an msp3 probe iden-tified a single band of ª 2.3 kb in D10 corresponding tothe endogenous gene. However, in D10Dm3-transfectedparasites, two hybridizing bands of ª 0.8 kb and 8 kb wereobtained, indicating that the plasmid had integrated intothe msp3 gene. Further restriction mapping of the inte-gration event confirmed that the pHDm3 plasmid had inte-grated via a single homologous cross-over to generatethe structure shown in Fig. 1B. This resulted in truncationof the msp3 gene in D10Dm3 by 111 bp. Expression of theDMSP3 protein from these parasites would lack 37 aminoacids, which encompasses the putative leucine zippersequence (Fig. 1A). Analysis of two independent clonalisolates derived from D10Dm3 parasites (designatedD10Dm3/1 and D10Dm3/2) showed an identical restrictionpattern. Similar analysis of the D10-m3/3¢ parasitesshowed that the pH-m3/3¢ plasmid had also integrated bya single homologous cross-over to replace the intactmsp3 gene (Fig. 1C). This recombination event places the3¢ termination sequence hsp86 3¢ at the 3¢ end of themsp3 gene, resulting in reconstitution of the full geneincluding the region encoding the putative leucine zipper(Fig. 1B). This parasite line was designed to express the full-length MSP3 and thus served as a control for subsequent experiments.
Disruption of msp3 leads to the expression of truncated MSP3
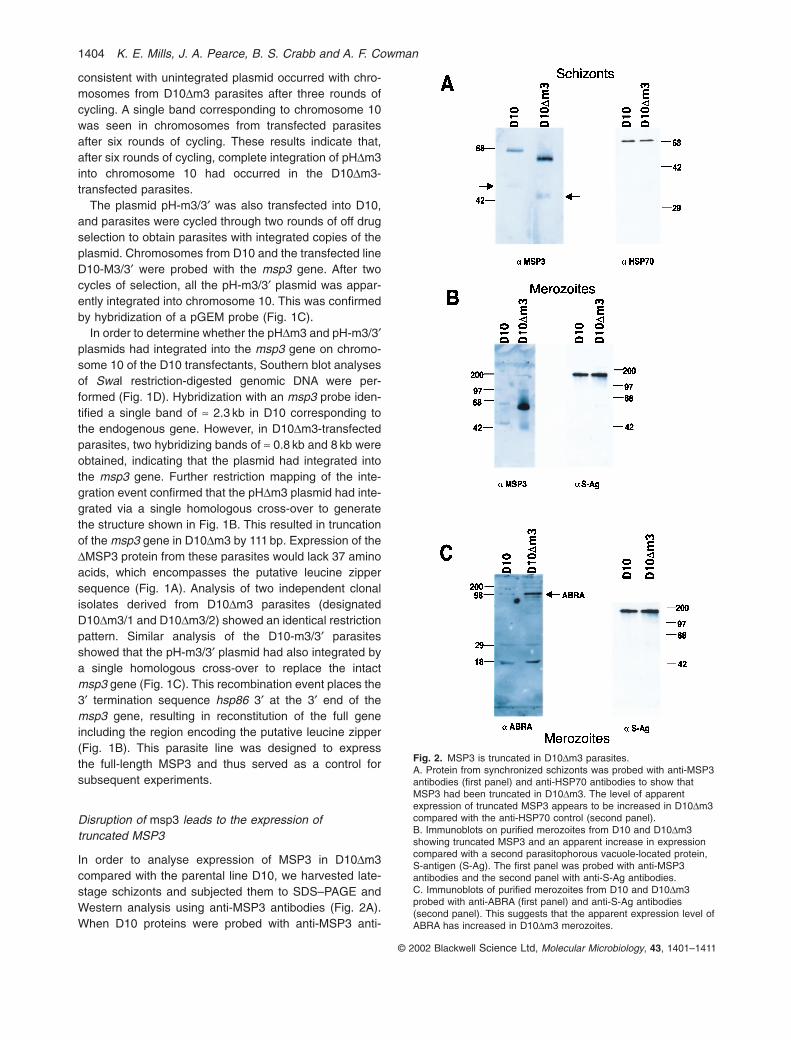
In order to analyse expression of MSP3 in D10Dm3 compared with the parental line D10, we harvested late-stage schizonts and subjected them to SDS–PAGE andWestern analysis using anti-MSP3 antibodies (Fig. 2A).When D10 proteins were probed with anti-MSP3 anti-
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
1404 K. E. Mills, J. A. Pearce, B. S. Crabb and A. F. Cowman
Fig. 2. MSP3 is truncated in D10Dm3 parasites.A. Protein from synchronized schizonts was probed with anti-MSP3antibodies (first panel) and anti-HSP70 antibodies to show thatMSP3 had been truncated in D10Dm3. The level of apparentexpression of truncated MSP3 appears to be increased in D10Dm3compared with the anti-HSP70 control (second panel).B. Immunoblots on purified merozoites from D10 and D10Dm3showing truncated MSP3 and an apparent increase in expressioncompared with a second parasitophorous vacuole-located protein,S-antigen (S-Ag). The first panel was probed with anti-MSP3antibodies and the second panel with anti-S-Ag antibodies.C. Immunoblots of purified merozoites from D10 and D10Dm3probed with anti-ABRA (first panel) and anti-S-Ag antibodies(second panel). This suggests that the apparent expression level ofABRA has increased in D10Dm3 merozoites.

Trafficking of MSP3 and ABRA to the parasitophorous vacuole 1405
bodies, the expected 62 kDa and 48 kDa protein bandswere detected (McColl et al., 1994). In contrast, Westernanalysis of the D10Dm3 parasites containing the dis-rupted msp3 gene revealed two MSP3 bands of ª 58 kDaand 43 kDa. The degree of reduction in the size of boththese bands compared with those seen in the D10parental line is consistent with the expected size of proteins resulting from the truncation of the msp3 gene.Interestingly, there appears to be an increase in theexpression of the truncated MSP3 in D10DM3 parasites,when compared with that observed for Hsp70 (Fig. 2A).
In order to determine whether truncated MSP3 inD10Dm3 parasites is associated with free merozoites,further Western analysis was performed. Merozoites puri-fied from the D10 parental and D10Dm3 parasites weresubjected to SDS–PAGE and Western blot analysis usinganti-MSP3 antibodies. The 62 kDa and 48 kDa bandswere obtained in the D10 merozoites; however, a moreintensely truncated 58 kDa was seen in D10Dm3(Fig. 2B). This is in contrast to the results observed with S-antigen, a protein that is also secreted into the parasitophorous vacuole of the schizont stage (Anderset al., 1983). The S-antigen protein was detected in bothD10 and D10Dm3 merozoites at approximately equallevels (Fig. 2B). We also analysed the expression of theacidic–basic repeat antigen (ABRA), which is alsoexpressed in the parasitophorous vacuole, and sur-prisingly found differences in D10Dm3 compared withD10. ABRA is a protease that is expressed in the para-sitophorous vacuole and has also been shown to be associated with the merozoite surface (Nwagwu et al.,1992; Kushwaha et al., 2000). Purified merozoites were analysed by SDS–PAGE and Western blots usingantibodies to the ABRA protein (Kushwaha et al., 2000).Interestingly, ABRA also displayed an apparent overex-pression in D10Dm3 compared with the parental D10 par-asites (Fig. 2C). ABRA appears to be present in D10Dm3merozoites at levels at least three or four times thatobserved with D10 relative to the S-antigen loadingcontrol. These results suggest that truncation of the MSP3protein is linked to an accumulation of ABRA withinD10Dm3 merozoites.
MSP3 and ABRA in D10Dm3 parasites do not localizeto the merozoite surface
Immunofluorescence microscopy was performed on D10and D10Dm3 schizont and merozoite stages in order todetermine the subcellular localization of the truncatedMSP3 protein and ABRA in D10Dm3 compared with theD10 parental parasites. Late schizonts and free mero-zoites were incubated with anti-MSP1, -MSP3 and -ABRAantibodies in immunofluorescence experiments (Fig. 3).In late D10 schizonts, MSP3 and ABRA had a localization
consistent with the parasitophorous vacuole and themerozoite surface, as described previously (Oeuvrayet al., 1994a; Kushwaha et al., 2000). This was similar to the subcellular localization of MSP1 (Holder andFreeman, 1984), which is present on the surface of thedeveloping merozoite as a glycosylphosphatidyl inositol-anchored protein (Holder et al., 1985). This subcellularlocalization was confirmed in free merozoites, in whichboth MSP3 and ABRA co-localized with MSP1 (Fig. 3,top). Localization of MSP3 and ABRA in D10Dm3 sch-izonts showed a very different pattern of localization compared with that observed in D10. Neither truncatedMSP3 nor ABRA fully co-localized with MSP1, suggestingthat these proteins were not able to traffic to the para-sitophorous vacuole in D10Dm3 (Fig. 3, lower). In orderto determine whether truncated MSP3 and ABRA werepresent within free merozoites, co-localization studieswere carried out with MSP1. A significant concentration ofboth MSP3 and ABRA proteins was observed in the formof small dots within the merozoites. These results suggestthat MSP3 and ABRA accumulate within the merozoite ofD10Dm3 during development and that they are unable tobe trafficked to the parasitophorous vacuole.
MSP3 localization in D10Dm3 free merozoites is notassociated with the endoplasmic reticulum
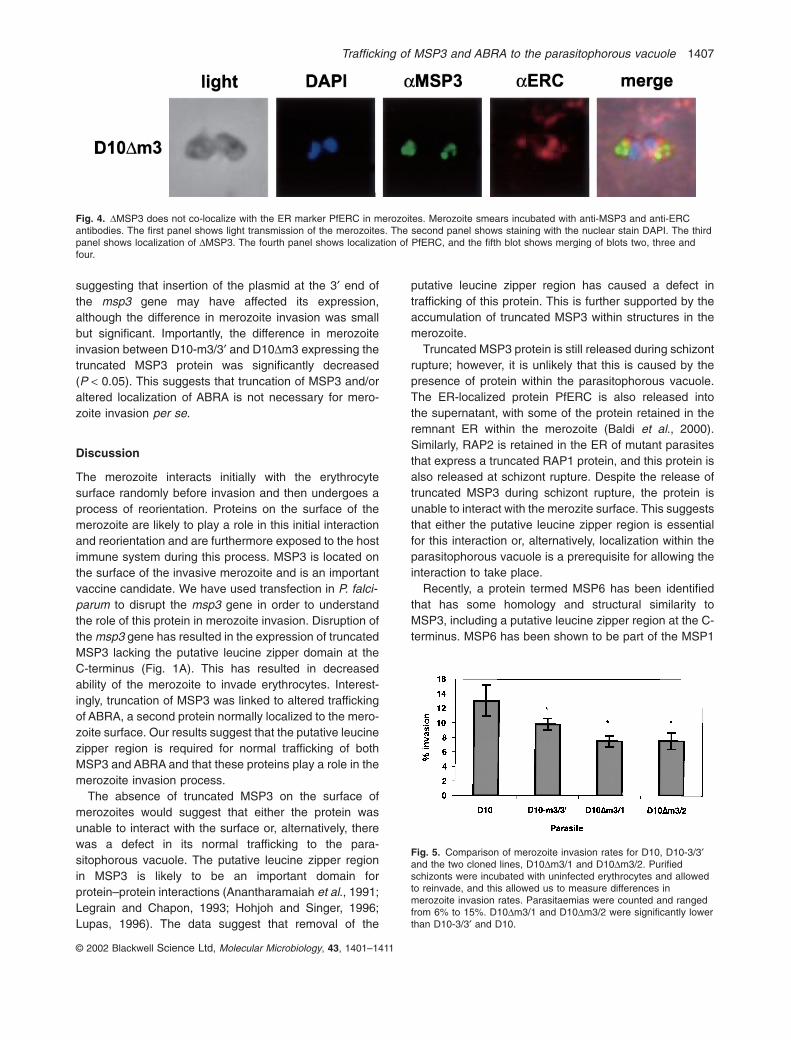
It is possible that the aberrant localization of MSP3 inD10Dm3 merozoites resulted from retention in the endo-plasmic reticulum (ER) as has been described for RAP2in parasites that express truncated RAP1 (Baldi et al.,2000). To determine whether this was the case in D10DM3merozoites, antisera were raised to the P. falciparum ERCprotein (La Greca et al., 1997). No co-localization ofMSP3 and ERC was observed with either D10 (data notshown) or D10DM3 parasites (Fig. 4). MSP3 appears tobe contained in vesicular-like structures or large aggre-gates at the apical end of the merozoite, and this was distinct from the ER and nucleus as shown by anti-ERCantibodies and DAPI staining.
Lack of MSP3 and ABRA on the merozoite surface hasan effect on merozoite invasion in vitro
To determine whether truncation of MSP3 had any effecton the ability of merozoites to invade human erythrocytes,D10, D10-m3/3¢ and the two D10Dm3 clones, D10DM3/1and D10DM3/2, were compared in merozoite invasionassays. Percoll-purified trophozoites were incubatedovernight with fresh human erythrocytes, and the numberof ring-stage parasites was counted. The invasion effi-ciency was calculated and expressed as percentage parasitaemia (Fig. 5). The parental D10 parasites invadedat a higher rate than the transfection control D10-m3/3¢,
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411

© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
1406 K. E. Mills, J. A. Pearce, B. S. Crabb and A. F. Cowman
Fig. 3. Subcellular localization of MSP3 andABRA shows that deletion of the putativeleucine zipper region is linked to incorrectlocalization of DMSP3 and ABRA in D10Dm3.Smears of late schizonts of D10 and D10Dm3were incubated with anti-MSP1 (located onthe surface of the merozoite) and anti-MSP3or anti-ABRA antibodies. Anti-MSP1 mouseantibodies were labelled with rhodamine-conjugated anti-mouse antibodies (red). Anti-MSP3 and anti-ABRA rabbit antibodies werelabelled with anti-rabbit antibodies conjugatedwith FITC (green). Co-localization is shown inthe row and indicated by yellow. The firstpanel is D10 smears labelled with anti-MSP1(first column) and anti-MSP3 or anti-ABRA(second column). The second panel isD10Dm3 labelled with anti-MSP1 (firstcolumn, red) and anti-MSP3 or anti-ABRAantibodies (green). D10Dm3 merozoites in thesecond and fourth panels have beenmagnified by a factor of two compared withthe other blots to enable increased detail tobe visualized.
Trafficking of MSP3 and ABRA to the parasitophorous vacuole 1407
suggesting that insertion of the plasmid at the 3¢ end ofthe msp3 gene may have affected its expression,although the difference in merozoite invasion was smallbut significant. Importantly, the difference in merozoiteinvasion between D10-m3/3¢ and D10Dm3 expressing thetruncated MSP3 protein was significantly decreased(P < 0.05). This suggests that truncation of MSP3 and/oraltered localization of ABRA is not necessary for mero-zoite invasion per se.
Discussion
The merozoite interacts initially with the erythrocytesurface randomly before invasion and then undergoes aprocess of reorientation. Proteins on the surface of themerozoite are likely to play a role in this initial interactionand reorientation and are furthermore exposed to the hostimmune system during this process. MSP3 is located onthe surface of the invasive merozoite and is an importantvaccine candidate. We have used transfection in P. falci-parum to disrupt the msp3 gene in order to understandthe role of this protein in merozoite invasion. Disruption ofthe msp3 gene has resulted in the expression of truncatedMSP3 lacking the putative leucine zipper domain at theC-terminus (Fig. 1A). This has resulted in decreasedability of the merozoite to invade erythrocytes. Interest-ingly, truncation of MSP3 was linked to altered traffickingof ABRA, a second protein normally localized to the mero-zoite surface. Our results suggest that the putative leucinezipper region is required for normal trafficking of bothMSP3 and ABRA and that these proteins play a role in themerozoite invasion process.
The absence of truncated MSP3 on the surface ofmerozoites would suggest that either the protein wasunable to interact with the surface or, alternatively, therewas a defect in its normal trafficking to the para-sitophorous vacuole. The putative leucine zipper region in MSP3 is likely to be an important domain forprotein–protein interactions (Anantharamaiah et al., 1991;Legrain and Chapon, 1993; Hohjoh and Singer, 1996;Lupas, 1996). The data suggest that removal of the
putative leucine zipper region has caused a defect in trafficking of this protein. This is further supported by theaccumulation of truncated MSP3 within structures in themerozoite.
Truncated MSP3 protein is still released during schizontrupture; however, it is unlikely that this is caused by thepresence of protein within the parasitophorous vacuole.The ER-localized protein PfERC is also released into the supernatant, with some of the protein retained in theremnant ER within the merozoite (Baldi et al., 2000). Similarly, RAP2 is retained in the ER of mutant parasitesthat express a truncated RAP1 protein, and this protein isalso released at schizont rupture. Despite the release oftruncated MSP3 during schizont rupture, the protein isunable to interact with the merozite surface. This suggeststhat either the putative leucine zipper region is essentialfor this interaction or, alternatively, localization within theparasitophorous vacuole is a prerequisite for allowing theinteraction to take place.
Recently, a protein termed MSP6 has been identifiedthat has some homology and structural similarity toMSP3, including a putative leucine zipper region at the C-terminus. MSP6 has been shown to be part of the MSP1
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
Fig. 4. DMSP3 does not co-localize with the ER marker PfERC in merozoites. Merozoite smears incubated with anti-MSP3 and anti-ERCantibodies. The first panel shows light transmission of the merozoites. The second panel shows staining with the nuclear stain DAPI. The thirdpanel shows localization of DMSP3. The fourth panel shows localization of PfERC, and the fifth blot shows merging of blots two, three andfour.
Fig. 5. Comparison of merozoite invasion rates for D10, D10-3/3¢and the two cloned lines, D10Dm3/1 and D10Dm3/2. Purifiedschizonts were incubated with uninfected erythrocytes and allowedto reinvade, and this allowed us to measure differences inmerozoite invasion rates. Parasitaemias were counted and rangedfrom 6% to 15%. D10Dm3/1 and D10Dm3/2 were significantly lowerthan D10-3/3¢ and D10.

complex on the merozoite surface. It is possible thatMSP3 interacts with a complex in a manner similar toMSP1–MSP6 and that interaction of MSP3 with other proteins may be required for correct targeting to the par-asitophorous vacuole. It has already been shown thattruncation of RAP1 disrupts formation of the RAP1–RAP2complex, resulting in retention of RAP2 in the ER ratherthan correct trafficking to the rhoptries (Baldi et al., 2000).Similarly, in Toxoplasma gondii, a transmembranemicronemal protein MIC6 has been shown to function asan escort for the accurate targeting of two soluble pro-teins, MIC1 and MIC4, to the rhoptries (Reiss et al., 2001).It is possible that MSP3 and ABRA interact with eachother and that deletion of the putative leucine zipperregion of MSP3 disrupts interaction with an escort protein,resulting in mislocalization of both MSP3 and ABRA. A number of genes have recently been identified in the P. falciparum genome that have structural similarities toMSP1, and the proteins encoded by these are located onthe merozoite surface (Black et al., 2001). These proteinsare candidates to act as binding partners for MSP3 andABRA in a complex on the merozoite surface.
The truncation of MSP3 and/or alterations in ABRAlocalization result in reduced invasion of merozoites inD10Dm3. The D10-3/3¢ control parasites also showed asmall decrease in merozoite invasion compared with theparental D10 line. One possibility to explain this result isthat there is a slight decrease in the level of expressionof MSP3 due to insertion of an altered 3¢ untranslatedregion. However, the important point is that the D10Dm3is significantly decreased in merozoite invasion comparedwith both the parental line D10 and D10-3/3¢. This is thefirst example to be described in which alteration of aprotein such as MSP3 results in reduction in invasion efficiency. For instance, truncation of the RAP-1 protein,which resulted in the loss of RAP2 from merozoites, hadno effect on invasion efficiency (Baldi et al., 2000), andtruncation of EBA-175, despite resulting in a switch ininvasion phenotype (Reed et al., 2000), similarly had nosignificant effect on invasion (Kaneko et al., 2000; Reedet al., 2000). Taken together, these results give weight tothe argument that MSP3 or an MSP3–ABRA complexplays a role in invasion. Furthermore, these results areconsistent with the demonstration that antibodies toMSP3 peptides in co-operation with monocytes can inhibitP. falciparum growth in an immunocompromised mousemodel (Badell et al., 2000) and the ability of anti-MSP3antibodies to inhibit merozoite invasion in an ADCI assay(Oeuvray et al., 1994a,b).
The requirement for MSP3 and/or ABRA for efficientmerozoite invasion of human erythrocytes supports these proteins as vaccine candidates. However, lack ofexpression of these on the surface of the merozoite does not ablate invasion. This suggests that they are
not absolutely essential and that there is some redun-dancy in the invasion process. Indeed, several differentpathways of invasion have been identified in P. falciparum(Orlandi et al., 1992; Dolan et al., 1994; Okoyeh et al.,1999). Furthermore, the same parasite strain is able to‘switch’ invasion pathways when a particular ligand isunavailable (Dolan et al., 1990; Reed et al., 2000). Given this redundancy, it is important that this be consid-ered in evaluating any P. falciparum protein as a vaccinecandidate.
These results suggest that MSP3 may be part of a complex on the surface of the merozoite and that interactions with other proteins are important for correcttargeting of both MSP3 and ABRA to the parasitophorousvacuole. Interestingly, MSP3 does not appear to beabsolutely essential for merozoite invasion, and it is very likely that it will be possible to construct P. falciparummutant parasites that do not express MSP3. Construc-tion of MSP3 null P. falciparum parasites would be useful to evaluate the role of antibodies to MSP3 in ADCIassays (Oeuvray et al., 1994a,b) and in P. falciparuminfections of immunocompromised mice (Badell et al.,2000).
Experimental procedures
Parasites and transfection
Plasmodium falciparum was cultivated (Trager and Jensen,1976) and synchronized as published previously (Lambrosand Vanderberg, 1979). Parasites of the cloned line D10 weretransfected as described previously (Crabb and Cowman,1996; Wu et al., 1996; Crabb et al., 1997) with 80 mg ofplasmid purified using Qiagen maxiprep columns. Trans-fected parasites were selected on 0.1mM pyrimethamine andsubjected to successive cycles of culture on and off drug forperiods of 4 weeks. Parasites containing integrated plasmidswere cloned by single-cell cloning.
pHDm3 and pH-m3/3¢ were created for transfection by thecloning of an 800 bp msp3 insert into the pHC1 transfectionvector (Crabb et al., 1997). This fragment was amplified fromD10 genomic DNA with the following oligonucleotides: pHDm3forward (5¢-GGTAACTCGAGGGAATTTGGAGAAGGTTTTAGTGCAG-3¢); pHDm3 reverse (5¢-TTTCCTCGAGTTCTTTTTTACGTTTTTCTCATTATTATTC-3¢); pH-m3/3¢ forward (5¢-GCTGCTCGAGCTGCTTTAAAGGC-3¢); pH-m3/3¢ reverse(5¢-TTAACTCGAGTTAATGATTTTTAAAATATTTGGATA-3¢).This amplification resulted in the introduction of XhoI restric-tion sites on both ends of each fragment (bold). These siteswere used for cloning into the pHC1 vector.
Nucleic acids and pulsed-field gel electrophoresis
Genomic DNA was extracted from late-stage parasites asdescribed previously (Corcoran et al., 1986). Analysis ofgenomic DNA was carried out using standard Southern blothybridization techniques. Pulsed-field gel electrophoresis
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
1408 K. E. Mills, J. A. Pearce, B. S. Crabb and A. F. Cowman

Trafficking of MSP3 and ABRA to the parasitophorous vacuole 1409
(Chu et al., 1986) was used for chromosome separation asdescribed previously (Corcoran et al., 1986).
Antisera and immunoblot assays
MSP3 antisera were raised in the following manner: full-length msp3 (excluding putative signal sequence) was ampli-fied from D10 genomic DNA using the oligonucleotides5¢-GCGGATCCAAAGAAATTGTAAAAAAATATAATC-3¢ and5¢-CGAATTCAATGATTTTTAAAATATTTGGATAATTC-3¢,introducing BamHI and EcoRI restriction sites respectively.The resulting fragment was cloned into pPro-EX-HTb (LifeTechnologies). Expression of this plasmid in the Escherichiacoli strain BL21 produced a protein in fusion with hexa-histidine. MSP3-hexa-his was purified on nickel–NTA beadsand by reversed-phase chromatography. Purified protein wasinoculated into NZ White rabbits. Serum generated was usedthroughout this study.
ERC antisera were raised in the following manner: ERCprotein (La Greca et al., 1997) was obtained from L. Tilley(La Trobe University, Melbourne, Australia) and inoculatedinto eight mice. Serum from each mouse was tested forcorrect localization by immunofluorescence microscopybefore being used throughout this study.
Proteins from late-stage parasites were separated onSDS–12% PAGE and transferred to nitrocellulose mem-branes as described previously (Cowman et al., 1991).MSP3, ABRA, S-antigen and HSP70 antisera were diluted1:500, 1:1000, 1:1000 and 1:5000, respectively, in 5% (w/v) skimmed milk in phosphate-buffered saline (PBS). Secondary antibody was sheep anti-rabbit Ig horseradishperoxidase (HRP)-conjugated antibody (Silenus). Antigen–antibody complexes were detected by ECL (Amersham) andautoradiography.
Immunofluorescence
Thin blood smears fixed at room temperature in methanolwere used for immunofluorescence. MSP1 was detectedusing the monoclonal anti-MSP1-19 antibody (Cooper et al.,1992). MSP3, ABRA and ERC were detected using the anti-bodies described above. Antibodies to MSP1, MSP3, ABRAand ERC were diluted 1:2000, 1:100, 1:10 and 1:75, respec-tively, in 5% (w/v) skimmed milk in PBS. Anti-MSP1 and -ERCantibodies were detected using goat anti-mouse rhodamine-labelled antibodies (Chemicon). Anti-MSP3 and -ABRA anti-bodies were detected using sheep anti-rabbit fluoresceinisothiocyanate (FITC)-labelled antibodies (Silenus). All para-sites were also labelled with 4¢,6-diamidino-2-phenylindole(DAPI) DNA stain (Molecular Probes).
Invasion assays
Merozoite invasion assays were conducted essentially asdescribed previously (Dolan et al., 1994). Briefly, 8 ¥ 105
synchronized, Percoll-purified trophozoites were mixed with4 ¥ 107 erythrocytes in a total volume of 200 ml with 50/50culture medium in a 96-well, flat-bottomed microtitre plate.Assays were performed in triplicate. After 28 h incubation,
wells were resuspended, and thin blood smears were madeand stained with Giemsa. Approximately 1000 cells werecounted per slide, and infected cells were expressed as a percentage of total cells. This allowed quantification ofmerozoite invasion of human erythrocytes.
Acknowledgements
We would like to thank Sonia Caruana for technical assis-tance, Leanne Tilley for the provision of ERC protein, Virander Chauhan for provision of the ABRA antibody, andRoss Coppel for the S-antigen antibody. We would like to acknowledge the Australian Red Cross Blood Bank (Melbourne, Australia) for supply of human erythrocytes and serum. K.E.M. is the recipient of a Melbourne ResearchScholarship. A.F.C. and B.S.C. are International ResearchFellows from the Howard Hughes Medical Institute. This work was funded by the National Health and MedicalResearch Council of Australia.
References
Anantharamaiah, G.M., Brouillette, C.G., Engler, J.A., De Loof, H., Venkatachalapathi, Y.V., Boogaerts, J., andSegrest, J.P. (1991) Role of amphipathic helixes in HDLstructure/function. Adv Exp Med Biol 285: 131–140.
Anders, R.F., Brown, G.V., and Edwards, A. (1983) Charac-terization of an S antigen synthesized by several isolatesof Plasmodium falciparum. Proc Natl Acad Sci USA 80:6652–6656.
Badell, E., Oeuvray, C., Moreno, A., Soe, S., van Rooijen, N.,Bouzidi, A., and Druilhe, P. (2000) Human malaria inimmunocompromised mice: an in vivo model to studydefense mechanisms against Plasmodium falciparum. J Exp Med 192: 1653–1660.
Baldi, D.L., Andrews, K.T., Waller, R.S., Roos, D., Crabb,B.S., and Cowman, A.F. (2000) RAP1 controls rhoptry targeting of RAP2 in the malaria parasite Plasmodium falciparum. EMBO J 19: 1–9.
Barnwell, J.W., and Galinski, M.R. (1998) Invasion of Vertebrate Cells: Erythrocytes. Washington, DC: AmericanSociety for Microbiology Press.
Black, C.G., Wu, T., Wang, L., Hibbs, A.R., and Coppel, R.L.(2001) Merozoite surface protein 8 of Plasmodium falci-parum contains two epidermal growth factor-like domains.Mol Biochem Parasitol 114: 217–226.
Blackman, M.J., Heidrich, H.-G., Donachie, S., McBride, J.S.,and Holder, A.A. (1990) A single fragment of a malariamerozoite surface protein remains on the parasite duringred cell invasion and is the target of invasion-inhibiting antibodies. J Exp Med 172: 379–382.
Chu, G., Vollrath, D., and Davis, R. (1986) Separation of largeDNA molecules by contour clamped homogeneous electricfields. Science 234: 1582–1585.
Cooper, J.A., Cooper, L.T., and Saul, A.J. (1992) Mapping ofthe region predominantly recognized by antibodies to thePlasmodium falciparum merozoite surface antigen MSA 1.Mol Biochem Parasitol 51: 301–312.
Corcoran, L.M., Forsyth, K.P., Bianco, A.E., Brown, G.V., andKemp, D.J. (1986) Chromosome size polymorphisms in
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411

Plasmodium falciparum can involve deletions and are frequent in natural parasite populations. Cell 44: 87–95.
Cowman, A.F., Karcz, S., Galatis, D., and Culvenor, J.G.(1991) A P-glycoprotein homologue of Plasmodium falci-parum is localized on the digestive vacuole. J Cell Biol113: 1033–1042.
Crabb, B.S., and Cowman, A.F. (1996) Characterization ofpromoters and stable transfection by homologous and nonhomologous recombination in Plasmodium falciparum.Proc Natl Acad Sci USA 93: 7289–7294.
Crabb, B.S., Cooke, B.M., Reeder, J.C., Waller, R.F.,Caruana, S.R., Davern, K.M., et al. (1997) Targeted genedisruption shows that knobs enable malaria-infected redcells to cytoadhere under physiological shear stress. Cell89: 287–296.
Dolan, S.A., Miller, L.H., and Wellems, T.E. (1990) Evidencefor a switching mechanism in the invasion of erythrocytesby Plasmodium falciparum. J Clin Invest 86: 618–624.
Dolan, S.A., Proctor, J.L., Alling, D.W., Okubo, Y., Wellems,T.E., and Miller, L.H. (1994) Glycophorin B as an EBA-175independent Plasmodium falciparum receptor of humanerythrocytes. Mol Biochem Parasitol 64: 55–63.
Escalante, A.A., Lal, A.A., and Ayala, F.J. (1998) Geneticpolymorphism and natural selection in the malaria parasitePlasmodium falciparum. Genetics 149: 189–202.
Galinski, M.R., Corredor-Medina, C., Povoa, M., Crosby, J.,Ingravallo, P., and Barnwell, J.W. (1999) Plasmodium vivaxmerozoite surface protein-3 contains coiled-coil motifs inan alanine-rich central domain. Mol Biochem Parasitol101: 131–147.
Hohjoh, H., and Singer, M.F. (1996) Cytoplasmic ribonucleo-protein complexes containing human LINE-1 protein andRNA. EMBO J 15: 630–639.
Holder, A.A., and Freeman, R.R. (1984) The three major anti-gens on the surface of Plasmodium falciparum merozoitesare derived from a single high molecular weight precursor.J Exp Med 160: 624–629.
Holder, A.A., Lockyer, M.J., Odink, K.G., Sandhu, J.S.,Riveros, M.V., Davey, L.S., et al. (1985) Primary structureof the precursor to the three major surface antigens ofPlasmodium falciparum merozoites. Nature 317: 270–273.
Hudson, D.E., Miller, L.H., Richards, R.L., David, P.H., Alving,C.R., and Gitler, C. (1983) The malarial merozoite surface:a 140,000 MW protein antigenically unrelated to othersurface components on Plasmodium knowlesi merozoites.J Immunol 130: 2886–2890.
Kaneko, O., Fidock, D.A., Schwartz, O.M., and Miller, L.H.(2000) Disruption of the C-terminal region of EBA-175 in the Dd2/Nm clone of Plasmodium falciparum does notaffect erythrocyte invasion. Mol Biochem Parasitol 110:135–146.
Kaslow, D.C., Hui, G., and Kumar, S. (1994) Expression andantigenicity of Plasmodium falciparum major merozoitesurface protein (MSP119) variants secreted from Saccha-romyces cerevisiae. Mol Biochem Parasitol 63: 283–289.
Kushwaha, A., Rao, P.P., Duttu, V.S., Malhotra, P., andChauhan, V.S. (2000) Expression and characterisation of Plasmodium falciparum acidic basic repeat antigenexpressed in Escherichia coli. Mol Biochem Parasitol106: 213–224.
La Greca, N., Hibbs, A.R., Riffkin, C., Foley, M., and Tilley, L.(1997) Identification of an endoplasmic reticulum-residentcalcium-binding protein with multiple EF-hand motifs inasexual stages of Plasmodium falciparum. Mol BiochemParasitol 89: 283–293.
Lambros, C., and Vanderberg, J.P. (1979) Synchronization of Plasmodium falciparum erythrocytic stages in culture. J Parasitol 65: 418–420.
Legrain, P., and Chapon, C. (1993) Interaction betweenPRP11 and SPP91 yeast splicing factors and characteri-zation of a PRP9–PRP11–SPP91 complex. Science 262:108–110.
Lupas, A. (1996) Coiled coils: new structures and new func-tions. Trends Biochem Sci 21: 375–382.
McColl, D.J., and Anders, R.F. (1997) Conservation of struc-tural motifs and antigenic diversity in the Plasmodium falciparum merozoite surface protein-3 (MSP-3). MolBiochem Parasitol 90: 21–31.
McColl, D.J., Silva, A., Foley, M., Kun, J.F.J., Favaloro, J.M.,Thompson, J.K., et al. (1994) Molecular variation in a novelpolymorphic antigen associated with Plasmodium falci-parum merozoites. Mol Biochem Parasitol 68: 53–67.
Marshall, V.M., Silva, A., Foley, M., Cranmer, S., Wang, L.,McColl, D.J., et al. (1997) A second merozoite surfaceprotein (MSP-4) of Plasmodium falciparum that contains an epidermal growth factor-like domain. Infect Immun65: 4460–4467.
Marshall, V.M., Tieqiao, W., and Coppel, R.L. (1998) Closelinkage of three merozoite surface protein genes on chromosome 2 of Plasmodium falciparum. Mol BiochemParasitol 94: 13–25.
Mulhern, T.D., Howlett, G.J., Reid, G.E., Simpson, R.J.,McColl, D.J., Anders, R.F., and Norton, R.S. (1995) Solution structure of a polypeptide containing four heptad repeat units from a merozoite surface antigen of Plasmodium falciparum. Biochemistry 34: 3479–3491.
Nwagwu, M., Haynes, J.D., Orlandi, P.A., and Chulay, J.D.(1992) Plasmodium falciparum: chymotryptic-like proteoly-sis associated with a 101-kDa acidic-basic repeat antigen.Exp Parasitol 75: 399–414.
Oeuvray, C., Bouharoun-Tayoun, H., Gras-Masse, H.,Bottius, E., Kaidoh, T., Aikawa, M., et al. (1994a) Merozoitesurface protein-3: a malaria protein inducing antibodiesthat promote Plasmodium falciparum killing by cooperationwith blood monocytes. Blood 84: 1594–1602.
Oeuvray, C., Bouharoun-Tayoun, H., Grass-Masse, H.,Lepers, J.P., Ralamboranto, L., Tartar, A., and Druilhe, P.(1994b) A novel merozoite surface antigen of Plasmodiumfalciparum (MSP-3) identified by cellular-antibody coopera-tive mechanism antigenicity and biological activity of anti-bodies. Mem Inst Oswaldo Cruz 89: 77–80.
Okoyeh, J.N., Pillai, C.R., and Chitnis, C.E. (1999) Plas-modium falciparum field isolates commonly use erythro-cyte invasion pathways that are independent of sialic acidresidues of glycophorin A. Infect Immun 67: 5784–5791.
Orlandi, P.A., Klotz, F.W., and Haynes, J.D. (1992) A malariainvasion receptor, the 175-kilodalton erythrocyte bindingantigen of Plasmodium falciparum recognizes the terminalNeu5Ac (alpha2–3) Gal- sequences of glycophorin A. J Cell Biol 116: 901–909.
Reed, M.B., Caruana, S.R., Batchelor, A.H., Thompson, J.K.,
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411
1410 K. E. Mills, J. A. Pearce, B. S. Crabb and A. F. Cowman

Trafficking of MSP3 and ABRA to the parasitophorous vacuole 1411
Crabb, B.S., and Cowman, A.F. (2000) Targeted disruptionof an erythrocyte binding antigen in Plasmodium falci-parum is associated with a switch toward a sialic acid independent pathway of invasion. Proc Natl Acad Sci USA97: 7509–7514.
Reiss, M., Viebig, N., Brecht, S., Fourmaux, M.N., Soete, M.,Di Cristina, M., et al. (2001) Identification and charact-erization of an escorter for two secretory adhesins in Toxoplasma gondii. J Cell Biol 52: 563–578.
Smythe, J.A., Coppel, R.L., Brown, G.V., Ramasamy, R.,Kemp, D.J., and Anders, R.F. (1988) Identification of twointegral membrane proteins of Plasmodium falciparum.Proc Natl Acad Sci USA 85: 5195–5199.
Trager, W., and Jensen, J.B. (1976) Human malaria parasitesin continuous culture. Science 193: 673–675.
Trucco, C., Fernandez-Reyes, D., Howell, S., Stafford, W.H.,Scott-Finnigan, T.J., Grainger, M., et al. (2001) The mero-zoite surface protein 6 gene codes for a 36 kDa proteinassociated with the Plasmodium falciparum merozoitesurface protein-1 complex. Mol Biochem Parasitol 112:91–101.
Wu, Y., Kirkman, L.A., and Wellems, T.E. (1996) Trans-formation of Plasmodium falciparum malaria parasites byhomologous integration of plasmids that confer resistanceto pyrimethamine. Proc Natl Acad Sci USA 93: 1130–1134.
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 1401–1411




















