
MK-Brh2+Blm-Mol. Micr. 2012
11
Brh2 domain function distinguished by differential cellular responses to DNA damage and replication stressMilorad Kojic and William K. Holloman* Department of Microbiology and Immunology, and Weill Cornell Cancer Center, Weill Cornell Medical College, New York, NY 10065, USA. Summary Mutants of the fungus Ustilago maydis defective in the RecQ helicase Blm are highly sensitive to killing by the DNA replication stressor hydroxyurea. This sensitivity or toxicity is dependent on the homologous recombi- nation (HR) system and apparently results from forma- tion of dead-end HR DNA intermediates. HU toxicity can be suppressed by deletion of the gene encoding Brh2, the BRCA2 orthologue that serves to regulate HR by mediating Rad51 filament formation on single- stranded DNA. Brh2 harbours two different DNA- binding domains that contribute to HR function. DNA- binding activity from a single domain is sufficient to provide Brh2 functional activity in HR, but to enable HU-induced killing two functional DNA-binding domains must be present. Despite this stringent requirement for dual functioning domains, the source of DNA-binding domains is less critical in that heter- ologous domains can substitute for the native endog- enous ones. The results suggest a model in which the nature of the DNA lesion is an important determinant in the functional response of Brh2 action. Introduction Repairing damaged DNA by homologous recombination (HR) is a mechanism for maintaining stability and restoring integrity of the genetic material common throughout the domains of life. The HR repair pathway can accommodate damaged DNA channelled from a number of different sources. DNA broken by the action of exogenous genotox- ins, ionizing radiation, replication-associated events or programmed cellular processes can be restored through the HR pathway (San Filippo et al., 2008; Budzowska and Kanaar, 2009; Mimitou and Symington, 2009). During DNA replication the HR system can be utilized for repair of a broken sister chromatid arising from fork passage over a nicked template and for processing stalled or collapsed replication forks. Repair of single-stranded DNA (ssDNA) gaps formed as a consequence of replication fork skipping past a lesion on the template strand can also be processed through the HR system (Lehmann and Fuchs, 2006; Lopes et al., 2006). Aberrant DNA structures formed under repli- cation stress or resulting from encounter of obstacles during DNA synthesis can be generated and also resolved by the HR system (Saintigny et al., 2001; Liberi et al., 2005; Mankouri et al., 2011). In eukaryotes DNA substrates directed into the HR system are engaged by Rad51, which assembles on exposed single-stranded regions to form orderly filamen- tous structures active for homologous pairing and DNA strand invasion (Heyer et al., 2010). Access of Rad51 to DNA is regulated by mediator proteins (San Filippo et al., 2008). In many taxa the primary mediator appears to be of the BRCA2 protein family, the eponymous member being the product of a hereditary breast cancer predisposition gene in human (Thorslund and West, 2007; Holloman, 2011). BRCA2 is an extremely large protein (> 3000 amino acids) with characteristic features that include crucial sites for interaction with Rad51 and DNA. BRCA2 interacts with Rad51 through the BRC repeat, a conserved motif of about 35 amino acid residues, reiterated eight times in the mam- malian proteins. An additional site of Rad51 interaction unrelated in sequence to the BRC motif is present at the extreme C-terminus of BRCA2. The BRC repeats promote the nucleation and growth of the Rad51 filament on ssDNA to stimulate DNA strand exchange (Carreira et al., 2009; Shivji et al., 2009), while the C-terminal Rad51-interaction site helps co-ordinate the filament assembly with the cell cycle (Ayoub et al., 2009). The C-terminal region of BRCA2 contains a conserved DNA-binding domain consisting of a helix-rich module followed by three tandem OB (oligonucleotide/oligosaccharide-binding) folds that are structurally related to the binding folds in the ubiquitous ssDNA-binding protein RPA (Yang et al., 2002), which is essential for numerous processes in DNA replication, recombination and repair. BRCA2 promotes the activity of Rad51 in HR through the co-ordinated action of these different domains. It enforces delivery of Rad51 to ssDNA, attenuates association of Rad51 with double-stranded DNA (dsDNA), and accelerates the RPA displacement from Accepted 21 November, 2011. *For correspondence. E-mail [email protected]; Tel. (+1) 212 746 6510; Fax (+1) 212 746 8587. Molecular Microbiology (2012) 83(2), 351–361 doi:10.1111/j.1365-2958.2011.07935.x First published online 15 December 2011 © 2011 Blackwell Publishing Ltd
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of MK-Brh2+Blm-Mol. Micr. 2012

Brh2 domain function distinguished by differential cellularresponses to DNA damage and replication stressmmi_7935 351..361
Milorad Kojic and William K. Holloman*Department of Microbiology and Immunology, and WeillCornell Cancer Center, Weill Cornell Medical College,New York, NY 10065, USA.
Summary
Mutants of the fungus Ustilago maydis defective in theRecQ helicase Blm are highly sensitive to killing by theDNA replication stressor hydroxyurea. This sensitivityor toxicity is dependent on the homologous recombi-nation (HR) system and apparently results from forma-tion of dead-end HR DNA intermediates. HU toxicitycan be suppressed by deletion of the gene encodingBrh2, the BRCA2 orthologue that serves to regulate HRby mediating Rad51 filament formation on single-stranded DNA. Brh2 harbours two different DNA-binding domains that contribute to HR function. DNA-binding activity from a single domain is sufficient toprovide Brh2 functional activity in HR, but to enableHU-induced killing two functional DNA-bindingdomains must be present. Despite this stringentrequirement for dual functioning domains, the sourceof DNA-binding domains is less critical in that heter-ologous domains can substitute for the native endog-enous ones. The results suggest a model in which thenature of the DNA lesion is an important determinant inthe functional response of Brh2 action.
Introduction
Repairing damaged DNA by homologous recombination(HR) is a mechanism for maintaining stability and restoringintegrity of the genetic material common throughout thedomains of life. The HR repair pathway can accommodatedamaged DNA channelled from a number of differentsources. DNA broken by the action of exogenous genotox-ins, ionizing radiation, replication-associated events orprogrammed cellular processes can be restored throughthe HR pathway (San Filippo et al., 2008; Budzowska andKanaar, 2009; Mimitou and Symington, 2009). During DNAreplication the HR system can be utilized for repair of a
broken sister chromatid arising from fork passage over anicked template and for processing stalled or collapsedreplication forks. Repair of single-stranded DNA (ssDNA)gaps formed as a consequence of replication fork skippingpast a lesion on the template strand can also be processedthrough the HR system (Lehmann and Fuchs, 2006; Lopeset al., 2006). Aberrant DNA structures formed under repli-cation stress or resulting from encounter of obstaclesduring DNA synthesis can be generated and also resolvedby the HR system (Saintigny et al., 2001; Liberi et al.,2005; Mankouri et al., 2011).
In eukaryotes DNA substrates directed into the HRsystem are engaged by Rad51, which assembles onexposed single-stranded regions to form orderly filamen-tous structures active for homologous pairing and DNAstrand invasion (Heyer et al., 2010). Access of Rad51 toDNA is regulated by mediator proteins (San Filippo et al.,2008). In many taxa the primary mediator appears to be ofthe BRCA2 protein family, the eponymous member beingthe product of a hereditary breast cancer predispositiongene in human (Thorslund and West, 2007; Holloman,2011). BRCA2 is an extremely large protein (> 3000 aminoacids) with characteristic features that include crucial sitesfor interaction with Rad51 and DNA. BRCA2 interacts withRad51 through the BRC repeat, a conserved motif of about35 amino acid residues, reiterated eight times in the mam-malian proteins. An additional site of Rad51 interactionunrelated in sequence to the BRC motif is present at theextreme C-terminus of BRCA2. The BRC repeats promotethe nucleation and growth of the Rad51 filament on ssDNAto stimulate DNA strand exchange (Carreira et al., 2009;Shivji et al., 2009), while the C-terminal Rad51-interactionsite helps co-ordinate the filament assembly with the cellcycle (Ayoub et al., 2009). The C-terminal region of BRCA2contains a conserved DNA-binding domain consistingof a helix-rich module followed by three tandem OB(oligonucleotide/oligosaccharide-binding) folds that arestructurally related to the binding folds in the ubiquitousssDNA-binding protein RPA (Yang et al., 2002), which isessential for numerous processes in DNA replication,recombination and repair. BRCA2 promotes the activity ofRad51 in HR through the co-ordinated action of thesedifferent domains. It enforces delivery of Rad51 to ssDNA,attenuates association of Rad51 with double-strandedDNA(dsDNA), and accelerates the RPAdisplacement from
Accepted 21 November, 2011. *For correspondence. [email protected]; Tel. (+1) 212 746 6510; Fax (+1) 212746 8587.
Molecular Microbiology (2012) 83(2), 351–361 � doi:10.1111/j.1365-2958.2011.07935.xFirst published online 15 December 2011
© 2011 Blackwell Publishing Ltd

ssDNA by Rad51 (Jensen et al., 2010). Through its abilityto promote and maintain formation of the Rad51 filament,BRCA2 performs a critical role in homology-directed repairof broken DNA molecules. Moreover, BRCA2 provides aprotective role for replication forks – outside of its repairfunction – in helping prevent degradation of newly repli-cated strands at stalled replication forks (Hashimoto et al.,2010; Schlacher et al., 2011). This protective functioninvolves recruitment of Rad51 to stalled, uncoupled repli-cation forks so as to inhibit nucleolytic processing ofnascent strands.
Brh2 is the BRCA2 orthologue in the fungus Ustilagomaydis but appears as a more streamlined version. It isone-third the size of BRCA2 and contains only a singleRad51-binding BRC motif located in the N-terminal regionof the protein (Kojic et al., 2002; 2003). Mapping studiesrevealed that a region at the extreme C-terminus structur-ally unrelated to the C-terminal Rad51-binding motif inBRCA2 also binds Rad51 (Zhou et al., 2007). In addition,genetic analysis in U. maydis and co-precipitation meth-odology established that an element related to a DMC1-interaction motif in BRCA2 also contributes to Rad51filament assembly (Kojic et al., 2011), DMC1 being ameiosis-specific RAD51-related strand exchange protein.In Brh2 two separate regions in the protein interactwith DNA. Within the C-terminus is the domain (CTD)that is structurally related to the DNA-binding domainof mammalian BRCA2 although it appears to lack thethird OB fold based on sequence alignment (Kojic et al.,2003). Within the N-terminal part of the protein is anempirically determined DNA-binding domain (NBD) thatpreferentially binds to D-loops or branched structureswith protruding single-stranded regions (Zhou et al.,2009a).
BLM, the Bloom’s syndrome RecQ family DNA heli-case, could be considered able to provide a counterbal-ance to BRCA2 function due to its ability to dissociate theRad51–ssDNA filament as well as D-loop strand invasionintermediates (van Brabant et al., 2000; Bachrati et al.,2006; Bugreev et al., 2007). But besides this anti-recombinational activity, BLM or orthologues such asSgs1 from budding yeast have been shown to performmultiple additional functions in overcoming obstacles toDNA replication (Liberi et al., 2005; Mankouri et al., 2011),preparing DNA ends for recombinational repair (Mimitouand Symington, 2009), and dissolving double Hollidayjunction intermediates (Wu and Hickson, 2003). InU. maydis, blm loss-of-function mutants are crippled ingrowth when cultured in the presence of the replicationstressor hydroxyurea (HU), but viability is substantiallyrestored when Brh2 is deleted (Mao et al., 2009). Thisobservation suggests that processing of HU-induced DNAstructures by HR in the course of repair can lead toformation of intermediates, which become toxic in the
absence of Blm action, as previously proposed in studieswith budding and fission yeasts (Gangloff et al., 2000;Hope et al., 2005).
Ustilago maydis mutants lacking Brh2 are defective indouble-strand break repair, heteroallelic recombinationand meiosis, and are extremely sensitive to DNA damag-ing agents including ionizing radiation, alkylating chemi-cals and UV light (Kojic et al., 2002). Since UV damageis thought to lead to single-stranded gaps in daughterstrands during DNA replication as the replication forkoccasionally skips past lesions, the UV sensitivity of brh2mutants is likely a consequence of genome instability dueto failed recombinational repair of the single-strandedgaps.
We showed previously that certain mutant variant formsof Brh2 with deletions of the entire C-terminal domain orwith a heterologous DNA-binding domain in place of theC-terminal domain provide a level of activity that permitscells expressing these variants to survive UV irradiationas well as wild type (Kojic et al., 2005). This observationimplies that these Brh2 variants are active in promotingrepair activity and enable processing UV damage-derivedDNA structures via the HR pathway. Nevertheless, theaction of these Brh2 variants should not be categorizedas functionally normal in that strains expressing thesevariants exhibit elevated levels of both double-strandgap repair and allelic recombination, pointing to somedysfunction in how recombination is executed. Takentogether, however, these findings suggest a model forBrh2 in which the N-terminal and C-terminal regions coop-erate in some as yet unknown way to promote recombi-nation in a precise and appropriately regulated manner(Zhou et al., 2009b).
Here we were interested in exploring how theN-terminal and C-terminal regions of Brh2 contribute tofunctional activity by studying mutant variants deleted ofeither the NBD or CTD, or else containing heterologousDNA-binding modules in place of the endogenousdomains (Kojic et al., 2011). The aim was to assess thecontribution of the Brh2 domains in response toUV-induced damage or HU-induced replication forkstalling.
Results
Recombination proficiency and HU toxicity in theabsence of Blm function
We reported previously that lethality of a U. maydis blmmutant strain caused by the DNA replication stressor HUcould be attenuated by deleting the gene encoding Brh2required for HR (Mao et al., 2009). To determine whetherthe observed attenuation was a general characteristic ofthe HR system of U. maydis or a peculiar response to
352 M. Kojic and W. K. Holloman �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

deletion of the Brh2 gene, as it is formally possible that aspecific HU-induced activity of Brh2 is abrogated by Blm,we tested survival of the blm mutant after deleting otherrecombination genes. We measured HU sensitivity, andalso resistance to UV as an indirect measure of recombi-national repair proficiency. UV is a potent inducer ofrecombination and inactivation of HR genes results inextreme sensitivity to UV in U. maydis (Holloman et al.,2008). When the gene for Brh2, Rad51, Rec2 (a Rad51paralogue) or Rad54 (a chromatin remodelling translo-case that stimulates Rad51) was deleted, there was lossof resistance to UV and, in every case, concomitant resis-tance to killing by HU (Fig. 1A). Therefore, in accord withother systems, it would appear that HR proficiency inU. maydis is a general feature that sets the stage forHU-induced lethality in a Blm-deficient mutant.
For simplicity in work described below, the phrase ‘HU-induced lethality’ or variations such as ‘HU toxicity’, etc.,will refer specifically to HU-induced toxicity in the contextof the Blm-deficient mutant.
DNA structure resulting from HU stress
Single-stranded breaks and gaps generated following UVirradiation, likely by replication fork-restart downstream ofsites of UV-induced DNA photodamage (Lehmann andFuchs, 2006; Lopes et al., 2006), are substrates for HRaction as evinced by the strong induction of HR by UV(Roman and Jacob, 1958) and the sensitivity of HRmutants to UV (Holloman et al., 2008). However, thenature of the structure of the putative HU-induced DNAintermediate that becomes toxic in the absence of func-tional Blm activity is unclear. HU specifically inhibits ribo-nucleotide reductase resulting in depletion of nucleotidepools and as a consequence DNA replication is disturbed(Timson, 1975). HU treatment is presumed to provokegeneration of DNA structures with single-stranded gapsas replication forks stall or as the co-ordination of leadingand lagging strand synthesis breaks down (Fabre et al.,2002), although certain off-target secondary events mightalso contribute to DNA damage (Davies et al., 2009). Butin contrast to the processing of UV-induced DNA damage,resolution of DNA intermediates caused by HU treatmentdepends on the action of Blm-related RecQ helicasesrather than through the homology-directed repair activityof the HR system (Liberi et al., 2005).
As an approach for gaining some insight into the natureof the DNA lesion(s) induced by HU stress, we examinedthe appearance of chromosomes in strains under investi-gation here by pulsed field gel electrophoresis (Fig. 1B).U. maydis has 23 chromosomes ranging in size from 0.34to 2.5 Mb (http://mips.helmholtz-muenchen.de/genre/proj/ustilago/). We were surprised to note that in all the strainsHU tested treatment for even a relatively brief duration
caused the disappearance of the entire complement ofchromosomal DNAs from the gel. However, the observa-tions that no extensive DNA fragmentation was detectablein the gels and that the chromosomes reappeared shortlyafter HU was removed from the medium suggest that theDNA had not been degraded. Rather, it seems that thechromosome-sized DNA molecules failed to enter the gel,presumably due to being rendered into conformationalstate that precludes electrophoresis. In the example ofwild type, after removal of HU and addition of fresh culturemedium, cells were able to resume growth after a shortrecovery period with little loss of viability (data not shown)
wild type
blm
brh2
HU recovery
0 1 2 3 0.5 1 2M
Time (hr)
blm brh2
control UV HUwild type
blm brh2
blm brh2
blm mus81
mus81
blm mus81 + RusA blm + RusA
MMS
HU recovery
0 1 2 3 0.5 1 2M
A
B
control UV HU
wild type
brh2 blmblm
rad51 blmrec2 blm
rad54 blm
C
Fig. 1. HU poisoning and homologous recombinationdependence.A. Survival of mutant strains after UV irradiation (120 J m-2) orexposure to HU (4 mM). Strains indicated were grown in liquidmedium, adjusted to a density of 2 ¥ 107 ml-1 spotted on solidmedium as serial 10-fold dilutions from left to right. Images wererecorded after 2–3 days of incubation.B. CHEF gels of genomic DNA complements of indicated strains.HU (20 mM) was added to exponentially growing cells at a densityof 2 ¥ 107 ml-1 for a period of 3 h. For recovery the cells werecollected, washed free of HU and resuspended in fresh medium.Aliquots removed at the indicated times were prepared for CHEFgel electrophoresis. M-chromosomal DNAs from Saccharomycescerevisiae were run as size standards.C. Mutant strains including those indicated expressing RusA weretested for survival after UV irradiation (120 J m-2), exposure toMMS (0.01%), or HU (4 mM).
Brh2 DNA-binding domain function 353
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

indicating that no lasting damage had been done. After arecovery period of 1–2 h the entire chromosome comple-ment once again became evident after electrophoresissuggesting that HU-induced lesions had been repaired(Fig. 1B). Similarly, the chromosome complement frombrh2 mutant cells disappeared from the gel after HU treat-ment and then reappeared, but even faster than in wildtype. In contrast, the chromosome complement from blmmutant cells became evident more slowly after recoveryfrom HU treatment, while chromosomes from the blm brh2double mutant appeared to be restored slightly faster thanthe blm single mutant. Two conclusions can be drawnfrom this analysis. First, at the gross level of DNA struc-ture discernible by pulsed field gel electrophoresis, itappears that HU treatment results in a global change inthe genome structure. Second, chromosomal integrity canbe restored upon HU removal but restoration appears lessefficient in blm mutant compared to brh2 mutant.
We considered the possibility that the HU-inducedchange causing disappearance of the chromosomalDNAs might be due to generation of Holliday junction-containing X-forms. Since Blm orthologues from yeastand human have been shown capable of removing doubleHolliday junctions (Wu and Hickson, 2003; Mankouriet al., 2011), the absence of such an activity mightaccount for the sensitivity of the blm mutant to HU.However, when the Holliday junction resolvase RusA wasexpressed in blm cells, there was little or only incrementalimprovement in survival after HU treatment (Fig. 1C). Onthe other hand, complementation of the extreme UV andmethylmethane sulphonate (MMS) phenotype of the blmmus81 double mutant by RusA was remarkably completeindicating that it could substitute for Mus81 in resolvingMMS-induced, but not HU-induced HR intermediates.Because it has been established in other systems thatMMS treatment causes formation of Holliday junctions orX-forms (Liberi et al., 2005; Mankouri et al., 2011), ourfindings suggest that some kind of HR DNA intermediateis generated during HU treatment, but evidently not con-taining just X-forms, if at all. Apparently something morethan generation of X-forms takes place. Stated anotherway, while an X-form structure could well be produced asan intermediate(s) during the HR process, it appears thatthe HU toxicity in the blm mutant does not result from aninability to resolve X-forms, but rather to some other asyet undefined dead-end structure.
Contribution of the DNA-binding domainsto HU-induced toxicity
Two different DNA-binding domains contribute to Brh2functional activity – the NBD located in the N-terminalregion, and the OB fold-containing CTD (Zhou et al.,2009a). This latter is the site for interaction with the regu-
lator protein Dss1 (Zhou et al., 2009b). A surprising obser-vation made previously was that a strain expressing atruncated form of Brh2 deleted of the entire CTD (e.g.Brh2DC551) was proficient in recombination (Kojic et al.,2005). It exhibited resistance to UV, proficiency in allelicrecombination and double-strand break repair, and inde-pendence of the requirement for Dss1 as a cofactor. Simi-larly, a strain expressing a chimeric Brh2, consisting of theN-terminal region fused to a heterologous DNA-bindingdomain comprised of the RPA70 single-strand bindingprotein subunit (Brh2DC551::RPA70) was resistant to UVand exhibited elevated recombination activity regardlessof Dss1 status (Kojic et al., 2005).
Given the recombination proficiency of these two differ-ent Brh2 variants, it seemed reasonable to conclude thatboth would cause HU-induced lethality in the absence ofBlm activity. Nevertheless, expression of these Brh2 vari-ants in the blm mutant resulted in different outcomes inresponse to DNAreplication stress (Fig. 2A). Expression ofmutant variant Brh2DC551 lacking the CTD did not reversethe HU resistance, while expression of the chimeric hybridBrh2DC551::RPA70 restored HU sensitivity causing cellkilling. Therefore, recombination proficiency per se doesnot necessarily lead to HU-induced toxicity in the absenceof Blm. In other words, it appears that Brh2 deleted of theCTD is still capable of providing sufficient functional activityfor recombinational repair of UV-induced lesions, butcannot enable generation of the presumed HR inter-mediate induced by replication stress that is toxic in theabsence of Blm activity. In contrast, coupling an additionalDNA-binding domain, as in the Brh2DC551::RPA70 hybrid,potentiates the activity of the N-terminal region andenables processing of putative HU-induced DNA interme-diates to a toxic form.
In the reciprocal experiment we examined whether thesame response was true for Brh2 forms lacking the NBDor containing a substitute DNA-binding domain. DNA-binding activity of the N-terminal region of Brh2 was pre-viously mapped by direct biochemical assay associatedwith overlapping polypeptides in conjunction with geneticcomplementation of the brh2 mutant phenotype by trun-cations of the N-terminal fragment (Zhou et al., 2009a).The region was localized approximately to residues359–503 although the precise boundaries remainuncertain in the absence of more rigorous structuralinformation. While the DNA-binding activity associatedwith the NBD had been previously determined to be moreavid than that associated with the isolated CTD, deletionof NBD from full-length Brh2 did not result in loss of repaircapacity as measured by UV resistance (Fig. 2B). TwoBrh2 forms (Brh2D362–493 and Brh2D362–541) differingby 49 residues at the distal endpoint of the internal dele-tion were tested and appeared comparable in conferr-ing DNA repair activity. However, brh2 mutant strains
354 M. Kojic and W. K. Holloman �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

expressing these two forms were not poisoned by HU inthe absence of Blm activity, similar to the finding withBrh2DC551, which retains the NBD but is deleted of theCTD. These observations suggest that a single DNA-binding domain suffices for Brh2 to promote proficiency inrecombinational repair after UV damage, but that both the
N-terminal and C-terminal DNA-binding domains of Brh2must be in place to establish the appropriate recombina-tion intermediate for HU-induced toxicity in the absence ofBlm activity.
In a test for the degree of stringency in DNA-binding-domain structure required for functional activity, the NBDregion from the two Brh2 mutant isoforms was replaced bya heterologous module consisting of OB-A and OB-B folds(OB-AB) from RPA70 (Fig. 2B). Complementation of UVsensitivity was conferred by expression of these chimericBrh2 variants. However, restored sensitivity to HU wasevident with only one of the Brh2 variants (Brh2D362–493::OB-AB), not the other (Brh2D362–541::OB-AB). Thisobservation shows that the OB-AB domain can substitutefor the NBD to a certain extent for functional proficiency, butthat the region between residues 493 and 541 contributessome additional property that is important for promotingformation of the HU-induced toxic intermediate. In theabsence of more concrete information about its structure orfunction, we shall designate this 49-amino-acid region asthe ‘spacer’ in discussion below because it links theN-terminal and C-terminal domains.
Two-DNA-binding-domain model for HU toxicity
With the spacer region present it seemed apparent thatsubstituting the NBD with the OB-AB module was suffi-cient to form a chimeric Brh2 active in generating aHU-inducible toxic intermediate in the blm mutant. Toexamine this spacer feature in an alternative context, wetested the requirement for the spacer in the reciprocalconfiguration by swapping the CTD for the RPA70 OB-foldarray retaining or deleting the spacer region (Fig. 2C).In agreement with the NBD replacement studies, theBrh2DC551::RPA70 and Brh2DC505::RPA70 chimericproteins both promoted recombinational repair activityafter UV damage, but the Brh2DC551::RPA70 chimericfused with the spacer region was more potent inHU-induced toxicity than the fusion without the spacer.The results suggest that severe toxicity caused by HUexposure to the blm mutant is caused when a Brh2isoform harbouring two DNA-binding domains separatedby the spacer region is present.
No protein motifs are recognizable within the spacersequence and there is considerable divergence evenamong fungal orthologues, although a sprinkling of con-served residues is evident (Fig. 2D). A noteworthy feature,however, is the excess number of basic residues to theextent that the calculated pI of the spacer sequence is 11.8.
Domain interplay
Previously, using pull-down methodology and proteincross-linking procedures, we obtained evidence that Brh2
Fig. 2. DNA-binding domain contribution to Brh2-promoted UVsurvival or HU poisoning.A–C. Survival of brh2 blm mutant strains expressing Brh2 variantsor hybrids with RPA70 OB-folds, as illustrated schematically, afterUV irradiation (120 J m-2) or exposure to HU (4 mM).D. Sequence alignment of the 49 residue spacer region of Brh2 infungal orthologues. As an anchor for orientation the conservedFVTP motif important for Dmc1 interaction is shown, but the spacerregion begins at U. maydis Brh2 K493. Accession numbers of Brh2orthologues: Coprinopsis cinerea EAU92556, Postia placentaEED79697, Rhodotorula glutinis EGU12633, Serpula lacrymansENG93090, Sporisorium reilianum CBQ73595.
Brh2 DNA-binding domain function 355
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

could interact with itself to form dimeric and higher orderstructures (Zhou et al., 2007). Intermolecular interactionappeared to be mediated through the region spanning thehelical domain and OB1 fold. Genetic evidence supportingprotein–protein interaction came from the observation thattwo different inactive mutant forms could cooperate tocomplement the UV phenotype when coexpressed in thebrh2 mutant. One inactive variant was deleted at theN-terminus to remove the Rad51-interacting BRC motifwhile the other was truncated at the C-terminus to removea short tract of residues encompassing the Rad51-interacting region there. Separately the mutant variantswere inactive, but when expressed together they wereable to restore UV resistance. This same cooperation wasevident also in the absence of Blm activity (Fig. 3A). Thesefindings suggest that Brh2 mutant forms cooperate throughintermolecular interaction in recombinational repair of DNAlesions caused by UV. However, when we tested whetherthe same inactive Brh2 variants could cooperate inresponse to HU, we found that the cells did not becomesensitive to HU challenge (Fig. 3A). These results suggestthat Brh2 cooperation, which presumably reflects domaininteraction, responds to the nature of the DNA substrate.Response of Brh2 to stalled replication forks appears more
demanding in terms of precise domain architecture thanresponse to UV-induced DNA lesions.
In all the different variants of Brh2 above in which theinterplay between two DNA-binding domains was testedfor activity in either the intramolecular or intermolecularconfiguration, the C-terminal binding domain was com-posed of an OB-fold array. To test whether an OB-fold wasa requirement, we constructed a Brh2 variant with a dif-ferent C-terminal DNA binding, namely the DNA-binding/self-association domain from Rad52 (Kojic et al., 2008).This fold bears no resemblance to the OB-fold (Kagawaet al., 2002; Singleton et al., 2002). Nevertheless, theBrh2DC551::Rad52DBD did not impair UV complementa-tion and was active in supporting the restoration of HUsensitivity (Fig. 3B). We conclude that there is plasticity inthe structural requirements for recognizing and presum-ably processing the HU-induced DNA intermediates, andthat tandem modular arrangement of two different bind-ing domains separated by a spacer region can meet aminimum standard.
Two DNA-binding domains in interhomolog versusintersister recombination
Resistance to UV and sensitivity of the blm mutant to HUmediated by Brh2 and variant isoforms as used above aremetrics of functional proficiency in HR, but are indirectmeasurements. We wished to determine by direct mea-surement the functional contribution of the Brh2 isoformsto HR (Fig. 4). For this purpose, two different reportersystems were utilized for direct readout of recombinationproficiency.
First, allelic recombination was assayed. In this assayrecombination between mutant alleles of nar1 (nitratereductase) residing on homologous chromosomes canreconstruct an active gene enabling growth on nitrate asthe sole nitrogen source (Holliday, 1966). Therefore, weexpressed the Brh2 variants in brh2 diploids and measuredheteroallelic recombination at nar1 in response to UVirradiation or HU block. Conditions of treatment, i.e. dose ofUV and time in HU medium, were standardized in pilotstudies using a wild-type control so that the level of induc-tion of Nar+ recombinants was about the same and killingwas kept to a minimum. The Brh2 variants deleted of eitherthe N-terminal NBD (Brh2D362–541) or C-terminal DBD(Brh2DC551) were both about a third as active as Brh2 inrestoring UV-induced recombination (Fig. 4B). The chi-meric Brh2D362–541::OB-AB with heterologous bindingdomain in place of NBD was also reduced to about thesame extent. However, the chimeric Brh2DC551::RPA70with CTD swapped out was hyperactive suggesting thatthe heterologous DNA-binding domain enabled recom-bination with UV damaged DNA to a significantly higherlevel than the endogenous DNA-binding domain. The
BRC HD CREOB1 OB2/towerBrh2
NBD
Brh2ΔN315
Brh2ΔC1034
wild type
+ Brh2
brh2 blm
control UV HU
+ Brh2ΔN315
+ Brh2ΔC1034
+ Brh2ΔN315/Brh2ΔC1034
Brh2ΔC551
BRC HD CREOB1 OB2/towerBrh2
NBD
wild type
+ Brh2
brh2 blm
+ Brh2ΔC551
A
+ Brh2ΔC551::Rad52 DBD
control UV HU
Brh2ΔC551::Rad52 DBD
Rad52 DBD
B
Fig. 3. Domain interplay.A. Intermolecular interaction. N- and C-terminal truncated forms ofBrh2 shown schematically were introduced into the brh2 blmmutant and tested for intermolecular complementation with UV(120 J m-2) and HU (4 mM).B. Survival of brh2 blm mutant strains expressing Brh2–Rad52DBD hybrid after UV irradiation (120 J m-2) or exposure to HU(4 mM).
356 M. Kojic and W. K. Holloman �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361
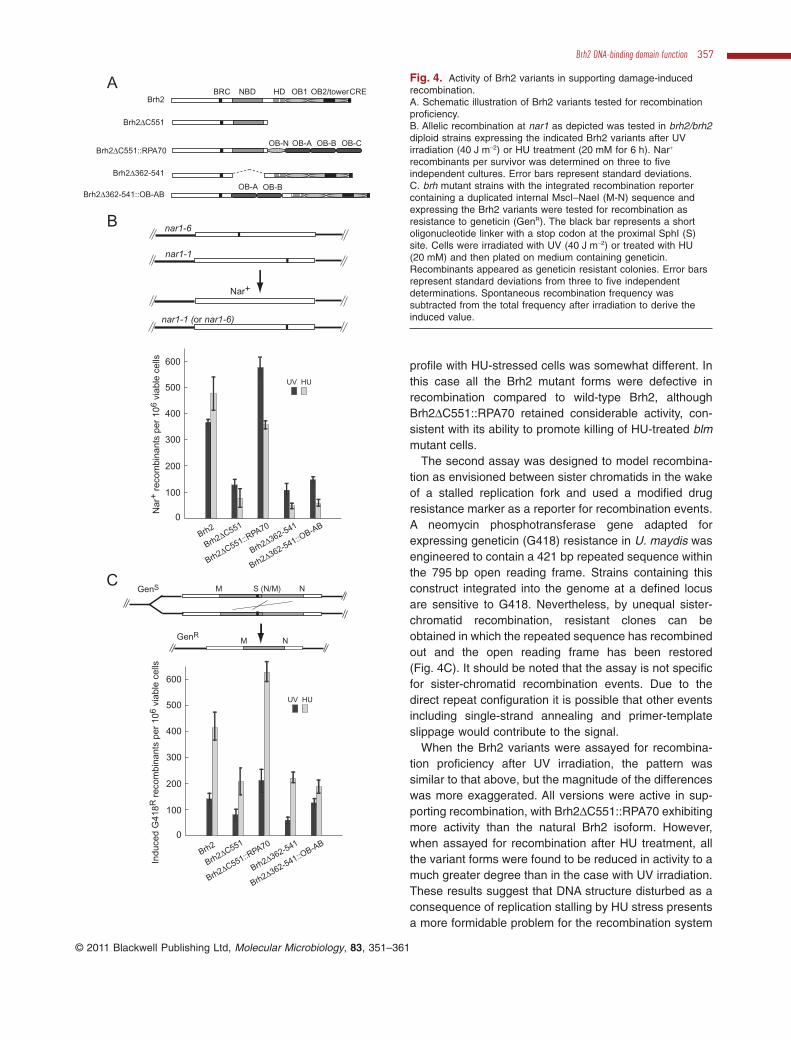
profile with HU-stressed cells was somewhat different. Inthis case all the Brh2 mutant forms were defective inrecombination compared to wild-type Brh2, althoughBrh2DC551::RPA70 retained considerable activity, con-sistent with its ability to promote killing of HU-treated blmmutant cells.
The second assay was designed to model recombina-tion as envisioned between sister chromatids in the wakeof a stalled replication fork and used a modified drugresistance marker as a reporter for recombination events.A neomycin phosphotransferase gene adapted forexpressing geneticin (G418) resistance in U. maydis wasengineered to contain a 421 bp repeated sequence withinthe 795 bp open reading frame. Strains containing thisconstruct integrated into the genome at a defined locusare sensitive to G418. Nevertheless, by unequal sister-chromatid recombination, resistant clones can beobtained in which the repeated sequence has recombinedout and the open reading frame has been restored(Fig. 4C). It should be noted that the assay is not specificfor sister-chromatid recombination events. Due to thedirect repeat configuration it is possible that other eventsincluding single-strand annealing and primer-templateslippage would contribute to the signal.
When the Brh2 variants were assayed for recombina-tion proficiency after UV irradiation, the pattern wassimilar to that above, but the magnitude of the differenceswas more exaggerated. All versions were active in sup-porting recombination, with Brh2DC551::RPA70 exhibitingmore activity than the natural Brh2 isoform. However,when assayed for recombination after HU treatment, allthe variant forms were found to be reduced in activity to amuch greater degree than in the case with UV irradiation.These results suggest that DNA structure disturbed as aconsequence of replication stalling by HU stress presentsa more formidable problem for the recombination system
100
200
300
400
500
600
Nar
+ r
ecom
bina
nts
per
106
viab
le c
ells
Brh2
Brh2ΔC551::RPA70
Brh2Δ362-541
Brh2ΔC551
Brh2Δ362-541::OB-AB
UV HU
0
Brh2Δ362-541::OB-AB
Brh2Δ362-541
OB-BOB-A
BRC HD CREOB1 OB2/towerBrh2
NBD
Brh2ΔC551
OB-BOB-A OB-COB-NBrh2ΔC551::RPA70
100
200
300
400
500
600
Indu
ced
G41
8R r
ecom
bina
nts
per
106
viab
le c
ells
Brh2
Brh2ΔC551::RPA70
Brh2Δ362-541
Brh2ΔC551
Brh2Δ362-541::OB-AB
UV HU
0
GenS
GenR
nar1-1 (or nar1-6)
nar1-6
Nar+
nar1-1
A
B
CM N(N/M)
M N
S
Fig. 4. Activity of Brh2 variants in supporting damage-inducedrecombination.A. Schematic illustration of Brh2 variants tested for recombinationproficiency.B. Allelic recombination at nar1 as depicted was tested in brh2/brh2diploid strains expressing the indicated Brh2 variants after UVirradiation (40 J m-2) or HU treatment (20 mM for 6 h). Nar+
recombinants per survivor was determined on three to fiveindependent cultures. Error bars represent standard deviations.C. brh mutant strains with the integrated recombination reportercontaining a duplicated internal MscI–NaeI (M-N) sequence andexpressing the Brh2 variants were tested for recombination asresistance to geneticin (GenR). The black bar represents a shortoligonucleotide linker with a stop codon at the proximal SphI (S)site. Cells were irradiated with UV (40 J m-2) or treated with HU(20 mM) and then plated on medium containing geneticin.Recombinants appeared as geneticin resistant colonies. Error barsrepresent standard deviations from three to five independentdeterminations. Spontaneous recombination frequency wassubtracted from the total frequency after irradiation to derive theinduced value.
Brh2 DNA-binding domain function 357
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

as mediated by Brh2 than damage resulting from UV. Foran appropriate response, both the NBD and CTD domainsof Brh2 must be intact.
Discussion
UV irradiation is a potent inducer of HR. It is thought thatphotoproducts in DNA are occasionally skipped overduring DNA replication leaving single-stranded gaps in thewake of the fork, and that these gaps are repaired by therecombination machinery using an undamaged homolo-gous sequence, preferably from the sister chromatid, as atemplate. HU, on the other hand, corrupts DNA replicationby inhibiting ribonucleotide reductase causing depletion ofnucleotide pools, which leads to uncoupling of leadingand lagging strand synthesis, accumulation of single-stranded gaps, replication fork stalling, and finallycollapse. DNA structures generated as a consequence ofHU treatment can be acted upon by the HR system andprocessed into intermediates. Normally these structurescan be resolved or removed by the action of additionaldownstream components of the HR system. However, inthe absence of their resolution or removal the structureshave the potential to trigger a pathological response.Such unprocessed dead-end intermediates can becometoxic presumably due to a failure in replication or in chro-mosome segregation during mitosis.
Here we assessed how the different domains of Brh2contribute to proficiency in recombinational repair inresponse to UV irradiation or to generating the pathologi-cal recombination intermediates manifested by HU poi-soning in the absence of Blm function. There are severalnoteworthy findings. First, Brh2 variants deleted of eitherthe NBD or CTD domains are competent for HR andsurvival after UV irradiation, but unlike the natural Brh2form do not cause a pathological state in blm mutant cellstreated with HU. This finding suggests that at least one ofthose Brh2 domains is required to support a thresholdactivity in recombinational repair proficiency, but that bothdomains synergistically contribute to processing interme-diates formed when DNA replication is stressed. Second,either of the domains can be replaced by a heterologousDNA-binding domain from RPA70 with little consequenceon survival after UV damage. However, any combinationof two DNA-binding domains arranged in an appropriateconfiguration is necessary for Brh2 to become pathologi-cal in generating toxic recombination intermediates duringreplication stress. That a small stretch of residues, orspacer region, connecting the DNA-binding regions isimportant for the poisonous response to HU replicationstress suggests the possibility that a precise architecturalarrangement of these domains is required. Third, twodefective Brh2 variants with inactivating truncations inthe N-terminal and C-terminal regions can cooperate
to enable survival after UV irradiation, but not to conferHU-induced killing. In the previously proposed model ofBrh2 as a dimer, it was suggested that proficiency inrecombinational repair requires two functional regions ofBrh2, which could be contributed from two different mol-ecules of Brh2 in a heterodimeric complex (Zhou et al.,2007). In the studies performed here, attainment of thepathological state for HU-induced killing could only beachieved when functional regions of Brh2 are arranged incis, implying that Brh2-mediated processing of certainDNA intermediates requires a more stringent constellationof functional domains.
The particular functional requirement for two DNA-binding domains in Brh2 to ensure successful processingof the HU-induced DNA structures raises an intriguingquestion of the specific molecular events involved in theprocess. A primary function of Brh2 in HR is to controlRad51 (Yang et al., 2005). This protein conducts thesearch for DNA sequence homology and promotes strandinvasion after polymerization on exposed ssDNA to forma nucleoprotein filament. As demonstrated in studieson human BRCA2, Rad51-filament status is regulatedthrough the BRC motif, which promotes assembly ofRad51 on ssDNA while slowing association on dsDNA,and which attenuates ATP hydrolysis of Rad51 whenbound to ssDNA, thereby stabilizing it in an extendedstate suitable for strand invasion (Carreira et al., 2009;Shivji et al., 2009; Jensen et al., 2010). A second Rad51-binding region located at the C-terminus of BRCA2 alsocontributes to filament stability through specific interactionwith the polymerized rather than monomeric form (Daviesand Pellegrini, 2007; Esashi et al., 2007).
Accumulating evidence from studies on Rad51 andBRCA2 in replication stressed cells using vertebratesystems (Saintigny et al., 2001; Lomonosov et al., 2003;Petermann et al., 2010) suggests two mechanistically dif-ferent situations in which these proteins function togetherin replication fork protection and DNA repair. During rep-lication it appears that BRCA2 is recruited to sites of forkstalling where it serves to stabilize the fork by promotingformation of Rad51 filaments on exposed ssDNA toprotect nascent strands from degradation (Hashimotoet al., 2010; Schlacher et al., 2011). After prolonged forkstalling resulting from continuous exposure to HU, forkscollapse coincident with DSB formation (Petermann et al.,2010). The HR system can be brought into action again toenable DSB repair and restore the integrity of thegenome. Mutational analysis of BRCA2 has revealed thatthe BRCA2 C-terminal Rad51-interaction site is essentialfor fork protection activity but is dispensable for HR(Schlacher et al., 2011).
Although it seems entirely possible that Brh2 wouldhave capabilities similar to BRCA2 in nucleating Rad51filament formation on ssDNA and stabilizing the filament,
358 M. Kojic and W. K. Holloman �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

these activities have not yet been formally distinguishedexperimentally with Brh2 nor have been ascribed to dif-ferent protein domains. Nonetheless, as investigatedhere, the responses to UV irradiation or HU treatment ofU. maydis cells expressing the variant forms of Brh2suggest different functional activities can be attributed tothe NBD and CTD protein domains. In contrast, however,to the findings with BRCA2 and the Rad51 filament, theobservations that Brh2 variants active in DNA repair andin promoting HU toxicity in the absence of Blm can becreated by substituting OB-fold DNA-binding modulesfrom RPA70 for either the NBD or CTD domains indicatethe primacy of DNA-binding activity, rather than Rad51filament stabilization, in mediating these responses.Perhaps the conformation or disposition of the Rad51filament along DNA requires precise ordering by the twoDNA-binding domains of Brh2 for optimal activity in pro-cesses that channel stalled replication forks through Blm.Alternatively, a maturation process for DNA intermediatesarising during fork stalling might explain the requirementfor two precisely arranged Brh2 DNA-binding regions.
We imagine that the nature of the DNA lesion is adeterminant in deployment of appropriate Brh2 elements.In the case of post-replication repair after UV irradiation,an actionable response requires only the stripped-down,fundamental mediator activity provided by a BRC motifcoupled to an ssDNA-binding domain. Stalled replicationforks, however, present a more formidable structuralproblem that requires the advent of two spatially co-ordinated DNA-binding domains for adequate response.This dual domain requirement suggests a two-stepprocess. The first might entail the mediator activity ofhelping Rad51 gain access to DNA while the second mightrequire a transition that involves DNA handling by the otherbinding domain for appropriate positioning suitable forsubsequent processing. In this scenario it is the DNA-binding-domain architecture of Brh2 rather than the dispo-sition of the Rad51 filament that establishes competence inelaborate processing of the stalled replication forks. Thespecific possibility of a transitional step is interesting tocontemplate. The additional major hurdle in understandingmolecular events performed by Brh2 during HU-inducedstress is, of course, to identify the specific DNAstructure(s)formed during the process. Our hope is that understandingthe genetic and molecular events will provide intellectualinsights into the broader question of why such an elaborateprocess is required for dealing with challenge to the mostpivotal cellular function of DNA replication.
Experimental procedures
Ustilago maydis strains and genetic methods
Manipulations with U. maydis, culture methods, gene dis-ruption and gene transfer procedures, survival after DNA
damage by UV or HU treatment, synthesis of diploids andmeasurement of allelic recombination, etc., have beendescribed previously [see Zhou et al. (2007), Kojic et al.(2011) and references therein]. For survival in brief, culturesof exponentially growing cells were diluted to 2 ¥ 107 ml-1,and then aliquots (10 ml) of serial 10-fold dilutions werespotted on solid medium, and irradiated with 254 nm UV(120 J m-2) or else were plated on medium containing HU.Plates were incubated at 30°C for 2–3 days until coloniesdeveloped. The blm mutant strain used in these studies wasinactivated for helicase activity by mutation in the Walker Abox (K443R) as described previously (Mao et al., 2009). Arad54 null mutant was constructed by standard gene disrup-tion methodology. The gene was identified as entry um02083in the annotated U. maydis database (see http://mips.helmholtz-muenchen.de/genre/proj/ustilago/) and the entireopen reading frame was replaced in haploid strain UCM350(pan1-1 nar 1–6 a1b1) by a cassette consisting of the strongconstitutive otef promoter from plasmid p123 (Aichingeret al., 2003) [kindly provided by Regine Kahmann (MaxPlanck Institute, Marburg)] fused to a neomycin phospho-transferease gene expressing resistance to geneticin (Gibco/BRL Life Technologies, Rockville, MD). Null mutants in thestructural genes for Brh2, Rad51 and Rec2 were previouslydescribed (Holloman et al., 2008). pan, met, nar and ab indi-cate auxotrophic requirements for pantothenate, inability toutilize nitrate, and mating type loci respectively. Multiplesequence alignment was performed using the T-Coffee align-ment tool (http://www.ebi.ac.uk/Tools/msa/tcoffee/). Molecu-lar karyotype analysis was performed by contour-clampedhomogeneous electric field (CHEF) gel electrophoresis usinga CHEF DRII drive module (Bio-Rad Laboratories) essentiallyas described (Kojic et al., 2002) for 15 h with a switch time of60 s followed by 11 h with a switch time of 90 s. Protoplastsfrom 109 cells were prepared using Glucanex (Sigma) inplace of Novozyme234, embedded in agarose plugs, andthen digested with Proteinase K at 30° for 24 h in a solutioncontaining 10 mM Tris-HCl, pH 8.0, 100 mM EDTA, 1%sodium N-lauroylsarcosine, 0.2% sodium deoxycholate.
Brh2 variants and plasmid construction
Self-replicating plasmids expressing genes encoding Brh2 orderivatives driven by gap promoter contained the hygromycinphosphotransferase gene or carboxin resistance gene forselection and maintenance. Brh2 mutant variants deleted offragments of the N-terminal DNA-binding domain were con-structed as described previously (Kojic et al., 2011). Internaldeletions and insertions were constructed so that the distalcoding region would be translated in frame. Brh2 variantswith RPA70 OB-AB modules in place of the entire NBD (resi-dues 362–541) were constructed using codons 141–434 ofRPA70 encoding the OB-AB fragment inserted in frame withBrh2 coding sequence. A Brh2–Rad52 DBD hybrid containingthe highly conserved DNA-binding/self-association domain ofRad52 (DBD, residues 79–317) was constructed by fusingthe 239-amino-acid Rad52 domain (Kojic et al., 2008) inframe to Brh2DC551. A vector for expression of RusA undercontrol of the gap promoter was constructed by transferringan NdeI–BamHI fragment containing the rusA gene fromplasmid pKR6980 (Bastin-Shanower et al., 2003) kindly pro-
Brh2 DNA-binding domain function 359
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

vided by Steve Brill (Rutgers University) into a self-replicatingplasmid expressing CbxR for selection in U. maydis.
Recombination assays
Allelic recombination at the nar1 locus was measured asdescribed previously (Holliday, 1966; Kojic et al., 2002) bydetermining Nar+ prototroph formation in brh2 homozygousdiploids expressing Brh2 mutant variants after induction witha UV dose of 40 J m-2 or else after holding in medium con-taining 20 mM HU for 7 h. A reporter for measuring sisterchromatid recombination was constructed by duplicating aninternal sequence in the transposon Tn5 neomycin phospho-transferase gene expressing geneticin resistance (GenR) inU. maydis. A 421 bp MscI–NaeI fragment from the middle ofthe open reading frame was introduced into the 795 bp geneopened at the NaeI site generating a reporter with an internaltandem duplication of this sequence. A short linker with a stopcodon in frame was inserted at the SphI site (at position 534in the orf) within the proximal MscI–NaeI repeat to ablateresidual drug resistance. This construct was placed undercontrol of the otef promoter in an integrative plasmid bearingan allele of the iron-sulphur subunit sdh2 of the succinicdehydrogenease gene (um00844) expressing resistance tocarboxin (CbxR) in U. maydis. CbxR transformed brh2 haploidstrains with the integrated recombination reporter that weresensitive to geneticin (100 mg ml-1) were chosen for analysis.For UV-induced recombination, strains were cultured over-night in rich medium, washed, then resuspended in water ata density of 2 ¥ 107 ml-1, spread directly on plates with solidmedium containing geneticin (Gibco/BRL Life Technologies)at 100 mg ml-1, and then irradiated with 40 J m-2 of UV light.For HU induction, freshly growing cells at 2 ¥ 107 ml-1 werecultured in medium spiked with 20 mM HU for 7 h, thenwashed, collected and plated on geneticin-containing solidmedium. GenR colonies were counted after incubating platesthree days at 28°.
Acknowledgements
We thank Lorraine Symington, Columbia University, forstimulating discussion and laboratory members QingwenZhou and Jeanette Sutherland for contributing help andideas. This work received financial support from NIH grantsGM042482 and GM079859.
References
Aichinger, C., Hansson, K., Eichhorn, H., Lessing, F.,Mannhaupt, G., Mewes, W., and Kahmann, R. (2003)Identification of plant-regulated genes in Ustilago maydisby enhancer-trapping mutagenesis. Mol Genet Genomics270: 303–314.
Ayoub, N., Rajendra, E., Su, X., Jeyasekharan, A.D., Mahen,R., and Venkitaraman, A.R. (2009) The carboxyl terminusof Brca2 links the disassembly of Rad51 complexes tomitotic entry. Curr Biol 19: 1075–1085.
Bachrati, C.Z., Borts, R.H., and Hickson, I.D. (2006) Mobiled-loops are a preferred substrate for the Bloom’s syndromehelicase. Nucleic Acids Res 34: 2269–2279.
Bastin-Shanower, S.A., Fricke, W.M., Mullen, J.R., and Brill,
S.J. (2003) The mechanism of Mus81-Mms4 cleavage siteselection distinguishes it from the homologous endonu-clease Rad1-Rad10. Mol Cell Biol 23: 3487–3496.
van Brabant, A.J., Ye, T., Sanz, M., German, I.J., Ellis, N.A.,and Holloman, W.K. (2000) Binding and melting of d-loopsby the Bloom syndrome helicase. Biochemistry 39: 14617–14625.
Budzowska, M., and Kanaar, R. (2009) Mechanisms ofdealing with DNA damage-induced replication problems.Cell Biochem Biophys 53: 17–31.
Bugreev, D.V., Yu, X., Egelman, E.H., and Mazin, A.V. (2007)Novel pro- and anti-recombination activities of the Bloom’ssyndrome helicase. Genes Dev 21: 3085–3094.
Carreira, A., Hilario, J., Amitani, I., Baskin, R.J., Shivji, M.K.,Venkitaraman, A.R., and Kowalczykowski, S.C. (2009) TheBRC repeats of BRCA2 modulate the DNA-binding selec-tivity of RAD51. Cell 136: 1032–1043.
Davies, B.W., Kohanski, M.A., Simmons, L.A., Winkler, J.A.,Collins, J.J., and Walker, G.C. (2009) Hydroxyurea induceshydroxyl radical-mediated cell death in Escherichia coli.Mol Cell 36: 845–860.
Davies, O.R., and Pellegrini, L. (2007) Interaction with theBRCA2 C terminus protects RAD51-DNA filaments fromdisassembly by BRC repeats. Nat Struct Mol Biol 14: 475–483.
Esashi, F., Galkin, V.E., Yu, X., Egelman, E.H., and West,S.C. (2007) Stabilization of RAD51 nucleoprotein filamentsby the C-terminal region of BRCA2. Nat Struct Mol Biol 14:468–474.
Fabre, F., Chan, A., Heyer, W.D., and Gangloff, S. (2002)Alternate pathways involving Sgs1/Top3, Mus81/Mms4,and Srs2 prevent formation of toxic recombination interme-diates from single-stranded gaps created by DNAreplication. Proc Natl Acad Sci USA 99: 16887–16892.
Gangloff, S., Soustelle, C., and Fabre, F. (2000) Homologousrecombination is responsible for cell death in the absenceof the Sgs1 and Srs2 helicases. Nat Genet 25: 192–194.
Hashimoto, Y., Chaudhuri, A.R., Lopes, M., and Costanzo, V.(2010) Rad51 protects nascent DNAfrom Mre11-dependentdegradation and promotes continuous DNA synthesis. NatStruct Mol Biol 17: 1305–1311.
Heyer, W.D., Ehmsen, K.T., and Liu, J. (2010) Regulation ofhomologous recombination in eukaryotes. Annu RevGenet 44: 113–139.
Holliday, R. (1966) Studies on mitotic gene conversion inUstilago. Genet Res 8: 323–337.
Holloman, W.K. (2011) Unraveling the mechanism of BRCA2in homologous recombination. Nat Struct Mol Biol 18: 748–754.
Holloman, W.K., Schirawski, J., and Holliday, R. (2008) Thehomologous recombination system of Ustilago maydis.Fungal Genet Biol 45 (Suppl. 1): S31–S39.
Hope, J.C., Maftahi, M., and Freyer, G.A. (2005) A postsyn-aptic role for Rhp55/57 that is responsible for cell death inDrqh1 mutants following replication arrest in Schizosaccha-romyces pombe. Genetics 170: 519–531.
Jensen, R.B., Carreira, A., and Kowalczykowski, S.C. (2010)Purified human BRCA2 stimulates RAD51-mediatedrecombination. Nature 467: 678–683.
Kagawa, W., Kurumizaka, H., Ishitani, R., Fukai, S., Nureki,O., Shibata, T., and Yokoyama, S. (2002) Crystal structure
360 M. Kojic and W. K. Holloman �
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361

of the homologous-pairing domain from the human Rad52recombinase in the undecameric form. Mol Cell 10: 359–371.
Kojic, M., Kostrub, C.F., Buchman, A.R., and Holloman, W.K.(2002) BRCA2 homolog required for proficiency in DNArepair, recombination, and genome stability in Ustilagomaydis. Mol Cell 10: 683–691.
Kojic, M., Yang, H., Kostrub, C.F., Pavletich, N.P., and Hollo-man, W.K. (2003) The BRCA2-interacting protein DSS1 isvital for DNA repair, recombination, and genome stability inUstilago maydis. Mol Cell 12: 1043–1049.
Kojic, M., Zhou, Q., Lisby, M., and Holloman, W.K. (2005)Brh2-Dss1 interplay enables properly controlled recombi-nation in Ustilago maydis. Mol Cell Biol 25: 2547–2557.
Kojic, M., Mao, N., Zhou, Q., Lisby, M., and Holloman, W.K.(2008) Compensatory role for Rad52 during recombina-tional repair in Ustilago maydis. Mol Microbiol 67: 1156–1168.
Kojic, M., Zhou, Q., Fan, J., and Holloman, W.K. (2011)Mutational analysis of Brh2 reveals requirements for com-pensating mediator functions. Mol Microbiol 79: 180–191.
Lehmann, A.R., and Fuchs, R.P. (2006) Gaps and forks inDNA replication: rediscovering old models. DNA Repair(Amst) 5: 1495–1498.
Liberi, G., Maffioletti, G., Lucca, C., Chiolo, I., Baryshnikova,A., Cotta-Ramusino, C., et al. (2005) Rad51-dependentDNA structures accumulate at damaged replication forks insgs1 mutants defective in the yeast ortholog of BLM RecQhelicase. Genes Dev 19: 339–350.
Lomonosov, M., Anand, S., Sangrithi, M., Davies, R., andVenkitaraman, A.R. (2003) Stabilization of stalled DNA rep-lication forks by the BRCA2 breast cancer susceptibilityprotein. Genes Dev 17: 3017–3022.
Lopes, M., Foiani, M., and Sogo, J.M. (2006) Multiple mecha-nisms control chromosome integrity after replication forkuncoupling and restart at irreparable UV lesions. Mol Cell21: 15–27.
Mankouri, H.W., Ashton, T.M., and Hickson, I.D. (2011) Hol-liday junction-containing DNA structures persist in cellslacking Sgs1 or Top3 following exposure to DNA damage.Proc Natl Acad Sci USA 108: 4944–4949.
Mao, N., Kojic, M., and Holloman, W.K. (2009) Role of Blmand collaborating factors in recombination and survival fol-lowing replication stress in Ustilago maydis. DNA Repair(Amst) 8: 752–759.
Mimitou, E.P., and Symington, L.S. (2009) Nucleases andhelicases take center stage in homologous recombination.Trends Biochem Sci 34: 264–272.
Petermann, E., Orta, M.L., Issaeva, N., Schultz, N., andHelleday, T. (2010) Hydroxyurea-stalled replication forksbecome progressively inactivated and require two different
RAD51-mediated pathways for restart and repair. Mol Cell37: 492–502.
Roman, H., and Jacob, F. (1958) A comparison of spontane-ous and ultraviolet-induced allelic recombinaiton withreference to the recombination of outside markers. ColdSpring Harb Symp Quant Biol 23: 155–160.
Saintigny, Y., Delacote, F., Vares, G., Petitot, F., Lambert,S., Averbeck, D., and Lopez, B.S. (2001) Characteriza-tion of homologous recombination induced by replicationinhibition in mammalian cells. EMBO J 20: 3861–3870.
San Filippo, J., Sung, P., and Klein, H. (2008) Mechanism ofeukaryotic homologous recombination. Annu Rev Biochem77: 229–257.
Schlacher, K., Christ, N., Siaud, N., Egashira, A., Wu, H., andJasin, M. (2011) Double-strand break repair-independentrole for BRCA2 in blocking stalled replication fork degra-dation by MRE11. Cell 145: 529–542.
Shivji, M.K., Mukund, S.R., Rajendra, E., Chen, S., Short,J.M., Savill, J., et al. (2009) The BRC repeats of humanBRCA2 differentially regulate RAD51 binding on single-versus double-stranded DNA to stimulate strand exchange.Proc Natl Acad Sci USA 106: 13254–13259.
Singleton, M.R., Wentzell, L.M., Liu, Y., West, S.C., andWigley, D.B. (2002) Structure of the single-strand anneal-ing domain of human RAD52 protein. Proc Natl Acad SciUSA 99: 13492–13497.
Thorslund, T., and West, S.C. (2007) BRCA2: a universalrecombinase regulator. Oncogene 26: 7720–7730.
Timson, J. (1975) Hydroxyurea. Mutat Res 32: 115–132.Wu, L., and Hickson, I.D. (2003) The Bloom’s syndrome
helicase suppresses crossing over during homologousrecombination. Nature 426: 870–874.
Yang, H., Jeffrey, P.D., Miller, J., Kinnucan, E., Sun, Y.,Thoma, N.H., et al. (2002) BRCA2 function in DNA bindingand recombination from a BRCA2-DSS1-ssDNA structure.Science 297: 1837–1848.
Yang, H., Li, Q., Fan, J., Holloman, W.K., and Pavletich, N.P.(2005) The BRCA2 homologue Brh2 nucleates RAD51filament formation at a dsDNA-ssDNA junction. Nature433: 653–657.
Zhou, Q., Kojic, M., Cao, Z., Lisby, M., Mazloum, N.A., andHolloman, W.K. (2007) Dss1 interaction with Brh2 as aregulatory mechanism for recombinational repair. Mol CellBiol 27: 2512–2526.
Zhou, Q., Kojic, M., and Holloman, W.K. (2009a) DNA-binding domain within the Brh2 N terminus Is the primaryinteraction site for association with DNA. J Biol Chem 284:8265–8273.
Zhou, Q., Mazloum, N., Mao, N., Kojic, M., and Holloman,W.K. (2009b) Dss1 regulates interaction of Brh2 with DNA.Biochemistry 48: 11929–11938.
Brh2 DNA-binding domain function 361
© 2011 Blackwell Publishing Ltd, Molecular Microbiology, 83, 351–361



![HC[Final]-BLM and Paul Kruger, Gauteng.pdf](https://static.fdokumen.com/doc/165x107/63236790be5419ea700e96f1/hcfinal-blm-and-paul-kruger-gautengpdf.jpg)
















