
Midbrain expression of Delta-like 1 homologue is regulated by GDNF and is associated with...
11
Midbrain expression of Delta-like 1 homologue is regulated by GDNF and is associated with dopaminergic differentiation Nicolaj S. Christophersen a,b, ⁎ , Mette Grønborg b , Thomas N. Petersen c , Lone Fjord-Larsen b , Jesper R. Jørgensen b , Bengt Juliusson b , Nikolaj Blom c , Carl Rosenblad b,1 , Patrik Brundin a a Neuronal Survival Unit, Department of Experimental Medical Science, Wallenberg Neuroscience Center, Lund University, SE-22184 Lund, Sweden b NsGene A/S, Baltorpvej 154, DK-2750 Ballerup, Denmark c Center for Biological Sequence Analysis, The Technical University of Denmark, DK-2800 Lyngby, Denmark Received 1 May 2006; revised 2 January 2007; accepted 8 January 2007 Available online 24 January 2007 Abstract Affymetrix GeneChip technology and quantitative real-time PCR (Q-PCR) were used to examine changes in gene expression in the adult murine substantia nigra pars compacta (SNc) following lentiviral glial cell line-derived neurotrophic factor (GDNF) delivery in adult striatum. We identified several genes that were upregulated after GDNF treatment. Among these, the gene encoding the transmembrane protein Delta-like 1 homologue (Dlk1) was upregulated with a greater than 4-fold increase in mRNA encoding this protein. Immunohistochemistry with a Dlk1- specific antibody confirmed the observed upregulation with increased positive staining of cell bodies in the SNc and fibers in the striatum. Analysis of the developmental regulation of Dlk1 in the murine ventral midbrain showed that the upregulation of Dlk1 mRNA correlated with the generation of tyrosine hydroxylase (TH)-positive neurons. Furthermore, Dlk1 expression was analyzed in MesC2.10 cells, which are derived from embryonic human mesencephalon and capable of undergoing differentiation into dopaminergic neurons. We detected upregulation of Dlk1 mRNA and protein under conditions where MesC2.10 cells differentiate into a dopaminergic phenotype (41.7 ± 7.1% Dlk1+ cells). In contrast, control cultures subjected to default differentiation into non-dopaminergic neurons only expressed very few (3.7 ± 1.3%) Dlk1-immunopositive cells. The expression of Dlk1 in MesC2.10 cells was specifically upregulated by the addition of GDNF. Thus, our data suggest that Dlk1 expression precedes the appearance of TH in mesencephalic cells and that levels of Dlk1 are regulated by GDNF. © 2007 Elsevier Inc. All rights reserved. Keywords: Delta-like 1 homologue; Fetal antigen 1; GDNF; Dopamine; Neural progenitor; Immortalization; Differentiation Introduction Several in vitro and in vivo studies have shown that glial cell line-derived neurotrophic factor (GDNF) exerts distinct neuro- trophic and pharmacological effects on both intact and injured nigrostriatal dopamine (DA) neurons. GDNF may therefore be a useful agent to prevent the neurodegeneration underlying Parkinson's disease (PD), although the results from the clinical trials are controversial (Sherer et al., 2006). Preservation or regeneration of striatal DA axon terminals is crucial for the recovery of motor function (Bjorklund et al., 2000) and is observed following striatal GDNF administration to the DA axon terminals but not when administrated to the cell bodies in the SNc (Kirik et al., 2000a,b; Oo et al., 2003). Delivery of GDNF by lentiviral vectors to the striatum has shown that stable long-term expression of high levels of GDNF can be achieved in rodents and in primates that are sufficient for almost full protection of the DA neurons in the SNc against toxic damage (Bjorklund et al., 2000; Georgievska et al., 2002; Kordower et al., 2000). In addition to providing protection against lesions, GDNF administration leads to increased DA turnover and to increased release of DA and its metabolites (Kirik et al., 2000b; Martin et al., 1996) and GDNF induces sprouting near the site of Experimental Neurology 204 (2007) 791 – 801 www.elsevier.com/locate/yexnr ⁎ Corresponding author. Neuronal Survival Unit, Wallenberg Neuroscience Center, Lund University, BMC A10, 221 84 Lund, Sweden. Fax: +46 46 2220531. E-mail address: [email protected] (N.S. Christophersen). 1 Current address: Department Neurology, Lund University Hospital, SE- 22185 Lund, Sweden. 0014-4886/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2007.01.014
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Midbrain expression of Delta-like 1 homologue is regulated by GDNF and is associated with...

04 (2007) 791–801www.elsevier.com/locate/yexnr
Experimental Neurology 2
Midbrain expression of Delta-like 1 homologue is regulated by GDNF and isassociated with dopaminergic differentiation
Nicolaj S. Christophersen a,b,⁎, Mette Grønborg b, Thomas N. Petersen c, Lone Fjord-Larsen b,Jesper R. Jørgensen b, Bengt Juliusson b, Nikolaj Blom c, Carl Rosenblad b,1, Patrik Brundin a
a Neuronal Survival Unit, Department of Experimental Medical Science, Wallenberg Neuroscience Center, Lund University, SE-22184 Lund, Swedenb NsGene A/S, Baltorpvej 154, DK-2750 Ballerup, Denmark
c Center for Biological Sequence Analysis, The Technical University of Denmark, DK-2800 Lyngby, Denmark
Received 1 May 2006; revised 2 January 2007; accepted 8 January 2007Available online 24 January 2007
Abstract
Affymetrix GeneChip technology and quantitative real-time PCR (Q-PCR) were used to examine changes in gene expression in the adultmurine substantia nigra pars compacta (SNc) following lentiviral glial cell line-derived neurotrophic factor (GDNF) delivery in adult striatum. Weidentified several genes that were upregulated after GDNF treatment. Among these, the gene encoding the transmembrane protein Delta-like 1homologue (Dlk1) was upregulated with a greater than 4-fold increase in mRNA encoding this protein. Immunohistochemistry with a Dlk1-specific antibody confirmed the observed upregulation with increased positive staining of cell bodies in the SNc and fibers in the striatum.Analysis of the developmental regulation of Dlk1 in the murine ventral midbrain showed that the upregulation of Dlk1 mRNA correlated with thegeneration of tyrosine hydroxylase (TH)-positive neurons. Furthermore, Dlk1 expression was analyzed in MesC2.10 cells, which are derived fromembryonic human mesencephalon and capable of undergoing differentiation into dopaminergic neurons. We detected upregulation of Dlk1 mRNAand protein under conditions where MesC2.10 cells differentiate into a dopaminergic phenotype (41.7±7.1% Dlk1+ cells). In contrast, controlcultures subjected to default differentiation into non-dopaminergic neurons only expressed very few (3.7±1.3%) Dlk1-immunopositive cells. Theexpression of Dlk1 in MesC2.10 cells was specifically upregulated by the addition of GDNF. Thus, our data suggest that Dlk1 expression precedesthe appearance of TH in mesencephalic cells and that levels of Dlk1 are regulated by GDNF.© 2007 Elsevier Inc. All rights reserved.
Keywords: Delta-like 1 homologue; Fetal antigen 1; GDNF; Dopamine; Neural progenitor; Immortalization; Differentiation
Introduction
Several in vitro and in vivo studies have shown that glial cellline-derived neurotrophic factor (GDNF) exerts distinct neuro-trophic and pharmacological effects on both intact and injurednigrostriatal dopamine (DA) neurons. GDNFmay therefore be auseful agent to prevent the neurodegeneration underlyingParkinson's disease (PD), although the results from the clinical
⁎ Corresponding author. Neuronal Survival Unit, Wallenberg NeuroscienceCenter, Lund University, BMC A10, 221 84 Lund, Sweden. Fax: +46 462220531.
E-mail address: [email protected] (N.S. Christophersen).1 Current address: Department Neurology, Lund University Hospital, SE-
22185 Lund, Sweden.
0014-4886/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.expneurol.2007.01.014
trials are controversial (Sherer et al., 2006). Preservation orregeneration of striatal DA axon terminals is crucial for therecovery of motor function (Bjorklund et al., 2000) and isobserved following striatal GDNF administration to the DAaxon terminals but not when administrated to the cell bodies inthe SNc (Kirik et al., 2000a,b; Oo et al., 2003). Delivery ofGDNF by lentiviral vectors to the striatum has shown that stablelong-term expression of high levels of GDNF can be achievedin rodents and in primates that are sufficient for almost fullprotection of the DA neurons in the SNc against toxic damage(Bjorklund et al., 2000; Georgievska et al., 2002; Kordower etal., 2000). In addition to providing protection against lesions,GDNF administration leads to increased DA turnover and toincreased release of DA and its metabolites (Kirik et al., 2000b;Martin et al., 1996) and GDNF induces sprouting near the site of

792 N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
administration i.e. globus pallidus in lesioned rats afterprolonged exposure in the striatum (Georgievska et al., 2002).Despite extensive studies of the functional effects of GDNF onDA neurons in vivo very little is understood about the molecularmechanisms underlying the observed effects. The transmem-brane protein Dlk1 is expressed in monoaminergic nuclei in theadult brain, including the SNc and the ventral tegmental area(VTA) (Jensen et al., 2001). The extracellular part of Dlk1corresponds to the soluble protein fetal antigen-1, which wasoriginally isolated from second trimester human amniotic fluid.The soluble form of Dlk1 has been shown to be involved inproliferation and differentiation of various non-neuronal celltypes in autocrine/paracrine/juxtacrine fashions (Jensen et al.,1997; Laborda et al., 1993; Smas and Sul, 1993). The role ofDlk1 in the SNc and VTA is of particular interest in relation toPD, which involves progressive degeneration of DA neurons inthese regions. In our study, we used Affymetrix GeneChiptechnology and Q-PCR to identify genes upregulated in themidbrain by intrastriatal injection of a viral vector expressingGDNF. The mRNA for the Dlk1 gene was identified as one ofthe most upregulated. The developmental expression andregulation of Dlk1 were further characterized in cells derivedfrom a human mesencephalic cell line (MesC2.10) (Lotharius etal., 2002) as they differentiated into DA neurons in the presenceof GDNF. Our results show that Dlk1 is expressed in midbrainneurons just prior to their differentiation into DA neurons andthat the expression is enhanced by GDNF.
Materials and methods
Animals and surgical procedures
All work involving experimental animals was conductedaccording to the guidelines set by the Danish Ethical Committee.Animals were housed in a 12 h light/darkness cycle with accessto food andwater. Female NMRImice (weighing 25 g at the timeof surgery) were used. VSV-G pseudotyped rLV vectors wereproduced as previously described (Rosenblad et al., 2003). Themice were anesthetized using Isofluoran and stereotaxicallyinjected with 0.5 μl rLV-GFP or rLV-GDNF of viral stocksdiluted to equals titers (2×108 TU/ml) along two tracts in theright striatum at the following coordinates (expressed in mmfrom bregma): (i) AP=+0.5, ML=−2.0, DV=−3.5, TB=0.0and (ii) AP=+0.5, ML=−2.0, DV=−3.0, TB=0.0. To preparetissue for Q-PCR, animals were deeply anesthetized withpentobarbital (70 mg/kg), decapitated and the brains quicklydissected out. Animals used for immunohistochemical analysiswere also deeply anesthetized using sodium pentobarbital andperfused transcardially with saline at room temperature for 1 minfollowed by 50 ml ice-cold phosphate-buffered 4% paraformal-dehyde (pH 7.2–7.4).
GDNF ELISA
After removing the brains as described above, tissue wasprepared and GDNF ELISA was carried out as describedpreviously (Georgievska et al., 2002).
Preparation of RNA
For the GeneChip analysis, RNA was prepared from tissuepieces dissected from the brain SNc region (see Fig. 1A). Afterdissection, the tissue pieces were instantly frozen in liquidnitrogen and stored at −80 °C. To generate material for themicroarray analysis, tissue from 3 animals was pooled and totalRNA was extracted using Trizol (Invitrogen) according to themanufacturer's instructions. To concentrate RNA and to removetraces of chromosomal DNA, RNase on column DNAsedigestion with RNase-Free DNase was used (Qiagen) andRNA subsequently eluted in ultra pure H2O. The quality ofRNA samples for GeneChip analysis was assessed using theAgilent Bioanalyzer 2100 and the RNA 6000LABChip. Toextract RNA derived from the ventral mesencephalon ofembryonic day (E) 11.5, E13.5, postnatal day (P) 1 and adultmice, tissue pieces dissected from the ventral mesencephalonwere instantly frozen in liquid nitrogen and stored at −80 °Cbefore using Trizol. For human cell cultures, total RNA wasextracted from either proliferating, 2- and 4-day differentiatedMesC2.10 cells plated in media conditions as indicated. Thequality of the RNA samples for Q-PCR was estimated fromOD260/280 ratio and Agilent Bioanalyzer 2100.
Microarray
RNA samples were amplified according to the Messa-geAmpTM aRNA protocol (Ambion). Briefly, total RNA wasused for first strand cDNA synthesis with a poly-dT primercontaining a T7 RNA-polymerase promoter sequence. Aftersecond strand synthesis, the resulting dsDNA was purified byphenol/chloroform extraction and used for in vitro transcriptionin the presence of biotin-labeled nucleotides to create labeledcRNA. The 2 μg total RNA used as starting material resulted inproduction of approximately 30 μg cRNA. After purificationand fragmentation, 15 μg biotinylated cRNAwas hybridized for16 h at 45 °C to the Affymetrix Mouse Genome 430A Chipcontaining almost 22,600 probe sets representing transcriptsfrom over 14,000 well-characterized mouse genes (www.affymetrix.com). The arrays were washed and subsequentlystained with a streptavidin–phycoerytrin conjugate using theGeneChip Fluidics Station 400. Scanning was done with theAffymetrix GeneArray Scanner, all according to manufacturer'sinstructions (Affymetrix, Santa Clara, CA, USA). Dataacquisition and primary analysis was performed as describedpreviously (Jorgensen et al., 2006). Briefly, two rounds ofhybridizations with independent RNA samples were performed.Each scanning generated a CEL file with measurements ofintensities corresponding to the hybridization of targets with therespective probes. Primary analysis of the datasets was carriedout using the software package GenePublisher (Knudsen et al.,2003). For each probe ID, the Excelfile contained signal values,fold change, p-value and gene annotation. To select genes forfurther analyses, a low-stringency filter was applied. First, theaverage signal values should be larger than 50 to ensure adetectable expression level. Second, the p-value should besmaller than 0.1 to exclude outliers in one of the experiments.
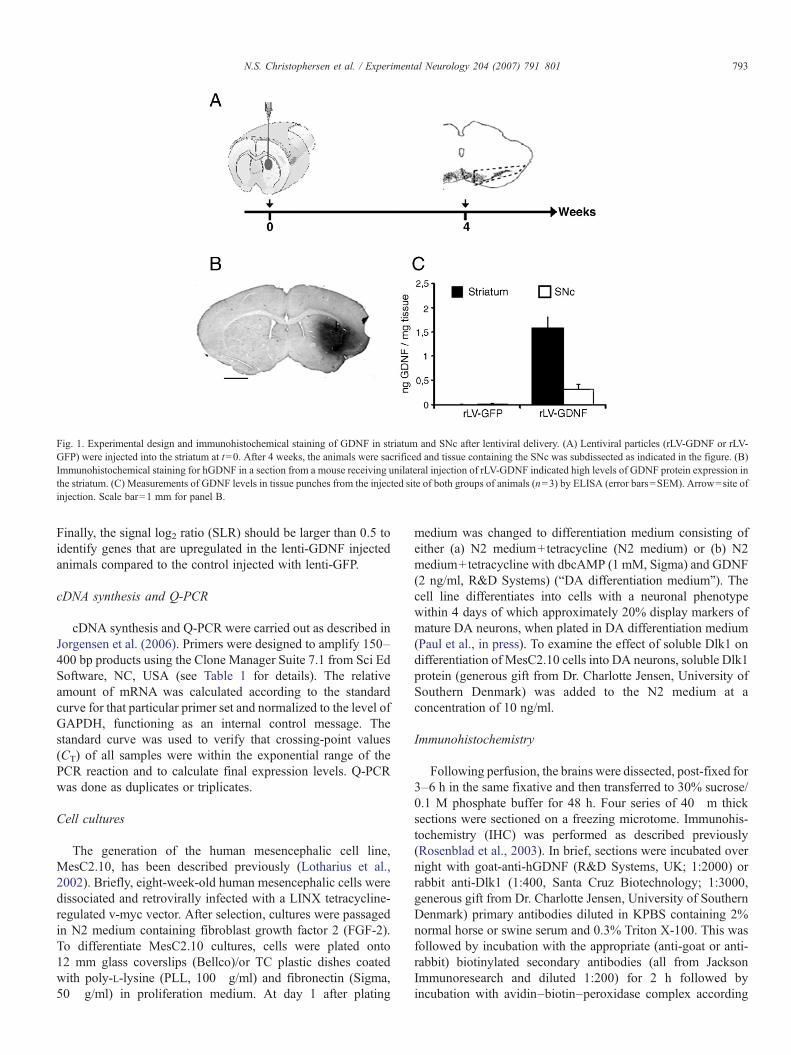
Fig. 1. Experimental design and immunohistochemical staining of GDNF in striatum and SNc after lentiviral delivery. (A) Lentiviral particles (rLV-GDNF or rLV-GFP) were injected into the striatum at t=0. After 4 weeks, the animals were sacrificed and tissue containing the SNc was subdissected as indicated in the figure. (B)Immunohistochemical staining for hGDNF in a section from a mouse receiving unilateral injection of rLV-GDNF indicated high levels of GDNF protein expression inthe striatum. (C) Measurements of GDNF levels in tissue punches from the injected site of both groups of animals (n=3) by ELISA (error bars=SEM). Arrow=site ofinjection. Scale bar=1 mm for panel B.
793N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
Finally, the signal log2 ratio (SLR) should be larger than 0.5 toidentify genes that are upregulated in the lenti-GDNF injectedanimals compared to the control injected with lenti-GFP.
cDNA synthesis and Q-PCR
cDNA synthesis and Q-PCR were carried out as described inJorgensen et al. (2006). Primers were designed to amplify 150–400 bp products using the Clone Manager Suite 7.1 from Sci EdSoftware, NC, USA (see Table 1 for details). The relativeamount of mRNA was calculated according to the standardcurve for that particular primer set and normalized to the level ofGAPDH, functioning as an internal control message. Thestandard curve was used to verify that crossing-point values(CT) of all samples were within the exponential range of thePCR reaction and to calculate final expression levels. Q-PCRwas done as duplicates or triplicates.
Cell cultures
The generation of the human mesencephalic cell line,MesC2.10, has been described previously (Lotharius et al.,2002). Briefly, eight-week-old human mesencephalic cells weredissociated and retrovirally infected with a LINX tetracycline-regulated v-myc vector. After selection, cultures were passagedin N2 medium containing fibroblast growth factor 2 (FGF-2).To differentiate MesC2.10 cultures, cells were plated onto12 mm glass coverslips (Bellco)/or TC plastic dishes coatedwith poly-L-lysine (PLL, 100 μg/ml) and fibronectin (Sigma,50 μg/ml) in proliferation medium. At day 1 after plating
medium was changed to differentiation medium consisting ofeither (a) N2 medium+tetracycline (N2 medium) or (b) N2medium+tetracycline with dbcAMP (1 mM, Sigma) and GDNF(2 ng/ml, R&D Systems) (“DA differentiation medium”). Thecell line differentiates into cells with a neuronal phenotypewithin 4 days of which approximately 20% display markers ofmature DA neurons, when plated in DA differentiation medium(Paul et al., in press). To examine the effect of soluble Dlk1 ondifferentiation of MesC2.10 cells into DA neurons, soluble Dlk1protein (generous gift from Dr. Charlotte Jensen, University ofSouthern Denmark) was added to the N2 medium at aconcentration of 10 ng/ml.
Immunohistochemistry
Following perfusion, the brains were dissected, post-fixed for3–6 h in the same fixative and then transferred to 30% sucrose/0.1 M phosphate buffer for 48 h. Four series of 40 μm thicksections were sectioned on a freezing microtome. Immunohis-tochemistry (IHC) was performed as described previously(Rosenblad et al., 2003). In brief, sections were incubated overnight with goat-anti-hGDNF (R&D Systems, UK; 1:2000) orrabbit anti-Dlk1 (1:400, Santa Cruz Biotechnology; 1:3000,generous gift from Dr. Charlotte Jensen, University of SouthernDenmark) primary antibodies diluted in KPBS containing 2%normal horse or swine serum and 0.3% Triton X-100. This wasfollowed by incubation with the appropriate (anti-goat or anti-rabbit) biotinylated secondary antibodies (all from JacksonImmunoresearch and diluted 1:200) for 2 h followed byincubation with avidin–biotin–peroxidase complex according
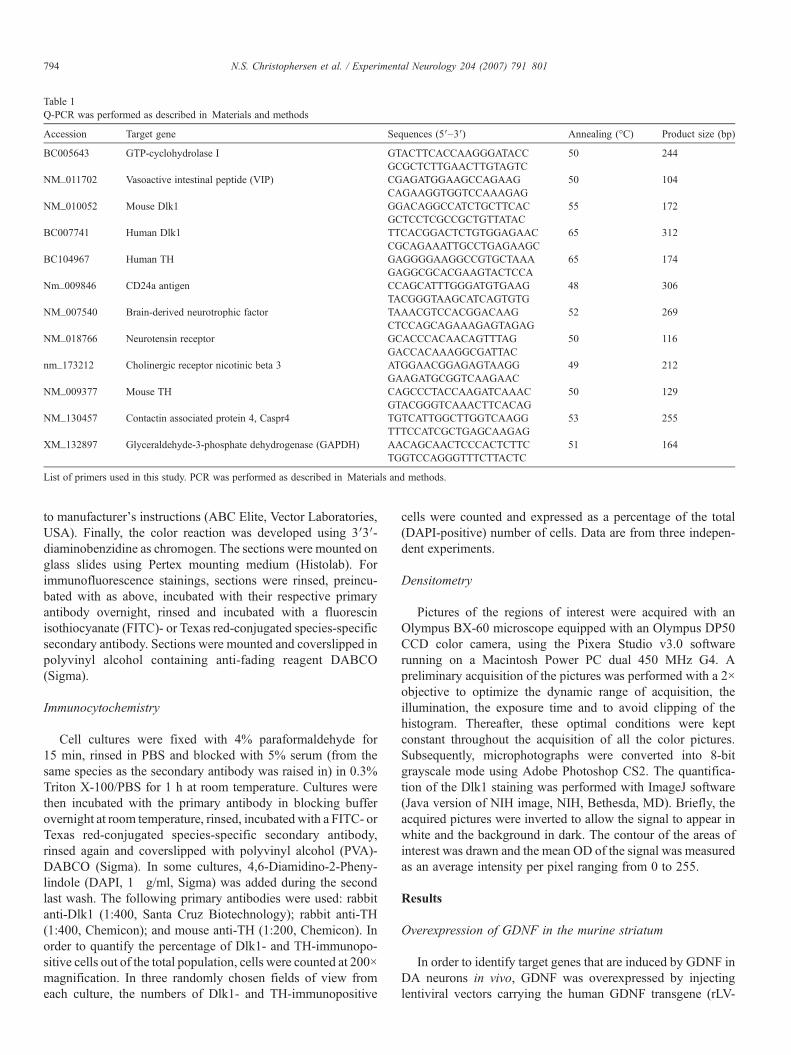
Table 1Q-PCR was performed as described in Materials and methods
Accession Target gene Sequences (5′–3′) Annealing (°C) Product size (bp)
BC005643 GTP-cyclohydrolase I GTACTTCACCAAGGGATACC 50 244GCGCTCTTGAACTTGTAGTC
NM_011702 Vasoactive intestinal peptide (VIP) CGAGATGGAAGCCAGAAG 50 104CAGAAGGTGGTCCAAAGAG
NM_010052 Mouse Dlk1 GGACAGGCCATCTGCTTCAC 55 172GCTCCTCGCCGCTGTTATAC
BC007741 Human Dlk1 TTCACGGACTCTGTGGAGAAC 65 312CGCAGAAATTGCCTGAGAAGC
BC104967 Human TH GAGGGGAAGGCCGTGCTAAA 65 174GAGGCGCACGAAGTACTCCA
Nm_009846 CD24a antigen CCAGCATTTGGGATGTGAAG 48 306TACGGGTAAGCATCAGTGTG
NM_007540 Brain-derived neurotrophic factor TAAACGTCCACGGACAAG 52 269CTCCAGCAGAAAGAGTAGAG
NM_018766 Neurotensin receptor GCACCCACAACAGTTTAG 50 116GACCACAAAGGCGATTAC
nm_173212 Cholinergic receptor nicotinic beta 3 ATGGAACGGAGAGTAAGG 49 212GAAGATGCGGTCAAGAAC
NM_009377 Mouse TH CAGCCCTACCAAGATCAAAC 50 129GTACGGGTCAAACTTCACAG
NM_130457 Contactin associated protein 4, Caspr4 TGTCATTGGCTTGGTCAAGG 53 255TTTCCATCGCTGAGCAAGAG
XM_132897 Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) AACAGCAACTCCCACTCTTC 51 164TGGTCCAGGGTTTCTTACTC
List of primers used in this study. PCR was performed as described in Materials and methods.
794 N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
to manufacturer's instructions (ABC Elite, Vector Laboratories,USA). Finally, the color reaction was developed using 3′3′-diaminobenzidine as chromogen. The sections were mounted onglass slides using Pertex mounting medium (Histolab). Forimmunofluorescence stainings, sections were rinsed, preincu-bated with as above, incubated with their respective primaryantibody overnight, rinsed and incubated with a fluorescinisothiocyanate (FITC)- or Texas red-conjugated species-specificsecondary antibody. Sections were mounted and coverslipped inpolyvinyl alcohol containing anti-fading reagent DABCO(Sigma).
Immunocytochemistry
Cell cultures were fixed with 4% paraformaldehyde for15 min, rinsed in PBS and blocked with 5% serum (from thesame species as the secondary antibody was raised in) in 0.3%Triton X-100/PBS for 1 h at room temperature. Cultures werethen incubated with the primary antibody in blocking bufferovernight at room temperature, rinsed, incubated with a FITC- orTexas red-conjugated species-specific secondary antibody,rinsed again and coverslipped with polyvinyl alcohol (PVA)-DABCO (Sigma). In some cultures, 4,6-Diamidino-2-Pheny-lindole (DAPI, 1 μg/ml, Sigma) was added during the secondlast wash. The following primary antibodies were used: rabbitanti-Dlk1 (1:400, Santa Cruz Biotechnology); rabbit anti-TH(1:400, Chemicon); and mouse anti-TH (1:200, Chemicon). Inorder to quantify the percentage of Dlk1- and TH-immunopo-sitive cells out of the total population, cells were counted at 200×magnification. In three randomly chosen fields of view fromeach culture, the numbers of Dlk1- and TH-immunopositive
cells were counted and expressed as a percentage of the total(DAPI-positive) number of cells. Data are from three indepen-dent experiments.
Densitometry
Pictures of the regions of interest were acquired with anOlympus BX-60 microscope equipped with an Olympus DP50CCD color camera, using the Pixera Studio v3.0 softwarerunning on a Macintosh Power PC dual 450 MHz G4. Apreliminary acquisition of the pictures was performed with a 2×objective to optimize the dynamic range of acquisition, theillumination, the exposure time and to avoid clipping of thehistogram. Thereafter, these optimal conditions were keptconstant throughout the acquisition of all the color pictures.Subsequently, microphotographs were converted into 8-bitgrayscale mode using Adobe Photoshop CS2. The quantifica-tion of the Dlk1 staining was performed with ImageJ software(Java version of NIH image, NIH, Bethesda, MD). Briefly, theacquired pictures were inverted to allow the signal to appear inwhite and the background in dark. The contour of the areas ofinterest was drawn and the mean OD of the signal was measuredas an average intensity per pixel ranging from 0 to 255.
Results
Overexpression of GDNF in the murine striatum
In order to identify target genes that are induced by GDNF inDA neurons in vivo, GDNF was overexpressed by injectinglentiviral vectors carrying the human GDNF transgene (rLV-

Table 2Genes with differential expression between GDNF and GFP groups successfullyconfirmed by Q-PCR
Accession Gene Microarray a Verification b
Mm.29742 CD24a antigen c 8.7–9.3 22±6.4Mm.10651 GTP cyclohydrolase Ic 3.9–5.0 3.8±1.1Mm.157069 Delta-like 1 homologue 4.34 5±0.5Mm.98916 Vasoactive intestinal polypeptide 2.7 1.5±0.4Mm.301712 Neurotensin receptor 1.9 1.9±0.6Mm.41580 RAB3B 1.8 2.5±0.6Mm.110444 Cholinergic receptor, nicotinic, beta
polypeptide 31.8 5.2±0.4
Mm.57199 Ret proto-oncogene 1.7 4.6±2.2Mm.1442 Brain derived neurotrophic factor 1.7 4±0.4Mm.209232 Contactin associated protein 4 1.5 2±0.5Mm.1292 Tyrosine hydroxylase 1.5 2±0.7Mm.88367 GDNF family receptor alpha 1 1.4 2.8±0.5a Ratio between normalized expression levels for GDNF groups and control
(GFP) groups (average values for two independent experiments).b Ratio between expression levels for GDNF groups and control (GFP) groups
(average values of duplicates) normalized to ratio between GAPDH expressionlevels (±SEM).c Gene is represented by more than one probe set.
795N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
GDNF) unilaterally into the striatum of NMRI mice (seeexperimental setup, Fig. 1A). Control animals receivedinjections of lentiviral vectors carrying the GFP transgene(rLV-GFP). Four weeks later the animals were sacrificed andGDNF levels and distribution were analyzed by GDNF ELISAand immunohistochemistry. As seen in Fig. 1B, immunoreac-tive GDNF was widely distributed within an area covering mostof the striatum around the injection site, whereas no stainingwas evident on the control (uninjected) side. We determinedaverage GDNF levels in tissue punches from the injected sideof both groups of animals (n=3) by ELISA. The striatal GDNFlevel in animals injected with the rLV-GDNF was 1.6 ngGDNF/mg tissue, which was significantly higher than thebarely detectable level (0.001 ng/mg) found in the striatum ofcontrol animals (Kruskal–Wallis one-way analysis, p<0.05)(Fig. 1C). Increased level of GDNF was also detected in theSNc of the rLV-GDNF injected animals (0.3 ng GDNF ng/mgtissue) compared to the SNc in the control animals (0.012 ngGDNF ng/mg tissue) confirming retrograde transport of GDNFalong the striatonigral pathway to SN (Fig. 1C). Together, thesedata demonstrated a significant overexpression of GDNF in thestriatum after lentiviral delivery.
Microarray analysis of GDNF-treated SNc
To gain insight into the molecular mechanisms of GDNF-induced effects on the dopaminergic neurons in the SNc, wecompared nigral gene expression profiles from animals injectedwith rLV-GDNF and control animals injected with rLV-GFP.Since we intended to use the microarray technology merely toidentify candidate genes for further studies, only two indepen-dent replicates were performed. As the array data were notstatistically significant after correction for multiple testing, weapplied a low stringency filter taking into account expressionlevel, fold change and significance. This approach has beendemonstrated to be an effective alternative analysis (Jorgensenet al., 2006). In our case, array data were sorted as follows: first,the average signal values should be larger than 50 to ensure adetectable expression level. Second, the p-value should besmaller than 0.1 to exclude the most extreme outliers. Finally,the signal log2 ratio (SLR) should be larger than 0.5 to identifygenes that are upregulated in the lenti-GDNF injected animalscompared to the control injected with lenti-GFP. Using thisapproach, 77 probes representing 70 genes were identified. Theprobes were arranged with descending fold changes and can befound as Supplementary data.
In order to validate the microarray analysis, eighteen geneswere selected for verification by Q-PCR. A number of thesewere considered to be relevant for the understanding of GDNFeffects in the nigrostriatal system and represent probes both fromthe top and the bottom of the list of regulated genes. Usingmaterial derived from independent biological samples, it waspossible to verify a difference in expression levels between thetwo groups for 12 of the selected genes (shown in Table 2).Verification of two of the selected genes failed because theexpression level was below the detection limit, whereas nodifference in expression levels could be detected for four of the
selected genes. Hence, approximately two-thirds of genesselected from the array could be verified validating the arrayexperiment. A complete list of the upregulated genes identifiedin this study is provided as Supplementary data.
Expression of Dlk1 in the lentiviral GDNF-treated adultmurine midbrain
As seen in Table 2, GeneChip analysis identified the mRNAfor the Dlk1 gene as one of the most upregulated (4.3-foldupregulation in the GDNF group). This finding was confirmedby Q-PCR showing 5-fold upregulation. Dlk1 has previouslybeen shown to be expressed in SNc (Jensen et al., 2001), but themolecular mechanisms regulating Dlk1 expression in the brainare unknown, so we chose to analyze further the expression andregulation of Dlk1. The effect of rLV-GDNF injections on Dlk1protein levels was analyzed by IHC. IHC with an anti-rat Dlk1antibody showed that unilateral overexpression of GDNF in thestriatum upregulated Dlk1 protein in fibers innervating thestriatum (Dlk1-immunoreactive fibers, Fig. 2A) as well asexpression in neurons of the SNc (Figs. 2B, D, cellularstaining). The upregulation of Dlk1 protein by rLV-GDNF wasconfirmed by densitometric measurements of the levels of Dlk1staining in cell bodies in the SNc (Fig. 2I). The Dlk1 proteinwas indeed co-expressed with TH in SNc neurons in the adultbrain (Figs. 2E–H).
Analysis of Dlk1 gene during the development of murineventral midbrain
In order to analyze the developmental regulation of Dlk1 invivo, we performed Q-PCR analysis of cDNAs derived fromdifferent developmental stages of the murine midbrain (Figs.3A–B). The expression of Dlk1 in the ventral mesencephalonreached the highest level between E13.5 and P1 in the mouse.

Fig. 2. Immunohistochemical staining of Dlk1-positive fibers in the striatum (A) and Dlk1-positive cell bodies in the SNc from an rLV-GDNF injected animal (B–D).(E–H) Double labeling with Dlk1 and TH showed co-localization in sections of mouse mesencephalon. (I) Optical density measurements of the levels of Dlk1 stainingin cell bodies found in the SNc in the ipsi- or contralateral side of injection of LV-GDNF or LV-GFP. *p<0.05 (Student's t-test). Arrow=site of injection. Scalebar=1 mm for panels A and B, 62.5 μm in panels C–F and 50 μm in panels G and H.
796 N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801

Fig. 3. Expression of Dlk1 mRNA and protein during embryonic midbrain development. Q-PCR analysis of Dlk1 (A) and TH (B) using cDNA derived from the ventralmesencephalon subdissected from different gestational ages (embryonic=E, postnatal=P, adult=Ad). Expression was normalized to GAPDH mRNA expression andshown as expression relative to the lowest level of expression detected. CT values ranged from 18.1 to 20.7 for Dlk1 and 18.2 to 23.1 for TH. Error bars=SEM. (C–F)Coronal sections through the VM of E11.5 and E13.5 mouse showed that Dlk1 protein co-localized with TH in developing midbrain DA neurons. Scale bar=1 mm.
797N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
For comparison, the developmental expression of TH was alsoanalyzed in the same material. As expected, this analysisrevealed that TH mRNA levels increase during embryonic andpostnatal development and then reach a plateau in the adultmouse ventral midbrain. Using IHC, we found that Dlk1 proteinwas co-localized with TH in developing midbrain DA neuronsin E11.5 and E13.5 mouse embryos (Figs. 3C–F).
Upregulation of the Dlk1 protein upon DA differentiation ofMesC2.10 cells
In order to further characterize the association of Dlk1expression with DA differentiation, we used a clonal cell line ofhuman mesencephalic DA progenitors immortalized with v-myc as a model system (Lotharius et al., 2002). In these cells, atetracycline-controlled transactivator activates transcription ofthe v-myc from a hybrid CMV promoter, allowing the culturedMesC2.10 cells to proliferate continuously in serum-free N2medium containing FGF-2. Proliferating MesC2.10 cells can be
differentiated into neurons by replating the proliferationmedium with N2 medium supplemented with tetracycline. IfdbcAMP and GDNF are also added to the culture medium,approximately 20% of MesC2.10 cells differentiate specificallyinto TH-expressing neurons (Fig. 4A). To determine whetherDlk1 expression is associated with TH expression, MesC2.10cells were analyzed for the expression of Dlk1 protein. Dlk1protein was detected in 41.7±7.1% of the MesC2.10 cellsgrown under conditions to promote differentiation into DAneurons (dbcAMP and GDNF present) (Fig. 4B). In contrast,cells differentiated in N2 medium alone expressed Dlk1 in only3.7±1.3% of the cells. Over 98% of the cells staining positivelyfor TH also expressed Dlk1 (Figs. 4C–F).
Time-course of Dlk1 expression in relation to dopaminergicdifferentiation
MesC2.10 cells were differentiated in DA differentiationmedium for 2–4 days and harvested for Q-PCR analysis of Dlk1

Fig. 4. Expression of TH and Dlk1 in MesC2.10 cell cultures after 4 days of differentiation. Cultures of the human MesC2.10 cell line were plated on PLL/fibronectincoated plates in N2 medium with dbcAMP and GDNF. Cells were stained for TH (red) (A) or Dlk1 (red) (B) and counterstained using the nuclear maker DAPI (blue).(C–F) All neurons that stained positive for mouse antibody against TH (green) were also Dlk1+ (red).
Fig. 5. Upregulation of Dlk1 mRNA in MesC2.10 cell cultures undergoing DAdifferentiation. (A) Q-PCR analysis of Dlk1 and TH expression in proliferating,2-day and 4-day differentiated MesC2.10 cells plated in DA differentiationmedium showed that Dlk1 was upregulated already after 2 days of DAdifferentiation of MesC2.10 cells as compared to proliferating cells and withhigher expression levels after 4 days of differentiation. *p<0.05 (Student's t-test). (B) Single addition of GDNF to the N2 medium resulted in significantupregulation of Dlk1 mRNA in MesC2.10 cells after 2 days of differentiationrelative to cultures following 2 days of differentiation in N2 medium only.Expression was normalized to GAPDH mRNA expression and shown asexpression relative to the lowest level of expression detected. *p<0.05(Student's t-test). CT values ranged from 20.3 to 29.9 for Dlk1 and 24.1 to35.3 for TH. Prol=proliferating, diff=DA differentiation medium, d=days ofdifferentiation, N2=N2 medium. Error bars=SEM.
798 N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
expression. Q-PCR analysis of differentiating MesC2.10 cellsshowed that Dlk1 is upregulated at day 2 of differentiationbefore the onset of TH expression at day 4 (Fig. 5A). WhenMesC2.10 cells were plated in N2 medium with the additionalsupplementation of GDNF (2 ng/ml) only, the level of Dlk1mRNA increased significantly (16-fold) compared to cellcultures differentiated in N2 medium only (Fig. 5B).
Discussion
In this study, we show that GDNF stimulates the expressionof Dlk1 both in vivo and in vitro. To understand the molecularmechanisms underlying GDNF-induced effects observed invivo, we compared gene expression profiles in the SNc betweenmice overexpressing GDNF in the striatum with control animalsusing Affymetrix GeneChip technology. To our knowledgegene profiling studies addressing the mechanisms of GDNF inthis neuronal population in vivo have not been published so far,although studies comparing the transcriptional events in intactand damaged SNc have been reported (Mandel et al., 2002;Miller et al., 2004). Using Affymetrix GeneChip technology, wefound Dlk1 to be one of the most upregulated genes in the SNcfollowing GDNF overexpression in the striatum, which wasverified also at the protein level. During development, the timeof onset of Dlk1 expression in midbrain DA neurons correlatedwith the onset of the expression of the DA-synthesizing enzymeTH. After birth, the expression of Dlk1 decreased dramaticallyin the mesencephalon, but it still remained detectable in theadult. In vitro, Dlk1 expression was upregulated before TH

799N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
could be detected in the MesC2.10 cell line, when the cells werecultured under conditions that promote differentiation into DAneurons. Thus, Dlk1 expression is associated with DAdifferentiation and precedes the appearance of TH neurons inmesencephalic cells. Earlier reports have described regulatedDlk1 expression to be associated with differentiation inneuronal and non-neuronal cell lines. Neuroblastoma cells candifferentiate into a neuronal phenotype in response to retinoicacid or nerve growth factor (NGF). Treatment with glucocorti-coids, on the other hand, promotes differentiation of neuro-blastoma cells into a neuroendocrine chromaffin-likephenotype. High Dlk1 expression is associated with differentia-tion of neuroblastoma cells into chromaffin cells (Van Limpt etal., 2003). Interestingly, glucocorticoids and treatment withdbcAMP both increase Dlk1 expression in neuroblastoma cells,whereas Dlk1 levels are decreased in neuroblastoma cellstreated with NGF or retinoic acid (Gaetano et al., 1992; Helmanet al., 1987). In pancreatic β-cells, growth hormone can induceDlk1 gene expression, cell proliferation and insulin production(Carlsson et al., 1997). The functional role of Dlk1 in the brain,however, is still not clear, and the molecular mechanisms ofhow Dlk1 exerts its biological effects are unknown. Thedevelopmental expression profile of Dlk1 in the SNc and theinduction of Dlk1 expression by GDNF suggest that the proteinmay play a role in mediating the regenerative and/orpharmacological effect of GDNF. We did not observe anyeffect on TH expression when adding soluble Dlk1 protein(10 ng/ml) to the N2 medium in MesC2.10 cells (data notshown), but this does not exclude a role of the membrane boundform(s) of Dlk1 in the differentiation of mesencephalic DAneurons. Alternatively, Dlk1 mediated effects in DA neuronsmay require cofactors co-induced by GDNF. In 3T3-L1preadipocytes undergoing differentiation into adipocytes,secreted splice variants of Dlk1 are downregulated while theexpression of the membrane-associated forms of Dlk1 isupregulated early in the differentiation process (Garces et al.,1999). Thus, the ratio between the soluble and membrane-associated forms is crucial in determining the effect of theprotein (Smas et al., 1997). In agreement with our data, a recentconference abstract reported that overexpression of full-lengthDlk1 does not have any effect on mouse DA progenitors or theirdifferentiated neurons (Bauer et al., 2005). On the other hand,Bauer and coworkers observed that silencing of Dlk1 expres-sion in undifferentiated DA progenitors resulted in decreasedTH mRNA, TH protein, and numbers of TH-immunopositiveneurons following differentiation. The expression of othermarkers of the DAergic lineage, including the GDNF receptorRet, DA transporter and vesicular monoamine transporter 2,was also decreased. These preliminary data indicate that Dlk1indeed contributes to DA maturation and has a functional role inthe generation of midbrain DA neurons. Interestingly, Mar-uyama et al. (Genbank accession no. D16847) have described agene encoding for stromal cell-derived protein-1 (SCP1) in theadipocytic stromal cell line PA6 that is identical to mouse Dlk1.The stromal cell line PA6 has the ability to induce differentia-tion of embryonic stem cells into DA neurons, and this effect isthought to be due to both soluble and cell-surface bound factor
(s) (Kawasaki et al., 2000). Although we have no directevidence, the fact that SCP1 is expressed in PA6 cells opens upthe possibility that Dlk1 could contribute to the effects of PA6cells on embryonic stem cells.
The majority of the observed effects of GDNF on geneexpression in the SNc are likely to be mediated by a receptorcomplex in the neurons consisting of GFRα1 (providing ligandbinding specificity) and Ret (providing intracellular signaling)(Airaksinen et al., 1999). Our data indicate an increasedexpression of mRNA levels of both receptor components in theanimals injected with rLV-GDNF. We speculate that Dlk1expression would be significantly downregulated in themidbrain of GDNF receptor knock out mice. Interestingly, ourdata indicate that BDNF may mediate some of the effects ofGDNF on nigrostriatal DA neurons. There was a 4-foldupregulation in BDNF mRNA levels in the rLV-GDNF injectedanimals. In cultures established from the ventral mesencepha-lon, BDNF can induce many of the effects also ascribed toGDNF, including those on differentiation, survival andincreases in DA release (Baquet et al., 2005; Studer et al.,1995, 1996). We observed 2-fold upregulation of TH mRNAlevel in the rLV-GDNF injected animals compared with controlanimals. We also performed immunostaining for TH, but did notfind any difference in TH protein levels between the rLV-GDNFinjected and untreated sides (data not shown). In previousstudies it has been reported that striatal overexpression ofGDNF using viral vectors in rats leads to downregulation of THmRNA and protein expression in the SNc and the striatum(Georgievska et al., 2004; Rosenblad et al., 2003). The down-regulation of TH is thought to occur as an adaptation to asustained activation of DA turnover induced by GDNFstimulation. The phenomenon can be observed both in animalswith intact and damaged nigrostriatal systems, when the level ofGDNF in the striatum exceeds 0.7 ng/mg tissue. We did not,however, find evidence of TH mRNA downregulation in ourstudy despite high striatal expression levels of GDNF. Theactivation of TH is critically dependent on tetrahydrobiopterin(BH4) as a cofactor. GTP cyclohydrolase I catalyzes the rate-limiting step of BH4 synthesis. Our data indicated a 4- to 5-foldupregulation of GTP cyclohydrolase I mRNA levels in rLV-GDNF injected mice. Thus, the increase in DA productioncaused by GDNF administration may be due to increased THactivity, in turn related to increased synthesis of the essentialcofactor BH4.
In other studies, oligonucleotide microarrays have been usedto analyze gene regulation by GDNF signaling in dorsal rootganglia (Linnarsson et al., 2001) and in immortalized striatalprogenitor cells (Pahnke et al., 2004). Linnarsson and coworkersshowed that embryonic and postnatal dorsal root ganglia differin their transcriptional program in response to GDNF stimula-tion, despite GDNF receptors being present at both stages. Mostinterestingly, this study showed a suppressive effect of GDNF onneurite growth and/or branching in this system (Linnarsson etal., 2001). On the other hand, Pahnke et al. observed that GDNFupregulated genes promoting axonal sprouting, neurite out-growth and spine formation in the immortalized striatal cell lineST14A (Pahnke et al., 2004). Comparison of the studies on

800 N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
GDNF-mediated transcriptional changes indicates that GDNF-induced changes depend on several factors such as the type ofneurons studied, the concentration of GDNF and the duration ofGDNF exposure. Our experimental setup was designedspecifically to identify GDNF-mediated transcriptional changesin midbrain SNc neurons in vivo. We think that the role of GDNFin this neuronal population is particularly interesting in relationto normal development and to human disease. Due to theimportance of duration of exposure, we cannot exclude that ourresults would have differed had we expressed GDNF for shorteror longer time periods.
Currently, no single, stage-specific marker exists that canidentify the progenitors of DA neurons and their progeny in amixed cell population. Preferably, such markers should bemembrane-localized proteins. Dlk1 might be a potentialcandidate for such a marker in addition to the cell surfaceglycoprotein CD24a antigen identified as upregulated in ourmicroarray study. In line with our results, a recent paper has useda microarray approach to identify genes highly expressed in SNcshowing high levels of CD24 RNA in adult SNc neurons (Chunget al., 2005). Development of protocols for fluorescenceactivated cell sorting using antibodies raised against Dlk1 mayenable enrichment of DA neurons derived from humanembryonic stem (ES) cells or to increase the proportion of DAneurons in donor tissue prior to intracerebral transplantation.
Acknowledgments
The work was supported by EU grants QLG3-CT-2002-01000 and LSHBCT-2003-503005 and the Swedish ResearchCouncil (K2002-99SX-14472-01A), the Crafoord Foundationand the Swedish Parkinson Foundation. Nicolaj S. Christo-phersen is supported by a grant from the Danish Academy ofTechnical Sciences.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.expneurol.2007.01.014.
References
Airaksinen, M.S., Titievsky, A., Saarma, M., 1999. GDNF family neurotrophicfactor signaling: four masters, one servant? Mol. Cell. Neurosci. 13,313–325.
Baquet, Z.C., Bickford, P.C., Jones, K.R., 2005. Brain-derived neurotrophicfactor is required for the establishment of the proper number ofdopaminergic neurons in the substantia nigra pars compacta. J. Neurosci.25, 6251–6259.
Bauer, M., Meixner, A., Ueffing, M., 2005. Investigation of Developmental andDisease-Related Genes in Primary Neuronal Systems using LentiviralVector-Based Technology. NGFN, Bonn, Germany.
Bjorklund, A., Kirik, D., Rosenblad, C., Georgievska, B., Lundberg, C.,Mandel, R.J., 2000. Towards a neuroprotective gene therapy for Parkinson'sdisease: use of adenovirus, AAV and lentivirus vectors for gene transfer ofGDNF to the nigrostriatal system in the rat Parkinson model. Brain Res. 886,82–98.
Carlsson, C., Tornehave, D., Lindberg, K., Galante, P., Billestrup, N.,Michelsen, B., Larsson, L.I., Nielsen, J.H., 1997. Growth hormone and
prolactin stimulate the expression of rat preadipocyte factor-1/delta-likeprotein in pancreatic islets: molecular cloning and expression pattern duringdevelopment and growth of the endocrine pancreas. Endocrinology 138,3940–3948.
Chung, C.Y., Seo, H., Sonntag, K.C., Brooks, A., Lin, L., Isacson, O., 2005. Celltype specific gene expression of midbrain dopaminergic neurons revealsmolecules involved in their vulnerability and protection. Hum. Mol. Genet.14, 1709–1725.
Gaetano, C., Matsumoto, K., Thiele, C.J., 1992. In vitro activation of distinctmolecular and cellular phenotypes after induction of differentiation in ahuman neuroblastoma cell line. Cancer Res. 52, 4402–4407.
Garces, C., Ruiz-Hidalgo, M.J., Bonvini, E., Goldstein, J., Laborda, J., 1999.Adipocyte differentiation is modulated by secreted delta-like (dlk) variantsand requires the expression of membrane-associated dlk. Differentiation 64,103–114.
Georgievska, B., Kirik, D., Rosenblad, C., Lundberg, C., Bjorklund, A., 2002.Neuroprotection in the rat Parkinson model by intrastriatal GDNF genetransfer using a lentiviral vector. NeuroReport 13, 75–82.
Georgievska, B., Kirik, D., Bjorklund, A., 2004. Overexpression of glial cellline-derived neurotrophic factor using a lentiviral vector induces time- anddose-dependent downregulation of tyrosine hydroxylase in the intactnigrostriatal dopamine system. J. Neurosci. 24, 6437–6445.
Helman, L.J., Thiele, C.J., Linehan, W.M., Nelkin, B.D., Baylin, S.B., Israel,M.A., 1987. Molecular markers of neuroendocrine development andevidence of environmental regulation. Proc. Natl. Acad. Sci. U. S. A. 84,2336–2339.
Jensen, C.H., Krogh, T.N., Stoving, R.K., Holmskov, U., Teisner, B., 1997. Fetalantigen 1 (FA1), a circulating member of the epidermal growth factor (EGF)superfamily: ELISA development, physiology and metabolism in relation torenal function. Clin. Chim. Acta 268, 1–20.
Jensen, C.H., Meyer, M., Schroder, H.D., Kliem, A., Zimmer, J., Teisner, B.,2001. Neurons in the monoaminergic nuclei of the rat and human centralnervous system express FA1/dlk. NeuroReport 12, 3959–3963.
Jorgensen, J.R., Juliusson, B., Henriksen, K.F., Hansen, C., Knudsen, S.,Petersen, T.N., Blom, N., Seiger, A., Wahlberg, L.U., 2006. Identification ofnovel genes regulated in the developing human ventral mesencephalon. Exp.Neurol. 198, 427–437.
Kawasaki, H., Mizuseki, K., Nishikawa, S., Kaneko, S., Kuwana, Y., Nakanishi,S., Nishikawa, S.I., Sasai, Y., 2000. Induction of midbrain dopaminergicneurons from ES cells by stromal cell-derived inducing activity. Neuron 28,31–40.
Kirik, D., Rosenblad, C., Bjorklund, A., 2000a. Preservation of a functionalnigrostriatal dopamine pathway by GDNF in the intrastriatal 6-OHDA lesionmodel depends on the site of administration of the trophic factor. Eur. J.Neurosci. 12, 3871–3882.
Kirik, D., Rosenblad, C., Bjorklund, A., Mandel, R.J., 2000b. Long-term rAAV-mediated gene transfer of GDNF in the rat Parkinson's model: intrastriatalbut not intranigral transduction promotes functional regeneration in thelesioned nigrostriatal system. J. Neurosci. 20, 4686–4700.
Knudsen, S., Workman, C., Sicheritz-Ponten, T., Friis, C., 2003. GenePublisher:automated analysis of DNA microarray data. Nucleic Acids Res. 31,3471–3476.
Kordower, J.H., Emborg, M.E., Bloch, J., Ma, S.Y., Chu, Y., Leventhal, L.,McBride, J., Chen, E.Y., Palfi, S., Roitberg, B.Z., Brown, W.D., Holden,J.E., Pyzalski, R., Taylor, M.D., Carvey, P., Ling, Z., Trono, D., Hantraye, P.,Deglon, N., Aebischer, P., 2000. Neurodegeneration prevented by lentiviralvector delivery of GDNF in primate models of Parkinson's disease. Science290, 767–773.
Laborda, J., Sausville, E.A., Hoffman, T., Notario, V., 1993. dlk, a putativemammalian homeotic gene differentially expressed in small cell lungcarcinoma and neuroendocrine tumor cell line. J. Biol. Chem. 268,3817–3820.
Linnarsson, S., Mikaels, A., Baudet, C., Ernfors, P., 2001. Activation by GDNFof a transcriptional program repressing neurite growth in dorsal root ganglia.Proc. Natl. Acad. Sci. U. S. A. 98, 14681–14686.
Lotharius, J., Barg, S., Wiekop, P., Lundberg, C., Raymon, H.K., Brundin, P.,2002. Effect of mutant alpha-synuclein on dopamine homeostasis in a newhuman mesencephalic cell line. J. Biol. Chem. 277, 38884–38894.

801N.S. Christophersen et al. / Experimental Neurology 204 (2007) 791–801
Mandel, S., Grunblatt, E., Maor, G., Youdim, M.B., 2002. Early and late genechanges in MPTP mice model of Parkinson's disease employing cDNAmicroarray. Neurochem. Res. 27, 1231–1243.
Martin, D., Miller, G., Cullen, T., Fischer, N., Dix, D., Russell, D., 1996.Intranigral or intrastriatal injections of GDNF: effects on monoamine levelsand behavior in rats. Eur. J. Pharmacol. 317, 247–256.
Miller, R.M., Callahan, L.M., Casaceli, C., Chen, L., Kiser, G.L., Chui, B.,Kaysser-Kranich, T.M., Sendera, T.J., Palaniappan, C., Federoff, H.J., 2004.Dysregulation of gene expression in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse substantia nigra. J. Neurosci. 24,7445–7454.
Oo, T.F., Kholodilov, N., Burke, R.E., 2003. Regulation of natural cell death indopaminergic neurons of the substantia nigra by striatal glial cell line-derived neurotrophic factor in vivo. J. Neurosci. 23, 5141–5148.
Pahnke, J., Mix, E., Knoblich, R., Muller, J., Zschiesche, M., Schubert, B.,Koczan, D., Bauer, P., Bottcher, T., Thiesen, H.J., Lazarov, L., Wree, A.,Rolfs, A., 2004. Overexpression of glial cell line-derived neurotrophic factorinduces genes regulating migration and differentiation of neuronalprogenitor cells. Exp. Cell Res. 297, 484–494.
Paul, G., Christophersen, N.S., Raymon, H., Kiaer, C., Smith, R., Brundin, P.,in press. Tyrosine hydroxylase expression is unstable in a humanimmortalized mesencephalic cell line - Studies in vitro and afterintracerebral grafting in vivo. doi:10.1016/j.mcn.2006.11.010.
Rosenblad, C., Georgievska, B., Kirik, D., 2003. Long-term striatal over-expression of GDNF selectively downregulates tyrosine hydroxylase in theintact nigrostriatal dopamine system. Eur. J. Neurosci. 17, 260–270.
Sherer, T.B., Fiske, B.K., Svendsen, C.N., Lang, A.E., Langston, J.W., 2006.Crossroads in GDNF therapy for Parkinson's disease. Mov. Disord. 21,136–141.
Smas, C.M., Sul, H.S., 1993. Pref-1, a protein containing EGF-like repeats,inhibits adipocyte differentiation. Cell 73, 725–734.
Smas, C.M., Chen, L., Sul, H.S., 1997. Cleavage of membrane-associated pref-1generates a soluble inhibitor of adipocyte differentiation. Mol. Cell. Biol. 17,977–988.
Studer, L., Spenger, C., Seiler, R.W., Altar, C.A., Lindsay, R.M., Hyman, C.,1995. Comparison of the effects of the neurotrophins on the morphologicalstructure of dopaminergic neurons in cultures of rat substantia nigra. Eur. J.Neurosci. 7, 223–233.
Studer, L., Spenger, C., Seiler, R.W., Othberg, A., Lindvall, O., Odin, P., 1996.Effects of brain-derived neurotrophic factor on neuronal structure ofdopaminergic neurons in dissociated cultures of human fetal mesencepha-lon. Exp. Brain Res. 108, 328–336.
Van Limpt, V.A., Chan, A.J., Van Sluis, P.G., Caron, H.N., Van Noesel, C.J.,Versteeg, R., 2003. High delta-like 1 expression in a subset of neuroblastomacell lines corresponds to a differentiated chromaffin cell type. Int. J. Cancer105, 61–69.




















