
Isolation and Characterization of Pluripotent Human Spermatogonial Stem Cell-Derived Cells

Robbins, Michael T. Longaker and Joseph C. WuFumi Ikeno, Cholawat Pacharinsak, Andrew J. Connolly, Sanjiv S. Gambhir, Robert C.
Han, Bruno C. Huber, Won Hee Lee, Yongquan Gong, Patricia E. de Almeida, Jennifer Lyons, Mingxia Gu, Patricia K. Nguyen, Andrew S. Lee, Dan Xu, Shijun Hu, Jordan R. Plews, Leng
Derived Endothelial Cells Improve Myocardial Function by Paracrine Activation−Microfluidic Single-Cell Analysis Shows That Porcine Induced Pluripotent Stem Cell
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2012 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.112.2690012012;111:882-893; originally published online July 19, 2012;Circ Res.
http://circres.ahajournals.org/content/111/7/882World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2012/07/19/CIRCRESAHA.112.269001.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

Integrative Physiology
Microfluidic Single-Cell Analysis Shows That PorcineInduced Pluripotent Stem Cell–Derived Endothelial Cells
Improve Myocardial Function by Paracrine ActivationMingxia Gu,* Patricia K. Nguyen,* Andrew S. Lee, Dan Xu, Shijun Hu, Jordan R. Plews, Leng Han,Bruno C. Huber, Won Hee Lee, Yongquan Gong, Patricia E. de Almeida, Jennifer Lyons, Fumi Ikeno,
Cholawat Pacharinsak, Andrew J. Connolly, Sanjiv S. Gambhir, Robert C. Robbins,Michael T. Longaker, Joseph C. Wu
Rationale: Induced pluripotent stem cells (iPSCs) hold great promise for the development of patient-specifictherapies for cardiovascular disease. However, clinical translation will require preclinical optimization andvalidation of large-animal iPSC models.
Objective: To successfully derive endothelial cells from porcine iPSCs and demonstrate their potential utility forthe treatment of myocardial ischemia.
Methods and Results: Porcine adipose stromal cells were reprogrammed to generate porcine iPSCs (piPSCs).Immunohistochemistry, quantitative PCR, microarray hybridization, and angiogenic assays confirmed thatpiPSC-derived endothelial cells (piPSC-ECs) shared similar morphological and functional properties asendothelial cells isolated from the autologous pig aorta. To demonstrate their therapeutic potential, piPSC-ECswere transplanted into mice with myocardial infarction. Compared with control, animals transplanted withpiPSC-ECs showed significant functional improvement measured by echocardiography (fractional shortening atweek 4: 27.2�1.3% versus 22.3�1.1%; P<0.001) and MRI (ejection fraction at week 4: 45.8�1.3% versus42.3�0.9%; P<0.05). Quantitative protein assays and microfluidic single-cell PCR profiling showed thatpiPSC-ECs released proangiogenic and antiapoptotic factors in the ischemic microenvironment, which promotedneovascularization and cardiomyocyte survival, respectively. Release of paracrine factors varied significantlyamong subpopulations of transplanted cells, suggesting that transplantation of specific cell populations mayresult in greater functional recovery.
Conclusions: In summary, this is the first study to successfully differentiate piPSCs-ECs from piPSCs anddemonstrate that transplantation of piPSC-ECs improved cardiac function after myocardial infarction viaparacrine activation. Further development of these large animal iPSC models will yield significant insights intotheir therapeutic potential and accelerate the clinical translation of autologous iPSC-based therapy. (Circ Res.2012;111:882-893.)
Key Words: induced pluripotent stem cells � large-animal models � paracrine activation � myocardial infarction �molecular imaging � ischemic heart disease � vascular biology
In recent years, induced pluripotent stem cells (iPSCs) havebecome a popular alternative to embryonic stem cells
(ESCs) for regenerative medicine.1,2 Not only do iPSCsprovide an unlimited source of pluripotent cells that are
capable of self-renewal, but they also circumvent the ethicalconcerns associated with ESC derivation and can be trans-planted autologously, limiting potential immunologic rejec-tion.3,4 To date, iPSCs have been generated from a growing
Original received March 11, 2012; revision received July 17, 2012; accepted July 19, 2012. In June 2012, the average time from submission to firstdecision for all original research papers submitted to Circulation Research was 13.35 days.
From the Department of Medicine, Division of Cardiology, Stanford University School of Medicine, Stanford, CA (M.G., P.K.N., A.S.L., D.X., S.H.,J.R.P., L.H., B.C.H., W.H.L., Y.G., P.E.d.A., J.L., F.I., J.C.W.); the Department of Radiology, Molecular Imaging Program, Stanford University, Stanford,CA (A.S.L., D.X., S.H., J.R.P., L.H., B.C.H., W.H.L., Y.G., P.E.d.A., S.S.G., J.C.W.); the Department of Physiology and Pathophysiology, KeyLaboratory of Molecular Cardiovascular Sciences of Education Ministry, Peking University Health Sciences Center, Beijing, China (M.G.); and theDepartment of Cardiothoracic Surgery (Y.G., R.C.R.), the Department of Comparative Medicine (C.P.), the Department of Pathology (A.J.C.), theDepartment of Plastic Surgery (M.T.L.), and the Institute of Stem Cell Biology and Regenerative Medicine, Stanford University, Stanford, CA (J.C.W.).
*These authors contributed equally.The online-only Data Supplement is available with this article at http://circres.ahajournals.org/lookup/suppl/doi:10.1161/CIRCRESAHA.112.
269001/-/DC1.Correspondence to Joseph C. Wu, MD, 265 Campus Dr, Room 1120B, Stanford, CA 94305-5344. E-mail [email protected]; Michael Longaker,
MD, 257 Campus Dr, Stanford, CA 94305-5148. E-mail [email protected]© 2012 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.112.269001
882 at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

list of species including mice,5,6 rats,7,8 monkeys,9 dogs,10,11
humans,12,13 and pigs,14,15 confirming the universality oftransgenes for reprogramming.12,13 Perhaps the most signifi-cant achievement is the development of porcine iPSCs, whichcan potentially fill the gap between the transplantation ofthese cells into mice and the initiation of the first clinicaltrials using iPSCs in humans.
In This Issue, see p 815Editorial, see p 824
Swine are an ideal large-animal model for the clinical trans-lation of experimental medical therapies due to their resem-blance to human organ size and physiology and their relativelylong lifespan.16 Consequently, several recent studies have uti-lized the porcine model as a preclinical tool for the clinicaltranslation of cell-based therapy. These studies, however, havebeen limited to porcine mesenchymal stem cells.17–20
It has long been appreciated that endothelial cells play animportant role in the survival and function of nearby cardio-myocytes by promoting vascular regeneration and angiogen-esis.21 In the normal heart, capillary networks can be foundnear almost every cardiomyocyte, modulating cardiac perfor-mance, contraction, and growth. Thus, it is not surprising thatin the ischemic myocardium, a deficit of blood flow leads toprogressive cardiomyocyte death and myocardial dysfunc-tion.22 To exploit the potential therapeutic benefits of endo-thelial cells, several investigators have differentiated endo-thelial cells from iPSCs generated from mice,23 dogs,11 andhumans,24 demonstrating that paracrine release of proangio-genic factors promote functional recovery in small animalmodels of ischemia. Prior studies, however, have analyzedentire cell populations. Because cell populations are hetero-geneous, especially those generated from exogenous factorssuch as iPSCs and their derivatives, it is critical to study theregulation of angiogenesis at the single-cell level.
In this study, we lay the foundation for the clinicalapplication of iPSCs for the treatment of ischemic heartdisease. After generation of iPSCs from adipose tissue ofadult miniswine, we successfully differentiated these cellsinto endothelial cells that share similar morphological andfunctional properties to their endogenous counterparts (ie,endothelial cells isolated from the porcine aorta). We thendemonstrated that porcine iPSC-derived endothelial cells(piPSC-ECs) could provide therapeutic benefit in a murinemodel of myocardial infarction (MI). To explore the biolog-ical mechanisms for functional recovery, we showed thatpiPSC-ECs released both proangiogenic and antiapoptoticfactors that promote neoangiogenesis and cardiomyocytesurvival, respectively, at the single-cell level using microflu-idic dynamic arrays.25 We then demonstrated that the releaseof paracrine factors varied among specific subpopulations ofiPSC-ECs, suggesting that transplantation of specific cellsubpopulations may result in greater functional recovery.Finally, cell delivery and engraftment are monitored usingpositron emission tomography/computed tomography (PET/CT) as well as MRI in a porcine model of MI, laying thegroundwork for the development of future imaging protocolsto monitor cell delivery and engraftment in humans.
MethodsDerivation of Porcine iPSCsPorcine adipose tissue was harvested and cultured, as described indetail in the Online Supplemental Methods section. Approximately1�104 porcine adipose stromal cells (pASCs) were seeded onto a24-well tissue culture dish and maintained with human ASC growthmedium. Cells were then transduced with 5 �g/mL polybrene(Sigma) plus individual lentivirus containing Oct4, Sox2, Klf4, andc-MYC in a 4:2:2:1 ratio.13 On day 6, cells were dissociated with0.05% trypsin-EDTA (Gibco) and counted with a hemocytometer.Reprogramming of piPSCs was performed on mouse embryonicfibroblast (MEF) feeder cells. Ten thousand cells were then trans-ferred onto an MEF feeder layer in a gelatin-coated 6-well plate andcultured with human ESC growth medium containing knockoutDMEM (Invitrogen), 20% knockout FBS (Invitrogen), 2 mmol/LL-glutamine (Invitrogen), 1� nonessential amino acids (Invitrogen),1�2-mercaptoethanol (Invitrogen), 100 U/mL penicillin, 100 �g/mLstreptomycin, and 10 ng/mL human �-fibroblast growth factor(bFGF) (Invitrogen). ESC-like colonies with distinct edges appearedon day 13 to 15. On day 15, positive colonies with ESC-likemorphologies were isolated with a glass needle and seeded onto new
Non-standard Abbreviations and Acronyms
bFGF human �-fibroblast growth factor
BLI bioluminescence imaging
CSF3 colony-stimulating factor 3
CT computed tomography
EBs embryoid bodies
ECs endothelial cells
ESC embryonic stem cell
FACS fluorescence activated cell sorting
FGF-� fibroblast growth factor-�
FGFR2 fibroblast growth factor receptor 2
FLuc firefly luciferase
HSVttk herpes simplex virus truncated thymidine kinase
IFN-� interferon gamma
IL interleukin
Ip10 interferon-inducible protein
iPSCs induced pluripotent stem cells
LAD left anterior descending
MEF mouse embryonic fibroblast
MI myocardial infarction
mRFP monomeric red fluorescent protein
pAorta-ECs endothelial cells harvested from the porcine aorta
pASC porcine adipose stromal cells
PBS phosphate-buffered saline
PCR polymerase chain reaction
PET positron emission tomography
piPSC-ECs endothelial cells derived from porcine induced pluripo-tent stem cells
qRT-PCR quantitative reverse-transcription polymerase chainreaction
TIMP1 tissue inhibitor of metalloproteinase 1
TIMP2 tissue inhibitor of metalloproteinase 2
TNF-� tumor necrosis factor-�
TTC triphenyltetrazolium chloride
VEGF vascular endothelial growth factor
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 883
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

MEFs. These pASC-derived iPSCs were maintained on MEF feederlayers with iPSC medium containing knockout DMEM (Invitrogen),20% ES-qualified FBS (Invitrogen), 2 mmol/L L-glutamine (Invit-rogen), 1� nonessential amino acids (Invitrogen), 1�2-mercaptoethanol (Invitrogen), 100 U/mL penicillin, 100 �g/mLstreptomycin, and 50 ng/mL human bFGF (Invitrogen). Each se-lected colony was then established as one individual pASC-iPSCline. A total of 10 lines from the 5 miniswine were generated.
Differentiation of piPSC-Derived ECsFor endothelial cell differentiation, well maintained piPSCs at passagenumber 25 were treated with type IV collagenase (Invitrogen, Carlsbad,CA) for 15 minutes at 37°C and transferred to ultra-low attachmentplates (Corning Life Sciences, Kennebunk, ME) containing differenti-ation medium for 2 days.26 The differentiation medium consisted ofDMEM (Invitrogen) containing 20% knockout serum (Invitrogen), 4.5g/L L-glutamine (Invitrogen), 1% nonessential amino acids (Invitro-gen), 0.1 mmol/L 2-mercaptoethanol (Invitrogen), 50 U/mL penicillin(Invitrogen), 50 �g/mL streptomycin (Invitrogen), 10 ng/mL Activin A(R&D Systems), and 20 ng/mL bone morphogenetic protein-4 (BMP-4)as described previously.26 The differentiation medium was supple-mented with 25 ng/mL vascular endothelial growth factor (VEGF)(R&D Systems), 8 ng/mL bFGF (R&D Systems), and 10 �mol/LTGF-� inhibitor SB431542 (Sigma). The 2-day embryoid bodies (EBs)were then seeded onto 0.5% gelatin-coated dishes and cultured foranother 8 days in differentiation media in absence of BMP-4. On day 10,plated EBs were digested and sorted by fluorescence activated cellsorting (FACS) for CD31-positive cells. The CD31-positive cells werecollected and seeded into one well of a 6-well plate with EGM-2medium (Lonza).
Quantitative Reverse-Transcription PolymeraseChain Reaction Analysis to Confirm PluripotentState in piPSCsQuantitative reverse-transcription polymerase chain reaction (qRT-PCR) was performed after isolation of total RNA and cDNA. UsingTaqman Gene Expression Assays, PCR reactions were performed witha StepOnePlus Realtime-PCR 7900 HT System (Applied Biosystems,Foster City, CA). Primers for genes defining pluripotency and confirm-ing germ layer differentiation are listed in Online Table I.
Microarray Hybridization and Data Analysis ofpiPSCs and piPSC-ECsTotal RNA samples were hybridized to Affymetrix GeneChipPorcine Genome Array, and then normalized and annotated by theAffymetrix Expression Console software. For comparison to ourgenerated lines, we obtained piPSC lines (piPSC-ID4, piPSC-ID6,and piPSC-IC1) from R. Michael Roberts and Toshihiko Ezashi(University of Missouri, Columbia, MO) and are grateful for theircontribution. We also obtained the expression data for piPSC linesand fibroblast from the Gene Expression Omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo, GSE15472).27 The Pearson correlationcoefficient was calculated for each pair of samples using theexpression level of transcripts. For hierarchical clustering, a Pearsoncorrelation for average linkage clustering was used.
Generation of a Murine Model of MI andIntramyocardial Delivery of Therapeutic CellsLigation of the mid left anterior descending (LAD) artery wasperformed in 8- to 10-week-old female NOD SCID mice (JacksonLaboratory) under anesthesia (2% to 3% inhaled isoflurane) by asingle experienced microsurgeon (Y.G.). Survival rate for eachsurgery group was �90%. Myocardial blanching and ECG changesconfirmed MI after ligation. Animals were randomized into 4groups: (1) phosphate-buffered saline (PBS) as control, (2) 1�106
pASCs, (3) 1�106 pAorta-ECs, and (4) 1�106 piPSC-ECs (n�20/group). Immediately after LAD artery ligation, animals were injectedintramyocardially near the peri-infarct zone at 2 different sites usinga 31-gauge Hamilton syringe (25 �L per injection). Study protocols
were approved by the Stanford Animal Research Committee. Animalcare was provided in accordance with the Stanford University Schoolof Medicine guidelines and policies for the use of laboratory animals.
Optical Bioluminescence Imaging and MicroPositron Emission Tomography for TrackingCell FatePlease see Online Data Supplement Methods.
Angiogenesis and Antiapoptosis Cytokine ArrayAfter Hypoxia ExposurePlease see Online Data Supplement Methods.
Single-Cell Gene Expression Profiling ofTransplanted piPSC-ECs and ASCsPlease see Online Data Supplement Methods.
Assessment of Paracrine Activation bySpecific SubpopulationsBecause cell populations of pASC and piPSC-ECs are heteroge-neous, we hypothesized that the release of paracrine factors varied byspecific cell subpopulations. To investigate this hypothesis, we usedsingle-cell expression profiling to analyze the release of paracrinefactors by subpopulations of pASCs and piPSC-ECs. One week aftertransplantation of pASCs and piPSC-ECs into mice undergoing LADartery infarction (n�5), we euthanized the mice and harvested anddissociated their hearts as described above. We then used FACS toisolate subpopulations of cells and sort them into a 96-well 0.2-mL PCRplate, based on their expression of endothelial specific markers, specif-ically CD31, CD34, and CD144. We then performed single-cell geneexpression analysis to determine the relative amounts of paracrinefactors released by each subpopulation of pASC and piPSC-ECs: (1)CD31�/CD34�/CD144�, (2) CD31�/CD34�/CD144�, and (3)CD31�/CD34�/CD144�. Very few cells expressed all three markersand this subpopulation was, therefore, not analyzed.
Tracking Transplanted Cells in Miniswine UsingPET and CT ImagingPlease see Online Data Supplement Methods.
Statistical AnalysisFor analysis of microarray data, a Pearson correlation coefficient wasutilized. A probability value of �0.05 was considered significant.ANOVA with post hoc testing was used to analyze the differences inqRT-PCR, single-cell PCR, angiogenesis, and antiapoptotic cytokinedata between groups, percent infarct size by triphenyltetrazoliumchloride (TTC), and density of dTd-mediated dUTP nick-end label-ing (TUNEL)-positive nuclei. The percent infarct size is calculatedby dividing the area of infarct by the total area of the left ventricle.Repeated measures of ANOVA were used to analyze MRI, biolu-minescence imaging (BLI), and echocardiographic data. All data areexpressed as mean�SD.
ResultsDerivation of piPSCs From Adult MiniswinePorcine adipose stromal cells (pASCs) were harvested andsuccessfully reprogrammed into piPSCs from 5 young adultYucatan miniswine pigs. Retroviral overexpression of Oct4,Sox2, Klf4, and c-Myc was used in a 4:2:2:1 ratio to inducepluripotency. Compact colonies of cells staining positive foralkaline phosphatase, a universal pluripotent marker (OnlineFigure IA), were initially observed under bright-field micros-copy 15 days after transduction (Figure 1A). Individual colonieswere mechanically dissociated using a pulled Pasteur pipette andtransferred onto irradiated MEF feeder layers for clonal expan-sion into distinct cell lines. A total of 10 piPSC lines weregenerated, 2 lines from each miniswine. piPSC colonies stained
884 Circulation Research September 14, 2012
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
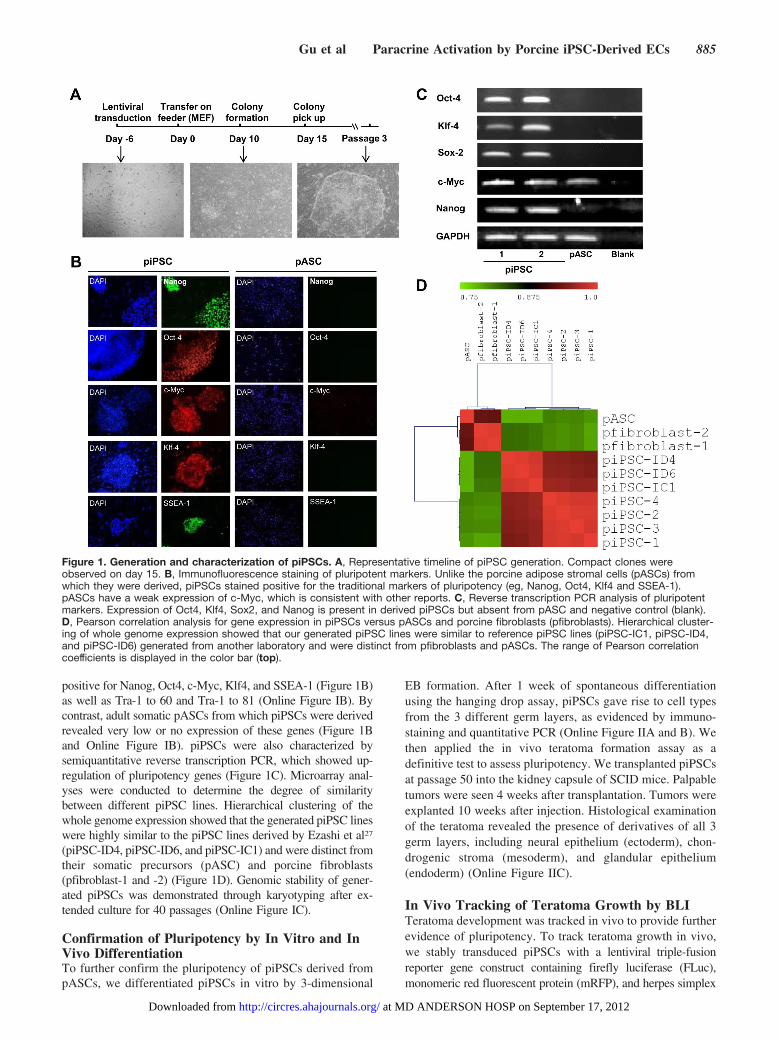
positive for Nanog, Oct4, c-Myc, Klf4, and SSEA-1 (Figure 1B)as well as Tra-1 to 60 and Tra-1 to 81 (Online Figure IB). Bycontrast, adult somatic pASCs from which piPSCs were derivedrevealed very low or no expression of these genes (Figure 1Band Online Figure IB). piPSCs were also characterized bysemiquantitative reverse transcription PCR, which showed up-regulation of pluripotency genes (Figure 1C). Microarray anal-yses were conducted to determine the degree of similaritybetween different piPSC lines. Hierarchical clustering of thewhole genome expression showed that the generated piPSC lineswere highly similar to the piPSC lines derived by Ezashi et al27
(piPSC-ID4, piPSC-ID6, and piPSC-IC1) and were distinct fromtheir somatic precursors (pASC) and porcine fibroblasts(pfibroblast-1 and -2) (Figure 1D). Genomic stability of gener-ated piPSCs was demonstrated through karyotyping after ex-tended culture for 40 passages (Online Figure IC).
Confirmation of Pluripotency by In Vitro and InVivo DifferentiationTo further confirm the pluripotency of piPSCs derived frompASCs, we differentiated piPSCs in vitro by 3-dimensional
EB formation. After 1 week of spontaneous differentiationusing the hanging drop assay, piPSCs gave rise to cell typesfrom the 3 different germ layers, as evidenced by immuno-staining and quantitative PCR (Online Figure IIA and B). Wethen applied the in vivo teratoma formation assay as adefinitive test to assess pluripotency. We transplanted piPSCsat passage 50 into the kidney capsule of SCID mice. Palpabletumors were seen 4 weeks after transplantation. Tumors wereexplanted 10 weeks after injection. Histological examinationof the teratoma revealed the presence of derivatives of all 3germ layers, including neural epithelium (ectoderm), chon-drogenic stroma (mesoderm), and glandular epithelium(endoderm) (Online Figure IIC).
In Vivo Tracking of Teratoma Growth by BLITeratoma development was tracked in vivo to provide furtherevidence of pluripotency. To track teratoma growth in vivo,we stably transduced piPSCs with a lentiviral triple-fusionreporter gene construct containing firefly luciferase (FLuc),monomeric red fluorescent protein (mRFP), and herpes simplex
Figure 1. Generation and characterization of piPSCs. A, Representative timeline of piPSC generation. Compact clones wereobserved on day 15. B, Immunofluorescence staining of pluripotent markers. Unlike the porcine adipose stromal cells (pASCs) fromwhich they were derived, piPSCs stained positive for the traditional markers of pluripotency (eg, Nanog, Oct4, Klf4 and SSEA-1).pASCs have a weak expression of c-Myc, which is consistent with other reports. C, Reverse transcription PCR analysis of pluripotentmarkers. Expression of Oct4, Klf4, Sox2, and Nanog is present in derived piPSCs but absent from pASC and negative control (blank).D, Pearson correlation analysis for gene expression in piPSCs versus pASCs and porcine fibroblasts (pfibroblasts). Hierarchical cluster-ing of whole genome expression showed that our generated piPSC lines were similar to reference piPSC lines (piPSC-IC1, piPSC-ID4,and piPSC-ID6) generated from another laboratory and were distinct from pfibroblasts and pASCs. The range of Pearson correlationcoefficients is displayed in the color bar (top).
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 885
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
virus truncated thymidine kinase (HSVttk) driven by ubiquitinpromoter. Cells expressing the reporter gene were selected bycell sorting for mRFP (Online Figure IIIA). After injection intothe kidney capsule of immunodeficient mice, teratoma growthwas confirmed by BLI (Online Figure IIIB).
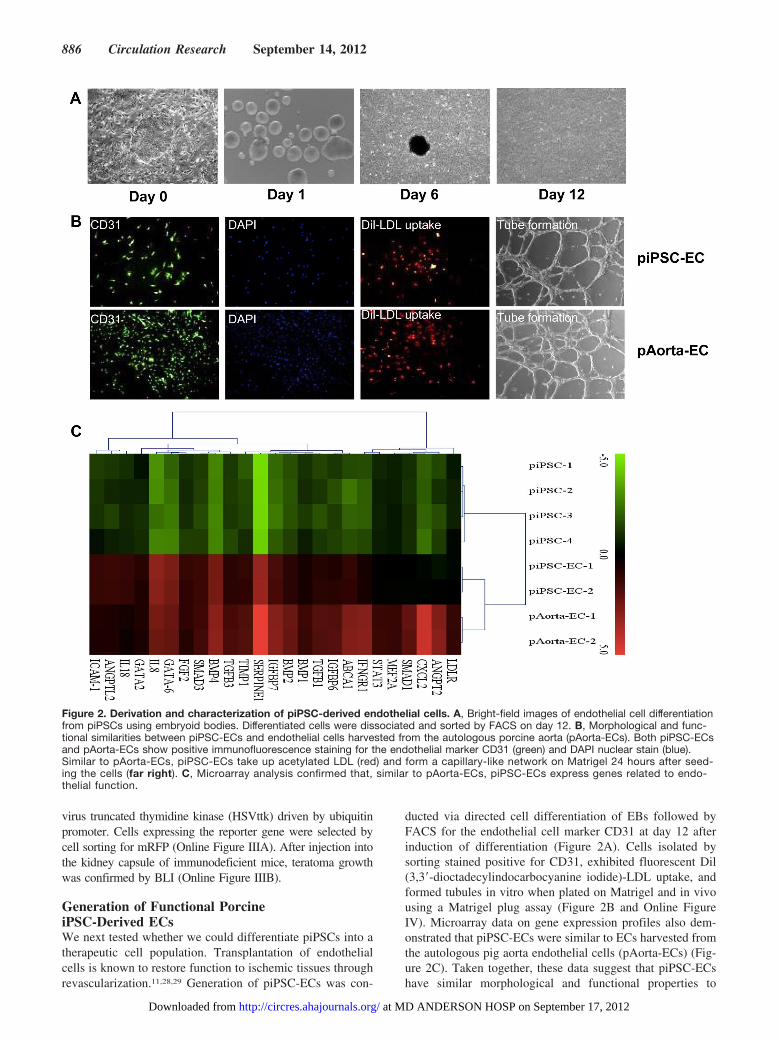
Generation of Functional PorcineiPSC-Derived ECsWe next tested whether we could differentiate piPSCs into atherapeutic cell population. Transplantation of endothelialcells is known to restore function to ischemic tissues throughrevascularization.11,28,29 Generation of piPSC-ECs was con-
ducted via directed cell differentiation of EBs followed byFACS for the endothelial cell marker CD31 at day 12 afterinduction of differentiation (Figure 2A). Cells isolated bysorting stained positive for CD31, exhibited fluorescent Dil(3,3�-dioctadecylindocarbocyanine iodide)-LDL uptake, andformed tubules in vitro when plated on Matrigel and in vivousing a Matrigel plug assay (Figure 2B and Online FigureIV). Microarray data on gene expression profiles also dem-onstrated that piPSC-ECs were similar to ECs harvested fromthe autologous pig aorta endothelial cells (pAorta-ECs) (Fig-ure 2C). Taken together, these data suggest that piPSC-ECshave similar morphological and functional properties to
Figure 2. Derivation and characterization of piPSC-derived endothelial cells. A, Bright-field images of endothelial cell differentiationfrom piPSCs using embryoid bodies. Differentiated cells were dissociated and sorted by FACS on day 12. B, Morphological and func-tional similarities between piPSC-ECs and endothelial cells harvested from the autologous porcine aorta (pAorta-ECs). Both piPSC-ECsand pAorta-ECs show positive immunofluorescence staining for the endothelial marker CD31 (green) and DAPI nuclear stain (blue).Similar to pAorta-ECs, piPSC-ECs take up acetylated LDL (red) and form a capillary-like network on Matrigel 24 hours after seed-ing the cells (far right). C, Microarray analysis confirmed that, similar to pAorta-ECs, piPSC-ECs express genes related to endo-thelial function.
886 Circulation Research September 14, 2012
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
endogenous ECs and, thus, can be used as an alternativetherapeutic option for vascular regeneration.
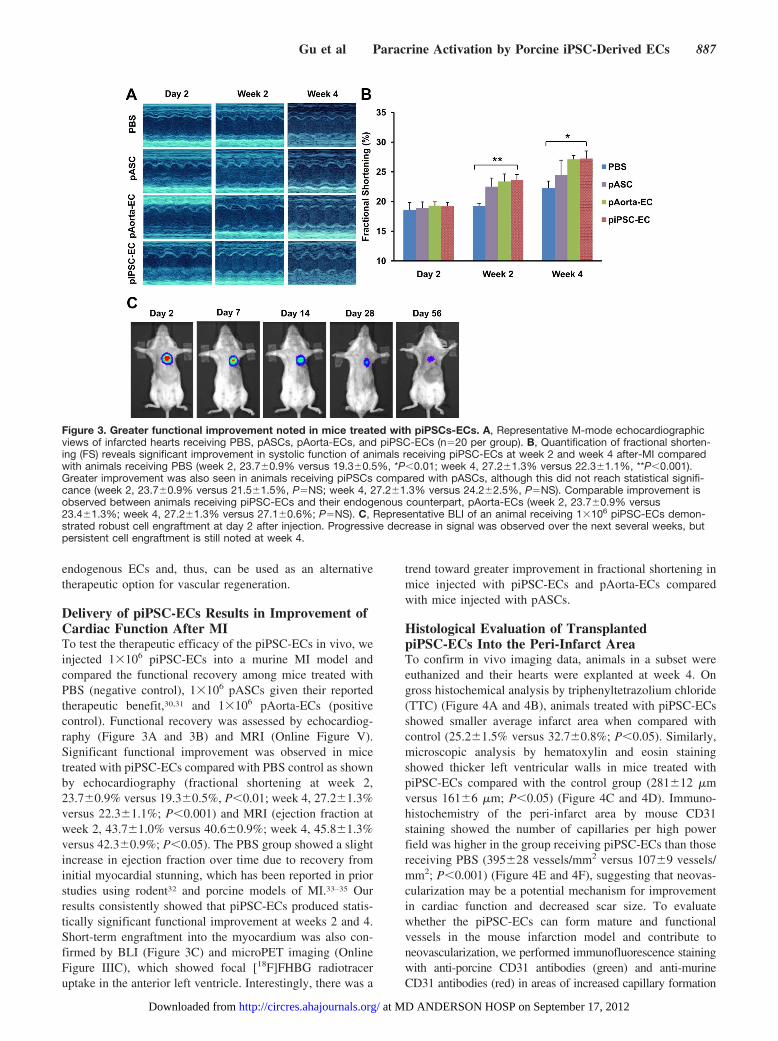
Delivery of piPSC-ECs Results in Improvement ofCardiac Function After MITo test the therapeutic efficacy of the piPSC-ECs in vivo, weinjected 1�106 piPSC-ECs into a murine MI model andcompared the functional recovery among mice treated withPBS (negative control), 1�106 pASCs given their reportedtherapeutic benefit,30,31 and 1�106 pAorta-ECs (positivecontrol). Functional recovery was assessed by echocardiog-raphy (Figure 3A and 3B) and MRI (Online Figure V).Significant functional improvement was observed in micetreated with piPSC-ECs compared with PBS control as shownby echocardiography (fractional shortening at week 2,23.7�0.9% versus 19.3�0.5%, P�0.01; week 4, 27.2�1.3%versus 22.3�1.1%; P�0.001) and MRI (ejection fraction atweek 2, 43.7�1.0% versus 40.6�0.9%; week 4, 45.8�1.3%versus 42.3�0.9%; P�0.05). The PBS group showed a slightincrease in ejection fraction over time due to recovery frominitial myocardial stunning, which has been reported in priorstudies using rodent32 and porcine models of MI.33–35 Ourresults consistently showed that piPSC-ECs produced statis-tically significant functional improvement at weeks 2 and 4.Short-term engraftment into the myocardium was also con-firmed by BLI (Figure 3C) and microPET imaging (OnlineFigure IIIC), which showed focal [18F]FHBG radiotraceruptake in the anterior left ventricle. Interestingly, there was a
trend toward greater improvement in fractional shortening inmice injected with piPSC-ECs and pAorta-ECs comparedwith mice injected with pASCs.
Histological Evaluation of TransplantedpiPSC-ECs Into the Peri-Infarct AreaTo confirm in vivo imaging data, animals in a subset wereeuthanized and their hearts were explanted at week 4. Ongross histochemical analysis by triphenyltetrazolium chloride(TTC) (Figure 4A and 4B), animals treated with piPSC-ECsshowed smaller average infarct area when compared withcontrol (25.2�1.5% versus 32.7�0.8%; P�0.05). Similarly,microscopic analysis by hematoxylin and eosin stainingshowed thicker left ventricular walls in mice treated withpiPSC-ECs compared with the control group (281�12 �mversus 161�6 �m; P�0.05) (Figure 4C and 4D). Immuno-histochemistry of the peri-infarct area by mouse CD31staining showed the number of capillaries per high powerfield was higher in the group receiving piPSC-ECs than thosereceiving PBS (395�28 vessels/mm2 versus 107�9 vessels/mm2; P�0.001) (Figure 4E and 4F), suggesting that neovas-cularization may be a potential mechanism for improvementin cardiac function and decreased scar size. To evaluatewhether the piPSC-ECs can form mature and functionalvessels in the mouse infarction model and contribute toneovascularization, we performed immunofluorescence stainingwith anti-porcine CD31 antibodies (green) and anti-murineCD31 antibodies (red) in areas of increased capillary formation
Figure 3. Greater functional improvement noted in mice treated with piPSCs-ECs. A, Representative M-mode echocardiographicviews of infarcted hearts receiving PBS, pASCs, pAorta-ECs, and piPSC-ECs (n�20 per group). B, Quantification of fractional shorten-ing (FS) reveals significant improvement in systolic function of animals receiving piPSC-ECs at week 2 and week 4 after-MI comparedwith animals receiving PBS (week 2, 23.7�0.9% versus 19.3�0.5%, *P�0.01; week 4, 27.2�1.3% versus 22.3�1.1%, **P�0.001).Greater improvement was also seen in animals receiving piPSCs compared with pASCs, although this did not reach statistical signifi-cance (week 2, 23.7�0.9% versus 21.5�1.5%, P�NS; week 4, 27.2�1.3% versus 24.2�2.5%, P�NS). Comparable improvement isobserved between animals receiving piPSC-ECs and their endogenous counterpart, pAorta-ECs (week 2, 23.7�0.9% versus23.4�1.3%; week 4, 27.2�1.3% versus 27.1�0.6%; P�NS). C, Representative BLI of an animal receiving 1�106 piPSC-ECs demon-strated robust cell engraftment at day 2 after injection. Progressive decrease in signal was observed over the next several weeks, butpersistent cell engraftment is still noted at week 4.
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 887
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

from explanted murine hearts. We found that microvessels wereonly immunoreactive to murine-specific antibodies (Online Fig-ure VIA and B). However, piPSC-ECs could be found surround-ing murine microvessels even though the cells themselves do notform mature vessels in hearts explanted 4 weeks after transplan-tation (Online Figure VIC and D). The coalescing of piPSC-ECsadjacent to murine microvessels supports our findings thatpiPSC-ECs release proangiogenic factors that increase nativevessel formation. The failure of piPSC-ECs to form discretemicrovessels in the mouse infarcted heart may be due to aninsufficient number of piPSC-ECs present in the infarctedmouse heart to support vessel formation. As shown by biolumi-nescence imaging (Figure 3C), the majority of transplantedpiPSC-ECs do not survive 4 weeks after-transplantation. Incontrast, formation of discrete microvessels was demonstratedby the in vivo Matrigel plug assay 7 days after implanta-tion, which is a very different environmental niche (OnlineFigure IV).
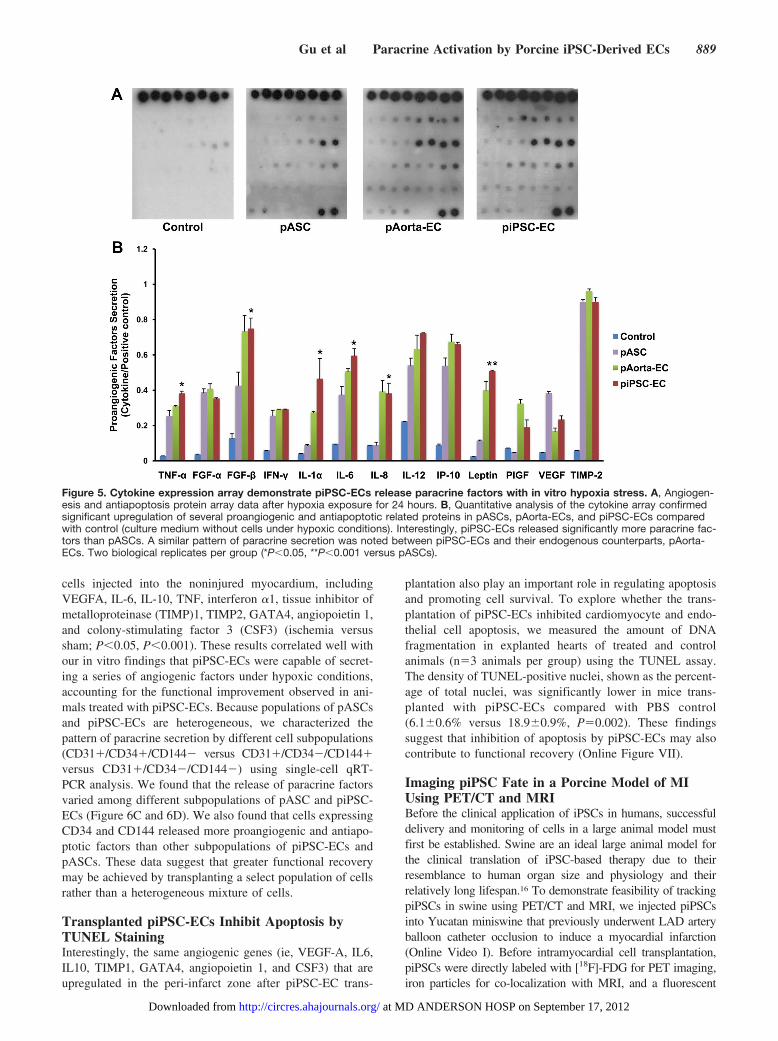
Assessment of Paracrine Factor Release In Vitroby piPSC-ECs in Response to HypoxiaGiven that the majority of cells do not survive 4 weeks aftertransplantation (Figure 3C), we hypothesize that piPSC-ECsmay release paracrine factors that promote angiogenesis inthe setting of ischemia, accounting for the longer-termfunctional benefit that was observed. To explore this hypoth-esis, we compared the secretion of proangiogenic cytokines inpASC, pAorta-ECs, and piPSC-ECs after 24 hours of hypoxia(Figure 5). The culture medium served as control. Both
pASCs and piPSC-ECs secreted more angiogenic factors thancontrol. In addition, piPSC-ECs secreted more tumor necrosisfactor-� (TNF-�), fibroblast growth factor-� (FGF-�), inter-leukin (IL)-1�, IL-6, IL-8, and leptin than pASCs (P�0.05,P�0.01 versus pASCs). The pattern of cytokine secretionwas similar between piPSC-ECs and pAorta-ECs. Takentogether, these in vitro data suggest that piPSC-ECs canprovide a framework for new vessel growth via secretion ofparacrine factors regulating angiogenesis.
Single-Cell Profiling of Transplanted piPSC-ECsReveals In Vivo Paracrine ActivationBecause of the heterogeneous nature of all cell populations,especially those derived from exogenous factors such aspiPSCs and their derivatives,36 it is important to study theexpression of proangiogenic genes at the single-cell level. Inaddition, it is also necessary to confirm that piPSC-ECsrelease paracrine factors under ischemic conditions in vivo.To meet these objectives, we injected the piPSC-ECs into theperi-infarct zone of murine hearts after MI or into the sameanatomic location of control mice with noninfarct shamsurgery. After enzymatic digestion of explanted hearts intosingle-cell suspension, we performed FACS for mRFP ex-pression by piPSC-ECs. Individually isolated piPSC-ECstransplanted into ischemic versus nonischemic hearts wereanalyzed by the expression of 13 common angiogenic genesusing the microfluidic single-cell PCR platform (Figure 6Aand 6B). A number of genes were highly expressed in thepiPSC-ECs injected into the peri-infarct zone compared with
Figure 4. Histological evaluation ofpiPSC-EC therapy in infarcted hearts atweek 4. A, Representative triphenyltetra-zolium chloride (TTC) gross histochemicalanalysis of infarcted hearts injected withPBS (control group) and piPSC-ECs. B,Quantitative analysis of the infarct sizeshowed that the percent infarct size issignificantly smaller in mice treated withpiPSC-EC compared with PBS(25.2�1.5% versus 32.7�0.8%; *P�0.05).C, Representative histology of infarctedhearts injected with PBS and piPSC-ECs(hematoxylin and eosin, magnification1.25� and 5� for the whole heart and leftventricular wall, respectively). D, Quantita-tive analysis of the left ventricular wallthickness showed thicker ventricular wallswere present in the piPSC-EC comparedwith PBS control group (281�12 �m ver-sus 161�6 �m; *P�0.01). E, Representa-tive immunofluorescence staining of themurine endothelial marker CD31 in theperi-infarct area of mice treated with PBS,pASCs, pAorta-ECs, and piPSC-ECs. F,Quantitative analysis of capillary density(vessels/mm2) showed a significantincrease in vessel density (number of ves-sels per high-power field) in animalstreated with piPSC-ECs compared withPBS (395�28 vessels/mm2 versus 107�9vessels/mm2, *P�0.001).
888 Circulation Research September 14, 2012
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
cells injected into the noninjured myocardium, includingVEGFA, IL-6, IL-10, TNF, interferon �1, tissue inhibitor ofmetalloproteinase (TIMP)1, TIMP2, GATA4, angiopoietin 1,and colony-stimulating factor 3 (CSF3) (ischemia versussham; P�0.05, P�0.001). These results correlated well withour in vitro findings that piPSC-ECs were capable of secret-ing a series of angiogenic factors under hypoxic conditions,accounting for the functional improvement observed in ani-mals treated with piPSC-ECs. Because populations of pASCsand piPSC-ECs are heterogeneous, we characterized thepattern of paracrine secretion by different cell subpopulations(CD31�/CD34�/CD144� versus CD31�/CD34�/CD144�versus CD31�/CD34�/CD144�) using single-cell qRT-PCR analysis. We found that the release of paracrine factorsvaried among different subpopulations of pASC and piPSC-ECs (Figure 6C and 6D). We also found that cells expressingCD34 and CD144 released more proangiogenic and antiapo-ptotic factors than other subpopulations of piPSC-ECs andpASCs. These data suggest that greater functional recoverymay be achieved by transplanting a select population of cellsrather than a heterogeneous mixture of cells.
Transplanted piPSC-ECs Inhibit Apoptosis byTUNEL StainingInterestingly, the same angiogenic genes (ie, VEGF-A, IL6,IL10, TIMP1, GATA4, angiopoietin 1, and CSF3) that areupregulated in the peri-infarct zone after piPSC-EC trans-
plantation also play an important role in regulating apoptosisand promoting cell survival. To explore whether the trans-plantation of piPSC-ECs inhibited cardiomyocyte and endo-thelial cell apoptosis, we measured the amount of DNAfragmentation in explanted hearts of treated and controlanimals (n�3 animals per group) using the TUNEL assay.The density of TUNEL-positive nuclei, shown as the percent-age of total nuclei, was significantly lower in mice trans-planted with piPSC-ECs compared with PBS control(6.1�0.6% versus 18.9�0.9%, P�0.002). These findingssuggest that inhibition of apoptosis by piPSC-ECs may alsocontribute to functional recovery (Online Figure VII).
Imaging piPSC Fate in a Porcine Model of MIUsing PET/CT and MRIBefore the clinical application of iPSCs in humans, successfuldelivery and monitoring of cells in a large animal model mustfirst be established. Swine are an ideal large animal model forthe clinical translation of iPSC-based therapy due to theirresemblance to human organ size and physiology and theirrelatively long lifespan.16 To demonstrate feasibility of trackingpiPSCs in swine using PET/CT and MRI, we injected piPSCsinto Yucatan miniswine that previously underwent LAD arteryballoon catheter occlusion to induce a myocardial infarction(Online Video I). Before intramyocardial cell transplantation,piPSCs were directly labeled with [18F]-FDG for PET imaging,iron particles for co-localization with MRI, and a fluorescent
Figure 5. Cytokine expression array demonstrate piPSC-ECs release paracrine factors with in vitro hypoxia stress. A, Angiogen-esis and antiapoptosis protein array data after hypoxia exposure for 24 hours. B, Quantitative analysis of the cytokine array confirmedsignificant upregulation of several proangiogenic and antiapoptotic related proteins in pASCs, pAorta-ECs, and piPSC-ECs comparedwith control (culture medium without cells under hypoxic conditions). Interestingly, piPSC-ECs released significantly more paracrine fac-tors than pASCs. A similar pattern of paracrine secretion was noted between piPSC-ECs and their endogenous counterparts, pAorta-ECs. Two biological replicates per group (*P�0.05, **P�0.001 versus pASCs).
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 889
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

carbocyanine dye for histological identification. We chose directlabeling of cells with [18F]-FDG rather than reporter geneimaging due to the robustness of this technique and its applica-tion in current clinical trials.37–39 After intramyocardial deliveryof 2�108 piPSCs into the peri-infarct area under direct visual-ization, PET/CT and MRI demonstrated that transplanted cellslocalized to the peri-infarct area of the apical lateral wall of theleft ventricle (Online Figure VIIIA and B). After imaging, heartswere explanted, and immunofluorescence staining demonstratedthe presence of cells expressing the carbocyanine dye (red) aswell as the pluripotent marker Oct-4 (green) in the peri-infarctarea (Online Figure VIIIC). Histology revealed piPSCs with alarge nucleus and abundant cytoplasm in the peri-infarct area(Online Figure VIIID).
DiscussionBecause swine have a close resemblance to humans in termsof anatomy and physiology, the results of this study have
several important implications for the clinical translation ofiPSC therapy. First, we have shown that piPSCs can besuccessfully differentiated into a therapeutic cell populationthat mimics their endogenous counterparts, paving the wayfor the development of additional therapeutic subtypes. Sec-ond, we have demonstrated that the therapeutic effectivenessof piPSC-ECs in repairing the damaged myocardium ismediated by the release of proangiogenic and antiapoptoticmodulating factors, using a novel microfluidic single-cell anal-ysis. Third, we have also shown that the release of paracrinefactors is highest among the subpopulation of piPSC-ECSexpressing the endothelial specific markers CD34 and CD144,suggesting that transplantation of a subpopulation of these cellsmay lead to greater functional recovery. Finally, we haveprovided further validation of in vivo noninvasive multimodalityimaging, facilitating the future development of imaging proto-cols to guide the delivery and monitoring of iPSC derivatives.
Figure 6. Microfluidic single-cell geneexpression profiling demonstratespiPSC-ECs can release paracrine fac-tors in vivo. A, Schematic outline ofsingle-cell expression profiling experimentdemonstrating that paracine factors arereleased in response to the ischemicmicroenvironment. One week after injec-tion into normal versus infarcted hearts,piPSC-ECs were harvested, sorted byFACS, and analyzed by single-cell qRT-PCR. B, Comparison of fold-changeobtained by single-cell qRT-PCR showeda significant increase in the expression of10 genes, which have proangiogenic and/or antiapoptotic effects, in the MI groupcompared with the sham group. Fold-change is defined as the Ct value of theMI group divided by the Ct value of thesham group. Results confirm that piPSC-ECs are able to respond to the local is-chemic milieu by secreting proangiogenicand antiapoptotic factors. C, Schematicoutline of single-cell expression experi-ment showing that the release of para-crine factors varied among different sub-populations of pASC and piPSC-ECs.One week after injection into infarctedhearts, injected cells were harvested, sub-populations of pASCs and piPSC-ECswere isolated using FACS, and single-cellqRT-PCR was performed. D, Comparisonof fold-change obtained by single-cellqRT-PCR showed patterns of paracrinerelease varied among 3 populations ofpiPSC-ECs and ASCs and cells: (1)CD31�/CD34�/CD144�, (2) CD31�/CD34�/CD144�, and (3) CD31�/CD34�/CD144�.
890 Circulation Research September 14, 2012
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

Large-animal iPSCs have been recently derived from anumber of species including monkeys,9 dogs,10,11 hu-mans,12,13 and pigs.14,15 Successful differentiation of thesecells into therapeutic cell populations, however, is necessaryto harness the clinical potential of iPSC technology. Surpris-ingly, except for human iPSCs, reports on large-animal iPSCdifferentiation into therapeutic cell types for the treatment ofcardiovascular disease have been limited. Using a modifiedprotocol designed for human iPSC-EC differentiation,26 wegenerated piPSC-ECs that were morphologically and func-tionally similar to endothelial cells harvested from the autol-ogous porcine aorta, based on standard in vitro and in vivoassays. Interestingly, these cells secrete paracrine factors atcomparable levels and improve cardiac function to a similarextent as their endogenous counterparts. Although cell transplan-tation with native endothelial cells has been shown to promoteangiogenesis and improve cardiac function,40 we believe thatiPSC derivatives are a superior alternative because they have apotentially unlimited source unlike native endogenous endothe-lial cells, which are challenging to harvest and have a restrictedsupply. However, further optimization and modification ofestablished human differentiation protocols are still needed forefficiently generating large quantities of piPSC derivatives suchas endothelial cells or cardiomyocytes.41,42
Although the debate continues over the optimal stem celltype for the clinical application of cardiac regenerativetherapy, iPSCs appear promising because of their unlimitedsupply, immune privilege via autologous transplantation, andlack of ethical or regulatory hurdles impeding their clinicalapplication.43 These cells may also be more effective thanother cell types, such as adult somatic stem cells, which havebeen most often applied in preclinical and clinical trials todate. The efficacy of ASCs, for example, has been demon-strated in several studies with reported benefits due to thesecretion of paracrine factors promoting angiogenesis andcell survival.30 Our head-to-head comparison revealed thatpiPSC-ECs secrete significantly more proangiogenic and anti-apoptotic modulating cytokines than pASCs in vitro, a findingthat is analogous to a recent report showing that iPSC-derivedmesenchymal cells release more paracrine factors than adultbone marrow stem cells.44 Findings from our in vivo studyconfirmed that transplantation of piPSC-ECs resulted in moreneovascularization in the peri-infarct area than transplantation ofpASCs. Additional studies directly comparing the efficacy ofdifferent stem cell types, however, are needed to determine thebest option for clinical translation.
The release of cytokines and growth factors that promoteneovascularization and cytoprotection has emerged as aleading mechanism to explain the observed functional im-provement after stem cell therapy,45 in light of results fromnumerous studies demonstrating poor long-term survival andengraftment of transplanted cells and a paucity of newlygenerated cardiomyocytes.46 Improvement in cardiac func-tion has been associated with stem cell induced release ofVEGF, IL1-�, IL6, TNF-�, FGF-�, TIMP1, TIMP2, andangiopoietin 1,45 all of which were significantly increased inour study. Importantly, in a porcine model of myocardialinfarction, Doyle et al found that pigs treated with conditionedmedia containing paracrine factors produced equivalent im-
provement in regional myocardial function and mass comparedwith those treated with transplanted cells; recovery was attenu-ated after administration of cytokine specific antibodies.34 Inter-estingly, Cho et al found that transplanted cells not only directlyrelease cytokines but also induced the release of these factorsfrom host tissues that is sustained for greater than 2 weeks, muchlonger than donor derived factors.47
Previous findings, however, have been performed in wholecell populations, which are heterogeneous, and may reflectthe average gene expression profile, not necessarily theexpression profile of the cells of interest. Because whole-cellpopulations of iPSCs and their derivatives are even moreheterogeneous than other stem cell types,36 perhaps as a resultof limitations in reprogramming, variations in differentiation,and/or persistent donor cell memory,48,49 we used a novelmicrofluidic platform to examine gene expression at a single-cell level. We confirmed that transplanted piPSC-ECs releaseproangiogenic and antiapoptotic modulators that lead to newvessel formation, decreased apoptosis, and significant func-tional recovery of injured myocardium.50 Despite these en-couraging results, further research is needed to define meth-ods to improve survival and engraftment of transplanted cells,which will not only enhance paracrine release of thesebeneficial factors, but also promote cardiac regeneration.
Because of the heterogeneous nature of iPSC derivatives, wealso compared the pattern of paracine release from differentsubpopulations, specifically cells expressing the endothelialspecific markers CD34 and CD144 (or VE-cadherin). Single-cell expression analysis revealed that piPSC-ECs expressingeither CD34 or CD144 released more proangiogenic and anti-apoptotic factors than other subpopulations of piPSC-ECs andpASCs. This is not surprising, given the function of theseproteins in vasculogenesis. CD34 is a transmembrane proteinthat is expressed on early hematopoietic precursors and vascularassociated structures.51 CD144 (VE-cadherin) is an endothelialspecific adhesion molecule that maintains and controls endothe-lial cell-to-cell adhesion and, thus, can regulate vascular perme-ability and leukocyte extravasation.52 In addition, it regulates cellproliferation, apoptosis, and the function of vascular endothelialgrowth factor, making it is essential for embryonic angiogenesis.Furthermore, previous studies have shown that endothelialprogenitor cells expressing CD34 and CD144 augment angio-genesis by release of paracrine factors.53 Given our findings thatcells expressing these markers release more proangiogenic andantiapoptotic factors than other subpopulations, future studiesshould consider transplantation of specific iPSC-derivative cellsubpopulations to further enhance functional recovery.
Finally, the development of a robust platform for noninva-sive in vivo imaging in large animals is critical to ensure thattherapeutic cells have been safely delivered near the targetarea of injury and have integrated into the target tissue.39 Inthe present study, we directly labeled cells with [18F]-FDGand iron particles for in vivo monitoring by PET/CT andMRI, respectively. Previous studies have reported successfulmonitoring of cell engraftment in large animals using theseapproaches, albeit with other stem cell subtypes.37–39 Simi-larly, we demonstrate the feasibility of this approach formonitoring the piPSC delivery and engraftment into theperi-infarct area. A major limitation of our study is that we
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 891
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

did not evaluate the functional efficacy of piPSC-ECs in aporcine model. This is primarily due to the large number ofanimals that would be needed to demonstrate robust statisticaldifferences, which would need to be carefully evaluated infollow-up studies. Future studies could also determinewhether a dual approach using direct labeling and reportergene expression could exploit the advantages of both tech-niques to monitor stem cell delivery in large animals.
In summary, we successfully derived piPSCs and differen-tiated them into piPSC-ECs, which share similar morpholog-ical and functional properties to their endogenous counter-parts. We found that piPSC-EC therapy promotesproangiogenic and antiapoptotic cytokine release in vitro andin vivo, leading to improved cardiac function in ischemicinjury models. We also used multimodality noninvasiveimaging to monitor cell engraftment into the peri-infarct areain small- and large-animal models of myocardial infarction.Taken together, our study provides preclinical validation andoptimization of piPSCs-ECs as an effective therapeutic cellpopulation, laying the groundwork for the eventual applica-tion of autologous iPSC-based therapy in the future.
AcknowledgmentsWe would like to thank C. Dana Bangs for performing the cytogeneticanalysis for this study. We are grateful to the Stanford FunctionalGenomics Facility for their assistance with the microarray experiment.
Sources of FundingThis study was supported by National Institutes of Health grants DP2OD004437, NIH RC1 HL099117, NIH R01 EB009689, FondationLeducq, NIH R33 HL0890267 (J.C.W.), NIH RC1 HL100490 (M.T.L.),and U01 HL099776 (R.C.R.) and the American College of CardiologyFoundation GE Healthcare Career Development Award (P.K.N.).
DisclosuresNone.
References1. Narsinh K, Narsinh KH, Wu JC. Derivation of human induced pluripotent
stem cells for cardiovascular disease modeling. Circ Res. 2011;108:1146–1156.
2. Yamanaka S. Induced pluripotent stem cells: past, present, and future.Cell Stem Cell. 2012;10:678–684.
3. Pearl JI, Lee AS, Leveson-Gower DB, Sun N, Ghosh Z, Lan F, RansohoffJ, Negrin RS, Davis MM, Wu JC. Short-term immunosuppressionpromotes engraftment of embryonic and induced pluripotent stem cells.Cell Stem Cell. 2011;8:309–317.
4. Zhao T, Zhang ZN, Rong Z, Xu Y. Immunogenicity of induced pluri-potent stem cells. Nature. 2011;474:212–215.
5. Aoi T, Yae K, Nakagawa M, Ichisaka T, Okita K, Takahashi K, Chiba T,Yamanaka S. Generation of pluripotent stem cells from adult mouse liverand stomach cells. Science. 2008;321:699–702.
6. Okita K, Nakagawa M, Hyenjong H, Ichisaka T, Yamanaka S. Generationof mouse induced pluripotent stem cells without viral vectors. Science.2008;322:949–953.
7. Hamanaka S, Yamaguchi T, Kobayashi T, Kato-Itoh M, Yamazaki S,Sato H, Umino A, Wakiyama Y, Arai M, Sanbo M, Hirabayashi M,Nakauchi H. Generation of germline-competent rat induced pluripotentstem cells. PLoS One. 2011;6:e22008.
8. Liao J, Cui C, Chen S, Ren J, Chen J, Gao Y, Li H, Jia N, Cheng L, XiaoH, Xiao L. Generation of induced pluripotent stem cell lines from adultrat cells. Cell Stem Cell. 2009;4:11–15.
9. Liu H, Zhu F, Yong J, et al. Generation of induced pluripotent stem cellsfrom adult rhesus monkey fibroblasts. Cell Stem Cell. 2008;3:587–590.
10. Luo J, Suhr ST, Chang EA, Wang K, Ross PJ, Nelson LL, Venta PJ,Knott JG, Cibelli JB. Generation of leukemia inhibitory factor and basic
fibroblast growth factor-dependent induced pluripotent stem cells fromcanine adult somatic cells. Stem Cells Dev. 2011;20:1669–1678.
11. Lee AS, Xu D, Plews JR, et al. Preclinical derivation and imaging ofautologously transplanted canine induced pluripotent stem cells. J BiolChem. 2011;286:32697–32704.
12. Park IH, Zhao R, West JA, Yabuuchi A, Huo H, Ince TA, Lerou PH,Lensch MW, Daley GQ. Reprogramming of human somatic cells topluripotency with defined factors. Nature. 2008;451:141–146.
13. Sun N, Panetta NJ, Gupta DM, Wilson KD, Lee A, Jia F, Hu S, CherryAM, Robbins RC, Longaker MT, Wu JC. Feeder-free derivation ofinduced pluripotent stem cells from adult human adipose stem cells. ProcNatl Acad Sci U S A. 2009;106:15720–15725.
14. Ezashi T, Matsuyama H, Telugu BP, Roberts RM. Generation of colonies ofinduced trophoblast cells during standard reprogramming of porcine fibro-blasts to induced pluripotent stem cells. Biol Reprod. 2011;85:779–787.
15. West FD, Terlouw SL, Kwon DJ, Mumaw JL, Dhara SK, Hasneen K,Dobrinsky JR, Stice SL. Porcine induced pluripotent stem cells producechimeric offspring. Stem Cells Dev. 2010;19:1211–1220.
16. Prather RS, Shen M, Dai Y. Genetically modified pigs for medicine andagriculture. Biotechnol Genet Eng Rev. 2008;25:245–265.
17. Amado LC, Saliaris AP, Schuleri KH, St John M, Xie JS, Cattaneo S,Durand DJ, Fitton T, Kuang JQ, Stewart G, Lehrke S, Baumgartner WW,Martin BJ, Heldman AW, Hare JM. Cardiac repair with intramyocardialinjection of allogeneic mesenchymal stem cells after myocardialinfarction. Proc Natl Acad Sci U S A. 2005;102:11474–11479.
18. Gyongyosi M, Blanco J, Marian T, et al. Serial noninvasive in vivopositron emission tomographic tracking of percutaneously intramyocar-dially injected autologous porcine mesenchymal stem cells modified fortransgene reporter gene expression. Circ Cardiovasc Imaging. 2008;1:94–103.
19. Lim SY, Kim YS, Ahn Y, Jeong MH, Hong MH, Joo SY, Nam KI, Cho JG,Kang PM, Park JC. The effects of mesenchymal stem cells transduced withAkt in a porcine myocardial infarction model. Cardiovasc Res. 2006;70:530–542.
20. Johnston PV, Sasano T, Mills K, Evers R, Lee ST, Smith RR, Lardo AC, LaiS, Steenbergen C, Gerstenblith G, Lange R, Marban E. Engraftment, differ-entiation, and functional benefits of autologous cardiosphere-derived cells inporcine ischemic cardiomyopathy. Circulation. 2009;120:1075–1083.
21. Narmoneva DA, Vukmirovic R, Davis ME, Kamm RD, Lee RT. Endo-thelial cells promote cardiac myocyte survival and spatial reorganization:implications for cardiac regeneration. Circulation. 2004;110:962–968.
22. Brutsaert DL. Cardiac endothelial-myocardial signaling: its role incardiac growth, contractile performance, and rhythmicity. Physiol Rev.2003;83:59–115.
23. Joo HJ, Kim H, Park SW, Cho HJ, Kim HS, Lim DS, Chung HM, KimI, Han YM, Koh GY. Angiopoietin-1 promotes endothelial differentiationfrom embryonic stem cells and induced pluripotent stem cells. Blood.2011;118:2094–2104.
24. Li Z, Hu S, Ghosh Z, Han Z, Wu JC. Functional characterization andexpression profiling of human induced pluripotent stem cell- andembryonic stem cell-derived endothelial cells. Stem Cells Dev. 2011;20:1701–1710.
25. Sanchez-Freire V, Ebert AD, Kalisky T, Quake SR, Wu JC. Microfluidicsingle-cell real-time PCR for comparative analysis of gene expressionpatterns. Nat Protoc. 2012;7:829–838.
26. James D, Nam HS, Seandel M, Nolan D, Janovitz T, Tomishima M,Studer L, Lee G, Lyden D, Benezra R, Zaninovic N, Rosenwaks Z,Rabbany SY, Rafii S. Expansion and maintenance of human embryonicstem cell-derived endothelial cells by TGFbeta inhibition is Id1dependent. Nat Biotechnol. 2010;28:161–166.
27. Ezashi T, Telugu BP, Alexenko AP, Sachdev S, Sinha S, Roberts RM.Derivation of induced pluripotent stem cells from pig somatic cells. ProcNatl Acad Sci U S A. 2009;106:10993–10998.
28. Li Z, Wu JC, Sheikh AY, Kraft D, Cao F, Xie X, Patel M, Gambhir SS,Robbins RC, Cooke JP. Differentiation, survival, and function ofembryonic stem cell derived endothelial cells for ischemic heart disease.Circulation. 2007;116:I46–I54.
29. Yamahara K, Sone M, Itoh H, Yamashita JK, Yurugi-Kobayashi T,Homma K, Chao TH, Miyashita K, Park K, Oyamada N, Sawada N, TauraD, Fukunaga Y, Tamura N, Nakao K. Augmentation of neovascularization inhindlimb ischemia by combined transplantation of human embryonic stemcells-derived endothelial and mural cells. PLoS One. 2008;3:e1666.
30. Gimble JM, Katz AJ, Bunnell BA. Adipose-derived stem cells for regen-erative medicine. Circ Res. 2007;100:1249–1260.
892 Circulation Research September 14, 2012
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

31. Schenke-Layland K, Strem BM, Jordan MC, Deemedio MT, HedrickMH, Roos KP, Fraser JK, Maclellan WR. Adipose tissue-derived cellsimprove cardiac function following myocardial infarction. J Surg Res.2009;153:217–223.
32. Fernandes S, Naumova AV, Zhu WZ, Laflamme MA, Gold J, Murry CE.Human embryonic stem cell-derived cardiomyocytes engraft but do notalter cardiac remodeling after chronic infarction in rats. J Mol CellCardiol. 2010;49:941–949.
33. Nakamura Y, Wang X, Xu C, Asakura A, Yoshiyama M, From AH,Zhang J. Xenotransplantation of long-term-cultured swine bone marrow-derived mesenchymal stem cells. Stem Cells. 2007;25:612–620.
34. Doyle B, Sorajja P, Hynes B, Kumar AH, Araoz PA, Stalboerger PG,Miller D, Reed C, Schmeckpeper J, Wang S, Liu C, Terzic A, Kruger D,Riederer S, Caplice NM. Progenitor cell therapy in a porcine acutemyocardial infarction model induces cardiac hypertrophy, mediated byparacrine secretion of cardiotrophic factors including TGFbeta1. StemCells Dev. 2008;17:941–951.
35. Xiong Q, Hill KL, Li Q, Suntharalingam P, Mansoor A, Wang X, JameelMN, Zhang P, Swingen C, Kaufman DS, Zhang J. A fibrin patch-basedenhanced delivery of human embryonic stem cell-derived vascular celltransplantation in a porcine model of postinfarction left ventricularremodeling. Stem Cells. 2011;29:367–375.
36. Narsinh KH, Sun N, Sanchez-Freire V, Lee AS, Almeida P, Hu S, Jan T,Wilson KD, Leong D, Rosenberg J, Yao M, Robbins RC, Wu JC. Singlecell transcriptional profiling reveals heterogeneity of human inducedpluripotent stem cells. J Clin Invest. 2011;121:1217–1221.
37. Doyle B, Kemp BJ, Chareonthaitawee P, Reed C, Schmeckpeper J,Sorajja P, Russell S, Araoz P, Riederer SJ, Caplice NM. Dynamictracking during intracoronary injection of 18F-FDG-labeled progenitorcell therapy for acute myocardial infarction. J Nucl Med. 2007;48:1708–1714.
38. Chen IY and Wu JC. Cardiovascular molecular imaging: focus on clinicaltranslation. Circulation. 2011;123:425–432.
39. Nguyen PK, Lan F, Wang Y, Wu JC. Imaging: guiding the clinicaltranslation of cardiac stem cell therapy. Circ Res. 2011;109:962–979.
40. Okada M, Payne TR, Zheng B, Oshima H, Momoi N, Tobita K, Keller BB,Phillippi JA, Peault B, Huard J. Myogenic endothelial cells purified fromhuman skeletal muscle improve cardiac function after transplantation intoinfarcted myocardium. J Am Coll Cardiol. 2008;52:1869–1880.
41. Ren Y, Lee MY, Schliffke S, Paavola J, Amos PJ, Ge X, Ye M, Zhu S,Senyei G, Lum L, Ehrlich BE, Qyang Y. Small molecule Wnt inhibitorsenhance the efficiency of BMP-4-directed cardiac differentiation ofhuman pluripotent stem cells. J Mol Cell Cardiol. 2011;51:280–287.
42. Burridge PW, Keller G, Gold JD, Wu JC. Production of de novo cardio-myocytes: human pluripotent stem cell differentiation and direct repro-gramming. Cell Stem Cell. 2012;10:16–28.
43. Sun N, Longaker MT, Wu JC. Human iPS cell-based therapy: consider-ations before clinical applications. Cell Cycle. 2010;9:880–885.
44. Lian Q, Zhang Y, Zhang J, Zhang HK, Wu X, Lam FF, Kang S, Xia JC,Lai WH, Au KW, Chow YY, Siu CW, Lee CN, Tse HF. Functionalmesenchymal stem cells derived from human induced pluripotent stemcells attenuate limb ischemia in mice. Circulation. 2010;121:1113–1123.
45. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mechanisms in adult stemcell signaling and therapy. Circ Res. 2008;103:1204–1219.
46. Laflamme MA, Chen KY, Naumova AV, et al. Cardiomyocytes derivedfrom human embryonic stem cells in pro-survival factors enhance function ofinfarcted rat hearts. Nat Biotechnol. 2007;25:1015–1024.
47. Cho HJ, Lee N, Lee JY, Choi YJ, Ii M, Wecker A, Jeong JO, Curry C, QinG, Yoon YS. Role of host tissues for sustained humoral effects afterendothelial progenitor cell transplantation into the ischemic heart. J ExpMed. 2007;204:3257–3269.
48. Ghosh Z, Wilson KD, Wu Y, Hu S, Quertermous T, Wu JC. Persistent donorcell gene expression among human induced pluripotent stem cells contributesto differences with human embryonic stem cells. PLoS One. 2010;5:e8975.
49. Kim K, Doi A, Wen B, et al. Epigenetic memory in induced pluripotentstem cells. Nature. 2010;467:285–290.
50. Quevedo HC, Hatzistergos KE, Oskouei BN, Feigenbaum GS, RodriguezJE, Valdes D, Pattany PM, Zambrano JP, Hu Q, McNiece I, HeldmanAW, Hare JM. Allogeneic mesenchymal stem cells restore cardiacfunction in chronic ischemic cardiomyopathy via trilineage differentiatingcapacity. Proc Natl Acad Sci U S A. 2009;106:14022–14027.
51. Furness SG, McNagny K. Beyond mere markers: functions for CD34family of sialomucins in hematopoiesis. Immunol Res. 2006;34:13–32.
52. Vestweber D. VE-cadherin: the major endothelial adhesion moleculecontrolling cellular junctions and blood vessel formation. ArteriosclerThromb Vasc Biol. 2008;28:223–232.
53. Sieveking DP, Buckle A, Celermajer DS, Ng MK. Strikingly differentangiogenic properties of endothelial progenitor cell subpopulations:insights from a novel human angiogenesis assay. J Am Coll Cardiol.2008;51:660–668.
Novelty and Significance
What Is Known?● Induced pluripotent stem cells (iPSCs) have been created from adult
somatic cells of many small and large animals.● More recently, iPSCs have been derived from swine, an animal that
shares a cardiovascular anatomy and physiology similar tohumans.
What New Information Does This Article Contribute?● We successfully created iPSCs from porcine adipose tissue.● For the first time, we also injected the iPSCs into the myocardium in a
porcine model of myocardial infarction and tracked their localizationto peri-infarct area by multimodality MRI and PET/CT imaging.
● In addition, we successfully differentiated porcine iPSCs into endo-thelial cells (iPSC-ECs) in vitro and demonstrated that intramyo-cardial injection of iPSC-ECs improved cardiac function in amurine model of myocardial infarction.
Although several groups have created iPSCs from porcineadult tissue, the generation of a therapeutic cell type from
these large animals has remained challenging. Using amodified protocol for the derivation of human endothelialcells from iPSCs, we successfully generated endothelial cellsfrom porcine iPSCs that shared similar morphological andfunctional properties as endothelial cells from the aorta.Transplantation of these iPSC-ECs into the murine myocar-dium after myocardial infarction resulted in significant im-provement in ejection fraction 4 weeks after transplantation.Using a novel microfluidic PCR technique to determine geneexpression at the single-cell level, we discovered thatiPSC-ECs are capable of releasing proangiogenic and anti-apoptotic factors in the ischemic environment. These para-crine factors promoted the formation of new blood vessels inthe peri-infarct area. We also demonstrated that the patternof paracrine release varied among different cell subpopula-tions. In summary, further development of these large-animaliPSC models may provide additional insight to facilitate thedevelopment of autologous iPSC-based therapy in humans.
Gu et al Paracrine Activation by Porcine iPSC-Derived ECs 893
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

1 !
SUPPLEMENTAL MATERIAL SUPPLEMENTAL METHODS Porcine adipose stem cell harvest and culture. Adipose tissue and skin were resected from five juvenile, female Yucatan pigs (Pork Power Farms, Turlock CA). Adipose tissue was separated from the overlying skin and digested with 0.2% collagenase II at 37oC on a shaker for 1 to 2 hours, followed by vigorous manually shaking for 5-10 seconds every 20 min. After digestion, the stromal vascular fraction was separated from lipid-filled adipocytes by centrifugation at room temperature at 300x g for 10 min. Each stromal vascular fraction pellet was then re-suspended thoroughly in 20 ml of PBS and filtered through a 100-micron pore size filter. All adherent cells acquired a fibroblast-like morphology within 5-7 days. Primary porcine adipose stromal cells (pASCs) were maintained in Dulbecco's Modified Eagle Medium (DMEM) (Invitrogen, Carlsbad, CA), containing 10% fetal bovine serum (FBS) (Invitrogen), Glutamax-I (Invitrogen), 4.5 g/L glucose (Sigma, St. Louis, MO), 110 mg/L sodium pyruvate (Invitrogen), 50 units/ml penicillin, and 50 µg/ml streptomycin (Invitrogen) at 37oC, 95% air, and 5% CO2 in a humidified incubator. All pASCs used for reprogramming were within passage 15. Lentivirus production and transduction. Lentivirus carrying the reprogramming transgenes (Oct4, Sox2, Klf4, and cMyc) or reporter genes (Fluc-mRFP-HSVttk) was generated as previously described.1, 2 Briefly, 293FT cells (Invitrogen) were transfected with lentiviral vectors carrying the appropriate gene (12 µg lentivirus/gene), packaging plasmids (8 µg), and VSVG plasmids (4 µg) using Lipofectamine 2000 (Invitrogen). Two days after transfection, the supernatant was collected, filtered, and mixed with PEG-it Virus Concentration Solution (System Biosciences) overnight at 4°C. The following day, viruses carrying the appropriate genes were precipitated at 1,500x g and resuspended with Opi-MEM medium (Invitrogen). Immunofluorescence, alkaline phosphatase staining, and triphenyltetrazolium chloride (TTC) staining. After fixation with 4% paraformaldehyde for 10 minutes, cells were blocked with 5% bovine serum albumin in PBS for 1 hour at room temperature. For histological sections requiring nuclear staining, cells were treated with 0.3% tritonX-100 in PBS for 10 min for membrane permeabilization prior to adding antibodies. The following primary antibodies were used: 1) markers of pluripotency: Oct3/4 (Santa Cruz, Santa Cruz, CA), Sox2 (Biolegend, San Diego, CA), Klf4 (Abcam, Cambridge, UK), c-MYC (Abcam), SSEA-1 (Millipore, Billeria, MA), Tra-1-60 (Millipore), Tra-1-81 (Millipore), and Nanog (Santa Cruz); 2) markers of germ layer differentiation: AFP (Santa Cruz), Sox17 (R&D Systems, Minneapolis, MN), SMA (Santa Cruz), Desmin (Sigma, St. Louis, MO), and Nestin (Millipore). AlexaFluor conjugated secondary antibodies (Invitrogen, Carlsbad, CA) were used after primary antibody incubation for detection by fluorescent microscopy. For alkaline phosphatase (AP) staining, the quantitative alkaline phosphatase ES characterization kit (Millipore Chemicon, Billeria, MA) was used according to manufacturer’s instructions. For histological analysis by TTC, H&E, and TUNEL staining, animals were sacrificed and their hearts explanted at week 4. For TTC staining (n=5 per group), tissue sections were incubated in TTC for 15-20 minutes. After living tissue sections turned a deep red and infarcted areas turned white, sections were fixed in 10% formalin for 20 min. NIH Image J software was used to contour the infarcted area. The percent infarct size was calculated as the area of the infarct divided by the total area of the left ventricle. For the TUNEL assay (Roche Applied Science) (n=3 per group), the in situ cell death detection kit was used according to manufacturer’s instructions. After staining, murine sections were examined by
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

2 !
confocal microscopy. Three heart tissue slides were examined per mouse and five different fields were examined per murine section. For capillary density analysis, 3 sections from each mouse heart were stained using a CD31 antibody (BD Pharmingen), followed by a secondary antibody (Alexafluor-594). Capillary density was assessed by counting the number of capillaries in 6 random high-powered fields in each of 3 tissue sections. Images were then analyzed using ImageJ analysis software (NIH) and the data were expressed as capillaries/mm2. Karyotype analysis. Karyotype analyses of pASC-derived piPSCs were performed at the Cytogenetics Laboratory in the Department of Pathology at Stanford Hospital and Clinics. Cells were treated with 0.1 mg/mL colcemid for induction of mitotic arrest and were subsequently harvested by trypsin dispersal, hypotonic shock, and fixation with 3:1 methanol:acetic acid. For each cell line, 20 metaphases were analyzed by the standard G-banding method. Angiogenesis Matrigel plug assay. Using appropriate anesthesia and under sterile conditions, 0.5 ml of Matrigel mixed with 1x106 piPSCs-ECs (n=2 mice) or PBS (n=2 mice) was injected subcutaneously near the dorsal midline of SCID mice, per manufacturer’s instructions. The Matrigel plug formed within seconds subcutaneously as Matrigel solidifies at body temperature. Seven days after implantation, the Matrigel plugs were explanted and prepared for histological and immunohistochemical analysis. Echocardiographic evaluation of murine cardiac function. Echocardiography was performed at 0 (day 2 post MI), 1, 2, and 4 weeks post LAD artery ligation, using a Siemens-Acuson Sequoia C512 system equipped with a multi-frequency (8-14 MHz) 15L8 transducer (n=20 per group). Two-dimensional and M-mode echocardiography of the left ventricular short-axis were used to estimate fractional shortening (FS), as previously described.3, 4 MRI evaluation of murine cardiac function. A subset of mice was randomly selected for MRI evaluation of cardiac function (n=5 per PBS and iPSC-EC group) at 2 and 4 weeks post MI. Using a Signa 3.0T Excite HD scanner (GE, Milwaukee, Wisconsin) equipped with a Mayo Clinic T/R MRI coil (Mayo Clinic Medical Devices, Rochester, Minnesota), mice were placed in the prone position after anesthesia (1.5% isoflurane with 1 ml/min of oxygen). Heart rate, respiratory rate, and body temperature were monitored (Small Animal Instruments, Stony Brook, New York). After cardiac localization using gradient recalled echo (GRE), fast spoiled GRE (FSPGR) was used to acquire 20 short axis cine frames of the heart over 1 cardiac cycle. The following imaging parameters were applied: TR = 10 ms, TE = 4.6 ms, number of excitations (NEX) = 10, field of view (FOV) = 40 × 49 mm, matrix = 256 × 256, flip angle (FA) = 45°, slice thickness 1.5 mm, spacing = 0 mm. Ejection fraction was calculated using a commercial analysis program (Osirix Version 3.81) that was used to trace the endocardial border of the left ventricle (LV) at end diastole and end systole.5 Optical bioluminescence imaging (BLI) and micro positron emission tomography (microPET) for tracking cell fate. To enable in vivo tracking, piPSCs were stably transduced with a triple fusion reporter gene consisting of firefly luciferase, monomeric red fluorescent protein, and herpes simplex virus truncated thymidine kinase (Fluc-mRFP-HSVttk). BLI of the reporter gene FLuc was used to monitor cell survival and engraftment (n=5). Briefly, following intravenous tail injection with 250 mg/kg of D-luciferin, animals were imaged using a Xenogen IVIS Spectrum (Xenogen, Alameda, CA). Image analysis was performed using Living Image 4.0 (Caliper Life Sciences), as previously described.3, 4 MicroPET imaging of the reporter gene HSVttk was performed using a R4 Concorde MicroPET system (n=5). The PET reporter probe 9-[4-[18F]fluoro-3-(hydroxymethyl)butyl]guanine ([18F]-FHBG) (150uCi/animal) was injected into mice. Sixty to seventy five minutes after injection, images were taken and then reconstructed
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

3 !
using a filtered back-projection algorithm. A set of serial microPET images was obtained for 90 minutes (18 frames, 5 min each) to assess tracer influx. Regions of interest (ROIs) were defined for each area of interest. To obtain an image ROI-derived [18F]-FHBG percentage injected dose per gram (% ID/g), the ROI counts per milliliter per minute were converted to counts per gram per minute (assuming a tissue density of 1 g/mL) and divided by the injected dose, as described previously.2 Angiogenesis cytokine array following hypoxia exposure. To determine whether differentiated piPSC-ECs are capable of producing paracrine factors under hypoxic exposure, angiogenesis cytokine array (Panomics) experiments were performed (2 biological replicates). One million pASCs, pAorta-ECs (positive control), and piPSC-ECs were seeded onto a 6-well plate and exposed to 5% O2 hypoxia for 24 hours. Culture medium was used as the negative control. Supernatants were collected for the angiogenesis protein array. The arrays were hybridized and developed using a Kodak X-OMAT 2000 camera. The total pixel intensity for each spot was combined to calculate the expression intensities. Inter-array data was normalized by using positive control spots (eight per array) on each array. Protein expression levels were also normalized to PBS controls. Single cell gene expression profiling of transplanted piPSC-ECs and ASCs: To determine the gene expression of different pro-angiogenic and anti-apoptotic cytokines in vivo at the single cell level, we harvested the transplanted piPSC-ECs and performed single cell gene expression profiling of piPSC-ECs injected into mice receiving LAD ligation and sham surgeries (n=5 per group). One week after transplantation, animals were sacrificed and hearts were explanted and dissociated using Liberase TH Research Grade high Thermolysin and TM Research Grade medium Thermolysin (Roche Applied Science) per manufacturer’s instructions. For single cell expression profiling, piPSC-ECs were first sorted by FACS into a 96 well 0.2-ml PCR plate for reverse transcription and specific target amplification using two different antibodies to prevent murine cell contamination. For negative selection, we used mouse Anti-H2kb monoclonal antibody, PE conjugated (BD 553570, BD Sciences). For selecting piPSCs, we used a mouse anti-porcine SLA Class I JM1E3 antibody (AM05880FC-N, Acris Antibodies GmbH). Each well contained a reverse transcription-specific target amplification master mix that included the following reagents: 1) 2x CellDirect Reaction Mix (CellsDirect One Step qRT-PCR kit, Invitrogen), 2) SUPERase-In (Applied Biosystems) for prevention of RNA degradation by RNases, 3) Taqman primers defining angiogenic factors (Applied Biosystems) for real time qPCR using the dynamic array, 4) SuperScript II Reverse Transcriptase for specific cDNA synthesis, and 5) Platinum Taq DNA polymerase (Invitrogen) for inhibiting polymerase activity and stopping the reaction at room temperature. Reverse transcription and specific target amplification was conducted on a thermocycler (Applied Biosystems Veriti) using the following parameters: 1) RNA release and reverse transcription at 50°C for 15 minutes, 2) reverse transcription reaction inhibition and activation of Platinum Taq DNA at 70°C for 2 minutes, and 3) specific target amplification for 18 cycles with each cycle consisting of denaturation (95°C for 15 minutes) followed by annealing and extension (60°C for 4 minutes). Quantitative real-time PCR was then conducted on a Fluidigm 48.48 Dynamic Array chip by placing the samples into a 48 microfluidic chamber with angiogenic genes and one housekeeping gene to be analyzed in the opposite chamber. Samples and genes were loaded into the reaction chamber using the Nanoflex 4-IFC controller. Single cell qPCR is performed on a BioMark HD reader in nanoliter reaction volumes, and analyzed using the Fluidigm Real-time PCR analysis software. Results are expressed as threshold cycles (CT), which measures the amount of transcript present in the sample. A total of thirteen genes were analyzed in parallel with three replicates per chip. Two chips were analyzed, Genes were selected based their role in the regulation of angiogenesis
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

4 !
and apoptosis. Genes included positive (VEGF, TNF-α, FGF-β, IFN-γ, IL-1α, IL6, IL8, GATA4, Leptin, PIGF, ANGT1, FGFR2, BCL2, and CSF3) and negative modulators of angiogenesis (IL-10, IL12, Ip10, IFNA1, TIMP1, TIMP2, and ANGT2), which are both required for normal vessel development.6 A subset of these genes also has apoptotic (i.e., TNF, IFN-γ, Ip10, TIMP2, and ANGPT2), and anti-apoptotic effects (i.e., VEGF, IL6, IL10, TIMP1, Leptin, GATA4, ANGPT1, BCL2, and CSF3). Generation of a porcine model of myocardial infarction and intramyocardial delivery of piPSC. After induction of anesthesia (5 mg/kg tiletamine/zolazepam with 4-5% inhaled isoflurane) and intubation, Yucatan miniswine underwent carotid balloon catheter occlusion of the proximal to mid left anterior descending artery under fluoroscopic guidance, as previously described.7 At 12 weeks post myocardial infarction, pigs underwent intramyocardial injection of piPSC (2x108 cells) via lateral thoracotomy. Animals were injected into the peri-infarct zone of the left ventricle at 8 different sites using a 25-gauge syringe (500 µl per injection). Study protocols were approved by the Stanford Animal Research Committee. Animal care was provided in accordance with the Stanford University School of Medicine guidelines and policies for the use of laboratory animals. Tracking transplanted piPSCs in miniswine using positron emission tomography / computed tomography (PET/CT) imaging and MRI. For in vivo PET/CT imaging, piPSCs were directly radiolabeled with [18F]-Fluorodeoxyglucose ([18F]-FDG).8 One hour prior to injection, piPSCs were incubated with 5 mCi of [18F]-FDG in a 10 ml test tube containing 10 U/mL heparin (Roche), 0.1 U/ml recombinant human insulin (Novo Nordisk), and serum-free PBS (pH 7.2) at room temperature. After a 30-minute incubation period, centrifugation was performed at 7x g, 27x g, and then 60x g for 120 seconds each, followed by washing in heparinized PBS. A dose calibrator (CRC 55-TR, Capintec Inc, Ramsey, NJ) was used to measure the radioactivity in the pellet and supernatant. Prior to injection, labeled cells were resuspended into 2 ml of heparinized PBS (10 U/mL). PET/CT imaging was performed with a clinical PET/CT scanner (Discovery LightSpeed Plus; GE Medical Systems, Waukesha, Wisconsin), with the animals in a supine position, as previously described.2 Briefly, an initial three-plane scout was obtained for cardiac localization (30 mAs, 120 kV), followed by a non-enhanced 4-detector CT scan using the following parameters: 5 mm thickness, 38 mm per rotation, 0.8-second gantry rotation time, 120 kV x-ray tube voltage, and 150 mA tube current. A 1-hour static non-gated PET scan was acquired two hours after intramyocardial injection of 2x108 piPSCs. CT data sets were then reconstructed in the transverse plane with 5 mm section thickness at a 4.25 mm interval. An iterative algorithm (ordered-subset expectation maximization, 2 iterative steps, 28 subsets) with CT-based attenuation correction was used to reconstruct the images. For cellular localization by MRI, piPSCs were magnetically labeled with iron particles (Resovist, Schering AG, Germany) at a concentration of 250 µg Fe/ml and protamine sulfate (5 µg/ml) for 24 h, as previously described.9 MR images were acquired by using a Signa 3.0 Tesla Excite HD Scanner (GE Medical Systems, Milwaukee, Wisconsin) and an 8-element cardiac phased array coil. For imaging cells labeled with iron, a T2 weighted GRE sequence was performed in two orthogonal views. 9, 10 Evaluation of left ventricular function and infarct size was also performed using steady state free precession and gadolinium delayed enhancement sequences, respectively.11 Finally, cells were identified by histology by labeling with a fluorescent carbocyanine dye (Vybrant CM-Dil, Invitrogen) that binds to phospholipid cell membranes. Labeling was performed prior to cell transplantation per manufacturer’s instructions.
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

5 !
ONLINE SUPPLEMENTAL FIGURE LEGEND Online Figure I: Successful derivation of piPSCs from pASCs. A) piPSC colonies stain positive for alkaline phosphatase (AKP), a universal marker of pluripotency for all types of pluripotent cells. B) Immunofluorescence staining shows expression of additional pluripotent markers (e.g., Tra1-60 and Tra1-81 shown in green) in piPSCs. DAPI nuclear staining shown in blue. C) Karyotyping of piPSCs at passage 20 shows normal porcine karyotype: 2n=38, XX. Online Figure II: piPSCs are capable of in vitro and in vivo differentiation into the three germ layers. A) Immunofluorescence staining of differentiated piPSCs. Differentiated cells are positive for endodermal (AFP and Sox-17), mesodermal (SMA and Desmin), and ectodermal (Nestin) markers. B) Quantitative-PCR analysis confirms that differentiated piPSCs express genes from all three germ layers. Spontaneously differentiated piPSCs (e.g., iPS1-EB, iPS2-EB, iPS3-EB) express genes from the endoderm (Foxa2, AFP), mesoderm (GATA4, GATA6, MLC, Nkx), and ectoderm (TJF, Gbx). These genes, however, are not expressed at detectable levels in piPSCs and pASCs. Foxa2: forkhead box protein A2; AFP: alpha-fetoprotein; GATA4: GATA-binding protein 4; GATA6: GATA-binding protein 6; MLC: myosin light chain; Gbx: gastrulation-brain-homeobox. C) Hematoxylin and eosin (H&E) staining of a teratoma derived from porcine iPSCs reveal the presence of tissues from all three germ layers, shown by the asterisk: 1) neural epithelium (ectoderm), chondrogenic stroma (mesoderm), and 3) glandular epithelium (endoderm). Online Figure III: Transplantation of cells expressing the triple fusion reporter gene for in vivo cell tracking by BLI and microPET. A) Diagram for the triple fusion reporter gene expressing firefly luciferase (Fluc), monomeric red fluorescent protein (mRFP), and herpes simplex virus truncated thymidine kinase (HSVttk) driven by the ubiquitin promoter (top). Expression of Fluc and HSVttk enables in vivo cell tracking by BLI and PET, respectively. Expression of mRFP ex vivo confirms stable integration of the construct in cells and identifies cell engraftment for post-mortem histology (bottom). B) Representative BLI of teratoma development in the kidney capsule of a SCID mouse after injection of 1x106 piPSCs. Continuous growth of the tumor is noted over 60 days as demonstrated by BLI. C) Representative microPET images of a SCID mouse after intramyocardial injection of 1x106 piPSC-ECs. Transverse and coronal images of [18F]-FHBG uptake in the anterior wall of the left ventricle. Online Figure IV: In vivo angiogenesis within Matrigel plugs mixed with piPSC-ECs. Ten days after implantation into the dorsum of SCID mice (n=2 per group), plugs were explanted. A) Representative images of Matrigel plugs mixed with piPSC-ECs showing formation of vascular-like structures. B) Similar structures are not observed in Matrigel plugs mixed with PBS. C) Representative immunofluorescence staining of Matrigel plugs mixed with piPSC-ECs. Vascular-like structures in the Matrigel plugs show strong expression of the endothelial marker CD31. D) Representative hematoxylin and eosin (H&E) sections from the Matrigel plugs mixed with piPSC-ECs confirm the presence of capillary formations. Online Figure V: Improvement of left ventricular function after delivery of piPSC-ECs confirmed by MRI. A) Representative MRI short-axis views taken at the level of the papillary muscles at week 2 and 4 post-MI in mice treated with piPSC-ECs and PBS control. Increased
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

6 !
wall thickening and contractility is noted at end systole in mice receiving piPSC-ECs compared to PBS control. B) Quantification of left ventricular ejection fraction (LVEF) reveals significant improvement at week 2 and 4 post MI (*P<0.05) Online Figure VI: CD31 staining of the microvasculature using anti-porcine and anti-murine antibodies revealed that porcine iPSC-ECs do not form discrete vasculature, but can be found adjacent to murine vessels. A) Immunofluorescence staining with anti-porcine CD31 antibodies (green) and B) anti-murine CD 31 antibodies (red) in areas of increased capillary formation from explanted hearts show that microvessels are only immunoreactive to murine-specific antibodies. Overlay images at low (C) and high (D) magnification demonstrate that piPSC-ECs can be found surrounding murine microvessels, supporting our findings that piPSC-ECs release pro-angiogenic factors that increase native vessel formation. Scale bars=200 µm at low magnification and µm at high magnification. Online Figure VII: Decreased apoptosis in murine hearts injected with iPSC-ECs compared to those injected with PBS. Four weeks after treatment, murine hearts were explanted and mouse frozen sections were examined for DNA fragmentation using TUNEL assay. A) Immunofluorescence staining with TUNEL demonstrates fewer cardiomyocytes with DNA fragmentation in the transplanted vs. control group (shown in green). DAPI nuclear staining shown in blue. Cardiac troponin T shown in red. Overlay of TUNEL, DAPI and cardiac troponin T (far right). B) Quantification of the density of TUNEL positive nuclei. Murine hearts transplanted with piPSC-ECs have a lower average density of apoptotic cells (6.1±0.6% compared to 18.9±0.9%,*P<0.005). Online Figure VIII: In vivo tracking of injected piPSCs by PET/CT and MRI in a porcine model of myocardial infarction. piPSCs were directly labeled with [18F]-FDG, iron oxide particles, and carbocyanine dye (i.e., Vybrant CM-Dil) for localization by PET/CT, MRI, and histological analysis, respectively. A) Representative axial non-enhanced PET-CT fusion image of piPSCs (left) with coronal reconstruction of PET data set (right) one hour after delivery of cells into the peri-infarct area. A strong signal (0.05±0.2 %ID/g) can be seen at the sites of the injection in the distal left ventricular wall (noted by single arrow). B) Representative T2 weighted GRE images by MRI shows iron-labeled cells as hypointense signals (dark areas noted by the single arrow) in the apical lateral wall (left), which corresponds to areas of strong signal noted on PET imaging. Cells were injected near the region of infarct, noted by areas of hyperenhancement on gadolinium enhanced MRI (bright white areas shown on right). C) Immunostaining reveals the expression of Vybrant CM-Dil fluorescent dye (red), Oct4 (green), and DAPI (blue) cells injected into the porcine myocardium. Overlay of Vybrant CM-Dil fluorescent dye, Oct4, and DAPI (far right). D) Histological sections of porcine hearts confirm the absence of cells in the adjacent, non-injected myocardium (left) and successful delivery of the piPSCs in the myocardium (center). piPSCs have a large nucleus (12-14 µm) with significant cytoplasmic volume (single white arrow on 100x high power field). Online Video I: Cine of MRI of left ventricular function in a porcine model of myocardial infarction. Representative short-axis steady-state free precession cine MRI of left ventricle in a pig that underwent balloon occlusion of the left anterior descending artery. Ejection fraction is moderately to severely depressed post-MI. Thinning and poor contractility of the mid to distal anterior wall and apex are noted. Online Table I: Primer sequences for evaluating pluripotency by RT-PCR. A list of primer sequences used to perform RT-PCR to confirm that generated porcine induced pluripotent stem
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

7 !
cells express pluripotency genes and can successful differentiate into the three embryonic germ layers.
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

8 !
Supplemental References 1. Sun N, Panetta NJ, Gupta DM, Wilson KD, Lee A, Jia F, Hu S, Cherry AM, Robbins RC,
Longaker MT, Wu JC. Feeder-free derivation of induced pluripotent stem cells from adult human adipose stem cells. Proc Natl Acad Sci U S A. 2009;106:15720-15725.
2. Willmann JK, Paulmurugan R, Rodriguez-Porcel M, Stein W, Brinton TJ, Connolly AJ, Nielsen CH, Lutz AM, Lyons J, Ikeno F, Suzuki Y, Rosenberg J, Chen IY, Wu JC, Yeung AC, Yock P, Robbins RC, Gambhir SS. Imaging gene expression in human mesenchymal stem cells: from small to large animals. Radiology. 2009;252:117-127.
3. Li Z, Wu JC, Sheikh AY, Kraft D, Cao F, Xie X, Patel M, Gambhir SS, Robbins RC, Cooke JP. Differentiation, survival, and function of embryonic stem cell derived endothelial cells for ischemic heart disease. Circulation. 2007;116:I46-54.
4. Esteban MA, Wang T, Qin B, Yang J, Qin D, Cai J, Li W, Weng Z, Chen J, Ni S, Chen K, Li Y, Liu X, Xu J, Zhang S, Li F, He W, Labuda K, Song Y, Peterbauer A, Wolbank S, Redl H, Zhong M, Cai D, Zeng L, Pei D. Vitamin C Enhances the Generation of Mouse and Human Induced Pluripotent Stem Cells. Cell Stem Cell. 2009.
5. Huang M, Nguyen P, Jia F, Hu S, Gong Y, de Almeida PE, Wang L, Nag D, Kay MA, Giaccia AJ, Robbins RC, Wu JC. Double knockdown of prolyl hydroxylase and factor-inhibiting hypoxia-inducible factor with nonviral minicircle gene therapy enhances stem cell mobilization and angiogenesis after myocardial infarction. Circulation. 2011;124:S46-54.
6. Pepper MS. Positive and negative regulation of angiogenesis: from cell biology to the clinic. Vasc Med. 1996;1:259-266.
7. Amado LC, Saliaris AP, Schuleri KH, St John M, Xie JS, Cattaneo S, Durand DJ, Fitton T, Kuang JQ, Stewart G, Lehrke S, Baumgartner WW, Martin BJ, Heldman AW, Hare JM. Cardiac repair with intramyocardial injection of allogeneic mesenchymal stem cells after myocardial infarction. Proc Natl Acad Sci U S A. 2005;102:11474-11479.
8. Hofmann M, Wollert KC, Meyer GP, Menke A, Arseniev L, Hertenstein B, Ganser A, Knapp WH, Drexler H. Monitoring of bone marrow cell homing into the infarcted human myocardium. Circulation. 2005;111:2198-2202.
9. Li Z, Suzuki Y, Huang M, Cao F, Xie X, Connolly AJ, Yang PC, Wu JC. Comparison of reporter gene and iron particle labeling for tracking fate of human embryonic stem cells and differentiated endothelial cells in living subjects. Stem Cells. 2008;26:864-873.
10. Li Z, Lee A, Huang M, Chun H, Chung J, Chu P, Hoyt G, Yang P, Rosenberg J, Robbins RC, Wu JC. Imaging survival and function of transplanted cardiac resident stem cells. J Am Coll Cardiol. 2009;53:1229-1240.
11. Kim RJ, Fieno DS, Parrish TB, Harris K, Chen EL, Simonetti O, Bundy J, Finn JP, Klocke FJ, Judd RM. Relationship of MRI delayed contrast enhancement to irreversible injury, infarct age, and contractile function. Circulation. 1999;100:1992-2002.
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

Online Figure I �
AP#
BTra'1'60#
Tra'1'81#
DAPI#
DAPI#
piPSC A AP#
pASC
Tra'1'60#
Tra'1'81#
DAPI#
DAPI#
C
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
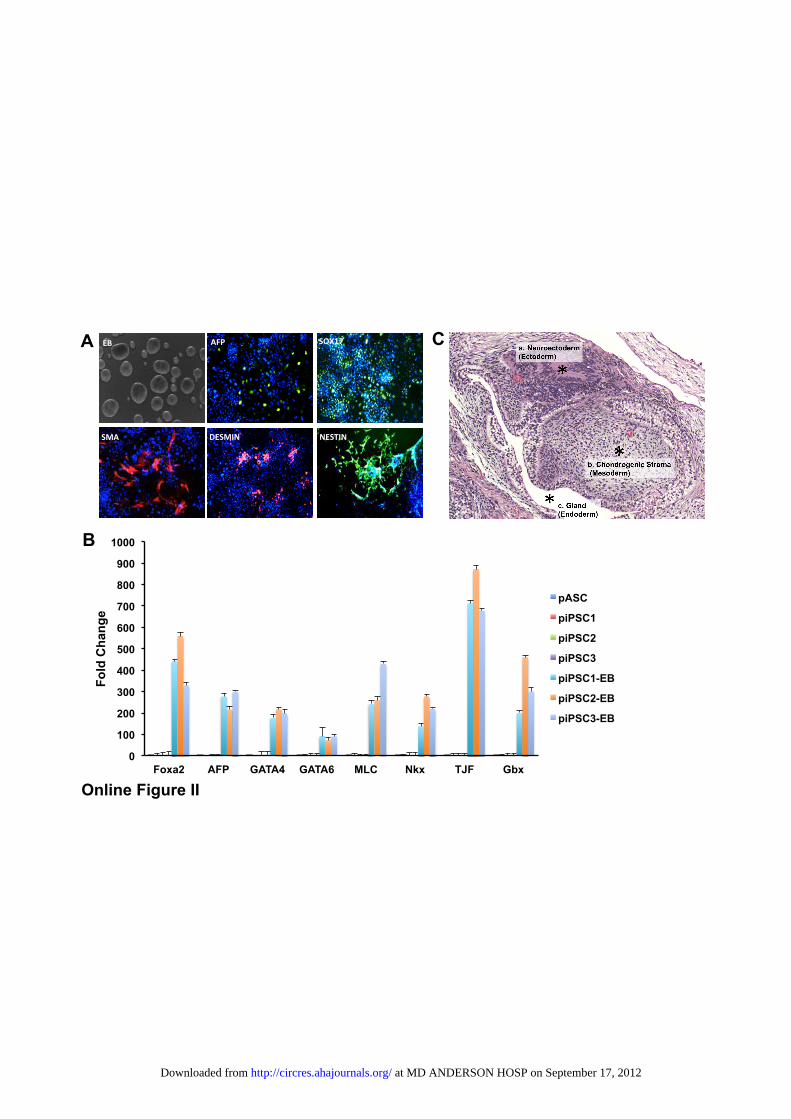
0
100
200
300
400
500
600
700
800
900
1000
Foxa2 AFP GATA4 GATA6 MLC Nkx TJF Gbx
Fold
Cha
nge
pASC
piPSC1
piPSC2
piPSC3
piPSC1-EB
piPSC2-EB
piPSC3-EB
B
AFP$#
A
DESMIN$#
DAPI$#
DAPI$#
SMA$#
SOX17#
$DESMIN$#
SOX17#AFP#
$NESTIN#
$EB# C
Online Figure II �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

Day 2 Week 2 Week 4 Week 8
p/s/
cm2 /s
r
1e5
1e9 B
A
transverse
coronal
piPSC-EC
C pUbiquitin Fluc mRFP HSVttk
Online Figure III �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
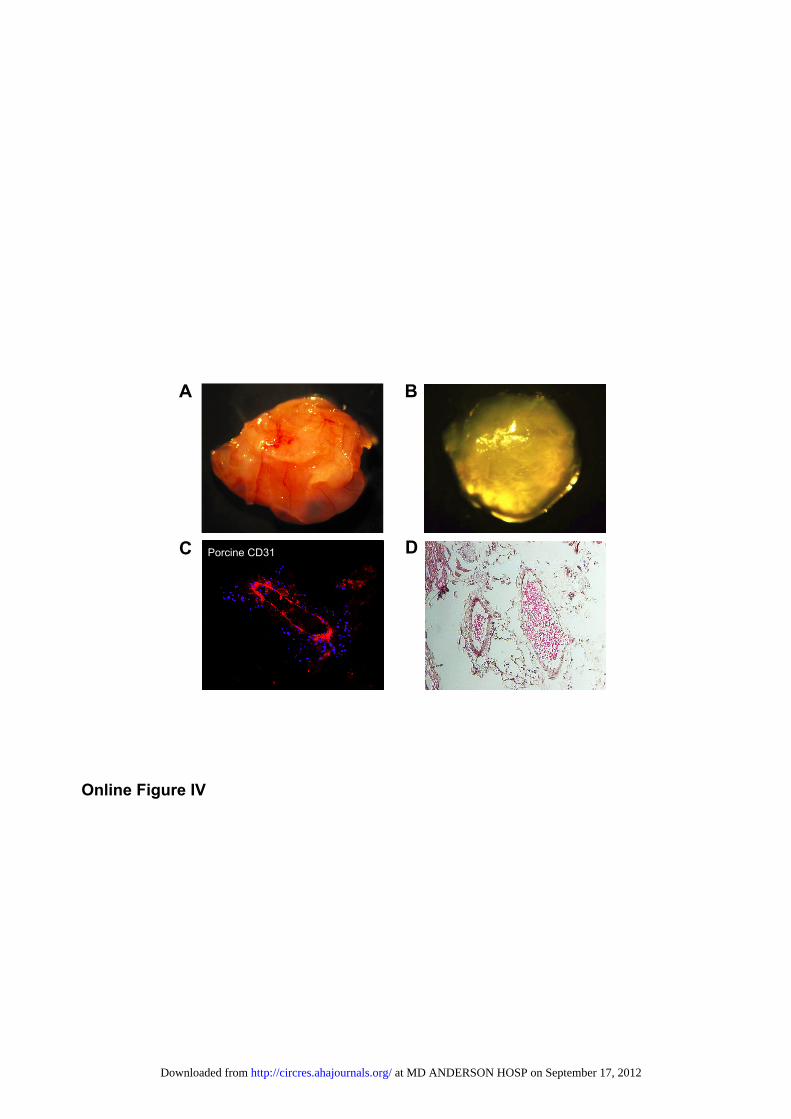
Porcine CD31
A
D C
B
Online Figure IV �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

30
35
40
45
50
Week 2 Week 4
LVEF
(%)
PBS
piPSC-EC
A$
B$*
*
• PB
S pi
PSC
-EC
Systole Systole Diastole Diastole Week 4 Week 2
Online Figure V �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from

Porcine CD31 A
Mouse CD31 B
C D
Online Figure VI �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
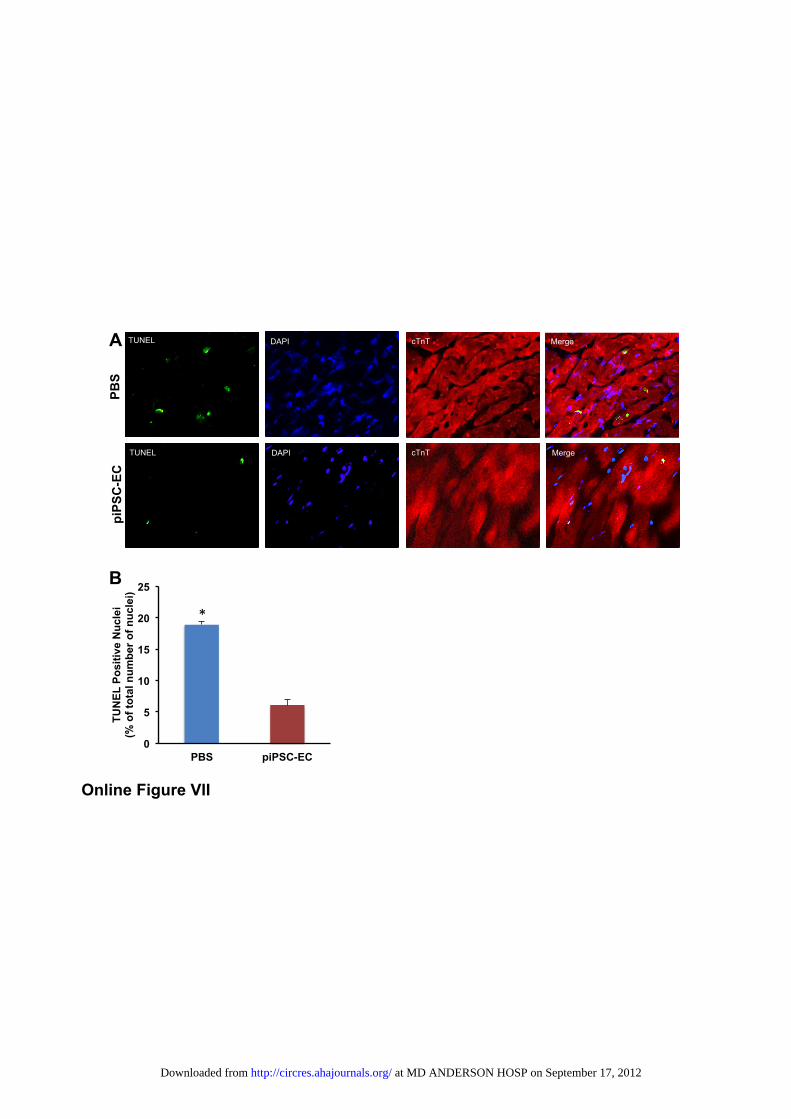
0
5
10
15
20
25
PBS piPSC-EC
TUN
EL P
ositi
ve N
ucle
i (%
of t
otal
num
ber o
f nuc
lei)
TUNEL DAPI cTnT Merge
TUNEL DAPI cTnT Merge
PBS
piPS
C-E
C
B
A
*#
TUNEL
Online Figure VII �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
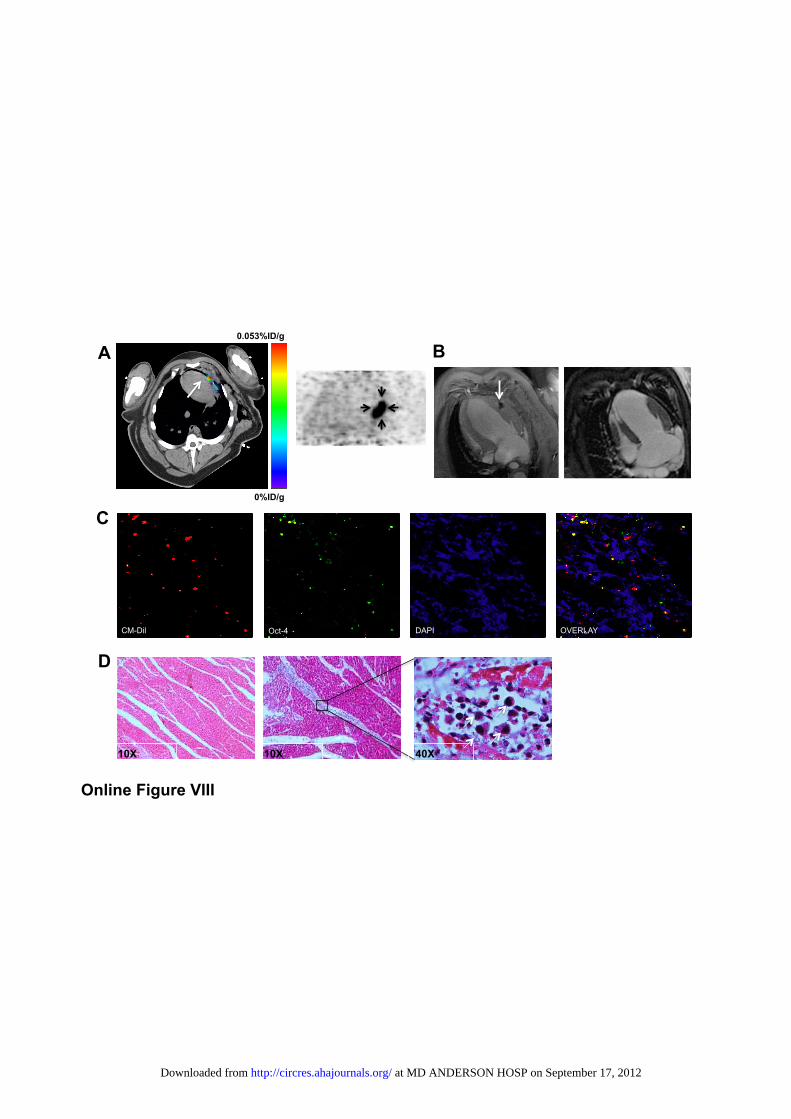
1.5#
B
D
A
C
Oct-4 DAPI OVERLAY CM-Dil
0.053%ID/g
0%ID/g
40X 10X 10X
Online Figure VIII �
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
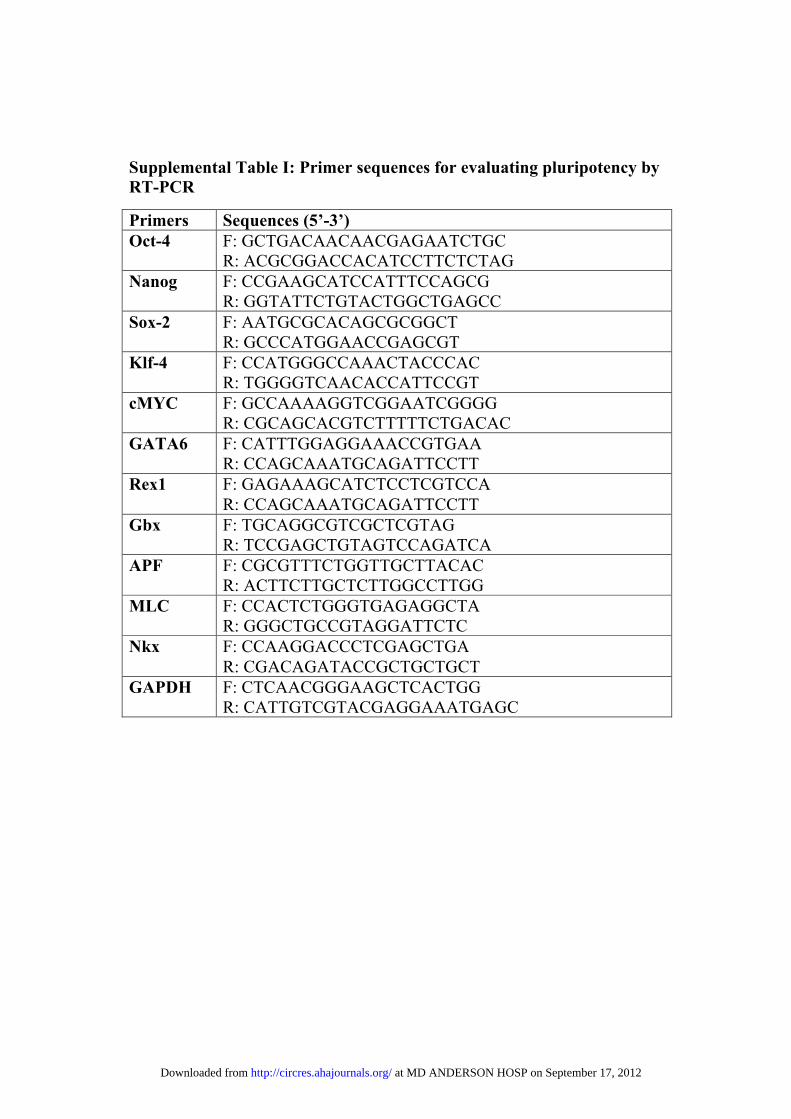
Supplemental Table I: Primer sequences for evaluating pluripotency by RT-PCR
Primers Sequences (5’-3’) Oct-4 F: GCTGACAACAACGAGAATCTGC
R: ACGCGGACCACATCCTTCTCTAG Nanog F: CCGAAGCATCCATTTCCAGCG
R: GGTATTCTGTACTGGCTGAGCC Sox-2 F: AATGCGCACAGCGCGGCT
R: GCCCATGGAACCGAGCGT Klf-4 F: CCATGGGCCAAACTACCCAC
R: TGGGGTCAACACCATTCCGT cMYC F: GCCAAAAGGTCGGAATCGGGG
R: CGCAGCACGTCTTTTTCTGACAC GATA6 F: CATTTGGAGGAAACCGTGAA
R: CCAGCAAATGCAGATTCCTT Rex1 F: GAGAAAGCATCTCCTCGTCCA
R: CCAGCAAATGCAGATTCCTT Gbx F: TGCAGGCGTCGCTCGTAG
R: TCCGAGCTGTAGTCCAGATCA APF F: CGCGTTTCTGGTTGCTTACAC
R: ACTTCTTGCTCTTGGCCTTGG MLC F: CCACTCTGGGTGAGAGGCTA
R: GGGCTGCCGTAGGATTCTC Nkx F: CCAAGGACCCTCGAGCTGA
R: CGACAGATACCGCTGCTGCT GAPDH F: CTCAACGGGAAGCTCACTGG
R: CATTGTCGTACGAGGAAATGAGC
at MD ANDERSON HOSP on September 17, 2012http://circres.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















