
Effect of Different Sprouting Conditions on Alpha Amylase ...

Experimental Neurology 178, 1–12 (2002)doi:10.1006/exnr.2002.8017
Metallothionein-III Inhibits Initial Neurite Formation in DevelopingNeurons as Well as Postinjury, Regenerative Neurite Sprouting
R. S. Chung,1 J. C. Vickers, M. I. Chuah, B. L. Eckhardt, and A. K. WestNeuroRepair Group, University of Tasmania, P.O. Box 252-58, Hobart, Tasmania 7001, Australia
Received January 29, 2002; accepted August 7, 2002
Human metallothionein-III (MT-III) is an inhibitoryfactor deficient in the Alzheimer’s disease (AD) brain.MT-III has been identified as an inhibitor of neuritesprouting, and its deficiency has been proposed to beinvolved in the formation of neurofibrillary tangles(NFT) in the neuropathology of AD. However, therehas been limited investigation of the proposed neuritegrowth inhibitory properties of MT-III. We have ap-plied recombinant human MT-III to both single cellembryonic cortical neurons (to investigate initial neu-rite formation), as well as mature (21 days postplating)clusters of cortical neurons (to investigate the regen-erative sprouting response following injury). We re-port that MT-III inhibited the initial formation of neu-rites by rat embryonic (E18) cortical neurons. This wasbased on both the percentage of neurite positive neu-rons and the number of neurites per neuron (45 and30% inhibition, respectively). Neurite inhibition wasonly observed in the presence of adult rat brain ex-tract, and was also reversible following replacement ofMT-III-containing medium. MT-III inhibited the for-mation and growth of both axons and dendrites. Ofmore physiological significance, MT-III also inhibitedthe regenerative neurite sprouting response followingaxonal transection. The morphology of sprouting neu-rites was also altered, with the distal tip often endingin bulb-like structures. Based on these results, we pro-pose that MT-III, in the presence of brain extract, is apotent inhibitor of neurite sprouting, and may be in-volved in abnormal sprouting potentially underlyingboth AD and epilepsy. © 2002 Elsevier Science (USA)
Key Words: metallothionein; growth inhibitory fac-tor; neurite development; neurite sprouting; reactivesprouting; Alzheimer’s disease.
INTRODUCTION
Alzheimer’s disease is defined pathologically by�-amyloid plaques and neurofibrillary tangles (NFTs),
1 To whom correspondence should be addressed. E-mail:[email protected].
which are accompanied by substantial neuronal lossand aberrant neurite sprouting (1). The relationshipbetween NFTs and �-amyloid deposits, and their rolein the neuropathy of AD, is unclear. However, it hasbeen noted that preclinical cases of AD are character-ized by the presence of �-amyloid deposits without theneurofibrillary changes associated with the latterstages of the disease (2). This suggests that the depo-sition of insoluble �-amyloid plaques precedes neurofi-brillay changes and the subsequent neuronal degener-ation observed in AD. Indeed, it has been hypothesizedthat �-amyloid deposition may result in structural in-jury to surrounding axons, resulting in an abnormalregenerative response and subsequent neurofibrillarychanges (2).
It has also been proposed that an increase in neuro-trophic activity of AD brain leads to neurofibrillarychanges (3). Indeed, Uchida et al. (4) identified aninhibitory factor deficient in the Alzheimer’s diseasebrain, which they named growth inhibitory factor. Thisprotein was subsequently found to resemble the metal-lothionein (MT) family of proteins, and hence has beenrenamed metallothionein-III (MT-III). MT-III exhibitsneuronal-inhibitory activity on cortical neuron cul-tures, as assayed by the level of anti-MAP-2 immuno-labeling of neurons quantified by ELISA (4). This assaymeasures primarily cell number (and hence neuronalsurvival), as discussed by Uchida and Tomonaga (3).Indeed, direct cell counting measurements support thefinding that MT-III (in the presence of brain extract)exhibits neurotoxicity on cultured cortical neurons (5,6). Uchida et al. (4) also proposed that MT-III exhibitsneurite growth inhibitory properties. Using a rat cor-tical neuron assay system, we have examined thisproperty of MT-III and found that MT-III can inhibitinitial neurite formation by cultured cortical neuronsin a dose-dependent manner, but only in the presenceof brain extract. Further, we have found that MT-III, inthe presence of brain extract, can inhibit regenerativeneurite sprouting following axonal transection of ma-ture cultured cortical neurons. Regenerating neuritesalso displayed altered morphology, often ending in
1 0014-4886/02 $35.00
© 2002 Elsevier Science (USA)All rights reserved.

bulb-like structures, as opposed to the growth cone-likestructures often observed in vehicle-treated neurons.
MATERIALS AND METHODS
Construction of the Recombinant Expression Vectorfor Human MT-III
Recombinant human MT-III was prepared essen-tially as reported previously (5), using RT-PCR sub-clones in the bacterial expression vector pET-3d (No-vagen). The structure of the construct was confirmedby DNA sequencing. Recombinant MT was induced inthe presence of ZnSO4 and was purified as describedpreviously (6). Following anion-exchange chromatogra-phy, purified MT was then acidified with dilute HCl topH 2.0, and bound Zn(II) ions were removed by apply-ing the sample to an ultrafiltration column. The apo-metallothionein was stored in aliquots at �80°C. Theapo-metallothionein was reconstituted with Zn(II) bythe addition of 7.5 mol equivalents followed by neutral-ization of the sample to pH 8.0 with 20 mM Tris-HCl.The sample was desalted by ultrafiltration.
Preparation of Rat Brain Extract
Rat brain extracts were prepared as described pre-viously (7). Briefly, a freshly removed whole brain fromadult male Hooded Wistar rat (200 g) was homogenizedwith a Potter-Elvjhem homogenizer in 5 vol of ice-coldPBS, followed by centrifugation of the suspension at30,000g, 4°C. The supernatant was stored at �80°C.We found no difference between frozen and freshlyprepared rat brain extract (data not shown). Followingthawing, the brain extract was filter-sterilized(0.22-�m filter, Millipore), and the protein concentra-tion determined by the Bradford method (8). Extractswere used immediately at a protein concentration of150 �g/ml. Over the course of experiments, a total of 6different rat brain extracts were used. However, withineach individual experiment, rat brain extract from thesame rat was used.
Single Cell Neuron Cultures
Cortical tissue was removed from Embryonic Day 18(sperm-positive day � E1) Hooded Wistar rat embryosand incubated in sterile 10 mM Hepes buffer (37°C).This was followed by trypsin digestion (2.5%, Sigma),followed by addition of trypsin inhibitor (40 BTEEunits/mg protein; Sigma). Three gentle washes of thecell pellet were made using fresh Hepes buffer. The cellsuspension was then triturated carefully using a 1-mlpipette. The cell suspension was filtered through gauzeof pore size 60 �m, and cells were counted by trypanblue exclusion. Cells were then plated onto glass cov-erslips (132 mm2) precoated overnight with poly-L-ly-sine (0.01%), at a cell density of 1 � 105 cells/well, with
150 �g/ml rat brain extract added. Unless stated oth-erwise, MT-III at the appropriate concentration (0.01,0.1, 1, and 5 �g/ml), or 0.1% PBS (vehicle) was added atthis time. Cultures were maintained at 37°C in humid-ified air containing 5% CO2 for 3 days after MT addi-tion. A specific culture medium developed for selectiveneuronal growth was used (9, 10), consisting of Neuro-basal medium (GIBCO), supplemented with 0.1% (f/c)B-27 supplement (GIBCO), 0.1 mM (f/c) L-glutamine(Sigma), and 200 U/ml gentamicin (DBL).
Neuron Cluster Cultures
Neurons were obtained as described above, exceptthat the neurons were plated directly from the unfil-tered cell suspension onto glass coverslips (254 mm2) ata cell density of 4.5 � 105 cells/well and incubatedovernight at 37°C, 5% CO2. The increased plating den-sity promotes neuronal cluster formation, with thickaxonal bundles forming between the clusters (13).These coverslips had been precoated overnight withpoly-L-lysine (0.01%). The next day, the culture me-dium was replaced with fresh medium, and the neu-rons were maintained for 21 days (initial plating is Day0), with the medium replaced with fresh medium every4 days. The medium used was as described previouslyfor single cell neuron cultures.
Transection of Axonal Bundles
Transection of axonal bundles was essentially asdescribed previously (13). Briefly, at 20 days after ini-tial plating, coverslips were moved to individual petridishes (approximately 2900 mm2) with 150 �g/ml ratbrain extract added. The following day, neurite bun-dles were transected on an inverted microscope (LeitzFluovert) using a fine goniotomy knife (Kaisers). Im-mediately following transection, 1 �g/ml of MT-III oran equivalent volume of 0.1% PBS was added, and thecultures were maintained for 12 or 18 h.
Fluorescent Immunohistochemistry of CulturedNeurons
At the appropriate time, cells were fixed with 4%paraformaldehyde, for 20 min at room temperature.Coverslips were then incubated with a variety of pri-mary antibodies (anti-tau, anti-MAP-2 and anti-�III-tubulin) diluted in 0.1% PBS, 0.03% Triton X-100.Anti-tau labels the microtubule-associated tau protein,found primarily in axons (11). Anti-MAP-2 labels mi-crotubule-associated protein-2, found primarily in den-drites (12). Anti-�III-tubulin labels the major buildingblock of microtubules, and is neuron specific (14). Cov-erslips were then incubated with two secondary anti-bodies (horse anti-mouse IgG conjugated to Alexafluor488 and goat anti-rabbit IgG conjugated to Alexafluor594, 1:1000 dilution; Molecular Probes), applied in
2 CHUNG ET AL.

0.1% PBS. Coverslips were mounted onto slides usingPermafluor mounting medium (Immunotech). In somecases nuclear yellow staining was performed. Follow-ing fixation, a 0.1% solution of nuclear yellow wasapplied to the coverslip and incubated at room temper-ature for 5 min, and the coverslip subsequently washed3 times with PBS. In some experiments, neuronal sur-vival was assessed by propidium iodide (Sigma) up-take.
Calculations and Analysis of Neuron Cultures
For quantatative analysis of the effect of MT-III oninitial neurite formation, 10 bright-field digital imageswere captured in a systematic manner (2 images perquadrant, and 2 from the central region of the cover-slip), for each timepoint, at a magnification of 400�(Olympus BL-51). Groups of three coverslips were usedfor each data point. Neuron viability was determinedby trypan blue exclusion, and only viable cells werecounted for all analysis. The total number of cellscounted per coverslip ranged from 300 to 500 cells. Inall cases, nuclear yellow staining was used to investi-gate neuronal survival. Cells which exhibited frag-mented or unusual nuclei were considered as dyingcells.
The percentage of surviving neurons was determinedby dividing the average number of neurons per field inthe presence of MT-III (or vehicle treated) by the av-erage number of neurons per field upon initial plating.Inhibition of neurite formation was investigated in twoways: the percentage of neurite-bearing neurons andthe number of neurites per neuron. A neurite (whichincludes both axons and dendrites) was defined as anyprocess extending from the cell body with a length ofgreater than 20 �m. The percentage of neurite-bearingneurons (or more accurately the percentage of neuronswith one or more neurites) was calculated by dividingthe average number of neurite-bearing neurons perfield by the average total number of neurons per field.Of the neurite-bearing neurons, the number of neuritesper cell was calculated by dividing the average numberof neurites per field by the average number of neurite-bearing neurons per field. ANOVA, Student’s t test,and standard error analysis of results was made usingSigmaStat (Jandel).
Neurite length measurements of developing neuritesover 3 days were performed using NIH Image analysissoftware, with at least 300 neurites being measuredper treatment group. To determine the rate of neuriteelongation between time points, the average length ofneurites at the earlier timepoint was subtracted fromthe average length of neurites at the later time point.
Cytoskeletal changes following axonal transectionwere identified by double-labeled immunohistochemis-try, using both anti-tau and anti-�III-tubulin primaryantibodies. For analysis of tau/�III-tubulin immunola-
beled coverslips at 12 and 18 h posttransection, 5 dig-ital images of different injury sites of the coverslipwere captured (Olympus BX-60). Each group, unlessotherwise stated, consisted of at least 3 coverslips. Allneurite measurements were performed using NIH Im-age analysis software. T test and standard error anal-ysis of results was made using SigmaStat.
Anti-�III-tubulin immunoreactivity was found toaccumulate punctately at the tips of retracting,transected axons. This was used to determine the prox-imal end of sprouting tau-immunoreactive neurites,which were measured from the proximal end (definedby the punctately distributed �III-tubulin) to the distaltip. At least 150 neurites were measured per group.
RESULTS
MT-III Inhibits Neuronal Survival at HighConcentrations
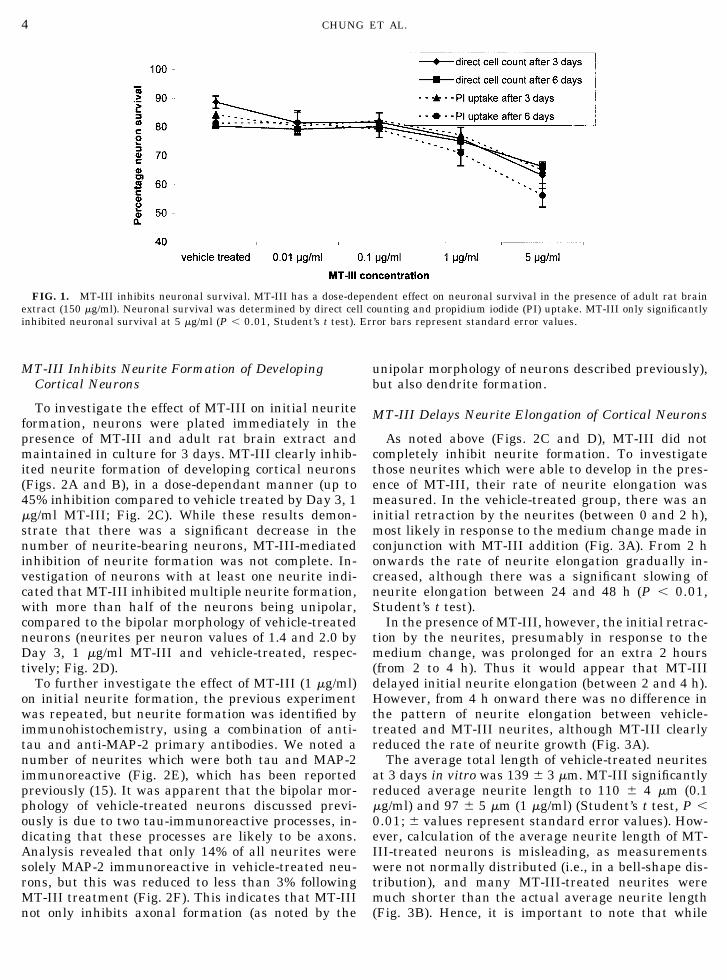
As reported previously (5), in the presence of brainextract, human MT-III has a dose-dependent inhibi-tory effect on neuron survival. However, to investigatethe neurite inhibitory properties of MT-III, it is impor-tant to use doses which are not overtly toxic. To deter-mine such doses, a range of MT concentrations wereapplied to 1 day-old neuron cultures (in the presence ofbrain extract), and the percentage of surviving neuronswas determined after a further 3 days in culture (Fig.1). At lower doses (0.01, 0.1, and 1 �g/ml), MT-III hadno significant effect on neuron survival, although thepercentage of surviving neurons at all of these concen-trations was lower than that of the vehicle-treatedgroup. At the highest dose (5 �g/ml), MT-III had asignificant inhibitory effect on neuron survival withneuron survival reduced to 63% compared to 88% neu-ron survival in vehicle-treated cultures (P � 0.01,Student’s t test). For subsequent studies investigatingthe neurite-inhibitory properties of MT-III, levelswhich had no significant neurotoxic effect (0.1 and 1.0�g/ml; less than 10% reduction in neuron survival com-pared to vehicle treated) were used. In the absence ofbrain extract, MT-III was not neurotoxic, and in factappeared to promote neuronal survival (results notshown). This is similar to findings by other researchers(5, 16), and suggests that MT-III-induced neuronaldeath is a specific action, rather than a nonspecifictoxic effect.
As a further indication of the effect of MT-III onneuronal survival, MT-III-treated cultures up to 6 daysin vitro were analyzed by propidium iodide uptake. At0.1 and 1 �g/ml concentrations of MT-III, neuronalsurvival was not significantly reduced compared to ve-hicle-treated neurons (Fig. 1). Neuronal degenerationwas also not observed by immunohistochemical stain-ing (results not shown).
3MT-III, AN INHIBITOR OF NEURITE GROWTH
MT-III Inhibits Neurite Formation of DevelopingCortical Neurons
To investigate the effect of MT-III on initial neuriteformation, neurons were plated immediately in thepresence of MT-III and adult rat brain extract andmaintained in culture for 3 days. MT-III clearly inhib-ited neurite formation of developing cortical neurons(Figs. 2A and B), in a dose-dependant manner (up to45% inhibition compared to vehicle treated by Day 3, 1�g/ml MT-III; Fig. 2C). While these results demon-strate that there was a significant decrease in thenumber of neurite-bearing neurons, MT-III-mediatedinhibition of neurite formation was not complete. In-vestigation of neurons with at least one neurite indi-cated that MT-III inhibited multiple neurite formation,with more than half of the neurons being unipolar,compared to the bipolar morphology of vehicle-treatedneurons (neurites per neuron values of 1.4 and 2.0 byDay 3, 1 �g/ml MT-III and vehicle-treated, respec-tively; Fig. 2D).
To further investigate the effect of MT-III (1 �g/ml)on initial neurite formation, the previous experimentwas repeated, but neurite formation was identified byimmunohistochemistry, using a combination of anti-tau and anti-MAP-2 primary antibodies. We noted anumber of neurites which were both tau and MAP-2immunoreactive (Fig. 2E), which has been reportedpreviously (15). It was apparent that the bipolar mor-phology of vehicle-treated neurons discussed previ-ously is due to two tau-immunoreactive processes, in-dicating that these processes are likely to be axons.Analysis revealed that only 14% of all neurites weresolely MAP-2 immunoreactive in vehicle-treated neu-rons, but this was reduced to less than 3% followingMT-III treatment (Fig. 2F). This indicates that MT-IIInot only inhibits axonal formation (as noted by the
unipolar morphology of neurons described previously),but also dendrite formation.
MT-III Delays Neurite Elongation of Cortical Neurons
As noted above (Figs. 2C and D), MT-III did notcompletely inhibit neurite formation. To investigatethose neurites which were able to develop in the pres-ence of MT-III, their rate of neurite elongation wasmeasured. In the vehicle-treated group, there was aninitial retraction by the neurites (between 0 and 2 h),most likely in response to the medium change made inconjunction with MT-III addition (Fig. 3A). From 2 honwards the rate of neurite elongation gradually in-creased, although there was a significant slowing ofneurite elongation between 24 and 48 h (P � 0.01,Student’s t test).
In the presence of MT-III, however, the initial retrac-tion by the neurites, presumably in response to themedium change, was prolonged for an extra 2 hours(from 2 to 4 h). Thus it would appear that MT-IIIdelayed initial neurite elongation (between 2 and 4 h).However, from 4 h onward there was no difference inthe pattern of neurite elongation between vehicle-treated and MT-III neurites, although MT-III clearlyreduced the rate of neurite growth (Fig. 3A).
The average total length of vehicle-treated neuritesat 3 days in vitro was 139 � 3 �m. MT-III significantlyreduced average neurite length to 110 � 4 �m (0.1�g/ml) and 97 � 5 �m (1 �g/ml) (Student’s t test, P �0.01; � values represent standard error values). How-ever, calculation of the average neurite length of MT-III-treated neurons is misleading, as measurementswere not normally distributed (i.e., in a bell-shape dis-tribution), and many MT-III-treated neurites weremuch shorter than the actual average neurite length(Fig. 3B). Hence, it is important to note that while
FIG. 1. MT-III inhibits neuronal survival. MT-III has a dose-dependent effect on neuronal survival in the presence of adult rat brainextract (150 �g/ml). Neuronal survival was determined by direct cell counting and propidium iodide (PI) uptake. MT-III only significantlyinhibited neuronal survival at 5 �g/ml (P � 0.01, Student’s t test). Error bars represent standard error values.
4 CHUNG ET AL.
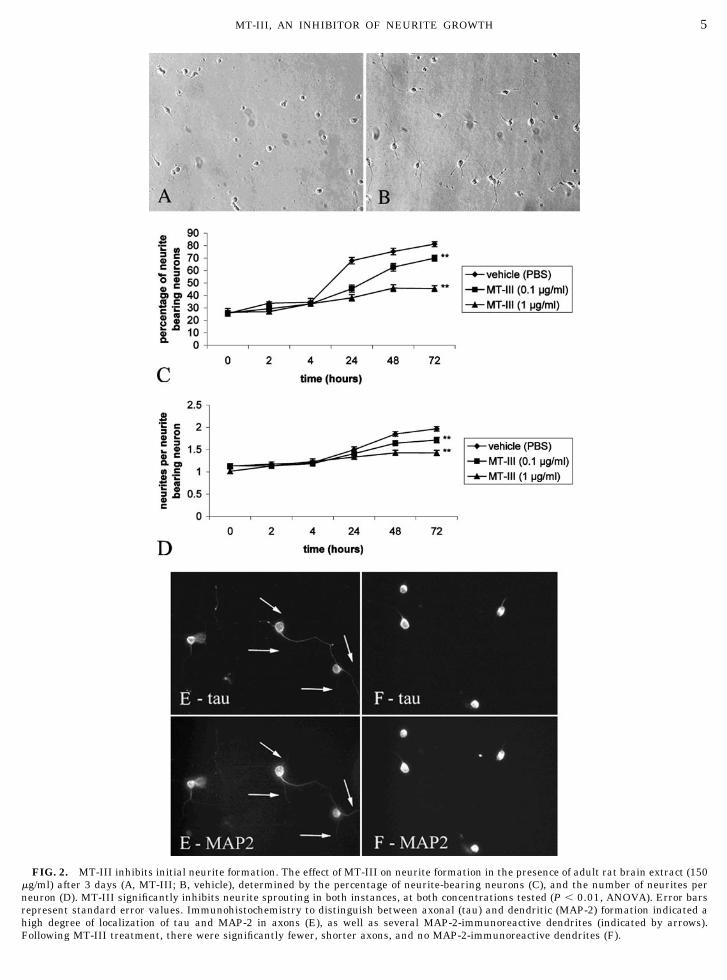
FIG. 2. MT-III inhibits initial neurite formation. The effect of MT-III on neurite formation in the presence of adult rat brain extract (150�g/ml) after 3 days (A, MT-III; B, vehicle), determined by the percentage of neurite-bearing neurons (C), and the number of neurites perneuron (D). MT-III significantly inhibits neurite sprouting in both instances, at both concentrations tested (P � 0.01, ANOVA). Error barsrepresent standard error values. Immunohistochemistry to distinguish between axonal (tau) and dendritic (MAP-2) formation indicated ahigh degree of localization of tau and MAP-2 in axons (E), as well as several MAP-2-immunoreactive dendrites (indicated by arrows).Following MT-III treatment, there were significantly fewer, shorter axons, and no MAP-2-immunoreactive dendrites (F).
5MT-III, AN INHIBITOR OF NEURITE GROWTH
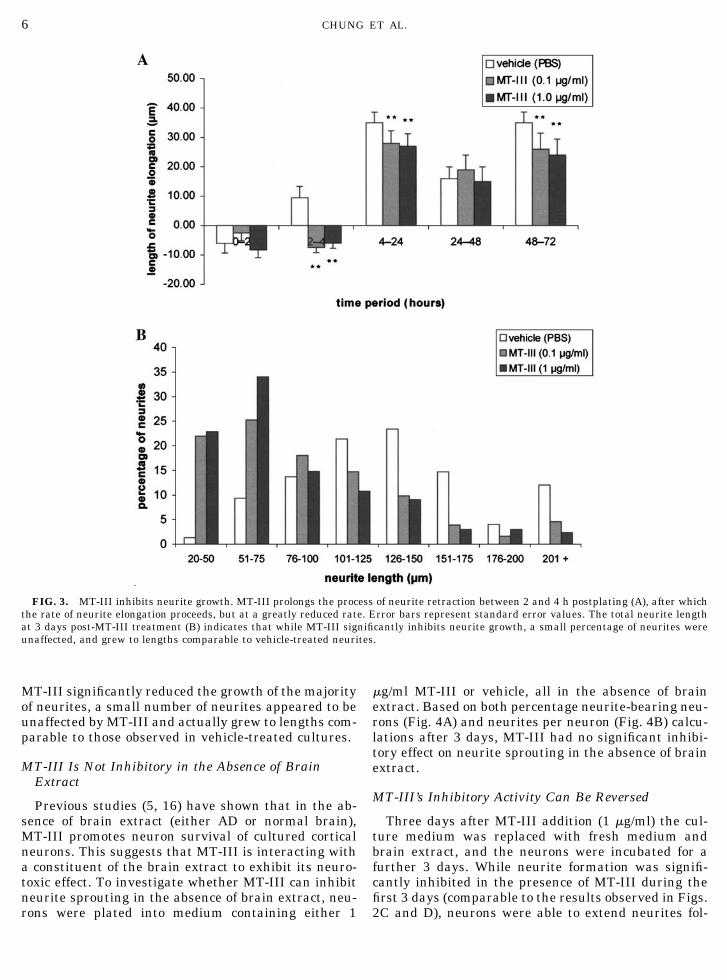
MT-III significantly reduced the growth of the majorityof neurites, a small number of neurites appeared to beunaffected by MT-III and actually grew to lengths com-parable to those observed in vehicle-treated cultures.
MT-III Is Not Inhibitory in the Absence of BrainExtract
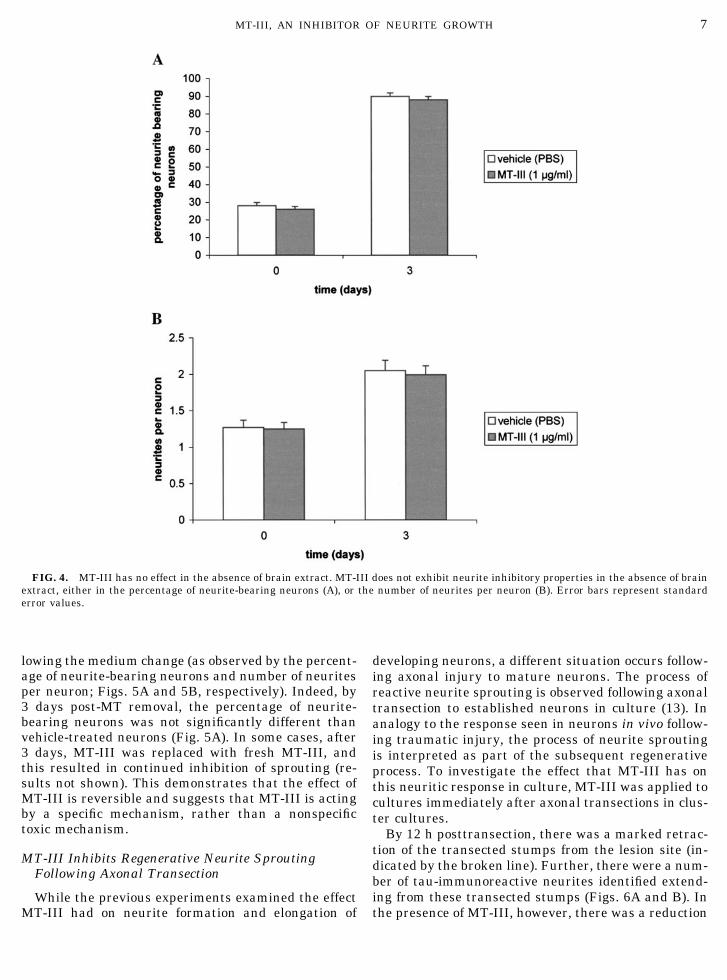
Previous studies (5, 16) have shown that in the ab-sence of brain extract (either AD or normal brain),MT-III promotes neuron survival of cultured corticalneurons. This suggests that MT-III is interacting witha constituent of the brain extract to exhibit its neuro-toxic effect. To investigate whether MT-III can inhibitneurite sprouting in the absence of brain extract, neu-rons were plated into medium containing either 1
�g/ml MT-III or vehicle, all in the absence of brainextract. Based on both percentage neurite-bearing neu-rons (Fig. 4A) and neurites per neuron (Fig. 4B) calcu-lations after 3 days, MT-III had no significant inhibi-tory effect on neurite sprouting in the absence of brainextract.
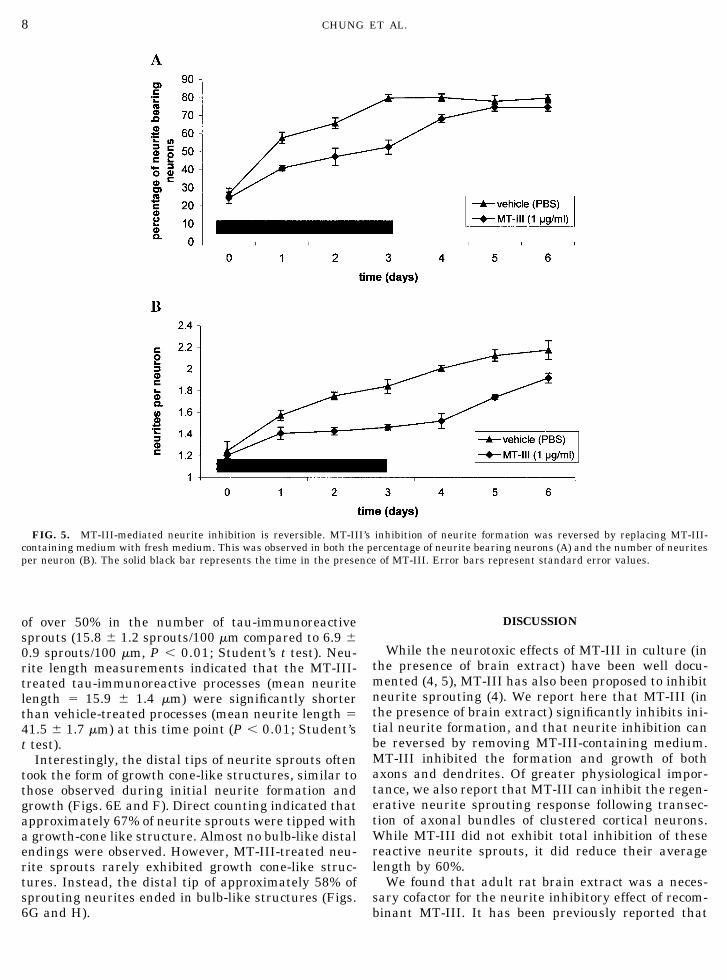
MT-III’s Inhibitory Activity Can Be Reversed
Three days after MT-III addition (1 �g/ml) the cul-ture medium was replaced with fresh medium andbrain extract, and the neurons were incubated for afurther 3 days. While neurite formation was signifi-cantly inhibited in the presence of MT-III during thefirst 3 days (comparable to the results observed in Figs.2C and D), neurons were able to extend neurites fol-
FIG. 3. MT-III inhibits neurite growth. MT-III prolongs the process of neurite retraction between 2 and 4 h postplating (A), after whichthe rate of neurite elongation proceeds, but at a greatly reduced rate. Error bars represent standard error values. The total neurite lengthat 3 days post-MT-III treatment (B) indicates that while MT-III significantly inhibits neurite growth, a small percentage of neurites wereunaffected, and grew to lengths comparable to vehicle-treated neurites.
6 CHUNG ET AL.
lowing the medium change (as observed by the percent-age of neurite-bearing neurons and number of neuritesper neuron; Figs. 5A and 5B, respectively). Indeed, by3 days post-MT removal, the percentage of neurite-bearing neurons was not significantly different thanvehicle-treated neurons (Fig. 5A). In some cases, after3 days, MT-III was replaced with fresh MT-III, andthis resulted in continued inhibition of sprouting (re-sults not shown). This demonstrates that the effect ofMT-III is reversible and suggests that MT-III is actingby a specific mechanism, rather than a nonspecifictoxic mechanism.
MT-III Inhibits Regenerative Neurite SproutingFollowing Axonal Transection
While the previous experiments examined the effectMT-III had on neurite formation and elongation of
developing neurons, a different situation occurs follow-ing axonal injury to mature neurons. The process ofreactive neurite sprouting is observed following axonaltransection to established neurons in culture (13). Inanalogy to the response seen in neurons in vivo follow-ing traumatic injury, the process of neurite sproutingis interpreted as part of the subsequent regenerativeprocess. To investigate the effect that MT-III has onthis neuritic response in culture, MT-III was applied tocultures immediately after axonal transections in clus-ter cultures.
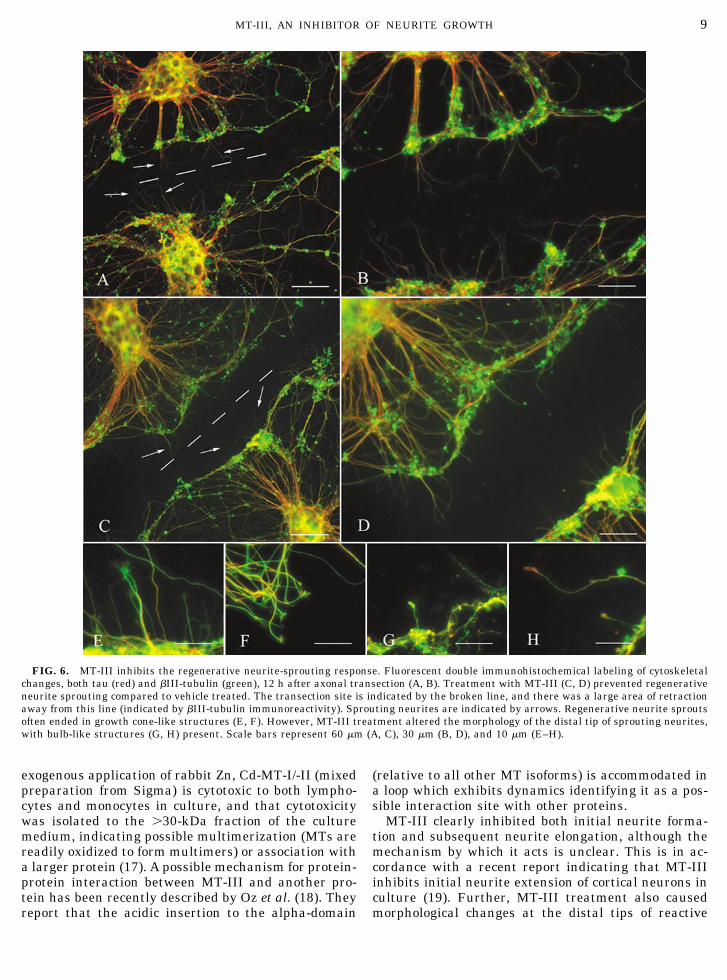
By 12 h posttransection, there was a marked retrac-tion of the transected stumps from the lesion site (in-dicated by the broken line). Further, there were a num-ber of tau-immunoreactive neurites identified extend-ing from these transected stumps (Figs. 6A and B). Inthe presence of MT-III, however, there was a reduction
FIG. 4. MT-III has no effect in the absence of brain extract. MT-III does not exhibit neurite inhibitory properties in the absence of brainextract, either in the percentage of neurite-bearing neurons (A), or the number of neurites per neuron (B). Error bars represent standarderror values.
7MT-III, AN INHIBITOR OF NEURITE GROWTH
of over 50% in the number of tau-immunoreactivesprouts (15.8 � 1.2 sprouts/100 �m compared to 6.9 �0.9 sprouts/100 �m, P � 0.01; Student’s t test). Neu-rite length measurements indicated that the MT-III-treated tau-immunoreactive processes (mean neuritelength � 15.9 � 1.4 �m) were significantly shorterthan vehicle-treated processes (mean neurite length �41.5 � 1.7 �m) at this time point (P � 0.01; Student’st test).
Interestingly, the distal tips of neurite sprouts oftentook the form of growth cone-like structures, similar tothose observed during initial neurite formation andgrowth (Figs. 6E and F). Direct counting indicated thatapproximately 67% of neurite sprouts were tipped witha growth-cone like structure. Almost no bulb-like distalendings were observed. However, MT-III-treated neu-rite sprouts rarely exhibited growth cone-like struc-tures. Instead, the distal tip of approximately 58% ofsprouting neurites ended in bulb-like structures (Figs.6G and H).
DISCUSSION
While the neurotoxic effects of MT-III in culture (inthe presence of brain extract) have been well docu-mented (4, 5), MT-III has also been proposed to inhibitneurite sprouting (4). We report here that MT-III (inthe presence of brain extract) significantly inhibits ini-tial neurite formation, and that neurite inhibition canbe reversed by removing MT-III-containing medium.MT-III inhibited the formation and growth of bothaxons and dendrites. Of greater physiological impor-tance, we also report that MT-III can inhibit the regen-erative neurite sprouting response following transec-tion of axonal bundles of clustered cortical neurons.While MT-III did not exhibit total inhibition of thesereactive neurite sprouts, it did reduce their averagelength by 60%.
We found that adult rat brain extract was a neces-sary cofactor for the neurite inhibitory effect of recom-binant MT-III. It has been previously reported that
FIG. 5. MT-III-mediated neurite inhibition is reversible. MT-III’s inhibition of neurite formation was reversed by replacing MT-III-containing medium with fresh medium. This was observed in both the percentage of neurite bearing neurons (A) and the number of neuritesper neuron (B). The solid black bar represents the time in the presence of MT-III. Error bars represent standard error values.
8 CHUNG ET AL.
exogenous application of rabbit Zn, Cd-MT-I/-II (mixedpreparation from Sigma) is cytotoxic to both lympho-cytes and monocytes in culture, and that cytotoxicitywas isolated to the �30-kDa fraction of the culturemedium, indicating possible multimerization (MTs arereadily oxidized to form multimers) or association witha larger protein (17). A possible mechanism for protein-protein interaction between MT-III and another pro-tein has been recently described by Oz et al. (18). Theyreport that the acidic insertion to the alpha-domain
(relative to all other MT isoforms) is accommodated ina loop which exhibits dynamics identifying it as a pos-sible interaction site with other proteins.
MT-III clearly inhibited both initial neurite forma-tion and subsequent neurite elongation, although themechanism by which it acts is unclear. This is in ac-cordance with a recent report indicating that MT-IIIinhibits initial neurite extension of cortical neurons inculture (19). Further, MT-III treatment also causedmorphological changes at the distal tips of reactive
FIG. 6. MT-III inhibits the regenerative neurite-sprouting response. Fluorescent double immunohistochemical labeling of cytoskeletalchanges, both tau (red) and �III-tubulin (green), 12 h after axonal transection (A, B). Treatment with MT-III (C, D) prevented regenerativeneurite sprouting compared to vehicle treated. The transection site is indicated by the broken line, and there was a large area of retractionaway from this line (indicated by �III-tubulin immunoreactivity). Sprouting neurites are indicated by arrows. Regenerative neurite sproutsoften ended in growth cone-like structures (E, F). However, MT-III treatment altered the morphology of the distal tip of sprouting neurites,with bulb-like structures (G, H) present. Scale bars represent 60 �m (A, C), 30 �m (B, D), and 10 �m (E–H).
9MT-III, AN INHIBITOR OF NEURITE GROWTH

sprouts. Such observations have been observed previ-ously using chemical agents such as taxol (unpublishedresults from our lab), which inhibits microtubule dy-namics. This suggests that MT-III may act to inhibitneurite sprouting and elongation by a similar mecha-nism. Investigation of the changes in cytoskeletal dy-namics caused by MT-III treatment will elucidate suchmechanisms.
There are numerous reports indicating the detec-tion of MT-III in the extracellular environment invivo (20 –22), and in culture (23). In this context, it ispossible that MT-III is released from astrocytes (theprimary location of MT-III in the brain) in the after-math of physical injury and modifies the subsequentneuronal response. Erickson et al. (5) have calcu-lated that human frontal cortex contains approxi-mately 20 �g/g MT-III in the brain and given thatnecrosis or physical damage could cause extensivelysis of cells, it is possible that local extracellularconcentrations of MT-III could reach the levels whichwe and others have shown to have significant effectson neuron survival and neurite sprouting (i.e., in thevicinity of 1–5 �g/ml). Further, MT-III is upregu-lated in reactive astrocytes surrounding areas of ce-rebral infarct, stab wound (24), or excitotoxic braindamage (25), and in culture, extracellular MT-III hasbeen shown to promote astrocyte proliferation andmigration (26). It is possible then, that the release ofMT-III by astrocytes is a contributing factor to themigration of new astrocytes into the lesion area, andsubsequent glial scar formation. Indeed, our reportthat MT-III inhibits the regenerative neurite sprout-ing response suggests that MT-III may be a compo-nent of the glial scar, which itself is very inhibitoryto regenerative neurite sprouting in the CNS. Fur-ther supporting this idea is the identification of me-tallothionein receptors on the cell surface of humanastrocytes (27), providing a possible mechanism forextracellular MT-III action on astrocytes.
The close resemblance between the neurofilamen-tous changes observed following physical damage toaxons and those observed in AD suggests that �-amy-loid plaques may cause prolonged damage to axons,resulting in reactive cytoskeletal alterations (28). Theability of MT-III to inhibit neurite sprouting andgrowth may be important in this regard, consideringits apparent deficiency in the Alzheimer’s disease brain(4). However, reports of MT-III levels in the AD brainare conflicting, with findings of both deficiency (4, 29,30) and no deficiency (5, 26). While this issue isclouded, there is further biochemical evidence for a linkbetween MT-III and AD. For instance, oxidative stresshas been implicated in the neuropathy of AD (31), andthus a deficiency in the potentially free radical scav-enging MT-III may be involved in the development ofAD. It has also recently been reported that MT-III caninhibit the formation of neurotoxic �-amyloid aggrega-
tions in vitro (32) and protect against free radical dam-age induced by the �-amyloid peptide 25–35 (33). Ourreport that MT-III inhibits tau-immunoreactive neu-rite sprouting (hypophosphorylated tau is the majorconstituent of NFTs) indicates that a deficiency in MT-III in the brain could indeed be a possible mechanisminvolved in NFT formation.
MT-III may also have a role to play in other neuro-logical disorders. For instance, the fact that MT-IIInull mice are highly susceptible to kainic acid seizures(34) suggests that MT-III is involved in the pathologyof epileptic seizures. Indeed, MT-III overexpressingmice exhibit much less kainic acid-induced brain dam-age than control mice (35). It is possible that the zinc-binding properties of MT-III would allow it to removeintracellularly, electrically stimulated synaptic zinc re-lease, preventing possible seizure occurence. However,Cole et al. (36) using ZnT3/MT-IIII double knockoutmice (which lack vesicular zinc) have found that MT-III is unlikely to protect against kainic acid-inducedseizures by modulating intracellular zinc release. In-terestingly, McKinney et al. (37) have reported abnor-mal neurite sprouting following axonal injury to CA3hippocampal neurons in brain slice culture (one of themost MT-III-rich regions in the brain). Further, theseinjured pyramidal neurons were hyperexcitable, andsynaptic responses often displayed unusual prolongedpolysynaptic components. Hence, it may be that MT-IIIprotects against seizures by preventing abnormal neu-rite sprouting following axonal injury.
While the mechanism by which MT-III is neuriteinhibitory is unclear, it is possible that this may be aphysiological function of MT-III. Indeed, MT-III maypossibly be involved in the development of AD, as wellas in the pathology of epileptic seizures. Excitingly,MT-III not only inhibits neurite formation of develop-ing neurons, but also regenerative neurite sprouting.Because inhibition of neurite formation caused by MT-III is reversible, it is possible that MT-III could be usedto delay regenerative neurite sprouting until a permis-sive environment is reached. Such therapeutic use forMTs has recently been explored by Hidalgo and co-workers, who have used intraperitoneal injections ofZn-MT-II (Sigma) to ameliorate the neurological dam-age which occurs during experimental autoimmune en-cephalomyelitis (EAE), an animal model of multiplesclerosis (38, 39).
ACKNOWLEDGMENTS
This work was supported by the Motor Accident and InsuranceBoard (MAIB) of Tasmania and the National Health and MedicalResearch Council (NHMRC). We also thank Jyoti Chuckowree,Graeme McCormack, and Marian Quilty for their technical help.
10 CHUNG ET AL.

REFERENCES
1. Masilah, E., M. Mallory, L. Hansen, M. Alford, T. Albright, R.DeTeresa, R. Terry, J. Baudier, and T. Saitoh. 1991. Patterns ofaberrant sprouting in Alzheimer’s disease. Neuron 6: 729–739.
2. Vickers, J. C., T. C. Dickson, P. A. Adlard, H. L. Saunders, C. E.King, and G. McCormack. 2000. The cause of neuronal degen-eration in Alzheimer’s disease. Prog. Neurobiol. 60: 139–165.
3. Uchida, Y., and M. Tomonaga. 1989. Neurotrophic action ofAlzheimer’s disease brain extract is due to the loss of inhibitoryfactors for survival and neurite formation of cerebral corticalneurons. Brain Res. 481: 190–193.
4. Uchida, Y., K. Takio, K. Titani, Y. Ihara, and M. Tomonaga.1991. The growth inhibitory factor that is deficient in the Alz-heimer’s disease brain is a 68 amino acid metallothionein-likeprotein. Neuron 7: 337–347.
5. Erickson, J. C., A. K. Sewell, L. T. Jensen, D. R. Winge, andR. D. Palmiter. 1994. Enhanced neurotrophic activity in Alzhei-mer’s disease cortex is not associated with down-regulation ofmetallothionein-III (GIF). Brain Res. 649: 297–304.
6. Chung, R. S., A. F. Holloway, B. L. Eckhardt, J. A. Harris, J. C.Vickers, M. I. Chuah, and A. K. West. 2002. Sheep have anunusual variant of the brain-specific metallothionein, MT-III.Biochem J. 365: 323–328.
7. Uchida, Y., Y. Ihara, and M. Tomonaga. 1988. Alzheimer’sdisease brain extract stimulates the survival of cerebral corticalneurons from neonatal rats. Biochem. Biophys. Res. Commun.150: 1263–1267.
8. Bradford, M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72: 248–254.
9. Brewer, G. J. 1995. Serum-free B27/neurobasal medium sup-ports differentiated growth of neurons from the striatum, sub-stantia nigra, septum, cerebral cortex, cerebellum, and dentategyrus. J. Neurosci. Res. 42: 674–683.
10. Brewer, G. J. 1997. Isolation and culture of adult rat hippocam-pal neurons. J. Neurosci. Methods 71: 143–155.
11. Goedart, M., C. M. Wischik, R. A. Crowther, J. E. Walker, andA. Klug. 1988. Cloning and sequencing of the cDNA encoding acore protein of the paired helical filament of Alzheimer’s dis-ease: Identification as the microtubule-associated protein tau.Proc. Natl. Acad. Sci. USA 85: 4051–4055.
12. Binder, A., A. Frankfurter, H. Kim, A. Caceres, M. R. Payne,and L. I. Febhun. 1984. Heterogeneity of microtubule-associ-ated protein 2 during rat brain development. Proc. Natl. Acad.Sci. USA 81: 5613–5617.
13. Dickson, T. C., P. A. Adlard, and J. C. Vickers. 2000. Sequenceof cellular changes following localized axotomy to cortical neu-rons in glia-free culture. J. Neurotrauma 17: 1095–1103.
14. Lee, M. K., J. B. Tuttle, L. I. Rebhun, D. W. Cleveland, and A.Frankfurter. 1990. The expression and posttranslational mod-ification of a neuron-specific beta-tubulin isotype during chickembryogenesis. Cell Motil. Cytol. 17: 118–132.
15. Kosik, K. S., and E. A. Finch. 1987. MAP2 and tau segregateinto dendritic and axonal domains after the elaboration of mor-phologically distinct neurites: An immunocytochemical study ofcultured rat cerebrum. J. Neurosci. 7: 3142–3153.
16. Masters, B. A., C. J. Quaife, J. C. Erickson, E. J. Kelly, G. J.Froelick, B. P. Zambrowicz, R. L. Brinster, and R. D. Palmiter.1994. Metallothionein III is expressed in neurons that seques-ter zinc in synaptic vesicles. J. Neurosci. 14: 5844–5857.
17. Borghesi, L. A., and M. A. Lynes. 1996. Nonprotective effects ofextracellular metallothionein. Toxicol. App. Pharmacol. 139:6–14.
18. Oz, G., K. Zangger, and I. M. Armitage. 2001. Three-dimensional structure and dynamics of a brain specificgrowth inhibitory factor: Metallothionein-3. Biochemistry 40:11433–11441.
19. Uchida, Y., F. Gomi, T. Masumizu, and Y. Miura. 2002. Growthinhibitory factor prevents neurite extension and the death ofcortical neurons caused by high oxygen exposure through hy-droxyl radical scavenging. J. Biol. Chem. ePub 10/6/2002.
20. Bremner, L., R. K. Mehra, and M. Sato. 1987. Metallothioneinin blood, bile and urine. Toxicology 108: 129–140.
21. Garvey, J. S. 1984. Metallothionein: Structure/antigenicity anddetection/quantitation in normal physiological fluids. Environ.Health Prosp. 54: 117–127.
22. Hidalgo, J., M. Giralt, J. S. Garvey, and A. Armario. 1988.Physiological role of glucocorticoids on rat serum and livermetallothionein basal and stress conditions. Am. J. Physiol.254: E71–78.
23. Trayhurn, P., J. S. Duncan, A. M. Wood, and J. H. Beattie.2000. Regulation of metallothionein gene expression and secre-tion in rat adipocytes differentiated from preadipocytes in pri-mary culture. Horm. Metab. Res. 32: 542–547.
24. Hozumi, I., T. Inuzuka, M. Hiraiwa, Y. Uchida, T. Anezaki, H.Ishiguro, H. Kobayashi, Y. Uda, T. Miyatake, and S. Tsuji.1995. Changes of growth inhibitory factor after stab wounds inrat brain. Brain Res. 688: 143–148.
25. Acarin, L., J. Carrasco, B. Gonzalez, J. Hidalgo, and B. Castel-lano. 1999. Expression of growth inhibitory factor (metallothio-nein-III) mRNA and protein following excitotoxic immaturebrain injury. J. Neuropathol. Exp. Neurol. 58: 389–397.
26. Carrasco, J., M. Giralt, A. Molinero, M. Penkowa, T. Moos,and J. Hidalgo. 1999. Metallothionein (MT)-III: Generationof polyclonal antibodies, comparison with MT-I�II in thefreeze lesioned rat brain and in a bioassay with astrocytes,and analysis of Alzheimer’s disease brains. J. Neurotrauma16: 1115–1129.
27. El Refaey, H., M. Ebadi, C. A. Kuszynski, J. Sweeney, F. M.Hamada, and A. Hamed. 1997. Identification of metallothioneinreceptors in human astrocytes. Neurosci. Lett. 231: 131–134.
28. Vickers, J. C. 1997. A cellular mechanism for the neuronalchanges underlying Alzheimer’s Disease. Neuroscience 78:629–639.
29. Tsuji, T., H. Kobayashi, Y. Uchida, Y. Ihara, and T. Miyatake.1992. Molecular cloning of human growth inhibitory factorcDNA and its down-regulation in Alzheimer’s disease. EMBO J.11: 4843–4850.
30. Yu, W. H., W. J. Lukiw, C. Bergeron, H. B. Niznik, and P. E.Fraser. 2001. Metallothionein III is reduced in Alzheimer’sdisease. Brain Res. 894: 37–45.
31. Hensley, K., J. M. Carney, M. P. Mattson, M. Aksenova, M.Harris, J. F. Wu, D. A. Floyd, and A. Butterfield. 1994. A modelfor beta-amyloid aggregation and neurotoxicity based on freeradical generation by the peptide: Relevance to Alzheimer’sdisease. Proc. Natl. Acad. Sci. USA 91: 3270–3274.
32. Irie, Y., and W. M. Keung. 2001. Metallothionein-III antago-nizes the neurotoxic and neurotrophic effects of amyloid betapeptides. Biochem. Biophys. Res. Commun. 282: 416–420.
33. Ren, H., Q. Ji, Y. Liu, and B. Ru. 2001. Different protective rolesin vitro of �- and �-domains of growth inhibitory factor (GIF) onneuron injuries caused by oxygen free radicals. Biochim. Bio-phys. Acta 1568: 129–134.
34. Erickson, J. C., G. Hollopeter, S. A. Thomas, G. J. Froelick, andR. D. Palmiter. 1997. Disruption of the metallothionein-III gene inmice: Analysis of brain zinc, behavior, and neuron vulnerability tometals, aging, and seizures. J. Neurosci. 17: 1271–1281.
11MT-III, AN INHIBITOR OF NEURITE GROWTH

35. Campagne, M. V., H. Thibodeaux, N. van Bruggen, B. Cairns,R. Gerlai, J. T. Palmer, S. P. Williams, and D. G. Lowe. 1999.Evidence for a protective role of metallothionein-1 in focal ce-rebral ischemia. Proc. Natl. Acad. USA 96: 12870–12875.
36. Cole, T. B., C. A. Robbins, H. J. Wenzel, P. A. Schwartzkroin,and R. D. Palmiter. 2000. Seizures and neuronal damage inmice lacking vesicular zinc. Epilepsy Res. 39: 153–169.
37. McKinney, R. A., D. Debanne, B. H. Gahwiler, and S. M.Thompson. 1997. Lesion-induced axonal sprouting and hyper-
excitability in the hippocampus in vitro: Implications for thegenesis of posttraumatic epilepsy. Nature Med. 3: 957–958.
38. Penkowa, M., and J. Hidalgo. 2000. Metallothionein I � IIexpression and their role in experimental autoimmune enceph-alomyelitis. GLIA 32: 247–263.
39. Penkowa, M., and J. Hidalgo. 2001. Metallothionein treatmentreduces proinflammatory cytokines IL-6 and TNF-a andapoptotic cell death during experimental autoimmune enceph-alomyelitis (EAE). Exp. Neurol. 170: 1–14.
12 CHUNG ET AL.
Copyright © 2022 FDOKUMEN




















