
Metabolic and Type 1 cannabinoid receptor imaging of a transgenic rat model in the early phase of...
10
Metabolic and Type 1 cannabinoid receptor imaging of a transgenic rat model in the early phase of Huntington disease C. Casteels a,b, ⁎, C. Vandeputte a,b , J.R. Rangarajan c , T. Dresselaers b,d , O. Riess e , G. Bormans b,f , F. Maes c , U. Himmelreich b,d , H. Nguyen e , K. Van Laere a,b a Division of Nuclear Medicine, KU Leuven and University Hospital Leuven, Belgium b IMIR, In vivo Molecular Imaging Research group, KU Leuven, Belgium c KU Leuven Medical Image Computing (ESAT/PSI) & Medical Imaging Research Center, University Hospital Leuven, Belgium d Biomedical NMR Unit, KU Leuven, Belgium e Department of Medical Genetics, University of Tübingen, Germany f Laboratory for Radiopharmacy, KU Leuven, Belgium abstract article info Article history: Received 17 January 2011 Revised 11 March 2011 Accepted 21 March 2011 Available online 31 March 2011 Keywords: Type 1 cannabinoid receptor Huntington disease [ 18 F]MK-9470 Small animal PET tgHD Pre-symptomatic Several lines of evidence imply early alterations in metabolic and endocannabinoid neurotransmission in Huntington disease (HD). Using [ 18 F]MK-9470 and small animal PET, we investigated for the first time cerebral changes in type 1 cannabinoid (CB1) receptor binding in vivo in pre-symptomatic and early symptomatic rats of HD (tgHD), in relation to glucose metabolism, morphology and behavioral testing for motor and cognitive function. Twenty-three Sprague–Dawley rats (14 tgHD and 9 wild-types) were investigated between the age of 2 and 11 months. Relative glucose metabolism and parametric CB1 receptor images were anatomically standardized to Paxinos space and analyzed voxel-wise. Volumetric microMRI imaging was performed to assess HD neuropathology. Within the first 10 months, bilateral volumes of caudate–putamen and lateral ventricles did not significantly differ between genotypes. Longitudinal- and genotype evolution showed that relative [ 18 F] MK-9470 binding progressively decreased in the caudate–putamen and lateral globus pallidus of tgHD rats (−8.3%, p ≤1.1×10 −5 at 5 months vs. −10.9%, p b 1.5 × 10 −5 at 10 months). In addition, relative glucose metabolism increased in the bilateral sensorimotor cortex of 2-month-old tgHD rats (+8.1%, p ≤1.5×10 −5 ), where it was positively correlated to motor function at that time point. TgHD rats developed cognitive deficits at 6 and 11 months of age. Our findings point to early regional dysfunctions in endocannabinoid signalling, involving the lateral globus pallidus and caudate–putamen. In vivo CB1 receptor measurements using [ 18 F]MK-9470 may thus be a useful early biomarker for HD. Our results also provide evidence of subtle motor and cognitive deficits at earlier stages than previously described. © 2011 Elsevier Inc. All rights reserved. Introduction Huntington disease (HD) is an autosomal, dominantly inherited neurodegenerative disease, characterized by progressive motor dysfunction, emotional changes and cognitive impairment. The disease is caused by an expanded CAG repeat in exon 1 of the gene encoding the protein huntingtin (htt) (The Huntington's Disease Collaborative Research Group, 1993). Adult onset is usually observed with trinucle- otide repeat blocks between 40 and 50 units, whereas more than 70 repeats results in a more severe and much less frequently observed juvenile form. The most striking pathophysiological feature of HD affected brains is the progressive atrophy of the caudate nucleus and the putamen, accompanied by a secondary enlargement of the lateral ventricles (Vonsattel et al., 1985). Despite progress in elucidating the molecular pathology of HD, therapeutic benefit for patients in terms of effective pharmacotherapy with either symptomatic or protective effects, has been scarce. The endocannabinoid system (ECS) (Katona and Freund, 2008) is likely involved in human HD. One of the earliest neurochemical alterations observed is the loss of type 1 cannabinoid (CB1) receptor binding from the basal ganglia nuclei (Richfield and Herkenham, 1994; Glass et al., 2000). This loss appears to occur prior to the loss of co- localised receptors such as the dopamine D 1 and D 2 receptors (Glass et al., 2000) and alterations in cerebral metabolism (Antonini et al., 1996). In the brain, CB1 receptors are found at high densities on GABA- ergic striatal terminals in the globus pallidus and substantia nigra that Experimental Neurology 229 (2011) 440–449 ⁎ Corresponding author at: Division of Nuclear Medicine, University Hospital Gasthuisberg, Herestraat 49 bus 7003, 3000 Leuven, Belgium. Fax: + 32 16 343759. E-mail address: [email protected] (C. Casteels). 0014-4886/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2011.03.014 Contents lists available at ScienceDirect Experimental Neurology journal homepage: www.elsevier.com/locate/yexnr
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Metabolic and Type 1 cannabinoid receptor imaging of a transgenic rat model in the early phase of...

Experimental Neurology 229 (2011) 440–449
Contents lists available at ScienceDirect
Experimental Neurology
j ourna l homepage: www.e lsev ie r.com/ locate /yexnr
Metabolic and Type 1 cannabinoid receptor imaging of a transgenic rat model in theearly phase of Huntington disease
C. Casteels a,b,⁎, C. Vandeputte a,b, J.R. Rangarajan c, T. Dresselaers b,d, O. Riess e, G. Bormans b,f, F. Maes c,U. Himmelreich b,d, H. Nguyen e, K. Van Laere a,b
a Division of Nuclear Medicine, KU Leuven and University Hospital Leuven, Belgiumb IMIR, In vivo Molecular Imaging Research group, KU Leuven, Belgiumc KU Leuven Medical Image Computing (ESAT/PSI) & Medical Imaging Research Center, University Hospital Leuven, Belgiumd Biomedical NMR Unit, KU Leuven, Belgiume Department of Medical Genetics, University of Tübingen, Germanyf Laboratory for Radiopharmacy, KU Leuven, Belgium
⁎ Corresponding author at: Division of Nuclear MGasthuisberg, Herestraat 49 bus 7003, 3000 Leuven, Bel
E-mail address: [email protected] (C.
0014-4886/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.expneurol.2011.03.014
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 January 2011Revised 11 March 2011Accepted 21 March 2011Available online 31 March 2011
Keywords:Type 1 cannabinoid receptorHuntington disease[18F]MK-9470Small animal PETtgHDPre-symptomatic
Several lines of evidence imply early alterations in metabolic and endocannabinoid neurotransmission inHuntington disease (HD). Using [18F]MK-9470 and small animal PET, we investigated for the first time cerebralchanges in type 1 cannabinoid (CB1) receptor binding in vivo in pre-symptomatic and early symptomatic rats ofHD (tgHD), in relation to glucose metabolism, morphology and behavioral testing for motor and cognitivefunction. Twenty-three Sprague–Dawley rats (14 tgHDand 9wild-types)were investigated between the age of 2and 11 months. Relative glucose metabolism and parametric CB1 receptor images were anatomicallystandardized to Paxinos space and analyzed voxel-wise. Volumetric microMRI imaging was performed to assessHD neuropathology.Within the first 10 months, bilateral volumes of caudate–putamen and lateral ventricles didnot significantly differ between genotypes. Longitudinal- and genotype evolution showed that relative [18F]MK-9470 binding progressively decreased in the caudate–putamen and lateral globus pallidus of tgHD rats(−8.3%, p≤1.1×10−5 at 5 months vs. −10.9%, pb1.5×10−5 at 10 months). In addition, relative glucosemetabolism increased in the bilateral sensorimotor cortex of 2-month-old tgHD rats (+8.1%, p≤1.5×10−5),where itwaspositively correlated tomotor function at that timepoint. TgHDrats developed cognitive deficits at 6and 11 months of age. Our findings point to early regional dysfunctions in endocannabinoid signalling, involvingthe lateral globus pallidus and caudate–putamen. In vivo CB1 receptor measurements using [18F]MK-9470 maythus be a useful early biomarker for HD. Our results also provide evidence of subtlemotor and cognitive deficits atearlier stages than previously described.
edicine, University Hospitalgium. Fax: +32 16 343759.Casteels).
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Huntington disease (HD) is an autosomal, dominantly inheritedneurodegenerative disease, characterized by progressive motordysfunction, emotional changes and cognitive impairment. The diseaseis caused by an expanded CAG repeat in exon 1 of the gene encoding theprotein huntingtin (htt) (The Huntington's Disease CollaborativeResearch Group, 1993). Adult onset is usually observed with trinucle-otide repeat blocks between 40 and 50 units, whereas more than 70repeats results in a more severe and much less frequently observedjuvenile form. The most striking pathophysiological feature of HD
affected brains is the progressive atrophy of the caudate nucleus and theputamen, accompanied by a secondary enlargement of the lateralventricles (Vonsattel et al., 1985). Despite progress in elucidating themolecular pathology of HD, therapeutic benefit for patients in terms ofeffective pharmacotherapy with either symptomatic or protectiveeffects, has been scarce.
The endocannabinoid system (ECS) (Katona and Freund, 2008) islikely involved in human HD. One of the earliest neurochemicalalterations observed is the loss of type 1 cannabinoid (CB1) receptorbinding from the basal ganglia nuclei (Richfield and Herkenham, 1994;Glass et al., 2000). This loss appears to occur prior to the loss of co-localised receptors such as the dopamine D1 and D2 receptors (Glasset al., 2000) and alterations in cerebral metabolism (Antonini et al.,1996). In the brain, CB1 receptors are found at high densities on GABA-ergic striatal terminals in the globus pallidus and substantia nigra that

441C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
are specifically affected by cellular dysfunction and death in HD(Richfield and Herkenham, 1994), and are involved in the control ofmotor behavior through modulation of neuronal firing and neurotrans-mitter release (Goutopoulos andMakriyannis, 2002). Hence, dysregula-tion of cannabinoid-mediated control of striatal function might play acritical role in the pathogenesis and symptom development of HD. Lossof CB1 receptor mRNA has also been observed in the striatum oftransgenic HD mice prior to the onset of motor related HD-likesymptoms and preceding neural degeneration (Denovan-Wright andRobertson, 2000; Lastres-Becker et al., 2002; Blazquez et al., 2011).Genetic ablation of CB1 receptors has been shown to accelerate theonset of HD-like symptoms in transgenic R6/2 mice (Blazquez et al.,2011). In symptomatic HD patients, we recently found a profoundcortical and subcortical loss of CB1 receptor availability in vivo (VanLaere et al., 2010).
So far, only in vitro and ex vivo data exist on the ECS in genetic HDmodels. Studies of CB1 receptor changes in transgenic models of HDhave focused on the basal ganglia, hippocampus and motor cortex, buthave not assessed other brain regions (Dowie et al., 2009;Dowie et al.,2010).Alsounknown iswhether CB1 receptor levels are changed in vivo.In vivo imaging of CB1 receptors in rat brain has become feasible due tothe development of CB1-selective PET radioligands, such as [18F]MK-9470 (Burns et al., 2007).
Our primary objective was to explore CB1 receptor changes in vivousing small animal PET at the pre-symptomatic and early symptomaticstage (defined up to 10 months using literature data (Nguyen et al.,2006)) of a transgenic rat model for HD. As a second objective, brainglucose metabolism and behavioral variables for cognitive and motorfunction, all shown to be altered early in patients and animal models ofHD (Antonini et al., 1996;Wang et al., 2005), were investigated in thesame animals and correlated to the regional CB1 receptor availability.
Materials and methods
Animals
In this study, we have used a transgenic HD (tgHD) rat model,previously characterized by an adult-onset neurological HD-likephenotype, with slowly progressive motor function, cognitiveimpairments and neuropathological abnormalities such as neuronalintranuclear inclusions and enlarged lateral ventricles (Nguyen et al.,2006). In total, 14 homozygous male Sprague–Dawley tgHD (+/+)rats and 9 wild-type littermates (WT) were included. Breeding wascarried out by mating hemizygous transgenic males with hemizygoustransgenic females so that offspring of the homozygous and WTgenotype were obtained within one litter.
The tgHD rats express 727 amino acids of the HD genewith 51 CAGrepeats (cDNA position 324–2321, corresponding to 22% full length),
Fig. 1. Experiment time-line. Experiment time-line for functional and structural imaging of aof the different modalities used per month is shown in B. m, month; d, day; h, hour.
under control of the endogenous rat htt promoter (von Horsten et al.,2003). Genotyping was carried out at the age of 3 weeks by QPCR. Allanimals were housed as pairs within a cage, at an average roomtemperature of 22 °C and a 12-h light/dark cycle. Food andwater weregiven ad libitum. The research protocol was approved by the localAnimal Ethics Committees and was according to European EthicsCommittee guidelines (decree 86/609/EEC).
Radiotracer preparation and imaging
Functional imaging of brain CB1 receptor binding and glucosemetabolism was performed in tgHD and WT animals at the age of 2, 5and 10months (for time-line overview, see Fig. 1). Brain CB1 receptorimagingwas done using the radioligand [18F]MK-9470 (N-[2-(3-cyano-phenyl)-3-(4-(2-[18F]fluorethoxy)-phenyl)-1-methylpropyl]-2-(5-methyl-2-pyridyloxy)-2-methylpropona-mide), which is characterizedbyhigh specificity andhighaffinity for theCB1 receptor (rat IC50 0.9nM).The precursor for [18F]MK-9470 was obtained from Merck ResearchLaboratories (MRL, West Point, USA). Radiolabeling was performed on-site by alkylation of the precursor with 2-[18F]fluoroethylbromide.Metabolic imaging was performed using [18F]-FDG. [18F]-FDG wasprepared using an IBA 18/9 cyclotron and a routine [18F]-FDG synthesismodule. Tracer preparation and characteristics have been describedpreviously (Casteels et al., 2006;Burns et al., 2007).
Prior to small animal PET imaging, rats were anesthetized using2.5% isoflurane in 2.0 L/min oxygen. Tail veins were catheterized forinjection of the radioligands ([18F]MK-9470 21.6±1.8 MBq; [18F]-FDG20.3±1.6 MBq; specific activity range 100–760 GBq/μmol). Theradioligands were diluted with saline to obtain a 5% ethanol solutionand injected in a total volume of approximately 500 μL. After overnightfasting, [18F]MK-9470 measurements were obtained during a 20 mininterval, starting 1 h post-injection, while [18F]-FDG acquisitions wereperformed between 60 and 100 min post-injection. [18F]-FDGdatawereacquired 4 days after [18F]MK-9470 measurements.
Small animal PET imaging was performed using a FOCUS 220tomograph (Siemens/Concorde Microsystems, Knoxville, TN), whichhas a transaxial resolution of 1.35 mm full-width at half-maximum.Data were acquired in a 128×128×95 matrix with a pixel width of0.475 mm and a slice thickness of 0.796 mm.
To assess neuropathological features of HD, manifested as loss of thecaudate–putamen volume and as enlargement of the lateral ventricles,anatomical magnetic resonance imaging (MRI) was performed 1 dayafter PET on a 9.4 Tesla Bruker Biospec Scanner (Bruker Biospin,Ettlingen, Germany) using a dedicated rat brain surface coil (RapidBiomedical, Rimpar, Germany) (Fig. 1). Three-dimensional coronal highresolution images of the entire rat brainwere obtained in 20 minusing aconventional 3D turboRARE sequence with following parameters:TR=1200 ms, TEeff=46 ms, rare-factor=12, matrix 256×156×77,
transgenic rat model in the early phase of Huntington disease (A). A detailed description

442 C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
isotropic voxel size of 195 μm. Rats were anesthetized with 2–3%isoflurane, the head fixed in a stereotaxic frame and body temperaturemaintained at 37±1 °C.
Behavioral testing
All rats were testedwithin the light phase of a 12 h light/dark cyclefor cognitive capability using the radial arm maze and for motorfunction using the Catwalk as described below. Cognitive capabilitywas assessed at the age of 3, 6 and 11 months (Fig. 1). Motor functionwas tested within 1 week from the small animal PET experiments andrepeated monthly until the age of 10 months (Fig. 1). Body weight asconfounding variable was measured at all test days.
Radial arm mazeSpatial learning and memory were assessed in an eight-arm radial
maze for rats. The radial maze consisted of an octagonal central areafrom which eight arms (55×15×22.5 cm; LxWxH) radiated out-wards. The maze was placed 70 cm above the floor in a room withfixed extra-maze visual cues. At each time point, rats were habituatedto the radial arm maze in a 10-min daily session on 3 successive days.The baits (sweetened cereal) were available towards the end of eacharm to encourage exploration. Following this training period, animals’weight was gradually reduced over 4 days and maintained atapproximately 85% of their free-feeding rate during the testingperiod. The testing period consisted of 2 days, on which explorativebehavior and reinforced alternation were evaluated. The test protocolbelow is based on Nguyen et al. (Nguyen et al., 2006).
Experiment 1 (day 1): exploratory behavior. Each animal wasindividually placed in the centre of the eight-armmaze, and was allowedto freely investigate the maze for 10 min. The frequency distribution ofangles between consecutively entered arms was evaluated. An arm entrywas scoredwhen all four paws of the ratwerewithin an armusing digitalvideo recording.One trial per animalwas given. Thearmswerenot baited.
Experiment 2 (day 2): reinforced alternation. The animal started in arandomly chosen arm. The remaining seven arms were baited with afood pellet. The trial was terminated if all seven pellets had beencollected or 10 min had elapsed. The number ofworkingmemory errors(i.e. re-entering an arm that was previously visited) were scored usingdigital video recording. One trial per animal was given.
CatwalkThe feasibility of the Catwalk to asses gait disturbances in animal
models of neurodegenerative diseases has been demonstrated before(Vandeputte et al., 2010). The Catwalk™ (Noldus, Wageningen, TheNetherlands) consists of a horizontal glass plate and video-capturingequipment placed below. It allows semi-automated quantification of alargenumber of locomotor parameters duringwalkway crossing, such asstride length, base of support, interlimb coordination, and swing/stancephases. The principle of thismethod is based on optical reflection of pawcontact points using a fluorescent tube. For correct locomotor analysis,three uninterrupted runswere usedwith aminimumof 4 step sequencepatterns. For experiments reported here between16 and28 strideswerecollected for every animal (mean=18.4±3.0). Locomotor speed, i.e. thecovered distance over the runway (cm) divided by the time needed tocross it, was additionally determined in each run as it may act asconfounder for results’ interpretation (Koopmans et al., 2007). Also,stride frequency was post hoc calculated as the number of strides persecond.
Image processing and data analysis
For quantification purposes, PET scans were reconstructed usingfiltered back projection. Parametric images based onmodified standarduptake values (mSUV) ( = activity concentration (MBq/ml)×((bodymass (g)+mean body mass control)/2)/injected dose (MBq) ) were
generated as measure of absolute [18F]MK-9470 binding. These werepreviously validated to give good estimates of total receptor volume ofdistribution as determined by full kinetic modelling (Sanabria-Bohor-quez et al., 2010).No significantdifferences inweight or injected activitywerepresent between tgHDandWT rats.We investigatedboth absoluteand relative [18F]MK-9470 binding. Relative [18F]MK-9470 binding wasexpressed asmSUVnormalized onwhole-brainmSUV. Relative regionalglucose metabolism was determined by normalizing [18F]-FDG data tothewhole-brain uptake. Thewithin subject test–retest variability of ourimaging procedure is consistent inmagnitudewith humandata, i.e. b 5%(Casteels et al., 2006;Goffin et al., 2008).
To obtain maximal use of information without a priori knowledge,PET data were analyzed voxel-wise using SPM2 (Statistical ParametricMapping, Welcome Department of Cognitive Neurology, London, UK).For spatial normalisation, individual PET-MRI data were non-linearlynormalized to custom-made rat brain templates in Paxinos stereo-tactic space (Casteels et al., 2006). This methodology allows reportingresults in coordinates directly corresponding to the Paxinos coordi-nate system for the rat brain. For SPM analysis, data were smoothedwith an isotropic Gaussian kernel of 1.2 mm and analyzed both in amultifactor design using genotype (tgHD vs. WT)×age (2 months vs.5 months vs. 10 months) (i.e. looking at differences over time be-tween genotypes), as well as in a single factor design using genotypeor age only (i.e. looking at cross-sectional and longitudinal differ-ences, respectively). For statistical analysis, T-maps were interrogatedat a peak voxel level of p=0.005 (uncorrected) and extent thresholdkEN200 voxels. Only clusters with pb0.05, corrected for multiplecomparisons were retained, as described previously (Casteels et al.,2010b). Exceptions on pcluster weremade for clusters which were bothneurobiologically plausible and relevant in the light of other findingsin this study. For analysis of relative receptor binding and [18F]-FDG,proportional scaling to themean voxel value was used and an analysisthreshold of 0.8 of the mean image intensity was applied.
In addition, a voxel-based correlation analysis between relative[18F]MK-9470 uptake / [18F]-FDG data and the behavioral outcomesfor cognitive and motor function was performed within the tgHDgroup. Literature data have previously shown that [18F]-FDG uptake ismainly coupled to glutamate- and GABA-mediated synaptic activity(Magistretti, 2006), which is also modulated by the ECS.
To assess the neuropathological features of HD in vivo, caudate–putamen volume and lateral ventricle size were quantified (Vonsattelet al., 1985). Bilateral caudate nucleus and putamen volumes weredetermined bymanual delineation on the transverse slices of MRI data.
A measure for the volume of the lateral ventricles was obtainedfrom the 3D turboRARE MRI images using an automated method, inorder to avoid the impact of possible manual segmentation inaccu-racies induced by MRI image ambiguity (e.g. due to partial volumeeffects that are likely to occur for the thin, elongated shape of thelateral ventricle). A volume of interest (VOI) around the left and rightlateral ventricles was first defined in the image through affine co-registration with an atlas template (Maes et al., 1997). Hyper-intensevoxels within this VOI, corresponding to ventricular cerebrospinal-fluid (CSF), were segmented by thresholding the intensity values inthis VOI above mean+1.5*SD and the volume of these CSF voxels wasreported as indicative for the lateral ventricular volume.
General statistics
Reported values inmanuscript and graphs are given as themean±SD.Conventional statistics were carried out using Statistica v8.0 (Statsoft,Tulsa, OK, USA). Behavioral outcomes and body weight were analyzedusing non-parametric Mann–Whitney U tests as time-variations in size,weight and skeletalmorphometryhavebeen shownto influence standardgait measurements (Wooley et al., 2009). Caudate–putamen volume andlateral ventricle size were subjected to repeated measures ANOVA withone between-subject factor, i.e. genotype andwith repeatedmeasures on
443C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
the factor age. Tukey HSD test was used for post hoc comparison.Significance was accepted at the 95% probability level.
Results
Small animal [18F]MK-9470 and [18F]-FDG PET imaging
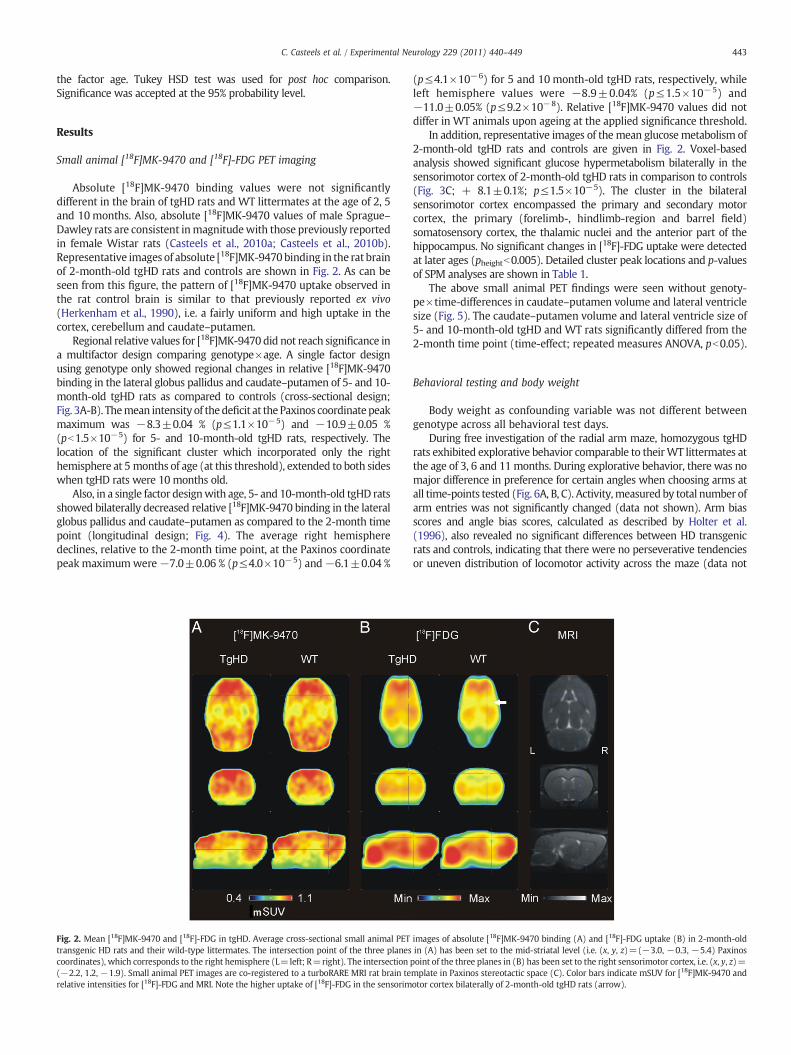
Absolute [18F]MK-9470 binding values were not significantlydifferent in the brain of tgHD rats and WT littermates at the age of 2, 5and 10months. Also, absolute [18F]MK-9470 values of male Sprague–Dawley rats are consistent inmagnitudewith those previously reportedin female Wistar rats (Casteels et al., 2010a; Casteels et al., 2010b).Representative images of absolute [18F]MK-9470binding in the rat brainof 2-month-old tgHD rats and controls are shown in Fig. 2. As can beseen from this figure, the pattern of [18F]MK-9470 uptake observed inthe rat control brain is similar to that previously reported ex vivo(Herkenham et al., 1990), i.e. a fairly uniform and high uptake in thecortex, cerebellum and caudate–putamen.
Regional relative values for [18F]MK-9470 did not reach significance ina multifactor design comparing genotype×age. A single factor designusing genotype only showed regional changes in relative [18F]MK-9470binding in the lateral globus pallidus and caudate–putamen of 5- and 10-month-old tgHD rats as compared to controls (cross-sectional design;Fig. 3A-B). Themean intensity of thedeficit at the Paxinos coordinate peakmaximum was −8.3±0.04 % (p≤1.1×10−5) and −10.9±0.05 %(pb1.5×10−5) for 5- and 10-month-old tgHD rats, respectively. Thelocation of the significant cluster which incorporated only the righthemisphere at 5 months of age (at this threshold), extended to both sideswhen tgHD rats were 10 months old.
Also, in a single factor designwith age, 5- and 10-month-old tgHD ratsshowed bilaterally decreased relative [18F]MK-9470 binding in the lateralglobus pallidus and caudate–putamen as compared to the 2-month timepoint (longitudinal design; Fig. 4). The average right hemispheredeclines, relative to the 2-month time point, at the Paxinos coordinatepeak maximumwere−7.0±0.06 % (p≤4.0×10−5) and−6.1±0.04 %
Fig. 2. Mean [18F]MK-9470 and [18F]-FDG in tgHD. Average cross-sectional small animal PETtransgenic HD rats and their wild-type littermates. The intersection point of the three planescoordinates), which corresponds to the right hemisphere (L=left; R=right). The intersection(−2.2, 1.2, −1.9). Small animal PET images are co-registered to a turboRARE MRI rat brain terelative intensities for [18F]-FDG and MRI. Note the higher uptake of [18F]-FDG in the sensorim
(p≤4.1×10−6) for 5 and 10 month-old tgHD rats, respectively, whileleft hemisphere values were −8.9±0.04% (p≤1.5×10−5) and−11.0±0.05% (p≤9.2×10−8). Relative [18F]MK-9470 values did notdiffer in WT animals upon ageing at the applied significance threshold.
In addition, representative images of themean glucose metabolism of2-month-old tgHD rats and controls are given in Fig. 2. Voxel-basedanalysis showed significant glucose hypermetabolism bilaterally in thesensorimotor cortex of 2-month-old tgHD rats in comparison to controls(Fig. 3C; + 8.1±0.1%; p≤1.5×10−5). The cluster in the bilateralsensorimotor cortex encompassed the primary and secondary motorcortex, the primary (forelimb-, hindlimb-region and barrel field)somatosensory cortex, the thalamic nuclei and the anterior part of thehippocampus. No significant changes in [18F]-FDG uptake were detectedat later ages (pheightb0.005). Detailed cluster peak locations and p-valuesof SPM analyses are shown in Table 1.
The above small animal PET findings were seen without genoty-pe×time-differences in caudate–putamen volume and lateral ventriclesize (Fig. 5). The caudate–putamen volume and lateral ventricle size of5- and 10-month-old tgHD and WT rats significantly differed from the2-month time point (time-effect; repeated measures ANOVA, pb0.05).
Behavioral testing and body weight
Body weight as confounding variable was not different betweengenotype across all behavioral test days.
During free investigation of the radial arm maze, homozygous tgHDrats exhibited explorative behavior comparable to theirWT littermates atthe age of 3, 6 and 11 months. During explorative behavior, there was nomajor difference in preference for certain angles when choosing arms atall time-points tested (Fig. 6A, B, C). Activity,measured by total number ofarm entries was not significantly changed (data not shown). Arm biasscores and angle bias scores, calculated as described by Holter et al.(1996), also revealed no significant differences between HD transgenicrats and controls, indicating that there were no perseverative tendenciesor uneven distribution of locomotor activity across the maze (data not
images of absolute [18F]MK-9470 binding (A) and [18F]-FDG uptake (B) in 2-month-oldin (A) has been set to the mid-striatal level (i.e. (x, y, z)=(−3.0, −0.3, −5.4) Paxinospoint of the three planes in (B) has been set to the right sensorimotor cortex, i.e. (x, y, z)=mplate in Paxinos stereotactic space (C). Color bars indicate mSUV for [18F]MK-9470 andotor cortex bilaterally of 2-month-old tgHD rats (arrow).
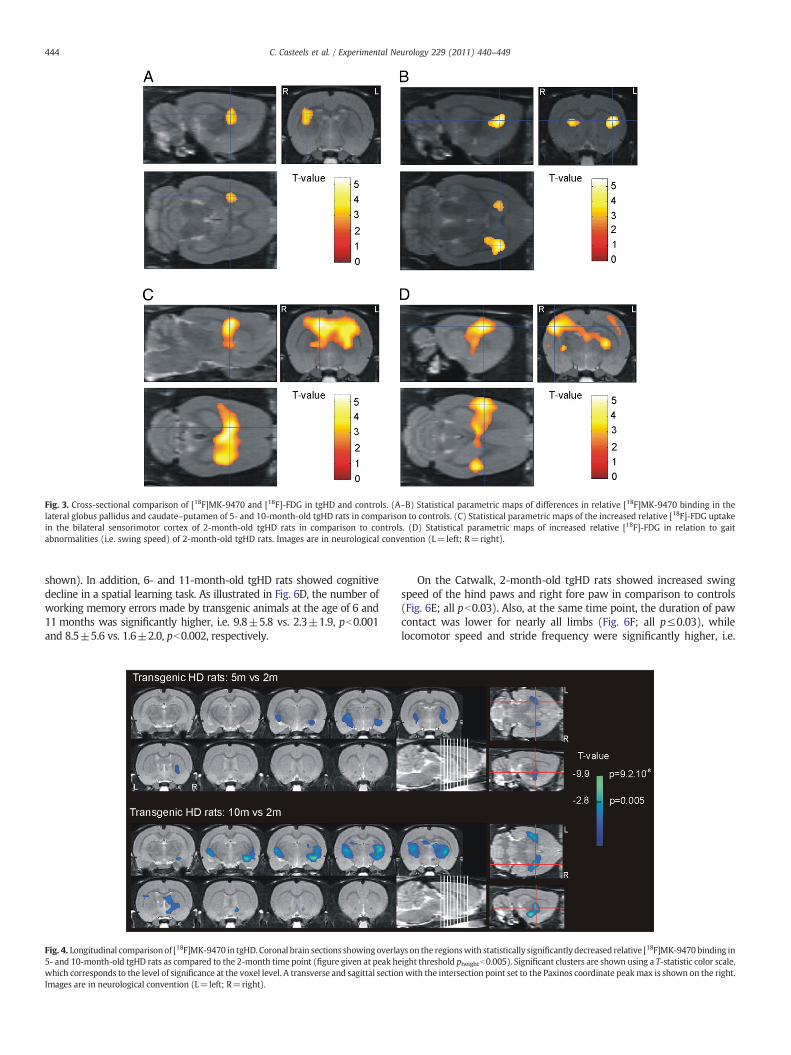
Fig. 3. Cross-sectional comparison of [18F]MK-9470 and [18F]-FDG in tgHD and controls. (A–B) Statistical parametric maps of differences in relative [18F]MK-9470 binding in thelateral globus pallidus and caudate–putamen of 5- and 10-month-old tgHD rats in comparison to controls. (C) Statistical parametric maps of the increased relative [18F]-FDG uptakein the bilateral sensorimotor cortex of 2-month-old tgHD rats in comparison to controls. (D) Statistical parametric maps of increased relative [18F]-FDG in relation to gaitabnormalities (i.e. swing speed) of 2-month-old tgHD rats. Images are in neurological convention (L=left; R=right).
444 C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
shown). In addition, 6- and 11-month-old tgHD rats showed cognitivedecline in a spatial learning task. As illustrated in Fig. 6D, the number ofworking memory errors made by transgenic animals at the age of 6 and11 months was significantly higher, i.e. 9.8±5.8 vs. 2.3±1.9, pb0.001and 8.5±5.6 vs. 1.6±2.0, pb0.002, respectively.
Fig. 4. Longitudinal comparisonof [18F]MK-9470 in tgHD. Coronal brain sections showingoverla5- and 10-month-old tgHD rats as compared to the 2-month time point (figure given at peak hewhich corresponds to the level of significance at the voxel level. A transverse and sagittal sectioImages are in neurological convention (L=left; R=right).
On the Catwalk, 2-month-old tgHD rats showed increased swingspeed of the hind paws and right fore paw in comparison to controls(Fig. 6E; all pb0.03). Also, at the same time point, the duration of pawcontact was lower for nearly all limbs (Fig. 6F; all p≤0.03), whilelocomotor speed and stride frequency were significantly higher, i.e.
ys on the regionswith statistically significantlydecreased relative [18F]MK-9470 binding inight threshold pheightb0.005). Significant clusters are shown using a T-statistic color scale,n with the intersection point set to the Paxinos coordinate peakmax is shown on the right.
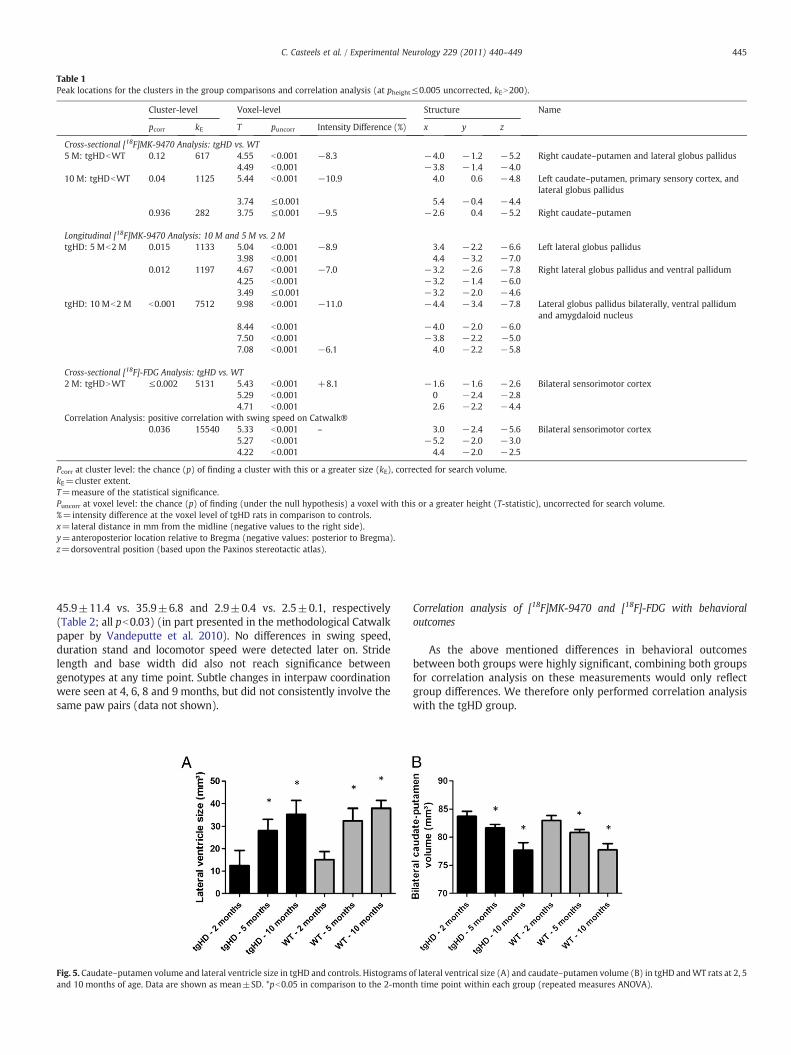
Table 1Peak locations for the clusters in the group comparisons and correlation analysis (at pheight≤0.005 uncorrected, kEN200).
Cluster-level Voxel-level Structure Name
pcorr kE T puncorr Intensity Difference (%) x y z
Cross-sectional [18F]MK-9470 Analysis: tgHD vs. WT5 M: tgHDbWT 0.12 617 4.55 b0.001 −8.3 −4.0 −1.2 −5.2 Right caudate–putamen and lateral globus pallidus
4.49 b0.001 −3.8 −1.4 −4.010 M: tgHDbWT 0.04 1125 5.44 b0.001 −10.9 4.0 0.6 −4.8 Left caudate–putamen, primary sensory cortex, and
lateral globus pallidus3.74 ≤0.001 5.4 −0.4 −4.4
0.936 282 3.75 ≤0.001 −9.5 −2.6 0.4 −5.2 Right caudate–putamen
Longitudinal [18F]MK-9470 Analysis: 10 M and 5 M vs. 2 MtgHD: 5 Mb2 M 0.015 1133 5.04 b0.001 −8.9 3.4 −2.2 −6.6 Left lateral globus pallidus
3.98 b0.001 4.4 −3.2 −7.00.012 1197 4.67 b0.001 −7.0 −3.2 −2.6 −7.8 Right lateral globus pallidus and ventral pallidum
4.25 b0.001 −3.2 −1.4 −6.03.49 ≤0.001 −3.2 −2.0 −4.6
tgHD: 10 Mb2 M b0.001 7512 9.98 b0.001 −11.0 −4.4 −3.4 −7.8 Lateral globus pallidus bilaterally, ventral pallidumand amygdaloid nucleus
8.44 b0.001 −4.0 −2.0 −6.07.50 b0.001 −3.8 −2.2 −5.07.08 b0.001 −6.1 4.0 −2.2 −5.8
Cross-sectional [18F]-FDG Analysis: tgHD vs. WT2 M: tgHDNWT ≤0.002 5131 5.43 b0.001 +8.1 −1.6 −1.6 −2.6 Bilateral sensorimotor cortex
5.29 b0.001 0 −2.4 −2.84.71 b0.001 2.6 −2.2 −4.4
Correlation Analysis: positive correlation with swing speed on Catwalk®0.036 15540 5.33 b0.001 – 3.0 −2.4 −5.6 Bilateral sensorimotor cortex
5.27 b0.001 −5.2 −2.0 −3.04.22 b0.001 4.4 −2.0 −2.5
Pcorr at cluster level: the chance (p) of finding a cluster with this or a greater size (kE), corrected for search volume.kE=cluster extent.T=measure of the statistical significance.Puncorr at voxel level: the chance (p) of finding (under the null hypothesis) a voxel with this or a greater height (T-statistic), uncorrected for search volume.%=intensity difference at the voxel level of tgHD rats in comparison to controls.x=lateral distance in mm from the midline (negative values to the right side).y=anteroposterior location relative to Bregma (negative values: posterior to Bregma).z=dorsoventral position (based upon the Paxinos stereotactic atlas).
445C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
45.9±11.4 vs. 35.9±6.8 and 2.9±0.4 vs. 2.5±0.1, respectively(Table 2; all pb0.03) (in part presented in the methodological Catwalkpaper by Vandeputte et al. 2010). No differences in swing speed,duration stand and locomotor speed were detected later on. Stridelength and base width did also not reach significance betweengenotypes at any time point. Subtle changes in interpaw coordinationwere seen at 4, 6, 8 and 9 months, but did not consistently involve thesame paw pairs (data not shown).
Fig. 5. Caudate–putamen volume and lateral ventricle size in tgHD and controls. Histograms oand 10 months of age. Data are shown as mean±SD. *pb0.05 in comparison to the 2-mon
Correlation analysis of [18F]MK-9470 and [18F]-FDG with behavioraloutcomes
As the above mentioned differences in behavioral outcomesbetween both groups were highly significant, combining both groupsfor correlation analysis on these measurements would only reflectgroup differences. We therefore only performed correlation analysiswith the tgHD group.
f lateral ventrical size (A) and caudate–putamen volume (B) in tgHD andWT rats at 2, 5th time point within each group (repeated measures ANOVA).
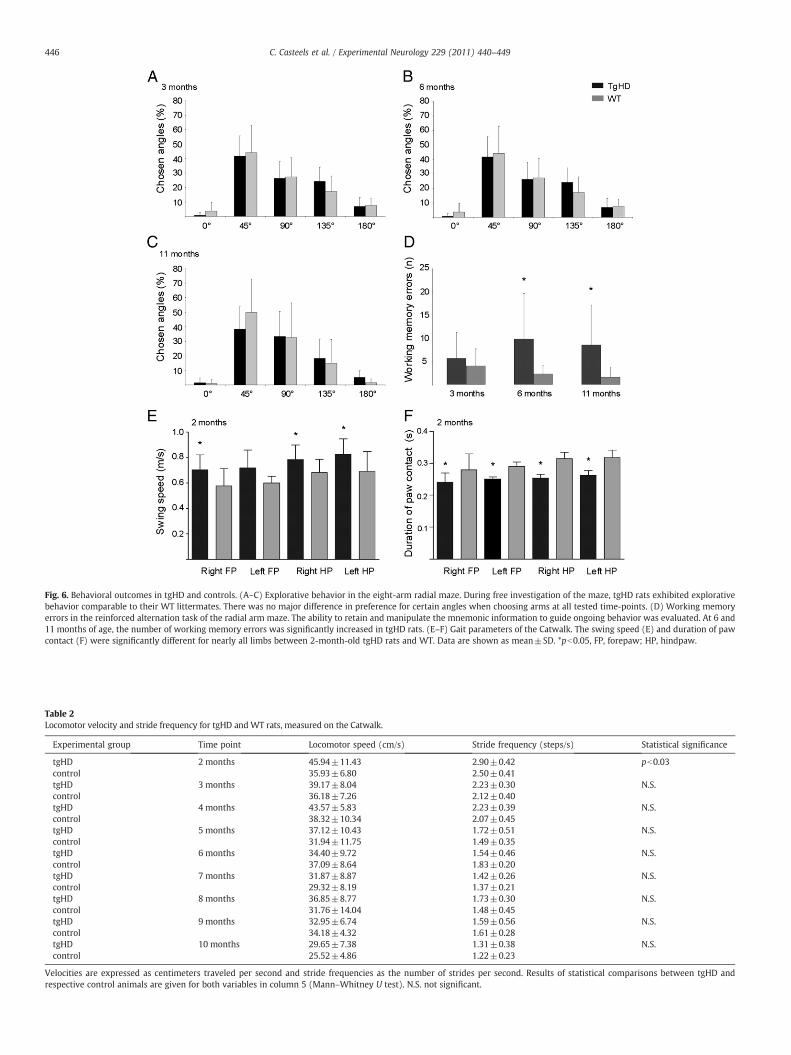
Fig. 6. Behavioral outcomes in tgHD and controls. (A–C) Explorative behavior in the eight-arm radial maze. During free investigation of the maze, tgHD rats exhibited explorativebehavior comparable to their WT littermates. There was no major difference in preference for certain angles when choosing arms at all tested time-points. (D) Working memoryerrors in the reinforced alternation task of the radial arm maze. The ability to retain and manipulate the mnemonic information to guide ongoing behavior was evaluated. At 6 and11 months of age, the number of working memory errors was significantly increased in tgHD rats. (E–F) Gait parameters of the Catwalk. The swing speed (E) and duration of pawcontact (F) were significantly different for nearly all limbs between 2-month-old tgHD rats and WT. Data are shown as mean±SD. *pb0.05, FP, forepaw; HP, hindpaw.
Table 2Locomotor velocity and stride frequency for tgHD andWT rats, measured on the Catwalk.
Experimental group Time point Locomotor speed (cm/s) Stride frequency (steps/s) Statistical significance
tgHD 2 months 45.94±11.43 2.90±0.42 pb0.03control 35.93±6.80 2.50±0.41tgHD 3 months 39.17±8.04 2.23±0.30 N.S.control 36.18±7.26 2.12±0.40tgHD 4 months 43.57±5.83 2.23±0.39 N.S.control 38.32±10.34 2.07±0.45tgHD 5 months 37.12±10.43 1.72±0.51 N.S.control 31.94±11.75 1.49±0.35tgHD 6 months 34.40±9.72 1.54±0.46 N.S.control 37.09±8.64 1.83±0.20tgHD 7 months 31.87±8.87 1.42±0.26 N.S.control 29.32±8.19 1.37±0.21tgHD 8 months 36.85±8.77 1.73±0.30 N.S.control 31.76±14.04 1.48±0.45tgHD 9 months 32.95±6.74 1.59±0.56 N.S.control 34.18±4.32 1.61±0.28tgHD 10 months 29.65±7.38 1.31±0.38 N.S.control 25.52±4.86 1.22±0.23
Velocities are expressed as centimeters traveled per second and stride frequencies as the number of strides per second. Results of statistical comparisons between tgHD andrespective control animals are given for both variables in column 5 (Mann–Whitney U test). N.S. not significant.
446 C. Casteels et al. / Experimental Neurology 229 (2011) 440–449

447C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
Correlation analysis revealed a positive correlation between swingspeed and glucose metabolism in the sensorimotor cortex bilaterally at2 months of age (Fig. 3D; p≤5.3×10−5). The cluster encompassed theprimary motor cortex, the primary (trunk-, dysgranular-region andbarrel field) somatosensory cortex, the thalamic nuclei and the anteriorpart of the hippocampus. No homologous regional correlations of [18F]-FDG, nor behavioral outcomes with relative [18F]MK-9470 wereobserved.
Discussion
In this study, we have for the first time characterized early CB1receptor alterations in vivo in a transgenic rat model of HD using [18F]MK-9470 and small animal PET. We showed that in pre-symptomaticand early symptomatic tgHD rats, CB1 receptor availability is selectivelyaltered in different basal ganglia regions depending on diseaseprogression. Specifically, in 5- and 10-month-old tgHD rats, CB1receptor binding was reduced in the caudate–putamen and lateralglobus pallidus, where cell bodies and axon terminals of CB1 receptor-positive neurons are located, respectively (Herkenham et al., 1990).
Reductions in CB1 receptor binding of these regions are in linewith ex vivo samples of R6/1 transgenic mice at the pre-symptomaticHD stage (Dowie et al., 2009). Dowie et al., demonstrated, in theabsence of neuronal loss, a 20% and 11% reduction in the striatal andpallidal CB1 receptor density of 12-week-old R6/1 mice using [3H]CP55,940 autoradiography. In line with Dowie et al. (2009) andDenovan-Wright and Robertson (2000), we also detected alterationsin CB1 receptor binding prior to the onset of major motor symptomsand in the presence of cognitive deficits. Moreover, the progressivenature of our findings—although modest—may be reminiscent ofprevious data showing that striatal CB1 mRNA progressively declinesin the pre-symptomatic phase to reach a steady-state low levelaround the onset of overt motor symptoms (Denovan-Wright andRobertson, 2000;Naver et al., 2003;McCaw et al., 2004). These findingsof an early in vivo loss of CB1 receptors suggests that [18F]MK-9470may be a useful biomarker for HD. To further determine its usefulness,preclinical in vivo studies tracking the evolution of CB1 levels untillater stages of HD are needed, as well as studies in pre-manifesthuman carriers of the HDmutation using the same imaging techniqueto confirm the clinical significance of our current finding.
The mechanism by which HD promotes this early loss of CB1receptors is suggested to be caused by interactions between mutant httand nuclear transcription factors. Blázquez and coworkers showed instriatal cells that mutant htt down-regulates CB1 receptors bycontrolling gene promoter activity via repressor element 1 silencingtranscription factor and sensitizes cells to excitotoxic damage (Blazquezet al., 2011). In experimental and human HD, CB1 mRNA is also one ofthe most consistently decreased transcripts in postmortem striataltissue (McCaw et al., 2004;Kuhn et al., 2007). Of special interest, tgHDrats display first polyQ recruitment sites around 6 months of age,temporally related to our findings (Nguyen et al., 2006). Also, theabsence of obvious loss in caudate–putamen volume and lateralventricle size, promotes rather involvement of neuronal dysfunctionthan cell death in the early disease process. Despite this agreement,however, other contributing mechanisms such as changes in receptoraffinity or in conformational state cannot be excluded.
Interestingly, in the current study, cross-sectional genotype differ-ences in levels of CB1 receptor evolved from the right hemisphere oftgHD rats to both sides upon disease progression. Using a longitudinaldesign, only bilateral deficits were seen for tgHD rats. These observa-tions together suggest that also in WT rats CB1 receptor changes occurunder our significance threshold, pointing to influence of normal ageingon CB1 receptormeasurements in vivo. The annual ex vivo rate of declinein rats has been estimated at 14% per year for basal ganglia regions(Romero et al., 1998). Slightly lower rates have been observed in brainregions other than extrapyramidal areas (Berrendero et al., 1998),
arguing against the possibility that our findings are related to anincreased rate of ageing in tgHD rats in comparison to controls. Besides,of all basal ganglia areas, the loss of CB1 receptors due to ageing seemspreferentially to occur in striatonigral and striatoentopenduncularpathways, but not in striatalpallidal neurons (Romero et al., 1998).
No correlation of the basal ganglia CB1 receptor deficiency withglucose metabolism measured by [18F]-FDG was observed. Inpre- symptomatic patients and animal models of HD, metabolicdata on striatal regions have been inconsistent, with some studiesdemonstrating normal (Young et al., 1987) but most reduced levels(Hayden et al., 1987;Antonini et al., 1996;Wang et al., 2005) ofglucose metabolism. Here, [18F]-FDG values of striatal regions didnot differ between genotypes, while [18F]-FDG uptake was higher inthe bilateral sensorimotor cortex of 2-month-old transgenic ratas compared to controls. The cortical [18F]-FDG values were alsopositively correlated to motor outcome, suggesting that neuronalactivity of the sensorimotor cortex directly underlies motorperformance on the Catwalk. In agreement with this finding,corticostriatal activity has been shown to be associated withmotor symptoms in YAC128 transgenic mice (Joshi et al., 2009). InYAC128 mice, synaptic responses have been reported to increase inyoung pre-symptomatic animals, consistent to our finding, whereasdevelopment of the motor phenotype coincided with an oppositeeffect. In addition, the 2-month motor phenotype seen on theCatwalk here is in line with the accelerod performance of this model(Nguyen et al., 2006). Improved rotarod performance or increasedgaits with respect to WTs are, however, still abnormal behaviors.R6/2 mice have shown hyperactivity in the open field at 3 weeks ofage (Luesse et al., 2001), but are profoundly dysfunctional on othertests of cognitive function (Lione et al., 1999). It has also beenshown that reduction of cortical BDNF causes early improvementsin rotarod performance, despite neuropathological changes (Baquetet al., 2004).
From our observations it also appears as if early deficits in spatiallearning and memory are not accompanied by in vivo alterations inhippocampal CB1 receptor binding. Although a prominent role ofendocannabinoid signaling has been suggested in controlling hippo-campal symptoms (Solowij and Michie, 2007), our finding isconsistent with previous ex vivo data in other HD models. Insymptomatic R6/1 mice, [3H]CP55,940 autoradiography revealeddecreased ligand binding in striatal projection areas, but no changein the hippocampus and the cortex in the presence of cognitivedysfunction (Dowie et al., 2010). Also, in R6/2 animals displayingcognitive and mnemonic impairments, endocannabinoid levels of thehippocampus did not differ from the levels of WTs at both early andlate stages (Bisogno et al., 2008). This contrasts our in vivo findings inearly symptomatic HD patients, where disease burden was inverselycorrelated with CB1 receptor availability in the prefrontal cortex (VanLaere et al., 2010). Also, in early HD patients, the widespread andrelatively uniform reduction of CB1 receptor availability throughoutthe gray matter is ad odds with the regional selectivity of the earlychanges here. Nonetheless, it cannot be ruled out that species-specificdifferences in receptor–response exist, as, for example, speciesdifferences in brain CB1 receptor distribution have been reported(Burns et al., 2007).
As the disease progresses, transgenic rats expressing a fragment ofthe rat HD genewith 51 CAG repeats have been reported to develop anHD-like phenotype from the age of ~8 months with regard to motorfunction (Nguyen et al., 2006) and cell loss (von Horsten et al., 2003).While we observed modest changes in interpaw coordination andwell-known alterations due to ageing (Walhovd et al., 2005) inparameters such as lateral ventrical size and caudate–putamenvolume, none of them clearly demonstrated a symptomatic HD-likephenotype. Many environmental enrichment studies have shown thathousing conditions can have dramatic effect on the onset andprogression of HD in R6 mice (van Dellen et al., 2000; Hockly et al.,

448 C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
2002). Slight differences in living conditions such as socialisation orbedding can thus potentially influence the course of the disease.Routine care of our rats included provision of normal feed andbedding,as well as the ethics committee's requirement for wooden blocks ineach cage.
In the design of this study, we have explicitly set the aim toinvestigate CB1 receptor network alterations by using both absoluteand relative [18F]MK-9470 measurements. It is known that there is ahigher physiologic interindividual variability of absolute regional [18F]MK-9470 determinations in the human brain, in the order of severaltens of percents (Barrero et al., 2005;Burns et al., 2007). Relativemeasurements have the advantage of being much more sensitive,allowing changes of 5%–10% to be measured using StatisticalParametric Mapping (Van Laere et al., 2002), as proven here.
It is also known that the use of anesthesia may significantly changetracer kinetics and uptake of [18F]-FDG. It has been reported thatisoflurane induces changes in brain glucose metabolism in rodents (e.g.cortical changes), but so do other anesthetics (Toyama et al., 2004).Moreover, also inmonkey, effects of isoflurane have been demonstrated(Noda et al., 2003). As only relative [18F]-FDG measurements wereperformed and the control group underwent exactly the sameprocedure, we have assumed that possible differential effects ofanesthesia could be neglected.
Conclusion
In vivo cerebralmappingof pre-symptomatic andearly symptomaticrats transgenic for HD using [18F]MK-9470 small animal PET points toearly regional dysfunctions in endocannabinoid signalling, incorporat-ing the lateral globus pallidus and caudate–putamen. In vivo CB1receptor measurements using [18F]MK-9470may thus be a useful earlybiomarker for HD. Translational studies in premanifest human carriersof the HD mutation using the same imaging technique are needed tofurther demonstrate the clinical significance of this finding. Our resultsalso provide evidence of (1) early metabolic upregulation in thesensorimotor cortex and (2) of subtle motor and cognitive deficits atearlier stages than previously described.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Acknowledgments
Merck & Co, Inc. is acknowledged for the availability of the [18F]MK-9470 precursor, and for their critical revision of this manuscriptand their improving suggestions. The authors thank Peter Vermae-len and Ann Van Santvoort for their assistance in data acquisition, aswell as the Leuven PET radiopharmacy team for tracer preparations.Financial support of the Research Council of the KatholiekeUniversiteit Leuven (OT/05/58), the Fund for Scientific Research,Flanders, Belgium (FWO/G.0548.06), the K.U.Leuven MolecularSmall Animal Imaging Center (KUL EF/05/08), and the Institute forthe Promotion of Innovation by Science and Technology in Flanders(SBO50151/SBO060819) is gratefully acknowledged. Part of thiswork is also performed under the European Commission FP6-project Diagnostic Molecular Imaging (DIMI), LSHB-CT-2005-512146. Cindy Casteels is supported by a post-doctoral mandateof the Flemish Fund for Scientific Research. Koen Van Laere is asenior clinical investigator of the Flemish Fund for ScientificResearch. Caroline Vandeputte is a doctoral fellow of the Institutefor the Promotion of Innovation through Science and Technology inFlanders (IWT).
References
Antonini, A., Leenders, K.L., Spiegel, R., Meier, D., Vontobel, P., Weigell-Weber, M.,Sanchez-Pernaute, R., de Yebenez, J.G., Boesiger, P., Weindl, A., Maguire, R.P., 1996.Striatal glucose metabolism and dopamine D2 receptor binding in asymptomaticgene carriers and patients with Huntington's disease. Brain 119 (Pt 6), 2085–2095.
Baquet, Z.C., Gorski, J.A., Jones, K.R., 2004. Early striatal dendrite deficits followed byneuron loss with advanced age in the absence of anterograde cortical brain-derivedneurotrophic factor. J. Neurosci. 24, 4250–4258.
Barrero, F.J., Ampuero, I., Morales, B., Vives, F., de Dios Luna Del Castillo, J., Hoenicka, J.,Garcia, Y.J., 2005. Depression in Parkinson's disease is related to a geneticpolymorphism of the cannabinoid receptor gene (CNR1). Pharmacogenomics J. 5,135–141.
Berrendero, F., Romero, J., Garcia-Gil, L., Suarez, I., De la Cruz, P., Ramos, J.A., Fernandez-Ruiz, J.J., 1998. Changes in cannabinoid receptor binding andmRNA levels in severalbrain regions of aged rats. Biochim. Biophys. Acta 1407, 205–214.
Bisogno, T., Martire, A., Petrosino, S., Popoli, P., Di Marzo, V., 2008. Symptom-relatedchanges of endocannabinoid and palmitoylethanolamide levels in brain areas of R6/2mice, a transgenic model of Huntington's disease. Neurochem. Int. 52, 307–313.
Blazquez, C., Chiarlone, A., Sagredo, O., Aguado, T., Pazos, M.R., Resel, E., Palazuelos, J.,Julien, B., Salazar, M., Borner, C., Benito, C., Carrasco, C., ez-Zaera, M., Paoletti, P., az-Hernandez, M., Ruiz, C., Sendtner, M., Lucas, J.J., de Yebenes, J.G., Marsicano, G.,Monory, K., Lutz, B., Romero, J., Alberch, J., Gines, S., Kraus, J., Fernandez-Ruiz, J.,Galve-Roperh, I., Guzman, M., 2011. Loss of striatal type 1 cannabinoid receptors isa key pathogenic factor in Huntington's disease. Brain 134, 119–136.
Burns, H.D., Van Laere, K., Sanabria-Bohorquez, S., Hamill, T.G., Bormans, G., Eng, W.S.,Gibson, R., Ryan, C., Connolly, B., Patel, S., Krause, S., Vanko, A., Van Hecken, A.,Dupont, P., De Lepeleire, I., Rothenberg, P., Stoch, S.A., Cote, J., Hagmann, W.K.,Jewell, J.P., Lin, L.S., Liu, P., Goulet, M.T., Gottesdiener, K., Wagner, J.A., de Hoon, J.,Mortelmans, L., Fong, T.M., Hargreaves, R.J., 2007. [18F]MK-9470, a positronemission tomography (PET) tracer for in vivo human PET brain imaging of thecannabinoid-1 receptor. Proc. Natl. Acad. Sci. U. S. A. 104, 9800–9805.
Casteels, C., Lauwers, E., Baitar, A., Bormans, G., Baekelandt, V., Van Laere, K., 2010a. Invivo type 1 cannabinoid receptor mapping in the 6-hydroxydopamine lesion ratmodel of Parkinson's disease. Brain Res. 1316, 153–162.
Casteels, C., Martinez, E., Bormans, G., Camon, L., de Vera, N., Baekelandt, V., Planas, A.M.,Van Laere, K., 2010b. Type 1 cannabinoid receptor mapping with [(18)F]MK-9470PET in the rat brain after quinolinic acid lesion: a comparison to dopaminereceptors and glucose metabolism. Eur. J. Nucl. Med. Mol. Imaging 37, 2354–2363.
Casteels, C., Vermaelen, P., Nuyts, J., Van Der Linden, A., Baekelandt, V., Mortelmans, L.,Bormans, G., Van Laere, K., 2006. Construction and evaluation of multitracer small-animal PET probabilistic atlases for voxel-based functional mapping of the rat brain.J. Nucl. Med. 47, 1858–1866.
Denovan-Wright, E.M., Robertson, H.A., 2000. Cannabinoid receptor messenger RNAlevels decrease in a subset of neurons of the lateral striatum, cortex andhippocampus of transgenic Huntington's disease mice. Neuroscience 98, 705–713.
Dowie, M.J., Bradshaw, H.B., Howard, M.L., Nicholson, L.F., Faull, R.L., Hannan, A.J., Glass,M., 2009. Altered CB1 receptor and endocannabinoid levels precede motorsymptom onset in a transgenic mouse model of Huntington's disease. Neuroscience163, 456–465.
Dowie, M.J., Howard, M.L., Nicholson, L.F., Faull, R.L., Hannan, A.J., Glass, M., 2010.Behavioural and molecular consequences of chronic cannabinoid treatment inHuntington's disease transgenic mice. Neuroscience 170, 324–336.
Glass, M., Dragunow, M., Faull, R.L., 2000. The pattern of neurodegeneration inHuntington's disease: a comparative study of cannabinoid, dopamine, adenosineand GABA(A) receptor alterations in the human basal ganglia in Huntington'sdisease. Neuroscience 97, 505–519.
Goffin, K., Bormans, G., Casteels, C., Bosier, B., Lambert, D.M., Grachev, I.D., VanPaesschen, W., Van Laere, K., 2008. An in vivo [(18)F]MK-9470 microPET study oftype 1 cannabinoid receptor binding in Wistar rats after chronic administration ofvalproate and levetiracetam. Neuropharmacology 54, 1103–1106.
Goutopoulos, A., Makriyannis, A., 2002. From cannabis to cannabinergics: newtherapeutic opportunities. Pharmacol. Ther. 95, 103–117.
Hayden, M.R., Hewitt, J., Stoessl, A.J., Clark, C., Ammann, W., Martin, W.R., 1987. Thecombined use of positron emission tomography and DNA polymorphisms forpreclinical detection of Huntington's disease. Neurology 37, 1441–1447.
Herkenham, M., Lynn, A.B., Little, M.D., Johnson,M.R., Melvin, L.S., de Costa, B.R., Rice, K.C.,1990. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. U. S. A. 87,1932–1936.
Hockly, E., Cordery, P.M.,Woodman, B.,Mahal, A., vanDellen, A., Blakemore, C., Lewis, C.M.,Hannan,A.J., Bates,G.P., 2002. Environmental enrichment slowsdiseaseprogression inR6/2 Huntington's disease mice. Ann. Neurol. 51, 235–242.
Holter, S.M., Tzschentke, T.M., Schmidt, W.J., 1996. Effects of amphetamine, morphineand dizocilpine (MK-801) on spontaneous alternation in the 8-arm radial maze.Behav. Brain Res. 81, 53–59.
Joshi, P.R., Wu, N.P., Andre, V.M., Cummings, D.M., Cepeda, C., Joyce, J.A., Carroll, J.B.,Leavitt, B.R., Hayden, M.R., Levine, M.S., Bamford, N.S., 2009. Age-dependentalterations of corticostriatal activity in the YAC128 mouse model of Huntingtondisease. J. Neurosci. 29, 2414–2427.
Katona, I., Freund, T.F., 2008. Endocannabinoid signaling as a synaptic circuit breaker inneurological disease. Nat. Med. 14, 923–930.
Koopmans, G.C., Deumens, R., Brook, G., Gerver, J., Honig,W.M., Hamers, F.P., Joosten, E.A.,2007. Strain and locomotor speed affect over-ground locomotion in intact rats.Physiol. Behav. 92, 993–1001.
Kuhn, A., Goldstein,D.R., Hodges, A., Strand, A.D., Sengstag, T., Kooperberg, C., Becanovic, K.,Pouladi, M.A., Sathasivam, K., Cha, J.H., Hannan, A.J., Hayden, M.R., Leavitt, B.R.,

449C. Casteels et al. / Experimental Neurology 229 (2011) 440–449
Dunnett, S.B., Ferrante, R.J., Albin, R., Shelbourne, P., Delorenzi, M., Augood, S.J.,Faull, R.L., Olson, J.M., Bates, G.P., Jones, L., Luthi-Carter, R., 2007. Mutant huntingtin'seffects on striatal gene expression in mice recapitulate changes observed in humanHuntington's disease brain and do not differ with mutant huntingtin length or wild-type huntingtin dosage. Hum. Mol. Genet. 16, 1845–1861.
Lastres-Becker, I., Berrendero, F., Lucas, J.J., Martin-Aparicio, E., Yamamoto, A., Ramos, J.A.,Fernandez-Ruiz, J.J., 2002. Loss ofmRNA levels, binding and activation of GTP-bindingproteins for cannabinoid CB1 receptors in the basal ganglia of a transgenic model ofHuntington's disease. Brain Res. 929, 236–242.
Lione, L.A., Carter, R.J., Hunt, M.J., Bates, G.P., Morton, A.J., Dunnett, S.B., 1999. Selectivediscrimination learning impairments in mice expressing the human Huntington'sdisease mutation. J. Neurosci. 19, 10428–10437.
Luesse, H.G., Schiefer, J., Spruenken, A., Puls, C., Block, F., Kosinski, C.M., 2001.Evaluation of R6/2 HD transgenic mice for therapeutic studies in Huntington'sdisease: behavioral testing and impact of diabetes mellitus. Behav. Brain Res. 126,185–195.
Maes, F., Collignon, A., Vandermeulen, D., Marchal, G., Suetens, P., 1997. Multimodalityimage registration by maximization of mutual information. IEEE Trans. Med.Imaging 16, 187–198.
Magistretti, P.J., 2006. Neuron-glia metabolic coupling and plasticity. J. Exp. Biol. 209,2304–2311.
McCaw, E.A., Hu, H., Gomez, G.T., Hebb, A.L., Kelly, M.E., Denovan-Wright, E.M., 2004.Structure, expression and regulation of the cannabinoid receptor gene (CB1) inHuntington's disease transgenic mice. Eur. J. Biochem. 271, 4909–4920.
Naver, B., Stub, C., Moller, M., Fenger, K., Hansen, A.K., Hasholt, L., Sorensen, S.A., 2003.Molecular and behavioral analysis of the R6/1 Huntington's disease transgenicmouse. Neuroscience 122, 1049–1057.
Nguyen, H.P., Kobbe, P., Rahne, H., Worpel, T., Jager, B., Stephan, M., Pabst, R., Holzmann,C., Riess, O., Korr, H., Kantor, O., Petrasch-Parwez, E., Wetzel, R., Osmand, A., vonHorsten, S., 2006. Behavioral abnormalities precede neuropathological markers inrats transgenic for Huntington's disease. Hum. Mol. Genet. 15, 3177–3194.
Noda, A., Takamatsu, H., Minoshima, S., Tsukada, H., Nishimura, S., 2003. Determinationof kinetic rate constants for 2-[18F]fluoro-2-deoxy-D-glucose and partitioncoefficient of water in conscious macaques and alterations in aging or anesthesiaexamined on parametric images with an anatomic standardization technique.J. Cereb. Blood Flow Metab. 23, 1441–1447.
Richfield, E.K., Herkenham, M., 1994. Selective vulnerability in Huntington's disease:preferential loss of cannabinoid receptors in lateral globus pallidus. Ann. Neurol.36, 577–584.
Romero, J., Berrendero, F., Garcia-Gil, L., De la Cruz, P., Ramos, J.A., Fernandez-Ruiz, J.J.,1998. Loss of cannabinoid receptor binding and messenger RNA levels andcannabinoid agonist-stimulated [35S]guanylyl-5'O-(thio)-triphosphate binding inthe basal ganglia of aged rats. Neuroscience 84, 1075–1083.
Sanabria-Bohorquez, S.M., Hamill, T.G., Goffin, K., De Lepeleire, I., Bormans, G., Burns, H.D.,Van Laere, K., 2010. Kinetic analysis of the cannabinoid-1 receptor PET tracer [(18)F]MK-9470 in human brain. Eur. J. Nucl. Med. Mol. Imaging 37, 920–933.
Solowij, N., Michie, P.T., 2007. Cannabis and cognitive dysfunction: parallels withendophenotypes of schizophrenia? J. Psychiatry Neurosci. 32, 30–52.
The Huntington's Disease Collaborative Research Group, 1993. A novel gene containing atrinucleotide repeat that is expanded and unstable on Huntington's disease chromo-somes. The Huntington's Disease Collaborative Research Group. Cell 72, 971–983.
Toyama, H., Ichise, M., Liow, J.S., Modell, K.J., Vines, D.C., Esaki, T., Cook, M., Seidel, J.,Sokoloff, L., Green, M.V., Innis, R.B., 2004. Absolute quantification of regionalcerebral glucose utilization in mice by 18F-FDG small animal PET scanning and 2-14C-DG autoradiography. J. Nucl. Med. 45, 1398–1405.
van Dellen, A., Blakemore, C., Deacon, R., York, D., Hannan, A.J., 2000. Delaying the onsetof Huntington's in mice. Nature 404, 721–722.
Van Laere, K., Casteels, C., Dhollander, I., Goffin, K., Grachev, I., Bormans, G.,Vandenberghe, W., 2010. Widespread decrease of type 1 cannabinoid receptoravailability in Huntington disease in vivo. J. Nucl. Med. 51, 1413–1417.
Van Laere, K.J., Versijpt, J., Koole, M., Vandenberghe, S., Lahorte, P., Lemahieu, I., Dierckx, R.A.,2002. Experimental performance assessment of SPM for SPECT neuroactivation studiesusing a subresolution sandwich phantom design. Neuroimage 16, 200–216.
Vandeputte, C., Taymans, J.M., Casteels, C., Coun, F., Ni, Y., Van Laere, K., Baekelandt, V.,2010. Automated quantitative gait analysis in animal models of movementdisorders. BMC Neurosci. 9, 11–92.
von Horsten, S., Schmitt, I., Nguyen, H.P., Holzmann, C., Schmidt, T., Walther, T., Bader,M., Pabst, R., Kobbe, P., Krotova, J., Stiller, D., Kask, A., Vaarmann, A., Rathke-Hartlieb, S., Schulz, J.B., Grasshoff, U., Bauer, I., Vieira-Saecker, A.M., Paul, M., Jones,L., Lindenberg, K.S., Landwehrmeyer, B., Bauer, A., Li, X.J., Riess, O., 2003. Transgenicrat model of Huntington's disease. Hum. Mol. Genet. 12, 617–624.
Vonsattel, J.P., Myers, R.H., Stevens, T.J., Ferrante, R.J., Bird, E.D., Richardson Jr., E.P.,1985. Neuropathological classification of Huntington's disease. J. Neuropathol. Exp.Neurol. 44, 559–577.
Walhovd, K.B., Fjell, A.M., Reinvang, I., Lundervold, A., Dale, A.M., Eilertsen, D.E., Quinn,B.T., Salat, D., Makris, N., Fischl, B., 2005. Effects of age on volumes of cortex, whitematter and subcortical structures. Neurobiol. Aging 26, 1261–1270.
Wang, X., Sarkar, A., Cicchetti, F., Yu, M., Zhu, A., Jokivarsi, K., Saint-Pierre, M., Brownell,A.L., 2005. Cerebral PET imaging and histological evidence of transglutaminaseinhibitor cystamine induced neuroprotection in transgenic R6/2 mouse model ofHuntington's disease. J. Neurol. Sci. 231, 57–66.
Wooley, C.M., Xing, S., Burgess, R.W., Cox, G.A., Seburn, K.L., 2009. Age, experience andgenetic background influence treadmill walking inmice. Physiol. Behav. 96, 350–361.
Young, A.B., Penney, J.B., Starosta-Rubinstein, S., Markel, D., Berent, S., Rothley, J., Betley,A., Hichwa, R., 1987. Normal caudate glucose metabolism in persons at risk forHuntington's disease. Arch. Neurol. 44, 254–257.




















