
Meiotic behaviour of chromosomes 1R, 2R and 5R in autotetraploid rye
8
Chromosoma (Berl) (1984) 89:143-150 CHROMOSOMA Springer-Verlag 1984 Meiotic behaviour of chromosomes 1R, 2R and 5R in autotetraploid rye Tomfis Naranjo 1 and Juan Orellana 2 1 Departamento de Gen~tica, Facultad de Biologia, Universidad de Oviedo, Oviedo, Spain; 2 Departamento de Gen6tica, Facultad de Biologia, Universidad Complutense de Madrid, Madrid, Spain Abstract. The meiotic behaviour of chromosomes IR, 2R and 5R was studied in C-banded preparations of autote- traploid rye. Analysis of pairing and chiasma formation was based on metaphase I configurations, using the model designed by Sybenga, with slight modifications. Frequencies of two modes of pairing (one quadrivalent or two bivalents) differed from those expected for random pairing. Although preferential pairing for some arm pairs of chromosome 2R was detected, this did not seem to be the cause of the in- creased bivalent pairing. This increase was attributed to either the spatial separation of the four homologous chro- mosomes in some premeiotic cells into two groups of two, or a correction of the synaptonemal complex, or both. The number of chiasmate associations showed variation be- tween chromosomes and between arms within the same chromosome. It was closely related to arm length, but dif- ferent after quadrivalent and bivalent pairing. This is sug- gested to be a consequence of partner exchange interfering with pairing and, consequently, with chiasma formation, and a different chiasma distribution after quadrivalent pair- ing. Variation between chromosomes in the frequencies of alternate and adjacent co-orientation in metaphase I quad- rivalents without interstitial chiasmata suggests that the rel- ative positions of the centromeres in the quadrivalent influ- ence their co-orientation. Introduction Chromosome pairing in autotetraploids can rarely be stud- ied at stages where pairing is at its maximum. Usually only later stages are accessible: diplotene and, more often, diaki- nesis and metaphase I. The distribution of the different kinds of chromosome configurations: multivalents, biva- lents and univalents, at diakinesis or metaphase I depends upon the patterns of chromosome pairing and the frequency and location of chiasmata between homologous chromo- somes (for reviews see: Sybenga 1975 a; Jackson and Casey 1980). Sybenga (1975b) has designed a model that allows an estimation of the frequency of the two modes of pairing involving four homologous chromosomes with two terminal pairing initiation points per chromosome (one quadrivalent or two bivalents) as well as an estimation of the frequency of being bound by chiasmata for each chromosome arm for both pairing types. The model concerns metacentric chromosomes but without the possibility of distinguishing between the arms at the stage of observation, and only considers organisms with pronounced distal chiasma local- ization, i.e., no chiasmata in interstitial segments. The fol- lowing metaphase I configuration types can then be distin- guished: ring quadrivalent (frequency rq); chain quadriva- lent (frequency cq); trivalent and univalent (frequency t); ring bivalent (frequency r); open bivalent (frequency o); and univalent pair (frequency u). The first three must neces- sarily derive from quadrivalent pairing; the ring bivalents are assumed to be derived from bivalent pairing and the open bivalents and univalent pairs can be formed both after quadrivalent and bivalent pairing. When the frequency of quadrivalent pairing is f, and the probabilities of chiasmate association of the end seg- ments of both chromosome arms are a' and b' in quadriva- lents, and a and b in bivalents, six equations can be derived. Application of these equations to observations on the pooled chromosomes of Tradescantia virginiana gave con- figuration frequencies ahnost exactly equal to those ob- served and demonstrated that the estimates obtained for f, a', b', a and b were quite acceptable. In rye where there are interstitial chiasmata in addition to distal ones, the model is also applicable by incorporation of the additional configurations in the categories of configu- rations without interstitial chiasmata (Sybenga 1975b). Even with this apparent complication the analysis can be considerably refined by identifying specific chromosomes and their respective arms by C-banding. Figure 1 presents pairing and metaphase I configura- tions for a theoretical rye chromosome set carrying a marker telomeric C-band in the short arm of the four homo- logues (Centromere co-orientation has not been taken into account). Three modes of pairing are considered. Qs and Q1 are quadrivalents with the point of partner exchange in the short and the long arm, respectively. B represents two pairs of bivalents. All metaphase I quadrivalents and trivalents plus univalents derive from Qs and Q1 (ql, q4, q6, ql0, q12, tl and t4 from Qs; q2, q5, q7, qll, q13, t2 and t5 from Q1 and q3, q8, q9 and t3 from both Qs and Q1). Ring bivalents, which in Sybenga's model only arise from bivalent pairing, can also originate after quadri- valent pairing. For example, bl can be formed from Qs or Q1 by chiasmata in L, L, Si and Si or S, S, Li and Li, respectively. Thus, this configuration may constitute a source of error in the model, its magnitude depending on chiasma frequency in interstitial and pairing segments.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Meiotic behaviour of chromosomes 1R, 2R and 5R in autotetraploid rye

Chromosoma (Berl) (1984) 89:143-150 CHROMOSOMA �9 Springer-Verlag 1984
Meiotic behaviour of chromosomes 1R, 2R and 5R in autotetraploid rye Tomfis Naranjo 1 and Juan Orellana 2
1 Departamento de Gen~tica, Facultad de Biologia, Universidad de Oviedo, Oviedo, Spain; 2 Departamento de Gen6tica, Facultad de Biologia, Universidad Complutense de Madrid, Madrid, Spain
Abstract. The meiotic behaviour of chromosomes IR, 2R and 5R was studied in C-banded preparations of autote- traploid rye. Analysis of pairing and chiasma formation was based on metaphase I configurations, using the model designed by Sybenga, with slight modifications. Frequencies of two modes of pairing (one quadrivalent or two bivalents) differed from those expected for random pairing. Although preferential pairing for some arm pairs of chromosome 2R was detected, this did not seem to be the cause of the in- creased bivalent pairing. This increase was attributed to either the spatial separation of the four homologous chro- mosomes in some premeiotic cells into two groups of two, or a correction of the synaptonemal complex, or both. The number of chiasmate associations showed variation be- tween chromosomes and between arms within the same chromosome. It was closely related to arm length, but dif- ferent after quadrivalent and bivalent pairing. This is sug- gested to be a consequence of partner exchange interfering with pairing and, consequently, with chiasma formation, and a different chiasma distribution after quadrivalent pair- ing. Variation between chromosomes in the frequencies of alternate and adjacent co-orientation in metaphase I quad- rivalents without interstitial chiasmata suggests that the rel- ative positions of the centromeres in the quadrivalent influ- ence their co-orientation.
Introduction
Chromosome pairing in autotetraploids can rarely be stud- ied at stages where pairing is at its maximum. Usually only later stages are accessible: diplotene and, more often, diaki- nesis and metaphase I. The distribution of the different kinds of chromosome configurations: multivalents, biva- lents and univalents, at diakinesis or metaphase I depends upon the patterns of chromosome pairing and the frequency and location of chiasmata between homologous chromo- somes (for reviews see: Sybenga 1975 a; Jackson and Casey 1980).
Sybenga (1975b) has designed a model that allows an estimation of the frequency of the two modes of pairing involving four homologous chromosomes with two terminal pairing initiation points per chromosome (one quadrivalent or two bivalents) as well as an estimation of the frequency of being bound by chiasmata for each chromosome arm for both pairing types. The model concerns metacentric
chromosomes but without the possibility of distinguishing between the arms at the stage of observation, and only considers organisms with pronounced distal chiasma local- ization, i.e., no chiasmata in interstitial segments. The fol- lowing metaphase I configuration types can then be distin- guished: ring quadrivalent (frequency rq); chain quadriva- lent (frequency cq); trivalent and univalent (frequency t); ring bivalent (frequency r); open bivalent (frequency o); and univalent pair (frequency u). The first three must neces- sarily derive from quadrivalent pairing; the ring bivalents are assumed to be derived from bivalent pairing and the open bivalents and univalent pairs can be formed both after quadrivalent and bivalent pairing.
When the frequency of quadrivalent pairing is f, and the probabilities of chiasmate association of the end seg- ments of both chromosome arms are a' and b' in quadriva- lents, and a and b in bivalents, six equations can be derived. Application of these equations to observations on the pooled chromosomes of Tradescantia virginiana gave con- figuration frequencies ahnost exactly equal to those ob- served and demonstrated that the estimates obtained for f, a', b', a and b were quite acceptable.
In rye where there are interstitial chiasmata in addition to distal ones, the model is also applicable by incorporation of the additional configurations in the categories of configu- rations without interstitial chiasmata (Sybenga 1975b). Even with this apparent complication the analysis can be considerably refined by identifying specific chromosomes and their respective arms by C-banding.
Figure 1 presents pairing and metaphase I configura- tions for a theoretical rye chromosome set carrying a marker telomeric C-band in the short arm of the four homo- logues (Centromere co-orientation has not been taken into account). Three modes of pairing are considered. Qs and Q1 are quadrivalents with the point of partner exchange in the short and the long arm, respectively. B represents two pairs of bivalents. All metaphase I quadrivalents and trivalents plus univalents derive from Qs and Q1 (ql, q4, q6, ql0, q12, tl and t4 from Qs; q2, q5, q7, q l l , q13, t2 and t5 from Q1 and q3, q8, q9 and t3 from both Qs and Q1). Ring bivalents, which in Sybenga's model only arise from bivalent pairing, can also originate after quadri- valent pairing. For example, bl can be formed from Qs or Q1 by chiasmata in L, L, Si and Si or S, S, Li and Li, respectively. Thus, this configuration may constitute a source of error in the model, its magnitude depending on chiasma frequency in interstitial and pairing segments.

144
L Si Si L " " I I �9 t ~ I �9 1='1 [
Os Ol Le B "
"Cll q2 q3 q4 q5
S I
L I
!
L I
q6
'Cl7 q8 q9 qlO (111 q12
q13 t l t 2 t3 t4 t5
"[ol b2 b3 b4 b5 b6
l" 00 0 , a b7 b8
i "
b9 blO ~ / . % /"
% / ,,.
q14
Fig. 1. Pairing and metaphase I configurations for four homolo- gous chromosomes of rye with a heterochromatic telomeric band in the short arm, Metaphase I configurations ql, q4, q6, q10, qt2, tl and t4 arise from quadrivalent pairing with partner exchange in the short arm (Qs). Configurations q2, q5, q7, ql t , q13, t2 and t5 arise from quadrivalent pairing with partner exchange in the long arm (QI). Configurations q3, q8, qg, and t3 can be formed from both Qs and QI. Configurations bl to bl0 can derive from all three pairing types Qs, Q1 and B. If, in addition to the two end initiation pairing points, there is one interstitial initiation pair- ing point, q14 can be formed
Open bivalents and univalent pairs are formed from both quadrivalent and bivalent pairing. Metaphase I quadriva- lent configuration q14, with one par tner exchange in the short a rm and another in the long arm, arise from a differ- ent type of quadrivalent pairing. It requires an additional interstitial initiation pairing point and will not be consid- ered.
For adjustment to Sybenga's model, different configura- tions must be included in the same class. Thus, class rq (" ring quadrivalents") includes quadrivalents with the four arm pairs associated and has the frequency rq = ql + q2 + q3.
Class cq ("chain quadrivalents") including quadriva- lents with three a rm pairs bound can be separated into two subclasses: one having bound two pairs of long arms, frequency cql=q5 + q 6 + q8, and the other with two pairs of short arms bound, frequency cqs = q4 + q7 + q9. Configu- rations q6 and q7 lack chiasmata in one segment Si and Se (Qs) and in one segment Li and Le (Q1), respectively. They have one whole a rm pair not bound.
Class t (" t r ivalents") contains multivalents with one pair of long arms and one pair of short arms associated, frequency t = q12 + ql 3 + tl + t2 + t3. Configurations q12
and tl have a similar origin; they arise by chiasmate associ- ation in one long ann pair (L in Qs) in addition to chiasmate associations in one interstitial segment (Si) and one end segment (Se) of the short arm. Likewise, configurations q13 and t2 are comparable.
Class r (" ring bivalents") contains only ring bivalents, and its frequency is r = 1/2(2bl + b2 + b3 + b4).
Class o (" open bivalents") includes open bivalents and the equivalent in open bivalents of multivalent configura- tions ql0, q11, t4 and t5. Configurations q l0 and q l l are comparable to configurations b5 and b6, respectively, since they show the same two arm pairs associated. In a similar way t4 and t5 are comparable to b8 and b9. As in cq, class o can be subdivided into two subclasses: one including open bivalents or multivalents with the long arm associated, frequency ol= 1/2(2ql I + t5 + b2 + 2b6 + b7 + b9), and the other those with the short a rm associated, frequency os = 1/2(2ql 0 + t4 + b3 + 2b5 + b7 + b8).
Class u (" univalent pa i rs") contains univalent pairs and the equivalent in univalent pairs of configurations t4 and t5, its frequency is u = 1/2(t4 + t5 + b4 + b8 + b9 + 2b10).
Subdivision of classes cq and o requires that Eqs. (2) and (5) of Sybenga (1975b) must be split up into:
cql = 2f" (a' .b '2 - - a '2" b '2) (2,1) cqs = 2f ' ( a ' 2 ' b' - a '2. b '2) (2.2) ol = f" ( b ' - 2a ' . b ' + a '2. b ') + (1 - f ) ( b - a. b) (5.1) os = f. (a' - 2a ' . b ' + a ' . b '2) + (1 - f)(a - a" b) (5,2)
which adds two degrees of freedom. The remaining equations remain as in Sybenga (1975 b)
rq = f ' a ' 2 . b '2 (1) t = 4f- (a' . b ' - a ' . b '2 - - a '2 ' b ' + a '2. b '2) (3) r = ( 1 - f ) . a . b (4) u = f . ( 1 - a ' - b ' + a ' 2 - b ' + a ' . b ' 2 - a ' 2 . b '2) (6)
+(1 - f ) ( a + b - 2 a - b )
The overall frequencies g and ~ of chiasmate associa- tions of short and long arms, respectively, can be estimated as follows:
= 1/2(2rq + cql + 2cqs + t + 2r + 2os) = 1/2(2rq + 2cql+ cqs + t + 2r + 2oi)
The possibility of distinguishing between chromosomes also allows the analysis of centromere co-orientation in quadrivalents at metaphase I. In the present study we ana- lyze pairing, bound arm frequencies and centromere co- orientation for chromosomes 1R, 2R and 5R of autotetra- ploid rye. These chromosomes are submetacentric, but with a different long arm: short a rm ratios. Chromosome 5R is the least metacentric and chromosome 2R is usually the most nearly metacentric. All of them can be recognized at meiosis by C-banding.
Material and methods
Plants belonging to the F 1 of the cross between two inbred lines of rye Secale cereale (P and E) were treated with col- chicine by the capping method. By self-fertilization of this F1, five autotetraploid plants, which formed the material of this study, were obtained among the offspring.
C-banding was performed according to GirAldez et al. (1979) and 100 pollen mother cells (PMCs) per plant were studied.

line P line E
1R 2R 5R 1R 2R 5R
Di nt ii tt DD F1
1R 2R 5R
Colchicine and self-fertilization
F 2 (4n)
Plants 1R 2R 5R
:oOlO-, DDii Dill Dill
:oloo:: DiDD ttDB tDOD
20,o. DDii tDtt DDD0 Fig, 2. Pedigree and C-banding pattern for chromosomes 1R, 2R and 5R of the five autotetraploid plants obtained by chromosome duplication and self-fertilization of the F 1 of the cross between the two rye inbred lines P and E
Results
C-banding
Chromosomes 1R, 2R and 5R of rye (see Sybenga 1983) have been previously identified by C-banding in inbred lines P and E and in their F 1 (Girfildez et al. 1979; Girfildez and Orellana 1979). The C-banding patterns they presented in five autotetraploid plants are shown in the pedigree of Figure 2. Chromosome 1R (Satellite chromosome) was easi- ly identifiable by the interstitial C-band adjacent to the nucleolar organizer region (NOR). Two of the four 1R ho- mologues presented a prominent telomeric band in the long arm in two plants, and only one was marked in this way in the remaining three plants. In the homologous 2R set two chromosomes carried a thick band and the other two a thin band at the telomere of the short arm in four plants. The remaining plant carried only one chromosome with the prominent band. (A still thinner band was also observed at the telomere of the long arm in most chromosomes.) The four 5R homologues showed, as expected, the same C-banding pattern in all plants, namely, a thick telomeric band in the short arm and a subterminal thin band in the long one.
Pairing
As both arms of these chromosomes could be distinguished, their respective metaphase I configurations were easily ac-
145
Fig. 3a-e. Metaphase I configurations in autotetraploid rye. a Chromosome 1R shows configuration b2 of Fig. 1; chromosome 2R, configuration bl; and chromosome 5R configuration qS. b Chromosomes 1R and 2R show configuration q8; and chromo- some 5R, configuration q3. e Chromosome 1R shows configuration b3; chromosome 2R, configuration ql i (arrows indicate the two chromosomes carrying the prominent telomeric band in the short arm); and chromosome 5R, configuration q5. Bar represents 5 ktm
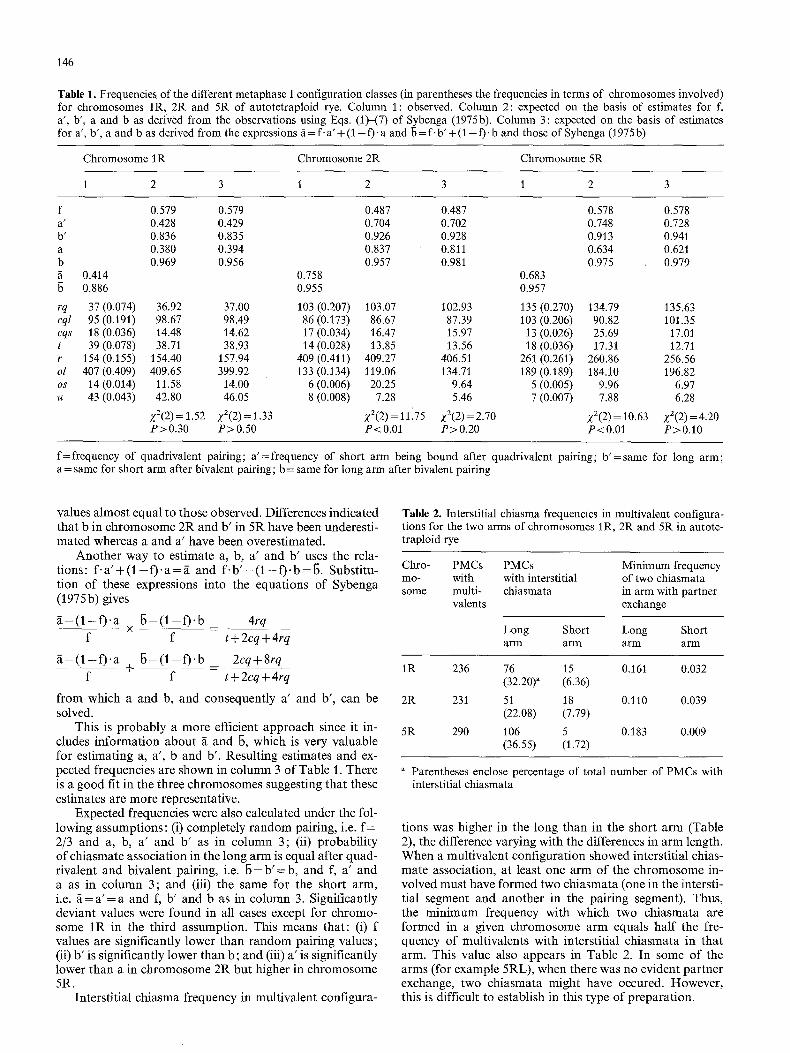
commodated to those of Figure 1. Examples of such config- urations are given in Figure 3. Four PMCs (two for chro- mosome 1R and two for chromosome 2R) that presented configuration q14 (Fig. 1) were not considered, The ob- served frequencies for each chromosome, which are clearly different, were pooled for the five plants (Table 1, column 1). Significant differences between plants were not detected. The average chiasmate association frequencies fi (for the short arm) and b (for the long arm) show some variation, especially for fi, between chromosomes with the lowest values for chromosome 1R.
Estimating f on the basis of Eq. (7) of Sybenga (1975b) yields values (0.579 for 1R, 0.487 for 2R, and 0.578 for 5R), which are slightly higher than the observed multivalent frequency for chromosomes 2R (0.442) and 5R (0.538) and obviously higher for chromosome 1R (0.379). The estimates of a', b', a and b, obtained using Eqs. (1)-(7) of Sybenga (1975b), vary between chromosomes, the most deviant esti- mates being those of chromosome 1R.
The expected configuration frequencies, obtained when introducing the estimates into Eqs. (1)-(6) to test such esti- mates, are shown in column 2 of Table 1. There are signifi- cant differences for chromosomes 2R and 5R. The lack of fit is obvious in classes ol and os for chromosome 2R, and cql and cqs for chromosome 5R. Observed values are higher in ol and cql and lower in os and cqs, respectively, than those expected. Pooling these classes gives expected
146
Table 1. Frequencies of the different metaphase I configuration classes (in parentheses the frequencies in terms of chromosomes involved) for chromosomes IR, 2R and 5R of autotetraploid rye. Column 1: observed. Column 2: expected on the basis of estimates for f, a', b', a and b as derived from the observations using Eqs. (t)-(7) of Sybenga (1975b). Column 3: expected on the basis of estimates for a', b', a and b as derived from the expressions g = f. a '+ ( 1 - f) 'a and b = f -b '+ ( 1 - f). b and those of Sybenga (1975 b)
Chromosome 1R Chromosome 2R Chromosome 5R
1 2 3 1 2 3 1 2 3
f 0.579 0.579 0.487 0.487 0.578 a' 0.428 0.429 0.704 0.702 0.748 b' 0.836 0.835 0.926 0.928 0.913 a 0.380 0.394 0.837 0.811 0.634 b 0.969 0.956 0.957 0.981 0.975 g 0.414 0.758 0.683
0.886 0.955 0.957
rq 37 (0.074) 36.92 37.00 103 (0.207) 103.07 102.93 135 (0.270) cql 95 (0.191) 98.67 98.49 86 (0.173) 86.67 87.39 103 (0.206) cqs 18 (0.036) 14.48 14.62 17 (0.034) 16.47 15.97 13 (0.026) t 39 (0.078) 38.71 38.93 14 (0.028) 13.85 13.56 18 (0,036) r 154 (0.155) 154.40 157.94 409 (0.411) 409.27 406.51 261 (0.261) ol 407 (0.409) 409.65 399.92 133 (0.134) 119.06 134.71 189 (0.189) os 14 (0.014) 11.58 14.00 6 (0.006) 20.25 9.64 5 (0.005) u 43 (0.043) 42.80 46.05 8 (0.008) 7.28 5.46 7 (0.007)
Z2(2) = 1.52 Z2(2) = 1.33 Z2(2) = 11175 Z2(2) = 2.70 P>0.30 P>0.50 P<O.01 P>0.20
0.578 0.728 0.941 0.621 0.979
134.79 135.63 90.82 101.35 25.69 17.01 17.31 12.71
260.86 256.56 184.10 196.82
9.96 6.97 7.88 6.28
Z2(2) = 10.63 Z2(2) =4.20 P<0.0I P>0.10
f= frequency of quadrivalent pairing; a' =frequency of short arm being bound after quadrivalent pairing; b ' = same for long arm; a = same for short arm after bivalent pairing; b = same for long arm after bivalent pairing
values almost equal to those observed. Differences indicated that b in chromosome 2R and b ' in 5R have been underesti- mated whereas a and a ' have been overestimated.
Another way to estimate a, b, a ' and b ' uses the rela- tions: f. a ' + (1 - f). a = ~ and f. b ' + (1 - f). b = b. Substitu- t ion of these expressions into the equations of Sybenga (1975b) gives
~- (1 -0 .a • -- (1 -- f). b 4rq
f f t+2cq+4rq
f i - ( 1 - f ) - a + b - ( 1 - f ) . b _ 2cq+8rq
f f t + 2cq + 4rq
from which a and b, and consequently a ' and b ' , can be solved.
This is p robably a more efficient approach since it in- cludes informat ion about ~ and b, which is very valuable for estimating a, a ' , b and b'. Resulting estimates and ex- pected frequencies are shown in column 3 of Table 1. There is a good fit in the three chromosomes sugges t ingthat these estimates are more representative.
Expected frequencies were also calculated under the fol- lowing assumptions: (i) completely random pairing, i.e. f = 2/3 and a, b, a ' and b ' as in column 3; (it) probabi l i ty of chiasmate association in the long arm is equal after quad- rivalent and bivalent pairing, i.e. b = b ' = b , and f, a ' and a as in column 3; and (iii) the same for the short arm, i.e. f i= a ' = a and f, b ' and b as in column 3. Significantly deviant values were found in all cases except for chromo- some I R in the third assumption. This means that : (i) f values are significantly lower than random pair ing values; (it) b ' is significantly lower than b; and (iii) a ' is significantly lower than a in chromosome 2R but higher in chromosome 5R.
Intersti t ial chiasma frequency in mult ivalent configura-
Table 2. Interstitial chiasma frequencies in multivalent configura- tions for the two arms of chromosomes 1R, 2R and 5R in autote- traploid rye
Chro- PMCs PMCs Minimum frequency mo- with with interstitial of two chiasmata some multi- chiasmata in arm with partner
valents exchange
Long Short Long Short arm arm arm arm
1R 236 76 15 0.161 0.032 (32.20) a (6.36)
2R 231 51 18 0.110 0.039 (22.08) (7.79)
5R 290 106 5 0.183 0.009 (36.55) (t.72)
a Parentheses enclose percentage of total number of PMCs with interstitial chiasmata
tions was higher in the long than in the short arm (Table 2), the difference varying with the differences in arm length. When a mult ivalent configurat ion showed intersti t ial chias- mate association, at least one arm of the chromosome in- volved must have formed two chiasmata (one in the intersti- tial segment and another in the pair ing segment). Thus, the min imum frequency with which two chiasmata are formed in a given chromosome arm equals half the fre- quency of multivalents with intersti t ial chiasmata in that arm. This value also appears in Table 2. In some of the arms (for example 5RL), when there was no evident par tner exchange, two chiasmata might have occured. However, this is difficult to establish in this type of preparat ion.

B |
o
, t I ,
..-Tq
-,,'-;71I' �9 i
�9 I
i
' b
o0 7q (7v
s @
,TI, -11'
Qs 2
-,,] I'
,TI,
Q s 3
,11 I !
o,311
�9 i
I
B 2 �9 I
i
OO 5A 5s"
' k /
Fig . 4a-e. The different associations of four homologous chromo- somes, two of which carry a marker telomeric C-band in the short arm, in a quadrivalent or in two bivalents and some of the resulting metaphase I configurations, a Homomorphic pairing, b Hetero- morphic pairing. There are two possibilities (2 and 3) for quadriva- lent pairing, e Some metaphase I configurations resulting from homomorphic pairing, d Some metaphase I configurations result- ing from heteromorphic pairing. The origin (Q2 or Q3) of the multivalent configurations cannot be determined using only this marker, e Metaphase I configurations that can derive from both homomorphic and heteromorphic pairing
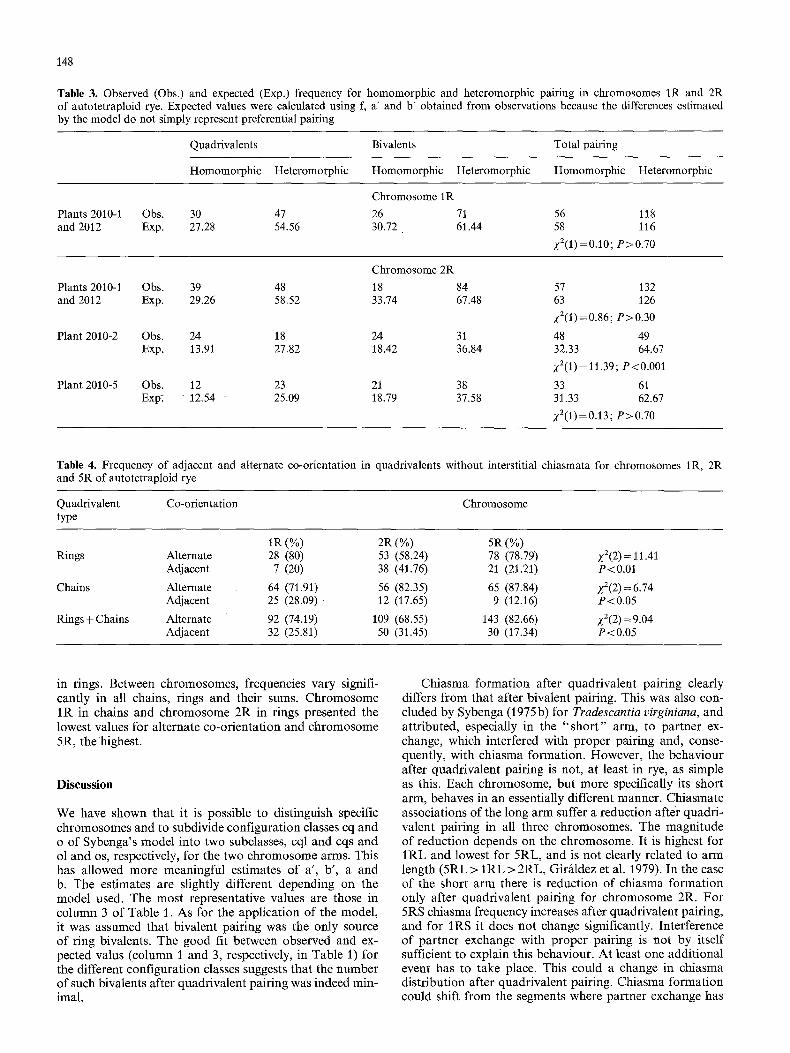
Another point of interest in the analysis of pairing is preferential association between particular chromosomes. We examined chromosome 1R in plants 2010-1 and 2012 and chromosome 2R in plants 2010-1, 2012, 2010-2 and 2010-5 for prefential association. In both cases only two chromosomes of the four homologues were marked with a prominent telomeric band (long arm for I R and short arm for 2R). Thus, two association types in quadrivalent and bivalent pairing could be distinguished: homomorphic, where the two marked chromosome arms pair with each other, and heteromorphic, where a marked arm pairs with an unmarked one. Quadrivalent and bivalent associations at pachytene belonging to each type are shown in Figure 4a and b. Two types of quadrivalent (2 and 3) are distin- guished with heteromorphic pairing. It is possible to esti- mate the frequencies of homomorphic and heteromorphic pairing from bivalent and multivalent frequencies at meta- phase I. Figure 4c gives representative configurations for homomorphic and Figure 4d, for heteromorphic pairing. Nevertheless, with only this marker it was not possible to ascertain if a given heteromorphic metaphase I multivalent (Fig. 4d) was derived from pachytene quadrivalent type
147
2 or 3. On the other hand configurations shown in Figure 4e, which could have formed after both homomorphic and heteromorphic pairing, were observed at a relatively low frequency (13% for chromosome IR and 4.52% for chro- mosome 2R), and were not considered in the analysis. Be- cause the multivalents that were included in classes ol and os of the model are among those of Figure 4e, configura- tions belonging to these classes with evidently homo- morphic or heteromorphic pairing consist only of open bi- valents.
Then, if there is no preferential association between chromosomes, and as the total expected frequencies for ho- momorphic and heteromorphic pairing are 1/3 and 2/3, re- spectively, metaphase I configurations are expected in the following frequencies:
homomorphic multivalents = 1/3f. a'. br. (a'. b ' - 2 a ' - 2b' + 4)
heteromorphic multivalents = 2/3f. a'- b ' . (a" b' - 2 a ' - 2b' + 4)
(the sum would equal rq + cql + cqs + t)
homomorphic bivalents = 1/3((1 - f ) + f" [1 - a ' .b ' . (a'. b ' - 2 a ' - 2 b ' +4)1 }
heteromorphic bivalents = 2/3{(1 - f) + f ' [ l - a ' . b ' - ( a ' - b ' - 2 a ' - 2b' + 4)]}
(the sum would equal r + ol + os + u)
The resulting expected values and those observed are given in Table 3. For chromosome 2R significant differ- ences between plants were found, and only two out of the four plants could be pooled. Estimates used for f, a ' and b' were not those of Table 1 but were calculated from the observations on the plant or plants analyzed.
In the case of chromosome 1R a slight prevalence ap- peared for homomorphic quadrivalents and heteromorphic bivalents. This behaviour might be a consequence of a slight preference for association between the short arms of a marked and an unmarked chromosome since the marked arm (the long one) paired at random (total frequencies for homomorphic and heteromorphic pairing are not signifi- cantly different from 1/3 and 2/3, respectively).
A similar but more evident preference is found for chro- mosome 2R in the two plants pooled. As in this chromo- some the short arm was marked and paired at random; preferential association took place between the long arms of one marked and one unmarked chromosome.
In plant 2010-2 the total homomorphic pairing fre- quency was significantly higher than 1/3 and, therefore, there was preferential association between the two marked short arms or the two unmarked ones.
The good fit in plant 2010-5 means there was no prefer- ence for pairing of chromosome 2R.
Co-orientation
In quadrivalents without interstitial chiasmata two types of centromere co-orientation, adjacent and alternate, could be distinguished. Frequencies with which they were ob- served in ring and chain quadrivalents are given in Table 4. Alternate co-orientation was the most frequent. Signifi- cant differences for chromosomes 1R and 5R were not found, but for chromosome 2R 0(1 z= 10.45) alternate co- orientation was more frequently observed in chains than
148
Table 3. Observed (Obs.) and expected (Exp.) frequency for homomorphic and heteromorphic pairing in chromosomes 1R and 2R of autotetraploid rye. Expected values were calculated using f, a' and b' obtained from observations because the differences estimated by the model do not simply represent preferential pairing
Quadrivalents Bivalents Total pairing
Homomorphic Heteromorphic Homomorphic Heteromorphic Homomorphic Heteromorphic
Chromosome 1R
Plants 2010-1 Obs. 30 47 26 71 56 118 and 2012 Exp. 27.28 54.56 30.72 61.44 58 116
2"2(1) = 0.10; P>0.70
Chromosome 2R
Plants 2010-1 Obs. 39 48 18 84 and 2012 Exp. 29.26 58.52 33.74 67.48
Plant 2010-2 Obs. 24 18 24 31 Exp. 13.91 27.82 18.42 36.84
Plant 2010-5 Obs. 12 23 21 38 Exp. 12.54 25.09 18.79 37.58
57 132 63 126 Z2(1) =0.86; P>0.30
48 49 32.33 64.67 X2(1) = 11.39; P<O.O01
33 61 31.33 62.67
2"2(I)=0.13; P>0.70
Table 4. Frequency of adjacent and alternate co-orientation in quadrivalents without interstitial chiasmata for chromosomes 1R, 2R and 5R of autotetraploid rye
Quadrivalent Co-orientation Chromosome type
1R (%) 2R (%) 5R (%) Rings Alternate 28 (80) 53 (58.24) 78 (78.79) Z2(2)=11.41
Adjacent 7 (20) 38 (41.76) 21 (21.21) P<0.01
Chains Alternate 64 (71.91) 56 (82.35) 65 (87.84) y2(2)=6.74 Adjacent 25 (28.09) 12 (17.65) 9 (12.16) P<0.05
Rings+Chains Alternate 92 (74.19) 109 (68.55) 143 (82.66) Z2(2)=9.04 Adjacent 32 (25.81) 50 (31.45) 30 (17.34) P<0.05
in rings. Between chromosomes, frequencies vary signifi- cantly in all chains, rings and their sums. Chromosome 1R in chains and chromosome 2R in rings presented the lowest values for alternate co-orientation and chromosome 5R, the highest.
Discussion
We have shown that it is possible to distinguish specific chromosomes and to subdivide configuration classes cq and o of Sybenga's model into two subclasses, cql and cqs and ol and os, respectively, for the two chromosome arms. This has allowed more meaningful estimates of a', b', a and b. The estimates are slightly different depending on the model used. The most representative values are those in column 3 of Table 1. As for the application of the model, it was assumed that bivalent pairing was the only source of ring bivalents. The good fit between observed and ex- pected valus (column 1 and 3, respectively, in Table 1) for the different configuration classes suggests that the number of such bivalents after quadrivalent pairing was indeed min- imal.
Chiasma formation after quadrivalent pairing clearly differs from that after bivalent pairing. This was also con- cluded by Sybenga (1975 b) for Tradescantia virginiana, and attributed, especially in the " s h o r t " arm, to partner ex- change, which interfered with proper pairing and, conse- quently, with chiasma formation. However, the behaviour after quadrivalent pairing is not, at least in rye, as simple as this. Each chromosome, but more specifically its short arm, behaves in an essentially different manner. Chiasmate associations of the long arm suffer a reduction after quadri- valent pairing in all three chromosomes. The magnitude of reduction depends on the chromosome. It is highest for 1RL and lowest for 5RL, and is not clearly related to arm length (5RL > 1RL > 2RL, Gir~ldez et al. 1979). In the case of the short arm there is reduction of chiasma formation only after quadrivalent pairing for chromosome 2R. For 5RS chiasma frequency increases after quadrivalent pairing, and for IRS it does not change significantly. Interference of partner exchange with proper pairing is not by itself sufficient to explain this behaviour. At least one additional event has to take place. This could a change in chiasma distribution after quadrivalent pairing. Chiasma formation could shift from the segments where partner exchange has

149
occurred towards segments that have paired, resulting in a notable increase of chiasmate associations in the latter. In chromosomes with quite different arm length, the long arm is most affected by interference of partner exchange. Then, the short arm might form more chiasmata than after bivalent pairing. This might be the case for chromosome 5R. Nevertheless, such an effect would decrease with the long arm short arm ratio. Thus, in almost metacentric chro- mosomes (2R) chiasma formation might be reduced in both arms after quadrivalent pairing. Chromosome 1R with an intermediate arm ratio presents an intermediate behaviour in the short arm: chiasma formation is almost equal after both bivalent and quadrivalent pairing.
Chiasma frequency was estimated earlier for arm I RL and arm 2RS in the diploid F 1 of inbred lines P and E by Gir/tldez and Orellana (1979) and Orellana and Girfildez (1981). F1 plants were heterozygous for telomeric bands in these two arms and the frequency of being bound at metaphase I and the frequency of recombined chromatids at anaphase I and metaphase II were estimated. Bound arm frequency in both chromosome arms was higher in diploids (0.998 for 1RL and 0.948 for 2RS) than in tetra- ploids (Table 1). However, the frequency with which two chiasmata were formed in the arm 1RL in diploids (0.172) was similar to that of tetraploids after quadrivalent pairing (minimum value 0.161, Table 2). For 2RS it was assumed to be 0 in diploids, and in tetraploids it showed a minimum value of 0.039 after quadrivalent pairing. This fact agrees with the assumed change in chiasma distribution after quadrivalent pairing.
The fact that the values of f are higher, especially for chromosome 1R, than the frequency of quadrivalents ob- served, indicates that a variable number of quadrivalents must have fallen apart because of lack of chiasmata. Yet, estimates for f do not reach the expected random pairing frequency (2/3) in any chromosome. This suggests that there is a tendency for the homologues to form bivalents at pa- chytene, as has already been reported by Timmis and Rees (1971). An excess of bivalents has also been found in other material. John and Henderson (1962) made a comparative study of multivalents and chiasma formation at diplotene in polyploid cells of several species of Acrididae and con- cluded that the number of quadrivalents formed by each chromosome of the complement is predominantly a func- tion of the relative chromosome length and the diploid chiasma frequencies of the bivalent concerned; short chro- mosomes rarely form quadrivalents.
Avivi (1976a) worked with Triticum longissimum, a wild diploid species with genotypes that are able to promote homoeologous pairing (intermediate pairing lines) and genotypes that have no such effect or suppress pairing (low pairing lines). Autotetraploids induced in both lines did not differ in the number of arms associated at metaphase I, but differed significantly in multivalent frequency. In in- termediate pairing lines the multivalent frequency corre- sponded with random pairing, whereas in low pairing lines multivalent frequency was significantly lower. It was sug- gested that low pairing genotypes do not affect meiotic pair- ing per se, but modify the pattern of homologous associa- tion, the homologues being spatially separated in premeiotic cells into two groups of two chromosomes. This assumption is supported by the results obtained on intermediate pairing lines of T. longissimum and the high pairing line of T. spel- toides after colchicine treatment of premeiotic cells (Avivi
1976b). A phenocopying effect by colchicine on the low pairing genes was found. When colchicine induced tetra- ploidy, bivalent formation would occur mostly between the newly replicated sister chromosomes because of their close proximity.
Another possible explanation for the excess of bivalents in autotetraploids is the existence of preferential pairing between two specific chromosomes. Preferential pairing has been detected in polyploids and aneuploids (Rhoades 1952; Doyle 1963, 1969; Snope 1967; Elci and Sybenga 1976; Gir~tldez and Santos 1981 ; Santos et al. 1983). In our mate- rial preferential pairing was indeed observed for chromo- somes 2R (Table 3). In two plants (2010-1 and 2012) this concerned the long but not the short arm of marked and unmarked chromosomes. In plant 2010-2 the short but not the long arm of the two marked and the two unmarked chromosomes was affected. There was, in addition, a fourth plant (2010-5) without pairing preferences. (For chromo- some 1R a slight preference among the long but not the short arms of marked and unmarked chromosomes might exist, although the results are not conclusive.) Whether or not preferential pairing exists, total multivalent and bivalent metaphase I configuration frequencies do not vary signifi- cantly between plants (Table 3). Preferential pairing in only one chromosome arm (short or long) is not enough to modi- fy frequencies of bivalent and quadrivalent pairing. These frequencies will be changed only when preferential pairing occurs in the two arms (short and long) of the same two chromosomes. As rye and T. longissimum are related spe- cies, spatial separation in some premeiotic cells of the four homologues, as suggested by Avivi (1976a, b), seems a real- istic cause of the excess of bivalents in autotetraploid rye. Preferential pairing would occur in meiotic cells where the four homologues remain relatively close to each other.
An attractive alternative explanation is the correction of the synaptonemal complex observed in triploid and tetra- ploid Bombyx oocytes (Rasmussen 1977; Rasmussen and Holm 1979, 1980) or as suggested for Triticum aestivum (Hobolth 1981). Without further analysis it is not possible to decide which of the two explanations is valid, or if per- haps both contribute in sequence.
Preferential pairing may be assumed to result from vari- ation in differences between the chromosome arms or the whole chromosomes involved. In our material this hypothe- sis cannot be tested. The constitution of the chromosomes is unknown because of the possibility of recombination be- tween homologous but not identical chromosomes derived from lines P and E in the meiosis of their F 1 (Fig. 2). Observations of Gir~ldez and Santos (1981) in a spontane- ous tetraploid/diploid chimaera involving meiotic cells of a male individual of Eucorthippus pulvinatus heterozygous for a C-heterochromatin segment in pair 9 confirm a prefer- ence for meiotic pairing between identical rather than ho- mologous chromosomes, but here the spatial distribution resulting from late doubling may be involved. Similar re- sults had been obtained for chromosome 11 in Eryprepocne- mis plorans by Santos et al. (1983). However, this was not so for the pairing of chromosome arms IRL and 2RS of rye, also analyzed by these authors. They found both a preference for homologous rather than identical pairing and random pairing between the four homologous chromo- somes. They concluded that pairing preferences do not de- pend exclusively on the similarities between chromosomes involved and suggest that genotypic or cryptic structural

150
differences between the homologous chromosomes of each pair might be responsible for some of the pairing prefer- ences found.
The analysis of centromere co-orientation in multiva- lents has usually been carried out in interchange heterozy- gores where the point of partner exchange is fixed. In poly- somics and autopolyploids the point o f partner exchange is variable, and the study of co-orientation becomes more difficult. Nevertheless, some conclusions can be derived from our results. As in interchange heterozygotes (Sybenga 1975 a), a predominance of alternate over adjacent co-orien- tation was observed in all three chromosomes (Table 4). Chains and rings showed similar frequencies for alternate and adjacent co-orientation in chromosomes 1R and 5R but not in chromosome 2R where alternate co-orientation was more frequent in chains. Variable behaviour for chain and ring quadrivalents has also been reported in inter- change heterozygotes. Sybenga (1968) showed that chains tend to have a lower frequency of alternate co-orientation than rings, whereas Naranjo and Lacadena (1979) observed alternate co-orientation more frequently in chains than in rings. In the same manner that different translocation heter- ozygotes show different behaviour, alternate and adjacent co-orientation frequencies differ between chromosomes in autotetraploids. The more metacentric the chromosome is, the higher is the adjacent co-orientation frequency. The rel- ative position of centromeres in the quadrivalent seems to play an important role in the co-orientation. Distance be- tween two centromeres at metaphase I had also been shown to be closely related to their co-orientation behaviour in translocation heterozygotes (Arana et al. 1980, 1982; Vos- selman 1981). Probably adjacent co-orientation would be- come more stable in ring quadrivalents with similar dis- tances between the four centromeres, as this quadrivalent type is expected to be formed more frequently by metacent- ric chromosomes.
Acknowledgements. The authors thank E. Perera for her valuable technical assistance.
References
Arana P, Santos JL, Giraldez R (1980) Chiasma interference and centromere co-orientation in a spontaneous translocation het- erozygote of Eucorthippus pulvinatus gallicus (Acrididae; Or- thoptera). Chromosoma 78 : 322340
Arana P, Santos JL, Henriques-Gil N, Gir/tldez R (1982) Centro- mere co-orientation in a spontaneous translocation heterozy- gote of Euchorthippus pulvinatus gallicus (Acrididae, Orthop- tera). Genetica 58:81-84
Avivi L (1976a) The effect of genes controlling different degrees of homoeologous pairing on quadrivalent frequency in induced autotetraploid lines of Triticum longissimum. Can J Genet Cy- tol 18:357 364
Avivi L (1976b) Colchicine-induced bivalent pairing of tetraploid microsporocytes in Triticum longissimum and T. speltoides. Can J Genet Cytol 18:731-738
Doyle GG (1963) Preferential pairing in structural heterozygotes of Zea mays. Genetics 48 : 1011-1027
Doyle GG (1969) Preferential pairing in trisomics of Zea mays. Chromosomes Today 2: 12-20
Elci S, Sybenga J (1976) Incomplete preferential pairing in a tetra- ploid Secale hybrid carrying translocations : multivalent config- uration frequencies and marker segregation. Genetica 46:177-182
Gilr/ddez R, Orellana J (1979) Metaphase I bonds, crossing-over frequency and genetic length of specific chromosome arms of rye. Chromosoma 72:377 385
Girfildez R, Santos JL (1981) Cytological evidence for preferences of identical over homologous but not-identical meiotic pairing. Chromosome 82: 447-451
Gir~ldez R, Cermefio MC, Orellana J (1979) Comparison of C- banding pattern in the chromosomes of inbred lines and open pollinated varieties of rye. Z Pflanzenz 83 : 40-48
Hobolth P (1981) Chromosome pairing in allohexaploid wheat vat. Chinese Spring. Transformation of multivalents into bivalents, a mechanism for exclusive bivalent formation. Carlsberg Res Commun 46:129-173
Jackson RC, Casey J (1980) Cytogenetics of polyploids. In: Lewis WH (ed) Polyploidy. Biological relevance. Plenum Press, New York London, pp 17-44
John B, Henderson SA (1962) Asynapsis and polyploidy in Schisto- cerca paranensis. Chromosoma 13 : 111-147
Naranjo T, Lacadena JR (1979) Analysis of centromere co-orienta- tion in a rye-wheat derivative by means of C-banding. Chromo- soma 73 : 227-235
Orellana J, Girfildez, R (1981) Metaphase I bound arms and cross- ing-over frequency in rye. I. Open pollinated varieties. Chromo- soma 84 : 439-449
Rasmussen SW (1977) Chromosome pairing in triploid females of Bombyx mori analyzed by three-dimensional reconstructions of synaptonemal complexes. Carlsberg Res Commun 42:163-197
Rasmussen SW, Holm PB (1979) Chromosome pairing in autote- traploid Bombyx females. Mechanism for exclusive bivalent formation. Carlsberg Res Commun 44:101-125
Rasmussen SW, Holm PB (1980) Mechanics of meiosis. Hereditas 93:187-216
Rhoades MM (1952) Preferential segregation in maize. In: Cowen JW (ed) Heterosis. Iowa State College Press, Ames, pp 66-80
Santos JL, Orellana J, Gir~ldez R (1983) Pairing competition be- tween identical and homologous chromosomes in rye and grass- hoppers. Genetics 104: 677-684
Snope AJ (1967) Meiotic behaviour in autotetraploid maize with abnormal chromosome 10. J Heredity 58:173-177
Sybenga J (1968) Orientation of interchange multiples in Secale cereale. Heredity 23: 73-79
Sybenga J (1975a) Meiotic configurations. Monographs on theo- retical and applied genetics, vol. 1. Springer, Berlin Heidelberg New York
Sybenga J (1975 b) The quantitative analysis of chromosome pair- ing and chiasma formation based on the relative frequencies of metaphase I configurations. VII. Autotetraploids. Chromo- soma 50: 211-222
Sybenga J (1983) Rye chromosome nomenclature and homoeolog2r relationships. Workshop report. Z Pflanzenz 90:297-304
Timmis JN, Rees H (1971) A pairing restriction at pachyteue upon multivalent formation in autotetraploids. Heredity 26:269-275
Vosselman L (1981) Meiotic segregation of five different reciprocal translocations in the onion fly Hylemya antiqna (Meigen). Chromosoma 81:727-738
Received May 31, 1983 / in revised form September 13, 1983 Accepted by J. Sybenga




















