
Media Konservasi Vol. XII, No. 2 Agustus 2007
42
Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66 57 KEANEKARAGAMAN ARTHROPODA TANAH DI HUTAN PENDIDIKAN GUNUNG WALAT (Diversity of Soil Arthropods in Gunung Walat Education Forest) LAILAN SYAUFINA 1 , NOOR FARIKHAH HANEDA 1 DAN ASRI BULIYANSIH 2 1 Departemen Silvikultur, Fakultas Kehutanan IPB 2 Departemen Manajemen Hutan, Fakultas Kehutanan IPB Diterima 15 Januari 2007 / Disetujui 18 Juni 2007 ABSTRACT A study was conducted to investigate the difference in abundance, richness, diversity and evenness of soil arthropods in Gunung Walat Education Forest, Sukabumi, West Java. Sampling was done in two difference habitats, e.g. burn and unburn habitat. A total of 1280 individuals from 40 families and 16 orders were collected by soil and litter extracting (Berlesse funnel). The biodiversity indices indicated that unburn habitat had a higher indices than that of burn habitat. Keywords: Diversity, Berlesse funnel, Forest Health Monitoring, unburn, soil arthropods PENDAHULUAN Penggunaan api secara intensif terutama dalam kegiatan konversi lahan menyebabkan semakin meluasnya hutan dan lahan kebakaran yang terjadi di Indonesia. Sejak November 1982-April 1983, Agustus 1990, Juni-Oktober 1991, Agustus-Oktober 1994 dan September-November 1997 (Februari-Mei 1998) kebakaran hutan dan lahan semakin luas dan menyebar. Kebakaran hutan dan lahan menyebabkan dampak seperti gangguan kesehatan, tercemarnya lingkungan, terganggunya aktivitas ekonomi serta hilangnya keanekaragaman hayati. Salah satu kerusakan yang terjadi adalah hilangnya biodiversiti dan rusaknya habitat makhluk hidup. Kerusakan tanah merupakan contoh kerusakan habitat yang terjadi. Hal ini berdampak pada keberadaan makhluk hidup di permukaan ataupun di bawah permukaan tanah. Kebakaran permukaan menyebabkan hilangnya vegetasi yang menutupi tanah. Jika suhu yang dihasilkan tinggi atau cukup tinggi untuk memanaskan tanah sampai ke bawah permukaannya dalam jangka waktu tertentu maka akan sangat berpengaruh pada sifat biologi tanah khususnya organisme tanah seperti arthropoda tanah. Suhu kebakaran yang melebihi suhu letal arthropoda-arthropoda tanah tersebut akan menyebabkan kematian. Hal ini dapat menyebabkan berkurang atau bahkan menghilangkan jenis-jenis arthropoda tanah tertentu. Hilangnya arthropoda-arthropoda tanah tersebut akan sangat berpengaruh terhadap keseimbangan ekosistem. Manfaat arthropoda tanah, khususnya serangga-serangga seperti pendekomposisi bahan organik, berperan dalam siklus nitrogen termasuk mineralisasi, denitrifikasi dan fiksasi N serta pengambilan nutrien seperti simbiosis mikoriza dengan akar tumbuhan yang membantu pengambilan P dan nutrien yang lain (DeBano et al., 1998). Jika serangga-serangga tanah ini terganggu sehingga berkurang atau hilang maka manfaat-manfaatnya pun akan hilang dan akan berdampak terhadap vegetasi sendiri. Untuk mengetahui sejauh mana gangguan yang terjadi terhadap hutan maka perlu ada monitoring terhadap kesehatan hutan. Salah satu metode yang dapat digunakan adalah dengan Forest Health Monitoring ( FHM ) dengan kerusakan terhadap sifat biologi tanah (organisme- organisme tanah) sebagai salah satu indikator kesehatan hutan. Pemantauan kondisi hutan dengan metode FHM ini sangat penting untuk dilakukan terutama pada areal hutan dan lahan bekas terbakar. Pentingnya penelitian dengan metode FHM ini juga disebabkan oleh hutan-hutan Indonesia yang mempunyai kondisi ekosistem berbeda sehingga pemantauan tidak cukup dilakukan di satu daerah saja. Perbedaan ekosistem akan menyebabkan perbedaan kondisi alam, biodiversitas serta responnya terhadap kebakaran. Metode FHM ini diharapkan mampu memantau kondisi hutan dari waktu ke waktu. Penelitian ini bertujuan mempelajari pengaruh kebakaran hutan yang terjadi terhadap arthropoda tanah dan memperoleh informasi yang akurat tentang jenis-jenis arthropoda tanah yang hilang, berkurang atau bertambah di suatu areal akibat kebakaran sehingga dapat digunakan sebagai pertimbangan dalam pengelolaan hutan bekas terbakar tersebut.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Media Konservasi Vol. XII, No. 2 Agustus 2007

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66
57
KEANEKARAGAMAN ARTHROPODA TANAH DI HUTAN PENDIDIKAN GUNUNG WALAT
(Diversity of Soil Arthropods in Gunung Walat Education Forest)
LAILAN SYAUFINA1, NOOR FARIKHAH HANEDA1 DAN ASRI BULIYANSIH2
1Departemen Silvikultur, Fakultas Kehutanan IPB 2Departemen Manajemen Hutan, Fakultas Kehutanan IPB
Diterima 15 Januari 2007 / Disetujui 18 Juni 2007
ABSTRACT
A study was conducted to investigate the difference in abundance, richness, diversity and evenness of soil arthropods in Gunung Walat Education Forest, Sukabumi, West Java. Sampling was done in two difference habitats, e.g. burn and unburn habitat. A total of 1280 individuals from 40 families and 16 orders were collected by soil and litter extracting (Berlesse funnel). The biodiversity indices indicated that unburn habitat had a higher indices than that of burn habitat. Keywords: Diversity, Berlesse funnel, Forest Health Monitoring, unburn, soil arthropods
PENDAHULUAN
Penggunaan api secara intensif terutama dalam kegiatan konversi lahan menyebabkan semakin meluasnya hutan dan lahan kebakaran yang terjadi di Indonesia. Sejak November 1982-April 1983, Agustus 1990, Juni-Oktober 1991, Agustus-Oktober 1994 dan September-November 1997 (Februari-Mei 1998) kebakaran hutan dan lahan semakin luas dan menyebar. Kebakaran hutan dan lahan menyebabkan dampak seperti gangguan kesehatan, tercemarnya lingkungan, terganggunya aktivitas ekonomi serta hilangnya keanekaragaman hayati.
Salah satu kerusakan yang terjadi adalah hilangnya biodiversiti dan rusaknya habitat makhluk hidup. Kerusakan tanah merupakan contoh kerusakan habitat yang terjadi. Hal ini berdampak pada keberadaan makhluk hidup di permukaan ataupun di bawah permukaan tanah. Kebakaran permukaan menyebabkan hilangnya vegetasi yang menutupi tanah. Jika suhu yang dihasilkan tinggi atau cukup tinggi untuk memanaskan tanah sampai ke bawah permukaannya dalam jangka waktu tertentu maka akan sangat berpengaruh pada sifat biologi tanah khususnya organisme tanah seperti arthropoda tanah. Suhu kebakaran yang melebihi suhu letal arthropoda-arthropoda tanah tersebut akan menyebabkan kematian. Hal ini dapat menyebabkan berkurang atau bahkan menghilangkan jenis-jenis arthropoda tanah tertentu.
Hilangnya arthropoda-arthropoda tanah tersebut akan sangat berpengaruh terhadap keseimbangan ekosistem. Manfaat arthropoda tanah, khususnya serangga-serangga seperti pendekomposisi bahan organik, berperan dalam siklus nitrogen termasuk mineralisasi, denitrifikasi dan
fiksasi N serta pengambilan nutrien seperti simbiosis mikoriza dengan akar tumbuhan yang membantu pengambilan P dan nutrien yang lain (DeBano et al., 1998). Jika serangga-serangga tanah ini terganggu sehingga berkurang atau hilang maka manfaat-manfaatnya pun akan hilang dan akan berdampak terhadap vegetasi sendiri.
Untuk mengetahui sejauh mana gangguan yang terjadi terhadap hutan maka perlu ada monitoring terhadap kesehatan hutan. Salah satu metode yang dapat digunakan adalah dengan Forest Health Monitoring ( FHM ) dengan kerusakan terhadap sifat biologi tanah (organisme-organisme tanah) sebagai salah satu indikator kesehatan hutan. Pemantauan kondisi hutan dengan metode FHM ini sangat penting untuk dilakukan terutama pada areal hutan dan lahan bekas terbakar.
Pentingnya penelitian dengan metode FHM ini juga disebabkan oleh hutan-hutan Indonesia yang mempunyai kondisi ekosistem berbeda sehingga pemantauan tidak cukup dilakukan di satu daerah saja. Perbedaan ekosistem akan menyebabkan perbedaan kondisi alam, biodiversitas serta responnya terhadap kebakaran. Metode FHM ini diharapkan mampu memantau kondisi hutan dari waktu ke waktu.
Penelitian ini bertujuan mempelajari pengaruh kebakaran hutan yang terjadi terhadap arthropoda tanah dan memperoleh informasi yang akurat tentang jenis-jenis arthropoda tanah yang hilang, berkurang atau bertambah di suatu areal akibat kebakaran sehingga dapat digunakan sebagai pertimbangan dalam pengelolaan hutan bekas terbakar tersebut.

Keanekaragaman Anthropoda Tanah
58
METODE PENELITIAN
Penelitian dilakukan bulan Juli 2005 di Hutan Pendidikan Gunung Walat pada areal bekas terbakar dan areal tidak terbakar. Ekstraksi dan identifikasi arthropoda tanah dilakukan di Laboratorium Entomologi Hutan Fakultas Kehutanan IPB. Penentuan petak pengamatan
dalam Forest Health Monitoring ( FHM ) dilakukan dengan membuat desain plot sampling. Desain plot sampling yang digunakan dalam INDO-FHM disebut Desain Cluster Plot (Gambar 1) (USDA Forest Servis, 1997 dalam Supriyanto et al., 2001a, b).
Gambar 1. Bentuk klaster FHM
Binatang arthropoda tanah yang dijumpai dan yang
dapat ditangkap dimasukkan ke dalam tabung-tabung plastik berisi alkohol 70% yang telah diberi label berdasarkan petak pengamatan dan tingkat kedalamannya. Arthropoda tanah yang tidak dapat ditangkap dilakukan ekstraksi tanah dengan menggunakan metode corong Barlese-Tullgren. Untuk mengidentifikasi arthropoda tanah digunakan beberapa buku seperti Pengenalan Pelajaran Serangga (Borror et al., 1996), Acarology, Mites And Human Welfare (Woolley, 1988) dan The pests of Crops In Indonesia (Kalshoven,1981). Pengamatan dilakukan di areal yang tidak terbakar dan satu tahun setelah terbakar.
Analisis keragaman, kekayaan dan kemerataan jenis pada lahan yang terbakar dan lahan yang tidak terbakar dengan menggunakan indeks kekayaan jenis Margalef (Richness Index), indeks keragaman Shannon-Wiener (Diversity Index) dan indeks kemerataan Pielou (Evenness Index) (Magurran, 1988). Uji t digunakan untuk membedakan variabel di plot yang terbakar dan tidak terbakar.
1
4 3
Annular plot Jari-jari 58.9’
(17.95 m)
Microplot Jari-jari 6.8’ (2.07 m) azimuth 900 dari
titik pusat subplot (3.66m)
Subplot
Jari-jari 24.0’ (7.32 m)
Azimuth 1-2 3600
Azimuth 1-3 1200 Azimuth 1-4 2400
Jarak antara tiap titik pusat plot : 120` (36,6 m)
2
Letak Petak Pengamatan Makrofauna Tanah

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66
59
HASIL DAN PEMBAHASAN
A. Kondisi Umum Lokasi Penelitian
Hutan Pendidikan Gunung Walat (HPGW) terletak sekitar 2,5 km ke arah selatan dari jalan Bogor-Sukabumi yang berjarak 55 km dari Bogor dan 15 km dari Sukabumi. Secara geografis kawasan ini terletak pada 060 53’35”-060 55’10” Lintang Selatan (LS) dan 106047’50”-106051’30” Bujur Timur (BT) dengan rata-rata ketinggian 557 meter di atas permukaan laut (m dpl) dengan luasan semula adalah 359 ha dan kini menjadi sekitar 349 ha. HPGW terletak pada ketinggian 500-700m dpl dengan topografi yang bervariasi dari landai sampai bergelombang terutama di bagian selatan. Di bagian utara memiliki kondisi topografi yang semakin berat. Kondisi topografi di kawasan HPGW ini adalah bergunung (98 ha), berbukit (42 ha), bergelombang (23 ha), berombak (9 ha) dan datar (4 ha).
Kondisi tanah di kawasan HPGW umumnya terdiri dari jenis tanah yang kompleks, diantaranya podsolik merah kuning, latosol dan litosol dari batuan endapan dan beku di daerah bukit. Berdasarkan data curah hujan Gunung Walat tahun 2000 – 2005 dari Laboratorium Pengaruh hutan Fakultas Kehutanan IPB diperoleh curah hujan rata-rata sebesar 2272.79 ml/bulan dengan rata-rata hari hujan 10 hari/bulan. Curah hujan rata-rata terendah terjadi pada tahun 2001 (1797.67 ml/bulan).
Kondisi penutupan lahan oleh vegetasi di kawasan HPGW sekitar 75 % adalah hutan tanaman yang ditanam sejak tahun 1958 dengan dominasi jenis damar (Agathis dammara), tusam-pinus (Pinus merkusii) dan mahoni (Swietenia macrophylla). Tumbuhan bawah yang dominan antara lain alang-alang (Imperata cylindrica), harendong (Melastoma malabathricum), pakis areuy (Nekania scanden), pakis rane (Selaginella plana) (Rahardjo, 2003). B. Sejarah Kebakaran
Kebakaran merupakan peristiwa yang telah berulang kali terjadi di HPGW. Berdasarkan buku catatan, HPGW pernah terjadi enam kali peristiwa kebakaran hutan selama tahun 2002. Kebakaran terjadi pada bulan Agustus, September dan Oktober pada lokasi dan waktu yang berbeda. Kebakaran terbesar terjadi pada jalur Cimenyan di tegakan Pinus pada tanggal 17 September 2002. Tipe kebakaran yang terjadi adalah kebakaran permukaan (surface fire) dan kebakaran tajuk (crown fire). Pada kebakaran ini api membakar bahan bakar yang ada di
permukaan seperti serasah, ranting-ranting pohon yang jatuh, batang pohon dan tajuk pohon.
Pada bulan Agustus dan September 2002 hanya ada satu hari hujan. Rendahnya curah hujan menyebabkan pengeringan bahan bakar (Pinus) sehingga kebakaran lebih mudah terjadi. Selain itu, kandungan resin pada Pinus membuat api lebih mudah menyala dan sulit untuk dipadamkan. Perubahan arah angin menyebabkan api menjalar ke arah tegakan yang belum terbakar. Angin merupakan pensuplai oksigen untuk pembakaran dan mempercepat pengeringan bahan bakar, sehingga kebakaran menjadi besar dan lebih sulit dikendalikan. Luasan lahan yang terbakar adalah ± 30 ha dan saat ini kondisi tegakan bekas terbakar cukup baik.
C. Fire Severity
Menurut DeBano et al. (1998) fire Severity adalah suatu penilaian yang menggambarkan respon ekosistem terhadap kebakaran, dapat digunakan untuk mendeskripsi-kan efek kebakaran pada sistem air dan tanah, ekosistem flora dan fauna, atmosfer dan manusia. Fire severity sangat tergantung pada bahan bakar alami yang tersedia dan perilaku api saat bahan bakar dibakar.
Berdasarkan kondisi tegakan saat ini maka kebakaran yang terjadi tahun 2002 tersebut termasuk low fire severity. Hal ini dibuktikan dengan kondisi tegakan dimana > 50 % pohon-pohonnya mengalami kerusakan yang tidak terlihat dan > 80% pohon-pohon yang rusak/terbakar dapat bertahan. Selain itu warna permukaan tanah pada plot terbakar sedikit lebih hitam dibandingkan plot yang tidak terbakar dengan kedalaman 0-5 cm. Kondisi lokasi juga menunjukkan bahwa kebakaran yang terjadi adalah low severity burn dimana < 2% areal terbakar berat, <15% areal terbakar sedang dan sisanya terbakar ringan atau tidak terbakar. D. Dampak Kebakaran terhadap Arthropoda Tanah
Keberadaan arthropoda tanah pada suatu habitat sangat dipengaruhi oleh kondisi habitat tersebut. Arthropoda tanah akan melimpah pada habitat yang mampu menyediakan faktor-faktor yang dapat mendukung kehidupan arthropoda tanah seperti ketersediaan makanan, suhu yang optimal, dan ada atau tidaknya musuh alami. Beberapa parameter yang dapat mempengaruhi keberadaan arthropoda tanah pada lokasi penelitian dapat dilihat pada Tabel 1.

Keanekaragaman Anthropoda Tanah
60
Tabel 1. Parameter yang mempengaruhi Arthropoda tanah
Parameter Mikro Habitat Terbakar Tidak Terbakar
Nitrogen rata-rata (%)* 0.11 0.08 Suhu serasah rata-rata (0C) 27.93 28.24 Tebal serasah rata-rata (cm) 3.89 2.11 Tumbuhan bawah Clidemia hirta, Patorium inufolium, Leucos
lavandulaefolia, Dimeria ornithopoda Curculigo latifolia Bukaan tajuk (%)** 44.38 21.43 Keterangan : * Priandi (2005) ** Wahyuningriyanti (2005) 1. Kelimpahan ArthropodaTanah
Kelimpahan arthropoda tanah mengacu pada jumlah total individu arthropoda tanah yang ditemukan pada lokasi penelitian yaitu pada plot terbakar dan plot yang tidak terbakar. Untuk menggambarkan hasil yang diperoleh dari kedua plot tersebut digunakan nilai rata-rata dari kelimpahan arthropoda tanah yang ada. a. Lapisan tanah (0-5 cm)
Kelimpahan arthropoda tanah sangat dipengaruhi oleh kondisi lingkungan sekitarnya sehingga faktor ini harus benar-benar diperhatikan. Perubahan pada lingkungan akan berdampak pada keberadaan arthropoda tanah baik secara langsung atau tidak langsung. Kondisi lingkungan pada plot
terbakar dan tidak terbakar di lokasi penelitian dapat digambarkan dengan parameter pada Tabel 1.
Tekstur tanah plot terbakar dan plot tidak terbakar adalah sama yaitu lempung liat berdebu, namun permukaan tanah pada plot tidak terbakar terdapat pasir-pasir halus. Tanah pada plot tidak terbakar ini juga lebih padat dan lebih lembab dibandingkan dengan tanah pada plot terbakar. Kelembaban tanah ini mempengaruhi suhu tanah dimana suhu tanah rata-rata pada plot terbakar lebih tinggi dibandingkan dengan plot tidak terbakar. Sementara, kandungan nitrogen sebagai salah satu unsur yang penting bagi makhluk hidup lebih tinggi pada plot terbakar.
Jumlah total individu yang ditemukan di tanah terbakar adalah 638 individu dan pada tanah tidak terbakar ditemukan 410 individu (Tabel 2).
Tabel 2. Kelimpahan Arthropoda tanah pada Plot Pengamatan
Ordo Famili Kelimpahan Plot Tidak terbakar Plot Terbakar
Araneida Araneidae 6 1 Oxyopidae 4 - Lycosidae 12 2 Salticidae 3 1 Tetragnathidae 1 1
Blattaria Blattidae 5 - Chelonethida - 44 1 Chilopoda Geophilidae 48 4 Coleoptera Oedemeridae 1 -
Tenebrionidae 1 - Staphylinidae 45 - Reduviidae 1 - Scydmaenidae 1 - Scarabaidae 44 -
Cryptostigmata Schlerobatidae 44 - Diplopoda Julidae 1 1
Polydesmidae 7 - Homoptera Cercopidae 3 - Hymenoptera Formicidae 69 513 Isopoda Trichoniscidae 36 13

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66
61
Tabel 2. (Lanjutan)
Ordo Famili Kelimpahan Plot Tidak terbakar Plot Terbakar
Isoptera Termitidae 1 12 Mantodea Mantidae 1 - Mesostigmata Tetranychidae 1 -
Laelapidae - 86 Orthoptera Acrididae 1 -
Grillidae 1 1 Jumlah Total 410 638
b. Lapisan Serasah
Pada Tabel 3 terlihat jumlah Collembola mendominasi pada plot terbakar dengan 112 individu karena serasahnya banyak. Pada plot tidak terbakar individu yang banyak
ditemukan adalah Collembola dan Acarina sejumlah 11 individu. Sementara kelimpahan ordo Coleoptera, Chilopoda, Araneida, Diplopoda, Thysanoptera dan Chelonethida adalah kecil.
Tabel 3. Kelimpahan Makrofauna Tanah pada Serasah
Ordo Family Kelimpahan Tidak Terbakar Terbakar
Araneida Thomisidae 1 1 Linyphiidae - 1
Chilopoda Geophilidae - 3 Coleoptera Nitidulidae - 1
Ellateridae 1 - Collembola Isotomidae 5 104
Podoridae - 4 Sminthuridae - 4
Chelonethida 1 - Cryptostigmata Zetorchestidae - 6
Schlerobatidae 4 15 Ephylohmaiidae - 6 Rhysotritiidae - 1
Diplopoda Polydesmidae - 1 Hymenoptera Formicidae 1 16 Mesostigmata Phytoseiidae - 4
Veigaiidae - 1 Laelapidae 5 22 Tetranychidae 2 -
Metastigmata Argasidae - 21 Thysanoptera Thripidae 1 - Total 21 211
Dari Tabel 3 ini dapat dilihat bahwa serasah pada plot terbakar lebih tebal dibandingkan serasah pada plot tidak terbakar. Ketebalan serasah ini berpengaruh terhadap jumlah serasah yang dapat terdekomposisi, semakin tebal serasah maka akan semakin banyak bahan organik yang dihasilkan. Bukaan tajuk pada plot terbakar yang lebih besar menyebabkan dekomposisi serasah pada lantai hutan lebih aktif dan cepat. Szujeki (1987) menyebutkan bahwa Colembolla merupakan invertebrata yang kelimpahannya
menunjukkan osilasi (kisaran) yang luas pada lahan terbakar.
Menurut Wallwork (1970), biasanya pada sebagian besar padang rumput, tanah mor dan tanah hutan, Collembola adalah fauna yang paling mewakili, dari segi jumlah individu dan spesies, pada lapisan organik di bagian atas 10-15 cm. Dalam skala kedalaman ini, populasi terbesar Colembola biasanya terdapat pada tingkat kedalaman dimana dekomposisi bahan organik secara aktif terjadi; pada profil tipe mor, tingkatan ini dikenal sebagai

Keanekaragaman Anthropoda Tanah
62
‘zona fermentasi’, yang berbeda dan terpisah dari lapisan litter dan lapisan humus.
Burgers dan Raw (1967) menyimpulkan, dari analisis usus Collembola bahwa jenis Collembola yang lebih besar memakan fungi, sebaliknya yang berukuran lebih kecil langsung memakan humus. Collembola juga mengkonsumsi bagian tanaman yang lapuk, spora, Collembola yang lain, bagian cacing tanah yang terdekomposisi dan kutikulanya sendiri. Sebagai dekomposer Collembola berperan menghancurkan feses arthropoda yang lebih besar, menghasilkan kitin agar tersedia di tanah dan memudahkan proses dekomposisi oleh dekomposer yang lain.
Sama seperti Collembola, Acarina juga merupakan mesofauna yang banyak ditemukan pada lapisan permukaan, lapisan fermentasi dan lapisan humus. Acarina mengkonsumsi tanaman yang lapuk, lumut, fungi dan alga. Acarina juga berperan sebagai dekomposer. Pada lahan hutan yang tidak kondusif bagi dekomposer yang lebih besar maka dekomposisi bagian tanaman dilakukan oleh Acarina. Ordo Cryptostigmata berperan dalam mencampurkan bahan organik pada lapisan tanah di bawah permukaan.
Berdasarkan hasil uji t, kelimpahan rata-rata dari tanah yang terbakar dengan tanah tidak terbakar tidak berbeda nyata (P < 0.205), demikian pula pada serasah yang terbakar dan serasah tidak terbakar ( P < 1.419 ). Hal ini menunjukkan bahwa perbedaan jumlah total individu yang ditemukan pada kedua plot tidak signifikan, artinya kebakaran yang terjadi memberikan dampak yang ringan.
Jumlah individu total di plot yang terbakar adalah 638 individu dan di plot tidak terbakar 410 individu. Meskipun dari hasil uji t kelimpahan pada kedua plot ini tidak berbeda nyata tetapi kecenderungan menunjukkan bahwa kelimpahan arthropoda tanah di plot terbakar lebih besar daripada kelimpahan arthropoda tanah di plot tidak terbakar.
2. Indeks Kekayaan (Richness Index) Arthropoda Tanah
Kekayaan jenis (Richness) arthropoda tanah mengacu pada banyaknya spesies yang ditemukan pada suatu ekosistem. Jumlah total spesies dalam suatu komunitas (S) tergantung pada ukuran sampel dan waktu. Beberapa indeks dianjurkan untuk mengukur spesies Richness yang tidak tergantung pada ukuran sampel tetapi berdasarkan hubungan antara S dan jumlah total individu yang diamati (N) yang meningkat dengan meningkatkan ukuran sampel (Ludwig dan Reynolds, 1988).
Richnesss Index yang digunakan dalam menganalisa kekayaan jenis arthropoda tanah pada penelitian ini adalah Margalef index karena kalkulasi atau perhitungannya lebih mudah. Kekayaan jenis arthropoda tanah ditunjukkan dalam banyaknya famili arthropoda tanah yang dapat ditemukan pada plot terbakar dan plot tidak terbakar. Nama famili dan jumlahnya dapat dilihat pada Tabel 2.
a. Pada Lapisan Tanah
Pada tanah tidak terbakar terdapat 14 ordo dengan 25 famili, sementara pada tanah terbakar terdapat 9 ordo dengan 12 famili. Perbedaan ini cukup besar dimana terdapat selisih 5 ordo dan 13 famili yang merupakan suatu lingkup taksonomi yang besar (Tabel 4).
Ordo yang terdapat pada tanah tidak terbakar tetapi tidak pada tanah terbakar adalah Blattaria, Coleoptera, Cryptostigmata, Diplopoda, Mantodea dan Homoptera. Sedangkan famili-famili yang tidak terdapat di tanah terbakar adalah Polydesmidae, Julidae, Mantidae, dan Cercopidae. Sebagian besar famili yang tidak ada adalah dari ordo Coleoptera, famili Mantidae dan Cercopidae dimana famili-famili ini adalah pemakan tumbuhan ataupun memakan bagian dari tumbuhan. Kebakaran menyebabkan pengurangan dan hilangnya stok makan mereka sehingga dapat mengganggu perkembangbiakan dan keberadaannya. Selain itu, kebakaran juga dapat mematikan larva serangga-serangga ini yang umumnya terdapat pada permukaan tanah.
Tabel 4. Keanekaragaman arhropoda tanah di Hutan Pendidikan Gunung Walat, Sukabumi
Lapisan Tanah Lapisan Serasah
Tidak terbakar Terbakar Tidak Terbakar Terbakar ∑ Ordo 14 9 8 9
∑ Famili 25 12 9 17 ∑ Individu 410 638 21 211 Richness 4.039 1.704 2.627 2.989
Diversity (H’) 2.394 0.709 1.948 1.838 Evenness 0.802 0.479 0.873 0.501

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66
63
Famili Blattidae, Polydesmidae dan Julidae merupakan famili yang sebagian besar hidup di permukaan tanah ataupun di dalam tanah yang dekat ke permukaan. Saat terjadinya kebakaran maka binatang yang ada di permukaan tanah ataupun yang dekat dengan permukaan tanah akan sangat mudah terkena dampak kebakaran walaupun kebakaran yang terjadi adalah low severity burn. Dampak kebakaran ini bisa berupa pengurangan jumlah arthropoda tanah maupun hilangnya famili atau jenis arthropoda tanah tertentu.
b. Pada Lapisan Serasah
Pada serasah di plot terbakar terdapat 9 ordo dengan 17 famili, sementara pada serasah di plot yang tidak terbakar terdapat 8 ordo dengan 9 famili (Tabel 4). Ordo-ordo yang tidak terdapat pada serasah tidak terbakar adalah Chilopoda dan Diplopoda. Sementara famili-famili yang tidak ada pada serasah tidak terbakar adalah Linyphiidae, Geophilidae, Nitidulidae, Podoridae, Sminthuridae, Zetorchestidae, Ephylohmaiidae, Rhysotriidae, Polydesmidae, Phytoseiidae, Veigaiidae dan Argasidae. Sebagian besar famili yang tidak terdapat pada serasah tidak terbakar merupakan Acarina. Banyaknya jumlah famili Acarina yang ditemukan menyebabkan serasah yang terbakar mempunyai famili-famili yang lebih beragam dibandingkan dengan serasah yang tidak terbakar.
Dalam Wallwork (1976) disebutkan bahwa Acarina adalah yang paling umum meskipun bukan yang paling mewakili pada sebagian besar tanah, dan banyak terdapat pada tanah organik tinggi dan lahan hutan. Pada kebanyakan tipe hutan khususnya konifer, bagian tumbuhan yang hancur membentuk lapisan yang terkadang satu kaki atau lebih dalam Brown dan Davis (1973). Kebakaran menyebabkan terbukanya akses ke lantai hutan sehingga dekomposisi bahan organik menjadi lebih cepat. Kondisi ini menyebabkan tersedianya habitat yang baik untuk perkembangan Acarina setelah kebakaran.
Richness Index rata-rata pada tanah terbakar dan tanah tidak terbakar memperoleh nilai P < 0.046, artinya kedua habitat tersebut mempunyai Richness Index yang berbeda nyata. Nilai Richness Index rata-rata pada serasah terbakar dan serasah tidak terbakar tidak menunjukkan perbedaan yang nyata. dengan nilai probability (P) sebesar 0.620.
3. Indeks Keragaman (Diversity Index) Arthropoda
tanah
Diversity index menyatukan spesies richness dan evenness dalam satu nilai (Ludwig dan Reynolds, 1988). Diversity index yang digunakan dalam analisis data adalah Diversity index Shannon –Winner (H’). Indeks ini mempunyai dua properti yang membuat indeks ini banyak digunakan untuk keragaman spesies, (1) H’=0 jika hanya jika ada satu spesies dalam sampel, (2) H’ maksimum hanya
saat (jumlah total spesies dalam komunitas/S) semua spesies diwakili oleh jumlah individu yang sama, yang merupakan distribusi kelimpahan yang sempurna (Ludwig dan Reynolds, 1988). a. Pada Lapisan Tanah
Pada tanah terbakar, nilai keragaman rata-ratanya adalah 0.709 keragamannya rendah (Tabel 4). Hal ini dapat terlihat dari sebaran kelimpahan jumlah individu pada setiap famili yang tidak merata. Ada beberapa famili yang mempunyai kelimpahan yang besar yaitu Formicidae dan Laelapidae.
Nilai keragaman rata-rata pada tanah tidak terbakar adalah 2.394, keragamannya lebih tinggi daripada tanah yang terbakar. Famili yang banyak ditemukan pada tanah tidak terbakar adalah Formicidae, Geophilidae, Staphylinidae, Scarabaidae, Chelonethida, Schlerobatidae, dan Trichoniscidae. Sementara, famili yang jarang ditemukan berjumlah 17 famili diantaranya Araneidae, Oxyopidae, Salticidae, Tetragnathidae, Blattidae, Oedemeridae, Tenebrionidae, Reduviidae, Scydmaenidae, Julidae, Termitidae, Mantidae dan Laelapidae.
Pada tanah yang terbakar Formicidae (semut) mempunyai kelimpahan terbesar sehingga kebakaran dapat meningkatkan populasi semut. Suatu studi di Australia terhadap vegetasi Sclerophyllous menunjukkan bahwa semut mengkonsumsi biji yang banyak dihasilkan pada awal suksesi tanaman setelah kebakaran (Andersen, 1988). Pinus merupakan salah satu spesies fire yang berkembang lebih baik setelah kebakaran. Di lokasi penelitian, biji pinus lebih banyak ditemukan pada areal yang terbakar dibandingkan areal yang tidak terbakar sehingga areal yang terbakar menjadi tempat yang lebih baik untuk perkembangan semut.
b. Pada Lapisan Serasah
Nilai keragaman rata-rata pada serasah terbakar adalah 1.838 dan pada serasah tidak terbakar bernilai 1.948 (Tabel 4). Family Isotomidae (ordo Collembola) merupakan famili yang paling banyak ditemukan pada serasah bekas terbakar. Dalam Wallwork (1970) disebutkan bahwa Collembola biasanya melimpah pada tanah di padang rumput dan tanah mor seperti pada lantai hutan dengan skala kepadatan 5000-50000/m2, terkadang lebih besar. Collembola merupakan invertebrata yang mempunyai penyebaran luas karena dapat tersebar terbawa angin, aliran air ataupun menempel pada kaki burung. Setelah terjadi kebakaran terdapat akumulasi bahan organik di permukaan tanah, sehingga jika ada Colembolla yang sampai di daerah tersebut akan mudah berkembang karena salah satu makanan alaminya adalah bagian tumbuhan yang melapuk.
Berdasarkan hasil uji t diketahui bahwa keragaman binatang pada lapisan tanah tidak berbeda nyata. Nilai

Keanekaragaman Anthropoda Tanah
64
keragaman rata-rata pada serasah terbakar dan serasah tidak terbakar juga tidak berbeda nyata. Hal ini menunjukkan bahwa perbedaan diversity atau keragaman famili arthropoda tanah pada plot terbakar dan plot tidak terbakar adalah tidak signifikan, meskipun nilai Diversity Index pada plot tidak terbakar cenderung lebih besar. 4. Indeks Kemerataan (Evenness Index) Arthropoda
Ludwig dan Reynolds (1988) mengatakan bahwa evenness (kemerataan) mengacu pada bagaimana kelimpahan spesies (jumlah individu, biomass, penutupan dan lain-lain) didistribusikan diantara spesies. Evenness (kemerataan ) ini merupakan salah satu komponen dari diversity. Nilai evenness index berkisar antara 0 - 1. Nilai satu menunjukkan semua jenis ada dalam kelimpahan yang sama (Magurran, 1988).
a. Pada Lapisan Tanah
Pada tanah tidak terbakar Formicidae berjumlah 16.83% dari total individu, kemudian Geophilidae (11.71%), Staphylinidae (10.97%), Scarabaidae dan Schlerobatidae sebesar 10.73% dan Trichonischidae (8.78%) dan famili lainnya sebesar 40.97%.
Sedangkan pada tanah terbakar famili Formicidae (semut) mempunyai jumlah individu terbanyak yaitu 80.41% dari jumlah total individu yang ada, kemudian famili Laelapidae (13.48%) dan 6.11% tersebar pada famili lainnya.
Famili-famili seperti Staphylinidae, Geophilidae dan Trichonischidae termasuk invertebrata permukaan yang kemungkinan berkurang atau hilang setelah terjadinya kebakaran. Selain itu hilangnya lapisan serasah menyebabkan berkurangnya stok makanan karena Trichonischidae termasuk ordo Isopoda yang makanannya adalah bahan organik yang mati atau lapuk. Sementara Geophilidae termasuk ordo Chilopoda yang dapat masuk ke dalam tanah beberapa inchi saja sehingga sangat rentan terhadap pemanasan tanah akibat kebakaran.
Famili-famili yang berasal dari ordo Coleoptera (Staphylinidae dan Scarabaidae) merupakan pemakan tumbuhan yang jumlahnya pasti berkurang setelah kebakaran karena hilangnya tumbuhan sebagai bahan makanan mereka dan matinya larva serangga ini yang terdapat di permukaan tanah.
Pada tanah yang terbakar Formicidae (semut) mempunyai jumlah individu yang besar. Besarnya jumlah semut ini terkait dengan produksi biji tanaman setelah terbakar. Pinus menghasilkan biji yang banyak setelah kebakaran sehingga bahan makanan bagi semut tersedia lebih banyak dibandingkan dengan biji yang dihasilkan pada tegakan yang tidak terbakar.
b. Pada Lapisan Serasah
Pada serasah di plot terbakar famili Isotomidae (Collembola) mempunyai jumlah individu 49.29% dari jumlah total individu yang ditemukan, kemudian Laelapidae (10,43%), Argasidae (9.95%), Schlerobatidae dan Formicidae (± 7.66%) dan 22.00% individu dari famili lainnya. Dari sebaran persentase jumlah individu pada setiap famili terlihat bahwa sebaran kelimpahan pada setiap famili tidak merata.
Banyaknya jumlah Collembola pada serasah plot terbakar dipengaruhi oleh kondisi tegakan setelah kebakaran. Bukaan tajuk pada plot terbakar ini sebesar 44.34%, lebih besar dibandingkan dengan plot tidak terbakar yang hanya 21.43% (Tabel 1). Terbukanya tajuk menyebabkan akses yang besar ke lantai hutan seperti cahaya matahari dan hujan, hal ini dapat mempercerpat proses dekomposisi tumbuhan atau bagian tumbuhan yang ada di lantai hutan seperti serasah sehingga bahan organik terbentuk lebih cepat.
Collembola merupakan invertebrata yang banyak tedapat pada lapisan tanah yang aktif terdekomposisi. Pada plot yang tidak terbakar, lapisan serasahnya lebih tipis dibandingkan dengan serasah pada plot yang terbakar karena sebagian serasah tertahan pada tumbuhan bawah sehingga tidak sampai ke tanah.
Pada serasah plot terbakar juga banyak terdapat Acarina. Eksperimen kultur menunjukkan bahwa macrochelids dan veigaiids dapat memakan beberapa varietas binatang, termasuk Collembola, Protura, Pauropoda, nematoda dan Enchytreids, dan telur Musca domestica (Wallwork, 1970). Dengan melimpahnya Collembola maka makanan bagi Acarina akan tersedia, sehingga populasinya dapat berkembang.
Nilai evenness index rata-rata pada tanah terbakar adalah 0.479 dan tanah tidak terbakar lebih besar yaitu sebesar 0.802 (Tabel 4). Nilai Evennes Index yang kurang dari 1 ini menunjukkan bahwa sebaran kelimpahan individu pada setiap famili tidak merata.
Nilai Evenness Index rata-rata pada serasah terbakar dan serasah yang tidak terbakar adalah 0.501 dan 0.873 (Tabel 4), dimana serasah yang tidak terbakar mempunyai Evenness Index rata-rata yang lebih tinggi. Nilai Evenness Index rata-rata sebesar 0.873, dimana nilai ini hampir mendekati 1, artinya, sebaran kelimpahan individu pada masing-masing famili hampir merata, jadi tidak ada famili yang jumlah individunya mendominasi.
Untuk mengetahui perpindahan arthropoda tanah secara vertikal maka dilakukan uji t antara lapisan tanah terbakar dengan serasah terbakar dan lapisan tanah tidak terbakar dengan serasah tidak terbakar. Dari uji t diperoleh nilai probabilitis untuk perbandingan serasah dan tanah pada plot terbakar sebesar 0.216, artinya kelimpahannya tidak berbeda nyata. Hal ini menunjukkan bahwa saat

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 57 – 66
65
kebakaran tidak terjadi perpindahan vertikal arthropoda tanah dari lapisan serasah ke dalam tanah.
Pada plot tidak terbakar, perbandingan antara serasah tidak terbakar dan tanah tidak terbakar mempunyai nilai probabilitis sebesar 0.016, artinya kelimpahannya berbeda nyata dimana arthropoda tanah yang terdapat pada plot tidak terbakar hidup pada habitatnya masing-masing (tanah atau serasah).
KESIMPULAN
1. Kebakaran yang terjadi di HPGW pada bulan September 2002 ini menyebabkan penurunan jumlah ordo dari 17 menjadi 10 ordo (41,18%), penurunan jumlah famili dari 31 menjadi 29 famili (6,45%), tetapi terjadi peningkatan jumlah individu dari 431 menjadi 849 individu (96,98%).
2. Kebakaran yang terjadi termasuk low fire severity dengan tipe kebakaran adalah kebakaran permukaan (surface fire) dan kebakaran tajuk (crown fire). Berdasarkan definisi hutan yang tidak sehat maka tegakan bekas terbakar ini masih dapat menjalankan fungsinya, walaupun ada perubahan dalam ekosistemnya tetapi tidak signifikan
3. Nilai kelimpahan, Richness, Diversity dan Evenness indices rata-rata pada kedua plot untuk tanah dan serasah menunjukkan bahwa hanya spesies richness antara kedua plot berbeda nyata, dan Evenness index untuk serasah pada kedua plot berbeda sangat nyata, sementara nilai rata-rata selainnya tidak berbeda nyata
4. Ada beberapa famili yang hilang setelah kebakaran yaitu Thripidae, Tetranichidae, Ellateridae, Staphylinidae, Scarabaidae, Julidae, Polydesmidae, Blattidae, Oedemeridae, Cercopidae, Mantidae, Tenebrionidae, Acrididae, Reduviidae, Scydmaenidae, Oxyopidae, Salticidae dan Tetragnathidae. Ordo-ordo yang tidak ditemukan pada plot bekas terbakar adalah Blattaria, Homoptera, Mantodea, dan Thysanoptera.
5. Famili yang hanya terdapat di plot terbakar adalah Linyphiidae, Podoridae, Sminthuridae, Zetorchestidae, Ephylohmalidae, Rhysotritiidae, Phytoseiidae, Argasidae, Veigaiidae dan Nitidulidae. Ordo yang hanya ditemukan pada plot bekas terbakar adalah Metastigmata.
6. Diperlukan pengamatan berkala terhadap plot terbakar sehingga dapat dilihat perkembangan kondisi biota tanah seperti kelimpahan, kekayaan, keragaman dan kemerataan jenisnya, apakah dapat kembali seperti semula atau tidak.
7. Pengelolaan atau penggunaan lahan bekas terbakar harus sangat diperhatikan agar tidak menghambat proses pemulihan kondisi tegakan.
DAFTAR PUSTAKA
Andersen. A.N. 1988. Immediate and longer-term effects of fire on seed predation by ants in sclerophyllous vegetation of southeastern Australia. Australian Journal of Ecology 13: 285-293.
Borror, Donald J., Charles A. Triplehorn dan Norman F. Johnson (1996). Pengenalan Pelajaran Serangga. Edisi ke-6. Gajah Mada University Press. Yogyakarta.
Brown, A.A dan K.P. Davis. 1973. Forest Fire Control Use. Mc. Grow – Hill Books Company. USA.
Burgers, A. and F. Raw. 1967. Soil Biology. Academic Press. London.
DeBano, C.F, D.G. Neay dan P.F. Folliot. 1998. Fire’s Effect On Ecosystems. New York. John Wiley and Sons. Inc.
Kalshoven, L.G.E. 1981. The Pests of Crops in Indonesia. Jakarta. PT. Ikhtiar Baru Van Hoeve.
Ludwig, J.A., and J.F. Reynolds. 1988. Statistical Ecology. New York. John Wiley and Sons.
Magurran, A. E. 1988. Ecological Diversity and Its Measurement. Cambridge. University Press.
Rahardjo, S. 2003. Komposisi Jenis dan Adaptasi Tumbuhan Bawah Pada Areal Bekas Kebakaran di Bawah Tegakan. [Tesis] Bogor. Sekolah Pascasarjana IPB.
Supriyanto, I.C. Stuckle, C.A. Siregar dan J. Kartana. 2001a. Forest Health Monitoring To Monitor The Sustainability Of Indonesian Tropical Rain Forest, Volume I. Bogor. ITTO-SEAMEO BIOTROP.
Supriyanto, I.C. Stuckle, C.A. Siregar dan J. Kartana. 2001b. Forest Health Monitoring To Monitor The Sustainability Of Indonesian Tropical Rain Forest, Volume II. Bogor. ITTO-SEAMEO BIOTROP.
Szujecki, A. 1987. Ecology of Forest Insect. Warzawa. PWN – Polish Scientific Publisher.
Wahyuningriyanti, R. 2005. Penilaian Dampak Kebakaran Hutan Terhadap Vegetasi Dengan Metode Forest Health Monitoring (Studi Kasus di Hutan Pendidikan Gunung Walat , Kabupaten Sukabumi). Skripsi Bogor. Fakultas Kehutanan IPB.
Wallwork, J.A. 1970. Ecology of Soil Animals. England. McGraw-Hill Publishing Company Limited.

Keanekaragaman Anthropoda Tanah
66
Wallwork, J.A. 1976. The Distribution and Diversity of Soil Fauna. San Franscisco. Academic Press.
Wooley, T. A. 1988. Acarology, Mites and Human Welfare. USA. John Wiley & Son, Inc.

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 67 – 76
67
APLIKASI SISTEM INFORMASI GEOGRAFIS (SIG) DAN PENGINDERAAN JAUH UNTUK MODEL HIDROLOGI ANSWERS
DALAM MEMPREDIKSI EROSI DAN SEDIMENTASI Studi Kasus : DTA Cipopokol Sub DAS Cisadane Hulu Kabupaten Bogor
(Application of Geographical Information System (GIS) and Remote Sensing
for ANSWERS Hydrological Model in Predicting Erosion and Sedimentation.
Case Study: Cipopokol Water Cathment Area, Cisadane Hulu Sub Watershed,
Bogor District)
DIAH IRAWATI DWI ARINI1), LILIK BUDI PRASETYO1) DAN OMO RUSDIANA2)
1)Departemen Konservasi Sumberdaya Hutan dan Ekowisata IPB Bogor 16680 2)Departemen Manajemen Hutan IPB Bogor 16680
Diterima 22 Januari 2007 / Disetujui 28 Mei 2007
ABSTRACT
Forest conversion in watershed may lead to erosion, sedimentation, and water quantity fluctuation. Prediction of erosion and sedimentation quantity is one way to describe condition of a watershed. ANSWERS is one of good hydrological model since it can be linked with map. It can formulated the best land use composition and distribution based on simulation. However, most of steps in ANSWERS are conducted manually, especially during thematic map development. The objective of research are (1) to combine the GIS and Remote Sensing in order to reduce technical error during thematic map development, (2) to predict the value of erosion and sedimentation to map the class of erosion and sedimentation range. Research is done in Cipopokol water cathment, Cisadane Hulu sub watershed, Bogor district at June untill October 2005. Keywords: Sedimentation, erosion, water catchment area, GIS, Spatial data
PENDAHULUAN
Daerah Aliran Sungai (DAS) merupakan suatu wilayah daratan yang menerima air hujan untuk kemudian mengalirkannya kembali melalui satu sungai utama menuju ke hilir. DAS bagian hulu seringkali menjadi fokus perencanaan pengelolaan DAS karena selain fungsinya yang sangat penting sebagai daerah tangkapan air (Water Catchment Area) juga adanya keterkaitan biofisik dengan daerah tengah dan hilir. Segala bentuk kerusakan yang terjadi di daerah hulu pada akhirnya tidak hanya akan membawa dampak bagi daerah hulu saja namun akhirnya juga berdampak pada daerah tengah dan terutama daerah hilir.
DAS Cisadene berhulu di Gunung Gede Pangrango dan Gunung Halimun. Saat ini kondisi peruntukan lahan di kawasan hulu ini telah mengalami banyak perubahan akibat pemanfaatan lahan yang kurang sesuai dengan fungsinya. Luas lahan kritis yang telah dipetakan oleh Balai Pengelolaan DAS Citarum-Ciliwiung pada tahun 2003 mencapai angka 12.723,20 Ha atau sekitar 8% dari luas keseluruhan DAS Cisadane (156.043 Ha).
Daerah Tangkapan Air Cipopokol merupakan bagian dari DAS Cisadane bagian hulu yang saat ini telah mengalami banyak perubahan penutupan lahan menjadi pemukiman, perkebunan, lahan pertanian dan sebagainya. Perubahan ini tentunya akan membawa pengaruh yang cukup besar bagi kemampuan lahan untuk menangkap air hujan yang jatuh untuk kemudian menyerap dan menyimpannya di dalam tanah (BAPEDA Prop. Jawa Barat 2004).
Prediksi tingkat bahaya erosi akibat perubahan penutupan lahan telah banyak dilakukan melalui pendekatan model hidrologi diantaranya USLE (Universal Soil Loss Equation). Dalam permodelan hidrologi metode USLE termasuk ke dalam model empiris yang bersifat lumped dimana parameter dan variabel masukan, keluaran dan besaran yang mewakilinya tidak memiliki variabilitas keruangan atau spatial (Harto, 1993). Namun sesuai dengan perkembangannya, saat ini model USLE telah dapat diaplikasikan dengan menggunakan Sistem Informasi Geografis (SIG). Model hidrologi lain yang banyak digunakan selain USLE yaitu ANSWERS (Areal Nonpoint Source Watershed Environment Response Simulation). Model hidrologi yang diperkenalkan oleh Beasley dan

Aplikasi Sisitem InformasiGeografis (GIS)
68
Huggins (1991) kemudian dikembangkan kembali oleh Environmental Protection Agency (EPA) di bawah Purdue Agricultural Experiment Station. ANSWERS merupakan model simulasi karakteristik DAS yang biasanya digunakan untuk mengevaluasi kondisi DAS khususnya di daerah pertanian. Keunggulan utama model ini adalah dapat digunakan untuk kegiatan simulasi perencanaan dan mengevaluasi strategi dalam mengendalikan erosi.
Penelitian ini bertujuan untuk mengkombinasikan aplikasi SIG dan Inderaja dengan model hidrologi ANSWERS. Kombinasi dari metode ini diharapkan dapat memprediksi besarnya nilai erosi dan sedimentasi di DTA Cipopokol untuk selanjutnya dapat dipetakan tingkat
penyebaran erosi dan sedimentasi. Informasi yang dihasilkan diharapkan dapat menjadi pertimbangan bagi kegiatan rehabilitasi DTA Cipopokol Sub DAS Cisadane Hulu.
METODE PENELITIAN
Penelitian diiaksanakan di Daerah Tangkapan Air Cipopokol yang merupakan bagian dari hulu DAS Cisadane di Kabupaten Bogor (Gambar 1). Waktu kegiatan dilaksana-kan selama 4 bulan dari bulan Juli sampai Oktober 2005.
Gambar 1. Peta Lokasi Penelitian. Metode penelitian di bagi ke dalam beberapa tahapan : 1. Tahap pengumpulan data.
Data yang dikumpulkan digunakan sebagai masukan atau input data untuk model hidrologi ANSWERS. Data yang dikumpulkan terdiri dari : a. Data peta, meliputi Citra ASTER/TERRA tahun 2004,
peta batas daerah tangkapan air Cipopokol, peta jenis tanah, peta topografi, peta sungai,
b. Data atribut, meliputi data curah hujan, tinggi muka air, sedimen, data parameter tanah, penggunaan lahan dan parameter karakteristik sungai.
c. Data lapangan, meliputi data titik-titik koordinat penutupan lahan di daerah tangkapan air Cipopokol.
Lo (1995) menjelaskan bahwa hasil pengamatan penutupan lahan, diharapkan dapat menduga kegiatan manusia serta penggunaan lahan. Data mengenai penutupan lahan DTA Cipopokol diperoleh dengan melakukan klasifikasi citra ASTER (Advanced Space Borne Thermal Emission and Reflection Radiometer) tahun 2004.

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 67 – 76
69
Klasifikasi dilakukan berdasarkan pada kenampakan bentuk penutupan lahan yang dapat dilihat secara jelas dari perbedaan warna piksel pada citra ASTER dengan menggunakan kombinasi band 2-3-1. Subsistem yang digunakan adalah VNIR (Visible and Near Infrared) yang memiliki resolusi 15x15 meter. Fungsi utama dari VNIR adalah mendeskripsikan sumberdaya air, tanah serta kerapatan tanaman.
2. Tahap Pembangunan Basis Data
a. Pembuatan Peta Digital. Pembuatan peta digital dilakukan dengan melakukan konversi peta-peta analog menjadi bentuk digital. Peralatan yang digunakan adalah scanner dan software ArcView GIS 3.3.
b. Pembuatan peta penutupan lahan,. Pemetaan penutupan lahan (land cover) merupakan suatu upaya dalam menyajikan informasi tentang pola penggunaan atau penutupan lahan di suatu wilayah secara spasial. Informasi penutupan lahan diperoleh dengan melakukan klasifikasi atau penafsiran citra satelit ASTER tahun 2004 dengan menggunakan software ERDAS Imagine 8.5 (ERSDAC, 2001).
c. Pembuatan Peta DEM (Digital Elevation Model) dan Arah Aliran (Flow Direction). DEM merupakan gambaran citra yang mampu memetakan ketinggian tempat dari permukaan bumi atau elevasi. Turunan dari peta DEM adalah peta ketinggian dan peta kelas lereng. Arah aliran merupakan arah dimana air hujan yang jatuh ke bumi mengalir menuju suatu outlet. Peta arah aliran diperoleh dari peta kontur yang selanjutnya dilakukan analisis secara spasial (Spatial Analyst).
d. Pembangunan Data model hidrologi ANWERS. Dalam model hidrologi ANSWERS suatu DAS diekspresikan sebagai kumpulan elemen bujursangkar yang diasumsikan homogen atau setiap areal mempunyai parameter hidrologi dan erosi yang sama. Masukan
model hidrologi ANSWERS terdiri dari lima bagian yaitu data intensitas hujan, data jenis dan parameter tanah, data jenis dan parameter penggunaan lahan, data nomor saluran (channel) dan parameter sungai serta data elemen (baris dan kolom, kemiringan lereng, arah aliran, jenis tanah, jenis penutupan lahan, sungai/ saluran dan parameternya, nomor stasiun penakar hujan dan data elevasi/ketinggian tempat). Penentuan luasan grid atau elemen dilakukan berdasarkan jumlah maksimun elemen yang diijinkan di dalam model yaitu sebanyak 1000 elemen. Jika luasan daerah tangkapan air Cipopokol mencapai 159,20 Ha maka jumlah elemennya adalah 995 elemen dengan ukuran gridnya 40 x 40 meter atau seluas 0,16 Ha.
e. Konversi peta vektor ke dalam format raster. Peta-peta dalam format vektor untuk selanjutnya dirubah ke dalam format raster (piksel).
f. Konversi raster ke dalam bentuk text. Pembangunan data untuk masukan input ke dalam model dilakukan dengan merubah data raster ke dalam bentuk *txt (text file) untuk selanjutnya data tersebut diinput ke dalam software ANSWER_PLUS V1.01.
3. Pemetaan penyebaran nilai erosi dan sedimentasi
Peta penyebaran nilai erosi dan sedimentasi dilakukan dengan cara merubah angka yang diperoleh dari hasil keluaran model ANSWERS menjadi bentuk raster dan diolah kembali menjadi bentuk peta. 4. Simulasi penggunaan lahan
Simulasi penggunaan lahan dilakukan dengan tujuan untuk menentukan besarnya nilai erosi dan sedimentasi yang dihasilkan pada tutupan lahan yang berbeda.
Secara keseluruhan alur penelitian seperti ditunjukkan pada Gambar 2.

Aplikasi Sisitem InformasiGeografis (GIS)
70
Gambar 2. Alur Penelitian
HASIL DAN PEMBAHASAN
Data Masukan ANSWERS
a. Penutupan Lahan
Hasil interpretasi tutupan lahan daerah penelitian dapat dikelompokkan ke dalam 6 kelas penutupan lahan terdiri dari hutan seluas 8,80 Ha, perkebunan 41,12 Ha, pertanian lahan kering 75,20 Ha, pemukiman 4,48 Ha, sawah 3,84 Ha dan semak belukar 25,76 Ha. Nilai akurasi hasil klasifikasi citra ASTER sebesar 81,82% untuk overall classification accuracy dan 72,45% untuk overall kappa statistic.
b. Jenis Tanah
Secara umum, Sub DAS Cisadane hulu didominasi oleh dua jenis tanah yaitu latosol coklat dan latosol coklat kemerahan. DTA Cipopokol termasuk ke dalam jenis tanah latosol coklat dengan sifat agak peka erosi, memiliki permeabilitas sedang sampai lambat, kendungan bahan organiknya berkisar 1,24 – 6,93%, tekstur liat dan struktur gumpal. c. Kondisi Saluran (Channel)
Daerah Tangkapan Air Cipopokol memiliki 3 saluran atau aliran sungai, terdiri dari satu sungai utama yaitu Sungai Cipopokol dengan lebar mencapai 1,80 meter dan 2 anak sungai dengan lebar masing-masing 1,30 meter dan
KONDISI UMUM DTA CIPOPOKOL
SUB DAS
CISADANEHULU
Data Fisik
Cuarah Hujan dan
Debit
Penggunaan lahan dan
parameter
Kelas Kelerengan
dan Elevasi
Arah
Aliran
Karakteristik Saluran dan Parameter
Jenis tanah dan
Parameter
Model Hidrologi
ANSWERS
Nilai erosi dan
sedimen
Uji Akurasi Diterima
?
Peta penyebaran erosi dan
sedimentasi
DTA Cipopokol
Simulasi
Penggunaan lahan
Tidak
Ya
Media Konservasi Vol. XII, No. 2 Agustus 2007 : 67 – 76
71
0,80 meter. Tipe sungainya adalah sungai alami dan terdapat sejumlah batu, rumput atau gulma. Karakteristik Sungai Cipopokol memiliki bentuk paralel dimana dua jalur pengaliran bersatu di bagian hilir dan banjir kadang-kadang terjadi pada pertemuan sungai. Sungai bagian hulu suatu DAS memiliki ukuran yang tidak terlalu besar jika dibandingkan dengan sungai di bagian tengah dan hilir yang dapat mencapai puluhan meter. Kedalaman sungainya pun dapat dikatakan dangkal. d. Kemiringan lereng dan Ketinggian tempat
Pengolahan data kontur/topografi menghasilkan nilai ketinggian tempat. Kelas ketinggian tempat di DTA Cipopokol dibagi ke dalam 4 kelas, terdiri dari ketinggian 500 – 600 mdpl dengan luas 1,60 Ha, ketinggian 600 – 700 mdpl seluas 56,32 Ha, ketinggian 700 – 800 mdpl seluas 79, 36 Ha dan ketinggian di atas 800 mdpl seluas 21,92 Ha.
DTA Cipopokol memiliki kelas kelerangan (slope) yang bervariasi, mulai dari datar sampai dengan sangat curam. Didominasi oleh kelas lereng agak curam (15-25%) dengan luas 59,52 Ha, kelas lereng landai (8-15%) seluas 40,32 Ha, kelas lereng curam (25-40%) seluas 38,72 Ha, kelas lereng sangat curam (>40%) seluas 10,88 Ha dan datar (0-8%) seluas 9,76 Ha. e. Arah Aliran (flow direction)
Aliran air mengalir dari tempat yang tinggi menuju ke tempat yang rendah, dan menuju ke arah sungai serta berlawanan dengan garis kontur. Sungai di kawasan Cipopokol mengalir dari arah timur menuju ke barat sehingga tidak ditemukan arah aliran yang menuju ke arah tenggara. Arah aliran terluas pada arah aliran utara atau 270o dengan luas 44,5 Ha selanjutnya arah barat atau 1800 seluas 42,4 Ha (Tabel 1).
Tabel 1. Arah Aliran (flow direction)
Arah aliran Keterangan Jumlah elemen
Luas (Ha)
0o Timur 1 0,16 45o Tenggara 0 0,00 90o Selatan 15 2,40
135o Barat Daya 165 26,40 180o Barat 265 42,40 225o Barat Laut 157 25,12 270o Utara 278 44,48 315o Timur Laut 114 18,24
Luas / Jumlah 995 159,20
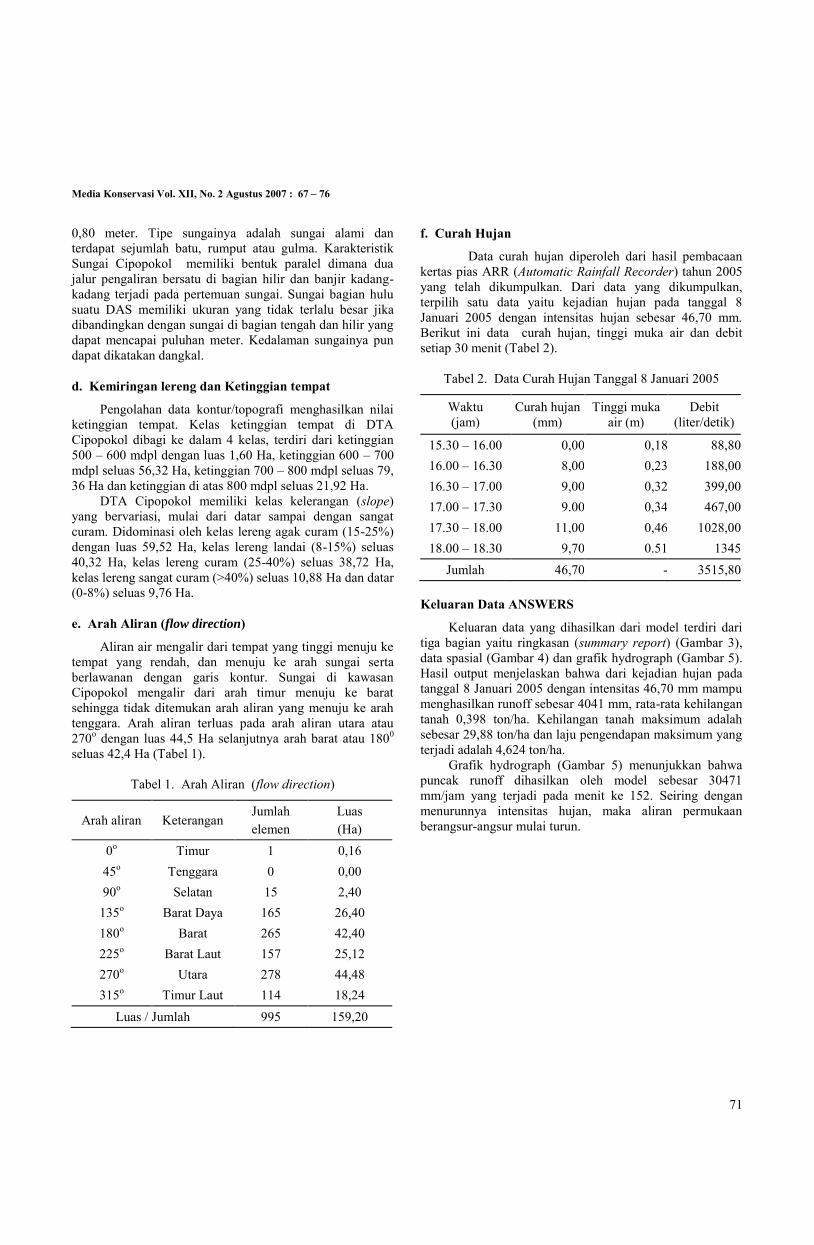
f. Curah Hujan
Data curah hujan diperoleh dari hasil pembacaan kertas pias ARR (Automatic Rainfall Recorder) tahun 2005 yang telah dikumpulkan. Dari data yang dikumpulkan, terpilih satu data yaitu kejadian hujan pada tanggal 8 Januari 2005 dengan intensitas hujan sebesar 46,70 mm. Berikut ini data curah hujan, tinggi muka air dan debit setiap 30 menit (Tabel 2).
Tabel 2. Data Curah Hujan Tanggal 8 Januari 2005
Waktu (jam)
Curah hujan (mm)
Tinggi muka air (m)
Debit (liter/detik)
15.30 – 16.00 0,00 0,18 88,80 16.00 – 16.30 8,00 0,23 188,00 16.30 – 17.00 9,00 0,32 399,00 17.00 – 17.30 9.00 0,34 467,00 17.30 – 18.00 11,00 0,46 1028,00 18.00 – 18.30 9,70 0.51 1345
Jumlah 46,70 - 3515,80 Keluaran Data ANSWERS
Keluaran data yang dihasilkan dari model terdiri dari tiga bagian yaitu ringkasan (summary report) (Gambar 3), data spasial (Gambar 4) dan grafik hydrograph (Gambar 5). Hasil output menjelaskan bahwa dari kejadian hujan pada tanggal 8 Januari 2005 dengan intensitas 46,70 mm mampu menghasilkan runoff sebesar 4041 mm, rata-rata kehilangan tanah 0,398 ton/ha. Kehilangan tanah maksimum adalah sebesar 29,88 ton/ha dan laju pengendapan maksimum yang terjadi adalah 4,624 ton/ha.
Grafik hydrograph (Gambar 5) menunjukkan bahwa puncak runoff dihasilkan oleh model sebesar 30471 mm/jam yang terjadi pada menit ke 152. Seiring dengan menurunnya intensitas hujan, maka aliran permukaan berangsur-angsur mulai turun.

Aplikasi Sisitem InformasiGeografis (GIS)
72
Gambar 3. Summary Report ANSWERS Gambar 4. Spasial data
Gambar 5. Grafik hydrograph
Prediksi Erosi dan Sedimentasi DTA Cipopokol
Prediksi nilai erosi dan sedimen bertujuan untuk memberikan gambaran secara menyeluruh terhadap erosi tanah yang terjadi, untuk DTA Cipopokol yang merupakan bagian hulu dari DAS Cisadane. Berdasarkan hasil perhitungan dengan menggunakan model hidrologi ANSWERS dapat diketahui bahwa jumlah elemen yang
mengalami erosi atau kehilangan tanah sebanyak 852 elemen atau sebesar 136,32 Ha dan sisanya sebanyak 103 elemen atau seluas 22,88 Ha mengalami sedimentasi atau pengendapan. Hasil prediksi nilai erosi dan sedimentasi pada setiap kelas penutupan lahan dapat dilihat dalam Tabel 3.
Tabel 3. Prediksi nilai erosi dan sedimentasi
Kelas tutupan lahan
Kelas Sedimentasi (Ton/Ha) Kelas Erosi (Ton/Ha)
0-0,5 0,5 – 1 >1 0-0,5 0,5 – 1 5-1 10-May >10 Hutan 0.32 0 0 1.12 3.68 3.68 0 0 Perkebunan 0 0 0 4.8 16.8 18.24 0.96 0.32 Pertanian Lahan Kering 0.16 0 0.16 0.64 24.8 47.36 2.08 0 Pemukiman 0 0 0 0.16 0.8 3.52 0 0 Sawah 2.56 0 0 1.28 0 0 0 0 Semak Belukar
16.8 2.24 0.8 6.08 0 0 0 0 Total 19.84 2.24 0.96 14.08 46.08 72.8 3.04 0.32

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 67 – 76
73
Erosi dan sedimentasi dapat dipengaruhi oleh berbagai macam faktor diantaranya iklim, vegetasi penutup tanah, topografi, dan jenis tanah. Daerah Tangkapan Air Cipopokol merupakan wilayah yang memiliki intensitas hujan bulanan yang cukup tinggi. Berdasarkan data yang diperoleh dari Balai Pengelolaan DAS Citarum-Ciliwung tahun 2003 sampai 2004, bulan basah terjadi selama 11 bulan (bulan September sampai dengan bulan Juli) dan bulan lembab terjadi selama satu bulan.
Selain iklim, vegetasi penutup tanah juga memberikan pengaruh terhadap terjadinya erosi dan sedimentasi. Pengaruh penutupan lahan terhadap terjadinya erosi dan sedimentasi di DTA Cipopokol. Sebagian besar elemen pada kelas penutupan lahan berupa pertanian lahan kering dan perkebunan, hampir seluruhnya mengalami kehilangan tanah yaitu masing-masing sebanyak 468 elemen atau sebesar 74,88 Ha dan 257 elemen atau sebesar 41,12 Ha. Kelas penutupan lahan berupa hutan menghasilkan erosi seluas 8,38 Ha, kelas penutupan lahan berupa sawah menghasilkan erosi seluas 1,28 Ha dan kelas penutupan lahan berupa semak/belukar menghasilkan erosi seluas 6,08 Ha.
Kisaran nilai erosi antara 1-5 Ton/Ha merupakan kisaran nilai erosi paling luas di wilayah DTA Cipopokol yaitu sebesar 72,80 Ha, dengan kelas penutupan lahan berupa pertanian lahan kering merupakan penyumbang erosi terbesar yaitu 47,36 Ha. Diikuti dengan kisaran kelas erosi 0,5 – 1 Ton/Ha seluas 46,08 Ha.
Kisaran nilai erosi terbesar yaitu lebih dari 10 Ton/Ha terjadi pada kelas penutupan lahan berupa perkebunan seluas 0,32 Ha yang berarti bahwa hanya dua elemen berada pada nilai kisaran kelas erosi tersebut. Kisaran kelas sedimentasi atau pengendapan yang paling luas terjadi pada 0 sampai dengan 0,5 Ton/Ha. Berdasarkan kelas penutupan lahannya, sedimen yang paling banyak terjadi adalah kelas penutupan semak/belukar seluas 19,68 Ha.
Kelas penutupan lahan berupa hutan dan semak belukar menghasilkan nilai erosi yang lebih kecil dibandingkan perkebunan dan pertanian lahan kering. Jenis vegetasi hutan yang ditemukan di DTA Cipopokol adalah hutan pinus dengan struktur penutupan tajuk jarang, namun kerapatan tumbuhan bawah yang ada di sekitar hutan pinus mampu menciptakan suatu stratifikasi tajuk yang berlapis sehingga dapat menurunkan kecepatan terminal air hujan serta memperkecil diameter tetesan air. Tumbuhan bawah merupakan stratum vegetasi terakhir yang menentukan besar kecilnya nilai erosi percikan. Hal yang sama
dikemukakan oleh Suwardjo et al. dalam Purnama (2005) yang menyatakan bahwa alang-alang yang bercampur semak akan lebih efektif dalam mencegah tanah longsor daripada alang-alang murni. Campuran antara Albizzia sp dengan semak belukar akan sama efektifnya dengan alang-alang murni namun apabila semaknya dibersihkan maka erosi akan meningkat menjadi 114 kali lipat. Sedangkan alang-alang yang dicangkul bersih setiap tahun akan meningkatkan erosi sebesar 84 kali lipat dibandingkan dengan erosi pada alang-alang murni.
Sedimen atau pengendapan yang terjadi sebagai akibat dari proses erosi di Daerah Tangkapan Air Cipopokol, sebagian besar terjadi pada kelas penutupan lahan berupa semak belukar. Keberadaan semak/belukar yang sebagian besar merupakan tumbuhan bawah di sepanjang sungai Cipopokol mampu memberikan perlindungan terhadap erosi dan aliran permukaan serta mampu menahan pengangkutan tanah yang tererosi untuk masuk ke dalam sungai. Sedimen yang mengendap akibat pengangkutan tanah oleh air memiliki kecenderungan berada tersebar di sepanjang sungai Cipopokol.
Selain disebabkan oleh jenis penutupan lahan, terjadinya erosi juga dipengaruhi oleh faktor topografi yakni kemiringan lereng dan panjang lereng. Kedua faktor tersebut akan sangat menentukan besarnya kecepatan air larian dan volume air larian. Lahan pertanian dan perkebunan di DTA Cipopokol sebagian besar berada pada lereng dengan kelas kemiringan curam (25 – 40%) dan agak curam (15 – 25%). Nilai sedimen banyak terjadi pada kelas kemiringan agak curam yaitu 15 – 25% dengan luas mencapai 8,16 Ha diikuti dengan kelas kemiringan landai seluas 4,80 Ha. Sedangkan nilai erosi lebih banyak terjadi pada kelas kemiringan 15 – 25% dengan luas mencapai 52,60 Ha.
Hasil overlay antara kemiringan lereng dengan kelas penutupan lahan diperoleh bahwa sedimentasi terjadi pada kelas penutupan lahan berupa semak belukar yang berada pada kelas kemiringan antara 15 – 25% dengan kriteria agak curam seluas 7,84 Ha. Sedangkan nilai erosi terjadi pada kelas penutupan lahan pertanian lahan kering seluas 29,12 Ha dengan kelas kemiringan 15 – 25% diikuti dengan kelas kemiringan 18,56 Ha pada kelas kemiringan 8 – 15%. Hal ini mengindikasikan bahwa sebagian besar dari penggarap lahan belum menerapkan teknik konservasi tanah dan air secara tepat.
Penyebaran erosi dan sedimentasi seperti pada Gambar 6.

Aplikasi Sisitem InformasiGeografis (GIS)
74
Gambar 6. Penyebaran Erosi dan Sedimentasi.
Simulasi Penggunaan Lahan
Simulasi penggunaan lahan dilakukan dengan melakukan penyusunan luas penggunaan lahan melalui beberapa skenario. Skenario yang disusun berdasarkan pada RTRW (Rencana Tata Ruang Wilayah) Kabupaten Bogor dan berdasarkan UU No 32 tahun 1990 yang mengatur tentang kawasan lindung.
Hasil overlay dengan peta Rencana Tata Ruang Wilayah Kabupaten Bogor, Daerah Tangkapan Air
Cipopokol merupakan kawasan yang diperuntukkan bagi budidaya tanaman tahunan atau perkebunan.
Menurut UU No.32 Tahun 1990 dijelaskan mengenai kriteria-kriteria tentang kawasan lindung. Beberapa point yang digunakan sebagai dasar simulasi penggunaan lahan diantaranya kawasan pada kelerengan > 40% dan buffer 50 meter untuk sungai dengan lebar kurang dari 30 meter. Simulasi penggunaan lahan berikut nilai sedimen, erosi dan runoff yang dihasilkan dapat dilihat dalam Tabel 4.
Tabel 4. Simulasi penggunaan lahan
Skenario Tutupan Lahan Luas Runoff (mm)
Rata-rata kehilangan tanah
(kg/Ha)
Erosi maksimum
(kg/Ha)
Pengendapan maksimum
(kg/ha) Kondisi Aktual Hutan 8,80
4014 398 29088 4624
Perkebunan 41,12 Pertanian lahan kering 75,20 Pemukiman 4,48 Sawah 3,84 Semak belukar 25,71
Skenario 1 Perkebunan 154.72 4194 432 30503 1941 Pemukiman 4.48

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 67 – 76
75
Tabel 4. (Lanjutan)
Skenario 2 Pertanian Lahan Kering 154.72 3864 546 27609 5298 Pemukiman 4.48 Skenario 3 Hutan (lereng >40%) 37.92
3972 383 29088 2017
Perkebunan 30.88 Pertanian Lahan Kering 63.84 Pemukiman 3.68 Sawah 3.84 Semak Belukar 19.04
Hasil simulasi dari skenario satu menunjukan bahwa
perubahan penutupan lahan menjadi kawasan perkebunan akan meningkatkan nilai runoff, nilai rata-rata kehilangan tanah dan erosi maksimum. Penurunan yang cukup signifikan terjadi pada pengendapan maksimum sebesar 1,941 Ton/Ha dibandingkan pada kondisi aktual sebesar 4,624 Ton/Ha. Hal yang sama terjadi pada skenario ketiga yaitu perubahan penutupan lahan menjadi kawasan budidaya dalam bentuk pertanian lahan kering. Peningkatan terjadi pada parameter rata-rata kehilangan tanah dan laju pengendapan maksimum, sedangkan nilai runoff, laju erosi maksimum mengalami penurunan dibandingkan pada kondisi aktual dan hasil skenario satu.
Perbandingan nilai runoff antara penutupan lahan berupa perkebunan dan pertanian lahan kering menunjukan bahwa perkebunan menghasilkan nilai runoff yang lebih besar dibandingkan pertanian lahan kering. Hal ini lebih disebabkan oleh adanya perbedaan nilai parameter-parameter penggunaan lahan. Besar kecilnya nilai runoff dipengaruhi oleh faktor pengelolaan lahan. Penutupan lahan pertanian lahan kering di daerah tangkapan air lebih banyak menerapkan teknik-teknik konservasi tanah diantaranya sistem teras dan guludan, berbeda dengan kelas perkebunan yang kurang menerapkan teknik konservasi (Arsyad, 1989).
Sesuai dengan Undang-Undang No. 32 tahun 1990, penetapan lokasi kawasan perlindungan setempat dilakukan dengan tujuan menjaga sempadan sungai dari kegiatan manusia yang dapat mengganggu dan merusak kualitas air sungai, kondisi fisik pinggir dan dasar sungai serta mengamankan aliran sungai. Simulasi ketiga dilakukan dengan melakukan revegetasi pada kawasan dengan kelerengan > 40% dan sepanjang kanan-kiri sungai. Hasil yang diperoleh menunjukan penurunan terjadi pada nilai parameter runoff, rata-rata kehilangan tanah, dan laju pengendapan maksimum, namun laju erosi maksimum tidak mengalami perubahan dari kondisi aktual.
Dari hasil simulasi di atas, maka dapat disimpulkan bahwa tindakan revegetasi untuk Daerah Tangkapan Air Cipopokol sangat diperlukan, terutama untuk kawasan yang telah ditetapkan menjadi kawasan perlindungan setempat (lahan pada kelerengan >40% dan kawasan sempadan sungai) yang pada kondisi aktual saat ini telah berubah fungsi menjadi lahan pertanian. Perlindungan terhadap
kawasan resapan air dilakukan dengan tujuan untuk memberikan ruang yang cukup bagi peresapan air hujan pada suatu daerah tertentu guna keperluan penyediaan kebutuhan air tanah serta penanggulangan banjir, baik kawasan bawahnya maupun kawasan yang bersangkutan.
Hasil simulasi menunjukan bahwa kawasan budidaya berupa pertanian lahan kering dan perkebunan menghasilkan nilai rata-rata kehilangan tanah yang cukup tinggi dibandingkan pada kondisi aktual. Hasil indeks sensitifitas terhadap parameter tanah maupun penggunaan/ penutupan lahan diperoleh bahwa parameter FC atau laju infiltrasi merupakan parameter yang memiliki tingkat sensitifitas tertinggi sehingga dalam hal ini pengolahan atau pemadatan tanah yang dilakukan akan meningkatkan rata-rata kehilangan tanah, sehingga perlu adanya upaya teknik konservasi yang dapat meningkatkan laju infiltrasi. Teknik konservasi yang sangat efektif diterapkan adalah tanaman penutup tanah atau cover crop khususnya pada kawasan perkebunan dan pemberian mulsa atau bahan organik pada kawasan pertanian lahan kering. Penggunaan tanaman penutup tanah/cover crop akan sangat berperan dalam melindungi tanah dari ancaman kerusakan oleh erosi. Hal ini akan menyebabkan berkurangnya dispersi air hujan dan mengurangi jumlah serta kecepatan aliran permukaan sehingga mampu mengurangi erosi dan memperbesar infiltrasi ke dalam tanah. Bahan organik dan mulsa akan berperan dalam peningkatan ketahanan struktur tanah, memperbesar kemampuan tanah untuk menyerap dan menahan air hujan serta mampu menambah unsur hara.
Daerah Tangkapan Air Cipopokol yang merupakan bagian dari Sub DAS Cisadane Hulu, selain berfungsi sebagai kawasan resapan air, juga memiliki potensi sebagai kawasan budidaya untuk pertanian dan merupakan salah satu penyangga sektor perekonomian Kabupaten Bogor. Berkaitan dengan hal tersebut, maka perlu diadakannya koordinasi dan kerjasama antar pihak-pihak terkait dalam hal kegiatan penataan ruang yang hendaknya disesuaikan dengan kondisi atau karakteristik wilayah daerah yang bersangkutan baik secara fisik maupun kondisi sosial masyarakatnya.

Aplikasi Sisitem InformasiGeografis (GIS)
76
KESIMPULAN
1. Aplikasi Sistem Informasi Geografis (SIG) dan Penginderaan Jauh dapat digunakan dan diaplikasikan ke dalam model hidrologi ANSWERS sebagai sebuah tools untuk memudahkan dalam memperoleh masukan data ke dalam model.
2. Hasil prediksi nilai erosi dan sedimentasi DTA Cipopokol dengan menggunakan model hidrologi ANSWERS dari kejadian hujan tanggal 8 Januari 2005 dengan intensitas 46,70 mm adalah sebesar 0,398 ton/Ha untuk rata-rata kehilangan tanah, pengendapan atau sedimen yang terbentuk sebesar 4,624 ton/Ha dan menghasilkan runoff sebesar 2428 mm/jam.
DAFTAR PUSTAKA
Arsyad, S. 1989. Konservasi Tanah dan Air. Bogor: IPB Press.
[BAPEDA Propinsi Jawa Barat] Badan Perencanaan Daerah Propinsi Jawa Barat. 2004. Kebijakan Pengelolaan Sumberdaya Air di SWS Ciliwung-Cisadane untuk Mengatasi Krisis Air Jakarta. Di dalam: Seminar Krisis Air Jakarta: Tinjauan Pengelolaan Sumber Daya Air Terpadu Ciliwung Cisadane; Jakarta, 29 Juni 2004. Jakarta: BAPEDA Propinsi Jawa Barat.
Beasley, D.B dan L.F. Huggins. 1991. ANSWERS (Areal Nonpoint Source Watershed Environment Respon Simulation) User’s Manual: 2th Edition.Chicago: US EPA Region V.
[BPDAS Citarum-Ciliwung] Balai Pengelolaan Daerah Aliran Sungai Citarum-Ciliwung. 2003. Rencana Teknik Lapangan Rehabilitasi Lahan dan Konservasi Tanah DAS Cisadane. Bogor: BPDAS Citarum-Ciliwung.
[ERSDAC] Earth Remmote Sensing Data and Analysis Center. 2001. ASTER User’s Guide Part I General Ver.3.1. Japan. ERSDAC.
Harto Br, S. 1993. Analisis Hidrologi. Jakarta. PT Gramedia Pustaka Utama.
Lillesand, T.M & F.W. Kiefer. 1990. Penginderaan Jauh dan Interpretasi Citra. R. Dubahri, penerjemah; Yogyakarta; Gadjah Mada University Press.
Lo, C.P. 1995. Penginderaan Jauh Terapan. B. Purbowaseso, penerjemah. Jakarta. Universitas Indonesia.
Purnama, A. 2005. Analisis Perubahan Penggunaan Lahan Menggunakan Citra Satelit dan Perencanaan Penggunaan Lahan yang Berkelanjutan di Daerah Aliran Sungai (DAS) Citarum Hulu [tesis]. Bogor. Program Pascasarjana, Institut Pertanian Bogor.
Setiyanto. 2005. Analisis Karakteristik Biofisik dan Hidrograf Aliran di Daerah Tangkapan Air Cipopokol Sub DAS Cisadane Hulu [skripsi]. Bogor. Departemen Manajemen Hutan, Fakultas Kehutanan, Institut Pertanian Bogor.

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 77 – 79
77
HUJAN ASAM PADA BEBERAPA PENGGUNAAN LAHAN DI KABUPATEN DAN KOTA BOGOR
(Acid Rain on Several Landuses in Bogor Regency and City)
R PURIPUSPITA SARI1, SITI BADRIYAH RUSHAYATI2 DAN RACHMAD HERMAWAN2
1Departemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan IPB 2Bagian Hutan Kota dan Jasa Lingkungan Departemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan IPB
Diterima 2 April 2007 / Disetujui 9 Juli 2007
ABSTRACT
Acid rain is caused by increasing air pollution. The emission of sulphur dioxide and nitrogen dioxide from fossil fuel combustion into the atmosphere, which is followed by a chemical transformation (hydrolysis) into sulphuric and nitric acid, causes acid rain. The study was conducted on four (4) locations with different landuses. The lowest pH of rainfall was on industrial area, whereas the highest pH of rainfall was on vegetation area. pH of rainfall in beginning of rainy season was lower than pH of rainfall in middle of rainy season. Keyword : Acid rain, pH, landuse, rainfall
PENDAHULUAN
Udara merupakan faktor yang penting bagi makhluk hidup, namun seiring dengan meningkatnya jumlah penduduk, perkembangan industri dan makin padatnya transportasi menyebabkan kualitas udara mengalami penurunan. Penurunan kualitas udara ini disebabkan adanya berbagai macam polutan gas hasil berbagai aktifitas antropogenik (Fakuara, 1986). Polutan gas yang berada di atmosfer akan menyebabkan komposisi gas di atmosfer berubah, sehingga juga akan merubah kualitas hujan yang turun di wilayah tersebut. Berbagai senyawa gas diantaranya SOx, NOx, CO2, NO3
-, Cl-, SO42- , di atmosfer
akan bereaksi dengan H2O dan menghasilkan berbagai asam dan mengakibatkan pH air hujan menjadi rendah (Harmantyo, 1989; Soedomo, 2001).
Kemasaman hujan ini dipengaruhi oleh sumber polutan yang sangat terkait dengan jenis aktifitas dan penggunaan lahan (Husin et al., 1991). Kota dan Kabupaten Bogor merupakan wilayah potensial yang akan terus berkembang karena berbatasan dengan Kota Jakarta. Perkembangan kota di wilayah ini akan menghasilkan dampak negatif terhadap lingkungan terutama peningkatan konsentrasi polutan udara dan potensial akan menyebabkan dampak hujan asam. Dari kondisi ini, perlu dilakukan analisis kualitas air hujan di beberapa wilayah dengan perbedaan penggunaan lahan yang tersebar di Kabupaten dan Kota Bogor.
Tujuan penelitian ini adalah mengkaji pengaruh perbedaan penggunaan lahan terhadap pH, konsentrasi karbondioksida (CO2), klorida (Cl-), nitrat (NO3
-), nitrit
(NO2-), ammonia (NH3) dan sulfat (SO4
2-) yang terkandung dalam air hujan. Hasil dari penelitian ini diharapkan dapat menjadi dasar dalam pengelolaan lingkungan kaitannya dengan antisipasi terjadinya hujan asam.
METODE PENELITIAN
Penelitian dilakukan selama lima bulan yaitu mulai bulan Oktober 2004 sampai bulan Februari 2005. Tempat penelitian yaitu di Kota dan Kabupaten Bogor dengan pemilihan titik berdasarkan perbedaan penggunaan lahan yang diduga akan mempengaruhi konsentrasi polutan dan kemasaman air hujan. Lokasi pengambilan sampel sebagai berikut : a. Cibinong (industri, jalan raya, permukiman) b. Ciawi (jalan raya, pasar, sarana pendidikan) c. Kebun Raya Bogor (hutan kota, perkantoran, pasar) d. Megamendung (hutan tanaman, perkebunan, per-
mukiman).
Variabel yang diukur di empat lokasi penelitian ini adalah : pH air hujan, konsentrasi ammonia (NH3), klorida (Cl-), nitrit (NO2
-), nitrat (NO3-), sulfat (SO4
2-) dan karbon dioksida (CO2). Pada masing-masing lokasi dilakukan pengukuran 6 kali.
Analisis data untuk melihat hubungan antara pH air hujan dengan konsentrasi ion-ion nitrat, nitrit, sulfat, amonia, dan klorida yang terkandung didalam air hujan digunakan model regresi linear berganda, dengan persamaan sebagai berikut :
Y = a + b1X1 + b2X2 + b3X3 + b4X4 + b5X5 + b6X6

Hujan Asam pada Beberapa Penggunaan Lahan
78
Keterangan : Y = pH air hujan X1 = Konsentrasi gas karbon dioksida (mg/l) X2 = Konsentrasi gas ammonia (mg/l) X3 = Konsentrasi ion nitrat (mg/l) X4 = Konsentrasi ion nitrit (mg/l) X5 = Konsentrasi ion sulfat (mg/l) X6 = Konsentrasi ion klorida (mg/l) a = Intersep bi = Koefisien regresi
HASIL DAN PEMBAHASAN
Konsentrasi Polutan dan pH Air Hujan
Secara keseluruhan nilai pH dan konsentrasi polutan rata-rata di dalam air hujan pada masing-masing lokasi disajikan pada Tabel 1.
Tabel 1. Nilai rata-rata pH dan konsentrasi polutan di dalam air hujan
No Lokasi pH CO2 (mg/l)
NH3 (mg/l)
NO3-
(mg/l) SO4
2-
(mg/l) Cl-
(mg/l)
1 Megamendung 6,53 7,53 1,10 0,31 5,11 2,06
2 KRB 6,40 8,66 1,45 0,65 7,05 5,03 3 Ciawi 5,88 34,96 1,42 0,39 8,37 2,91 4 Cibinong 5,56 8,85 1,62 0,73 9,95 2,73
Konsentrasi CO2, NH3, NO3-, SO4
2- dan Cl- yang terkandung di dalam air hujan, terendah terukur di Megamendung. Hal ini disebabkan sumber polutan udara di kawasan tersebut relatif rendah (hutan tanaman, perkebunan, permukiman) bila dibandingkan dengan 3 kawasan lain. Rendahnya konsentrasi polutan udara ini menyebabkan senyawa asam yang terbentuk karena adanya reaksi kimia dengan H2O juga akan rendah sehingga pH air hujan agak tinggi (tidak terjadi hujan asam).
Cibinong sebagai kawasan industri, permukiman dan transportasi menghasilkan polutan udara tertinggi terutama NH3, NO3
- dan SO4
2-. Ketiga senyawa ini banyak dihasilkan dari berbagai jenis industri. Sedangkan Ciawi dengan kepadatan transportasi tinggi, CO2 yang terukur juga sangat tinggi. Alat transportasi dengan bahan bakar fosil (bensin, solar) akan menghasilkan limbah gas CO2 yang tinggi. Meskipun beberapa parameter kualitas udara yang terukur di sekitar Kebun Raya relatif tinggi, tetapi karena terdapat banyak vegetasi maka seiring dengan waktu maka vegetasi tersebut dapat mengabsorbsi dan menjerap polutan sehingga pH air hujan masih aman (belum terjadi hujan asam).
Pengaruh Konsentrasi Polutan terhadap pH Air Hujan
Frohlinger dan Kane (1975) dalam Husin et al. (1991) menjelaskan bahwa secara teoritis penurunan pH dalam air hujan terutama disebabkan oleh larutnya polutan SO4
2-, CO2 dan NOx, sedangkan peningkatan pH disebabkan oleh terlarutnya kalsium (Ca) dan ammonium (NH3) dalam air hujan. Untuk melihat hubungan antara konsentrasi polutan dalam air hujan dengan pH air hujan pada bulan Oktober
2004 – Februari 2005, maka dilakukan analisis regresi linear berganda. Hasil analisis regresi linear berganda hubungan antara pH (Y) dengan polutan, yaitu CO2 (X1), NH3 (X2), NO3
- (X3), SO42- (X4) dan Cl- (X5) menghasilkan
persamaan sebagai berikut :
Y = 6.724811 – 0.008074X1 + 0.207894X2 - 0.411138X3 – 0.073305X4 + 0.039120X5
Hasil uji statistik menunjukkan bahwa konsentrasi polutan di udara berpengaruh terhadap pH air hujan pada taraf nyata 5%. Berdasarkan nilai koefisien regresi maka dapat diketahui bahwa konsentrasi nitrat (NO3
-) paling berpengaruh dalam menurunkan pH air hujan, setelah itu diikuti oleh konsentrasi sulfat (SO4
2-) dan konsentrasi karbon dioksida (CO2). Ion NO3
- lebih berpengaruh dalam
menurunkan pH air hujan karena bersifat sulit larut dalam air sehingga lebih sulit tercuci oleh hujan, sedangkan ion SO4
2- bersifat mudah larut dalam air, sehingga mudah tercuci oleh hujan, selain itu masa aktif NO3
- di udara lebih lama daripada SO4
2-. Parameter kualitas udara khususnya NO3
- an SO42- yang terukur di Cibinong sangat tinggi,
hasilnya pH air hujan yang terukur rendah (sudah terjadi hujan asam karena pH air hujan < 5,6).
Kaitan antara Periode Musim Hujan dengan pH
Bulan Oktober (awal musim hujan) polutan yang terukur masih tinggi sehingga ketika terjadi hujan maka senyawa polutan gas itu bereaksi dengan H2O dan menghasilkan pH rendah. Berbeda dengan saat pertengahan musim hujan, karena berulangkali sudah terjadi hujan dan pencucian polutan di atmosfer, maka pH yang terukur

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 77 – 79
79
makin tinggi (banyak senyawa pembentuk asam tercuci oleh hujan). Dari Gambar 1 terlihat dengan jelas dari bulan
Oktober sampai dengan bulan Fabruari, di keempat lokasi penelitian pH-nya cenderung meningkat.
Gambar 1. pH air hujan di empat lokasi penelitian pada bulan Oktober-Desember tahun 2004 dan bulan Februari 2005.
KESIMPULAN
a. Cibinong dengan kondisi lingkungan penggunaan lahan daerah industri, transportasi dan permukiman, memiliki konsentrasi SO4
2-, NO3-, dan NH3 air hujan yang lebih
tinggi daripada Ciawi, daerah sekitar Kebun Raya dan Megamendung. Megamendung dengan penggunaan lahan hutan tanaman, dan perkebunan memiliki konsentrasi polutan di dalam air hujan paling rendah. Kawasan padat transportasi (Ciawi) memiliki konsentrasi CO2 paling tinggi yang dihasilkan dari sisa pembakaran bahan bakar kendaraan bermotor. Nilai pH rata-rata paling rendah dihasilkan oleh kawasan industri (Cibinong) dan di kawasan ini telah terjadi hujan asam, sedangkan pH tertinggi dihasilkan di kawasan hutan tanaman.
b. Ion nitrat (NO3-) lebih berpengaruh dalam menurunkan
pH air hujan dibandingkan ion sulfat (SO42-) Cl- dan
CO2. c. Konsentrasi polutan didalam air hujan pada awal musim
hujan (bulan Oktober-November) lebih tinggi daripada
pertengahan musim hujan (bulan Desember-Februari), sedangkan pH rata-rata air hujan pada awal musim hujan cenderung lebih rendah daripada pertengahan musim hujan.
DAFTAR PUSTAKA
Fakuara, M.Y. 1986. Beberapa usaha mengatasi dampak hujan Asam. Media Konservasi. Jurusan Konservasi Sumberdaya Hutan. Bogor. Fakultas Kehutanan. IPB. Vol. 1. No. 2.
Harmantyo, D. 1989. Studi tentang hujan masam di Wilayah Jakarta dan Sekitarnya. Disertasi. Bogor. Program Pasca Sarjana. IPB. Bogor.
Husin, Y. A, H. Suharsono, dan S. Effendy. 1991. Studi tingkat pencemaran udara dan hujan asam di daerah Bogor. Laporan Penelitian. Bogor. LP-PPLH-IPB.
Soedomo, M. 2001. Kumpulan karya ilmiah pencemaran udara. Bandung. Penerbit ITB.
5.44 5.796.53 6.435.97 6.25 6.7
7.17
5.985.995.615.28345678
Cibinong Ciawi KRB Megamendung
Lokasi
pH a
ir hu
jan
Oktober Desember Februari

Pola Reproduksi Burung Tekukur
80
POLA REPRODUKSI BURUNG TEKUKUR (Streptopelia chinensis) DAN PUTER (Streptopelia risoria) DI PENANGKARAN
(Reproductive Pattern of Spotted Dove (Streptopelia chinensis) and Ringdove (Streptopelia risoria) in Captivity)
BURHANUDDIN MASYUD
Laboratorium Konservasi Eksitu/Penangkaran Satwaliar, Departemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan IPB, Kampus IPB Darmaga Bogor 16680 Indonesia
Email: [email protected]
Diterima 16 April 2007 / Disetujui 18 Juni 2007
ABSTRACT
This study was carried out to know reproductive pattern of spotted dove (Streptopelia chinensis) and ringdove (Streptopelia risoria) in captivity. Fifteen pairs of spotted dove and seventeen pairs of ringdove were used to identify reproductive pattern. Each pairs of the birds were cared in cage (1.5 x 0.75 x 2.0 m). Result of this research showed that both types of the birds had the same pattern of reproduction, i.e : age of puberty (first mating) or onset of first egg production was 6.5 -7 months, avarage clutch size was two eggs, hatching period was 14.47 ± 0.74 days, egg production period was 27.11 ± 6.72 days to 48.79 ± 3.53 days, and hatching rate was 55% to 68%. The length of sperm head was 13.00 ± 0.62 µm to 14.73 ± 0.46 µm and the width of sperm head was 1.00 µm and the length of sperm tail was 87.00 ± 9.59 µm to 104.00 ± 6.22 µm. Sperm motility and concentration were relatively the same i.e 50 – 60% and 270 x 106 to 710 x 106 respectively. The birds were identified as temporal monogamous mating type. Key words: Streptopelia chinensis, Streptopelia risoria, reproductive pattern, matting type, captivity.
PENDAHULUAN
Burung tekukur (Streptopelia chinensis) dan burung puter (Streptopelia risoria) merupakan dua diantara aneka jenis burung bernyanyi yang banyak digemari. Dewasa ini usaha untuk menyilangkan kedua jenis burung ini terus dikembangkan terutama untuk mendapatkan keturunan dengan kualitas suara yang bagus.
Keberhasilan reproduksi dan/atau penyilangan kedua burung ini di penangkaran sangat berkaitan erat dengan pengetahuan dasar tentang pola reproroduksinya. Karena informasi ini penting sebagai acuan didalam pengaturan program reproduksi dan penyilangannya. Selain itu, informasi ini juga mempunyai arti penting bagi pengembangan ilmu pengetahuan dan teknologi.
Berdasarkan hal tersebut, penelitian ini dilakukan dengan tujuan, untuk mempelajari anatomi reproduksi dan pola reproduksi dari burung tekukur dan burung puter di penangkaran.
METODE PENELITIAN
Penelitian dilakukan di Laboratorium Penangkaran Satwaliar, Departemen Konservasi Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan IPB. Penelitian berlangsung
selama kurang lebih sepuluh bulan, April 2002 sampai Februari 2003.
Penelitian menggunakan masing-masing 15 pasang burung tekukur dan 17 pasang burung puter. Semua hewan percobaan dipelihara dalam kandang penangkaran (1,5 x 0,75 x 2,0 m) berlantai campuran pasir dan sekam padi, dengan kondisi cahaya alami (12 T/12G). Hewan percobaan diberi pakan yang diramu sendiri terdiri atas gabah padi, jagung kuning (giling), beras merah, ketan hitam, kacang hijau, millet merah, millet putih, dengan kadar protein sekitar 12%. Air minum diberikan ad libitum. Untuk keperluan penganalisaaan sperma digunakan NaCl fisiologis sebagai bahan pengencer semen. Umur burung puter bervariasi, mulai satu minggu (sejak menetas) sampai berumur sekitar tiga sampai empat bulan (remaja). Sedangkan burung tekukur berumur sekitar satu sampai dua bulan, dan umumnya merupakan burung-burung hasil tangkapan langsung dari alam.
Peubah yang ditelaah untuk menggambarkan pola reproduksi burung, meliputi : (1) anatomi reproduksi burung; (2) umur pertama kali kawin dan/atau bertelur, (3) spermatozoa burung, (4) telur burung, (5) clutch size, masa inkubasi dan daya tetas telur, (6) jarak waktu antar dua periode bertelur, dan (7) perilaku seksual. Prosedur dan cara kerja dalam penelaahan masing-masing peubah, sebagai berikut:

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 80 – 88
81
Anatomi Reproduksi Burung. Anatomi saluran reproduksi burung yang diamati meliputi saluran reproduksi burung jantan dan betina. Peubah yang ditelaah meliputi (a) panjang saluran reproduksi burung jantan (vas deferens) dan betina (uterus atau kelenjar kerabang), (b) testes (bentuk, warna, berat, panjang dan lebar testes kiri dan kanan), (c) ovarium dan folikel (bentuk, jumlah dan ukuran folikel). Pengamatan dan pengukuran dilakukan setelah burung dimatikan, dibedah dan anatomi serta saluran reproduksi dikeluarkan. Data pengukuran yang terkumpul ditabulasi kemudian dihitung nilai rataan dan simpangan baku.
Umur Pertama Kali Kawin dan/atau Bertelur. Umur mulai pertama kali kawin dihitung saat hari pertama kali terlihat pasangan burung mulai memperlihatkan tanda-tanda kawin yang dimulai dengan perilaku seksual sampai terjadi penunggangan dan kopulasi. Sedangkan umur pertama kali bertelur dihitung saat hari pertama kali burung bertelur. Umur mulai kawin dan bertelur ini dijadikan sebagai indikator dewasa kelamin.
Spermatozoa Burung. Pengamatan spermatozoa burung dilakukan dengan cara pengambilan contoh semen dari saluran reproduksi (epididimis dan vas deferens) segera setelah burung-burung contoh dimatikan. Epididimis dan vas deferens dipisahkan dari testes, kemudian bagian saluran reproduksi ini digerus dan dicampur dengan pengencer NaCl fisiologis sebanyak 1 ml. Selanjutnya dibuatkan preparat ulas dan diamati morfometrik serta diambil gambarnya. Peubah morfologi spermatozoa yang ditelaah meliputi bentuk dan ukuran spermatozoa (panjang kepala, lebar kepala, panjang ekor dan perbandingan panjang kepala dan panjang ekor); juga dihitung persentase motilitas dan konsentrasi sperma (Toelihere 1985). Jumlah hewan contoh yang digunakan masing-masing 7 ekor burung tekukur dan 7 ekor burung puter.
Telur Burung. Peubah yang ditelaah meliputi bentuk, warna dan ukuran telur (bobot, panjang dan lebar telur, berat/persentase dari bagian-bagian telur yakni kerabang telur, kuning telur dan putih telur). Jumlah telur yang digunakan masing-masing 15 butir telur untuk tekukur dan 30 butir telur untuk puter.
Clutch Size, Masa Inkubasi dan Daya Tetas Telur. Clutch size atau jumlah telur per sarang adalah banyaknya telur (butir) yang dihasilkan secara berturutan dalam satu masa bertelur (reproduksi). Masa inkubasi atau lama pengeraman telur adalah lamanya waktu mulai telur dierami sampai menetas. Sedangkan daya tetas telur adalah banyaknya telur yang menetas dari jumlah telur yang dierami dikalikan 100%.
Jarak Waktu Bertelur. Jarak waktu bertelur adalah banyaknya hari antara dua periode bertelur atau antara dua
clutch. Jarak waktu ini diukur dengan menghitung banyaknya hari dimulai saat telur pertama dihasilkan secara berurutan pada satu clutch (sarang) sampai dengan hari dimana telur pertama yang dihasilkan pada clutch (sarang) berikutnya. Jarak ini ditelaah pada dua keadaan, yakni: (a) keadaan normal yaitu mulai telur dierami, menetas sampai anak disapih, dan (b) keadaan tidak normal, yaitu keadaan dimana telur tidak menetas lalu diambil (karena infertil) atau pecah.
Perilaku Seksual (sexual behaviour). Perilaku seksual yang diamati meliputi perilaku kawin, perilaku bersarang dan perilaku mengerami telur. Perilaku ini diamati secara kasuistik pada pasangan burung yang memperlihatkan tanda-tanda kawin, bersarang dan mengerami telur. Metode pengambilan data yang digunakan terdiri dari dua metode, yakni: (1) Metode Time Sampling, yaitu mencatat perilaku state (perilaku yang berlangsung cukup lama yakni perilaku bersarang dan mengerami telur) yang terjadi pada setiap interval dua menit setiap jam; (2) Metode Continous Sampling yaitu mencatat frekuensi terjadinya perilaku event (event behaviour), yaitu perilaku yang kejadiannya singkat (perilaku kawin), pada selang waktu satu jam, selama 6 jam setiap hari, mulai pukul 06.00-07.00, 07.00-08.00 dan seterusnya sampai pukul 16.00-17.00, kemudian dilanjutkan pada hari berikutnya mulai pukul 07.00-08.00, 08.00-09.00 dan seterusnya sampai pukul 17.00-18.00. Pengamatan dilakukan selama 14 hari. Data yang terkumpul dianalisis dan disajikan secara deskriptif untuk menggambarkan pola perilaku seksual dari kedua burung tersebut.
HASIL DAN PEMBAHASAN
Anatomi Organ Reproduksi Burung
Burung Jantan Berdasarkan hasil pembedahan terhadap contoh
burung untuk menelaah anatomi reproduksi burung, diperoleh gambaran adanya variasi ukuran, bentuk dan warna anatomi organ reproduksi antara burung tekukur jantan dan burung puter jantan (Tabel 1, Gambar 1).
Ukuran (panjang, lebar dan berat) testes kiri relatif lebih besar dibanding dengan testes kanan, baik pada burung tekukur maupun burung puter. Meskipun ukuran testes kiri lebih besar daripada testes kanan, namun hasil analisis perbandingan rata-rata antara testes kiri dan testes kanan ternyata tidak berbeda nyata (P > 0.05). Etches (1996) mengemukakan bahwa pada burung biasanya testes kiri lebih besar 0.5 – 3 gram daripada testes kanan.

Pola Reproduksi Burung Tekukur
82
Tabel 1. Ukuran, bentuk dan warna anatomi reproduksi burung jantan dari burung tekukur (Streptopelia chinensis) dan burung puter (streptopelia risoria)
No Anatomi Reproduksi Jantan Burung Tekukur (n=7)
Burung Puter (n=9)
1. Berat testes kanan (g) 0.31 ± 0.04 0.36 ± 0.09 2. Berat testes kiri (g) 0.43 ± 0.06 0.44 ± 0.04 3. Panjang testes kanan (mm) 9.71 ± 3.45 14.78 ± 3.49 4. Panjang testes kiri (mm) 11.43 ± 3.69 17.22 ± 3.93 5. Lebar testes kanan (mm) 4.86 ± 1.07 5.44 ± 1.33 6. Lebar testes kiri (mm) 5.86 ± 1.07 6.33 ± 1.50 7. Panjang epididimis (mm) 12.10 ± 4.18 13.80 ± 2.11 8. Panjang vas deferens (mm) 34.30 ± 3.99 37.10 ± 1.05 9. Bentuk testes Bulat kacang kedelai Bulat kacang kedelai
10. Warna testes Putih kekuningan sampai coklat kehitaman
Putih kekuningan sampai coklat kehitaman
Gambar 1. Anatomi reproduksi burung jantan tekukur dan puter (1) testis; (2) epididimis; (3) vas
deferens (C = mm)
Ukuran berat testes kedua burung ini jauh lebih kecil
bila dibandingkan dengan berat testes bangsa unggas lainnya. Bahr dan Bakst (1987) menyatakan bahwa pada burung berat testes antara 14-60 gram tergantung jenis burung. Etches (1996) mengemukakan bahwa pada masa dewasa kelamin ukuran berat testes biasanya meningkat dari 2-4 gram menjadi 25-35 gram.
Burung Betina
Hasil telaahan hewan contoh menunjukkan bahwa ukuran, bentuk dan warna anatomi reproduksi pada burung tekukur dan burung puter betina menunjukkan adanya variasi (Tabel 2). Ukuran anatomi organ reproduksi burung tekukur betina relatif lebih kecil daripada burung puter,
meskipun secara statistik tidak berbeda nyata (P > 0.05). Seperti diketahui, pada burung hanya satu ovarium dan saluran reproduksi (uterus) yang aktif (sebelah kiri), sedang ovarium sebelah kanan mengalami degenerasi (Parker, 1969; Sturkie, 1970; Bahr dan Bakst, 1987; Etches, 1996).
Ovarium pada burung ini berupa rangkaian folikel yang tersusun seperti buah murbei dengan jumlah rata-rata bervariasi, masing-masing pada burung tekukur sebanyak 9.71 ± 1.38 folikel dan burung puter 10.44 ± 1.33 folikel, dengan rata-rata ukuran diameter setiap folikel masing-masing burung tekukur 2.71 ± 0.49 mm dan burung puter 2.78 ± 0.44 mm. Berat ovarium bervariasi dan relatif berbeda antara burung tekukur dan burung puter.
a b
1
1 1 2
3

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 80 – 88
83
Tabel 2. Ukuran anatomi organ reproduksi burung betina dari burung tekukur dan burung puter
No Anatomi Organ Reproduksi Betina Burung Tekukur (n = 7)
Burung Puter (n = 7)
1. Berat ovarium (g) 0.31 ± 0.02 0.32 ± 0.01 2. Panjang ovarium (mm) 8.79 ± 1.35 16.11 ± 0.93 3. Lebar ovarium (mm) 57 ± 0.03 6.00 ± 1.00 4. Panjang uterus (mm) 71.41 ± 6.11 75.67 ± 1.00 5. Diameter folikel (mm) 2.71 ± 0.49 2.78 ± 0.44 6. Jumlah folikel (butir) 9.71 ± 1.38 10.44 ± 1.33 7. Bentuk ovarium Rangkaian sel telur (folikel)
seperti buah murbei Rangkaian sel telur (folikel)
seperti buah murbei 8. Warna ovarium Putih sampai kekuningan Putih sampai kekuningan
Perbedaan ini juga terjadi karena berbeda umur. Berat
ovarium pada burung tekukur muda adalah 0.09 ± 0.03 gram dan pada burung tekukur dewasa adalah 0.31 ± 0.01 gram. Sedangkan pada burung puter muda berat ovariumnya 0.15 ± 0.01 gram dan pada burung dewasa beratnya 0.32 ± 0.02 gram. Variasi jumlah dan diameter folikel serta berat ovarium (indung telur) pada kedua burung ini dipengaruhi oleh beberapa faktor diantaranya umur dan status reproduksi burung. Burung yang berumur lebih tua relatif memiliki jumlah folikel lebih banyak, ukuran setiap folikel lebih besar, serta berat ovarium yang lebih besar. Burung-burung yang sedang aktif bereproduksi atau memasuki masa reproduksi, juga memiliki berat ovarium dan ukuran serta diameter folikel telur yang lebih banyak dan besar. Ukuran diameter folikel dan berat ovarium pada kedua burung ini jauh lebih kecil bila dibandingkan dengan unggas lain. Sebagai contoh diameter folikel pada ayam mencapai 35 mm dengan berat ovarium pada ayam dewasa yang sedang aktif bertelur mencapai 40-60 gram dan pada kalkun sekitar 125-200 gram (Toelihere, 1985). Jelas bahwa ukuran folikel pada burung ditentukan oleh beberapa faktor diantaranya status reproduksi dan umur burung.
Pola Reproduksi Burung
Umur Kawin atau Bertelur Pertama Kali
Hasil pengamatan terhadap contoh burung menunjukkan bahwa umur kawin dan/atau umur bertelur pertama kali antara burung tekukur dan puter relatif sama. Untuk burung tekukur jantan 6,8 + 0,13 bulan dan burung betina 6,9 + 0,19 bulan, sedangkan burung puter jantan 6,6 + 0,13 bulan dan burung betina 6,8 + 0,07 bulan.
Tanda-tanda mulai kawin pertama kali biasanya didahului oleh perilaku membentuk pasangan, membuat sarang dengan mencari dan membawa bahan sarang berupa rumput-rumput atau ranting kecil ke dalam sarang. Burung jantan mulai mengeluarkan bunyi untuk menarik pasangannya dan frekuensi keluar masuk sarang meningkat sejalan dengan mendekatnya waktu kawin.
Berdasarkan pengamatan terhadap ukuran anatomi reproduksinya, khususnya ukuran testes dan ovarium, diperoleh gambaran bahwa ukuran testes dan ovarium pada burung tekukur dan puter yang baru pertama kali kawin ternyata relatif lebih kecil daripada burung dewasa. Dari pengukuran ukuran testes burung contoh diperoleh berat testes burung yang baru pertama kali kawin sebesar 0,01-0,03 gram dengan panjang 4-7 mm dan lebar 3-3,5 mm. Sedangkan ukuran ovarium, berat 0,05-0,07 gram dengan panjang 11-13 mm dan lebar 3-5 mm berupa rangkaian sel telur (buah murbei). Jumlah untaian folikel berkisar 8-10 butir dengan ukuran diameter folikel juga masih relatif kecil (1-3 mm).
Perkembangan organ reproduksi burung untuk mencapai tahap fungsional yang ditandai oleh adanya aktivitas perkawinan dan produksi sperma dan sel telur dipengaruhi oleh banyak faktor baik internal maupun eksternal. Faktor internal yang penting adalah rangsangan hormon (FSH & LH) disamping kesiapan organ reproduksi betina yang secara tidak langsung memberikan andil dalam kerja hormon FSH dalam proses pematangan folikel ataupun hormon LH dalam proses spermatogenesis. Sedangkan faktor eksternal antara lain adalah faktor asupan pakan dengan kualitas dan keseimbangan gizi yang cukup (Parker, 1969; Toelihere, 1985; Grimes, 1994; Etches, 1996). Nur (2001) melaporkan bahwa burung puyuh yang memperoleh asupan vitamin E dengan dosis lebih tinggi (50 IU) ternyata lebih awal mencapai usia pertama kali bertelur yakni 47,58 hari dibanding burung yang memperoleh asupan vitamin E dengan dosis lebih rendah (25 IU) yakni pada usia 51,17 hari.
Spermatozoa
Hasil pengukuran spermatozoa dari hewan-hewan contoh menunjukkan bahwa kedua jenis burung ini memiliki ukuran, bentuk dan konsentrasi serta motilitas sperma yang relatif sama. Rataan panjang kepala sperma 14 µm dan lebar kepala sperma 1.00 µm dengan perbandingan panjang dan lebar kepala sperma sekitar 6-7:1. Konsentrasi

Pola Reproduksi Burung Tekukur
84
sperma berkisar 270 x106 sampai 710 x 106 sperma per ml, dan motilitas sperma berkisar 50-60%. Sebagai perbandingan dapat dilihat persen sperma hidup dan konsentrasi sperma dari burung bangau seperti yang dilaporkan Gee dan Temple (1978) bahwa dari hasil koleksi semen burung bangau yang ditangkarkan (dengan cara masage), ternyata untuk pejantan produktif diperoleh jumlah spermatozoa hidup sebanyak 90% dengan konsentrasi spermatozoa mencapai 360 x 106 sperma per ml semen.
Telur Burung
Ukuran, Bentuk dan Warna Telur
Hasil pengukuran terhadap contoh telur burung tekukur dan puter menunjukkan bahwa ukuran telur burung puter relatif lebih besar dibanding ukuran telur burung tekukur, akan tetapi hasil analisis perbandingan rataan ukuran telur antara kedua burung ini tidak berbeda nyata (P>0.05) (Tabel 3).
Tabel 3. Ukuran telur burung tekukur (Streptopelia chinensis) dan burung puter (Streptopelia risoria)
Ukuran Telur Burung Tekukur
(n=10) Burung Puter
(n =17)
Berat (gram) 5.81 ± 0.57a 6.57 ± 0.61a Panjang (mm) 26.78 ± 0.74a 28.47 ± 1.37a Lebar (mm) 21.10 ± 1.37a 22.24 ± 1.03a Berat cangkang (gram) 0.63 ± 0.10a 0.83 ± 0.40a Berat putih telur (gram) 3.27 ± 0.46a 3.43 ± 0.41a Berat kuning telur (gram) 2.01 ± 0.42a 2.29 ± 0.33a Tebal kulit cangkang (mm) 0.61 ± 0.09a 0.68 ± 0.09a
Keterangan: Huruf yang sama pada angka dari baris yang sama tidak berbeda nyata (P>0.05)
Secara umum ukuran panjang dan lebar telur kedua burung ini tidak jauh berbeda dengan telur burung puyuh (Nur 2001) ataupun jalak bali (Masy’ud 1992) namun berat telur kedua burung ini yakni telur tekukur (5.81 ± 0.57 gram) dan telur puter (6.57 ± 0.61 gram) sedikit lebih kecil jika dibandingkan dengan berat telur puyuh 8.0.67 – 9.0 gram (Nur 2001), dan berat telur burung jalak bali (Leucopsar rotschildii) 7.9 ± 0.1 gram (Masy’ud 1992).
Adapun bentuk telur burung tekukur dan puter relatif sama, yakni berbentuk oval (bulat panjang) dengan warna putih terang sampai putih kekuning-kuningan. Untuk telur-telur yang tidak menetas, hasil pengamatan menunjukkan bahwa ukuran telur umumnya lebih ringan, lebih kecil dengan warna lebih pucat.
Jumlah Telur Per Sarang (Clutch Size)
Hasil pengamatan terhadap jumlah telur dalam satu irama bertelur (ukuran sarang – clutch size) antara burung tekukur dan puter relatif sama yakni 1.70 ± 0.48 (1-2) butir untuk burung tekukur dan 2.07 ± 0.59 (1-3) butir untuk burung puter. Hasil yang relatif sama juga pernah dilaporkan oleh beberapa penulis seperti Zaini et al. (1997), Soemarjoto dan Raharjo (2000), Soejoedono (2001), Ehrlich (2004 a, b).
Ada beberapa faktor yang diduga berpengaruh terhadap jumlah telur dalam satu irama bertelur (clutch size), diantaranya umur burung, berat badan, makanan, kondisi kesehatan dan lingkungan kandang (luas, suhu dan kelembaban serta ada tidaknya gangguan) (Parker 1969; Etches 1996). Nalbandov (1990) mengemukakan bahwa variasi jumlah telur yang dihasilkan burung dalam satu masa irama bertelur juga dipengaruhi oleh susunan genetik kelenjar pituitari, terutama pada jumlah gonadotropin yang dihasilkannya. Pada percobaan yang dilakukan pada ayam yang diberikan preparat gonadotropin ternyata dapat merubah irama bertelur ayam dari dua atau tiga butir menjadi tiga sampai empat butir telur setiap ritmenya. Menurut Short (1993), jumlah telur yang dihasilkan suatu jenis burung dalam suatu irama bertelur (clutch size) ditentukan oleh seleksi alam dari berbagai faktor yang berkaitan dengan kehidupan burung. Ada tiga faktor utama yang diketahui berpengaruh terhadap evolusi yang menentukan ukuran sarang (clutch size), meskipun ada peluang individual dan variasi geografik yang terjadi. Ketiga faktor tersebut adalah : Pertama, kemampuan induk bekerja dengan kapasitas maksimum dalam membangun sarang dan menyiapkan makanan untuk anaknya setiap hari. Makin banyak telur yang dihasilkan maka makin besar usaha yang harus dilakukan induk untuk memelihara telur

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 80 – 88
85
tersebut. Kedua, besarnya peluang telur menjadi anak dan bertahan secara relatif tidak diganggu predator. Makin kecil peluang hidup dan makin besar gangguan predator maka jumlah telur yang dihasilkan cenderung lebih banyak. Ketiga, daya hidup induk burung untuk memelihara dirinya dan anaknya. Dikatakan pula bahwa ukuran sarang (clutch size) juga ditentukan oleh metode burung mencari makan. Secara umum burung-burung yang dapat mencari makan sepanjang hari namun cenderung lebih suka makan selama jam-jam terang cahaya pada pagi dan malam hari, biasanya bertelur dua butir. Letak lintang (latitude) dimana burung itu biasa bertelur dan membuat sarangnya juga menyebabkan perbedaan clutch size. Umumnya burung-burung yang kawin di daerah iklim dingin (temparate) di sebelah utara dan selatan tropis bertelur lebih banyak daripada burung-burung di daerah tropis. Short (1993) juga menyatakan satu teori yang dipercaya bahwa ukuran sarang dari suatu jenis burung berhubungan dengan kelimpahan makanan untuk burung tersebut. Dalam hal ini untuk jenis-jenis burung di daerah tropis dimana kelimpahan makanan stabil sepanjang tahun dan jumlah hari cahaya lebih baik sepanjang tahun, sehingga lebih banyak waktu untuk makan setiap hari, maka perkembangbiakannya cenderung menyebar sepanjang tahun. Akibatnya jumlah anak yang lebih kecil setiap masa reproduksinya lebih disukai yakni sekitar dua butir (ekor). Keadaan ini tampaknya juga berlaku pada burung tekukur dan burung puter yang lebih menunjukkan dominasi aktivitasnya pada siang hari, sehingga jumlah telur yang dihasilkan setiap kali bertelur adalah dua butir.
Menurut Sibley dan Ahlquist (1990), ada beberapa peubah demografi yang berhubungan seperti hubungan antara usia hidup (longevity) dengan ukuran sarang (clutch size). Umumnya burung-burung yang kawin pertama kali pada usia satu tahun atau burung-burung yang berumur pendek, bertelur lebih dari dua butir per sarang dan memiliki lebih dari satu sarang per tahun, dengan daya hidup telur dan anak relatif rendah. Sebaliknya burung-burung yang usia kawin pertamanya lebih lambat sampai empat tahun atau lebih, cenderung bertelur satu atau dua butir telur per sarang dan hanya satu kali dalam satu tahun, dan mempunyai daya tahan hidup yang lebih tinggi dan berumur panjang. Hal ini tampaknya juga berlaku pada burung tekukur dan puter yang memasuki kawin pertama pada usia satu tahun atau kurang dengan jumlah telur per sarang rata-rata dua butir dan memiliki beberapa sarang (dapat bertelur beberapa kali) dalam satu tahun.
Lama Pengeraman Telur (Inkubasi)
Hasil pengamatan lama pengeraman telur terhadap beberapa sarang diperoleh gambaran bahwa lama pengeraman telur pada burung tekukur dan burung puter relatif sama yakni sekitar 14 hari masing-masing 14.50 ±
0.76 hari untuk burung tekukur dan 14.47 ± 0.74 hari untuk burung puter.
Hasil uji perbandingan nilai rataan lama pengeraman telur antara kedua jenis burung ini ternyata tidak berbeda nyata (P > 0.05). Ada beberapa faktor yang diketahui berpengaruh terhadap lama masa pengeraman telur burung, diantaranya faktor suhu dan kelembaban. Menurut Short (1993), pada musim panas dimana suhu relatif lebih tinggi untuk daerah-daerah panas, periode pengeraman telur lebih pendek. Sebaliknya pada musim dingin atau daerah dimana suhu lebih rendah (dingin) maka lama waktu pengeraman telur relatif lebih lama. Disamping faktor suhu, variasi kebutuhan induk untuk makan (mencari makan) dan mempertahankan diri juga berpengaruh secara nyata terhadap variasi lama pengeraman telur. Meskipun ada pengecualian namun secara umum telah diketahui biasanya jenis burung dengan telur berukuran lebih kecil dan ukuran sarangnya (clutch size) kecil mempunyai masa pengeraman telur lebih pendek yakni sekitar 11 hari.
Berdasarkan pandangan tersebut, dalam banyak laporan diketahui bahwa jenis-jenis burung dengan ukuran dan jumlah telur per sarang seperti halnya burung tekukur dan puter yakni dua butir, mempunyai rata-rata lama pengeraman telur kurang lebih 14 hari, seperti terlihat pada burung jalak bali –Leucopsar rotschildii (Masyud, 1992), beo nias – Gracula religiosa robusta (Thohari, 1998), Cucak rawa (Ardhani, 1998), perkutut (Sarwono, 2000). Adanya variasi satu sampai dua hari dalam waktu lama pengeraman telur antara lain disebabkan adanya variasi suhu dan kelembaban lingkungan. Umumnya hasil pengamatan menunjukkan bahwa pada musim penghujan, masa pengeraman telur relatif lebih lama satu sampai dua hari dibanding pada musim panas.
Tugas Pengeraman Telur
Hasil pengamatan menunjukkan bahwa tugas pengeraman telur terutama dilakukan oleh induk betina dengan alokasi waktu sekitar lebih 95%. Induk jantan juga terlihat ikut dalam pengeraman telur terutama menggantikan peran betina pada saat-saat induk betina makan dan menggerakkan otot tubuh dengan cara mengepakkan sayapnya. Frekuensi penggantian peran pengeraman telur ini sekitar dua sampai tiga kali per hari, yakni pada pagi hari (08.00 – 10.00 WIB), siang hari (12.00 – 14.00) dan sore hari (16.00 – 18.00), antara lain sangat ditentukan oleh kondisi suhu lingkungan. Jika suhu lingkungan rendah, maka frekuensi penggantian lebih sedikit, sebaliknya jika suhu lingkungan tinggi maka frekuensinya lebih sering mencapai 2-3 kali per hari. Lama penggantian tugas pengeraman telur tersebut hanya berlangsung sekitar 5-10 menit, yakni waktu yang diperlukan oleh induk betina untuk makan dan minum serta mengepakkan sayap untuk pergerakan tubuh (exersice).

Pola Reproduksi Burung Tekukur
86
Menurut Short (1993), biasanya induk betina yang bertanggungjawab dalam proses pengeraman telur. Meskipun demikian, pada beberapa jenis burung, induk jantan juga ikut mengerami telur. Namun pada jenis-jenis passerina atau burung-burung bertengger, induk jantan hanya berfungsi dalam melindungi telur tetapi tidak memiliki kemampuan untuk mengerami telur.
Berdasarkan pandangan tersebut, tugas pengeraman telur pada burung tekukur dan burung puter lebih utama dilakukan oleh induk betina sementara induk jantan lebih bertugas melindungi atau menjaga agar kondisi suhu pada sarang relatif tetap selama induk betina meninggalkan sarang untuk makan dan minum serta pergerakan tubuh (exersice).
Jarak Waktu Periode Bertelur (Nest Period)
Hasil pengamatan terhadap jarak waktu antara dua masa bertelur (periode bertelur) pada burung tekukur dan burung puter memperlihatkan ada dua kondisi yang dapat dibedakan untuk menghitung jarak waktu bertelur, yakni (1) kondisi normal, artinya pada keadaan mulai bertelur, mengeram, menetas sampai anak disapih (alamiah), dan (2) kondisi tidak normal, yakni pada keadaan dimana telur busuk, tidak menetas dan pecah.
Jarak waktu bertelur pada keadaan normal, masing-masing pada burung tekukur adalah 48,79 ± 3.53 hari relatif lebih lama dibanding pada burung puter yakni 43,22 ± 1,39 hari. Sedangkan pada keadaan tidak normal jarak waktu bertelur relatif lebih cepat baik pada burung tekukur (31,22 ± 5,63 hari) maupun pada burung puter (27,11 ± 6,72 hari).
Adanya variasi individual dalam hal jarak waktu bertelur (nest period) pada keadaan normal antara lain berhubungan dengan jumlah anak per sarang (brood size). Pada induk-induk burung dengan jumlah anak lebih banyak (dua ekor) maka jarak waktu bertelur antar dua periode bertelur relatif lebih lama dibanding induk burung dengan jumlah anak lebih kecil (satu ekor). Sedangkan pada keadaan tidak normal dimana telur pecah, busuk atau tidak menetas, maka jarak waktu bertelur pada burung tekukur dan burung puter menjadi lebih singkat yakni kurang dari 40 hari. Pada pengamatan yang lebih spesifik diperoleh gambaran secara umum bahwa pada keadaan telur tidak menetas, burung akan segera kembali bertelur jika telur yang tidak menetas itu segera diambil. Artinya setelah melewati hari ke-16 sampai hari ke-18 dari masa pengeramannya, telur yang tidak menetas itu segera diambil, maka dalam waktu singkat sekitar 7-14 hari setelah telur diambil induk burung akan segera bertelur kembali.
Berdasarkan kondisi jarak bertelur seperti itu, maka dalam keadaan normal seekor induk burung tekukur atau burung puter dalam satu tahun dapat bertelur empat sampai enam kali. Ini berarti bahwa di penangkaran pola reproduksi (bertelur) burung tekukur dan burung puter dapat
berlangsung sepanjang tahun, berbeda dengan di alam bebas yang cenderung hanya berlangsung pada bulan-bulan tertentu saja atau lebih dikenal sebagai hewan pekawin bermusim (seasonal breeder). Salah satu faktor yang kuat berpengaruh terhadap perubahan pola reproduksi antara di alam bebas dengan di penangkaran adalah faktor makanan terutama yang berkaitan dengan kontinuitas ketersediaan pakan (energi) untuk memenuhi kebutuhan reproduksinya. Ruang gerak yang terbatas dalam seluruh aktivitas burung di penangkaran juga membawa implikasi pada efisiensi pemanfaatan energi yang relatif tinggi, sehingga ketersediaan energi tersebut selain untuk memenuhi kebutuhan hidup pokok secara optimal juga dapat digunakan untuk menunjang proses reproduksi.
Daya Tetas Telur
Hasil pengamatan terhadap tujuh sarang dari masing-masing pasang burung diperoleh gambaran daya tetas telur pada burung tekukur lebih rendah (35.71%) daripada burung puter (72.41%).
Diantara faktor yang diduga berpengaruh terhadap daya tetas telur adalah umur induk, suhu dan kelembaban kandang dan kualitas pakan (Kosin, 1969; Etches, 1996). Selain itu tingkat gangguan lingkungan kandang juga sangat berpengaruh terhadap daya tetas telur, terutama untuk pasangan burung tekukur. Burung tekukur jika dibanding dengan burung puter maka sifat liarnya masih relatif tinggi sehingga kepekaannya terhadap gangguan faktor lingkungan masih sangat tinggi. Dalam pengamatan diketahui bahwa jika ada gangguan maka cenderung induk betina tekukur yang sedang mengerami telur akan meninggalkan telurnya bahkan seringkali telurnya dimakan atau dipecahkan. Berbeda halnya dengan induk burung puter yang tampak lebih jinak dan adaptif dengan setiap perubahan atau gangguan dari lingkungannya, sehingga menunjukkan performans reproduksi yang lebih baik.
Perilaku Kawin
Hasil pengamatan terhadap perilaku kawin pada burung tekukur dan burung puter, dari keseluruhan rangkaian perilaku (Alcock, 1989) maka dapat dibedakan ke dalam tiga tahap (fase), yakni pra-kopulasi, kopulasi dan pasca kopulasi.
(1) Perilaku Pra Kopulasi
Perilaku pra kopulasi adalah perilaku yang dilakukan sebelum kopulasi. Tujuan perilaku ini adalah untuk menarik pasangannya agar siap/mau melakukan kopulasi. Perilaku menarik pasangan ini biasanya dilakukan oleh pejantan, yakni dengan cara bersuara secara berulang (degku.. kuukkur.… untuk tekukur dan kukkuurr…. kukkuurr… kukkuur untuk puter) sambil mengangguk-anggukkan

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 80 – 88
87
(menggerakan) kepala lalu bergerak mendekati betina, mematuk-matuk atau menyelisik bulu. Suara yang dikeluarkan bersifat khas dan lazim dikenal sebagai suara seksual (sexual calling – sexual vocal). Perilaku pra kopulasi pada burung jantan juga ditunjukkan dengan aktivitas menyiapkan sarang, yakni dengan sering keluar masuk sarang sambil membawa rumput atau ranting-ranting kecil ke dalam atau keluar sarang, diam sesaat di dalam sarang sambil mengeluarkan suara – sex calling. Pada burung betina yang belum siap secara fisiologis biasanya selalu terbang menghindar/menjauh jika didekati atau dicumbu oleh pejantan. Perilaku pra kopulasi ini dapat berlangsung singkat (beberapa jam) sampai beberapa hari (2–3 hari) bahkan kadang-kadang mencapai satu minggu atau lebih, tergantung tingkat kematangan dan kesiapan fisiologis dari burung betina. Pada betina yang terlihat mulai cocok dan siap kawin, tampak diam jika pejantan mulai mendekati, mencumbui dan belajar menungganginya, serta memberikan respons siap dikawini. Frekuensi penunggangan bisa terjadi beberapa kali (3 – 4 kali bahkan lebih). Betina yang sudah siap juga tampak intensif keluar masuk sarang atau belajar diam sesaat di dalam sarang untuk mengenal sarangnya sambil mengeluarkan suara khas …degkku…ku..kuuu (tekukur) atau kruk kukuu… kuuu…. Krukkuu….kuuu.
Keseluruhan rangkaian perilaku pra kopulasi tersebut pada dasarnya bertujuan untuk mempertinggi efektivitas pertemuan sperma dan sel telur atau memungkinkan agar perkawinan yang terjadi dapat berhasil dan efektif menghasilkan keturunan. Jadi perilaku pra kopulasi pada dasarnya berfungsi sebagai proses sinkronisasi kondisi fisiologis diantara pejantan dan betina agar proses kopulasi dapat berlangsung optimal dan efektif. Dalam hal ini faktor penting adalah kondisi hormonal seks di dalam tubuh satwa jantan dan betina.
(2) Perilaku Kopulasi
Perilaku kopulasi ditunjukkan oleh naiknya burung jantan ke atas punggung burung betina lalu memasukkan semen/spermatozoa ke dalam saluran reproduksi betina, ditandai oleh terangkatnya bulu ekor burung betina. Kopulasi berlangsung sangat singkat yakni 2-3 detik. Umumnya kopulasi berlangsung di lantai kandang, meskipun pada beberapa kasus ditemukan kopulasi berlangsung di tempat tenggeran. Waktu kopulasi umumnya terjadi pada pagi hari jam 09.00-11.00 WIB, siang hari (sekitar jam 12.00-13.30 WIB) dan menjelang sore hari sekitar jam 15.00-16.00 WIB. Frekuensi kopulasi terbanyak berlangsung pada siang hari. Dalam satu hari sepasang burung tekukur dapat melakukan kopulasi 4-5 kali, sedangkan burung puter dapat mencapai 5-6 kali dengan jarak waktu antar dua kopulasi secara berurutan dalam satu waktu sekitar satu sampai dua jam.
(3) Perilaku Pasca Kopulasi
Segera setelah kopulasi burung jantan turun dari punggung betina sambil mengepakkan sayap, diam sesaat kemudian terbang ke sarang atau tenggeran. Begitu pula halnya dengan burung betina. Setelah itu burung jantan kembali bersuara, terbang keluar masuk sarang dan mencoba kembali mendekati betina.
Jika kopulasi yang terjadi itu efektif, biasanya diikuti dengan aktivitas bersama antara jantan dan betina dalam mempersiapkan sarang bagi betina untuk meletakkan telurnya. Dari beberapa kasus yang diamati diketahui bahwa jarak waktu antara kopulasi dengan saat peletakkan telur oleh betina sekitar 3 – 5 hari, kadang-kadang mencapai 7-10 hari. Segera setelah telur semua diletakkan (biasanya dua butir) maka betina mulai mengerami telur tersebut. Dari pengamatan diketahui bahwa secara umum kedua telur itu diletakkan secara berurutan dengan jarak waktu mencapai 23-24 jam, namun dari beberapa kasus juga diketahui bahwa telur kedua diletakkan sekitar 40-48 jam kemudian, pada pagi maupun sore hari. Pengeraman telur dilakukan segera setelah telur kedua diletakkan. Tugas utama pengeraman telur dilakukan oleh induk betina, sedangkan induk jantan hanya membantu terutama dalam meng-amankan dan menjaga kestabilan kondisi sarang pada saat induk betina keluar sarang untuk makan dan minum serta menggerakkan tubuh.
Dari segi pasangan kawin dalam satu periode kawin/bertelur, hasil pengamatan menunjukkan bahwa kedua jenis burung ini dapat dikategorikan kedalam tipe monogamus temporalis. Artinya dalam satu masa kawin (reproduksi) burung tekukur dan puter hanya memiliki satu pasangan saja, dan pada masa kawin berikutnya pasangan tersebut dapat dipisahkan dan kawin dengan pasangan yang lain.
KESIMPULAN
Burung tekukur dan burung puter memiliki pola reproduksi yang relatif sama. Kedua jenis burung ini dapat dikategorikan ke dalam hewan dengan tipe monogamus temporalis, yakni hanya kawin dengan satu pasangan paling tidak dalam satu periode kawin (bertelur). Gambaran morfometrik spermatozoanya juga relatif sama, yakni untuk burung tekukur berturut-turut, panjang kepala sperma, lebar kepala sperma dan panjang ekor adalah 13.44 ± 0.73 µm, 1.00 µm, 104.00 ± 6.22 µm, sedangkan pada burung puter berturut-turut panjang kepala sperma 13.00 ± 0.62 µm, lebar kepala sperma 1,00 µm, dan panjang ekor 99.33 ± 8.02 µm. Konsentrasi spermatozoa, burung tekukur berkisar 270-700 x 106/ml pengencer (NaCl fisiologis) sedangkan burung puter berkisar 280-710 x 106/ml pengencer, dengan motilitas spermatozoa pada kedua jenis burung ini berkisar 50% sampai 60%.

Pola Reproduksi Burung Tekukur
88
DAFTAR PUSTAKA
Alcock J. 1989. Animal Behavior. An Evolutionary Approach. Sunderland. Massachusetts. Ainauer Associates Inc, Publisher.
Ardhani J. 1998. Memilih burung untuk penangkaran dan menjodohkan burung cucakrawa. Makalah pada Lokakarya Penangkaran Cucakrawa untuk Menunjang Kelestariannya, 25 Juli 1998 di Taman Burung TMII, Jakarta. Kerjasama PBI Pusat dan PBI Cabang Semarang dengan Taman Burung TMII.
Bahr JM & MR Bakst. 1987. Poultry. Dalam Reproduction in Farm Animals. 5th Edt. Editor ESE Hafez. Philadelphia. Lea and Febiger. pp 379-395.
Ehrlich P. 2004a. Spotted dove –Streptopelia chinensis. Article on Feral Birds Nature Ali Publication California (http//natureali.org/spottedove.htm)
------------. 2004b. Ringed turtle dove – Streptopelia risoria. Article on Feral Birds Nature Ali Publication California (http//natureali.org/ringeddove.ht).
Etches RJ. 1996. Reproduction in Poultry. Cab International. Canada.
Gee GF & SA Temple. 1978. Artificial insemintaion for breeding non-domestic birds. Dalam Artificial Breeding of Non-Domestic Animals. Edt. Watson P.F. Publish for The Zoologist Society of London. Academic Press London, NY and Sanfrancisco. pp. 51-72.
Grimes, J.L. 1994. The effect of protein level fed during the prebreeder period on performance of Large White Tukey Breeder hens after an induced molt. J. Poultry Sci., 73: 37-44.
Masy’ud B. 1992. Penampilan reproduksi dan karakteristik genetik jalak bali (Leucopsar rotschildi) hasil penangkaran. Thesis. Bogor. Program Pascasarjana, Institut Pertanian Bogor.
Nalbandov AV. 1990. Fisiologi Reproduksi pada Mamalia dan Unggas. Edisi Ketiga. Jakarta. UI Press.
Nur H. 2000. Peranan konsentrasi vitamin E dan Selenium dalam ransum terhadap reproduksi puyuh. Disertasi. Bogor. Program Pascasarjana Institut Pertanian Bogor.
Parker JE. 1969. Reproduction Physiology in Poultry. Dalam Reproduction in Farm Animals. Second Edition. Editor ESE. Hafez. Lea & Febiger, Philadelphia. Pp235-254.
Sarwono B. 2000. Perkutut. Cetakan XVII. Jakarta. Penebar Swadaya.
Short LL. 1993. The Lives of Bird. Birds of The World and Their Behavior. New York. Henry Honlt and Company.
Sibley CG & JE Ahlquist. 1990. Phylogeny and Clasification of Birds. A Study in Molecular Evolution. New Haven & London. Yale University Press.
Soejoedono R. 2001. Sukses Memelihara Derkuku dan Puter. Jakarta. Penebar Swadaya.
Soemarjoto R & RIB Rahardjo. 2000. Sinom dan Kelantan, Derkuku Unggul untuk Lomba. Jakarta. Penebar Swadaya.
Sturkie PD. 1976. Avian Physiology. New York. Third Edition. Springer-Verlag.
Thohari M. 1998. Pengembangan teknik pengembangbiakan beo nias (Gracula religiosa robusta) secara konvensional dan modern. Laporan Riset Unggulan Terpadu III. Bidang Teknologi Perlindungan Lingkungan. Jakarta. Kantor Menteri Negara Riset dan Teknologi. Dewan Riset Nasional.
Toelihere MR. 1985. Inseminasi Buatan Pada Ternak. Bandung. Penerbit Angkasa.
Zaini MA, RMK Wibhowo, Z Arifin & HMB Ilyas. 2000. Derkuku. Tim Penyusun Paguyuban Pelestari Derkuku Sukoharjo. Cetakan III. Surabaya. PT Trubus Agrisarana.

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 89 – 95
89
1 Disampaikan pada Seminar Nasional Herpet dan Mamalia Air di Bogor tanggal 23 November 2006.
KONSERVASI AMFIBI DI INDONESIA: MASALAH GLOBAL DAN TANTANGAN1)
(Conservation of Amphibian in Indonesia: Global Problems and Challenges)
MIRZA D KUSRINI
Departemen Konservasi Sumberdaya Hutan dan Ekosiwata Fakultas Kehutanan IPB, Kampus Darmaga PO Box 168 Bogor 16000
Telp: +62-251-621947; Email: [email protected]
Diterima 20 Mei 2007/Disetujui 30 Juni 2007
ABSTRACT
This paper reviews potential causes of amphibian population declines in Indonesia in relation to global amphibian declines. At least six factors were identified: over-harvesting, loss of forest and wetland habitats, pollution, disease, introduction of exotic species and deformities. The conservation of Indonesian amphibian faces many challenges, among other the low perception and knowledge about amphibian among general public, also low number and low quality of local herpetologist . Key word: global amphibian declines; amphibian; frog; Indonesia
PENDAHULUAN
Martha L Crump – peneliti katak, yang sedang melakukan penelitian di Costa Rica tahun 1989, menyadari bahwa subyek penelitiannya sulit ditemukan bahkan kemudian menghilang sama sekali. Hilangnya katak emas Atelopus zeteki di Costa Rica merupakan salah satu sinyal terjadinya masalah pada populasi katak di dunia. Berbagai laporan mengenai hilang atau turunnya populasi katak di berbagai dunia menyadarkan para peneliti bahwa keberadaan amfibi di dunia kini terancam (Barinaga 1990; Blaustein dan Wake 1990). Kecemasan ini beralasan karena dari dokumentasi diketahui kepunahan terjadi di daerah-daerah yang terpencil dan dilindungi (misalnya dalam taman nasional yang sedikit mendapat pengaruh manusia) serta tanpa sebab-sebab yang jelas.
Tahun 2004 IUCN melakukan evaluasi terhadap 5743 jenis amfibi di dunia yang dikenal dengan nama Global Amphibian Assessment (GAA) dengan melibatkan tidak kurang dari 500 peneliti dari 60 negara termasuk Indonesia. Hasilnya menunjukkan bahwa sedikitnya 1,856 jenis (32%) terancam punah, sedikitnya 9 jenis punah sejak 1980, 113 species tidak ditemukan lagi akhir-akhir ini dan 43% dari semua jenis mengalami penurunan populasi (Stuart et al. 2005) . Laporan ini tentunya mencemaskan para peneliti karena amfibi diketahui memberikan manfaat tak terhitung bagi manusia baik manfaat langsung sebagai sumber utama protein hewani ataupun manfaat tak langsung sebagai bagian dari rantai makanan. Paha katak beku, misalnya
merupakan salah satu komoditi penting yang diekspor dari negara miskin atau berkembang ke berbagai negara maju (Kusrini dan Alford 2006). Amfibi sangat penting dalam penelitian mengenai anatomi vertebrata, neurologi, fisiologi, embriologi, genetika, biologi evolusi, perilaku hewan dan ekologi komunitas (Stebbins dan Cohen, 1995; Pough et al. 1998). Telur dan larva amfibi telah digunakan secara ekstensif dalam studi toksikologi untuk melihat dampak kontaminan kimiawi yang dapat mempengaruhi kesehatan manusia (Sparling et al. 2000). Sekresi kulit dari beberapa jenis kini juga dikembangkan sebagai antibiotika dan obat penghilang rasa sakit (Stebbins dan Cohen 1995). Amfibi juga penting untuk mengontrol hama serangga seperti nyamuk (Pough et al. 1998). Selain itu, karena kehidupannya yang kompleks dimana mereka sangat tergantung pada habitat tertentu untuk kawin, mencari makan dan istirahat, serta kondisi morfologinya yang khas (kulit dan membran telur memiliki permeabilitas tinggi) maka beberapa jenis merupakan bioindikator kesehatan lingkungan yang berharga (Duellman dan Trueb 1986; Welsh dan Ollivier 1998; Tyler 1999).
Selama lebih dari satu dekade, berbagai penelitian dilakukan untuk mencari penyebab berkurangnya populasi amfibi secara global. Para peneliti kini mengetahui bahwa kelestarian amfibi terancam oleh satu atau kombinasi dari berbagai penyebab yaitu penangkapan, pengurangan habitat, pencemaran, introduksi spesies eksotik, serta penyakit dan parasit (Ferraro dan Burgin 1993; Pechman dan Wilbur 1994; Green 1997; McTaggart 1998; Alford dan Richards

Konservasi Amfibi di Indonesia
90
1999; Bury 1999; Kiesecker et al. 2001; Alford et al. 2001, Carrey et al. 2001). Data mengenai kepunahan amfibi lebih banyak dilaporkan dari negara-negara maju di benua Amerika, Australia dan Eropa. Hal ini antara lain disebabkan oleh kurangnya penelitian mengenai amfibi di Asia dan Afrika dibandingkan di negara-negara ke-3 benua lainnya.
Indonesia merupakan salah satu negara yang minim dalam penelitian amfibi. Saat ini data mengenai keberadaan dan status amfibi di Indonesia sangat sedikit. Tidak saja terkait berkurangnya jumlah jenis, namun akibat semakin intensifnya survey amfibi di berbagai tempat di Indonesia terutama di Sulawesi dan Papua, maka jumlah jenis di Indonesia pun dilaporkan bertambah dengan ditemukannya berbagai spesies baru. Hal ini tidak berarti jenis amfibi di Indonesia tidak terancam. Paling tidak 39 species kini telah masuk ke dalam Red List IUCN Tahun 2006 dengan kategori terancam, dua diantaranya dari Jawa yaitu Limnonectes macrodon (Vulnerable atau rentan) dan Leptophryne cruentata (Critically Endangered atau kritis). Limnonectes macrodon atau katak batu adalah katak berukuran relatif besar yang umum dijumpai di sungai-sungai di Jawa dan terutama ditangkap untuk konsumsi. Sementara Leptophryne cruentata adalah kodok berukuran relatif kecil, endemik Jawa Barat dengan populasi diketahui berada antara lain di Taman Nasional Gunung Gede Pangrango (Iskandar 1998; Kusrini et al. 2005). Makalah ini mencoba mengulas berbagai faktor yang dapat mengancam kelestarian amfibi di Indonesia dan tantangan ke depan dalam konservasi amfibi Indonesia.
FAKTOR PENYEBAB PENURUNAN POPULASI AMFIBI
1. Penangkapan lebih
Katak terutama ditangkap untuk dikonsumsi. Sedikitnya ada 4 jenis katak yang diperjualbelikan untuk konsumsi di Indonesia antara lain Fejervarya cancrivora, Fejervarya limnocharis, Limnonectes macrodon dan Rana catesbeiana. Jenis-jenis yang ditangkap ini bervariasi tergantung pulau, walaupun ada kesamaan dari jenis yang dikonsumsi ini yaitu berukuran relatif besar dan memiliki kulit tanpa kelenjar racun.
Indonesia adalah negara pengeskpor terbesar paha katak beku di dunia. Setiap tahun rata-rata sekitar 4 juta kg paha katak beku Indonesia diekspor ke berbagai negara terutama ke negara-negara di Eropa dimana lebih dari 80% merupakan hasil penangkapan dari alam (Kusrini dan Alford 2006). Sebelum Indonesia, India dan Bangladesh adalah negara pengekspor katak beku terbesar. Namun dengan makin berkurangnya populasi katak konsumsi di negara tersebut, katak-katak tersebut kemudian statusnya menjadi dilindungi dan dimasukkan dalam Appendix II
CITES. Indonesia kemudian mengambil alih posisi sebagai pengekspor katak beku terbesar di dunia.
Data mengenai ekspor Indonesia hanya dalam ukuran volume, sehingga tidak diketahui dengan pasti jumlah katak yang ditangkap per tahun untuk kegiatan ini. Diperkirakan jumlah katak yang ditangkap untuk kepentingan ekspor mencapai tidak kurang dari 57 juta ekor pada periode tahun 1999-2002 (Kusrini dan Alford 2006). Jumlah ini belum termasuk yang ditangkap untuk kepentingan konsumsi dalam negeri, diduga jumlahnya juga cukup besar. Walaupun penelitian terkini menunjukkan bahwa penangkapan katak konsumsi di Jawa yang berasal dari habitat persawahan masih di bawah batas penangkapan berlebih, namun selalu ada kekhawatiran bahwa suatu saat Indonesia akan melakukan penangkapan berlebih terutama untuk katak yang hidup di luar habitat persawahan (Kusrini 2005). Turunnya populasi katak akibat penangkapan berlebih katak konsumsi di Amerika Serikat pada tahun 1800-an telah didokumentasikan dengan baik oleh peneliti di negara tersebut (Jennings dan Hayes 1985).
Selain digunakan sebagai bahan makanan, amfibi diperjualbelikan antar negara sebagai binatang peliharaan (pet) dan juga digunakan sebagai binatang percobaan di laboratorium atau sebagai bahan percobaan di kelas serta kulitnya dimanfaatkan untuk kerajinan. Jenis-jenis amfibi yang diperjualbelikan untuk hewan peliharaan umumnya merupakan jenis-jenis amfibi yang berwarna cerah dan cenderung "jinak". Saat ini jenis-jenis yang laris diperjualbelikan di pasar adalah salamander dan katak dendrobatid dari Amerika Selatan (Spellerberg 1971; Gorzula 1996). Walaupun memelihara amfibi mungkin tidak populer bagi kebanyakan masyarakat Indonesia, namun ada juga jenis-jenis amfibi Indonesia yang diekspor keluar negeri untuk kepentingan pet. Beberapa kali tim peneliti dari IPB menjumpai pengumpul katak di lapang yang mencari katak bukan untuk konsumsi melainkan untuk pet. Jenis yang ditangkap umumnya katak pohon yang berwarna indah dan relatif besar seperti Rhacophorus reinwardtii, R. javanus, Nictyxalus margaritifer dan katak serasah yang berbentuk unik semisal Megophrys montana. Whitten et al. (1996) mencatat bahwa pasar utama katak peliharaan adalah Jepang, Amerika Utara dan Eropa. Sayangnya saat ini data tentang jenis dan volume amfibi yang diekspor dari Indonesia untuk kepetingan peliharaan sangat minim. Sejauh ini Soehartono dan Mardiastuti (1995) melaporkan bahwa jumlah jenis amfibi yang diekspor dari Indonesia tahun 1992-1995 meningkat empat kali yaitu dari 10 jenis tahun 1992 menjadi 40 jenis pada tahun 1995. Kebanyakan jenis yang diambil berasal dari sawah dan hutan di sekitar Jawa dan pulau-pulau besar lainnya seperti Kalimantan, Sulawesi dan Papua.
Secara tradisional katak digunakan dalam penelitian sebagai hewan percobaan atau bahan praktikum di kelas. Sebuah penelitian di India mengungkap banyaknya jumlah

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 89 – 95
91
katak yang digunakan sebagai bahan praktikum di kelas (Pandian dan Marian 1986) . Pengalaman masa sekolah di Indonesia biasanya menggunakan katak sawah Fejervarya cancrivora sebagai bahan praktikum anatomi. Merebaknya kasus penyakit katak chytridiomycosis di dunia disinyalir disebabkan oleh perjualbelian katak Xenopus (yang berasal dari Afrika) yang digunakan sebagai bahan percobaan laboratorium (Weldon et al. 2004).
Walaupun tidak dalam jumlah yang sangat besar, kulit amfibi dimanfaatkan sebagai bahan membuat kerajinan seperti tas. Di Indonesia jenis yang dimanfaatkan ini umumnya dari jenis kodok buduk Bufo melanostictus yang berkulit kasar sehingga saat disamak menghasilkan tekstur yang menarik. Katak ini sangat melimpah dan umum dijumpai di sekitar pemukiman di seluruh Indonesia. Oleh karena itu pemanfaatan Bufo melanostictus untuk diambil kulitnya kemungkinan besar tidak akan mempengaruhi populasinya.
2. Hilangnya Hutan dan Lahan Basah
Beberapa jenis amfibi terrestrial, misalnya Lepto-brachium hasseltii dan Megophrys montana yang menyebar di Jawa adalah penghuni hutan. Hilangnya hutan dapat memusnahkan jenis-jenis yang sangat tergantung kepada keberadaan habitat ini. Hasil penelitian. Utama et al. (2003), menunjukkan keanekaragaman amfibi di hutan bekas tebangan dan hutan yang belum ditebang sangat berbeda nyata. Hutan bekas tebangan memiliki jumlah jenis lebih sedikit daripada hutan yang masih alami. Hal yang sama juga dikonfirmasi oleh penelitian Ul-Hasanah (2006) yang menunjukkan berubahnya komposisi jenis amfibi di hutan yang masih baik dengan hutan yang telah terbuka. Jenis-jenis yang hilang adalah katak-katak penghuni serasah dan pepohonan.
Kebanyakan amfibi berbiak di lahan basah. Di berbagai negara, telah terjadi kehilangan lahan basah yang sangat menonjol, karena digunakan untuk kepentingan lain - umumnya untuk pembangunan. Hilangnya lahan basah sama dengan hilangnya amfibi. Selain hilangnya lahan basah, perubahan kualitas lahan basah melalui eutrofikasi, pencemaran, pemasukan ikan, hilangnya hutan dan padang di sekitarnya juga dapat menghilangkan populasi amfibi. Sebagai tambahan, banyak spesies amfibi memerlukan lahan basah temporer yang hanya muncul pada saat musim hujan. Sebagai contoh, genangan atau kubangan yang timbul pada musim hujan di dalam hutan atau kebun-kebun memiliki peran penting bagi pembesaran berudu katak pohon atau jenis lain (Yazid 2006). Penimbunan, kontrol nyamuk dan kerapihan manusia telah menghilangkan hampir semua lingkungan seperti itu. Hilangnya lingkungan lahan basah temporer bisa berdampak lebih berat daripada akibat kehilangan kolam permanen.
3. Pencemaran
Lahan basah dan habitat memijah amfibi lainnya seringkali menjadi tempat pembuangan, penampungan dan pengakumulasi bahan pencemar. Hasil penelitian menunjukkan bahwa amfibi rentan terhadap senyawa-senyawa seperti logam berat, produk petroleum, herbisida dan pestisida (Sparling et al. 2000). Penelitian laboratorium secara konsisten menunjukkan berudu lebih rentan terhadap pestisida daripada ikan, meskipun ikan merupakan organisme uji akuatik standar. Ribuan senyawa baru pertanian dan industri muncul di pasar dan lingkungan. Uji toksisitas terhadap tanaman dan hewan kurang cukup untuk menentukan dampak senyawa tersebut kepada lingkungan dimana kombinasi senyawa-senyawa yang tak diduga bisa menimbulkan efek tambahan atau sinergis. Selain itu kombinasi antara kemasaman perairan dan radiasi UV-B seringkali meningkatkan toksisitas kontaminan (Linder et al. 2003). 4. Penyakit
Berbagai jenis penyakit yang disebabkan oleh berbagai sebab (virus, bakteria, dan jamur) diduga berperan dalam penurunan populasi amfibi di dunia. Beberapa penelitian terakhir menunjukkan bahwa jamur Batrachochytrium dendrobatidis (Bd) sebagai penyebab penyakit chytridiomycosis diduga menjadi penyebab kematian masal amfibi di Amerika Tengah dan Australia dan kini telah diteliti secara intensif (Berger et al. 1998, 1999). Saat ini, Bd telah ditemukan di hampir seluruh populasi amfibi di dunia kecuali di Asia, yang tampaknya lebih disebabkan oleh kurangnya penelitian bukan oleh absennya patogen ini.
Walaupun Bd belum dijumpai di Indonesia, potensi ancaman Bd tdak boleh dikesampingkan, karena kondisi klimat di beberapa daerah seperti Jawa Barat, terutama di lokasi-lokasi dataran tinggi sangat potensial bagi perkembangan Bd. Sebagai contoh suhu di Taman Nasional Gede Pangrango (TNGP) dan TN Gunung Halimun Salak (TNGHS) di siang hari berkisar antara 13.5–28 oC dengan kelembaban relatif 63–100% yang merupakan kondisi optimal bagi Bd. Ron (2005) mengembangkan model penyebaran global Bd berdasarkan variabel lingkungan dan memprediksi hutan-hutan pegunungan di Jawa dan Sumatera sangat mungkin menjadi penyebaran Bd. Selain itu Indonesia juga mengintroduksi katak lembu Rana catesbeiana, yang diketahui seringkali terinfeksi Bd di kolam-kolam penangkaran di Amerika Selatan (Mazzoni et al. 2003) sehingga berpotensi menyebarkan Bd.
Masalah parasit pada katak merupakan masalah yang paling sedikit dipelajari dibandingkan kelas vertebrata lainnya. Paling tidak 41 spesies dari 35 genera dan 21 famili trematoda digenea menggunakan larva amfibi sebagai inang perantara (misalnya menyelinap ke dalam jaringannya untuk membentuk kista metacercaria) dan merupakan resiko alami

Konservasi Amfibi di Indonesia
92
populasi amfibi yang kurang diteliti (Smyth and Smyth 1980). Walaupun keberadaan patogen ini mungkin tidak mengakibatkan kematian namun paling tidak dapat mengurangi kesehatan individu yang terjangkit dan pada gilirannya akan mengurangi kemampuan bertahan satu jenis (Kiesecker dan Blaustein 1999; Berven dan Boltz 2001). 5. Spesies Introdusir
Amfibi di berbagai habitat danau dan sungai seringkali hilang karena dimakan oleh ikan atau jenis katak lain. Contoh, katak lembu Rana catesbeiana dimana bentuk dewasa maupun berudunya adalah predator, yang sengaja diintrodusir manusia untuk keperluan konsumsi. Ketidaktahuan atau pengabaian merupakan salah satu penyebab utama introduksi jenis eksotis ini. Kebanyakan introduksi spesies-spesies eksotis dilakukan oleh Departemen Perikanan, orang-orang yang bergerak di bidang perikanan dan masyarakat umum yang membeli benih berdasarkan promosi, atau masyarakat yang bosan akan peliharaan mereka dan membuangnya ke alam. Di Indonesia, paling tidak diketahui satu jenis katak yang diintrodusir yaitu katak lembu Rana catesbeiana (Iskandar 1998) yang berasal dari Amerika Selatan dan terutama ditujukan untuk penangkaran. Sejauh ini tidak diketahui dampak introdusir dari jenis ini di Indonesia, terutama jika jenis ini lepas di alam, namun dilaporkan bahwa Rana catesbeiana menjadi salah satu kompetitor utama katak asli di Amerika Serikat dan kini dianggap sebagai hama (Moyle 1973; Hayes and Jennings 1986; Kiesecker and Blaustein 1997).
6. Kecacatan pada katak
Kecacatan dapat terjadi pada semua makhluk hidup. Isu kecacatan pada amfibi baru muncul pada beberapa tahun. Walaupun kecacatan pada katak sudah dilaporkan dalam berbagai literatur sekitar 100 tahun yang lalu, namun jumlah lokasi dan proporsi abnormalitas saat itu tidak diketahui dengan jelas. Diperkirakan amfibi mempunyai laju kecacatan normal pada angka sekitar 5% (Oullett 2000). Frekuensi kecacatan tertinggi biasanya terdapat pada katak-katak yang baru saja mengalami metamorfosis dari berudu. Penyebab dari kecatatan ini sangat beragam antara lain berhubungan dengan bahan pencemar, parasit (trematoda), radiasi UV-B, genetik dan trauma, maupun sinergi diantara faktor-faktor tersebut (Blaustein et al. 1997; Johnson et al. 1999; Julian 2000; Oullett 2000; Leong 2001; Gray et al. 2002; Kiesecker 2002). Penelitian mengenai kecacatan sangat kurang dilakukan di Indonesia. Tahun 2001 Leong melaporkan temuan kecacatan pada tungkai anakan Rana chalconota dari Sukabumi yang diduga oleh keberadaan parasit. Kecacatan juga dilaporkan pada beberapa individu katak dewasa di berbagai lokasi di Jawa Barat lainnya (Radiansyah et al. 2003; Kusrini et al.
2004), walaupun umumnya frekuensi kecacatan rendah dan lebih banyak disebabkan oleh trauma. Walaupun kecacatan mungkin tidak mengakibatkan punahnya populasi suatu jenis, namun frekuensi kecacatan yang tinggi dikhawatirkan akan mengurangi kesehatan dan selanjutnya daya tahan hidup jenis tersebut. Selain itu, bila kecacatan disebabkan oleh kontaminan kimiawi, hal ini menggambarkan adanya resiko bagi kesehatan manusia.
TANTANGAN KONSERVASI AMFIBI DI INDOESIA
Amfibi adalah salah satu biota yang kurang mendapat perhatian dalam penelitian di Indonesia. Hal ini antara lain karena kurang dikenalnya hewan ini di masyarakat umum maupun di kalangan peneliti, seperti yang ditengarai oleh Iskandar dan Erdelen (2006). Adanya persepsi negatif bahwa katak beracun atau menjijikan (Kusrini et al. 2003) membuat amfibi dijauhi oleh masyarakat. Salah satu catatan mengenai diabaikannya amfibi secara politis adalah tidak adanya amfibi di Indonesia yang masuk ke dalam daftar satwa liar yang dilindungi oleh undang-undang. Oleh karena itu, tidaklah mengherankan jika lokasi-lokasi penting bagi spesies atau komunitas amfibi langka yang menjadi rusak atau berubah karena tidak ada yang tahu mengenai hewan yang hidup di dalamnya.
Salah satu cara untuk mengubah persepsi masyarakat adalah dengan pendidikan konservasi amfibi melalui berbagai cara antara lain penyuluhan bagi anak-anak sekolah dan masyarakat umum baik secara langsung (di kelas) maupun melalui media lainnya (misalkan penyebaran poster dan leaflet), pelatihan bagi guru sekolah, maupun pelatihan khusus bagi peneliti muda mengenai metode penelitian amfibi. Diharapkan dengan adanya pendidikan ini dapat a) meningkatkan pengetahuan masyarakat akan kekayaan hayati Indonesia, b) meningkatkan simpati dan dukungan publik bagi konservasi amfibi, dan c) meningkatkan efektivitas dari aksi konservasi dan kampanye konservasi amfibi.
Pendidikan herpetologi diharapkan juga dapat menginspirasi beberapa orang agar dapat menjadi peneliti amfibi di masa depan. Laporan-laporan mengenai penelitian amfibi di Indonesia lebih banyak ditulis oleh peneliti asing daripada peneliti lokal yang juga mengindikasikan kurangnya jumlah peneliti lokal (Iskandar dan Erdelen 2006). Saat ini jumlah peneliti amfibi di Indonesia sangatlah sedikit (yang dikenal kurang dari 10 orang, dimana sebagian besar bekerja di MZB-LIPI). Namun, perlahan-lahan banyak mahasiswa dan peneliti yang mulai tertarik melakukan penelitian amfibi. Untuk itu perlu diupayakan adanya suatu standar metode inventarisasi dan penelitian amfibi sehingga penelitian dapat diperbandingkan.
Salah satu kelemahan penelitian amfibi di Indonesia adalah hampir tidak ada kerja sama antar instansi terutama

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 89 – 95
93
dalam hal membagi informasi tentang hal yang pernah, sedang atau akan dilakukan. Laporan-laporan sering dibuat hanya untuk disimpan dalam laci dan tidak tersebar luas. Hal ini mengakibatkan adanya kemungkinan duplikasi penelitian, pengeluaran tidak efisien, dan kurangnya konsesus pada teknik dan pendekatan masalah. Selain itu penelitian yang ada umumnya berupa survey jangka pendek sehingga tidak bisa diketahui dinamika populasi suatu jenis yang mungkin terancam turun populasinya. Oleh karena itu perlu ditingkatkan kerja sama jaringan (networking) di antara para peneliti lokal untuk kemajuan penelitian pada masa datang.
Mengingat rendahnya informasi mengenai jenis dan status amfibi di Indonesia, diharapkan penelitian amfibi di Indonesia bisa lebih dikembangkan. Dalam rangka menjawab kemungkinan penurunan populasi amfibi di Indonesia maka perlu dilakukan tindakan antara lain: 1) mengidentifikasi jenis dan populasi yang mengalami ancaman penurunan populasi, 2) mengidentifikasi faktor penyebab dari penurunan populasi tersebut, 3) program monitoring populasi, 4) merevisi daftar jenis yang perlu masuk dalam undang-undang perlindungan, 5) analisis masalah dan rencana manajemen, dan 6) meningkatkan program pendidikan konservasi amfibi bagi masyarakat.
KESIMPULAN
1. Paling tidak enam faktor yang dapat menjadi penyebab turunnya populasi amfibi di Indonesia yaitu penangkapan lebih, hilangnya hutan dan lahan basah, pencemaran, penyakit, spesies introduksi dan kecacatan.
2. Konservasi amfibi di Indonesia masih menghadapi banyak tantangan antara lain disebabkan oleh kurangnya pengetahuan masyarakat tentang amfibi dan rendahnya kuantitas serta kualitas peneliti Indonesia di bidang amfibi.
3. Untuk menjawab kemungkinan penurunan populasi amfibi di Indonesia, diperlukan berbagai tindakan (aksi), antara lain: mengidentifikasi jenis dan populasi yang mengalami penurunan populasi serta faktor penyebab ancamannya; revisi daftar jenis yang perlu masuk dalam undang-undang perlindungan, dan peningkatan program pendidikan konservasi amfibi bagi masyarakat.
DAFTAR PUSTAKA
Alford, R. A. and S. J. Richards. 1999. Global amphibian declines: A problem in applied ecology. Ann.Rev. Ecol. Syst. 30: 133-165.
Alford, R. A., P. M. Dixon and J. H. K. Pechmann. 2001. Global amphibian population declines. Nature 412: 499-500.
Barinaga, M. 1990. Where have all the froggies gone? Science 247: 1033-1034.
Berger, L., R. Speare, P. Daszak, D. E. Green, A. A. Cunningham, C. L. Goggin, R. Stratcombe, M. A. Ragan, A. D. Hyatt, K. R. McDonald, H. B. Hines, K. R. Lips, G. Farantelli and H. Parkes. 1998. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forest of australia and central america. Proc.Natl.Acad.Sci. USA 95: 9031-9036.
Berger, L., R. Speare and A. Hyatt. 1999. Chytrid fungi and amphibian declines: Overview, implications and future directions.In: A. Campbell (eds) Declines and dissapearances of australian frogs. Canberra, Environment Australia: 23-33 pp.
Berven, K. A. and R. S. Boltz. 2001. Interactive effects of leech (desserobdella picta) infection on wood frog (rana sylvatica) tadpole fitness traits. Copeia 2001(4): 907-915.
Blaustein, A. R. and D. B. Wake. 1990. Declining amphibian population: A global phenomenon? Trends in Ecology and Evolution 5(7): 203-204.
Blaustein, A. R., J. M. Kiesecker, D. P. Chivers and R. G. Anthony. 1997. Ambient UV-B radiation causes deformities in amphibian embryos. Proc. Natl. Acad. Sci. USA. 94: 13735–13737.
Bury, R. B. 1999. A historical perspective and critique of the declining amphibian crisis. Wildlife Society Bulletin 27(4): 1064-1068.
Carey, C., W. R. Heyer, J. Wilkinson, R. A. Alford, J. W. Arntzen, T. Halliday, L. Hungeford, K. R. Lips, E. M. Middleton, S. A. Orchard and A. S. Rand. 2001. Amphibian declines and environmental change: Use of remote-sensing data to identify environmental correlates. Conservation Biology 15(4): 903-913.
Duellman, W. E. and L. Trueb. 1994. Biology of amphibian. The Johns Hopkins University Press. Baltimore670. pp.
Ferraro, T. J. and S. Burgin. 1993. Review of environmental factors influencing the decline of australian frogs. Dalam: D. Lunney and D. Ayers (eds) Herpetology in Australia: A diverse discipline, Mosman: Royal Zoological Society of New South Wales: 205-218. pp.
Gorzula, S. 1996. The trade in dendrobatid frogs from 1986 to 1993. Herpetological Review 27(3): 116-123.
Gray, H. M., M. Ouelett and D. M. Green. 2002. Traumatic injuries in two neotropical frogs, Dendrobates auratus

Konservasi Amfibi di Indonesia
94
and Physalaemus pustulosus. Journal of Herpetology 36(1): 117-121.
Green, D. M. 1997. Perspectives on amphibian population declines: Defining the problem and searching for answer.In: D. M. Green (eds) Amphibians in decline: Canadian studies of a global problem, Herpetological Conservation. 1: 291-308 pp.
Hayes, M. P. and M. R. Jennings. 1986. Decline of ranid frog species in western north America: Are bullfrogs (Rana catesbeiana) responsible? Journal of Herpetology 20: 490 - 509.
Iskandar, D. T. 1998. Amfibi Jawa dan Bali. 1. Puslitbang Biologi-LIPI. Bogor. 132 hal.
Iskandar, D. T and W.R. Erdelen. 2006. Conservation of amphibians and reptiles in Indonesia: issues and problems. Amphib. Reptile Conserv. 4(1):60-93
IUCN. 2006. IUCN Red List of Threatened Species. http://www.redlist.org/ (accessed on 18 November 2006).
Jennings, M. R. and M. P. Hayes. 1985. Pre-1900 overharvest of california red-legged frogs (Rana aurora draytonii): The inducement for bullfrog (Rana catesbeiana) introduction. Herpetologica 41: 94-103.
Julian, S. E. 2000. Effects of chronic pesticide exposure on larval amphibians. Master of Science thesis. Applied Ecology and Conservation Biology, Frostburg. 98 pp.
Johnson, P. T. J., K. B. Lunde, E. G. Ritchie and A. E. Launer. 1999. The effect of trematode infection on amphibian limb development and survivorship (trematode parasite caused increase in abnormality frequency). Science 284(5415): 802-807
Kiesecker, J. M. and A. R. Blaustein. 1997. Population differences in responses of red-legged frogs (rana aurora) to introduced bullfrogs. Ecology 78(6): 1752-1760.
Kiesecker, J. M. and A. R. Blaustein. 1999. Pathogen reverses competition between larval amphibians. Ecology 80(7): 2442.
Kiesecker, J. M., A. R. Blaustein and L. K. Belden. 2001. Complex causes of amphibian population declines. Nature 410: 681-684.
Kiesecker, J. M. 2002. Synergism between trematode infection and pesticide exposure: A link to amphibian limb deformities in nature? PNAS 99(15): 9900–9904.
Kusrini, M. D., A. Mardiastuti and A. Fitri. 2003. Promoting frog conservation through environmental education and research experience: Pilot project in
west java, indonesia. Dalam: M. D. Kusrini, A. Mardiastuti and T. Harvey (eds) Prosiding seminar hasil penelitian konservasi amfibi dan reptil di indonesia. Bogor, 8 Mei 2003. Bogor, Departemen Konservasi Sumberdaya Hutan.Institut Pertanian: hal 45-51.
Kusrini, M. D., R. A. Alford, A. Fitri, D. M. Nasir and S. Rahardyansah. 2004. Morphological abnormalities in frogs of west java, indonesia. Froglog 64.
Kusrini, M. D., A. Fitri, H. Utama, D. M. Nasir, D. Ardiansyah, V. Lestari and R. Rachmadi. 2005. Project 202404: Ecology and conservation of frogs of mount gede pangrango national park. Bogor, Institut Pertanian Bogor: 23 hal.
Kusrini, M. D. 2005. Edible frog harvesting in indonesia: Evaluating its impact and ecological context. PhD thesis. School of Tropical Biology, James Cook University, Towsnville. 239 hal.
Kusrini, M. D. and R. A. Alford. 2006. Indonesia’s exports of frogs’ legs. Traffic Bull. 21(1): 13-24.
Leong, T. M. 2001. Parasitic copepods responsible for limb abnormalities. FROGLOG 46(3).
Linder, G., S. K. Krest and D. W. Sparling. 2003. Amphibian decline: An integrated analysis of multiple stressor effects. Society of Environmental Toxicology and Chemistry (SETAC). Pensacola, FL, USA. 368 hal.
Mazzoni, R., A. A. Cunningham, P. Daszak, A. Apolo, E. Perdomo and G. Speranza. 2003. Emerging pathogen of wild amphibians in frogs (Rana catesbeiana) farmed for international trade. Emerging Infectious Diseases 9(8): 995-998.
McTaggart, I. 1998. Fatal fungus linked to frog declines. Ecos(98): 8.
Moyle, P. B. 1973. Effects of introduced bullfrogs, Rana catesbeiana, on the native frogs of the San Joaquin valley, California. Copeia 1973: 18-22.
Ouellet, M. 2000. Amphibian deformities: Current status of knowledge. Dalam: D. W. Sparling, G. Linder and C. A. Bishop (eds) Ecotoxicology of amphian and reptiles. Pensacola, Society of Environmental Toxicology and Chemistry (SETAC): hal. 617-696.
Pandian, T. J. and M. P. Marian. 1986. Production and utilization of frogs: An ecological view. Proc. Indian Acad. Sci. (Anim. Sci.) 95(3): 289 - 301.
Pechman, J. H. K. and H. M. Wilbur. 1994. Putting declining amphibians populations in perspective:

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 89 – 95
95
Natural fluctuations and human impacts. Herpetologica 50: 65-84.
Pough, F. H., R. M. Andrews, J. E. Cadle, M. L. Crump, A. H. Savitzky and K. D. Wells. 2004. Herpetology. Third Edition. Pearson Prentice Hall. London. 726 hal.
Radiansyah, S., A. Priyono dan M.D.Kusrini. 2003. Keanekaragaman Spesies Amfibi Di Sungai Cilember Dalam Kawasan Wana Wisata Curug Cilember, Bogor – Jawa Barat. Dalam: M. D. Kusrini, A. Mardiastuti and T. Harvey (eds) Prosiding seminar hasil penelitian konservasi amfibi dan reptil di indonesia. Bogor, 8 Mei 2003. Bogor, Departemen Konservasi Sumberdaya Hutan.Institut Pertanian: hal. 85 - 104
Ron, S. R. 2005. Predicting the distribution of the amphibian pathogen Batrachochytrium dendrobatidis in the new world. Biotropica 37(2): 209-221.
Soehartono, T. and A. Mardiastuti. 1995. An overview of wildlife trade in Indonesia. 2nd Meeting of the Conference of the Parties to the United Nation Convention on Biological Diversity, Jakarta.
Sparling, D. W., G. Linder and C. A. Bishop. 2000. Ecotoxicology of amphibians and reptiles. SETAC Technical Publications. Columbia. 877 hal.
Spellerberg, I. F. 1971. The amphibian and reptile trade with particular reference to collecting in europe. Biological Conservation 10: 221-232.
Smyth, J. D. and M. M. Smyth. 1980. Frogs as host-parasite systems I. First edition. The Macmillan Press Ltd. London.
Stuart, S. N., J. S. Chanson, N. A. Cox, B. E. Young, A. S. L. Rodrigues, D. L. Fischman and R. W. Waller. 2005. Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786.
Stebbins, R. C. and N. W. Cohen. 1995. A natural history of amphibian. Princeton University Press. New Jersey. 316 hal.
Tyler, M. J. 1999. Australian frogs: A natural history. Reed New Holland. Sydney. 192 hal.
Utama, H. A. Priyono Dan M.D. Kusrini. 2003. Studi Keanekaragaman Amfibi (Ordo Anura) Di Areal Pt Intracawood Manufacturing, Kalimantan Timur. Dalam: M. D. Kusrini, A. Mardiastuti And T. Harvey (Eds) Prosiding Seminar Hasil Penelitian Konservasi Amfibi Dan Reptil Di Indonesia. Bogor, 8 Mei 2003. Bogor, Departemen Konservasi Sumberdaya Hutan. Institut Pertanian: hal. 105-129.
Ul-hasanah, A. U. 2006. Amphibian diversity in Bukit Barisan Selatan National Park, Lampung - Bengkulu. Skripsi S1 tidak dipublikasikan.Departemen Konservasi Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan, Institut Pertanian Bogor. 56 hal.
Weldon, C., L. H. d. Preez, A. D. Hyatt, R. Muller and R. Speare. 2004. Origin of the amphibian chytrid fungus. Emerging Infectious Diseases 10(12).
Welsh, H. H., Jr. and L. M. Ollivier. 1998. Stream amphibians as indicators of ecosystem stress: A case study from california's redwoods. Ecological Applications 8(4): 1118-1132.
Whitten, A. J., R. E. Soeriaatmadja and S. A. Afiff. 1996. The ecology of Java and Bali. Periplus Editions (HK) Ltd. Singapore. 1028 hal.
Yazid, M. 2006. Perilaku berbiak Katak pohon hijau (Rhacophorus reinwardtii Kuhl & Van Hasselt, 1822) di kampus IPB Darmaga. Skripsi S1 tidak dipublikasikan.Departemen Konservasi Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan, Institut Pertanian Bogor. 50 hal.

Status and Recommendations for in situ
96
STATUS AND RECOMMENDATIONS FOR IN SITU ANOA (Bubalus sp.) WITH SUGGESTED IMPLICATIONS FOR THE CONSERVATION BREEDING POPULATION
(Status dan Rekomendasi Anoa (Bubalus sp.) In Situ dengan Implikasi untuk
Konservasi Populasi Berbiak)
JA BURTON1,2, AH MUSTARI 3 DAN ALASTAIR MACDONALD1
1Veterinary Biomedical Sciences, Royal (Dick) School of Veterinary Studies, The University of Edinburgh, Edinburgh EH9 1QH, U.K. 2Centre for Research and Conservation, Royal Zoological Society of Antwerp, Koningin Astridplein 26, 2018 Antwerp, Belgium.
3 Department of Forest Resources Conservation and Ecotourism,Faculty of Forestry, Bogor Agricultural University,PO Box 168, Bogor 16001, Indonesia, E-mail: [email protected]
Diterima 29 Januari 2007/Disetujui 21 Mei 2007
ABSTRAK
Ringkasan ini menguraikan status konservasi in situ kerbau kerdil Sulawesi. Trend populasi yang dilaporkan dalam dua belas tahun terakhir mengarahkan penulis dalam menyoroti arti penting konservasi populasi berbiak di lokasi ex situ pada masa mendatang. Anoa terdiri dari dua jenis kerbau kerdil Bubalus depressicornis & B. quarlesi endemik Pulau Sulawesi, Indonesia. Kajian distribusi anoa berdasarkan laporan historis dan sebagai hasil data lapang terbaru (1990-an sampai 2002) menyoroti terjadinya penurunan di keseluruhan pulau, terutama di semenanjung selatan dan timur laut. Penurunan terjadi akibat perburuan lokal untuk daging dan kehilangan habitat. Sebagian besar populasi secara cepat mengalami fragmentasi. Konservasi populasi viable pada akhirnya akan membutuhkan pengelolaan metapopulasi dan peran yang lebih besar dari populasi di kebun binatang. Kata Kunci: in situ, anoa, konservasi, populasi berbiak.
INTRODUCTION
Anoas are dwarf buffaloes, endemic to the Indonesian island of Sulawesi (and offshore islands). They are the smallest of the Bovini, standing about a meter tall at the shoulder. Two species are presently recognized, the lowland anoa Bubalus depressicornis and the mountain anoa Bubalus quarlesi (Groves, 1969; Corbet and Hill, 1992; Wilson and Reeder, 1993). Both anoa species are classified as Endangered by IUCN (http://www.redlist.org/, August 2004), are legally protected under Indonesian law (Jahja, 1987), and are included in Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (UNEP-WCMC, 2000). The anoas are an important species for locally and internationally defining the identity of the wildlife in the Wallacea bioregion, known for its high bio-diversity and endemism.
The two species taxonomic status is still under debate (Mohr, 1921; Groves, 1969; Weise, 1979; Kakoi, et al., 1994; Schreiber, et al., 1999). Work on this is on-going and will be reported when the analyses are completed. The data presented below has been complied by the authors from field surveys and a literature review, including information from 1990 to 2002. Further details have recently been published by Burton, et al. (2005).
CONSERVATION STATUS
Key areas for future conservation of anoa have been identified where sufficiently large numbers are present that are likely to maintain a viable population in the long-term. These include Lore Lindu National Park, Bogani Nani-Wartabone National Park and environs, the Upper Paguyaman / Nantu area, Morowali Nature Reserve, Tanjung Peropa Wildlife Reserve, Lambu Sango Wildlife Reserve or Buton Utara Nature Reserve.
Hunting (mainly for meat) and the loss of suitable habitat are the major threats to the anoa, and recent reports indicate that hunting is by far the more serious of the two. Anoa meat was usually sold for Rp5000–7500 per kilogram (ca. £0.50 per kg), but one hunter said he could sell anoa meat for Rp22,500/kg (ca. £1.60/kg) (J. Burton, personal observations, 2001). In rural areas, there appears to be little awareness of the protected status of anoas, and villagers readily recounted their hunting experiences to researchers (Schreiber and Nötzold, 1995). Recently anoas have been extirpated from Tangkoko Batuangus – Dua Saudara NR (J. Riley, personal communication 2001). During the Anoa Population and Habitat Viability Analysis Workshop (PHVA) available data on life history was used with the VORTEX Program to estimate the population viability of different population sizes under three levels of hunting

Media Konservasi Vol. XII, No. 2 Agustus 2007 : 96 – 98
97
pressure (Manansang, et al., 1996). These simulations highlight the threat that even the largest anoa populations face from hunting levels that could now be occurring throughout Sulawesi.
Habitat degradation and loss is the second most significant threat to the anoas. Many anoa populations are becoming isolated as the forest around the protected areas is cleared or converted to plantations. Total forested area remaining on Sulawesi in 1985 was estimated to be between 42 and 49% of the land area of the island (Holmes, 2000; Mathews, 2002). Although 11.9% of the island is protected, loss of suitable anoa habitat within protected areas is as much a problem as in unprotected forest (Aden, et al., 2001), such as Lore Lindu National Park and Bogani Nani-Wartabone NP.
The fragmentation of these once contiguous forests will reduce the natural movement of anoa between forest patches. This may cause the loss of genetic diversity and increase the chance of population extinctions (Gilpin, 1991; Wang and Caballero, 1999). However, assessing the scale of the threat posed to the anoa by shifting agriculture and fires is hampered by the lack of knowledge of their habitat requirements. This has recently been altered with research on anoa’s diet and habitat preferences by Mustari (2004).
The conservation breeding population in zoological collections doubled in size in the 10 years prior to the studbook’s publication; there were 125 animals in European, North American, and Asian zoos in December 1998. However, management of the captive breeding program is still hampered by the uncertain taxonomic status of the anoa, and the insecure classification of those animals currently in zoos (Nötzold, 1999). The genetic study of wild anoa presently underway will lay the foundations for the urgently needed clarification of the genetic representation of animals in the ex situ population.
CONSERVATION RECOMMENDATIONS
Anoa need to be protected from hunting by enforcing the existing laws of the Republic of Indonesia, particularly the Conservation of Natural Resources and Ecosystems Act (UURI No. 5). Law enforcement should be combined with an environmental education campaign that stresses that the anoas are unique to Sulawesi and in danger of being lost forever. Successful anti-poaching activities require frequent patrols of key protected areas. In order to make the selling of illegal meat difficult, the Indonesian authorities should target bush meat sellers at village and town/city markets (Lee, 1999).
While the work conducted to date has done much to bring up to date our knowledge of the distribution of anoa on Sulawesi and its offshore islands, we still know rather little about the details of regional population sizes and population trends. For the majority of sites we merely know
that anoa are present or are absent/likely to be absent. This makes it difficult to focus conservation efforts effectively. There is, therefore, still a need for an island-wide survey to quantify major anoa populations. In order to devise the most representative conservation strategy, the surveys should aim to locate populations throughout the island in order to preserve the full biogeographic variation exhibited by these species. In addition, populations from the full range of vegetation types and elevations used by the anoa need to be identified for inclusion in an effective anoa conservation strategy. This is especially significant due to the near total loss of lowland forest in Sulawesi.
On paper Sulawesi boasts an impressive system of protected areas. Unfortunately many of these areas are poorly managed, understaffed, and lack the basic resources required for effective conservation. The priority areas for conservation management should also be assessed. The largest areas of habitat are likely to be the top priorities, because these areas are likely to contain the populations most resilient to demographic and genetic problems and the effects of hunting (see Conservation Status, above).
For effective management of wildlife it is necessary for protected area staff to be adequately trained in surveying and monitoring techniques. It is necessary to explain to people living around protected areas why their activities need to be controlled and why wildlife, which they might otherwise utilise, should be conserved. Conservation-oriented NGOs need to determine whether conservation-education schemes exist in the communities around the most important protected areas in Sulawesi; if not then these should be initiated as a high priority.
By combining the above-mentioned recommendations with the forthcoming publication of the genetic study of the anoa’s taxonomy, the in situ and ex situ population will form a feasible conservation strategy to maintain a viable anoa population in the future. The ex situ population should be developed and maintained as a vital alternative gene pool as well as a source of individuals for future reintroduction. The conservation breeding population should be as genetically representative of the wild population as possible. Results from the in situ genetic study will inform detailed genetic analysis of the ex situ animals and facilitate effective breeding.
ACKNOWLEDGMENTS
This work was supported by The University of Edinburgh and The Dutch Zoo Foundation in collaboration with IPB University, Bogor, Indonesia and The Nature Conservancy, Indonesia.

Status and Recommendations for in situ
98
REFERENCES
Aden, J., G. Dore, J. Vincent and T. Walton. 2001. Indonesia: Environmental and the natural resource management in a time of transition, World Bank Report.
Burton, J. A., S. Hedges and A. H. Mustari. 2005. The taxonomic status, distribution and conservation needs of the lowland anoa Bubalus depressicornis and mountain anoa Bubalus quarlesi. Mammal Review 35(1): 25-50.
Corbet, G. B. and J. E. Hill. 1992. The mammals of the Indomalayan Region: a systematic review, Natural History Museum Publications and Oxford University Press, Oxford, UK.
Gilpin, M. 1991. The genetic effective size of a metapopulation. Biological Journal of the Linnean Society 42: 165-176.
Groves, C. P. 1969. Systematics of the anoa (Mammalia, Bovidae). Beaufortia 17: 1-12.
Holmes, D. 2000. Deforestation in Indonesia: A view of the situation in 1999, Jakarta Indonesia: World Bank Draft Report July 3.
http://www.redlist.org/ (August 2004) The 2003 IUCN Red List of Threatened Species: Bubalus quarlesi and Bubalus depressicornis, The World Conservation Union.
Jahja, M. M. 1987. The possibility of breeding anoa in captivity: an alternative for conservation of the species. BIOTROP, Special Publication 30: 101-108.
Kakoi, H., T. Namikawa, O. Takenaka, A. Takenaka, T. Amano and H. Martojo. 1994. Divergence between the anoas of Sulawesi and the Asiatic water buffaloes, inferred from their complete amino acid sequences of hemoglobin ß chains. Zeitschrift für Zoologische Systematik und Evolutionsforschung 32(1): 1-10.
Lee, R. J. 1999. Impact of subsistance hunting in North Sulawesi, Indonesia, and conservation options. In Hunting for Sustainability in Tropical Forests. Eds: J. G. Robinson and E. L. Bennett. Columbia University Press, New York, USA: pp.455-472.
Manansang, J., S. Hedges, S. Dwiatmo, P. Miller and U. S. Seal. 1996. Population and Habitat Viability Assessment Workshop for the Anoa (Bubalus
depressicornis and Bubalus quarlesi) Report. Population and Habitat Viability Assessment Workshop for the Anoa (Bubalus depressicornis and Bubalus quarlesi), Bogor Indonesia, IUCN/SSC Conservation Breeding Specialist Group, Apple Valley, MN., USA.
Mathews, E. 2002. State of the Forest: Indonesia, Forest Watch Indonesia, Global Forest Watch, World Resources Institute.
Mohr, E. 1921. Die Geographische Verbreitung der Anoa-Arten auf Celebes. Arch. Nauturgeschichte 87(6): 208-214.
Mustari, A. H. 2004. Ecology and Conservation of Lowland Anoa in Southeast Sulawesi, Indonesia. [PhD thesis], University of New England, Australia.
Nötzold, G. 1999. International Studbook: Anoa, (Bubalus [Anoa] depressicornis Smith, 1827), Leipzig Zoo, Germany.
Schreiber, A. and G. Nötzold. 1995. One EEP, but how many anoas? In EEP Yearbook 1994/95. F. Rietkerk, Brouwer, K., and Smits, S., EAZA/EEP Executive Office, Amsterdam, The Netherlands: pp.419-424.
Schreiber, A., I. Seibold, G. Nötzold and M. Wink. 1999. Cytochrome b gene haplotypes characterize chromosomal lineages of anoa, the Sulawesi dwarf buffalo (Bovidae: Bubalus sp.). The Journal of Heredity 90: 165-176.
UNEP-WCMC. 2000. UNEP-WCMC Species Database: CITES-Listed Species, UNEP-WCMC, http://www.cites.org/eng/resources/species.html (May 2004).
Wang, J. and A. Caballero. 1999. Developments in predicting the effective size of subdivided populations. Heredity 82: 212-226.
Weise, R. 1979. Untersuchungen zur innerartlichen Variabilität beim Anoa (Anoa depressicornis). Staatsexamenarbeit, Universität Kiel, Germany.
Wilson, D. E. and D. M. Reeder. 1993. Mammal Species of the World: A Taxonomic and Geographic Reference, Smithsonian Institution Press, Washington, USA and London, UK.




















