
MECHANISMS OF K+ UPTAKE ACROSS THE BASAL MEMBRANE OF MALPIGHIAN TUBULES OF FORMICA POLYCTENA: THE...
23
123 J. exp. Biol. 195, 123–145 (1994) Printed in Great Britain © The Company of Biologists Limited 1994 MECHANISMS OF K + UPTAKE ACROSS THE BASAL MEMBRANE OF MALPIGHIAN TUBULES OF FORMICA POLYCTENA: THE EFFECT OF IONS AND INHIBITORS A. LEYSSENS, S. DIJKSTRA, E. VAN KERKHOVE AND P. STEELS Laboratory of Physiology, Limburgs Universitair Centrum, B3590 Diepenbeek, Belgium Accepted 7 June 1994 Summary In the presence of 6 mmol l 21 Ba 2+ , known to block the K + channels in the basal membrane, a rise in bath [K + ] ([K + ]bl) induced an increase in intracellular K + concentration ([K + ]i) similar in amount and in time course to that obtained in the absence of Ba 2+ . The presence of active and passive (other than through K + channels) K + uptake mechanisms across the basal membrane was investigated in different bath K + concentrations. Dihydro-ouabain (10 23 mol l 21 ), a blocker of the Na + /K + -ATPase, tested in low bath [K + ], and Sch28080 (10 24 mol l 21 ), a K + /H + -ATPase inhibitor, were without effect on fluid secretion. Dihydro-ouabain was also without effect on electrical potential differences either in the absence or in the presence of Ba 2+ . Vanadate (10 23 mol l 21 ), in contrast, strongly reduced fluid secretion not only in control solution but also in high-K + , Na + -free medium and reduced the transepithelial and the apical membrane potential differences but not the basal membrane potential difference or [K + ]i. Omitting Na + from the bathing medium, replacing Cl 2 by Br 2 or applying bumetanide (10 25 mol l 21 ) inhibited fluid secretion only in a low-K + (10 mmol l 21 ) medium. In 51 mmol l 21 [K + ]bl, omitting Na + was without effect and 10 24 mol l 21 bumetanide was needed to inhibit secretion. Replacing Cl 2 by Br 2 stimulated fluid secretion at this K + concentration. Bumetanide (10 24 mol l 21 ) had no effect in 113 mmol l 21 [K + ]bl. Bumetanide (10 24 mol l 21 ) in 51 mmol l 21 [K + ]bl did not affect membrane potentials, did not lower [K + ]i and did not affect the rise in [K + ]i observed on an increase in [K + ]bl. The results were summarized in a model proposing that K + channels play a dominant role in high-K + (113 mmol l 21 ) bathing medium. A K + /Cl 2 cotransporter may become more important in 51 mmol l 21 [K + ]bl and a K + /Na + /2Cl 2 cotransporter may gain in importance in 10 mmol l 21 [K + ]bl. Active mechanisms for K + uptake across the basal membrane seem to play no detectable role in sustaining fluid secretion. The response to vanadate might be due to an effect on the apical electrogenic H + pump. Introduction Malpighian tubules of forest ants (Formica polyctena) secrete primary urine. As in Key words: Malpighian tubules, K + uptake mechanisms, fluid secretion, electrical potential differences, intracellular and luminal K + concentrations, ion substitution, Ba 2+ , dihydro-ouabain, Sch28080, vanadate, bumetanide.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of MECHANISMS OF K+ UPTAKE ACROSS THE BASAL MEMBRANE OF MALPIGHIAN TUBULES OF FORMICA POLYCTENA: THE...

123J. exp. Biol. 195, 123–145 (1994)Printed in Great Britain © The Company of Biologists Limited 1994
MECHANISMS OF K+ UPTAKE ACROSS THE BASALMEMBRANE OF MALPIGHIAN TUBULES OF FORMICAPOLYCTENA: THE EFFECT OF IONS AND INHIBITORS
A. LEYSSENS, S. DIJKSTRA, E. VAN KERKHOVE AND P. STEELS
Laboratory of Physiology, Limburgs Universitair Centrum, B3590 Diepenbeek,Belgium
Accepted 7 June 1994
SummaryIn the presence of 6 mmol l21 Ba2+, known to block the K+ channels in the basal
membrane, a rise in bath [K+] ([K+]bl) induced an increase in intracellular K+
concentration ([K+]i) similar in amount and in time course to that obtained in the absenceof Ba2+. The presence of active and passive (other than through K+ channels) K+ uptakemechanisms across the basal membrane was investigated in different bath K+
concentrations. Dihydro-ouabain (1023 mol l21), a blocker of the Na+/K+-ATPase, testedin low bath [K+], and Sch28080 (1024 mol l21), a K+/H+-ATPase inhibitor, were withouteffect on fluid secretion. Dihydro-ouabain was also without effect on electrical potentialdifferences either in the absence or in the presence of Ba2+. Vanadate (1023 mol l21), incontrast, strongly reduced fluid secretion not only in control solution but also in high-K+,Na+-free medium and reduced the transepithelial and the apical membrane potentialdifferences but not the basal membrane potential difference or [K+]i. Omitting Na+ fromthe bathing medium, replacing Cl2 by Br2 or applying bumetanide (1025 mol l21)inhibited fluid secretion only in a low-K+ (10 mmol l21) medium. In 51 mmol l21 [K+]bl,omitting Na+ was without effect and 1024 mol l21 bumetanide was needed to inhibitsecretion. Replacing Cl2 by Br2 stimulated fluid secretion at this K+ concentration.Bumetanide (1024 mol l21) had no effect in 113 mmol l21 [K+]bl. Bumetanide(1024 mol l21) in 51 mmol l21 [K+]bl did not affect membrane potentials, did not lower[K+]i and did not affect the rise in [K+]i observed on an increase in [K+]bl. The resultswere summarized in a model proposing that K+ channels play a dominant role in high-K+
(113 mmol l21) bathing medium. A K+/Cl2 cotransporter may become more important in51 mmol l21 [K+]bl and a K+/Na+/2Cl2 cotransporter may gain in importance in10 mmol l21 [K+]bl. Active mechanisms for K+ uptake across the basal membrane seem toplay no detectable role in sustaining fluid secretion. The response to vanadate might bedue to an effect on the apical electrogenic H+ pump.
Introduction
Malpighian tubules of forest ants (Formica polyctena) secrete primary urine. As in
Key words: Malpighian tubules, K+ uptake mechanisms, fluid secretion, electrical potential differences,intracellular and luminal K+ concentrations, ion substitution, Ba2+, dihydro-ouabain, Sch28080,vanadate, bumetanide.

many insect species, they preferentially transport K+ into the lumen in controlconditions, Cl2 and water following passively (see Phillips, 1981, for a review). Thefluid secretion rate is strongly dependent on the bath K+ concentration (Van Kerkhoveet al. 1989).
Since K+ is always transported across the epithelium against its electrochemicalgradient, K+ transport must occur transcellularly. The active step is primarily situated inthe apical membrane, where the extrusion of K+ into the lumen is believed to be realizedthrough the combination of an electrogenic H+ pump and an electroneutral K+/H+
antiporter acting in parallel (Weltens et al. 1992; Zhang et al. 1994; Dijkstra, 1993;Leyssens et al. 1993a,b). In Malpighian tubules of Formica polyctena, it was found thatthe bath K+ concentration determines the intracellular K+ content available for apical K+
transport. This may be an intrinsic regulatory mechanism of fluid secretion (Leyssenset al. 1992, 1993a).
K+ transport systems must exist across the basal membrane to ensure a sufficientlylarge K+ uptake to maintain the rate of fluid secretion observed in the different bathingK+ concentrations and to account for the change in intracellular K+ concentration inresponse to the surrounding bath K+ level. K+ entry may occur passively through K+
channels. Ion substitution and K+-selective measurements have demonstrated theappreciable K+ permeability of the basal membrane (Leyssens et al. 1992, 1993a). Thehyperpolarization of the basal membrane potential difference (Vbl) observed at all bathK+ concentrations when Ba2+ was used to block the K+ channels (Weltens et al. 1992)suggested the existence of an electrochemical K+ gradient favourable for K+ uptake.However, the calculated electrochemical gradient for K+ across the basal membrane wasvery small when the bath K+ concentration was decreased and could even reverse(Leyssens et al. 1993a). The fluid secretion was also drastically, but not completely,blocked after 30 min in the presence of Ba2+ (Weltens et al. 1992). This suggested that,as in other K+-transporting Malpighian tubules (for a review, see Nicolson, 1993),alternative pathways for K+ uptake may be present in the basal membrane and that therelative rates of these hypothetical K+ transport systems may vary under differenttransport conditions.
The present study focuses on these alternative routes for K+ transport across the basalmembrane at different bath K+ concentrations. The effects of ion substitutions andblockers of primary and secondary active transport systems on fluid secretion, onintracellular and luminal K+ concentration and on electrical potential differences wereinvestigated.
It is shown that the rise in intracellular K+ concentration when the bath [K+] isincreased still occurs after the basal K+ channels have been blocked. The existence of abasal coupled entry mechanism for Cl2 and K+ is demonstrated and its contribution toK+ uptake under different transport conditions is discussed. The role of Na+ intranscellular ion transport is also investigated. The presence of a primary active K+
uptake system is critically evaluated. On the basis of these and previous findings, atransport model for transepithelial ion transport in the Malpighian tubule of Formicapolyctena is proposed.
124 A. LEYSSENS AND OTHERS

Materials and methods
Preparation
Worker ants of the species Formica polyctena Förster were collected from natural nestsat the periphery of woods and kept in an artificial nest at a constant temperature of 20 ˚C.The animals were fed with sugar and water.
After dissection, a single Malpighian tubule (1–2 mm) was transferred to the stage ofan inverted microscope into a bathing droplet (50 ml). The bathing droplet was coveredwith paraffin oil to avoid evaporation (see Van Kerkhove et al. 1989).
Experimental arrangement
Fluid secretion experiments
The technique used for measuring fluid secretion rate has been described in detail byVan Kerkhove et al. (1989). A fluid secretion experiment consisted of a control (threetimes 10 min), a test and a wash-out period (3–4 times 10 min each). The bathing fluidcould be renewed using a perfusion and suction pipette. When a change of bathingsolution was needed, the new solution was added via the perfusion pipette at a rate of150 ml min21 over a 3 min period. Perfusion was then stopped until the next change inbath solution.
The effects of ions and inhibitors on fluid secretion rate were tested in 143 mmol l21
Cl2 Ringer’s solutions, containing varying K+ concentrations (see Artificial salines andResults). As described previously (Weltens et al. 1992), in each experiment, secretionrate was expressed as a relative rate, i.e. as a percentage of the rate during the last controlperiod before the change to that experimental solution. Furthermore, to account for adecrease in fluid secretion rate with time, for each collection period the ratio of the meanvalue of the experimental series over the mean value of the same period of fluid secretionexperiments was calculated, where the tubules were bathed continuously in the controlsolution.
In order to test the statistical significance of the effect of a drug, the experimentalrelative rate was compared with the control relative rate at the same moment (unpairedStudent’s t-test). The drug effect was considered to be fully reversible when nostatistically significant difference (unpaired Student’s t-test) was found between therelative secretion rate during a wash-out period and the corresponding control period.Although we realize that this method was not ideal (the change in fluid secretion withtime in the absence of a test solution had to be evaluated on a different set of tubules), itseemed to be the most suitable way to distinguish drug effects from time effects.Corrected results, taking into account the spontaneous decline of the fluid secretion ratewith time are shown in the figures and tables (see Results).
Electrical potential difference measurements
The method has been described in detail previously (Leyssens et al. 1992).Measurements were performed with microelectrodes filled with 0.1 mol l21 KCl andconnected to a high-impedance electrometer (Duo 773, WPI) via a Ag/AgCl wire. Thereference electrode was a low-resistance (1 MV) 3 mol l21 KCl electrode earthed via a
125Effects of ions and inhibitors on Malpighian tubules

Ag/AgCl wire. In order to facilitate impalements, the Malpighian tubule was immobilizedin the bathing droplet by two holding pipettes. The secreted fluid remained in the tubule.The measurement of the basal membrane potential difference (Vbl) was accepted if asudden negative potential deflection occurred and was stable for at least a few minutesand if the electrode potential differed maximally by 3 mV from the baseline afterelectrode withdrawal. The transepithelial potential difference (Vte) was measured byadvancing the microelectrode through the cell layer into the lumen of the tubule; Vbl andVte were expressed with reference to the bath side. The apical potential difference (Vap)was calculated as the difference between the transepithelial and transbasal potentialdifferences and expressed with reference to the lumen [Vap=2(Vte2Vbl)].
The bathing droplet was continuously perfused at a constant rate of 150 ml min21.
K+-selective measurements
The technique used for the construction of double-barrelled ion-sensitive electrodeshas been described previously (Leyssens et al. 1993a). The tip of the ion-selective barrelwas filled with a short column of the ion-selective liquid K+ exchanger (Corning 477317),and the rest of the barrel was backfilled with 1 mol l21 KCl. The reference barrel wasbackfilled with 1 mol l21 sodium acetate + 10 mmol l21 KCl. The filled electrodes werebevelled in a rotating polishing alumina solution. After bevelling, the ion-sensitive andreference barrels had tip resistances ranging from 50 to 100 GV and from 0.5 to 1 GV,respectively. Three tracings were recorded on a pen recorder (Sefram or Linseis): thepotential of the reference barrel, the potential of the ion-selective barrel and the differencebetween them, representing the K+-sensitive signal.
The K+-sensitive signal was calibrated in a 143 mmol l21 Cl2 Ringer’s solution, inwhich the K+ concentration was varied from 5 to 113 mmol l21, using Na+ as a substitute.Interference by Na+ can be ignored (see Leyssens et al. 1993a). The measuredintracellular ([K+]i) or luminal ([K+]l) K+ ‘signal’ was always expressed as the K+
concentration that would be needed in a 143 mmol l21 Cl2 Ringer’s solution to result inthe same reading of the K+-selective barrel. It should be kept in mind that the value for[K+]i should, in fact, be corrected if the activity coefficient in the cell was different fromthat in the bath. Once the electrode has impaled the cell, it should, however, be possible toobserve changes in [K+]i without too much additional error. Other possible errors arisingfrom tip and liquid junction potentials of the reference electrode have been discussed byLeyssens et al. (1993a). Since the calibration curve has a logarithmic scale, in the highconcentration range (>100 mmol l21), small changes in intracellular and luminalconcentrations (i.e. less than 10 mmol l21) correspond to minor changes in the K+-sensitive signal (i.e. 1–2 mV). On evaluating the results, one has to take into account thatthis approaches the limit of detection.
The ion-sensitive electrodes were accepted for use if a stable potential signal wasobtained for both the ion-sensitive and the reference barrel, if the K+-sensitivity of thereference barrel was less than 1 mV and if the calibration curve could be fitted by astraight line with a slope of at least 48 mV per decade. Except when the effect of a certaintest solution on the fluid secretion rate or on the electrical potentials or concentrations wasnot clearly reversible, the value of a specific electrophysiological variable, registered
126 A. LEYSSENS AND OTHERS

before and after ion substitution or drug application, was averaged and taken as thecontrol value. In this way, possible time-dependent changes in potentials orconcentrations were minimized.
Artificial salines
The different control solutions containing varying K+ and Na+ concentrations (seeResults) were obtained by mixing a K+-containing, Na+-free and a Na+-containing, K+-free standard Ringer’s solution (2 mmol l21 CaCl2, 13 mmol l21 MgCl2, 12.1 mmol l21
Hepes, 2.8 mmol l21 alanine, 10.6 mmol l21 trehalose, 11.7 mmol l21 maltose,139 mmol l21 glucose and 113 mmol l21 KCl or NaCl, respectively; pH was adjusted to7.20 with KOH or NaOH, respectively; the osmolality of the solution was375 mosmol kg21 H2O).
Solutions were freshly prepared each week, filtered through 0.45 mm Millipore filtersand kept at 2 ˚C until use.
In some experiments, the following alterations in ion composition were made:6 mmol l21 BaCl2 was added to the bath solution; Cl2 was completely replaced by Br2;Na+ was completely replaced by N-methyl-D-glucamine (NMDG+, Sigma). On replacingNaCl by NMDG+, the alkaline solution was titrated with an appropriate amount of HClfor pH adjustment; as a result, the final Cl2 concentration remained virtually unchanged(i.e. 143 mmol l21).
The following pharmacological substances were tested: dihydro-ouabain (Sigma),sodium orthovanadate (Janssen Chimica), bumetanide (Leo Pharmaceutical Products),Schering 28080 (Schering-Plough; solvent, 0.05 % methanol and 0.05 %dimethylsulphoxide without any effect on fluid secretion; unpublished results).
Statistics
Results are presented as mean values ± S.E.M. Statistical computations were made usingStatview II (Abacus Concepts Inc., Berkeley, CA, USA, 1987). The fluid secretionmeasurements and electrophysiological measurements were evaluated using unpairedand paired two-tailed Student’s t-test, respectively, except when indicated differently. Avalue of P<0.05 was accepted as indicating statistical significance.
Results
Effects of Ba2+
K+-selective measurements
Ba2+ is an effective blocker of the K+ channels in the basal membrane of theMalpighian tubule cells of Formica polyctena. It probably reduces an inward K+ currentand, because of the activity of the electrogenic apical H+ pump, both apical and basalmembranes hyperpolarize (Weltens et al. 1992). In the present study, we compared theeffects of a change in the bath K+ concentration ([K+]bl) on [K+]i in the absence andpresence of 6 mmol l21 Ba2+. An example is shown in Fig. 1. In the absence of Ba2+, anincrease in bath K+ concentration ([K+]bl) from 5 to 113 mmol l21 resulted in a promptdepolarization of Vbl and a concomitant rise in [K+]i (see also Leyssens et al. 1993a).
127Effects of ions and inhibitors on Malpighian tubules

When [K+]bl was decreased to 51 mmol l21 [K+]bl, Vbl hyperpolarized but [K+]i remainedat the same value. On returning to 5 mmol l21, Vbl and [K+]i changed: [K+]i decreased butto a slightly higher value than the control and Vbl was also slightly more negative than atthe start of the experiment. In the presence of 6 mmol l21 Ba2+, the response of Vbl to anincrease in [K+]bl was greatly reduced: Vbl depolarized by only 7 mV for an increase in[K+]bl from 5 to 113 mmol l21. In contrast, the rise in [K+]i on increasing [K+]bl wascomparable to the response of [K+]i to a change in [K+]bl in the absence of Ba2+.
In the experiment shown, the addition of 6 mmol l21 Ba2+ itself caused a considerabledecrease of [K+]i in both 5 and 113 mmol l21 [K+]bl. This type of behaviour was observedin two out of the four experiments. In the other two, [K+]i increased. On average, in both 5and 113 mmol l21 K+, [K+]i was not altered significantly on adding Ba2+ to the bathsolution (Table 1). Furthermore, the summarized results demonstrate that, although theK+-sensitivity of Vbl was lost under Ba2+ treatment, suggesting a blockage of basal K+
channels, the rise in [K+]i on increasing [K+]bl was not affected.
Effects of substitution of Na+ by NMDG+
Fluid secretion measurements
In order to detect any Na+-dependent transport system (the Na+ site of the Na+/K+/2Cl2
cotransporter, for instance, cannot transport large cations such as NMDG+, see Hedge and
128 A. LEYSSENS AND OTHERS
5113 51
0
5 113
6
200
10050
20105 1 min
[K+] i
(mm
ol l−1
)V
bl (
mV
)
−120
−100
−80
0
[Ba2+] (mmol l−1)[K+] (mmol l−1)
Cell
−20
−40
−60
Fig.1 at 50%Fig. 1. Effect of a change in bath K+ concentration on the basal membrane potential (Vbl,upper trace) and the intracellular K+ concentration ([K+]i, lower trace) in the absence andpresence of 6 mmol l21 Ba2+. On changing the bath solution, electrical disturbances wererecorded by the K+-selective (i.e. the most sensitive) barrel and these have been omitted fromthe figure for clarity.
Palfrey, 1992), Na+ was completely replaced by NMDG+. In 51 mmol l21 [K+]bl, therewas no significant effect (Table 2A). In a low-K+ solution (10 mmol l21 [K+]bl), thesubstitution of Na+ by NMDG+ caused a small but reversible inhibition of fluid secretion(Table 2A).
Effects of substitution of Cl2 by Br2
Fluid secretion experiments
It is known that the Na+/K+/2Cl2 and K+/Cl2 cotransport systems have a differentanion-dependence (Palfrey and Greengard, 1981; Ellory and Hall, 1988). When Cl2 wasomitted from the bath solution and replaced by Br2, fluid secretion rate was significantlystimulated in 51 mmol l21 [K+]bl but inhibited in 10 mmol l21 [K+]bl containing a highNa+ concentration (Table 2A).
Effects of dihydro-ouabain
Fluid secretion experiments
As a high K+ concentration is competitive with ouabain (see Baker and Willis, 1970),the effect of dihydro-ouabain (DHO) was tested in a low [K+]bl (5 mmol l21). Nosignificant effect of a high dose of dihydro-ouabain (1023 mol l21) was observed duringthe 30 min application of the drug (Table 2B). However, in this low [K+]bl, fluid secretionrate was very slow before DHO was applied (i.e. 76±19 pl min21, N=8) and the timecourse was highly variable. Consequently, it was difficult to distinguish a small drugeffect from the time-dependent changes.
Electrical potential difference measurements
The effect of dihydro-ouabain was also tested on Vbl (Fig. 2A): if dihydro-ouabainwere to reduce [K+]i, a depolarization of Vbl would be expected. However, in 5 mmol l21
[K+]bl, ouabain treatment (1023 mol l21) for at least 10 min did not depolarize Vbl. If theputative pump were electrogenic, its effect on Vbl might be masked by the high K+
permeability of the basal membrane. Therefore, the potential measurements were also
129Effects of ions and inhibitors on Malpighian tubules
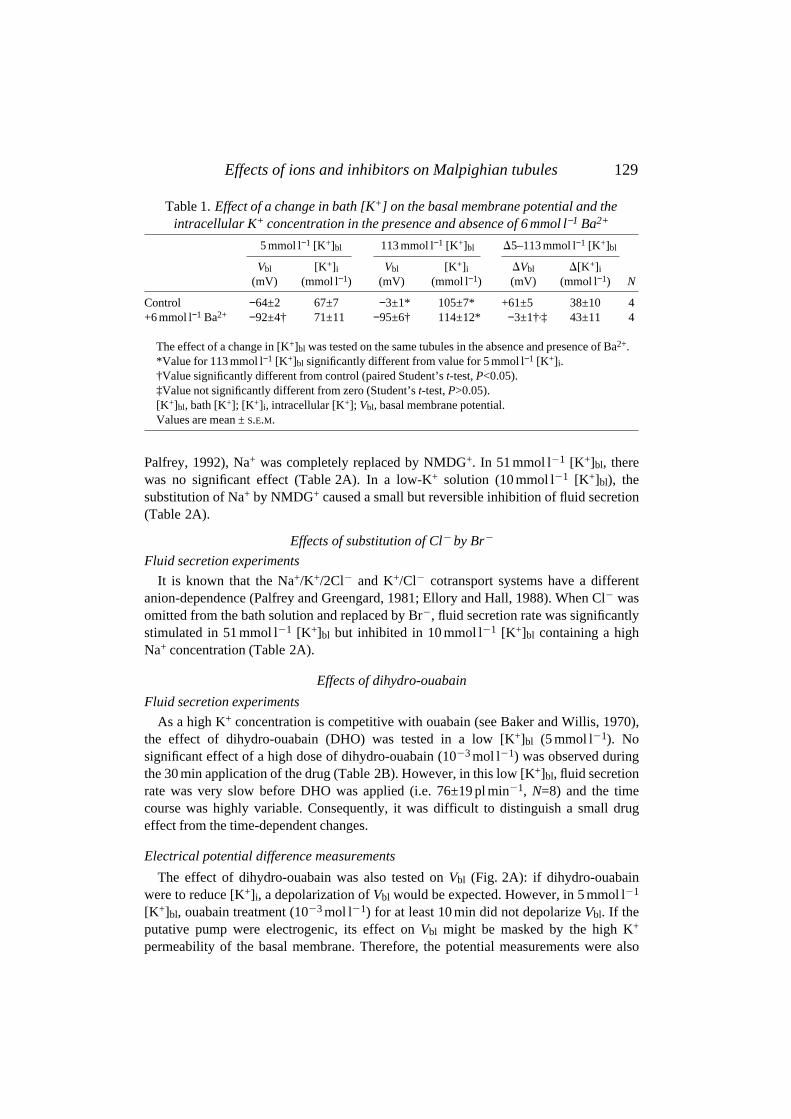
Table 1. Effect of a change in bath [K+] on the basal membrane potential and theintracellular K+ concentration in the presence and absence of 6 mmol l−1 Ba2+
5 mmol l−1 [K+]bl 113 mmol l−1 [K+]bl D5–113 mmol l−1 [K+]bl
Vbl [K+]i Vbl [K+]i DVbl D[K+]i
(mV) (mmol l−1) (mV) (mmol l−1) (mV) (mmol l−1) N
Control −64±2 67±7 −3±1* 105±7* +61±5 38±10 4+6 mmol l−1 Ba2+ −92±4† 71±11 −95±6† 114±12* −3±1†,‡ 43±11 4
The effect of a change in [K+]bl was tested on the same tubules in the absence and presence of Ba2+.*Value for 113 mmol l−1 [K+]bl significantly different from value for 5 mmol l−1 [K+]i.†Value significantly different from control (paired Student’s t-test, P<0.05).‡Value not significantly different from zero (Student’s t-test, P>0.05).[K+]bl, bath [K+]; [K+]i, intracellular [K+]; Vbl, basal membrane potential.Values are mean ± S.E.M.

performed in the presence of 6 mmol l21 Ba2+ (Fig. 2B) (for theoretical considerations,see also Weltens et al. 1992). The addition of 6 mmol l21 Ba2+ hyperpolarized Vbl. In thepresence of Ba2+ and following exposure to 1023 mol l21 dihydro-ouabain, Vbl decreasedslightly; but after wash-out, Vbl declined a further 1 mV, suggesting a small time-dependent decay of Vbl. After wash-out of Ba2+, Vbl depolarized to approximately itsinitial control value. The potential measurements are summarized in Table 3A.Significant effects of 1023 mol l21 dihydro-ouabain on Vbl were detected neither in theabsence nor in the presence of 6 mmol l21 Ba2+.
130 A. LEYSSENS AND OTHERS
Table 2. Effect of ion substitution and drugs on fluid secretion
Fluid secretion rateBath [K+] experimental/control
Ion substitution (mmol l−1) N (%)
A Effect of ion substitutionNa+ by NMDG+ 10 9/8 70±11*
51 6/9 111±15
Cl− by Br− 10 7/8 69±10*51 8/8 157±38*
Fluid secretion rate[Drug] Bath [K+] experimental/control
Drug (mol l−1) (mmol l−1) N (%)
B Effect of drugsDihydro-ouabain 10−3 5 8/8 74±24
Sch28080 10−4 10 6/9 130±1551 8/8 129±18
Vanadate 10−4 51 8/9 108±1810−3 51 6/9 8±3*
113 6/7 30±12*
Bumetanide 10−5 5 8/8 37±11*51 8/9 111±17
10−4 5 8/8 22±8*,†51 7/9 46±8*
113 8/7 97±1310−3 51 6/9 7±3*,†
N, number of tubules experimental/control.The control bath Na+ concentration was 0, 62, 103 or 108 mmol l−1 when the bath K+ concentration
was 113, 51, 10 or 5 mmol l−1, respectively; the control bath Cl− concentration was always 143 mmol l−1.*Statistically significant difference between the experimental relative rate and the control relative rate
during the same time period (unpaired Student’s t-test, P<0.05).†Statistically significant difference (unpaired Student’s t-test) between the relative secretion rate of
the experimental and the control series during a wash-out period; this means that the effect was notcompletely reversible.
Values are mean ± S.E.M.

Effects of Sch28080
Fluid secretion experiments
At 1024 mol l21, Sch28080 would only affect a K+/H+-ATPase, if present, or a V-typeH+-ATPase and not the activity of a Na+/K+-ATPase (see Froissart et al. 1992). Itsinhibitory action increases at lower bath K+ concentrations (see Scott et al. 1987). In ourhands, Sch28080 (1024 mol l21) had no effect on fluid secretion rate either in a high(51 mmol l21) or in a low (10 mmol l21) [K+]bl (Table 2B).
Effects of vanadate
Fluid secretion experiments
In 51 mmol l21 [K+]bl, 1024 mol l21 vanadate, known to be an inhibitor of E1E2 (P-type) ATPases (i.e. the Na+/K+- and K+/H+-ATPase) (see Nechay et al. 1986), did notaffect the fluid secretion rate; but at 1023 mol l21, fluid secretion was almost completelyabolished within 40 min (Fig. 3A, Table 2B). In a high-K+, Na+-free solution, vanadate(1023 mol l21) caused a similar inhibitory effect on fluid secretion (Fig. 3B, Table 2B).There was no statistically significant difference between the inhibitory effects in 51 or113 mmol l21 [K+]bl. In both cases, the effect of vanadate was reversible.
Electrical potential difference measurements
The effect of vanadate was tested on intracellular and luminal potentials in a differentseries of tubules (Table 3A). In 5 and 51 mmol l21 [K+]bl, at a concentration of
131Effects of ions and inhibitors on Malpighian tubules
A
[DHO] (mol l−1)
Vbl
(m
V)
0
50
51
10−3
Cell
1 min
Vbl
(mV
)
B
0−20
−40−60
−80−100
Cell5
00 10−3
6 00
1 min
[Ba2+] (mmol l−1)
[K+] (mmol l−1)
[DHO] (mol l−1)
[K+] (mmol l−1)
−20
−40
−60
Fig2 at 50%
Fig. 2. Effect of 1023 mol l21 dihydro-ouabain (DHO) on Vbl in the absence (A) and presence(B) of 6 mmol l21 Ba2+ (5 mmol l21 [K+]bl).

132 A. LEYSSENS AND OTHERST
able
3.E
ffec
t of d
rugs
on
elec
trop
hysi
olog
ical
var
iabl
es
Bat
hC
ontr
ol+
drug
[Dru
g]so
lutio
nV
blV
teV
apV
blV
teV
ap
Dru
g(m
oll−
1 )(m
mol
l−1 )
(mV
)(m
V)
(mV
)(m
V)
(mV
)(m
V)
AE
ffec
t on
cellu
lar a
nd lu
min
al e
lect
rica
l pot
entia
l mea
sure
men
tsD
ihyd
ro-o
uaba
in10
−35
[K+] b
l−6
1±1
(8)
24±
8 (5
)−8
5−6
0±1
(8)
23±
8 (5
)−8
35
[K+] b
l−9
3±4
(8)
14±
1 (6
)−1
07−9
0±4
(8)
13±
2 (6
)−1
03+
6 B
a2+
Van
adat
e10
−451
[K+] b
l−
25±
4 (5
)−
−22
±2
(5)
−10
−35
[K+] b
l−6
1±2
(4)
14±
3 (7
)−7
5−6
2±3
(4)
8±1*
(7)
−70
51 [K
+] b
l−1
6±1
(8)
23±
2 (9
)−3
9−1
7±2
(8)
12±
1* (9
)−2
951
[K+] b
l−5
4±4
(10)
−−
−30±
7* (1
0)−
−+
6 B
a2+
Bum
etan
ide
10−4
5 [K
+] b
l−
29±
6 (4
)−
−27
±5
(4)
−51
[K+] b
l−
35±
3 (1
1)−
−33
±3
(11)
−
Con
trol
+dr
ug
Bat
hC
ell
Lum
enC
ell
Lum
en
[Dru
g]so
lutio
nV
bl[K
+] i
Vte
[K+] l
Vbl
[K+] i
Vte
[K+] l
Dru
g(m
oll−
1 )(m
mol
l−1 )
(mV
)(m
mol
l−1 )
(mV
)(m
mol
l−1 )
(mV
)(m
mol
l−1 )
(mV
)(m
mol
l−1 )
BE
ffec
t on
cellu
lar a
nd lu
min
al K
+-s
elec
tive
mea
sure
men
tsV
anad
ate
10−3
51 [K
+] b
l−2
1±2
(4)
119±
13 (4
)22
±3
(7)
113±
23 (7
)−2
1±1
(4)
120±
12 (4
)12
±1
(7)*
120±
16 (7
)
Bum
etan
ide†
10−4
5 [K
+] b
l−5
2±3
(5)
52±
5 (5
)−
−−5
5±5
(5)
59±
7 (5
)−
−51
[K+] b
l−1
1±1
(5)
88±
6 (5
)−
−−1
2±2
(5)
89±
4 (5
)−
−
*Sig
nific
ant d
rug
effe
ct (p
aire
d St
uden
t’s
t-te
st, P
<0.
05).
†The
eff
ect o
f bum
etan
ide
was
test
ed o
n th
e sa
me
tubu
les
in 5
and
51
mm
oll−
1[K
+] b
l.V
alue
s ar
e m
ean
±S.
E.M
.(N
).[K
+] i,
intr
acel
lula
r [K
+];
[K+] b
l, ba
th [K
+];
[K+] l,
lum
inal
[K+];
Vbl
, bas
al m
embr
ane
pote
ntia
l; V
te, t
rans
epith
elia
l mem
bran
e po
tent
ial.

133Effects of ions and inhibitors on Malpighian tubules
6
51
10−30
[Ba2+] (mmol l−1)
−40
−20
0
Vbl
(m
V)
1 min
Cell
C
6/9 ( )
[Vanadate] (mol l−1)
[K+] (mmol l−1)
0 0
51
N = 8/9 ( )
0 20 40 60 80 100Time (min)
20
40
60
80
100
120
140
Flui
d se
cret
ion
rate
,ex
peri
men
tal/c
ontr
ol (
%)
**
0
0 010−3
113
N = 6/7
0 20 40 60 80 100Time (min)
0
20
40
60
80
100
120
Flui
d se
cret
ion
rate
,ex
peri
men
tal/c
ontr
ol (
%) 140
** *
51
A B
51 51
0
−60
−60
−80
−40
−20
15010050
20105
10−30
0
Vbl
(m
V)
Cell
[K+] i
(m
mol
l−1)
1 min
D
15010050
20105 1 min
5
Lumen
40
Vte
(m
V)
20
0
10−30
E
[Vanadate] (mol l−1)
[K+] (mmol l−1)
[K+] (mmol l−1)
[Vanadate] (mol l−1)
[K+] (mmol l−1)
[Vanadate] (mol l−1)
[K+] (mmol l−1)
[Vanadate] (mol l−1)
[K+] l
(m
mol
l−1)
Fig. 3. Effect of vanadate on the fluid secretion rate (A) in 51 mmol l21 [K+]bl [at 1024 (s) and1023 (d) mol l21] and (B) in 113 mmol l21 [K+]bl (Na+-free) (at 1023 mol l21) [K+]bl. Meanvalues ± S.E.M. *Statistically significant difference between the experimental relative fluidsecretion rate and the rate in the control series at the same moment (unpaired Student’s t-test,P<0.05). There was no statistically significant difference between the inhibitory effect ofvanadate in 51 and 113 mmol l21 [K+]bl (unpaired Student’s t-test, P>0.05). (C) Effect of1023 mol l21 vanadate on Vbl in the presence of 6 mmol l21 Ba2+ (51 mmol l21 [K+]bl). (D)Effect of 1023 mol l21 vanadate on Vbl and [K+]i and (E) on Vte and the luminal K+
concentration ([K+]l) in 51 mmol l21 [K+]bl. The intracellular and luminal K+-selectivemeasurements were performed on different tubules.

1023 mol l21, vanadate did not affect Vbl but significantly depolarized Vte; within 5 min, asteady state was reached. 1024 mol l21 vanadate had no significant effect on Vte. Thedepolarizing effect of 1023 mol l21 vanadate on Vte suggested the inhibition of anelectrogenic transport system (for theoretical considerations, see also Weltens et al.1992). Therefore, it was of interest to examine the effects of vanadate on the Ba2+-induced hyperpolarization of Vbl. A typical example is shown in Fig. 3C. Vanadatecaused a depolarization (i.e. a reduction of the Ba2+-induced hyperpolarization) of Vbl
within 5 min. The effect was statistically significant (Table 3A). The effects of vanadateon electrical potentials were not easily reversible within the experimental period (i.e.approximately 15 min wash-out).
K+-selective measurements
Since 1023 mol l21 vanadate did not affect Vbl in the absence of Ba2+ within 5 min, adrastic effect on [K+]i was not expected. This was verified in 51 mmol l21 [K+]bl.Examples of intracellular and luminal measurements are given in Fig. 3D,E. Theaddition of 1023 mol l21 vanadate did not cause significant changes in Vbl or [K+]i. In thelumen of another tubule, the reversible response of Vte and [K+]l to a decrease in [K+]bl
in control conditions was first confirmed. Upon administation of 1023 mol l21 vanadate,Vte began to decrease within the first minute; after 6 min, Vte had decreased by 18 mV.Within the same period, [K+]l was not affected. The experimental data are summarizedin Table 3B. There was no significant effect on [K+]i and [K+]l. A similar lack of effecton [K+]i (N=2) and [K+]l (N=3) was observed (not shown) in 5 mmol l21 [K+]bl.
Effects of bumetanide
Fluid secretion experiments
The loop diuretic bumetanide inhibits K+/Cl2 and Na+/K+/2Cl2 cotransport systems(see Ellory and Hall, 1988). A concentration of 1025 mol l21 is usually sufficient formaximal inhibition of the Na+/K+/2Cl2 cotransporter, but much higher concentrationsare generally needed to affect the K+/Cl2 cotransporter (see Ellory and Hall, 1988;Palfrey and O’Donnell, 1992). Bumetanide was tested in 5, 51 and 113 mmol l21 [K+]bl.In 51 mmol l21 [K+]bl, 1024 mol l21 bumetanide significantly diminished fluid secretionrate; the application of 1023 mol l21 almost completely abolished it (Fig. 4A, Table 2B).Only at 1024 mol l21 was the effect reversible. In the lower [K+]bl (5 mmol l21),bumetanide reversibly inhibited fluid secretion when applied at 1025 mol l21. Theinhibitory effect of 1024 mol l21 bumetanide was more pronounced and the fluidsecretion did not recover within the 30 min wash-out period (Fig. 4B, Table 2B). In thehigh-K+, Na+-free solution, in contrast, bumetanide (1024 mol l21) had no significanteffect (Fig. 4C, Table 2B).
Electrical potential measurements
Since bumetanide (1024 mol l21) diminished the fluid secretion rate in 51 and5 mmol l21 [K+]bl, electrophysiological effects were tested in these bath solutions. As isshown in Fig. 4D, in 51 mmol l21 [K+]bl, Vte remained constant after the addition of1024 mol l21 bumetanide; in 5 mmol l21 [K+]bl, Vte was 4 mV higher than the control
134 A. LEYSSENS AND OTHERS

value. On average (see Table 3A), neither in 5 nor 51 mmol l21 [K+]bl could a significanteffect on Vte be detected. In 2 out of the 11 experiments performed in 51 mmol l21 [K+]bl,the drug was applied for more than 20 min without any clear change in potential
135Effects of ions and inhibitors on Malpighian tubules
A B
[K+] (mmol l−1)
**
[Bumetanide] (mol l−1) 0 0
51
N = 8/9 ( ) 7/9 ( ) 6/9 ( )
0 20 40 60 80 100Time (min)
0
2040
6080
100
120
Flui
d se
cret
ion
rate
,ex
peri
men
tal/c
ontr
ol(%
)
***
*
** * *
* * *
0 0
5
0
N = 8/8
40 60 80Time (min)
0
2040
100
120
20
* ** * * *
6080
C D
E
0
20
40
5 51
0
Vte
(m
V)
1 min
Lumen
0 0
113
N = 8/7
0 20 40 60 80 100Time (min)
0
2040
60
80
100
120
140
Flui
d se
cret
ion
rate
,ex
peri
men
tal/c
ontr
ol(%
)
10−410−45
5 1 min
0−20−40
5 51
0
Vbl
(m
V)
[K+] i
(mm
ol l−1
)
−60
5 51 5
0
102050
100
Cell
−80
10−4
[K+] (mmol l−1)
[Bumetanide] (mol l−1)
[K+] (mmol l−1)
[Bumetanide] (mol l−1)[K+] (mmol l−1)
[Bumetanide] (mol l−1)
[K+] (mmol l−1)
[Bumetanide] (mol l−1)
Fig. 4. Effect of bumetanide on the fluid secretion rate (A) in 51 mmol l21 [K+]bl at 1025 (s),1024 (d) and 1023 (n) mol l21, (B) in 5 mmol l21 [K+]bl at 1025 (s) and 1024 (d) mol l21
and (C) in 113 mmol l21 [K+]bl (Na+-free) at 1024 mol l21. Mean values ± S.E.M. *Statisticallysignificant difference between the experimental relative fluid secretion rate and the rate in thecontrol series at the same moment (unpaired Student’s t-test, P<0.05). Effect of 1024 mol l21
bumetanide (D) on Vte and (E) on Vbl and [K+]i in 51 and 5 mmol l21 [K+]bl.

difference being observed.
K+-selective measurements
The effect of 1024 mol l21 bumetanide was tested on Vbl and [K+]i in 5 and 51 mmol l21
[K+]bl on the same tubule (Fig. 4E). In control conditions, an increase in [K+]bl from 5 to51 mmol l21 caused a depolarization of Vbl and an increase in [K+]i. On introduction ofthe bumetanide-containing bath solution in 51 mmol l21 [K+]bl, Vbl increased slightly and[K+]i remained virtually unaffected. When [K+]bl was lowered to 5 mmol l21 in thepresence of the drug, Vbl hyperpolarized and [K+]i decreased. On returning to51 mmol l21 [K+]bl in the presence of bumetanide, Vbl depolarized again, while [K+]i
approached its previous value in 51 mmol l21 [K+]bl. Thus, bumetanide did not inhibit therise of [K+]i on increasing [K+]bl. Washing out bumetanide in 5 mmol l21 [K+]bl did notchange the Vbl and [K+]i values measured in the presence of bumetanide. The small(irreversible) changes in Vbl and [K+]i occurring in this experiment during administrationof bumetanide were not systematically observed in other experiments. On average, therewas no statistically significant effect of 1024 mol l21 bumetanide on Vbl or [K+]i
(Table 3B). The lack of an effect on Vbl was confirmed in eight experiments performed in51 mmol l21 [K+]bl with 57 mmol l21 Cl2 in the solution (Cl2 substituted by citrate): evenafter 20 min of application of 1023 mol l21 bumetanide, a concentration which effectivelyinhibited fluid secretion (Verhulst et al. 1988), Vbl did not change significantly (R.Weltens, personal communication).
Combined effect of Ba2+ and bumetanide
Both Ba2+ and bumetanide are able to impair fluid secretion substantially, probably byaffecting different mechanisms. Neither Ba2+ nor bumetanide seemed to block a changein intracellular [K+] when applied separately. It was of interest, therefore, to observe theircombined action on fluid secretion and K+-selective measurements.
Fluid secretion experiments
After three periods of 10 min in control Ringer containing 51 mmol l21 [K+]bl and143 mmol l21 Cl2, 6 mmol l21 Ba2+ was added during the following three 10 min periods.This caused a decrease in fluid secretion to 18±10 % (N=3) of that in the previous controlperiod. Over the next 30 min, 1024 mol l21 bumetanide was applied in the presence ofBa2+. Fluid secretion was completely inhibited within 20 min. The effect was only poorlyreversible within the wash-out period (23±12 % recovery after 30 min). In a controlseries, 6 mmol l21 Ba2+ alone was added for 60 min (N=3). This treatment also resulted incomplete inhibition of fluid secretion.
Electrical potential and K+-selective measurements
In another series of experiments, the effects of a change in [K+]bl on Vbl and [K+]i wereexamined in the same cell of an isolated tubule, first in the presence of 6 mmol l21 Ba2+
and then in the presence of Ba2+ together with 1024 mol l21 bumetanide. The change in[K+]bl from 113 to 5 mmol l21 was carried out in both directions. The results aresummarized in Table 4. Surprisingly a change in [K+]i was still observed when both
136 A. LEYSSENS AND OTHERS

bumetanide and Ba2+ were present in the medium, although it was statistically smaller by6±1.5 mmol l21 (paired t-test, N=6) than in the presence of Ba2+ alone (P<0.01, paired t-test).
In this series of experiments, the tubules were less sensitive to Ba2+. Ba2+ still causedsome hyperpolarization of Vbl (compare Vbl with the control values in Table 1 and inWeltens et al. 1992), but the effect of [K+]bl on Vbl was only partly abolished by Ba2+
(compare the effect in Tables 1 and 4).
Discussion
Basal K+ channels are not the only K+ entry mechanism in Malpighian tubules ofFormica polyctena
In the present study, we have shown that in the presence of 6 mmol l21 Ba2+ in responseto a rise in bath [K+], [K+]i increased to the same extent and with a similar time course asin the absence of Ba2+ (Fig. 1 and Table 1). This result and previous findings in Formicapolyctena (see Introduction for references) strongly argue for the role of other K+ uptakesystems, besides entry through conductive channels, in maintaining fluid secretion and inadapting the cytosolic K+ concentration to the surrounding K+ concentration.
Ba2+ itself seemed to have contradictory effects on [K+]i: in some experiments, [K+]i
increased in the presence of Ba2+; in others, it decreased. As Ba2+ slows down both K+
entry across the basal membrane and K+ exit across the apical membrane (see Leyssenset al. 1993b), a difference in balance between these two effects might explain the rise in[K+]i in some tubules and the decrease in others.
Basal primary active K+-transporting ATPases
A Na+/K+-ATPase has been found in insect Malpighian tubules, but only in a fewspecies does its activity affect fluid secretion significantly (for a review, see Nicolson,1993).
In Malpighian tubules of Formica polyctena fluid secretion continued at a very highrate in a high-K+ (113 mmol l21) but completely Na+-free solution (Van Kerkhove et al.
137Effects of ions and inhibitors on Malpighian tubules
Table 4. Effect of 10−4 mol l−1 bumetanide on the change in Vbl and [K+]i induced by anincrease in bath K+ concentration in the presence of Ba2+
5 mmol l−1 [K+]bl 113 mmol l−1 [K+]bl D5–113 mmol l−1 [K+]bl
Vbl [K+]i Vbl [K+]i DVbl D[K+]i
Treatment (mV) (mmol l−1) (mV) (mmol l−1) (mV) (mmol l−1) N
6 mmol l−1 Ba2+ −75±4 63±7 −40±9 96±4 35±8 33±6 86 mmol l−1 Ba2+ + −73±7 59±8 −37±10 85±5 36±9 26±7 6
10−4 mol l−1 bumetanide
In six of the eight tubules, the change in [K+]bl was tested on the same tubules in the absence andpresence of bumetanide.
[K+]bl, bath [K+]; Vbl, basal membrane potential; [K+]i, intracellular [K+].

1989), and omission of Na+ from a 51 mmol l21 K+ solution had no effect (Table 2A).The latter findings do not support a significant role for any Na+-dependent K+ uptakemechanism in fluid secretion by Malpighian tubules of Formica polyctena, at least in thepresence of relatively high K+ concentrations.
In low K+ concentrations, omission of Na+ from the bath decreased fluid secretion(Table 2A), but the Na+/K+-ATPase inhibitor ouabain was without effect in the presenceof 5 mmol l21 K+ on either fluid secretion (Table 2B) or Vbl in the presence or absence ofBa2+ (Fig. 2A,B, Table 3A). Thus, although the insensitivity of Na+/K+-ATPase toouabain in Formica polyctena tubules, or even excretion and sequestration of ouabain(for a review, see Vaughan and Jungreis, 1977; Anstee and Bowler, 1979), cannot beexcluded, our results suggest that, even in a low-K+ medium, an active Na+/K+ pump is oflittle importance for transepithelial K+ transport in Malpighian tubules of Formicapolyctena. The small Na+-dependence of the fluid secretion rate in low [K+]bl canprobably be explained by the presence of another Na+-dependent transport system, i.e. aNa+/K+/2Cl2 cotransport system (see below).
Another candidate for active K+ uptake, the K+/H+-ATPase, if present, does not seemto play any role in fluid secretion: no effect of its blocker Sch28080 could be detected.
Since vanadate interacts with both Na+/K+- and K+/H+-ATPases, no significant effecton fluid secretion was expected on adding vanadate to the bath solution. Surprisingly,when applied at 1023 mol l21, vanadate strongly inhibited the fluid secretion rate, theinhibitory effect becoming statistically significant after 20 min (Fig. 3A). In both 5 and51 mmol l21 [K+]bl, Vte significantly depolarized to a new steady-state level within 5 minof the addition of vanadate. The observation that Vbl, [K+]i and [K+]l remained unaffected(Fig. 3D and Table 3A,B) seems to indicate that K+(Cl2?) and water transport wereaffected to the same extent: the speed of passage of K+ and water was slowed downwithout changing the actual K+ concentrations. This, in itself, seems to exclude an effectof vanadate on a Na+/K+-ATPase, as this would have altered the K+ and Na+
concentrations in the cell and, consequently, Vbl. Furthermore, since the inhibitory effectsof vanadate on fluid secretion rate in Na+-containing (i.e. 51 mmol l21 [K+]bl) and Na+-free (i.e. 113 mmol l21 [K+]bl) conditions were comparable (Fig. 3A,B, Table 2B), itsstrong inhibitory effect was probably not due to an interaction with a hypotheticalNa+/K+-ATPase. These results are different from the response of Malpighian tubules ofLocusta migratoria, where both vanadate and ouabain caused a slow depolarization ofboth Vbl and Vap (Baldrick et al. 1988).
Because in Formica polyctena there was no evidence for the presence of a K+/H+-ATPase either (see above), we had to consider other possible sites of action. A complexindirect effect of vanadate was suggested for Malpighian tubules of Drosophilamelanogaster (Bertram et al. 1991). A multitude of stimulatory and inhibitoryintracellular mechanisms for vanadate have been reviewed by Chasteen (1983) andNechay et al. (1986). The effects on the fluid secretion rate and electrophysiologicalvariables of Formica polyctena Malpighian tubules were comparable to, and followed asimilar time course to, the effects of the V-type ATPase inhibitors bafilomycin A1 and N-ethylmaleimide (NEM) reported by Weltens et al. (1992) on the same preparation. In thelatter study, the reduction of the Ba2+-induced hyperpolarization was attributed to an
138 A. LEYSSENS AND OTHERS

electrogenic current generated by the apical H+-ATPase and blocked by bafilomycin A1
and NEM. Unlike ouabain (in 5 mmol l21 [K+]bl), vanadate (1023 mol l21 in 51 mmol l21
[K+]bl) did significantly reduce the Ba2+-induced hyperpolarization of Vbl (Fig. 3C, Table3A).
Reports of a direct effect of vanadate on the apical H+ pump are controversial.According to Forgac (1989), vacuolar-type H+-ATPases are insensitive up to at least1 mmol l21. Chatterjee et al. (1992), however, described the presence of a novel V-typeH+-ATPase in osteoclast plasma membrane vesicles with a unique pharmacology andwith specific isoforms of two subunits in the catalytic portion of the enzyme; the H+-ATPase activity was completely inhibited by 1023 mol l21 vanadate. 531024 mol l21
vanadate caused 50 % inhibition of the ATP-dependent proton uptake of isolatedchromaffin granules and yeast vacuoles (Beltrán and Nelson, 1992). This is in line with asmall effect (10–15 % inhibition only) of a lower concentration of vanadate(1024 mol l21) on the cation-stimulated ATPase activity in purified goblet cell apicalmembranes of Manduca sexta (Wieczorek et al. 1986), later identified as a V-type H+-ATPase (Wieczorek et al. 1989). The effect was not significant and was considered to benegligible by the authors at that time, but might represent the threshold for vanadate-sensitivity of this ATPase.
In summary, no conclusive evidence was found for the existence of basal active K+
uptake mechanisms in exchange for either Na+ or H+. Their role in maintaining anasymmetric ion distribution across the basal membrane and a particular rate of fluidsecretion is apparently minimal. The strong inhibitory effect of vanadate on fluidsecretion can probably be explained by an effect on the apical H+ pump.
Coupled entry mechanisms for K+, Cl2 (and Na+) in the basal membrane
The existence of coupled entry mechanisms for K+, Na+ and Cl2 has been investigatedin Malpighian tubules of other insects, but only in a low bath K+ concentration (for areview, see Nicolson, 1993). In Malpighian tubules of Formica polyctena a moreextensive study was carried out to find evidence for an electroneutral coupled transport ofK+ in different bath K+ (and Na+) concentrations.
In high-K+ (113 mmol l21) Na+-free conditions, K+ entry via a Na+/K+/2Cl2
cotransporter can be excluded. K+ uptake via a K+/Cl2 cotransporter is not very likelyeither, as bumetanide had no effect at a concentration as high as 1024 mol l21 (Fig. 4C,Table 2B).
In 51 mmol l21 [K+]bl, the fluid secretion rate was not affected by omitting Na+ fromthe bath solution (Table 2A). Although bumetanide binding to a Na+/K+/2Cl2
cotransporter should be favoured in the 51 mmol l21 K+, 62 mmol l21 Na+ bath solution(see Haas and Forbush, 1986), no change in fluid secretion rate was observed after theaddition of 1025 mol l21 bumetanide (Fig. 4A, Table 2B). These findings do not favourK+ entry via a Na+/K+/2Cl2 cotransporter. However, the significant reduction of the fluidsecretion rate after the addition of 1024 mol l21 bumetanide suggests that a large portionof the basal K+ uptake may occur via a K+/Cl2 cotransporter (Fig. 4A, Table 2B). Since,at this concentration, no effect of bumetanide was observed in 113 mmol l21 [K+]bl,specific effects are unlikely. The observation that Br2 has a stimulatory effect on fluid
139Effects of ions and inhibitors on Malpighian tubules

secretion in 51 mmol l21 [K+]bl corroborates the hypothesis of a K+/Cl2 cotransportsystem (Table 2A; see Ellory and Hall, 1988). At 1023 mol l21, the inhibitory effect ofbumetanide was much more pronounced. However, at this high concentration, interactionwith other Cl2-dependent transport mechanisms cannot be excluded; in whole-cellrecordings of the rat lacrimal gland, it was shown that 431024 mol l21 bumetanideinhibited Cl2 channels, for instance (Evans et al. 1986).
In a low-K+, high-Na+ medium (i.e. 10 or 5 mmol l21 [K+]bl), both Na+ omission andCl2 substitution by Br2 caused a reversible reduction of the secretion rate (Table 2A),and bumetanide clearly inhibited fluid secretion at 1025 mol l21 (see Fig. 4B,Table 2B). These findings argue for the presence of a Na+/K+/2Cl2 cotransporter(Ellory and Hall, 1988). Evidence for a Na+/K+/2Cl2 cotransporter (Maddrell, 1969;O’Donnell and Maddrell, 1984; Hegarty et al. 1991) and/or Cl2-dependent K+ uptake(Wessing et al. 1987; Baldrick et al. 1988) has also been found for Malpighian tubulesof other species.
That bumetanide (and/or another loop diuretic, furosemide, for other species) affectselectroneutral transport mechanisms is confirmed by the lack of a significant effect onelectrical potential differences (Table 3A,B) (cf. Vbl in Locusta migratoria, Baldricket al. 1988, and Onymacris, Nicolson and Isaacson, 1987; Vap in Aedes aegypti,Hegarty et al. 1991). Long-term application, however (several minutes), might changethe intracellular K+, Cl2 and/or Na+ concentrations and hence Vbl (cfr. Rhodniusprolixus, O’Donnell and Maddrell, 1984; Aedes aegypti, Hegarty et al. 1991). InMalpighian tubules of Formica polyctena, [K+]i did not change much, as Vbl did notalter.
An important observation was that bumetanide did not hamper the response of [K+]i toa change in [K+]bl from 5 to 51 mmol l21 (Fig. 4E, Table 3B). This means that otherpathways, probably K+ channels, can ensure basal K+ uptake and adapt the intracellularK+ content to the surrounding K+ concentration. This finding indirectly points to theexistence of an inwardly directed basal electrochemical K+ gradient in the latterconditions.
To summarize, in a high-K+ (113 mmol l21) Na+-free solution, coupled entry of K+ andCl2 seems to be of little importance. In 51 mmol l21 [K+]bl, a substantial portion of K+
entry can occur via a K+/Cl2 cotransporter. In low [K+]bl, a Na+/K+/2Cl2 cotransporterseems to become important.
Combined effects of Ba2+ and bumetanide
When both Ba2+ and bumetanide were present, fluid secretion was completely blocked.It was possible, however, that this was due solely to the continued presence of Ba2+. Also,the effect of [K+]bl on [K+]i was reduced, but not abolished. This could mean either thatthe proposed model (see below) is still incomplete and that other K+ uptake mechanismsare present in the basal membrane or that Ba2+ does slow down K+ entry, but does notcompletely block it (see, for instance, the difference in sensitivity to Ba2+ of the series oftubules in Tables 1 and 4). Dijkstra et al. (1994) found that Ba2+ always increased thebasal membrane resistance, but that the hyperpolarization of Vbl was smaller in the anttubules in their study than in those investigated by Weltens et al. (1992). It would be
140 A. LEYSSENS AND OTHERS

interesting to study the Ba2+-sensitivity of single K+ channels using the patch-clamptechnique. The observation that Ba2+ can completely block fluid secretion if applied forlong enough can be explained by its concomitant hyperpolarizing effect on the apicalmembrane. This will severely reduce the turnover of the electrogenic H+ pump and thebuild-up of the proton concentration gradient that is needed to drive K+ into the lumen(see Leyssens et al. 1993b).
Transport model for transcellular K+ transport in Malpighian tubules of Formicaployctena
Several hypothetical transport mechanisms (Fig. 10 in Leyssens et al. 1992) have nowbeen investigated. Previous results have shown that, at the apical side, K+ extrusion intothe lumen is realized via the combination of an electrogenic H+ pump and anelectroneutral K+/H+ antiporter (see Weltens et al. 1992; Zhang et al. 1994; Leyssenset al. 1993a). The present study shows that the pathways for basal K+ entry seem to beconductive channels and secondary active cotransporters and that their relativeimportance varies depending on the bath K+ (and Na+) concentration. Primary active K+
uptake systems, i.e. a Na+/K+- or K+/H+-ATPase, if present, do not seem to contributesignificantly to transepithelial transport or to the maintenance of intracellular K+ (andNa+) levels. Fig. 5 shows the proposed transport model for Malpighian tubules ofFormica polyctena.
In high-K+, Na+-free conditions, a coupled entry of K+ and Cl2 seems to be of littleimportance. K+ uptake probably occurs via basal K+ channels, driven by a favourableelectrochemical gradient (Fig. 5C) (see also Leyssens et al. 1993a). In morephysiological conditions, i.e. 51 mmol l21 [K+]bl, when the basal electrochemical K+
gradient is smaller (see Leyssens et al. 1993a), K+ entry via channels may become lessimportant and a K+/Cl2 cotransport system probably accounts for a large portion of K+
uptake (Fig. 5B). In 5 or 10 mmol l21 [K+]bl (and high-Na+ conditions), little K+ isavailable for transepithelial transport; the basal electrochemical K+ gradient is evensmaller than in 51 mmol l21 [K+]bl and possibly outwardly directed (see Leyssens et al.1993a). The fluid secretion rate is low but still present. In this condition, a Na+/K+/2Cl2
cotransporter may take over a large part of the basal K+ uptake (Fig. 5A).Furthermore, the different basal transport systems for K+ entry may function at
different rates depending on the basal K+ (and Na+) concentration and may therebydetermine the amount of K+ available for transport across the apical membrane via theK+/H+ antiporter. The rise in the intracellular [K+] in response to an increased basal K+
concentration could occur either via K+ channels or via a coupled K+/Cl2 entrymechanism.
Questions remaining to be answered
In a low-K+, high-Na+ solution, the intracellular K+ content is markedly reduced (seeLeyssens et al. 1993a). Consequently, intracellular K+ has to be replaced by anothercation, probably Na+. A Na+ conductance could not be detected (see Leyssens et al.1992), but the uptake of Na+ might occur via a Na+/K+/2Cl2 cotransporter. The luminalK+ concentration also decreased when [K+]bl was lowered, but not to the same extent as
141Effects of ions and inhibitors on Malpighian tubules

142 A. LEYSSENS AND OTHERS
–
Vanadate (10−3 mol l−1)
K+
BathCellLumen
+
H+
H+
K+ transport
+ –[K+]i[K+]l
Ouabain (10−3 mol l−1)no effect
Bumetanide(10−5 mol l−1)
Br−
Ba2+ (6 mmol l−1)K+
K+Cl−Cl−Na+
–
–
–
–Na+-free
Sch28080 (10−4 mol l−1)no effect
A
B
C
+
K+
Ba2+ (6 mmol l−1)
K+
–
H+
H+
K+ transport
+ –
[K+]i
K+
Cl−[K+]l
–
+
–
Na+-freeno effect
Bumetanide(10−4 mol l−1)
Br−
–Vanadate (10−3 mol l−1)
Sch28080 (10−4 mol l−1)no effect
Ba2+ (6 mmol l−1)
K+H+
H+
K+ transport
+ –
[K+]i
K+
[K+]l
–
Bumetanide(10−4 mol l−1)
no effect
–
5 or 10 mmol l−1 [K+]bl
51 mmol l−1 [K+]bl
113 mmol l−1 [K+]bl
Fig. 5. Model for transcellular K+ transport in 5 or 10 (A), 51 (B) and 113 mmol l21 (C) [K+]bl,summarizing the results in the present study. It has previously been shown (1) that K+
transport increases (larger arrows) at a higher [K+]bl (Van Kerkhove et al. 1989); (2) that theapical and basal membrane potential differences decrease (smaller + and 2 signs in the figure)when [K+]bl increases (Leyssens et al. 1992); (3) that [K+]l and [K+]i increase (indicated bylarger symbols in the figure) when [K+]bl increases (Leyssens et al. 1993a); and (4) that an H+
pump is present in the apical membrane in parallel with a K+/H+ antiporter (Weltens et al.1992; Zhang et al. 1994; Leyssens et al. 1993a,b).

[K+]i (see Leyssens et al. 1993a). This implies that K+ is still the major cation secreted.Consequently, although the apical K+/H+ antiporter may have some affinity for Na+ whenintracellular Na+ levels rise, secretion of K+ still predominates. This means that net basalK+ uptake must exceed net Na+ uptake. Additional uptake via a K+/Cl2 cotransporterseems unlikely; it would need a very low intracellular Cl2 concentration. Preliminaryresults show that this is not the case (unpublished observations). As no evidence wasfound for a Na+/K+-ATPase, we have no indication of how intracellular [Na+] is regulatedin our preparation. Theoretically, in view of the high intracellular pH found in Malpighiantubules of Formica polyctena (Zhang et al. 1994), a Na+/H+ antiporter in the basalmembrane, working in reverse mode and unexpectedly functioning at an alkaline pH,might extrude Na+ in exchange for H+.
The transepithelial Cl2 pathway awaits further investigation. In high [K+]bl
(113 mmol l21), the electroneutral K+/Cl2 cotransporter did not seem to be functional andno relative Cl2 conductance was found. This suggests that Cl2 transport is primarilyparacellular. The lack of a relative Cl2 conductance in the basal membrane (Leyssenset al. 1992) points to a paracellular pathway for Cl2 down an electrical gradient in113 mmol l21 [K+]bl. In 51 and 5 mmol l21 [K+]bl, K+/Cl2 and Na+/K+/2Cl2
cotransporters, respectively, seem to be functional and at least part of the Cl2 transportmay occur transcellularly; hence, Cl2 channels would be required to ensure Cl2 exitacross the apical membrane. In a few experiments in 5 mmol l21 [K+]bl, a smallhyperpolarization of Vap was observed when bumetanide was applied (Fig. 4D) (cf.Rhodnius prolixus, O’Donnell and Maddrell, 1984; Locusta migratoria, Baldrick et al.1988). If a Cl2 conductance were present in the apical membrane, this might be explainedby a drop in intracellular Cl2 concentration due to inhibition of basal Cl2 influx. Theexistence of a transcellular Cl2 pathway was also suggested by a study on symmetricallyperfused Formica polyctena tubules (Dijkstra et al. 1994). Cl2 channels in the apicalmembrane have been detected by a patch-clamp study in Aedes aegypti (Wright andBeyenbach, 1987) and in Drosophila melanogaster tubules (J. Dow and B. Harvey,personal communication). Apical Cl2 channels (as in rectal gland; Greger et al. 1989)and/or bumetanide-sensitive basal cotransporters (as in Aedes tubules, Hegarty et al.1991) could be a site of regulation by hormones and second messengers to accelerate fluidsecretion rate even at lower bath K+ concentrations.
Preliminary results obtained from intracellular and luminal ion-selective measurementsin different bath K+ concentrations have demonstrated that the calculated gradients forCl2 and K+ are consistent with the transport mechanisms for K+ and Cl2 proposed in themodel.
The authors wish to thank Mr P. Pirotte for making the electrodes and performing partof the fluid secretion measurements, Mrs J. Vanderhallen for preparing the solutions, MrR. Van Werde for help with the electronics, Mr W. Leyssens for administrative tasks,Ms K. Ungricht for typing the tables and Mr and Mrs Withofs for artwork. This workwas supported by a grant from NFWO (Nationaal Fonds voor WetenschappelijkOnderzoek, Belgium) and by a grant from the EC (European Community: SC1-CT90-0480).
143Effects of ions and inhibitors on Malpighian tubules

ReferencesANSTEE, J. H. AND BOWLER, K. (1979). Ouabain-sensitivity of insect epithelial tissues. Comp. Biochem.
Physiol. 62A, 763–769.BAKER, P. F. AND WILLIS, J. S. (1970). Potassium ions and the binding of cardiac glycosides in
mammalian cells. Nature 226, 521–523.BALDRICK, P., HYDE, D. AND ANSTEE, J. H. (1988). Microelectrode studies on Malpighian tubule cells of
Locusta migratoria: effects of external ions and inhibitors. J. Insect Physiol. 34, 963–975.BELTRAN, C. AND NELSON, N. (1992). The membrane of vacuolar H+-ATPases by itself is impermeable
to protons. Acta physiol. scand. 146, 41–47.BERTRAM, G., SCHLEITHOFF, L., ZIMMERMANN, P. AND WESSING, A. (1991). Bafilomycin A1 is a potent
inhibitor of urine formation by Malpighian tubules of Drosophila hydei: is a vacuolar-type ATPaseinvolved in ion and fluid secretion? J. Insect Physiol. 37, 201–209.
CHASTEEN, N. D. (1983). The biochemistry of vanadium. Structure Bonding 53, 105–138.CHATTERJEE, D., CHAKRABORTY, M., LEIT, M., NEFF, L., JAMSAKELLOKUMPU, S., FUCHS, R.,
BARTKIEWICZ, M., HERNANDO, N. AND BARON, R. (1992). The osteoclast proton pump differs in itspharmacology and catalytic subunits from other vacuolar H+-ATPases. J. exp. Biol. 172, 193–204.
DIJKSTRA, S. (1993). Cl2-transport and H+-pumping in the primary urine formation in the ant. PhDthesis. Limburgs Universitair Centrum.
DIJKSTRA, S., LOHRMANN, E., STEELS, P. AND GREGER, R. (1994). Electrical properties of the isolated, invitro perfused Malpighian tubule of the ant, the Cl-pathway. Cell. Physiol. Biochem. 4, 19–30.
ELLORY, J. C. AND HALL, A. C. (1988). Human red cell volume regulation in hypotonic media. Comp.Biochem. Physiol. 90A, 533–537.
EVANS, M. G., MARTY, A., TAN, Y. P. AND TRAUTWEIN, A. (1986). Blockage of Ca-activated Clconductance by furosemide in rat lacrimal glands. Pflügers Arch. 406, 65–68.
FORGAC, M. (1989). Structure and function of vacuolar class of ATP-driven proton pumps. Physiol. Rev.69, 765–796.
FROISSART, M., BORENSZTEIN, P., HOUILLIER, P., LEVIEL, F., POGGIOLI, J., MARTY, E., BICHARA, M. AND
PAILLARD, M. (1992). Plasma membrane Na+–H+ antiporter and H+-ATPase in the medullary thickascending limb of rat kidney. Am. J. Physiol. 262, C963–C970.
GREGER, R., SCHLATTER, E. AND GÖGELEIN, E. (1989). Cl2 channels in the apical cell membrane of therectal gland ‘induced’ by cyclic AMP. Pflügers Arch. 406, 446–448.
HAAS, M. AND FORBUSH III, B. (1986). 3H-bumetanide binding to duck red cells: correlation withinhibition of (Na+K+2Cl) co-transport. J. biol. Chem. 261, 8834–8441.
HEDGE, R. S. AND PALFREY, H. C. (1992). Ionic effects on bumetanide binding to the activated Na/K/2Clcotransporter: selectivity and kinetic properties of ion bindings sites. J. Membr. Biol. 126, 27–37.
HEGARTY, J. L., ZHANG, B., PANNABECKER, T. L., PETZEL, P. H., BAUSTIAN, M. D. AND BEYENBACH,K. W. (1991). Dibutyryl cyclic AMP activates bumetanide-sensitive electrolyte transport inMalpighian tubules. Am. J. Physiol. 261, C521–C529.
LEYSSENS, A., STEELS, P., LOHRMANN, E., WELTENS, R. AND VAN KERKHOVE, E. (1992). Intrinsicregulation of K+ transport in Malpighian tubules (Formica): electrophysiological evidence. J. InsectPhysiol. 38, 431–446.
LEYSSENS, A., VAN KERKHOVE, E., ZHANG, S.-L., WELTENS, R. AND STEELS, P. (1993a). Measurement ofintracellular and luminal K+ concentrations in a Malpighian tubule (Formica): Estimate of basal andluminal electrochemical K+ gradients. J. Insect Physiol. 39, 945–958.
LEYSSENS, A., ZHANG, S.-L., VAN KERKHOVE, E. AND STEELS, P. (1993b). Both dinitrophenol and Ba2+
reduce KCl and fluid secretion in Malpighian tubules of Formica: the role of the apical H+ and K+
concentration gradient. J. Insect Physiol. 39, 1061–1073.MADDRELL, S. H. P. (1969). Secretion by the Malpighian tubules of Rhodnius. The movement of ions
and water. J. exp. Biol. 51, 71–97.NECHAY, B. R., NANNINGA, L. B., NECHAY, P. S. E., POST, R. L., GRANTHAM, J. J., MACARA, I. G.,
KUBENA, L. F., PHILLIPS, T. D. AND NIELSEN, F. H. (1986). Role of vanadium in biology. Fedn Proc.Fedn Am. Socs exp. Biol. 45, 123–132.
NICOLSON, S. W. (1993). The ionic basis of fluid secretion in insect Malpighian tubules: advances in thelast ten years. J. Insect Physiol. 39, 451–458.
NICOLSON, S. W. AND ISAACSON, L. C. (1987). Transepithelial and intracellular potentials in isolatedMalpighian tubules of tenebrionid beetle. Am. J. Physiol. 252, F645–F653.
144 A. LEYSSENS AND OTHERS

O’DONNELL, M. J. AND MADDRELL, S. H. P. (1984). Secretion by the Malpighian tubules of Rhodniusprolixus Stål: electrical events. J. exp. Biol. 110, 275–290.
PALFREY, H. C. AND GREENGARD, P. (1981). Hormone-sensitive ion transport systems in erythrocytes asmodels for epithelial ion pahtways. Ann. N.Y. Acad. Sci. 372, 291–308.
PALFREY, H. C. AND O’DONNELL, M. E. (1992). Characteristics and regulation of the Na/K/2Clcotransporter. Cell. Physiol. Biochem. 2, 293–307.
PHILLIPS, J. (1981). Comparative physiology of insect renal function. Am. J. Physiol. 241, R241–R257.SCOTT, C. K., SUNDELL, E. AND CASTROVILLY, L. (1987). Studies on the mechanism of action of the
gastric microsomal (H++K+)-ATPase inhibitors Sch 32651 and Sch 28080. Biochem. Pharmac. 36,97–104.
VAN KERKHOVE, E., WELTENS, R., ROINEL, N. AND DE DECKER, N. (1989). Haemolymph composition inFormica (Hymenoptera) and urine formation by the short isolated Malpighian tubules:electrochemical gradients for ion transport. J. Insect Physiol. 35, 991–1003.
VAUGHAN, G. L. AND JUNGREIS, A. M. (1977). Insensitivity of lepidopteran tissues to ouabain:physiological mechanisms for protection from cardiac glycosides. J. Insect Physiol. 24, 585–589.
VERHULST, H., VAN KERKHOVE, E. AND DE DECKER, N. (1988). KCl secretion by Malpighian tubules ofFormica: evidence for the presence of a KCl cotransport system. Archs int. Physiol. Biochem. 96,P8–P9.
WELTENS, R., LEYSSENS, A., ZHANG, S.-L., LOHRMANN, E., STEELS, P. AND VAN KERKHOVE, E. (1992).Unmasking of the apical electrogenic H pump in isolated Malpighian tubules (Formica polyctena) bythe use of barium. Cell. Physiol. Biochem. 2, 101–116.
WESSING A., HEVERT F. AND RÖNNAU K. (1987). Ion transport and intracellular activity of ions inMalpighian tubules of Drosophila hydei. Zool. Beitr. N.F. 30, 297–314.
WIECZOREK, H., WEERTH, S., SCHLINDBECK, M. AND KLEIN, U. (1989). A vacuolar-type proton pump in avesicle fraction enriched with potassium transporting plasma membranes from tobacco hornwormmidgut. J. biol. Chem. 264, 11143–11148.
WIECZOREK, H., WOLFERSBERGER, M. G., CIOFFI, M. AND HARVEY, W. R. (1986). Cation stimulatedATPase activity in purified plasma membranes from tobacco hornworm midgut. Biochim. biophys.Acta 857, 271–281.
WRIGHT, J. M. AND BEYENBACH, K. W. (1987). Chloride channels in apical membranes of mosquitoMalpighian tubules. Fedn Proc. Fedn Am. Socs exp. Biol. 46, 1270.
ZHANG, S.-L., LEYSSENS, A., VAN KERKHOVE, E., WELTENS, R., VAN DRIESSCHE, W. AND STEELS, P.(1994). Electrophysiological evidence for the presence of an apical H+ ATPase in Malpighian tubulesof Formica polyctena: intracellular and luminal pH measurements. Pflügers Arch. 426, 288–295.
145Effects of ions and inhibitors on Malpighian tubules




















