
Differential regulation of B/K protein expression in proximal and distal tubules of rat kidneys with...
27
Han et al. F-00009-2006.R1 (Final Accepted Version) 1 Differential regulation of B/K protein expression in proximal and distal tubules of rat kidneys with ischemia-reperfusion injury Ki-Hwan Han 1 , U-Young Lee 2,5 , Yoon-Seong Jang 3,5 , Yoon Mi Cho 3,5 , Young Min Jang 3,5 In-A Hwang 1 , Jung Yeon Ghee 4,5 , Sun-Woo Lim 4,5 , Wan-Young Kim 2,5 , Chul Woo Yang 4,5 , Jin Kim 2,5 , and Oh-Joo Kwon 3,5 1 Department of Anatomy, College of Medicine, Ewha Womans University, Seoul, Korea; 2 Departments of Anatomy, 3 Biochemistry and 4 Internal Medicine, and 5 MRC for Cell Death Disease Research Center, College of Medicine, The Catholic University of Korea, Seoul, Korea Running Head: B/K protein in the kidney Address Correspondence to: Jin Kim, M.D., Ph.D., Department of Anatomy, College of Medicine, The Catholic University of Korea, 505 Banpo-dong, Seocho-gu, Seoul 137-701, Korea, Phone: 82-2- 590-1153, Fax: 82-2-536-3110, e-mail: [email protected] Address Co-correspondence to: Oh-Joo Kwon, M.D., Ph.D., Department of Biochemistry, College of Medicine, The Catholic University of Korea, 505 Banpo-dong, Seocho-gu, Seoul 137-701, Korea, Phone: 82-2-590-1179, FAX: 82-2-596-4435, e-mail: [email protected] Page 1 of 27 Articles in PresS. Am J Physiol Renal Physiol (August 8, 2006). doi:10.1152/ajprenal.00009.2006 Copyright © 2006 by the American Physiological Society.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential regulation of B/K protein expression in proximal and distal tubules of rat kidneys with...

Han et al. F-00009-2006.R1 (Final Accepted Version)
1
Differential regulation of B/K protein expression in proximal and distal tubules of
rat kidneys with ischemia-reperfusion injury
Ki-Hwan Han1, U-Young Lee2,5, Yoon-Seong Jang3,5, Yoon Mi Cho3,5, Young Min
Jang3,5 In-A Hwang1, Jung Yeon Ghee4,5, Sun-Woo Lim4,5, Wan-Young Kim2,5, Chul
Woo Yang4,5, Jin Kim2,5, and Oh-Joo Kwon3,5
1Department of Anatomy, College of Medicine, Ewha Womans University, Seoul,
Korea; 2Departments of Anatomy, 3Biochemistry and 4Internal Medicine, and 5MRC for
Cell Death Disease Research Center, College of Medicine, The Catholic University of
Korea, Seoul, Korea
Running Head: B/K protein in the kidney
Address Correspondence to:
Jin Kim, M.D., Ph.D., Department of Anatomy, College of Medicine, The Catholic
University of Korea, 505 Banpo-dong, Seocho-gu, Seoul 137-701, Korea, Phone: 82-2-
590-1153, Fax: 82-2-536-3110, e-mail: [email protected]
Address Co-correspondence to:
Oh-Joo Kwon, M.D., Ph.D., Department of Biochemistry, College of Medicine, The
Catholic University of Korea, 505 Banpo-dong, Seocho-gu, Seoul 137-701, Korea,
Phone: 82-2-590-1179, FAX: 82-2-596-4435, e-mail: [email protected]
Page 1 of 27Articles in PresS. Am J Physiol Renal Physiol (August 8, 2006). doi:10.1152/ajprenal.00009.2006
Copyright © 2006 by the American Physiological Society.

Han et al. F-00009-2006.R1 (Final Accepted Version)
2
ABSTRACT
Brain/kidney (B/K) protein is a novel double C2-like-domain protein that is highly
expressed in rat brain and kidney, but its cellular localization and functional role in the
kidney are still undetermined. We examined the cellular localization of B/K protein in
the rat kidney under normal and ischemic conditions. Ischemia-reperfusion (IR) injury
was induced by clamping both renal arteries for 45 minutes, and animals were
sacrificed at 1 and 6 hours and 1, 2, 3, 5, 7, 14, and 28 days after the reperfusion.
Kidney tissues were processed for immunohistochemistry and immunoblot analyses
using rabbit anti-B/K polyclonal antibodies. In control kidneys, B/K protein was
expressed primarily in the distal tubules including the thick ascending limb, distal
convoluted and connecting tubules, and collecting duct. Notably, B/K protein was also
expressed in the straight portion (S3 segment), but not in the S1 or S2, of the proximal
tubules, and podocytes of the glomerulus. In rat kidneys with IR injury, expression of
B/K protein was differentially regulated according to anatomical location. In the distal
tubules, overall expression of B/K protein was markedly decreased. On the other hand,
IR injury significantly increased B/K protein expression in the S3 segment of the outer
medulla as well as in rat proximal tubular epithelial cell line NRK-52E in vitro.
Furthermore, B/K protein was strongly expressed in many exfoliated cells in the lumen
and urine. These findings suggest that B/K protein is closely associated with cell death
of the proximal tubules which are vulnerable to IR injury in kidney.
Key words: brain/kidney (B/K) protein, ischemia-reperfusion (IR) injury, cell death
Page 2 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
3
INTRODUCTION
Acute renal failure represents a significant and persistent problem in clinical medicine
with serious consequences. Renal ischemia-reperfusion (IR) injury is a major cause of
acute renal failure in the native kidney and an invariable occurrence in the transplanted
kidney. Understanding the molecular mechanisms underlying renal cell injury is critical
for the prevention or treatment of acute renal failure, and considerable attention and
efforts have been directed towards identification of the therapeutic targets or
biomarkers involved in the tubular cell damage following IR injury. Global changes in
renal gene expression by the ischemic renal injury have been recently reviewed (7).
However, the intracellular and molecular mechanisms leading to acute renal failure by
IR injury remain incompletely understood.
We previously isolated a novel protein with two C2-like domains from the rat brain and
named it “B/K” based on its predominant expression in the brain and kidney (18). B/K is
a 474 amino acid protein and has several unique features. Although B/K protein does
not contain a transmembrane domain, it can bind to the plasma membrane via the N-
terminal cysteine cluster (9). Three negatively charged amino acids in the C2a domains
that have been suggested to be necessary for calcium binding (28) are substituted.
Moreover, B/K has three consensus sequences for protein kinase A (PKA). We
recently reported the distribution pattern of B/K protein. In the brain, B/K protein is
prominently expressed in neurosecretory areas such as the hypothalamic neurons, the
circumventricular organs, and some endocrine cells of the adenohypophysis (20). B/K
protein is expressed in most ganglion cells, a few amacrine cells, and the retinal fibers
of Müller cells in the rat retina (19). Interestingly, expression of B/K protein was induced
in the vulnerable regions of the hippocampus by kainate-induced excitotoxic injury (13)
as well as in the retina by IR injury (14). These findings suggest the possibility that B/K
protein may play a role in the mechanism of cell death in some pathological conditions
Page 3 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
4
including IR injury.
Besides the brain, B/K protein is most abundantly expressed in the kidney. Recently,
we reported the vasopressin-induced and PKA-dependent phosphorylation of B/K
protein in LLC-PK1, a porcine renal proximal tubule cell line (3). However, in spite of its
very high level of expression, its candidature as a substrate for PKA, and its possible
involvement in vasopressin-mediated signaling, the role of B/K protein in the kidney
has remained unknown.
In the present study, we investigated the expression and the cellular localization of B/K
protein in the rat kidney under normal and IR injury conditions.
Page 4 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
5
MATERIALS AND METHODS
Animals
Male Sprague Dawley rats (250-300 g, n = 6 per each group) were housed at 21 , 12℃ -
hour light/dark cycle, and allowed free access to food and water. Animals were
anesthetized with an intraperitoneal injection of sodium pentobarbital (50 mg/kg body
weight). After opening the abdomen through a midline incision, both renal pedicles
were exposed and cleaned by blunt dissection. Microvascular clamps were placed on
both renal arteries to completely block renal blood flow, core body temperature was
maintained by placing the animals on a homeothermic table (29). After 45 minutes, the
clamps were removed and blood flow returned to the kidneys. Animals were sacrificed
at 1, and 6 hours and 1, 2, 3, 5, 7, 14, and 28 days after reperfusion. Control animals
received sham treatment. The sham operation was performed in a similar manner,
except for the clamping of the renal vessels.
Tissue Preservation
The kidneys were first perfused briefly through the abdominal aorta with phosphate-
buffered saline (PBS) to rinse out all blood and subsequently perfused with the fixative
solution, periodate-lysine-2% paraformaldehyde (PLP), for 5 min. The kidneys were
removed and cut transversely into 1- to 2-mm thick slices that were immersed in PLP
overnight at 4 . After being rinsed in PBS, sections of tissue were cut transversely ℃
through the entire kidney on a vibratome (Lancer Vibratome Series 1000; Technical
Products International, St. Louis, MO) at a thickness of 50 µm and processed for
immunohistochemical studies using a horseradish peroxidase preembedding technique.
Immunohistochemistry
Fifty-micrometer vibratome sections were processed for immunohistochemistry using
Page 5 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
6
an indirect preembedding immunoperoxidase method, as previously described (10, 11).
All sections were washed with 50 mM NH4Cl in PBS three times for 15 min. Before
incubation with the primary antibody, the all tissue sections were incubated for 3 hours
with PBS containing 1% BSA, 0.05% saponin, and 0.2% gelatin (solution A). The tissue
sections were then incubated overnight at 4 with the antibody against B/K protein℃
(20) diluted 1:1,000 in 1% BSA-PBS (solution B). Control incubations were performed
in solution B without primary antibody. After several washes with solution A, the tissue
sections were incubated for 2 hours in peroxidase-conjugated goat anti-rabbit IgG Fab
fragment (Jackson ImmunoResearch Laboratories, West Grove, PA), diluted 1:50 in
solution B. The tissues were then rinsed, first in solution A and subsequently in 0.05 M
Tris buffer, pH 7.6. For the detection of horseradish peroxidase, the sections were
incubated in 0.1% 3,3’-diaminobenzidine in 0.05 M Tris buffer for 5 min, after which
H2O2 was added to a final concentration of 0.01% and the incubation was continued for
10 min. After washing with 0.05 M Tris buffer three times, the sections were dehydrated
in a graded series of ethanol and embedded in Epon-812. From all animals, 50-µm-
thick vibratome sections through the entire kidney were mounted in Epon-812 between
polyethylene vinyl sheets. Sections were examined and photographed on an Olympus
photomicroscope equipped with differential-interference contrast (DIC) optics.
In Vitro IR Injury
A normal rat proximal tubular epithelial cell line (NRK-52E) was obtained from
American Type Culture Collection (Manassas, VA), and maintained in DMEM
supplemented with 5% fetal bovine serum (FBS) containing 0.2% gentamycin at 37°C
in a humidified atmosphere of 5% CO2 / 95% air. In vitro ischemia-reperfusion injury
was induced as previously described (30) with some modifications. In brief, when the
density of the cells (5 × 105 cells in 100 mm culture dishes) reached to around 70%
Page 6 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
7
confluency, the culture media was replaced with low serum media (DMEM/0.5% FBS)
and further incubated for 1 day. Then, the dishes were transferred to the anaerobic
chamber (ThermoForma model 1025, Marietta, OH) which was saturated with 85%
N2/10% H2/5% CO2, and incubated for 6 hours at 37°C. At the end of the ischemic
incubation, the dishes were removed from the anaerobic chamber and the culture
media was replaced with 10 ml complete media, and further incubated in a humidified
incubator with 5% CO2 at 37°C for additional 0, 12 and 24 hours.
For the cytotoxicity analysis, the cells were harvested at the indicated time and treated
with reagents containing Alexa Fluor 488-labeled annexin Ⅴ and propidium iodide (PI)
(Vybrant Apoptosis Assay Kit, Molecualr Probes, Eugene, OR) for 15 min at room
temperature. The apoptotic cells were analyzed by fluorescence-activated cell sorting
analysis (FACSCalibur; Becton Dikinson, Franklin Lakes, NJ). The total apoptotic cells
positive to both PI and annexin V were counted.
Immunoblot Analysis
For immunoblot analysis of in vivo experiments, the kidneys were isolated from rats
with IR injury at 1, and 6 hours and 1, 3, 5, 7, 14, and 28 days after the reperfusion and
perfused with PBS. The kidneys from sham operated rat were used as a control. The
cortex and medulla of the kidney were dissected carefully and homogenized in 10
volumes of 20 mM Tris-HCl (pH 7.4) containing 1% Triton X-100, 150 mM NaCl, 0.5%
sodium deoxycholate, 0.1% SDS, 1 mM EDTA, 0.02% sodium azide, 1 µM leupeptin,
and 1 mM phenylmethylsulfonylfluoride (Sigma, St. Louis, MO). After centrifugation
(12,000 xg, 10 min, 4℃), the supernatant was isolated and the protein concentration
was determined using Bradford method (Pierce, Rockford, IL). Protein concentrations
were measured by bicinchonic acid assay kit (Bio-Rad, Hercules, CA), and 25 µg of
each protein was separated by 7.5% SDS-PAGE and transferred onto a nitrocellulose
Page 7 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
8
membrane (Schleicher & Schell, Keene, NH). The membrane were blocked with 5%
skim milk for 1 hour, and incubated with the rabbit anti-B/K polyclonal antiserum
(1:5,000 dilution) overnight at 4 . Antibo℃ dy binding was detected using horseradish
peroxidase-conjugated goat anti-rabbit IgG (1:1,000 dilution) (Sigma) and the
immunoreactive bands were visualized with ECL method (Amersham Pharmacia
Biotech, Upsala, Sweden). For in vitro IR injury samples, the cell lysates from treated
or untreated cells were resuspended in RIPA cell lysis buffer (25 mM Tris-HCl, pH 7.5,
0.1% SDS, 0.1% Triton X-100, 1% sodium deoxycholate, 150 mM NaCl, 1 mM EDTA,
10% glycerol, 1 mM orthovanadate, 5 mM NaF, 1 mM phenylmethanesulfonyl fluoride,
and proteinase inhibitor). After incubation on ice for 5 min, the homogenate was
centrifuged at 13,200 xg for 30 min at 4°C, and 4 µg of total protein of the clarified
supernatants was separated by 12% SDS-PAGE.
Urine Cytology
Fresh urine samples from control or IR injury model were obtained by bladder puncture
and centrifuged at 4℃ for 10 min at 1,500 xg and fixed in PLP. The resuspended
aliquots were cytocentrifuged (Cytospin 2, Shandon, Frankfurt, Germany) onto
microscope slides for 5 min at 500 xg, and processed for immunocytochemistry using a
commercially available kit (Dako LSAB2Kit, Dako Corporation, Carpinteria, CA). B/K
protein antibody was used at 1:500 dilution.
Page 8 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
9
RESULTS
In control animals, B/K protein was immunostained in the renal cortex and outer
medulla (Fig. 1a). In the cortex, it was mainly located in distal tubules including the
cortical thick ascending limb, distal convoluted tubule and cortical collecting duct.
Some interstitial cells and podocytes of the glomerulus also showed positive
immunostaining. However, B/K protein immunoreactivity was not observed in proximal
tubules (S1 and S2 segments) in the cortex (Fig. 1b).
In the outer medulla, B/K protein immunostaining was strong in the medullary thick
ascending limb and the outer medullary collecting duct (Figs. 1c-e). Interestingly, the
labeling intensity appeared more prominent in type A intercalated cells than in principal
cells in the outer medullary collecting duct and the initial portion of the inner medullary
collecting duct (Figs. 1d and f). Moderate immunoreactivity was also observed in the
cells of proximal straight tubules (S3 segment) in the outer stripe of the outer medulla
(Fig. 1c). B/K protein was not immunostained in vascular bundles (Fig. 1e).
After IR injury, there was a notable change of B/K protein immunostaining in both the
cortex and the outer medulla (Fig. 2). In the cortex, the overall intensity of
immunostaining was decreased in distal tubules including the cortical thick ascending
limb, distal convoluted tubule and the cortical collecting duct, and it reached the lowest
level at day 1 after IR injury (Figs. 2c and 3a). The moderately decreased
immunostaining was also observed in podocytes of the glomerulus (Fig. 3a). Although
B/K protein immunoreactivity was not observed in proximal tubules in the cortex, some
damaged cells of the S2 segments showed B/K protein immunolabeling (Fig. 3a).
In the outer medulla, B/K protein immunostaining was rapidly decreased in the
medullary thick ascending limb and outer medullary collecting duct at 6 hours after IR
injury, which was easily recognizable in the inner stripe of the outer medulla (Fig. 2b).
The decreased immunoreactivity in the distal nephron segments began to be restored
Page 9 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
10
between 5 and 7 days and recovered fully at 14 days after the reperfusion (Fig. 2). On
the other hand, there was a marked increase of B/K protein expression in proximal
tubules (S3 segment) located in the outer stripe of the outer medulla (Fig. 2c). In
injured proximal tubules, B/K protein immunostaining was strongest in the many
exfoliated cells in the lumen at day 1 after IR injury and gradually decreased until 5
days after IR injury (Figs. 3b-f). Although B/K protein immunoreactivity increased in
severely damaged cells, it was very weak in intact tubular epithelial cells of the S3
segment (Fig. 4). Results from immunoblot analysis showed the early reduction of B/K
protein expression at 6 hours after IR injury, and rapid recovery at day 1 in the medulla
(Fig. 5). However, there were no significant changes of B/K protein expression in the
cortex (not shown). IR injury-induced increase of B/K protein expression in proximal
tubules was confirmed by in vitro experiments. As shown in Fig. 6a, B/K protein
expression in a rat proximal tubular epithelial cell line NRK-52E was strongly induced
by the 6 hour-ischemia, and further increased time-dependently by the reperfusion. In
the same experimental condition, IR strongly induced the apoptosis of NRK-52E cells
(Fig. 6b). Fraction of total apoptotic cells which were reactive to both propidium iodide
and annexin V were increased three-fold by 6 hour-ischemia (3.41%), and were
significantly and time-dependently increased by reperfusion (12 hour reperfusion,
17.54%; 24 hour reperfusion, 30.52%).
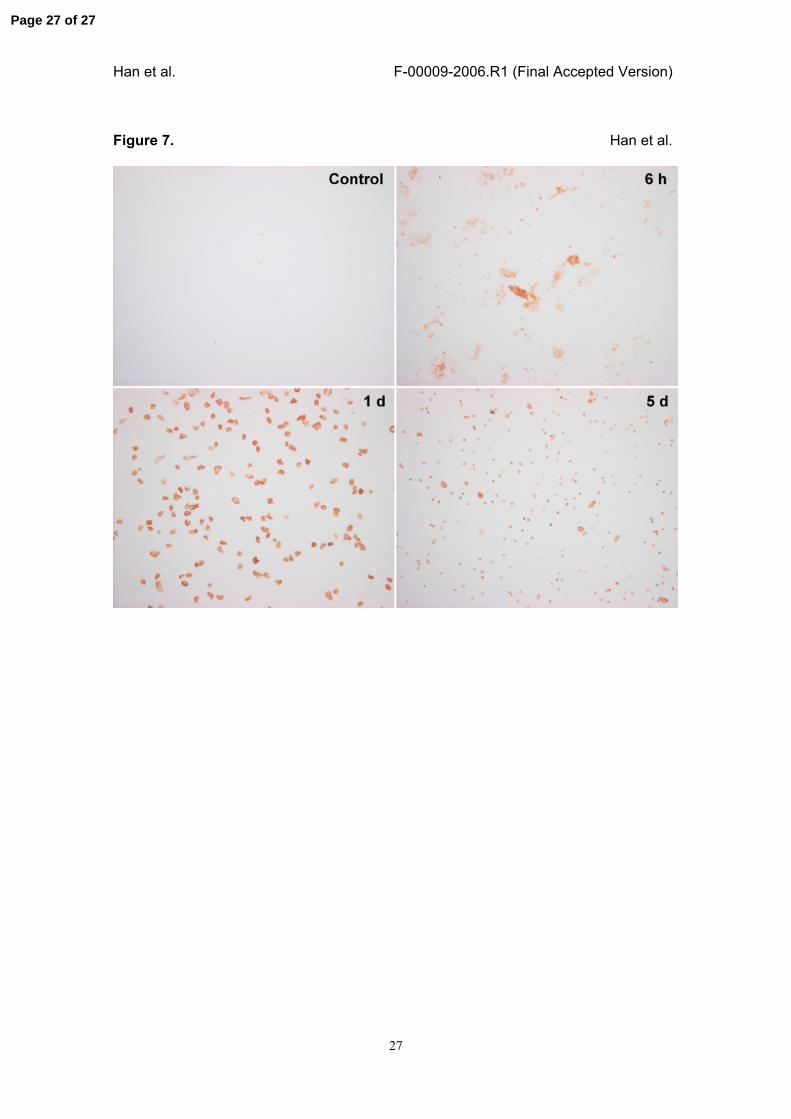
Cells immunopositive to B/K protein was also detected in the urine after IR injury. The
immunocytological investigations showed the strongest B/K protein immunostaining in
many urinary renal cells at day 1 and a gradual decrease until 5 days after IR injury.
However, few or no positive cells were found in the urine from control animal (Fig. 7).
Page 10 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
11
DISCUSSION
Renal ischemia-reperfusion (IR) injury is the major cause of acute renal failure, and
attempts at unraveling the molecular basis of cell damages induced by IR injury have
been facilitated by recent advances in functional genomics. Expression of many genes
has been found to be changed by IR injury (21, 22, 26), and recently, novel biomarkers
for early renal IR injury were identified by cDNA microarray analysis (7). In this study,
we examined the cellular localization of B/K protein in the rat kidney under normal
conditions and observed striking changes after IR injury, and suggest B/K protein as a
candidate marker for renal cell death.
In normal kidney, B/K protein was localized primarily in distal tubules including the thick
ascending limb, distal convoluted tubule and collecting duct. Moderate immunostaining
was also observed in the glomerulus, interstitium, and proximal straight tubule (S3
segment), suggesting a possible involvement of B/K protein in the cAMP-mediated
signaling mechanism. In the kidney, isoforms of adenylyl cyclase (AC) are expressed in
many nephron segments. To date, at least five AC isoforms (AC6, AC5, AC4, AC7, and
AC9) have been demonstrated in the rat kidney (2, 6, 12). Among them, AC4 was
expressed exclusively in the glomerulus, and both AC5 and AC6 were abundant in
some cells probably podocytes of glomeruli, and in the interstitium. AC6 was highly
expressed in the thick ascending limbs and weakly in the S3 segment, while AC5
mRNA was not detected in proximal tubules or the thick ascending limb. In collecting
ducts, AC5 was expressed only in intercalated cells, and AC6 was observed in both
intercalated and principal cells. Although we did not examine the expression of AC and
B/K proteins by coimmunostaining, it is possible that B/K protein is colocalized with
specific AC isoforms, especially AC5 and AC6, in the kidney.
cAMP produced by AC activates protein kinase A (PKA), and vasopressin is one of the
most important ligands for cAMP-PKA-mediated signaling mechanisms in the kidney.
Page 11 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
12
Vasopressin upregulates the expression of apical Na+-K+-2Cl−-cotransporter (BSC-1) in
the thick ascending limb (15, 16), and it increases water reabsorption in collecting duct
principal cells through a redistribution of the water channel aquaporin 2 by a
mechanism involving cAMP increase and PKA activation (23, 24). Apical K+ secretion
by ROMK in the thick ascending limb and the collecting duct segments also occurred
via a vasopressin-stimulated PKA pathway (8, 16). In our data, a significantly high level
of B/K protein expression was observed in the thick ascending limb, collecting duct and
distal convoluted tubules where vasopressin-stimulated cAMP-PKA pathways are well
established. Moreover, complete conservation of the consensus sequence for PKA
phosphorylation in B/K proteins among rat, mouse, and human genes, and
vasopressin- and PKA-dependent phosphorylation of B/K protein in a porcine kidney
cell line LLC-PK1 (3) strongly suggest the involvement of B/K protein in the
vasopressin-mediated cAMP-PKA pathways.
The S3 segment of the proximal tubule is a major target of hypoxic injury (21, 27). In
our experiments, IR injury induced B/K protein expression in many exfoliating cells but
not in intact epithelial cells in the S3 segment. Moreover, cells strongly positive to B/K
protein were detected in urine after IR injury. In addition, results from in vitro studies
showed that IR injury significantly induced B/K protein expression in a rat proximal
tubular cell line (NRK-52E), and also resulted in the apoptosis of the cells in the
manner nearly proportional to the level of B/K protein in the cells. These findings
strongly support our hypothesis that B/K protein may play a role in the cellular damage
of this segment after IR injury. Consistent with this result, our previous findings showed
a high level of expression of B/K protein in the most vulnerable regions of the
hippocampus in the kainate seizure model (13).
Many cases of IR-induced cellular damage are explained by apoptosis for which the
mitochondria play a crucial role in inducing apoptosis. Recently, the endoplasmic
Page 12 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
13
reticulum (ER)-mediated apoptosis mechanism has been suggested (25). Prolonged or
more substantial stress to ER (ER stress) may lead to cell death via apoptosis, and
many reports has also suggested ER stress as mechanisms for pathogenesis of
various diseases (1). In the kidney, ER stress responses also have been characterized
in experimental models of glomerulonephritis (5) and IR injury (17). For example,
phosphorylation of PERK and eIF2, important mediators for ER stress response, has
been reported in IR-dependent glomerular epithelial cell injury (4). In relation to ER
stress response, we also reported the ER-specific localization and the ER stress-
induced expression of B/K protein (13). These findings indicate the possible
involvement of B/K protein in an ER-mediated cell death mechanism in the IR injury
model of rat kidney.
In summary, B/K protein is expressed mainly in distal tubules in the normal kidney, and
IR injury remarkably changed the expression of B/K protein in both proximal and distal
tubules. We suggest a possible role for B/K protein in renal tubular cells under
conditions of cellular adaptation or damage after IR injury. However, the functional
significance of B/K protein expression in relation to the cAMP-mediated signaling
pathway or ER stress response remains to be established.
ACKNOWLEDGEMENTS
This work was supported by Korea Science & Engineering Foundation (KOSEF)
through the MRC for Cell Death Disease Research Center at the Catholic University of
Korea (R13-2002-005-01001-0) to J Kim, CW Yang and OJ Kwon.
Page 13 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
14
REFERENCES
1. Breckenridge D. Regulation of apoptosis by endoplasmic reticulum pathway.
Oncogene 22: 8608-8618, 2003.
2. Chabardes D, Firsov D, Aarab L, Clabecq A, Bellanger AC, Siaume-Perez S, and
Elalouf JM. Localization of mRNAs encoding Ca2+-inhibitable adenylyl cyclases along
the renal tubule. Functional consequences for regulation of the cAMP content. J Biol
Chem 271: 19264-19271, 1996.
3. Chin H, Choi SH, Jang YS, Cho SM, Kim HS, Lee JH, Jeong SW, Kim IK, Kim GJ,
Kwon OJ. Protein kinase A-dependent phosphorylation of B/K protein. Exp Mol Med
38: 144-152, 2006.
4. Cybulsky AV, Takano T, Papillon J, and Bijian K. Role of the endoplasmic reticulum
unfolded protein response in glomerular epithelial cell injury. J Biol Chem 280: 24396-
24403, 2005.
5. Cybulsky AV, Takano T, Papillon J, Khadir A, Liu J, and Peng H. Complement C5b-9
membrane attack complex increases expression of endoplasmic reticulum stress
proteins in glomerular epithelial cells. J Biol Chem 277: 41342-41352, 2002.
6. Defer N, Best-Belpomme M, and Hanoune J. Tissue specificity and physiological
relevance of various isoforms of adenylyl cyclase. Am J Physiol 279: F400-F416, 2000.
7. Devarajan P, Mishra J, Supavekin S, Patterson LT, and Potter SS. Gene expression
in early ischemic renal injury: clues towards pathogenesis, biomarker discovery, and
novel therapeutics. Mol Genet Metabol 80: 365-376, 2003.
8. Ecelbarger CA, Kim GH, Knepper MA, Liu J, Tate M, Welling PA, and Wade JB.
Regulation of potassium channel Kir 1.1 (ROMK) abundance in the thick ascending
limb of Henle's loop. J Am Soc Nephrol 12: 10-18, 2001.
9. Fukuda M and Mikoshiba K. The N-terminal cysteine cluster is essential for
membrane targeting of B/K protein. Biochem J 360: 441-448, 2001.
Page 14 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
15
10. Han KH, Lim JM, Kim WY, Kim H, Madsen KM, Kim J. Expression of endothelial
nitric oxide synthase in developing rat kidney. Am J Physiol Renal Physiol 288: F694-
F702, 2005.
11. Han KH, Woo SK, Kim WY, Park SH, Cha JH, Kim J, Kwon HM. Maturation of
TonEBP expression in developing rat kidney. Am J Physiol Renal Physiol 287: F878-
F885, 2004.
12. Helies-Toussaint C, Aarab L, Gasc JM, Verbavatz JM, and Chabardes D. Cellular
localization of type 5 and type 6 ACs in collecting duct and regulation of cAMP
synthesis. Am J Physiol 279: F185-F194, 2000.
13. Jang YS, Lee MY, Choi SH, Kim MY, Chin H, Jeong SW, Kim IK, and Kwon OJ.
Expression of B/K protein in the hippocampus of kainate-induced rat seizure model.
Brain Res 999: 203-211, 2004.
14. Ju WK, Choi SH, Kwon JS, Kwon OJ, Lee MY, Oh SJ, Moon JL, and Chun MH.
Expression of brain/kidney protein in Muller cells of rat retina following transient
ischemia. Neurosci Lett 293: 53-56, 2000.
15. Kim GH, Ecelbarger CA, Mitchell C, Packer RK, Wade JB, and Knepper MA.
Vasopressin increases Na-K-2Cl cotransporter expression in thick ascending limb of
Henle's loop. Am J Physiol 276: F96-F103, 1999.
16. Knepper MA, Kim GH, Fernandez-Llama P, and Ecelbarger CA. Regulation of thick
ascending limb transport by vasopressin. J Am Soc Nephrol 10: 628-634, 1999.
17. Kuznetsov G, Bush KT, Zhang PL, and Kigam SK. Perturbations in maturation of
secretory proteins and their association with endoplasmic reticulum chaperones in a
cell culture model for epithelial ischemia. Proc Natl Acad Sci USA 93: 8584-8589, 1996.
18. Kwon OJ, Gainer H, Wray S, and Chin H. Identification of a novel protein containing
two C2 domains selectively expressed in the rat brain and kidney. FEBS Lett 378: 135-
139, 1996.
Page 15 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
16
19. Kwon OJ, Ju WK, Choi SH, Lee MY, Oh SJ, and Chun MH. The expression and
cellular localization of brain/kidney protein in the rat retina. Brain Res 860: 178-180,
2000.
20. Lee MY, Choi SH, Shin SL, Chin H, and Kwon OJ. Distribution of B/K protein in rat
brain. Cell Tissue Res 303: 47-56, 2001.
21. Lieberthal W and Nigam SK. Acute renal failure. I. Relative importance of proximal
vs. distal tubular injury. Am J Physiol 275: F623-F631, 1998.
22. Megyesi J, DiMari J, Udvarhelyi N, Price PM, and Safirstein R. DNA synthesis is
dissociated from the immediate-early gene response in the post-ischemic kidney.
Kidney Int 48: 1451-1458, 1995.
23. Nielsen S, Kwon TH, Christensen BM, Promeneur D, Frøkiær J, and Marples D.
Physiology and pathophysiology of renal aquaporins. J Am Soc Nephrol 10: 647-663,
1999.
24. Promeneur D, Kwon TH, Frøkiær J, Knepper MA, and Nielsen S. Vasopressin V2-
receptor-dependent regulation of AQP2 expression in Brattleboro rats. Am J Physiol
279: F370-F382, 2000.
25. Rutkowski T and Kaufman RJ. A trip to ER: coping with stress. Trends Cell Biology
14: 20-28, 2004.
26. Safirstein R. Gene expression in nephrotoxic and ischemic acute renal failure. J Am
Soc Nephrol 4: 1387-1395, 1994.
27. Shanley PF, Brezis M, Spokes K, Silva P, Epstein FH, and Rosen S. Hypoxic injury
in the proximal tubule of the isolated perfused rat kidney. Kidney Int 29: 1021-1032,
1986.
28. Shao X, Davletov BA, Sutton RB, Sudhof TC, and Rizo J. Bipartite Ca2+-binding
motif in C2 domains of synaptotagmin and protein kinase C. Science 273: 248-251,
1996.
Page 16 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
17
29. Yang CW, Lim SW, Han KW, Ahn HJ, Park JH, Kim YH, Kirsh M, Cha JH, Park JH,
Kim YS, Kim J, Bang BK. Upregulation of ciliary neurotrophic factor (CNTF) and CNTF
receptor α in rat kidney with ischemia-reperfusion injury. J Am Soc Nephrol 12: 749-
757, 2001.
30. Zager RA, Johnson ACM, Hanson SY, Lund S. Ischemic proximal tubular injury
primes mice to endotoxin-induced TNF-α generation and systemic release. Am J
Physiol Renal Physiol 289: F289-F297, 2005.
Page 17 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
18
FIGURE LEGENDS
Figure 1. Light micrographs of immunostaining for B/K protein in the normal kidney.
Fifty micrometer sections of the kidney were cut on a vibratome and immunostained
with rabbit anti-rat B/K protein antibody as described in the Materials and Methods. In
the low magnification view (a), B/K protein is strongly expressed in outer and inner
stripes of the outer medulla and weakly in the cortex. In high-magnification views (b-f),
B/K protein is highly expressed in the podocyte (b, arrow), distal convoluted tubule (b,
DT), S3 segment of proximal tubule (c, S3), cortical and medullary thick ascending limb
(d, mTAL), and collecting duct (d and f). Note the strong immunoreactivity for B/K
protein in the intercalated cells (arrowheads) in the outer and inner medullary collecting
duct (d and f). No B/K immunoreactivity is observed in the proximal convoluted tubule
(b, PT) or vascular bundle (e, VB). Cx, cortex; OSOM, outer stripe of outer medulla;
ISOM, inner stripe of outer medulla; IM, inner medulla; IMi, initial part of inner medulla.
Magnifications: a, ⅹ20; b-f, ⅹ320.
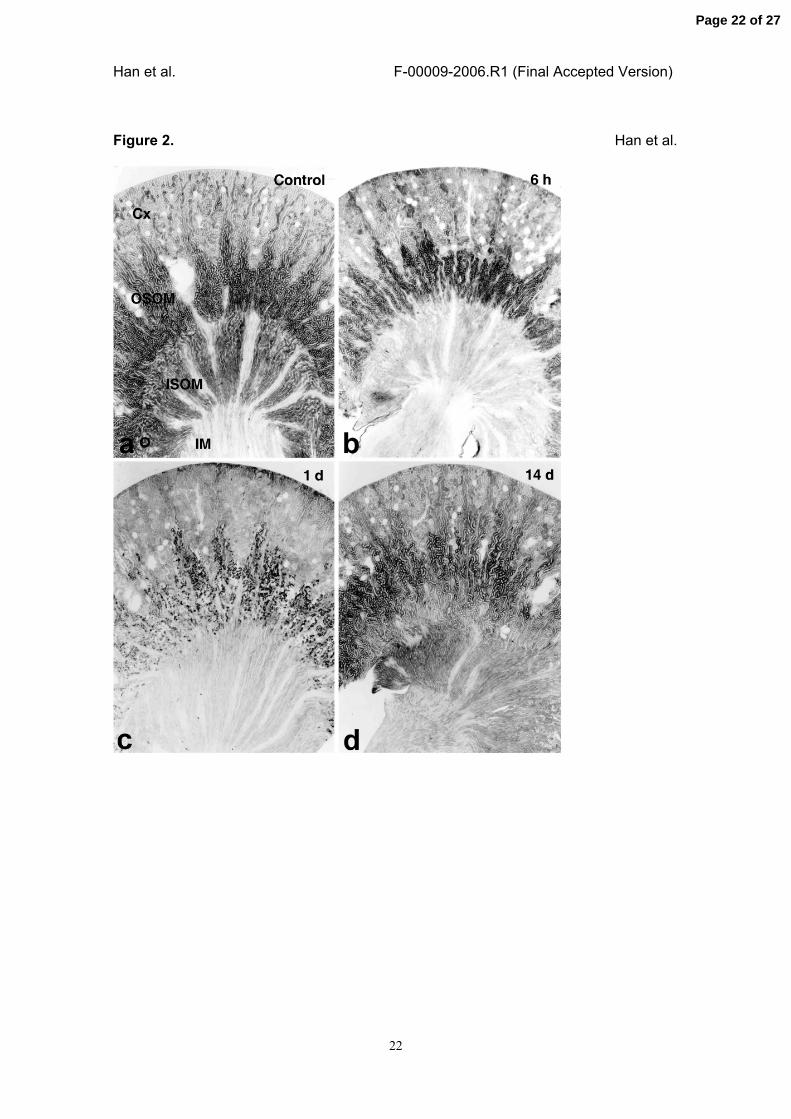
Figure 2. IR injury-induced change of B/K protein expression in the cortex (Cx), outer
stripe of outer medulla (OSOM), and inner stripe of outer medulla (ISOM).
Fifty micrometer sections of the kidney from the rats with sham operation (a) or IR-
injury for 6 hours (b) and 1 (c), and 14 days (d) were cut on a vibratome and
immunostained with rabbit anti-rat B/K protein antibody as described in Materials and
Methods. Immunostaining for B/K protein was decreased by IR injury in the distal
nephron including thick ascending limb from 6 h, reached the lowest level at day 1, but
was restored at 14 days after the injury. Magnifications: ⅹ20.
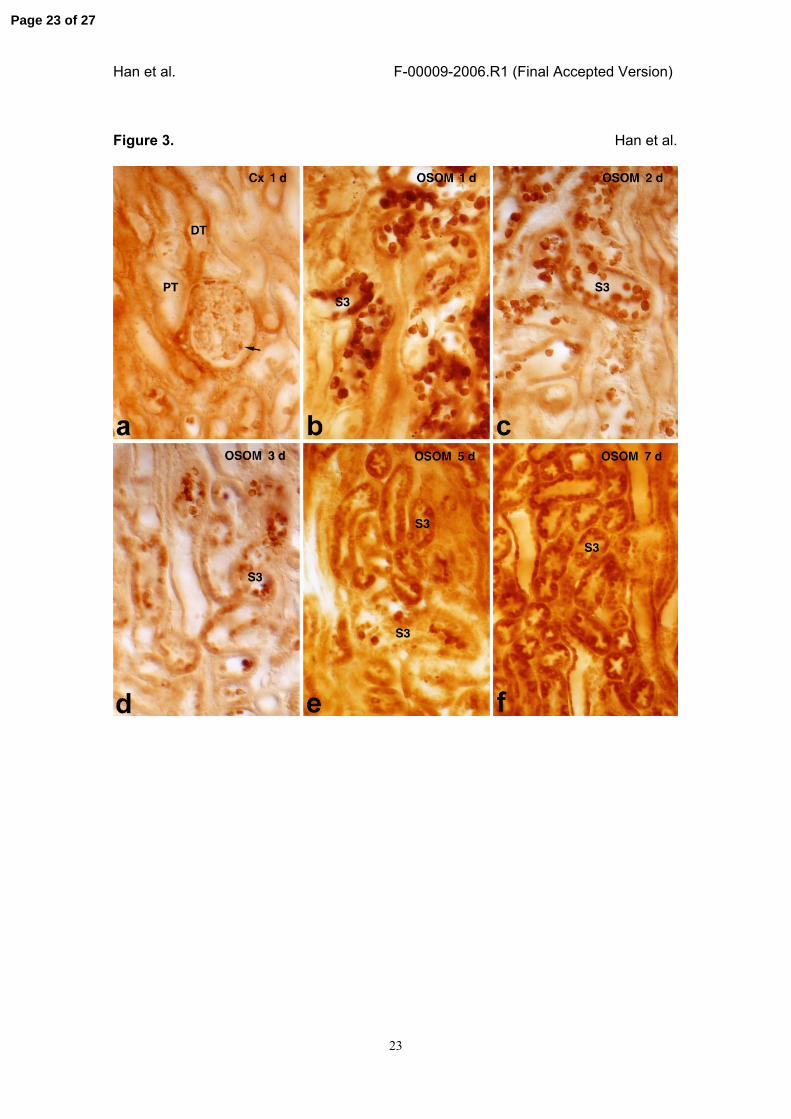
Figure 3. High-magnification views of light micrographic findings showing IR injury-
induced B/K protein expression in the cortex (Cx) and the outer stripe of the outer
Page 18 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
19
medulla (OSOM).
IR injury was induced for 1 (a and b), 2 (c), 3 (d), 5 (e), and 7 days (f), and the
expression of B/K protein in the cortex (a) or OSOM (b-f) was examined using
immunohistochemical staining methods. Immunoreactivity to B/K protein was reduced
in the podocyte (arrow) and distal convoluted tubule (DT) in the cortex (a). In contrast,
B/K expression was significantly increased in many exfoliating cells of the proximal
tubule S3 segment in the OSOM between 1 and 3 days after the injury (b-d). Note that
B/K protein in the S3 segment of the proximal tubule was restored from 5 days after the
reperfusion (e and f). Magnifications: ⅹ320.
Figure 4. Light micrograph of IR injury-induced B/K immunoreactivity in the outer stripe
of outer medulla.
IR injury was induced for 3 days, and the expression of B/K protein in the outer stripe of
outer medulla was examined using immunohistochemical staining methods in higher
magnification view. Note the cells detached in the tubular lumen, which were heavily
stained with B/K antibody. Magnification: ⅹ960.
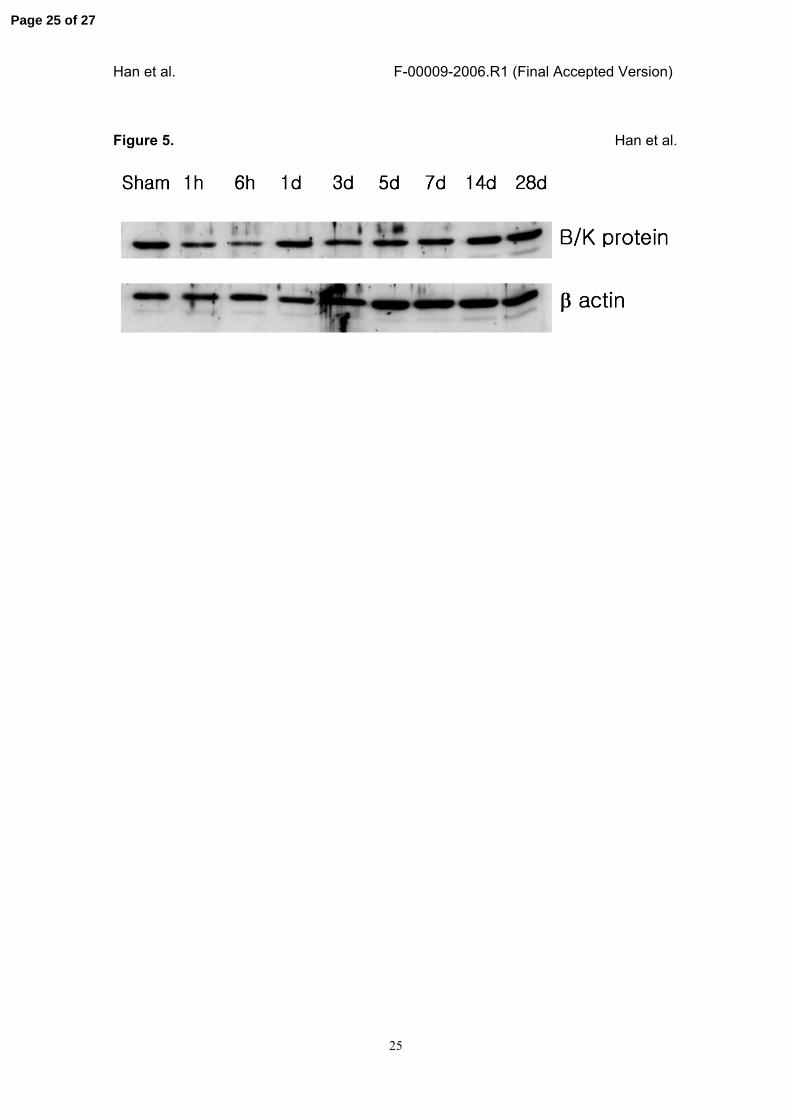
Figure 5. Temporal pattern of IR injury-induced B/K protein expression in the renal
medulla.
IR injury was induced in the rat kidney, and the medulla was dissected at the indicated
times after the reperfusion. The kidneys from sham-operated rats were used as a
control (Sham). Protein was extracted from each tissue, and the expression of B/K
protein was examined using immunoblot analysis as described in the Materials and
Methods. B/K protein immunoreactivity in the medulla was reduced immediately after
the reperfusion, reached the lowest level at 6 hours, and then returned to the control
level after 1 day. Beta actin was used as a loading control.
Page 19 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
20
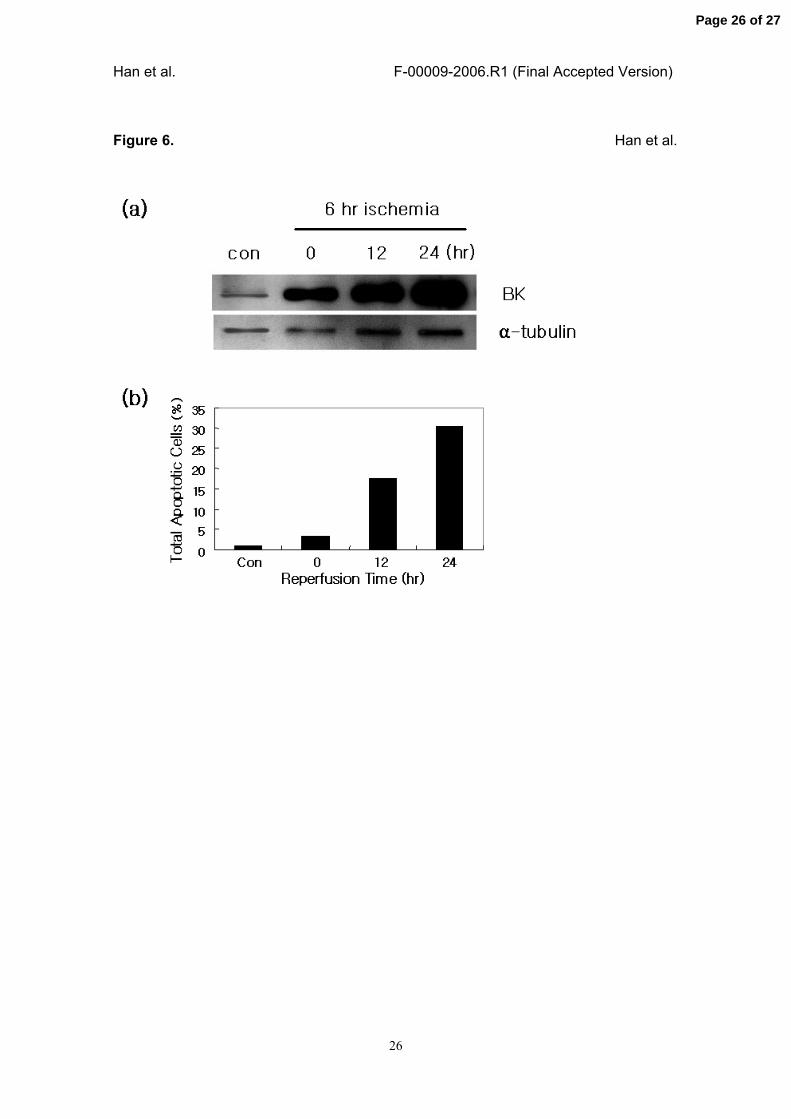
Figure 6. IR inury-induced increase of B/K protein expression in NRK-52E cells.
In vitro IR injury was induced in a rat proximal tubular epithelial cell line (NRK-52E) by
6-hour ischemia and followed by the reperfusion for the indicated times. (a) The
expression of B/K protein in each sample was examined using immunoblot analysis.
Control cells (Con) showed very low level of B/K protein expression, and it was strongly
induced by the exposure of the cells to 6-hour ischemia and further increased time-
dependently by the reperfusion. Alpha-tubulin was used as a loading control. (b) IR
injury-induced apoptosis of NRK-52E cells were analyzed using FACS methods, and
expressed as total apoptotic cells as described in Materials and Methods. Fraction of
total apoptotic cells was increased by ischemia and reperfusion, and it was almost
proportional to the level of B/K protein in the cells.
Figure 7. Immunocytochemical staining of B/K protein-positive cells in urine after IR
injury.
Fresh urine samples from control or IR injury model were obtained at the indicated
times by bladder puncture, and the cells immunopositive to B/K protein were stained as
described in Materials and Methods. Cells positive to B/K protein in urine samples were
strongly increased by IR injury, reached the peak at day 1 and then gradually
decreased until day 5. Urine collected from control animals showed few or no B/K
protein positive cells.
Page 20 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
21
Figure 1. Han et al.
Page 21 of 27
Han et al. F-00009-2006.R1 (Final Accepted Version)
22
Figure 2. Han et al.
Page 22 of 27
Han et al. F-00009-2006.R1 (Final Accepted Version)
23
Figure 3. Han et al.
Page 23 of 27

Han et al. F-00009-2006.R1 (Final Accepted Version)
24
Figure 4. Han et al.
Page 24 of 27
Han et al. F-00009-2006.R1 (Final Accepted Version)
25
Figure 5. Han et al.
Page 25 of 27
Han et al. F-00009-2006.R1 (Final Accepted Version)
26
Figure 6. Han et al.
Page 26 of 27
Han et al. F-00009-2006.R1 (Final Accepted Version)
27
Figure 7. Han et al.
Page 27 of 27




















