Late Quaternary rapid morphological evolution of an endemic diatom in Yellowstone Lake, Wyoming

Abundance and distribution of organic mound-building antsof the Formica rufa group in Yellowstone National ParkA. C. Risch1, M. F. Jurgensen2, A. J. Storer2, M. D. Hyslop2 & M. Schutz1
1 Swiss Federal Institute for Forest, Snow and Landscape Research, Birmensdorf, Switzerland
2 Ecosystem Science Center, Michigan Technological University, School of Forest Resources and Environmental Sciences, Houghton, MI, USA
Introduction
Red wood ants (Formica rufa group) are ubiquitous
in many conifer and mixed-conifer forests of Europe
and Asia and build large, organic nests (mounds)
that can be 2 m in height and 4 m in diameter (e.g.
Gosswald 1989a; b). Because of their wide occur-
rence in Eurasian forested ecosystems, these ants
have been the focus of extensive research on their
social structure (e.g. Crozier and Pamilo 1996;
Pamilo et al. 1997), geographical distribution and
density (e.g. Kissling 1985), population dynamics
Keywords
ant mounds, Formicidae, North America, red
wood ants
Correspondence
Anita C. Risch, Swiss Federal Institute for
Forest, Snow and Landscape Research,
Zuercherstrasse 111, 8903 Birmensdorf,
Switzerland. E-mail: [email protected]
Received: May 31, 2007; accepted: September
26, 2007.
doi: 10.1111/j.1439-0418.2007.01243.x
Abstract
Red wood ants (Formica rufa group) are ubiquitous in many conifer
and mixed-conifer forests of northern Europe and Asia. In contrast,
relatively little is known about the abundance and distribution of the
24 North American F. rufa group species. As ants are important com-
ponents of most soil invertebrate communities and are considered eco-
system engineers that alter the flow of energy and nutrients through
terrestrial systems, it is important to gain information on their distribu-
tion and abundance. We conducted a survey for red wood ant mounds
in Yellowstone National Park (YNP), Wyoming/Montana, USA, where
human disturbance has been kept to a minimum for over 130 years.
We found a total of 85 red wood ant mounds (0.11 to 0.17 mounds/
ha) on 327 km of roads and 180 km of the hiking trails we surveyed.
The occurrence of ant mounds was higher then expected by random
distribution at elevations between 1600 and 2400 m, annual precipita-
tion of 250 to 760 mm, middle and late successional lodgepole pine,
late successional Douglas fir forest and non-forested grassland/sage-
brush prairie vegetation. Additionally, mounds were clustered in gently
sloped not north-exposed locations and in areas that had not recently
burned. Most of the mounds detected were inhabited by Formica ob-
scuripes Forel, which occupied 94% of the mounds sampled. Based on
a multi-criteria binary Geographic Information System model that we
developed, we found that ant mounds were to be expected with a
high probability in less then 1% of the YNP area. These results
together with the detected low density and small size of the red wood
ant mounds within the study area suggest that these insects have a
much lower impact on invertebrate biodiversity and ecosystem pro-
cesses, such as forest productivity and carbon and nutrient cycling on
the ecosystem scale compared with their counterparts in European or
Asian systems.
J. Appl. Entomol.
326J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin

and behaviour (e.g. Klimetzek 1981), and effect on
invertebrate biodiversity (e.g. Laakso and Setala
1997, 1998, 2000; Hawes et al. 2002; Reznikova and
Dorosheva 2004). Other studies have investigated
the impacts of these ants on tree growth through
their interactions with defoliators and aphids (e.g.
Laakso and Setala 2000), and assessed their role in
forest ecosystem carbon and nutrient cycling (e.g.
Frouz et al. 1997; Lenoir et al. 2001; Risch et al.
2005; Domisch et al. 2006; Kilpelainen et al. 2007).
Twenty-four Formica species are placed within the
North American F. rufa group (Hedlund 2002), five of
which build large organic mounds like the Eurasian
species, while the others use smaller and variable
amounts of ‘thatch’ (organic debris) to cover their
nests (cf. Jurgensen et al. 2005). In contrast to the
Eurasian red wood ants, relatively little is known
about the abundance, distribution and ecology of the
F. rufa ants in North America, which occur in various
vegetation types ranging from meadows to sagebrush
and subalpine forests (e.g. Weber 1935; Gregg 1963;
Wheeler and Wheeler 1986; Jurgensen et al. 2005).
However, ants are important components of most
invertebrate communities (Holldobler and Wilson
1990; Folgrait 1998; Agosti et al. 2000) and are con-
sidered ecosystem engineers that alter the flow of
energy and nutrients through terrestrial systems
(Jones et al. 1994). Thus, it is important to gain a bet-
ter understanding of the distribution of red wood ants
and the role they play in North American ecosystems.
Some studies have investigated the habitat conditions
for some F. rufa group species (e.g. Weber 1935; Con-
way 1996, 1997; Lindgren and MacIsaac 2002). How-
ever, this information is not sufficient to develop
models to predict under which conditions these ants
would possibly occur and to estimate the role they
might play in North American ecosystems. To our
knowledge, no baseline information is available on
environmental factors that might constrain or benefit
the occurrence of these mound-building ants in
North America. Consequently, we conducted a sur-
vey on red wood ants in one of the most pristine eco-
systems in the United States, Yellowstone National
Park (YNP) in the states of Wyoming and Montana,
where human disturbance has been kept to a mini-
mum for over 130 years. More specifically, we were
interested in: (i) the density of red wood ant mounds
along selected trails and roads in YNP; (ii) the envi-
ronmental factors that correlate with mound occur-
rence and size; (iii) the occurrence of different red
wood ant species in YNP; and (iv) predicting which
areas of YNP would be most suitable for ant occur-
rence by developing a GIS-based habitat model.
Study area
Yellowstone National Park is located in the north-
western corner of Wyoming and the southwestern
corner of Montana, USA. Founded in 1872, YNP
encompasses nearly 9000 km2 of land. Fifty-five per-
cent of the land is covered by different aged lodge-
pole pine (Pinus contorta var. latifolia Engelm.)
stands, and approximately 26% by different aged
Douglas fir [Pseudotsuga menziesii (Mirb.) Franco],
subalpine fir [Abies lasiocarpa (Hook.) Nutt.], Engel-
mann spruce (Picea engelmannii Parry) and/or white-
bark pine (Pinus albicaulis Engelm.) stands. Thirteen
per cent of the land is non-forested vegetation
mainly dominated by grasslands or sagebrush prairie,
and the rest is covered by water.
Elevations within Yellowstone range from roughly
1500 to 3500 m. The climate features long cold win-
ters and short warm summers with an average
annual temperature of approximately 1�C. The aver-
age annual precipitation ranges from 250 mm per
year at the north boundary to 2050 mm per year in
the south west corner of the Park. Wild fires are fre-
quent in YNP and have return intervals of roughly
100 to 300 years. In the extremely dry year of 1988,
over 200 000 ha of forest were destroyed represent-
ing the largest fire event in the past 100 years.
Methods
Ant mound survey
During the summer of 2005, we surveyed for red
wood ant mounds along YNP’s roads and trails. All
paved roads with the exception of the two stretches
between Fishing Bridge Campground and the East
entrance and between Grant Village and the South
entrance were surveyed. The trails were randomly
chosen, but stratified by accessibility from the North
entrance, which lead to a higher proportion of the
trails surveyed in the northern and central part of
the park. Overall, we surveyed 327 of 431 km of the
paved roads (76%) and 180 of the 1634 km hiking
trails (11%). The roads were approximately 7 m, the
trails 1.5-m wide, and we surveyed a strip of 2.5 to
5 m on both sides of the roads and trails, respec-
tively. Consequently, a total area of 510–765 ha was
surveyed within the park. Road surveys were con-
ducted by car with a travel speed not exceeding
30 km/h. One observer in addition to the driver was
always present to ensure that all mounds were
found on both sides of the road. Upon locating
a mound along a road or a trail, we recorded its
A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz Red wood ants in Yellowstone National Park
J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin 327

coordinates with a Garmin eTrek Summit Global
Positioning System (GPS) unit (Garmin International
Inc., Olathe, Kansas, USA). Two perpendicular diam-
eters and four heights (in all four cardinal directions)
were measured on each mound. In addition, we
collected 10–15 large worker ants from each mound
and put them in 100% alcohol to preserve them for
species identification. When more than three mounds
were present in a radius of 15 m, we assumed them
to be colonies and only two of the mounds were
sampled for species identification. The ants were
identified to species, and selected specimens were
sent to Dr Andrew Suarez at the University of Illinois,
Champagne, Urbana, for confirmation. Six mound
sightings were called in by other researchers working
in YNP, and only GPS coordinates and elevation were
recorded for these mounds. No diameter measure-
ments were conducted on these mounds and no
specimens were collected.
Data analysis
The coordinates of each mound were imported into
ArcGIS 9.1 (ESRI, Redlands, California, http://
www.esri.com/) as text data and converted to a point
layer. Existing Geographic Information System (GIS)
layers were provided by the National Park Service
(NPS). These layers contained information on the
park boundary (1995, 1:62 500), 30-year average of
annual precipitation (1990; 1:125 000), elevation
above sea level [from digital elevation model (DEM),
30 m], slope and aspect (derived from DEM, 30 m),
vegetation type (1999, 1:62 500 m), and fire history
perimeters for fires larger then 0.4 km2 (2004;
1:62 500). All layers were then converted into 30-m
raster cells. The average annual precipitation was
divided into five groups (250–510 mm/year, 510–
760 mm/year, 760–1010 mm/year, 1010–1270 mm/
year, >1270 mm/year). The overstory vegetation was
grouped into ‘vegetation types’: (i) non-forest/sage-
brush; (ii) post-disturbance lodgepole stands (natural
regeneration after fire); (iii) early successional lodge-
pole pine stands; (iv) middle successional lodgepole
pine stands; (v) late successional lodgepole pine
stands; (vi) Douglas fir forest; (vii) white-bark pine
forest; (viii) Engelmann spruce forest; and (ix)
subalpine fir stands. Using this data, we calculated
the total hectares and frequency (in %) of each of
the classes for our variables: (i) average annual
precipitation; (ii) elevation; (iii) slope; (iv) aspect;
and (v) vegetation type for the entire park.
In a next step, we performed a spatial join of the
mound locations with each of the layers provided by
the NPS. This allowed us to determine the habitat
characteristics at each mound location. Following
this join, we calculated the frequencies (in %) of ant
mounds in each class of each feature. To test
whether the ant mounds were clustered or distrib-
uted randomly, we compared the frequencies of
each feature class (in %) with the frequencies of ant
mounds in each feature class (in %) using X2-test
statistics. Additionally, we used linear regression
analyses to assess whether there were relationships
between the dependent variables mean mound
height, mound diameter and mound volume (calcu-
lated from diameter and height measurements using
the equation of half an ellipsoid; Risch et al. 2005),
and each of the independent continuous variables
average annual precipitation, elevation and slope.
Analyses of variance (anova) followed by Tukey post-
hoc test for pair-wise comparisons were used to
assess whether there were relationships between the
dependent variables mean mound height, mound
diameter and mound volume and the independent
nominal variables aspect and vegetation type.
We determined the precipitation, the ranges of
elevation, slope, aspect, vegetation types and time
since last fire where ant mounds were clustered and
constructed a multi-criteria binary model for predict-
ing where red wood ants would most likely occur
within YNP. Areas that did not meet the criteria
were excluded from consideration, and areas that
met the criteria were combined to determine areas
most likely to contain ant mounds.
Results
Mound density and distribution
We located a total of 85 mounds along the 327 km
of road (51 mounds) and 180-km trail (34 mounds)
covered in our survey (fig. 1). As we surveyed a 5–
10 m wide strip along trails and roads (2.5 to 5 m on
each side), we covered an area of 510 to 765 ha.
Consequently, the density of mounds were estimated
to be roughly 0.11–0.17 mounds/ha. The mounds
showed a non-random distribution related to the
average annual precipitation (X2 = 135, df = 4,
P < 0.001; fig. 2a) with 73% of the mounds (62
mounds) located in areas with average annual pre-
cipitation of 250 to 760 mm. None of the mounds
was found in areas that received more then 1270-
mm precipitation per year. All mounds were located
at elevations between 1600 and 2650 m, with 47
(55%) of the 85 mounds detected between 2200 and
2400 m. The ant mounds also were clustered with
Red wood ants in Yellowstone National Park A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz
328J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin

regard to elevation as significantly more mounds
were found between 1600 and 2400 m elevations
than would be expected by random distribution
(X2 = 474, df = 12, P < 0.001; fig. 2b).
Eighteen (21%) of the mounds were located in
flat terrain (slope 0%); overall, only 15 (18%) of the
mounds were found on slopes steeper than 15%
(fig. 2c). Roughly, half of the mounds were located
in south-eastern to western exposures (44 mounds
or 51%); a total of 28 mounds (33%) on north-west,
north or north-east exposures (fig. 2c). The mounds
also showed a clustered distribution with regard to
slope and aspect. They were found more frequently
then expected by chance in terrain with slope
between 0% and 15% (X2 = 28, df = 5, P < 0.001)
and in locations with south-eastern to western and
flat conditions (X2 = 29, df = 8, P < 0.001). No infor-
mation on slope and aspect was available for six
(7%) of the 85 mounds.
Fifty-one of the 85 mounds (59%) were found in
forested habitat: six mounds (7%) in late succes-
sional Douglas fir stands, one in post-disturbance,
two in early, 24 in middle and 18 in late succes-
sional lodgepole pine stands (fig. 2d). No mounds
were detected in Engelmann spruce, subalpine fir or
Fig. 1 Locations of ant mounds (dots) found within Yellowstone
National Park. The surveyed roads (solid black lines) and trails (dashed
black lines) and non-surveyed roads (solid grey lines) and trails (dashed
grey lines), respectively, are shown. A total of 510 to 765 ha (2.5–5 m
on both sides of the roads and trails) were surveyed in our study.
(a) (b)
(c) (d)
Fig. 2 Frequency of the mounds related to the frequencies of different biotic and abiotic parameters: (a) average annual precipitation (mm/year);
(b) elevation above sea level (m); (c) slope (%) and aspect; and (d) vegetation type.
A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz Red wood ants in Yellowstone National Park
J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin 329

white-bark pine stands. Thirty-one mounds (36%)
were located in non-forested areas. For four mounds
(5%), no information on vegetation type was avail-
able. Again, the occurrence of ant mounds was clus-
tered with regard to vegetation type: significantly,
more mounds then expected were detected in late
successional Douglas fir, middle and late-successional
lodgepole pine and non-forested areas (X2 = 133,
df = 7, P < 0.001). In particular, post-disturbance
lodgepole pine stands seemed to be avoided by the
insects. A total of 59 mounds (69%) were located in
areas that were not subject to forest fires or did burn
before 1988, while 26 mounds (31%) were located
in forests that were subject to the large 1988 fires
that burned 240 km2 of the YNP area. Ten and 14 of
these 26 mounds were, however, less then 1000 and
2000 m, respectively, from unburned habitat.
The model we developed to predict where red
wood ant mounds would occur with the highest
likelihood within YNP showed that only limited
areas would be suitable habitat for these insect spe-
cies (fig. 3). Locations that met all six criteria (pre-
cipitation, elevation slope, aspect, vegetation, fire
history) totalled 7969 ha or 0.89% of the park’s
889 394 ha only.
Mound size and volume
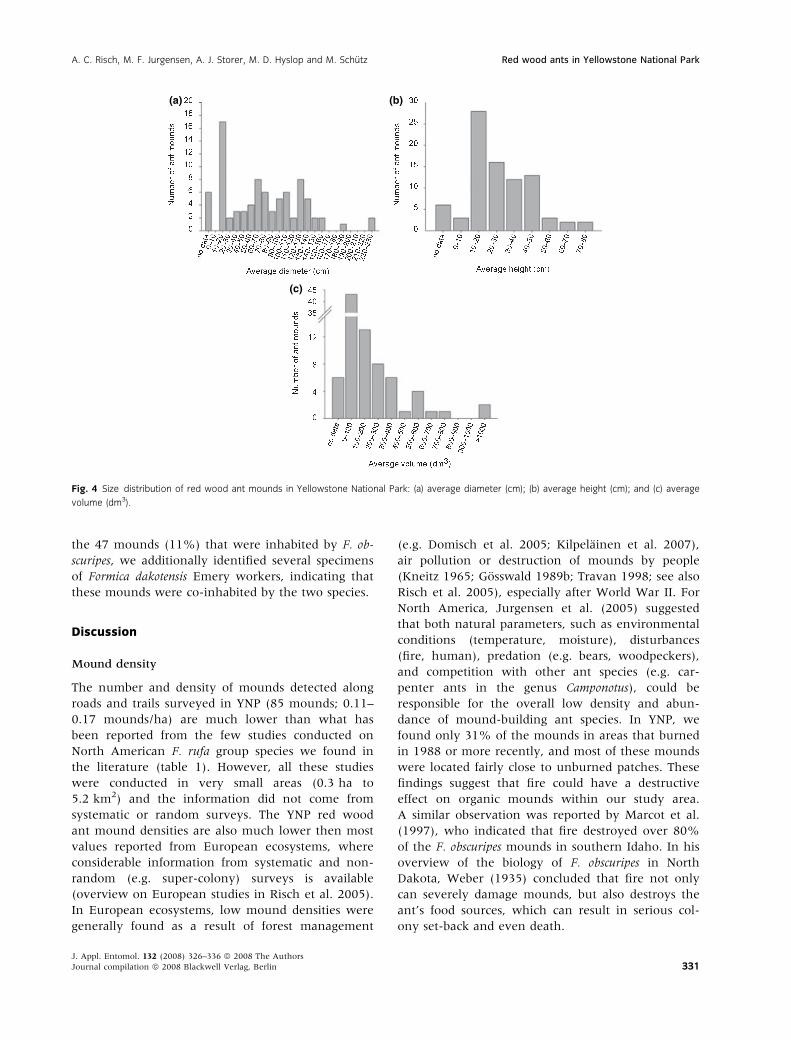
For six (7%) of the 85 mounds, no information on
mound size was collected. The diameter of the
remaining 79 mounds ranged from 10 to 228 cm,
the height from 5 to 76 cm, and volume from 1 and
1960 dm3, respectively (fig. 4). The average mound
diameter of the 79 mounds was 78 cm, average
height 28 cm and average volume 175 dm3 (table 1).
No relationships were detected between the size of
the mound (height, diameter or volume) and the
environmental parameters, such as precipitation, ele-
vation, slope, aspect and vegetation type (P > 0.05
for all linear regressions and anova).
Ant species
Overall, we collected worker ants from a total of 50
of the 85 mounds for species identification. Forty-
seven (94%) of these 50 mounds were inhabited by
Formica obscuripes Forel. Two mounds (4%) were
inhabited by Formica oreas Wheeler and one mound
(2%) by Formica puberula Emery, which is not con-
sidered a F. rufa group species, but placed within the
Formica sanguinea group (Hedlund 2002). In five of
Fig. 3 Potential habitat map for mound-building red wood ants in Yellowstone National Park. The black areas (0.89% or 7969 ha of the entire park)
represent the locations where all six input criteria were met and where ant mound occurrence were expected with a maximum likelihood. The
input criteria were the following: (i) elevation 250–760 mm/year (28.5% or 635 538 ha of the park); (ii) elevation 1600–2400 m (40.6% or
361 271 ha); (iii) slope 0–15% (59% or 364 644 ha); (iv) aspect flat, southeast–southwest (SE–SW) (24.5% or 218 370 ha); (v) vegetation type, middle
and late successional lodgepole pine, late successional Douglas fir forest and non-forested grassland/sagebrush prairie vegetation (40.2% or
532 076 ha); and (vi) time since last fire; never burned or burned before 1988 (44.1% or 391998 ha).
Red wood ants in Yellowstone National Park A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz
330J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin
the 47 mounds (11%) that were inhabited by F. ob-
scuripes, we additionally identified several specimens
of Formica dakotensis Emery workers, indicating that
these mounds were co-inhabited by the two species.
Discussion
Mound density
The number and density of mounds detected along
roads and trails surveyed in YNP (85 mounds; 0.11–
0.17 mounds/ha) are much lower than what has
been reported from the few studies conducted on
North American F. rufa group species we found in
the literature (table 1). However, all these studies
were conducted in very small areas (0.3 ha to
5.2 km2) and the information did not come from
systematic or random surveys. The YNP red wood
ant mound densities are also much lower then most
values reported from European ecosystems, where
considerable information from systematic and non-
random (e.g. super-colony) surveys is available
(overview on European studies in Risch et al. 2005).
In European ecosystems, low mound densities were
generally found as a result of forest management
(e.g. Domisch et al. 2005; Kilpelainen et al. 2007),
air pollution or destruction of mounds by people
(Kneitz 1965; Gosswald 1989b; Travan 1998; see also
Risch et al. 2005), especially after World War II. For
North America, Jurgensen et al. (2005) suggested
that both natural parameters, such as environmental
conditions (temperature, moisture), disturbances
(fire, human), predation (e.g. bears, woodpeckers),
and competition with other ant species (e.g. car-
penter ants in the genus Camponotus), could be
responsible for the overall low density and abun-
dance of mound-building ant species. In YNP, we
found only 31% of the mounds in areas that burned
in 1988 or more recently, and most of these mounds
were located fairly close to unburned patches. These
findings suggest that fire could have a destructive
effect on organic mounds within our study area.
A similar observation was reported by Marcot et al.
(1997), who indicated that fire destroyed over 80%
of the F. obscuripes mounds in southern Idaho. In his
overview of the biology of F. obscuripes in North
Dakota, Weber (1935) concluded that fire not only
can severely damage mounds, but also destroys the
ant’s food sources, which can result in serious col-
ony set-back and even death.
(a) (b)
(c)
Fig. 4 Size distribution of red wood ant mounds in Yellowstone National Park: (a) average diameter (cm); (b) average height (cm); and (c) average
volume (dm3).
A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz Red wood ants in Yellowstone National Park
J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin 331

Ta
ble
1V
eg
eta
tio
nty
pe
an
de
lev
ati
on
of
mo
un
dlo
cati
on
s,n
um
be
ro
fm
ou
nd
s/h
a,
nu
mb
er
of
mo
un
ds
inst
ud
y,
av
era
ge
he
igh
t,d
iam
ete
ra
nd
vo
lum
eo
fre
dw
oo
da
nt
mo
un
ds
inY
ello
wst
on
e
Na
tio
na
lP
ark
an
din
oth
er
No
rth
Am
eri
can
eco
syst
em
s
Ve
ge
tati
on
typ
e(s
)
Ele
va
tio
n
(m)
Re
dw
oo
da
nt
mo
un
d
So
urc
e
No
.m
ou
nd
s
pe
rh
a
No
.m
ou
nd
s
inst
ud
y
He
igh
t
(cm
)D
iam
ete
r(c
m)
vo
lum
e(m
3)
Sp
eci
es
Th
isst
ud
y,
Ye
llow
sto
ne
Na
tio
na
lP
ark
Lod
ge
po
lep
ine
,D
ou
gla
s
fir,
Sa
ge
bru
sh
21
78
(25
)0
.11
–0
.17
85
28
(2)
78
(6)
0.1
7(0
.03
)F
.o
bsr
uci
pe
s,
F.
da
kote
nsi
s,
F.
ore
as,
F.
pu
be
rula
Oth
er
stu
die
s
De
cid
uo
us
shru
bs,
pra
rie
––
–2
01
60
–1
10
10
.02
51
F.
ob
scu
rpie
sW
eb
er
19
35
;N
ort
hD
ako
ta
Lod
ge
po
lep
ine
––
11
00
20
0–
25
0F
.o
bsc
uri
pe
sLi
nd
gre
na
nd
Ma
cIsa
ac
20
02
;B
riti
shC
olu
mb
ia
Alp
ine
me
ad
ow
24
23
–4
02
1�
58
4�
20
–9
5�
23
0.2
5�
0.0
9F
.o
bsc
uri
pe
sM
cCa
ho
na
nd
Loo
kwo
od
19
90
;W
yo
min
g
Mix
ed
con
ife
r(g
ran
d
an
dD
ou
gla
sfir
)
–7
32
93
––
0.0
01
–>
1F
.o
bsc
uri
pe
sM
cIv
er
et
al.
19
97
;O
reg
on
Sa
ge
bru
sh1
29
0–
13
55
1.2
12
––
–F
.o
bsc
uri
pe
sM
cIv
er
an
dY
an
de
ll1
99
8;
Ore
go
n
Sa
ge
bru
sh2
56
01
15
85
26
26
52
–F
.o
bsc
uri
pe
sC
on
wa
y1
99
6,
19
97
;C
olo
rad
o
Sa
ge
bru
sh1
30
0–
4–
––
F.
ob
scu
rip
es
McI
ve
ra
nd
Ste
en
19
94
;O
reg
on
Sa
ge
bru
sh1
40
0–
5–
––
F.
ob
scu
rip
es
McI
ve
ra
nd
Loo
mis
19
93
;O
reg
on
Sa
ge
bru
sh2
30
0–
15
––
–F
.o
bsc
uri
pe
sJe
nks
an
dR
eit
he
l2
00
4;
Co
lora
do
Ab
an
do
ne
dra
ilro
ad
be
ad
14
60
–4
0–
––
F.
ob
scu
rip
es
O’N
eil
an
dK
em
p1
99
0;
Mo
nta
na
Sh
rub
-ste
pp
eh
ab
ita
t1
60
0–
6–
––
F.
ob
scu
rip
es
He
ikki
ne
n1
99
9;
Uta
h
Bla
cksp
ruce
/larc
h–
7–
17
4–
5–
37
4–
80
–F
.d
ako
ten
sis
Fra
nco
eu
ra
nd
Pe
pin
19
75
;Q
ue
be
c
Sa
ge
bru
sh2
00
0–
17
––
–F
.p
lan
ipili
s
Cre
igh
ton
McI
ve
ra
nd
Loo
mis
19
93
;C
alif
orn
ia
Ye
llow
pin
ea
nd
fir2
10
05
.71
08
––
–F
.h
ae
mo
rrh
oid
alis
Em
ery
Ma
cKa
ya
nd
Ma
cKa
y1
98
4;
Ca
lifo
rnia
Ab
an
do
ne
dfie
ld–
–8
91
73
50
3–
F.
sub
nit
en
s
Cre
igh
ton
Ay
re1
95
7;
Bri
tish
Co
lum
bia
Cle
ari
ng
inh
ard
wo
od
fore
st–
3.7
19
––
–F
.o
bsc
ruri
ve
ntr
is
Ma
yr
Ta
lbo
t1
96
3;
Mic
hig
an
F.,
Fo
rmic
a.
1a
ve
rag
es
of
en
tire
stu
dy
,h
eig
ht
ran
ge
s2
.5–
45
.7cm
,d
iam
ete
rra
ng
es
30
.5–
34
3cm
.2A
ve
rag
es
of
en
tire
stu
dy
,h
eig
ht
ran
ge
s6
–5
8cm
,d
iam
ete
rra
ng
es
19
–1
42
cm.
3A
ve
rag
es
of
en
tire
stu
dy
,h
eig
ht
ran
ge
s3
–3
6cm
,d
iam
ete
rra
ng
es
15
–1
17
cm;
sta
nd
ard
err
or
inp
are
nth
esi
s,
sta
nd
ard
de
via
tio
ng
ive
na
fte
r�
.
Red wood ants in Yellowstone National Park A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz
332J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin

Similar to fire, the sizeable grizzly bear population
in YNP [350 individuals in 2005; Interagency Grizzly
Bear Study Team (IGBST) 2005] might affect the
abundance of the mound-building ants. Mattson
(2001) showed that F. rufa group ants contributed
between 4.2% and 4.7% in July, and 22–27% in
August, respectively, to the grizzly bears’ diet (aver-
age values for the years 1977 to 1992). In 1977, they
even reported 28% of the bear’s diet to be composed
of red wood ants for July and 35% for August.
North American black bears have also been shown
to consume red wood ants (Colorado: Conway 1997;
Minnesota: Noyce et al. 1997; Oregon: Bull et al.
2001; Utah: Auger et al. 2004); however, no data on
black bears diet are available for YNP.
Our results obtained from a survey method that
used non-randomly distributed linear features
instead of random or systematically selected plots,
may not mirror the ‘true’ density of red wood ant
mounds in YNP. However, given the large area sur-
veyed and talking with YNP employees who work
and hike on and off-trail, we believe that the general
results of our survey are representative of F. rufa
group ants within the park. Our low estimate of
mound density rather is an over- than an underesti-
mation, as YNP fire crews would extinguish more
fires along the roads, and bears are disturbed by
tourists along the road side. Both these factors could
lead to ‘low disturbance’ zones for the ants, and
actually favour the formation and survival of
mounds. Further, ants have been shown to favour
forest edges (e.g. Sudd et al. 1977; Punttila 1996), as
more sunlight can penetrate the ground and the cli-
matic condition are more favourable compared with
the closed forest. Consequently, it is possible that
the nests of mound-building ants in YNP were clus-
tered along trails and roads, which would again lead
to an overestimation of mound density in our study.
Overall, the low densities of F. rufa group ant
mounds that we detected in YNP suggest that these
ants do not play a similar dominant role within this
ecosystem compared with European and Asian spe-
cies that have been reported to have a large effect
on: (i) the spatial heterogeneity of carbon and nutri-
ent stores and fluxes (e.g. Frouz et al. 1997; Lenoir
et al. 2001; Risch et al. 2005; Domisch et al. 2006;
Kilpelainen et al. 2007); (ii) the invertebrate biodi-
versity (e.g. Laakso and Setala 2000; Hawes et al.
2002; Reznikova and Dorosheva 2004); and (iii) tree
growth (e.g. Rosengren and Sundstrom 1991). How-
ever, in a next step, we would like to survey those
areas within YNP for which our model predicted ants
to occur with a high probability. This would help to
provide knowledge on whether these insects occur
in high abundances at specific locations. If they do
so, it is possible that they affect the ecosystem prop-
erties and processes at very small spatial scales, simi-
lar to findings by Heikkinen (1999). This author
showed that F. obscuripes had a negative effect on
the spider population of the sagebrushes located
immediately around mounds in a shrub-steppe habi-
tat in Utah.
Mound distribution and size
We detected that the ant mounds found within YNP
were clearly clustered with regard to average annual
precipitation, elevation, slope, aspect and vegetation
type. These conditions likely provide a favourable
temperature and moisture regime and/or better food
resources for the insects under study. Above-ground
mounds have the benefit of heating up more rapidly
on a daily and/or seasonal basis; however, they may
be vulnerable to temperature extremes (Holldobler
and Wilson 1990). Thus, building nests within a cer-
tain range of elevation and exposure might help
optimize temperature conditions for the mounds.
Similarly, building mounds in areas that receive
a certain amount of precipitation might also help the
insects to maintain an environment that is not too
dry, but also to assure that the mounds do not get
flooded. Weber (1935), for instance, suggested that
by placing thatch over the nest, F. obscuripes may
reduce the need for reliable rainfall; however, it is
not known to what extent.
Little information is available on the conditions in
which red wood ant mounds have been detected in
North America. Our literature survey revealed that
elevation ranges from 1300 to 2600 m seemed to be
dominating (table 1); however, some taxonomists
also detected mounds at elevations of up to 3200 m
(Gregg 1963; Wheeler and Wheeler 1986). Weber
(1935) reported F. obscuripes to occur in locations
with mean annual precipitation of 130 to 890 mm,
and some authors showed that F. obscuripes prefers
open areas and were rarely detected in dense woods
(Weber 1935; MacKay and MacKay 1984; McCahon
and Lookwood 1990; Conway 1996). However, for
all these studies, no qualitative or quantitative data
was presented. Only MacKay and MacKay (1984)
showed that most of their 108 Formica haemorrhoidal-
is Emery mounds detected were located on warmer
east, south-east and south exposed sites, which is
similar to our results. Furthermore, European F. rufa
group species have been reported to prefer warmer
south-southeast exposed slopes over slopes with
A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz Red wood ants in Yellowstone National Park
J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin 333

cooler northern exposures, as temperature
conditions are more favourable (Forel 1920;
Klimetzek 1970; Bretz 1971; Sossna 1973; Travan
1998). Most of the studies listed in table 1 were con-
ducted in either sagebrush habitat or other open
areas, and only a few in forests. Yet, our results
suggest that besides the non-forested habitat, the
older forest stands are also preferred by the ants.
This would be similar to what has been reported
from Europe, where red wood ant mounds were
numerous in late- compared to early-succession
Swiss conifer forests, and in 100- compared with
5-year old Norway spruce stands in Finland (Risch
et al. 2005; Kilpelainen et al. in press).
Similar sized F. rufa group mounds to those we
located were found in studies conducted in other
North American ecosystems (see table 1). However,
we could not find any relationship between the ant
mound size and the environmental parameters we
tested. From Europe, it is known that mound sizes
tended to be somewhat larger in older forests, sug-
gesting that mound size might be positively corre-
lated to the lack of disturbance (Domisch et al. 2005;
Risch et al. 2005; Kilpelainen et al. 2007). In Euro-
pean forests, researchers also found a tendency
towards larger mounds on north compared with
east, west and south slopes (Klimetzek 1970; Sossna
1973; Gosswald 1989a; Risch et al. 2005). The likely
reason is that, in these locations, temperatures tend
to be cooler, and more insulation is needed for the
mound to maintain temperature.
Ant species
Formica obscuripes was the most abundant F. rufa
group species in YNP. This result is similar to what
Jurgensen et al. (2005) indicated in their review of
North American F. rufa group species, and is also
mirrored by the studies summarized in table 1.
Formica dakotensis is also reported to have a wide
distribution in the United States (e.g. Hedlund
2002; Jurgensen et al. 2005). However, in our
study, we did not find any F. dakotensis mounds,
but only specimen of this species as co-inhabitants
of F. obscuripes mounds. It is known that some
European F. rufa group species can form mixed
mounds when one species takes over the mound
of another species, but does not kill the former
queen(s) (Collingwood 1979; pers. comm. P. Pun-
tilla, L. Sundstrom). Czechowski and Radchenko
(2006) suggested that the mixed mounds inhabited
by Formica aquilonia Yarrow, Formica polyctena Forst
and F. rufa L. they found in southern Finland
could have been formed in two ways: (i) an
already mixed group of fundatrices (colony foun-
ders) was present when a colony of Formica fusca
(slave species) was taken over; or (ii) individuals
of one species were adopted by an existing mound
of another species, especially if that colony has lost
its own queen(s). We could not find any literature
that would have described such mixed-mound
behaviour for the two species that were associated
with each other in YNP. Yet, given that we found
11% of the mounds sampled to be housed by the
two species, this phenomenon may be fairly fre-
quent.
The other two species detected in our study,
F. oreas and F. puberula, inhabited only three of
the mounds. Based on Hedlund (2002), F. oreas’
range of distribution is in most states of the north-
western USA, while F. puberula can occur all over
the western USA from Washington State to Texas.
However, we did not find any information on the
distribution or ecology of these two species in the
literature.
Conclusions
Overall, our results suggest that red wood ants
likely would not dominate the invertebrate commu-
nity at the ecosystem scale within YNP, and there-
fore, would have a lower impact on the overall
biodiversity compared with their counterparts in
Eurasian ecosystems. Further, they are likely to be
less important for increasing the spatial heterogene-
ity of carbon and nutrient flows as has been shown
from European ecosystems. Based on our review of
the literature, it is likely that this would be similar
for other temperate ecosystems in North America.
Acknowledgements
We are grateful for the support by the YNP Service
administration. We also like to thank Heidi Anderson,
Vicky Regula, Kerey Barnowe-Meyer, Georgina Ob-
rist, Monika Wysser, Vincent Green and Christine
Smith for reporting GPS coordinates from mounds
they found during their work. Justin Rosemier and
Andrew Suarez assisted with species identification.
This research was founded by Swiss-National Science
Foundation fellowships PBEZ-104320 and PBEZA-
104320 grant, by funding provided by Michigan
Technological University, School of Forest Resources
and Environmental Sciences and by the vegetation-
soils sub-project of the Swiss-NFI (National Forest
Inventory).
Red wood ants in Yellowstone National Park A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz
334J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin

References
Agosti D, Majer JD, Alonso LE, Schultz TR, 2000. Ants:
standard methods for measuring and monitoring biodi-
versity. Smithsonian Inst., Washington DC.
Auger J, Ogborn GL, Pritchett CL, Black HL, 2004. Selec-
tion of ants by the American black bear (Ursus americ-
anus). W. North Am. Nat. 64, 166–174.
Ayre GL, 1957. Ecological notes on Formica subnitens
Creighton (Hymenoptera: Formididae). Insect. Soc. 5,
173–175.
Bretz D, 1971. Wie bauen Waldameisen ihr Nest? Kos-
mos 67, 200–204.
Bull EL, Toegersen TT, Wertz T, 2001. The importance
of vegetation, insects and neonate ungulates in black
bear diet in northeastern Oregon. Northw. Sci. 75,
244–253.
Collingwood CA, 1979. The Formicidae (Hymenoptera) of
Fennoscandia and Denmark. Fauna Entomol. Scand. 8,
1–174.
Conway JR, 1996. A field study of the nesting ecology of
the thatching ant, Formica obscuripes Forel, at high alti-
tude in Colorado. Great Basin Nat. 56, 326–332.
Conway JR, 1997. Foraging activity, trails, food sources
and predators of Formica obscuripes Forel (Hymenoptera:
Fomicidae) at high altitude in Colorado. Pan-Pac. Ento-
mol. 73, 172–183.
Crozier RH, Pamilo P, 1996. Evolution of social insect
colonies. Oxford University Press, Oxford.
Czechowski W, Radchenko A, 2006. Do permanently
mixed colonies of wood ants (Hymenoptera: Formicidae)
really exist? Ann. Zool. Warszawa 56, 667–673.
Domisch T, Finer L, Jurgensen MF, 2005. Red wood ant
mound densities in managed boreal forests. Ann. Zool.
Fenn. 42, 277–282.
Domisch T, Finer L, Ohashi M, Risch AC, Sundstrom L,
Niemela P, Jurgensen MF, 2006. Contribution of red
wood ant mounds to forest floor CO2 efflux in boreal
coniferous forests. Soil Biol. Biochem. 38, 2425–2433.
Folgrait PJ, 1998. Ant biodiversity and its relationship to
ecosystem functioning: a review. Biodivers. Conserv. 7,
1221–1244.
Forel A, 1920. Les fourmis de la Suisse. Second edition.
Soc. Helv. Sci. Nat., La Chaux-des-Fonds.
Francoeur A, Pepin D, 1975. Productivite de la fourmi
Formica dakotensis dans la pressiere tourbeuse. 1. Den-
site observee et densite estimee des colonies. Insect.
Soc. 22, 135–150.
Frouz J, Santruckova H, Kalcik J, 1997. The effect of
wood ants (Formica polyctena Foerst.) on the transfor-
mation of phosphorus in a spruce plantation. Pedobio-
logia 41, 437–447.
Gosswald K, 1989a. Die Waldameise. Band 1. Biologische
Grundlagen, Okologie und Verhalten. AULA,
Wiesbaden.
Gosswald K, 1989b. Die Waldameise. Band 2. Die
Waldameise im Okosystem Wald, ihr Nutzen und ihre
Hege. AULA, Wiesbaden.
Gregg RE, 1963. The ants of Colorado with reference to
their ecology, taxonomy and geographic distribution.
Univ. Colorado, Fort Collins.
Hawes C, Stewart AJA, Evans HF, 2002. The impact of
wood ants (Formica rufa) on the distribution and abun-
dance of ground beetles (Coleoptera carabidae) in a Scots
pine plantation. Oecologia 131, 612–619.
Hedlund KS, 2002. Genus Formica. [WWW document]
http://www.cs.unc.edu/~hedlund/ants/catalog/
Heikkinen MW, 1999. Negative effects of the western
thatching ant (Formica obscuripes) on spiders (Araneae)
inhabiting big sagebrush (Artemisia tridentate). Great
Basin Nat. 59, 380–385.
Holldobler B, Wilson EO, 1990. The Ants. Harvard, Cam-
bridge.
IGBST, 2005. Interagency Grizzly Bear Study Team
annual report.
Jenks K, Reithel J, 2004. Impact of the western thatching
ant (Formica obscuripes) on invertebrate abundance and
diversity. URBEE 1, 3 URL http://www.urbee.org/vol1/
iss1/art3
Jones CG, Lawton JH, Shachak M, 1994. Organisms as
ecosystem engineers. Oikos 69, 373–386.
Jurgensen MF, Storer AJ, Risch AC, 2005. Red wood ants
in North America. Ann. Zool. Fenn. 42, 235–242.
Kilpelainen J, Finer L, Niemla P, Domisch T, Neuvonen
S, Ohashi M, Risch AC, Sundstrom L, 2007. Carbon,
nitrogen and phosphorus dynamics of ant mounds
(Formica rufa group) in managed boreal forests of dif-
ferent successional stages. Appl. Soil Ecol. 36, 156–163.
Kilpelainen J, Punttila P, Finer L, Niemla P, Domisch T,
Neuvonen S, Ohashi M, Risch AC, Sundstrom L, 2008.
Mound and species distribution of mound-building For-
mica ants in manage spruce stands of different age in
eastern Finland. J. Appl. Entomol, in press.
Kissling EM, 1985. Untersuchungen uber die Biot-
opanspruche und einen allfalligen Ruckgang von roten
Waldameisen aus der Formica rufa-Gruppe in der
Schweiz. Ph.D. thesis, ETH Zurich.
Klimetzek D, 1970. Zur Bedeutung des Kleinstandortes
fur die Verbreitung hugelbauender Waldameisen der
Formica rufa-Gruppe (Hymenoptera: Formicidae). Zeitsch.
Angew. Entomol. 66, 45–48.
Klimetzek D, 1981. Population studies on hill building
wood-ants of the Formica rufa-Group. Oecologia 48,
418–421.
Kneitz G, 1965. Formica-Arten mit vegetabilischem Nest-
bau in den Gurktaler Alpen (Karnten). Waldhygiene 5,
240–250.
Laakso J, Setala H, 1997. Nest mounds of red wood ants
(Formica aquilonia): hot spots for litter-dwelling earth-
worms. Oecologia 111, 565–569.
A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz Red wood ants in Yellowstone National Park
J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin 335

Laakso J, Setala H, 1998. Composition and trophic
structure of detrital food web in ant nest mounds of
Formica aquilonia and in the surrounding forest soil.
Oikos 81, 266–278.
Laakso J, Setala H, 2000. Impacts of wood ants (Formica
aquilonia Yarr.) on the invertebrate food web of the
boreal forest floor. Ann. Zool. Fenn. 37, 93–100.
Lenoir L, Persson T, Bengtsson J, 2001. Wood ant nests
as potential hot spots for carbon and nitrogen minerali-
zation. Biol. Fert. Soil 34, 235–240.
Lindgren SB, MacIsaac AM, 2002. A preliminary study of
ant diversity and of ant dependence on dead wood in
central interior British Columbia. USDA Forest Service,
Gen. Tech. Rep. PSW-GTR-181, 111–119.
MacKay E, MacKay WP, 1984. Biology of the thatching
ant Formica haemorrhoidalis Emery (Hymenoptera: Form-
icidae). Pan-Pac. Entomol. 60, 79–87.
Marcot BG, Castellano MA, Christy JA, Croft LK, Le-
hmkuhl JF, Naney RH, Rosentreter RE, Sandquist RE,
Zieroth E, 1997. Terrestrial ecology assessment. In: An
assessment of ecosystem components in the interior
Columbia Basin and portions of the Klamath and Great
Basins. Vol. III. Ed. by Quigley TM, Arbelbide SJ.
USDA Forest Service Gen. Tech. Rep. PNW-GTR-405.
USDA Forest Service Pac. North. Res. Stn., Portland,
OR, 1497–1713.
Mattson DJ, 2001. Myrmecophagy by Yellowstone grizzly
bear. Can. J. Zool. 79, 779–793.
McCahon TJ, Lookwood JA, 1990. Nest architecture and
pedoturbation of Formica obscuripes Forel (Hymenoptera:
Formicidae). Pan-Pac. Entomol. 66, 147–156.
McIver JD, Loomis C, 1993. A size-distance relation in
Homoptera-tending thatch ants (Formica obscuripes, For-
mica planipilis). Ins. Soc. 40, 207–218.
McIver JD, Steen T, 1994. Use of secondary nest in great
basin desert thatch ants (Formica obscuripes Forel). Great
Basin Nat. 54, 359–365.
McIver JD, Yandell K, 1998. Honeydew harvest in the
western thatching ant (Hymenoptera: Formicidae). Am.
Entomol. Spring, 30–35.
McIver JD, Torgersen TR, Cimon NJ, 1997. A supercol-
ony of the thatch ant Formica obscuripes Forel (Hyme-
noptera: Formicidae) from the Blue Mountains of
Oregon. Northw. Sci. 71, 18–29.
Noyce KV, Kannowski PB, Riggs MR, 1997. Black bears
as ant-eaters: seasonal associations between bear
myrmecophagy and ant ecology in north-central
Minnesota. Can. J. Zool. 75, 1671–1686.
O’Neil KM, Kemp WP, 1990. Worker response to thermal
constraints in the ant Formica obscuripes (Hymenoptera:
Formicidae). J. therm. Biol. 15, 133–140.
Pamilo P, Gertsch P, Thoren P, Seppa P, 1997. Molecular
population genetics of social insects. Ann. Rev. Ecol.
Syst. 28, 1–25.
Punttila P, 1996. Succession, forest fragmentation, and
the distribution of wood ants. Oikos 75, 291–298.
Reznikova Z, Dorosheva H, 2004. Impacts of red wood
ants Formica polyctena on the spatial distribution and
behavioural patterns of ground beetles (Carabidae).
Pedobilogia 48, 15–21.
Risch AC, Jurgensen MF, Schuetz M, Page-Dumroese DS,
2005. The contribution of red wood ants to soil C and
N pools and CO2 emissions in subalpine forests. Ecol-
ogy 86, 419–430.
Rosengren R, Sundstrom L, 1991. The interaction between
red wood ants, Cinara aphids and pines. A ghost of
mutualism past? In: Ant-plant interactions. Ed. By
Huxlea CR, Cuttler DF, Oxford Univ., Oxford, 80–91.
Sossna H, 1973. Untesuchungen zur Abhangigkeit des
Nestbauverhaltens der Waldameise Formica polyctena
von den Umweltfaktoren Warme, Licht und Feu-
chtigkeit im Labor. Zulassungsarbeit, Univ. Wurzburg.
Sudd JH, Douglas JM, Gaynard T, Murray DM, Stockdale
JM, 1977. The distribution of wood-ants (Formica lugu-
bris Zetterstedt) in a northern English forest. Ecol.
Entomol. 2, 301–313.
Talbot M, 1963. Nest structure and flights of the ant For-
mica obscuriventris Mayr. Anim. Behav. 12, 154–158.
Travan J, 1998. Uber den Einfluss von Standortsfaktoren
auf die Besiedlung des bayerischen Hochgebirges durch
Waldameisen (Formica spp.) (Hymen., Formicidae). Anz.
Schadlingskunde, Pflanzenschutz, Umweltschutz 71,
105–109.
Weber NA, 1935. The biology of the thatching ant For-
mica obscuripes Forel in North Dakota. Ecol. Monog. 5,
165–206.
Wheeler GC, Wheeler JN, 1986. The ants of Nevada. Nat
Hist Museum Los Angeles County, Los Angeles, CA.
Red wood ants in Yellowstone National Park A. C. Risch, M. F. Jurgensen, A. J. Storer, M. D. Hyslop and M. Schutz
336J. Appl. Entomol. 132 (2008) 326–336 ª 2008 The Authors
Journal compilation ª 2008 Blackwell Verlag, Berlin
Copyright © 2022 FDOKUMEN