
legislative mandates guiding the greater yellowstone network

q 2006 The Paleontological Society. All rights reserved. 0094-8373/06/3201-0003/$1.00
Paleobiology, 32(1), 2006, pp. 38–54
Late Quaternary rapid morphological evolution of an endemicdiatom in Yellowstone Lake, Wyoming
Edward C. Theriot, Sherilyn C. Fritz, Cathy Whitlock, and Daniel J. Conley
Abstract.—The centric diatom Stephanodiscus yellowstonensis Theriot and Stoermer is endemic to Yel-lowstone Lake, where it can be an important component of the summer phytoplankton assemblage.Its close relative, Stephanodiscus niagarae Ehrenberg, is abundant in nearby lakes and regional res-ervoirs. We used the stratigraphic record of Yellowstone Lake to investigate the evolution of S. nia-garae to S. yellowstonensis and to describe the limnologic and climatic conditions associated with itsevolution. A dramatic morphological shift took place between about 13.7 and 10.0 Ka, but mor-phology remained stable from 10 Ka to the present. Coincident with morphological change in theS. niagarae/S. yellowstonensis complex were changes in the diatom species assemblage, biogenic sil-ica concentrations, sediment lithology, and regional vegetation. These changes suggest an envi-ronment that experienced progressive warming following the retreat of continental glaciers. Wecould not identify a specific selective factor driving evolution. Nevertheless, nonrandom morpho-logical evolution strongly associated with continuous environmental change suggests that direc-tional selection is a reasonable hypothesis to account for evolution of S. yellowstonensis. Protists arepresumed to evolve gradually after speciation events because of large population size, high dis-persal capacity, and low reproductive barriers. However, published diatom examples and the evo-lution of S. yellowstonensis suggest that it is premature to generalize about rates of evolution in pro-tists, or at least to include diatoms in this generalization.
Edward C. Theriot. Texas Memorial Museum, 2400 Trinity Street, University of Texas, Austin, Texas78705. E-mail: [email protected]
Sherilyn C. Fritz. Department of Geosciences and School of Biological Sciences, University of Nebraska,Lincoln, Nebraska 68588-0340
Cathy Whitlock. Department of Earth Sciences, Montana State University, Bozeman, Montana 59715Daniel J. Conley. Department of Marine Ecology, National Environmental Research Institute, Post Office
Box 358, DK-4000 Roskilde, Denmark
Accepted: 15 June 2005
Introduction
Benton and Pearson (2001) reviewed avail-able records of post-speciation anagenesis inprotists and concluded that the usual tempo ofsuch evolutionary change is gradual and slowrelative to other kinds of organism. They alsonoted that, while protistan evolutionary tem-po parallels environmental change rate, in-trinsic biological properties of protists limittheir evolutionary rates. We present a newcase from the fossil record consistent withBenton and Pearson’s (2001) generalizationthat protistan evolutionary rates parallel thepace of environmental change and that stasismirrors environmental stability. However, ourstudy and the literature also suggest that di-atoms, at least, can evolve at much higher ratesthan are evidenced in other protistan studies.
Stephanodiscus yellowstonensis Theriot andStoermer was described from the sediments ofYellowstone Lake, Wyoming (Theriot and
Stoermer 1984a). Distributional studies, a par-tial stratigraphic record from the YellowstoneLake basin, and cladistic analysis each predictthat S. yellowstonensis evolved from a diatomwith S. niagarae Ehrenberg morphology (Ther-iot 1992). The latter is known from lakesacross temperate North America, from sedi-ments along the margin of Yellowstone Lakedeposited during an early Holocene lake high-stand, and from fossil deposits several millionyears old, but is absent from modern Yellow-stone Lake. Stephanodiscus niagarae is also ple-siomorphic with respect to S. yellowstonensisand several other species in an unresolved po-lytomy (Theriot 1992). This paper describes acore of Yellowstone Lake sediments, support-ing the hypothesis that S. yellowstonensisevolved from S. niagarae and finds that periodsof rapid morphological change and stasis arecorrelated with periods of rapid environmen-tal change and relative environmental stabili-ty, respectively.

39RAPID DIATOM MORPHOLOGICAL EVOLUTION
Environmental Setting
Yellowstone Lake is a large (340 km2) lakeon the Central Plateau of Yellowstone NationalPark, Wyoming (Fig. 1). The main body of thelake basin straddles the eastern margin of acaldera formed during late-Cenozoic volca-nism, but the southern part of the lake lies out-side the caldera (Locke and Meyer 1994; Goodand Pierce 1996). A large ice cap covered theentire Yellowstone Plateau during the Pine-dale Glaciation and retreated from the regionabout 14 Ka (Porter et al. 1983). The modernvegetation surrounding the lake is lodgepolepine (Pinus contortus) forest on infertile rhyo-litic soils, with Engelmann spruce (Picea en-gelmannii), subalpine fir (Abies lasiocarpa), andwhite bark pine (Pinus albicaulis) growing onthe andesitic soils of the east and south mar-gins (Despain 1990). The lake consists of alarge central basin, moderately deep (maxi-mum depth 5 107 m), with three shallowerarms. The lake usually freezes sometime be-tween early December and mid-January, withice-off between early May and early June. Nu-trient concentrations and diatom assemblagestrack seasonal changes in predictable ways,producing diatom blooms with species com-position and abundances reflecting local cli-mate change (Kilham et al. 1996; Theriot et al.1997; Interlandi et al. 1999).
Methods
An 8.55 m long core (FD2-92) was retrievedfrom the central basin of Yellowstone Lake(Fig. 1) in 82 m of water with a Kullenberg cor-ing system (Kelts et al. 1986). The coring lo-cation was a topographic high with horizontalstratigraphic units distinctly evident in seis-mic reflection profiles (Tiller 1995). In thefield, the core was cut into sections each ap-proximately one meter long. The sections werebrought to the University of Minnesota Lim-nological Research Center (LRC) and stored at48C until processed and described. Each sec-tion was split lengthwise, one half-section ar-chived, and the other half-section used foranalysis.
Samples for diatom community analysiswere taken at 16 cm intervals. 1–2 cm3 weretaken at each sample level with a scoop ap-
proximately 2 mm wide. Samples werecleaned in cold hydrogen peroxide, and oxi-dation by-products removed in a series of dis-tilled water rinses. A weighed subsample ofcleaned sediment was settled onto cleanedcoverslips in Battarbee trays (Battarbee 1973).Coverslips were mounted with Naphrax. Atleast 500 diatom valves were identified persample. Additional transects were scanneduntil 50 valves of the relatively rare (typically,5% relative abundance) S. niagarae-S. yellow-stonensis complex were counted or the entirecover slip examined.
Valve diameter, the number of costae pervalve, and the number of spines per valve ofspecimens in the S. niagarae-S. yellowstonensiscomplex first were recorded from 16 cm inter-vals. Additional slides were studied fromsamples at 4–8 cm intervals in core sectionsshowing the greatest evolutionary change.Abundance permitting, 50 specimens weremeasured at each level. In cases where therewere fewer than 50 measurable specimens ona slide, additional slides were made from theremaining sample until either 50 specimenswere obtained or the entire sample at a levelwas consumed. We compared specimens fromYellowstone Lake with modern and fossil S.niagarae from a wide variety of habitats, in-cluding specimens collected at Lewis Lakeand Heart Lake in Yellowstone Park, the twolakes closest to Yellowstone Lake having S.niagarae. We also included two other relatedspecies (S. reimerii Theriot and Stoermer, andS. superiorensis Stoermer and Theriot: Theriot1984; Theriot and Stoermer 1981, 1984a,b,c).For all samples, morphometric data were gath-ered from any valve identified as a member ofthe easily diagnosed S. niagarae complex(Theriot et al. 1988; Theriot 1992), avoidingbias or error of identification in morphometricanalysis. In addition to quantitative observa-tions, as many as 250 valves, abundance per-mitting, were qualitatively examined on eachslide.
The core was also sampled at 4 cm intervalsfor the measurement of biogenic silica. Bio-genic silica was measured by using a time-course extraction with 1% Na2CO3 in a modi-fication of the technique of DeMaster (1981).
Samples ranging in size from 0.5 to 1.0 cm3

40 EDWARD C. THERIOT ET AL.
FIGURE 1. Map of study lakes in Grand Teton and Yellowstone National Parks indicating coring site FD2-92.
were processed at regular intervals for pollenanalysis (Faegri et al. 1989). Residues weremounted in silicone oil and examined at mag-nifications of 400 and 10003. Identification ofpollen types was based on published atlasesand the pollen reference collection at MontanaState University. Between 300 and 500 terres-trial pollen grains were identified and talliedfor each stratigraphic level. The source of theHaploxylon-type Pinus pollen is probably P.albicaulis, and the Diploxylon-type pollen wasattributed to P. contortus. The pollen ratio wasbased on a denominator of total terrestrialpollen grains.
ResultsSediment Lithology and Chronology. There is
little evidence for sediment reworking or other
processes that might lead to nonconformity.Basal sediments are clay-rich muds, gradinginto diatom- and sulfide-rich laminated mudsabove 7.80 m core depth (Tiller 1995). Above7.30 m core depth, the sediments are diatom-rich and laminated. The laminations are gen-erally fine (10–20 laminations per cm) below6.30 m core depth and coarser above (5–10laminations per cm). The uppermost 4.90 m ofthe diatomaceous muds are banded or weaklylaminated.
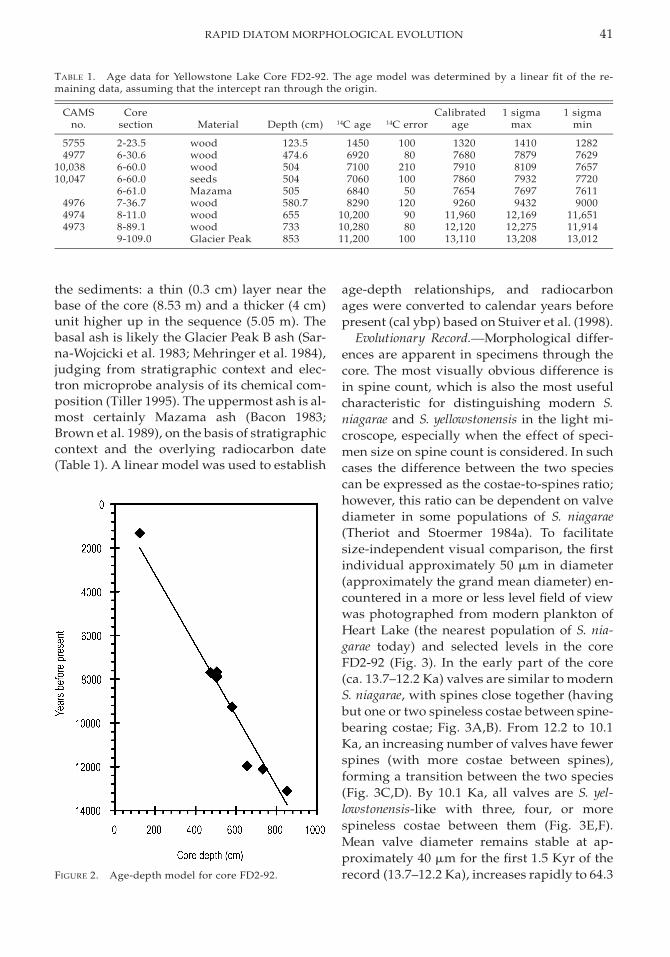
Estimates of a chronology for the Yellow-stone Lake sediments were constructed on thebasis of age-depth relationships derived fromAMS radiocarbon dates on terrestrial macro-fossils and known ages of two volcanic ashes(Table 1, Fig. 2). Two ash units are visible in
41RAPID DIATOM MORPHOLOGICAL EVOLUTION
TABLE 1. Age data for Yellowstone Lake Core FD2-92. The age model was determined by a linear fit of the re-maining data, assuming that the intercept ran through the origin.
CAMSno.
Coresection Material Depth (cm) 14C age 14C error
Calibratedage
1 sigmamax
1 sigmamin
5755 2-23.5 wood 123.5 1450 100 1320 1410 12824977 6-30.6 wood 474.6 6920 80 7680 7879 7629
10,038 6-60.0 wood 504 7100 210 7910 8109 765710,047 6-60.0 seeds 504 7060 100 7860 7932 7720
6-61.0 Mazama 505 6840 50 7654 7697 76114976 7-36.7 wood 580.7 8290 120 9260 9432 90004974 8-11.0 wood 655 10,200 90 11,960 12,169 11,6514973 8-89.1 wood 733 10,280 80 12,120 12,275 11,914
9-109.0 Glacier Peak 853 11,200 100 13,110 13,208 13,012
FIGURE 2. Age-depth model for core FD2-92.
the sediments: a thin (0.3 cm) layer near thebase of the core (8.53 m) and a thicker (4 cm)unit higher up in the sequence (5.05 m). Thebasal ash is likely the Glacier Peak B ash (Sar-na-Wojcicki et al. 1983; Mehringer et al. 1984),judging from stratigraphic context and elec-tron microprobe analysis of its chemical com-position (Tiller 1995). The uppermost ash is al-most certainly Mazama ash (Bacon 1983;Brown et al. 1989), on the basis of stratigraphiccontext and the overlying radiocarbon date(Table 1). A linear model was used to establish
age-depth relationships, and radiocarbonages were converted to calendar years beforepresent (cal ybp) based on Stuiver et al. (1998).
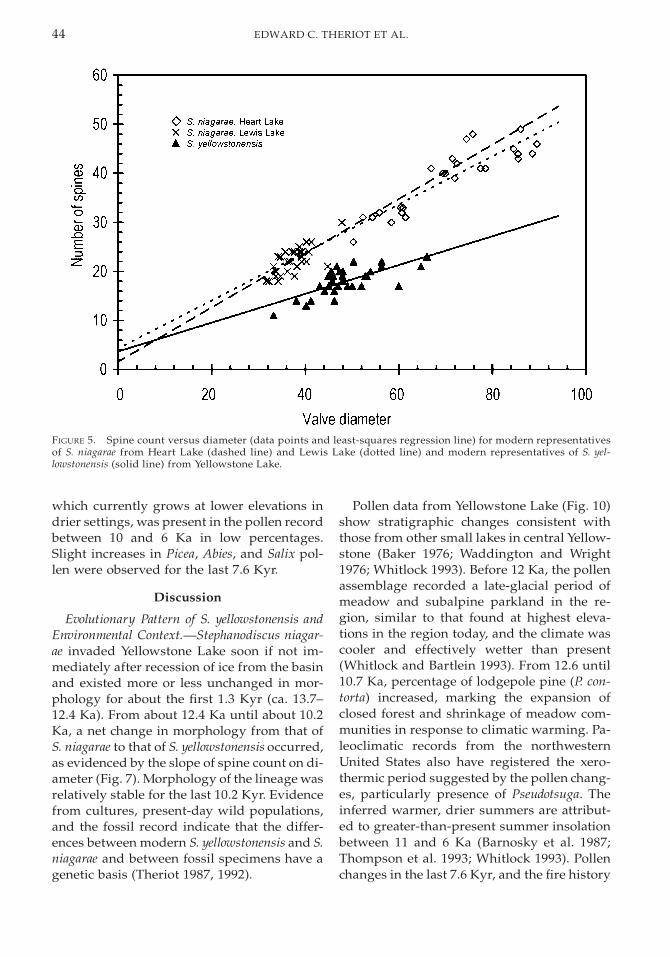
Evolutionary Record. Morphological differ-ences are apparent in specimens through thecore. The most visually obvious difference isin spine count, which is also the most usefulcharacteristic for distinguishing modern S.niagarae and S. yellowstonensis in the light mi-croscope, especially when the effect of speci-men size on spine count is considered. In suchcases the difference between the two speciescan be expressed as the costae-to-spines ratio;however, this ratio can be dependent on valvediameter in some populations of S. niagarae(Theriot and Stoermer 1984a). To facilitatesize-independent visual comparison, the firstindividual approximately 50 mm in diameter(approximately the grand mean diameter) en-countered in a more or less level field of viewwas photographed from modern plankton ofHeart Lake (the nearest population of S. nia-garae today) and selected levels in the coreFD2-92 (Fig. 3). In the early part of the core(ca. 13.7–12.2 Ka) valves are similar to modernS. niagarae, with spines close together (havingbut one or two spineless costae between spine-bearing costae; Fig. 3A,B). From 12.2 to 10.1Ka, an increasing number of valves have fewerspines (with more costae between spines),forming a transition between the two species(Fig. 3C,D). By 10.1 Ka, all valves are S. yel-lowstonensis-like with three, four, or morespineless costae between them (Fig. 3E,F).Mean valve diameter remains stable at ap-proximately 40 mm for the first 1.5 Kyr of therecord (13.7–12.2 Ka), increases rapidly to 64.3

42 EDWARD C. THERIOT ET AL.
FIGURE 3. Light micrographs of modern S. niagarae and S. yellowstonensis and of transitional forms. A, Modern S.niagarae from plankton of Heart Lake (ECT 3160). B, S. niagarae morphology from core FD2-92 section 9, depth 108cm (13,728 Ka). Specimen fd292c9s108otu1b.jpg. C, Transitional morphology (one or more regions of three spinelesscostae) from core FD2-92 section 8, depth 84 cm (11,730 Ka). Specimen fd292c8s84otu1a.jpg. D, Transitional mor-phology (one or more regions of three spineless costae) from core FD2-92 section 8, depth 52 cm (11,215 Ka). Spec-imen fd292c8s52otu1.jpg. E, S. yellowstonensis morphology (a majority of segments with three or more spinelesscostae) core FD2-92 section 4, depth 20 cm (4254 Ka). Specimen fd292c4s20otu1.jpg. F, Modern S. yellowstonensisfrom isotype slide (ECT 56) from surface sediments of Yellowstone Lake, Wyoming. Scale bar, 10 mm for all.
mm by 10.1 Ka, declines to 41.3 mm by 3.4 Ka,and finally increases slowly to reach 49.3 mmin the surface sediments (Fig. 4). However,mean spine count peaks much earlier thanmean diameter, at 11.2 Ka, and decreases to alocal minimum when diameter is at its maxi-mum (10.1 Ka).
Spine count is significantly dependent ondiameter in each measured population of S.niagarae and S. yellowstonensis (e.g., Fig. 5) andin each sample from FD2-92 (typically at p K
0.001 but never worse than p 5 0.01). This cor-relation pattern is variable across lake and fos-sil assemblages of S. niagarae, and is different

43RAPID DIATOM MORPHOLOGICAL EVOLUTION
FIGURE 4. Sample means and sample standard devia-tions for diameter, costa count, and spine count for spec-imens from core FD2-92 against time.
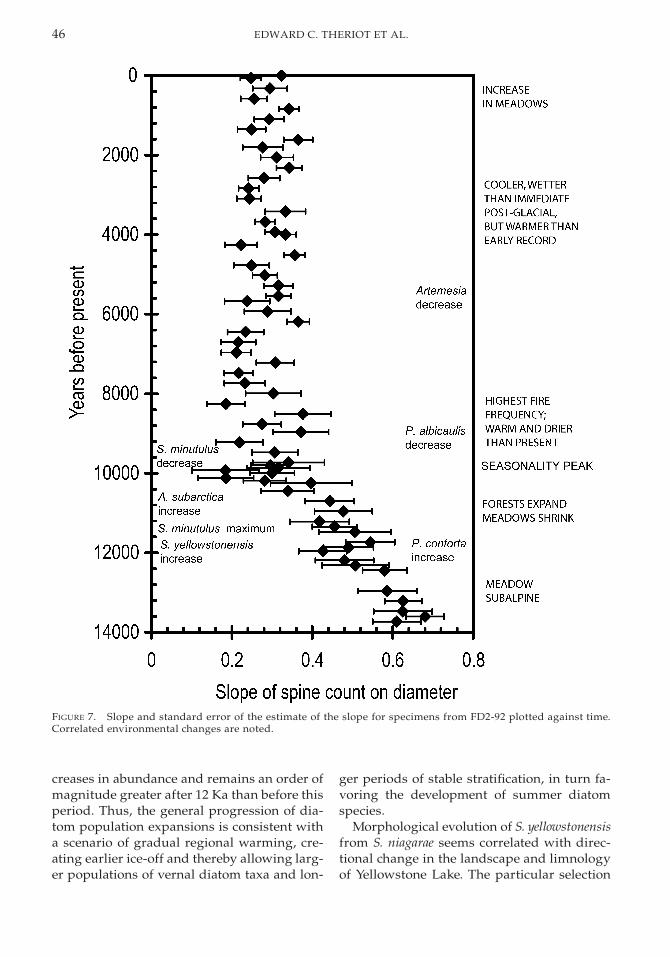
between S. yellowstonensis and S. niagarae. Thelatter has a much lower slope than the former(Fig. 6). The correlation pattern is also variablein the core. Analysis of covariance (ANCOVA)reveals a significant interaction between di-ameter and core-level on spine count (p ,0.0001). Slope of spine count on diameter de-creases more or less steadily from about 0.68to about 0.30, from 13.2 Ka to 10 Ka (Fig. 7).After that, slope fluctuates around 0.30. Stan-dard error of the slope is generally stable, per-haps slightly increasing, during the period ofrapid directional change, and then decreasessteadily until about 3.4 Ka when it was about0.03, or half that of the earliest samples.
Qualitatively, there was no obvious bimo-dality in the distribution of S. niagarae and S.yellowstonensis morphotypes anywhere in thecore, including the transitional phase. We usedLilliefors test to test for normality of distri-bution of the costae-to-spines ratio using the.Because of the use of ratios and because theratio itself can be dependent on diameter, re-
sults should be interpreted with caution, per-haps only as a heuristic indicator of near nor-mal distribution or as a numerical expressionof our qualitative conclusion about the lack ofbimodality. Only nine samples showed signif-icantly non-normal distribution for costae-to-spines ratio. Those samples occurred sporad-ically throughout the column (9797 Ka, 7219Ka, 6187 Ka, 5672 Ka, 4254 Ka, 3996 Ka, 2836Ka, 1804 Ka and 1611 Ka). Most importantly,all samples older than 9800 Ka, during theevolution of S. yellowstonensis morphology andat the completion of S. yellowstonensis evolu-tion, were normally distributed for costae-to-spines ratio.
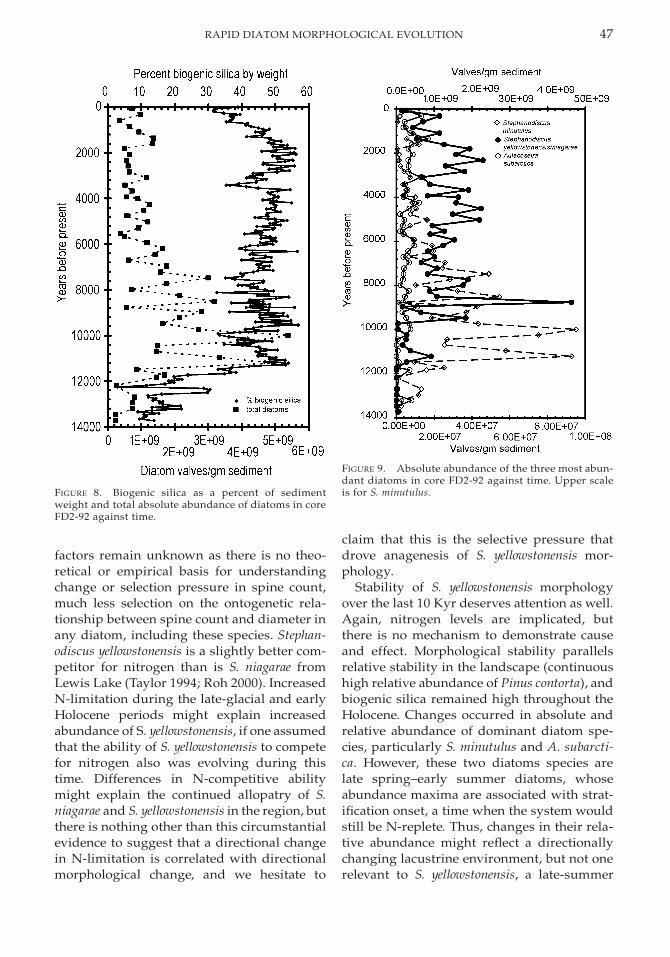
Environmental Record. Total diatom con-centrations are relatively low in the basal sed-iments of the core, increase from the base ofthe core to a high at 11.2 Ka, fluctuate forabout 1 Kyr, reach a maximum at 10 Ka, anddecline irregularly after that (Fig. 8). Biogenicsilica as a percent of sediment dry weighttracks the increase in overall diatom abun-dance in the early part of the core, but after 10Ka it remains high throughout the core in-stead of tracking the decrease in diatom con-centrations. The mismatch between total dia-tom count and biogenic silica after 10 Ka isprobably due to a relative increase in abun-dance in two larger and more heavily silicifieddiatoms, Aulacoseira subarctica (O. Muller)Haworth and S. yellowstonensis, over one smalland lightly silicified species, S. minutulus(Kutzing) Round starting at about 10–11 Ka(Fig. 9). Of special interest to this study, S. yel-lowstonensis occurs in very low percentages inthe lower levels of the core, during the firsthalf of its transition from S. niagarae morphol-ogy. Halfway through this transition, it be-comes dramatically more abundant in thecore, and its lowest abundance after 11.2 Ka isan order of magnitude greater than the high-est pre-11.7 Ka levels (Fig.9).
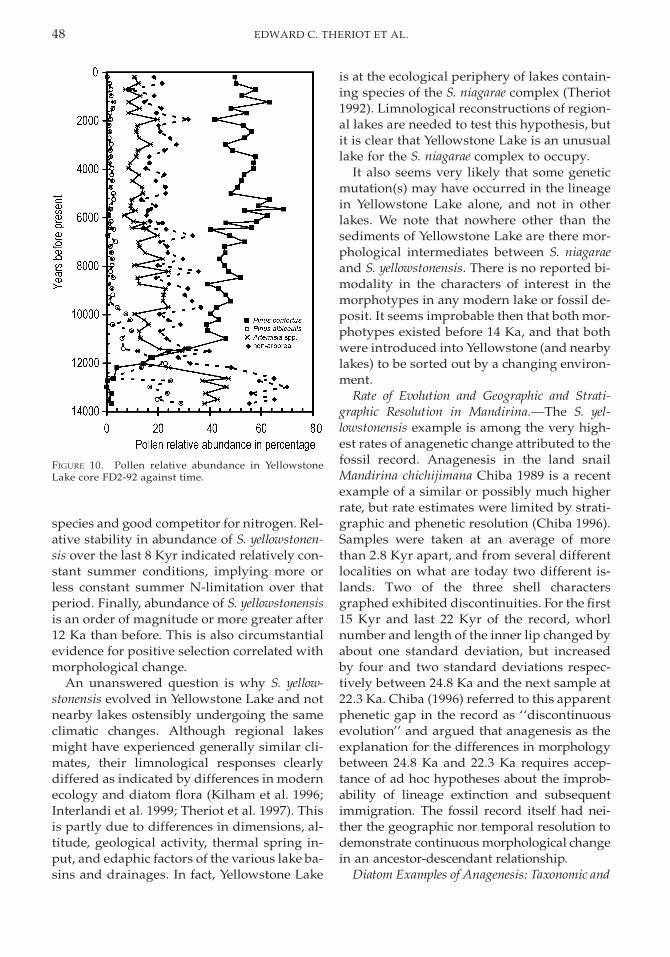
Four pollen types dominated the Yellow-stone Lake pollen record (Fig. 10). From 12.7to 10.7 Ka, Pinus contortus exchanged domi-nance with P. albicaulis and with Artemisia andother non-arboreal types (other Asteraceae,Poaceae and Chenopodiaceae). Abies pollen,Juniperus-type pollen, and low amounts of Pi-cea pollen also were present. Pseudotsuga,
44 EDWARD C. THERIOT ET AL.
FIGURE 5. Spine count versus diameter (data points and least-squares regression line) for modern representativesof S. niagarae from Heart Lake (dashed line) and Lewis Lake (dotted line) and modern representatives of S. yel-lowstonensis (solid line) from Yellowstone Lake.
which currently grows at lower elevations indrier settings, was present in the pollen recordbetween 10 and 6 Ka in low percentages.Slight increases in Picea, Abies, and Salix pol-len were observed for the last 7.6 Kyr.
Discussion
Evolutionary Pattern of S. yellowstonensis andEnvironmental Context. Stephanodiscus niagar-ae invaded Yellowstone Lake soon if not im-mediately after recession of ice from the basinand existed more or less unchanged in mor-phology for about the first 1.3 Kyr (ca. 13.7–12.4 Ka). From about 12.4 Ka until about 10.2Ka, a net change in morphology from that ofS. niagarae to that of S. yellowstonensis occurred,as evidenced by the slope of spine count on di-ameter (Fig. 7). Morphology of the lineage wasrelatively stable for the last 10.2 Kyr. Evidencefrom cultures, present-day wild populations,and the fossil record indicate that the differ-ences between modern S. yellowstonensis and S.niagarae and between fossil specimens have agenetic basis (Theriot 1987, 1992).
Pollen data from Yellowstone Lake (Fig. 10)show stratigraphic changes consistent withthose from other small lakes in central Yellow-stone (Baker 1976; Waddington and Wright1976; Whitlock 1993). Before 12 Ka, the pollenassemblage recorded a late-glacial period ofmeadow and subalpine parkland in the re-gion, similar to that found at highest eleva-tions in the region today, and the climate wascooler and effectively wetter than present(Whitlock and Bartlein 1993). From 12.6 until10.7 Ka, percentage of lodgepole pine (P. con-torta) increased, marking the expansion ofclosed forest and shrinkage of meadow com-munities in response to climatic warming. Pa-leoclimatic records from the northwesternUnited States also have registered the xero-thermic period suggested by the pollen chang-es, particularly presence of Pseudotsuga. Theinferred warmer, drier summers are attribut-ed to greater-than-present summer insolationbetween 11 and 6 Ka (Barnosky et al. 1987;Thompson et al. 1993; Whitlock 1993). Pollenchanges in the last 7.6 Kyr, and the fire history

45RAPID DIATOM MORPHOLOGICAL EVOLUTION
FIGURE 6. Slope and standard error of the estimate ofslope for assemblages of S. niagarae, S. reimerii, and S. yel-lowstonensis from across North America.
reconstruction from nearby Cygnet Lake(Millspaugh et al. 2000), indicate that sum-mers were cooler and wetter than between10.7 and 7.6 Ka. The slight increase in Salixpollen after 7 Ka indicates expansion of ripar-ian communities along the margin of the lake,perhaps also suggesting wetter summers.Overall, terrestrial vegetation changes of thelast 10 Ka were on a much smaller scale thanthose between 13.7 Ka and 10 Ka.
Long-term climate change would have pre-dictable effects on limnological and diatom as-semblage change in Yellowstone Lake. Factorslinking weather, limnological conditions, anddiatom abundance are qualitatively wellenough understood to infer particular limno-logical changes that may have occurred dur-ing the evolution of S. yellowstonensis. Seasonaland annual changes in the diatom assemblagetoday are driven by changes in external nutri-ent loadings and lake thermal structure. Inturn, nutrient loadings and thermal structure
are driven by local weather, which ultimatelydepend on changes to regional climate sys-tems (Whitlock 1993; Whitlock and Bartlein1993; Kilham et al. 1996; Theriot et al. 1997;Interlandi et al. 1999). Modern diatom abun-dance maxima are typically low under the iceand increase to their annual maxima at the on-set of stratification (Interlandi et al. 1999). Ear-ly Yellowstone Lake was subject to summersmarkedly cooler and wetter than present day,reducing the period of stratification and per-haps even resulting in some summers of con-tinuous circulation. This would have prevent-ed high abundances of diatoms through lightlimitation. In contrast, increasingly warm con-ditions would have increased water columnstability and allowed spring–early summerdiatom blooms to occur. Winter and summerdrought conditions probably also increasedlate summer N-limitation in Yellowstone Lake,thus altering the interspecific competitive re-gime in the lake (Kilham et al. 1996; Theriot etal. 1997; Interlandi et al. 1999) and producingchanges in the absolute and relative abun-dances of diatom species observed in the core.
Diatom changes over the long term followexpectations based on observations of season-al variation. The second highest peak of S.minutulus and the long-term minimum of Au-lacoseira subarctica both occur at about 11.2 Ka,at a time when overall diatom assemblagesalso showed a dramatic increase (as indicatedby total diatom counts and biogenic silica con-centrations), indicating a period when annualstratification was increasingly frequent andbetter developed. This coincides with the on-set of strong seasonal amplification of insola-tion in the region (Whitlock 1993; Whitlockand Bartlein 1993), probably driving more in-tense seasonal stratification. Stephanodiscusminutulus blooms during the vernal mixingperiod, though the bloom often extends pastthe onset of summer stratification (Kilham etal. 1996). Aulacoseira subarctica expands duringtimes of water column mixing in both springand fall and persists at the thermocline duringsummer months (Interlandi et al. 1999). Incontrast, S. yellowstonensis is a summer diatomand develops larger populations during yearsof extended stratification. In the core record,the S. yellowstonensis-S. niagarae lineage in-
46 EDWARD C. THERIOT ET AL.
FIGURE 7. Slope and standard error of the estimate of the slope for specimens from FD2-92 plotted against time.Correlated environmental changes are noted.
creases in abundance and remains an order ofmagnitude greater after 12 Ka than before thisperiod. Thus, the general progression of dia-tom population expansions is consistent witha scenario of gradual regional warming, cre-ating earlier ice-off and thereby allowing larg-er populations of vernal diatom taxa and lon-
ger periods of stable stratification, in turn fa-voring the development of summer diatomspecies.
Morphological evolution of S. yellowstonensisfrom S. niagarae seems correlated with direc-tional change in the landscape and limnologyof Yellowstone Lake. The particular selection
47RAPID DIATOM MORPHOLOGICAL EVOLUTION
FIGURE 8. Biogenic silica as a percent of sedimentweight and total absolute abundance of diatoms in coreFD2-92 against time.
FIGURE 9. Absolute abundance of the three most abun-dant diatoms in core FD2-92 against time. Upper scaleis for S. minutulus.
factors remain unknown as there is no theo-retical or empirical basis for understandingchange or selection pressure in spine count,much less selection on the ontogenetic rela-tionship between spine count and diameter inany diatom, including these species. Stephan-odiscus yellowstonensis is a slightly better com-petitor for nitrogen than is S. niagarae fromLewis Lake (Taylor 1994; Roh 2000). IncreasedN-limitation during the late-glacial and earlyHolocene periods might explain increasedabundance of S. yellowstonensis, if one assumedthat the ability of S. yellowstonensis to competefor nitrogen also was evolving during thistime. Differences in N-competitive abilitymight explain the continued allopatry of S.niagarae and S. yellowstonensis in the region, butthere is nothing other than this circumstantialevidence to suggest that a directional changein N-limitation is correlated with directionalmorphological change, and we hesitate to
claim that this is the selective pressure thatdrove anagenesis of S. yellowstonensis mor-phology.
Stability of S. yellowstonensis morphologyover the last 10 Kyr deserves attention as well.Again, nitrogen levels are implicated, butthere is no mechanism to demonstrate causeand effect. Morphological stability parallelsrelative stability in the landscape (continuoushigh relative abundance of Pinus contorta), andbiogenic silica remained high throughout theHolocene. Changes occurred in absolute andrelative abundance of dominant diatom spe-cies, particularly S. minutulus and A. subarcti-ca. However, these two diatoms species arelate spring–early summer diatoms, whoseabundance maxima are associated with strat-ification onset, a time when the system wouldstill be N-replete. Thus, changes in their rela-tive abundance might reflect a directionallychanging lacustrine environment, but not onerelevant to S. yellowstonensis, a late-summer
48 EDWARD C. THERIOT ET AL.
FIGURE 10. Pollen relative abundance in YellowstoneLake core FD2-92 against time.
species and good competitor for nitrogen. Rel-ative stability in abundance of S. yellowstonen-sis over the last 8 Kyr indicated relatively con-stant summer conditions, implying more orless constant summer N-limitation over thatperiod. Finally, abundance of S. yellowstonensisis an order of magnitude or more greater after12 Ka than before. This is also circumstantialevidence for positive selection correlated withmorphological change.
An unanswered question is why S. yellow-stonensis evolved in Yellowstone Lake and notnearby lakes ostensibly undergoing the sameclimatic changes. Although regional lakesmight have experienced generally similar cli-mates, their limnological responses clearlydiffered as indicated by differences in modernecology and diatom flora (Kilham et al. 1996;Interlandi et al. 1999; Theriot et al. 1997). Thisis partly due to differences in dimensions, al-titude, geological activity, thermal spring in-put, and edaphic factors of the various lake ba-sins and drainages. In fact, Yellowstone Lake
is at the ecological periphery of lakes contain-ing species of the S. niagarae complex (Theriot1992). Limnological reconstructions of region-al lakes are needed to test this hypothesis, butit is clear that Yellowstone Lake is an unusuallake for the S. niagarae complex to occupy.
It also seems very likely that some geneticmutation(s) may have occurred in the lineagein Yellowstone Lake alone, and not in otherlakes. We note that nowhere other than thesediments of Yellowstone Lake are there mor-phological intermediates between S. niagaraeand S. yellowstonensis. There is no reported bi-modality in the characters of interest in themorphotypes in any modern lake or fossil de-posit. It seems improbable then that both mor-photypes existed before 14 Ka, and that bothwere introduced into Yellowstone (and nearbylakes) to be sorted out by a changing environ-ment.
Rate of Evolution and Geographic and Strati-graphic Resolution in Mandirina. The S. yel-lowstonensis example is among the very high-est rates of anagenetic change attributed to thefossil record. Anagenesis in the land snailMandirina chichijimana Chiba 1989 is a recentexample of a similar or possibly much higherrate, but rate estimates were limited by strati-graphic and phenetic resolution (Chiba 1996).Samples were taken at an average of morethan 2.8 Kyr apart, and from several differentlocalities on what are today two different is-lands. Two of the three shell charactersgraphed exhibited discontinuities. For the first15 Kyr and last 22 Kyr of the record, whorlnumber and length of the inner lip changed byabout one standard deviation, but increasedby four and two standard deviations respec-tively between 24.8 Ka and the next sample at22.3 Ka. Chiba (1996) referred to this apparentphenetic gap in the record as ‘‘discontinuousevolution’’ and argued that anagenesis as theexplanation for the differences in morphologybetween 24.8 Ka and 22.3 Ka requires accep-tance of ad hoc hypotheses about the improb-ability of lineage extinction and subsequentimmigration. The fossil record itself had nei-ther the geographic nor temporal resolution todemonstrate continuous morphological changein an ancestor-descendant relationship.
Diatom Examples of Anagenesis: Taxonomic and

49RAPID DIATOM MORPHOLOGICAL EVOLUTION
Stratigraphic Resolution. A few examples ofdiatom anagenesis with demonstrated mor-phological continuity exist, but none were asfast as that of S. yellowstonensis. Two did notinvolve lineage splitting, and so were not rel-evant to discussion of post-cladogenetic ana-genesis (Fenster et al. 1989; Koizumi and Yan-agisawa 1990). It is not clear if a third, thetransformation of Rhizosolenia bergonii Pera-gallo into R. bergonii var. robusta Burckle andTrainer, involved lineage splitting or not(Burckle and Trainer 1979).
In a case involving cladogenesis, Rhizosolen-ia praebergonii Mukhina apparently evolvedfrom R. bergonii Peragallo in the equatorial Pa-cific Ocean (Sorhannus et al. 1988). Bentonand Pearson (2001) made a particular exampleof this case, claiming it to be an example ofpost-speciation gradualistic change. Althoughit did take 50–350 Kyr for these two species tobecome statistically separate for measuredcharacters (in terms of no overlap of standarddeviation [Sorhannus et al. 1988]), there is ev-idence that the lineages split earlier and per-haps rather rapidly. Published illustrationsand descriptions suggest that R. bergoniivalves are finely areolate with strongly flaredvalves whereas R. praebergonii valves arecoarsely areolate with roughly parallel sides(cf. Cupp 1943; Schrader 1973, 1974; Sund-strom 1986; Sorhannus 1990a). Sorhannus(personal communication) confirmed that thespecies were initially distinguished by thesefeatures. Evidently, initial separation of theancestor and descendant species is not entire-ly resolved in the equatorial Pacific fossil re-cord.
Related investigations more strongly sug-gest stratophenetic gaps in ancestor-descen-dant relationships. In one, Sorhannus (1990b)reported that forms identical to R. praebergoniisuddenly appeared in the Indian Ocean200,000 years earlier than such forms ap-peared in the equatorial Pacific, or before thetime that R. praebergonii and R. bergonii weremorphometrically separate in the equatorialPacific Ocean. In the Indian Ocean, there wereno intermediates between R bergonii and R.praebergonii as there had been in the equatorialPacific example. Benton and Pearson (2001)cited Sorhannus (1990b) and concluded only
that R. praebergonii later invaded the IndianOcean. However, Sorhannus (1990b) had of-fered alternative hypotheses and concludedthat it would be difficult to expect deep oceansediments to provide the spatial and temporalresolution necessary to explain the large mor-phological discontinuity observed. In a sec-ond example of morphological discontinuityin the marine sedimentary record of diatomspeciation, Sorhannus et al. (1991) hypothe-sized that R. sigmoida Sorhannus evolved fromR. bergonii, this time in the eastern equatorialPacific Ocean. However, these two specieswere already distinct qualitatively and quan-titatively at the earliest occurrence of R. sig-moida, and they diverged little afterwards(Sorhannus et al. 1991).
We note that all of these species do appearto be closely related, although the evidence isrevealed through informal cladistic analysis.All bear distinctive apical processes best de-scribed as ‘‘flute-shaped.’’ We know of no oth-er species of Rhizosolenia bearing such an api-cal process (Sundstrom 1986; Cupp 1943;Burckle and Trainer 1974; Schrader 1974; Sor-hannus 1990a,b; Sorhannus et al. 1988, 1991).This would suggest a synapomorphy for R.bergonii, R. praebergonii (both the Pacific andIndian Ocean lineages), R. praebergonii v. ro-busta, and R. sigmoida. Recent advances intransforming morphometric variables intocladistic characters may make cladistic anal-ysis of these taxa possible as it has for otherdiatoms (Edgar and Theriot 2004). Perhapscladistic analysis can yield additional supportfor the supposition of ancestor-descendant re-lationships in Rhizosolenia as it did for the re-lationship between S. niagarae and S. yellow-stonensis (Theriot 1992).
Mechanisms Controlling Rates of Protistan Evo-lution. Benton and Pearson (2001) suggestedthat intrinsic properties are responsible forslow rates of evolution, arguing that marineprotists are characterized by large popula-tions with low dispersal barriers and low bar-riers to reproduction. Similarly, Pearson et al.(1997) connected physical scale of branchingevents to large population size and broad dis-tribution, claiming ‘‘branching events tend tooccur in large populations over very large ar-eas, rather than in small geographical iso-

50 EDWARD C. THERIOT ET AL.
lates.’’ However, population size affects evo-lutionary rate differently under different en-vironmental conditions, and we know littleabout the reproductive behavior of protists.Population size interacts with selection (orlack thereof) and environmental constancy (orlack thereof) in different ways. In a changingenvironment, large populations may respondproportionately faster to directional selectionthan small populations because of increasedgenetic variance (Crow and Kimura 1970; Fal-coner 1989; Lande 1986). Directional selectionwas invoked as a causal mechanism by Sor-hannus et al. (1988) as well as in several stud-ies of foraminiferan and radiolarian evolution(e.g., Kellogg and Hays 1975; Malmgren et al.1983; Lazarus et al. 1995) and so a simpler al-ternative hypothesis is that rate of evolution inthese examples is limited by rate of environ-mental change, not by population size. Whenthe environment is stable, large populationsize is not a complete obstacle to rapid changedue to drift from one adaptive peak to anotherin a character under selection (Lande 1985).Protists may or may not have behavioral bar-riers to reproduction, but many different or-ganisms in many different phyla have chemi-cal gamete recognition systems and this haslong been suspected for diatoms (Round et al.1990). It may well be that protists have gameterecognition signals analogous to behavioraldifferences in promoting recognition andelimination of possible mates.
Geographic isolation, speciation pattern,and morphological change in deep sea pro-tists may be more complex and subtle than theliterature on protistan evolutionary mechanismsrecognizes. On the basis of molecular, morpho-logical, and paleontological evidence, de Vargaset al. (2001) argued for relatively rapid and re-cent lineage divergence in the foraminiferanGloborotalia truncatulinoides (d’Orbigny). Alsofinding correlations between genetic divergenceand subtle morphological differentiation in theforaminiferan Orbulina universa d’Orbigny, deVargas et al. (1999) speculated that ‘‘the pat-tern of planktonic foraminiferal speciationmay be similar to that of other high-dispersaltaxa in which gamete interactions, wide-spread genome incompatibility, a complexmating system, or reproductive behavior play
key roles in species formation and mainte-nance.’’ The local evolutionary events report-ed by Sorhannus (1990b) and Sorhannus et al.(1991) may be diatom examples resultingfrom such phenomena, particularly the highlylocalized evolution of R. sigmoida in one smallarea of the eastern Pacific.
One might presume that lacustrine protistswould face even greater barriers to dispersalthan do oceanic protists, but it is not clear justhow much of a barrier they face. Some authorsbelieve that endemism is much more prevalentin diatoms than is presently recognized andthat diatoms cannot easily disperse (Mannand Droop 1996; Kociolek and Spaulding2000). However, Finlay et al. (2002) claimedthat diatoms disperse readily at a scale andrate related to their global abundance. In fact,there is indirect evidence that the physical iso-lation of S. niagarae and S. yellowstonensis is notas complete as their allopatric distributionmight suggest. A diatom with S. niagarae mor-phology obviously invaded Yellowstone Lakeimmediately after glaciers receded, and thereis no reason to expect that some distributionalbarrier has arisen since the time the S. yellows-tonensis morphology began to evolve. Stephan-odiscus niagarae is found from Pleistocene sed-iments around Lago de Isabol, Guatemala, Ji-lin Province, China, and in modern lakes fromAlaska to California to Pennsylvania andacross the Canadian Shield (Theriot 1984;Theriot et al. 1988). It is abundant in numerousreservoirs for which there is no apparent up-stream source. Thus, strong circumstantial ev-idence suggests that S. niagarae has high dis-persal ability and that the evolution of S. yel-lowstonensis is an example of extremely rapidpost-speciation change in the face of at leastmoderate dispersal capability.
A robust generalization about mechanismscontrolling rates of protistan evolution re-quires unbiased examples across a spectrumof possible dispersal conditions and differentorganisms. The examples where the sedimen-tary record has provided evidence for conti-nuity of form over time are few for diatoms,and fewer still for lacustrine diatoms. With theS. yellowstonensis example and bringing nochallenge to Sorhannus et al. (1998) as an ex-ample of sympatric speciation, there are only

51RAPID DIATOM MORPHOLOGICAL EVOLUTION
two records of diatom speciation events com-plete enough (morphometric continuity be-tween ancestor and descendant) to say thatthe fossil record alone, without independentphylogenetic analysis, establishes anagenesisfollowing cladogenesis. There are not manymore examples of strictly anagenetic changefrom one morphospecies to another within alineage. This is a vanishingly small percentageof the 10,000 to more than 1,000,000 diatomsspecies that may exist (Round et al. 1990).Even if the percentage is a bit higher in plank-tonic foraminifera and radiolarians, it is stillvery small. As Pearson et al. (1997) wrote, ‘‘Agreat many cladogenetic events must have oc-curred to generate the biospecies diversity ofmodern mineralized plankton, and that whichis inferred to have been present in the past.However, relatively few studies have beenconducted which clearly demonstrate thesebranching events using morphometric tech-niques.’’ Investigators may need to renew in-terest in a much older question: ‘‘Why aremore evolutionary transitions (particularlythose involving cladogenesis) not found?’’
There are at least five nonexclusive expla-nations for the relative rarity of observationson morphological and stratigraphic continuitybetween lineages from the fossil record: (1)Special creation occurred and so there simplywas no evolution to record in the fossil record.(2) The fossils are common in most sedimen-tary deposits, but no one has bothered tostudy them. (3) Evolution occurred in an en-vironment that did not allow for fossilization.(4) Evolution happened in a very restricted lo-cality that is yet to be found or whose fossilswere subsequently destroyed. (5) Evolutionhappened faster than depositional rates canresolve. Lack of data is not logical evidence oflack of pattern or process, so investigation ofthe first reason is beyond the purview of thisarticle. The second seems unlikely given theremaining interest in the topic, no doubt inpart due to the desire to overwhelm the firstargument with data.
The remaining possibilities are traditionalexplanations for gaps in the fossil record,translating into insufficient spatial and/ortemporal resolution. The explanation usuallygiven for rapid post-cladogenetic anagenesis
is that of speciation through peripheral isola-tion, which can conflate these problems. Ja-blonski (2000) argued that an appropriateprotocol for assessment of the distribution ofstasis and change requires six components, in-cluding a stratigraphic record whose resolv-ing capacity will not limit the inferred maxi-mum rate of evolution, and an independentphylogenetic analysis of species. A phyloge-netic analysis is necessary particularly whenthe stratigraphic record cannot resolve contin-uous change between ancestor and descen-dant. An assessment of apomorphic and ple-siomorphic conditions on the phylogenetictree is critical to understanding ancestor-de-scendant relationships in such cases (Wiley1981; Theriot 1992; Frey 1993). The high re-solving power of Yellowstone Lake sedimentsmight arguably obviate the need for cladisticanalysis. However, because of cladistic anal-ysis, we could predict, rather than assume,that the ancestor of S. yellowstonensis wouldhave S. niagarae morphology. This avoids thecircularity exhibited in the R. sigmoida, IndianOcean R. praebergonii, and Mandarina snail ex-amples cited above. Those studies explicitlyrely on the ad hoc assumption that the directancestor is present in the fossil samples stud-ied to explain discontinuity in the fossil re-cord. Our study only demonstrates the appar-ent time at which certain plesiomorphic char-acters gave way to apomorphic in the S. yel-lowstonensis lineage.
Evolutionary transformations as rapid asthat in the S. yellowstonensis lineage would bedifficult, if not impossible, to recover in mostdeposits that are purported to record protis-tan anagenesis (Kellogg 1975; Kellogg andHays 1975; Malmgren and Kennett 1981; Ar-nold 1983; Baker 1983; Malmgren et al. 1983;Lazarus 1986; Hunter et al. 1988; Wei and Ken-nett 1988; Lazarus et al. 1995; Norris et al.1996; Pearson et al. 1997; Kucera and Malm-gren 1998). Whereas inferred sedimentationrates in Yellowstone Lake are about 0.6–0.7mm/yr, mean sedimentation rates publishedor calculated from data in those papers rangefrom about 0.008 to 0.100 mm/yr, nearly oneto three orders of magnitude slower than Yel-lowstone Lake. Although there are some ex-amples in which sedimentation rates in ocean

52 EDWARD C. THERIOT ET AL.
or nearshore environments are higher thanthat recorded in many lakes (Wells andShanks 1987; Sancetta 1989), we know of noexamples of anagenetic change in microfossilsfrom these environments. This lack of obser-vation is not evidence that evolution does notproceed more rapidly that has been assumed;rather we believe that it underscores the gulfbetween the great number of possible specia-tion events that could be recovered for micro-fossils and the very few reported cases.
Phylogenetic analysis is required when thefossil record does not demonstrate continuitybetween species. Even for cases with conti-nuity between forms, such as in S. yellowsto-nensis, independent phylogenetic analysisprovides evidence for ancestor-descendant re-lationships. We do not argue that absence oftransitions is proof that rapid transitions arecommon. Rather, we believe that the absenceof transformational data for the many thou-sands of mineralized plankton species that ex-ist or have existed, the lack of independentphylogenetic analyses, and the lack of knowl-edge about the population genetics and struc-ture of most protists should generate cautionabout any generalization regarding tempoand mode of evolution in diatoms, if not in allprotists.
Summary. We provide an empirical exam-ple of diatom evolution occurring more rap-idly than previously demonstrated in the min-eralized protistan fossil record. We examineseveral cases purported to show cladogenesisin the marine plankton diatom record anddemonstrate that the fossil record alone pro-vides insufficient resolution to demonstrateancestor-descendant relationships. In ourstudy, we provide independent stratigraphicand phylogenetic assessment of ancestor-de-scendant relationships and a detailed corre-lation of environmental and morphologicalchange, which together suggest that change inS. yellowstonenis was driven by directional se-lection. A direct cause remains unknown.Thus, this study provides an unusually wellcorroborated and highly resolved record ofevolution purported to document anagenesisassociated with evolution of a new species.Even if there are sufficient cases in other min-eralized plankton groups to support Benton
and Pearson’s (2001) hypotheses that post-spe-ciation evolution in some protists is gradual,published diatom examples and this studyclearly show that the generalization shouldnot yet be extended to diatoms.
Acknowledgments
We thank the late K. Kelts for directing thecoring of Yellowstone Lake, and B. Valero, M.Abbott, E. Truze, and B. Lorry for assistancewith coring. We gratefully acknowledge M.Abbott for radiocarbon dating of the core, C.Tiller for summarizing the lithologic descrip-tion of the core, D. Sea for some of the pollencounts, and S. Kilham and S. Interlandi fordiscussions on diatom ecology in YellowstoneLake. This research was funded by NationalScience Foundation grant BSR-91-07278 to E.C. Theriot.
Literature Cited
Arnold, A. J. 1983. Phyletic evolution in the Goborotalia crassa-formis (Galloway and Wissler) lineage: a preliminary report.Paleobiology 9:390–398.
Bacon, C. R. 1983. Eruptive history of Mount Mazama and Cra-ter Lake Caldera, Cascade Range, USA. Journal of Volcanol-ogy and Geothermal Research 18:57–115.
Baker, C. 1983. Evolution and hybridization in the radiolariangenera Theocorythium and Lamprocyclas. Paleobiology 9:341–354.
Baker, R. G. 1976. Late Quaternary vegetation history of the Yel-lowstone Lake Basin, Wyoming. U.S. Geological Survey Pro-fessional Paper 729-E:E1–E48.
Barnosky, C. W., P. M. Anderson and P. J. Bartlein. 1987. Thenorthwestern U. S. during deglaciation; vegetational historyand paleoclimatic implications. Pp. 289–321 in H. E. Wrightand W. F. Ruddiman, eds. North America and adjacent oceansduring the last deglaciation. Geology of North America, Vol.K-3. Geological Society of America, Boulder, Colo.
Battarbee, R. W. 1973. A new method for the estimation of ab-solute microfossil numbers with reference especially to dia-toms. Limnology and Oceanography 18:647–653.
Benton, M. J., and P. N. Pearson. 2001. Speciation in the fossilrecord. Trends in Ecology and Evolution 16:405–411.
Brown, T. A., D. E. Nelson, R. W. Mathews, J. S. Vogel, and J. R.Southan. 1989. Radiocarbon dating of pollen by acceleratormass spectrometry. Quaternary Research 32:302–212.
Burckle, L. H., and J. Trainer. 1979. Middle and Late Pliocenediatom datum levels from the Central Pacific. Micropaleon-tology 25:281–293.
Chiba, S. 1996. A 40,000-year record of discontinuous evolutionof island snails. Paleobiology 22:177–188.
Crow, J. F., and M. Kimura. 1970. An introduction to populationgenetics theory. Harper and Row, New York.
Cupp, E. E. 1943. Marine plankton diatoms of the west coast ofNorth America. Bulletin of the Scripps Institution of Ocean-ography 5:1–237.
de Vargas, C., R. Norris, L. Zaninetti, S. W. Gibb, and J. Paw-lowski. 1999. Molecular evidence of cryptic speciation inplanktonic foraminifers and their relation to oceanic provinc-

53RAPID DIATOM MORPHOLOGICAL EVOLUTION
es. Proceedings of the National Academy of Sciences USA 96:2864–2868.
de Vargas, C., S. Renaud, H. Hillbrecht, and J. Pawlowski. 2001.Pleistocene adaptive radiation in Globorotalia truncatulinoides:genetic, morphologic, and environmental evidence. Paleobi-ology 27:104–125.
DeMaster, D. J. 1981. The supply and accumulation of silica inthe marine environment. Geochimica et Cosmochimica Acta45:1715–1732.
Despain, D. G. 1990. Yellowstone vegetation; consequences ofenvironment and history in a natural setting. Roberts Rine-hart, Boulder, Colo.
Edgar, S. M., and E. C. Theriot. 2004. Phylogeny of Aulacoseira(Bacillariophyta) based on molecules and morphology. Jour-nal of Phycology 40:772–788.
Faegri, K., P. E. Kaland, and K. Krzywinski. 1989. Textbook ofpollen analysis. Wiley, New York.
Falconer, D. S. 1989. Introduction to quantitative genetics, 3d ed.Wiley, New York.
Fenster, E. J., U. Sorhannus, L. H. Burckle, and A. Hoffman. 1989.Patterns of morphological change in the Neogene diatomNitzschia jouseae Burckle. Historical Biology 2:197–211.
Finlay, B. J., E. B. Monaghan, and S. C. Maberly. 2002. Hypoth-esis: the rate and scale of dispersal of freshwater diatom spe-cies is a function of their global abundance. Protist 153:261–273.
Frey, J. K. 1993. Modes of peripheral isolate formation and spe-ciation. Systematic Biology 42:373–381.
Good, J. M., and K. L. Pierce. 1996. Interpreting the landscape:Recent and ongoing geology of Grand Teton and YellowstoneNational Parks. Grand Teton Natural History Association,Grand Teton National Park, Moose, Wyo.
Hunter, R. S. T., A. J. Arnold, and W. C. Parker. 1988. Evolutionand homeomorphy in the development of the Paleocene Plan-orotalites pseudomenardii and the Miocene Globorotalia (Globor-otalia) margaritae lineages. Micropaleontology 34:181–192.
Interlandi, S. J., S. S. Kilham, and E. C. Theriot. 1999. Responsesof phytoplankton to varied resource availability in large lakesof the Greater Yellowstone Ecosystem. Limnology and Ocean-ography 44:668–682.
Jablonski, D. 2000. Micro- and macroevolution: scale and hier-archy in evolutionary biology and paleobiology. In D. H. Er-win and S. L. Wing, eds. Deep time: Paleobiology’s perspective.Paleobiology 26(Suppl. to No. 4):15–52.
Kellogg, D. E. 1975. The role of phyletic change in the evolutionof Pseudocubus vema (Radiolaria). Paleobiology 1:359–370.
Kellogg, D. E., and J. D. Hays. 1975. Microevolutionary patternsin late Cenozoic radiolaria. Paleobiology 1:150–160.
Kelts, K. R., U. Briegel, K. Ghilard, and K. J. Hsu. 1986. The lim-nogeology ETH coring system. Schweiz Zeitschrift fur Hy-drologie 48:104–115.
Kilham, S. S., E. C. Theriot, and S. C. Fritz. 1996. Linking plank-tonic diatoms and climate change using resource theory in thelarge lakes of the Yellowstone ecosystem. Limnology andOceanography 41:1052–1062.
Kociolek, J. P., and S. A. Spaulding. 2000. Freshwater diatom bio-geography. Nova Hedwigia 71:223–241.
Koizumi, I., and Y. Yanagisawa. 1990. Evolutionary change indiatom morphology—an example from Nitzschia fossilis toPseudoeunotia doliolus. Transactions and Proceedings of the Pa-laeontological Society of Japan, new series 157:347–359.
Kucera, M., and B. A. Malmgren. 1998. Differences between evo-lution of mean form and evolution of new morphotypes: anexample from Late Cretaceous Planktonic Foraminifera. Pa-leobiology 24:49–63.
Lande, R. 1985. Expected time for random genetic drift of a pop-ulation between stable phenotype states. Proceedings of theNational Academy of Sciences USA 82:7641–7645.
———. 1986. The dynamics of peak shifts and the pattern ofmorphological evolution. Paleobiology 12:343–354.
Lazarus, D. 1986. Tempo and mode of morphologic evolutionnear the origin of the radiolarian lineage Pterocanium pris-matum. Paleobiology 12:175–189.
Lazarus, D., H. Hilbrecht, C. Spencer-Cervato, and H. Thier-stein. 1995. Sympatric speciation and phyletic change in Glo-borotalia truncatulinoides. Paleobiology 21:28–51.
Locke, W. W., and G. A. Meyer. 1994. A 12,000-year record ofvertical deformation across the Yellowstone caldera margin:the shorelines of Yellowstone Lake. Journal of Geophysical Re-search 99:20079–20094.
Malmgren, B. A., and J. P. Kennett. 1981. Phyletic gradualism ina Late Cenozoic planktonic foraminiferal lineage: DSDP site284, southwest Pacific. Paleobiology 7:230–240.
Malmgren, B. A., W. A. Berggren, and G. P. Lohman. 1983. Ev-idence for punctuated gradualism in the Late Neogene Glo-borotalia tumida lineage of planktonic foraminifera. Paleobi-ology 9:377–389.
Mann, D. G., and S. J. M. Droop. 1996. Biodiversity, biogeogra-phy and conservation of diatoms. Hydrobiologia 336:19–32.
Mehringer, P. J., Jr., J. C. Sheppard, and F. F. Foit. 1984. The ageof Glacier Peak tephra in west-central Montana. QuaternaryResearch 21:36–41.
Millspaugh, S. H., C. Whitlock, and P. J. Bartlein. 2000. Varia-tions in fire frequency and climate over the past 17000 yr incentral Yellowstone National Park. Geology 28:211–214.
Norris, R. D., R. M. Corfield, and J. Cartlidge. 1996. What isgradualism? Cryptic speciation in globorotalid foraminifera.Paleobiology 22:386–405.
Pearson, P. N., N. J. Shackleton, and M. A. Hall. 1997. Stable iso-tope evidence for the sympatric divergence of Globigerinoidestrilobus and Orbulina universa (planktonic foraminifera). Jour-nal of the Geological Society, London 154:295–302.
Porter, S. C., K. L. Pierce, and T. D. Hamilton. 1983. Late Wis-consin mountain glaciation in the western United States. Pp.71–111 in S. C. Porter, ed. Late Quaternary environments ofthe United States. University of Minnesota Press, Minneapo-lis.
Roh, H. 2000. Resource requirements and competitive abilitiesof diatoms from the greater Yellowstone ecosystem for siliconand nitrogen. Ph.D. dissertation. Drexel University, Philadel-phia.
Round, F. E., R. M. Crawford, and D. G. Mann. 1990. The dia-toms: biology and morphology of the genera. Cambridge Uni-versity Press, Cambridge.
Sancetta, C. 1989. Processes controlling the accumulation of di-atoms in sediments: a model derived from British Columbiafjords. Paleoceanography 4:235–251.
Sarna-Wojcicki, A. M., D. E. Champion, and J. O. Davis. 1983.Holocene volcanism in the conterminous United States andthe role of silicic volcanic ash layers in the correlation of latestPleistocene and Holocene deposits. Pp. 52–77 in H. E. Wright,ed. Late Quaternary environments of the United States, Vol.2. University of Minnesota Press, Minneapolis.
Schrader, H. J. 1973. Cenozoic diatoms from the Northeast Pa-cific, Leg 18. In L. D. Kulm and R. von Huene, eds. Initial Re-ports of the Deep Sea Drilling Project 18:673–765.
———. 1974. Cenozoic marine planktonic diatom stratigraphyof the tropical Indian Ocean. In R. L. Fisher and E. T Bunce,eds. Initial Reports of the Deep Sea Drilling Project 24:887–941.
Sorhannus, U. 1990a. A new planktonic diatom species, Rhizo-solenia sigmoida, from upper Pliocene sediments of the equa-torial Pacific. Micropaleontology 36:102–103.
———. 1990b. Punctuated morphological change in a Neogenediatom lineage: ‘‘local’’ evolution or migration? Historical Bi-ology 3:241–247.

54 EDWARD C. THERIOT ET AL.
Sorhannus, U., E. J. Fenster, L. H. Burckle, and A. Hoffman. 1988.Cladogenetic and anagenetic changes in the morphology ofRhizosolenia praebergonii Mukhina. Historical Biology 1:185–205.
———. 1991. Iterative evolution in the diatom genus Rhizosolen-ia. Lethaia 24:39–44.
Stuiver, M., P. J. Reimer, and T. F. Braziunas. 1998. High-preci-sion radiocarbon age calibration for terrestrial and marinesamples. Radiocarbon 40:1127–1151.
Sundstrom, Bo. 1986. The marine diatom genus Rhizosolenia: anew approach to the taxonomy. Ph.D. dissertation. Lund Uni-versity, Lund, Sweden.
Taylor, S. M. 1994. Resource requirements and competitive in-teractions of two closely related Stephanodiscus species for ni-trogen and silica. M.S. thesis. Drexel University, Philadelphia.
Theriot, E. 1984. Morphological variation in Stephanodiscus nia-garae (Bacillariophyceae). Ph.D. dissertation. University ofMichigan, Ann Arbor.
———. 1987. Principal component analysis and taxonomic in-terpretation of environmentally related variation in silicifi-cation in Stephanodiscus (Bacillariophyceae). British Phycolog-ical Journal 22:359–373.
———. 1992. Clusters, species concepts and morphological evo-lution of diatoms. Systematic Biology 41:141–157.
Theriot, E., and E. F. Stoermer. 1981. Some aspects of morpho-logical variation in Stephanodiscus niagarae (Bacillariophy-ceae). Journal of Phycology 17:64–72.
———. 1984a. Principal component analysis of Stephanodiscus:observations on two new species from the Stephanodiscus nia-garae complex. Bacillaria 7:37–58.
———. 1984b. Principal component analysis of variation in Ste-phanodiscus rotula and S. niagarae (Bacillariophyceae). System-atic Botany 9:53–59.
———. 1984c. Principal component analysis of character varia-tion in Stephanodiscus niagarae Ehrenb.: morphological varia-
tion related to lake trophic status. Pp. 97–111 in D. G. Mann,ed. Proceedings of the seventh international diatom sympo-sium. Koeltz, Koenigstein.
Theriot, E., Y. Z. Qi, J. R. Yang, and L. Y. Ling. 1988. Taxonomyof the diatom Stephanodiscus niagarae from a fossil depositin Jingyu County, Jilin Province, China. Diatom Research 3:145–153.
Theriot, E. C., S. C. Fritz, and R. E. Gresswell. 1997. Long-termlimnological data from the larger lakes of Yellowstone Na-tional Park. Journal of Arctic and Alpine Research 29:304–314.
Thompson, R. S., C. Whitlock, P. J. Bartlein, S. P. Harrison, andW. G. Spaulding. 1993. Climatic changes in the western Unit-ed States since 18,000 yr B. P. Pp. 468–513 in H. E. Wright., ed.Global climates since the last glacial maximum. University ofMinnesota Press, Minneapolis.
Tiller, C. C. 1995. Postglacial sediment stratigraphy of largelakes in greater Yellowstone. M.S. thesis. University of Min-nesota, Minneapolis.
Waddington, J. C. B., and H. E. Wright Jr. 1974. Late Quaternaryvegetational changes on the east side of Yellowstone NationalPark, Wyoming. Quaternary Research 4:175–184.
Wei, K.-Y., and J. P. Kennett. 1988. Phyletic gradualism andpunctuated equilibrium in the late Neogene planktonic fora-miniferal clade Globoconella. Paleobiology 14:345–363.
Wells, J. T., and A. L. Shanks. 1987. Observations and geologicalsignificance of marine snow in a shallow-water, partially en-closed marine embayment. Journal of Geophysical Research92:185–190.
Whitlock, C. 1993. Postglacial vegetation and climate of GrandTeton and southern Yellowstone National Parks. EcologicalMonographs 63:173–198.
Whitlock, C., and P. J. Bartlein. 1993. Spatial variations of Ho-locene climatic change in the Yellowstone region. QuaternaryResearch 39:231–238.
Wiley, E. O. 1981. Phylogenetics: the theory and practice of phy-logenetic systematics. Wiley, New York.
Copyright © 2022 FDOKUMEN




















