


Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in...
39
Accepted Manuscript Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in comparison with curcumin - An in vitro and in silico approach Kumaravel Mohankumar, Sankar Pajaniradje, Subhashree Sridharan, Vivek Kumar Singh, Larance Ronsard, Akhil C. Banerjea, Chellakkan S. Benson, Mohane Selvaraj Coumar, Rukkumani Rajagopalan PII: S0009-2797(13)00336-0 DOI: http://dx.doi.org/10.1016/j.cbi.2013.12.006 Reference: CBI 6959 To appear in: Chemico-Biological Interactions Received Date: 18 September 2013 Revised Date: 20 November 2013 Accepted Date: 16 December 2013 Please cite this article as: K. Mohankumar, S. Pajaniradje, S. Sridharan, V.K. Singh, L. Ronsard, A.C. Banerjea, C.S. Benson, M.S. Coumar, R. Rajagopalan, Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in comparison with curcumin - An in vitro and in silico approach, Chemico-Biological Interactions (2013), doi: http://dx.doi.org/10.1016/j.cbi.2013.12.006 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an analog of curcumin in...

Accepted Manuscript
Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by ananalog of curcumin in comparison with curcumin - An in vitro and in silicoapproach
Kumaravel Mohankumar, Sankar Pajaniradje, Subhashree Sridharan, VivekKumar Singh, Larance Ronsard, Akhil C. Banerjea, Chellakkan S. Benson,Mohane Selvaraj Coumar, Rukkumani Rajagopalan
PII: S0009-2797(13)00336-0DOI: http://dx.doi.org/10.1016/j.cbi.2013.12.006Reference: CBI 6959
To appear in: Chemico-Biological Interactions
Received Date: 18 September 2013Revised Date: 20 November 2013Accepted Date: 16 December 2013
Please cite this article as: K. Mohankumar, S. Pajaniradje, S. Sridharan, V.K. Singh, L. Ronsard, A.C. Banerjea,C.S. Benson, M.S. Coumar, R. Rajagopalan, Mechanism of apoptotic induction in human breast cancer cell, MCF-7,by an analog of curcumin in comparison with curcumin - An in vitro and in silico approach, Chemico-BiologicalInteractions (2013), doi: http://dx.doi.org/10.1016/j.cbi.2013.12.006
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting proof before it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

Mechanism of apoptotic induction in human breast cancer cell, MCF-7, by an
analog of curcumin in comparison with curcumin - An in vitro and in silico
approach
Kumaravel Mohankumara, Sankar Pajaniradjea, Subhashree Sridharana, Vivek Kumar Singhb,
Larance Ronsardc, Akhil C Banerjeac, Chellakkan S Benson d, Mohane Selvaraj Coumarb,
Rukkumani Rajagopalana*.
aDepartment of Biochemistry and Molecular Biology, School of Life Sciences, Pondicherry
University, Puducherry 605 014. India. bCentre for Bioinformatics, School of Life Sciences, Pondicherry University, Puducherry 605
014. India. cVirology Laboratory-II, National Institute of Immunology, New Delhi, India, dDepartment of Endocrinology, Dr. ALM PG Institute of Basic Medical Sciences, University of
Madras, Chennai-600 013. India
* Corresponding Author:
Dr. R. Rukkumani,
Assistant Professor,
Department of Biochemistry and Molecular Biology,
School of Life Sciences,
Pondicherry University,
Puducherry - 605 014.
India.
E-mail: [email protected], [email protected]
Ph: +91 9677847337, +91 413 2654537

1
Abstract
In Developing countries, survival rates for breast cancer are poor and it accounts for 22.9% of all
cancers in women. Curcumin, a major constituent from turmeric, is one of the well-known
chemopreventive agents. Reports have shown that curcumin induces apoptosis in breast cancer
cells. We synthesized an ortho-hydroxy substituted analog of curcumin (BDMC-A) and analyzed
its cytotoxicity. The BDMC-A inhibited MCF-7 at a dose equivalent to that of curcumin (30
µM). The present study was aimed at delineating the apoptotic mechanism of BDMC-A in
comparison to that of curcumin. In our study, BDMC-A exerted more potent effect on the
modulation of selective apoptotic markers (intrinsic pathway: p53, Bcl-2, Bax, cyt c, Apaf-1,
caspase-9, 3, PARP; extrinsic pathway: FasL, Caspase 8) compared to curcumin. mRNA
expression studies for Bcl2/Bax also supported the increased efficacy of BDMC-A. An in silico
molecular docking study with PI3K revealed that the docking of BDMC-A was more potent
compared to curcumin. Increased apoptotic induction by BDMC-A compared to curcumin was
also demonstrated by Annexin V, Rh123(∆Ψm), PI, Hoechst 33258, AO/EB fluorescent staining
studies which showed characteristic apoptotic features like nuclear fragmentation and chromatin
condensation. Moreover, BDMC-A treated cells effectively induced apoptosis through ROS
intermediates compared to curcumin, as measured by 2'-7'dichloro dihydro fluorescein diacetate
(DCFH-DA). Hence our overall results showed that BDMC-A induced apoptosis more
effectively compared to curcumin and the activity can be attributed to the presence of hydroxyl
group in the ortho position in its structure. Further researches are going on to delineate its
molecular targets to evaluate its effect as a potent anticancer agent.
Key words: MCF-7; BDMC-A; Curcumin; Apoptosis; Western Blotting; Molecular
Docking

2
1. Introduction
Cancer is a multifactorial disease that involves modulation of multiple pathways and targets. In
India, breast cancer is the second leading cause of cancer deaths in women and the risk of its
incidence is increasing every year. Several researches are going on to identify chemotherapeutic
and chemopreventive agents that can act on multiple signaling targets [1]. Identification of
pharmacologically safe phytochemicals that have multitargeting effect is a ‘hotspot’ in cancer
research. One such phytochemical is curcumin, the principle ingredient from the yellow spice
turmeric. It is a well-known plant derived chemopreventive agent, common in Southeast Asian
countries and used as a folk medicine and traditional food for several centuries [2]. This
pleiotropic molecule has many pharmacological properties and popularly known for its
anticarcinogenic, antioxidant and anti inflammatory activities [3-4].
Previous studies have shown the anticancer effect of curcumin against various cancers like
colorectal, pancreatic, liver, lung, prostate, ovarian, head and neck cancer, leukemia, multiple
myeloma and breast cancer [5-6]. Enormous studies have indicated the multitargeting efficacy of
curcumin to combat cancer. The chemopreventive properties are also attributed to the induction
of apoptosis, which is considered to be the hallmark in cancer treatment [7-8]. Apoptosis, a form
of programmed cell death is involved in maintenance of tissue homeostasis and provides a
controlled cell deletion to balanced cell proliferation through a biochemical cascade of
mechanism [9]. Curcumin’s ability to induce apoptosis in cancer cells without affecting the
healthy cells establishes an understanding of the anticancer potential of curcumin. Reports have
shown that many curcumin analogs are well known for their anti-tumour promoting and
antiproliferative activities [10] and thereby induce apoptosis in many cancer cells [11], but the
mechanism underlying these effects are not well understood [12-13].
Our compound is one such analog, BDMC-A, which has antitumor, anti mutagenic [14] and
antioxidant [15] properties. The protective role of BDMC-A in alcohol and ∆PUFA induced
oxidative stress [16], hyperlipidemia [17], circulatory lipid profiles [18], matrix
metalloproteinases [19] in experimental rats have already been reported from our lab. BDMC-A
has also been analysed for the anticarcinogenic effect in DMH-induced colon cancer [20]. We

3
have reported earlier the antiproliferative effect of BDMC-A through cell cycle arrest at G2/M
phases in MCF-7 cells [28]. Our present study is aimed at evaluating the apoptosis inducing
ability of BDMC-A in comparison with curcumin.
There are two major apoptosis signaling pathways. The death receptor (extrinsic) pathway and
the mitochondria (intrinsic) mediated pathway. The extrinsic pathway is initiated by death
receptors which are commonly activated by TNF-α and FasL. Once these receptors are activated,
they oligomerize and form scaffolding complexes that recruit caspase family of death protease,
classically caspase-8. This activated initiator caspase cleaves downstream effector caspases,
particularly caspase-3, which culminates in apoptosis.
Induction of intrinsic pathway is through some anticancer drugs and hypoxia. Permeabilization
of outer mitochondrial membrane is induced by these stimuli. This results in the activation of
mitochondrial pathway by releasing apoptogenic factors like cytochrome c from mitochondrial
intermembrane space to cytosol. The release of cytochrome c into the cytosol triggers activation
of caspase-3 through cytochrome c/Apaf-1/caspase 9 apoptosome complex [21].
To identify the possible molecular target of BDMC-A and curcumin, we investigated the
PI3K/Akt signaling axis in silico as possible targets. PI3K/Akt signaling axis has been already
suggested as target for curcumin for its anti-cancer effect [22-24]. Here we investigated the
interaction of curcumin and its analog (BDMC-A) to these two targets using docking, to
rationalize their anti-cancer activity.
2. Materials and Methods
2.1. Chemicals
Curcumin (1,7 – bis - (4- hydroxy -3-methoxyphenyl ) -1,6- hepatadiene -3,5- dione;
diferuloylmethane), Dulbecco's Modified Eagle Medium (DMEM), antibiotics and fetal bovine
serum (FBS), annexin V cy3 kit, acridine orange (AO), ethidium bromide (EtBr), DMSO,
Hoechst 33258, propidium iodide (PI), rhodamine 123, 2'-7'-Dichlorodihydrofluorescein

4
diacetate (DCFH-DA) were purchased from Sigma–Aldrich, Bangalore, India. Primary
antibodies against Bcl-2, Bax, caspase-3, caspase-8, caspase-9, cytochrome c, p53, FasL and
PARP, PI3K, pAkt were purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA) and
cell signaling technology, USA. The secondary antibodies were obtained from santacruz
Biotechnology, USA.
2.2. Curcumin analog
Curcumin analog, BDMC-A, was synthesized as per the method described by Dinesh Babu and
Rajasekharan [25].
2.3. Cell lines and culture conditions
MCF-7 cell line was obtained from NCCS, Pune, India. The cells were maintained in
1×Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% FBS at 37oC in CO2
incubator in an atmosphere of humidified 5% CO2 and 95% air. The cells were maintained by
routine sub culturing in tissue culture flasks. The culture medium was changed every 48 h and
the cells were split when they reached confluence.
2.4. Western blotting
Western blotting was carried out as described by Fido et al, [26]. Briefly the human breast cancer
cells, MCF-7, were grown in 6 well plate and selected concentration of curcumin and BDMC-A
were added. After 24 hours, nuclear proteins and cytosolic proteins (cytochrome c) were
extracted and protein concentrations were determined by using Bio-Rad Bradford protein assay.
Lysate samples were resolved by 12% sodium dodecyl sulphate (SDS)-polyacrylamide gel
electrophoresis and then proteins were electro-transferred to nitrocellulose membrane. The
primary antibodies of apoptotic markers were added and incubated at 4°C overnight. Membranes
were incubated with corresponding secondary antibodies for 1 hour. Protein abundance of β-
actin antibody served as a control for protein loading. Protein bands were revealed by enhanced
chemiluminescence method and the signals were detected by ECL, (Pierce, Rockford, IL, USA)
and Chemi Doc Imaging System (Bio-Rad, Hercules, CA, USA).

5
2.5. RT-PCR
Briefly, MCF-7 cells were grown in 6 well plate and selected concentrations of curcumin and
BDMC-A were added. After 12 h treatment, total RNA was extracted by using TRIzol extraction
method [27] according to the manufacturer’s instructions. Reverse transcription was performed
with 2 µg of total RNA using gene specific upstream and downstream primers for Bcl-2
(Upstream - 5’- attgggaagtttcaaatcagc - 3’, Downstream - 5’-tgcattcttggacgaggg-3’) and Bax
(Upstream - 5’- gctggacattggacttcctc - 3’, Downstream - 5’- ctcagcccatcttcttccag – 3’) . Relative
target mRNA expression level was normalized by GAPDH (Upstream - 5’- cgaccactttgtcaagctca
- 3’, Down straem - 5’ - cccctcttcaaggggtctac – 3’) in the same sample. PCR was run on optimum
conditions and the products were analyzed by running in 2% agarose gel with ethidium bromide
staining and captured in gel documentation system.
2.6. Morphological changes in cell apoptosis by Hoechst 33258 staining
Cell nuclear morphology was evaluated by fluorescence microscopy following Hoechst 33258
DNA staining. MCF-7 cells were seeded in 24-well plates at a density of (1X105 cells/ml) and
were grown for 24h. The cells were treated with BDMC-A (15 µM) and curcumin (15 µM) for
24h. IC50 concentrations and the cytostatic dose were fixed from our previous reports [28]. The
cells were washed with phosphate buffered saline, fixed with methanol, resuspended in 8 µg/mL
Hoechst 33258, and incubated for 30 min at 37°C in dark. The cells were then washed with PBS
and examined under a Nikon fluorescent microscope.
2.7. Detection of apoptotic nuclei by AO/EB staining
Acridine Orange/Ethidium Bromide (AO/EB) staining was carried out to detect morphological
evidence of apoptosis. MCF-7 cells were treated with BDMC-A and curcumin for 24 h, washed
with PBS and trypsinized. 25 µl of cell suspension (1X104 cells/ml) was mixed with 1 µl of
acridine orange/ ethidium bromide solution (one part each of 100 µg /ml of acridine orange and
100 µg/ml of ethidium bromide in PBS) just prior to microscopy. Ten microliters of gently
mixed suspension was placed on a microscope slide covered with glass slip and examined under
fluorescent microscope (Olympus) connected to a digital imaging system.

6
2.8. Propidium iodide staining
DNA integrity was detected using PI staining [29]. Briefly, MCF-7 (3x106 cells/mL) cells were
grown in 6 well plates with BDMC-A and curcumin for 24 h at 37 ºC. Cells were washed with
ice cold PBS and fixed in 70% ethanol. Cells were then washed twice with PBS. 0.5 mL of PI
buffer containing 0.1% Triton X-100, 0.1% sodium citrate, 5 µL of RNase A (1mg/mL) and 5 µL
of PI (50µg/mL in PBS) were added and incubated at 37°C for one hour and the fluorescence
was observed under fluorescent microscope (Olympus).
2.9. Annexin V Cy3 staining for early apoptosis
Annexin V Cy3TM (APO–AC, Sigma) kit was used to measure the early stage of apoptosis. The
kit contains two fluorescent labels to measure cell viability and apoptosis. Non-fluorescent 6
carboxyfluorescein diacetate (6-CFDA) is hydrolyzed to green fluorescent compound, 6-
carboxyfluorescein (6-CF) by esterase present in the live cells. AnnexinV-Cy3.18 (AnnCy3)
binds to phosphatidylserine that is present on the outer leaflet of the plasma membrane of cells
that are in the initial stages of apoptosis (observed as red fluorescence). Live cells are stained
only with 6-CF (green) while the cells in the early stage of apoptosis are stained with both
AnnCy3 (red) and 6-CF (green). MCF-7 cells after treatment with BDMC-A and curcumin for
24 h were stained with these fluorescent compounds and observed under fluorescent microscope
(Olympus).
2.10. Observation of mitochondrial membrane potential (∆Ψm) by Rhodamine 123
Rhodamine 123 is a fluorescent dye that accumulates as aggregates in polarized mitochondrial
membranes of normal cells. This cationic dye binds to metabolically active mitochondria.
Curcumin and BDMC-A treated cells were fixed with ice cold methanol for 30 mins and
incubated with 5 µg/ml rhodamine 123 at 37 ℃ for 30 min. After incubation, the cells were
washed once with PBS. The intensity of rhodamine 123 staining was measured by fluorescence

7
microscopy at excitation wave length 505 nm with emission filter 534 nm (Nikon fluorescent
microscope).

8
2.11. Intracellular reactive oxygen species (ROS) imaging
ROS generation was measured by the oxidative-sensitive fluorescent probe 2'-7'-
Dichlorodihydrofluorescein diacetate (DCFH-DA). To determine the role of ROS in cytotoxicity
caused by BDMC-A and curcumin, qualitative fluorescence microscopic analysis was done by
oxidative-sensitive fluorescent probe using DCFH-DA stain. Briefly, 0.2 x 106 cells were
pre‐cultured in a 6‐well plate for 48 h in DMEM and subsequently exposed to BDMC-A and
curcumin for 24 h. Following incubation, the cells were washed once with PBS and then stained
with 10 µM DCFH-DA for 30 min at 37°C in dark. Cells were again washed twice with chilled
PBS. Stain positive areas were analyzed and photographed by Nikon Fluorescence Microscope
(DCFH-DA Ex 488nm, Em 520 nm).
2.12. Molecular Docking
2.12.1. Ligands used
1. Curcumin
2. BDMC-A (Curcumin analog)
2.12.2. Ligand preparation
The ligand structures were sketched using Chemdraw software and the files stored in sdf format.
These structures were imported into the Schrödinger suite 2010 (v9.1) environment and
subjected to ligand preparation to generate possible conformations and ionization states using the
LigPrep module, keeping default values.
2.12.3. 3D protein structures used
1. PI3K (PDB: 1E8Z )
2. Akt (PDB: 4GV1)
2.12.4. Protein preparation
The protein structures were prepared using Protein preparation wizard of Schrodinger 2010
(v9.1). For this purpose first all the bond orders in the protein were assigned, hydrogen atoms
were added and all the crystallographic water molecules were removed. Subsequently, missing
residues and loops were added using Prime module of Schrodinger 2010. Finally the protein

9
complex was subjected to energy minimization using OPLS-2005 force field with implicit
solvation.
2.12.5. Docking
Docking studies of the ligands were performed using Glide Module of Schrodinger. The protein-
ligand complex prepared as described above was used to build energy grids using the default
values of protein atom scaling (1.0) within a cubic box around the centroid of the co-crystallized
ligand. The boundary box dimensions (within which the centroid of a docked pose is confined)
were set to 12Å *12Å* 12Å. Next, the grid files generated above were used for docking the
ligands in Glide Xtra Precision (XP) mode. Default parameters were utilized except for the
energy minimization steps (n = 200) for carrying out the docking of the ligand to the protein in
flexible mode.
2.12.6. Binding free energies calculations
The Molecular Mechanics /Generalized Born Surface Area (MM/GBSA) for the calculation of
binding free energies was employed using PRIME module of Schrodinger 2010 (v9.1). The
Prime/MM–GBSA method uses docked pose file of the receptor and ligand to calculate the
binding-free energy (∆Gbind) of each ligand. The simulation was performed based on the
receptor–ligand complex structure obtained from molecular docking. The obtained ligand poses
were minimized using the local optimization feature in Prime, whereas the energies of complex
were calculated with the OPLS-2005 force field and Generalized-Born/Surface Area continuum
solvent model. During the simulation process, the ligand strain energy was also considered.
2.13. Statistical Analysis
All the data were analyzed using the SPSS 7.5-Windows Students version software (SPSS
Inc., Chicago, IL, USA). For all the measurements, one-way ANOVA followed by Tukey’s
test was used to assess the statistical significance between groups. A statistically significant
difference was considered at the level of p≤0.05.

10
3. Results and Discussion
BDMC-A (Figure 1A) exhibited potent apoptotic effect by modulating both intrinsic and
extrinsic apoptotic markers in MCF-7 cells compared to curcumin (Figure 1B). In our previous
study we have reported the antiproliferative effects of BDMC-A in MCF-7 cells through cell
cycle arrest at G2/M phase. Also we showed an equivalent cytotoxic effect of BDMC-A
compared to curcumin by MTT assay and the IC50 value was 30 µM for both curcumin and
BDMC-A [28]. In the present study we have investigated the putative markers for apoptosis
induced cell death - both the intrinsic (mitochondrial) and the extrinsic (death receptor) pathway.
3.1. PI3K/Akt and Intrinsic pathway apoptotic markers
PI3K/Akt pathway is a cell survival pathway that is important for normal cell growth and
proliferation. Numerous researches in breast cancer have shown that deregulation of this
pathway is implicated in tumorigenesis and hence this has become an important target for breast
cancer treatment [30]. In our present report significant downregulation of pAKT and PI3K were
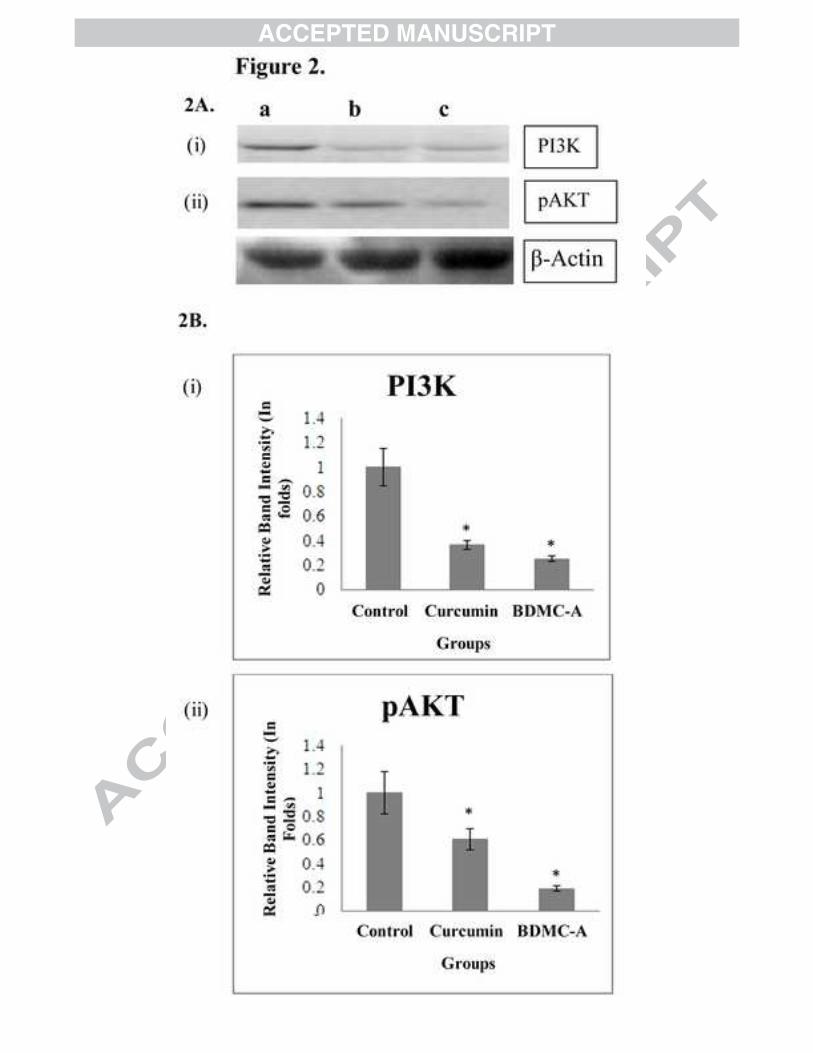
observed on treatment with BDMC-A compared to curcumin (Figure 2A, 2B (i, ii)).
Overexpression of cyclin D1 was reported with many cancers including breast cancer [31]. It has
been reported that when there is an inhibition in Akt phosphorylation, there is an increase in
GSK-3β that in turn cause inhibition of cyclin D1 protein expression that leads to cell cycle
arrest [32]. We have previously reported a decrease in cyclin D1 expression upon treatment with
BDMC-A [28], that inhibited cell cycle progression. This may be due to inhibition of Akt
phosporylation as observed in our study.
MCF-7 is more sensitive to curcumin and several reports have shown the induction of apoptosis
by curcumin through intrinsic pathway [33]. In our present study, we analyzed the intrinsic
apoptotic markers viz. p53, Bcl-2, Bax, cytochrome c, Apaf-1, caspase 9, caspase 3 and PARP .
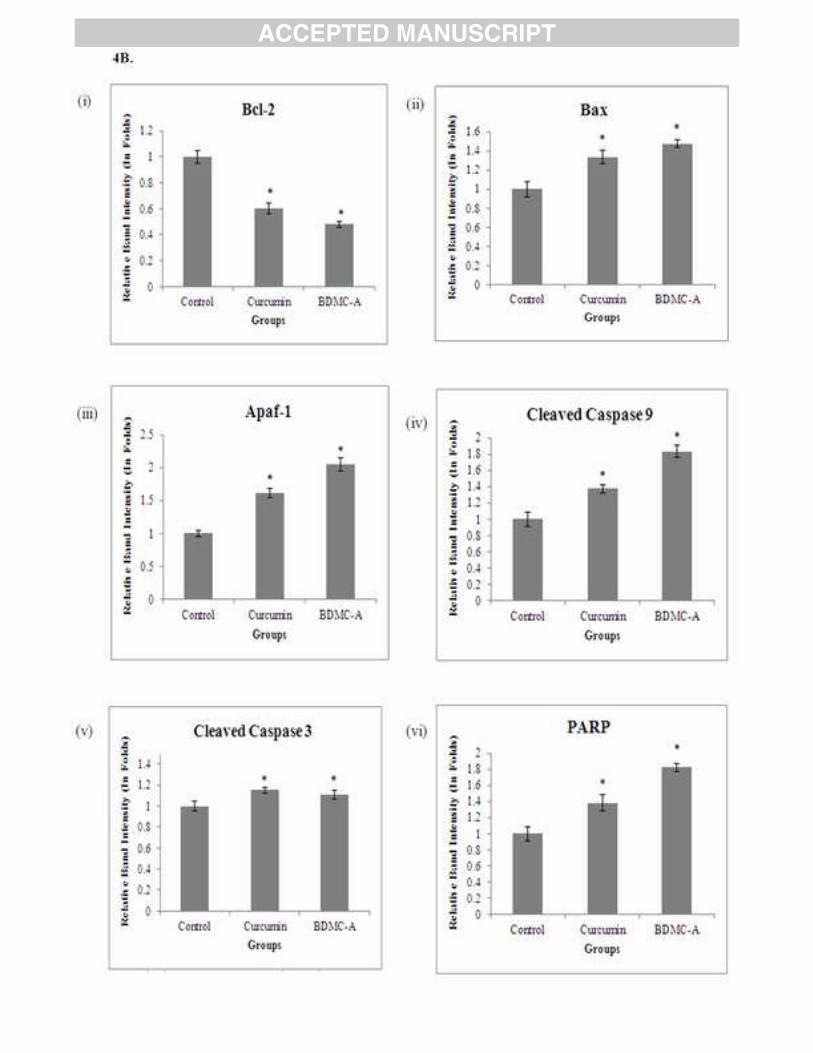
Treatment with BDMC-A significantly down regulated the Bcl-2 protein expression and
upregulated the expressions of p53, Bax, cytochrome c, Apaf-1, Cleaved caspase-9 and 3 and
PARP compared to curcumin. In many cancers, PI3K activates Akt, which in turn activates
Mdm2 that inhibits the downstream protein p53 thereby inhibiting apoptosis [34]. Functioning of
p53, upregulates Bax expression and induces apoptosis [35]. In normal cells, p53 is considered

11
as a ‘master regulator’ whereby the mutant form is seen mostly in all forms of cancer. An
important function of p53 is to act as a transcription factor that particularly binds to apoptosis
responsive genes which are expected to increase the synthesis of p21cip1or Bax [36]. Bcl-2 family
members are critical determinants of cellular homeostasis. Bcl-2 family proteins are structurally
related molecules, which positively or negatively regulate apoptosis. Bax, a member of Bcl-2
family, is a proapoptotic signaling protein, which antagonizes Bcl-2, an antiapoptotic signaling
protein, and induces apoptosis in many types of cancers [37]. Balance between Bcl-2/ Bax ratio
confirms the apoptotic status of the cell. When the Bax/Bcl-2 ratio is increased, the protective
effect of Bcl-2 on the mitochondrial membrane is disrupted and permeability increases, allowing
cytochrome c to leak into the cytosol as shown in our present report.
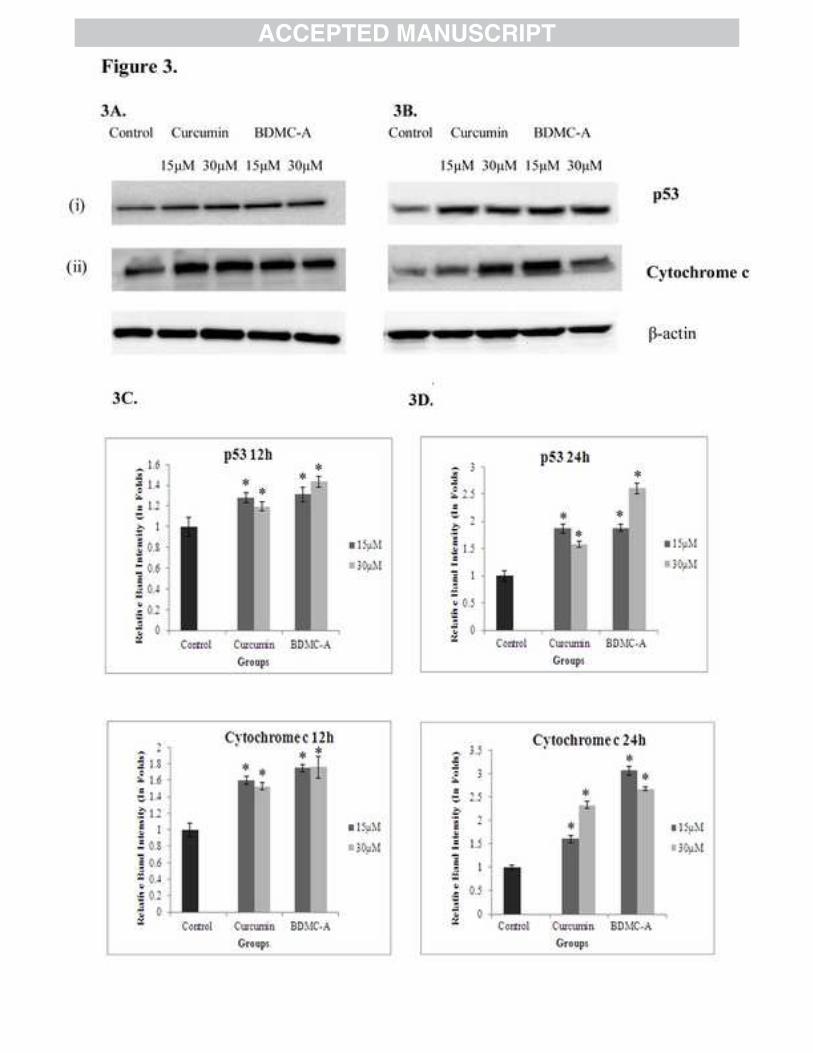
Our findings suggested that, on treatment with BDMC-A, p53 was activated (Figure 3A, 3B (i)),
which upregulated cytochrome c (Figure 3A, 3B (ii)) through Bax and acted as a master switch
for inducing apoptosis in a time dependent fashion. The downregulation of PI3K and
upregulation of p53 evidenced from our results can be corroborated to the significant
upregulation of Bax and downregulation of Bcl-2 protein expressions (Figure 4A & 4B (i & ii))
in BDMC-A treated cells.
Cytochrome c is an essential component of the electron transport chain in mitochondria that is
involved in initiation of apoptosis [38]. Several reports have shown that the leakage of
cytochrome c (hallmark for intrinsic apoptotic pathway) directly activates caspase 3 and induces
apoptosis in wide range of cancer with curcumin treatment [39]. In our study the expression of
cytochrome c was comparatively higher in BDMC-A treatment, compared to curcumin.
Cytochrome c binds to Apaf-1 (apoptotic protease activating factor-1) and caspase 9 to form an
apoptososme complex, which initiates the caspase cascade via activation of caspase-3 and
induces apoptosis in many cancers [40]. In our study BDMC-A proved to be more effective in
upregulating Apaf-1 expression (Figure 4A & 4B (iii)) and activation of initiator caspase,
caspase-9 (Figure 4A & 4B (iv)) than curcumin. Along with cytochrome c, Apaf-1 and activated
caspase-9 might have formed the apoptososme complex and activated the effector caspase,
caspase-3 (Figure 4A & 4B (v)). Activated caspase-3 might have proceeded to a cascade of

12
subsequent changes responsible for the apoptosis, exemplified by fragmentation of Poly ADP-
ribose Polymerase (PARP).
PARP is a family of proteins involved in many cellular processes that leads to apoptosis. In our
study, BDMC-A treated cells significantly upregulated the cleavage of PARP thereby inducing
programmed cell death better than curcumin (Figure 4A & 4B (vi)). mRNA expression analysis
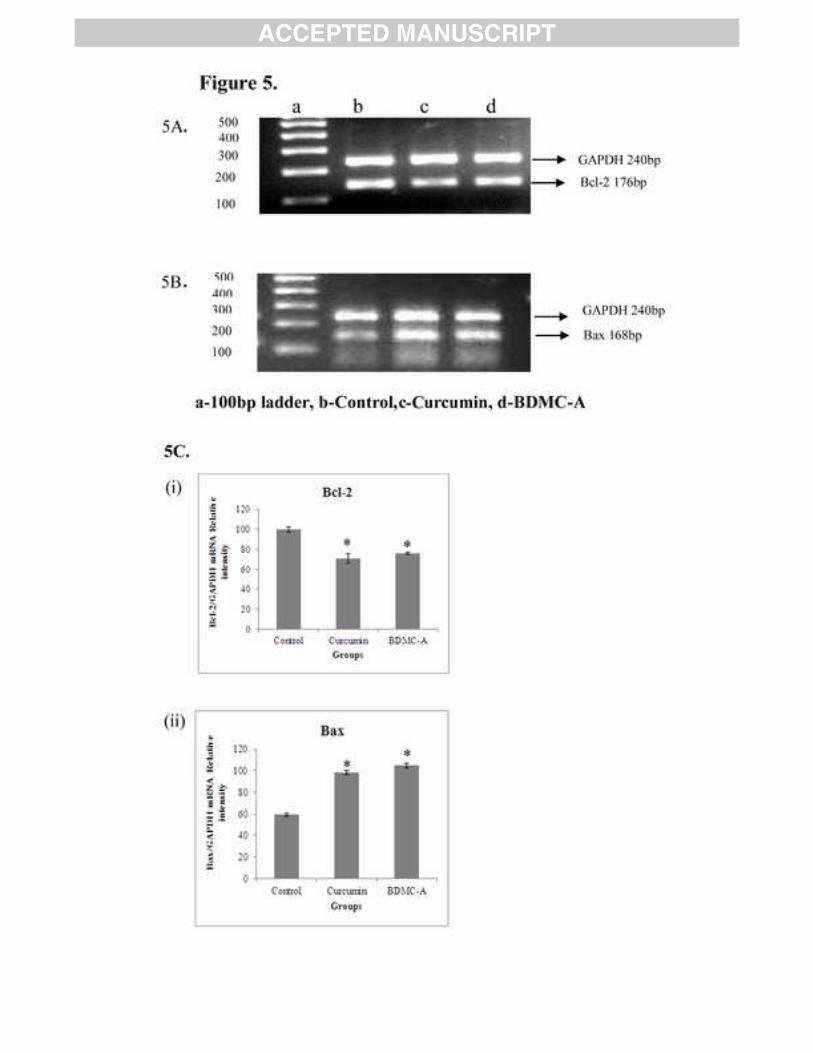
also confirmed downregulation of Bcl-2 (Figure 5A) and upregulation of Bax (Figure 5B) in
BDMC-A treatment. Our results suggest that BDMC-A disrupts mitochondrial integrity and
triggers apoptosis and leads to Bax dependent outer membrane pore formation in mitochondria.
Induction of apoptosis through mitochondria by BDMC-A is further confirmed by the leakage of
cytochrome c. Hence the result with the intrinsic pathway provides the evidence that apoptosis
happened via mitochondrial pathway and proved that BDMC-A was more potent than curcumin
in inducing apoptosis.
3.2. Extrinsic apoptotic pathway markers
Many reports have shown the stimulatory effect of curcumin on extrinsic apoptotic pathway,
which is triggered by “Death activators” such as TNF-α and Fas ligand to their corresponding
cell surface receptors. Activation of death receptor pathway plays a major role in apoptosis and
results in activation of caspase-8 and initiates caspase cascade. Upregulation of death receptors
by chemopreventive agents like curcumin has been shown to sensitize cancer cells and aggregate
Fas receptors thereby increasing the levels of caspase-8 and 3 in many cancer cells [41]. Our
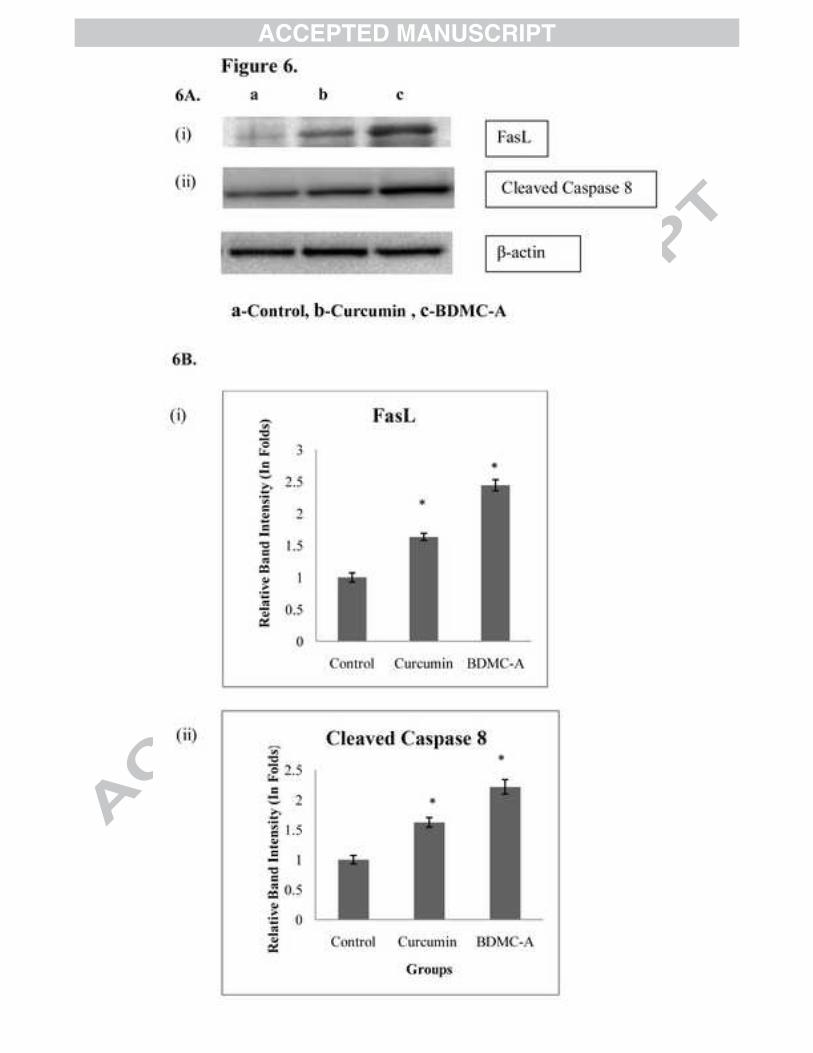
results showed that there was a significant increase in FasL expression (Figure 6A & 6B (i)) in
BDMC-A treated cells compared to curcumin. Death initiator caspase 8 are caspases that activate
effector caspases and lead to apoptosis. In our study, BDMC-A treated cells induced the
upregulation of cleaved caspase-8 (Figure 6A & 6B (ii)) which in turn activated caspase-3 and
induced apoptosis more potently compared to curcumin

13
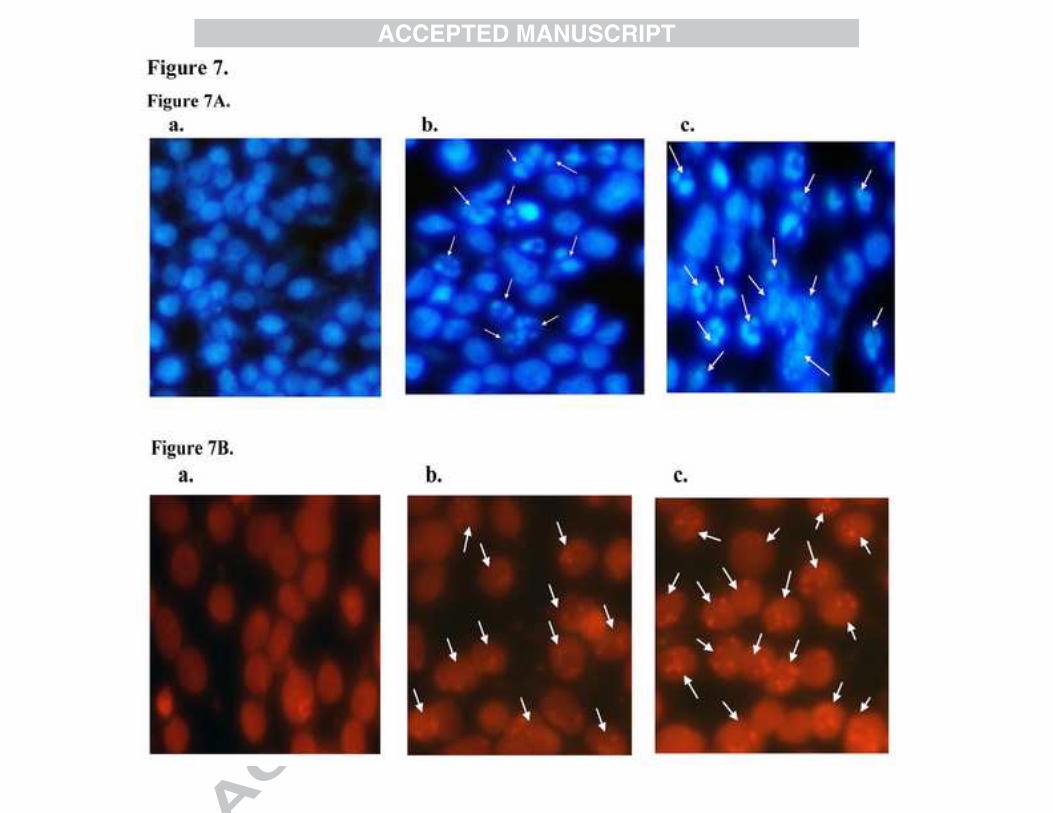
Morphological analysis for detection of apoptosis
Apoptotic cells can be differentiated from necrotic cells in several ways.
3.3. Fluorescent staining with Hoechst 33258
To investigate cellular apoptosis by BDMC-A, MCF-7 cells were treated with BDMC-A and
curcumin for 24h. After 24h, the cells were stained with Hoechst staining 33258. A clear evident
with morphological changes in cell apoptosis such as chromatin condensation and nuclear
fragmentation were seen in BDMC-A treated cells. The changes were significant compared to
curcumin and the apoptotic index was positively correlated (Figure7A).
3.4. Fluorescent staining with Propidium Iodide
Nuclear fragmentation was characterized for the identification of apoptotic cell death. We
examined the morphology of nucleus using a fluorescent DNA-binding agent, PI. PI was taken
up by apoptotic cells and the fragmented apoptotic bodies were seen. MCF-7 cells were treated
with respective IC50 concentrations of BDMC-A and curcumin and the fragmented DNA
content was analysed. The results of our study clearly revealed more fragmented apoptotic
bodies upon treatment with BDMC-A than curcumin. BDMC-A induced DNA damage like
chromatin condensation and nuclear fragmentation, which are the indicators of apoptotic death,
in cancer cells (Figure7B).
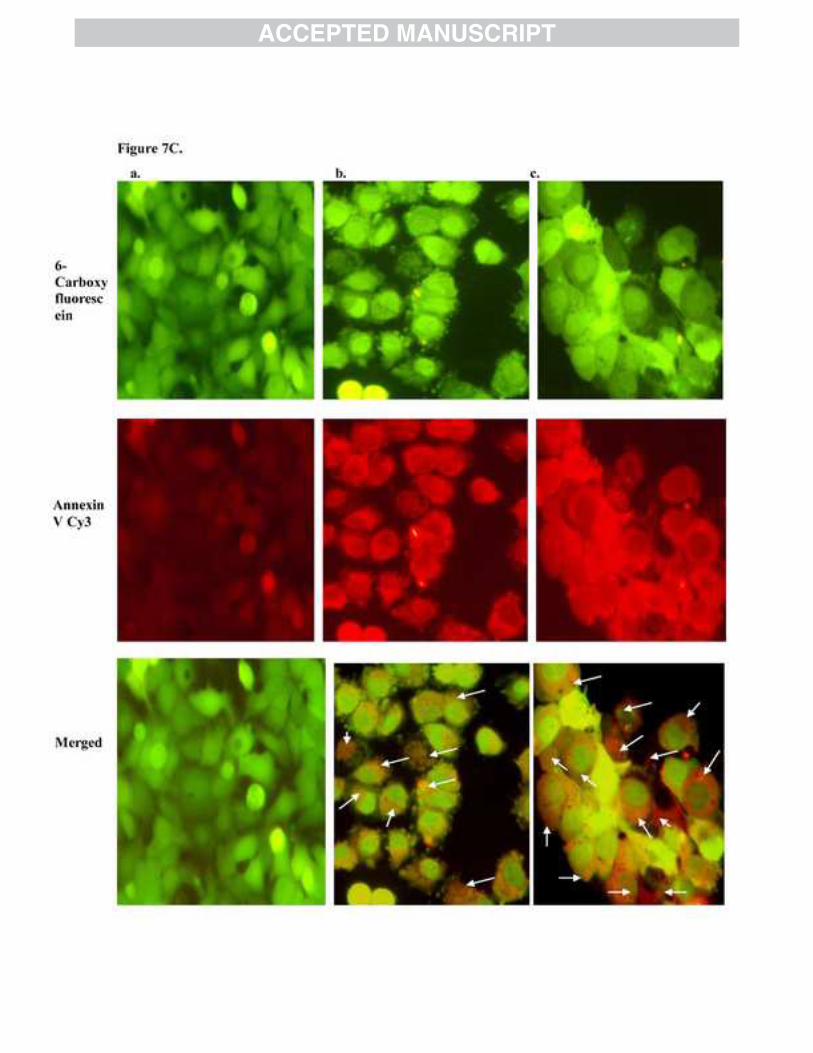
3.5. Study of phosphatidylserine externalization
Early apoptosis is marked by annexin proteins. Annexins are a family of Calcium-dependent
phospholipid-binding proteins, which bind to phosphatidylserine (PS). PS is predominantly
accumulated in the inner leaflet of plasma membrane of living cells. Upon initiation of apoptosis,
PS loses its asymmetric distribution in the phospholipid bilayer and gets translocated from inner
to outer leaflet of plasma membrane and will be available for binding. Cell surface PS is detected
by a preferential binding of PS with protein Annexin V conjugated with Cy3 (AnnCy3), using
annexinV-Cy3 detection kit. The live untreated cells are stained with 6-Carboxy fluorescein
Diacetate (6-CFDA) and emit green fluorescence. The results for treatment of MCF-7 cells with
BDMC-A indicated an early onset of apoptosis and clearly showed an increased externalization

14
of phosphatidylserine in comparison to curcumin treated cells which were double stained with
fluorescently labeled AnnCy3 and 6-CFDA, emitting red and green fluorescence (Figure7C).
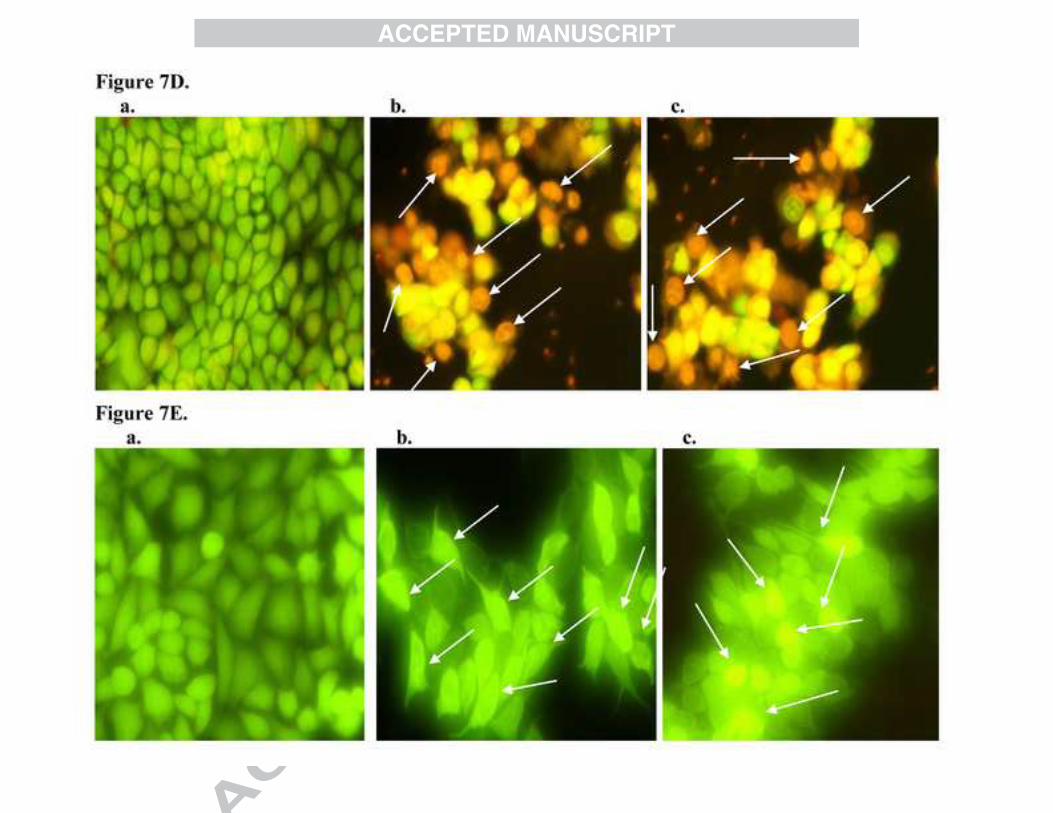
3.6. Fluorescent staining with Acridine Orange/ Ethidium Bromide (AO/EB)
Dual staining with Acridine Orange/Ethidium Bromide is used to evaluate the nuclear
morphology of apoptotic cells. Acridine orange is a vital dye that stains both live and dead cells,
whereas ethidium bromide will stain cells when membrane damage occurs. Cells that stain green
indicates viable cells, yellow indicate early apoptosis and orange/red indicate late apoptosis. In
control, uniformly green live cells were observed with normal and large nucleus. Whereas in
BDMC-A treated cells, a comparative yellow, orange/red staining were observed (Figure7D).
These results confirmed that BDMC-A significantly induced apoptosis in breast cancer cells than
curcumin.
3.7. Reactive oxygen species
Accumulation of ROS can induce apoptosis through both extrinsic and intrinsic pathways. It has
been reported that curcumin can induce apoptosis by inducing ROS formation, which is an
important signal for cytochrome c release from mitochondria into the cytoplasm and subsequent
recruitment of Fas associated protein and caspase-8, that trigger caspase mediated apoptosis in
many types of cancers particularly MCF-7 [42]. In our study, BDMC-A treated cells showed
more ROS production than curcumin which proved that there was a significant induction of
apoptosis in BDMC-A treatment compared to curcumin (Figure7E).
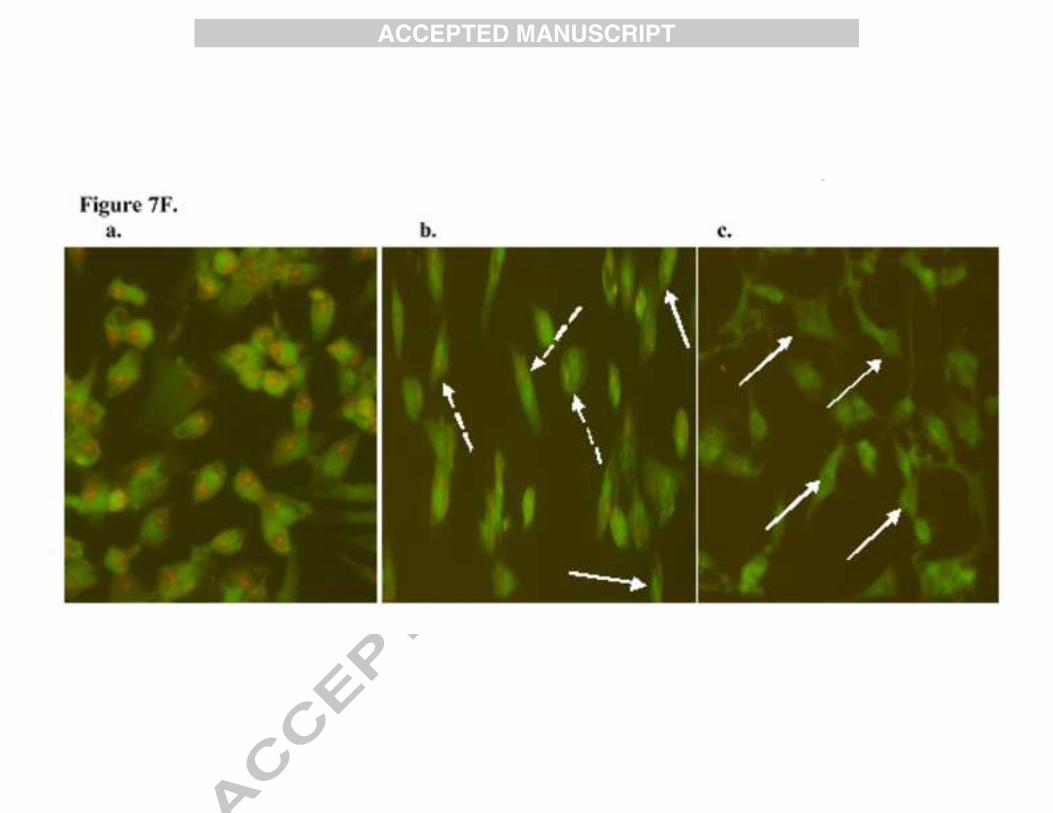
3.8. Assessment of mitochondrial membrane potential (∆Ψm)
Alteration in mitochondrial membrane potential (∆Ψm) (depolarization) is an indication of early
stages of apoptosis. Exit of many apoptogenic factors to the cytosol is facilitated by
mitochondrial depolarization. Rhodamine 123 (Rh123) is a lipophilic cationic dye used as an
indicator of mitochondrial membrane potential and is highly specific for mitochondria.
Depolarized mitochondria are marked by green fluorescence and polarized mitochondria are
marked by orange-red fluorescence. The BDMC-A and curcumin treatment to MCF-7 cells

15
resulted in loss of mitochondrial membrane potential, in which BDMC-A was effective in
inducing membrane depolarization than curcumin (Figure7F).
3.9. Molecular docking of BDMC-A and curcumin with PI3K
To validate our docking protocol, we initially docked the co-crystallized ligand into the protein
and found that the docked conformation generated by the docking program is quite close to the
original co-crystallized ligand conformation. This was quantified by superimposition of the
docked conformation generated by the program to the co-crystallized ligand conformation
extracted from the PDB structure of the protein, and calculating the RMSD (root mean square
deviation) value. For both the targets, the RMSD values were less than 2 Å (Table 1), confirming
that the docking program is able to generate the bioactive conformation of the co-crystallized
ligands successfully and could be reliably used to identify bioactive conformations of other
ligands.
Once the docking protocol was validated using the co-crystallized ligand, docking of curcumin
and BDMC-A was carried out to identify the possible targets for these compounds in the
PI3K/Akt signaling axis. Docking results (Glide score, number of H-Bond and free energy of
binding (ΔG) of curcumin and BDMC-A to the two targets - PI3K & Akt are given in Table 2. It
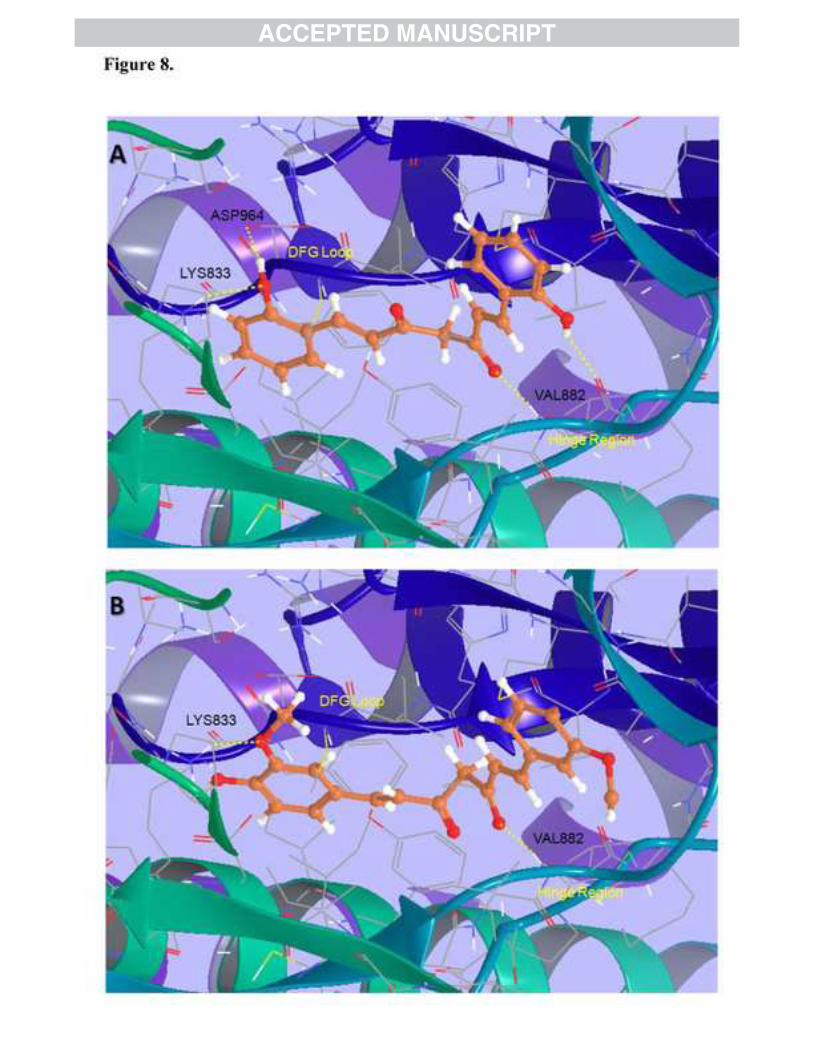
was found that BDMC-A was able to dock to PI3K with a higher Glide Score -9.79 in a manner
similar to that of the co-crystallized ligand (Staurosporine) [43], by making two H-bond
interactions with the back bone carbonyl (-C=O) and NH functions of hinge region residue
VAL882 through the hydroxyl (-OH) and carbonyl groups of the BDMC-A. In addition, the
analog also formed another H-bond with ASP964 similar to co-crystallized ligand (Figure 8A).
On the other hand, curcumin docked to PI3K with a lower Glide Score of -9.69 and formed only
one H-bond with the hinge region residue VAL882 (Figure 8B). However, unlike BDMC-A and
Staurosporine, no H-bond was observed between curcumin and ASP964, which is part of the
DFG loop controlling the conformation of the kinase. Other known PI3K inhibitors such as
wortmannin (PDB: 1E7U) and quercetin (PDB: 1E8W) [40] also interact with hinge region
VAL882 and DFG loop ASP964 residues, in a manner similar to that observed for BDMC-A in
the docking experiments. Hence the docking results with PI3K protein suggests that the

16
BDMC-A interacted with PI3K in a manner similar to known PI3K inhibitors and could inhibit it
better than curcumin, which is further confirmed from the higher binding free energy for BDMC-
A (∆Gbind = -36.34), compared to curcumin (∆Gbind = -21.98), calculated using the MM/PBSA
method (more negative the ∆Gbind value, stronger the binding affinity) (Table 2).
However analysis of the docking pose of curcumin and BDMC-A to Akt protein revealed that
both of them were not able to make the important H-bond interaction with the hinge residue
GLU228 and ALA230 similar to the co-crystallized ligand AZD5363 [44] suggesting that both
of them may not inhibit this target successfully. Moreover, the glide docking score for curcumin
and BDMC-A (-8.03 and -7.04, respectively) were considerably lower, when compared to the
co-crystallized ligand AZD5363 (-13.61). In conclusion, docking results suggest that BDMC-A
could target PI3K better than curcumin and this interaction could be responsible for the
upregulation of the downstream apoptotic proteins leading to potent anti-cancer effect of the
BDMC-A.
Hence in our study, BDMC-A induced apoptosis essentially by engaging in key mitochondrial
events peculiar to intrinsic pathway that resulted in translocation of Bax to mitochondria,
alteration in Bcl-2/Bax ratio, production of reactive oxygen species, drop in mitochondrial
membrane potential, release of mitochondrial proteins like cytochrome c and activation of
caspase-3. The disruption of mitochondrial homeostasis by BDMC-A suggests that it can engage
cell-intrinsic pathway but is not limited to intrinsic pathway. In the extrinsic pathway, BDMC-A
induces apoptosis through death effector domain with caspase 8, an initiator caspase that directly
activates effector caspase importantly caspase-3, that cleaves numerous cell death substrates
resulting in cellular dysfunction and destruction. The present study shows that BDMC-A
induced activation of caspase-3 is mediated by initiation of both extrinsic and intrinsic apoptotic
pathways and proves that BDMC-A is more potent than curcumin in inducing apoptosis.

17
4. Conclusion
Overall, BDMC-A significantly regulates PI3k/Akt, both intrinsic and extrinsic apoptotic
pathways by inhibiting Bcl-2 and inducing p53, Bax, cytochrome c, Apaf-1, FasL, Caspases – 8,
9, 3 and PARP cleavage . Further BDMC-A was efficient than curcumin in regulating most of
the markers studied. This may be due to the presence of ortho hydroxy group in its structure.
These findings suggest that BDMC-A can be a potential therapeutic agent for breast cancer.
Further study on the molecular mechanism is warranted.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgement
The authors acknowledge the financial support by University Grants Commission [F.No 37-
309/2009(SR)] and Dr. Mohane Selvaraj Coumar thank UGC [F.No 41-981/2012 (SR)]. The
authors also thank DST-FIST, UGC-SAP and DBT-IPLS for providing the infrastructural
support.
References
[1] C.P. Prasad, G. Rath, S. Mathur, D. Bhatnagar, R. Ralhan, Potent growth suppressive activity of curcumin in human breast cancer cells: Modulation of Wnt/beta-catenin signaling. Chem. Biol. Interact. 181 (2009) 263-271.
[2] G. Sa, T. Das, Anti cancer effects of curcumin: cycle of life and death, Cell. Div. 3 (2008) 14.
[3] B.B. Aggarwal, C. Sundaram, N. Malani, H. Ichikawa, Curcumin: the Indian solid gold, Adv. Exp. Med. Biol. 595 (2007) 1-75.
[4] W.D. Lu, Y. Qin, C. Yang, L. Li, Effect of curcumin on human colon cancer multidrug resistance in vitro and in vivo, Clinics. 68 (2013) 5. doi: 10.6061/clinics/2013(05)18.
[5] P. Anand, C. Sundaram, S. Jhurani, A.B. Kunnumakkara, B.B. Aggarwal, Curcumin and cancer: an "old-age" disease with an "age-old" solution, Cancer. Lett. 267 (2008) 133- 164.
[6] J. Ravindran, S. Prasad, B.B. Aggarwal, Curcumin and cancer cells: how many ways can curry kill tumor cells selectively?, AAPS J. 11 (2009) 495-510.

18
[7] N.A Abdullah Thani, B. Sallis, R. Nuttall, F.R. Schubert, M. Ahsan, D. Davies, S. Purewal, A. Cooper, H.K. Rooprai, Induction of apoptosis and reduction of MMP gene expression in the U373 cell line by polyphenolics in Aronia melanocarpa and by curcumin, Oncol. Rep. 28 (2012) 1435-1442.
[8] J.M. Brown, B.G. Wouters, Apoptosis, p53, and tumor cell sensitivity to anticancer agents, Cancer. Res. 59 (1999) 1391-1399.
[9] S. Elmore, Apoptosis: a review of programmed cell death, Toxicol.Pathol. 35 (2007) 495-516.
[10] P. Anand, S.G. Thomas, A.B. Kunnumakkara, C. undaram, K.B. Harikumar, B. Sung, S.T. Tharakan, K. Misra, I.K. Priyadarsini, K.N. Rajasekharan, B.B. Aggarwal, Biological activities of curcumin and its analogues (Congeners) made by man and Mother Nature. Biochem Pharmacol, 76 (2008) 1590-1611.
[11] Y.L. Liu, H.P. Yang, L. Gong, C.L. Tang, H.J. Wang, Hypomethylation effects of curcumin, demethoxycurcumin and bisdemethoxycurcumin on WIF-1 promoter in non-small cell lung cancer cell lines, Mol Med Rep. 4 (2011) 675-679.
[12] B. Yadav, S. Taurin, L. Larsen, R.J. Rosengren, RL71, a second-generation curcumin analog, induces apoptosis and downregulates Akt in ER-negative breast cancer cells, Int J Oncol. 41 (2012) 1119-1127.
[13] F. Faião-Flores, J.A. Suarez, P.C. Pardi, D.A. Maria, DM-1, sodium 4-[5-(4-hydroxy-3-methoxyphenyl)-3-oxo-penta-1,4-dienyl]-2-methoxy-phenolate: a curcumin analog with a synergic effect in combination with paclitaxel in breast cancer treatment, Tumour Biol. 33 (2012) 775-785.
[14] R.J. Anto, J. George, K.V. Babu, K.N. Rajasekharan, R. Kuttan, Antimutagenic and anticarcino-genic activity of natural and synthetic curcuminoid, Mutat Res. 370 (1996) 127-131.
[15] R.J. Anto, G. Kuttan, K.V.D. Babu , K.V. Rajasekharan, R. Kuttan, Anti-tumour and free radical scavenging activity of synthetic curcuminoids, Inter J of Pharm. 131 (1996) 1-7.
[16] R. Rukkumani, K, Aruna, P.S. Varma, K.N. Rajasekaran, V.P. Menon, Comparative effects of curcumin and an analog of curcumin on alcohol and PUFA induced oxidative stress, J Pharm Pharm Sci. 7 (2004) 274-283.
[17] R. Rukkumani, K. Aruna, P.S. Varma, K.N. Rajasekaran, V.P. Menon, Comparative effects of curcumin and its analog on alcohol and polyunsaturated fatty acid-induced alterations in circulatory lipid profiles, J Med Food. 8 (2005) 256-260.
[18] R. Rukkumani, K. Aruna, P.S. Varma, P. Viswanathan, K.N. Rajasekaran, V.P. Menon, Protective Role of a Novel Curcuminoid on Alcohol and PUFA-Induced Hyperlipidemia Toxicol Mech Methods, 15 (2005) 227-234.
[19] R. Rajagopalan, S. Sridharan, V.P. Menon, Hepatoprotective role of bis-demethoxy curcumin analog on the expression of matrix metalloproteinase induced by alcohol and polyunsaturated fatty acid in rats. Toxicol Mech Methods. 20 (2010) 252-259.
[20] T. Devasena, K.N. Rajasekaran, V.P. Menon, Bis-1,7-(2-hydroxyphenyl)-hepta-1,6-diene-3,5-dione (a curcumin analog) ameliorates DMH-induced hepatic oxidative stress during colon carcinogenesis, Pharmacol Res. 46 (2002) 39-45.
[21] M.M. Hill, C Adrain, P.J. Duriez, E.M. Creagh, S.J. Martin, Analysis of the composition, assembly kinetics and activity of native Apaf-1 apoptosomes, EMBO. J. 23 (2004) 2134-2145.

19
[22] M. Li, Z. Zhang, D.L. Hill, H. Wang, R. Zhang, Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway, Cancer. Res. 67 (2007) 1988-1996.
[23] M.S. Squires, E.A. Hudson, L. Howells, S. Sale, C.E. Houghton, J.L. Jones, L.H. Fox, M. Dickens, S.A. Prigent, M.M. Manson, Relevance of mitogen activated protein kinase (MAPK) and phosphotidylinositol-3-kinase/protein kinase B (PI3K/PKB) pathways to induction of apoptosis by curcumin in breast cells. Biochem. Pharmacol. 65 (2003) 361-376.
[24] L.R.Chaudhary, K.A. Hruska, Inhibition of cell survival signal protein kinase B/Akt by curcumin in human prostate cancer cells, J. Cell. Biochem. 89 (2003) 1-5.
[25] K.V. Dinesh Babu, K.N. Rajasekharan, Simplified conditions for the synthesis of curcumin I and other curcuminoids, Org.Prep.Proc.Int. 24 (1994) 674-677.
[26] R.J. Fido, A.S. Tatham, P.R. Shewry,Western blotting analysis. Methods in Molecular Biology: Plant gene transfer and expression protocols, Jones H. (Ed.). 49 (1995) 423-437.
[27] P. Chomczynski, N. Sacchi, Single-step method of RNA isolation by acid Guanidinium thiocyanate–phenol–chloroform extraction, Anal. Biochem. 162. (1987) 156–159.
[28] M. Kumaravel, P. Sankar, R. Rukkumani, Antiproliferative effect of an analog of curcumin bis-1,7-(2-hydroxyphenyl)-hepta-1,6-diene-3,5-dione in human breast cancer cells, Eur. Rev. Med. Pharmacol. Sci. 16 (2012) 1900-1907.
[29] C. Brana, C. Benham, A method for characterising cell death in vitro by combining propidium iodide staining with immunoistochemistry, Brain. Res.Protoc. 10 (2002) 109-114.
[30] C.A.Castaneda,H. Cortes-Funes, H.L. Gomez, E.M. Ciruelos, The phosphatidyl inositol 3-kinase/AKT signaling pathway in breast cancer, Cancer. Metastasis. Rev. 29 (2010) 751-759.
[31] X.L. Xu, S.Z. Chen, W. Chen, W.H. Zheng, X.H. Xia, H.J. Yang, B. Li, W.M. Mao, The impact of cyclin D1 overexpression on the prognosis of ER-positive breast cancers: a meta-analysis, Breast. Cancer. Res. Treat. 139 (2013) 329-339.
[32] R. Arunkumar, G. Sharmila, P. Elumalai, K. Senthilkumar, S. Banudevi, D.N. Gunadharini, C.S. Benson, P. Daisy, J. Arunakaran, Effect of diallyl disulfide on insulin like growth factor Signaling molecules involved in cell survival and proliferation of human prostate cancer cells in vitro and in silico approach through docking analysis, Phytomed. 19 (2012) 912-923.
[33] N. Singh, D. Zaidi, H. Shyam, R. Sharma, A.K. Balapure, Polyphenols sensitization potentiates susceptibility of MCF-7 and MDA MB-231 cells to Centchroman, PLoS One. 7 (2012) e37736. doi: 10.1371/journal.pone.0037736.
[34] S. Haupt, M. Berger, Z. Goldberg, Y. Haupt, Apoptosis - the p53 network, J. Cell. Sci. 116 (2003) 4077-4085.
[35] T. Miyashita, J.C. Reed, Tumor suppressor p53 is a direct transcriptional activator of the human bax gene, Cell. 80 (1995) 293-299.
[36] K. Polyak, T. Waldman, T.C. He, K.W. Kinzler, B. Vogelstein B, Genetic determinants of p53-induced apoptosis and growth arrest, Genes. Dev. 10 (1996) 1945-1952.

20
[37] S. Kondo, Y. Tamura, J.W. Bawden, S. Tanase, The immunohistochemical localization of Bax and Bcl-2 and their relation to apoptosis during amelogenesis in developing rat molars, Arch. Oral. Biol. 46 (2001) 557-568.
[38] Jr. Reiners, J.A. Caruso, P. Mathieu, B. Chelladurai, X.M. Yin, D. Kessel, Release of cytochrome c and activation of pro-caspase-9 following lysosomal photodamage involves Bid cleavage, Cell. Death. Differ. 9 (2002) 934-944.
[39] S. Uddin, A.R. Hussain, P.S. Manogaran, K. Al-Hussein, L.C. Platanias, M.I. Gutierrez, K.G. Bhatia, Curcumin suppresses growth and induces apoptosis in primary effusion lymphoma, Oncogene. 24 (2005) 7022-7030.
[40] K.W. Sun, Y.Y. Ma, T.P. Guan, Y.J. Xia, C.M. Shao, L.G. Chen, Y.J. Ren, H.B. Yao, Q. Yang, X.J. He, Oridonin induces apoptosis in gastric cancer through Apaf-1, cytochrome c and caspase-3 signaling pathway, World. J. Gastroenterol. 18 (2012) 7166-7174.
[41] H.F. Lu, K.C. Lai, S.C. Hsu, H.J Lin, M.D. Yang, Y.L. Chen, M.J. Fan, J.S. Yang, P.Y. Cheng, C.L. Kuo, J.G. Chung, Curcumin induces apoptosis through FAS and FADD, in caspase-3-dependent and -independent pathways in theN18 mouse-rat hybrid retina ganglion cells. Oncol. Rep. 22 (2009) 97-104.
[42] T. Ozben, Oxidative stress and apoptosis: impact on cancer therapy, J. Pharm. Sci. 96 (2007) 2181-2196.
[43] E.H. Walker, M.E. Pacold, O. Perisic, L. Stephens, P.T. Hawkins, M.P. Whymann, R.L.Williams, Structural determinants of phosphoinositide 3-kinase inhibition by wortmannin, LY294002, quercetin, myricetin, and staurosporine, Mol. Cell. 6 (2000) 909-919.
[44] M. Addie, P. Ballard, D. Buttar, C. Crafter, G. Currie, B.R. Davies, J. Debreczeni, H. Dry, P. Dudley, R. Greenwood, P.D. Johnson, J.G. Kettle, C. Lane, G. Lamont, A. Leach, R.W. Luke, J. Morris, D. Ogilvie, K. Page, M. Pass, S. Pearson, L. Ruston, Discovery of 4-amino-N-[(1S)-1-(4-chlorophenyl)-3-hydroxypropyl]-1-(7H-pyrrolo[2,3-d]pyrimidin-4-yl)piperidine-4-carboxamide (AZD5363), an orally bioavailable, potent inhibitor of Akt kinases, J. Med. Chem. 56 (2013) 2059-2073.

21
Figure and Table Captions
Table 1. Validation of docking protocol for the two targets - PI3K & Akt.
Table 2. Comparison of docking results of curcumin and analog (BDMC-A) to the two targets - PI3K & Akt.
Figure 1: (A) Structure of BDMC-A (B) Structure of Curcumin.
Figure 2: PI3K/AKT markers (A) Effect of BDMC-A and Curcumin on expression of PI3K/AKT markers. Cells were plated and incubated with 15 µM curcumin and 15 µM BDMC-A for 24h. After 24h whole cell extracts were prepared and resolved on 12% SDS-PAGE, electro transferred and probed with antibody. β-Actin is used as internal loading control. (B) Densitometry. Protein levels were quantified using densitometry analysis and expressed in relative band intensity. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments. ANOVA followed by Tukey’s test was used to assess the statistical significance between groups. *p≤0.05 level, Significance relative to control group.
Figure 3: (A&B) Time and Dose dependent expression of p53 and Cytochrome c. MCF-7 cells were plated and incubated with 15 µM & 30 µM of curcumin and 15 µM & 30 µM of BDMC-A for 12 (A) and 24 hours (B). After 12h & 24h whole cell extracts were prepared and resolved on 12% SDS-PAGE, electrotransferred and probed with antibody. β-Actin is used as internal loading control. (C and D) Densitometry for 12h and 24h treatment respectively. Protein levels were quantified using densitometry analysis and expressed in relative band intensity. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments. ANOVA followed by Tukey’s test was used to assess the statistical significance between groups. *p≤0.05 level, Significance relative to control group.
Figure 4: Western Blot for Intrinsic apoptotic markers (A) Effect of BDMC-A and curcumin on expression of intrinsic apoptotic proteins. Cells were plated and incubated with 15 µM curcumin and 15 µM BDMC-A for 24h. After 24h whole cell extracts were prepared and resolved on 12% SDS-PAGE, electro transferred and probed with antibody. β-Actin is used as internal loading control. (B) Densitometry. Protein levels were quantified using densitometry analysis and expressed in relative band intensity. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments considered at the p ≤ 0.05 level. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments. ANOVA followed by Tukey’s test was used to assess the statistical significance between groups. *p≤0.05 level, Significance relative to control group.
Figure 5: Reverse Transcriptase-PCR (A&B) Effect of BDMC-A and curcumin on mRNA expression of Bcl-2 and Bax. Cells were plated and incubated with 15 µM curcumin and 15 µM BDMC-A for 24h. After 24h, whole cell extracts were prepared using TRI solution. The mRNA

22
expression for antiapoptotic Bcl-2 and pro-apoptotic Bax were analysed by Reverse transcriptase PCR (RT-PCR) with Agarose Gel electrophoresis. GAPDH is used an internal loading control. (C) Densitometry. mRNA levels were quantified using densitometry analysis and expressed as relative intensity. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments. ANOVA followed by Tukey’s test was used to assess the statistical significance between groups. *p≤0.05 level, Significance relative to control group.
Figure 6: Extrinsic apoptotic markers (A) Effect of BDMC-A and curcumin on expression of extrinsic apoptotic proteins. Cells were plated and incubated with 15 µM curcumin and 15 µM BDMC-A for 24h. After 24h whole cell extracts were prepared and resolved on 12% SDS-PAGE, electrotransferred and probed with antibody. β-Actin is used as internal loading control. (B) Densitometry. Protein levels were quantified using densitometry analysis and expressed in relative band intensity. Values are expressed as mean ± Standard Deviation (SD) of three independent experiments. ANOVA followed by Tukey’s test was used to assess the statistical significance between groups. *p≤0.05 level, Significance relative to control group.
Figure 7: Morphological Analysis for Detection of Apoptosis
Figure 7A: Hoechst 33258 staining: Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a-Control, b-Curcumin and c-BDMC-A. Morphologic analysis of nuclear chromatin of cell undergoing apoptosis and fragmented apoptotic bodies were examined by Hoechst 33258 staining. Condensed or fragmented nuclei were observed under a fluorescence microscope.
Figure 7B: Propidium Iodide staining: Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a. Control, b. Curcumin and c. BDMC-A. Morphologic analysis of nuclear chromatin of cell undergoing apoptosis and fragmented apoptotic bodies were examined by Propidium Iodide staining. Condensed or fragmented nuclei were observed under a fluorescence microscope. Figure 7C: Annexin V staining: Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a. Control, b. Curcumin and c. BDMC-A. Phosphatidyl externalization was observed with Annexin V. 6-Carboxyfluorescein stains green, cy-3 stains red and merged stain orange red denotes early apoptotic stage. Figure 7D: Acridine Orange / Ethidium Bromide Staining: Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a. Control, b. Curcumin and c. BDMC-A. Morphologic analysis of nuclear chromatin of cell undergoing apoptosis. Live cells stain green, yellow- the early apoptotic and orange - the late apoptotic stages were examined by AO/EB staining.
Figure 7E: Production of Reactive oxygen species (ROS): Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a. Control, b.

23
Curcumin and c. BDMC-A. Production of reaction oxygen species were examined by DCFH-DA stain.
Figure 7F: Rhodamine 123 for mitochondrial membrane potential: Induction of apoptosis by BDMC-A and curcumin in MCF-7 cells at 24h in fixed concentration. a. Control, b. Curcumin and c. BDMC-A. Depolarization of mitochondrial membrane potential was observed as green fluorescent and polarized mitochondria as orange/red fluorescent. Dashed arrows indicate cells containing polarized mitochondria and Solid arrows indicate cells containing depolarized mitochondria.
Figure 8: Molecular Docking (A) Docking pose of BDMC-A to PI3K (PDB: 1E8Z). Total of four H-bonds are formed between the protein and BDMC-A: Two H-bonds with hinge region residue VAL882, one with DFG loop ASP964 and another with Lys833 residue. Glide Score: -9.79. (B) Docking pose of Curcumin to PI3K (PDB: 1E8Z). Total of two H-bonds are formed between the protein and Curcumin: One H-bond with hinge region residue VAL882 and one with Lys833 residue. Glide Score: -9.69.
24
Table 1.
Sl. No. Target PDB ID Co-Crystallized Ligand Glide Score RMSD (Å)
1 PI3K 1E8Z Staurosporine -10.22 0.88
2 Akt 4GV1 AZD5363 -13.61 1.87
Table 2.
Sl. No. Target Parameters Curcumin BDMC-A
Glide score -9.69 -9.79
No of H-bond 2 4
1 PI3K
(PDB: 1E8Z)
ΔGbind -21.98 -36.34
Glide score -8.03 -7.04
No of H-bond 1 1
2 Akt
(PDB: 4GV1)
ΔGbind -21.90 -11.62

25
Highlights:
1.Anticancer effect of BDMC-A was more potent than curcumin in breast cancer
cells
2. BDMC-A induced apoptosis through both intrinsic and extrinsic pathways
3. We demonstrated the interactions of BDMC-A and curcumin with PI3K in silico




















