
Diffusion alterations in corpus callosum of patients with HIV
Upload
independentCategory
view
0download
0
The nature and extent of callosal morphological alterations inschizophrenia remain unresolved. A parametric surface modelingapproach using magnetic resonance (MR) images was employed.This provided spatially accurate representations of midsagittalcallosal surfaces in schizophrenic patients (n = 25; 15 males) andnormal controls (n = 28; 15 males). Areas of functionally relevantcallosal channels and measures reflecting callosal shape werevisualized and compared across groups. To register neuroanatomicallandmarks surrounding the corpus callosum, each three-dimensionalMR volume was scaled according to Talairach AC–PC distance, andraw distances included as covariates in multivariate analyses.Results revealed: (i) a marked vertical displacement of the corpuscallosum in patients (P < 0.01); (ii) increases in curvature ofsuperior and inferior callosal surfaces (P < 0.001); and (iii) signif-icant increases in maximum widths in anterior and posterior regionsin male patients compared to male controls; as well as (iv) increasedpatterns of callosal variability in female patients but no effects ofdiagnosis between female groups. These findings demonstrate aclear index of structural neuropathology in male schizophrenicpatients. Displacement and curvature increases were highly cor-related with structural differences in surrounding neuroanatomicalregions, including increased volume of the lateral ventricles (P <0.01).
IntroductionThe corpus callosum plays an integral role in relaying sensory,
motor and cognitive information from homologous regions in
the two cerebral hemispheres. Given that cognitive impairments
and both bilateral and unilateral neuroanatomic structural
abnormalities are characteristic in schizophrenia, the mor-
phology of the corpus callosum has stimulated much research
(Nasrallah et al., 1986; Uematsu et al., 1988; Rossi et al., 1989;
Woodruff et al., 1993, 1997; Jacobsen et al., 1997). Most
investigations have used ‘region of interest’ analyses to assess
differences in midsagittal area, callosal length and width
(Woodruff et al., 1995), while few have investigated group
differences in callosal shape or in three-dimensional location
(Casanova et al., 1990; DeQuardo et al., 1996). Volumetric
methods are less sensitive to across-subject variability and less
likely capture subtle differences in anatomy between groups.
Probabilistic brain atlases using magnetic resonance (MR)
images retain quantitative information on interindividual neuro-
anatomical variability. In these approaches three-dimensional
brain volumes are registered (realigned) in a standardized
coordinate system to allow cross-group comparisons while
preserving subtle intragroup variability patterns (Mazziotta et
al., 1995; Thompson et al., 1997, 1999a,b).
Several observations motivate the study of the corpus
callosum in schizophrenia (Thompson et al., 1999a): (i) callosal
pruning and myelination as well as interhemispheric coherence
continue to develop into early adulthood, a factor that may be
relevant to age of onset in schizophrenia (Njiokiktjien et al.,
1994); (ii) impairments in callosal transfer have been reported in
patients (David et al., 1994), implicating alterations in callosal
connectivity; (iii) structural alterations in asymmetric peri-
sylvian regions linked by the callosum have been reported
(Kikinis et al., 1994; Barta et al., 1997); (iv) callosal myelination
begins prenatally and is susceptible to malnutrition, asphyxia
and toxins of infectious origin; also, these same events are linked
with aberrant neurodevelopmental events in schizophrenia
(Njiokiktjien et al., 1994; Bishop and Wahlsten, 1997; Hoek et
al., 1998); (v) the corpus callosum forms the roof of the superior
horns of the lateral ventricles, which are enlarged in schizo-
phrenic patients (Sharma et al., 1997, 1998). In spite of evidence
linking callosal abnormality to schizophrenia, imaging studies
assessing alterations in callosal morphometry as well as in other
cortical and subcortical structures have produced surprisingly
mixed results (Woodruff et al., 1995). Interpretation of these
results is further obscured by the failure of many studies to
control for brain size in patient and normal populations. Since
callosal size and brain size are correlated (Rauch and Jinkins,
1994; Jäncke et al., 1997), and brain size is reported to be
decreased in MR studies of schizophrenic patients (Zipursky et
al., 1992; Lawrie and Abukmeil, 1998), it is clear headsize
corrections are necessary.
A surface deformation-based approach was employed in this
study to allow quantification of standard callosal parameters
such as area, length and width in addition to other parameters
indexing shape (Thompson et al., 1996a,b, 1997). This approach
was aimed at providing information about structural alterations
in surrounding brain regions in addition to assessing callosal
abnormalities. After some initial scaling to bring callosal surfaces
into register and to correct for differences in overall brain
dimensions, curves and points from the callosal surfaces of
subjects in each group were matched and variability information
from each individual surface warp retained. The scaling ratios
used to align the corpus callosum across subjects can be factored
out either by converting measures back into native space or by
using them as covariates in a multivariate analysis, thereby
ensuring that callosal measures are not affected by brain volume.
These techniques allow a comprehensive analysis of callosal
structure in schizophrenic patients and normal controls and
provide the ability to quantify and visualize any structural
differences as they relate to diagnosis and sex in a three-
dimensional coordinate system in standardized and native
scanner space. Furthermore, callosal shape renderings from
Alzheimer’s patients (n = 10 males) acquired using the same
methods in an earlier study (Thompson et al., 1998) were
compared with the callosal shape renderings of male schizo-
phrenic patients and controls to evaluate whether callosal shape
profiles in schizophrenia are disease specific.
Cerebral Cortex Jan 2000;10:40–49; 1047–3211/00/$4.00
Mapping Morphology of the CorpusCallosum in Schizophrenia
Katherine L. Narr, Paul M. Thompson, Tonmoy Sharma1,
Jacob Moussai, Andrew F. Cannestra and Arthur W. Toga
Laboratory of Neuro Imaging, Department of Neurology,
Division of Brain Mapping, UCLA School of Medicine, Los
Angeles, CA, USA and 1Section of Cognitive
Psychopharmacology, Institute of Psychiatry, Denmark Hill,
London, UK
© Oxford University Press 2000

Hypotheses
We set out to detect a modest decrease in midsagittal area in
schizophrenic patients, looking specifically for area differences
implicating involvement of specific callosal channels as defined
by Witelson et al. (Witelson et al., 1989) and Clarke et al. (Clarke
et al., 1994). Furthermore, we were interested in assessing
alterations in callosal shape measured by curvature and dis-
placement in three-dimensional space in patients versus
controls, which would provide further hypotheses concerning
structural differences in surrounding anatomical regions.
Specifically, relationships between callosal displacement and
lateral and third ventricular enlargement were assessed.
Materials and Methods
Subjects
Twenty-five chronic schizophrenic patients (15 male/10 female; mean
age 31.1 ± 5.6 years) and 28 normal controls (15 male/13 female; mean
age 30.5 ± 8.7 years) were included in the study. Subjects were scanned at
the Institute of Psychiatry, London, UK. Groups did not differ significantly
in age, years of education, height or parental socio-economic class.
Socio-economic status was derived from UK census data (Office of
Population Censuses and Surveys, 1991), using details of ‘best-ever’
parental occupation. There were two left-handed subjects in each group
as defined by the Annett Handedness Scale (Annett, 1970).
All schizophrenic patients met DSM-III-R (American Psychiatric
Association, 1987) criteria and were receiving regular antipsychotic
medication. Control subjects were screened for any personal or family
history of psychiatric illness. Exclusion criteria for both patients and
controls included head trauma, drug abuse and hereditary neurological
disorders. All subjects gave informed written consent for their
participation in this study with ethical permission obtained from the
Bethlem and Maudsley Ethical Committee (Research).
Image Analysis Procedures
High-resolution three-dimensional SPGR MR images (256 × 256 matrix;
20 cm FOV) were acquired using a GE Signa 1.5 T scanner for each
subject as a series of 124 contiguous 1.5 mm coronal slices. Images were
resliced in the sagittal plane at 0.5 mm. To correct for head position and
brain length and to allow for morphometric comparison of the corpus
callosum between subjects, each brain volume was scaled and placed into
the standardized Talairach and Tournoux stereotaxic coordinate system
(Talairach and Tournoux, 1988). In the scaling procedure x, y and z pixel
coordinates were chosen for the anterior commisure (AC), posterior
commisure (PC) and midline. The scan is digitally realigned to place the
AC at the origin (0,0,0) and the PC at (–23.5,0,0) with the inter-
hemispheric fissure aligned vertically at midline. This process brings
midline structures into register according to Talairach AC–PC distance
and provides sufficient alignment to compare corpus callosum para-
meters, and derive precise statistical information about positional and
geometric variability (Thompson et al. 1996a,b, 1997) (Figs 1–5).
The corpus callosum was contoured by technicians blind to diagnosis
in each magnified midsagittal slice by following tissue boundaries using a
mouse-driven cursor. Midsagittal sections were defined by identifying the
interhemispheric fissure in native contiguous 1.5 mm coronal slices and
again in the sagittal plane when volumes were resliced at 0.5 mm after
AC–PC scaling. Coronal midlines were chosen independently by two
investigators and confirmed by the presence of the falx cerebri and
septum pellucidum in the sagittal planes. The reslicing procedure
ensured that callosi were contoured at midline within the limits of our
high-resolution MR data. Reliability of contouring was evaluated by
repeatedly outlining the corpus callosum (six times) in a randomly
chosen brain (contouring r.m.s. error < 1 mm).
Callosal surfaces were reconstructed and within- and between-group
variations in callosal shape parameters quantified using a surface-based
anatomical modeling approach (Thompson et al., 1996a,b, 1997). Brief ly,
algorithms generate a parametric grid after uniformly redigitizing points
from the callosal outline of each three-dimensional data set to form a
regular rectangular mesh that is stretched over the callosal surface
contour. To obtain average parametric meshes and measures of variability
in the shape of the callosal surfaces within each group, the parametric
grids from each surface are matched at corresponding points and the
mean and variation between these sets of points are calculated. The
surface mesh algorithms thus create average representations of callosal
surfaces in three-dimensional space and index the amount of variability
present within each group (Figs 1–4; Fig. 6). To visualize and quantify
regional displacement of average callosal surfaces between groups,
algorithms were used whereby corresponding grid points were matched
and the discrepancy in distance calculated as three-dimensional dis-
placement vectors between points (Fig. 5). Callosum meshes were
divided into superior and inferior pieces to generate surface mesh varia-
bility and displacement maps. Measures of length, width, curvature and
all three-dimensional extents of the corpus callosum were obtained from
the uniformly redigitized grid points in pixel coordinates translated into
millimeters.
Finally, midsagittal callosal renderings where divided into five vertical
partitions representing the (i) anterior third, (ii) anterior body, (iii)
posterior body, (iv) isthmus and (v) splenium (Fig. 7) (Witelson et al.,
1989; Clarke and Zaidel, 1994). Midsagittal areas were obtained in mm2
for the entire callosum and for each midsagittal segment.
Brain and Ventricle Volume
Total brain volume and volumes of the lateral and third ventricles were
acquired by segmenting each three-dimensional image set into gray and
white matter, cerebrospinal f luid (CSF) and background. Brief ly, image
data (16 bit) was processed using a number of steps that included: (i)
correction for signal intensity inhomogeneities; (ii) tissue classification by
choosing representative signal intensity values for the different tissue
types from 120 predefined regions across the entire brain volume (Sowell
et al., 1999); (iii) removal of the skull, cerebellum and extracortical CSF;
and (iv) calculation of brain volume in native space. Reliability of
selection for representative tissue type intensity values was evaluated by
classifying 10 different brains and computing intensity errors (reliability
> 94% for all tissue types). Volume of the lateral and third ventricles were
measured by outlining the regions of interest in consecutive tissue
segmented coronal slices by following anatomical landmarks and
restricted to boundaries between CSF and other tissue types (Fig. 8).
Alzheimer’s Data
To compare the shape profiles of midsagittal callosal surface renderings
between male schizophrenic patients (n = 15), adult controls (n = 15) and
Alzheimer’s patients (n = 10), Alzheimer’s data was included from a
study assessing cortical variability in normal aging and Alzheimer’s
disease (Thompson et al., 1998). Image analysis procedures employed
in this study were identical to those used here and callosal shape para-
meters were measured in the same stereotaxic coordinate system. The
rationale for including this data was to evaluate whether Alzheimer’s
patients exhibit similar differences in callosal shape profiles given that
Alzheimer’s patients show ventricular enlargement and decreases in brain
size that are similarly reported in schizophrenia (Lawrie and Abukmeil,
1998). Callosal renderings from all male groups are overlaid for com-
parison in Figure 9.
Statistical Analyses
Differences in discrete callosal areas and morphometric shape parameters
obtained from the mesh pieces were compared within and between
groups using multivariate analyses of covariance (MANCOVAs) and, when
appropriate, followed by univariate analyses. Multivariate test statistics
control for type I error by adopting a strict criterion for significance. In
order to control for type I error, tests of individual dependent variables
were only performed if the corresponding multivariate tests were found
significant. In these analyses Sex and Diagnosis were used as independent
variables, while curvature and superior and inferior extents of top and
bottom callosal surfaces, length, width and area measures were used as
dependent variables. Talairach scaling ratios used to normalize AC–PC
distance and brain volume were assessed as potential covariates. Results
of these analyses, summarized in Table 1, are described below.
ResultsSeveral striking trends emerged in the callosal morphometric
Cerebral Cortex Jan 2000, V 10 N 1 41
analyses between groups: (i) different patterns of variability
were observed in each group with female schizophrenic patients
showing the highest variability overall; (ii) significant vertical
displacement of the corpus callosum was observed in male
patients compared to controls; (iii) male patients exhibited a
significant increase in curvature for superior and inferior
callosal surfaces; and (iv) increases in callosal thickness were
identified in anterior and posterior callosal regions in male
patients versus male controls. Finally, callosal shape measures
were significantly related to lateral ventricle enlargement.
Table 1Summary of significant results
Dependent variable Effect Mean ± SEM Probability
Raw brain volume (univariate effects) Sex F: 1161.5 ± 18.1 M: 1270.9 ± 21.5 <0.001w/ covariate Sex <0.01AC–PC distance (univariate effects) Sex M: 26.83 ± 0.24 F: 25.68 ± 0.29 <0.006Midsagittal area (multivariate effects) AC–PC covariate <0.001Shape parameters (multivariate effects) Diagnosis <0.053
Diagnosis × Shape variable <0.001Diagnosis × Sex × Shape <0.008AC–PC <0.001AC–PC by Shape <0.001
Length (univariate effects) AC–PC Distance <0.001Top superior boundary a AC-PC distance <0.001
Diagnosis NC: 24.84 ± 0.62 SZ: 26.78 ± 0.58 <0.009Diagnosis × Sex <0.005Male patients > male controls NC M: 23.27 ± 0.56 SZ M: 27.38 ± 0.67 <0.001Diagnosis × Sex w/o Covariate <0.05
Bottom superior boundarya AC-PC distance <0.001Diagnosis NC: 19.67 ± 0.57 SZ: 21.44 ± 0.55 <0.016Diagnosis × Sex <0.008Male patients > male controls NC M: 18.29 ± 0.52 SZ M: 22.03 ± 0.73 <0.001
Inferior rostrum Diagnosis NC: 2.78 ± 0.28 SZ: 2.84 ± 0.34 <0.033Top curvaturea Diagnosis × Sex <0.001
Male patients > male controls NC M: 1.29 ± 0.01 SZ M: 1.33 ± 0.01 <0.001Bottom curvaturea Diagnosis × Sex <0.004
Male patients > male controls NC M: 1.50 ± 0.02 SZ M: 1.63 ± 0.02 <0.001Width (multivariate effects) AC–PC distance <0.001
Diagnosis × Sex <0.004Diagnosis × Width <0.009
Anterior segment (univariate effects) Diagnosis NC: 19.88 ± 0.72 SZ: 21.94 ± 0.82 <0.006Diagnosis × Sex <0.016Male patients > male controls NC M: 18.63 ± 0.76 SZ M: 22.71 ± 0.73 <0.05
Splenium Male patients > male controls NC M: 15.05 ± 0.16 SZ M: 16.70 ± 0.56 <0.001Posterior midbody Male patients > male controls NC M: 5.97 ± 0.25 SZ M: 7.07 ± 0.26 <0.006Ventricle volume (multivariate effects) Diagnosis × Sex <0.01w/ brain volume covariate Diagnosis × Sex × Volume <0.02
Brain volume covariate <0.05Lateral ventriclesa (univariate effects) Diagnosis × Sex <0.01
Male patients > male controls NC M: 10.70 ± 1.60 SZ M: 20.17 ± 2.65 <0.001Third ventricle Diagnosis × Sex <0.03w/o covariate Male patients > male controls NC M: 1.86 ± 0.13 SZ M: 2.58 ± 0.24 <0.01
SZ, schizophrenic patients; NC; normal controls; M, males; F, females. Multivariate analyses were performed with Sex and Diagnosis as factors, and included callosal areas, shape parameters or ventricularvolumes as dependent variables using AC–PC distance or raw brain volume as covariates. Univariate analyses were only performed if the corresponding multivariate test was significant.
Measures of volumes are reported in cm3, and distance measures are in mm.aSignificant correlations between lateral ventricle volume and callosal shape parameters (P < 0.01).
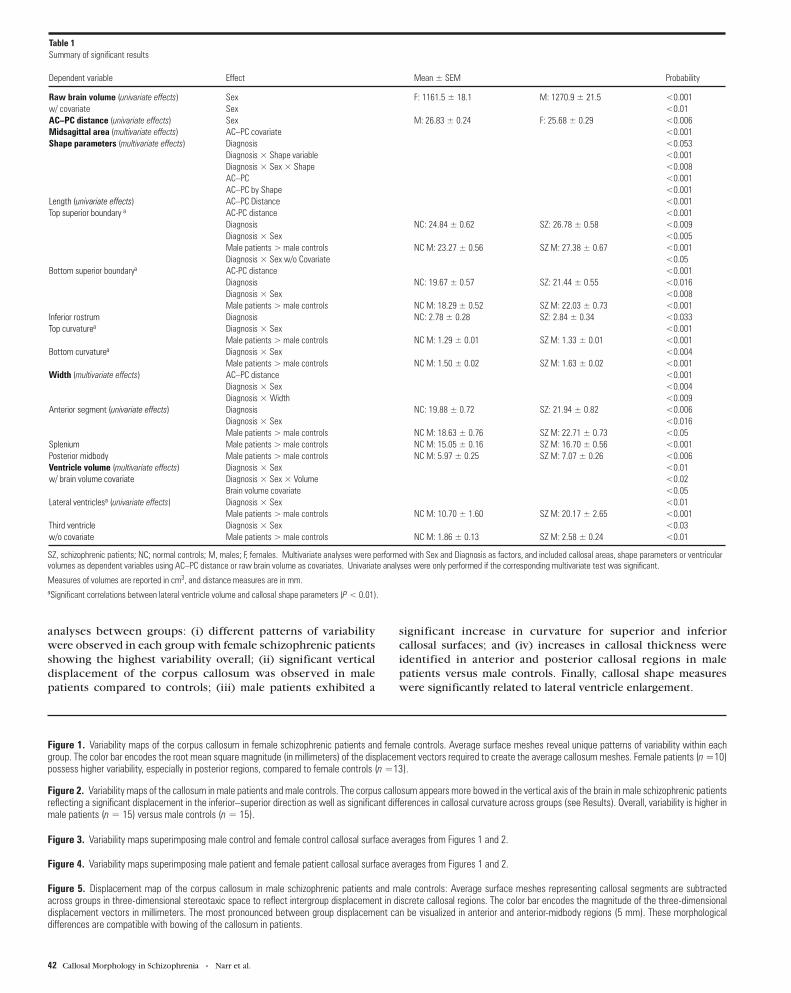
Figure 1. Variability maps of the corpus callosum in female schizophrenic patients and female controls. Average surface meshes reveal unique patterns of variability within eachgroup. The color bar encodes the root mean square magnitude (in millimeters) of the displacement vectors required to create the average callosum meshes. Female patients (n =10)possess higher variability, especially in posterior regions, compared to female controls (n =13).
Figure 2. Variability maps of the callosum in male patients and male controls. The corpus callosum appears more bowed in the vertical axis of the brain in male schizophrenic patientsreflecting a significant displacement in the inferior–superior direction as well as significant differences in callosal curvature across groups (see Results). Overall, variability is higher inmale patients (n = 15) versus male controls (n = 15).
Figure 3. Variability maps superimposing male control and female control callosal surface averages from Figures 1 and 2.
Figure 4. Variability maps superimposing male patient and female patient callosal surface averages from Figures 1 and 2.
Figure 5. Displacement map of the corpus callosum in male schizophrenic patients and male controls: Average surface meshes representing callosal segments are subtractedacross groups in three-dimensional stereotaxic space to reflect intergroup displacement in discrete callosal regions. The color bar encodes the magnitude of the three-dimensionaldisplacement vectors in millimeters. The most pronounced between group displacement can be visualized in anterior and anterior-midbody regions (5 mm). These morphologicaldifferences are compatible with bowing of the callosum in patients.
42 Callosal Morphology in Schizophrenia • Narr et al.
Cerebral Cortex Jan 2000, V 10 N 1 43

Variability and Displacement Maps
Variability maps revealed distinct differences in the patterns of
local variability and shape in each group (Figs 1–4). The average
callosal surface mesh appeared considerably more bowed in the
male schizophrenic group (Fig. 2). Overall, female schizophrenic
patients displayed higher intragroup variability in callosal
parameters than male patients or controls (∼4.5 mm) (Figs 1 and
4). Male patients exhibited higher callosal variability compared
with male controls (Fig. 2). Maximal displacement of callosal
variability averages was visualized in anterior, superior and
inferior midbody regions between male patients and controls,
approaching 5 mm (Fig. 5).
Covariates
Raw AC–PC distances (anterior–posterior axis) and brain volume
were evaluated as potential covariates to remove the effects of
head size and of AC–PC scaling from the data. Preliminary
analyses found AC–PC scaling to be highly correlated with
midsagittal callosal area and shape parameters after normal-
ization (average r = 0.48, P < 0.005). Brain volume, however, did
not correlate significantly with any of the AC–PC scaled
dependent variables (correlations did not exceed 0.15 for any
variable, P > 0.3); thus only raw AC–PC distance was used as a
covariate in subsequent analyses of callosal parameters.
To determine whether the four groups differed in average
values on the AC–PC covariate or brain volume, the AC–PC
scaling measure and brain volumes in native space were used as
dependent variables in separate 2 (Sex) × 2 (Diagnosis) ANOVAs.
There was a significant effect of Sex on AC–PC distance [F(1,49)
= 8.42, P < 0.006], with males (mean ± SEM = 26.84 ± 0.24) having
larger raw AC–PC distances compared to females (mean = 25.72
± 0.29). There were no Diagnosis or Sex × Diagnosis interactions
for brain volume. Brain volume, however, was significantly
dependent upon Sex (males larger, P < 0.001).
Midsagittal Area
A 2 (Sex) × 2 (Diagnosis) MANCOVA analysis confirmed signif-
icance of the AC–PC covariate [F(1,48) = 29.68, P < 0.001]. There
were no further main effects. When AC–PC was excluded as a
covariate the Sex effect approached significance [F(1,50) = 3.63,
P = 0.062], indicating the covariate corrected for the difference
in raw AC–PC distances across Sex.
Shape Parameters
A similar MANCOVA was performed to determine whether
group differences existed in parameters characterizing the
shape of the corpus callosum, including top and bottom curvat-
ure, apices of the superior and inferior callosal surfaces, the most
inferior part of the rostrum and callosal length as dependent
variables. This analysis revealed a marginal multivariate effect of
Diagnosis [F(1,48) = 3.925, P = 0.053] and a marginal multi-
variate Diagnosis × Sex interaction for all shape parameters (P =
0.096). The Diagnosis × Sex interaction, however, differed
significantly for the shape variables [F(6,288) = 2.968, P < 0.008].
The effect of Diagnosis also differed for the dependent measures
[F(6,288) = 62.489, P < 0.001]. These results justified conducting
univariate F-tests to examine Diagnosis × Sex interactions
separately for each variable listed above. Finally, the AC–PC
multivariate effect was highly significant [F(1,48) = 25.34, P <
0.001], but the covariate interacted with the different shape
parameters [F(6,288) = 13.386, P < 0.001].
Thus univariate ANOVAs were first run to determine which of
the shape parameters were significantly affected by the co-
variate. Of these, length [F(1,48) = 22.79, P < 0.001] and the most
superior limits of the top [F(1,48) = 22.24, P < 0.001] and bottom
callosal surfaces, [F(1,48) = 15.76, P < 0.001], were significantly
affected by AC–PC distance. Since the covariate did not account
for a significant portion of the variance for curvature or inferior
surface measures, use of raw AC–PC distance as a covariate was
considered inappropriate for these dependent variables and they
were reanalyzed using ANOVAs without the covariate, again
assessing Diagnosis × Group effects.
Effects of Diagnosis
There were significant effects of Diagnosis for the top and
bottom callosal surface meshes (most superior points) [F(1,48) =
7.34, P < 0.001 and F(1,48) = 6.25, P < 0.016 respectively], and
for the rostrum of the corpus callosum [F(1,48) = 4.84, P < 0.033]
with callosal location displaced in the vertical axis (y-domain) in
three-dimensional space in patients versus controls for all three
measures. Diagnosis × Sex interactions were apparent for the
curvature of top [F(1,49) = 19.49, P < 0.001] and bottom [F(1,49)
= 9.30, P < 0.004] callosal surface meshes. The superior
boundaries of bottom and top callosal surfaces showed Diagnosis
× Sex interactions even after covariate correction [F(1,48) =
7.69, P < 0.008 and F(1,48) = 8.85, P < 0.005 respectively]. In all
cases male patients had higher means ref lecting callosal dis-
placement in midbody regions. Finally, in univariate tests
assessing the assumption of equality for the effect of the
covariate on the four groups, there was a significant Diagnosis ×
Sex × Slope interaction for the superior extent of the bottom
callosal surface. This measure was reanalyzed without the
covariate, but again produced a significant Diagnosis × Sex
interaction [F(1,49) = 4.00, P < 0.05].
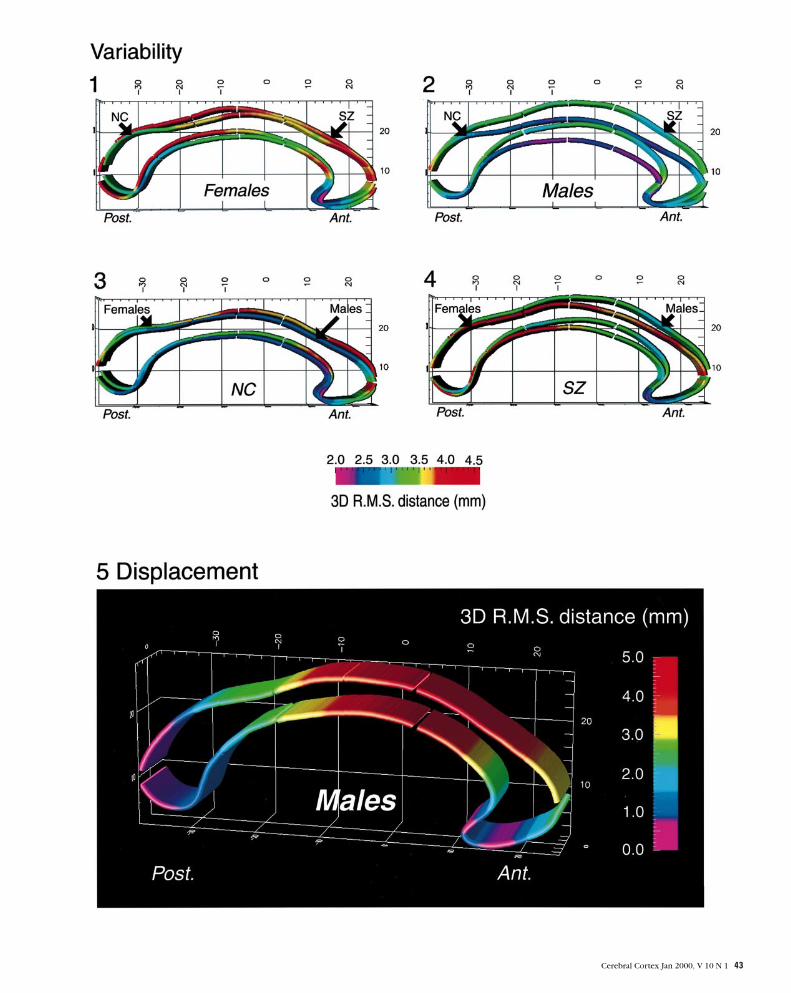
Figure 6. Parametric mesh construction. Digital points representing boundaries of the corpus callosum are redigitized to render each respective set of callosal points uniform. Analgorithm generates a parametric grid by creating a rectangular mesh over the uniform points from each callosal surface. Points from each two-dimensional surface mesh are pulledinto correspondence to represent an average callosal surface and retain intragroup variability profiles.
Figure 7. Corpus callosum partitioning protocol. Many partitioning protocols have been applied to generate comprehensive analyses of callosal area. The partitioning schemeemployed in this study divided midsagittal callosal area into anterior third, anterior midbody, posterior midbody, isthmus and splenium (adapted from Witelson et al., 1989; Clarke etal., 1994). Measures of maximal callosal width were obtained for each subdivision.
Figure 8. Brain volume and segmentation of the ventricles. To acquire brain volumes and volumes of the lateral and third ventricles, raw image data was processed in a number ofsteps. Different tissue types were classified according to signal intensity values and brain volumes were calculated. Volumes of the lateral and third ventricles were obtained by fillingin the regions of interest in native space according to neuroanatomic landmarks and CSF boundaries.
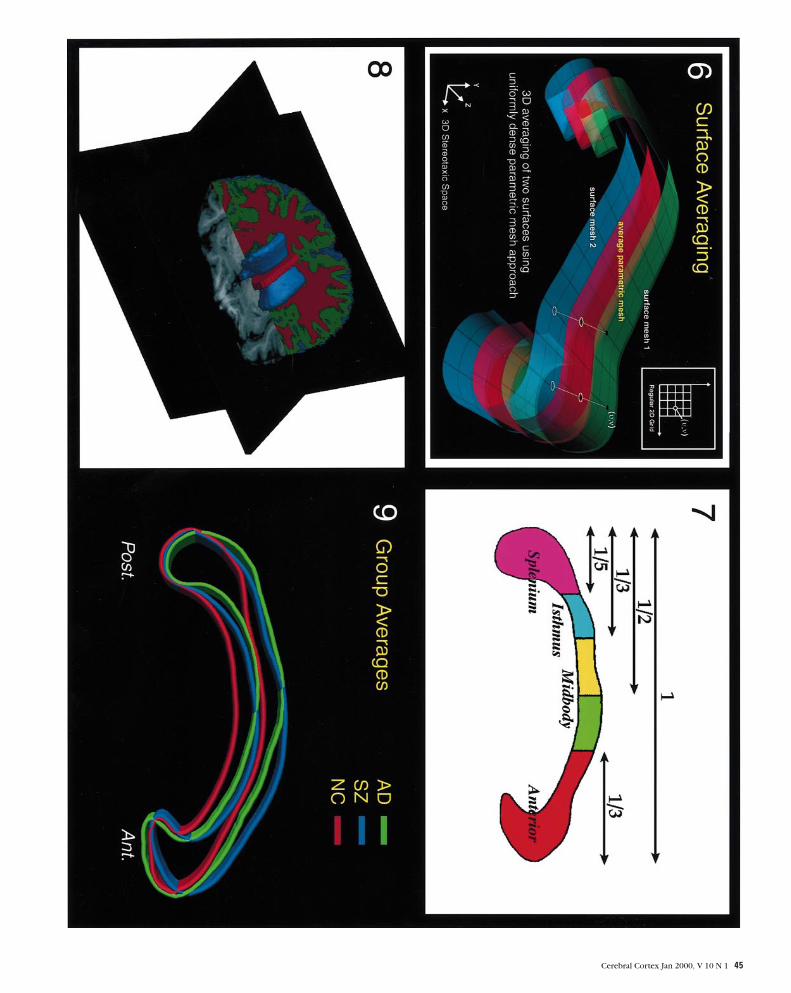
Figure 9. Color shape averages. Different shape profiles of the corpus callosum are seen in male Alzheimer’s patients (AD: green), male schizophrenic patients (SZ: blue) and maleadult controls (NC: red). The callosal renderings are taken from variability maps that represent the mean shape profiles within each group after AC–PC distance normalization instereotaxic space. The following composite was aimed at demonstrating that Alzheimer’s patients possess a mean shape profile of the corpus callosum (Thompson et al., 1998) thatis different from that of schizophrenic patients in spite of ventricular enlargement.
44 Callosal Morphology in Schizophrenia • Narr et al.
Cerebral Cortex Jan 2000, V 10 N 1 45

Diagnosis × Sex Interactions
Finally, of measures that revealed significant Diagnosis × Sex
interactions, post hoc tests of simple effects showed differences
in shape parameters between male patients and controls only. In
male groups there were significant differences in curvature for
bottom and top callosal surfaces [F(1,28) = 30.862 and 13.959
respectively, P < 0.001] with surface curvature substantially
increased in male patients. Significant differences in superior
points of top and bottom callosal surfaces [F(1,27) = 26.72, P <
0.001 and F(1,27) = 19.351, P < 0.001] also ref lected vertical
displacement of the callosum in three-dimensional space in male
patients.
Width
Measures of callosal thickness were obtained by taking the
maximum vertical width from each callosal subdivision used in
the area analysis (Fig. 7). The MANCOVA again confirmed the
significance of the AC–PC covariate [F(1,48) = 31.96, P < 0.001].
A significant multivariate Diagnosis × Sex interaction was found
[F(1,48) = 9.20, P < 0.004]. The multivariate effect of Diagnosis
differed for the dependent measures [F(4,192) = 3.47, P < 0.009].
Univariate ANOVAs revealed a significant difference across
Diagnosis for the width of the anterior callosal segment [F(1,48)
= 8.17, P < 0.006], with the width increased in schizophrenic
patients even after covarying for AC–PC distance. The Sex ×
Diagnosis interaction was significant for the splenial and anterior
callosal regions [F(1,48) = 4.58 and 6.19, P < 0.03 and P < 0.016
respectively]. A univariate F-test of simple effects in male
patients and controls showed significant group differences for
the anterior callosal segment, posterior midbody and splenium
[F(1,27) = 4.5, 8.78 and 14.44, P < 0.05, 0.006 and 0.001 respect-
ively]. In these regions male patients exhibited an increase in
callosal thickness compared to controls. There were no
significant differences in callosal widths between female groups.
Ventricular Volumes
In a 2 (Sex) × 2 (Diagnosis) MANCOVA using brain volume as
a covariate and raw lateral and third ventricle volume as
dependent variables there was a trend towards an effect of
Diagnosis [F(1,48) = 3.29, P < 0.08], but a significant Diagnosis ×
Sex interaction [F(1,48) = 4.07, P < 0.01] that also interacted
significantly with the dependent variables [F(1,48) = 5.91, P <
0.02]. There was significant effect of the covariate, but again this
differed for the dependent variables [F(1,48) = 4.06, P < 0.05].
Univariate analyses showed brain volume interacted with lateral
ventricle volume [F(1,48) = 4.09, P < 0.05], but not with third
ventricle volume. There remained, however, a significant Sex ×
Diagnosis interaction for third ventricle volume without the
covariate [F(1,48) = 4.86, P < 0.03]; covarying for brain volume,
this Sex × Diagnosis interaction was present for lateral ventricle
volumes [F(1,28) = 6.80, P < 0.01]. Tests of simple effects
revealed male patients had larger lateral ventricle volumes
[F(1,27) = 10.43, P < 0.001] and third ventricle volumes [F (1,27)
= 6.80, P < 0.01] than male controls.
Correlations
Correlations were performed between callosal shape parameters
(including curvature and vertical extents of the superior and
inferior callosal surfaces), and the volume of the lateral and third
ventricles. Significant two-tailed probability values were found
for correlations between lateral ventricle volumes and callosal
parameters, ref lecting displacement from the above analyses
(top and bottom curvature and superior limits, P < 0.01, r = 0.36,
0.52, 0.64, 0.70 respectively). Ventricular volume increases are
therefore related to increases in callosal surface curvature and
displacement. To ensure that significant correlations between
the shape measures and lateral ventricle volume were not due to
effects of AC–PC scaling or brain volume, partial correlations
between each measure, controlling separately for the effect of
AC–PC distance and for brain volume, were carried out. In all
cases the correlations remained significant upon removal of
variance associated with either AC–PC distance or brain volume.
There were no significant correlations between third ventricle
volumes and the callosal shape parameters.
ConclusionClear differences in patterns of variation and callosal shape were
observed in this morphometric analysis of the corpus callosum
between patients and controls (Figs 1–5). These differences
were clearly modulated by sex, suggesting different structural
patterns in male and female patients. Statistical tests confirmed
group differences in callosal shape parameters in three-
dimensional stereotaxic space, especially between male patients
and controls. Significant correlations ref lecting a relationship
between ventricular enlargement and callosal displacement
were evident.
Area and Width
Studies assessing standard callosal parameters in schizophrenia
have produced mixed results. MR studies report smaller (Rossi et
al., 1989; Woodruff et al., 1993), larger (Nasrallah et al., 1986;
Jacobsen et al., 1997), or no differences in callosal areas in
patients (Uematsu and Kaiya, 1988; Casanova et al., 1990; Raine
et al., 1990; Gunther et al., 1991; Woodruff et al., 1997). Studies
measuring callosal thickness have similarly reported increases
(Nasrallah et al., 1986), decreases (DeQuardo et al., 1996) and
no difference in callosal width across groups. Since the corpus
callosum connects functionally related regions, efforts have
been made to divide this structure into different areas to isolate
malformation of specific callosal channels (Thompson et al.,
1999a). Roughly equivalent estimates of the anterior corpus cal-
losum have been reported as thicker or larger in area (Nasrallah
et al., 1986) in patients, a finding compatible with ours where
maximum anterior callosal widths were greater in patients
compared to controls although were not different in area.
Gender Differences
Differences in callosal area and thickness have been found to
interact with sex. Hoff et al. (Hoff et al., 1994), for example,
reported that first-episode female patients had smaller callosal
area than female controls, partially replicating data (Hauser et
al., 1989) where chronic female schizophrenic patients were
shown to have smaller anterior callosal widths. In contrast,
Nasrallah (Nasrallah et al., 1986) found increased thickness of
anterior and middle callosal areas in females in some
concordance with Raine et al. (Raine et al., 1990) who found
anterior and posterior callosal widths smaller in male versus
female patients with the opposite effects in controls. A trend
towards a reversed sex difference in anterior and posterior
callosal size was also reported by Colombo et al. (Colombo et al.,
1993). The Diagnosis × Sex interaction in this study revealed
significant increases in widths of anterior and posterior callosal
regions in male patients, but no difference or reversal in females.
These effects appear unrelated to brain size given no effects of
diagnosis were evident and sex differences in brain volume
did not ref lect callosal width differences in female groups.
46 Callosal Morphology in Schizophrenia • Narr et al.

Moreover, our analyses show that AC–PC correction sufficiently
controls for relative differences in callosal sizes across sex.
Interestingly Highley et al. have observed an increase in callosal
fiber density in male patients versus controls and the opposite in
female groups in all regions but the posterior midbody and
splenium (Highley et al., 1999). It is possible that these changes
in fiber density could affect callosal thickness, especially for
small diameter fibers (Aboitiz et al., 1992). It is more likely,
however, that increased widths here ref lect bowing of the
callosum in male patients as anterior and posterior portions
appear more extended in three-dimensional space (Fig. 2).
Finally, many studies of callosal morphology have used low-
resolution scans that cannot represent true midline, different
partitioning protocols, and have sometimes neglected to use
head size corrections, all reasons making it difficult to compare
results across studies.
Shape
Overall, clear vertical displacements of the corpus callosum
were observed in the superior–inferior axis of three-dimensional
space in male schizophrenic patients, especially in midbody
callosal regions (Fig. 5). These results partially replicate one of
the few studies measuring callosal shape in addition to standard
morphometric parameters in schizophrenia. Specifically,
Casanova et al. used a statistical analysis of the coefficients of a
Fourier expansion series to demonstrate differences in callosal
shape in twins discordant for schizophrenia (Casanova et al.,
1990). A bowing of the corpus callosum was reported in the
superior–inferior dimension in affected twins compared to
unaffected co-twins, thought to ref lect enlargement of the
midbody of the lateral ventricle superior horns and/or third
ventricle enlargement. Furthermore using a two-dimensional
skeletonization technique, Frumin et al. report shape differ-
ences in the posterior corpus callosum (increased curvature) in
chronic schizophrenic patients that was not apparent in bipolar
patients or normal controls (Frumin et al., 1998). In our sample,
we found significant increases in lateral and third ventricular
volumes in male patients with increases in lateral ventricular
volume significantly relating to increases in callosal displace-
ment and curvature. It appears that ventricular enlargement
results in vertical displacements of the superior horn in male
patients that have been shown in stereotaxic space (Moussai et
al., 1998). In addition, a study using the same shape analyses in
pediatric temporal lobe epilepsy, autism, attention deficit
hyperactivity disorder and juvenile onset schizophrenia detected
similar displacements and bowing of the corpus callosum in the
schizophrenia group only (R. Blanton et al., personal com-
munication) even though ventricular enlargement was not
specific. To further establish whether bowing of the corpus
callosum is specific to male schizophrenic patients, we
compared callosal surface renderings from our male patients and
controls with callosal averages from Alzheimer’s patients in the
same stereotaxic space. The male Alzheimer’s patients exhibited
ventricular enlargement as well as a 24.5% decrease in posterior
midbody callosal regions (Thompson et al., 1998) (Fig. 9). While
the corpus callosum is bowed in both Alzheimer’s and schizo-
phrenic patients, distinct patterns of shape are found in each
group, indicating that the pattern of callosal displacement in
schizophrenia may indeed be disease specific.
Gender
There has been an ongoing controversy concerning sex differ-
ences in callosal morphology with an emphasis on splenial area
in normal populations (De Lacoste-Utamsing et al., 1982;
Holloway and De Lacoste, 1996; Bishop and Wahlsten, 1997;
Davatzikos and Resnick, 1998). Furthermore, callosal size has
been shown to correlate with small brain size, but not with
large brain size (Rauch and Jinkins, 1994; Jäncke et al., 1997).
In either case, gender and/or head size clearly inf luence
normal callosal morphometry, making gender interactions with
morphometric abnormalities in schizophrenic populations of
considerable interest.
There are a number of reasons why differences in callosal
shape may be more apparent in male populations. There appear
to be distinct gender differences in schizophrenic populations.
For example, negative symptoms are more prominent in male
patients and males have an earlier age of onset, a poorer quality
of life and a worse course of illness (DeLisi et al., 1989;
Waddington, 1993; Gur et al., 1996). Furthermore, not only do
clinical features in schizophrenia appear sexually dimorphic but
there appear to be relationships between neuroanatomical
volumes and specific clinical dimensions across the sexes
(Cowell et al., 1996). The curvature of the corpus callosum has
also been noted to increase with age in male populations at a
higher rate than in females (Rajapakse et al., 1996).
Clinical and psychopathological heterogeneity in schizo-
phrenic patients may also account for inconsistencies in results
when assessing structural morphology, including morphology of
the callosum (Colombo et al., 1993). For example, patients with
negative symptoms show smaller callosal sizes (Gunther et al.,
1991; Woodruff et al., 1993), thicker callosa (Coger and
Serafetinides, 1990) or no difference relative to other groups.
Similarly, early-onset schizophrenic patients have been shown to
have larger total, anterior and posterior callosal areas compared
to controls (Bigelow et al., 1983; Coger and Serafetinides, 1990;
Jacobsen et al., 1997). Clearly more information is needed to
establish the relationship of these factors to callosal morph-
ometry in schizophrenia.
Variability Maps
Results mentioned above and those from other studies have
indicated an enormous range of morphometric findings in the
corpus callosum in schizophrenic populations. Different types
of head size correction or failure to include these corrections
may contribute to discrepancies in results (Jäncke et al., 1997).
Failure to replicate findings across studies may also result from
patient heterogeneity and differences in callosal structure
between the sexes across diagnosis. In our study, variability and
displacement maps (Figs 1–5) show clear differences between
populations in patterns of callosal variation. Overall, the vari-
ability and displacement maps indicate a marked upward shift of
the corpus callosum in three-dimensional space in schizophrenic
patients that bears a relationship to lateral but not third ventricle
enlargement. Many studies have reported no inf luence of age in
their findings of callosal pathology (Thompson et al., 1998), but
few have looked at shape across age. Even though the effects of
aging and callosal development are still under investigation,
these results and those from other studies have controlled for age
to some degree. Finally, in our sample, groups were matched for
handedness. Handedness appears to be a predictor of neuro-
anatomical asymmetry and thus bears a relationship to callosal
morphology (Clarke and Zaidel, 1994).
In sum, the methods employed in this study revealed unique
differences in callosal shape and patterns of variability between
schizophrenic patients and normal controls with clear gender
differences. Further investigations are in progress, relating
Cerebral Cortex Jan 2000, V 10 N 1 47

callosal parameters to other neuroanatomical regions shown to
possess structural alterations in schizophrenic patients, such
as asymmetric perisylvian cortices and prefrontal cortices.
Furthermore, structural alterations in surrounding regions such
as thalamus and cingulate cortices may also be related to callosal
displacements. Finally, it is clear that gender inf luences callosal
morphology in schizophrenia. Larger sample sizes and homo-
geneous patient populations matched closely with control
subjects as well as correlations with symptom complexes are
required. This is necessary as it appears callosal morphology in
schizophrenic patients is tempered by a number of clinical
variables, including symptomatology, disease course and age of
onset in addition to sex, handedness and age.
NotesWe especially thank Tonmoy Sharma and his colleagues at the Institute of
Psychiatry for having the foresight to collect this data and we are grateful
to Janice M. Rayman for contributing her statistical expertise.This work
was supported by a Human Brain Project grant to the International
Consortium for Brain Mapping, funded jointly by NIMH and NIDA (P20
MH/DA52176), by a P41 Resource Grant from the NCRR (13642), by
research grants from the National Library of Medicine (LM/MH05639),
the National Science Foundation (BIR 93–22434), the NCRR (RR05056)
and NINCDS (NS38253), and research grants from the Howard Hughes
Medical Institute, United States Information Agency, and the US–UK
Fulbright Commission, London (to P.T.).
Address correspondence to Dr Arthur W. Toga, Laboratory of Neuro
Imaging, Department of Neurology, Division of Brain Mapping, UCLA
School of Medicine, 710 Westwood Plaza, Los Angeles, CA 90095–1769,
USA. Email: [email protected].
ReferencesAboitiz F, Scheibel AB, Fisher RS, Zaidel E (1992) Fiber composition of the
human corpus callosum. Brain Res 598:143–153.
American Psychiatric Association (1987) Diagnostic and statistical manual
for mental disorders (3rd edn, revised). Washington, DC: American
Psychiatric Association Press.
Annett M (1970) A classification of hand preference by association. Br J
Psychol 61:303–321.
Barta PE, Pearlson GD, Brill LB, Royall R (1997) Planum temporale
asymmetry reversal in schizophrenia: replication and relationship to
gray matter abnormalities. Am J Psychiat 154:661–667.
Bigelow LB, Nasrallah HA, Rausher FP (1983) Corpus callosum thickness
in chronic schizophrenia. Arch Gen Psychiat 142:284–287.
Bishop KM, Wahlsten, D (1997) Sex differences in the human corpus
callosum: myth or reality? Neurosci Biobehav Rev 21:581–601.
Casanova MF, Sanders RD, Goldberg TE, Bigelow LB, Christison G, Torrey
EF, Weinberger DR (1990) Morphometry of the corpus callosum in
monozygotic twins discordant for schizophrenia: a magnetic
resonance imaging study. J Neurol Neurosurg Psychiat 53:416–421.
Clarke JM, Zaidel E (1994) Anatomical–behavioral relationships: corpus
callosum morphometry and hemispheric specialization. Behav Brain
Res 64:185–202.
Colombo C, Bonfanti, Livian S, Abbruzzese M, Scarone S (1993) Size of
the corpus callosum and auditory comprehension in schizophrenics
and normal controls. Schizophr Res 11:63–70.
Coger RW, Serafetinides EA (1990) Schizophrenia, corpus callosum, and
interhemispheric transfer: a review. Psychiat Res 34:163–184.
Cowell PE, Kostianovsky DJ, Gur RC, Turetsky BI, Gur RE (1996) Sex
differences in neuroanatomical and clinical correlations in schizo-
phrenia. Am J Psychiat 153:799–805.
David AS (1994) Schizophrenia and the corpus callosum: developmental,
structural and functional relationships. Behav Brain Res 64:203–211.
Davatzikos C, Resnick SM (1998) Sex differences in anatomic measures of
interhemispheric connectivity: Correlations with cognition in women
but not men. Cereb Cortex 8:635–640.
De Lacoste-Utamsing MC, Holloway RL (1982) Sexual dimorphism in the
human corpus callosum. Science 216:1431–1432.
DeLisi LE, Dauphinais ID, Hauser P (1989) Gender differences in the
brain: are they relevant to the pathogenesis of schizophrenia. Comp
Psychiat 30:197–208.
DeQuardo JR, Bookstein FL, Green WD, Brunberg JA, Tandon R (1996)
Spatial relationships of neuroanatomic landmarks in schizophrenia.
Psychiat Res 61:81–95.
Frumin M, Golland P, McCarley RW, Hirayasu Y, Salisbury DF, Kikinis R,
Shenton M (1998) Shape differences in the corpus callosum in first
episode schizophrenia and affective disorder. ACNP meeting
(abstract).
Gunther W, Petsch R, Steinberg R, Moser E, Streck P, Heller H, Kurtz G,
Hippius H (1991) Brain dysfunction during motor activation and
corpus callosum alterations in schizophrenia measured by cerebral
blood f low and magnetic resonance imaging. Biol Psychiat
29:535–555.
Gur RE, Petty RG, Turetsky BI, Gur RC (1996) Schizophrenia throughout
life: sex differences in severity and profile of symptoms. Schizophr
Res 21: 1–12.
Hauser P, Dauphinais ID, Berrettini W, DeLisi LE, Gelernter J, Post RM
(1989) Corpus callosum dimensions measured by magnetic resonance
imaging in bipolar affective disorder and schizophrenia. Biol Psychiat
26:659–668.
Highley JR, Esiri MM, McDonald B, Cortina-Borja M, Herron BM, Crow TJ
(1999) The size and fibre composition of the corpus callosum with
respect to gender and schizophrenia: a post-mortem study. Brain
122:99–110.
Hoek HW, Brown AS, Susser E (1998) The Dutch famine and schizo-
phrenia spectrum disorders. Soc Psychiat Psychiat Epidemiol 33:
373–379.
Hoff AL, Neal C, Kushner M, DeLisi LE (1994) Gender differences in
corpus callosum size in first-episode schizophrenics. Biol Psychiat
35:913–919.
Holloway RL, De Lacoste MC (1996) Sexual dimorphism in the corpus
callosum: an extension and replication study. Hum Neurobiol
5:87–91.
Jacobsen LK, Giedd JN, Rajapakse JC, Hamberger SD, Vaituzis AC, Frazier
JA, Lenane MC, Rapoport JL (1997) Quantitative magnetic resonance
imaging of the corpus callosum in childhood onset schizophrenia.
Psychiat Res 68:77–86.
Jäncke L, Staiger JF, Schlaug G, Huang Y, Steinmetz H (1997) The
relationship between corpus callosum size and forebrain volume.
Cereb Cortex 7:48–56.
Kikinis R, Shenton ME, Gerig G, Hokama H, Haimson J, O’Donnell BF,
Wible CG, McCarley RW, Jolesz FA (1994) Temporal lobe sulco-gyral
pattern anomalies in schizophrenia: an in vivo MR three-dimensional
surface rendering study. Neurosci Lett 182:7–12.
Lawrie SM, Abukmeil SS (1998) Brain abnormality in schizophrenia. Br J
Psychiat 172:110–120.
Office of Population Censuses and Surveys (1991) Classification of
Occupations. Standard Occupational Classification (SOS). London:
HMSO.
Mazziotta JC, Toga AW, Evans AC, Fox P, Lancaster J (1995) A probabilistic
atlas of the human brain: theory and rationale for its development.
NeuroImage 2:89–101.
Moussai J, Sharma T, Anvar BA, Narr KL, Cannestra AF, Thompson PM,
Toga AW (1998) 3-Dimensional analysis of lateral ventricles in
schizophrenia. 4th International Conference on Functional Mapping
of the Human Brain. NeuroImage 7:S505.
Nasrallah HA, Andreasen NC, Coffman JA, Olson SC, Dunn VD, Ehrhardt
JC, Champman SM (1986) A controlled magnetic resonance imaging
study of corpus callosum thickness in schizophrenia. Biol Psychiat
21:3:274–282.
Njiokiktjien C, de Donneville L, Vaal J (1994) Callosal size in children with
learning disabilities. Behav Brain Res 64:213–218.
Raine A, Harrison GN, Reynolds GP, Sheard C, Cooper JE, Meddley I
(1990) Structural and functional characteristics of the corpus
callosum in schizophrenics, psychiatric controls, and in normals.
Arch Gen Psychiat 471060–1064.
Rajapakse JC, Geidd JN, Rumsey JM, Vaituzis AC, Hamburger SD,
Rapoport JL (1996) Regional MRI measurements of the corpus
callosum: a methodological and developmental study. Brain Devel
18:379–388
Rauch R A, Jinkins JR (1994) Analysis of cross-sectional area
measurements of the corpus callosum adjusted for brain size in male
and female subjects from childhood to adulthood. Behav Brain Res
64:65–78.
48 Callosal Morphology in Schizophrenia • Narr et al.

Rossi A, Stratta P, Gallucci M, Passariello R, Casacchia M (1989)
Quantification of corpus callosum and ventricles in schizophrenia
with nuclear magnetic resonance imaging: a pilot study. Am J Psychiat
146:99–101.
Sharma T, du Boulay G, Lewis S, Sigmundsson T, Gurling H, Murray R
(1997). The Maudsley Family Study. I: Structural brain changes on
magnetic resonance imaging in familial schizophrenia. Progr
Neuro-Psychopharmacol Biol Psychiat 21:1297–1315.
Sharma T, Lancaster E, Lee D, Lewis S, Sigmundsson T, Takei N, Gurling
H, Barta P, Pearlson G, Murray R (1998). Brain changes in
schizophrenia volumetric MRI study of families multiply affected
with schizophrenia — the Maudsley Family study 5. Br J Psychiat
173:132–138.
Sowell ER, Thompson PM, Holmes CJ, Batth R, Jernigan TL, Toga AW
(1999) Localizing age-related changes in brain structure between
childhood and adolescence using statistical parametric mapping.
NeuroImage 9:587–597.
Talairach J, Tournoux P (1988) Co-planar stereotaxic atlas of the human
brain. New York: Thieme.
Thompson P, Toga AW (1996a) A surface-based technique for warping
three-dimensional images of the brain. IEEE Trans Med Imag 15:
402–417.
Thompson PM, Schwartz C, Toga AW (1996b) High-resolution random
mesh algorithms for creating a probabilistic 3D surface atlas of the
human brain. NeuroImage 3:19–34.
Thompson PM, MacDonald D, Mega MS, Holmes CJ, Evans AC, Toga AW
(1997) Detection and mapping of abnormal brain structure with a
probabilistic atlas of cortical surfaces. J Comput Assist Tomogr
21:567–581.
Thompson PM, Moussai J, Zohoori S, Goldkorn A, Khan AA, Mega MS,
Small GW, Toga AW (1998) Cortical variability and asymmetry in
normal aging and Alzheimer’s disease. Cereb Cortex 8:492–509.
Thompson PM, Narr KL, Blanton RE, Toga AW (1999a) In: Mapping
structural alterations of the corpus callosum during brain develop-
ment and disease (Iacoboni M, Zaidel E, eds). Cambridge, MA: MIT
Press (in press) [review].
Thompson PM, Woods RP, Mega MS, Toga AW (1999) Mathematical/
computational challenges in creating deformable and probabilistic
brain atlases. Hum Brain Map 9 (in press).
Uematsu M, Kaiya H (1988) The morphology of the corpus callosum in
schizophrenia: a MRI study. Schizophr Res 1:391–398.
Waddington JL (1993) Neurodynamics of abnormalities in cerebral
metabolism and structure in schizophrenia. Schizophr Bull 19:55–58.
Witelson SF (1989) Hand and sex differences in the isthmus and genu of
the human corpus callosum: a postmortem morphological study.
Brain 112:799–835.
Woodruff PWR, Pearlson GD, Geer MJ, Barta PE, Chilcoat HD (1993) A
computerized imaging study of corpus callosum morphology in
schizophrenia. Psychol Med 23:45–56.
Woodruff PWR, McManus IC, David AS (1995) Meta-analysis of corpus
callosum size in schizophrenia. J Neurol Neurosurg Psychiat
58:457–461.
Woodruff PWR, Phillips ML, Rushe T, Wright RC, Murray RM, David AS
(1997) Corpus callosum size and inter-hemispheric function in
schizophrenia. Schizophr Res 23:189–196.
Woodruff PWR, Wright IC, Shurique N, Russouw H, Rushe T, Howard RJ,
Graves M, Bullmore ET, Murray RM (1997b) Structural brain abnor-
malities in male schizophrenics ref lect fronto-temporal dissociation.
Psychol Med 27:1257–1266.
Zaidel DW, Esiri MM, Harrison PJ (1997) Size, shape, and orientation of
neurons in the left and right hippocampus: investigation of normal
asymmetries and alterations in schizophrenia. Am J Psychiat
154:812–818.
Zipursky RB, Lim KO, Sullivan EV, Brown BW, Pfefferbaum A (1992)
Widespread cerebral gray matter volume deficits in schizophrenia.
Arch Gen Psychiat 49:195–205.
Cerebral Cortex Jan 2000, V 10 N 1 49
Copyright © 2022 FDOKUMEN




















